
Qri2/Nse4, a component of the essential Smc5/6 DNA repair complex: Nse4, an essential cell cycle...
16
Molecular Microbiology (2005) 55(6), 1735–1750 doi:10.1111/j.1365-2958.2005.04531.x © 2005 Blackwell Publishing Ltd Blackwell Science, LtdOxford, UKMMIMolecular Microbiology0950-382XBlackwell Publishing Ltd, 2005 ? 200555617351750Original ArticleNse4, an essential cell cycle proteinB. Hu et al. Accepted 14 December, 2004. *For correspondence. E-mail [email protected]; Tel. (+44) 207 848 4003; Fax (+44) 207 848 4003. Qri2/Nse4, a component of the essential Smc5/6 DNA repair complex Bin Hu, 1 Chunyan Liao, 1 Stefan H. Millson, 2 Mehdi Mollapour, 2 Chrisostomos Prodromou, 3 Laurence H. Pearl, 3 Peter W. Piper 2 and Barry Panaretou 1 * 1 Department of Life Sciences, King’s College London, London SE1 9NN, UK. 2 Department of Molecular Biology and Biotechnology, The University of Sheffield, Sheffield S10 2TN, UK. 3 Section of Structural Biology, Chester Beatty Laboratories, The Institute of Cancer Research, London SW3 6JB, UK. Summary We demonstrate a role for Qri2 in the essential DNA repair function of the Smc5/6 complex in Saccharo- myces cerevisiae . We generated temperature- sensitive ( ts ) mutants in QRI2 and characterized their properties. The mutants arrest after S phase and prior to mitosis. Furthermore, the arrest is dependant on the Rad24 checkpoint, and is also accompanied by phosphorylation of the Rad53 checkpoint effector kinase. The mutants also display genome instability and are sensitive to agents that damage DNA. Two- hybrid screens reveal a physical interaction between Qri2 and proteins that are n on-S mc e lements of the Smc5/6 DNA repair complex, which is why we pro- pose the name NSE4 for the open reading frame pre- viously known as QRI2 . A key role for Nse4 in Smc5/ 6 function is likely, as overexpressing known subunits of the Smc5/6 complex suppresses nse4 ts cell cycle arrest. The nse4 ts growth arrest is non-lethal and unlike the catastrophic nuclear fragmentation pheno- type of smc6 ts mutants, the nucleus remains intact; replicative intermediates and sheared DNA are not detected. This could imply a role for Nse4 in mainte- nance of higher order chromosome structure. Introduction In the budding yeast Saccharomyces cerevisiae around 120 genes are expressed towards the end of G1. This group of genes is referred to as the CLN2 cluster, after one of its most conspicuous members (Spellman et al ., 1998). This cluster includes sets of genes involved in DNA synthesis ( CDC6 , POL1 , RNR1 ), DNA repair ( SMC6 , RAD27 ), sister chromatid cohesion ( SMC1 , SMC3 ), spindle pole body duplication ( TUB4 , SPC42 ) and bud emergence ( SVS1 , BN14 ) (Spellman et al ., 1998). QRI2 is an essential S. cerevisiae open reading frame (ORF) of unknown function that is a member of the CLN2 cluster. This ORF’s name reflects its location next to QRI1 on chromosome 4, and is not a descriptor of the function of the corresponding protein. We show that Qri2 is essential for cell cycle progression, and that it is part of the essential Smc5/6 DNA repair complex. The SMC (s tructural m aintainance of c hromosomes) family of pro- teins play essential roles in genome integrity. There are six classes of Smc protein in eukaryotes, forming three pairs of heterodimers. Each pair associates with addi- tional non-Smc proteins to form high molecular mass complexes. Qri2 is a n on-S mc e lement that associates with the Smc5/6 complex, and in keeping with the estab- lished nomenclature we have renamed this ORF as NSE4 , and refer to the ORF using this name from now on. The name change has been agreed with the group that originally deleted QRI2 (Simon et al ., 1994). We have registered NSE4 with the Saccharomyces Genome Data- base (SGD) – as an alias for QRI2 . NSE4 is an essential S. cerevisiae ORF that is con- served throughout the eukaryotic lineage. Like many genes expressed at the G1/S transition, NSE4 contains a perfect MCB ( Mlu I cell cycle box) element in its promoter. Transcription from MCB elements is controlled by MBF (MCB binding factor), a heterodimeric transcription factor composed of Swi1, which is tethered to DNA by Swi4. MBF mediated transcription activation is cell cycle regu- lated as MBF itself is activated post-translationally by Cln3-Cdc28 (Valdivieso et al ., 1993). Also, microarray pro- filing shows that NSE4 transcription is extensively repressed in a time-dependent manner when cells are arrested in G1 by a -factor mating pheromone (Roberts et al ., 2000). Periodic variation of transcript levels through the cell cycle, and rapid transcriptional inhibition after cell cycle arrest, imply that Nse4 is required for passage through the cell cycle. Furthermore, the recent proteome wide localization of yeast proteins reveals that Nse4 is a nuclear protein (Huh et al ., 2003). This is in keeping with the location of most of the proteins encoded by the CLN2
-
Upload
independent -
Category
Documents
-
view
1 -
download
0
Transcript of Qri2/Nse4, a component of the essential Smc5/6 DNA repair complex: Nse4, an essential cell cycle...

Molecular Microbiology (2005)
55
(6), 1735–1750 doi:10.1111/j.1365-2958.2005.04531.x
© 2005 Blackwell Publishing Ltd
Blackwell Science, LtdOxford, UKMMIMolecular Microbiology0950-382XBlackwell Publishing Ltd, 2005
? 2005
55
617351750
Original Article
Nse4, an essential cell cycle proteinB. Hu
et al.
Accepted 14 December, 2004. *For correspondence. [email protected]; Tel. (
+
44) 207 848 4003; Fax (
+
44)207 848 4003.
Qri2/Nse4, a component of the essential Smc5/6 DNA repair complex
Bin Hu,
1
Chunyan Liao,
1
Stefan H. Millson,
2
Mehdi Mollapour,
2
Chrisostomos Prodromou,
3
Laurence H. Pearl,
3
Peter W. Piper
2
and Barry Panaretou
1
*
1
Department of Life Sciences, King’s College London, London SE1 9NN, UK.
2
Department of Molecular Biology and Biotechnology, The University of Sheffield, Sheffield S10 2TN, UK.
3
Section of Structural Biology, Chester Beatty Laboratories, The Institute of Cancer Research, London SW3 6JB, UK.
Summary
We demonstrate a role for Qri2 in the essential DNArepair function of the Smc5/6 complex in
Saccharo-myces cerevisiae
. We generated temperature-sensitive (
ts
) mutants in
QRI2
and characterized theirproperties. The mutants arrest after S phase and priorto mitosis. Furthermore, the arrest is dependant onthe Rad24 checkpoint, and is also accompanied byphosphorylation of the Rad53 checkpoint effectorkinase. The mutants also display genome instabilityand are sensitive to agents that damage DNA. Two-hybrid screens reveal a physical interaction betweenQri2 and proteins that are non-Smc elements of theSmc5/6 DNA repair complex, which is why we pro-pose the name
NSE4
for the open reading frame pre-viously known as
QRI2
. A key role for Nse4 in Smc5/6 function is likely, as overexpressing known subunitsof the Smc5/6 complex suppresses
nse4
ts
cell cyclearrest. The
nse4
ts
growth arrest is non-lethal andunlike the catastrophic nuclear fragmentation pheno-type of
smc6
ts
mutants, the nucleus remains intact;replicative intermediates and sheared DNA are notdetected. This could imply a role for Nse4 in mainte-nance of higher order chromosome structure.
Introduction
In the budding yeast
Saccharomyces cerevisiae
around120 genes are expressed towards the end of G1. Thisgroup of genes is referred to as the
CLN2
cluster, after
one of its most conspicuous members (Spellman
et al
.,1998). This cluster includes sets of genes involved inDNA synthesis (
CDC6
,
POL1
,
RNR1
), DNA repair(
SMC6
,
RAD27
), sister chromatid cohesion (
SMC1
,
SMC3
), spindle pole body duplication (
TUB4
,
SPC42
)and bud emergence (
SVS1
,
BN14
) (Spellman
et al
.,1998).
QRI2
is an essential
S. cerevisiae
open readingframe (ORF) of unknown function that is a member of the
CLN2
cluster. This ORF’s name reflects its location nextto
QRI1
on chromosome 4, and is not a descriptor of thefunction of the corresponding protein. We show that Qri2is essential for cell cycle progression, and that it is part ofthe essential Smc5/6 DNA repair complex. The SMC(structural maintainance of chromosomes) family of pro-teins play essential roles in genome integrity. There aresix classes of Smc protein in eukaryotes, forming threepairs of heterodimers. Each pair associates with addi-tional non-Smc proteins to form high molecular masscomplexes. Qri2 is a non-Smc element that associateswith the Smc5/6 complex, and in keeping with the estab-lished nomenclature we have renamed this ORF as
NSE4
, and refer to the ORF using this name from now on.The name change has been agreed with the group thatoriginally deleted
QRI2
(Simon
et al
., 1994). We haveregistered
NSE4
with the
Saccharomyces
Genome Data-base (SGD) – as an alias for
QRI2
.
NSE4
is an essential
S. cerevisiae
ORF that is con-served throughout the eukaryotic lineage. Like manygenes expressed at the G1/S transition,
NSE4
contains aperfect MCB (
Mlu
I cell cycle box) element in its promoter.Transcription from MCB elements is controlled by MBF(MCB binding factor), a heterodimeric transcription factorcomposed of Swi1, which is tethered to DNA by Swi4.MBF mediated transcription activation is cell cycle regu-lated as MBF itself is activated post-translationally byCln3-Cdc28 (Valdivieso
et al
., 1993). Also, microarray pro-filing shows that
NSE4
transcription is extensivelyrepressed in a time-dependent manner when cells arearrested in G1 by
a
-factor mating pheromone (Roberts
et al
., 2000). Periodic variation of transcript levels throughthe cell cycle, and rapid transcriptional inhibition after cellcycle arrest, imply that Nse4 is required for passagethrough the cell cycle. Furthermore, the recent proteomewide localization of yeast proteins reveals that Nse4 is anuclear protein (Huh
et al
., 2003). This is in keeping withthe location of most of the proteins encoded by the
CLN2

1736
B. Hu
et al.
© 2005 Blackwell Publishing Ltd,
Molecular Microbiology
,
55
, 1735–1750
cluster genes, as they play roles predominantly in DNAreplication and repair.
The systematic yeast gene deletion project showed that
Nse4
is essential (Simon
et al
., 1994). In this report, wepresent functional analysis of Nse4. We show that cellsrequire this protein for the metaphase to anaphase tran-sition. More specifically, we demonstrate that cells lackingfunctional Nse4 arrest in a manner that is dependent onactivation of a DNA damage checkpoint. We show thatNse4 plays a role in repairing chromosomal aberrationsthat appear during normal progression of the cell cycle,at a point after DNA replication. Furthermore, we identifythe DNA repair complex that it is abrogated by loss ofNse4 function.
NSE4
has been conserved throughout the eukaryoticlineage, with murine, worm, fly and plant orthologuesidentified by
BLAST
searches. This is not surprising giventhe overall conservation of all three SMC heterodimersand their non-SMC components. In humans there are twoNse4 orthologues – the product of the AAH27612 locus(NCBI Accession AAH27612) and FLJ20003 (NCBIAccession NP_060085). EST databases reveal the latteris expressed in most (if not all) human tissues; an align-ment of Nse4 with FLJ20003 (revealing 22% similarity) ispresented in Fig. 1 (the other human orthologue and amurine orthologue are also included in the alignment).Analysis of the primary sequence of Nse4 does not revealinformation about its biochemical properties. Our func-
tional characterization of this protein was performed byassessing the arrest caused by conditional lethal allelesof
NSE4
.
