
Precipitation variability does not affect soil respiration and nitrogen dynamics in the understorey...
17
REGULAR ARTICLE Precipitation variability does not affect soil respiration and nitrogen dynamics in the understorey of a Mediterranean oak woodland Marjan Jongen & Xavier Lecomte & Stephan Unger & David Fangueiro & João Santos Pereira Received: 24 January 2013 / Accepted: 11 April 2013 # Springer Science+Business Media Dordrecht 2013 Abstract Background and aims Future climate scenarios for the Mediterranean imply increasing precipitation variability. This study presents a large-scale water manipulation experiment simulating changes in precipitation variabil- ity, aiming at a better understanding of the effects of rainfall patterns on soil C and N cycling and understorey productivity in a Mediterranean oak woodland. Methods We used rain-out shelters to achieve (1) a normal dry period (7 days), and (2) a dry period increased three-fold (21 days), without altering total annual precipitation inputs. Results The temporal patterns of soil respiration (R s ) and soil inorganic N were not affected by treatment. However, water infiltration and N leaching increased with large infrequent watering events. R s and soil NH 4 + -N correlated with soil temperature, with soil NO 3 - -N being influenced by leaching. Conclusions The lack of significant treatment effects on either R s or soil inorganic N can be explained by (1) minor differences in plant productivity between the treatments, suggesting equal plant N demand, and (2) the absence of moisture dependence of R s and soil NH 4 + -N. Increased N leaching with large infrequent precipitation events may have longer-term conse- quences for ecosystem functioning. Our results con- tribute to an improved understanding of possible climate change effects on key ecosystem processes in Mediterranean ecosystems. Keywords Soil CO 2 flux . Soil inorganic N . Understorey vegetation . Mediterranean ecosystem . Precipitation manipulation . Climate change Introduction Anthropogenic activities continue to change atmospheric greenhouse gas concentrations, thereby exerting strong impacts on climate and biogeochemical cycles (Raupach and Canadell 2010). Future climate scenarios for the Iberian Peninsula predict increasing temperatures and an overall decrease in precipitation (IPCC 2007). In addition, scenarios predict an increased temporal vari- ability of precipitation regimes (Luterbacher et al. 2006), with a lower frequency of rainfall days and longer intervening dry periods (Easterling et al. 2000). The increase in precipitation variability is expected to extend the periods of soil moisture deficit (Jackson et al. 2001), which might have important consequences for productivity, biodiversity and the matter cycles of many Plant Soil DOI 10.1007/s11104-013-1728-7 Responsible Editor: Stefano Manzoni. M. Jongen (*) : X. Lecomte : D. Fangueiro : J. S. Pereira Instituto Superior de Agronomia, Universidade Técnica de Lisboa, Tapada da Ajuda, 1349-017 Lisbon, Portugal e-mail: [email protected] S. Unger Department of Experimental and Systems Ecology, University of Bielefeld, Universitätsstr. 25, 33615 Bielefeld, Germany
Transcript of Precipitation variability does not affect soil respiration and nitrogen dynamics in the understorey...

REGULAR ARTICLE
Precipitation variability does not affect soil respirationand nitrogen dynamics in the understoreyof a Mediterranean oak woodland
Marjan Jongen & Xavier Lecomte & Stephan Unger &
David Fangueiro & João Santos Pereira
Received: 24 January 2013 /Accepted: 11 April 2013# Springer Science+Business Media Dordrecht 2013
AbstractBackground and aims Future climate scenarios for theMediterranean imply increasing precipitation variability.This study presents a large-scale water manipulationexperiment simulating changes in precipitation variabil-ity, aiming at a better understanding of the effects ofrainfall patterns on soil C and N cycling and understoreyproductivity in a Mediterranean oak woodland.Methods We used rain-out shelters to achieve (1) anormal dry period (7 days), and (2) a dry periodincreased three-fold (21 days), without altering totalannual precipitation inputs.Results The temporal patterns of soil respiration (Rs)and soil inorganic N were not affected by treatment.However, water infiltration and N leaching increasedwith large infrequent watering events. Rs and soilNH4
+-N correlated with soil temperature, with soilNO3
−-N being influenced by leaching.Conclusions The lack of significant treatment effectson either Rs or soil inorganic N can be explained by (1)
minor differences in plant productivity between thetreatments, suggesting equal plant N demand, and (2)the absence of moisture dependence of Rs and soilNH4
+-N. Increased N leaching with large infrequentprecipitation events may have longer-term conse-quences for ecosystem functioning. Our results con-tribute to an improved understanding of possibleclimate change effects on key ecosystem processes inMediterranean ecosystems.
Keywords Soil CO2 flux . Soil inorganic N .
Understorey vegetation .Mediterranean ecosystem .
Precipitationmanipulation . Climate change
Introduction
Anthropogenic activities continue to change atmosphericgreenhouse gas concentrations, thereby exerting strongimpacts on climate and biogeochemical cycles(Raupach and Canadell 2010). Future climate scenariosfor the Iberian Peninsula predict increasing temperaturesand an overall decrease in precipitation (IPCC 2007). Inaddition, scenarios predict an increased temporal vari-ability of precipitation regimes (Luterbacher et al.2006), with a lower frequency of rainfall days andlonger intervening dry periods (Easterling et al. 2000).The increase in precipitation variability is expected toextend the periods of soil moisture deficit (Jackson et al.2001), which might have important consequences forproductivity, biodiversity and the matter cycles of many
Plant SoilDOI 10.1007/s11104-013-1728-7
Responsible Editor: Stefano Manzoni.
M. Jongen (*) :X. Lecomte :D. Fangueiro : J. S. PereiraInstituto Superior de Agronomia,Universidade Técnica de Lisboa, Tapada da Ajuda,1349-017 Lisbon, Portugale-mail: [email protected]
S. UngerDepartment of Experimental and Systems Ecology,University of Bielefeld, Universitätsstr. 25,33615 Bielefeld, Germany

terrestrial ecosystems (Austin et al. 2004; Huxman et al.2004; Schwinning and Sala 2004), and particularly fordrought-susceptive grasslands (Knapp et al. 2002; Fayet al. 2003; Harper et al. 2005).
In the Iberian Peninsula, evergreen oak woodlands—montados in Portugal and dehesas in Spain—are ofgreat ecological and socio-economic importance, keyto biodiversity conservation and ecosystem services(Bugalho et al. 2011). These savanna-type agro-silvo-pastoral ecosystems are characterized by evergreenoaks, a shrub layer of varying diversity, and an herba-ceous understorey. The diverse community of C3 annualspecies that constitutes the understorey plays a signifi-cant role for ecosystem productivity (Pereira et al. 2007;Aires et al. 2008; Unger et al. 2009; Jongen et al. 2011),by exerting a large influence on soil nutrient cycling andthe ecosystem water balance (Moreno Marcos et al.2007; Otieno et al. 2011).
Due to the semi-arid nature of these Mediterraneanecosystems, water availability strongly influences eco-system productivity and functioning (Pereira et al.2007; Bugalho et al. 2011), including the biogeochem-ical cycles of carbon (C) and nitrogen (N). Moreover,in these semi-arid systems even discrete precipitationpulses can trigger crucial ecosystem processes, such assoil respiration (Rs) and nitrogen mineralization(Huxman et al. 2004). Rs is one of the main determi-nants of the carbon balance of terrestrial ecosystems(Valentini et al. 2000) and is composed of a multitudeof sources, which can be of autotrophic (Ra), hetero-trophic (Rh), or inorganic origin (Kuzyakov andGavrichkova 2010). For semi-arid Mediterranean eco-systems, where Rs can constitute 64-90 % of ecosys-tem C losses (Unger et al. 2012), soil water availabilityhas been shown to be the main determinant of Rs on anannual basis (Correia et al. 2012), with a strong de-pression of Rs during drought periods (Reichstein et al.2003; Unger et al. 2010a) and rapidly increasing soilCO2 efflux in response to rewetting of previously drysoil (Unger et al. 2010b). However, several otherstudies have shown that in these ecosystems, soilmoisture and soil temperature controls on Rs alternate(Curiel Yuste et al. 2007; Almagro et al. 2009; Rey etal. 2011), with soil moisture being the predominantdriver throughout the dry season, while soil tempera-ture controls Rs throughout the rest of the year.
Similar to Rs, mineralization of soil N by soil mi-crobes is often determined by water availability(Borken and Matzner 2009). Generally, precipitation
stimulates N mineralization from accumulated organicmatter (Austin et al. 2004) and facilitates N flow toplant roots (James and Richards 2006). However,plant N uptake, microbial immobilization of N, anddecreasing microbial activity and N flow with decreas-ing soil moisture limit the duration of increased Navailability after a precipitation pulse (Cui andCaldwell 1997; Hodge et al. 1999; Dijkstra et al.2012). Additionally, N losses by leaching (Yahdjianand Sala 2010), increased denitrification rates (e.g.Tiemann and Billings 2012) and increased fluxes ofN2O, NO and NH3 (Kim et al. 2012) after precipitationpulses can diminish the positive effects of the soilmoisture increases on N availability. Thus, in contrastto soil C cycling, which is generally stimulated byincreased soil water content, the effects of precipita-tion pulses on soil N availability and cycling dependon the balance of mineralization and N losses, poten-tially having a direct effect on productivity in nutrientpoor systems.
Developing a better understanding of the effects ofaltered precipitation regimes on soil C and N cyclingis crucial, as the predicted increase in precipitationvariability will affect both precipitation pulse fre-quency and magnitude. Although several studiesaddressed the effect of precipitation manipulationon Rs (Li et al. 2005; Zhou et al. 2006; McCulleyet al. 2007; Chou et al. 2008; Talmon et al. 2011;Schindlbacher et al. 2012; Selsted et al. 2012; Suseelaet al. 2012) or N mineralization (Yahdjian et al. 2006;Patrick et al. 2009; Yahdjian and Sala 2010), only afew studies have reported on the effects of alterationsin the precipitation regime on these processes, with-out changing total annual precipitation inputs. Forexample, in a C4 grassland, alterations in the timingof growing season precipitation, with less frequentbut larger rain events, resulted in a decrease in Rs
(Harper et al. 2005; Fay et al. 2011). However, in anarid grassland, an increase in precipitation variabilityresulted in increasing Rs rates (Thomey et al. 2011;Vargas et al. 2012). In relation to mineralization,increasing precipitation variability, with a shift tomore extreme rainfall patterns, increased soil N avail-ability in a C4 grassland and a semi-arid steppe, buthad no influence on soil N in a mixed grass prairie(Heisler-White et al. 2009).
For the herbaceous understorey in Mediterraneanevergreen oak woodlands, to date no studies have beenpublished on the effect of precipitation variability on
Plant Soil

