
Potential and limitations of current concepts regarding the response of clonal plants to...
16
Vegetatio 127:55 70, 1996. 55 (~) 1996 Kluwer Academic Publishers. Printed in Belgium. Potential and limitations of current concepts regarding the response of clonal plants to environmental heterogeneity Josef F. Stuefer Department of Plant Ecology & Evolutionary Biology, Utrecht University, Utrecht, The Netherlands Key words: Clonal growth, Constraints, Habitat patchiness, Morphological plasticity, Physiological integration, Plant-environment interactions Abstract Plant ecologists have spent considerable effort investigating the physiological mechanisms and ecological con- sequences of clonal growth in plants. One line of research is concerned with the response of clonal plants to envir- onmental heterogeneity. Several concepts and hypotheses have been formulated so far, suggesting that intra-clonal resource translocation, morphological plasticity on different organizational levels (e.g. leaves, ramets, fragments), and other features of clonal plants may represent potentially adaptive traits enabling stoloniferous and rhizomatous species to cope better with habitat patchiness. Although each of these concepts contributes substantially to our understanding of the ecology of clonal species, it is difficult to combine them into a consistent theoretical frame- work. This apparent lack of conceptual coherence seems partly be caused by an uncritical use of the term 'habitat heterogeneity'. Researchers have not always acknowledged the fact that 'heterogeneity' may refer to a number of fundamentally different aspects of environmental variability (i.e. scale, contrast, predictability, temporal vs. spatial heterogeneity), and that each of these aspects may, on one hand, allow for the evolution of specific plant responses to heterogeneity and, on the other, severely constrain the viability of potentially adaptive traits. Since adaptive responses are operational only in a narrow range of conditions (delimited by external environmental conditions and constraints internal to plants) it seems imperative to clearly define the context and the limits within which concepts regarding clonal plants' responses to heterogeneity are valid. In this paper an attempt is made to review a number of these concepts and to try and identify the necessary conditions for them to be operational. Special attention is paid (1) to different aspects of environmental heterogeneity and how they may affect clonal plants, and (2) to possible constraints (e.g. sectoriality, perception of environmental signals, morphological plasticity) on plant responses to patchiness. Introduction Over the last decade the clonal growth of plants and its ecological consequences have received increasing attention from plant ecologists (Jackson et al. 1985; de Kroon 1990; van Groenendael & de Kroon 1990; Callaghan et al. 1992; Dong 1994; Herben et al. 1994; Oborny & Podani 1995). At the same time there has been a growing awareness of the significance of small-scale environmental heterogeneity for the eco- logy of clonal plants, leading to the formulation of con- cepts and hypotheses regarding possible adaptations of clonal plants to a non-uniform distribution of resources and other environmental factors (Grime 1979; Pitelka & Ashmun 1985; Salzman 1985; Hutchings & Slade 1988; Hutchings & de Kroon 1994). These concepts are based on the idea that clonal plants, especially those with long spacers between ramets, are likely to experience environmental heterogeneity under field conditions and that specific traits may have evolved enhancing fitness-relevant processes such as resource uptake, clonal expansion and offspring establishment in heterogeneous habitats. Clonal 'strategies' proposed so far range from avoidance and tolerance responses based on plastic changes in clone architecture (Hutch- ings & de Kroon 1994) and physiological integration
Transcript of Potential and limitations of current concepts regarding the response of clonal plants to...

Vegetatio 127:55 70, 1996. 55 (~) 1996 Kluwer Academic Publishers. Printed in Belgium.
Potential and limitations of current concepts regarding the response of clonal plants to environmental heterogeneity
J o s e f F. S tue fe r Department of Plant Ecology & Evolutionary Biology, Utrecht University, Utrecht, The Netherlands
Key words: Clonal growth, Constraints, Habitat patchiness, Morphological plasticity, Physiological integration, Plant-environment interactions
Abstract
Plant ecologists have spent considerable effort investigating the physiological mechanisms and ecological con- sequences of clonal growth in plants. One line of research is concerned with the response of clonal plants to envir- onmental heterogeneity. Several concepts and hypotheses have been formulated so far, suggesting that intra-clonal resource translocation, morphological plasticity on different organizational levels (e.g. leaves, ramets, fragments), and other features of clonal plants may represent potentially adaptive traits enabling stoloniferous and rhizomatous species to cope better with habitat patchiness. Although each of these concepts contributes substantially to our understanding of the ecology of clonal species, it is difficult to combine them into a consistent theoretical frame- work. This apparent lack of conceptual coherence seems partly be caused by an uncritical use of the term 'habitat heterogeneity'. Researchers have not always acknowledged the fact that 'heterogeneity' may refer to a number of fundamentally different aspects of environmental variability (i.e. scale, contrast, predictability, temporal vs. spatial heterogeneity), and that each of these aspects may, on one hand, allow for the evolution of specific plant responses to heterogeneity and, on the other, severely constrain the viability of potentially adaptive traits. Since adaptive responses are operational only in a narrow range of conditions (delimited by external environmental conditions and constraints internal to plants) it seems imperative to clearly define the context and the limits within which concepts regarding clonal plants' responses to heterogeneity are valid. In this paper an attempt is made to review a number of these concepts and to try and identify the necessary conditions for them to be operational. Special attention is paid (1) to different aspects of environmental heterogeneity and how they may affect clonal plants, and (2) to possible constraints (e.g. sectoriality, perception of environmental signals, morphological plasticity) on plant responses to patchiness.
Introduction
Over the last decade the clonal growth of plants and its ecological consequences have received increasing attention from plant ecologists (Jackson et al. 1985; de Kroon 1990; van Groenendael & de Kroon 1990; Callaghan et al. 1992; Dong 1994; Herben et al. 1994; Oborny & Podani 1995). At the same time there has been a growing awareness of the significance of small-scale environmental heterogeneity for the eco- logy of clonal plants, leading to the formulation of con- cepts and hypotheses regarding possible adaptations of clonal plants to a non-uniform distribution of resources
and other environmental factors (Grime 1979; Pitelka & Ashmun 1985; Salzman 1985; Hutchings & Slade 1988; Hutchings & de Kroon 1994). These concepts are based on the idea that clonal plants, especially those with long spacers between ramets, are likely to experience environmental heterogeneity under field conditions and that specific traits may have evolved enhancing fitness-relevant processes such as resource uptake, clonal expansion and offspring establishment in heterogeneous habitats. Clonal 'strategies' proposed so far range from avoidance and tolerance responses based on plastic changes in clone architecture (Hutch- ings & de Kroon 1994) and physiological integration

56
(Pitelka & Ashmun 1985), respectively, to exploitation strategies due to a developmentally fixed (Callaghan 1976, 1988; J6nsd6ttir & Callaghan 1988) or environ- mentally inducible 'division of labour' among inter- connected ramets (Birch & Hutchings 1994; Stuefer et al. 1996).
All of these concepts are concerned with func- tional interactions between external, environmental conditions (i.e. habitat heterogeneity) and specific physiological, morphological and/or developmental processes internal to plants (e.g. phenotypic plasti- city, intra-clonal translocation of resources). However, little attention has been paid to the limitations and to the necessary pre-conditions for the proposed plant- environment interactions to be operational and, hence, to be considered as possibly adaptive traits. This makes it difficult to generalize results and conclusions from experimental studies and field observations, and to assess the ecological and evolutionary significance of responses of clonal plants to different types of envir- onmental heterogeneity.
This conceptual problem may partly arise from the use of unclear definitions, and partly from insufficient mechanistic thinking about what concerns both differ- ent aspects of environmental heterogeneity and pos- sible plant responses to them. Many studies on clonal plants have used vaguely defined concepts of envir- onmental heterogeneity, omitting explicit statements about which aspect of heterogeneity is being invest- igated. On the other hand, little attention has been paid to the fact that plant responses to heterogeneity are subject to various inherent constraints (e.g. percep- tion ability, size of plant, response time, sectoriality) which may significantly restrict the ability of a clonal plant to respond to habitat patchiness. The aim of this paper is to (a) make an explicit distinction between different aspects of environmental heterogeneity, i.e. to break down this general concept into its functional components, and (b) to survey possible constraints on responses of clonal plants to habitat heterogeneity.
Habitat heterogeneity
Habitat heterogeneity refers to a non-uniform distri- bution of resources or other biotic and abiotic envir- onmental conditions in the natural surroundings of an organism. Environmental patchiness (in space and in time) relevant to plants may occur above and below ground, and it can be caused by an uneven distri- bution of, for instance, light, nutrients, water, tox-
ic substances, pathogens, herbivore activity, physical disturbance and others. Patchiness is virtually ubiquit- ous under field conditions and it affects many ecolo- gically important processes and phenomena, includ- ing responses of populations (cf. Wiens 1976; Fowler 1988, Levin 1992) and of individuals and parts of indi- viduals (cf. Shorrocks & Swingland 1990; Caldwell & Pearcy 1994). Many ecological studies, however, view habitat heterogeneity in a rather uncritical way. Des- pite the availability of good conceptual frameworks for the study of heterogeneity (Wiens 1976, 1989; Addicott et al. 1987; Kotliar & Wiens 1990; Li & Reyn- olds 1994, 1995), ecologists have paid surprisingly little attention to the fact that environmental patchi- ness comprises a number of fundamentally different aspects such as scale, contrast, aggregation, predictab- ility, spatial covariance, and that patchiness can occur in space and/or in time. 'Habitat heterogeneity' is a general term which encompasses very different phe- nomena. It is important to realize that each of these phenomena offers specific opportunities for plants to respond to heterogeneity, and that each of them may exert specific constraints on the viability of putative adaptations of clonal plants to patchy environments.
Scale
Scale refers to the spatial and temporal dimensions of patches in a heterogeneous environment. In natural sys- tems heterogeneity occurs at different scales in space and in time (for temporal aspects of scale, see section Temporal versus spatial aspects of heterogeneity), and patterns and processes present at one scale may not occur or may not be functional on others (Addicott et al. 1987; Wiens 1989; Kotliar & Wiens 1990; Levin 1992). For example, an increase in nutrient availability within the rooting area of a single ramet may prompt specific responses such as a local stimulation of root growth (cf. Drew & Saker 1975; Fitter 1994; Robin- son 1994) or an enhanced uptake capacity for nutrients (Caldwell 1994), but on the level of ramet groups or whole clones, such small-scale heterogeneity may not provoke any response. On the other hand, heterogen- eity will not induce plant responses if it occurs on such a large scale that the whole plant lives within a single patch and hence perceives the environment as homo- geneous.
Heterogeneity in environmental conditions which is present in a given natural system may be called structural heterogeneity (Kolasa & Rollo 1991; see Figure la). This term refers to 'variability of a system

