
The 1900 World's Fair or the Attraction of the Senses: The Case of the Maréorama

Posr-Pnleozorc Reen-BurLDrNc CNroenre
THE RECOVERY OF REEF-BUILDING COMMUNITIES AND THE ROLEOF CNIDARIANS IN CARBONATE SEQUENCES OF THE MIDDLE
TRIASSIC (ANISIA}T) IN THE ITALIAN DOLOMITES
ElrsenerrA Fors eNo Mnunrzro GRernNt
Dipartimento di Scienze della Terra, Milano, Italy
ABSTRACT
Late Anisian carbonate sequences in the Dolomites (Italy) give the best case history of the earlier Triassic "reeff" communitiesin the Southern Alps. Small (few cubic meters) mud mounds developed in a deep infralitîoral environment not far from islands.A pioneer community is made of Olangocoelia otti BechsÈrdt and Brandner growing on peloidal packstones, and of blue-greenalgae laminae and/or Tubiphytes obscuns Maslov developing on muddy substrata. Over the latter a second stage characterizedby abundant encrusting Sphinctozoan Celyphia zoldana Ott might develop. A third stage marked by more diversified communities(i.e., trepostomatid bryozoans, sphinctozoans/inozoans, porostromate blue-green algae, solenoporaceans) characterizes the fulldevelopment ofthe mounds. Scanty scleractinians appear at this stage, characterized by fairly high evolutionary complexity anddiversitv in contrast with the low densitv.
l 9 l
INTRODUCTION
The recovery of the "reef" building communitiesafter the Permo-Triassic crisis in the Southern Alpsoccured during the Anisian. In order to illustrate thosecommunities and the pattern ofthe recovery, a numberof Anisian units and sequences in the Southern Alpshave been studied. In situ mud mounds and bouldersembedded in the topmost Dont Formation, as well asthe very base of the Cernera Dolomite in the Mt. Cer-nera and Valdaora areas (Eastern Dolomites) are therichest and most diversified sites. The age is Late An-isian (Illyrian, Trinodosus Zone).
The main characters of the microfacies of the Val-daora area have been described by Bechstàdt andBrandner (1970). Other localized occurrences are incarbonate lenses in the Agordo Formation (Ott et al.,1980), and in the Upper Serla Formation (Pisa et al.,1979; Farabegoli and Guasti, I 980; Gaetani et al., l98l)in the Dolomites. Similar facies are also present in theUpper Serla Formation ofCarnia (Jadoul, pers. comm.).Consequently the present paper deals with the outcropswe consider more suitable for this case history.
ACKNOWLEDGMENTS
We are indebted to Profs. R. Boardman, N. FantiniSestini and G. Sîanley, and E. Ott for useful discussionson bryozoan, scleractinian and calcareous sponge tax-onomy, respectively; to Prof. W. Schlager for usefuldiscussions on the general model. Financial supportwas by C.N.R. CT.81.01982. Thin sections by C. Mal-inverno, photographs by G. Chiodi, drawings by S.Antico.
GEOLOGICAL SETTING
The paleogeographic setting ofthe Dolomites duringthe Anisian has been described by Assereto a al. (1977\.Land was located westward and possibly north in theform of islands, and deltas were intermittently spread-ing clastics on an open platform. Towards the begin-ning of the Trinodosus Zonet}l.e terrigenous input tem-porarily decreased and small micritic moundsdeveloped (Text-fig. l). The tectonic framework wasunstable and limited displacement ofthe mounds maybe noted. The decrease and end of the clastic supplyallowed the mud mounds to coalesce and form a con-tinuous subtidal carbonate bank (Cernera Dolomite;the informal name is introduced for the first Illyriancarbonate bank which grows onto the Dont Formationand is interrupted by the Richthofen Conglomerate(see Text-fig. l). Renewed uplifting caused an increasedterrigenous input from the neighboring islands(Richthofen Conglomerate). In the Valdaora area theuplifting also disrupted the Cernera Dolomite, andboulders from it and the topmost Dont Formation areembedded in breccias and conglomerates.
Consequently three different settings are present(Text-fig.2):
A) In si/a mud mounds (Text-fig. 2A). They formlenticular bodies varying in size from about I to l0cubic meters. Exceptionally several small lenses havebeen observed to coalesce forming a wall about 15 mhigh and 8-10 m wide. They have been interpreted asin situ mounds because: l) most of the organisms arein growth position; 2) geopetal infillings, including cav-ities within the organisms, are concordant with thepresent orientation ofthe outcrop; 3) basal and lateral

rt92 PelesoNrocRApHrcA AvrpnrcRNe, NuNrssn 54
contacts with the marly limestone of the Dont For-mation are gradual, the mound growth being preceded
by irregularly nodular facies (up fo l-2 m thick) madeof blue-green algae bindstones with calcareous spongesand solenoporaceans increasing upwards; the same
nodular facies characterizes the flanks of the mounds.The mounds normally develop on planar marly
limestone of the Dont Formation. which some 2 mbefore the true mound show a distinct increase in CaCO3due to the activity ofcalcareous blue-green algae andsubordinate calcareous sponges; the increasing bindingproduces first planar nodular limestone (outlined bydiagenetic processes) passing upwards to very irregu-larly nodular facies (Text-fig. 2A).