Results
Isolation of
NSE4
temperature-sensitive (
ts
) mutants
We used hydroxylamine mutagenesis and the plasmidshuffle technique to isolate point mutations in
NSE4
thatconfer a
ts
growth phenotype. We initially constructed ahaploid yeast strain (BP12 in Table 1) in which
NSE4
wasdeleted from the genome.
NSE4
is an essential gene, soviability was maintained by a copy of this gene controlledby its native promoter, on a centromeric
URA3
vector. Thisstrain was used to select for
ts nse4
mutants, generatedby hydroxylamine treatment of a centromeric
LEU2
vectorbearing
NSE4
expressed from its native promoter. Eight
ts
mutants were isolated – and the Nse4 coding regionswere sequenced. Some of the mutants were isolated morethan once, giving four unique mutants (
nse4-[1–4]
ts
). Asexpected, the mutants resulted from G to A or C to Ttransitions, giving the following amino acid substitutions:
nse4-1
ts
D103N, R127K and S195F;
nse4-2
ts
S*329D;
nse4-3
ts
G*224D;
nse4-4
ts
G*224S and S231D (the resi-dues marked with an asterisk are conserved in the
S.cerevisiae
, murine and human Nse4 orthologues). All fourmutants were isolated by virtue of their inability to grow at
Fig. 1.
Structural and functional conservation of Nse4.A. Alignment of Nse4 from
S. cerevisiae,
against murine (NCBI Accession NP_079775) and two human orthologues: (i) FLJ20003; NCBI Accession NP_060085 and (ii) product of locus AAH27612 (NCBI Accession AAH27612). The amino acid sequence of the yeast orthologue is numbered. Sites mutated in
ts
mutants are underlined [
nse4-1
ts
D103N R127K and S195F;
nse4-2
ts
S329D;
nse4-3
ts
G224D;
nse4-4
ts
G224S and S231D; (*) conserved residues; (:) strongly similar residues; (.) residues of weak similarity].

Nse4, an essential cell cycle protein
1737
© 2005 Blackwell Publishing Ltd,
Molecular Microbiology
,
55
, 1735–1750
37
∞
C (Fig. 2A). At 25
∞
C, however,
nse4-3
ts
and
nse4-4
ts
cells grew at the same rate as cells bearing the wild-typegene, whereas
nse4-1
ts
and
nse4-2
ts
exhibited a slowgrowth phenotype, with doubling times of 3 and 4 h,respectively, in contrast to the 2 h doubling time of wild-type. For each mutant, the
ts
phenotype does not resultin cell death at the non-permissive temperature, as return-ing the cells to 25
∞
C results in restoration of growth – thephenotype at the non-permissive temperature is thereforea growth arrest (Fig. 2B).
nse4
ts
mutants arrest at G2/M
We characterized the growth defect by flow cytometry[fluorescence activated cell sorting (FACS)], examiningthe DNA content of
nse4
ts
mutant cells at non-permissiveconditions. Asynchronous log phase cultures grown at25
∞
C were shifted to 37
∞
C and samples were taken every3 h. Cells were stained with propidum iodide and cell cycledistribution was assessed by FACS. After 3 h at 37
∞
C, allfour mutants arrested with a 2C DNA content, indicatingarrest between late S phase and prior to the exit frommitosis (Fig. 3A). We subsequently assessed the cellulardistribution of DNA, the state of the mitotic spindle and
cell morphology. Exponential phase cultures of wild-typeand all four
nse4
ts
mutants grown at 25
∞
C were shifted to37
∞
C for 6 h;
>
80% of
nse4
ts
cells were arrested with largebuds and a short bipolar mitotic spindle, the 2C nucleuswas located near the bud neck (Fig. 3B).
Flow cytometry cannot be used to distinguish betweena block in late S phase from a block at G2/M boundary(Chakraverty
et al
., 2001). TOP3, CDC17, HYS2, RFC2are all genes involved in DNA synthesis, but conditionalmutations in these genes are ‘leaky’ and FACS analysisshows they arrest with a G2/M content of DNA under non-permissive conditions, instead of the expected arrest in(or just prior to) S phase (Chakraverty et al., 2001; Sug-imoto et al., 1995; Hennessy et al., 1991; Noskov et al.,1998). Although our FACS data suggests that the bulk ofDNA appears to be replicated in nse4ts mutants undernon-permissive conditions, there could be significantamounts of unreplicated DNA, sufficient to cause cellcycle arrest via activation of the replication checkpoint orvia a defect in DNA replication per se. To determinewhether DNA was completely replicated, chromosomes ofarrested nse4ts cells were analysed by pulse f ield gelelectrophoresis (PFGE). In this assay, only fully replicatedDNA enters a gel – whereas incompletely replicated DNA,
Table 1. S. cerevisiae strains used in this study.
Strain Genotype Source
BP12 MATa; ura3-52; leu2D1; nse3::kanMX4; pBP01 (NSE3/URA3) This studyBH01 MATa; ura3-52; leu2D1; nse3::kanMX4; pBH1 (nse3-1ts/URA3) This studyBH02 MATa; ura3-52; leu2D1; nse3::kanMX4; pBH2 (nse3-2ts/URA3) This studyBH03 MATa; ura3-52; leu2D1; nse3::kanMX4; pBH3 (nse3-3ts/URA3) This studyBH04 MATa; ura3-52; leu2D1; nse3::kanMX4; pBH4 (nse3-4ts/URA3) This studyBH21 MATa; ura3D0; leu2D0; his3D1; nse3::HygB; pBH1 (nse3-1ts/URA3) This studyBH30 MATa; ura3D0; leu2D0; his3D1; lys2D0; rad24::kanMX4; nse3::HygB; pBH1 (nse3-1ts/URA3) This studyBH31 MATa; ura3-52; leu2D1; nse3::kanMX4; pBP01 (NSE3/URA3); YEplac181 (LEU2) This studyBH32 MATa; ura3-52; leu2D1; nse3::kanMX4; pBH1 (nse3-1ts/URA3); YEplac181 (LEU2) This studyBH33 MATa; ura3-52; leu2D1; nse3::kanMX4; pBH1 (nse3-1ts/URA3); pBH101 (YDR288W/LEU2) This studyBH34 MATa; ura3-52; leu2D1; nse3::kanMX4; pBH1 (nse3-1ts/URA3); pBH102 (MMS21/LEU2) This studyBH35 MATa; ura3-52; leu2D1; nse3::kanMX4; pBH1 (nse3-1ts/URA3); pBH103 (NSE1/LEU2) This studyBH36 MATa; ura3-52; leu2D1; nse3::kanMX4; pBH1 (nse3-1ts/URA3); pBH104 (SMC5/LEU2) This studyBH37 MATa; ura3-52; leu2D1; nse3::kanMX4; pBH1 (nse3-1ts/URA3); pBH104 (RHC18(SMC6)/LEU2) This studyBH38 MATa; ura3-52; leu2D1; nse3::kanMX4; pBH4 (nse3-4ts/URA3); YEplac181 (LEU2) This studyBH39 MATa; ura3-52; leu2D1; nse3::kanMX4; pBH4 (nse3-4ts/URA3); pBH10 1 (YDR288W/LEU2) This studyBH40 MATa; ura3-52; leu2D1; nse3::kanMX4; pBH4 (nse3-4ts/URA3); pBH102 (MMS21/LEU2) This studyBH41 MATa; ura3-52; leu2D1; nse3::kanMX4; pBH4 (nse3-4ts/URA3); pBH103 (NSE1/LEU2) This studyBH42 MATa; ura3-52; leu2D1; nse3::kanMX4; pBH4 (nse3-4ts/URA3); pBH104 (SMC5/LEU2) This studyBH43 MATa; ura3-52; leu2D1; nse3::kanMX4; pBH4 (nse3-4ts/URA3); pBH104 (RHC18(SMC6)/LEU2) This studyBH45 MATa; ura3-52; leu2D1; nse3::kanMX4; pBH2 (nse3-2ts/URA3); YEplac181 (LEU2) This studyBH46 MATa; ura3-52; leu2D1; nse3::kanMX4; pBP01 (NSE3/URA3) pDK243 (1ARS/LEU2/ADE3) This studyBH47 MATa; ura3-52; leu2D1; nse3::kanMX4; pBP01 (NSE3/URA3) pDK368-7 (7ARS/LEU2/ADE3) This studyBH48 MATa; ura3-52; leu2D1; nse3::kanMX4; pBH2 (nse3-2ts/URA3); pDK243 (1ARS/LEU2/ADE3) This studyBH49 MATa; ura3-52; leu2D1; nse3::kanMX4; pBH2 (nse3-2ts/URA3); pDK368-7 (7ARS/LEU2/ADE3) This studyPJ69-4a MATaaaa; trp1-901; leu2-3112; ura3-52; his3-200; gal4D; gal80D; LYS2::GAL1-HIS3; GAL2-ADE2;
met2::GAL7-lacZUetz et al. (2000)
10590D Mata/a; ura3-52/ura3-52; his3D200/HIS3; leu2D1/LEU2; LYS2/LYS2; trp1D63/TRP1; nse4(YDL105W)::kanMX4/YDL105w
EUROSCARF
Y23802 Mata/a; his3D1/his3D1; leu2D0/leu2D0; lys2D0/LYS2; MET15/met15D0; ura3D0/ura3D0; nse4(YDL105W)::kanMX4/NSE4(YDL105W)
EUROSCARF
Y16169 Mata; his3D1; leu2D0; lys2D0; ura3D0; YER173W(rad24)::kanMX4 EUROSCARF
EUROSCARF, European Saccharomyces cerevisiae Archive for Functional analysis.
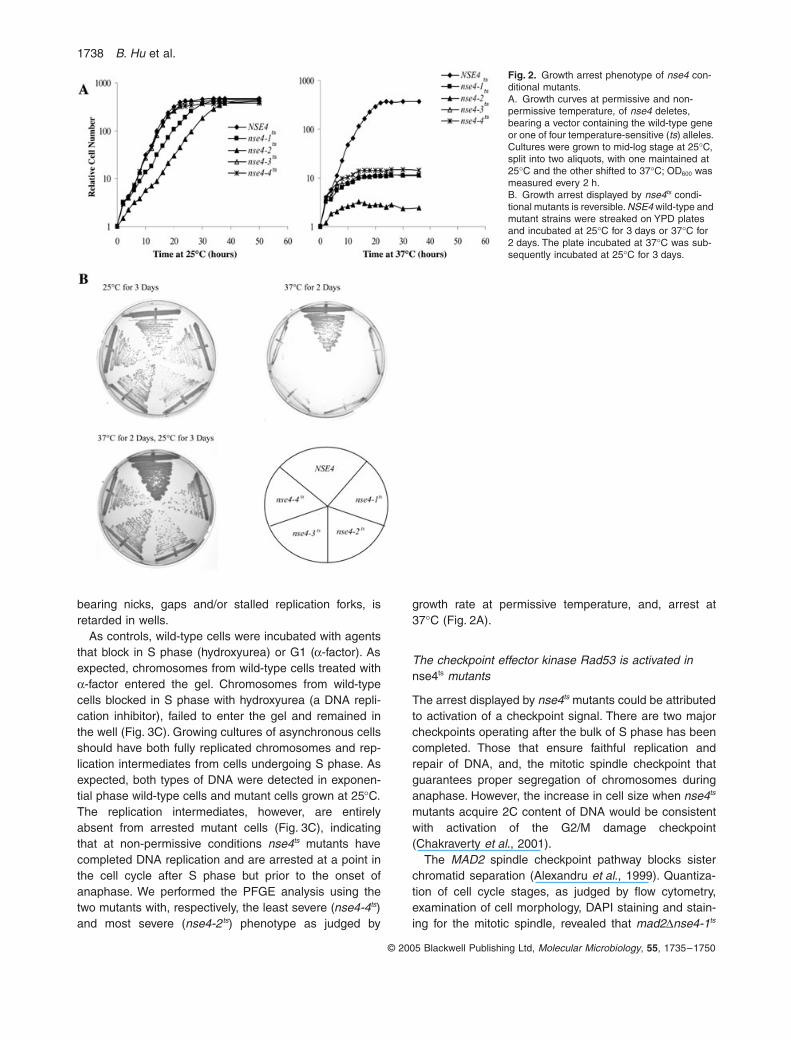
1738 B. Hu et al.
© 2005 Blackwell Publishing Ltd, Molecular Microbiology, 55, 1735–1750
bearing nicks, gaps and/or stalled replication forks, isretarded in wells.
