
Potential effects of plant protease inhibitors, oryzacystatin I and soybean Bowman-Birk inhibitor,...
11
Journal of Insect Physiology 51 (2005) 941–951 Potential effects of plant protease inhibitors, oryzacystatin I and soybean Bowman-Birk inhibitor, on the aphid parasitoid Aphidius ervi Haliday (Hymenoptera, Braconidae) H. Azzouz a,c, , E.D.M. Campan a , A. Cherqui a , J. Saguez a , A. Couty a , L. Jouanin b , P. Giordanengo a , L. Kaiser c,1 a BPCIR, EA 3900 Universite´de Picardie Jules Verne, 33 rue Saint Leu, 80039 Amiens Cedex 1, France b INRA-CNRS, Laboratoire de Biologie Cellulaire, route de St Cyr, 78026 Versailles Cedex, France c Laboratoire de Neurobiologie Compare´e des Inverte´bre´s, INRA, Bures-sur-Yvette, France Received 22 November 2004; received in revised form 15 April 2005; accepted 18 April 2005 Abstract Protease inhibitors (PIs) have been shown to cause lethal and sublethal effects on aphids depending on the kind of PI and aphid species. Therefore, these proteins might affect aphid parasitoids directly by inhibiting their digestive proteolysis or indirectly via their development in a less suitable host. In our study, the risk of exposure and the potential effects of soybean Bowman-Birk inhibitor (SbBBI) and oryzacystatin I (OCI) on the aphid endoparasitoid Aphidius ervi were investigated using artificial diet to deliver PIs. Immunoassays showed that both SbBBI and OCI were detected in the honeydew of aphids reared on artificial diet containing these recombinant proteins at 100 mg/mL. However, only SbBBI was detected in parasitoid larvae, while this PI could not be detected in adult parasitoids emerged from PI-intoxicated aphids. Enzymatic inhibition assays showed that digestive proteolytic activity of larvae and adults of A. ervi predominantly relies on serine proteases and especially on chymotrypsin-like activity. Bioassays using SbBBI and OCI on artificial diet were performed. A. ervi that developed on intoxicated aphids had impaired fitness. Thus development and parasitism success of parasitoids exposed to OCI were severely affected. On the contrary, SbBBI only altered significantly female size and sex ratio. Direct exposure to PIs through adult food intake did not affect female’s longevity, while SbBBI and OCI (100 mg/mL) induced 69% and 30% inhibition of digestive protease activity, respectively. These studies made it possible to estimate the risk of exposure to plant PIs and the sensitivity of the aphid parasitoid A. ervi to these entomotoxins, by combining immunological, biochemical and biological approaches. First it pointed out that only immature stages are affected by PIs. Secondly, it documented two different modes of effect, according to the nature of the PIs and both host and parasitoid susceptibility. OCI prevented the development of A. ervi mainly due to the host susceptibility, whereas SbBBI only induced sublethal effects on the parasitoid, possibly due to both direct action on the parasitoid susceptible proteases, and host-mediated action through size reduction. r 2005 Elsevier Ltd. All rights reserved. Keywords: Aphid parasitoid; Protease inhibitors; Tritrophic interactions; Risk assessment; Transgenic plants 1. Introduction Protease inhibitors (PIs) from plants exhibiting insecticide effects emerged as an interesting strategy for insect pests control (Jouanin et al., 1998; Carlini and Grossi-de-Sa, 2002; Lawrence and Koundal, 2002; Ranjekar et al., 2003). Several serine and cysteine PIs ARTICLE IN PRESS www.elsevier.com/locate/jinsphys 0022-1910/$ - see front matter r 2005 Elsevier Ltd. All rights reserved. doi:10.1016/j.jinsphys.2005.04.009 Corresponding author. BPCIR, EA 3900 Universite´ de Picardie Jules Verne, 33 rue Saint Leu, 80039 Amiens Cedex 1, France. Tel.: +33 322 82 7547; fax: +33-322-82-7547. E-mail address: [email protected] (H. Azzouz). 1 Present address : De´ veloppement, Evolution et Plasticite´ du Syste`me Nerveux, IAF, CNRS, 91198 Gif/Yvette Cedex.
-
Upload
independent -
Category
Documents
-
view
5 -
download
0
Transcript of Potential effects of plant protease inhibitors, oryzacystatin I and soybean Bowman-Birk inhibitor,...

ARTICLE IN PRESS
Journal of Insect Physiology 51 (2005) 941–951
0022-1910/$ - se
doi:10.1016/j.jin
�CorrespondJules Verne, 33
+33 322 82 754
E-mail addr1Present add
Systeme Nerveu
www.elsevier.com/locate/jinsphys
Potential effects of plant protease inhibitors, oryzacystatin I andsoybean Bowman-Birk inhibitor, on the aphid parasitoid Aphidius
ervi Haliday (Hymenoptera, Braconidae)
H. Azzouza,c,�, E.D.M. Campana, A. Cherquia, J. Sagueza, A. Coutya, L. Jouaninb,P. Giordanengoa, L. Kaiserc,1
aBPCIR, EA 3900 Universite de Picardie Jules Verne, 33 rue Saint Leu, 80039 Amiens Cedex 1, FrancebINRA-CNRS, Laboratoire de Biologie Cellulaire, route de St Cyr, 78026 Versailles Cedex, France
cLaboratoire de Neurobiologie Comparee des Invertebres, INRA, Bures-sur-Yvette, France
Received 22 November 2004; received in revised form 15 April 2005; accepted 18 April 2005
Abstract
Protease inhibitors (PIs) have been shown to cause lethal and sublethal effects on aphids depending on the kind of PI and aphid
species. Therefore, these proteins might affect aphid parasitoids directly by inhibiting their digestive proteolysis or indirectly via
their development in a less suitable host. In our study, the risk of exposure and the potential effects of soybean Bowman-Birk
inhibitor (SbBBI) and oryzacystatin I (OCI) on the aphid endoparasitoid Aphidius ervi were investigated using artificial diet to
deliver PIs. Immunoassays showed that both SbBBI and OCI were detected in the honeydew of aphids reared on artificial diet
containing these recombinant proteins at 100 mg/mL. However, only SbBBI was detected in parasitoid larvae, while this PI could notbe detected in adult parasitoids emerged from PI-intoxicated aphids. Enzymatic inhibition assays showed that digestive proteolytic
activity of larvae and adults of A. ervi predominantly relies on serine proteases and especially on chymotrypsin-like activity.
Bioassays using SbBBI and OCI on artificial diet were performed. A. ervi that developed on intoxicated aphids had impaired
fitness. Thus development and parasitism success of parasitoids exposed to OCI were severely affected. On the contrary, SbBBI only
altered significantly female size and sex ratio. Direct exposure to PIs through adult food intake did not affect female’s longevity,
while SbBBI and OCI (100mg/mL) induced 69% and 30% inhibition of digestive protease activity, respectively. These studies made
it possible to estimate the risk of exposure to plant PIs and the sensitivity of the aphid parasitoid A. ervi to these entomotoxins, by
combining immunological, biochemical and biological approaches. First it pointed out that only immature stages are affected by
PIs. Secondly, it documented two different modes of effect, according to the nature of the PIs and both host and parasitoid
susceptibility. OCI prevented the development of A. ervi mainly due to the host susceptibility, whereas SbBBI only induced sublethal
effects on the parasitoid, possibly due to both direct action on the parasitoid susceptible proteases, and host-mediated action
through size reduction.
r 2005 Elsevier Ltd. All rights reserved.
Keywords: Aphid parasitoid; Protease inhibitors; Tritrophic interactions; Risk assessment; Transgenic plants
e front matter r 2005 Elsevier Ltd. All rights reserved.
sphys.2005.04.009
ing author. BPCIR, EA 3900 Universite de Picardie
rue Saint Leu, 80039 Amiens Cedex 1, France. Tel.:
7; fax: +33-322-82-7547.
ess: [email protected] (H. Azzouz).
ress : Developpement, Evolution et Plasticite du
x, IAF, CNRS, 91198 Gif/Yvette Cedex.
