
Production and scavenging of reactive oxygen species in Fagus sylvatica seeds during storage at...
13
Journal of Plant Physiology 162 (2005) 873—885 Production and scavenging of reactive oxygen species in Fagus sylvatica seeds during storage at varied temperature and humidity Stanis$awa Pukacka , Ewelina Ratajczak Seed Biochemistry Laboratory, Institute of Dendrology, Polish Academy of Sciences, Parkowa 5, 62-035 Ko ´rnik, Poland Received 30 July 2004; accepted 6 October 2004 Summary The accumulation of reactive oxygen species (ROS) in seed tissues plays an important role in the loss of seed viability during storage. In the present study, we examined whether the loss of germination capacity and viability of beech (Fagus sylvatica L.) seeds during storage under different temperatures (4, 20 and 30 1C) and relative humidity levels (45% and 75% RH) is associated with: (1) an increase in the level of ROS, such as superoxide radical (O 2 d ), oxygen peroxide (H 2 O 2 ); and, (2) changes in low molecular antioxidants (ascorbate and glutathione) and enzymatic scavengers such as ascorbate peroxidase dehydroascorbate reductase, glutathione reductase, catalase, superoxide dismutase and guaiacol peroxidase. Beech seeds progressively lost their ability to germinate during 9 weeks of storage under the above conditions. The deleterious effects of temperature treatments increased with growing seed moisture content at higher humidity. The loss of seed viability was correlated with the generation of ROS during storage, which was more intensive at higher temperatures and humidity levels. The ascorbate content significantly increased in seeds stored in all temperature and humidity variants, when the seeds lost the ability to germinate to a large degree. At the same time, glutathione content dramatically decreased, but it was possible to observe a defensive reaction in seeds stored at 20 1C. Activities of all scavenging enzymes, measured after slow imbibition of seeds, significantly increased in comparison to the non-treated control (8–9% MC, 10 1C). This increase was higher in embryo axes than in cotyledons. Our results suggest that the loss of viability of beech seeds during storage at different temperatures, above zero, and at different humidity levels is closely related to ROS production, and that the antioxidative system is not sufficient to protect them. & 2005 Elsevier GmbH. All rights reserved. ARTICLE IN PRESS www.elsevier.de/jplph KEYWORDS Ascorbate; Beech; Enzymatic scaven- gers; Glutathione; Hydrogen peroxide; Seed germination; Superoxide radical; Viability 0176-1617/$ - see front matter & 2005 Elsevier GmbH. All rights reserved. doi:10.1016/j.jplph.2004.10.012 Corresponding author. Tel.: +48 061 8170 033; fax: +48 061 8170 166. E-mail address: [email protected] (S. Pukacka).
-
Upload
independent -
Category
Documents
-
view
0 -
download
0
Transcript of Production and scavenging of reactive oxygen species in Fagus sylvatica seeds during storage at...

ARTICLE IN PRESS
Journal of Plant Physiology 162 (2005) 873—885
KEYWORDAscorbateBeech;Enzymaticgers;GlutathionHydrogenSeed germSuperoxideViability
0176-1617/$ - sdoi:10.1016/j.
�CorrespondE-mail addr
www.elsevier.de/jplph
Production and scavenging of reactive oxygenspecies in Fagus sylvatica seeds during storageat varied temperature and humidity
Stanis$awa Pukacka�, Ewelina Ratajczak
Seed Biochemistry Laboratory, Institute of Dendrology, Polish Academy of Sciences, Parkowa 5, 62-035 Kornik, Poland
Received 30 July 2004; accepted 6 October 2004
S;
scaven-
e;peroxide;ination;radical;
ee front matter & 200jplph.2004.10.012
ing author. Tel.: +48 0ess: [email protected]
SummaryThe accumulation of reactive oxygen species (ROS) in seed tissues plays an importantrole in the loss of seed viability during storage. In the present study, we examinedwhether the loss of germination capacity and viability of beech (Fagus sylvatica L.)seeds during storage under different temperatures (4, 20 and 30 1C) and relativehumidity levels (45% and 75% RH) is associated with: (1) an increase in the level ofROS, such as superoxide radical (O2
d�), oxygen peroxide (H2O2); and, (2) changes inlow molecular antioxidants (ascorbate and glutathione) and enzymatic scavengerssuch as ascorbate peroxidase dehydroascorbate reductase, glutathione reductase,catalase, superoxide dismutase and guaiacol peroxidase. Beech seeds progressivelylost their ability to germinate during 9 weeks of storage under the above conditions.The deleterious effects of temperature treatments increased with growing seedmoisture content at higher humidity. The loss of seed viability was correlated with thegeneration of ROS during storage, which was more intensive at higher temperaturesand humidity levels. The ascorbate content significantly increased in seeds stored inall temperature and humidity variants, when the seeds lost the ability to germinate toa large degree. At the same time, glutathione content dramatically decreased, but itwas possible to observe a defensive reaction in seeds stored at 20 1C. Activities of allscavenging enzymes, measured after slow imbibition of seeds, significantly increasedin comparison to the non-treated control (8–9% MC, �10 1C). This increase was higherin embryo axes than in cotyledons. Our results suggest that the loss of viability ofbeech seeds during storage at different temperatures, above zero, and at differenthumidity levels is closely related to ROS production, and that the antioxidative systemis not sufficient to protect them.& 2005 Elsevier GmbH. All rights reserved.
5 Elsevier GmbH. All rights reserved.
61 8170 033; fax: +48 061 8170 166.oznan.pl (S. Pukacka).

