Polyphenol rich botanicals used as food supplements interfere with EphA2-ephrinA1 system
7
Pharmacological Research 64 (2011) 464–470 Contents lists available at ScienceDirect Pharmacological Research journa l h o me pa ge: www.elsevier.com/locate/yphr s Polyphenol rich botanicals used as food supplements interfere with EphA2–ephrinA1 system Iftiin Hassan Mohamed a,1 , Carmine Giorgio a,1 , Renato Bruni b,∗ , Lisa Flammini a , Elisabetta Barocelli a , Damiano Rossi c , Giuseppe Domenichini a , Ferruccio Poli d , Massimiliano Tognolini a a Dipartimento di Scienze Farmacologiche, Biologiche e Chimiche Applicate, Università di Parma, Viale G.P. Usberti 27a, 43124 Parma, Italy b Dipartimento di Biologia Evolutiva e Funzionale, Università di Parma, Viale G.P. Usberti 11a, 43124 Parma, Italy c Dipartimento di Biologia ed Evoluzione, Sez. Risorse Agrotecnologiche e Farmaceutiche, AgriUnife, Università degli Studi di Ferrara, C.so Ercole I d’Este 32, 44121 Ferrara, Italy d Dipartimento di Biologia Evolutiva e Sperimentale, Università di Bologna, Via Irnerio 42, 40126 Bologna, Italy a r t i c l e i n f o Article history: Received 4 April 2011 Received in revised form 7 June 2011 Accepted 10 June 2011 Keywords: Eph kinases Ephrin Dietary phytochemicals Herbal supplements Cancer Screening a b s t r a c t The Eph tyrosine kinase receptors and their ephrin ligands play a central role in several human cancers and their deregulated expression or function promotes tumorigenesis, inducing aggressive tumor phe- notypes. Green tea extracts (GTE) have been recently found to inhibit Eph-kinase phosphorylation. In order to evaluate the potential contribution of edible and medicinal plants on EphA2–ephrinA1 modula- tion, 133 commercially available plant extracts used as food supplements, essential and fixed oils were screened with an ELISA-based binding assay. Nine plant extracts, rich of polyphenols, reversibly inhibited binding in a dose-dependent manner (IC 50 0.83–24 g/ml). Functional studies on PC3 prostate adeno- carcinoma cells revealed that active extracts antagonized ephrinA1-Fc-induced EphA2-phosphorylation at non-cytotoxic concentrations (IC 50 0.31–11.3 g/ml) without interfering with EGF-induced EGFR acti- vation, suggesting a specific effect. These findings could furnish an interesting starting point regarding the potential relationship between diet, edible plant secondary metabolites and Eph–ephrin system, suggesting their possible involvement in cancer development modulation. © 2011 Elsevier Ltd. All rights reserved. 1. Introduction A number of human epidemiological studies have outlined an inverse correlation between the adherence to specific diets, the consumption of fruits, vegetables, whole foods, edible medici- nal plants, herbs or spices and various human chronic diseases including oral, prostate, liver, colon, lung, stomach, bladder and breast cancers [1–4]. Notwithstanding these evidences, a conclu- sive and unambiguous pattern has not yet emerged regarding the mechanisms involved in food-related reduction of cancer risk development. It is now increasingly evident that the sole actions of antioxidants and anti-inflammatory nutrients, initially credited as preeminent, are not sufficient to explain the advantages ascribed to dietary habits rich in phytochemicals. In fact, no clear and direct correlation between these activities and cancer prevention has been established in large cohort studies or in meta analyses, thus leading to hypothesize that further mechanisms may be involved [5–8]. At present, the effects of dietary plant secondary metabo- lites on human health are not completely elucidated and it is ∗ Corresponding author. Tel.: +39 0521 906004; fax: +39 0521 905403. E-mail address: [email protected] (R. Bruni). 1 These authors contributed equally to this work. likely the existence of multiple mechanisms by which this highly chemodiverse array of substances elicits a protective effect against cancer development. In order to define the potential contribution of phytochemicals, multiple hypothesis are under investigation (proper modulation of signal transduction pathways, inhibition of cell proliferation, induction of apoptosis, inhibition of endogenous nitrosation and antimutagenic activity among others). In the quest for relevant mechanisms involved in this multifaceted process, fur- ther hypotheses may be worth an evaluation, including systems involved in angiogenesis and metastasis like Eph–ephrin system [9–11]. With 16 members arranged into 2 classes (A and B) according to sequence homology of extracellular domain and to their affinity for ligands, Eph-receptors constitute the largest family of recep- tor tyrosine kinases. Ephrins-A are glycosylphosphatidyl-inositol (GPI)-linked proteins anchored to cell membrane and preferentially bind to EphA receptors, while ephrins B are transmembrane pro- teins that preferentially bind to EphB receptors. Upon Eph–ephrin interaction a bidirectional signaling is triggered: a forward signals into the cells expressing Eph receptors go along with reverse signals into the cells bearing ephrin ligand [12]. This system is involved in multiple morphogenetic processes as embryonic patterning, gastrulation, segmentation, axonal path finding, neural crest cell migration and topographic organization of 1043-6618/$ – see front matter © 2011 Elsevier Ltd. All rights reserved. doi:10.1016/j.phrs.2011.06.008
Transcript of Polyphenol rich botanicals used as food supplements interfere with EphA2-ephrinA1 system

PE
IDa
b
c
d
a
ARRA
KEEDHCS
1
icnibstdaptcbl[l
1d
Pharmacological Research 64 (2011) 464– 470
Contents lists available at ScienceDirect
Pharmacological Research
journa l h o me pa ge: www.elsev ier .com/ locate /yphr s
olyphenol rich botanicals used as food supplements interfere withphA2–ephrinA1 system
ftiin Hassan Mohameda,1, Carmine Giorgioa,1, Renato Brunib,∗, Lisa Flamminia, Elisabetta Barocelli a,amiano Rossi c, Giuseppe Domenichinia, Ferruccio Polid, Massimiliano Tognolinia
Dipartimento di Scienze Farmacologiche, Biologiche e Chimiche Applicate, Università di Parma, Viale G.P. Usberti 27a, 43124 Parma, ItalyDipartimento di Biologia Evolutiva e Funzionale, Università di Parma, Viale G.P. Usberti 11a, 43124 Parma, ItalyDipartimento di Biologia ed Evoluzione, Sez. Risorse Agrotecnologiche e Farmaceutiche, AgriUnife, Università degli Studi di Ferrara, C.so Ercole I d’Este 32, 44121 Ferrara, ItalyDipartimento di Biologia Evolutiva e Sperimentale, Università di Bologna, Via Irnerio 42, 40126 Bologna, Italy