1. Introduction
Protease inhibitors (PIs) from plants exhibitinginsecticide effects emerged as an interesting strategyfor insect pests control (Jouanin et al., 1998; Carlini andGrossi-de-Sa, 2002; Lawrence and Koundal, 2002;Ranjekar et al., 2003). Several serine and cysteine PIs

ARTICLE IN PRESSH. Azzouz et al. / Journal of Insect Physiology 51 (2005) 941–951942
have been expressed in transgenic plants belonging todifferent families to enhance their resistance againstLepidoptera (Hilder et al., 1987; De Leo et al., 2001;Falco and Silva-Filho, 2003) and Coleoptera (Lecar-donnel et al., 1999; Alfonso-Rubı et al., 2003). Theinsecticidal effects of PIs against sap-sucking phytopha-gous insects was established recently (Annadana et al.,2002; Ceci et al., 2003; Rahbe et al., 2003a; Rahbe et al.,2003b).Aphicidal effects of PIs, especially of some serine and
cysteine PIs, have been established by diet incorporationassays. The pea and soybean trypsin–chymotrypsininhibitors (PsTI-2, SbBBI) belonging to the Bowman-Birk family (Rahbe et al., 2003b) and the mustard-typetrypsin–chymotrypsin variant Chy8 (Ceci et al., 2003)induced significant mortality and growth inhibition ofthe pea aphid Acyrthosiphon pisum. In addition, SbBBIhad deleterious effect on the life history parameters ofthe adult potato aphid Macrosiphum euphorbiae (Az-zouz et al., 2005). The phytocystatin oryzacystatin I(OCI) isolated from rice seeds (Abe et al., 1987)significantly reduced the nymphal survival of M.
euphorbiae and prevented aphids from reproducing(Azzouz et al., 2005). In addition, Rahbe et al. (2003a)showed that OCI significantly reduced adult weight andfecundity of the aphid Myzus persicae. Genes encodingPIs are therefore regarded as candidates for expressionin transgenic crops and appear a promising strategy toconfer resistance against homopteran pests and espe-cially aphids (Deraison, 2002; Ceci et al., 2003; Rahbeet al., 2003b; Azzouz et al., 2005). Except for Btendotoxin expressing plants (Rice and Pilcher, 1998;Kumar and Kumar, 2004), target insects are not usuallycompletely controlled by transgenic plants (Stoger et al.,1999). Consequently aphid limitation by sublethaleffects of plant expressing PIs could be complementedby the use of beneficial insects (van Emden, 1999). Thus,it is important to investigate the potential negative sideeffects of this new technology on non-target insects suchas predators and parasitoids.Several studies investigated the effects of PIs on insect
predators (Ashouri et al., 1998; Overney et al., 1998;Walker et al., 1998; Bouchard et al., 2003; Ferry et al.,2003) and parasitoids (Down et al., 1999; Ashouri et al.,2001; Bell et al., 2001; Schuler et al., 2001; Cowgill et al.,2004). They reported that the major digestive proteolyticactivity of adult and larvae ladybirds Adalia bipunctata
(Walker et al., 1998) and Harmonia axyridis (Ferryet al., 2003), and the stinkbug Perillus bioculatus
(Overney et al., 1998) was cysteine protease based. It isonly known that OCI reduced the fitness of P. bioculatus
(Ashouri et al., 1998) but did not affect H. axyridis
(Ferry et al., 2003). Regarding parasitoids, only thedigestive protease activities in the larval stage ofectoparasitoid Eulophus pennicornis (Down et al.,1999) and the larval and adult stage of endoparasitoid
Aphelinus abdominalis (Azzouz et al., 2005) werereported. No deleterious effect of OCI was found inthe aphid parasitoids Aphidius spp. (Ashouri et al., 2001;Cowgill et al., 2004) and Diaeretiella rapae (Schuleret al., 2001) parasitising aphids reared on OCI-expres-sing plants. However, in laboratory bioassay using M.
euphorbiae reared on artificial diets supplemented withOCI and SbBBI, we demonstrated that developmentand parasitism success of A. abdominalis were adverselyaffected (Azzouz et al., 2005).The objective of this study was to characterise and
analyse the effects of oryzacystatin I (OCI) and soybeanBowman-Birk inhibitor (SbBBI) on the aphid parasitoidAphidius ervi Haliday (Hymenoptera: Braconidae) incontrolled exposure conditions. A. ervi is a solitaryendoparasitoid commercialised to control several aphidspecies in greenhouses. Risks of exposure of A. ervi toOCI and SbBBI were investigated first. Secondly, thedigestive protease activities of larval and adult stageswere determined. Finally, effects of PIs enrichedartificial diets were investigated on parasitoid develop-ment and parasitism success. Effect of PIs on A. ervi
adult survival was also studied.
2. Materials and methods
2.1. Reagents
Trans-epoxysuccinyl-L-leucylamido-(4-guanidino)-butane(E-64) and soybean trypsin–chymotrypsin Bowman-Birk Inhibitor (SbBBI) (495% of purity) were obtainedfrom Sigma. Pepstatin A and chymostatin were suppliedby Alexis biochemicals. Ethylenediamine Tetra-AceticAcid (EDTA) was obtained from Prolabo and pefablockfrom Interchim. The EnzChekR Protease Assay Kit(fluorescent protein BODIPYR-FL casein) was pur-chased from Molecular Probes. Oryzacystatin I (OCI)(495% of purity) was obtained as previously describedby Leple et al. (1995). The polyclonal anti-OCI (Leple etal., 1995) or anti-SbBBI (Bonade-Bottino, unpublishedresults) were raised in rabbits and used as primaryantibodies. The secondary antibody peroxidase-coupledgoat anti-rabbit IgG (A 0545) was purchased from Sigma.ECL kit and nitrocellulose membrane HybondTM–ECLTM were obtained from Amersham Biosciences.
2.2. Artificial diets
As a control and a basis for the protein dilution, astandard diet adapted for M. euphorbiae was preparedas described by Febvay et al. (1988) and modified byDown et al. (1996). OCI or SbBBI were incorporated tothe standard diet to obtain the following diets: OCI-100,OCI-250 contained 100 and 250 mg/mL of OCI, respec-tively; SbBBI-100 and SbBBI-500, contained 100 and

ARTICLE IN PRESSH. Azzouz et al. / Journal of Insect Physiology 51 (2005) 941–951 943
500 mg/mL of SbBBI, respectively. The diet sachets wereprepared as described in Couty et al. (2001b) underaseptic conditions, sterilised by filtration through0.45 mm filters (Millipore Corp., Bedford, MA) thenkept frozen at �20 1C until used.To expose adult parasitoids to PIs through food
intake, sugar solution of 70% (w/v) 1:1 glucose–fructosemixture was used as a sugar source for A. ervi females.Twenty mg/mL of Bovine Serum Albumin (BSA) wasadded to this sugar solution to stimulate the proteaseactivity of this parasitoid. This control solution was thensupplemented with BSA, OCI or SbBBI at 100 mg/mL(BSA100, OCI100 and SbBBI100, respectively).
2.3. Insects
The LB05 Sm clone of the potato aphid M. euphorbiae
provided by Yvan Rahbe (INRA-INSA Villeurbanne,France) was reared in the laboratory on potato plants(Solanum tuberosum. cv Desiree) in a controlled environ-ment room (T ¼ 20� 1 1C; RH ¼ 6075%;16L/8D).Neonate nymphs (12h old) used in the experiments wereissued from females synchronised on an excised potatoleaf isolated in a plastic box.Mummies of A. ervi purchased from Biobest (Bel-
gium) were placed in plastic boxes (17� 11� 7 cm) untiladult emergence under the same conditions as describedabove. Emerging male and female parasitoids were kepttogether and fed ad libitum on cottonwool imbibed withsugar solution (glucose and fructose 70% (w/v)) during48 h before infesting aphids.
2.4. Insect bioassays
2.4.1. Effects of protease inhibitors on M. euphorbiae size
In order to assess the effect of PIs on M. euphorbiae
size, neonate aphids were fed for four days on OCI-100,SbBBI-100 and control diets. Thirty five-days-oldnymphs were individually measured from the tip ofthe head to the base of the cauda (length) and acrossthe metathorax (width), under a microscope usingan eyepiece graticule. Experiment was conducted ina controlled environment room (T ¼ 20� 1 1C; RH ¼
6075%;16L/8D).The expressed level of PIs in plant foliage is from
0.05% to 2.5% of total soluble proteins (Leple et al.,1995; De Leo et al., 1998; Charity et al., 1999; Lee et al.,1999). The level of total soluble proteins in the phloem isless than 5mg/mL in non-cucurbit species (Fisher et al.,1992; van Helden et al., 1994; Ishiwatari et al., 1995;Schobert et al., 1998; Foissac et al., 2002). According toRahbe et al. (2003a) the amounts expressed into leaftissue seems to be close to the amount present inthe phloem sap. Therefore we chose to deliver toaphids a PI concentration of 100 mg/mL of protein,which is consistent with our knowledge of PI expression
in transgenic plants and protein content in plantphloem sap.