Rs and N mineralization and availability. To close thisknowledge gap and to gain better insights into thelinkage of C and N cycles and their relation with plantproductivity under altered precipitation regimes, westudied Rs and N dynamics during the growing seasonof the understorey vegetation in a Mediterranean oakwoodland under experimental rainfall manipulation.We hypothesized that, with no net changes in rainfallquantity, large infrequent precipitation events, withlonger intervening dry periods, will decrease both (1)Rs and (2) N availability, as compared to small fre-quent precipitation events, due to water deficit duringprolonged inter-pulse periods. Further, water deficitduring the inter-pulse periods will cause (3) decreasingplant productivity, which will feed back negatively onsoil N and Rs. The presented results contribute to abetter understanding of climate change mediated ef-fects on the soil C and N cycles in Mediterraneanecosystems, which is crucial for developing adaptationand mitigation strategies to ensure future sustainabilityand productivity.
Material and methods
Site description
The study was conducted at the Herdade daMachoqueirado Grou (39º08′16″N, 8º20′03″W), 30 km northeast ofCoruche, Portugal. The soil is a Cambisol (FAO), with81% sand, 14 % silt and 5 % clay. Volumetric soil watercontent (SWC) at field capacity and at permanentwilting point are 19.3 % and 7.6 %, respectively. Theclimate is Mediterranean, characterized by wet and mildwinters, and dry and hot summers. Long-term meanannual temperature is ~15.9 °C (Inst. de Meteorologia,Lisbon). However, the Mediterranean climate is alsocharacterized by a high variability in annual precipita-tion and a high frequency of drought periods (Xoplaki etal. 2004).
The study site is an evergreen oak woodland withQuercus suber being the only tree species. The treecover is about 50 %, with a tree density of 177 treesha−1. Average tree height is 7.9 m. The understoreyvegetation consists of a mixture of C3 annual species(e.g. Tuberaria guttata, Tolpis barbata, Agrostispourretii), emerging after the first rains in autumn andsenescing in late spring. Aboveground productivity ofthe understorey vegetation in May 2009 was 64±
7.7 g m−2. Until October 2009, the experimental sitewas intermittently grazed with a stocking density of0.16 cattle ha−1. In October 2010, the site wasploughed and seeded with a legume-rich mixture(Charneca 650, S 07874, Fertiprado, Vaiamonte,Portugal), a common agricultural practice inPortugal, to improve productivity and soil fertility(Crespo 2010).
Experimental design and rainfall manipulation
In December 2009, eight rainfall manipulation shelters(‘Fraga’, Prilux, Ponte de Vagos, Portugal) wereconstructed within a fenced area of ~3500 m2, enablingmanipulation of the precipitation received by theunderstorey. The distance between two shelters was aminimum of 8 m, and shelters were randomly assignedto one of the two treatments. Each shelter covered anarea of 6×5 m (30 m2), with an eave height of 1.6 m anda ridge height of 2.5 m. The shelter roofs were coveredin November 2010 by a clear, 2 mm, UV-transparentpolyethylene greenhouse film (Plásticos F. Matos,Massamá, Portugal). For additional information on shel-ter design see Jongen et al. (2013).
The water manipulation treatment was based onhistorical precipitation data (1955–2007) for the ex-perimental site (Inst. de Meteorologia, Lisbon): aver-age annual precipitation is 680±210 mm, with 87 %(594 mm) of the precipitation being confined to theunderstorey growing season (October 1 and May 31),and the median length of the dry periods is 7 days. Inaddition, historical precipitation data gives a mediannumber of rain events during the growing season of24, with the median size of these events being23.9 mm. The two water manipulation treatmentswere: ‘weekly watering treatment’, where natural con-ditions were simulated with a dry period of 7 days, and‘3-weekly watering treatment’, with the dry periodincreased three-fold to 21 days. Each of the two ma-nipulation treatments had four replicate experimentalplots. To avoid a treatment effect on germination andseedling establishment, all experimental plots weresubjected to natural precipitation, ensuring equal waterinputs, until the end of November, receiving 148 mm ofwater. From November 30 onwards, the weeklywatering treatment received one application of 16 mmof water every week and the 3-weekly watering treat-ment received one application of 48 mm every 3 weeks.In total, precipitation inputs during the growing season
Plant Soil

of October 2010 to the end of May 2011 amounted to590 mm for both treatments.
Microclimate
Soil temperature (Ts) at a depth of 5 cm was continu-ously measured in four of the experimental plots usingECT temperature sensors (Decagon Devices, Pullman,USA). Air temperature and humidity were continuouslymeasured using EHT sensors with radiation shields(Decagon Devices, Pullman, USA). Water vapour pres-sure deficit (VPD) was calculated from the temperatureand humidity data according to Goudriaan and vanLaar (1994), and presented as the average values forthe 11.00 h–18.00 h period. SWC at a depth of10 cm was measured continuously in each of theexperimental plots using EC-5 soil moisture sensors(Decagon Devices, Pullman, USA). All above-mentioned sensors were connected to EM-50 dataloggers (Decagon Devices, Pullman, USA), recordinghalf-hourly means. Data on SWC at a depth of 10,20, 30, 40, 60 and 100 cm were obtained in allexperimental plots using soil profile probes (PR1,Delta-T Devices, Cambridge, UK). Measurementswere made 1 h before and 24 h after watering events,only on dates when both weekly and 3-weekly wateringtreatments were irrigated. PR1 data were used to esti-mate infiltration, calculated as the increase in theamount of water (l m−2) in the top 35 cm soil layerwithin 24 h after watering as compared to pre-wateringvalues. Precipitation was measured with a RG2 raingauge (Delta-T Devices, Burwell, Cambridge, UK),and stored as half-hourly means on a DL2 data logger(Delta-T Devices, Burwell, Cambridge, UK).
Primary productivity
Germination of the herbaceous understorey in ourmontado ecosystem occurs in autumn after the firstsubstantial precipitation event, with biomass develop-ment primarily confined to the period betweenFebruary and May. Aboveground net primary produc-tivity (ANPP) was estimated on February 1, April 5,May 17 and June 6, the latter date corresponding withthe onset of sward senescence. All plant material intwo 40×40 cm quadrats within each experimental plotwas harvested, and subsequently divided into live andsenescent biomass, sorted into the individual species,oven dried at 60 °C for 72 h, and weighed. Belowground
net primary productivity (BNPP) was estimated at peakbiomass (May 17). In each of the experimental plots twosoil cores of 8 cm diameter and a depth of up to 20 cmwere taken. Roots were washed out, oven dried at 60 °Cfor 72 h, and weighed.
Soil CO2 fluxes
Short-term soil CO2 fluxes were measured using anSRS-2000 portable soil respiration system (ADCBioScientific, Hertfordshire, UK). PVC collars, witha height of 7 cm, were installed in the experimentalplots 1 week prior to the first measurements. At thetime of measurement, the soil chamber was fitted tothe collars, with subsequent measurements of soilrespiration (Rs) between 11.00 h and 13.00 h, at twosample points in each plots. Measurements were madeat 3-weekly intervals in all experimental plots, imme-diately prior to the watering events, and 1 day after thewatering events. In addition, measurements wereconducted in all experimental plots prior to some ofthe weekly watering events.
Soil N dynamics
Soil sampling, for determination of soil NO3−-N and
NH4+-N, was done in all experimental plots at 3-
weekly intervals, immediately prior to, and 1 day afterthe watering events. From each of the experimentalplots, four duplicate soil cores (2 cm diameter, 0–10 cm) were pooled, mixed to achieve uniformity,and sieved. NO3
−-N and NH4+-N were extracted by
shaking 8 g of soil with 20 ml 2 M KCl for 1 h.Extracts were analyzed by automated segmented-flow spectrophotometry (SanPlus, Skalar AnalyticalB.V., Breda, the Netherlands) using the Berthelot andhydrazinium reduction method, and the sulfanilamidediazotizing method for NH4
+-N and NO3−-N, respec-
tively (Houba et al. 1989). Plant available NO3−-N and
NH4+-N was measured with PST-1 ion-exchange resin
capsules (Unibest Inc., Bozeman, USA) filled withmixed anion-cation resins, providing an index of over-all inorganic N availability in the soil during a givenincubation period (Binkley and Hart 1989). Capsuleswere installed in October 2010, into the main rootingzone at 10 cm depth, with three replicates per exper-imental plot. Capsules were collected in June 2011,with subsequent stripping of adsorbed ions by sequen-tial shaking in two batches of 20 ml 2 M KCl. Extracts
Plant Soil