ARTICLE IN PRESS
S. Pukacka, E. Ratajczak874
Introduction
Seeds deteriorate and lose their ability togerminate after prolonged storage. During the last25 years, extensive research has been conducted tounderstand better the physiology of seed aging, butprimary processes of aging damage are still notexplicitly defined (Walters, 1998; Mc Donald, 1999).Seed aging processes are controlled by temperatureand moisture conditions, and may be associatedwith various chemical reactions and metabolicalterations (Priestley, 1986; Vertucci and Ross,1990; Walters, 1998). The accumulation of reactiveoxygen species (ROS) is often indicated as theprime cause of seed deterioration, as they initiatereactions with polyunsaturated fatty acids, leadingto lipid peroxidation and destruction of cellularmembranes (Senaratna et al., 1988; Pukacka,1991). Apart from the techniques applied tomeasure lipid peroxidation, there are some meth-ods for detecting radicals themselves. Theseinclude biochemical methods, facilitating the de-termination of the level of superoxide (O2
d�),hydrogen peroxide (H2O2), hydroxyl radical (OH
d),or other radicals (Doke, 1983; Chaitanya andNaithani, 1994; Bailly et al., 2004); detection ofradicals by electron spin resonance (ESR) is alsopossible. In seeds of some species, the ESR responsedid not support the involvement of free radicals inseed deterioration (Priestley et al., 1980; Hepburnet al., 1986; Girard and Le Meste, 1992; Khan et al.,1996), but there have been also many positivecorrelations (Buchvarov and Gantcheff, 1984; Hen-dry et al., 1992; Hendry, 1993). To counteract toxicevents evoked through ROS, antioxidative defencesystems are present in plant tissues. These protec-tive systems are composed of low molecular andenzymatic scavengers, such as superoxide dismu-tase (SOD), catalase (CAT) and enzymes of theHalliwell–Asada pathway: ascorbate peroxidase(APO), dehydroascorbate reductase (DHAR) andglutathione reductase (GR), connected with anti-oxidant compounds: ascorbate and glutathione(Asada, 1992). The cooperative activities of theseenzymes and high levels of the low molecularantioxidants may increase the resistance to oxida-tive injury and minimize cell damage.
The common beech (Fagus ylvatica L.) is one ofthe most important broadleaved species in Eur-opean forestry. Beech is propagated by seeds, butseed set is irregular, with five to ten years betweencrops (Suszka et al., 1996). Consequently, it isnecessary to store the seeds. Beechnuts arecharacterized by a greater sensitivity to dryingand storage conditions compared to seeds ofanother species (Pukacka and Wojkiewicz, 2003).
Their viability can decline after shedding, whenthey are stored at temperatures above 0 1C, beforethe drying process. In addition, beechnuts are noteasy to store, or long-lived in storage. They exhibitstorage behaviour between those of ‘orthodox’ and‘recalcitrant’ seeds, the characteristics of whichwere described by Roberts (1973). Seeds withorthodox storage behaviour tolerate desiccationto moisture contents below 5%, and reducedstorage temperature and moisture content increasetheir longevity during storage. Recalcitrant seedsare characterised by an inability to withstanddesiccation below relatively high moisture content(40–20%) and they can be stored only for a shorttime. Beechnuts were classified as intermediate, orsuborthodox, by Bonner (1990) and Gosling (1991).However, Poulsen (1993) classified them in theorthodox category. Recently, Leon-Lobos andEllis (2002), on the basis of investigations of theirviability during storage at different moistureand temperature conditions, established that theoptimal temperature and moisture for storageof these seeds is 7.8�11.5% of water content and�10 to �20 1C. Pukacka et al. (2003) confirmedthese optimal conditions through investigationsof glass formation in embryo axes of beech seeds,and found that water sorption by these organsis greater than in cotyledons. The results indicatedthat the water relations in beech seeds arethe main cause of their sensitivity to storageconditions. The present study was initiated todetermine the relationship between the ROSproduction, the behaviour of the detoxifyingsystem, and the deterioration of the beech seedsduring storage under various temperature andhumidity conditions.
Materials and methods
Plant material and storage conditions
Beech seeds were collected in the autumn of2000 in the Krucz Forest District (NW Poland). Afterdesiccation to 9% MC, they were stored at �10 1C ina seed-bank in Bia"ogard. From this location, weobtained the seed lot (7 kg), collected from trees ofone population, and of equal age, for our experi-ments. Seeds were transported to our laboratory inplastic bags in a cooling box and stored inlaminated aluminium foil bags at �10 1C. Theywere of good quality and their germinabilityreached 93–96%.
To investigate the effect of temperature andhumidity, aliquots of seed samples were stored for

ARTICLE IN PRESS
Reactive oxygen species and seeds variability 875
1 week at two humidity levels: 45% and 75% RH(established above respective saturated salt solu-tions of K2CO3 and NaCl) (Scande et al., 2000), andat three temperatures: 4, 20 and 30 1C. After the 1week period, seeds were placed in laminatedaluminum foil bags, hermetically sealed and storedfor an additional 8 weeks at respective tempera-tures. After 3, 6 and 9 weeks of storage, seeds wereused for analyses. Untreated seeds stored at �10 1Cwere the control. Their viability maintained on thissame high level over a period of 2 years.
Germination test
Samples of 2� 100 seeds from each variant wereallowed to germinate at 3 1C, after careful imbibi-tion, between moist rolled paper towels in separateboxes. Normal germination was assessed accordingto ISTA (1999).