2.4.2. Effects of protease inhibitors on A. ervi
development
To study the effect of PIs on A. ervi development,neonate aphids were fed for four days on OCI-100,SbBBI-100 and control diets. Host infestation wasperformed by placing a single mated A. ervi female ina cylinder of artificial diet with 25 five-days-old nymphsfor 4 h, then aphids were kept on their respective dietuntil mummification. Mummies were collected on theday of formation and isolated into gelatin capsules untiladult emergence. Fifteen replicates were set up for eachtreatment.We recorded the percentage of mummification (i.e.
the ratio between the number of mummies formed andthe number of aphid nymphs submitted to parasitisa-tion) and of emergence (i.e. the ratio between thenumber of emerged parasitoids and the number ofmummies formed) were calculated. Larval and nymphaldevelopment time were defined as the period from hostinfestation to mummification, and from mummificationto adult emergence, respectively. Newly emerged adultparasitoids were sexed. They were then isolated in Petridishes (5.5 cm diameter) provided with a filter paperimbibed with water until death to evaluate theirresistance to starvation. The tibia length was measuredunder a binocular microscope to estimate the adult size.Experiments were carried out under the same conditionsas described above.
2.4.3. Effects of protease inhibitors on A. ervi adult
survival
Seventeen one day-old females were placed inventilated plastic boxes (17� 11� 7 cm) and allowedto feed for 4 h daily on a cotton wool imbibed withsugar-control or BSA100 or OCI100 or SbBBI100. Deadinsects were counted and removed every day. Threereplicates were carried out under controlled conditions(T ¼ 20� 0:5 1C; RH ¼ 6075%; 16L/8D).
2.5. Determination of A. ervi digestive protease activity
2.5.1. Enzyme preparation
Four day-old aphids issued from synchronisedfemales were placed on cut potato leaves and parasitisedby A. ervi females. Third instar parasitoid larvae wereextracted by dissecting aphids in 0.15M NaCl under abinocular microscope at 4 1C. Ten whole parasitoidlarvae or 10 dissected larval or adult guts were placed in30 mL 0.15M NaCl buffer. Each sample was immedi-ately homogenised on ice then centrifuged at 5000g at4 1C for 15min. Supernatants were aliquoted and storedat �20 1C. Adult guts were dissected out of cold-anaesthetised females and extracts made according to

ARTICLE IN PRESSH. Azzouz et al. / Journal of Insect Physiology 51 (2005) 941–951944
the above procedure. Total protein content of parasitoidextracts was determined by the Bradford (1976) methodusing BSA (1 mg/mL) as standard protein.
2.5.2. Electrophoresis assay
Whole larvae, larval or adult gut extracts were diluted2-fold in Laemmli buffer containing 2% (v/v) b-mercaptoethanol. The samples were separated in a10% sodium dodecyl sulphate polyacrylamide gel(SDS-PAGE) copolymerised with 0.05% gelatin. Afterseparation at 100V at 4 1C, the gel was washed in 2.5%Triton X-100 for 1 h and rinsed twice with distilledwater. Subsequently the gel was incubated overnight at37 1C in 50mM Tris–HCl buffer pH 8 containing150mM NaCl, 5mM CaCl2 and 2mM DTT thenstained with Coomassie R-250 brilliant blue solution.Indeed, preliminary experiments (data not shown)revealed that the optimal pH range for the proteaseactivity was 8–9 for the larvae and 7–8 for the adults.
2.5.3. Inhibition assays
Electrophoresis assay showed that whole larva andlarval gut extracts presented the same proteolytic profile(Fig. 1). Thus, protease activity of both A. ervi adultguts and whole larvae was quantified using thefluorescent protein casein as substrate, at a finalconcentration of 0.15 mg/mL. Inhibition assays werecarried out in 0.1M Tris–HCl pH 8. An equivalent of1.5 larvae or 5 adult guts per well was incubated for15min at ambient temperature with inhibitors, beforeadding the casein substrate. Fluorescence was measuredevery 2min during 60min at 37 1C on a LabSystems
Fig. 1. Zymogram of the gelatinolytic activity of whole larvae, larval
and adult gut extracts from Aphidius ervi (6 mg of protein of each
sample was submitted to electrophoresis).
Fluoroskan II fluorimeter at 538 nm after an excitationat 485 nm. The class of chemical and protein inhibitors,and final assay concentrations tested are presented inTable 4.All assays were performed in triplicate for larvae and
duplicate for adults, with appropriate controls. Theinhibition of the proteolytic activity was expressedrelatively to the control. Inhibitors were dissolved inmilli-Q water (E-64, pefablock, EDTA, SbBBI and OCI)or DMSO (pepstatin A and chymostatin).
2.6. Transfer of protease inhibitors from artificial diet to
parasitoids and aphid honeydew
2.6.1. Aphids and parasitoids on artificial diet
Aphids were reared on the following diets OCI-100,OCI-250, SbBBI-100, SbBBI-500 and control. On thefifth day, aphids were parasitised as described before. Atthe beginning of the aphid cuticle melanisation (corre-sponding to the 3rd instar of the parasitoid larva), agroup of aphids were dissected to collect parasitoidlarvae. The remaining aphids were kept on theirrespective diet until mummification. Mummies werecollected on the day of formation and placed in a Petridish until adult parasitoid emergence.
2.6.2. Honeydew collection
Nitrocellulose membrane was placed during 24 hunder plastic cylinder containing 20 aphids fed duringthree days with OCI-100, SbBBI-100 or control. Thesame number of aphids was kept in each plastic cylinder(two repetitions per diet). Nitrocellulose membraneswere then placed between two blotting papers and keptat 4 1C until use.
2.6.3. Immunoassay blot
Detection of OCI and SbBBI in protein extracts fromparasitoids was performed by western analysis usinganti-OCI and anti-SbBBI antibodies, respectively. Pro-tein samples were extracted by homogenising parasitoids(larvae and adults) in 50mM Tris–HCl, (pH 6.8)containing 1% SDS (3 mL of buffer per individual).Laemmli buffer containing 2% b-mercaptoethanol wasthen added and the samples were heated at 95 1C during5min. Samples were centrifuged for 15min at 5000g at4 1C. The supernatants were electrophoresed in 10%SDS–PAGE then blotted onto nitrocellulose membraneaccording to Towbin et al. (1979). OCI and SbBBIdiluted in sample buffer were used as controls. Afterelectrotransfer at 100mA, the membrane was blockedfor 1 h at room temperature with a solution of 5% (w/v)skimmed milk in TBS (10mM Tris–HCl pH 7.5, 0.1MNaCl) containing 0.25% (v/v) Tween-20. OCI or SbBBIantibodies (1:500) and the peroxidase-coupled goat anti-rabbit IgG (1:50,000) were used to hybridise themembrane. Immunostained bands were detected by
ARTICLE IN PRESSH. Azzouz et al. / Journal of Insect Physiology 51 (2005) 941–951 945
chemiluminescence (ECL) according to the manufac-turer’s instructions. The nitrocellulose membrane con-taining honeydew droplets excreted by aphids weretreated as described before excepted that the secondaryantibody concentration used was 1:1000. In our condi-tions the detection limit of the western blots was 10 ngfor SbBBI and 0.1 ng for OCI.
2.7. Statistical analysis
Statistical analyses were done using STATISTICA 5.5software (StatSofts, Tulsa, Oklahoma, USA). Theeffects of PIs on M. euphorbiae size and parasitismsuccess of A. ervi were analysed using Kruskal-Wallis-ANOVA. When a significant effect was found theMann-Whitney-U test was performed to compare themeans. The significance level ða0Þ of the Mann-Whitney-U test was corrected according to the number ofcomparisons, using the Bonferroni adjustment: a0 ¼a=k with a ¼ 5% nd k ¼ number of comparisons. Theeffects of PIs on A. ervi development were analysedusing Mann-Whitney-U test. The results are reported asmean7standard error (SE).