As controls, wild-type cells were incubated with agentsthat block in S phase (hydroxyurea) or G1 (a-factor). Asexpected, chromosomes from wild-type cells treated witha-factor entered the gel. Chromosomes from wild-typecells blocked in S phase with hydroxyurea (a DNA repli-cation inhibitor), failed to enter the gel and remained inthe well (Fig. 3C). Growing cultures of asynchronous cellsshould have both fully replicated chromosomes and rep-lication intermediates from cells undergoing S phase. Asexpected, both types of DNA were detected in exponen-tial phase wild-type cells and mutant cells grown at 25∞C.The replication intermediates, however, are entirelyabsent from arrested mutant cells (Fig. 3C), indicatingthat at non-permissive conditions nse4ts mutants havecompleted DNA replication and are arrested at a point inthe cell cycle after S phase but prior to the onset ofanaphase. We performed the PFGE analysis using thetwo mutants with, respectively, the least severe (nse4-4ts)and most severe (nse4-2ts) phenotype as judged by
growth rate at permissive temperature, and, arrest at37∞C (Fig. 2A).
The checkpoint effector kinase Rad53 is activated in nse4ts mutants
The arrest displayed by nse4ts mutants could be attributedto activation of a checkpoint signal. There are two majorcheckpoints operating after the bulk of S phase has beencompleted. Those that ensure faithful replication andrepair of DNA, and, the mitotic spindle checkpoint thatguarantees proper segregation of chromosomes duringanaphase. However, the increase in cell size when nse4ts
mutants acquire 2C content of DNA would be consistentwith activation of the G2/M damage checkpoint(Chakraverty et al., 2001).
The MAD2 spindle checkpoint pathway blocks sisterchromatid separation (Alexandru et al., 1999). Quantiza-tion of cell cycle stages, as judged by flow cytometry,examination of cell morphology, DAPI staining and stain-ing for the mitotic spindle, revealed that mad2Dnse4-1ts
Fig. 2. Growth arrest phenotype of nse4 con-ditional mutants.A. Growth curves at permissive and non-permissive temperature, of nse4 deletes, bearing a vector containing the wild-type gene or one of four temperature-sensitive (ts) alleles. Cultures were grown to mid-log stage at 25∞C, split into two aliquots, with one maintained at 25∞C and the other shifted to 37∞C; OD600 was measured every 2 h.B. Growth arrest displayed by nse4ts condi-tional mutants is reversible. NSE4 wild-type and mutant strains were streaked on YPD plates and incubated at 25∞C for 3 days or 37∞C for 2 days. The plate incubated at 37∞C was sub-sequently incubated at 25∞C for 3 days.
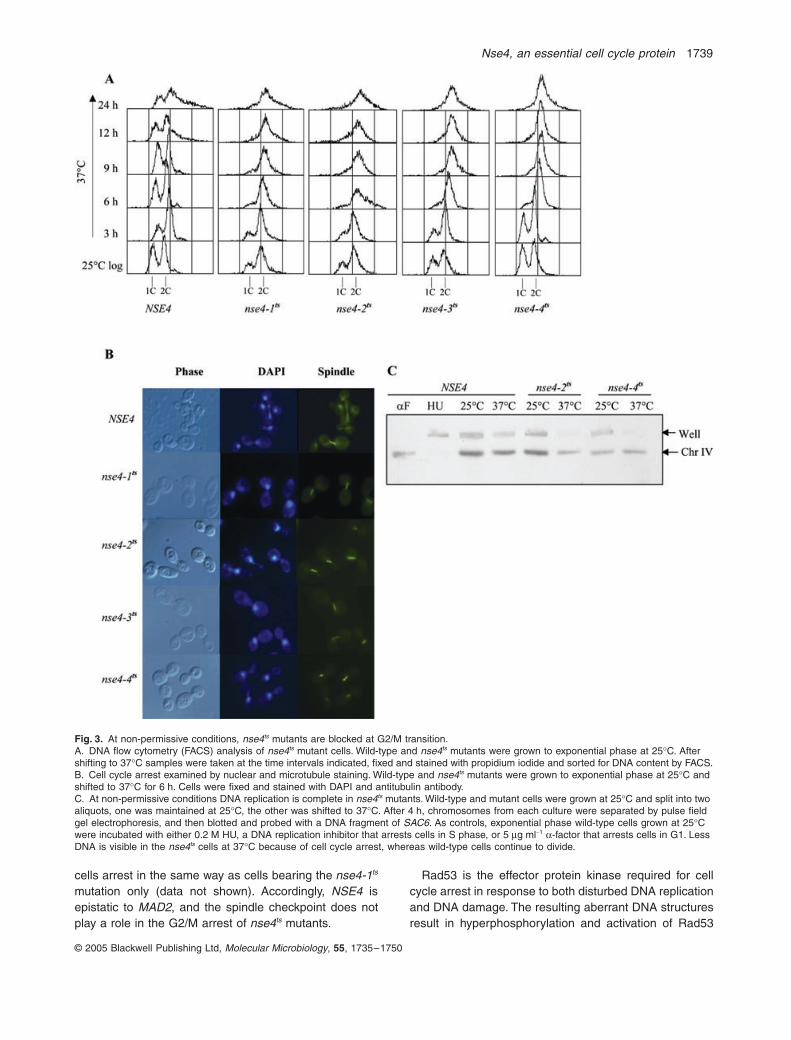
Nse4, an essential cell cycle protein 1739
© 2005 Blackwell Publishing Ltd, Molecular Microbiology, 55, 1735–1750
cells arrest in the same way as cells bearing the nse4-1ts
mutation only (data not shown). Accordingly, NSE4 isepistatic to MAD2, and the spindle checkpoint does notplay a role in the G2/M arrest of nse4ts mutants.
Rad53 is the effector protein kinase required for cellcycle arrest in response to both disturbed DNA replicationand DNA damage. The resulting aberrant DNA structuresresult in hyperphosphorylation and activation of Rad53
Fig. 3. At non-permissive conditions, nse4ts mutants are blocked at G2/M transition.A. DNA flow cytometry (FACS) analysis of nse4ts mutant cells. Wild-type and nse4ts mutants were grown to exponential phase at 25∞C. After shifting to 37∞C samples were taken at the time intervals indicated, fixed and stained with propidium iodide and sorted for DNA content by FACS.B. Cell cycle arrest examined by nuclear and microtubule staining. Wild-type and nse4ts mutants were grown to exponential phase at 25∞C and shifted to 37∞C for 6 h. Cells were fixed and stained with DAPI and antitubulin antibody.C. At non-permissive conditions DNA replication is complete in nse4ts mutants. Wild-type and mutant cells were grown at 25∞C and split into two aliquots, one was maintained at 25∞C, the other was shifted to 37∞C. After 4 h, chromosomes from each culture were separated by pulse field gel electrophoresis, and then blotted and probed with a DNA fragment of SAC6. As controls, exponential phase wild-type cells grown at 25∞C were incubated with either 0.2 M HU, a DNA replication inhibitor that arrests cells in S phase, or 5 mg ml-1 a-factor that arrests cells in G1. Less DNA is visible in the nse4ts cells at 37∞C because of cell cycle arrest, whereas wild-type cells continue to divide.
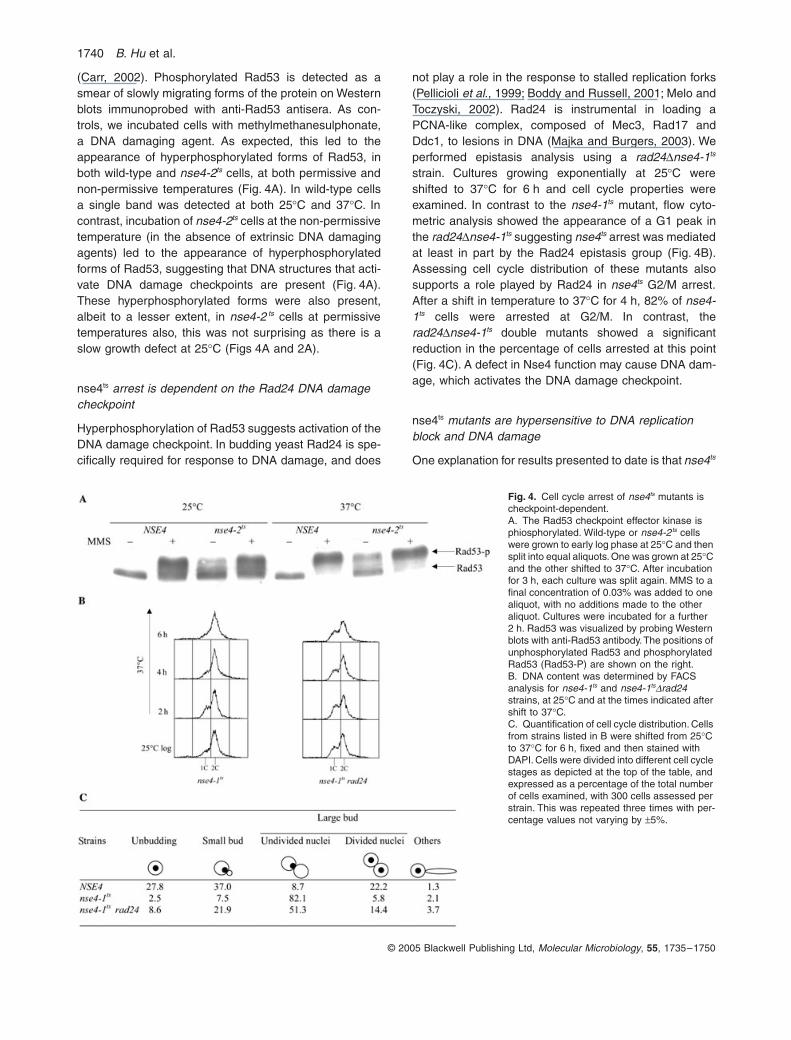
1740 B. Hu et al.
© 2005 Blackwell Publishing Ltd, Molecular Microbiology, 55, 1735–1750
(Carr, 2002). Phosphorylated Rad53 is detected as asmear of slowly migrating forms of the protein on Westernblots immunoprobed with anti-Rad53 antisera. As con-trols, we incubated cells with methylmethanesulphonate,a DNA damaging agent. As expected, this led to theappearance of hyperphosphorylated forms of Rad53, inboth wild-type and nse4-2ts cells, at both permissive andnon-permissive temperatures (Fig. 4A). In wild-type cellsa single band was detected at both 25∞C and 37∞C. Incontrast, incubation of nse4-2ts cells at the non-permissivetemperature (in the absence of extrinsic DNA damagingagents) led to the appearance of hyperphosphorylatedforms of Rad53, suggesting that DNA structures that acti-vate DNA damage checkpoints are present (Fig. 4A).These hyperphosphorylated forms were also present,albeit to a lesser extent, in nse4-2 ts cells at permissivetemperatures also, this was not surprising as there is aslow growth defect at 25∞C (Figs 4A and 2A).