The mounds consist of several smaller micritic lenseswelded together and enveloped by veneers of marlylimestone, which frequently correspond to tiny patches
of different organisms. In the case of the biggest moundrecorded, among the smaller lenses are veneers of cri-noidal calcarenites, likely formed by in situ remains ofcrinoids which could attach onto the other organisms.A comparison to the present day Halimeda sands fill-ing the hollows in patch reefs might be suggested. Themound when fully developed must have reached a re-lief of at least I m over the sea bottom. They are cappedby crinoidal calcarenites (few meters) rapidly passing
into a thick sequence of marly limestone and marls ofthe topmost Dont Formation. The best preserved
mounds crop out in the Mt. Cernera area (Col Toront).In the Valdaora area the base of the Cernera Dolomiteis made of mud lenses welded together, separated bymarly veneers, displaying a binding community ingrowth position.
B) Displaced mud mounds (Text-fig. 2A-B). Theyconsist of lenticular carbonate bodies comparable insize and composition to the in silz mud mounds. Theyhave been interpreted as displaced mounds because of:I ) tilting of geopetal infillings in relation to the present
orientation of the mound; 2) presence of slump struc-tures in the basal and surrounding beds; 3) occasionaltilting of the internal bedding of the mound.
The degree ofdisplacement is variable; in some casesit consists of a gentle sliding of the mound on its flatbase, thus not affecting the original orientation, buttruncating the lateral connections (Text-fig. 2A top).More intense displacement may cause tilting, breakingup of the mounds and formation of swarms of mon-ogenic carbonate lenses (Text-fig. 2B). We do not con-sider them as boulders which slid from an active mar-gin ofthe Cernera Dolomite because the latter is younger
than the mounds and forms a tabular body. They cropout throughout the area examined.
C) Boulders (Text-fig. 2C). They are carbonate blocksup to some tens of cubic meters in size derived from
the erosion of the Cernera Dolomite and Contrin For-mation. They are interbedded in the topmost part ofthe Dont Formation associated with conglomeratic-arenitic layers announcing the Richthofen Conglom-erate event. They crop out in the area of Piz Tre Dita(Valdaora area) where no Illyrian carbonate bodies arerecorded in the sequence. As the Cernera Dolomitecrops out in the neighboring regions, the area is inter-preted as a paleofurrow into which the clastics werefunneled with erosion ofthe adjacent carbonate banks.
The three different settings yield similar microfaciesand may be considered as part of the same stage ofevolution of the recovery of the "reefv" communitiesin the Late Anisian.
Dobbraco
. ' C o r l r n : d A
o
oa A g o r d o
E e l l u n o
B l A g D o S e
Text-figure l.-Top: location map. Center: stratigraphic scheme;Ag : Agordo Fm.; Se : Upper Serla Fm.; Do : Dont Fm.; Ce:Cernera Dolomite; Ri : RichthoÈn Conglomerate; Co : ContrinFm. Capital letters A, B, C refer to the îhree settings of Anisianmounds and boulders illustrated in Text-fig. 2. Bottom: block-dia-gram showing the paleogeographic setting during the late Anisian.The mounds and the Cernera Dolomite developed on a continentalshelf polluted by clastics from the islands of the Western Dolomites.Bl : Bellerophon Fm.; We : Werlen Fm.; Ag : Agordo Fm.; Do =
Dont Fm.; Se : Upper Serla Fm.

IPosr-Pelrozorc Rrer-BurLDrNG CNmenra r93
Text-figure 2.-Mounds and boulders setting. l. Col Toront (W side of Mt. Cernera). The lower belt of mounds is in growth position,reaching a thickness greater than l0 m. Note the irregularly nodular basal part corresponding to stages I and II of the growth model. Communiîiesare well developed and diversified. In the topmost part the mounds appear to be stightly displaced, sliding for a short distance on its flat base;communities do not reach the highest diversification. .8. Col Vallaccia (Valdaora area) NW side. l,arge mounds are displaced and even tilted,originating from a neighboring area within the topmost part of the Dont Fm. They are interpreted as noî derived from the Cernera Dolomite,which forms the bank at the upper side of the sketch and is younger in age. Communities are dominated by Olangocoelia olli. C. NE side ofPiz Tre Dita 1:s""11on D of Bechstiidt and Brandner, 1970). Carbonate boulders are embedded in a marly-silty sequence which laterallysubstitutes for the Cernera Dolomite. Blocks dominated by the Olangocoelia community may be derived both from the topmost Donî Fm.,and from an active/passive margin of the Cernera Dolomite. Conglomeratic layers (Richthofen Conglomerate) increase upwards.