r t i c l e i n f o
rticle history:eceived 4 April 2011eceived in revised form 7 June 2011ccepted 10 June 2011
eywords:ph kinases
a b s t r a c t
The Eph tyrosine kinase receptors and their ephrin ligands play a central role in several human cancersand their deregulated expression or function promotes tumorigenesis, inducing aggressive tumor phe-notypes. Green tea extracts (GTE) have been recently found to inhibit Eph-kinase phosphorylation. Inorder to evaluate the potential contribution of edible and medicinal plants on EphA2–ephrinA1 modula-tion, 133 commercially available plant extracts used as food supplements, essential and fixed oils werescreened with an ELISA-based binding assay. Nine plant extracts, rich of polyphenols, reversibly inhibited
phrinietary phytochemicalserbal supplementsancercreening
binding in a dose-dependent manner (IC50 0.83–24 �g/ml). Functional studies on PC3 prostate adeno-carcinoma cells revealed that active extracts antagonized ephrinA1-Fc-induced EphA2-phosphorylationat non-cytotoxic concentrations (IC50 0.31–11.3 �g/ml) without interfering with EGF-induced EGFR acti-vation, suggesting a specific effect. These findings could furnish an interesting starting point regardingthe potential relationship between diet, edible plant secondary metabolites and Eph–ephrin system,suggesting their possible involvement in cancer development modulation.
. Introduction
A number of human epidemiological studies have outlined annverse correlation between the adherence to specific diets, theonsumption of fruits, vegetables, whole foods, edible medici-al plants, herbs or spices and various human chronic diseases
ncluding oral, prostate, liver, colon, lung, stomach, bladder andreast cancers [1–4]. Notwithstanding these evidences, a conclu-ive and unambiguous pattern has not yet emerged regardinghe mechanisms involved in food-related reduction of cancer riskevelopment. It is now increasingly evident that the sole actions ofntioxidants and anti-inflammatory nutrients, initially credited asreeminent, are not sufficient to explain the advantages ascribedo dietary habits rich in phytochemicals. In fact, no clear and directorrelation between these activities and cancer prevention haseen established in large cohort studies or in meta analyses, thus
eading to hypothesize that further mechanisms may be involved5–8]. At present, the effects of dietary plant secondary metabo-ites on human health are not completely elucidated and it is
∗ Corresponding author. Tel.: +39 0521 906004; fax: +39 0521 905403.E-mail address: [email protected] (R. Bruni).
1 These authors contributed equally to this work.
043-6618/$ – see front matter © 2011 Elsevier Ltd. All rights reserved.oi:10.1016/j.phrs.2011.06.008
© 2011 Elsevier Ltd. All rights reserved.
likely the existence of multiple mechanisms by which this highlychemodiverse array of substances elicits a protective effect againstcancer development. In order to define the potential contributionof phytochemicals, multiple hypothesis are under investigation(proper modulation of signal transduction pathways, inhibition ofcell proliferation, induction of apoptosis, inhibition of endogenousnitrosation and antimutagenic activity among others). In the questfor relevant mechanisms involved in this multifaceted process, fur-ther hypotheses may be worth an evaluation, including systemsinvolved in angiogenesis and metastasis like Eph–ephrin system[9–11].
With 16 members arranged into 2 classes (A and B) accordingto sequence homology of extracellular domain and to their affinityfor ligands, Eph-receptors constitute the largest family of recep-tor tyrosine kinases. Ephrins-A are glycosylphosphatidyl-inositol(GPI)-linked proteins anchored to cell membrane and preferentiallybind to EphA receptors, while ephrins B are transmembrane pro-teins that preferentially bind to EphB receptors. Upon Eph–ephrininteraction a bidirectional signaling is triggered: a forward signalsinto the cells expressing Eph receptors go along with reverse signals
into the cells bearing ephrin ligand [12].This system is involved in multiple morphogenetic processesas embryonic patterning, gastrulation, segmentation, axonal pathfinding, neural crest cell migration and topographic organization of

acolog
tamtfaooegsctoisgiTpoadwsat[
iiluwtpipgf[wmlAenptasppmbom
2
2
RCM
I. Hassan Mohamed et al. / Pharm
issues. Further investigations have highlighted Eph–ephrin systems a crucial and rate-limiting determinant also in the physiologicalodulation of angiogenesis. Finally, increasing evidences suggest
hat Eph–ephrin system plays a critical role in tumor and vascularunctions during carcinogenesis, while its deregulated expressionnd/or function may promote tumorigenesis and the developmentf more aggressive and metastatic phenotypes in a large varietyf human cancers [11,13]. In particular, EphA2 kinase is over-xpressed in several cancer types, including those affecting theastrointestinal tract and its levels have been correlated with tumortage and progression [14–18]. Unfortunately, it is not completelylear whenever the activation or inhibition of Eph–ephrin sys-em forward or reverse signaling, could have tumor suppressingr tumor enhancing effect. For instance, EphA2 forward signal-ng promotes angiogenesis and cell migration [19,20] but at theame time EphA2 overexpression in cancer cells stimulates onco-enic signaling pathways in a ligand-independent manner, whichs suppressed by ephrinA1-induced EphA2 phosphorylation [18].herefore, the ephrinA1–EphA2 system has opposite roles in therogression of cancer depending on tissues and cell types. Despiteur limited mechanistic understanding of the dichotomous Ephctivities, Eph–ephrin system could be a promising target for theevelpment of therapeutic agents either intended to interfereith their tumor-promoting effects or to enhance their tumor-
uppressing effects [21–24]. Both EphA1, EphA2 and ephrins A1nd A2 are abundantly expressed in healthy colon tissues, wherehey control the positioning of cell types along the crypt-villus axis13,25,26].
