
Protective immunity against porcine circovirus 2 in mice induced by a gene-based combination...
14
THE JOURNAL OF GENE MEDICINE RESEARCH ARTICLE J Gene Med 2009; 11: 288–301. Published online 4 February 2009 in Wiley InterScience (www.interscience.wiley.com) DOI: 10.1002/jgm.1300 Protective immunity against porcine circovirus 2 in mice induced by a gene-based combination vaccination Kandan Aravindaram 1 Tsun-Yung Kuo 2† Chun-Wen Lan 1† Hsiu-Hui Yu 1 Pei-Hsueh Wang 1 Yu-San Chen 2 Gabriel Hsu-Chung Chen 2 Ning-Sun Yang 1 * 1 Agricultural Biotechnology Research Center, Academia Sinica, Taipei, Taiwan 2 Department of Animal Science, National Ilan University, Ilan, Taiwan *Correspondence to: Ning-Sun Yang, Agricultural Biotechnology Research Center, Academia Sinica, no. 128, Section 2, Academia Road, Nangang, Taipei 115, Taiwan. E-mail: [email protected] † These two investigators (T.Y.K and C.W.L.) contributed equally to this work. (K.A. is the first and senior author). Received: 21 May 2008 Revised: 21 October 2008 Accepted: 23 December 2008 Abstract Background Porcine circovirus type 2 (PCV2) is the primary cause of an emerging swine disease, postweaning multisystemic wasting syndrome, that is responsible for economic losses. To develop an effective vaccine for PCV2, we evaluated a heterologous prime-boost vaccine approach, using a gene gun-mediated naked DNA vector as a priming and modified vaccinia virus ankara (MVA) as a booster, in Balb/c mice. Methods Three open reading frames (ORF) of PCV2 viral samples from infected pigs were amplified, and gene gun-mediated DNA priming vaccination was performed followed by boosts with MVA vectors expressing the same ORFs of PCV2. After vaccination, mice were challenged with PCV2 virus, and virus titers in the lungs and lymph nodes were measured. Results The combination of ORF-2 and -3 in this gene-based vaccine strategy resulted in high antibody titers and virus neutralization activity in serum, reduced PCV2 virus load, and reduced levels of apoptosis in the lungs. No cross-reaction was observed between ORF-1 and -2, but weak cross-reaction was observed between ORF-1 and -3, and between ORF-2 and -3. Following vaccination, expression of chemokines, macrophage inflammatory protein- 1β and regulated upon activation, normal T cell expressed and secreted, increased significantly. The expression of T helper 1-type cytokine (interferon- γ ) and specific lysis of PCV2-infected cells increased; concomitantly, the level of T helper 2-type cytokine (interleukin-10) decreased in test mice. The expression of tumor necrosis factor-α and granulocyte-macrophage colony- stimulating factor increased significantly in mice vaccinated with ORF-2/-3, and with ORF-1/-2/-3. Conclusions This prime-boost vaccination strategy, using a gene gun for DNA priming and recombinant MVA for boosts, may be an attractive vaccine strategy against PCV2 infection in swine. Copyright 2009 John Wiley & Sons, Ltd. Keywords cellular immunity; gene-gun; humoral immunity; porcine circovirus; prime-boost; vaccine Introduction Porcine circovirus (PCV), a member of the Circoviridae, is a non-enveloped, single-stranded DNA virus containing a circular genome [1]. PCV was Copyright 2009 John Wiley & Sons, Ltd.
-
Upload
independent -
Category
Documents
-
view
0 -
download
0
Transcript of Protective immunity against porcine circovirus 2 in mice induced by a gene-based combination...

THE JOURNAL OF GENE MEDICINE R E S E A R C H A R T I C L EJ Gene Med 2009; 11: 288–301.Published online 4 February 2009 in Wiley InterScience (www.interscience.wiley.com) DOI: 10.1002/jgm.1300
Protective immunity against porcine circovirus 2 inmice induced by a gene-based combinationvaccination
Kandan Aravindaram1
Tsun-Yung Kuo2†
Chun-Wen Lan1†
Hsiu-Hui Yu1
Pei-Hsueh Wang1
Yu-San Chen2
Gabriel Hsu-Chung Chen2
Ning-Sun Yang1*
1Agricultural Biotechnology ResearchCenter, Academia Sinica, Taipei,Taiwan2Department of Animal Science,National Ilan University, Ilan, Taiwan
*Correspondence to:Ning-Sun Yang, AgriculturalBiotechnology Research Center,Academia Sinica, no. 128, Section 2,Academia Road, Nangang, Taipei115, Taiwan. E-mail:[email protected]
†These two investigators (T.Y.K andC.W.L.) contributed equally to thiswork. (K.A. is the first and seniorauthor).
Received: 21 May 2008Revised: 21 October 2008Accepted: 23 December 2008
Abstract
Background Porcine circovirus type 2 (PCV2) is the primary cause of anemerging swine disease, postweaning multisystemic wasting syndrome, thatis responsible for economic losses. To develop an effective vaccine for PCV2,we evaluated a heterologous prime-boost vaccine approach, using a genegun-mediated naked DNA vector as a priming and modified vaccinia virusankara (MVA) as a booster, in Balb/c mice.
Methods Three open reading frames (ORF) of PCV2 viral samplesfrom infected pigs were amplified, and gene gun-mediated DNA primingvaccination was performed followed by boosts with MVA vectors expressingthe same ORFs of PCV2. After vaccination, mice were challenged with PCV2virus, and virus titers in the lungs and lymph nodes were measured.
Results The combination of ORF-2 and -3 in this gene-based vaccine strategyresulted in high antibody titers and virus neutralization activity in serum,reduced PCV2 virus load, and reduced levels of apoptosis in the lungs. Nocross-reaction was observed between ORF-1 and -2, but weak cross-reactionwas observed between ORF-1 and -3, and between ORF-2 and -3. Followingvaccination, expression of chemokines, macrophage inflammatory protein-1β and regulated upon activation, normal T cell expressed and secreted,increased significantly. The expression of T helper 1-type cytokine (interferon-γ ) and specific lysis of PCV2-infected cells increased; concomitantly, the levelof T helper 2-type cytokine (interleukin-10) decreased in test mice. Theexpression of tumor necrosis factor-α and granulocyte-macrophage colony-stimulating factor increased significantly in mice vaccinated with ORF-2/-3,and with ORF-1/-2/-3.
Conclusions This prime-boost vaccination strategy, using a gene gun forDNA priming and recombinant MVA for boosts, may be an attractive vaccinestrategy against PCV2 infection in swine. Copyright 2009 John Wiley &Sons, Ltd.
Keywords cellular immunity; gene-gun; humoral immunity; porcine circovirus;prime-boost; vaccine
Introduction
Porcine circovirus (PCV), a member of the Circoviridae, is a non-enveloped,single-stranded DNA virus containing a circular genome [1]. PCV was
Copyright 2009 John Wiley & Sons, Ltd.

Gene-based porcine circovirus vaccine 289
originally identified as a contaminant from porcinekidney cell cultures (PK15, ATCC CCL-33). Infection withPCV type 2 (PCV2) but not PCV1 is associated withpostweaning multisystemic wasting syndrome (PMWS) inyoung weaned pigs. The disease was first recognized inCanada in 1991 [2] and subsequently has been describedin virtually all regions of the world [3–7]. Clinical andlaboratory studies showed that PCV2 is etiologicallyinvolved in PMWS [8–12]. PMWS is characterizedclinically by enlarged lymph nodes, chronic pneumonia,progressive weight loss, and, less commonly, icterus,pallor, or diarrhea [13]. Morbidity and mortality withPMWS are severe in acute outbreaks, usually resulting inthe death of 70–80% of affected animals [14].
A previous study showed that the pathogenic PCV2 andnonpathogenic PCV1 share similar genomic organizationsbut only approximately 76% nucleotide sequence identity[5]. To date, three open reading frames (ORF) of PCV2have been characterized: ORF-1 encodes the rep proteinsrequired for viral replication [15,16], ORF-2 encodes themajor immunogenic capsid protein of 27.8 kDa [15,17],and an ORF-3 protein plays an important role in viralpathogenesis by its apoptotic activity [12,18].
PCV2 is a very stable virus and apparently can persistin the environment of infected herds; thus, eradication ofthe virus is very difficult. PCV2 vaccine studies in miceand/or pigs have reported on an attenuated chimericPCV1-2 live virus as well as chimeric PCV1-2 infectiouscDNA vectors [11], ORF-1 and ORF-2 subunit vaccines[19], PCV2 structural protein vaccines [20], and arecombinant adenovirus expressing the capsid protein ofPCV2 [21]. However, most of these previous studies havenot systematically characterized the immune responses.In addition, an ORF-3 gene-based vaccine has not beenstudied in vivo.
Modified vaccinia virus ankara (MVA) is considered anexcellent alternative vaccine vector candidate, becauseit is highly attenuated and has limited ability toreplicate in mammalian cell lines. Genes encodingproteins with immunomodulatory functions and hostrange determinants were found to be lost during serialpassages of the virus through chick embryo fibroblasts(CEFs), which led to attenuated replication and virulence;on the 516th passage, the virus was renamed MVA [22].More than 15% of the original vaccinia genome were lost[23,24], but most genes encoding structural proteins wereretained, which suggests that key antigenic determinantshave been preserved in MVA. Therefore, the MVA vectoris both safer and more immunogenic than the replication-competent vaccinia virus currently licensed for smallpoxvaccination and in development as a vaccine vector formalaria, human immunodeficiency virus, tuberculosis,and cancer [25–33]. Some advantages of MVA vector-based vaccines include their established safety profile inhumans, their efficient delivery of heterologous antigensin clinical trials, and the availability of technologies fortheir large-scale production [31,34–36]. MVA also hasgood immunogenicity, extreme host-range restriction,possibility of long-term storage (stockpiling), and easy
production at biosafety level 1 conditions in CEFs andbaby hamster kidney (BHK-21) cells [36–39].
The order of vaccination is critically important. Themain limiting factor in the development of vaccinesfor intracellular infectious diseases and chronic diseasesis the ability to induce very strong and long-lastingcell-mediated immunity. The rationale behind ourcurrent consecutive vaccination strategy was that DNAimmunization, which is believed to elicit a relativelylow-level but persistent immunity, may prime a greatlyenhanced response following boosting with anothervector such as the MVA, which can express higherlevels of the targeted vaccine antigen [28,40,41]. DNAplasmid vector-based vaccines can prime efficiently butoften cannot boost a very strong CD8+ response aswell as a CD4+ T-cell response. Heterologous prime-boost immunization, an approach employing two differentvector systems for delivery of the same or relatedgene constructs, induces higher levels of effector T-cell responses in animals and humans than homologousboosting with the same vaccine vector [28,40,41].