3. Results
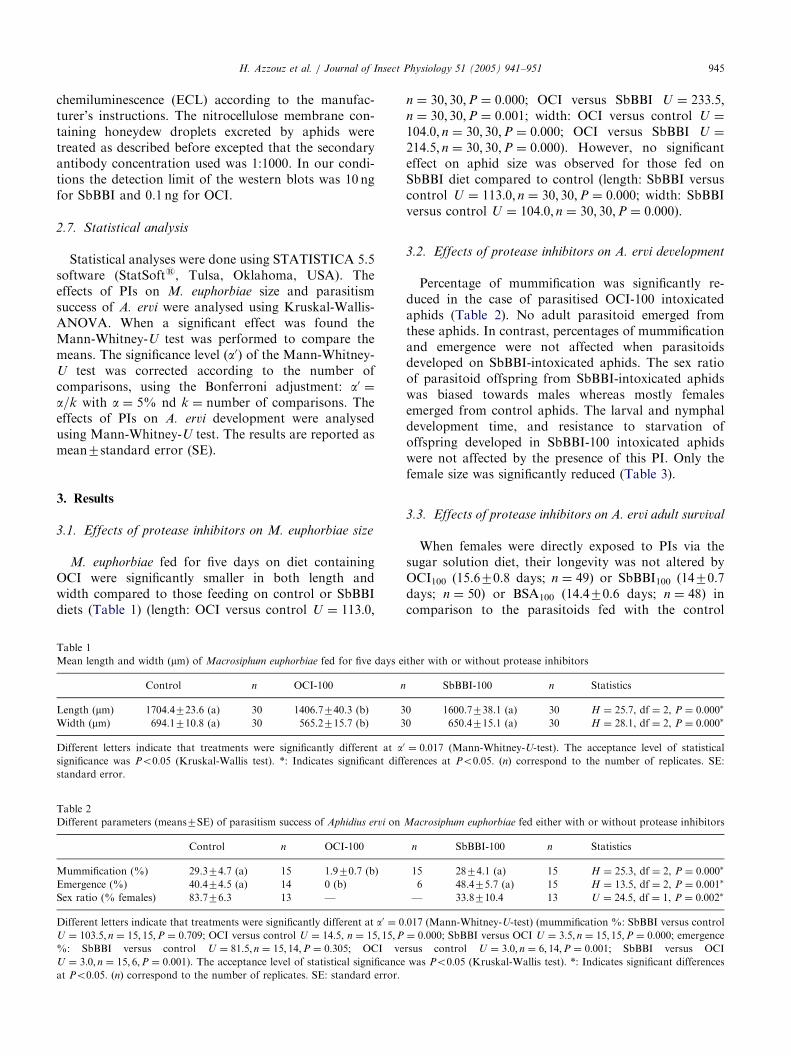
3.1. Effects of protease inhibitors on M. euphorbiae size
M. euphorbiae fed for five days on diet containingOCI were significantly smaller in both length andwidth compared to those feeding on control or SbBBIdiets (Table 1) (length: OCI versus control U ¼ 113:0;
Table 1
Mean length and width (mm) of Macrosiphum euphorbiae fed for five days e
Control n OCI-100 n
Length (mm) 1704.4723.6 (a) 30 1406.7740.3 (b) 3
Width (mm) 694.1710.8 (a) 30 565.2715.7 (b) 3
Different letters indicate that treatments were significantly different at a0
significance was Po0:05 (Kruskal-Wallis test). *: Indicates significant diff
standard error.
Table 2
Different parameters (means7SE) of parasitism success of Aphidius ervi on
Control n OCI-100
Mummification (%) 29.374.7 (a) 15 1.970.7 (b)
Emergence (%) 40.474.5 (a) 14 0 (b)
Sex ratio (% females) 83.776.3 13 —
Different letters indicate that treatments were significantly different at a0 ¼ 0
U ¼ 103:5; n ¼ 15; 15;P ¼ 0:709; OCI versus control U ¼ 14:5, n ¼ 15; 15;P%: SbBBI versus control U ¼ 81:5; n ¼ 15; 14;P ¼ 0:305; OCI ve
U ¼ 3:0; n ¼ 15; 6;P ¼ 0:001). The acceptance level of statistical significanceat Po0:05. ðnÞ correspond to the number of replicates. SE: standard error.
n ¼ 30; 30;P ¼ 0:000; OCI versus SbBBI U ¼ 233:5;n ¼ 30; 30;P ¼ 0:001; width: OCI versus control U ¼
104:0; n ¼ 30; 30;P ¼ 0:000; OCI versus SbBBI U ¼
214:5; n ¼ 30; 30;P ¼ 0:000). However, no significanteffect on aphid size was observed for those fed onSbBBI diet compared to control (length: SbBBI versuscontrol U ¼ 113:0; n ¼ 30; 30;P ¼ 0:000; width: SbBBIversus control U ¼ 104:0; n ¼ 30; 30;P ¼ 0:000).
3.2. Effects of protease inhibitors on A. ervi development
Percentage of mummification was significantly re-duced in the case of parasitised OCI-100 intoxicatedaphids (Table 2). No adult parasitoid emerged fromthese aphids. In contrast, percentages of mummificationand emergence were not affected when parasitoidsdeveloped on SbBBI-intoxicated aphids. The sex ratioof parasitoid offspring from SbBBI-intoxicated aphidswas biased towards males whereas mostly femalesemerged from control aphids. The larval and nymphaldevelopment time, and resistance to starvation ofoffspring developed in SbBBI-100 intoxicated aphidswere not affected by the presence of this PI. Only thefemale size was significantly reduced (Table 3).
3.3. Effects of protease inhibitors on A. ervi adult survival
When females were directly exposed to PIs via thesugar solution diet, their longevity was not altered byOCI100 (15.670.8 days; n ¼ 49) or SbBBI100 (1470.7days; n ¼ 50) or BSA100 (14.470.6 days; n ¼ 48) incomparison to the parasitoids fed with the control
ither with or without protease inhibitors
SbBBI-100 n Statistics
0 1600.7738.1 (a) 30 H ¼ 25:7, df ¼ 2, P ¼ 0:000�
0 650.4715.1 (a) 30 H ¼ 28:1, df ¼ 2, P ¼ 0:000�
¼ 0:017 (Mann-Whitney-U-test). The acceptance level of statistical
erences at Po0:05. ðnÞ correspond to the number of replicates. SE:
Macrosiphum euphorbiae fed either with or without protease inhibitors
n SbBBI-100 n Statistics
15 2874.1 (a) 15 H ¼ 25:3, df ¼ 2, P ¼ 0:000�
6 48.475.7 (a) 15 H ¼ 13:5, df ¼ 2, P ¼ 0:001�
— 33.8710.4 13 U ¼ 24:5, df ¼ 1, P ¼ 0:002�
:017 (Mann-Whitney-U-test) (mummification %: SbBBI versus control
¼ 0:000; SbBBI versus OCI U ¼ 3:5; n ¼ 15; 15;P ¼ 0:000; emergencersus control U ¼ 3:0; n ¼ 6; 14;P ¼ 0:001; SbBBI versus OCI
was Po0:05 (Kruskal-Wallis test). *: Indicates significant differences
ARTICLE IN PRESS
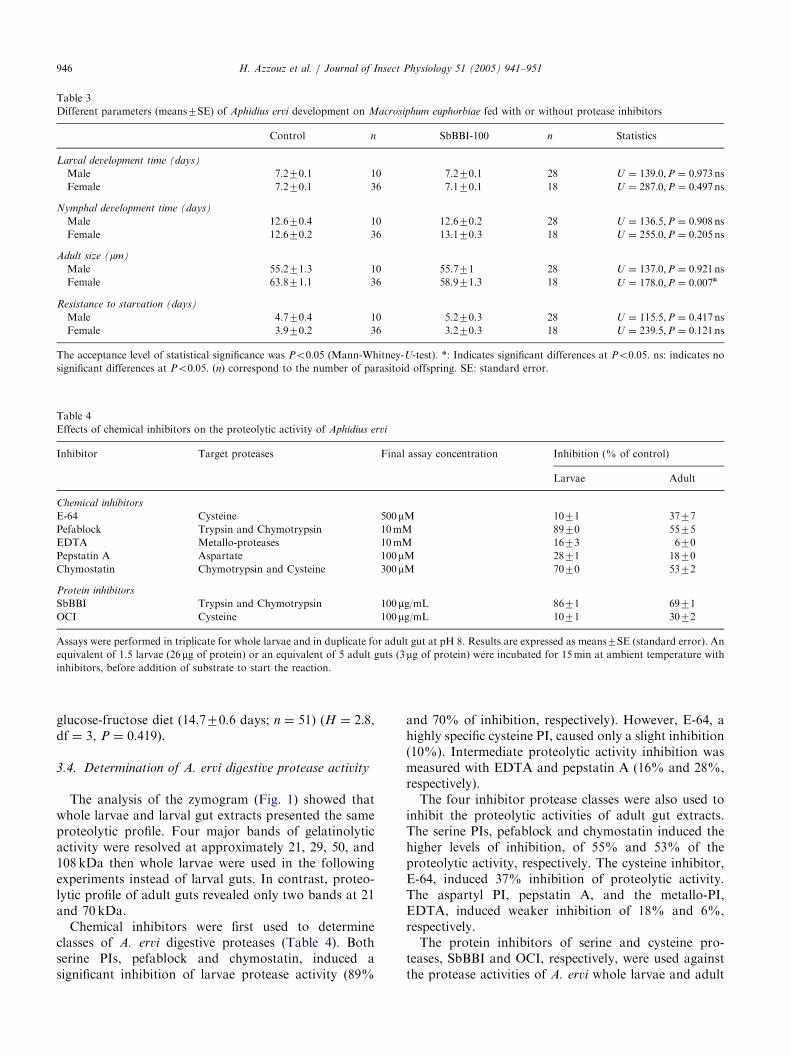
Table 3
Different parameters (means7SE) of Aphidius ervi development on Macrosiphum euphorbiae fed with or without protease inhibitors
Control n SbBBI-100 n Statistics
Larval development time (days)
Male 7.270.1 10 7.270.1 28 U ¼ 139:0;P ¼ 0:973nsFemale 7.270.1 36 7.170.1 18 U ¼ 287:0;P ¼ 0:497ns
Nymphal development time (days)
Male 12.670.4 10 12.670.2 28 U ¼ 136:5;P ¼ 0:908nsFemale 12.670.2 36 13.170.3 18 U ¼ 255:0;P ¼ 0:205ns
Adult size (mm)
Male 55.271.3 10 55.771 28 U ¼ 137:0;P ¼ 0:921nsFemale 63.871.1 36 58.971.3 18 U ¼ 178:0;P ¼ 0:007n
Resistance to starvation (days)
Male 4.770.4 10 5.270.3 28 U ¼ 115:5;P ¼ 0:417nsFemale 3.970.2 36 3.270.3 18 U ¼ 239:5;P ¼ 0:121ns
The acceptance level of statistical significance was Po0:05 (Mann-Whitney-U-test). *: Indicates significant differences at Po0:05. ns: indicates nosignificant differences at Po0:05. ðnÞ correspond to the number of parasitoid offspring. SE: standard error.