This scenario renders valuable the search for chemical entitiesnterfering with the Eph–ephrin system and on this regard fewnformation is available on the influence of secondary metabo-ites of plant origin, including those coming from sources alreadysed for food and/or medicinal purposes. Protein inhibitors areidespread within plant secondary metabolism products, where
hey either modulate homeostatic pathways or cause harm toathogens or competitors. A plethora of phytochemicals have been
solated so far and utilized to modulate protein-mediated signalingathways, also in human pathological processes such as carcino-enesis. Specific classes of plant proteins (phytocyanins) have beenound to share topological similarity with ephrin-B2 ectodomain27]. In plants, phytocyanins are physiologically prone to interactith small molecular weight compounds, rather than with proteinediators and this may be an indicator of the presence of some
ow-weight molecules that could influence also the ephrin activity. good prodrome on this regard is represented by green tea cat-chins, recently found to affect ephrin-induced angiogenesis in ao better-explained way, reinforcing this hypothesis towards theotential of plant foods [28]. Given the limited knowledge on theopic of edible phytochemicals, we have chosen to screen, through
proper ELISA-like binding assay, the widest possible array of plantources, including both lipophilic and hydrophilic extracts of ediblelants known to contain flavonoids and other polyphenols, ter-enes, terpenoids, glycosides and other classes of plant secondaryetabolites. The rationale behind the library building was the edi-
le or health-related use of the plants by means of oral intake andur goal was to verify if further plant sources, other than green tea,ay inhibit ephrinA1-Fc-induced EphA2 activation.
. Materials and methods
.1. Reagents
All culture media and supplements were purchased from Lonza.ecombinant proteins and antibodies were from R&D systems.ells were purchased from ECACC. Leupeptin, aprotinin, NP40,TT, tween20, BSA and salts for solutions were from Applichem;
ical Research 64 (2011) 464– 470 465
analytical grade extraction solvents, bile acids, EDTA and sodiumorthovanadate were from Sigma.
2.2. Plant material
A complete list of plant extracts, their relative botanical sources,origin of retrieval and voucher details is available as Table A in sup-plementary material. Vouchers of the crude extracts, both directlyobtained in our labs or commercially available via ACEF (Fioren-zuola, Italy), were deposited at the Dipartimento di BiologiaEvolutiva e Funzionale of the University of Parma with codes listedin supplementary material.
2.3. Preparation of crude plant extracts
For essential oil isolation, the coarsely ground plant drugs werehydrodistilled according to European Pharmacopoeia methods andessential oil samples obtained were stored in glass vials withTeflon-sealed caps at 2 ± 0.5 ◦C in the absence of light. Fixed oilswere obtained by Soxhlet extraction with n-hexane. For hydroal-coholic, water and chloroform extracts, the finely ground plantdrugs were extracted in 80% ethanol, distilled water and chloro-form, respectively, and the extracts obtained were then filtered andfreeze-dried in an Edwards E-C Modulyo lyophilizer and thus storedin the dark at −20 ◦C. The resulting powder was then re-dissolvedin DMSO according to the corresponding assay conditions.
2.4. Total polyphenols, flavonoids and procyanidin content
The determination of the total polyphenolic, flavonoidic andprocyanidin content in the active extracts has been performedusing a ThermoSpectronic Helios-y spectrophotometer, accordingto previously described methods [29–31]. Total polyphenols areexpressed as gallic acid, flavonoids as quercetin and procyanidinsas cyanidin chloride.
2.5. Phytochemical fingerprinting and NMR spectroscopy
The extracts evidencing the best results in terms of perturba-tion of the Eph–ephrin system (Arctostaphylos uva-ursi, Azadirachtaindica, Embelia ribes, Ginkgo biloba, Lagerstroemia speciosa, Phyllan-thus emblica, Punica granatum, Terminalia bellerica, and Terminaliachebula) were fingerprinted by means of 1H NMR in order to acquirea first basic set of information about their chemical composition(see supplementary material). Their expanded spectra are avail-able as supplementary material. The 1H spectrum was recorded ona Varian Gemini-400 spectrometer operating at 399.97 MHz and ata temperature of 303 K. Each extract (15 mg/1.0 ml) was dissolvedin deuterated water or mixture of deuterated water/methanol intoa 5 mm NMR tube and the solvent signal of deuterated water wasused for spectral calibration (1H 4.75 ppm). 1H spectrum was runusing a standard pulse sequence “s2pul”, 3.00 s of acquisition time,6400 Hz spectral width, 0.33 Hz Fid resolution.