NATURE OF THE REEF COMMUNITIES
The organisms involved, notwithsîanding local vari-ations, form a single broad facies. An assemblage ofbuilding organisms in which blue-green algae, sphinc-tozoans and bryozoans play the main role has beendetected. Solenoporacean algae, inozoans,. vermetidgastropods, annelids, sessile foraminifers and sclerac-tinians are subordinate forms. Several, often exclusivecommunities dominated by one or two groups of or-ganisms, have been recognized, which develop at dif-ferent stages of the sediment colonization.
r. Olansocoelia C!f-""ttt
O. otti Bechstiidt & Brandner appears to form anexclusive community rarely associated with other or-ganisms (Celyphia zoldana Ott, spongiostromate lam-inae, Tubiphytes obscurus Maslov, a gglutinating worms)which if present are totally subordinate; sometimesthose forms are well developed too, but colonize ad-jacent but separate areas.
Growth morphology.-O. otti acted as sediment sta-bilizer and baffier. Its pearl necklace shaped skeleton
Text-figure 3.-Olangocoelia community. O. otri colonizes finepeloidal packstone creaîing irregular cavities with geopetal infrlling.Spongiostromate laminae are subordinate. Burrow at the topmostleft. Scale bar 5 mm. Thin section EF 173. Piz Tre Dita, Valdaora

r94 PeleeoNrocRApHrcA AvrsnrceNe. Nulasrn 54
with the wandering growth direction created many ir-regular cavities later infilled by peloidal packstone. Theproblematic sphinctozoan must have formed a fairlyrigid framework as most of the cavities show geopetalinfilling. Nevertheless rare burrowings in the sedimentsurrounding the sponge skeleton rule out early lithifi-cation of the whole rock. That community normallygrows on fine peloidal packstones, where it seems torepresent the first and last stage ofcolonization ofthesediment (Text-fig. I l, below).
Location.-The community is fairly widespread inthe area east of Piz da Peres (case C) where it is themain constituent of the smallest boulders. It is sub-ordinate west of Piz da Peres and in the Cernera areain the upper displaced mounds (case B).
2. Blue-Green Algae, Celyphia zoldana,Tubiphytes Community
(Text-figs. 4-5)
Blue-green algae (spongiostromate laminae and veryrare porostromate algae, i. e., C ladogirvanel/a-like tufts)and C. zoldana form irregular encrustinglayers; Tubi-phytes obscurus forms tiny erect tufts when not veryabundant while it grows as irregular nodules when itis a main constituent of the community. Locally oneof the organisms becomes predominant. Associatedforms are vermiform gastropods (Spirorbis sp.), agglu-tinating wonns and occasionally trepostomatid bryo-zoans, solenoporaceans, other sphinctozoans (Denin-geria-typ forms) and inozoans. Sessil foraminifers (i.e.,Alpinophragmium sp.) are secondary encrusters.
Text-figure 4.-Spongiostromate/Celyphia community. Spongiostromate laminae (A) create a network of irregularcavities with frequentgeopetal infilling; C. zoldana (B) grows onto the firm surfaces formed by the algal laminae. In the drawing, spongiostromate laminae are ingrey. Scale bar 3 mm. Thin section EF 170 c. Piz Tre Dita, Valdaora area.
Text-figure 5.-Spongiostromate community. Blue-green algae laminae create a framework of cavities with geopetal infrlling. Celyphiazoldana (A) and agglutinating worms (B) are secondary forms. In the drawing, spongiostromate laminae are in grey. Scale bar l0 mm. Thinsection EF I 55. Piz da Peres. Valdaora area.

G rowt h morp hology. - The spongiostromata andlorTubiphytes colonize mud substrata forming hard or atleast firm surfaces; they created a network ofirregularcavities on and inside which C. zoldana developed,intergrowing with following spongiostromate laminae.However occasionally, C. zoldana appears to be ableto grow directly on mud as well. The community cre-ated a framework with several tiny cavities (the largestconnected to spongiostromate laminae) in which thesediment was trapped as secondary geopetal infilling.The community is the pioneer colonizer of muddysubstrata and if following stages are present, it repre-sents the base the other organisms could grow on (Text-fig. I l). In the succeeding communities dominated by
Text-figure 6. - Trepostomatid bryozoan communiîy. Bryozoansgrow laterally and downward in a framework partially built andstrengthened by spongiostromate laminae. They are the preferentialsubstratum for boring bivalves (A) and secondary encrusters suchas Celyphia zoldana (B) and Ladinella porala (C). Scale bar 5 mm.Thin section EF 190. Piz da Peres. Valdaora area.
Posr-Paleozorc REEF-BurLDrNG CNroanle 1 9 5
other organisms (stage III) it helps strengthen theframework.
Location.-The community is common throughoutthe area wherever a muddy substratum develops. Itcontributes to form the base of mud mounds in theCernera area (case A); in the Valdaora area it normallyoccurs in boulders (case C) except for a mud lens crop-ping out in a gully under Piz da Peres (case B) and thebase of the Cernera Dolomite (case A).