(a) e n v i r o n m e n t
' B :
space
(b) p l a n t s
i / i @ i ......
, 0 ............ space
(e) interaction
= ........ ; . . . . . . . / ;
space
Figure 1. Schematic representation of the range of scales in space (x-axes) and in time (y-axes), (a) at which habitat patchiness occurs in the two environments A and B (i.e. structural heterogeneity), (b) which can be perceived and responded to by plants 1, 2, and 3 (i.e. organisms' grain and extent), and (c) at which functional plant- environment interactions may be operational. The shaded area in (c) indicates the range of scales in time and space at which functional heterogeneity occurs. Plant 2 can not show functional responses to heterogeneity in environments A and B, because its perception ability (i.e. its grain and extent) lies outside the range of scales at which patchiness is present in these habitats. The scaling of axes, and the shape of zones (circles) at which patchiness occurs, or can be perceived by plants, are arbitrary.
property without reference to any functional effects' (Li & Reynolds 1995). Plants or functional plant parts can perceive spatial heterogeneity only within a cer- tain range of scales (Figure lb). The lower and the upper limits of this range of scales, termed 'grain' and 'extent', respectively (Kotliar & Wiens 1990; Wiens 1989, 1990), depend mainly on the size, the spa- tial structure and the physiological perception abilit- ies of plants or plant parts. Outside the limits of grain and extent every organism 'functionally perceives its environment as homogeneous and does not respond to any structure that might actually exist' (Kotliar & Wiens 1990). Functional responses of plants to hetero- geneity are only operational if structural heterogeneity occurs within an organism's limits of grain and extent ('overlap' area in Figure lc). Patchiness occurring in this range of scales is called.~nctional heterogeneity (Li & Reynolds 1995). It follows that the scale of environmental heterogeneity without reference to the organism or to the process under study has no direct meaning in the study of plant responses to patchiness
57
(i.e. heterogeneity should be organism- rather than observer-defined; Wiens 1976; Addicott et al. 1987; Levin 1992).
The range of scales within which organisms can perceive and respond to heterogeneity has been giv- en different terms by different authors. Addicot et al. (1987) have proposed the ambiguous term ecological neighborhood (other definitions exist for this term; cf. Antonovics & Levin 1980), which corresponds roughly to the organism's grain and extent (Kotliar & Wiens 1990) and, for sessile organisms like most plants, also to the resource depletion zone sensu Harper (1985; Addicott et al. 1987). It is important to note that any of these definition should include not only the surround- ings of a plant which provide resources for it, but also the environment which can be sensed by the perception of non-resource signals, such as changes in the red/far- red ratio of transmitted or reflected light (Aphalo & Ballar6 1995, and studies quoted there).
In clonal plant research the explicit recognition of scale seems especially important, because of the hier- archical organization of clones (cf. Tuomi & Vuorisalo 1989 a,b; Eriksson & Jerling 1990). Individual plant organs (e.g. leaves, roots), single ramets and groups of ramets (clonat fragments) are likely to show specific responses to spatial patchiness within their own limits of grain and extent, but these responses will not neces- sarily be present at other organizational levels. Indi- viduals of non-clonal plants and (similar-sized) ramets of clonal plants can be expected to show comparable responses to small-scale heterogeneity, such as loc- al proliferation of roots and shoots (Hutchings & de Kroon 1994; Ballar6 1994) or physiological plasticity in the uptake capacity for above and below ground resources (Jackson et al. 1990; Caldwell 1994; Pearcy & Sims 1994; Pearcy et al. 1994). However, if het- erogeneity occurs at the scale of interconnected ramet groups, such responses are not functional and may, hence, be substituted by other processes specifically linked to the organizational level of ramet groups, such as selective ramet placement (cf. Sutherland & Still- man 1988), clonal integration (cf. Pitelka & Ashmun 1985) or division of labour (cf. Callaghan 1988; Stue- fer et al. 1996). Heterogeneity at a larger scale than that of whole clones can not be responded to by indi- vidual plants, but it can affect population processes such as seed dispersal and germination. It should be emphasized that it is the spatial scale corresponding to interconnected clonal fragments which is of primary interest to clonal plant ecologists, because adaptive

58
responses which are unique to clonal plants may be found on this scale.
Despite its crucial importance for the viability of adaptive responses to patchiness, the match of scales between structural heterogeneity and the limits of grain and extent (cf. Figure 1) of clonal plants has rarely been tested explicitly. The model of Sutherland & Stillman (1988), for instance, makes clear predictions about optimal patch sizes for selective ramet placement to be beneficial, but no published study is available to date which compares patch size distributions in natural hab- itats of clonal plants with such theoretical predictions. Moreover, hypotheses on clonal plant adaptations to heterogeneity may not be valid if patch size distribu- tions and aggregation patterns (ranging from clumped to regular and random; Kotliar & Wiens 1990) present in the field differ markedly from those assumed in experimental studies and mathematical models. Con- siderable amounts of data are available on the spatial scales and pattern of habitat patchiness in different systems (cf. Eriksson 1986; Silvertown et al. 1988; Silvertown & Smith 1988, 1989; Kelly & Canham 1990; Diaz Barradas et al. 1992; Herben et al. 1995; Robertson & Gross 1994; Gross et al. 1995; Miller et al. 1995, and many others). However, very few stud- ies have specifically investigated the functional corres- pondence between naturally occurring habitat hetero- geneity and potential plant responses to it.
Contrast
Heterogeneous habitats are characterized by transitions between areas differing in environmental quality. The degree of difference between patches or between a patch and its surrounding matrix is called 'contrast' (Kotliar & Wiens 1990). The absence of contrast with- in the species-specific perception limits of plants or plant parts is equivalent to functional homogeneity of the environment (Figure 2). The level of contrast which is necessary to induce a response to heterogen- eity depends on the perception abilities of the plant under study with respect to the environmental factor causing heterogeneity.
For clonal plants, the contrast of patchiness plays a prominent role as the main external driving force behind integration effects. It is the degree of differ- ence between patches in terms of resource supply (Pitelka & Ashmun 1985; Marshall 1990), salt con- centration (Salzman & Parker 1985; Evans & Whitney 1992), and other factors like defoliation (J6nsd6ttir & Callaghan 1989.; Price & Hutchings 1992b), which is
high contrast
no contrast Figure 2. Schematic representation of differences in contrast between adjacent patches (vertical blocks within box) of a heterogeneous environment. The contrast between patches increases gradually from the bottom to the top of the figure. The dashed horizontal line indic- ates the limits of perception ability for an arbitrary organism growing in the patchy habitat. Below and above this line, the organism per- ceives the environment as uniform (functionally homogeneous) and non-uniform (functionally heterogeneous), respectively.
responsible for the establishment of source-sink rela- tions between adult interconnected ramets (Marshall 1990). It should be added that in many cases intra- clonal resource translocation may not be caused by environmental patchiness, but is a result of develop- mental differences between sinks (e.g. establishing ramets) and sources (adult clone parts) within clones (Marshall 1990; J6nsd6ttir & Callaghan 1990).
Physiological integration may be enhanced under higher levels of contrast, as predicted by theoretic- al studies (Caraco & Kelly 1991; Eriksson & Jer- ling 1990). Yet experimental evidence for this posit- ive relationship between contrast and the strength of clonal integration is rather scarce. In a greenhouse study on Glechoma hederacea, Stuefer & Hutchings (1994) aimed at testing this hypothesis, but strong internal constraints on carbon and nutrient integration prevented translocation of resources. An experiment by Wijesinghe & Handel (1994) provided evidence for the general hypothesis that the beneficial effects of integration increase with 'increasing heterogeneity'.