Table 4
Effects of chemical inhibitors on the proteolytic activity of Aphidius ervi
Inhibitor Target proteases Final assay concentration Inhibition (% of control)
Larvae Adult
Chemical inhibitors
E-64 Cysteine 500mM 1071 3777
Pefablock Trypsin and Chymotrypsin 10mM 8970 5575
EDTA Metallo-proteases 10mM 1673 670
Pepstatin A Aspartate 100mM 2871 1870
Chymostatin Chymotrypsin and Cysteine 300mM 7070 5372
Protein inhibitors
SbBBI Trypsin and Chymotrypsin 100mg/mL 8671 6971
OCI Cysteine 100mg/mL 1071 3072
Assays were performed in triplicate for whole larvae and in duplicate for adult gut at pH 8. Results are expressed as means7SE (standard error). An
equivalent of 1.5 larvae (26mg of protein) or an equivalent of 5 adult guts (3mg of protein) were incubated for 15min at ambient temperature with
inhibitors, before addition of substrate to start the reaction.
H. Azzouz et al. / Journal of Insect Physiology 51 (2005) 941–951946
glucose-fructose diet (14.770.6 days; n ¼ 51) (H ¼ 2:8,df ¼ 3, P ¼ 0:419).
3.4. Determination of A. ervi digestive protease activity
The analysis of the zymogram (Fig. 1) showed thatwhole larvae and larval gut extracts presented the sameproteolytic profile. Four major bands of gelatinolyticactivity were resolved at approximately 21, 29, 50, and108 kDa then whole larvae were used in the followingexperiments instead of larval guts. In contrast, proteo-lytic profile of adult guts revealed only two bands at 21and 70 kDa.Chemical inhibitors were first used to determine
classes of A. ervi digestive proteases (Table 4). Bothserine PIs, pefablock and chymostatin, induced asignificant inhibition of larvae protease activity (89%
and 70% of inhibition, respectively). However, E-64, ahighly specific cysteine PI, caused only a slight inhibition(10%). Intermediate proteolytic activity inhibition wasmeasured with EDTA and pepstatin A (16% and 28%,respectively).The four inhibitor protease classes were also used to
inhibit the proteolytic activities of adult gut extracts.The serine PIs, pefablock and chymostatin induced thehigher levels of inhibition, of 55% and 53% of theproteolytic activity, respectively. The cysteine inhibitor,E-64, induced 37% inhibition of proteolytic activity.The aspartyl PI, pepstatin A, and the metallo-PI,EDTA, induced weaker inhibition of 18% and 6%,respectively.The protein inhibitors of serine and cysteine pro-
teases, SbBBI and OCI, respectively, were used againstthe protease activities of A. ervi whole larvae and adult

ARTICLE IN PRESS
Fig. 2. (A) Western blot analysis of parasitoid developed in aphids fed with SbBBI diets. 100 ng of SbBBI (lane 1). Parasitoid larvae developed in
aphids reared on control diet (lane 2), on SbBBI-100 diet (lane 4) and on SbBBI-500 diet (lane 6). Parasitoid adults emerged from aphids reared on
control diet (lane 3), on SbBBI-100 diet (lane 5) and on SbBBI-500 diet (lane 7). 50mg of protein of larvae and adults parasitoid extracts were
submitted to electrophoresis under reducing conditions. (B) Western blot analysis of parasitoid developed in aphids fed with OCI diets. 10 ng of OCI
(lane 1). Parasitoid larvae developed in aphids reared on control diet (lane 2), on OCI-100 diet (lane 4) and on OCI-250 diet (lane 5). Parasitoid adults
emerged from aphids reared on control diet (lane 3). 50mg of protein of larvae and adults parasitoid extracts were submitted to electrophoresis underreducing conditions.
Fig. 3. Dot blot analysis of honeydew excreted by aphids fed with or
without protease inhibitors.
H. Azzouz et al. / Journal of Insect Physiology 51 (2005) 941–951 947
gut extracts. The differences of inhibition levels matchedthose observed with the corresponding chemical inhibi-tors. Indeed, the assays performed with these PIsshowed that SbBBI induced a much higher inhibitoryeffect compared to OCI. The trypsin–chymotrypsininhibitor SbBBI (100 mg/mL) inhibited 86% and 69%of larvae and adult protease activities, respectively. Thecysteine PI OCI (100 mg/mL), presented only 10% and30% inhibition against whole larvae and adult gutdigestive protease extracts, respectively.
3.5. Detection of protease inhibitors by immunoassays
analysis
Western blot analysis showed that the primaryantibodies recognised SbBBI (0.1mg) and recombinantEscherichia coli OCI (0.01mg) (Fig. 2A and B lane 1). Asexpected, in extracts from parasitoids developed in aphidsfed with control diet no hybridised band could beobserved (lanes 2 and 3). Extracts of parasitoid larvaedissected out from aphids fed with SbBBI-100 and SbBBI-500 showed the presence of this PI (8 kDa, Fig. 2A lanes 4and 6). Whereas, no hybridisation occurred in the adultextracts (lanes 5 and 7). When host aphids were fed withOCI diets, western blots did not reveal any staining inextracts of parasitoid larvae (Fig. 2B). No adult emergedfrom OCI-intoxicated aphids.Dot blot analysis showed the presence of SbBBI and
OCI in the honeydew produced by aphids fed withSbBBI-100 and OCI-100 diets (Fig. 3).
4. Discussion
Our study demonstrated that SbBBI had a sublethaleffect on A. ervi development and offspring (sex ratio,female size), whereas oryzacystatin I OCI preventedparasitoid development (mortality). However, the directexposure to either PI during adult food intake did notaffect female’s survival.
The comparison of the larval and adult proteaseprofiles showed that four different gelatin hydrolysisbands were observed in the extracts of larval gut, andonly two gelatinolysis band in adult gut. The parasitoidlarvae have a larger proteolytic activity spectrumcompared to adults. This difference corresponds tocontrasted lifestyles of both stages of A. ervi. The larvadevelops entirely into the aphid body, consuming hostfluids and tissues exclusively (de Eguileor et al., 2001),whereas the adult feeds only on carbohydrate-richsources like floral and extrafloral nectar and honeydew(Hagvar and Hofsvang, 1991). As the result of differentphysiological needs and ecology between larvae andadults, the risk and the consequence of exposure to PIsare different.SbBBI was detected in parasitoid larvae developed on
M. euphorbiae reared on artificial diet containing thisentomotoxin, and in honeydew. Therefore, A. ervi wasexposed to this serine PI during both larval developmentand adult stage. Nevertheless, this PI was not revealed inadult parasitoids. SbBBI ingested by parasitoid larvaecould have been metabolised due to high proteolyticactivity known in larval instars of endoparasitoids.Alternatively, SbBBI could have been excreted in theform of meconial pellets just prior to pupation. This factwas reported for A. ervi developed on M. persicae rearedon lectin-enriched artificial diet (Couty et al., 2001b).The study of SbBBI direct effect on A. ervi showed
that the protease activities revealed in the whole larvae

ARTICLE IN PRESSH. Azzouz et al. / Journal of Insect Physiology 51 (2005) 941–951948
or adult gut extracts were highly inhibited by this PI. A.