In the present study, we used a heterologous prime-boosting regime with all three ORFs of PCV2, alone orin combination, as recombinant plasmid DNA constructsand as MVA vectors, and studied their combinedimmune responses and related mechanisms. The use ofa combination of both ORF-2 and -3 enhanced T helper(Th) 1-mediated immune responses and neutralized PCV2viral titers better than all other individual or combinedtreatments examined.
Materials and methods
Viruses and cells
The immortalized PK-15 cell line, free from PCV contam-ination, was maintained in Dulbecco’s minimal Eagle’smedium (Gibco, Gaithersburg, MD, USA) supplementedwith 5% heat inactivated fetal bovine serum (FBS), 5%L-glutamine, penicillin (100 U/ml), and streptomycin(100 µg/ml) (Invitrogen, Carlsbad, CA, USA) at 37 ◦Cin 5% CO2. The wild-type PCV2 virus used in the studywas originally isolated from lung tissue of a pig naturallyaffected by PMWS.
Animals
Female Balb/cBYJ mice (6–8 weeks of age) werepurchased from the National Laboratory Animal Breedingand Research Center, Taipei, Taiwan. All mice weremaintained in a laminar air-flow cabinet in an animalroom kept at 24 ± 2 ◦C and 40% to 70% humidityunder a 12 : 12 h light/dark cycle and specific pathogen-free conditions. All facilities were approved by theAcademia Sinica Institutional Animal Care and UtilizationCommittee, and all animal experiments were conducted
Copyright 2009 John Wiley & Sons, Ltd. J Gene Med 2009; 11: 288–301.DOI: 10.1002/jgm

290 K. Aravindaram et al.
under the institutional guidelines established for theAnimal Core Facility in Academia Sinica, Taipei, Taiwan.
Construction and expression of cDNAexpression vectors for DNA vaccine
In the present study, we used plasmid pIBSY1, a p3224mammalian expression vector containing the marker geneof kanamycin resistance for selection in Escherichia coliand a SV40 intron A sequence immediately follow-ing the cytomegalovirus (CMV) immediate early genepromoter for vector construction and gene-based immu-nization. Plasmids p3224-ORF-1, -2 and -3, containingthe cDNA genes encoding ORF-1, -2 and -3 of PCV2,respectively, were constructed by subcloning the ORF-1, -2 and -3 gene fragments into the BamHI andBglII sites of the pIBSY1 vector by standard molecularcloning procedures. Plasmid pIBSY1 was constructed bycloning a 74-bp DNA fragment coding for the murineimmunoglobulin IgG κ-chain leader sequence into theEcoRI-BamHI site of the pIBSY1 vector. The double-stranded 74-bp DNA fragment was obtained by anneal-ing the two synthetic oligodeoxynucleotides: IgκLf, 5′-GGCCATGGAGACAGACACACTCCTGCTATGGGTACTGC-TGCTCTGGGTTCCAGGTTCCACTGGTGACGCGGCCG-3′,and IgκLr, 5′-AATTCGGCCGCGTCACCAGTGGAACCTGG-AACCCAGAGCAGCAGTACCCATAGCAGGAGTGTGTCTG-TCTCCAT-3′. All plasmid constructs were fully sequencedto confirm their authenticity. For DNA immunization,each DNA plasmid was amplified in E. coli strain XL10-Gold (Stratagene, La Jolla, CA, USA) and purified using anEndofree Plasmid Giga kit (Qiagen, Valencia, CA, USA).The purity of DNA preparations was determined by opticaldensity readings at 260 and 280 nm. In vitro transgenegenome expression was tested in transient expressionexperiments with PK15 cells grown in six-well plates andtransfected with the above cDNA constructs (4 µg of plas-mid DNA) with use of Lipofectamine 2000 (Invitrogen)as described in the manufacturer’s protocol. At 48 h post-transfection, cells were collected for reverse transcriptase-polymerase chain reaction (RT-PCR). Transgenic proteinexpressions were tested by immuno-peroxidase mono-layer assay (IPMA) as described previously [42].
Recombinant MVA construction
The three cDNA fragments of ORF-1, -2 and 3 wereamplified and inserted into the SalI-PstI site of the pLW44transfer vector (kindly provided by Dr Bernard Moss,Chief, Laboratory of viral diseases, National Instituteof Allergy and Infectious Diseases, National Institutesof Health, Bethesda, MD, USA), bringing them underthe control of the vaccinia virus-modified H5 earlylate promoter [43]. cDNA encoding enhanced greenfluorescent protein regulated by the vaccinia virus P11late promoter was placed adjacent to this gene. AnIgG κ-chain leader sequence was subcloned into the
SmaI-SalI site of pLW44 for secretion purposes. Thecorrect sequence of the ORF-1, -2 and -3 cDNA insertwas confirmed by sequencing. Recombinant MVAs weremade by transfecting transfer plasmids into BHK-21 cellsinfected with 0.05 plaque forming units (pfu) of MVA percell. Fluorescent plaques were cloned by eight successiverounds of plaque isolation and propagated in BHK-21, andtiters of recombinant MVA were determined by stainingplaques with anti-vaccinia virus rabbit antibody [44].
Immunization and PCV2 viruschallenge
We used the Helios gene-gun system (Bio-Rad, FosterCity, CA, USA) with slight modifications as describedpreviously [45]. Briefly, 35 mg of gold was added to 0.05M spermidine and sonicated prior to incubation with theplasmid DNA and 2.5 M CaCl2. The DNA-coated gold waswashed with absolute ethanol prior to loading into thecompletely dry tubing, each half-inch segment of whichallowed the delivery of 0.5 mg of gold and 1 µg of plasmidDNA per transfection. When two or three plasmids wereused in combination, each plasmid was precipitated ontoseparate gold particles, which were then mixed togetherto deliver 1 µg of each plasmid DNA per transfection. ForDNA vaccination, mouse abdominal skin was depilatedwith Nair depilatory cream (Carter-Wallace, New York,NY, USA), and transfected in vivo with the cDNA vectorswith a 380-psi helium gas pulse. For DNA prime-MVAboost experiments, each mouse (ten per group) receiveda dose of 3 µg of ORF-1, -2 or -3 cDNA, either individuallyor in combination. The ‘empty vector control’ group micewas gene gun-bombarded with vector backbone that didnot contain the target gene insert (i.e. empty vector,either as recombinant plasmid DNA or a MVA viral vector,which was not engineered with any of the PCV2 ORFcDNA sequence).
For boosting with recombinant MVA, mice wereimmunized twice with MVA-ORF-1, -2 or -3, aloneor in combination, 2 weeks after gene transfer at14-day intervals. Recombinant MVA was inoculatedintraperitoneally (i.p.) at 1 × 107 pfu per immunization.The mice intraperitoneal infection model is generallyaccepted for poxvirus infection and evaluation of viralimmunogenicity [46], T-cell epitope mapping [47], andrational enhancement of MVA vector immunogenicity[48].
Two weeks after the second boost with recombinantMVA, immunized mice were challenged intramuscularly(i.m.) with 5 × 104.5 TCID50/ml of PCV2 as describedpreviously [49]. One week later, lungs and draininglymph nodes of two test animals in each group wereremoved and the PCV2 titers determined. Sera werecollected before immunization and at various times upto 10 weeks after immunization. Blood was allowed torest for 16–24 h at room temperature, when serumwas then collected from the coagulated blood bymicrocentrifugation at 8000 r.p.m. 6200 g for 15 min.
Copyright 2009 John Wiley & Sons, Ltd. J Gene Med 2009; 11: 288–301.DOI: 10.1002/jgm

Gene-based porcine circovirus vaccine 291
Antibody responses
The levels of serum IgG antibody and the subtypes (IgG2aand IgG1) against all three individual ORF-1, -2 and -3 ofPCV2 were measured as described previously [50], withslight modifications. In brief, 96-well microtiter plateswere coated with 20 µg/ml of recombinant ORF-1, -2 or-3 proteins, separately, in PBS (50 µl/well) overnight at4 ◦C, and serum samples in serial dilutions were addedto test wells. After 1 h of incubation at 37 ◦C and threewashes with PBS containing 0.1% Tween-20, titers of IgGantibody against specific PCV2 antigens were measuredusing biotinylated rat anti-mouse IgG1 (1 : 1000 dilution)or biotinylated rat anti-mouse IgG2a (1 : 2000 dilution)antibodies (R&D Systems, Minneapolis, MN, USA). Theantibody titer was expressed as the value of the reciprocalof the dilution that gave 50% of the maximum enzyme-linked immunosorbent assay (ELISA) (OD492) reading.Cross reaction tests were also performed for sera of miceimmunized with PCV2 ORF vaccines and challenged withPCV2 virus, using IPMA as described above (Table 1).
Virus neutralization assay
Serum levels of neutralizing antibodies were measuredat weeks 4 and 8 after the first immunization by virus-neutralizing activity assay. Briefly, an equal volume ofPCV2 (1000 TCID50/ml) was added to the titrated serumsamples and incubated for 1 h at 37 ◦C. The mixturewas then inoculated into the 96-well plate containingconfluent PK15 cells as described previously [42], withslight modification. After 48 h, the culture plate was fixedin 4% paraformaldehyde for 1 h at 37 ◦C, and cells werethen washed with PBS and incubated with anti-rabbitPCV2 ORF-1, -2 or -3 with respective treatments for 1 hat 37 ◦C. After 1 h, cells were washed and incubatedwith a peroxidase-conjugated secondary antibody, thenstained with the enzyme substrate at room temperaturefor 15 min, then washed again and viewed under whitelight microscopy.
Real-time PCR
Three days after challenge with PCV2, total RNA extractedfrom inguinal lymph nodes of immunized and con-trol mice was reverse transcribed using an Invitro-gen RT kit, with an input of 500 ng used for eachreaction. Subsequent PCR to detect expression of themouse chemokines regulated upon activation, normal Tcell expressed and secreted (RANTES) and macrophageinflammatory protein-1β (MIP-1β) mRNA was carried outfor 40 cycles at the appropriate annealing temperaturefor the following pairs of primers: RANTES forward, 5′-CGCGGATCCCCACGTCAAGGAGTATTTCTACACC-3′ andreverse, 5′-CGGGATCCCTGGTTTCTTGGGTTTGCTGTG-3′; and MIP-1β forward, 5′-CGCGGA TCCCGGAAGATTC-CACGCCAATTC-3′ and reverse, 5′-GGATCCGGTTGAGG-
AA CGTGTCCTGAAG-3′. Real-time PCR used the LightCy-cler 2.0 Real-Time PCR System (Roche Diagnostics, Basel,Switzerland) and the results were normalized to mouseglyceraldehyde 3-phosphate dehydrogenase (Gapdh) asan internal control. The Gapdh primers were: forwardGapdh-F, 5′-CATCACTGCCACCCAGAAGACTGTGGA-3′and reverse Gapdh-R, 5′-TACTCCTTGGAGGCCATGTAGG-CCATG-3′. Two independent experiments were conductedin triplicate.