nse4ts arrest is dependent on the Rad24 DNA damage checkpoint
Hyperphosphorylation of Rad53 suggests activation of theDNA damage checkpoint. In budding yeast Rad24 is spe-cifically required for response to DNA damage, and does
not play a role in the response to stalled replication forks(Pellicioli et al., 1999; Boddy and Russell, 2001; Melo andToczyski, 2002). Rad24 is instrumental in loading aPCNA-like complex, composed of Mec3, Rad17 andDdc1, to lesions in DNA (Majka and Burgers, 2003). Weperformed epistasis analysis using a rad24Dnse4-1ts
strain. Cultures growing exponentially at 25∞C wereshifted to 37∞C for 6 h and cell cycle properties wereexamined. In contrast to the nse4-1ts mutant, flow cyto-metric analysis showed the appearance of a G1 peak inthe rad24Dnse4-1ts suggesting nse4ts arrest was mediatedat least in part by the Rad24 epistasis group (Fig. 4B).Assessing cell cycle distribution of these mutants alsosupports a role played by Rad24 in nse4ts G2/M arrest.After a shift in temperature to 37∞C for 4 h, 82% of nse4-1ts cells were arrested at G2/M. In contrast, therad24Dnse4-1ts double mutants showed a significantreduction in the percentage of cells arrested at this point(Fig. 4C). A defect in Nse4 function may cause DNA dam-age, which activates the DNA damage checkpoint.
nse4ts mutants are hypersensitive to DNA replication block and DNA damage
One explanation for results presented to date is that nse4ts
Fig. 4. Cell cycle arrest of nse4ts mutants is checkpoint-dependent.A. The Rad53 checkpoint effector kinase is phiosphorylated. Wild-type or nse4-2ts cells were grown to early log phase at 25∞C and then split into equal aliquots. One was grown at 25∞C and the other shifted to 37∞C. After incubation for 3 h, each culture was split again. MMS to a final concentration of 0.03% was added to one aliquot, with no additions made to the other aliquot. Cultures were incubated for a further 2 h. Rad53 was visualized by probing Western blots with anti-Rad53 antibody. The positions of unphosphorylated Rad53 and phosphorylated Rad53 (Rad53-P) are shown on the right.B. DNA content was determined by FACS analysis for nse4-1ts and nse4-1tsDrad24 strains, at 25∞C and at the times indicated after shift to 37∞C.C. Quantification of cell cycle distribution. Cells from strains listed in B were shifted from 25∞C to 37∞C for 6 h, fixed and then stained with DAPI. Cells were divided into different cell cycle stages as depicted at the top of the table, and expressed as a percentage of the total number of cells examined, with 300 cells assessed per strain. This was repeated three times with per-centage values not varying by ±5%.
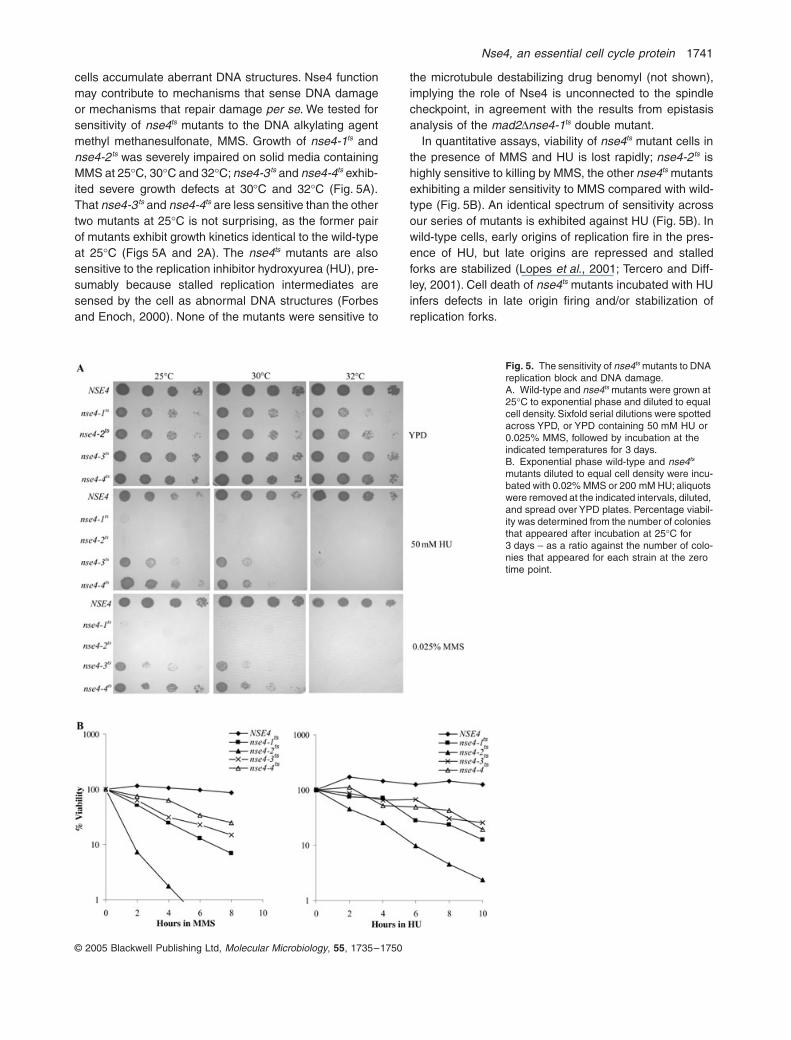
Nse4, an essential cell cycle protein 1741
© 2005 Blackwell Publishing Ltd, Molecular Microbiology, 55, 1735–1750
cells accumulate aberrant DNA structures. Nse4 functionmay contribute to mechanisms that sense DNA damageor mechanisms that repair damage per se. We tested forsensitivity of nse4ts mutants to the DNA alkylating agentmethyl methanesulfonate, MMS. Growth of nse4-1ts andnse4-2ts was severely impaired on solid media containingMMS at 25∞C, 30∞C and 32∞C; nse4-3ts and nse4-4ts exhib-ited severe growth defects at 30∞C and 32∞C (Fig. 5A).That nse4-3ts and nse4-4ts are less sensitive than the othertwo mutants at 25∞C is not surprising, as the former pairof mutants exhibit growth kinetics identical to the wild-typeat 25∞C (Figs 5A and 2A). The nse4ts mutants are alsosensitive to the replication inhibitor hydroxyurea (HU), pre-sumably because stalled replication intermediates aresensed by the cell as abnormal DNA structures (Forbesand Enoch, 2000). None of the mutants were sensitive to
the microtubule destabilizing drug benomyl (not shown),implying the role of Nse4 is unconnected to the spindlecheckpoint, in agreement with the results from epistasisanalysis of the mad2Dnse4-1ts double mutant.
In quantitative assays, viability of nse4ts mutant cells inthe presence of MMS and HU is lost rapidly; nse4-2ts ishighly sensitive to killing by MMS, the other nse4ts mutantsexhibiting a milder sensitivity to MMS compared with wild-type (Fig. 5B). An identical spectrum of sensitivity acrossour series of mutants is exhibited against HU (Fig. 5B). Inwild-type cells, early origins of replication fire in the pres-ence of HU, but late origins are repressed and stalledforks are stabilized (Lopes et al., 2001; Tercero and Diff-ley, 2001). Cell death of nse4ts mutants incubated with HUinfers defects in late origin firing and/or stabilization ofreplication forks.
Fig. 5. The sensitivity of nse4ts mutants to DNA replication block and DNA damage.A. Wild-type and nse4ts mutants were grown at 25∞C to exponential phase and diluted to equal cell density. Sixfold serial dilutions were spotted across YPD, or YPD containing 50 mM HU or 0.025% MMS, followed by incubation at the indicated temperatures for 3 days.B. Exponential phase wild-type and nse4ts mutants diluted to equal cell density were incu-bated with 0.02% MMS or 200 mM HU; aliquots were removed at the indicated intervals, diluted, and spread over YPD plates. Percentage viabil-ity was determined from the number of colonies that appeared after incubation at 25∞C for 3 days – as a ratio against the number of colo-nies that appeared for each strain at the zero time point.
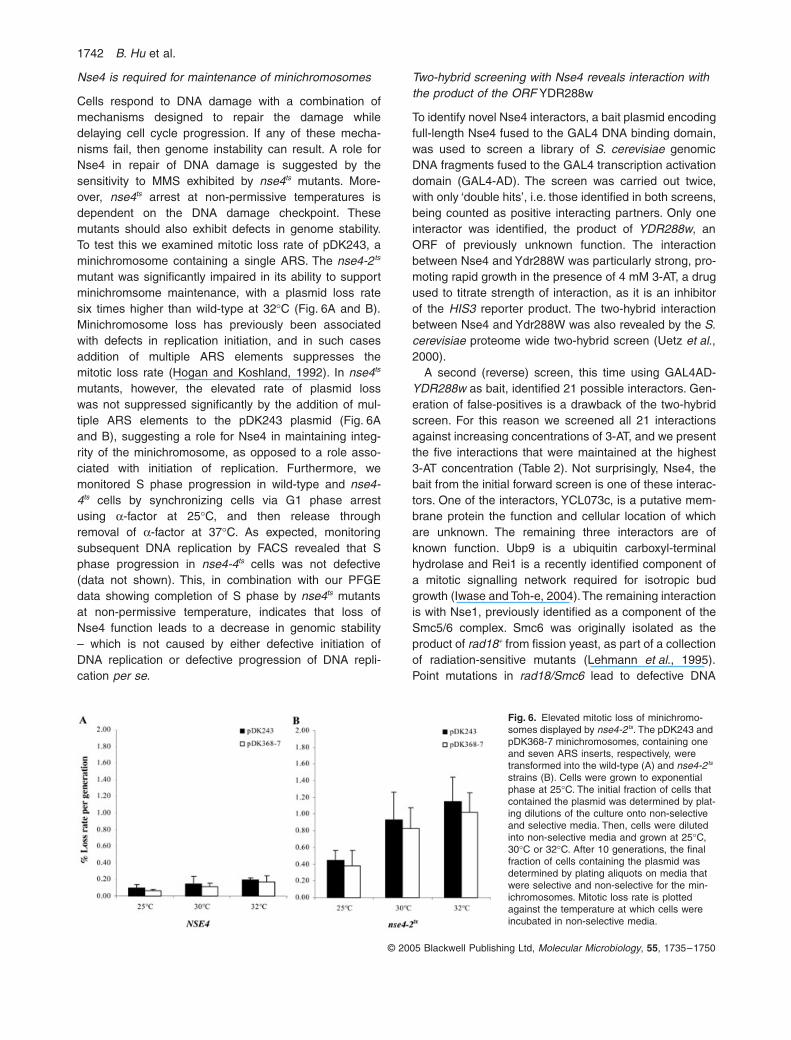
1742 B. Hu et al.
© 2005 Blackwell Publishing Ltd, Molecular Microbiology, 55, 1735–1750
Fig. 6. Elevated mitotic loss of minichromo-somes displayed by nse4-2ts. The pDK243 and pDK368-7 minichromosomes, containing one and seven ARS inserts, respectively, were transformed into the wild-type (A) and nse4-2ts strains (B). Cells were grown to exponential phase at 25∞C. The initial fraction of cells that contained the plasmid was determined by plat-ing dilutions of the culture onto non-selective and selective media. Then, cells were diluted into non-selective media and grown at 25∞C, 30∞C or 32∞C. After 10 generations, the final fraction of cells containing the plasmid was determined by plating aliquots on media that were selective and non-selective for the min-ichromosomes. Mitotic loss rate is plotted against the temperature at which cells were incubated in non-selective media.