Superoxide determination
Superoxide production by seeds was assayed inaccordance with the method of Doke (1983), by thecapacity to reduce of nitroblue tetrazolium in thedark at room temperature. The absorbance of theend product was measured at 530 nm. O2
d� forma-tion was expressed as DA530/g DW of the sample.
Evaluation of hydrogen peroxide content
Seed samples of five seeds each were ground to afine powder in liquid nitrogen. Next, they werehomogenized with 5mL of 5% trichloroacetic acid(TCA) containing 10mmol/L EDTA. The homogenatewas filtered through two layers of cheesecloth andthen centrifuged at 4 1C at 12000g for 15min. Thetotal amount of supernatant was analysed using theferrithiocyanate method according to Sagisaka(1976).
Ascorbic acid (ASA) assay
ASA content was assayed according to Kampfen-kel et al. (1995). Samples of 15 embryo axes andfive cotyledons were taken for analyses. Theextraction was performed in 6% TCA (w/v) in anice bath. The assay is based on the reduction ofFe3+–Fe2+ by ASA in an acidic solution. Fe2+ formscomplexes with bipyridyl, giving a pink color withthe maximum absorbance at 525 nm.
Glutathione assay
Glutathione in the reduced (GSH) and oxidized(GSSG) forms were assayed according to Smith(1985). The samples of 15 embryo axes and fivecotyledons were homogenized in 5% (w/v) sulpho-salicylic acid on an ice bath and centrifuged at10,000g for 20min. A 1mL aliquot of the super-natant was removed and neutralized by adding1.5mL of 0.5mol/L K-phosphate buffer pH 7.5. Thissample was used for the determination of totalglutathione (GSH+GSSG). Another 1mL of neutra-lized supernatant was pretreated with 0.2mL of2-vinylpyridine for 1.5 h at 25 1C to mask GSH and toallow determination of GSSG alone. Both sampleswere extracted twice with 5mL diethylether. Theincubation mixture contained: 0.5mL of 0.1mol/Lsodium phosphate buffer (pH 7.5) containing5mmol/L EDTA, 0.2mL of 6mmol/L 5,50-dithiobis-(2-nitrobenzoic acid), 0.1mL of 2mmol/L NADPH,0.1mL (1 unit) of glutathione reductase type III(Sigma Chemical Co), and 0.1 mL of extract. Thechange in A at 412 nm was followed at 25 1C. Astandard curve was prepared by using the GSHstandard.
Enzyme extraction and assay
All enzyme assays were performed in the samecrude extract. All extraction procedures werecarried out at 4 1C. The samples of 50 embryo axesand 10 cotyledons were ground in liquid nitrogenand homogenized in 50mmol/L sodium phosphatebuffer, pH 7.0 containing 0.2mmol/L EDTA and 20%polyvinylpolypyrrolidone (PVPP) for 10min. Homo-genates were filtered through two layers of cheese-cloth and centrifuged at 4 1C at 15,000g for 20min.Supernatant was desalted on Sephadex G50 columnaccording to Helmerhorst and Stokes (1980).
Ascorbate peroxidase (APO, EC.1.11.1.11) activ-ity was measured by following the decreasein absorbance at 290 nm due to ascorbic acidoxidation for 5–10min, according to Nakano andAsada (1981). The reaction mixture contained: 1mLof 0.68mmol/L ascorbic acid and 0.1mmol/L EDTAin 0.1mol/L phosphate buffer pH 7.0, 1mL of4mmol/L H2O2 and 50–100 mL of enzyme extract.APO activity was expressed as nmol ASA min/mg/protein.
Dehydroascorbate reductase (DHAR, EC. 1.8.5.1)activity was measured according to Arrigoni et al.(1992). The reaction mixture contained: 0.9mL of0.05M potassium phosphate buffer pH 6.3, 100 mLof 13.5mmo/L dehydroascorbate (DHASA), 100 mLof 13.5mmol/L reduced glutathione (GSH) and

ARTICLE IN PRESS
Figure 1. The decline in seed germination after storageof 9 weeks at temperatures of 4, 20, 30 1C and humiditiesof 45% and 75% RH.
S. Pukacka, E. Ratajczak876
100–200 mL of enzyme extract. The DHAR activitywas monitored by following the formation ofascorbate at 265 nm for 5min. Enzyme activitywas expressed as nmol ASA/min/mg protein.
Glutathione reductase (GR, EC. 1.6.4.2) activitywas determined according to Esterbauer and Grill(1978), by following the rate of NADPH oxidation at340 nm. The assay mixture contained: 0.5mmol/LNADPH, 10mmol/L oxidized glutathione (GSSG),3mmol/L MgCl2 in 0.1mol/L phosphate buffer pH7.8, and 50–100 mL of enzyme extract. GR activitywas expressed as nmol NADPH/min/mg protein.
Superoxide dismutase (SOD, EC. 1.15.11) activitywas determined according to Giannopolitis and Ries(1977), by measuring its ability to inhibit thephotochemical reduction of nitroblue tetrazolium(NBT), as described in Pukacka and Pukacki (2000).
Catalase (CAT, EC 1.11.1.6) activity was esti-mated according to Chance and Maehly (1955). Thereaction mixture contained 1mL of 0.1mol/Lphosphate buffer pH 7.0, 1mL of 30mmol/L H2O2,and 50–100 mL enzyme extract. Catalase activitywas estimated by decrease in absorbance at 240 nmand was expressed as mmol H2O2/min/mg protein.