Determination of virus titers
One week after immunized mice had been challenged withPCV2, viral titers in lungs and inguinal lymph nodes weredetermined by use of IPMA as described previously [42],with slight modification. Briefly, lungs and lymph nodeswere collected from test mice and homogenized in 1 ml ofPBS by use of a hand-held homogenizer. The homogenateswere placed on ice for 15 min to allow cell debris tosettle. Clarified lysates were then diluted in serum-freemedium (Invitrogen), added to PK15 cells cultured in48-well plates, and incubated for 1 h at 37 ◦C, followedby the addition of Dulbecco’s modified Eagle’s mediumsupplemented with 10% FBS. At 48 h post-infection, testmedium was carefully removed and IPMA performed asdescribed above.
Murine interferon (IFN)-γenzyme-linked immunosorbent spot(ELISPOT) assay
PCV2-specific IFN-γ -secreting spleen cells were enumer-ated by ELISPOT assay, essentially as described previously[51]. Briefly, microwell plates (MultiScreenHTS; Millipore,Billerica, MA, USA) were coated with capture antibody(anti-mouse IFN-γ capture antibody; R&D Systems) andincubated for 16–18 h at 4 ◦C, then blocked with 1%bovine serum albumin and 5% sucrose in PBS for 2 h atroom temperature. Spleen cells from immunized (PCV2-ORF) and empty vector treated mice (i.e. with emptyvector of plasmid DNA or MVA viral vector constructedwithout any insertion of PCV2 ORF cDNA genes) were col-lected 2 weeks after the last immunization step. After lysisof red blood cells with ACK lysis buffer, spleen cells wereresuspended to a final density of 5 × 105 cells/well, and100 µl of cell suspension was cultured with or withoutPCV2 recombinant ORF proteins (20 µg/ml) (for stim-ulation) and medium only for 24 h at 37 ◦C. The plateswere washed, and detection antibody (1 : 60) (anti-mouseIFN-γ detection antibody; R&D Systems) was added andincubated overnight at 4 ◦C. After further washes, plateswere developed with streptavidin-alkaline phosphataseconjugate (R&D Systems) and 5-bromo-4-chloro-3′-indolylphosphate-nitroblue tetrazolium substrate. IFN-γspots were counted with an Immunospot analyser andconfirmed by the computer-based Immunospot software(Cellular Technology, Shaker Heights, OH, USA). Results
Copyright 2009 John Wiley & Sons, Ltd. J Gene Med 2009; 11: 288–301.DOI: 10.1002/jgm
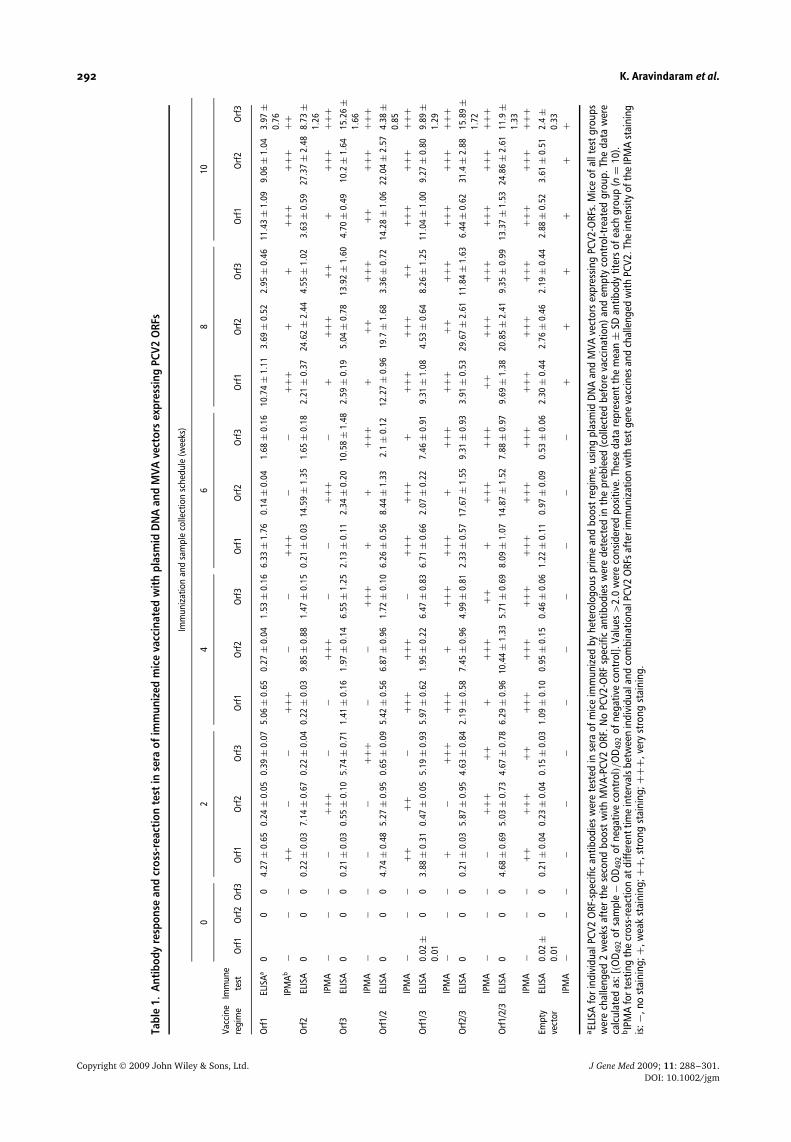
292 K. Aravindaram et al.
Tabl
e1.
Antib
ody
resp
onse
and
cros
s-re
actio
nte
stin
sera
ofim
mun
ized
mic
eva
ccin
ated
with
plas
mid
DN
Aan
dM
VAve
ctor
sex
pres
sing
PCV2
ORF
s
Imm
uniz
atio
nan
dsa
mpl
eco
llect
ion
sche
dule
(wee
ks)
02
46
810
Vacc
ine
regi
me
Imm
une
test
Orf
1O
rf2
Orf
3O
rf1
Orf
2O
rf3
Orf
1O
rf2
Orf
3O
rf1
Orf
2O
rf3
Orf
1O
rf2
Orf
3O
rf1
Orf
2O
rf3
Orf
1EL
ISAa
00
04.
27±
0.65
0.24
±0.
050.
39±
0.07
5.06
±0.
650.
27±
0.04
1.53
±0.
166.
33±
1.76
0.14
±0.
041.
68±
0.16
10.7
4±
1.11
3.69
±0.
522.
95±
0.46
11.4
3±
1.09
9.06
±1.
043.
97±
0.76
IPM
Ab−
−−
++−
−++
+−
−++
+−
−++
++
+++
+++
+++
Orf
2EL
ISA
00
00.
22±
0.03
7.14
±0.
670.
22±
0.04
0.22
±0.
039.
85±
0.88
1.47
±0.
150.
21±
0.03
14.5
9±
1.35
1.65
±0.
182.
21±
0.37
24.6
2±
2.44
4.55
±1.
023.
63±
0.59
27.3
7±
2.48
8.73
±1.
26IP
MA
−−
−−
+++
−−
+++
−−
+++
−+
+++
+++
+++
+++
Orf
3EL
ISA
00
00.
21±
0.03
0.55
±0.
105.
74±
0.71
1.41
±0.
161.
97±
0.14
6.55
±1.
252.
13±
0.11
2.34
±0.
2010
.58
±1.
482.
59±
0.19
5.04
±0.
7813
.92
±1.
604.
70±
0.49
10.2
±1.
6415
.26
±1.
66IP
MA
−−
−−
−++
+−
−++
++
+++
++
++++
+++
+++
+++
Orf
1/2
ELIS
A0
00
4.74
±0.
485.
27±
0.95
0.65
±0.
095.
42±
0.56
6.87
±0.
961.
72±
0.10
6.26
±0.
568.
44±
1.33
2.1
±0.
1212
.27
±0.
9619
.7±
1.68
3.36
±0.
7214
.28
±1.
0622
.04
±2.
574.
38±
0.85
IPM
A−
−−
++++
−++
+++
+−
+++
+++
+++
+++
+++
+++
+++
+++
Orf
1/3
ELIS
A0.
02±
0.01
00
3.88
±0.
310.
47±
0.05
5.19
±0.
935.
97±
0.62
1.95
±0.
226.
47±
0.83
6.71
±0.
662.
07±
0.22
7.46
±0.
919.
31±
1.08
4.53
±0.
648.
26±
1.25
11.0
4±
1.00
9.27
±0.
809.
89±
1.29
IPM
A−
−−
+−
+++
+++
+++
+++
++
+++
+++
++++
+++
+++
+++
+O
rf2/
3EL
ISA
00
00.
21±
0.03
5.87
±0.
954.
63±
0.84
2.19
±0.
587.
45±
0.96
4.99
±0.
812.
33±
0.57
17.6
7±
1.55
9.31
±0.
933.
91±
0.53
29.6
7±
2.61
11.8
4±
1.63
6.44
±0.
6231
.4±
2.88
15.8
9±
1.72
IPM
A−
−−
−++
+++
+++
+++
+++
+++
+++
+++
+++
+++
+++
+++
Orf
1/2/
3EL
ISA
00
04.
68±
0.69
5.03
±0.
734.
67±
0.78
6.29
±0.
9610
.44
±1.
335.
71±
0.69
8.09
±1.
0714
.87
±1.
527.
88±
0.97
9.69
±1.
3820
.85
±2.
419.
35±
0.99
13.3
7±
1.53
24.8
6±
2.61
11.9
±1.
33IP
MA
−−
−++
+++
++++
+++
+++
+++
+++
+++
+++
+++
+++
+++
+++
+++
+Em
pty
vect
orEL
ISA
0.02
±0.
010
00.
21±
0.04
0.23
±0.
040.
15±
0.03
1.09
±0.
100.
95±
0.15
0.46
±0.
061.
22±
0.11
0.97
±0.
090.
53±
0.06
2.30
±0.
442.
76±
0.46
2.19
±0.
442.
88±
0.52
3.61
±0.
512.
4±
0.33
IPM
A−
−−
−−
−−
−−
−−
−+
++
++
+
a ELI
SAfo
rind
ivid
ualP
CV2
ORF
-spe
cific
antib
odie
sw
ere
test
edin
sera
ofm
ice
imm
uniz
edby
hete
rolo
gous
prim
ean
dbo
ostr
egim
e,us
ing
plas
mid
DNA
and
MVA
vect
ors
expr
essin
gPC
V2-O
RFs.
Mic
eof
allt
estg
roup
sw
ere
chal
leng
ed2
wee
ksaf
ter
the
seco
ndbo
ost
with
MVA
-PCV
2O
RF.N
oPC
V2-O
RFsp
ecifi
can
tibod
ies
wer
ede
tect
edin
the
preb
leed
(col
lect
edbe
fore
vacc
inat
ion)
and
empt
yco
ntro
l-tre
ated
grou
p.Th
eda
taw
ere
calc
ulat
edas
:[(O
D 492
ofsa
mpl
e−
OD 4
92of
nega
tive
cont
rol)/
OD 4
92of
nega
tive
cont
rol].