were analyzed for NH4+-N and NO3
−-N as describedabove.
Using pore water samplers (SK20 suction cups,UMS GmbH, München, Germany) in connection witha vacuum system (VS-pro, UMS GmbH, München,Germany), soil pore water at a depth of 30 cm wascollected in the period of March to May, by applying aconstant vacuum (between 25 kPa and 30 kPa,depending on SWC and plot elevation) for a 24 hperiod, starting at the time of the respective 3-weeklywatering events. A total of eight pore water samplerswere installed; two pore water samplers in half of theexperimental plots. Subsequently, the water collectedwas analyzed for NO3
−-N and NH4+-N as described
above. N leaching was estimated as the product ofwater infiltration to depths >35 cm and the concentra-tion of NO3
−-N in soil pore water collected at 30 cmdepth.
Data analysis
Statistical analyses was performed using Sigmaplot11.0 (Systac Software, Inc., San Jose, USA).Significance level was set to p<0.05. Data on seasonalvariation of Rs, soil inorganic N and pore water NO3
−-N failed to meet the normality assumptions for analy-sis of variance (ANOVA) despite transformations, andwere analyzed using non-parametric Mann–Whitneyrank sum tests for all-over treatment effects, andFriedman repeated measures ANOVA on ranks forall-over date effects. Data on Rs and soil inorganic Naround individual watering events, and data on waterinfiltration and ANPP were analyzed using a two-wayrepeated measures ANOVA, with treatment and dateas main effects. Data on BNPP were analyzed using aone-way ANOVA, with treatment being the only fac-tor. When a significant difference was found for amain effect, Fisher’s LSD post-hoc pairwise compar-ison was applied to determine individual differencesbetween means. Linear regression analysis was used torelate Rs with SWC.
The temperature dependency of Rs and soil inor-ganic NH4
+-N was calculated using the van’t Hoffempirical exponential equation (Lloyd and Taylor1994): y=aebTs, where y represents Rs in μmol m−2 s−1
or soil inorganic NH4+-N in μg g−1, Ts is the soil tem-
perature at 5 cm depth and a and b are model coeffi-cients. Subsequently, the temperature sensitivity of Rs,the Q10, was calculated as Q10=e
10b.
Results
Microclimate and soil water
Daily average Ts over the course of the study rangedfrom 5.0 °C to 22.4 °C (Fig. 1a). December andJanuary were characterized by several relatively warmspells, with average daily Ts in the period of December6 until January 21 exceeding 9 °C on 35 occasions. InApril and May, daily maximum Ts exceeded 20 °C on50 occasions, with VPD, presented as the averagevalues for the 11.00 h–18.00 h period, exceeding 1.5kPa for most of these ‘warm’ days.
The experimental precipitation pattern (Fig. 1b),without concurrent changes in total precipitation,resulted in significant differences in temporal soilmoisture dynamics between the two treatments,with peaks in SWC at the time of watering events(Fig. 1c). At the end of November, when the watermanipulation treatment started, SWC was 12.6 %.From the beginning of January until the end ofMarch, SWC did not go below 14.1 %. However,from April onwards, higher temperatures in com-bination with increasing biomass resulted in a rap-id decrease in SWC, with values as low as 10.3 %and 9.0 % being recorded in the weekly and 3-weekly watering treatments, respectively. AverageSWC at 10 cm depth for the duration of watermanipulation was 16.0±0.11 % and 14.9±0.12 %in the weekly and 3-weekly watering treatments,respectively.
Water infiltration in the top 35 cm (Fig. 2), assessed24 h after the watering events, was significantly af-fected by date (p<0.001). When watered, the 3-weekly watering treatment received three times theamount of water (48 l m−2), as compared to the weeklywatering treatment (16 l m−2). However, the differencein water infiltration in the top 35 cm between the twotreatments was relatively small. Up to the end ofMarch, infiltration was 11.4±0.45 l m−2 and 12.6±0.41 l m−2 in the weekly and 3-weekly watering treat-ments, respectively. In May and June water retentionincreased, due to lower SWC, with infiltration of13.6±0.94 l m−2 and 19.7±1.2 l m−2 in the weeklyand 3-weekly watering treatments, respectively(Fig. 2). In May and June, infiltration in the 3-weekly watering treatment was significantly higher,as compared to the weekly watering treatment.Infiltration in the top 35 cm in the 3-weekly watering
Plant Soil

treatment only accounted for 12 to 21 l m−2, indicatingthat a large amount of the applied water infiltrated tosoil layers >35 cm, and hence this water was notavailable to the shallow-rooted understorey species.
In contrast, in the weekly watering treatment, infiltra-tion in the top 35 cm accounted, on average, for 75 %of the applied water, with 1.5 to 6.8 l m−2 infiltratingdeeper soil layers.
Fig. 1 (a) Daily average soil temperature (°C, black line), dailymaximum and minimum soil temperature (°C, grey line) anddaytime (11.00 h–18.00 h) water vapour pressure deficit (VPDin kPa, grey area) at the experimental site over the course of thestudy. (b) Quantity of water received by the experimental plotsduring the growing season between October 2010 and June2011. All plots received equal water inputs until the end ofNovember, when the water manipulation treatment started.Black bar=natural precipitation, light grey bar=weeklywatering treatment, dark grey bar=3-weekly watering
treatment. (c) Volumetric soil water content (%) at a depth of10 cm in the experimental plots for the period between October2010 and June 2011. Black line=natural precipitation, light greyline=weekly watering treatment, dark grey line=3-weeklywatering treatment. Data represent 12-hourly values, with n=4. Error bars are removed for clarity. Dashed horizontal linesshow the upper and lower thresholds for soil water stress,representing the soil water content at field capacity and perma-nent wilting point
Plant Soil

Primary productivity
As low temperatures in winter inhibit plant growth in thestudied ecosystem, development of the herbaceous veg-etation is primarily confined to the period betweenFebruary and May. Over the growing season, ANPPincreased significantly (p<0.001), reaching a peak in latespring (Table 1). Subsequently, the sward senesced dueto increasing temperatures and decreasing soil wateravailability. Precipitation variability, with a shift to fewer,but larger rain events, had no effect on ANPP or BNPP.
Soil CO2 fluxes
The temporal pattern of Rs over the course of thegrowing season was significantly (p<0.001) affectedby date (Fig. 3), with Rs increasing over time. Mean Rs
in the period of December to February was0.7 μmol m−2 s−1, with irrigation having no effect onRs. In April and May, Rs rates increased, averaging2.5 μmol m−2 s−1. In this period, irrigation led tohigher Rs in the 3-weekly watering treatment on alloccasions, while Rs in the weekly watering treatment
Table 1 Aboveground net primary productivity (ANPP), per-centage senescence of biomass and belowground net primaryproductivity (BNPP) in the weekly and 3-weekly watering
treatments. Data represent mean±SE, n=4. No significant dif-ferences (at p<0.05) were found between the two treatments
ANPP (g m−2) % Senescence BNPP (g m−2)
Weekly watering 3-weekly watering Weekly watering 3-weekly watering Weekly watering 3-weekly watering
Feb 1 111±7.6 124±15.6 – – – –
April 5 215±16.6 211±38.2 17±3.0 15±2.1 – –
May 17 375±64.3 390±63.0 38±3.0 41±2.8 214±18.3 230±33.9
June 6 361±46.6 409±70.4 – – – –
Fig. 2 Water infiltration in the top 35 cm of the soil profile,assessed 24 h after the watering events. Measurements onlyperformed on dates when both the weekly and 3-weeklywatering treatments were irrigated. Bars (black=weeklywatering treatment, grey=3-weekly watering treatment)
represent mean±SE, n=4. Asterisks mark significant differences(*p<0.05, **p<0.01) between treatments. The white bars indi-cate the total amount of water given to the experimental plots(16 l m-2 for the weekly watering treatment, 48 l m-2 for the 3-weekly watering treatment). Data are not available for 20 April
Plant Soil