Nse4 is required for maintenance of minichromosomes
Cells respond to DNA damage with a combination ofmechanisms designed to repair the damage whiledelaying cell cycle progression. If any of these mecha-nisms fail, then genome instability can result. A role forNse4 in repair of DNA damage is suggested by thesensitivity to MMS exhibited by nse4ts mutants. More-over, nse4ts arrest at non-permissive temperatures isdependent on the DNA damage checkpoint. Thesemutants should also exhibit defects in genome stability.To test this we examined mitotic loss rate of pDK243, aminichromosome containing a single ARS. The nse4-2ts
mutant was significantly impaired in its ability to supportminichromsome maintenance, with a plasmid loss ratesix times higher than wild-type at 32∞C (Fig. 6A and B).Minichromosome loss has previously been associatedwith defects in replication initiation, and in such casesaddition of multiple ARS elements suppresses themitotic loss rate (Hogan and Koshland, 1992). In nse4ts
mutants, however, the elevated rate of plasmid losswas not suppressed significantly by the addition of mul-tiple ARS elements to the pDK243 plasmid (Fig. 6Aand B), suggesting a role for Nse4 in maintaining integ-rity of the minichromosome, as opposed to a role asso-ciated with initiation of replication. Furthermore, wemonitored S phase progression in wild-type and nse4-4ts cells by synchronizing cells via G1 phase arrestusing a-factor at 25∞C, and then release throughremoval of a-factor at 37∞C. As expected, monitoringsubsequent DNA replication by FACS revealed that Sphase progression in nse4-4ts cells was not defective(data not shown). This, in combination with our PFGEdata showing completion of S phase by nse4ts mutantsat non-permissive temperature, indicates that loss ofNse4 function leads to a decrease in genomic stability– which is not caused by either defective initiation ofDNA replication or defective progression of DNA repli-cation per se.
Two-hybrid screening with Nse4 reveals interaction with the product of the ORF YDR288w
To identify novel Nse4 interactors, a bait plasmid encodingfull-length Nse4 fused to the GAL4 DNA binding domain,was used to screen a library of S. cerevisiae genomicDNA fragments fused to the GAL4 transcription activationdomain (GAL4-AD). The screen was carried out twice,with only ‘double hits’, i.e. those identified in both screens,being counted as positive interacting partners. Only oneinteractor was identified, the product of YDR288w, anORF of previously unknown function. The interactionbetween Nse4 and Ydr288W was particularly strong, pro-moting rapid growth in the presence of 4 mM 3-AT, a drugused to titrate strength of interaction, as it is an inhibitorof the HIS3 reporter product. The two-hybrid interactionbetween Nse4 and Ydr288W was also revealed by the S.cerevisiae proteome wide two-hybrid screen (Uetz et al.,2000).
A second (reverse) screen, this time using GAL4AD-YDR288w as bait, identified 21 possible interactors. Gen-eration of false-positives is a drawback of the two-hybridscreen. For this reason we screened all 21 interactionsagainst increasing concentrations of 3-AT, and we presentthe five interactions that were maintained at the highest3-AT concentration (Table 2). Not surprisingly, Nse4, thebait from the initial forward screen is one of these interac-tors. One of the interactors, YCL073c, is a putative mem-brane protein the function and cellular location of whichare unknown. The remaining three interactors are ofknown function. Ubp9 is a ubiquitin carboxyl-terminalhydrolase and Rei1 is a recently identified component ofa mitotic signalling network required for isotropic budgrowth (Iwase and Toh-e, 2004). The remaining interactionis with Nse1, previously identified as a component of theSmc5/6 complex. Smc6 was originally isolated as theproduct of rad18+ from fission yeast, as part of a collectionof radiation-sensitive mutants (Lehmann et al., 1995).Point mutations in rad18/Smc6 lead to defective DNA

Nse4, an essential cell cycle protein 1743
© 2005 Blackwell Publishing Ltd, Molecular Microbiology, 55, 1735–1750
repair pathways; in addition, rad18/Smc6 is required tomaintain cell cycle arrest in the presence of DNA damage(Verkade et al., 1999). Our data suggest a role for Nse4in DNA damage repair, so our subsequent work focusedon determining if there was a functional interactionbetween Nse4 and the known members of the Smc5/6DNA repair complex.
Functional interactions between members of the Smc5/6 complex
Overexpressing Smc5, one of the components of the coreSmc5/6 heterodimer, rescued the ts phenotype of nse4-1ts, though overexpression of other members of the Smc5/6 complex (Mms21, Nse1 and Smc6) did not. Werepeated this analysis, using the nse4-4ts mutant. In thiscase, the growth defect at 37∞C was rescued by Nse1(Fig. 7A). The product of the YDR288w ORF is also aknown component of the S. cerevisiae Smc5/6 complex(Hazbun et al., 2003). Overexpresssion of YDR288w res-cues the ts phenotype of both nse4-1ts and nse4-4ts.
Overexpression of NSE1, SMC5 or YDR288w did notrescue the ts phenotype of nse4-2ts, most likely becauseof this allele bearing the mutation with the most potentdebilitating effect on function, as evidenced by nse4-2ts
exhibiting the poorest growth rate at permissive tempera-ture (Fig. 2A).
Discussion
Nse4 is essential for progression of the cell cycle
Under non-permissive conditions, conditional nse4mutants arrest as large budded cells with a 2C nucleus atthe bud neck and a short mitotic spindle. The 2C nucleusrepresents a genuine post-S-phase arrest, as replicativeintermediates are not detected when chromosomes areresolved by PFGE. In addition, there is no chromosome
breakage as we do not detect sheared DNA by PFGE.The phenotype of nse4ts mutants at non-permissive tem-peratures has the hallmarks of an arrest caused by acti-vation of the DNA damage checkpoint. In budding yeastdamage induced arrest occurs prior to anaphase but isoften termed G2/M arrest.
Phosphorylation of the effector checkpoint kinaseRad53 is clearly detectable in arrested nse4ts cells. Wedemonstrated a requirement for an intact RAD24 check-point for the nse4ts cell cycle arrest. This in combinationwith our FACS and PFGE data, showing complete repli-cation of DNA in arrested cells, suggests Rad53 activationin nse4ts cells is more likely a consequence of activationof the damage checkpoint and not the intra-S checkpoint.Rad53 phosphorylation infers that loss of Nse4 functionleads to disturbance in chromosomal structure. Not sur-prisingly all four of our nse4ts mutants are hypersensitiveto agents that generate aberrant DNA structures, namelystalled replicative intermediates generated by the replica-tion inhibitor HU, and DNA damage caused by the meth-ylation agent, MMS.
Physical interaction of Nse4 with components of the Smc5/Smc6 DNA repair complex
There are numerous complexes involved in DNA repair.Nse4 may play a role in the function of one of these, andwe anticipated that two-hybrid screening would identifythe complex. Use of Nse4 as bait reveals a strong inter-action with Ydr288w, a protein of unknown function. Asexpected, the reverse screen using Ydr288w as bait,picks up Nse4 and a strong interaction with Nse1 (Non-Smc element 1) a component of a known DNA repaircomplex, Smc5/6 (Lehmann et al., 1995; Verkade et al.,1999). As is the case for nse4ts alleles, all mutants in thecore Smc5/6 complex plus mutants in the known non-Smc elements, confer sensitivity to DNA damagingagents. This includes mutations in: S. cerevisiae SMC6/RHC18 and its S. pombe orthologue, rad18 (Lehmannet al., 1995; Onoda et al., 2004), the S. cerevisiae and S.pombe NSE1 orthologues (Fujioka et al., 2002;McDonald et al., 2003) and NSE2 orthologues (Prakashand Prakash, 1977; McDonald et al., 2003). Additionally,like nse4-2ts cells, smc6ts mutants also exhibit a mitoticinstability phenotype (Onoda et al., 2004). Consistentwith a nuclear role, GFP tagged Smc5, Smc6, Nse4,Ydr288w and Mms21 (the S. cerevisiae orthologue of S.pombe Nse2) all localize to the nucleus, as does Nse1(Fujioka et al., 2002; Huh et al., 2003; McDonald et al.,2003).
The mutant phenotype data plus data from our two-hybrid screens suggest a role for Nse4 in Smc5/6 function,which was corroborated by functional interaction betweenNse4 and members of the Smc5/6 complex. Growth arrest
Table 2. Strong two-hybrid interactions with Ydr288w/Nse3.
ORF Gene Function
YCL073C Protein of unknown functionYDL105W QRI2 (NSE4) Member of SMC5–6 DNA repair
complexYER098W UBP9 Ubiquitin carboxyl-terminal hydrolaseYLR007W NSE1 Forms a complex with Smc5 and
Rhc18 (Smc6); required for DNA repair
YBR267W REI1 Required for isotropic bud growth
A ‘bait’ fusion to the Gal4 DNA binding domain (BD) was screenedagainst an array of yeast Gal4 activation domain (AD) protein fusionsthat represent the entire yeast proteome. Interacting partners wereidentified as reproducible two-hybrid positives that were observedtwice. Only interactors that allowed growth in the presence of high 3-AT levels are shown (3-AT is an inhibitor of the reporter product forthis screen).

1744 B. Hu et al.
© 2005 Blackwell Publishing Ltd, Molecular Microbiology, 55, 1735–1750
of nse4-1ts was suppressed by overexpression of SMC5;nse4-4ts growth arrest was suppressed by overexpressionof NSE1. That neither SMC5 nor NSE1 overexpressionrescued both mutants is intriguing, undoubtedly reflectingthe different positions of the amino acid substitutions inthe two nse4ts mutants and consequently the role thesesites play in the function of the Smc5/6 holocomplex. Bothnse4ts mutants were rescued by overexpression ofYDR288w. A very recent publication has identified the S.pombe orthologue of Ydr288w as a component of theSmc5/6 complex; this S. pombe orthologue has beennamed Nse3 (Pebernard et al., 2004). This protein is
essential in both budding and fission yeast. In S. pombe,Nse3 is necessary for mitotic chromosome segregationand cellular resistance to genotoxic agents. We haveshown that all of these properties are shared by cellslacking Nse4, the S. cerevisiae binding partner forYdr288w/Nse3. Furthermore, the interaction betweenNse4 and Ydr288w/Nse3 is conserved in eukaryotes asthe recently completed two-hybrid interaction map forDrosophila melanogaster reveals an interaction betweenthe fly orthologues of these proteins (Giot et al., 2003).We propose renaming YDR288w as NSE3 (non-SMC ele-ment 3), in keeping with the name proposed for the S.
Fig. 7. Nse4 is a component of the Smc5–6 complex.A. YDR288w/NSE3, NSE1 and SMC5 are muticopy suppressors of nse4ts mutants. The nse4-1ts and nse4-4ts mutants were trans-formed with multicopy vectors bearing the ORFs indicated under the control of the MET25 promoter. Gene expression via MET25 was induced by inoculating on selective media that did not contain methionine; the plates were incubated at 37∞C for 3 days.B. Composite schematic combining our two-hybrid data with two-hybrid and TAP-tag inter-action data described elsewhere. Two-hybrid interactions identified from work presented here are indicated by double-headed solid lines, with the two-hybrid bait represented by a solid circle. Proteins identified by copurification of TAP tagged Ydr288w/Nse3 are circled (Hazbun et al., 2003). The two-hybrid interaction between Nse4 (used as bait) and Ydr288W/Nse3 has also been described previously (Uetz et al., 2000). Functional interactions from Fig. 7A are represented by solid bars.