Valu
es>
2.0
wer
eco
nsid
ered
posit
ive.
Thes
eda
tare
pres
entt
hem
ean
±SD
antib
ody
titer
sof
each
grou
p(n
=10
).b I
PMA
fort
estin
gth
ecr
oss-
reac
tion
atdi
ffere
nttim
ein
terv
alsb
etw
een
indi
vidu
alan
dco
mbi
natio
nalP
CV2
ORF
saft
erim
mun
izat
ion
with
test
gene
vacc
ines
and
chal
leng
edw
ithPC
V2.T
hein
tens
ityof
the
IPM
Ast
aini
ngis:
−,no
stai
ning
;+,w
eak
stai
ning
;++,
stro
ngst
aini
ng;+
++,v
ery
stro
ngst
aini
ng.
Copyright 2009 John Wiley & Sons, Ltd. J Gene Med 2009; 11: 288–301.DOI: 10.1002/jgm

Gene-based porcine circovirus vaccine 293
for all wells were averaged and normalized by comparingthe number of antigen-specific spots to negative controlspots.
Cytotoxicity assays
Test PCV2 vaccines were also assayed for cytotoxicityT lymphocytes activity using nonradioactive DELFIAEuTDA cytotoxicity reagents [52,53]. Briefly, plated PK15cells infected with PCV2 were scraped free, washedonce and labelled with DELFIA BATDA reagent (PerkinElmer, Waltham, MA, USA) for 30 min at 37 ◦C. Afterthree additional washes, 5 × 103 target cells (per well)were incubated with test spleen cells in 96-well flat-bottom plates. The amount of specific cell lysis wascalculated as: 100 × [experimental release (counts)− spontaneous release (counts)]/[maximum release(counts) − spontaneous release (counts)].
ELISA for cytokine expression
Two weeks after the last immunization, spleens wererecovered from mice immunized with different ORFgenes. After tissue homogenates were passed througha sieve, erythrocytes were lysed with ACK lysis buffer.Spleen cells (5 × 106) were then incubated in 1 mlwith PCV2 recombinant ORF proteins (20 µg/ml) at37 ◦C for 48 h. The quantities of IFN-γ , interleukin(IL)-10, tumor necrosis factor (TNF)-α and granulocyte-macrophage colony-stimulating factor (GM-CSF) in thesupernatants from these cultures were measured by ELISA(R&D Systems).
Terminal deoxynucleotidyltransferase-mediated nick endlabelling (TUNEL) assay
Empty vector or PCV2 ORF-1, -2 or -3 vaccine-immunizedmice were challenged with PCV2 virus 2 weeks afterthe last immunization. One week later, the lung tissuesof test mice were collected and fixed overnight with4% paraformaldehyde in PBS. Test lung tissues werethen washed with 30% sucrose in PBS three times andcryoprotected in 30% sucrose solution overnight. Tissueswere then embedded in optimal cutting temperaturecompound and cut into 7-µm sections with a microtome.Cryostat tissue sections were placed onto precleanedslides (Fisher Scientific Co., Pittsburgh, PA, USA) andstained with use of the In Situ Cell Death Detectionperoxidase kit (Roche Diagnostics). After blocking,permeabilization and proteinase digestion, tissue slideswere incubated with 100 µl of TUNEL reaction mixtureand covered with a lid for 60 min at 37 ◦C in a humidifiedchamber in the dark. Finally, slides were washed threetimes with 1 × PBS, mounted, and examined under afluorescence microscope.
Statistical analysis
The data are presented as the mean ± SD. The significanceof any differences between group means was examinedusing the nonpaired two-tailed Student’s t-test, assumingequal variance. p < 0.05 was considered statisticallysignificant.
Results
Construction and expression of PCV2transgenes
All three ORFs of PCV2 and ORF-1, -2 and -3 werePCR amplified from lung tissues of PCV2-infected pigsand sub-cloned under the control of the cytomegaloviruspromoter. To ensure secretion of transgenic ORF viralproteins, all constructs were engineered with an IgGκ leader sequence, a strong signal peptide for proteinsecretion, immediately upstream of the ORF genes.We observed a partial secretion of transgenic PCV2ORF proteins in the conditioned medium collected 48 hafter PK15 cells were transfected with the respectiveplasmid DNA encoding PCV2 ORFs (data not shown).Transgenic expression of PCV2-ORF RNA transcriptsin PK-15 cells is shown in Figure 1A. All constructswere further confirmed by sequencing. We then usedIPMA to further test in PK-15 cells the expression oftransgenic proteins encoded by PCV2 ORFs. Figure 1Bshows expression of transgenic ORF proteins at thecellular level, with no protein expression in mock-transfected PK-15 cells. At the subcellular level, thesetransgenic proteins were located in the nucleoli andrarely in the nucleoplasm of the transfected PK-15 cells(data not shown). These results demonstrate that therecombinant cDNA plasmids encoding various ORFs ofPCV2 can be effectively expressed in transfected PK-15 cells at both RNA and protein levels. Importantly,the transgenic expression of recombinant MVA vectorsencoding ORF-1, -2 and -3 was also confirmed (data notshown).
Gene-based vaccination with PCV2 ORFinduces protective immunity in mice
Mice were subjected to a gene-based PCV2 vaccinationregime (Figure 1C) via priming with a plasmid DNA ORFvector and boosting with a MVA vector expressing thecorresponding specific PCV2 ORF. Significantly enhancedlevels of PCV2 antibodies were detected in mice treatedwith ORF-2/-3 or -1/-2/-3 vectors in combinationcompared to other treatments. The PCV2 antibody levelsincreased at weeks 6–10 in all PCV2-immunized mice,whereas no or only background immune activity wasdetected in animals treated with the empty-vector regime(Table 1). An antibody response to PCV2 ORF was firstseen in immunized mice from the second week, after
Copyright 2009 John Wiley & Sons, Ltd. J Gene Med 2009; 11: 288–301.DOI: 10.1002/jgm

294 K. Aravindaram et al.
Figure 1. Analysis of transgenic expression of PCV2-ORF cDNA constructs by RT-PCR and IPMA. (A) Expression of PCV2-ORF RNAtranscripts in PK-15 cells. M, 100 bp ladder; lanes 1, 4, 7, transient (48 h post-transfection) expression of ORF-1, -2 and -3 inPK15 cells; lanes 2, 5, 8, positive control; lanes 3, 6, 9, Mock control. (B) Transgenic protein expression of ORF-1, -2 and -3cDNAs in PK15 cells transfected for 48 h, as mock-transfected (a), ORF-1 (b), ORF-2 (c) and ORF-3 (d) analysed using IPMA.(C) Prophylactic model. Vaccination schedule consisted of three immunizations at 2-week intervals, followed by i.m. challenge with5 × 104.5 TCID50/ml of PCV2 in test mice
which the antibody titers gradually increased until week8–10 in all immunized mice (i.e. at 14 days after thesecond boost with respective MVA vectors encodingcorresponding PCV2 ORFs). After challenge with PCV2,we were able to detect weak antibody responses for all testPCV2 ORFs vaccines, which was also confirmed by IPMA.We also obtained a very weak antibody cross-reactionbetween the ORF-2 and -3 treated mice and the ORF-1and -3 treated mice, whereas no cross reaction was foundbetween the ORF-1 and -2 treated mice (Table 1). Todetermine the effect of PCV2 ORF antigens on antibodyisotype profiles of anti-PCV2 antibodies, the serum IgG1and IgG2a antibody titers of test gene-immunized orcontrol mice were analysed (Figure 2A). The humoralresponses of this PCV2 gene-based vaccination wereenhanced IgG2a level and concomitant decreased IgG1level. The shift in switching between Th1 and Th2
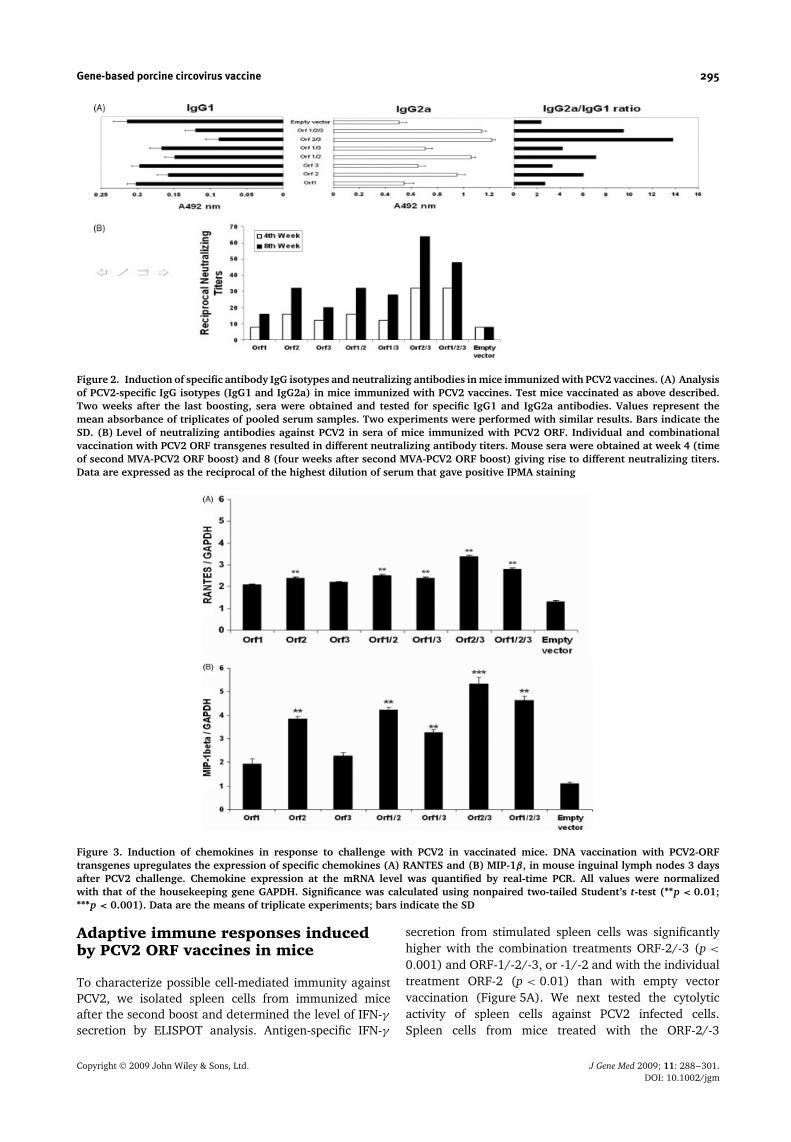
responses against PCV2-ORF antigens, reflected indirectlyby the altered IgG2a/IgG1 ratio, were found to be higherfollowing the combination treatment with ORF-2/-3 or -1/-2/-3, than with other tested individual or combinationtreatments (Figure 2A). The capacity of different ORF-expressing vectors to produce neutralizing antibodies intest mice was measured by serum titer (Figure 2B). Thegroups that received a boost with a MVA expressingthe same specific PCV2 ORF showed 50% antibodyneutralizing effect at titers of 1 : 8–1 : 32 from week 4.A second boost effectively enhanced this ratio to 1 : 64with the ORF-2/-3 and ORF-1/-2/-3 treatments at week8 (Figure 2B).