only increased with irrigation in April. No significantdifferences were found between the weekly and 3-weekly watering treatment, except for May 11.
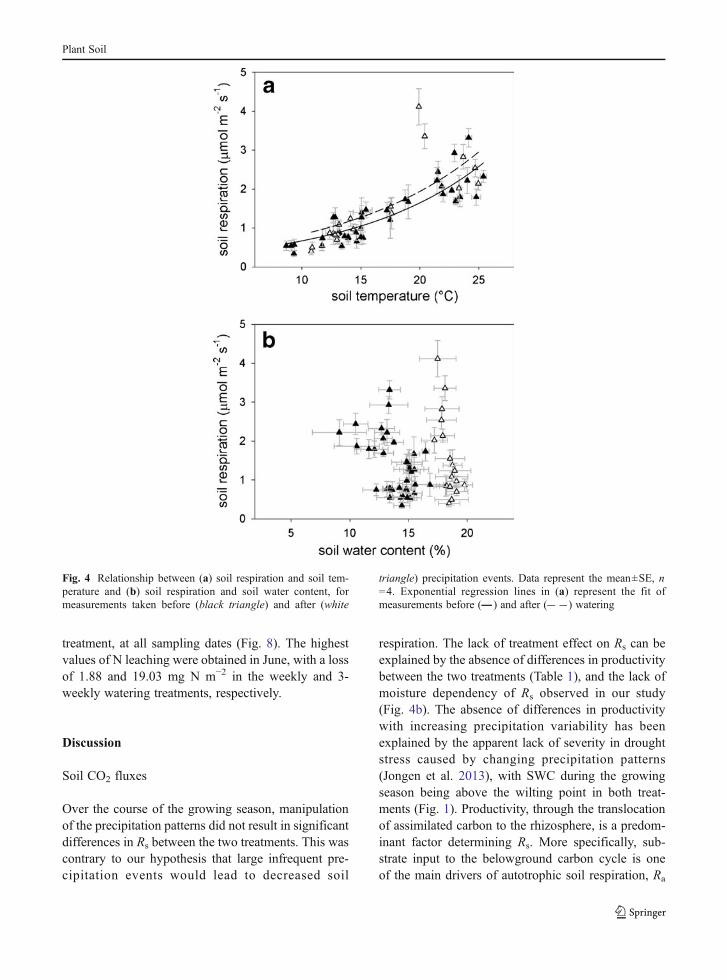
Over the course of the growing season, based onmeasurements taken prior to precipitation events, varia-tion in Rs was highly correlated with Ts (Fig. 4a), withchanges in Ts explaining 77% of the variation inRs (Rs=0.27e0.090Ts). After watering events, with all measure-ments taken at high soil moisture, the dependence of Rson Ts exhibited a similar slope (Rs=0.36e
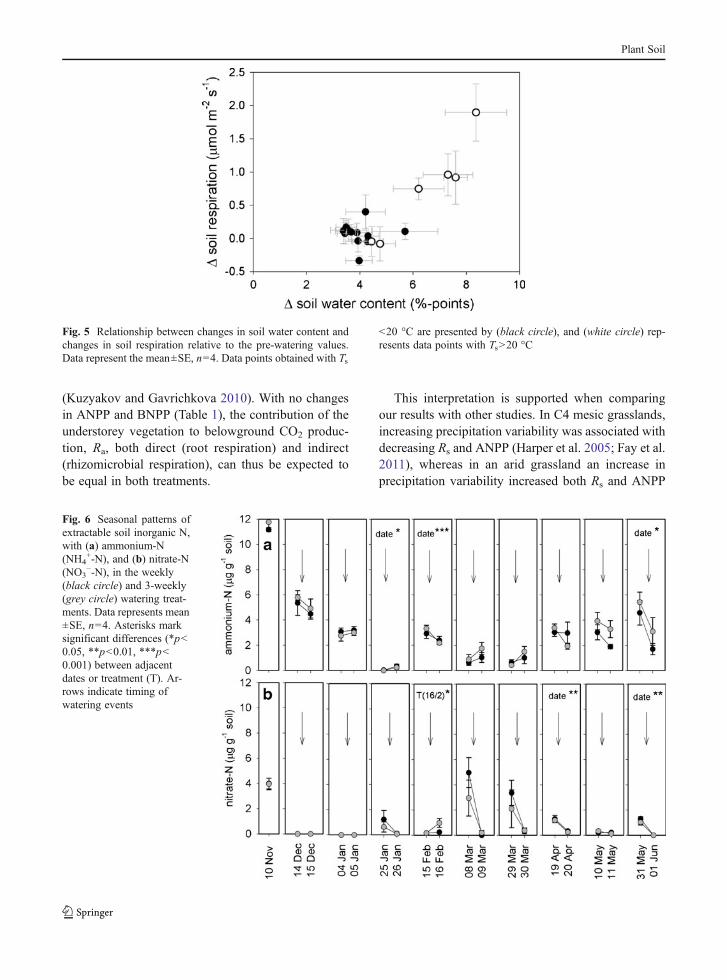
0.084Ts) ascompared to measurements taken before wateringevents, with a minor difference in intercepts, whichwas solely due to two high Rs values on April 20 (3.4and 4.1 μmol m−2 s−1 in the weekly and 3-weeklytreatment, respectively), which deviated from the expo-nential regression curve. The temperature sensitivity ofrespiration (Q10) was 2.46 and 2.32, based on measure-ments taken before and after watering, respectively.There was no significant correlation between Rs andSWC (Fig. 4b). However, absolute changes in Rs withirrigation was related to the magnitude of the change inSWC (Fig. 5). This was most pronounced in April, witha change in SWC of 8.4 %-points in the 3-weeklywatering treatment, which resulted in an increase of Rsby 1.90 μmol m−2 s−1.
Soil N dynamics
Seasonal dynamics of soil ammonium-N (NH4+-N)
and nitrate-N (NO3−-N), as shown in Fig. 6, indicated
a substantial available N pool in November. However,soil available NH4
+-N and NO3−-N rapidly decrease in
winter, with a subsequent significant (p<0.001)
increase in NH4+-N during spring. On several occa-
sions, irrigation resulted in significant differences insoil NH4
+-N or NO3−-N, as compared to soil inorganic
N concentrations prior to the watering treatment.However, the magnitude and direction of this changediffered considerably. For the period of mid January toJune, soil NH4
+-N correlated well with Ts (beforewatering: NH4
+-N=0.15e0.15Ts, r2 = 0.66, afterwatering: NH4
+-N=0.60e0.06Ts, r2=0.38), with no cor-relation between Ts and soil NO3
−-N (Fig. 7).No significant differences were found between the
two watering treatments in soil NH4+-N or NO3
−-N onany of the sampling dates, except for NO3
−-N onFebruary 16 (p=0.022). Similarly, cumulative soil Navailability over the growing season, measured usingion-exchange resin capsules, was not found to differsignificantly between the watering treatments, with onaverage 0.63±0.06 mg NH4
+-N capsule−1 and 0.81±0.05 mg NO3
−-N capsule−1 in both treatments.Analysis of soil pore water samples for NH4
+-Nand NO3
−-N showed that nitrate was the only formof inorganic N present in pore water samples. Theconcentration of NO3
−-N in soil pore water was notsignificantly different between the watering treat-ments, but was affected by sampling date (p<0.001),with a higher concentration of NO3
−-N in June, ascompared to the three other sampling dates (Fig. 8).
Leaching of nitrate from the herbaceous rootingzone was estimated using data on water infiltration,as depicted in Fig. 2, and soil pore water NO3
−-Nconcentration (Fig. 8). Results show a significant (p<0.001) increase in leaching in the 3-weekly wateringtreatment, as compared to the weekly watering
Fig. 3 Seasonal patterns of soil respiration in the weekly (blackcircle) and 3-weekly (grey circle) watering treatments. Data rep-resents mean±SE, n=4. Asterisks mark significant differences (*p
<0.05, **p<0.01) between adjacent dates, treatments (T), or in-teraction between date and treatment (D×T). Arrows indicatetiming of watering events
Plant Soil
treatment, at all sampling dates (Fig. 8). The highestvalues of N leaching were obtained in June, with a lossof 1.88 and 19.03 mg N m−2 in the weekly and 3-weekly watering treatments, respectively.
Discussion
Soil CO2 fluxes
Over the course of the growing season, manipulationof the precipitation patterns did not result in significantdifferences in Rs between the two treatments. This wascontrary to our hypothesis that large infrequent pre-cipitation events would lead to decreased soil
respiration. The lack of treatment effect on Rs can beexplained by the absence of differences in productivitybetween the two treatments (Table 1), and the lack ofmoisture dependency of Rs observed in our study(Fig. 4b). The absence of differences in productivitywith increasing precipitation variability has beenexplained by the apparent lack of severity in droughtstress caused by changing precipitation patterns(Jongen et al. 2013), with SWC during the growingseason being above the wilting point in both treat-ments (Fig. 1). Productivity, through the translocationof assimilated carbon to the rhizosphere, is a predom-inant factor determining Rs. More specifically, sub-strate input to the belowground carbon cycle is oneof the main drivers of autotrophic soil respiration, Ra
Fig. 4 Relationship between (a) soil respiration and soil tem-perature and (b) soil respiration and soil water content, formeasurements taken before (black triangle) and after (white
triangle) precipitation events. Data represent the mean±SE, n=4. Exponential regression lines in (a) represent the fit ofmeasurements before ( ) and after ( ) watering
Plant Soil
(Kuzyakov and Gavrichkova 2010). With no changesin ANPP and BNPP (Table 1), the contribution of theunderstorey vegetation to belowground CO2 produc-tion, Ra, both direct (root respiration) and indirect(rhizomicrobial respiration), can thus be expected tobe equal in both treatments.
This interpretation is supported when comparingour results with other studies. In C4 mesic grasslands,increasing precipitation variability was associated withdecreasing Rs and ANPP (Harper et al. 2005; Fay et al.2011), whereas in an arid grassland an increase inprecipitation variability increased both Rs and ANPP
Fig. 6 Seasonal patterns ofextractable soil inorganic N,with (a) ammonium-N(NH4
+-N), and (b) nitrate-N(NO3
−-N), in the weekly(black circle) and 3-weekly(grey circle) watering treat-ments. Data represents mean±SE, n=4. Asterisks marksignificant differences (*p<0.05, **p<0.01, ***p<0.001) between adjacentdates or treatment (T). Ar-rows indicate timing ofwatering events
Fig. 5 Relationship between changes in soil water content andchanges in soil respiration relative to the pre-watering values.Data represent the mean±SE, n=4. Data points obtained with Ts
<20 °C are presented by (black circle), and (white circle) rep-resents data points with Ts>20 °C
Plant Soil