3. Bryozoan-Rich Community(Text-fig. 6)
The bryozoans can either create an almost exclusivecommunity or are abundant components of a moreheterogeneous assemblage. All the specimens belongto a new genus and new species of Trepostomata. As-sociate forms are spongiostromate laminae, sphinc-tozoans (C. zoldana and new genus l), inozoans andsolenoporaceans. Secondary encrusters are Tubip hytesobscurus, vermiform gastropods, agglutinating wormsand Ladinella porata Ott.
Growth morphology.-The substratum the bryozo-ans grow on consists either of encrusting forms (i.e.,spongiostromates, sphinctozoans) or of scattered bio-clasts in muddy peloidal sediment (i.e., crinoid frag-ments, pelecypod shells, calcareous sponges). Theynormally become abundant in an advanced stage ofdevelopment of the building community. It appearsthat when they found a favorable environment, thebryozoans rapidly outpaced the other organisms, form-ing almost exclusive communities. The digitiform toplaty colonies grew laterally and downward in shelteredspaces (created by shells and the algal-sponge com-munity) presently very difficult to detect because theyare totally filled by sediment. On the basis of recent
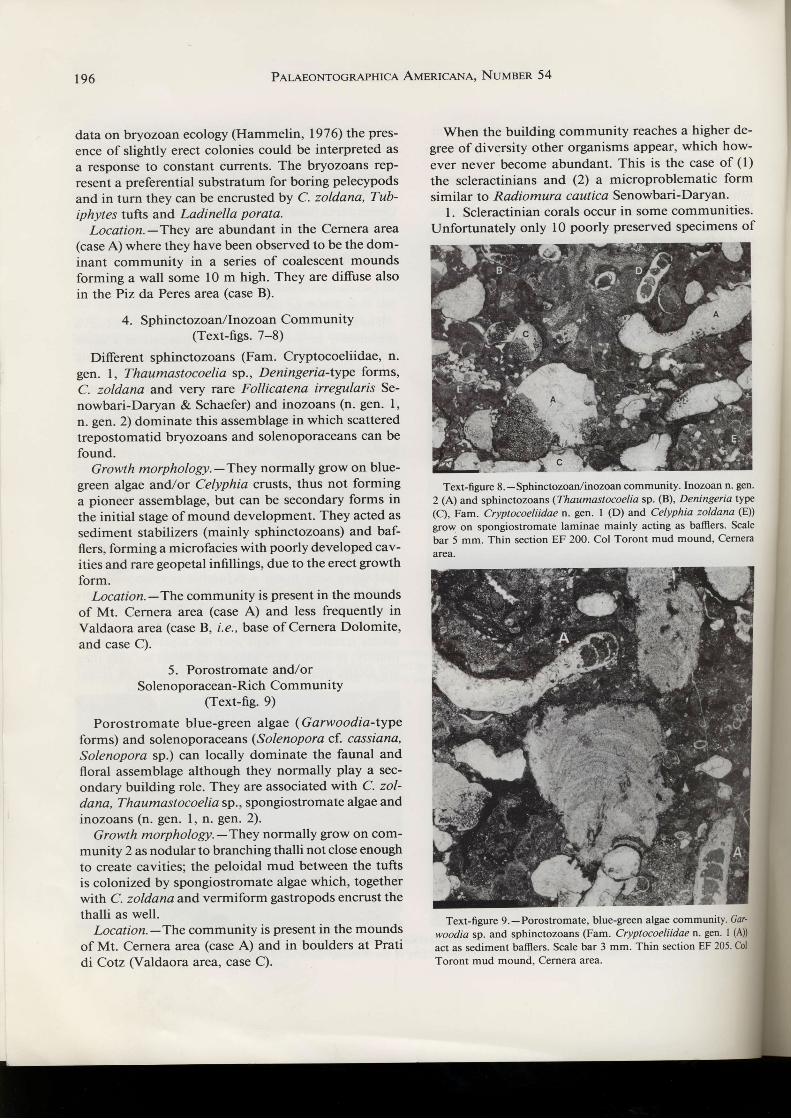
Text-figure 7.-Sphinctozoan/inozoan community. Sphinctozoans (Fam. Cryptocoeliidaen.gen. I (A), Celyphia zoldana(B)) and inozoans(n. gen. 2) (C)) grow in a framework of spongiosîromale laminae (in grey in the drawing). Scale bar 6 mm. Thin section EF 199. Col Torontmud mound. Cernera area.
r96 Pe,r-nEoNTocRAPHICA
data on bryozoan ecology (Hammelin, 1976) the pres-
ence of slightly erect colonies could be interpreted as
a response to constant currents. The bryozoans rep-
resent a preferential substratum for boring pelecypods
and in turn they can be encrusted by C. zoldana, Tub-
iphvtes tufts and Ladinella porata.
Location.-They are abundant in the Cernera area(case A) where they have been observed to be the dom-
inant community in a series of coalescent mounds
forming a wall some l0 m high. They are diffuse also
in the Piz da Peres area (case B).