In this study, however, heterogeneity did not refer to the contrast of patchiness as defined above, but to a combination of other aspects of heterogeneity such as scale, spatial frequency, and aggregation of nutrient- rich patches in the experimental environment.
Spatial covariance
The vast majority of both experimental (Hartnett & Bazzaz 1983; Salzman & Parker 1985; Slade & Hutch- ings 1987a; Evans 1988; Alpert 1991; Dong 1993) and theoretical (Sutherland & Stillman 1988; Caraco & Kelly 1991; Oborny 1994 a,b; Cain 1994) stud- ies on the responses of clonal plants to environmental patchiness have made use of a simple mono-factorial model of spatial heterogeneity, in which patchiness is caused by a spatially uneven distribution of one single resource, e.g. light availability (Hartnett & Bazzaz 1983) or nutrient supply (Evans 1988). The resulting habitat consists of two types of micro-sites, resource- rich ('favourable') and resource-poor ('unfavourable') patches, comparable to the black and white squares on a chess board (Figure 3a). A qualitatively similar environment is created if patchiness is caused by two or more resources (e.g. light, water, nutrients), and if high-supply areas for different resources coincide spa- tially. In natural systems, such a positive covariance in resource supply may occur after disturbance (e.g. animal activity in grasslands, cf. Waite 1994) when resources become locally available for a limited period of time. It creates a chess board type of heterogen- eity with a multifactorial contrast between favourable and unfavourable patches (Figure 3b). If the supply of different resources covaries negatively in space a fundamentally different type of heterogeneity will be created (Figure 3c). In this case, patches rich in one resource are deficient in another and vice versa, and a general characterization of patches as 'favourable' and as 'unfavourable' is not possible (de Kroon et al. 1994; Stuefer et al. 1994; Stuefer & Hutchings 1994).
Under field conditions a spatially negative covari- ance in resource supply may be caused by inherently antagonistic relationships in the availability of dif- ferent resources. High-light availability, for instance, tends to reduce water availability to plants by creat- ing increased evapo-transpiration rates (c.f. Young & Smith 1979, 1980; Schulze & Hall 1981). As a con- sequence, high-light patches are favourable in terms of light supply but may be unfavourable in terms of water economy (Stuefer et al. 1994). Such a negative cov- ariance in the availability of water and light may also
59
be created by increased salt concentrations in open spots of salt marshes (Bertness et al. 1992; Shumway 1995). The availability of nutrients and light may also show a negative spatial covariance in natural systems because nutrient-rich microsites tend to support more dense vegetation than nutrient-poor patches (Stuefer & Hutchings 1994). A negative relationship between light and nutrient supply is also reported from a dune sys- tem referred to by Alpert & Mooney (1986, 1996) and Friedman & Alpert (1991 ). In this case nitrogen-fixing bushes locally increase soil nutrient levels and, at the same time, decrease light availability underneath their canopy. Evans & Whitney (1992) speculate about a possibly negative spatial covariance between the avail- ability of nitrogen and the presence of salt in their study system.
In habitats with a strict negative covariance in resource supply, simple search-avoidance responses of clonal plants can not be considered functional, because of the absence of both clearly unfavourable patches which could be avoided, and favourable patches which could be searched for. In this situation plant responses to heterogeneity, such as selective ramet placement in 'good' patches (Sutherland & Stillman 1988; Oborny 1994; i.e. 'habitat selection' sensu Salzman 1985) are not feasible because an aggregation of ramets in any one patch type is unlikely to be beneficial (de Kroon et al. 1994; Stuefer et al. 1996). Instead, oth- er responses such as local specialization in resource uptake combined with reciprocal translocation of cap- tured resources within clones ('spatial division of labour') may represent a viable strategy for clonal plants to exploit this specific type of environmental heterogeneity (Callaghan 1988; Lau & Young 1988; Evans 1991 ; Friedman & Alpert 1991 ; Hutchings & de Kroon 1994; de Kroon et al. 1994; Stuefer & Hutchings 1994). Tentative evidence for such responses comes from the experimental studies of Alpert & Mooney (1986), Friedman & Alpert (1991) and Stuefer et al.
(1994). A recent greenhouse study has provided clear experimental evidence for 'spatial division of labour' in clones of Trifolium repens (Stuefer et al. 1996).
The concept of 'selective ramet placement' by plastic changes in clone architecture and that of a 'spatial division of labour' through specialization and cooperation are based on different notions of environ- mental heterogeneity. Both types of patchiness, visu- alized in Figure 2a, b and 2c, respectively, may occur under field conditions. It should be emphasized that predictions and conclusions associated with each of

60
(a) / . / J , . / / / / /
(b) / " /
~,/fl /
(c) _ / /
Figure 3. Schematic representation of different types of spatial heterogeneity caused by a patchy distribution of one (a) or two (b, c) resources. The availability of the two resources may show a positive (b) or negative (c) covariance in space. Open and shaded areas indicate favourable (no resource limitation) and unfavourable (resource limitation) patches, respectively. The two types of shading refer to a low availability of two different resources (e.g. water and light).
these concepts are restricted to situations encompassed by the underlying model of environmental patchiness.
A spatially negative covariance in resource supply also has important consequences for the application of cost-benefit analyses to results of integration stud- ies. This type of analysis, introduced to clonal plant research by Salzman & Parker (1985), is based on a simple mono-factoriai model of heterogeneity (cf. Figure 2a, b) where clone parts can function either as donor or as recipient for the shared resource. If a negat- ive covariance in resource supply occurs, all parts of a clone may simultaneously import and export different resources. Ordinary cost-benefit analyses applied to such cases may yield surprising results, such as signi- ficantly negative cost terms, which fall outside the pre- dictions of mono-factorial cost-benefit models (Stuefer et al. 1994). Cost-benefit analyses are also applicable to multi-factorial cases, but assumptions and predic- tions would have to be re-defined for such situations (Stuefer & Hutchings 1994).
Temporal versus spatial aspects of heterogeneity
Habitat heterogeneity can refer to patchiness both in space and in time. Temporal heterogeneity refers to the dynamics of patches and (like spatial heterogen- eity) it comprises different aspects such as scale of temporal heterogeneity (i.e. patch longevity), contrast, frequency of patches, temporal aggregation, and pre- dictability through time. Most general statements made in foregoing sections about spatial aspects of hetero- geneity and potential plant responses to it, are - in an analogous way - also valid for environmental patchi- ness in time.
The range of temporal scales which can be per- ceived and responded to by plants is determined by the response time of the induced processes (i.e. lower limit or temporal grain), and by the life time of the organ- ism (i.e. upper limit or temporal extent). If the tem- poral scale of heterogeneity in the environment does not fall into the range between temporal 'grain' and

'extent' of the expected process or organism, no spe- cific response to heterogeneity is to be expected (see Figure 1). If, for instance, small-scale temporal het- erogeneity in resource supply occurs, 'slow' morpho- logical responses like plasticity in root-shoot alloc- ation or changes in internode length and branching frequency are unlikely to enhance resource uptake of clones. In this case 'fast' physiological responses like rapid changes in the uptake capacity of roots (Cald- well 1994) and leaves (Pearcy et al. 1994), some- times termed physiological plasticity (cf. Hutchings & de Kroon 1994), may be more effective responses to heterogeneity. It can be assumed that physiologic- al processes, including resource integration, are usu- ally faster in response than morphological changes, because they do not rely on the inherently 'slow' pro- cess of growth (i.e. tissue production). Large scale temporal variation exceeding the average life span of genetical individuals can not be responded to by onto- genetic processes (phenotypic plasticity), but only by evolution through natural selection.
Up to now clonal plant research has mainly locused on spatial aspects of heterogeneity, probably because the architecture of clones, especially those with long internodes between ramets, makes them very likely to experience patchiness in space. With a few exceptions, most studies were based on stationary models of habitat heterogeneity which (implicitly) assume temporal het- erogeneity to be absent. Theoretical considerations and mathematical models, however, suggest that temporal instability can exert severe constraints on the viability of potentially adaptive plant responses to heterogen- eity (cf. Obomy 1994 a,b). On the contrary, tempor- al variability on different scales may also have lead to the evolution of specific responses of clonal plants (e.g. 'sit-and-wait' strategies to capture short resource pulses; cf. Hutchings & de Kroon 1994). Experimental and theoretical studies, explicitly including temporal aspects of heterogeneity, may give further insight in the viability of specific plant responses in temporally variable environments.
Predictability
Predictability is probably both the most complex and the least understood aspect of environmental hetero- geneity. It can be expected to play a major role in the evolution of plant responses to heterogeneity, and it is also likely to constrain the viability of adaptive plant responses to patchiness. With the notable exception of two theoretical papers (Oborny 1994 a,b), however,
61
this aspect of habitat heterogeneity has only occasion- ally been included in studies on clonal plants (cf. Wijes- inghe & Handel 1994).
Predictability is the result of a complex interplay between spatial and temporal aspects of scale and aggregation of patches, and of their frequency in time and in space (Oborny 1994 a,b). The phenomenon of predictability comprises three different aspects, (a) spatial, (b) temporal, and (c) spatio-temporal predict- ability. For a good conceptual understanding of the ecological significance of environmental predictability it is necessary to study the viability and the effective- ness of plant responses, such as morphological plasti- city in clonal architecture, with respect to changes in each of these components (Obomy 1994b).
The spatial predictability of an environment increases with decreasing uncertainty of encountering a patch of specific environmental quality at a given dis- tance from any point in space. Spatial predictability is a function of patch size and patch frequency (Obomy 1994b). It measures the accuracy with which environ- mental quality (e.g. light availability) can be predicted in space. It attains maximum values in environments which are either homogeneous or show a regular pat- tern (i.e. a systematic change in environmental qual- ity) in space. Temporal predictability can be defined in an analogous way. It measures the accuracy with which the quality of the environment (i.e. the quality of one and the same point in space) can be predicted through time (Oborny 1994b). Highest and lowest val- ues of temporal predictability are reached in constant or systematically changing, and in randomly fluctuat- ing environments, respectively. Spatio-temporal pre- dictability measures the accuracy with which one can predict the quality of the environment at a given spa- tial distance and after a certain period of time from an arbitrary reference point in space and in time (Oborny 1994b). All three aspects of environmental predictab- ility may be quantified by autocorrelation studies (cf. Herben et al. 1995; During & Lloret 1996), or by the use of other methods of pattern analysis (cf. Upton & Fingleton 1985, 1989; Baldocchi & Collineau 1994).
Environmental predictability is strongly scale- dependent (Levin 1992). It only affects plant responses within the range of temporal and spatial scales which can be perceived by the plant or plant part under consid- eration. This implies that different hierarchical levels of a clonal system (sensu Tuomi & Vuorisalo 1989a, b; e.g. individual leaves and roots, ramets, integrated clonal fragments) may be exposed to environments dif-