2.6. ELISA screening and IC50 determination
96 well ELISA high binding plates (Costar #2592) were incubatedovernight at 4 ◦C with 100 �l/well of 1 �g/ml EphA2-Fc (R&D 639-A2) diluted in sterile PBS (0.2 g/l KCl, 8.0 g/l NaCl, 0.2 g/l KH2PO4,1.15 g/l Na2HPO4, pH 7.4), washed three times with washing buffer(PBS + 0.05% tween20, pH 7.5) and blocked with 300 �l of block-ing solution (PBS + 0.5% BSA) for 1 h at 37 ◦C. Phytocomplexes were
added to the wells at proper concentration and incubated at 37 ◦Cfor 1 h. Biotinylated ephrinA1-Fc (R&D BT602) was added for 4 hat 30 ng/ml in displacement studies. Wells were washed threetimes and incubated with 100 �l/well streptavidin-HRP (Sigma
4 acolog
Spttoattrms
2
Fp9
2
1ctdar5pa
2
laBitwpairf
2
CtaTamwc
3
3
ec
plant extracts completely abolished EphA2–ephrinA1 interaction.In order to calculate the inhibitory concentration reducing bind-ing of 50% (IC50), we built a displacement curve (Fig. 1A) using
Fig. 1. Nine extracts reversibly inhibited EphA2–ephrinA1 binding. 96 well ELISAhigh binding plates were incubated O/N with EphA2-Fc and the following daywashed and blocked with PBS + 0.5% BSA for 1 h at 37 ◦C. Compounds were added inthe wells at proper concentrations 1 h before the addition of biotinylated ephrinA1-Fc. GTE was used as natural reference. After 4 h wells were washed and incubatedwith a streptavidin-HRP solution for 20 min at room temperature. Wells werewashed again and incubated with tetra-methylbenzidine. The reaction was stoppedwith 3 N HCl and the absorbance was measured at 450 nm. (A) Nine extractsdose-dependently displaced binding of ephrin-A1-Fc ectodomain from immobilized
66 I. Hassan Mohamed et al. / Pharm
5512) solution (0.05 �g/ml in PBS supplemented with 0.5% BSA,H 7.4) for 20 min at room temperature, then washed again forhree times and incubated at room temperature with 0.1 mg/mletra-methylbenzidine (Sigma T2885) reconstituted in stable per-xide buffer (11.3 g/l citric acid, 9.7 g/l sodium phosphate, pH 5.0)nd 0.02% H2O2 (30%, m/m, in water) added immediately beforehe use. The reaction was stopped with 3 N HCl 100 �l/well andhe absorbance at 450 nm was measured using an ELISA plateeader (Sunrise, TECAN, Switzerland). The IC50 value was deter-ined using one-site competition non-linear regression with Prism
oftware (GraphPad Software Inc.).
.7. Cell cultures
PC3 human prostate adenocarcinoma cells were grown in Ham12 supplemented with 7% fetal bovine serum (FBS) and 1%enicillin–streptomycin and grown in a humidified atmosphere of5% air, 5% CO2 at 37 ◦C.
.8. Cell lysates
Cells were seeded in 12-well plates at concentration of05 cells/ml in complete medium until they reached about 40%onfluence and serum starved overnight. The day after cells werereated with phytocomplexes under study, vehicle or standardrug, stimulated with the proper agonist, rinsed with sterile PBSnd solubilized in lysis buffer. The lysates were suspended andocked at 4 ◦C for 30 min and then centrifuged at 14,000 × g for
min. The protein content of supernatant was measured with BCArotein assay kit (Thermo scientific), standardized to 200 �g/mlnd transferred into a clean test tube ready to be used.
.9. Phosphorylation assays
EphA2- and EGFR-phosphorylation were measured in cellysates using DuoSet® IC Sandwich ELISA (RnD Systems, #DYC4056nd #DYC1095, respectively) following manufacturer’s protocol.riefly, 96 well ELISA high binding plates (Costar 2592) were
ncubated overnight at room temperature with 100 �l/well ofhe specific capture antibody diluted in sterile PBS to the properorking concentrations, washed and blocked for 1 h at room tem-erature. Wells were washed and 100 �l/well of lysates were addedt room temperature for 2 h, then wells were washed again andncubated with antibody detection at room temperature for 2 h. Theeceptor phosphorylation was revealed utilizing a standard HRPormat with a colorimetric reaction read at 450 nm.
.10. MTT assay
Cell viability was evaluated using the MTT colorimetric assay.ells were plated in 96-well plates at a density of 105 cells/ml andreated with proper concentrations of extracts for 48 h. MTT wasdded at the final concentration of 1 mg/ml and incubated for 2 h.he resulting formazan crystals were washed with PBS 100 �l/wellnd then solubilized with DMSO 100 �l/well. The absorbance waseasured at 550 nm using an ELISA plate reader and the resultsere expressed as the percent ratio between absorbance of the
ells treated with extracts and untreated cells.
. Results
.1. Nine extracts inhibited EphA2–ephrinA1 binding
Nine extracts (A. uva-ursi, A. indica, E. ribes, G. biloba, L. speciosa, P.mblica, P. granatum, T. bellerica, and T. chebula) out of 133 phyto-omplexes dose-dependently inhibited EphA2–ephrinA1 binding
ical Research 64 (2011) 464– 470
with a potency in the �g/ml range. To identify extracts or essen-tial oils interfering with EphA2–ephrinA1 binding we immobilizedEphA2-Fc ectodomain on proper ELISA plates and binding ofbiotinylated-ephrin-A1-Fc was detected using the colorimetricreaction developed by streptavidine-HRP and tetramethylbenzi-dine as previously described [32]. Selectivity and specificity of theassay were tested using non-biotinylated ephrin-A1-Fc as a ligandof the EphA2-Fc receptor. As expected, non-biotinylated ephrin-A1-Fc competitively inhibited binding between EphA2-Fc-ectodomainand biotinylated-ephrin-A1-Fc, with a Ki of 102 ng/ml and a Hillcoefficient of 1.19 (see supplementary material). Furthermore, Fcalone (AG714, Millipore) did not interfere with the binding pro-cess in the range of concentration tested. All the extracts and oilswere screened at 5 mg/ml incubating them for 1 h before addingbiotinylated-ephrinA1-Fc at a concentration corresponding to itsKD. Extracts displacing more then 40% binding were further stud-ied. None of the essential and fixed oils of our library interferedwith EphA2–ephrinA1 binding, whereas the above-mentioned
EphA2-Fc ectodomain. Data are the mean of at least three independent experiments.Displacement curves were fitted using one-site competition non-linear interpola-tion with Prism 4.0. (B) EphA2–ephrinA1 binding in presence of 20 �g/ml extractwith or without washing three times with PBS with the sole exception of E. ribeswhich, being less potent, was incubated at 80 �g/ml.

I. Hassan Mohamed et al. / Pharmacological Research 64 (2011) 464– 470 467
Table 1Inhibition of EphA2–ephrinA1 binding and EphA2-phorphorylation (IC50, �g/ml)induced by most active plant extracts and reference compounds.