(Thomey et al. 2011; Vargas et al. 2012). Thus, al-though these studies show the contrasting sensitivity ofdifferent ecosystems to increased precipitation variability,the linkage between Rs and ANPP was parallel to ourstudy.
During the growing season, Rs rates ranged between0.3 and 4.1 μmol m−2 s−1, with a Q10 of 2.32–2.46,which is similar to values reported in otherMediterranean systems. Ramos (2010) reported Rs
values between 1.6 and 5.1 μmol m−2 s−1 for theNovember to June period, and a Q10 of 1.4, in a nearbystudy-site. Comparing Mediterranean ecosystems,Correia et al. (2012) reported springtime Rs in openforests, closed forests and grasslands (3.1, 2.8 and2.7 μmol m−2 s−1, respectively), with our study site
(2.8 μmol m−2 s−1) ranging at the lower margin for openMediterranean forests. Site-specific soil parameters,with lower levels of soil organic matter (C/N=20.9) inour site compared to more productive sites (C/N=11.5–16.4) can explain the observed differences.
Our results showed that, even though the temporalvariability in SWC in the 3-weekly watering treatmentwas more pronounced, the seasonal pattern of Rs
closely followed Ts (Fig. 4a), as previously reportedfor other Mediterranean ecosystems (Joffre et al. 2003;Curiel Yuste et al. 2007; Almagro et al. 2009;Gavrichkova et al. 2010). This observed high temper-ature sensitivity is another indicator for the close rela-tion between understorey productivity and Rs, as wasdescribed in a meta-analysis by Wang et al. (2010),where Q10 was found to be closely related to vegeta-tion activity.
From April onwards, increasing Ts and low SWC atthe end of the dry periods resulted in Rs rates increas-ing in response to irrigation (Fig. 3), with differencesobserved between the two manipulation treatments.This increase in Rs following a precipitation pulse, aphenomenon referred to as the ‘Birch effect’ (Jarvis etal. 2007), is related to a stimulation of microbialactivity in response to wetting of dry soils (Birch1964; Fierer and Schimel 2002; Kim et al. 2012;Unger et al. 2012). Austin et al. (2004) suggested thatsmall precipitation events would only stimulate micro-bial activity in the top layer of soil, while large
Fig. 7 Relationship between (a) ammonium-N (NH4+-N) and
soil temperature and (b) nitrate-N (NO3−-N) and soil tempera-
ture, for measurements taken before (black triangle) and after(white triangle) precipitation events. Data represent the mean±SE, n=4. Exponential regression lines in (a) represent the fit ofmeasurements before ( ) and after ( ) watering
Fig. 8 Concentration of nitrate-N (NO3−-N) in soil pore water
collected at a depth of 30 cm 24 h after the watering events inthe weekly (black circle) and 3-weekly (grey circle) wateringtreatments. Bars represent N leaching, quantified by combiningdata on soil water dynamics (see Fig. 2) and NO3
−-N in soil porewater in the weekly (black bar) and 3-weekly (grey bar)watering treatments. Data represents mean±SE, n=4. Data for20 April are missing
Plant Soil

precipitation events could reach the rooting zone andtrigger assimilation processes (Reynolds et al. 2004),subsequently transferring more labile carbon sub-strates via roots to microorganisms (Aanderud et al.2011). The precipitation pulses in our two treatmentswere large enough to increase SWC in the top 35 cm(Fig. 2), encompassing the rooting zone, subsequentlyinfluencing aboveground carbon uptake, as confirmedby an increase in net CO2 exchange and CO2 assimi-lation by the understorey vegetation in April and Mayafter watering (Jongen et al. 2013).
Our results showed that the size of the Rs burst after aprecipitation pulse was related to the magnitude of thechange in soil moisture (Fig. 5), as has previously beenshown in other studies (Cable et al. 2008; Fischer 2009;Unger et al. 2010b). This indicates that prior to irriga-tion, soil moisture was already limiting microbialgrowth and metabolic activity, even with SWC abovethe wilting point. The particularly sandy nature of thesoil, which inhibits diffusion of soluble substrates evenunder moderate moisture stress conditions (Olesen et al.2001), can explain this finding. Increases in Rs in re-sponse to watering were only observed when the changein SWC was more that 6 %-points (Fig. 5), explainingthe differential treatment responses of Rs to precipitationin May (Fig. 3). However, compared to other studies(Xu and Baldocchi 2004; Jarvis et al. 2007; Cable et al.2008; Inglima et al. 2009; Unger et al. 2010b; Unger etal. 2012), which were carried out during the summerdrought, the observed CO2 pulses in our study in re-sponse to precipitation were small, which may beexpected, as during the growing season moisturechanges and antecedent soil moisture were moderate.Thus, although the ‘Birch effect’ may exert a largeinf luence on ecosys tem carbon balance inMediterranean systems (Unger et al. 2012), altered pre-cipitation regimes during the growing period of theherbaceous understorey will probably only have aminor impact on soil carbon losses in the studiedsystem.
Soil N dynamics
Increased variability in precipitation regimes is expectedto alter N cycling through its effects on plant productivity(N demand), microbial activity (N mineralization) andabiotic processes, such as leaching (N losses). In contrastto our hypothesis that large infrequent precipitationevents would decrease soil N availability, our results
showed that precipitation manipulation had no signifi-cant effect on temporal variation of soil NH4
+-N orNO3
−-N over the course of the growing season. Thiscan be explained by the lack of a treatment effect on(1) Rs (Fig. 3), indicating no changes in mineralizationprocesses; and on (2) ANPP (Table 1), suggesting equalN demand by the vegetation. The lack of a treatmenteffect on mineralization processes is supported by theresults on cumulative N availability, showing no differ-ences between the weekly and 3-weekly watering treat-ment, thus suggesting no treatment effect on netmineralization rates. In a mixed grass prairie, Heisler-White et al. (2009) reported no difference in soil Navailability in response to larger, infrequent precipitationevents, which is in agreement with our results. However,with no data on Rs in the above-mentioned study, and alarge increase in ANPP, the lack of response in N avail-ability cannot be explained following our rationale, asoutlined above. A lab incubation study withMediterranean soils by Fierer and Schimel (2002) foundthat increasing drying-wetting frequency reflected nega-tively on soil N availability although the nitrificationpotential dramatically increased, which was explainedby increased losses of NO3
−-N through leaching.Further, simulating different precipitation patterns, with-out altering total water input, Yahdjian and Sala (2010)reported an increase in NO3
−-N leaching with largeprecipitation events. Thus, both size and frequency ofthe pulses might reflect on soil N availability. In ourstudy, the concentration of NO3
−-N in soil pore waterdid not differ between the watering treatments. However,water infiltration to deeper (> 35 cm) soil layers wassignificantly higher in the 3-weekly watering treatment,and as a result, the large, infrequent watering eventsincreased leaching of NO3
−-N. However, this NO3−-N
loss for the herbaceous understorey by leaching fromupper soil layers might be beneficial to Q. suber trees,as their extensive root system enables water acquisitionfrom deeper soil layers (Cubera and Moreno 2007).
As the majority of grassland species were shown topreferentially take up NO3
−-N (Austin et al. 2006),and species of neutral soils perform best with NO3
−-N (Falkengren-Gerup 1995), we assume a preferentialplant uptake of NO3
−-N in our ecosystem. This pref-erence, together with the higher mobility of NO3
−-N,which promotes leaching of this ion (Marschner2012), can explain the seasonal patterns of NH4
+-Nand NO3
−-N, and the effects of precipitation events onthese two soil inorganic N species.
Plant Soil