Guaiacol peroxidase (POX, EC 1.11.1.7) activitywas measured in the reaction of oxidation ofguaiacol at 470 nm (e ¼ 26.6mM/cm) according toChance and Maehly (1955). The reaction mixturecontained 1mL of 0.1mol/L phosphate buffer pH7.0, 1mL of 1% guaiacol, 1mL of 0.2mol/L H2O2
and 50–100 mL of enzyme extract.Protein content of crude enzyme extracts was
estimated according to Bradford (1976), usingbovine serum albumin (BSA) as a standard.
Statistical analysis
Data are presented as means7standard devia-tion of six replicates (two parallel treatmentsof three replicates). The statistical differencesbetween seeds stored at different temperature andhumidity conditions and the control (non-treated)were tested using an analysis of variance (ANOVA).Levels of significance are indicated as �Po0:05 and��Po0:01:
Results
Seed viability and ROS production
Beech seed viability was assessed on the basis ofthe ability of the seeds to germinate. Fig. 1 showsthat this clearly depended on storage conditions.Seed germinability decreased during storage, with
a greater fall as storage temperature increased.As soon as post-3 weeks of storage at 30 1C, viabilitydecreased to 25�30%, but after 9 weeks of storage,seeds nearly completely lost the ability to germi-nate. Initially, a small decrease in viability wasobserved at 4 1C; however, later, the decreaseintensified to 46–58%. At each temperature, viabilitydecreased more strongly at the higher humidity.
Production of the superoxide radical (O2�d)
clearly increased with time (Fig. 2). The increasewas earlier and stronger at higher temperature andhumidity levels. At the same time, we alsoobserved a very intensive generation of hydrogenperoxide (H2O2) (Fig. 3), which was also clearlycoincident with temperature and humidity levels. Astrong correlation was found between seed viabilityand ROS production (see Table 1).
Status of low-molecular antioxidants
Among low-molecular antioxidants, we assayedascorbic acid (ASA) and glutathione as compoundsdirectly involved in the Halliwell–Asada pathway,which plays an important role in the removal of ROSfrom cells (Asada, 1992). Changes in ASA, DHASAand the total pool of ascorbate content duringbeech seed storage under the above conditions arepresented in Fig. 4A–D. In the embryo axes, after aslight decrease in the first 3 weeks of incubation(from 1.08mg/g DW in the control to 0.57–0.83mg/g DW in incubated seeds), ASA level returned to theinitial level and even slightly exceeded it in week 6,reaching significantly higher values in week 9 (1.55,2.4mg/g DW and 2.4, 2.66mg/g DW, respectively,for 45% and 75% RH), especially at 20 and 30 1C

ARTICLE IN PRESS
Figure 2. Production of superoxide radical in beech (Fagus sylvatica) seeds during storage of 9 weeks at 4, 20, 30 1C and45%, 75% RH.
Figure 3. Production of hydrogen peroxide in beech (F. sylvatica) seeds during storage of 9 weeks at 4, 20, 30 1C and45%, 75% RH.
Reactive oxygen species and seeds variability 877
(Fig. 4A and C). In cotyledons, a marked increase inASA content was observed after week 6 at highertemperatures, and after week 9 at 4 1C (Fig. 4B andD), from 0.21mg/g DW in the control to 0.41, 0.42and 0.31mg/g DW for respectively 4, 20, and 30 1Cand 45% RH and to 0.36, 0.34, and 0.38mg/g DW,respectively, for 75% RH. The DHASA level wasgenerally higher in embryo axes than in cotyledonscompared to respective controls. It is also note-worthy that the level of the reduced form clearlydominated over the oxidized form of this compoundduring storage of seeds.
Glutathione occurred in beech seeds in thereduced (GSH) or oxidized (GSSG) form. In contrastto other plant organs, in seeds GSSG dominatedover GSH. On the basis of dry weight, much moreglutathione was present in embryo axes than in
cotyledons. The glutathione content during seedincubation in the studied conditions is presented inFig. 5A–D. During seed storage at 30 1C, weobserved a stronger decrease in both GSH andGSSG forms than in 4 and 20 1C. In embryo axes asignificant increase of both forms, from 130 and375 mg/g DW in control to 249 and 696 mg/g DW,respectively, at 20 1C and 45% RH, after 3 weeks ofincubation, was recorded. This seems to reflect adefensive reaction. The reaction observed incotyledons was not so pronounced.