PCV2 ORF gene vaccinations increasechemokine expression and decreaseviral titers
The levels of the CC chemokines RANTES and MIP-1β
were enhanced in mice immunized with test PCV2-ORFvaccine after challenge with PCV2. The level of RANTESmRNA was significantly higher (approximately 2.5-fold)in the inguinal lymph nodes of mice vaccinated with ORF-2/-3 genes than in mice vaccinated with the empty vectorcontrol. Mice immunized with ORF-2 and combinationaltreatment either with ORF-1/-2, -1/-3 and -1/-2/-3also had significantly increased (p < 0.01) RANTESmRNA levels (Figure 3A). The expression of another CCchemokine, MIP-1β, was higher (approximately 4.9-foldhigher) in mice when vaccinated with ORF-2/-3 as wellas ORF-1/-2/-3 (approximately 4.27-fold higher) than inmice immunized with an empty vector. Of the individualtreatments, ORF-2 led to MIP-1β levels 3.5-fold higherthan with the empty vector (Figure 3B).
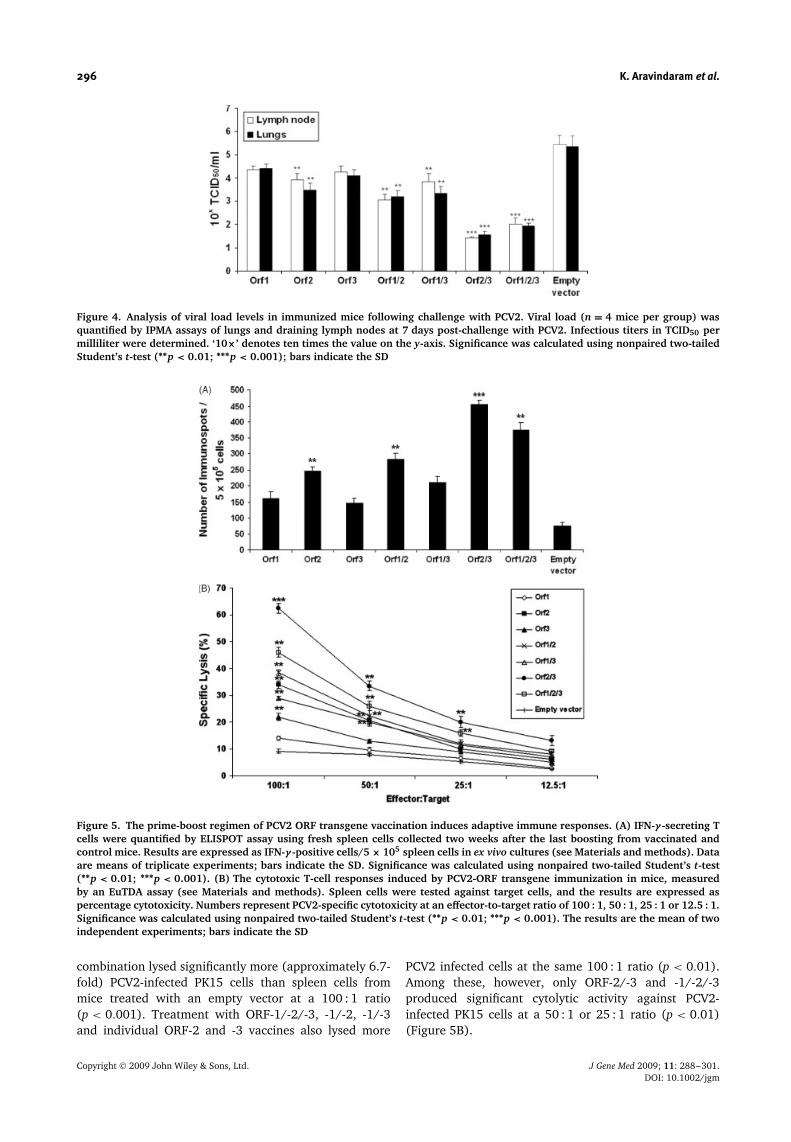
Next, we assessed the efficacy of PCV2-ORF vaccines forprotection against PCV2 infection. Immunized mice werechallenged i.m. with 5 × 104.5 TCID50/ml of PCV2. Oneweek after challenge, mice that were immunized withPCV2 ORF-2/-3 and -1/-2/-3 vaccines exhibited lowerPCV2 viral titers in lung and inguinal lymph nodes of testmice compared to other vaccine treatments (Figure 4).Immunization with PCV2 ORF-2 and the combinationaltreatment with ORF-1/-2 or -1/-3 led to significantly lowerPCV2 viral titers (p < 0.01) than mice vaccination withthe empty vector set (Figure 4).
Copyright 2009 John Wiley & Sons, Ltd. J Gene Med 2009; 11: 288–301.DOI: 10.1002/jgm
Gene-based porcine circovirus vaccine 295
Figure 2. Induction of specific antibody IgG isotypes and neutralizing antibodies in mice immunized with PCV2 vaccines. (A) Analysisof PCV2-specific IgG isotypes (IgG1 and IgG2a) in mice immunized with PCV2 vaccines. Test mice vaccinated as above described.Two weeks after the last boosting, sera were obtained and tested for specific IgG1 and IgG2a antibodies. Values represent themean absorbance of triplicates of pooled serum samples. Two experiments were performed with similar results. Bars indicate theSD. (B) Level of neutralizing antibodies against PCV2 in sera of mice immunized with PCV2 ORF. Individual and combinationalvaccination with PCV2 ORF transgenes resulted in different neutralizing antibody titers. Mouse sera were obtained at week 4 (timeof second MVA-PCV2 ORF boost) and 8 (four weeks after second MVA-PCV2 ORF boost) giving rise to different neutralizing titers.Data are expressed as the reciprocal of the highest dilution of serum that gave positive IPMA staining
Figure 3. Induction of chemokines in response to challenge with PCV2 in vaccinated mice. DNA vaccination with PCV2-ORFtransgenes upregulates the expression of specific chemokines (A) RANTES and (B) MIP-1β, in mouse inguinal lymph nodes 3 daysafter PCV2 challenge. Chemokine expression at the mRNA level was quantified by real-time PCR. All values were normalizedwith that of the housekeeping gene GAPDH. Significance was calculated using nonpaired two-tailed Student’s t-test (∗∗p < 0.01;∗∗∗p < 0.001). Data are the means of triplicate experiments; bars indicate the SD
Adaptive immune responses inducedby PCV2 ORF vaccines in mice
To characterize possible cell-mediated immunity againstPCV2, we isolated spleen cells from immunized miceafter the second boost and determined the level of IFN-γsecretion by ELISPOT analysis. Antigen-specific IFN-γ
secretion from stimulated spleen cells was significantlyhigher with the combination treatments ORF-2/-3 (p <
0.001) and ORF-1/-2/-3, or -1/-2 and with the individualtreatment ORF-2 (p < 0.01) than with empty vectorvaccination (Figure 5A). We next tested the cytolyticactivity of spleen cells against PCV2 infected cells.Spleen cells from mice treated with the ORF-2/-3
Copyright 2009 John Wiley & Sons, Ltd. J Gene Med 2009; 11: 288–301.DOI: 10.1002/jgm
296 K. Aravindaram et al.
Figure 4. Analysis of viral load levels in immunized mice following challenge with PCV2. Viral load (n = 4 mice per group) wasquantified by IPMA assays of lungs and draining lymph nodes at 7 days post-challenge with PCV2. Infectious titers in TCID50 permilliliter were determined. ‘10×’ denotes ten times the value on the y-axis. Significance was calculated using nonpaired two-tailedStudent’s t-test (∗∗p < 0.01; ∗∗∗p < 0.001); bars indicate the SD
Figure 5. The prime-boost regimen of PCV2 ORF transgene vaccination induces adaptive immune responses. (A) IFN-γ -secreting Tcells were quantified by ELISPOT assay using fresh spleen cells collected two weeks after the last boosting from vaccinated andcontrol mice. Results are expressed as IFN-γ -positive cells/5 × 105 spleen cells in ex vivo cultures (see Materials and methods). Dataare means of triplicate experiments; bars indicate the SD. Significance was calculated using nonpaired two-tailed Student’s t-test(∗∗p < 0.01; ∗∗∗p < 0.001). (B) The cytotoxic T-cell responses induced by PCV2-ORF transgene immunization in mice, measuredby an EuTDA assay (see Materials and methods). Spleen cells were tested against target cells, and the results are expressed aspercentage cytotoxicity. Numbers represent PCV2-specific cytotoxicity at an effector-to-target ratio of 100 : 1, 50 : 1, 25 : 1 or 12.5 : 1.Significance was calculated using nonpaired two-tailed Student’s t-test (∗∗p < 0.01; ∗∗∗p < 0.001). The results are the mean of twoindependent experiments; bars indicate the SD
combination lysed significantly more (approximately 6.7-fold) PCV2-infected PK15 cells than spleen cells frommice treated with an empty vector at a 100 : 1 ratio(p < 0.001). Treatment with ORF-1/-2/-3, -1/-2, -1/-3and individual ORF-2 and -3 vaccines also lysed more
PCV2 infected cells at the same 100 : 1 ratio (p < 0.01).Among these, however, only ORF-2/-3 and -1/-2/-3produced significant cytolytic activity against PCV2-infected PK15 cells at a 50 : 1 or 25 : 1 ratio (p < 0.01)(Figure 5B).