62
fering in predictability. They are hence also likely to show different responses to habitat patchiness.
The growth of clones or clonal fragments is an inherently spatio-temporal process, because it leads to a change in the spatial position of plant parts (e.g. ramets, leaves, roots)through time. Plastic changes in the growth and elongation of spacers alter the spatio-temporal positioning of resource-acquiring plant parts in response to environmental signals. It thus seems reasonable to assume that the benefits of plastic changes in clone architecture are related to the spatio-temporal predictability of the environment. This view gains support from a simulation study (Oborny 1994a, b) which has shown that spatio-temporal pre- dictability within the range of scales perceivable by clonal fragments is tightly linked to 'the effectiveness of a plastic response of spacers to environmental qual- ity' (Oborny 1994b).
Environmental predictability also plays a central role in theoretical considerations about the evolution and effectiveness of plastic responses under differ- ent environmental conditions. According to Scbeiner (1993), plasticity is favoured in the course of evolution if (and only if) 'the environment is variable but predict- able' (Scheiner 1993, p.56). In this context variability may be interpreted as non-constancy of the environ- ment in time and/or in space, which is a precondition for plastic responses to evolve. Environmental pre- dictability relates to the accuracy with which benefi- cial effects of plastic responses can be predicted. In the case of plastic growth and elongation responses of plants and plant parts this is a function of the spatio- temporal predictability of the environment on the scale (in time and space) at which the responses operate. In a recent study Huber (1996) has suggested that envir- onmental (un-)predictability may explain the fact that in clonal species the length of (prostrate) stolon inter- nodes usually shows much lower degrees of plasticity in response to shading than the length of (vertical) petioles, or than vertically-oriented stem internodes of congeneric erect species (Huber 1996; see also Hutch- ings & de Kroon 1994; Dong 1995). The reason for this may lie in the spatio-temporal predictability of light supply which can be assumed to be rather high in the vertical plane of herbaceous canopies (Fliervoet & Werger 1984; Hirose & Werger 1995), i.e. leaves and branches on elongating stems of erect species as well as leaf blades on petioles of clonal ramets have a reasonable chance of encountering increased light levels after elongation (temporal aspect), which moves them higher up into the canopy (spatial aspect). In the
horizontal plane, light availability may be much less predictable. Data on spatial and temporal patterns of resource supply in natural habitats could be used to test this hypothesis. Unfortunately, no published papers are available to date which explicitly link ecological and evolutionary hypotheses about clonal plants to meas- ured patterns of resource availability in natural envir- onments.
In a variable environment with low degrees of spatio-temporal predictability, random expansion of clonal fragments may be the best strategy to adopt. Patches of increased resource availability encountered accidentally by clone parts may be exploited by a local adjustment of the uptake capacity for above and below ground resources. In this situation plastic responses are expected on the organizational level of individual ramets or parts of ramets, which are responsible for the opportunistic exploitation of locally and/or temporally increased resource supplies. This conclusion parallels the statement of de Kroon & Hutchings (1995) that in many instances plastic responses on the ramet level may be more profitable than on the fragment level. Under the assumption that plasticity requires a cer- tain level of spatio-temporal predictability for being profitable (and hence for being selected through evol- ution), this statement would imply that most habit- ats of clonal plants are spatio-temporally predictable on a small (spatial and temporal) scale, allowing for adaptive plasticity of individual ramets, but not on lar- ger scales, thereby preventing the evolution of plastic responses of clonal fragments. It should be emphasized that similar degrees of spatio-temporal predictability may occur on different spatial and/or temporal scales (cf. Oborny 1994b), which may lead to very different responses of plants or plant parts to habitat patchi- ness (e.g. morphological plasticity on different scales, physiological plasticity, reversible and non-reversible responses, etc.).
In habitats which (on the spatio-temporal scale of clone expansion) are too unpredictable to provoke adaptive plasticity in the architecture of clonal frag- ments, physiological integration combined with plastic responses on a lower organizational level than the frag- ment, may play a crucial role. Integration allows for a rapid re-distribution of locally captured resources over adjacent clone parts. This idea gains support from an experimental study on Glechoma hederacea (Birch & Hutchings 1994) which has demonstrated the potential of developing clones to opportunistically exploit loc- alized soil resources through a strong increase in the nutrient uptake capacity of ramets (situated in a rich

patch), and to distribute these resources over clone parts situated outside the patch.
Plastic responses at low, and strong resource integ- ration at higher organizational levels have also been described for clonal sedges from resource-poor envir- onments and arctic-alpine tundra habitats (e.g. Carex bigelowii; cf. Callaghan 1976, 1988; J6nsd6ttir 1989). In these systems patches of increased resource availab- ility are generally infrequent (in time and in space) and of a relatively short duration. The chance of encoun- tering favourable resource conditions at a single point in space and time, and the chance of being able to exploit patches by slow morphological responses, is consequently rather low. In this situation plants can enhance their resource acquisition through an increase in the number of sampling points which are active dur- ing extended periods of time, and which can rapidly respond to resource pulses. This may partly explain the success of large and long-lived, integrated clonal sys- tems in resource-poor environments, which apparently adopt a 'random-growth' and 'sit-and-wait' strategy (cf. J6nsd6ttir 1989; Hutcbings & de Kroon 1994).
It is clear that natural environments may strongly differ in the degree of environmental predictability and in the spatial and/or temporal scale at which pre- dictability occurs. This may exert both strong con- straints on specific plant responses to patchiness, but it may also offer possibilities for alternative traits to develop. Detailed theoretical and experimental studies are needed to unravel the complex interplay between aspects of predictability, scale and patch frequency, in shaping and constraining the evolution and expression of clonal plants' responses to heterogeneity in differ- ent environments. The results of such studies could be linked to general theories on plasticity and envir- onmental variability (cf. Grime et al. 1986; Campbell et al. 1991; Hutchings & de Kroon 1994).
Constraints on plant responses
Morphological plastici~
(a) Selective ramet placement Plastic changes in the length of internodes and petioles, in branching frequency and biomass allocation to roots and shoots form the basis for several concepts of how clonal plants may respond to environmental patchiness. The idea of selective ramet placement (sensu Suther- land & Stillman 1988), is based on environmentally
63
inducible changes in spacer length and branching fre- quency (cf. Hutchings & Slade 1988). Numerous stud- ies have quantified these parameters for various spe- cies under different environmental conditions (literat- ure reviewed by Hutchings & de Kroon 1994; de Kroon & Hutchings 1995) but only a few of them (e.g. Dong & de Kroon 1994) have reported degrees of plasticity which can reasonably be expected to result in a sig- nificant ramet concentration in favourable patches of a heterogeneous environment. The majority of clonal species studied so far, however, showed degrees of plasticity which are considerably below model predic- tions (cf. Sutherland & Stillman 1988) for success- ful ramet placement in patchy habitats. Based on this evidence, de Kroon & Hutchings (1995) have recently concluded that 'many clonal species will be unable to forage selectively for favourable patches within their environment because they lack the appropriate mor- phological plasticity'. In general terms this means that the lack of correspondence between external condi- tions (heterogeneity) and the degree of plant response (plasticity in internode length) constrains the applic- ability of the 'foraging'-concept in the sense of select- ive ramet placement, as it was originally proposed for Glechoma hederacea (Slade & Hutchings 1987b). The recognition of the limitations of this concept has sub- stantially contributed to the awareness among clonal plant ecologists that both external and internal con- straints may restrict the validity of adaptionist hypo- theses to a well-definable, narrow set of conditions.
(b) Directional meristem initiation Directional growth of clonal plants could be a result of directional meristem initiation, i.e. of a selective outgrowth of buds directed towards more favourable patches in a heterogeneous environment. Studies on non-clonal plants have convincingly shown the poten- tial of directional bud outgrowth in producing specific responses to heterogeneous light environments (Jones 1985; Jones & Harper 1987 a,b; Novoplansky et al. 1990). It seems reasonable to expect similar responses in clonal plants growing in habitats with sharp bor- ders between open sites and the surrounding veget- ation, such as sea and river shores, where parts of the vegetation are regularly destroyed by flooding. In such situations selective bud outgrowth (or inhibition) could lead to directional growth of stolons towards open sites and, hence to a rapid colonisation of new territory. Solangaarachchi & Harper (1989) provide evidence that bud development and meristem activity