Nse4, an essential cell cycle protein 1745
© 2005 Blackwell Publishing Ltd, Molecular Microbiology, 55, 1735–1750
pombe orthologue of this protein by Pebernard andcoworkers. We have registered the name NSE3 as analias for YDR288w, at the SGD.
Pebernard and coworkers point out that Nse3/Ydr288wis orthologous to the melanoma antigen (MAGE) family inhumans. The function of MAGE has been obscure, untilthe work presented by Pebernard et al. proposed a rolein the Smc5/6 DNA repair complex. Humans contain over20 members of the MAGE family, though Drosophila andyeast contain one (Pebernard et al., 2004). In humans,MAGE has been implicated in cell cycle regulation andinhibition of apoptosis (Barker and Salehi, 2002).
Recent data from proteomic studies substantiates therole we suggest for Nse4 in Smc5/6 function. TAP taggedNse3/Ydr288w copurifies with a complex containingNse4, Smc5, Smc6, Nse1 and Mms21 (the S. cerevisiaeorthologue of S. pombe Nse2) (Hazbun et al., 2003).Some of these proteins are undoubtedly the orthologuesof the unidentified non-Smc proteins in S. pombe thatbound to epitope tagged Smc6 (Fousteri and Lehman,2000). Our two-hybrid data, plus two-hybrid and TAP-taginteraction data generated by Hazbun and coworkers,demonstrate that Nse4 and Nse3/Ydr288w are physicallypart of the Smc5/6 complex. We have also shown thatNse4 plays a functional role in this complex, as overex-pression of Smc5/6 components suppresses the pheno-type of nse4 conditional mutants. A composite schematiccombining our two-hybrid data with two-hybrid and TAP-tag interaction data described elsewhere, plus our func-tional interaction data, is shown in Fig. 7B. Recent dataon Rad62, the S. pombe orthologue of NSE4 are in agree-ment with our results, indicating a role in DNA repair. TheS. pombe rad62-1 mutant is hypersensitive to genotoxicagents, and is defective in repair of double-strand breaks;in addition, Rad62 coimmunoprecipitates with epitopetagged Smc5 (Morikawa et al., 2004).
Our two-hybrid screen also revealed interactionsbetween Nse3/Ydr288w and a putative membrane protein(YCL073c) a ubiquitin carboxyl-terminal hydrolase (Ubp9)and a component of a mitotic signalling network requiredfor isotropic bud growth (Rei1). None of these proteins areinvolved in DNA repair or maintenance of higher-orderchromosome structures, so functional interaction withNse3/Ydr288w is not immediately apparent. Our two-hybrid screen using Nse4 as bait identified Nse3/Ydr288was the only binding partner. This interaction was alsorevealed by a genome-wide two-hybrid screen, which alsoidentified seven other binding partners (Uetz et al., 2000).Three of these, Hsc82, Sti1 and Cpr6, are components ofthe Hsp90 chaperosome (Pearl and Prodromou, 2000).This chaperosome is responsible for creating and main-taining the active conformation of key regulatory proteins,and may play a similar role in maintenance of Smc5/6holocomplex activity. The remaining Nse4 interactors were
a repressor of transcription (Tup1) a component of a MAPkinase cascade that responds to osmotic shock (Ssk2)and an enzyme that catalyses the first step of GMP bio-synthesis (Imd2). A functional interaction between theseproteins and Nse4 is not immediately apparent.
Overall, the function of SMC complexes is to establishthe higher-order structure of chromosomes. Smc1/3(cohesin) is the glue that ensures sister chromatids aremaintained as a pair; Smc2/4 (condensin) mediatesmitotic chromosome condensation (Hirano, 2000).Higher-order function of the Smc5/6 heterodimerremains poorly understood. Overall, it seems mainte-nance of higher-order structures is of importance toDNA repair and replication. Mutations in cohesin sub-units lead to increased sensitivity to genotoxic agents(Tatebayashi et al., 1998; Walowsky et al., 1999). Thesame is true for condensin subunits (Aono et al., 2002;Chen et al., 2004). Studies in S. pombe indicate a rolefor Smc5/6 in homologous recombination based DNArepair (Lehman et al., 1995). A conditional mutant of S.cerevisiae SMC6 is unable to induce interchromosomalrecombination in response to DNA damage (Onodaet al., 2004). However, both SMC5 and SMC6 (likeNSE4) are essential for progression of the undisturbedcell cycle, in the absence of extrinsic agents that dam-age DNA. It has been suggested that the Smc5/6 com-plex holds together broken DNA molecules in the vicinityof double-strand breaks, so that repair by recombinationis allowed to take place (Fousteri and Lehman, 2000).This is plausible given that occasional double-strandbreaks occur during DNA replication (Muris et al., 1996).It is not surprising that a complex involved in maintain-ing higher-order chromosome structure plays a role inrecombination. In human cells, Smc1/3 promotes repairof DNA lesions by homologous recombination (Jess-berger et al., 1996). Indeed, Smc5/6 seems to facilitatevarious DNA repair processes, which would explain whyS. pombe smc6 mutants are sensitive to a wide rangeof DNA damaging agents (Harvey et al., 2004). Further-more mutants in smc6 are synthetically lethal with amutation in DNA topoisomerase 2 (Verkade et al.,1999). All of this suggests that Smc5/6, like the Smc1/3and Smc2/4 complexes, plays a role in chromatin orga-nization. In cells treated with the genotoxic agents HUand MMS, nse4ts cells lose viability. It is not activationof a checkpoint that is lost here, because the Rad53effector kinase is activated in the mutants, even atnon-permissive temperatures. HU causes stalling of rep-lication forks, which also occurs when S phase cellsencounter DNA alkylated by MMS (Tercero et al., 2003).Cell death under these circumstances implies a role forstabilization of stalled forks by Nse4, and by the Smc5/6complex as a whole. The association of human Smc5/6with DNA during interphase, and its exclusion from DNA

1746 B. Hu et al.
© 2005 Blackwell Publishing Ltd, Molecular Microbiology, 55, 1735–1750
during mitosis, is in agreement with this (Taylor et al.,2001).
Unlike mutants in smc5 and smc6, nse4 mutants arrest at a discrete stage in the cell cycle
In S. pombe, cells deleted for either Smc5 or Smc6 dis-play a range of lethal terminal phenotypes such as elon-gated cells, many with several septa with a singlenucleus, or a dispersed distribution of DNA or a cut phe-notype. Chromosome missegregation has also beenreported for a conditional smc6 mutant in S. cerevisiae(Onoda et al., 2004). Similar phenotypes are exhibited bythe ts mutants in the non-Smc subunits, Nse1 and Nse2(Fujioka et al., 2002; McDonald et al., 2003). Our nse4ts
mutants, however, display a terminal phenotype that is farless extreme, namely a growth arrest prior to onset ofanaphase. Moreover, this arrest is not lethal, even whenthe cells are incubated at non-permissive conditions for3 days. Also, the nucleus is not fragmented at all, and isarrested at a stage prior to anaphase. In FACS analysisof smc6ts mutants, the G2 peak becomes broader andflatter after 10 h at non-permissive conditions, reflectingthe chromosome missegregation that occurs (Onodaet al., 2004). This does not occur in nse4ts mutants, theG2 peak persisting for at least 24 h. If the Smc5/6 com-plex repairs a distinct type of genomic lesion, character-ization of this lesion will be easier in nse4ts mutants.Moreover, the catastrophic terminal phenotype of Smc5/6 mutants implies a collapse in the structure of the com-plex itself, leading to amorphous distribution of nuclearDNA and ‘cut’ morphology. This is not surprising becauseSmc5/6 forms the core of the complex. In nse4ts mutants,loss of function does not result in large-scale chromo-somal aberrations, implying the central role of the Smc5/6 core remains intact. The nse4ts mutant phenotype may
represent a ‘snapshot’ of the Smc5/6 mechanism ofaction.
In S. pombe, Smc5/6 functions in tandem with Rad60(Morishita et al., 2002; Boddy et al., 2003). The buddingyeast orthologue of Rad60 is Esc2, named for its role inchromatin silencing (Dhillon and Kamakaka, 2000). Wehave noticed that we are unable to synchronize the mostsevere nse4ts mutant (nse4-2ts) with a-factor. This couldbe because of a silencing defect, giving rise to simulta-neous expression of MATa and MATa genes. It is possiblethat Nse4 is involved in repair of damage, such as double-strand breaks, perhaps through assembly of newlyrepaired DNA into chromatin. We note that mutants incomponents of the Smc5/6 complexes exhibit deficienciesin recombination. PFGE of these cells would revealsheared DNA. However, we do not see sheared DNA inarrested nse4ts cells. This is why we suggest a possiblerole for Nse4 in chromatin assembly post DNA repair.Moreover, S. cerevisiae which lack Asf1, a protein thatmediates chromatin assembly, are – like nse4ts cells –sensitive to MMS and HU, and cannot be synchronizedwith a-factor (Tyler et al., 1999; Hu et al., 2001). The nextkey issue to be addressed is the exact nature of thechromosomal aberration that exists in arrested nse4ts
cells. We propose to do this by investigating genetic andbiochemical interaction between Nse4 and proteinsinvolved in recombination repair and chromatin assembly.
Experimental procedures
Yeast strains, media and plasmid construction
All yeast strains and plasmids used in this study are listed inTables 1 and 3 respectively. Combinations of mutants in thesame strain were constructed by standard procedures, yeaststrains were cultured on rich (YPD) or synthetic (SD) glucosesupplemented with the appropriate amino acids and nucleic
Table 3. Vectors used in this study.