ervi proteases were also inhibited by the chymostatin,which is a common inhibitor of both chymotrypsin andcysteine proteases. Nevertheless, E-64 did not inhibit thedigestive proteolytic activity, suggesting that the majorA. ervi proteases belong to the serine protease class andare chymotrypsin-like. Such serine protease-based di-gestive metabolism was previously shown in the aphidparasitoid A. abdominalis (Azzouz et al., 2005), in thelarval stage of the eulophid ectoparasitoid Eulophus
pennicornis (Down et al., 1999) and in the honeybee(Burgess et al., 1996). Our results about digestiveproteases of the parasitoid and their sensitivity toSbBBI indicate a potential risk of toxicity of SbBBI atthe 100 mg/mL concentration (0.1% of total solubleproteins).When A. ervi developed on SbBBI-intoxicated aphids,
percentages of mummification and emergence were notaffected. The former depends on percentage of para-sitised aphids (host-acceptance) and on successful larvaldevelopment. Thus, parasitoid females accepted SbBBI-intoxicated aphids as well as control ones, at least in ournon-choice experimental procedure. The larval andnymphal developmental times of both males and femaleswere not affected by the presence of SbBBI in the aphid.The resistance to starvation of parasitoids emerged fromaphids reared on SbBBI supplemented diets wasunaffected, although the emerging females were sig-nificantly smaller. In addition, the proportion of femaleprogeny developed on SbBBI-intoxicated aphids wassignificantly lower compared to the control. Lowerfemale size and male-biased sex-ratio could be explainedby the reduction of aphid size (Cloutier et al., 1981;King, 1987). In fact, a 6% reduction in aphid size wasobserved in a complementary experiment. Host-sizemediated effect on parasitoid size and sex-ratio was alsoreported by Couty et al. (2001a), when A. abdominalis
developed in GNA (Galanthus nivalis agglutinin) in-toxicated aphids. In addition, our results on A. ervi
proteases indicate that SbBBI may directly affect theparasitic larvae by binding to digestive proteases, thusimpairing insect protein digestion. It could also affecthaemolymph composition, and thus its nutritionalquality for the endoparasitic larvae.The direct exposure to SbBBI100 during adult food
intake did not affect female’s longevity, while this PIinduced 69% inhibition of adult digestive proteaseactivity. Even highest concentration of SbBBI (500 mg/mL) did not reduce A. ervi female’s survival (data notshown). Fecundity was not directly assessed in thepresent study, but the reduction of female size couldhave important effects on the fecundity (Visser, 1994;Lauziere et al., 2000). In addition, the lower proportionof female in the progeny could affect the parasitoidpopulation (Cloutier et al., 2000a). In another case, a5% size reduction of the aphid Metopolophium dir-
rhodum induced by partially resistant wheat cultivars ledto decreased weight of the braconid parasitoid Aphidius
rhopalosiphi by 34% and fecundity by 26% (van Emden,1995). So, sublethal effects of SbBBI-intoxicated aphidson A. ervi induced an alteration of parasitoid fitness.Regarding OCI, it could not be detected in parasitoid
larvae whatever the OCI concentration used to feed hostaphids. The same result was found with A. abdominalis
developed in OCI-intoxicated aphids (Azzouz et al.,2005). Previous study revealed that OCI bound to aphidproteins and the free form was not detected anymore inthis insect (Azzouz et al., 2005). This explains theabsence of OCI detection in endoparasitoid larvae. Wefound OCI in honeydew excreted by M. euphorbiae
reared on artificial diet supplemented by this PI. This isin accordance with the detection of OCI in honeydew ofM. persicae reared on OCI-expressing transgenic oilseedrape (Rahbe et al., 2003a). The study of OCI directeffect on A. ervi showed that the digestive proteolyticactivities revealed in the whole larvae or adult gutextracts were slightly inhibited by this PI (10% and 30%for larvae and adult, respectively). These results indicatethat the direct risk of exposure of A. ervi to the cysteinePI OCI was a priori low. Adult A. ervi survival was notaffected after exposure to PIs during food intake.Nevertheless, our bioassays showed that A. ervi couldnot develop to the adult stage in OCI-intoxicated aphids(few mummified aphids and no adult emergence). Suchdrastic effects on A. ervi can be explained by reducedhost survival and quality. Indeed, a complementaryexperiment showed that the size of M. euphorbiae rearedon OCI supplemented diet (OCI-100) was significantlyreduced by 17% and the nymphal survival wassignificantly altered (80% of mortality) (Azzouz et al.,2005). The adverse effects were not the consequence of adirect action of OCI on A. ervi by binding to digestiveproteases, since the parasitoid has a low cysteinedigestive proteases activity. However, detrimental ef-fects of OCI could come partly from the disruption ofnon-digestive parasitoid proteases (Faktor and Raviv,1997). Indeed, OCI could reach non-digestive proteasesof the immature parasitoid because immuno-histologicalstudies performed on M. persicae showed that OCIcrosses the midgut epithelium and reaches the hemo-lymph, where it accumulates in oenocytes and bacter-iocytes. As protease activity is intense duringmetamorphosis of the parasitoid larvae, it might beimpaired even by very small amount of the PIs, leadingto nymphal mortality.We investigated effects in the case of controlled
exposure to PIs in laboratory systems. What happenswith PI-expressing transgenic plants? Neither the fitnessof A. ervi females, nor the sex ratio of their offspringwere affected when the parasitoid developed on aphidsfeeding on OCI-potatoes (Cowgill et al., 2004). The lackof effects on parasitoid could be due to a very low level

ARTICLE IN PRESSH. Azzouz et al. / Journal of Insect Physiology 51 (2005) 941–951 949
of the OCI variant (OCIDD86) in the phloem sap. Infact, immunoassay experiments showed that OCI wasnot detected in aphids that fed on OCI-potatoes(Cowgill et al., 2004). Another study showed thatAphidius nigripes exhibited an increased size andfecundity when developed on M. euphorbiae feeding onOCI-expressing potatoes (Kennebec-OCI) (Ashouri etal., 2001). But this probiotic effect may not be related toOCI because the authors did not check OCI level in theirplants, and transgene expression is difficult to control intransgenic potatoes due to tetraploidy. The risk assess-ment study by Schuler et al. (2001) using M. euphorbiae
reared on OCI-expressing oilseed rape (Drakkar) didnot reveal any adverse effect on another Aphidiidaeparasitoid, Diaeretiella rapae. Nevertheless, the OCIexpression level in the phloem of the tested plants wastwice higher (0.2% of total protein) than the dose usedin our experiment (Rahbe et al., 2003a). These datahighlight the difficulty to control PI expression intransgenic plants, which can vary according to trans-genic line, plant physiology and tissue nature (De Leo etal., 1998, 2001; Cloutier et al., 2000b; Marchetti et al.,2000; Cowgill et al., 2004). With regard to a risk of PIseffect when adult parasitoids feed on honeydew, it canbe considered as negligible, because we found that PIsingestion by adult A. ervi was not harmful. However,honeydew that contains transgene products such GNA(Galanthus nivalis agglutinin) could pose a risk tohoneydew feeding parasitoids (Romeis et al., 2003; Bellet al., 2004). PIs could not be detected in the nectar oftransgenic plants containing genes encoding theseproteins, especially when the gene was under the controlof the CaMV 35S promoter (Malone and Burgess, 2000;Couty et al., 2002). Thus, adult parasitoids could not beintoxicated by nectar of PIs transgenic plants.Considering unreliability of PIs expression in cur-
rently available transgenic potatoes, we worked withcontrolled amounts of PIs delivered via artificial diets, inthe objective to get reliable effects on the parasitoid. Itallowed us to estimate the consequence of an exposureto plant PIs and the sensitivity of the aphid parasitoid A.
ervi to these entomotoxins, by combining immunologi-cal, biochemical and biological approaches. First itpointed out that only immature stages are affected byPIs. Secondly, it documented two different modes ofeffect, according to the nature of the PIs and both hostand parasitoid susceptibility. A. ervi developmentdisruption appears the consequence of its host suscept-ibility to OCI, whereas SbBBI only induced sublethaleffects on the parasitoid, possibly due to both directaction on the parasitoid susceptible proteases, and host-mediated action through size reduction.Now, in order to better predict side effects of PIs
transgenic plants on the third trophic level, analysingeffects of a range of PIs doses is one way to face variableexpression in planta. Our study shows that such analyse
can be restricted to the preimaginal development andoffspring fitness parameters. To predict effects on alonger term, it will be necessary to characterise effects onoffspring fitness, by studying the reproductive potentialat the second generation of parasitoids. This data,together with the improvement of PIs transgenic plants,are needed to sort out a key issue for sustainableagriculture, i.e. the compatibility between transgenicinsect-resistant plants and other components of inte-grated pest management (IPM), to get durable andeffective pest resistance.
Acknowledgements
We thank Dr. Celine Deraison and Dr. TatianaGorojankina for their help in the determination of A.
ervi digestive protease activity (Laboratoire de BiologieCellulaire, INRA-Versailles). Dr. Yvan Rahbe isthanked for advice and discussion (laboratoire deBiologie Fonctionnelle Insectes et Interactions, INRA-INSA de Lyon). The research was supported by aNational Tunisian grant to Hichem Azzouz, theMinistere Franc-ais de la Recherche, the ConseilRegional de Picardie and the Fonds Social Europeen.