The absence of soil NO3−-N in December and
January probably resulted from nitrate losses throughleaching and low nitrification rates, with nitrificationpotential decreasing with low soil temperature (Sabey1956; Gallardo et al. 2000). In March, with a modestplant N demand, prior to the precipitation event NO3
−-N was the largest component of the extractable inor-ganic N. In April and May, plant biomass of theherbaceous understorey rapidly increased (Table 1),with simultaneously increased N demand, thereby de-pleting N reserves in the soil, indicating that N is alimiting factor for biomass development. This isreflected in soil NO3
−-N decreasing >90 % prior towatering during this period (4.0 μg g−1 on March 8–0.3 μg g−1 on May 10), which was correlated withnitrate concentrations of soil pore water (r2=0.73,p=0.03). In contrast, Otieno et al. (2011), reportedan increase in the soil N pool between February andMay in a Portuguese montado ecosystem, suggesting thatthe herbaceous layer did not deplete soil N. However,these differences can be explained by the substantiallylower ANPP reported in Otieno et al. (2011), i.e.180 g m−2 as compared to 385 g m−2 in our study.
At the end of May, increasing temperatures anddecreasing SWC resulted in the understorey vegeta-tion rapidly senescing. Plant senescence, generatinglarge inputs of fresh plant material, has been shownto increase mineralization rates and soil N (Augustineand McNaughton 2004; Dujardin et al. 2012). Indeed,as vegetation senesced, we observed high concentra-tions of NO3
−-N in soil pore water after wateringevents, indicating an increased leaching of this ion.
The substantial NH4+-N pool present in November
originated from mineralization processes that occurredin autumn. Subsequently, the high levels of NH4
+-Nfound in December and January can be explained bythe low N demand by plants, which germinate inautumn but do not exhibit significant growth ratesuntil February. For the period of mid January toJune, soil NH4
+-N ranged from 0 to 5 μg g−1, whichis comparable to values reported in other montadoecosystems (Otieno et al. 2011; Shvaleva et al.2011). Similar to Rs, soil NH4
+-N correlated well withTs, with low temperatures in the December-Februaryperiod inhibiting mineralization. During spring, whentemperatures rapidly increased, enhanced microbialactivity and N mineralization was reflected in in-creased Rs and soil NH4
+-N. Focusing on the differ-ences in NH4
+-N around subsequent precipitation
events, the direction of response, with soil NH4+-N
increasing after precipitation pulses in March and de-creasing in April and May, can be explained by in-creased plant N demand (Jamieson et al. 1998;Dujardin et al. 2012). As ANPP and N demand rapidlyincreased between the beginning of April and mid-May (Table 1), and nitrate became increasingly scarcedue to leaching effects after watering events, plantsmost probably increased their ammonium uptake.
Synchrony versus asynchrony
Seasonal dynamics of soil N availability have beenstudied at length in a variety of ecosystems, withrecent research attention being given to the phenome-non of synchrony/asynchrony between nitrogenmineralization and plant productivity (Augustineand McNaughton 2004; Yahdjian and Sala 2010;Giese et al. 2011; Kaiser et al. 2011; Dijkstra etal. 2012).
Our results pinpoint to a synchronized pattern be-tween Rs, soil inorganic N and plant productivity ofthe herbaceous understorey. This presence of synchronybetween N supply by microbes and N demand by plantsis in agreement with previous studies in semi-arid grass-lands (Giese et al. 2011; Dijkstra et al. 2012). Similarly,in mesic ecosystems, studies report synchrony betweenmicrobial and plant processes (e.g. Vitousek et al. 1998;McCulley et al. 2009; Bobbink et al. 2010), these eco-systems being characterized by minimal activity duringautumn-winter and a rapid increase of microbial activity,nitrogen mineralization and plant growth in spring.However, several studies report asynchronicity betweenN supply and demand in arid, semi-arid andMediterranean-type ecosystems (Jackson et al. 1988;Augustine and McNaughton 2004), due to the largefluctuations in soil moisture, with differences in thehierarchy, intricately linked to differences in thresholds,of the responses of plants and microbes to changes insoil water (Schwinning and Sala 2004; Collins et al.2008). Moreover, increased precipitation variabilitycould amplify the asynchronicity in N processing, asdecoupling N supply and demand may affect the Nretention capacity of the ecosystem, with the excess Nbeing lost through leaching, emissions, or both (deSchrijver et al. 2008). In our ecosystem, increasingprecipitation variability did not affect synchronicity, aslarge, infrequent watering events did not affect the tem-poral dynamics of the studied below- and aboveground
Plant Soil

ecosystem variables. However, the increase in Nleaching with large infrequent precipitation events mayresult in a shift from a closed internal N cycle to an‘open’ cycle, with long-term consequences for nutrientdynamics and plant productivity.
Conclusions
This large-scale rainfall manipulation study contributesto an improved understanding of the responses of atypical Mediterranean ecosystem to the predictions ofclimate change. We showed that, in contrast to ourhypothesis, changes in precipitation regimes, with largeinfrequent precipitation events and longer interveningdry periods, without altering total annual precipitationinputs, are not likely to exert significant effects on eitherbelowground C or N cycling during the understoreygrowing season in the studied ecosystem. Althoughthe applied treatments induced distinctly different tem-poral patterns in SWC, all measured parameters, includ-ing productivity, Rs and soil inorganic N concentration,were influenced by temperature rather than soil mois-ture, indicating that the precipitation manipulation nevercaused substantial stress for microbial and plant activity.Our results showed that increased system carbon lossesby the ‘Birch effect’, as expected from the changes inprecipitation variability, only played a minor role duringthe growing season. In addition, over the course of thegrowing season, the synchrony between Rs, soil inor-ganic N and plant productivity of the herbaceousunderstorey was not affected by increased precipitationvariability, indicating ecosystem resilience to these cli-matic changes. However, increased infiltration and Nleaching to deeper soil layers, as observed with largeprecipitation events, might have long-term conse-quences for microbial activity and understorey plantperformance, as nitrogen might be increasingly lostfrom the system, although this effect on the ecosystemmight be buffered by increased N uptake by Q. subertrees. The possible long-term effects of increased pre-cipitation variability on soil C and N require furtherstudy, as understanding climate change effects on eco-system processes in Mediterranean biomes is crucial fordeveloping mitigation approaches and future manage-ment strategies.
Acknowledgements The authors acknowledge the financialsupport of FCT (Fundação para a Ciência e Tecnologia),
through the project PTDC/CLI/64480/2006 and throughpostdoctoral fellowships to Marjan Jongen (SFRH/BPD/79662/2011) and David Fangueiro (SFRH/BPD/84229/2012). The authors wish to thank António GonçalvesFerreira for providing the field site at Machoqueira doGrou, the staff of Machoqueira do Grou, especially Mr.Pirra, for continuous support to the work, Filipe Costa eSilva, Joaquim Mendes, Francesco Minunno and AlexandraCorreia for assisting with the construction of the rain-outshelters, Prof. Fernando Monteiro for the measurements ofwilting point, Marta Pintó-Marijuan and Elsa Breia forvaluable support in the laboratory, and Peter Fay for sug-gestions on the manuscript.
References
Aanderud ZT, Schoolmaster DR Jr, Lennon JT (2011) Plantsmediate the sensitivity of soil respiration to rainfall vari-ability. Ecosystems 14:156–167
Aires LMI, Pio CA, Pereira JS (2008) Carbon dioxide exchangeabove a Mediterranean C3/C4 grassland during two climato-logically contrasting years. Glob Change Biol 14:539–555
Almagro M, López J, Querejeta JI, Martínez-Mena M (2009)Temperature dependence of soil CO2 efflux is strongly mod-ulated by seasonal patterns of moisture availability in aMediterranean ecosystem. Soil Biol Biochem 41:594–605
Augustine DJ, McNaughton SJ (2004) Temporal asynchrony insoil nutrient dynamics and plant production in a semiaridecosystem. Ecosystems 7:829–840
Austin AT, Sala OE, Jackson RB (2006) Inhibition of nitrifica-tion alters carbon turnover in the Patagonian steppe.Ecosystems 9:1257–1265
Austin AT, Yahdjian L, Stark JM, Belnap J, Porporato A, NortonU, Ravetta DA, Schaeffer SM (2004) Water pulses andbiogeochemical cycles in arid and semiarid ecosystems.Oecologia 141:221–235
Binkley D, Hart SC (1989) The components on nitrogen avail-ability assessments in forest soils. Adv Soil Sci 10:57–112
Birch HF (1964) Mineralisation of plant nitrogen followingalternate wet and dry conditions. Plant Soil 20:43–49
Bobbink R, Hicks K, Galloway J, Spranger T, Alkemade R,Ashmore M, Bustamante M, Cinderby S, Davidson E,Dentener F, Emmett B, Erisman J-W, Fenn M, Gilliam F,Nordin A, Pardo L, de Vries W (2010) Global assessmentof nitrogen deposition effects on terrestrial plant diversity:A synthesis. Ecol Appl 20:30–59
Borken W, Matzner E (2009) Reappraisal of drying and wettingeffects on C and N mineralization and fluxes in soils. GlobChange Biol 15:808–824
Bugalho MN, Caldeira MC, Pereira JS, Aronson J, Pausas JG(2011) Mediterranean cork oak savannas require humanuse to sustain biodiversity and ecosystem services. FrontEcol Environ 9:278–286
Cable JM, Ogle K, Williams DG, Weltzin JF, Huxman TE(2008) Soil texture drives responses of soil respiration toprecipitation pulses in the sonoran desert: Implications forclimate change. Ecosystems 11:961–979
Plant Soil

