
Поливариантность онтогенеза и жизненные формы лесных...
12
БЮЛЛЕТЕНЬ МОСКОВСКОГО ОБЩЕСТВА ИСПЫТАТЕЛЕЙ ПРИРОДЫ Основан в 1829 г. ОТДЕЛ БИОЛОГИЧЕСКИЙ Том 96, вып. 4 ИЮЛЬ — АВГУСТ Выходит 6 раз в год ИЗДАТЕЛЬСТВО МОСКОВСКОГО УНИВЕРСИТЕТА 1991
-
Upload
independent -
Category
Documents
-
view
0 -
download
0
Transcript of Поливариантность онтогенеза и жизненные формы лесных...

БЮЛЛЕТЕНЬ
МОСКОВСКОГО ОБЩЕСТВА
ИСПЫТАТЕЛЕЙ ПРИРОДЫ
Основан в 1829 г.
ОТДЕЛ БИОЛОГИЧЕСКИЙ
Том 96, вып. 4 ИЮЛЬ — АВГУСТ
Выходит 6 раз в год
ИЗДАТЕЛЬСТВО МОСКОВСКОГО УНИВЕРСИТЕТА 1991

БЮЛ. MOCK. О-ВА ИСПЫТАТЕЛЕЙ ПРИРОДЫ. ОТД. БИОЛ. 1991. Т. 96, ВЫП. 4
УДК 58
ПОЛИВАРИАНТНОСТЬ ОНТОГЕНЕЗА И ЖИЗНЕННЫЕ ФОРМЫ ЛЕСНЫХ КУСТАРНИКОВ
И. И. Истомина, Н. Н. Богомолова
В популяционной биологии растений сравнительно недавно сформировалась концепция поливариантности развития особей (Зауголь- нова и др., 1988а). В рамках этой концепции онтоморфогенез рассматривается не как ряд последовательных однозначно определяемых этапов, а как сложно организованная совокупность рядов со взаимными переходами. Неоднозначность онтоморфогенеза в разных эколого-це- нотических ситуациях приводит к становлению различных жизненных форм у одного и того же вида. Это явление описано для некоторых трав и деревьев (Жукова, 1986; Чистякова, 1986, 1988; Заугольнова и др., 1988а, б), но практически не рассматривалось у кустарников.
В связи с этим в работе была поставлена задача — выявить основные направления развития лесных кустарников, оценить степень морфологической поливариантности и определить полный набор жизненных форм у следующих видов: лещина обыкновенная (Corylus avel- lana L.), бересклет бородавчатый (Euonymus verrucosa Scop.), бересклет европейский (E. europaea L.), жимолость лесная (Lonicera xylo- steum L.), крушина ломкая (Frangula alnus Mill.), свидина кроваво- красная (Thelycrania sanguinea (L.) Fourr.), бузина красная и черная (Sambucus racemosa L., S. nigra L.).
Материал и методы исследованияМатериал по онтогенезу исследованных видов собран в грабовых и дубово-гра
бовых лесах Украины (Каневский заповедник, Львовский лесхоззаг), в дубово-липовых лесах Центральной России (Сурское лесничество Пензенской обл.), в хвойно-широколиственных лесах (Истринское лесничество Московской обл.).
При выделении возрастных состояний у кустарников использовали общепринятые методики (Работнов, 1964; Заугольнова и др., 1988а), жизненные формы у них выделены и описаны в соответствии с разработками И. Г. Серебрякова (1964).
Результаты исследования
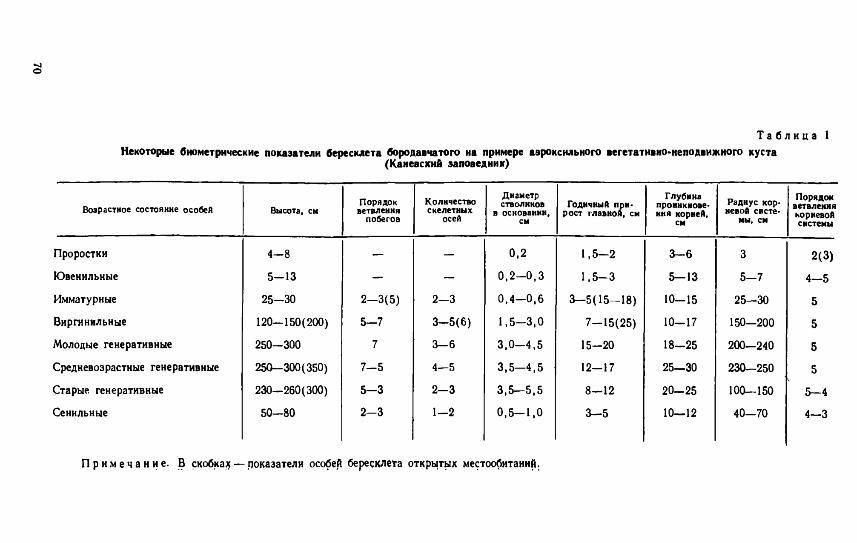
Согласно классификации И. Г. Серебрякова (1962), перечисленные выше лесные кустарники относятся к группам аэроксильных (бересклет бородавчатый, жимолость лесная, крушина ломкая) или гео- ксильных (лещина). Однако детальное последование онтогенезов этих видов показало, что каждому из них свойственна не одна, а определенный набор жизненных форм. Морфогенез каждой из этих форм является результатом соответствующего пути онтогенеза. Рассмотрим эти пути на примере бересклета бородавчатого (рис. 1, табл. 1). В онтогенезе этого вида можно выделить два направления развития (два пути онтогенеза): семенной и вегетативный.
Р а з в и т и е о с о б е й с е м е н н о г о п р о и с х о ж д е н и я
Проростки. Семена бересклета бородавчатого обычно прорастают на вторую весну после созревания. Тип прорастания надземный (Чистяков, 1935; Николаева, 1956; Леонова, 1974). Семядоли овальные»
68