4. Sphinctozoan/lnozoan Community(Text-figs. 7-8)
Different sphinctozoans (Fam. Cryptocoeliidae, n.gen. I, Thaumastocoelia sp., Deningeria-type forms,
C. zoldana and very rare Follicatena irregularls Se-
nowbari-Daryan & Schaefer) and inozoans (n. gen. l,
n. gen. 2) dominate this assemblage in which scattered
trepostomatid bryozoans and solenoporaceans can be
found.Growth morphology. -They normally grow on blue-
green algae and/or Celyphia crusts, thus not forming
a pioneer assemblage, but can be secondary forms in
the initial stage of mound development. They acted as
sediment stabilizers (mainly sphinctozoans) and baf-
flers, forming a microfacies with poorly developed cav-ities and rare geopetal infillings, due to the erect growth
form.Location.-The community is present in the mounds
of Mt. Cernera area (case A) and less frequently in
Valdaora area (case B, i.e., base of Cernera Dolomite,
and case C).
5. Porostromate and/orSolenoporacean-Rich CommunitY
(Text-fig. 9)
Porostromate blue-green algae (Garwoodia-type
forms) and solenoporaceans (Solenopora cf. cassiana,
Solenopora sp.) can locally dominate the faunal and
floral assemblage although they normally play a sec-
ondary building role. They are associated with C. zol-
dana, Thaumastocoelia sp., spongiostromate algae and
inozoans (n. gen. l, n. gen. 2).Growth morphology. -They normally grow on com-
munity 2 as nodular to branching thalli not close enough
to create cavities; the peloidal mud between the tufts
is colonized by spongiostromate algae which, togetherwith C. zoldana and vermiform gastropods encrust the
thall i as well.Location.-The community is present in the mounds
of Mt. Cernera area (case A) and in boulders at Prati
di Cotz (Valdaora area, case C).
AvreprcRNR, Nuvssn 54
When the building community reaches a higher de-
gree of diversity other organisms appear, which how-
ever never become abundant. This is the case of (l)
the scleractinians and (2) a microproblematic form
similar fo Radiomura cautica Senowbari-Daryan.l. Scleractinian corals occur in some communities.
Unfortunately only l0 poorly preserved specimens of
Text-figure 8.-Sphinctozoan/inozoan community. Inozoan n. gen'
2 (A) and sphinctozoans (Thaumastocoelic sp. (B), Deningeria fype
(C), Fam. Cryptocoeliidae n. gen. I (D) and Celyphia zoldana (E))
grow on spongiostromate laminae mainly acting as baffiers. Scale
bar 5 mm. Thin section EF 200. Col Toronl mud mound, Cernera
afea.
Text-figure 9.-Porostromale, blue-green algae community. Gar-
woodia sp. and sphinctozoans (Fam. Cryptocoeliidae n. gen. t (A))
act as sediment baffiers. Scale bar 3 mm. Thin section EF 205. Col
Toront mud mound, Cernera area.

scleractinian colonies have been found. One specimenis possibly related to the group of Isastrea profundaReuss (Text-fig. l0left) while other small massive col-onies show striking similarities to the genus Cassi-anastrea Voltz (Text-fig. l0 center). Two other speci-mens are referable to the "Thecosmilia" group,belonging to two species and possibly different generatoo, based on the difference of wall organization.
They appear when the building community is welldifferentiated and form tiny (hand size) colonies whichgrow in sheltered spaces among the previously de-scribed communities. Thus they do not play a primaryrole in the building of the mud mound, being more asguests, settling only when the environment is partic-ularly suitable and protected niches have been createdby other organisms.
Corals have been found only in the Cernera area andthey are generally different from the forms quoted inBechstàdt and Brandner (1970) as Calamophyllia sp.,cf. Thecosmilia sp. except for the form referred to astabulate incertae sedls. Specimens must be rare as wesearched at length in the localities sampled in Valdaoraarea, but no other corals were obtained.
Huge coral colonies (meter size) have been found inthe Contrin Formation (Latest Anisian) in the Mar-molada area; they show strong similarities with The-cosmilia cyclica Senowbari-Daryan & Schaefer in themorphology of septa and dissepiments with ring-likearrangement, the thick wall and the general size ofcorallites which average 7 mm diameter (Text-fig. l0right).
The density of scleractinians is very low; howeverthe diversity is fairly high, with 5 genera and at least4 families.
Posr-Pelsozorc Reer'-BurLDrNG CNroRnre 197
2. The problematic microorganism similar to Ra-diomura cautica can create very rare communities ap-pearing in a later stage of the mound development. Itis associated with rare C. zoldana and grows on mudforming a network of tiny cavities in which the sedi-ment settled creating geopetal infillings. It has beenfound only in the Cernera area (case A).