Plasmid Insert/Marker Promoter Copy no. Source
pBP01 NSE3/URA3 NSE3 CEN This studypBH1 nse3-1ts/URA3 NSE3 CEN This studypBH2 nse3-2ts/URA3 NSE3 CEN This studypBH3 nse3-3ts/URA3 NSE3 CEN This studypBH4 nse3-4ts/URA3 NSE3 CEN This studypBH10 NSE3/LEU2 NSE3 CEN This studypBH101 YDR288W/LEU2 MET25 2 mm This studypBH102 MMS21/LEU2 MET25 2 mm This studypBH103 NSE1/LEU2 MET25 2 mm This studypBH104 SMC5/LEU2 MET25 2 mm This studypBH105 RHC18(SMC6)/LEU2 MET25 2 mm This studypBH106 FLJ20003/LEU2 MET25 2 mm This studypBH107 FLJ20003/LEU2 NSE3 2 mm This studypDK243 1ARS/LEU2/ADE3 Hogan and Koshand (1992)pDK368-7 7ARS/LEU2/ADE3 Hogan and Koshand (1992)pBDC-Nse3 YDL105W/NSE3 2 mm This studyPbdc-288w YDR288W/NSE4 2 mm This study

Nse4, an essential cell cycle protein 1747
© 2005 Blackwell Publishing Ltd, Molecular Microbiology, 55, 1735–1750
acid bases (Rose et al., 1990). NSE4 was expressed via itsnative promoter by sequential cloning of DNA fragments intothe S. cerevisiae/Escherichia coli shuttle vector YCplac33(Gietz and Sugino, 1988). First, a 414 bp Pst I-HindIII frag-ment containing the ADH1 transcription terminator frompGBT9 (Clontech) was ligated into Pst I-HindIII-cleavedYCplac33, to give YCplac33T. The NSE4 ORF and its pro-moter were amplified separately from S. cerevisiae genomicDNA and cloned sequentially into YCplac33T, to give pBP01.The NSE4 promoter and ORF from this vector were thensubcloned into the centromeric LEU2 vector YCplac111T, togive pBP02; construction of YCplac111T itself is describedelsewhere (Panaretou et al., 1999)
Isolation of ts alleles of NSE4
The centromeric plasmid bearing a LEU2 marker and theNSE4 ORF ligated to its native promoter (pB02), was sub-jected to hydroxylamine mutagenesis in vitro (Sikorski andBoeke, 1991). Pools of mutants, were transformed into ahaploid S. cerevisiae strain (BP12) deleted for NSE4. Viabilityof this strain was maintained by a centromeric plasmid (pB01)bearing a URA3 marker and the NSE4 ORF ligated to itsnative promoter. More than 8000 transformants were pickedand incubated overnight at 25∞C in 384 well plates containingminimal (SD) media without leucine and uracil. Using a 384-pin high density replica tool (Fisher Scientific) cells weretransferred to two SD plates without leucine containing 5-FOA (Rose et al., 1990). One plate was incubated at 25∞C,the other at 37∞C. The 5-FOA permitted growth of cells thatacquired the mutated plasmid but had lost the wild-type plas-mid. Transformants that did not grow at 37∞C but grew at 25∞Cwere selected. To ensure these ts phenotypes were attributedto mutated DNA, plasmids from ts mutants were recovered,transformed into the BP12 S cerevisiae strain andrescreened for the ts phenotype. Nse4 coding sequences,from plasmids that continued to confer the ts phenotyopeafter rescreening, were subcloned into a centromeric URA3plasmid, followed by transformation into a diploid strain(10590D, from the EUROSCARF collection) that is a het-erozygous delete at the NSE4 locus. Following sporulationand tetrad dissection, haploids bearing the mutated plasmidin the NSE4 delete background were screened for the tsphenotype. The coding regions of nse4ts mutants weresequenced with the Taq DyeDeoxy Terminator CycleSequencing kit (Applied Biosystems) and a model 370A auto-mated sequencer (Applied Biosystems).
Sensitivity against DNA damaging agents
To determine sensitivity to HU or MMS, exponential cultureswere adjusted so that they were at equal cell density (1 ¥ 107
cells ml-1) and four successive sixfold serial dilutions werespotted on YPD, or YPD containing either 50 mM HU or0.025% MMS. Plates were incubated at 25∞C, 30∞C or 32∞Cfor 3 days.
To determine viability in HU, cells were grown at 25∞C tolog phase in YPD, and HU was added to a final concentrationof 200 mM. Aliquots were removed and spread on YPD platesevery 2 h. Plates were incubated at 25∞C for 3 days. The
percent survival compared with that at time zero was deter-mined by counting the resulting colonies. For viability aftertreatment with MMS, cells grown in YPD to log phase wereincubated in 0.025% MMS, aliquots were removed every 2 hand the MMS was inactivated by addition of sodium thiosul-phate to a final concentration of 5%. Aliquots were spread onYPD, followed by incubation at 25∞C. Colony numbers weredetermined after 3 days. Survival was expressed as a per-centage of the number of colonies obtained from cells thatwere not treated with MMS.
Fluorescence activator cell sorter (FACS) analysis
Standard procedures were used for FACS analysis, usingpropidium iodide to stain DNA followed by analysis via FAC-Scan (Becton Dickinson) (Chakraverty et al., 2001).
Pulse field gel analysis
Isogenic strains bearing the ts mutant or wild alleles of NSE4were grown in YPD to log phase at 25∞C and then incubatedat 37∞C for 4 h. Control samples were (i) aliquots of the samecultures maintained at 25∞C or (ii) wild-type cells treated witheither a-factor (5 mg ml-1) for 3 h or HU (200 mM) for 2 h.Yeast chromosomal DNA was prepared by a standard proce-dure, involving embedding cells in agarose followed by treat-ment of the agarose plugs with proteinase K and lyticase(Hennessy et al., 1991). PFGE was carried out in 1%megabase-agarose (Bio-Rad) in a CHEF-DRTM II electro-phoresis cell (Bio-Rad). Electrophoresis was performed for24 h with a switching time of 70 s at 170 V in 0.5% Tris-borate-EDTA. The separated chromosomes were then acid-nicked and transferred to nylon membrane (Hybond N+,Amersham-Pharmacia) (Hennessy et al., 1991). The blot wasprobed with DIG-labelled SAC6 fragment and detected withCDP-StarTM kit (Boehringer-Mannheim). This probe to chro-mosome IV was chosen because analysis of replication stateby PFGE is most sensitive when probes are derived fromlarge chromosomes (Hennessy et al., 1991).
Immunofluorescence microscopic analysis
Cells were processed for fluorescence and indirect fluores-cence (Bowers et al., 2000). Fixed cells were stained for DNAusing DAPI (25 mg ml-1), and for the mitotic spindle, using ratmonoclonal anti-tubulin antibody (YOL1/34, abcam) andFITC-conjugated goat anti-rat antibody (abcam) using estab-lished methods (Hisamoto et al., 1994).
Protein extraction and Rad53 detection
Yeast protein extracts were prepared from TCA-treated cellsas described previously (Pellicioli et al., 1999). Rad53 wasdetected with a rabbit polyclonal antibody provided by J.Diffley (Tercero et al., 2003).
Assay for mitotic loss of minichromosomses
Plasmid loss assays were performed using the minichromo-

1748 B. Hu et al.
© 2005 Blackwell Publishing Ltd, Molecular Microbiology, 55, 1735–1750
some vectors pDK243 or pDK368-7, bearing one or sevenARS elements respectively (Hogan and Koshland, 1992).Selective pressure was maintained by presence of a LEU2marker. The vectors were transformed into (i) the haploidBP12 S. cerevisiae strain, which is deleted for NSE4 butbears a plasmid with NSE4 and the URA3 marker and (ii) anisogenic strain except the vector bears the nse4-2ts mutant.The initial fraction of cells that contained the plasmid wasdetermined by plating dilutions of the culture onto SDmedium with or without leucine. Cells were subsequentlydiluted into YPD and grown at 25∞C, 30∞C or 32∞C. After 8–10 generations, the final fraction of cells containing the plas-mid was determined by plating aliquots on media that wereselective (SD media without leucine) and non-selective(YPD) for the minichromosomes. The loss rate was calcu-lated as previously described (Hogan and Koshland, 1992).
Yeast two-hybrid screen
NSE4 and [Nse3/YDR288w] C-terminal ‘bait’ fusions to theGal4 DNA binding domain (BD) of the pBDC TRP1 vectorwere generated by homologous recombination within theyeast strain PJ69-4a as described previously (Uetz et al.,2000; Millson et al., 2003). Presence of a BD fusion wasconfirmed by Western blotting, using anti-Gal4p BD antise-rum (Clontech). The bait fusions in PJ69-4a did not self-activate transcription of the integrated GAL1 promoter/HIS3fusion reporter, as shown by lack of growth on medium con-taining 2 mM 3-amino-1,2,4-triazole (3-AT), an inhibitor of theHIS3 reporter product.
PJ69-4a bearing (BD) bait fusions were mated by pinningonto the previously described 16-plate, 384 format array ofyeast Gal4 activation domain (AD) protein fusions, all ofwhich are carried in a strain of the opposite mating type(PJ69-4a) (Uetz et al., 2000). Protein–protein interactionswere identified by pinning the diploids onto medium lackingleucine, tryptophan and histidine, supplemented with 4 mM3-AT (Uetz et al., 2000). Colonies were scored for growthafter 10 days at 30∞C. Putative interacting partners wereidentified as reproducible two-hybrid positives that wereobserved twice from duplicate screens. It was assumed thatinteracting proteins identified only once were the result of anon-reproducible false-positive.
Assessing functional interaction between putative members of the Smc5/6 complex by multicopy suppression
Candidate ORFs were overexpressed from the vectorYEp81Met, which is a modification of the episomal LEU2vector YEplac181. The vector bears the MET25 induciblepromoter and the transcription termination site from PGK1,separated by a multiple cloning site. Candidate ORFs wereamplified from S. cerevisiae genomic DNA and ligated 3¢ tothe MET25 promoter. The resulting vectors were transformedinto the BP12 strain, or an isogenic strain containing a vectorborne nse4ts allele instead of the wild-type gene. Transfor-mants were streaked on SD plates without uracil and leucine,and without methionine to induce overexpression of the can-didate ORF.
Acknowledgements
We thank Douglas Koshland (Carnegie Institution ofWashington) for providing the pDK243 and pDK368-7 vec-tors; Frank Cooke (University College London) for providingthe YEp81Met vector and John Diffley (CRUK, Clare Hall) forproviding antisera to Rad53. We thank Katherine Bowers(CIMR, Cambridge) for assistance with immunofluorescence,Lucy Drury (CRUK, Clare Hall) for assistance with detectionof phosphorylated forms of Rad53 and David Barford and LoriPassmore (Institute of Cancer Research, London) for valu-able comments. This work was supported by the CentralResearch Fund of the University of London, the Royal Societyand the BBSRC (Grant No. BBS/B/04951). B.H. was partlysupported by a doctoral scholarship funded by the KC WongFoundation and the China Scholarship Council.
References
Alexandru, G., Zachariae, W., Schleiffer, A., and Nasmyth,K. (1999) Sister chromatid separation and chromosomere-duplication are regulated by different mechanisms inresponse to spindle damage. EMBO J 18: 2707–2721.
Aono, N., Sutani, T., Tomonaga, T., Mochida, S., andYanagida, M. (2002) Cnd2 has dual roles in mitotic con-densation and interphase. Nature 417: 197–202.
Barker, P.A., and Salehi, A. (2002) The MAGE proteins:emerging roles in cell cycle progression, apoptosis, andneurogenetic disease. J Neurosci Res 67: 705–712.
Boddy, M.N., and Russell, P. (2001) DNA replication check-point. Curr Biol 11: R953–R956.
Boddy, M.N., Shanahan, P., Mcdonald, W.H., Lopez-Girona,A., Noguchi, E., Yates, J.R., and Russell, P. (2003). Rep-lication checkpoint kinase Cds1 regulates recombinationalrepair protein Rad60. Mol Cell Biol 23: 5939–5946.
Bowers, K., Levi, B.P., Patel, F.I., and Stevens, T.H. (2000)The sodium/proton exchanger Nhx1p is required for endo-somal protein trafficking in the yeast Saccharomyces cer-evisiae. Mol Biol Cell 11: 4277–4294.
Carr, A.M. (2002) DNA structure dependent checkpoints asregulators of DNA repair. DNA Repair 1: 983–994.
Chakraverty, R.K., Kearsey, J.M., Oakley, T.J., Grenon, M.,De La Torre Luiz, M.A., Lowndes, N.F., and Hickson, I.D.(2001) Topoisomerase III acts upstream of Rad53p in theS-Phase DNA damage checkpoint. Mol Cell Biol 21: 7150–7162.
Chen, E.S., Sutani, T., and Yanagida, M. (2004) Cti1/C1Dinteracts with condensin SMC hinge and supports the DNArepair function of condensin. Proc Natl Acad Sci USA 101:8078–8083.
Dhillon, N., and Kamakaka, R.T. (2000) A histone variant,Htz1p, and a Sir1p-like protein, Esc2p, mediate silencingat HMR. Mol Cell 6: 769–780.