64
in Trifolium repens can show plastic responses to the quality of their immediate environment.
Directional growth through selective bud initiation is constrained by the number and position of viable meristems on the ramets. This morphological con- straint is expected to be strongest in clonal plants with only one or two buds per tamer (e.g. Trifolium, Glechoma, Hydrocotyle, Oxalis) and it may be virtu- ally absent in adult ramets of rosette-forming plants (e.g. Potentilla, Fragaria), where numerous buds posi- tioned around the centre of the rosette are available for stolon formation. Up to now, however, little spe- cific information has been collected on directional bud outgrowth in clonal plants and its potential ecological significance in heterogeneous environments.
Resource translocation
(a) Vascular constraints Translocation of resources between ramets plays a central role in the response of clonal plants to envir- onmental heterogeneity. Numerous studies have been carried out to quantify translocation streams by using tracers (see Pitelka & Ashmun 1985; Marshall 1990 for references), or to estimate integration effects on growth through an analysis of the costs and benefits of resource sharing (Salzman & Parker 1985; Slade & Hutchings 1987a; Stuefer & Hutchings 1994; Stuefer et al. 1994). However, little attention has been paid to potential con- straints on translocation processes due to the vascular organization of plants. So far, very few studies (Price & Hutchings 1992 a,b; Price et al. 1992) have systemat- ically investigated the way in which patterns of clonal integration are shaped by constraints, such as sectorial- ity and the gradual disintegration of developing clones into physically connected but functionally independ- ent fragments (IPUs sensu Watson 1984, and Watson & Casper 1984). In Glechoma hederacea, for instance, the vascular architecture of shoots strongly affects pat- terns of resource integration, mainly because stolon branches (including secondary and higher order sto- lons and ramets) originating from the same node lack direct vascular contacts and are hence unable to share resource (Price & Hutchings 1992 a,b).
Similar constraints on intra-clonal resource move- ment are likely to occur in other clonal taxa within the Lamiaceae family, because many of them show similar vascularization patterns to Glechoma, characterized by an opposite decussate leaf insertion, a 'closed' vascu- lar system and a strongly reduced number of vascular
bundles in the cross section of stems (Strasburger 1991 ; Gifford & Foster 1989). All of these characteristics can be expected to favour sectoriality in resource transport within clones. Although morphologists, taxonomists and developmental biologists have studied and metic- ulously described patterns of vascularization in many plant families (Dormer 1972; Zimmermann & Tomlin- son 1972; Gifford & Foster 1989; Strasburger 1991), the implications of their findings in terms of sectorial constraints on resource movement in clonal plants have never been analyzed in a comprehensive way.
Sectorial constraints arise from constructional properties of the vascular system such as the stelar architecture (e.g. number and arrangement of vascu- lar bundles), the organization of nodes (e.g. 'lacun- arity', 'branching' of bundles), the type of vascular system ( 'open' vs. 'closed'), and others (cf. Dormer 1972; Gifford & Foster 1989). Many of these traits are likely to be linked to certain phylogenetic groups, and may hence differ strongly between higher taxa such as between monocots and dicots, or also between taxo- nomic groups within dicots (e.g. Lamiidae vs. Rosid- ae). Integration studies on grasses and sedges suggest that strong vascular constraints on intra-clonal resource translocation may be more or less absent monocots (cf. Pitelka & Ashmun 1985; Steeves & Sussex 1989; de Kroon & van Groenendaal 1990; Marshall 1990). In contrast to Glechoma, dicotyledonous species such as Trifolium repens show very little sectorial effects on assimilate integration as demonstrated by various tracer studies (cf. Chapman et al. 1991 a,b, 1992; Kemball & Marshall 1995). Comparative experimental studies and spatially explicit models of resource integ- ration, taking into account the vascular architecture of different phylogenetic groups, could provide further insight in the ecological significance of vascular con- straints in clonal plants.
(b) Directional constraints on resource transport Tracer studies have shown that in several clonal spe- cies assimilates are predominantly moved acropetally (i.e. towards developmentally younger clone parts), and to a much lesser extent in a basipetal direction (Pitelka & Ashmun 1985; Marshall 1990). At least in some species, such as in Glechoma hederacea, this pat- tern of 'asymmetric' resource movement does not only reflect simple source-sink relationships (i.e. growing parts are stronger sinks than older parts), as the acro- petal translocation stream is not fully reversible by shading or defoliation (Marshall 1990, literature cited

there; Price et al. 1992; Slade & Hutchings 1987a). In a recent study, Stuefer & Hutchings (1994) found no evidence for assimilate translocation between inter- connected fragments of Glechoma hederacea, sub- jected to spatially heterogeneous light (and nutrient) environments. The lack of integration was not due to vascular constraints. Since assimilate sharing would have required basipetal translocation in this experi- mental set-up, the absence of integration effects was most probably caused by an irreversible acropetal flow of photosynthates (Stuefer & Hutchings 1994). This shows that such asymmetry in intra-clonal resource translocation may strongly constrain the viability of integration responses to environmental patchiness.
In dicotyledonous species like Tri~lium repens, Fragaria chiloensis, Potentilla anserina and P reptans (cf. AIpert & Mooney 1986; Marshall 1990; Chap- man et al. 1991 a,b, 1992; Stuefer et al. 1994), as well as in grasses and sedges (cf. Callaghan 1977; J6nsd6ttir 1989; Marshall 1990), directional constraint on assimilate movement within clones appear to be less severe which means that virtually all interconnected clone parts can share assimilates according to internal source-sink relationships. The proximate and ultimate causes of strong asymmetric assimilate movement in some species, and the striking absence of this constraint in others, are still largely unexplored.
Directional constraints are less likely to affect water transport in plants because this is a passive process driven by water potential gradients, which arise from intra-clonal differences in water loss and water uptake (Marshall 1990, and literature quoted there).
(c) Pathways of resource transport Translocation of the three main resources for plant growth (i.e. water, assimilates, nutrients) relies on two different transport systems. The xylem and the phloem represent specific transport systems for water and assimilates, respectively, and they are largely inde- pendent from each other with respect to source-sink relationships and the direction of resource transloca- tion (Marshall 1990). In contrast, transport of nutrients is always coupled to, and thus constrained by, the flow of other resources. This interdependency affects nutri- ent translocation patterns in clonal plants, and it may have important consequences for the interpretation of nutrient integration studies, and also for the applicab- ility of ecological concepts, such as intra-clonal 'divi- sion of labour' in resource uptake (cf. Callaghan 1988; Hutchings & de Kroon 1994) which are based on bid-
65
irectional translocation of nutrients and assimilates (or nutrients and water) in clonal plants.
It appears difficult to assess the significance of such pathway constraints for the interpretation of existing studies on nutrient integration in clonal plants. Most papers fail to consider that nutrients may be assimilated in different organs in different plants (cf. Pate 1980, 1983; Peoples & Giff'ord 1990), and consequently be translocated in different systems. The latter depends on whether they are free ions moving in the xylem mainly, or chemically bound to assimilates and hence transported in the phloem. In the case of nitrogen the pattern of intra-clonal translocation and its dependence on water and assimilate movement may be rather com- plex, as it can be taken up in different forms by plants. The two main forms, nitrate and ammonium, may be assimilated in different organs, and may thus be sub- ject to different transport constraints. Moreover, these two forms of nitrogen may not be present or may not be taken up in equal amounts by different plant species occurring in different systems (cf. Lee & Stewart 1978; J6nsd6ttir & Callaghan 1990), which further complic- ates the exact pattern of nutrient sharing within clonal systems.
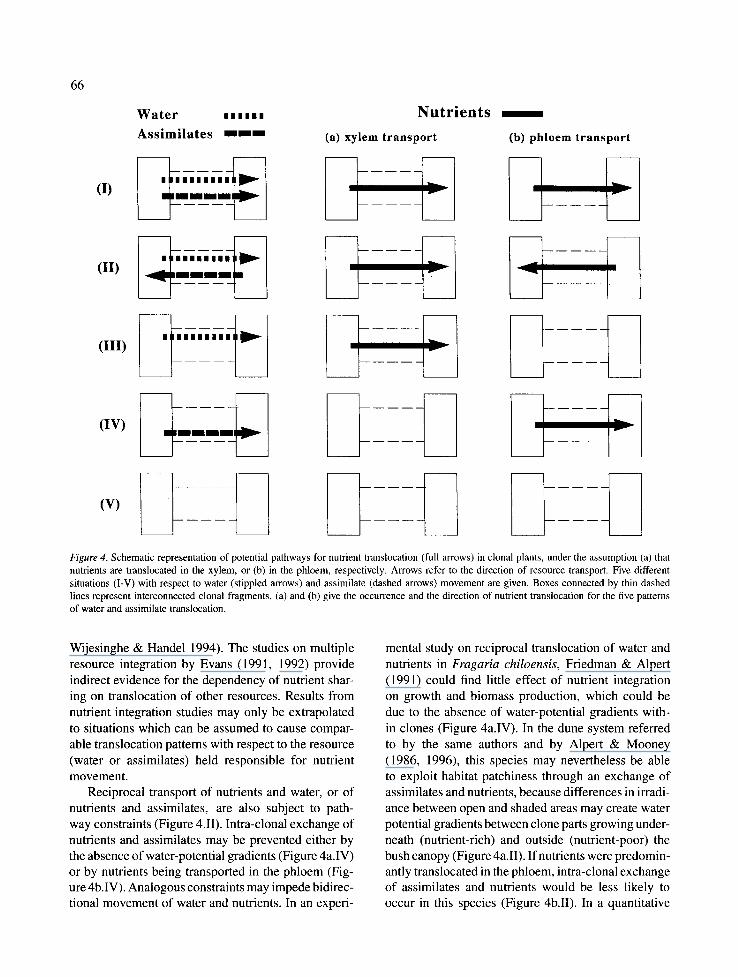
The inherent coupling of nutrient movement to one of the two main transport systems in plants can lead to different constraints under different assumptions (Fig- ure 4). For some plants mass flow of nutrients takes place in the xylem and hence follows water potential gradients from absorbing roots to transpiring leaves (cf. Marshall 1990; Figure 4a). In this case nutrient sharing is likely to be effective only in the presence of paral- lel gradients in plant water potential between nutrient sources and sinks, respectively (Figure 4.I-III). Stuefer & Hutchings (1994)concluded that the lack of nutrient sharing in their experiment was due to this constraint. However, the situation depicted in Figure 4b (III or V), i.e. nutrient transport coupled to assimilate movement, could be another interpretation for the same result. For many clonal species (including Glechoma hederacea) it is not clear yet, which transport system is responsible for the mass-flow of nutrients.
Since intra-clonal gradients in assimilates and/or water supply are a pre-condition for nutrient translo- cation to occur, pathway constraints are unavoidable in studies on nutrient sharing in clonal plants. Only a minority of papers (J6nsd6ttir & Callaghan 1990) have incorporated such constraints in their predictions and conclusions, while most others did not explicitly take them into account (cf. Slade & Hutchings 1987a; Evans 1988; Alpert 1991; Birch & Hutchings 1994;
66
(I)
( [ [)
( I I l )
(IV)
N u t r i e n t s
(a) xylem transport (b) phloem transPort
Water mnmmuu
A s s i m i l a t e s nnmmmmm
~ l l l l l l i l ] ~
(v, ,
Figure 4. Schematic representation of potential pathways for nutrient translocation (full arrows) in clonal plants, under the assumption (a) that nutrients are translocated in the xylem, or (b) in the phloem, respectively. Arrows refer to the direction of resource transport. Five different situations (I-V) with respect to water (stippled arrows) and assimilate (dashed arrows) movement are given. Boxes connected by thin dashed lines represent interconnected clonal fragments. (a) and (b) give the occurrence and the direction of nutrient translocation for the five patterns of water and assimilate translocation.
Wijesinghe & Handel 1994). The studies on multiple resource integration by Evans (1991, 1992) provide indirect evidence for the dependency of nutrient shar- ing on translocation of other resources. Results from nutrient integration studies may only be extrapolated to situations which can be assumed to cause compar- able translocation patterns with respect to the resource (water or assimilates) held responsible for nutrient movement.
Reciprocal transport of nutrients and water, or of nutrients and assimilates, are also subject to path- way constraints (Figure 4.II). Intra-clonal exchange of nutrients and assimilates may be prevented either by the absence of water-potential gradients (Figure 4a.IV) or by nutrients being transported in the phloem (Fig- ure 4b.IV). Analogous constraints may impede bidirec- tional movement of water and nutrients. In an experi-
mental study on reciprocal translocation of water and nutrients in Fragaria chiloensis, Friedman & Alpert (1991) could find little effect of nutrient integration on growth and biomass production, which could be due to the absence of water-potential gradients with- in clones (Figure 4a.IV). In the dune system referred to by the same authors and by Alpert & Mooney (1986, 1996), this species may nevertheless be able to exploit habitat patchiness through an exchange of assimilates and nutrients, because differences in irradi- ance between open and shaded areas may create water potential gradients between clone parts growing under- neath (nutrient-rich) and outside (nutrient-poor) the bush canopy (Figure 4a.II). If nutrients were predomin- antly translocated in the phloem, intra-clonal exchange of assimilates and nutrients would be less likely to occur in this species (Figure 4b.II). In a quantitative