Copyright 2009 John Wiley & Sons, Ltd. J Gene Med 2009; 11: 288–301.DOI: 10.1002/jgm

Gene-based porcine circovirus vaccine 297
Effect of PCV2 ORF vaccination oncytokine expression profile
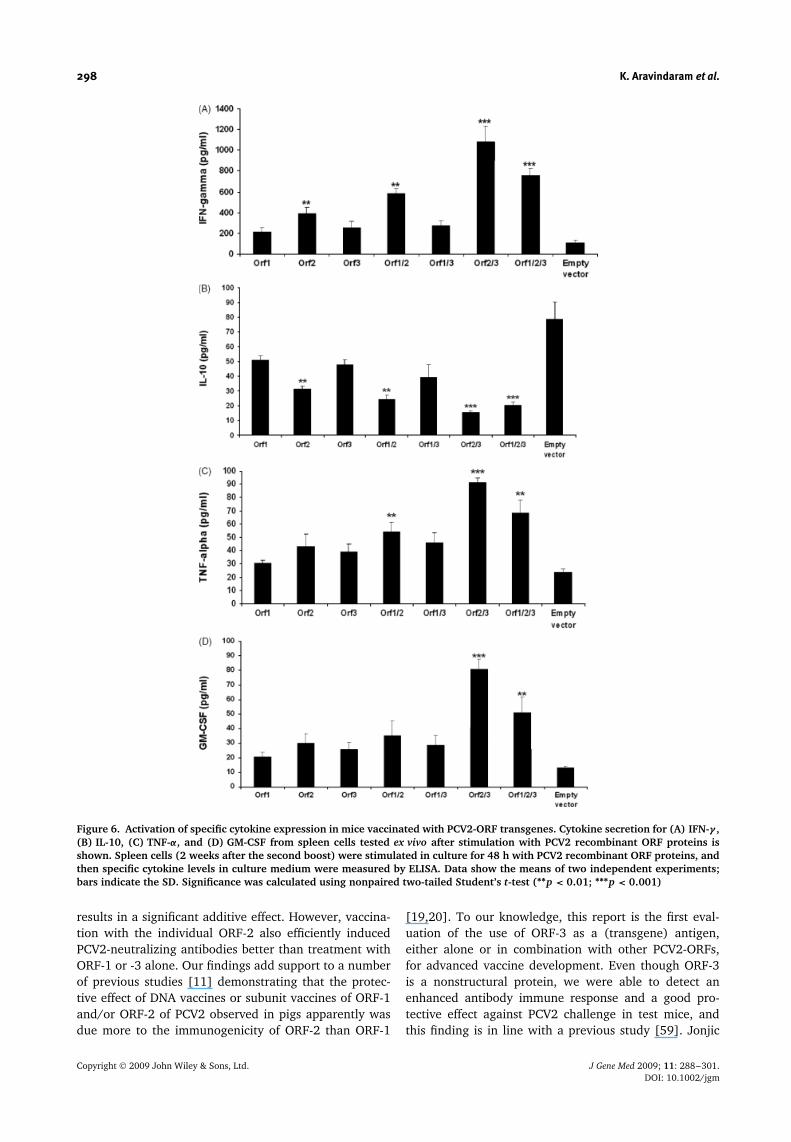
To further characterize the immune response of PCV2ORF-vaccinated mice, we analysed (using ELISA) theexpression/secretion patterns of the cytokines IFN-γ , IL-10, TNF-α and GM-CSF, 48 h after stimulation of spleencells with PCV2 recombinant ORF proteins. Treatmentwith ORF-2/-3 and -1/-2/-3 produced significantly more(p < 0.001) IFN-γ secretion, followed by treatment withORF-1/-2 and -2 (p < 0.01) than empty-vector controltreatment (Figure 6A). To determine the shift betweenTh1 versus Th2 response preference, we also analysedthe effect on cytokine IL-10 secretion. Interestingly, IL-10 release was substantially decreased in sera of micetreated with ORF-2/-3 and -1/-2/-3 transgene vectors(p < 0.001), and with ORF-1/-2 and -2 vectors (p < 0.01)compared to the empty vector-treated control mice(Figure 6B). This cytokine profile (with increased Th1and decreased Th2 responses) in PCV2-ORF-vaccinatedmice was tightly associated with an increased IgG2a/IgG1ratio and decreased viral titers in lungs and inguinallymph nodes of test mice. This suggests that Th1-typecytokines are apparently involved in the PCV2-ORFvaccine-mediated immunity against PCV2. Because theTh1 immune response is known to induce class switchingto an IgG2a isotype in B cells, our results suggest thatthe PCV2-ORF transgene vaccines apparently can elicit animmune response preferentially tilted towards the Th1,but not the Th2 type pathway.
The level of two other important cytokines, TNF-αand GM-CSF, was also elevated in level more by theORF-2/-3 and -1/-2/-3 combinational treatments thanwith individual ORF vaccination (Figures 6C and 6D).Secretion of the pro-inflammatory cytokine TNF-α wassignificantly higher in mice treated with ORF-2/-3 (p <
0.001) and by treatment with ORF-1/-2/-3 and -1/-2(p < 0.01) than in mice immunized with empty vectoronly (Figure 6C). Secretion of GM-CSF was significantlyincreased with ORF-2/-3 (p < 0.001) and ORF-1/-2/-3treatment (p < 0.01) (Figure 6D).
Vaccination with PCV2 ORF genevectors reduces apoptosis in lungs ofPCV2-challenged mice
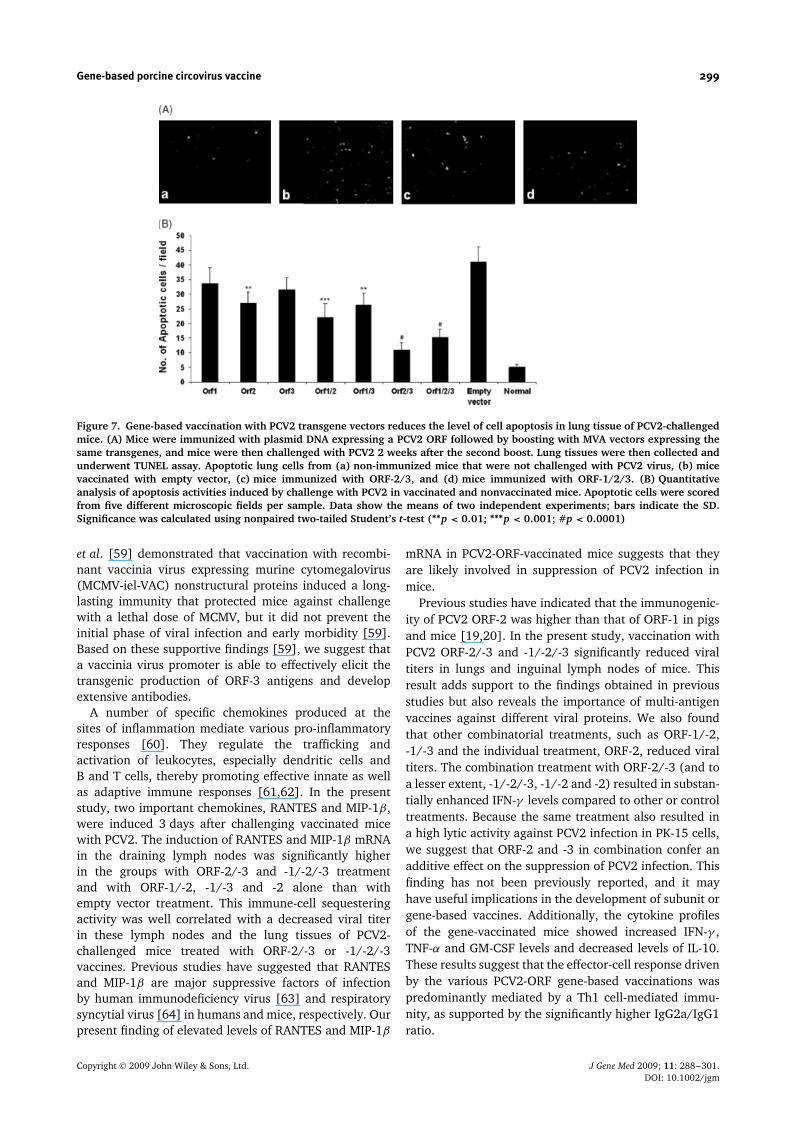
To investigate a possible role of test PCV2-ORF vaccinein protecting lung tissues of mice challenged with PCV2,we quantified the level of apoptotic cells in mouse lungsby TUNEL assay (Figure 7A). Mice vaccinated with ORF-2/-3 and -1/-2/-3 had significantly fewer apoptotic cellsin the lungs, which was associated with reduced viralload, and suggests that this combined ORF vaccinationwas highly protective against PCV2 disease in test mice.The number of apoptotic cells was lower in ORF-2/-3 and-1/-2/-3-treated mice (p < 0.0001) than in mice treatedwith empty vector only (Figure 7B), with no significantdifference between these two treatment groups.
Discussion
In the present study, we developed a recombinant plasmidDNA vector system for expressing PCV2-ORFs -1, -2and -3, to prime host mice, followed by boosting withthe same antigen or antigen combinations constructedinto recombinant MVA vectors, to initiate a completeimmune response. Mice immunized with the specific gene-based vaccines containing plasmid DNA/MVA-PCV2 ORFconstructs generated significant levels of PCV2-specificantibodies, especially from vaccination with PCV2 ORF-2/-3 and -1/-2/-3: the PCV2 ORF-2/-3 combination wasadditive, and had a significantly better effect than othertreatments. To date, little information is available onthe host range of PCV, but antibodies have indeed beendetected in various species other than pigs, includinghumans, mice, and cattle [54,55]. Kiupel et al. [56]further demonstrated that PCV2 was capable of replicatingin Balb/c mice and caused microscopic lesions similarto those resulting from PCV2 infection in pigs. Bycontrast, Quintana et al. [57] reported that no microscopiclesions compatible with the PCV2 infections in pigswere detectable in inoculated mice, but this result wasconsidered to be due to the dosage of the inoculumand administration route used [57]. In our current PCV2vaccination study, we used Balb/c mice and found thatinduction of antibody responses was readily observed.This apparently plays a key role in the protection againstPCV2, especially in mice vaccinated with PCV2 ORF-2/-3and -1/-2/-3 vectors, which showing greatly reduced viraltiters in the inguinal lymph nodes and lungs of test miceupon challenge with PCV2. Antibody isotypes play a majorrole in the mechanism of protection against PCV2 in testmice: we found the IgG2a/IgG1 ratio to be higher in all themice vaccinated with PCV2-ORFs, which suggests that themajor induced IgG isotype was IgG2a. We believe that theobserved immune response against PCV2 was mediatedmainly by Th1 but not Th2 cells, because we also observeda tight positive correlation between the IgG switchingtowards IgG2a (Th1) and other immune-related activities,including enhanced Th1 cytokine (IFN-γ ) expression anddecreased IgG1 (Th2) isotype levels with correspondingdecreased Th2 cytokine (IL-10) expression in PCV2 ORFtransgene immunized mice.