References
Abe, K., Emori, Y., Kondo, H., Suzuki, K., Arai, S., 1987. Molecular
cloning of a cysteine proteinase inhibitor of rice (oryzacystatin).
Journal of Biological Chemistry 262, 16793–16797.
Alfonso-Rubı, J., Ortego, F., Castanera, P., Carbonero, P., Dıaz, I.,
2003. Transgenic expression of trypsin inhibitor CMe from barley
in indica and japonica rice, confers resistance to the rice weevil
Sitophilus oryzae. Transgenic Research 12, 23–31.
Annadana, S., Peters, J., Gruden, K., Schipper, A., Outchkourov,
N.S., Beekwilder, M.J., Udayakumar, M., Jongsma, M.A., 2002.
Effects of cysteine protease inhibitors on oviposition rate of the
western flower thrips, Frankliniella occidentalis. Journal of Insect
Physiology 48, 701–706.
Ashouri, A., Overney, S., Michaud, D., Cloutier, C., 1998. Fitness and
feeding are affected in the two-spotted stinkbug, Perillus bioculatus,
by the cysteine proteinase inhibitor, oryzacystatin I. Archives of
Insect Biochemistry and Physiology 38, 74–83.
Ashouri, A., Michaud, D., Cloutier, C., 2001. Recombinant and
classically selected factors of potato plant resistance to the
Colorado potato beetle, Leptinotarsa decemlineata, variously affect
the potato aphid parasitoid Aphidius nigripes. BioControl 46,
401–418.
Azzouz, H., Cherqui, A., Campan, E., Rahbe, Y., Duport, G.,
Jouanin, L., Kaiser, L., Giordanengo, P., 2005. Effects of plant
protease inhibitors, oryzacystatin I and soybean Bowman-Birk
Inhibitor, on the aphid Macrosiphum euphorbiae (Homoptera,
Aphididae) and its parasitoid Aphelinus abdominalis (Hymenop-
tera, Aphelinidae). Journal of Insect Physiology 51, 75–86.
Bell, H.A., Fitches, E.C., Down, R.E., Ford, L., Marris, G.C.,
Edwards, J.P., Gatehouse, J.A., Gatehouse, A.M.R., 2001. Effect
of dietary cowpea trypsin inhibitor (CpTI) on the growth and
development of the tomato moth Lacanobia oleracea (Lepidoptera:
Noctuidae) and on the success of the gregarious ectoparasitoid

ARTICLE IN PRESSH. Azzouz et al. / Journal of Insect Physiology 51 (2005) 941–951950
Eulophus pennicornis (Hymenoptera: Eulophidae). Pest Manage-
ment Science 57, 57–65.
Bell, H.A., Kirkbride-Smith, A.E., Marris, G.C., Edwards, J.P.,
Gatehouse, A.M.R., 2004. Oral toxicity and impact on fecundity
of three insecticidal proteins on the gregarious ectoparasitoid
Eulophus pennicornis (Hymenoptera: Eulophidae). Agricultural and
Forest Entomology 6, 215–222.
Bouchard, E., Michaud, D., Cloutier, C., 2003. Molecular interactions
between an insect predator and its herbivore prey on transgenic
potato expressing a cysteine proteinase inhibitor from rice.
Molecular Ecology 12, 2429–2437.
Bradford, M., 1976. A rapid and sensitive method for the quantifica-
tion of microgram quantities of protein using the principle of dye-
binding. Analytical Biochemistry 72, 248–254.
Burgess, E.P.J., Malone, L.A., Christeller, J.T., 1996. Effects of two
proteinase inhibitors on the digestive enzymes and survival of
honey bees (Apis mellifera). Journal of Insect Physiology 42,
823–828.
Carlini, C.R., Grossi-de-Sa, M.F., 2002. Plant toxic proteins with
insecticidal properties. A review on their potentialities as bioinsec-
ticides. Toxicon 40, 1515–1539.
Ceci, L.R., Volpicella, M., Rahbe, Y., Gallerani, R., Beekwilder, J.,
Jongsma, M.A., 2003. Selection by phage display of a variant
mustard trypsin inhibitor toxic against aphids. The Plant Journal
33, 557–566.
Charity, J.A., Anderson, M.A., Bitismch, D.J., Whitecross, M.,
Higgins, T.J.V., 1999. Transgenic tobacco and peas expressing a
protease inhibitor from Nicotiana alata increased insect resistance.
Molecular Breeding 5, 357–365.
Cloutier, C., McNeil, J.N., Regniere, J., 1981. Fecundity, longevity,
and sex ratio of Aphidius nigripes (Hymenoptera: Aphidiidae)
parasitizing different stages of its host, Macrosiphum euphorbiae
(Homoptera: Aphididae). The Canadian Entomologist 113,
193–198.
Cloutier, C., Duperron, J., Tertuliano, M., McNeil, J.N., 2000a. Host
instar, body size and fitness in the koinobiotic parasitoid Aphidius
nigripes. Entomologia Experimentalis et Applicata 97, 29–40.
Cloutier, C., Jean, C., Fournier, M., Yelle, S., Michaud, D., 2000b.
Adult Colorado potato beetles, Leptinotarsa decemlineata compen-
sate for nutritional stress on oryzacystatin I-transgenic potato
plants by hypertrophic behavior and over-production of insensitive
proteases. Archives of Insect Biochemistry and Physiology 44,
69–81.
Couty, A., de la Vina, G., Clark, S.J., Kaiser, L., Pham-Delegue,
M.H., Poppy, G.M., 2001a. Direct and indirect sublethal effects of
Galanthus nivalis agglutinin (GNA) on the development of a
potato-aphid parasitoid, Aphelinus abdominalis (Hymenoptera:
Aphelinidae). Journal of Insect Physiology 47, 553–561.
Couty, A., Down, R.E., Gatehouse, A.M.R., Kaiser, L., Pham-
Delegue, M.H., Poppy, G.M., 2001b. Effects of artificial diet
containing GNA and GNA-expressing potatoes on the develop-
ment of the aphid parasitoid Aphidius ervi Haliday (Hymenoptera:
Aphidiidae). Journal of Insect Physiology 47, 1357–1366.
Couty, A., Jouanin, L., Pham-Delegue, M.H., 2002. Impact de
proteines vegetales exprimees dans des plantes transgeniques, sur
des insectes pollinisateurs et auxiliaires. In: Regnault-Roger, C.,
Philogene, B.J.R., Vincent, C. (Eds.), Biopesticides d’Origine
Vegetale. Lavoisier, Paris, pp. 243–264.
Cowgill, S.E., Danks, C., Atkinson, H.J., 2004. Multitrophic interac-
tions involving genetically modified potatoes, nontarget aphids,
natural enemies and hyperparasitoids. Molecular Ecology 13,
639–647.
de Eguileor, M., Grimaldi, A., Tettamanti, G., Valvassori, R.,
Leonardi, M.G., Giordana, B., Tremblay, E., Digilio, M.C.,
Pennacchio, F., 2001. Larval anatomy and structure of absorbing
epithelia in the aphid parasitoid Aphidius ervi Haliday (Hymenop-
tera, Braconidae). Arthropod Structure and Development 30,
27–37.
De Leo, F., Bonade-Bottino, M.A., Ceci, L.R., Gallerani, R., Jouanin,
L., 1998. Opposite effects on Spodoptera littoralis larvae of high
expression level of a trypsin proteinase inhibitor in transgenic
plants. Plant Physiology 118, 997–1004.
De Leo, F., Bonade-Bottino, M., Ceci, L.R., Gallerani, R., Jouanin,
L., 2001. Effects of a mustard trypsin inhibitor expressed in
different plants on three lepidopteran pests. Insect Biochemistry
and Molecular Biology 31, 593–602.
Deraison, C., 2002. Isolation, characterization and targets of new
proteinase inhibitors for building transgenic plants resistant to
aphids. Thesis, University of Paris XI Orsay.
Down, R.E., Gatehouse, A.M.R., Hamilton, W.D.O., Gatehouse,
J.A., 1996. Snowdrop lectin inhibits development and decreases
fecundity of the glasshouse potato aphid (Aulacorthum solani)
when administered in vitro and via transgenic plants both in
laboratory and glasshouse trials. Journal of Insect Physiology 42,
1035–1045.
Down, R.E., Ford, L., Mosson, H.J., Fitches, E., Gatehouse, J.A.,
Gatehouse, A.M.R., 1999. Protease activity in the larval stage of
the parasitoid wasp, Eulophus pennicornis (Nees) (Hymenoptera:
Eulophidae); effects of protease inhibitors. Parasitology 119,
157–166.
Faktor, O., Raviv, M., 1997. Inhibition of molt in Spodoptera littoralis
larvae treated with soybean Bowman-Birk protease inhibitor.
Entomologia Experimentalis et Applicata 82, 109–113.