Reaction of enzymatic antioxidants toconditions of seed storage
The activity of enzymes taking part in theremoval of ROS from cells was studied after an

ARTICLE IN PRESS
S. Pukacka, E. Ratajczak878
initial slow rehydration of seeds at 3 1C for 24 h.Among enzymes of the Halliwell–Asada pathway,we assayed the activity of APO, DHAR, and GR. Wealso assayed the activity of other enzymaticscavengers, like POX, SOD, and CAT. The activityof all enzymes was measured in week 1 (i.e. after aweek of incubation), and subsequently in weeks 3and 6, but only for 75% RH. This was because weassumed that at the higher humidity level, thedefensive reactions of enzymatic scavengers wouldbe more conspicuous. Enzymes of the Halliwell–A-
Figure 4. Changes in ascorbate (ASA) and dehydroascorbate (seeds stored at 45% (A,B) and 75% (C,D) RH, and at 4, 20 and
Table 1. The correlation (r2) between germinationcapacity and ROS production (O2
d� and H2O2) in beech(Fagus sylvatica) seeds under various storage conditions
Seed storageconditionsTemp/RH (1C/%)
Germinationcapacity andO2d�
Germinationcapacity andH2O2
4/45 0.9700 0.9087p-0.0000 p-0.0000
20/45 0.9286 0.9277p-0.0000 p-0.0000
30/45 0.9054 0.8758p-0.0001 p-0.0002
4/75 0.9670 0.9159p-0.0000 p-0.0000
20/75 0.8786 0.9083p-0.0002 p-0.0000
30/75 0.9302 0.8733p-0.0000 p-0.0002
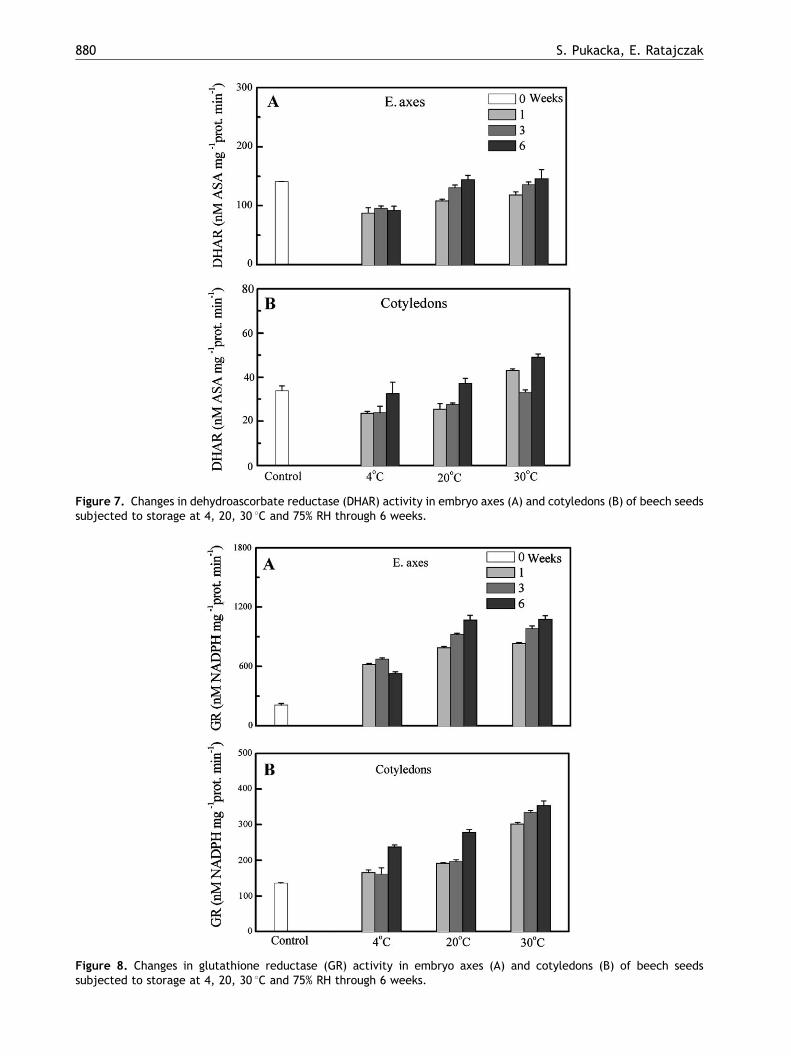
sada pathway (Figs. 6–8) are clearly involved inscavenging of the excess of ROS from beech seedsafter their rehydration. This is reflected in theirincreased activity in relation to the control as soonas after only 1 week of incubation (APO, GR), and afurther increase was observed after the next fewweeks of storage. The activity of CAT was high,whereas that of SOD was relatively low. SOD alsoweakly changed its activity during prolongedstorage of seeds (Fig. 9). Moreover, it is noteworthythat APO, DHAR, GR and CAT (Figs. 6–8 and 10)showed a similar run of changes in activityin embryo axes as well as in cotyledons. Theiractivity increased with increasing temperature andtime of seed incubation. POX activity (Fig. 11) incotyledons, significantly increased after one weekof storage, but clearly decreased in the nextweeks.
In general, the activity of all enzymes exceptPOX was higher in embryo axes than in cotyledons.
Discussion
This paper presents the results of research onROS production and effects of low molecular andenzymatic antioxidants on the germinability ofbeech seeds stored at different temperatures andhumidity levels. Our previous research (Pukacka etal., 2003) showed that beech seeds stored at 45%
DHASA) contents in embryo axes and cotyledons of beech30 1C through 9 weeks.

ARTICLE IN PRESS
Reactive oxygen species and seeds variability 879
and 75% RH differ substantially in moisture contentof embryo axes. In this study, the water content ofembryo axes was 7.8–8.2% for 45% RH and12.4–12.8% for 75% RH, while in cotyledons it
Figure 5. Changes in reduced (GSH) and oxidized (GSSG) formseeds stored at 45% (A,B) and 75% (C,D) RH, and at 4, 20 and
Figure 6. Changes in ascorbate peroxidase (APO) activitysubjected to storage at 4, 20, 30 1C and 75% RH through 6 w
reached 5.8% and 7.8%, respectively. This suggeststhat embryo axes are much more sensitive tostorage conditions than are cotyledons (Walters,1998). This is confirmed by the fast loss of seed
s of glutathione in embryo axes and cotyledons of beech30 1C through 9 weeks.
in embryo axes (A) and cotyledons (B) of beech seedseeks.