An absence of PCV2-neutralizing antibodies is cor-related with PCV2 replication and symptoms of PCV2-related disease (PMWS), which suggests that increasingthe level of PCV2-neutralizing antibodies could enhanceprotection against PCV2 infection [58]. In the presentstudy of outcomes of PCV2 infection in our test experi-mental mice, the result is nonlethal, in agreement withprevious findings [49]. Mice immunized with variousPCV2-ORFs showed PCV2-neutralizing antibody levels:mice treated with the combinations ORF-2/-3 and -1/-2/-3, and to a lesser extent ORF-1/-2 and -2, hadsubstantially more neutralizing antibodies than mice inoc-ulated with individual ORF-1 or -3. Thus, we suggestthat the use of ORF-2/-3 or -1/-2/-3 in combination
Copyright 2009 John Wiley & Sons, Ltd. J Gene Med 2009; 11: 288–301.DOI: 10.1002/jgm
298 K. Aravindaram et al.
Figure 6. Activation of specific cytokine expression in mice vaccinated with PCV2-ORF transgenes. Cytokine secretion for (A) IFN-γ ,(B) IL-10, (C) TNF-α, and (D) GM-CSF from spleen cells tested ex vivo after stimulation with PCV2 recombinant ORF proteins isshown. Spleen cells (2 weeks after the second boost) were stimulated in culture for 48 h with PCV2 recombinant ORF proteins, andthen specific cytokine levels in culture medium were measured by ELISA. Data show the means of two independent experiments;bars indicate the SD. Significance was calculated using nonpaired two-tailed Student’s t-test (∗∗p < 0.01; ∗∗∗p < 0.001)
results in a significant additive effect. However, vaccina-tion with the individual ORF-2 also efficiently inducedPCV2-neutralizing antibodies better than treatment withORF-1 or -3 alone. Our findings add support to a numberof previous studies [11] demonstrating that the protec-tive effect of DNA vaccines or subunit vaccines of ORF-1and/or ORF-2 of PCV2 observed in pigs apparently wasdue more to the immunogenicity of ORF-2 than ORF-1
[19,20]. To our knowledge, this report is the first eval-uation of the use of ORF-3 as a (transgene) antigen,either alone or in combination with other PCV2-ORFs,for advanced vaccine development. Even though ORF-3is a nonstructural protein, we were able to detect anenhanced antibody immune response and a good pro-tective effect against PCV2 challenge in test mice, andthis finding is in line with a previous study [59]. Jonjic
Copyright 2009 John Wiley & Sons, Ltd. J Gene Med 2009; 11: 288–301.DOI: 10.1002/jgm
Gene-based porcine circovirus vaccine 299
Figure 7. Gene-based vaccination with PCV2 transgene vectors reduces the level of cell apoptosis in lung tissue of PCV2-challengedmice. (A) Mice were immunized with plasmid DNA expressing a PCV2 ORF followed by boosting with MVA vectors expressing thesame transgenes, and mice were then challenged with PCV2 2 weeks after the second boost. Lung tissues were then collected andunderwent TUNEL assay. Apoptotic lung cells from (a) non-immunized mice that were not challenged with PCV2 virus, (b) micevaccinated with empty vector, (c) mice immunized with ORF-2/3, and (d) mice immunized with ORF-1/2/3. (B) Quantitativeanalysis of apoptosis activities induced by challenge with PCV2 in vaccinated and nonvaccinated mice. Apoptotic cells were scoredfrom five different microscopic fields per sample. Data show the means of two independent experiments; bars indicate the SD.Significance was calculated using nonpaired two-tailed Student’s t-test (∗∗p < 0.01; ∗∗∗p < 0.001; #p < 0.0001)
et al. [59] demonstrated that vaccination with recombi-nant vaccinia virus expressing murine cytomegalovirus(MCMV-iel-VAC) nonstructural proteins induced a long-lasting immunity that protected mice against challengewith a lethal dose of MCMV, but it did not prevent theinitial phase of viral infection and early morbidity [59].Based on these supportive findings [59], we suggest thata vaccinia virus promoter is able to effectively elicit thetransgenic production of ORF-3 antigens and developextensive antibodies.
A number of specific chemokines produced at thesites of inflammation mediate various pro-inflammatoryresponses [60]. They regulate the trafficking andactivation of leukocytes, especially dendritic cells andB and T cells, thereby promoting effective innate as wellas adaptive immune responses [61,62]. In the presentstudy, two important chemokines, RANTES and MIP-1β,were induced 3 days after challenging vaccinated micewith PCV2. The induction of RANTES and MIP-1β mRNAin the draining lymph nodes was significantly higherin the groups with ORF-2/-3 and -1/-2/-3 treatmentand with ORF-1/-2, -1/-3 and -2 alone than withempty vector treatment. This immune-cell sequesteringactivity was well correlated with a decreased viral titerin these lymph nodes and the lung tissues of PCV2-challenged mice treated with ORF-2/-3 or -1/-2/-3vaccines. Previous studies have suggested that RANTESand MIP-1β are major suppressive factors of infectionby human immunodeficiency virus [63] and respiratorysyncytial virus [64] in humans and mice, respectively. Ourpresent finding of elevated levels of RANTES and MIP-1β
mRNA in PCV2-ORF-vaccinated mice suggests that theyare likely involved in suppression of PCV2 infection inmice.
Previous studies have indicated that the immunogenic-ity of PCV2 ORF-2 was higher than that of ORF-1 in pigsand mice [19,20]. In the present study, vaccination withPCV2 ORF-2/-3 and -1/-2/-3 significantly reduced viraltiters in lungs and inguinal lymph nodes of mice. Thisresult adds support to the findings obtained in previousstudies but also reveals the importance of multi-antigenvaccines against different viral proteins. We also foundthat other combinatorial treatments, such as ORF-1/-2,-1/-3 and the individual treatment, ORF-2, reduced viraltiters. The combination treatment with ORF-2/-3 (and toa lesser extent, -1/-2/-3, -1/-2 and -2) resulted in substan-tially enhanced IFN-γ levels compared to other or controltreatments. Because the same treatment also resulted ina high lytic activity against PCV2 infection in PK-15 cells,we suggest that ORF-2 and -3 in combination confer anadditive effect on the suppression of PCV2 infection. Thisfinding has not been previously reported, and it mayhave useful implications in the development of subunit orgene-based vaccines. Additionally, the cytokine profilesof the gene-vaccinated mice showed increased IFN-γ ,TNF-α and GM-CSF levels and decreased levels of IL-10.These results suggest that the effector-cell response drivenby the various PCV2-ORF gene-based vaccinations waspredominantly mediated by a Th1 cell-mediated immu-nity, as supported by the significantly higher IgG2a/IgG1ratio.
Copyright 2009 John Wiley & Sons, Ltd. J Gene Med 2009; 11: 288–301.DOI: 10.1002/jgm

300 K. Aravindaram et al.
In conclusion, the present study has shown well-coordinated immune responses to an advanced gene-based vaccine strategy, such as enhanced levels of PCV2-specific antibodies, elevated levels of chemokines RANTESand MIP-1β, and secretion of cytokines, all of which ledto decreased PCV2 viral titers and reduced the numberof apoptotic cells in the lungs of vaccinated mice. Thecombination of ORF-2 and -3 is apparently additive inthe protection of test mice against PCV2 infection, andenhances both the humoral and cell-mediated immuneresponses. The use of this specific prime-boost regimenwith a plasmid DNA and a MVA vector system incombination for expressing specific PCV2-ORF-2 and -3antigens could be an attractive vaccine strategy forpreventing PCV2 infection and the resulting PMWSdisease, and its effectiveness should be systematicallyexamined in the swine system, sooner rather than later.
Acknowledgements
This work was supported by a grant (95N-1003) from theNational Science and Technology Program for AgriculturalBiotechnology, Taiwan. We also thank Dr Harry Wilson andMs Laura Heraty for professional editing of this manuscript.
References
1. Tischer I, Gelderblom H, Vettermann W, Koch MA. A very smallporcine virus with circular single-stranded DNA. Nature 1982;295: 64–66.
2. Clark EG. Post-weaning multisystemic wasting syndrome. ProcAm Assoc Swine Pract 1997; 28: 499–501.
3. Allan GM, McNeilly F, Kennedy S, et al. Isolation of porcinecircovirus-like viruses from pigs with a wasting disease in theUSA and Europe. J Vet Diagn Invest 1998; 10: 3–10.
4. Choi C, Chae C, Clark EG. Porcine postweaning multisystemicwasting syndrome in Korean pig: detection of porcine circovirus2 infection by immunohistochemistry and polymerase chainreaction. J Vet Diagn Invest 2000; 12: 151–153.
5. Fenaux M, Halbur PG, Gill M, Toth TE, Meng XJ. Geneticcharacterization of type 2 porcine circovirus (PCV-2) from pigswith postweaning multisystemic wasting syndrome in differentgeographic regions of North America and development of adifferential PCR-restriction fragment length polymorphism assayto detect and differentiate between infections with PCV-1 andPCV-2. J Clin Microbiol 2000; 38: 2494–2503.
6. Mankertz A, Domingo M, Folch JM, et al. Characterization ofPCV2 isolates from Spain, Germany, and France. Virus Res 2000;66: 65–77.
7. Kim J, Chung HK, Jung T, Cho WS, Choi C, Chae C.Postweaning multisystemic wasting syndrome of pigs in Korea:prevalence, microscopic lesions and coexisting microorganisms.J Vet Med Sci 2002; 64: 57–62.
8. Balasch M, Segales J, Rosell C, et al. Experimental inoculationof conventional pigs with tissue homogenates from pigs withpostweaning multisystemic wasting syndrome. J Comp Pathol1999; 121: 139–148.
9. Albina E, Truong C, Hutet E, et al. An experimental modelfor postweaning multisystemic wasting syndrome (PMWS) ingrowing piglets. J Comp Pathol 2001; 125: 292–303.
10. Bolin SR, Stoffregen WC, Nayar GP, Hamel AL. Postweaningmultisystemic wasting syndrome induced after experimentalinoculation of cesarean-derived, colostrum-deprived piglets withtype 2 porcine circovirus. J Vet Diagn Invest 2001; 13: 185–194.
11. Fenaux M, Opriessnig T, Halbur PG, Elvinger F, Meng XJ. Achimeric porcine circovirus (PCV) with the immunogenic capsidgene of the pathogenic PCV type 2 (PCV2) cloned intothe genomic backbone of the nonpathogenic PCV1 induces
protective immunity against PCV2 infection in pigs. J Virol2004; 78: 6297–6303.
12. Liu J, Chen I, Du Q, Chua H, Kwang J. The ORF-3 protein ofporcine circovirus type 2 is involved in viral pathogenesis in vivo.J Virol 2006; 80: 5065–5073.
13. Opriessnig T, Yu S, Gallup JM, et al. Effect of vaccination withselective bacterins on conventional pigs infected with type 2porcine circovirus. Vet Pathol 2003; 40: 521–529.
14. Segales J, Domingo M. Porcine dermatitis and nephropathysyndrome: a porcine circovirus type 2 infection disease. PMWSSymposium, 16th International Pig Veterinarians Society Congress.Melbourne, Australia, 2000; 21–31.
15. Cheung AK. Transcriptional analysis of porcine circovirus.Virology 2003; 305: 168–180.
16. Mankertz A, Mankertz J, Wolf K, Buhk HJ. Identification of aprotein essential for replication of porcine circovirus. J Gen Virol1998; 79: 381–383.
17. Nawagitgul P, Morozov I, Bolin SR, Harms PA, Sorden SD. Openreading frame 2 of porcine cicovirus type 2 encodes a majorcapsid protein. J Gen Virol 2000; 81: 2281–2287.
18. Liu J, Chen I, Kwang J. Characterization of a previouslyunidentified viral protein in porcine circovirus type 2-infectedcells and its role in virus-induced apoptosis. J Virol 2005; 79:8262–8274.