Forbes, K.H., and Enoch, T. (2000) Cell cycle checkpoints.In The Yeast Nucleus. Fantes, P., and Beggs, J. (eds).Oxford: Oxford University Press, pp. 106–133.
Fousteri, M.I., and Lehmann, A.R. (2000) A novel SMCprotein complex in Schizosaccharomyces pombecontains the Rad18 DNA repair protein. EMBO J 19:1691–1702.

Nse4, an essential cell cycle protein 1749
© 2005 Blackwell Publishing Ltd, Molecular Microbiology, 55, 1735–1750
Fujioka, Y., Kimata, Y., Nomaguch, I.K., Watanabe, K., andKohno, K. (2002) Identification of a novel non-structuralmaintenance of chromosomes (SMC) component of theSMC5–SMC6 complex involved in DNA repair. J Biol Chem277: 21585–21591.
Gietz, R.D., and Sugino, A. (1988) New yeast-Escherichiacoli shuttle vectors constructed with in vitro mutagenizedyeast genes lacking 6-base pair restriction sites. Gene 74:527–534.
Giot, L., Bader, J.S., Brouwer, C., Chaudhuri, A., Kuang, B.,Li, Y., et al. (2003) A protein interaction map of Drosophilamelanogaster. Science 302: 1727–1736.
Harvey, S.H., Sheedy, D.M., Cuddihy, A.R., and O’Connell,M.J. (2004) Coordination of DNA damage responses viathe Smc5/Smc6 complex. Mol Cell Biol 24: 662–674.
Hazbun, T.R., Malmstrom, I., Anderson, S., Graczyk, B.J.,Fox, B., Riffle, M., et al. (2003) Assigning function to yeastproteins by integration of technologies. Mol Cell 12: 1353–1365.
Hennessy, K.M.A., Lee, A., Chen, E., and Botstein, D. (1991)A group of interacting yeast DNA-replication genes. GeneDev 5: 958–969.
Hirano, T. (2000) Chromosome cohesion, condensation, andseparation. Ann Rev Biochem 69: 115–144.
Hisamoto, N.K., Sugimoto, K., and Matsumoto, K. (1994) TheGLC7 type-1 protein phosphatase of Saccharomyces-cerevisiae is required for cell-cycle progression in G(2)/M.Mol Cell Biol 14: 3158–3165.
Hogan, E., and Koshland, D. (1992) Addition of extra originsof replication to a minichromosome suppresses its mitoticloss in cdc6 and cdc14 mutants of Saccharomyces-cerevisiae. Proc Natl Acad Sci USA 89: 3098–3102.
Hu, F.H., Alcasabas, A.A., and Elledge, S.J. (2001) Asf1 linksRad53 to control of chromatin assembly. Gene Dev 15:1061–1066.
Huh, W.K., Falvo, J.V., Gerke, L.C., Carroll, A.S., Howson,R.W., Weissman, J.S., and O’Shea, E.K. (2003) Globalanalysis of protein localization in budding yeast. Nature425: 686–691.
Iwase, M., and Toh-e, A. (2004) Ybr267w is a newcytosplasmic protein belonging to the mitotic signallingnetwork of Saccharomyces cerevisiae. Cell Struct Funct29: 1–15.
Jessberger, R., Riwar, B., Baechtold, H., and Akhmedov,A.T. (1996) SMC proteins constitute two subunits of themammalian recombination complex RC-1. EMBO J 15:4061–4068.
Lehmann, A.R., Walicka, M., Griffiths, D.J.F., Murray, J.M.,Watts, F.Z., McCready, S., and Carr, A.M. (1995) Therad18 gene of Schizosaccharomyces-pombe defines anew subgroup of the smc superfamily involved in DNA-repair. Mol Cell Biol 15: 7067–7080.
Lopes, M., Cotta-Ramusino, C., Pellicioli, A., Liberi, G., Ple-vani, P., Muzi-Falconi, M., et al. (2001) The DNA replicationcheckpoint response stabilizes stalled replication forks.Nature 412: 557–561.
McDonald, W.H., Pavlova, Y., Yates, J.R., and Boddy, M.N.(2003) Novel essential DNA repair proteins Nse1 and Nse2are subunits of the fission yeast Smc5–Smc6. J Biol Chem278: 45460–44546.
Majka, J., and Burgers, P.M.J. (2003) Yeast Rad17/Mec3/
Ddc1: a sliding clamp for the DNA damage checkpoint.Proc Natl Acad Sci USA 100: 2249–2254.
Melo, J., and Toczyski, D. (2002) A unified view of the DNA-damage checkpoint. Curr Opin Cell Biol 14: 237–245.
Millson, S.H., Truman, A.W., and Piper, P.W. (2003) Vectorsfor N- or C-terminal positioning of the yeast Gal4p DNAbinding or activator domains. Biotechniques 35: 60–64.
Morikawa, H., Morishita, T., Kawane, S., Iwasaki, H., Carr,A.M., and Shinagawa, H. (2004) Rad62 protein functionallyand physically associates with the Smc5/Smc6 proteincomplex and is required for chromosome integrity andrecombination repair in fission yeast. Mol Cell Biol 24:9401–9413.
Morishita, T., Tsutsui, Y., Iwasaki, H., and Shinagawa, H.(2002) The Schizosaccharomyces pombe rad60 gene isessential for repairing double-strand DNA breaks sponta-neously occurring during replication and induced by DNA-damaging agents. Mol Cell Biol 22: 3537–3548.
Muris, D.F.R., Vreeken, K., Carr, A.M., Murray, J.M., Smit,C., Lohman, P.H.M., and Pastink, A. (1996) Isolation of theSchizosaccharomyces pombe RAD54 homologue, rhp54+,a gene involved in the repair of radiation damage andreplication fidelity. J Cell Sci 109: 73–81.
Noskov, V.N., Araki, H., and Sugino, A. (1998) The RFC2gene, encoding the third-largest subunit of the replicationfactor C complex, is required for an S-phase checkpointin Saccharomyces cerevisiae. Mol Cell Biol 18: 4914–4923.
Onoda, F., Takeda, M., Seki, M., Maeda, D., Tajima, J., Ui,A., et al. (2004) SMC6 is required for MMS-induced inter-chromosomal and sister chromatid recombinations in Sac-charomyces cerevisiae. DNA Repair 3: 429–439.
Panaretou, B., Sinclair, K., Prodromou, C., Johal, J., Pearl,L., and Piper, P.W. (1999) The Hsp90 of Candida albicanscan confer Hsp90 functions in Saccharomyces cerevisiae:a potential model for the processes that generate immuno-genic fragments of this molecular chaperone in C-albicansinfections. Microbiology 145: 3455–3463.
Pearl, L.H., and Prodromou, C. (2000) Structure and in vivofunction of Hsp90. Curr Opin Struct Biol 10: 46–51.
Pebernard, S., McDonald, W.H., Pavlova, Y., Yates, J.R., andBoddy, M.N. (2004) Nse1, Nse2, and a novel subunit ofthe Smc5–Smc6 complex, Nse3, play a crucial role inmeiosis. Mol Biol Cell 15: 4866–4876.
Pellicioli, A., Lucca, C., Liberi, G., Marini, F., Lopes, M.,Plevani, P., et al. (1999) Activation of Rad53 kinase inresponse to DNA damage and its effect in modulatingphosphorylation of the lagging strand DNA polymerase.EMBO J 18: 6561–6572.
Prakash, S., and Prakash, L. (1977) Increased spontaneousmitotic segregation in MMS-sensitive mutants of Saccha-romyces cerevisiae. Genetics 87: 229–236.
Roberts, C.J., Nelson, B., Marton, M.J., Stoughton, R.,Meyer, M.R., Bennett, H.A., et al. (2000) Signaling andcircuitry of multiple MAPK pathways revealed by a matrixof global gene expression profiles. Science 287: 873–880.
Rose, M.D., Winston, F., and Hieter, P. (1990) Methods inYeast Genetics: A Cold Spring Harbor Laboratory CourseManual. Cold Spring Harbor, NY: Cold Spring Harbor Lab-oratory Press.
Sikorski, R.S., and Boeke, J.D. (1991) In vitro mutagenesis

1750 B. Hu et al.
© 2005 Blackwell Publishing Ltd, Molecular Microbiology, 55, 1735–1750
and plasmid shuffling – from cloned gene to mutant yeast.Methods Enzymol 194: 302–318.
Simon, M., Benit, P., Vassal, A., Dubois, C., and Faye, G.(1994) Sequence of the PHO2-POL3 (CDC2) region ofchromosome-IV of Saccharomyces cerevisiae. Yeast 10:1653–1656.
Spellman, P.T., Sherlock, G., Zhang, M.Q., Iyer, V.R.,Anders, K., Eisen, M.B., et al. (1998) Comprehensive iden-tification of cell cycle-regulated genes of the yeast Saccha-romyces cerevisiae by microarray hybridization. Mol BiolCell 9: 3273–3297.
Sugimoto, K., Sakamoto, Y., Takahashi, O., and Matsumoto,K. (1995) HYS2, an essential gene required for DNA rep-lication in Saccharomyces cerevisiae. Nucleic Acids Res23: 3493–3500.
Tatebayashi, K., Kato, J., and Ikeda, H. (1998) Isolation of aSchizosaccharomyces pombe rad21 (ts) mutant that isaberrant in chromosome segregation, microtubule func-tion, DNA repair and sensitive to hydroxyurea: possibleinvolvement of Rad21 in ubiquitin-mediated proteolysis.Genetics 148: 49–57.
Taylor, E.M., Moghraby, J.S., Lees, J.H., Smit, B., Moens,P.B., and Lehmann, A.R. (2001) Characterization of anovel human SMC heterodimer homologous to theSchizosaccharomyces pombe Rad18/Apr18 complex. MolBiol Cell 12: 1583–1594.
Tercero, J.A., and Diffley, J.F.X. (2001) Regulation of DNA
replication fork progression through damaged DNA by theMec1/Rad53 checkpoint. Nature 412: 553–557.
Tercero, J.A., Longhese, M.P., and Diffley, J.F.X. (2003) Acentral role for DNA replication forks in checkpoint activa-tion and response. Mol Cell 11: 1323–1336.
Tyler, J.K., Adams, C.R., Chen, S.R., Kobayashi, R.,Kamakaka, R.T., and Kadonaga, J.T. (1999) The RCAFcomplex mediates chromatin assembly during DNA repli-cation and repair. Nature 402: 555–560.
Uetz, P., Giot, L., Cagney, G., Mansfield, T.A., Judson, R.S.,Knight, J.R., et al. (2000) A comprehensive analysis ofprotein–protein interactions in Saccharomyces cerevisiae.Nature 403: 623–627.
Valdivieso, M.H., Sugimoto, K., Jahng, K.-Y., Fernandes,P.M.B., and Wittenberg, C. (1993) FAR1 is required forposttranscriptional regulation of CLN2 gene expression inresponse to mating pheromone. Mol Cell Biol 13: 1013–1022.
Verkade, H.M., Bugg, S.J., Lindsay, H.D., Carr, A.M., andO’Connell, M.J. (1999) Rad18 is required for DNA repairand checkpoint responses in fission yeast. Mol Biol Cell10: 2905–2918.
Walowsky, C., Fitzhugh, D.J., Castano, I.B., Ju, J.Y., Levin,N.A., and Christman, M.F. (1999) The topoisomerase-related function gene TRF4 affects cellular sensitivity to theantitumor agent camptothecin. J Biol Chem 274: 7302–7308.




