study of reciprocal transport of water and nutrients in a clonal Carex species, de Kroon et al. (subm.) have demonstrated that only a negligible amount of nitrate- nitrogen was moved against intra-clonal water gradi- ents in their two-ramet system. This study provides a clear example of the interdependency of nutrient and water transport in a clonal plant. The results, however, may not be generalizable to other species (with phloem transport of nutrients) or to other forms of nutrients.
Bi-directional transport of water and assimilates is unlikely to be affected by such constraints. A number of experimental studies provide evidence for this view. Alpert & Mooney (1986) grew interconnected ramet pairs of Fragaria chiloensis in extreme conditions of either drought or quasi-darkness. Disconnecting the two ramets lead to the death of plants, whereas intact pairs survived, suggesting that ramets were able to exchange water and assimilates. The studies by Stuefer et al. (1996) and Stuefer (1995) provide strong circum- stantial evidence for reciprocal transport of assimilates and water between interconnected fragments of Trifo- lium repens and Potentilla anserina.
Conclusion
Various concepts and models are available which aim at interpreting and predicting functional responses of clonal plants to heterogeneous environments. Most of these concepts have (often implicitly) made specific assumptions about the type of variability occurring in the environment, and about the ability of clonal plants to perceive and to respond to habitat patchiness. It is a challenge for future research on clonal plants to com- bine these concepts into a coherent theoretical frame- work, and to assess their potential in explaining func- tional interactions between environmental patterns in space and time, and processes internal to clonal plants. This paper may contribute to the awareness that for this it is necessary to sharply define the limitations of con- cepts, and to identify potential constraints operating on them.
Acknowledgements
I am very grateful to Heinjo During, Tina D'Hertefeldt, Milena Holmgren, Heidrun Huber, Mike Hutchings, Ingibj6rg J6nsd6ttir, M. Esth6r P6rez Corona, Marinus Werger, Thomas Wilhalm, and to all participants of the Seili meeting for stimulating discussions and valuable
67
suggestions on topics raised in this paper. I thank the editors for their patience (in space and in time), an anonymous and a half-anonymous referee for critical comments on an earlier version of the manuscript, and Be~ita Oborny for her functional response to unpredict- able requests about section Predictability. I am grate- ful to Johanna Wagner, Walter Latchet and Vladimir Onipchenko for providing information used in section Vascular constraints. This work was financially sup- ported by the Faculty of Biology of the Utrecht Uni- versity.
References
Addicott, J. F., Aho, J. M., Antolin, M. E, Padilla, D. K., Richardson, J. S. & Soluk, D. A. 1987. Ecological neighborhoods: scaling environmental patterns. Oikos 49: 340-346.
Alpert, P & Mooney, H. A. 1996. Resource heterogeneity generated by shrubs and topography on coastal sand dunes. Vegetatio 122: 83-93.
Alpert, E 1991. Nitrogen sharing among ramets increases clonal growth in Fragaria chiloensis. Ecology 72: 69-80.
Alpert, E & Mooney, H. A. 1986. Resource sharing among ramets in the clonal herb, Fragaria chiloensis. Oecologia 70: 227-233.
Antonovics, J. & Levin, D. A. 1980. The ecological and genetic consequences of density-dependent regulation in plants. Ann. Rev. Ecol. Syst. 1 h 411-452.
Aphalo, P. J. & Ballar6, C. L. 1995. On the importance of information-acquiring systems in plant-plant interactions. Funct. Ecol. 9: 5-14.
Baldocchi, D. & Collineau, S. 1994. The physical nature of solar radiation in heterogeneous canopies: spatial and temporal attrib- utes. Pp. 21-71. In: Caldwell M. M. & Pearcy R. W. (eds), Exploitation of environmental heterogeneity by plants. Academic Press, London.
Ballar6, C. L. 1994. Light gaps: sensing the light opportunities in highly dynamic canopy environments. Pp. 73-110. In: Caldwell M. M. & Pearcy R. W. (eds), Exploitation of environmental het- erogeneity by plants. Academic Press, London.
Bertness, M. D., Gough, L. & Shumway, S. W. 1992. Salt tolerance and the distribution of vascular plants in a New England salt marsh. Ecology 73:1842-1851.
Birch, C. P. D. & Hutchings, M. J. 1994. Exploitation of patchily distributed soil resources by the clonal herb Glechoma hederacea. J. Ecol. 82: 653~164.
Cain, M. L. 1994. Consequences of foraging in clonal plant species. Ecology 75: 933-944.
Caldwell, M. M. 1994. Exploiting nutrients in fertile soil microsites. Pp. 325-347. In: Caldwell M. M. & Pearcy R. W. (eds), Exploit- ation of environmental heterogeneity by plants. Academic Press, London.
Caldwell, M. M. & Pearcy, R. W. (eds) 1994. Exploitation of envir- onmental heterogeneity by plants. Academic Press, London.
Callaghan, T. V. 1976. Growth and population dynamics of Carex bigelowii in an alpine environment. Oikos 27: 402-413.
Callaghan, T. V. 1977. Adaptive strategies in the life cycles of South Georgian graminoid species. Pp. 981-10f12. In: Llano, G. A. (ed.), Adaptations within Antarctic ecosystems. Smithsonian Inst., Washington.