ARTICLE IN PRESS
Figure 7. Changes in dehydroascorbate reductase (DHAR) activity in embryo axes (A) and cotyledons (B) of beech seedssubjected to storage at 4, 20, 30 1C and 75% RH through 6 weeks.
Figure 8. Changes in glutathione reductase (GR) activity in embryo axes (A) and cotyledons (B) of beech seedssubjected to storage at 4, 20, 30 1C and 75% RH through 6 weeks.
S. Pukacka, E. Ratajczak880

ARTICLE IN PRESS
Figure 9. Changes in superoxide dismutase (SOD) activity in embryo axes (A) and cotyledons (B) of beech seedssubjected to storage at 4, 20, 30 1C and 75% RH through 6 weeks.
Reactive oxygen species and seeds variability 881
germinability, proportional to temperature andhumidity level (Fig. 1).
The intensity of ROS (superoxide anion radicaland hydrogen peroxide) production proved todepend on storage conditions (Figs. 2 and 3), andwas strongly correlated with seed germinability(Table 1). Correlation coefficients suggest that thestudied ROS can cause a decline in seed viability,not only by lipid peroxidation (after 3 weeks ofincubation changes in membrane permeability andcomposition were insignificant; (Ratajczak andPukacka, 2005), but also by affecting enzymaticsystems and DNA. For example, the toxic effects ofH2O2 consist in generation of the most active OHd
radical through the Haber–Weiss reaction. Itattacks all macromolecules, causing degradationof cell components and leading to cell death(Scandalios, 1993). Our results showed an increasedROS production after 3 weeks of seed storage at30 1C and after 6 weeks in all variants of theexperiment, i.e. also at 4 1C (Figs. 2 and 3), ascompared to the control.
Plant cells may contain many antioxidants, i.e.compounds scavenging ROS. Among them, ascor-bate and glutathione play an important role. Ingeneral, dry seeds of the orthodox category lack
ASA. This was confirmed, for example, in Vicia faba(Arrigoni et al., 1992), Heliantus annuus (De Paulaet al., 1996), Pinus pinea, and Avena sativa(Tommasi et al., 1999). Our research showed thatASA is present in dry beech seeds, and this seems tosupport their intermediate character. ASA concen-tration is c. 5 times higher in embryo axes than incotyledons. ASA is a very important antioxidant,reacting directly with OHd, O2
�d, and atomic oxygen(Noctor and Foyer, 1989). It cooperates with APO inscavenging H2O2 (Nakano and Asada, 1987). Itparticipates in the reduction of a-tocopherol – animportant antioxidant associated directly withmembranes (Padh, 1990). The removal of H2O2
from cells takes place in the Halliwell–Asadapathway, which involves ASA, glutathione, andenzymes, such as APO DHAR, GR, and MDHAR(monodehydroascorbate reductase, not analysedin this study). In the storage conditions tested inthis study, the increase in ASA content of embryoaxes and cotyledons during storage may result fromits synthesis de novo, as a result of enhancedrespiration in those parts of seeds (Siendones et al.,1999; Millar et al., 2003), or from a low activity ofenzymatic decomposition. Nevertheless, ASA accu-mulation in seeds, beside its positive, antioxidative

ARTICLE IN PRESS
Figure 10. Changes in catalase (CAT) activity in embryo axes (A) and cotyledons (B) of beech seeds subjected tostorage at 4, 20, 30 1C and 75% RH through 6 weeks.
S. Pukacka, E. Ratajczak882
function, may have a negative effect, becauseunder conditions of water deficit, ASA is more likelyto react with iron ions (Fenton’s reaction), releas-ing OHd radicals (De Tulio and Arrigoni, 2003).
The role of glutathione as an antioxidant exists indirect reactions with free radicals, or in coopera-tion with ASA in the Halliwell–Asada pathway. It ischaracteristic of beech seeds to contain much moreGSSG (oxidized form of glutathione) thanGSH (reduced form). Such a strong domination ofGSSG has not been observed in seeds of otherspecies, such as H. annuus (De Paula et al., 1996)and Citrullus vulgaris (Hsu and Sung, 1997).Such a shift in redox status may decreasethe concentration of –SH groups in cells (Alscher,1989; Kranner and Grill, 1993). The ratio ofthiol groups to disulphide groups plays an importantrole in stabilization of enzymatic proteins indry seeds. At 30 1C in beech seeds this ratio iseven more favourable for disulphide groups. A fastdecrease in GSH at 30 1C may be associatedwith ROS accumulation. At 20 1C, we recorded amarked increase in GSH level in embryo axes after3 weeks of storage, but it was followed by a slowdecline. This most likely suggests to a defensivereaction and involvement of enzymes generatingGSH.
Our results suggest that the Halliwell–Asadapathway and other enzymes are involved in thedefensive reaction against ROS after seed imbibi-tion. This system was active although the seeds lostthe ability to germinate. This indicates that thechanges that took place in cells under the influenceof ROS are irreversible, and lead to a loss ofgerminability and seed death. The role of low-molecular and enzymatic antioxidants in preserva-tion of seed viability has been investigated mainlyduring accelerated aging of orthodox seeds ofspecies like H. annuus (Bailly et al., 1996; De Paulaet al., 1996), C. vulgaris (Hsu and Sung, 1997),Vigna radiata (Murthy et al., 2002) and Gossypiumhirsutum (Goel et al., 2003). In all the studies, ahigh temperature (33–47 1C) and humidity level(80–100% RH) caused a fast decline in seed viability(within 8–11 days), and a decrease in the activity ofenzymatic scavengers. Our results showed that inbeech seeds stored even at relatively low tempera-ture (4 1C), excessive production of ROS appearsafter 6 weeks of incubation, which consequentlyleads to the 40% decrease of their viability(Figs. 1–3). The important characteristic of beechseeds is the substantial difference in sorptionproperties of embryo axes (Pukacka et al., 2003),which may result in a faster loss of viability at a

ARTICLE IN PRESS
Figure 11. Changes in peroxidase (POX) activity in embryo axes (A) and cotyledons (B) of beech seeds subjected tostorage at 4, 20, 30 1C and 75% RH through 6 weeks.