IC50 (95% confidence interval) (�g/ml)
Binding EphA2-phosphorylation
Azadirachta indica 0.83 (0.49–1.4) 0.88 (0.73–1.1)Ginkgo biloba 1.1 (0.75–1.6) 3.9 (2.4–6.4)Camellia sinensis 1.1 (0.62–2.0) 0.78 (0.41–1.5)Terminalia chebula 1.4 (0.65–2.9) 0.31 (0.24–0.38)Terminalia bellerica 1.9 (0.93–3.9) 1.2 (0.63–2.2)A. uva-ursi 2.2 (1.4–3.4) 1.1 (0.89–1.4)Punica granatum 2.7 (2.0–3.7) 0.95 (0.58–1.5)Lagerstroemia speciosa 3.5 (2.7–4.7) 1.2 (0.65–2.0)Phyllanthus emblica 8.9 (4.9–16) 1.9 (1.4–2.5)
deLdv(iwteecf(ewrtb
3n
arabsp(op((twegsecef
3
t
Fig. 2. Extracts dose-dependently inhibited EphA2-kinase phosphorylation. Serumstarved PC3 cells were pretreated for 20 min with PBS or the indicated concentra-tions (�g/ml) of the extracts and stimulated for 20 min with 0.25 �g/ml ephrinA1-Fcor Fc alone as control. GTE was used as natural reference. Phosphorylation levelsof EphA2 are relative to ephrinA1-Fc + PBS (assumed as 100%). Data are the meanof at least three independent experiments. Displacement curves were fitted usingone-site competition non-linear interpolation with Prism 4.0.
Fig. 3. The extracts were not cytotoxic. PC3 cells were grown in the presence of40 �g/ml of each extract for 48 h, MTT was added for 2 h, solubilized with DMSO
Embelia ribes 24 (19–30) 11.3 (6.0–21)Dasatinib Inactive 0.012 (0.008–0.018)
ecreasing concentration of each extract towards biotinylated-phrinA1-Fc. The extracts of A. uva-ursi, A. indica, E. ribes, G. biloba,. speciosa, P. emblica, P. granatum, T. bellerica, and T. chebula dose-ependently inhibited EphA2–ephrinA1 binding (Fig. 1A) with IC50alues ranging from 0.83 �g/ml (A. indica) to 24 �g/ml (E. ribes)Table 1). Interestingly, green tea extract, previously reported tonhibit EphA2 phosphorylation upon ephrinA1-Fc activation [28],
as also able to inhibit EphA2–ephrinA1 protein–protein interac-ion with an IC50 of 1.1 �g/ml. Moreover, we repeated displacementxperiments incubating for 1 h each extract at 20 �g/ml with thexception of E. ribes incubated at 80 �g/ml, being less potent. Theseoncentrations displaced at least 80% of biotinylated-ephrinA1-Fcrom EphA2. Half of the wells treated with the extracts were washed3 times with PBS, 5 min) before the addition of biotinylated-phrinA1-Fc. In this experiment the displacement action of extractsas detected only when washing was not performed, suggesting a
eversible binding of the extracts to EphA2 receptor (Fig. 1B). Greenea showed a longer persistence, being able to partially displace theinding after the washing.
.2. The extracts inhibited Eph-kinase phosphorylation aton-cytotoxic concentrations
Functional studies were performed in cultured cells to evalu-te agonist or antagonist properties of the extracts towards EphA2eceptor. We used PC3 human prostate adenocarcinoma cells as
model for their known ability to naturally express EphA2 andecause they are a well established model to study the Eph–ephrinystem [32,33]. None of the extracts was able to induce EphA2 phos-horylation on PC3 cells when incubated for 30 min at 40 �g/mldata not shown), thus evidencing a non-agonist behavior. On thether hand, all the extracts dose-dependently inhibited EphA2hosphorylation induced by 0.25 �g/ml ephrinA1-Fc on PC3 cellsFig. 2), showing IC50 values (Table 1) ranging from 0.31 �g/mlT. chebula) to 11.3 �g/ml (E. ribes), comparable with concentra-ion obtained from green tea extract (0.78 �g/ml). Dasatinib 1 �Mas used as reference compound, being a multikinase inhibitor
ndowed with a high potency towards Eph kinases [34], whilereen tea extract was used as a dietary reference. MTT assay demon-trated that inhibitory effect of extracts were not due to any toxicffect on cells, albeit green tea extract exhibited a slightly higherytotoxicity if compared with other plants. In fact, none of thextracts decreased PC3 cell viability when incubated up to 40 �g/mlor 48 h (Fig. 3).
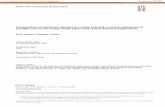
.3. Extracts did not affect EGFR activity
All active discovered extracts were completely inactive whenested towards EGFR phosphorylation, induced by 30 ng/ml EGF
and read at 550 nm. PBS was used as control and relative cell viability is referredto PBS. GTE was used as natural reference. Data are the mean of three independentexperiments ± standard errors.
in PC3 cells, at doses at least 4 times higher then those active inEphA2 phosphorylation (Fig. 4). The EGFR kinase inhibitor gefi-tinib (10 �M), used as reference compound, completely abolishedresponse to EGF in this functional study. Green tea extract (con-taining 44% EGCG) did not interfere with EGFR phosphorylation upto 5 �g/ml accordingly with previous observations in which EGCGprovided a significant activity only above 50 �M, corresponding to4.8 �M in our case [35].