68
Callaghan, T. V. 1988. Physiological integration and demograph- ic implications of modular construction in cold environments. Pp. 111-135. In: Davy A. J., Hutchings M. J. & Watkinson A. R. (eds), Plant population biology. Blackwell Scientific Publica- tions, Oxford.
Callaghan, T. V., Carlsson, B. ,~., J6nsd6ttir, I. S., Svensson, B. M. & Jonasson, S. 1992. Clonal plants and environmental change: introduction to the proceedings and summary. Oikos 63: 341- 347.
Campbell, B. D., Grime, J. P. & Mackey, J. M. L. 1991. A trade-off between scale and precision in resource foraging. Oecologia 87: 532-538.
Caraco, T. & Kelly, C. K. 1991. On the adaptive value of physiolo- gical integration in clonal plants. Ecology 72:81-93.
Chapman, D. E, Robson, M. J. & Snaydon, R. W. 1991a. The influence of leaf position and defoliation on the assimilation and translocation of carbon in white clover (Trifolium repens L. ). I. Carbon distribution patterns. Ann. Bot. 67: 295-302.
Chapman, D. F., Robson, M. J. & Snaydon, R. W. 1991b. The influence of leaf position and defoliation on the assimilation and translocation of carbon in white clover (Trifolium repens L. ). 2. Quantitative carbon movement. Ann. Bot. 67: 303-308.
Chapman, D. E, Robson, M. J. & Snaydon, R. W. 1992. Physiologic- al integration in the perennial herb Trifolium repens L. Oecologia 89: 338-347.
de Kroon, H. 1990. In search of a foraging plant. The clonal growth of Brachypodium pinnatum and Carexflacca. Ph. D. Dissertation, Utrecht University. 107p.
de Kroon, H. & Hutchings, M. J. 1995. Morphological plasticity in clonal plants: the foraging concept reconsidered. J. Ecol. 83: 143-152.
de Kroon, H., Stuefer, J. E, Dong, M. & During, H. J. 1994. On plastic and non-plastic variation in clonal plant morphology and its ecological significance. Folia Geobot. Phytotax. 29:123-138.
de Kroon, H., van der Zalm, E., van Rheenen, J. W. A., van Dijk,A. & Kreulen, R. (submitted). The interaction between water and nutrient translocation in a rhizomatous sedge (Carexflacca).
de Kroon, H. & van Groenendaal, J. 1990. Regulation and function of clonal growth in plants: an evaluation. Pp. 177-186. In: van Groenendael J. & de Kroon H. (eds), Clonal growth in plants: regulation and function. SPB Academic Publishing, The Hague.
Diaz Barradas, M. C., During, H. J. & Terlou, M. 1992. The struc- ture of bryophyte communities in the dunes of Meijendel, Neth- erlands. Pp. 313-323. In: Carter R. W. G., Curtis T. G. F. & Sheehy-Skeffington M. J. (eds), Coastal dunes. Geomorphology, ecology and management for conservation. Balkema, Rotterdam.
Dong, M. 1993. Morphological plasticity of the clonal herb Lami- astrum galeobdolon (L.) Ehrend. & Polatschek in response to partial shading. New Phytol. 124: 291-300.
Dong, M. 1994. Foraging through morphological plasticity in clonal herbs. Ph. D. Dissertation, Utrecht University. 96 p.
Dong, M. 1995. Morphological responses to local light conditions in clonal herbs from contrasting habitats, and their modification due to physiological integration. Oecologia 101: 282-288.
Dong, M. & de Kroon, H. 1994. Plasticity in morphology and bio- mass allocation in Cynodon dactylon, a grass forming stolons and rhizomes. Oikos 70: 99-106.
Dormer, K. J. 1972. Shoot organization in vascular plants. Chapman & Hall. London.
Drew, M. C. & Saker, L. R. 1975. Nutrient supply and the growth of the seminal root system in barley. II. Localized, compensatory increase in lateral root growth and rates of nitrate uptake when nitrate supply is restricted to only part of the root system. J. Exper. Bot. 26: 79-90.
During, H. J. & Lloret, E 1996. Permanent grid studies in bryophyte communities. 1. Pattern and dynamics of individual species. J. Hattori Bot. Lab. 79: 1-41.
Eriksson, O. 1986. Mobility and space capture in the stoloniferous plant Potentilla anserina. Oikos 46: 82-87.
Eriksson, O. & Jerling, L. 1990. Hierarchical selection and risk spreading in clonal plants. Pp. 79-94. In: van Groenendael J. & de Kroon H. (eds), Clonal growth in plants: regulation and function. SPB Academic Publishing, The Hague.
Evans, J. P. 1988. Nitrogen translocation in a clonal dune perennial, Hydrocotyle bonariensis. Oecologia 77: 64q58.
Evans, J. P. 1991. The effect of resource integration on fitness related traits in a clonal dune perennial, Hydroco~le bonariensis. Oeco- logia 86: 268-275.
Evans, J. P. 1992. The effect of local resource availability and clonal integration on ramet functional morphology in Hydrocotyle bon- ariensis. Oecologia 89: 265-276.
Evans, J. P. & Whitney, S. 1992. Clonal integration across a salt gradient by a non-halophyte, Hydrocolyle bonariensis (Api- aceae). Amer. J. Bot. 79: 1344-1347.
Fitter, A. H. 1994. Architecture and biomass allocation as compon- ents of the plastic response of root systems to soil heterogeneity. Pp. 305-323. In: Caldwell M. M. & Pearcy R. W. (eds), Exploit- ation of environmental heterogeneity by plants. Academic Press, London.
Fliervoet, L. M. & Werger, M. J. A. 1984. Canopy structure and microclimate of two wet grassland communities. New Phytol. 96:115-130.
Fowler, N. 1988. The effects of environmental heterogeneity in space and time on the regulation of populations and communities. Pp. 249-269. In: Davy A. J., Hutchings M. J. & Watkinson A. R. (eds), Plant population ecology. Blackwell Scientific Publica- tions, Oxford.
Friedman, D. & Alpert, P. 1991. Reciprocal transport between ramets increases growth of Fragaria chiloensis when light and nitrogen occur in separate patches but only if patches are rich. Oecologia 86: 76-80.
Gifford, E. M. & Foster, A. S. 1989. Morphology and evolution of vascular plants. Freeman & Co., New York.
Grime, J. P. 1979. Plant strategies and vegetation processes. Wiley & Sons, Chichester.
Grime, J. P., Crick, J. C. & Rincon, J. E. 1986. The ecological signi- ficance of plasticity. Pp. 4-29. In: Jennings D. H. & Trewavas A. J. (eds), Plasticity in plants. Cambridge University Press, Cam- bridge.
Gross, K. L., Pregitzer, K. S. & Burton, A. J. 1995. Spatial variation in nitrogen availability in three successional plant communities. J. Ecol. 83: 357-367.
Harper, J. L. 1985. Modules, branches, and the capture of resources. Pp. 1-33. In: Jackson J. B. C., Buss L. W. & Cook R. E. (eds), Population biology and evolution of clonal organisms. Yale Uni- versity Press, New Haven.
Hartnett, D. C. & Bazzaz, E A. 1983. Physiological integration among intraclonal ramets in Solidago canadensis. Ecology 64: 779-788.
Herben, T., During, H. J. & Krahulec, E 1995. Spatiotemporal dynamics in mountain grasslands: species autocorrelations in space and time. Folia Geobot. Phytotax. 30: 185-196.
Herben, T., Hara, T., Marshall, C. & Soukupov~i, L. 1994. Plant clonality: biology and diversity. Folia Geobot. Phytotax. 29: l 13- 122.
Hirose, T. & Werger, M. J. A. 1995. Canopy structure and photon flux partitioning among species in a herbaceous plant community. Ecology 76: 466-474.