REEF COMMUNITY GROWTH MODEL ANDMOUND DEVELOPMENT
Field observations and microfacies analysis suggestthe following interpretation for the Anisian "reefy"
community.l. The substratum the building community grows
on consists of muddy peloidal sediment (wackestoneversus packstone). The sand size sediment (bioclastic/grapestone packstone versus grainstone) is never col-onized by organic communities possibly because it wastoo mobile to offer a suitable substratum and poor innutrients. In contrast modern examples of non-skeletalblue-green algae growing on muddy bottom in FloridaBay and the Bahamas suggest colonization of muddysubstrata by blue-green algae is not unusual; moreoverthe Anisian Spongiostromata were calcifying forms,thus rapidly forming a firm substratum for hard bot-tom organisms.
These patterns contrast with the development ofbet-ter known mud mounds (Ordovician-Silurian) whichoften grow on biocalcarenites (Pitcher, 1964; Alber-stadt et al., 19741, Crowley, 1973) nevertheless growthon muddy sediment has occasionally been recorded(Scoffin, 197 l). However, it is worth remembering thatthe Anisian pioneer community is mainly made ofblue-green algae while the Middle Paleozoic ones con-
Text-figure 10.-Scleractinian corals. l-eft: colony referable to the Isastrea profunda group; thin section EF 203, Col Toront mud mound,Cernera area. Center: colony showing striking similarities to the genus Cassianastea; thin section EF 202, Col Toront mud mound, Cerneraarea. Right: colony showing similarities with Thecosmilia cyclica for the ringJike arrangement of septa; thin section G 1414, Contrin Fm.,Marmolada area. Scale bars 2 mm.

1 9 8
sist oftabulate, stromatoporoids, and bryozoans which
seemed to prefer coarser and above all firmer substrata.
This interpretation is supported also for the Anisian
bryozoans and corals that grew either on algallspongecnrsts or on single bioclasts, i.e., a frlm bottom.
2. No evidence ofgrowth on identifiable topograph-ic highs has been detected but growth on flat bottoms,
not protected from moderate mud input, did not pre-
vent the algal growth as previously discussed.3. Growth sequence. A model of growth for the
"reefy" community has been hypothesized on the basis
of observations of mud mounds (Cernera and Piz da
Peres areas case A and B) and boulders (Piz Tre Dita-
Col Vallaccia area, case B and C) yielding similar fau-
nal and floral assemblages.Stage I.-A first stage of sediment colonization has
been detected in which two different communities can
develop. A. Olangocoelia otti stabilizes fine peloidal
packstones and creates an irregular framework with
several cavities. Secondary encrusting forms such as
blue-green algae, rare Celyphia zoldana and Tubi-phytes contibute in strengthening the framework. This
community has never been observed to form the sub-
stratum for the succeeding more diversified assem-
blages. However scattered O. otti may be present in
other communities. (Text-fig. I lA). B. Blue-green al-gae and/or Tubiphytes nodular masses start binding
the muddy sediment and form hard, at least rubbery
irregular crusts which create a framework with several
cavities. This community is generally strictly con-
nected to the development of stage II (Text-fig. I lB).
Stage II.-It is marked by the appearance, closely
related to stage IB, of encrusting sphinctozoans (i.e.,
C. zoldana) which are interlayered with spongiostro-mates. The second stage is characterized by the de-
velopment of a true framework in which the mud is
trapped mainly as secondary infilling often with geo-
petal structures. Secondary encrusters contribute to
build the framework. Scattered different organisms such
as bryozoans, inozoans and other sphinctozoans can
be present but always subordinate. Stages IB and II
form irregularly nodular facies which create the base
and the flanks of the lenticular mounds.Stage lll.-Diverse and often mutually exclusive
groups of organisms (i.e., sphinctozoan/inozoan com-
munity, porostromate blue-green algae and soleno-poracean community, bryozoan community, corals)
develop on a substratum formed during the previous
stages. Depending on the particular growth morphol-
ogy of the organism involved (erect tufts for soleno-poraceans, sphinctozoans and sometimes bryozoans
too, platy thickets for inozoans and bryozoans) either
a framework or a sequence of crusts can form. In both
cases blue-green algae laminae and sometimes Cely-
PelesoNrocRApHIcA AlasRIceNe, NuNasrn 54
phia too are still growing between and on the other
organisms. Secondary encrusting forms such as Ladi-
nella porata, tiny Tubiphytes tufts and Vermetids,
commonly grow onto the other organisms. No bur-
rowers have been detected, while several borings,mainly affecting the bryozoan colonies, testify that the
organic framework was not rapidly covered by sedi-
ment (Text-fig. I l). No early cementation has been
detected although the presence of micritic cement can-
not be excluded.4. The mound attained some relief (0.50-l m, ex-
ceptionally more) over the bottom which prevented it
from being covered by the constant input of sediments.However, the reliefwas not enough to protect the com-
munity from larger inputs of onlapping sandy calcar-
enites, and of marly limestone and marl of the topmost
Dont Formation.5. Death ofthe mound. Data are obviously obtained
from the analysis of case A. Apparently the growth of
the mound is intern-rpted by inputs of sand size sedi-
ment, either crinoidal packstones/grainstones or siltypackstones, thus confirming that calcarenites do not
represent a suitable substratum for the reef commu-
nity. On the contrary a moderate input of mud does
not definitely intemrpt the mound development. The
C o r a l s
B r y o z o a n s
S o l e n o p o r a c e a n s
I n o z o a n s
O t h e r S p h i n c t o z o a n s
C e l y p h i a
O l a n g o c o e l i aB l u e g r e e n a l g a l l a m i n a ea n d T u b i p h y t e s . J - / t
P e l e c y p o d s
B o r i n g s
C a v i t i e s w i l hg e o p e t a li n f i l l i n g
J 6
r!