Chou WW, Silver WL, Jackson RD, Thompson AW, Allen-DiazB (2008) The sensitivity of annual grassland carbon cy-cling to the quantity and timing of rainfall. Glob ChangeBiol 14:1382–1394
Collins SL, Sinsabaugh RL, Crenshaw C, Green L, Porras-AlfaroA, Stursova M, Zeglin LH (2008) Pulse dynamics and mi-crobial processes in aridland ecosystems. J Ecol 96:413–420
Correia AC, Minunno F, Caldeira MC, Banza J, Mateus J,Carneiro M, Wingate L, Shvaleva A, Ramos A, JongenM, Bugalho MN, Nogueira C, Lecomte X, Pereira JS(2012) Soil water availability strongly modulates soilCO2 efflux in different Mediterranean ecosystems: Modelcalibration using the Bayesian approach. Agric EcosystEnviron 161:88–100
Crespo D (2010) Species diversity: David crespo takes C3pastures the next step to boost soil carbon. Aust Farm J20:44–47
Cubera E, Moreno G (2007) Effect of single Quercus ilex treesupon spatial and seasonal changes in soil water content indehesas of central western Spain. Ann For Sci 64:355–364
Cui MY, Caldwell MM (1997) A large ephemeral release ofnitrogen upon wetting of dry soil and corresponding rootresponses in the field. Plant Soil 191:291–299
Curiel Yuste J, Baldocchi DD, Gershenson A, Goldstein A,Misson L, Wong S (2007) Microbial soil respiration andits dependency on carbon inputs, soil temperature andmoisture. Glob Change Biol 13:1–18
de Schrijver A, Verheyen K, Mertens J, Staelens J, Wuyts K,Muys B (2008) Nitrogen saturation and net ecosystemproduction. Nature 451:E1. doi:10.1038/nature06578
Dijkstra FA, Augustine DJ, Brewer P, von Fischer JC (2012)Nitrogen cycling and water pulses in semiarid grasslands:are microbial and plant processes temporally asynchronus?Oecologia 170:799–808
Dujardin G, Bureau F, Vinceslas-Akpa M, Decaëns T, LangloisE (2012) Soil functioning in a mosaic of herbaceous com-munities of a chalky environment: Temporal variations ofwater availability and N dynamics. Plant Soil 360:197–213
Easterling DR, Evans JL, Groisman PY, Karl TR, Kunkel KE,Ambenje P (2000) Observed variability and trends in ex-treme climate events: A brief review. Bull Am MeteorolSoc 81:417–442
Falkengren-Gerup U (1995) Interspecies differences in the pref-erence of ammonium and nitrate in vascular plants.Oecologia 102:305–311
Fay PA, Blair JM, Smith MD, Nippert JB, Carlisle JD, Knapp AK(2011) Relative effects of precipitation variability andwarming on tallgrass prairie ecosystem function.Biogeosciences 8:3053–3068, Biogeosciences 8:3052–3068
Fay PA, Carlisle JD, Knapp AK, Blair JM, Collins SL (2003)Productivity responses to altered rainfall patterns in a C4-dominated grassland. Oecologia 137:245–251
Fierer N, Schimel JP (2002) Effects of drying-rewetting fre-quency on soil carbon and nitrogen transformations. SoilBiol Biochem 34:777–787
Fischer T (2009) Substantial rewetting phenomena on soil res-piration can be observed at low water availability. Soil BiolBiochem 41:1577–1579
Gallardo A, Rodríguez-Saucedo JJ, Covelo F, Fernández-Alés R(2000) Soil nitrogen heterogeneity in a dehesa ecosystem.Plant Soil 222:71–82
Gavrichkova O, Moscatelli MC, Kuzyakov Y, Grego S,Valentini R (2010) Influence of defoliation on CO2 effluxfrom soil and microbial activity in a Mediterranean grass-land. Agric Ecosyst Environ 136:87–96
Giese M, Gao YZ, Lin S, Brueck H (2011) Nitrogen availabilityin a grazed semi-arid grassland is dominated by seasonalrainfall. Plant Soil 340:157–167
Goudriaan J, van Laar HH (1994) Modelling potential cropgrowth processes. Kluwer Academic Publishers, Dordrecht
Harper CW, Blair JM, Fay PA, Knapp AK, Carlisle JD (2005)Increased rainfall variability and reduced rainfall amountdecreases soil CO2 flux in a grassland ecosystem. GlobChange Biol 11:322–334
Heisler-White JL, Blair JM, Kelly EF, Harmoney K, Knapp AK(2009) Contingent productivity responses to more extremerainfall regimes across a grassland biome. Glob ChangeBiol 15:2894–2904
Hodge A, Stewart J, Robinson D, Griffiths BS, Fitter AH (1999)Plant, soil fauna and microbial responses to N-rich organicpatches of contrasting temporal availability. Soil BiolBiochem 31:1517–1530
Houba VJ, van der Lee JJ, Novozamsky I, Walinga I (1989) Soiland plant analysis. Part 5—Soil analysis procedures.Wageningen Agricultural University, Wageningen
Huxman TE, Snyder KA, Tissue D, Leffler AJ, Pockman W,Ogle K, Sandquist D, Potts DL, Schwinning S (2004)Precipitation pulses and carbon balance in semi-arid andarid ecosystems. Oecologia 141:254–268
Inglima I, Alberti G, Bertoloni T, Vaccari FP, Giolo B, MigliettaF, Cortufo MF, Peressortti A (2009) Precipitation pulseenhance respiration of Mediterranean ecosystems: the bal-ance between organic and inorganic components of in-creased soil CO2 efflux. Glob Change Biol 15:1289–1301
IPCC (2007) Climate change 2007: Synthesis report. In: CoreWriting Team, Pachauri RK, Reisinger A (eds)Contribution of working groups I, II and III to the fourthassessment report of the intergovernmental panel on cli-mate change. IPCC, Geneva
Jackson LE, Strauss RB, Firestone MK, Bartolome JW (1988)Plant and soil nitrogen dynamics in California annualgrassland. Plant Soil 110:9–17
Jackson RB, Carpenter SR, Dahm CN, McKnight DM, NaimanRJ, Postel SL, Running SW (2001) Water in a changingworld. Ecol Appl 11:1027–1045
James JJ, Richards JH (2006) Plant nitrogen capture in pulse-driven systems: Interactions between root responses andsoil processes. J Ecol 94:765–777
Jamieson N, Barraclough D, Unkovich M, Monaghan R (1998)Soil N dynamics in a natural calcareous grassland underchanging climate. Biol Fert Soils 27:267–273
Jarvis PG, Rey A, Petsikos C, Wingate L, Rayment M, PereiraJS, Banza J, David JS, Miglietta F, Borgetti M, Manca G,Valentini R (2007) Drying and wetting of Mediterraneansoils stimulates decomposition and carbon dioxide emis-sion: The “birch effect”. Tree Physiol 27:929–940
Joffre R, Ourcival J-M, Rambal S, Rocheteau A (2003) Thekey-role of topsoil moisture on CO2 efflux from aMediterranean Quercus ilex forest. Ann For Sci 60:519–526
Jongen M, Lecomte X, Unger S, Pintó-Marijuan M, Pereira JS(2013) The impact of changes in the timing of precipitation
Plant Soil