Huber, H. 1996. Plasticity of internodes and petioles in prostrate and erect Potentilla species. Funct. Ecol. 10, in press.
Hutchings, M. J. & de Kroon, H. 1994. Foraging in plants: the role of morphological plasticity in resource acquisition. Adv. Ecol. Res. 25: 159-238.
Hutchings, M. J. & Slade, A. J. 1988. Morphological plasticity, foraging and integration in clonal perennial herbs. Pp. 83-109. In: Davy A. J., Hutchings M. J. & Watkinson A. R. (eds), Plant population biology. Blackwell Scientific Publications, Oxford.
Jackson, J. B. C., Buss, L. W. & Cook, R. E. (eds) 1985. Population biology and evolution of clonal organisms. Yale University Press, New Haven.
Jackson, R. B., Manwaring, J. H. & Caldwell, M. M. 1990. Rapid physiological adjustment of roots to localized soil enrichment. Nature 344: 58~60.
Jones, M. 1985. Modular demography and form in silver birch. Pp. 223-237. In: White, J. (ed), Studies on plant demography. A Festschrift for John L. Harper. Academic Press, London.
Jones, M. & Harper, J. L. 1987a. The influence of neighbours on the growth of trees. I. The demography of buds in Betula pendula. Proc. Roy. Soc. Lond., Ser. B, 232: 1-18.
Jones, M. & Harper, J. L. 1987a. The influence of neighbours on the growth of trees. II. The fate of buds on long and short shoots in Betula pendula. Proc. Roy. Soc. Lond., Series B, 232: 19-33.
J6nsd6ttir, I. S. 1989. The population dynamics, intraclonal physiology and grazing tolerance of Carex bigelowii. PhD Dis- sertation. Lund University, 86p.
J6nsd6ttir, I. S. & Callaghan, T. V. 1988. Interrelationships between different generations of interconnected tillers of Carex bigelowii. Oikos 52: 120-128.
J6nsd6ttir, I. S. & Callaghan, T. V. 1989. Localized defoliation stress and the movement of 14C-photoassimilates between tillers of Carex bigelowii. Oikos 54:211-219.
J6nsd6ttir, I. S. & Callaghan, T. V. 1990. Intraclonal translocation of ammonium and nitrate in Carex bigelowii using 15N and nitrate reductase assays. New Phytol. 114: 419-428.
Kelly, V. R. & Canham, C. D. 1992. Resource heterogeneity in oldfields. J. Veg. Sci. 3: 545-552.
Kemball, W. D. & Marshall, C. 1995. Clonal integration between parent and branch stolon in white clover: a developmental study. New Phytol. 129: 513-521.
Kolasa, J. & Rollo, C. D. 1991. The heterogeneity of heterogeneity: a glossary. Pp. 1-23. In: Kolasa, J. & Pickett, S. T. A. (eds), Ecological heterogeneity. Springer, New York.
Kotliar, N. B. & Wiens, J. A. 1990. Multiple scale of patchiness and patch structure: a hierarchical framework tbr the study of heterogeneity. Oikos 59: 253-260.
Lau, R. R. & Young, D. R. 1988. Influence of physiological integra- tion on survivorship and water relations in a clonal herb. Ecology 69: 215-219.
Lee, J. A. & Stewart, G. R. 1978. Ecological aspects of nitrogen assimilation. Adv. Bot. Res. 6: 1-43.
Levin, S. A. 1992. The problem of pattern and scale in ecology. Ecology 73: 1943-1967.
Li, H. & Reynolds, J. E 1994. A simulation experiment to quantify spatial heterogeneity in categorical maps. Ecology 75: 2446- 2455.
Li, H. & Reynolds, J. E 1995. On definition and quantification of heterogeneity. Oikos 73: 280-284.
Miller, R. E., Ver Hoef, J. M. & Fowler, N. L. 1995. Spatial hetero- geneity in eight central Texas grasslands. J. Ecol. 83: 919-928.
Marshall, C. 1990. Source-sink relations of interconnected ramets. Pp. 23-41. In: van Groenendael J. & de Kroon H. (eds), Clonal
69
growth in plants: regulation and function. SPB Academic Pub- lishing, The Hague.
Novoplansky, A., Cohen, D. & Sachs, T. 1990. How Portulaca seedlings avoid their neighbours. Oecologia 82: 490-493.
Oborny, B. 1994a. Growth rules in clonal plants and environmental predictability - a simulation study. J. Ecol. 82:341-351.
Oborny, B. 1994b. Spacer length in clonal plants and the efficiency of resource capture in heterogeneous environments: a Monte Carlo simulation. Folia Geobot. & Phytotax. 29: 139-158.
Oborny, B. & Podani, J. (eds) 1995. Clonality in plant communities. Opulus Press, Uppsala.
Pate, J. S. 1980. Transport and partitioning of nitrogenous solutes. Ann. Rev. PI. Phys. 31: 313-340.
Pate, J. S. 1983. Patterns of nitrogen metabolism in higher plants and their ecological significance. Pp. 225-255. In: Lee J. A., McNeill S. & Rorison I. H. (eds), Nitrogen as an ecological factor. Blackwell Scientific Publications, Oxford.
Pearcy, R. W. & Sims, D. A. 1994. Photosynthetic acclimation to changing light environments: scaling from the leaf to the whole plant. Pp. 145-174. In: Caldwell M. M. & Pearcy R. W. (eds), Exploitation of environmental heterogeneity by plants. Academic Press, London.
Pearcy, R. W., Chazdon, R. L., Gross, L. J. & Mott, K. A. 1994. Pho- tosynthetic utilization of sunflecks: a temporally patchy resource on a time scale of seconds to minutes. Pp. 175-208. In: Cald- well M. M. & Pearcy R. W. (eds), Exploitation of environmental heterogeneity by plants. Academic Press, London.
Peoples, M. B. & Gifford, R. M. 1990. Long-distance transport of nitrogen and carbon from sources to sinks in higher plants. Pp. 434-447. In: Dennis D. T. & Turpin D. H. (eds), Plant physiology, biochemistry and molecular biology. Longman, Hat- low.
Pitelka, L. E & Ashmun, J. W. 1985. Physiology and integration of ramets in clonal plants. Pp. 399-435. In: Jackson J. B. C., Buss L. W. & Cook R. E. (eds), Population biology and evolution of clonal organisms. Yale University Press, New Haven.
Price, E. A. C. & Hutchings, M. J. 1992a. The causes and develop- mental effects of integration and independence between different parts of Glechoma hederacea clones. Oikos 63: 376-386.
Price, E. A. C. & Hutchings, M. J. 1992b. Studies of growth in the clonal herb Glechoma hederacea. II. The effects of selective defoliation. J. Ecol. 80: 39-47.
Price, E. A. C., Marshall, C. & Hutchings, M. J. 1992. Studies of growth in the clonal herb Glechoma hederacea. I. Patterns of physiological integration. J. Ecol. 80: 25-38.
Robertson, G. E & Gross, K. L. 1994. Assessing the heterogeneity of belowground resources: quantifying pattern and scale. Pp. 237- 253. In: Caldwell M. M. & Pearcy R. W. (eds), Exploitation of environmental heterogeneity by plants. Academic Press, London.
Robinson, D. 1994. The response of plants to non-uniform supplies of nutrients. New Phytol. 127: 635~674.
Salzman, A. G. 1985. Habitat selection in a clonal plant. Science 228: 603-604.
Salzman, A. G. & Parker, M. A. 1985. Neighbors ameliorate local salinity stress for a rhizomatous plant in a heterogeneous envir- onment. Oecologia 65: 273-277.
Scheiner, S. M. 1993. Genetics and evolution of phenotypic plasti- city. Ann. Rev. Ecol. Syst. 24: 35~8 .
Schulze, E. -D. & Hall, A. E. 1981. Stomatal responses, water loss and CO2 assimilation rates of plants in contrasting environments. Encycl. PI. Phys., New Series, 12B: 181-230.
Shorrocks, B. & Swingland, R. (eds) 1990. Living in a patchy envir- onment. Oxford University Press, Oxford.

70
Shumway, S. W. 1995. Physiological integration among clonal ramets during invasion of disturbance patches in a New England salt marsh. Ann. Bot. 76: 225-233.
Silvertown, J. & Smith, B. 1988. Gaps in the canopy: the missing dimension in vegetation dynamics. Vegetatio 77: 57-60.
Silvertown, J. & Smith, B. 1989. Mapping the microenvironment for seed germination in the field. Ann. Bot. 63: 163-167.
Silvertown, J., Prince, S. D. & Smith, B. 1988. A field-portable instrument for mapping the micro environment within grass can- opies. Funct. Ecol. 2: 263-268.
Slade, A. J. & Hutchings, M. J. 1987a. An analysis of the costs and benefits of physiological integration between ramets in the clonal perennial herb Glechoma hederacea. Oecologia 73:425-431.
Slade, A. J. & Hutchings, M. J. 1987b. The effect of nutrient availab- ility on foraging in the clonal herb Glechoma hederacea. J. Ecol. 75:95-112.
Solangaarachchi, S. M. & Harper, J. L. 1989. The growth and asym- metry of neighbouring plants of white clover (Trifolium repens L.). Oecologia 78: 208-213.
Steeves, T. A. & Sussex, I. M. 1989. Patterns in plant development. 2nd edition. Cambridge University Press, Cambridge.
Strasburger, E. 1991. Lehrbuch der Botanik. 33rd edition. G. Fischer, Stuttgart.
Stuefer, J. E 1996. Separating the effects of assimilate and water integration in clonal fragments by the use of steam-girdling. Abstr. Bot. 19: 75-81.
Stuefer, J. E, de Kroon, H. & During, H. J. 1996. Exploitation of environmental heterogeneity by spatial division of labour in a clonal plant. Funct. Ecol. 10, in press.
Stuefer, J. E, During, H. J. & de Kroon, H. 1994. High benefits of clonal integration in two stoloniferous species, in response to heterogeneous light environments. J. Ecol. 82:511-518.
Stuefer, J. E & Hutchings, M. J. 1994. Environmental heterogeneity and clonal growth: a study of the capacity for reciprocal translo- cation in Glechoma hederacea L. Oecologia 100: 302-308.
Sutherland, W. J. & Stillman, R. A. i988. The foraging tactics of plants. Oikos 52: 239-244.
Tuomi, J. & Vuorisalo, T. 1989a. Hierarchical selection in modular organisms. Tr. Ecol. Evol. 4: 209-213.
Tuomi, J. & Vuorisalo, T. 1989b. What are the units of selection in modular organisms? Oikos 54: 227-233.
Upton, G. J. G. & Fingleton, B. 1985. Spatial data analysis by example. I. Point pattern and quantitative data. Wiley & Sons, Chichester.
Upton, G. J. G. & Fingleton, B. 1989. Spatial data analysis by example. II. Categorical and directional data. Wiley & Sons, Chichester.
van Groenendael, J. & de Kroon, H. (eds) 1990. Clonal growth in plants: regulation and function. SPB Academic Publishing, The Hague.
Waite, S. 1994. Field evidence of plastic growth responses to habitat heterogeneity in the clonal herb Ranunculus repens. Ecol. Res. 9: 311-316.
Watson, M. A. 1984. Developmental constraints: effect on popula- tion growth and patterns of resource allocation in a clonal plant. Amer. Nat. 123:411--426.
Watson, M. A. & Casper, B. B. 1984. Morphogenetic constraints on patterns of carbon distribution in plants. Ann. Rev. Ecol. Syst. 15: 233-258.
Wiens, J. A. 1976. Population responses to patchy environments. Ann. Rev. Ecol. Syst. 7: 81-120.
Wiens, J. A. 1989. Spatial scaling in ecology. Funct. Ecol. 3: 385- 397.
Wiens, J. A. 1990. On the use of 'grain' and 'grain size' in ecology. Funct. Ecol. 4: 720.
Wijesinghe, D. K. & Handel, S. N. 1994. Advantages of clonal growth in heterogeneous habitats: an experiment with Potentilla simplex. J. Ecol. 82: 495-502.
Young, D. R. & Smith, W. K. 1979. Infuence of sunflecks on the temperature and water relations of two subalpine understorey congeners. Oecologia 43: 195-205.
Young, D. R. & Smith, W. K. 1980. Influence of sunlight on pho- tosynthesis, water relations, and leaf structure in the understorey species Arnica cordifolia. Ecology 61 : 1380-1390.
Zimmermann, M. H. & Tomlinson, P. B. 1972. The vascular con- struction of monocotyledonous stems. Bot. Gaz. 133:141-155.




