4. Discussion
Our data suggest a specific interaction between nine phyto-complexes from edible plants and EphA2–ephrinA1 system. We
468 I. Hassan Mohamed et al. / Pharmacolog
Fig. 4. None of the extracts affected EGFR activity. Serum starved PC3 cells were pre-treated for 20 min with PBS, 10 �M gefitinib or the indicated concentrations (�g/ml)of the extracts and stimulated for 20 min with 30 ng/ml EGF. GTE was used as nat-uoc
otwtdwtarEronspt(flsipl
TP
ral reference. Phospho-EGFR levels are relative to EGF + PBS. Data are the meanf at least three independent experiments ± standard error. T-test was performedomparing all the columns to EGF + PBS. **p < 0.01.
btained dose-response relationships both in binding and func-ional assays and the active concentrations in the binding studyere comparable with the working concentrations in the func-
ional assay. Active concentrations may be easily obtained throughietary supplementation, as the in vitro active dose may be attainedith 2–5 g of crude drug. Moreover, we showed that the interac-
ion between extracts and EphA2-Fc ectodomain was reversiblend extracts were completely inactive towards EGFR, anothereceptor tyrosine kinase, suggesting a preferential interaction withph–ephrin system. Finally, the extracts inhibited EphA2 phospho-ylation induced by ephrinA1 at non-cytotoxic concentration: nonef the extracts modified cell viability up to 40 �g/ml. Total polyphe-ol, flavonoid and procyanidin content of the extracts (Table 2)howed mixed results when compared to binding and phos-horylation data. A moderate correlation was detected betweenotal polyphenolic content and both binding and phosphorilationR2 = 0.7, see supplemental material), while the correlation withavonoid and procyanidin content was negligible. Nevertheless,
ome of the most active extracts (e.g. A. indica, G. biloba) were richn procyanidins, but their action was similar to those provided byhytocomplexes with much smaller amounts, like T. chebula, T. bel-erica or almost devoid of this class of polyphenols, like P. granatum.
able 2hytochemical profiles of most active plant extracts.
Total polyphenols
Azadirachta indica 53.81 ± 0.14
Ginkgo biloba 61.1 ± 0.10
Terminalia chebula 88.02 ± 0.21
Terminalia bellerica 59.84 ± 0.22
A. uva-ursi 68.36 ± 0.13
Punica granatum 87.08 ± 0.16
Lagerstroemia speciosa 56.88 ± 0.12
Phyllanthus emblica 47.08 ± 0.21
Embelia ribes 9.79 ± 0.05
ical Research 64 (2011) 464– 470
At the same time E. ribes extract, providing the lowest IC50 both inbinding and in phospohorylation assays, presented a considerableamount of procyanidins. This could lead to suggest that differentpolyphenolic substances may be potentially involved in the per-turbation of the ephrin system and may warrant further distinctphytochemical evaluation of some extracts.
Interestingly, most active plants (T. chebula, T. bellerica, P.emblica, E. ribes and A. indica) are listed as ayurvedic Rasayanaherbal food-medicines, whose habitual intake through the diet isreputed as a way to “restore normal function”, also under the per-spective of cancer prevention [36,37]. Moreover, it must be alsonoticed that these specific extracts were obtained by simple decoc-tion, thus exactly representing the dietary form by which theseplants are ingested. In particular, the decoctions of three of them(P. emblica, T. chebula and T. bellerica) are the main constituents ofTriphala, MAK-4 and MAK-5 food-medicines, widely used in cancertreatment by the ayurvedic medicine and known to reduce metas-tasis in vivo and increase animal life expectancy in cancer-affectedmice in a no-better explained way [38–40]. Given the involvementof the ephrin system in angiogenesis and cell migration [20,25],such activities could be the object of further evaluation. Besidesthese plants, other sources of extracts active in our study, namelypomegranate fruit and ginkgo leaves, are increasingly supportedas functional food ingredients due to their purported health ben-efits and possess an antiangiogenic background from a preclinicalstandpoint [41–43]. Moreover, A. indica and G. biloba are traditionalanticancer remedies and different constituents isolated from themhave shown the capacity to reduce different kind of cancers. Theirantiangiogenic effect seems to be performed by means of somekind of regulation of cell proliferation and differentiation, all factorsthat involve the core function of the ephrin system, albeit a com-plete description of their mechanism of action is not available [44].L. speciosa and Arctostaphylos uva-ursi, instead, offer limited evi-dences in terms of antitumoral activity, but as the other seven activeplants they are known to be rich in tannins and polyphenols as evi-denced also by their NMR fingerprints, thus leading to hypothesizethat this class of secondary metabolites may be somehow involvedin the modulation of the ephrin system. Interestingly, these sub-stances are deemed responsible or co-responsible of the not yetcompletely explained correlation between their intake and reducedcancer risk. In particular, all the nine extracts that displayed anactivity in our assay are known to contain polymeric polyphenolsand tannins, which often act as non-specific inhibitors of enzymaticsystems and proteins. In our case, however, the reversibility of theinteraction and the absence of a nonspecific response may lead tosuggest the presence of at least a class of substances that may act asa true modulator of the Eph–ephrin system. Their action on ephrinsmay complement previously reported apoptotic and chemopreven-tive mechanisms. This property, coupled with their other effects,
may render them interesting candidates against both the initiationand promotion/progression stages of carcinogenesis.Considering the role of the Eph–Ephrin system in tumor angio-genesis, cell migration and metastasis, the screening for edible
Total flavonoids (g/100 g) Total procyanidins
5.86 ± 0.07 1.718 ± 0.01125.13 ± 0.09 0.678 ± 0.010
5.64 ± 0.06 0.069 ± 0.0044.87 ± 0.07 0.073 ± 0.0044.27 ± 0.05 0.171 ± 0.009
27.32 ± 0.04 0.004 ± 0.00117.13 ± 0.04 0.268 ± 0.003
1.48 ± 0.02 0.108 ± 0.0081.7 ± 0.05 0.449 ± 0.012

acolog
ptoEconastLmtbpmdhsphm
iimtuceftat
A
ofd
A
t
R
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[[
[
[
[
[
[
[
[
[
[
[
[
[
I. Hassan Mohamed et al. / Pharm
lants able to interfere with this system may help to explainhe correlation between foods, polyphenols and cancer devel-pment. While the discovery of a lead compound binding tophA2–ephrinA1 system may offer a basic scaffold for the chemi-al manipulations needed by pharmaceutical applications, the rolef dietary phytochemicals may be further evaluated also from autraceutical point of view, as some polyphenol-rich plants mayffect this system. In the intestine, in particular, the Eph/ephrinystem is strictly connected with the renewal of the epitheliumhrough the activity of staminal cells located at the bottom of theieberkühn crypts [25,26]. Notably, these proteins regulate celligration patterns on the crypt-villus axis and may act as potential
umor suppressors if properly modulated, thus their perturbationy food-related phytocomplexes may suggest intriguing patho-hysiological implications [45,46]. In this case the active extractsay also act without a mandatory involvement of a systematic
istribution and could play an unpredicted role in the intestinalomeostasis. Therefore, our findings could be useful for furthertudies aimed to explain the correlation between the intake oflant secondary metabolites and the incidence of colorectal cancer,ighlighted by several epidemiology studies, but whose molecularechanisms are far to be clear [47].The novelty of gathered data, according to the quality of the
nteractions detected, could thus warrant further developmentsn the knowledge of Eph-kinases and their ligands: to determine
echanism, to elucidate which chemical skeletons are more proneo interact with the ephrin system and to provide further toolsseful to evaluate the role of ephrins also from a nutraceuti-al standpoint. According to data hereby presented, some plantxtracts may retain a promising future as a source of molecules orunctional foods able to modulate the Eph–ephrin system, shouldheir activity be confirmed by in vivo systems, taking into properccount the biotransformation of plant secondary metabolites byhe gut flora, their bioavailability and kinetics.