Reactive oxygen species and seeds variability 883
higher humidity, as compared with seeds of otherspecies. Embryo axes are protected against thedestructive effects of ROS to some extent, as theyare characterized by a higher concentration of low-molecular antioxidants and a higher activity ofenzymatic scavengers than cotyledons. This protec-tion is insufficient even at 4 1C. In summary, weconclude that the loss of germinability of beechseeds during storage at temperatures above 0 1Cwas due to a strong oxidative stress and aninefficient system of scavengers of free radicals.
Acknowledgements
This research was supported by the StateCommittee for Scientific Research (Poland) – GrantNo. 5PO6H06119.
References
Alscher RG. Biosynthesis and antioxidant function ofglutathione in plants. Physiol Plant 1989;77:457–64.
Arrigoni O, De Gara L, Tommasi F, Liso R. Changes inascorbate system during seed development of Viciafaba L. Plant Physiol 1992;99:235–8.
Asada K. Ascorbate peroxidase – a hydrogen peroxidescavenging enzyme in plants. Physiol Plant 1992;85:235–41.
Bailly C, Benamar A, Corbineau F, Come D. Changes inmalondialdehyde content and in superoxide dismu-tase, catalase and glutathione reducrase activities insunflower seeds as related to deterioration duringaccelerated aging. Physiol Plant 1996;97:104–10.
Bailly C, Leymarie J, Lehner J, Rousseau S, Come D,Corbineau F. Catalase activity in developing sunflowerseeds as related to drying. J Exp Bot 2004;55:475–83.
Bonner FT. Storage of seeds: potential and limitation forgermplasm conservation. Forest Ecol Manage 1990;35:35–43.
Bradford MM. A rapid and sensitive method for thequantitation of microgram quantities of proteinutilizinh the principle of protein-dye binding. AnalBiochem 1976;72:248–54.
Buchvarov P, Gantcheff TS. Influence of accelerated andnatural aging on free radical levels in soybean seeds.Physiol Plant 1984;60:53–6.
Chaitanya KSK, Naithani SC. Role of superoxide, lipidperoxidation and superoxide dismutase in membraneperturbation during loss of viability in seeds of Shorearobusta Gaertn.f. New Phytol 1994;126:623–7.

ARTICLE IN PRESS
S. Pukacka, E. Ratajczak884
Chance B, Maehly AC. Assay of catalases and peroxidases.Collowick SP, Kapplan NO, editors. Methods inenzymology, vol. 2. New York: Academic Press; 1955.p. 764–75.
De Paula M, Perez-Otaola M, Darder M, Torres M, FrutosG, Martınez-Honduvilla CJ. Function of the ascorba-te–glutathione cycle in aged sunflower seeds. PhysiolPlanat 1996;96:543–50.
De Tulio MC, Arrigoni O. The ascorbic acid system inseeds: to protect and to serve. Seed Sci Res2003;13:249–60.
Doke N. Generation of superoxide anion by potato tuberprotoplasts during the hyper-sensitive response tohyphal wall components of Phytophthora infestansand specific inhibition of the reaction by suppressorsof hypersensitivity. Physiol Plant Pathol 1983;23:359–67.
Esterbauer H, Grill D. Seasonal variation of glutathioneand glutathione reductase in needles of Picea abies.Plant Physiol 1978;61:119–21.
Giannopolitis CN, Ries SK. Superoxide dismutase. I.Occurrence in higher plants. Plant Physiol 1977;59:309–14.
Girard J, Le Meste M. Absence the relation entre taux deradicau libre mesure par RPE viabilite des semencesde ble. Com Ren Acad Sci Paris 1992;Serie III:417–22.
Goel A, Goel AK, Sheoran IS. Changes in oxidative stressenzymes during artificial aging in cotton (Gossypiumhirsutum L.) seeds. J Plant Physiol 2003;160:1093–100.
Gosling PG. Beechnuts storage: a review and practicalinterpretation of the scientific literature. Forestry1991;64:51–9.
Helmerhorst E, Stokes GB. Microcentrifuge desalting: arapid, quantitative method for desalting smallamounts of protein. Anal Biochem 1980;104:130–5.
Hendry GA. Oxygen free radical processes and seedlongevity. Seed Sci Res 1993;3:141–53.
Hendry GAF, Finch-Savage WE, Thorpe PC, Atherton NM,Buckland SM, Nilsson KA, Seel WE. Free radicalprocesses and loss of seed viability during desiccationin the recalcitrant species Quercus robur L. NewPhytol 1992;122:273–9.
Hepburn HA, Goodman BD, McPhail DB, Matthews S,Powell AA. An evaluation of EPR measurements of theorganic free radical content of individual seeds in thenon-destructive testing of seed viability. J Exp Bot1986;37:1675–84.
Hsu JL, Sung JM. Antioxidant role of glutathioneassociated with accelerated aging and hydration oftriploid watermelon seeds. Physiol Plant 1997;100:967–74.