ассимилирующие, 0,8—1,0 см; первые настоящие листья по форме почти ничем не отличаются от листьев взрослых растений, но имеют меньшие размеры— 2,0—2,7 см. Общая высота проростков 4—8 см. Главный корень проникает в почву на глубину 3—6 см, ветвление боковых корней незначительно и достигает второго, реже третьего порядка.
Рис. 1. Варианты онтогенеза бересклета бородавчатого (семенной ряд): А — аэроксиль- ный вегетативно-неподвижный кустарник, Б — деревце, В — геоксильный эпигеогенно-
корневищный кустарник, Г — формирование стланика
Ювенильные растения одноосные, нарастают моноподиально или при сильном затенении неустойчиво моноподиально. В зависимости от жизненности высота ювенильных растений колеблется от 5 до 13 см. Главный корень проникает в почву «а глубину 6—13 см, порядок ветвления корневой системы 4—5.
Имматурные растения характеризуются началом ветвления. Именно в этом возрастном состоянии начинает проявляться поливариантность онтогенеза особей семенного происхождения. Растения, имеющие одну главную ось, которая ветвится в верхней части, дают начало д р е в о в и д н о й ф о р м е (деревцу) бересклета бородавчатого. Древовидные имматурные особи бересклета имеют небольшой порядок ветвления (2—3), высота их достигает 60—70 см, диаметр кроны 12—25 см, высота прикрепления кроны 15—20 (до 30) см. Другие особи в результате раннего усыхания верхушечной почки не только начинают интенсивно ветвиться, но и образуют из спящих почек базальной части новые (дочерние) оси возобновления, т. е. переходят в фазу кущения и формируют а э р о к с и л ь н ы й в е г е т а т и в н о - н е п о д в и ж н ы й куст. К у с т о в и д н ы е имматурные особи бересклета бородавчатого имеют 2—3 скелетные оси, достигающие 25—30 (до 50, а иногда и более) см в высоту, с порядком ветвления 3—4 (до 5).
В корневой системе как древовидных, так и кустовидных имматур- ных особей наряду с дальнейшим развитием стержневого корня появляются боковые скелетные корни, растущие горизонтально, посте-
зсВщеннесть
69
о
Т а б л и ц а 1Некоторые биометрические показатели бересклета бородавчатого на примере аэроксильного вегетативно-неподвижного куста
(Каневский заповедник)
Возрастное состояние особей Высота, смПорядокветвления
побегов
Количествоскелетных
осей
Диаметрстволиков Годичный прн-
в основании, рост главной, см см
Глубина проникновения корней,
см
Радиус корневой систе
мы, см
Порядокветвлениякорневойсистемы
Проростки 4—8 0,2 1,5 - 2 3—6 3 2(3)
Ювенильные 5—13 0 ,2—0,3 1,5 - 3 5—13 5—7 4—5
Имматурные
Виргиннльные
Молодые генеративные
25—30
120—150(200)
250—300
2—3(5)
5—7
7
2- 3
3- 5(6)
3—6
0 ,4—0,6
1,5—3,0
3 ,0—4,5
3- 5( 15- 18)
7- 15(25)
15-20
10-15
10-17
18-25
25—30
150—200
200—240
5
5
5
Средневозрастные генеративные
Старые генеративные
Сенильные
250—300(350)
230—260(300)
50—80
7 - 5
5 - 3
2 - 3
4—5
2 - 3
3 .5— 4,5 12—17
3 .5— 5,5 8—12
1 -2 0 ,5- 1,0 3 - 5
25—30
20-25
10—12
230—250
100—150
40-70
5
5—4
4—3
П р и м е ч а н и е . В скобка^ — показатели особей бересклета открытых местообитаний.