cknowledgements
This work was supported by “My First AIRC Grant” number 6181f the “Associazione Italiana Ricerca sul Cancro” (www.airc.it). Theunder had no role in study design, data collection and analysis,ecision to publish, or preparation of the manuscript.
ppendix A. Supplementary data
Supplementary data associated with this article can be found, inhe online version, at doi:10.1016/j.phrs.2011.06.008.
eferences
[1] Riboli E, Norat T. Epidemiologic evidence of the protective effect of fruit andvegetables on cancer risk. Am J Clin Nutr 2003;78:559S–69S.
[2] Pavia M, Pileggi C, Nobile CG, Angelillo IF. Association between fruit and veg-etable consumption and oral cancer: a meta-analysis of observational studies.Am J Clin Nutr 2006;83:1126–34.
[3] Whu AH, Yu MC, Tseng CC, Pike MC. Epidemiology of soy exposures and breastcancer risk. Brit J Cancer 2008;98:9–14.
[4] Sofi F, Cesari F, Abbate R, Gensini GF, Casini A. Adherence to mediterranean dietand health status: meta-analysis. Brit Med J 2008;337:344–51.
[5] Myung SK, Kim Y, Ju W, Choi HJ, Bae WK. Effects of antioxidant supplements oncancer prevention: meta-analysis of randomized controlled trials. Ann Oncol2009;21:166–79.
[6] Bardia A, Platz EA, Yegnasubramanian S, De Marzo AM, Nelson WG. Anti-inflammatory drugs, antioxidants, and prostate cancer prevention. Curr OpinPharmacol 2009;9:419–26.
[7] Bardia A, Tleyjeh IM, Cerhan JR, Sood AK, Limburg PJ, Erwin PJ, MontoriVM. Efficacy of antioxidant supplementation in reducing primary cancer inci-
dence and mortality: systematic review and meta-analysis. Mayo Clin Proc2008;83:23–34.[8] Mekary RA, Wu K, Giovannucci E, Sampson L, Fuchs C, Spiegelman D, et al. Totalantioxidant capacity intake and colorectal cancer risk in the Health Profession-als Follow-up Study. Cancer Causes Contr 2010;21:1315–21.
[
[
ical Research 64 (2011) 464– 470 469
[9] Liu RH. Potential synergy of phytochemicals in cancer prevention: mechanismof action. J Nutr 2004;134:3479S–85S.
10] Stan SD, Kar S, Stoner GD, Singh SV. Bioactive food components and cancer riskreduction. J Cell Biochem 2008;104:339–56.
11] Surawska H, Ma PC, Salgia R. The role of ephrins and Eph receptors in cancer.Cytokine Growth Factor Rev 2004;15:419–33.
12] Gale NW, Yancopoulos GD. Ephrins and their receptors: a repulsive topic? CellTiss Res 1997;290:227–41.
13] Hafner C, Schmitz G, Meyer S, Bataille F, Haum P, Langmann T, et al. Differentialgene expression of Eph receptors and ephrins in benign human tissues andcancers. Clin Chem 2004;50:490–9.
14] Ogawa K, Pasqualini R, Lindberg RA, Kain R, Freeman AL, Pasquale EB. Theephrin-A1 ligand and its receptor EphA2, are expressed during tumor neovas-cularization. Oncogene 2000;19:6043–52.
15] Walker-Daniels J, Hess AR, Hendrix MJ, Kinch MS. Differential regulation ofEphA2 in normal and malignant cells. Am J Pathol 2003;162:1037–42.
16] Zelinski DP, Zantek ND, Stewart JC, Irizarry AR, Kinch MS. EphA2 overexpressioncauses tumorigenesis of mammary epithelial cells. Cancer Res 2001;61:2301–6.
17] Sugimura H, Wang JD, Mori H, Tsuboi M, Nagura K, Igarashi H, et al. Eph–ephrinin human gastrointestinal cancers. World J Gastrointest Oncol 2010;2:421–8.
18] Wykosky J, Debinski W. The EphA2 receptor and ephrinA1 ligand in solidtumors: function and therapeutic targeting. Mol Cancer Res 2008;6:1795–806.
19] Cheng N, Brantley DM, Liu H, Lin Q, Enriquez M, Gale N, et al. Blockade of EphAreceptor tyrosine kinase activation inhibits vascular endothelial cell growthfactor-induced angiogenesis. Mol Cancer Res 2002;1:2–11.
20] Fang WB, Brantley-Sieders DM, Parker MA, Reith AD, Chen J. A kinase-dependent role for EphA2 receptor in promoting tumor growth and metastasis.Oncogene 2005;24:7859–68.