@Ms@qp
afq , €
g
,é
Text-frgure I I . - Mud mound growth model. A pioneer stage (stage
I) is recognizable in which either Olangocoelia otti (A) or spongio'
stromate laminae and Tubiphytes obscurus(B) stabilize the sediment.
O. otti seems to prefer fine peloidal packstones and has never been
observed to create the substratum for succeeding communities. It
creates a neîwork ofcavities with geopetal infilling. In the case B, a
second and third stage might be present growing on stage I. Stage II
is characterized by the appearance of the encrusting sphinctozoan
Celyphia zoldana which, together with the spongiostromate laminae,
create a true framework. Stage III is marked by a higher diversity,
with communities of bryozoans, porostromate blue-green algae, so-
lenoporaceans, sphinctozoans/inozoans and scattered scleractinian
corals.

Posr-Pelsozorc REEF-BurLDrNG CNroenra r99
interpretation is strengthened by the absence ofcolo-nization in sequences made ofbiocalcarenites through-out (southern side of Mt. Cernera).
6. On the basis of the following elements a deepinfralittoral environment is inferred for the Anisianmounds. l) No evidence that the mounds were growingin a protected area, thus the mud is related to fairlydeep water. 2) Absence of dasycladacean algae whichare present in under- and overlying units of similarlithology (marly limestone of the Agordo Formation :Algenwellenkalk of Bechstiidt and Brandner, 1970; siltylimestone of Calcari di Pontebba, Fois and Jadoul,1983). 3) Community morphology and organismsgrowth form. Some similarities with the Squamari-acean facies of the "Biocenose du Détritique cótier"of the present Mediterranean at the limit between in-fra- and circalittoral zones (Perès and Picard, 1964)have been noticed. Obviously the various roles in thecommunity are played by different organisms but inboth cases an initial mud encrusting stage is present.Blue-green algae and recent squamariaceans crusts andblades form special microenvironments for the suc-ceeding cryptic and epibiontic communities (red algae,bryozoans). Among the Anisian organisms the bryo-zoan growth morphology (erect to platy colonies) mightbe indicative ofan environment dominated by currentsbut with mud input still present (Hammelin, 1976). 4)Parallel laminations in the biocalcarenites indicate cur-rent influence.
On the other side, the fairly high calcium carbonateproductivity and the general trend leading upwards toa continuous carbonate bank (Cernera Dolomite), sug-gest a depth not higher than 30 m, possibly 50 m.
DISCUSSION OF THE SIGNIFICANCEOF THE SCLERACTINIANS
In the Late Anisian mud mounds of the SouthernAlps, scleractinians play a subordinate building role.In fact only a few specimens confined to a few areasand appearing in the most diversified communitieshave been found. Despite the low number, the coralassemblage shows surprisingly high diversity and evo-lutionary advancement. At present five families, sevengenera and ten species have been detected also on thebasis of similar assemblages reported from the Sturialimestone of the Recoaro area, which is time-equiva-lent to the sequence of the Dolomites (Fliigel, 196l;De Zanche and Mietto, l98l).
Although they are the oldest scleractinians found inthe Southern Alps up to now, the high diversity sug-gests the existence of an environmentally optimal areapossibly located far eastward along the Tethys. Fromthat area the coral planulae occasionally reached theDolomites, situated at the "cul-de-sac" of the Tethyan
seaways. The distance from the source area was a toolfor selection, explaining the striking ratio of density todiversity.
The coral occumence which is confrned to few areas,and the low density suggest that most of the Dolomiteswere too polluted by clastics and had very muddy sub-strata. Moreover other groups such as blue-green algae,calcareous sponges and bryozoans still were able to outcompete the scleractinians.
The same scenery persists during the Ladinian andonly in the Carnian the role of scleractinians in thereef-building communities becomes more important(Fùrsich and Wendt, 1977).
CONCLUSIONS
A fairly homogeneous "ree$" community has beenidentified in the Anisian of the Dolomites (SouthernAlps, Italy). At the moment similar microfacies havenot been detected in coeval units outcropping west-wards (Lombardian Alps) possibly because the latterarea was more protected and muddy. In the Dolomites,constant characters of the building community havebeen detected in different Upper Anisian units (Ott eral., l98O; Farabegoli and Guasti, 1980; Bechstàdt andBrandner, 1970). The community consists ofblue-greenalgal laminae and encrusting sphinctozoans playing thepioneer role, followed by sphinctozoans, inozoans, tre-postomatid bryozoans, porostromate blue-green algae,solenoporaceans and rare scleractinians, together withsubordinate encrusters (vermetids, microproblemat-ics, sessile foraminifers). Corals are thought to havereached the area occasionally, drifted as planulae froma climax area situated far away.