ISTA. International rules for seed testing. Rules 1999.Seed Sci Technol 1999;27(Suppl.):1–133.
Kampfenkel K, Van Montagu M, Inze D. Extraction anddetermination of ascorbate and dehydroascorbatefrom plant tissue. Anal Biochem 1995;225:165–7.
Khan MM, Hendry GAF, Atherton NM, Vertucci-WaltersCW. Free radical accumulation and lipid peroxidationin testas of rapidly aged soybean seeds: a light-promoted processes. Seed Sci Res 1996;6:101–7.
Kranner I, Grill D. Content of low-molecular-thiols duringthe imbibition of pea seeds. Physiol Plant 1993;88:557–62.
Leon-Lobos P, Ellis RH. Seed storage behaviour of Fagussylvatica and Fagus crenata. Seed Sci Res 2002;12:31–7.
Mc Donald MB. Seed deterioration: physiology, repair andassessment. Seed Sci Technol 1999;27:177–237.
Millar AH, Mittova V, Kiddle G, Heazelwood JL, BartoliCG, Theodoulou FL, Foyer CH. Control of ascorbatesynthesis by respiration and its implications for stressresponses. Plant Physiol 2003;133:443–7.
Murthy NUM, Liang Y, Kumar PP, Sun WQ. Non-enzymaticprotein modification by the Maillard reaction reducesthe activity scavenging enzymes in Vigna radiata.Physiol Plant 2002;115:213–20.
Nakano Y, Asada K. Hydrogen peroxide is scavenged byascorbate peroxidase in spinach chloroplasts. PlantCell Physiol 1981;22:867–80.
Nakano Y, Asada K. Purification of ascorbate peroxidasein spinach chloroplasts: its inactivation in ascorbate-depleted medium and reactivation by monodehy-droascorbate radical. Plant Cell Physiol 1987;28:131–40.
Noctor G, Foyer CH. Ascorbate and glutathione: keepingactive oxygen under control. Ann Rev Plant PhysiolPlant Mol Biol 1989;49:249–79.
Padh H. Cellular function of ascorbic acid. Biochem CellBiol 1990;68:1166–73.
Poulsen KM. Predicting the storage life of beech nuts.Seed Sci Technol 1993;21:327–37.
Priestley DA. Seed aging. Ithaca, NY, London: ComstockPublishing Associates; 1986.
Priestley DA, McBride MB, Leopold AC. Tocopherol andorganic free radical levels in soybean seeds duringnatural and accelerated aging. Plant Physiol 1980;66:715–9.
Pukacka S. Changes in membrane lipid componentsand antioxidant levels during natural aging ofseeds of Acer platanoides. Physiol Plant 1991;82:306–10.
Pukacka S, Pukacki PM. Seasonal changes in antioxidantlevel of Scots pine (Pinus sylvestris L.) needlesexposed to industrial pollution. II. Enzymatic scaven-gers activities. Acta Physiol Plant 2000;22:457–64.
Pukacka S, Wojkiewicz E. The effect of temperature ofdrying on viability and some factors affecting stor-ability of Fagus sylvatica seeds. Acta Physiol Plant2003;25:163–9.
Pukacka S, Hoffmann SK, Goslar J, Pukacki PM, Wojkie-wicz E. Water and lipid reactions in beech (Fagussylvatica L.) seeds and its effect on storage behaviour.Bioch Biophys Acta 2003;1621:48–56.
Ratajczak E, Pukacka S. Decrease in beech (Fagussylvatica) seed viability caused by temperature andhumidity conditions as related to membrane damageand lipid composition. Acta Physiol Plant 2005;27:3–12.
Roberts EH. Predicting the storage life of seeds. Seed SciTechnol 1973;1:499–514.

ARTICLE IN PRESS
Reactive oxygen species and seeds variability 885
Sagisaka S. The occurrence of peroxide in aperennial plant Populus gelrica. Plant Physiol 1976;57:309.
Scandalios JG. Oxygen stress and superoxide dismutase.Plant Physiol 1993;101:7–12.
Scande M, Buitink J, Hoekstra FA. A study of waterrelations in neem (Azadirachta indica) seed that ischaracterized by complex storage behaviour. J Exp Bot2000;51:635–43.
Senaratna T, Gusse JF, Mc Kersie BD. Age-induced changesin cellular membranes of imbibed soybean axes.Physiol Plant 1988;73:85–91.
Siendones E, Gonzales-Reyes JA, Santos-Ocana Navas P,Cordoba F. Biosynthesis of ascorbic acid in kidneybean. L-galactono-gamma-lactone dehydrogenase is
an intrinsic protein located at the mitochondrial innermembrane. Plant Physiol 1999;120:907–12.
Smith I. Stimulation of glutathione synthesis in photo-respiring plants by catalase inhibitors. Plant Physiol1985;79:1044–7.
Suszka B, Muller C, Bonnet-Masimbert M. Seeds of forestbroadleaves: from harvest to sowing. Paris: INRA;1996.
Tommasi F, Paciolla C, Arrigoni O. The ascorbate systemin recalcitrant and orthodox seeds. Physiol Plant1999;105:193–8.
Vertucci CW, Ross EE. Theoretical basis of protocols forseed storage. Plant Physiol 1990;94:1019–23.
Walters C. Understanding the mechanisms and kinetics ofseed aging. Seed Sci Res 1998;8:223–44.




