Falco, M.C., Silva-Filho, M.C., 2003. Expression of soybean
proteinase inhibitors in transgenic sugarcane plants: effects on
natural defense against Diatraea saccharalis. Plant Physiology and
Biochemistry 41, 761–766.
Febvay, G., Delobel, B., Rahbe, Y., 1988. Influence of amino acid
balance on the improvement of an artificial diet for a biotype of
Acyrthosiphon pisum (Homoptera, Aphididae). Canadian Journal
of Zoology 66, 2449–2453.
Ferry, N., Raemaekers, R.J.M., Majerus, M.E.N., Jouanin, L., Port,
G., Gatehouse, J.A., Gatehouse, A.M.R., 2003. Impact of oilseed
rape expressing the insecticidal cysteine protease inhibitor oryza-
cystatin on the beneficial predator Harmonia axyridis (multi-
coloured Asian ladybeetle). Molecular Ecology 12, 493–504.
Fisher, D., Wu, Y., Ku, M., 1992. Turnover of soluble proteins in the
wheat sieve tube. Plant Physiology 100, 1433–1441.
Foissac, X., Edwards, M.G., Du, J.P., Gatehouse, A.M.R., Gate-
house, J.A., 2002. Putative protein digestion in a sap-sucking
homopteran plant pest (rice brown plant hopper; Nilaparvata
lugens: Delphacidae)—identification of trypsin-like and cathepsin
B-like proteases. Insect Biochemistry and Molecular Biology 32,
967–978.
Hagvar, E.B., Hofsvang, T., 1991. Aphid parasitoids (Hymenoptera,
Aphidiidae): biology, host selection and use in biological control.
Biocontrol News and Information 12, 13–41.
Hilder, V.A., Gatehouse, A.M.R., Sheerman, S.E., Barker, R.F.,
Boulter, D., 1987. A novel mechanism of insect resistance
engineered into tobacco. Nature 300.
Ishiwatari, Y., Honda, C., Kawashima, I., Nakamura, S., Hirano, H.,
Mori, S., Fujiwara, T., Hayashi, H., Chino, M., 1995. Thioredoxin
H is one of the major proteins in rice phloem sap. Planta 195,
456–463.
Jouanin, L., Bonade-Bottino, M., Girard, C., Morrot, G., Giband, M.,
1998. Transgenic plants for insect resistance. Plant Science 131,
1–11.
King, B.H., 1987. Offspring sex ratios in parasitoid wasps. The
Quarterly Review of Biology 62, 367–396.
Kumar, H., Kumar, V., 2004. Tomato expressing Cry1A(b) insec-
ticidal protein from Bacillus thuringiensis protected against
tomato fruit borer, Helicoverpa armigera (Hubner) (Lepidoptera:

ARTICLE IN PRESSH. Azzouz et al. / Journal of Insect Physiology 51 (2005) 941–951 951
Noctuidae) damage in the laboratory, greenhouse and field. Crop
Protection 23, 135–139.
Lauziere, I., Perez-Lachaud, G., Brodeur, J., 2000. Effect of female
body size and adult feeding on the fecundity and longevity of the
parasitoid Cephalonomia stephanoderis Betrem (Hymenoptera:
Bethylidae). Annals of the Entomological Society of America 93,
103–109.
Lawrence, P.K., Koundal, K.R., 2002. Plant protease inhibitors in
control of phytophagous insects. Electronic Journal of Biotechnol-
ogy 5, 93–109.
Lecardonnel, A., Chauvin, L., Jouanin, L., Beaujean, A., Prevost, G.,
Sangwan-Norreel, B.S., 1999. Effects of rice cystatin I expression in
transgenic potato on Colorado potato beetle larvae. Plant Science
140, 71–79.
Lee, S., Koo, J., Chun, H., Lim, C., Mun, J., Song, Y., Cho, M., 1999.
Soybean Kunitz trypsin inhibitor (SKTI) confers resistance to the
brown planthopper (Nilaparvata lugens Stal) in transgenic rice.
Molecular Breeding 5, 1–9.
Leple, J., Bonade-Bottino, M., Augustin, S., Pilate, G., Letan, V.,
Delplanque, A., Cornu, D., Jouanin, L., 1995. Toxicity to
Chrysomela tremulae (Coleoptera: Chrysomelidae) of transgenic
poplars expressing a cysteine proteinase inhibitor. Molecular
Breeding 1, 319–328.
Malone, L.A., Burgess, E.P.J., 2000. Interference of proteinase
inhibitors on non-target organisms. In: Michaud, D. (Ed.),
Recombinant Protease Inhibitors in Plants. EUREKAH.COM,
pp. 91–108.
Marchetti, S., Delledonne, M., Fogher, C., Chiaba, C., Chiesa, F.,
Savazzini, F., Giordano, A., 2000. Soybean Kunitz, C-II and PI-IV
inhibitor genes confer different levels of insect resistance to tobacco
and potato transgenic plants. Theoretical Applied Genetic 101,
519–526.
Overney, S., Yelle, S., Cloutier, C., 1998. Occurrence of digestive
cysteine proteases in Perillus bioculatus, a natural predator of the
Colorado potato beetle. Comparative Biochemistry and Physiology
Part B 120, 191–196.
Rahbe, Y., Deraison, C., Bonade-Bottino, M., Girard, C., Nardon, C.,
Jouanin, L., 2003a. Effects of the cysteine protease inhibitor
oryzacystatin (OC-I) on different aphids and reduced performance
of Myzus persicae on OC-I expressing transgenic oilseed rape. Plant
Science 164, 441–450.
Rahbe, Y., Ferrasson, E., Rabesona, H., Quillien, L., 2003b. Toxicity
to the pea aphid Acyrthosiphon pisum of anti-chymotrypsin
isoforms and fragments of Bowman-Birk protease inhibitors from
pea seeds. Insect Biochemistry and Molecular Biology 33, 299–306.
Ranjekar, P.K., Patankar, A., Gupta, V., Bhatnagar, R., Bentur, J.,
Kumar, P.A., 2003. Genetic engineering of crop plants for insect
resistance. Current Science 84, 321–329.
Rice, M.E., Pilcher, C.D., 1998. Potential benefits and limitations of
transgenic Bt corn for management of the European corn borer
(Lepidoptera: Crambidae). American Entomologist 44, 75–78.
Romeis, J., Babendreier, D., Wackers, F.L., 2003. Consumption of
snowdrop lectin (Galanthus nivalis agglutinin) causes direct effects
on adult parasitic wasps. Oecologia 134, 528–536.
Schobert, C., Baker, L., Szederkenyi, J., Grobmann, P., Komor, E.,
Hayashi, H., Chino, M., Lucas, W.J., 1998. Identification of
immunologically related proteins in sieve-tube exudate collected
from monocotyledonous and dicotyledonous plants. Planta 206,
245–252.
Schuler, T.H., Denholm, I., Jouanin, L., Clark, S.J., Clark, A.J.,
Poppy, G., 2001. Population-scale laboratory studies of the effect
of transgenic plants on nontarget insects. Molecular Ecology 10,
1845–1853.
Stoger, E., Williams, S., Christou, P., Down, R.E., Gatehouse, J.A.,
1999. Expression of the insecticidal lectin from snowdrop
(Galanthus nivalis agglutinin; GNA) in transgenic wheat plants:
effects on predation by the grain aphid Sitobion avenae. Molecular
Breeding 5, 65–73.
Towbin, H., Staehelin, T., Gordon, J., 1979. Electrophoretic transfer
of proteins from polyacrylamide gels to nitrocellulose sheets:
procedure and some applications. Proceedings of National Aca-
demic Science of USA 76, 4350–4354.
van Emden, H.F., 1995. Host plant-Aphidophaga interactions.
Agriculture, Ecosystems and Environment 52, 3–11.
van Emden, H.F., 1999. Transgenic host plant resistance to insects—
some reservations. Annals of the Entomological Society of America
92, 788–797.
van Helden, M., Tjallingii, W.F., van Beek, T.A., 1994. Phloem sap
collection from lettuce (Lactuca sativa L.): chemical comparison
among collection methods. Journal of Chemical Ecology 20,
3191–3206.
Visser, M.E., 1994. The importance of being large: the relationship
between size and fitness in females of the parasitoid Aphaereta
minuta (Hymenoptera: Braconidae). Journal of Animal Ecology 63,
963–978.
Walker, A.J., Ford, L., Majerus, M.E.N., Geoghegan, I.E., Birch, N.,
Gatehouse, J.A., Gatehouse, A.M.R., 1998. Characterisation of the
mid-gut digestive proteinase activity of the two-spot ladybird
(Adalia bipunctata L.) and its sensitivity to proteinase inhibitors.
Insect Biochemistry and Molecular Biology 28, 173–180.




