21] Brantley DM, Cheng N, Thompson EJ, Lin Q, Brekken RA, Thorpe PE, et al. SolubleEph A receptors inhibit tumor angiogenesis and progression in vivo. Oncogene2002;21:7011–26.
22] Carles-Kinch K, Kilpatrick KE, Stewart JC, Kinch MS. Antibody targetingof the EphA2 tyrosine kinase inhibits malignant cell behavior. Cancer Res2002;62:2840–7.
23] Dobrzanski P, Hunter K, Jones-Bolin S, Chang H, Robinson C, Pritchard S, et al.Antiangiogenic and antitumor efficacy of EphA2 receptor antagonist. CancerRes 2004;64:910–9.
24] Pasquale EB. Eph receptors and ephrins in cancer: bidirectional signalling andbeyond. Nat Rev Cancer 2010;10:165–80.
25] Kosinski C, Li VS, Chan AS, Zhang J, Ho C, Tsui WY, et al. Gene expression patternsof human colon tops and basal crypts and BMP antagonists as intestinal stemcell niche factors. Proc Natl Acad Sci USA 2007;104:15418–23.
26] Moore KA, Lemischka R. Stem cells and their niches. Science 2006;31:1880–5.27] Toth J, Cutforth T, Gelinas AD, Bethoney KA, Bard J, Harrison CJ. Crystal structure
of an ephrin ectodomain. Dev Cell 2001:83–92.28] Tang FY, Chiang EP, Shih CJ. Green tea catechin inhibits ephrin-A1-mediated
cell migration and angiogenesis of human umbilical vein endothelial cells. JNutr Biochem 2007;18:391–9.
29] Chaovanalikit A, Wrolstad RE. Total antocyanins and total phenolics of freshand processed cherries and their antioxidant properties. Food Chem Toxicol2004;69:67–72.
30] Porter LJ, Hrstich LN, Chan BC. The conversion of procyanidins and prodelphini-dins to cyanidins and delphinidins. Phytochemistry 1986;25:225–30.
31] Luximon-Ramma A, Bahorun T, Soobrattee MA, Aruoma OI. Antioxidant activ-ities of phenolic, proanthocyanidin, and flavonoid components in extracts ofCassia fistula. J Agric Food Chem 2002;50:5042–7.
32] Giorgio C, Hassan Mohamed I, Flammini L, Barocelli E, Incerti M, Lodola A, et al.Lithocholic acid is an Eph–ephrin ligand interfering with Eph-kinase activation.PLoS One 2011;6:e18128.
33] Miao H, Burnett E, Kinch M, Simon E, Wang B. Activation of EphA2 kinase sup-presses integrin function and causes focal-adhesion-kinase dephosphorylation.Nat Cell Biol 2000;2:62–9.
34] Tanaka M, Kamata R, Sakai R. EphA2 phosphorylates the cytoplasmictail of claudin-4 and mediates paracellular permeability. J Biol Chem2005;280:42375–82.
35] Chang Q, Jorgensen C, Pawson T, Hedley DW. Effects of dasatinib on EphA2receptor tyrosine kinase activity and downstream signalling in pancreatic can-cer. Brit J Cancer 2008;99:1074–82.
36] Sah JF, Balasubramanian S, Eckert RL, Rorke EA. Epigallocatechin-3-gallateinhibits epidermal growth factor receptor signaling pathway. Evidencefor direct inhibition of ERK1/2 and AKT kinases. J Biol Chem 2006;279:12755–62.
37] Aggarwal BB, Ichikawa H, Garodia P, Weerasinghe P, Sethi G, Bhatt ID, et al. Fromtraditional ayurvedic medicine to modern medicine: identification of thera-peutic targets for suppression of inflammation and cancer. Expert Opin TherTargets 2006;10:87–118.
38] Balachandran P, Govindarajan R. Cancer – an ayurvedic perspective. Pharm Res2005;51:19–30.
39] Menon LG, Kuttan R, Kuttan G. Effect of rasayanas in the inhibition oflung metastasis induced by B16F-10 melanoma cells. J Exp Clin Canc Res1997;16:365–8.
40] Baliga MS. Triphala, ayurvedic formulation for treating and preventing cancer:a review. J Altern Complement Med 2010;16:1301–8.
41] Vimal KP, Wang J, Shen RN, Sharma HM, Brahmi Z. Reduction of metastasesof Lewis lung carcinoma by an ayurvedic food supplement in mice. Nutr Res1992;12:51–61.

4 acolog
[
[
[
[
[
70 I. Hassan Mohamed et al. / Pharm
42] Lansky EP, Newman RA. Punica granatum (pomegranate) and its potentialfor prevention and treatment of inflammation and cancer. J Ethnopharmacol2007;109:177–206.
43] DeFeudis FV, Papadopoulos V, Drieu K. Ginkgo biloba extracts and
cancer: a research area in its infancy. Fund Clin Pharm 2003;17:405–17.44] Kumar S, Sureah PK, Vijayababu MR, Arunkumar A, Arunakaran J. Anticancereffects of ethanolic neem leaf extract on prostate cancer cell line (PC-3). JEthnopharmacol 2005;105:246–50.
[
ical Research 64 (2011) 464– 470
45] Cortina C, Palomo-Ponce S, Iglesias M, Fernandez-Masip JL, Vivancos A, WhissellG, et al. EphB–ephrin-B interactions suppress colorectal cancer progression bycompartmentalizing tumor cells. Nat Genet 2007;11:1376–83.
46] Batlle E, Henderson JT, Beghtel H, van den Born MMW, Sancho E, Hols GH,
et al. �-Catenin and TCF mediate cell positioning in the intestinal epitheliumby controlling the expression of EphB/ephrinB. Cell 2002;11:251–63.47] Michels KB, Giovannucci E, Chan AT, Singhania R, Fuchs CS, Willett WC. Fruit andvegetable consumption and colorectal adenomas in the Nurses’ Health Study.Cancer Res 2006;66:3942–53.