19. Blanchard P, Mahe D, Cariolet R, et al. Protection of swineagainst post-weaning multisystemic wasting syndrome (PMWS)by porcine circovirus type 2 (PCV2) proteins. Vaccine 2003; 21:4565–4575.
20. Kamstrup S, Barfoed AM, Frimann TH, Mikkelsen ASL,Bøtner A. Immunization against PCV2 structural protein by DNAvaccination of mice. Vaccine 2004; 22: 1358–1361.
21. Wang X, Jiang P, Li Y, Jiang W, Dong X. Protection of pigsagainst post-weaning multisystemic wasting syndrome by arecombinant adenovirus expressing the capsid protein of porcinecircovirus type 2. Vet Microbiol 2007; 121: 215–224.
22. Stickl H, Hochstein-Mintzel V, Mayr A, Huber HC, Schafer H,Holzner A. MVA vaccination against smallpox: clinical testswith an attenuated live vaccinia virus strain (MVA) [author’stranslation]. Dtsch Med Wochenschr 1974; 99: 2386–2392.
23. Meyer H, Sutter G, Mayr A. Mapping of deletions in the genomeof the highly attenuated vaccinia virus MVA and their influenceon virulence. J Gen Virol 1991; 72: 1031–1038.
24. Moss B. Replicating and host-restricted non-replicating vacciniavirus vectors for vaccine development. Dev Biol Stand 1994; 82:55–63.
25. Parrino J, McCurdy LH, Larkin DB, et al. Safety, immunogenicityand efficacy of modified vaccinia Ankara (MVA) against Dryvaxchallenge in vaccinia-naive and vaccinia-immune individuals.Vaccine 2007; 25: 1513–1525.
26. Amara RR, Villinger F, Altman JD, et al. Control of a mucosalchallenge and prevention of AIDS by a multiprotein DNA/MVAvaccine. Science 2001; 292: 69–74.
27. Feng CG, Blanchard TJ, Smith GL, Hill AV, Britton WJ.Induction of CD8+ T-lymphocyte responses to a secreted antigenof Mycobacterium tuberculosis by an attenuated vaccinia virus.Immunol Cell Biol 2001; 79: 569–575.
28. McConkey SJ, Reece WH, Moorthy VS, et al. Enhanced T-cell immunogenicity of plasmid DNA vaccines boosted byrecombinant modified vaccinia virus Ankara in humans. NatMed 2003; 9: 729–735.
29. McShane H. Developing an improved vaccine againsttuberculosis. Expert Rev Vaccines 2004; 3: 299–306.
30. Mwau M, Cebere I, Sutton J, et al. A human immunodeficiencyvirus 1 (HIV-1) clade A vaccine in clinical trials: stimulation ofHIV-specific T-cell responses by DNA and recombinant modifiedvaccinia virus Ankara (MVA) vaccines in humans. J Gen Virol2004; 85: 911–919.
31. Sutter G, Staib C. Vaccinia vectors as candidate vaccines: thedevelopment of modified vaccinia virus Ankara for antigendelivery. Curr Drug Targets Infect Disord 2003; 3: 263–271.
32. Vollmar J, Arndtz N, Eckl KM, et al. Safety and immunogenicityof IMVAMUNE, a promising candidate as a third generationsmallpox vaccine. Vaccine 2005; 24: 2065–2070.
33. Harrop R, Connolly N, Redchenko I, et al. Vaccination ofcolorectal cancer patients with modified vaccinia Ankaradelivering the tumor antigen 5T4 (TroVax) induces immuneresponses which correlate with disease control: a phase I/IItrial. Clin Cancer Res 2006; 12: 3416–3424.
Copyright 2009 John Wiley & Sons, Ltd. J Gene Med 2009; 11: 288–301.DOI: 10.1002/jgm

Gene-based porcine circovirus vaccine 301
34. Drexler I, Staib C, Sutter G. Modified vaccinia virus Ankara asantigen delivery system: how can we best use its potential? CurrOpin Biotechnol 2004; 15: 506–512.
35. Cosma A, Nagaraj R, Buhler S, et al. Therapeutic vaccinationwith MVA-HIV-1 nef elicits Nef-specific T-helper cell responsesin chronically HIV-1 infected individuals. Vaccine 2003; 22:21–29.
36. Takada A, Kuboki N, Okazaki K, et al. A virulent avian influenzavirus as a vaccine strain against a potential human pandemic. JVirol 1999; 73: 8303–8307.
37. Staib C, Drexler I, Sutter G. Construction and isolationof recombinant MVA. Methods Mol Biol 2004; 269:77–100.
38. Moss B, Carroll MW, Wyatt LS, et al. Host range restricted, non-replicating vaccinia virus vectors as vaccine candidates. Adv ExpMed Biol 1996; 397: 7–13.
39. Drexler I, Heller K, Wahren B, Erfle V, Sutter G. Highlyattenuated modified vaccinia virus Ankara replicates in babyhamster kidney cells, a potential host for virus propagation, butnot in various human transformed and primary cells. J Virol1998; 79: 347–352.
40. Schneider J, Gilbert SC, Blanchard TJ, et al. Enhancedimmunogenicity for CD8+ T cell induction and completeprotective efficacy of malaria DNA vaccination by boosting withmodified vaccinia virus Ankara. Nat Med 1998; 5: 397–402.
41. McShane H, Brookes R, Gilbert SC, Hill AVS. Enhancedimmunogenicity of CD4+ T-cell responses and protectiveefficacy of a DNA-modified vaccinia virus Ankara prime-boostvaccination regimen for murine tuberculosis. Infect Immun 2001;69: 681–686.
42. McNair I, Marshall M, McNeilly F, et al. Interlaboratory testingof porcine sera for antibodies to porcine circovirus type 2. J VetDiagn Invest 2004; 16: 164–166.
43. Wyatt LS, Shors ST, Murphy BR, Moss B. Development of areplication-deficient recombinant vaccinia virus vaccine effectiveagainst parainfluenza virus 3 infection in an animal model.Vaccine 1996; 14: 1451–1458.
44. Earl PL, Moss B, Wyatt LS, et al. Generation of recombinantvaccinia virus. In Current Protocols in Molecular Biology,Ausubel FM, Brent R, Kingston RE, et al (eds), Vol. 2. GreenePublishing Associates & Wiley Interscience: New York, NY, 1998;16.17.1–16.17.19.
45. Rakhmilevich AL, Turner J, Ford MJ, et al. Gene gun-mediatedskin transfection with interleukin 12 gene results in regressionof established primary and metastatic murine tumors. Proc NatlAcad Sci USA 1996; 93: 6291–6296.
46. Harrington LE, Most Rv R, Whitton JL, Ahmed R. Recombinantvaccinia virus-induced T-cell immunity: quantitation of theresponse to the virus vector and the foreign epitope. J Virol2002; 76: 3329–3337.
47. Tscharke DC, Karupiah G, Zhou J, et al. Identification ofpoxvirus CD8+ T cell determinants to enable rational designand characterization of smallpox vaccines. J Exp Med 2005;201: 95–104.
48. Staib C, Kisling S, Erfle V, Sutter G. Inactivation of the viralinterleukin 1β receptor improves CD8+ T-cell memory responseselicited upon immunization with modified vaccinia virus Ankara.J Gen Virol 2005; 86: 1997–2006.
49. Kiupel M, Stevenson GW, Choi J, Latimer KS, Kanitz CL,Mittal SK. Viral replication and lesions in BALB/c miceexperimentally inoculated with porcine circovirus isolated froma pig with post-weaning multisystemic wasting syndrome. VetPathol 2001; 38: 74–82.
50. Liu C, Ihara T, Nunoya T, Veda S. Development of an ELISAbased on the baculovirus-expressed capsid protein of porcinecircovirus type 2 as antigen. J Vet Med Sci 2004; 66: 237–242.
51. Herd KA, Harvey T, Khromykh AA, Tindle RW. RecombinantKunjin virus replicon vaccines induce protective T-cell immunityagainst human papillomavirus 16 E7-expressing tumor. Virology2004; 319: 237–248.
52. Stevenson PG, Doherty PC. Kinetic analysis of the specific hostresponse to a murine gamma herpesvirus. J Virol 1998; 72:943–949.
53. Acres B, Gantzer M, Remy C, et al. Fusokine interleukin-2/interleukin-18, a novel potent innate and adaptive immunestimulator with decreased toxicity. Cancer Res 2005; 65:9536–9546.
54. Nayar GP, Hamel AL, Linn L, Sachvie C, Grudeski E,Spearman G. Evidence for circovirus in cattle with respiratorydisease and from aborted bovine fetuses. Can Vet J 1999; 40:277–278.
55. Tischer I, Bode L, Apodaca J, et al. Presence of antibodiesreacting with porcine circovirus in sera of humans, mice, andcattle. Arch Virol 1995; 140: 1427–1439.
56. Kiupel M, Stevenson GW, Choi J, Latimer KS, Kanitz CL,Mittal SK. Viral replication and lesions in BALB/c miceexperimentally inoculated with porcine circovirus isolated froma pig with post-weaning multisystemic wasting syndrome. VetPathol 2001; 38: 74–82.
57. Quintana J, Balasch M, Segale’s J, et al. Experimentalinoculation of porcine circoviruses type 1 (PCV1) and type 2(PCV2) in rabbits and mice. Vet Res 2002; 33: 229–237.
58. Meerts P, Misinzo G, Lefebvre D, et al. Correlation between thepresence of neutralizing antibodies against porcine circovirustype 2 (PCV2) and protection against replication of the virusand development of PCV2-assoicated disease. BMC Vet Res 2006;30: 6–9.
59. Jonjic S, Val MD, Keil GM, Reddehase MJ, Koszinowski UH.A nonstructural viral protein expressed by a recombinantvaccinia virus protects against lethal cytomegalovirus infection.J Virol 1988; 62: 1653–1658.
60. Schall TJ. Biology of the RANTES/SIS cytokine family. Cytokine1991; 3: 165–183.
61. Ward SG, Westwick J. Chemokines: understanding their role inT-lymphocyte biology. J Biochem 1998; 333: 457.
62. Rossi D, Zlotnik A. The biology of chemokines and theirreceptors. Annu Rev Immunol 2000; 18: 217–42.
63. Cocchi F, DeVico AL, Garzino-Demo A, Arya SK, Gallo RC,Lusso P. Identification of RANTES, MIP-1α, and MIP-1β as themajor HIV-suppressive factors produced by CD8+ T cells. Science1995; 270: 1811–1815.
64. Power UF, Huss T, Michaud V, Plotnicky-Gilquin H, Bon-nefoy JY, Nguyen TN. Differential histopathology andchemokine gene expression in lung tissues following respiratorysyncytial virus (RSV) challenge of formalin-inactivated RSV- orBBG2Na-immunized mice. J Virol 2001; 75: 12421–12430.
Copyright 2009 John Wiley & Sons, Ltd. J Gene Med 2009; 11: 288–301.DOI: 10.1002/jgm



