Distinctive elements which differentiate the UpperAnisian building assemblage from the topmost Ani-sian/Lower Ladinian one (Gaetani et al., 1981) are:the encrusting sphinctozoan Celyphia zoldana, theproblematic sphinctozoan Olangocoelia otti and thetrepostomatid bryozoans. On the whole the Anisiancommunity is not less differentiated than the Ladinianone; it is just smaller in the number of specimens, lesswidespread, and appears to be able to create small mudmounds in a deep infralittoral environment.
REFERENCES CITED
Alberstadt, L. P., Walker, K. R., and Zurawski, R. P.1974. Patch reefs in the Carters Limestone (Middle Ordovician)
in Tennessee and vertical zonation in Ordovician reefs.Geol. Soc. America Bull., vol. 85, pp. I l7l-l 182.
Assereto, R., Brusca, C., Gaetani, M., and Jadoul, F.1977. Le mineralizzazioni Pb-Zn nel Trias delle Dolomiti.
Quadro geologico e interpretazione genetica. Industria Mi-neraria, vol. 28, no. 66, pp. 367-402.

200 PeI-aeor.rrocRApHrcA ArrlenrcnNn. NuÀasrn 54
Bechstiidt T., and Brandner, R.1970. Das Anis zwischen Sr. Vigil und dem Hóhlensteintal
(Pragser und Olanger Dolomiten, Síidtyro\. Festband.Geol. Inst., 300 Jahr Feier Univ., pp. 9-103, l8 pls., 3maps.
Crowley, D. J.1973. Middle Silurian patch reefs in Gasport Member (Lockport
Fm.), New York. Am. Assoc. Pet. Geologists Bull., vol.s7, pp. 283-300.
De Zanche, V., and Mietto, P.1981. Review ofthe Triassic sequence ofRecoaro (Italy) and re-
lated problems. Rend. Soc. Geol. It., vol. 4, pp.25-28.
Farabegoli, E., and Guasti, M.1980. Anisian lithostratigraphy and paleogeography of Mt. Rite
(Cadore, SE Dolomites). Riv. Ital. Paleont. Strat., vol. 85,no. 3-4, pp. 909-930.
Flúgel, E.1961. Eine Koralle aus dem schwarzen Sturia-Kalk (Aniil des
Tretto (Vicentin). Yerh. Geol. Bundesanst., vol. l-3, pp.79-83.
Fois, E., and Jadoul, F.1983. La Dorsale Paleocarnica anisica di Pontebba. Riv. Ital.
Paleont. Strat., vol. 89, no. I, pp.3-29.
Fùrsich, F. T., and Wendt J.1977. Biostratinomy and paleoecology of the Cassian Fm.
(Triassic) of the Southern Alps. Palaeogeogr. Palaeoclim.Palaeoecol., vol. 22, pp. 257 -323.
Gaetani, M., Fois, E., Jadoul, F., and Nicorao A.198 l. Nature and evolution of Middle Triassic carbonate build-
ups in the Dolomites (Italy). Iù,.lariîe Geol., vol. 44, pp.25-57.
Hammelin, J. G.1976. I* Sous-Ordre des Tubuliporina (Bryozoaires Cyclo-
stomes) en Méditerranée. Ecologie et sistématique. Iùfem.Inst. Ocean., no. 10, pp. l-326.
Ott, E., Pisa, G., and Farabegoli, E.1980. Celyphia zoldana sp. n., a reef building Sphinctozoan
sponge in Anisian Limestones of the Southeastern Dolo-mites. Riv.Ital. Paleont. Strat., vol. 85, no. 3-4, pp. 829-842.
Perès, J. M., and Picard, J.1964. Nouveau manuel de bionomie bentique de la Mer Médi-
terranée. Rec. Trav. St. Mar. Endoume, vol. 31, no. 47,pp. 5-137.
Pisq G., Farabegoli, E., and Ott E.1979. Stratigrafa e paleogeografia dei terreni anisici della conca
di Agordo e dell'alta Val di Zoldo (Dolomiti Sudorientali).Mem. Soc. Geol. It., vol. 18, pp.63-92.
Pitcher. M.1964. Evolution of Chazyan (Ordovician) reefs of eastern United
States and Canada. Bull. Canadian Petroleum Geology,vol. 12, pp.632-691.
Scoffin, T. P.1971. The conditions of growth of the Wenlock reefs of Shrop-
shire (England). Sedimentology, vol. 17, pp. 173-219.
Copyright © 2022 FDOKUMEN




