on the herbaceous understorey of Mediterranean oakwoodlands. Agr Forest Meteorol 171–172:163–173
Jongen M, Pereira JS, Aires LM, Pio CA (2011) The effects ofdrought and timing of precipitation on the inter-annual vari-ation in ecosystem-atmosphere exchange in a Mediterraneangrassland. Agr Forest Meteorol 151:595–606
Kaiser C, Fuchslueger L, Koranda M, Gorfer M, Stange CF,Kitzler B, Rasche F, Strauss J, Sessitsch A, Zechmeister-Boltenstern S, Richter A (2011) Plants control the seasonaldynamics of microbial N cycling in a beech forest soil bybelowground C allocation. Ecology 92:1036–1051
Kim D-G, Vargas R, Bond-Lamberty B, Turetsky MR (2012)Effects of soil rewetting and thawing on soil gas fluxes: areview of current literature and suggestions for future re-search. Biogeosciences 9:2459–2483
Knapp AK, Fay PA, Blair JM, Collins SL, Smith MD, CarlisleJD, Harper CW, Danner BT, Lett MS, McCarron JK (2002)Rainfall variability, carbon cycling, and plant species di-versity in a mesic grassland. Science 298:2202–2205
Kuzyakov Y, Gavrichkova O (2010) Time lag between photosyn-thesis and carbon dioxide efflux from soil: A review ofmechanisms and controls. Glob Change Biol 16:3386–3406
Li SG, Asanuma J, Eugster W, Kotani A, Liu JJ, Urano T,Oikawa T, Davaa G, Oyunbaatar D, Sugita M (2005) Netecosystem carbon dioxide exchange over grazed steppe incentral Mongolia. Glob Change Biol 11:1941–1955
Lloyd J, Taylor JA (1994) On the temperature dependence ofsoil respiration. Funct Ecol 8:315–323
Luterbacher J, Xoplaki E, Casty C, Wanner H, Pauling A, KüttelM, Rutishauser T, Brönnimann S, Fischer E, Fleitmann D,Gonzalez-Rouco FJ, García-Herrera R, Barriendos M,Rodrigo F, Gonzalez-Hidalgo JC, Saz MA, Gimeno L,Ribera P, Brunet M, Paeth H, Rimbu N, Felis T, JacobeitJ, Dünkeloh A, Zorita E, Guiot J, Türkes M, AlcoforadoMJ, Trigo R, Wheeler D, Tett S, Mann ME, Touchan R,Shindell DT, Silenzi S, Montagna P, Camuffo D, MariottiA, Nanni T, Brunetti M, Maugeri M, Zerefos C, De Zolt S,Lionello P, Nunes MF, Rath V, Beltrami H, Garnier E,Ladurie ELR (2006) Mediterranean climate variability overthe last centuries; a review. In: Lionello P, Malanotte-Rizzoli P, Boscolo R (eds) The Mediterranean climate:An overview of the main characteristics and issues.Elsevier, Amsterdam, pp 27–148
Marschner P (2012) Marschner’s mineral nutrition of higherplants. Academic, London
McCulley RL, Boutton TW, Archer SR (2007) Soil respirationin a subtropical savanna parkland: Response to water ad-ditions. Soil Sci Soc Am J 71:820–828
McCulley RL, Burke IC, Lauenroth WK (2009) Conservation ofnitrogen increases with precipitation across a major grass-land gradient in the central great plains of North America.Oecologia 159:571–581
Moreno Marcos G, Obrador JJ, García E, Cubera E, MonteroMJ, Pulido F, Dupraz C (2007) Driving competitive andfacilitative interactions in oak dehesas through manage-ment practices. Agroforest Syst 70:25–40
Olesen T, Moldrup O, Yamaguchi T, Rolston DE (2001) Constantslope impedance factor model for predicting the solute diffu-sion coefficient in unsaturated soil. Soil Sci 166:89–96
Otieno DO, Mirzaei H, Hussain MZ, Li YL, Schmidt MWT,Wartinger M, Jung E, Ribeiro N, Pereira JS, Tenhunen J
(2011) Herbaceous layer development during spring doesnot deplete soil nitrogen in the Portuguese montado. J AridEnviron 75:231–238
Patrick LD, Ogle K, Bell CW, Zak J, Tissue D (2009)Physiological responses of two contrasting desert plantspecies to precipitation variability are differentially regu-lated by soil moisture and nitrogen dynamics. Glob ChangeBiol 15:1214–1229
Pereira JS, Mateus JA, Aires LM, Pita G, Pio C, David JS,Andrade V, Banza J, David TS, Paço TA, Rodrigues A(2007) Net ecosystem carbon exchange in three contrastingMediterranean ecosystems – the effect of drought.Biogeosciences 4:791–802
Ramos AG (2010) Estudo dos factores que influenciam arespiração do solo de um montado de sobro. InstitutoSuperior de Agronomia. MSc thesis. Universidade Técnicade Lisboa, Lisbon, p 51
Raupach MR, Canadell JG (2010) Carbon and the Anthropocene.Curr Opin Env Sust 2:210–218
ReichsteinM, ReyA, Freibauer A, Tenhunen J, Valentini R, BanzaJ, Casals P, Cheng Y, Grünzweig J, Irvine J, Joffre R, Law B,Loustau D, Miglietta F, Oechel W, Ourcival JM, Pereira JS,Peressotti A, Ponti F, Qi Y, Rambal S, RaymentM, RomanyaJ, Rossi F, Tedeschi V, Tirone G, Xu M, Yakir D (2003)Modelling temporal and large-scale spatial variability of soilrespiration from soil water availability, temperature and veg-etation productivity indices. Glob Biogeochem Cy 17:1104
Rey A, Pegoraro E, Oyonarte C,Were A, Escribano P, Raimundo J(2011) Impact of land degradation on soil respiration in asteppe (Stipa tenacissima L.) semi-arid ecosystem in the SEof Spain. Soil Biol Biochem 43:393–403
Reynolds JF, Kemp PR, Ogle K, Fernandez RJ (2004)Modifying the ‘pulse-reserve’ paradigm for deserts ofNorth America: precipitation pulses, soil water, and plantresponses. Oecologia 141:194–210
Sabey BR (1956) Influence of temperature on nitrification insoils. Soil Sci Soc Am J 20:357–360
Schindlbacher A, Wunderlich S, Borken W, Kitzler B,Zechmeister-Boltenstern S, Jandl R (2012) Soil respirationunder climate change: prolonged summer drought offsetssoil warming effects. Glob Change Biol 18:2270–2279
Schwinning S, Sala OE (2004) Hierarchy of responses to re-source pulses in arid and semi-arid ecosystems. Oecologia141:211–220
Selsted MB, van der Linden L, Ibrom A, Michelsen A, LarsenKS, Pedersen JK, Mikkelsen TN, Pilegaard K, Beier C,Ambus P (2012) Soil respiration is stimulated by elevatedCO2 and reduced by summer drought: three years of mea-surements in a multifactor ecosystem manipulation exper-iment in a temperate heathland (CLIMAITE). Glob ChangeBiol 18:1216–1230
Shvaleva A, Lobo-do-Vale R, Cruz C, Castaldi S, Rosa AP, ChavesMM, Pereira JS (2011) Soil-atmosphere greenhouse gases(CO2, CH4 and N2O) exchange in evergreen oak woodlandin southern Portugal. Plant Soil Environ 57:471–477
Suseela VA, Conant RT, Wallenstein MD, Dukes JS (2012)Effects of soil moisture on the temperature sensitivity ofheterotrophic respiration vary seasonally in an old-fieldclimate change experiment. Glob Change Biol 18:336–348
Talmon Y, Sternberg M, Grünzweig JM (2011) Impact of rain-fall manipulations and biotic controls on soil respiration in
Plant Soil

Mediterranean and desert ecosystems along an aridity gra-dient. Glob Change Biol 17:1108–1118
ThomeyML, Collins SL, Vargas R, Johnson JE, Brown RF, NatvigDO, Friggens MT (2011) Effect of precipitation variability onnet primary production and soil respiration in a ChihuahuanDesert grassland. Glob Change Biol 17:1505–1515
Tiemann LK, Billings SA (2012) Tracking C and N flowsthrough microbial biomass with increased soil moisturevariability. Soil Biol Biochem 49
Unger S, Máguas C, Pereira JS, Aires LM, David TS, Werner C(2009) Partitioning carbon fluxes in a Mediterranean oakforest to disentangle changes in ecosystem sink strengthduring drought. Agr Forest Meteorol 149:949–961
Unger S, Máguas C, Pereira JS, Aires LM, David TS, Werner C(2010a) Disentangling drought-induced variation in eco-system and soil respiration using stable carbon isotopes.Oecologia 163:1043–1057
Unger S, Máguas C, Pereira JS, David TS, Werner C (2010b)The influence of precipitation pulses on soil respiration—Assessing the “Birch effect” by stable carbon isotopes. SoilBiol Biochem 42:1800–1810
Unger S, Máguas C, Pereira JS, David TS, Werner C (2012)Interpreting postdrought rewetting effects on soil and eco-system carbon dynamics in a Mediterranean oak savannah.Agr Forest Meteorol 154–155:9–18
Valentini R, Matteucci G, Dolman AJ, Schulze E-D, RebmannC, Moors EJ, Granier A, Gross P, Jensen NO, Pilegaard K(2000) Respiration as the main determinant of carbon bal-ance in European forests. Nature 404:861–865
Vargas R, Collins SL, Thomey ML, Johnson JE, Brown RF,Natvig DO, Friggens MT (2012) Precipitation variabilityand fire influence the temporal dynamics of soil CO2 effluxin an arid grassland. Glob Change Biol 18:1401–1411
Vitousek PM, Hedin LO, Matson PA, Fownes JH, Neff J (1998)Within-system element cycles, input–output budgets, andnutrient limitation. In: Pace ML, Groffmann PM (eds)Successes, limitations, and frontiers in ecosystem science.Springer, New York, pp 432–451
Wang X, Piao S, Ciais P, Janssens IA, Reichstein M, Peng S,Wang T (2010) Are ecological gradients in seasonal Q10 ofsoil respiration explained by climate or by vegetation sea-sonality? Soil Biol Biochem 42:1728–1734
Xoplaki E, Gonzalez-Rouco JF, Luterbacher J, Wanner H (2004)Wet seasonMediterranean precipitation variability: influenceof large-scale dynamics and trends. Clim Dynam 23:63–78
Xu L, Baldocchi DD (2004) Seasonal variation in carbon diox-ide exchange over a Mediterranean annual grassland inCalifornia. Agr Forest Meteorol 123:79–96
Yahdjian L, Sala OE (2010) Size of precipitation pulses controlsnitrogen transformation and losses in an arid Patagonianecosystem. Ecosystems 13:575–585
Yahdjian L, Sala OE, Austin AT (2006) Differential controls ofwater input on litter decomposition and nitrogen dynamicsin the Patagonian steppe. Ecosystems 9:128–141
Zhou X, Sherry RA, An Y, Wallice LL, Luo Y (2006) Main andinteractive effects of warming, clipping, and doubled pre-cipitation on soil CO2 efflux in a grassland ecosystem.Glob Biogeochem Cy 20, GB1003
Plant Soil




