женно главный корень теряет свое лидирующее 'положение и корневая •система становится поверхностной.
У виргинильных растений отмечено дальнейшее увеличение разнообразия габитуса. Так, в лесах, в условиях сильного бокового затенения, продолжаются усиленный рост верхушки и дальнейшее развитие .древовидной жизненной формы. Виргинильные особи древовидной формы роста характеризуются увеличением порядка ветвления до 5—6, годичный прирост главной оси достигает 15, реже 20 ем и более. Высота этих растений 105—150 см, высота прикрепления кроны 40—50 см.
В больших окнах, на опушках леса и вырубках начинается усиленное ветвление в базальной части оси 1-го порядка на небольшой высоте (5—10 см) над поверхностью почвы и формируется типичный аэроксильный вегетативно-неподвижный кустарник. Виргинильные аэроксильные кусты бересклета имеют от 2 до 6 скелетных осей высотой 120—150 (до 200) см, ветвление которых достигает 6—7-го порядка.
Под пологом леса, в условиях бокового и верхушечного затенения, у бересклета бородавчатого к 15—20 годам жизни главная ось накло- яяется и полегает в результате резкого ослабления камбиальной деятельности, вызванного низкой освещенностью (Серебряков, 1962). Вертикальной остается только верхняя часть побега. На засыпанном лесной подстилкой участке оси появляются придаточные корни, формируется эпигеогенное корневище (Серебряков, Серебрякова, 1965). Это укоренение стимулирует пробуждение спящих почек по всей длине полегшего побега (Серебряков, 1962). Из опящих почек полегших осей возобновления возникают новые побеги, интенсивно растущие 5—7 лет, л затем замедляющие овой рост и развивающиеся по типу материнских. Таким образом формируется геоксильный эпигеогенно-корневищный кустарник. Полегание главной и боковых осей приводит к формированию достаточно развитой системы придаточных корней, вследствие чего функциональное значение главного корня снижается и он быстро отмирает.
Виргинильные эпигеогенно-геоксильные кусты бересклета имеют 4 —7 осей возобновления высотой 75—110 см с диаметром стволиков €.8—1,5 см. Ветвление побегов достигает 6—7-го порядка. Годичный прирост осей невелик — 5—7 см. Корневая система поверхностная, глубина проникновения в почву 10—15 ом, радиус 100—150 ем. В лесных ценозах бересклет бородавчатый начинает плодоносить с 8— 16 лет. В литературе отмечено более раннее начало плодоношения {Леонова, 1974).
Молодые генеративные растения. Древовидные особи бересклета в этом состоянии имеют высоту 150—200 (до 300) см, диаметр стволика в основании 4,0—5,5 см, порядок ветвления 7. Высота прикрепления кроны 90—130 см, а ее диаметр в широкой части ПО—160 см. Радиус корневой системы 150—200 см, глубина проникновения в почву 18— 20 см. Аэроксильные вегетативно-неподвижные кусты бересклета в молодом генеративном состоянии могут достигать 250—300 см высоты, имеют обычно 3—6 осей возобновления с диаметром в основании 3,0— 4,5 см. Корневая система поверхностная, радиус ее достигает 200— 240 см.
У особей бересклета бородавчатого, относящихся к зпигеогенно- геоксильной жизненной форме, продолжаются на этом этапе онтогенеза процессы разрастания, укоренения осей, а также образования новых осей возобновления следующих порядков. Они развиваются по типу материнской оси, которая в свою очередь постепенно усыхает. Вы-
71

сота этих растений 110—130 см, диаметр стволиков в основании 1,5— 2,0 см, порядок ветвления 7, годичный прирост 5—7 см. Корневая система в основном придаточная, поверхностная, глубина ее проникновения в почву 12—17 см, радиус 130—150 см.
Средневозрастные генеративные растения бересклета бородавчатого характеризуются максимальным плодоношением, но процессы новообразования осей замедляются, появляются первые признаки старения. У особей древовидной формы усыхают побеги последних порядков, в связи е этим порядок ветвления уменьшается до 5. Высота растений 170—200 см, крона приобретает зонтиковидную форму, высота прикрепления ее ПО—140 см, а диаметр в широкой части 130—160 см.
У средневозрастных генеративных аэроксильных вегетативно-неподвижных кустов также наблюдается снижение порядка ветвления до 6—5 в связи с усыханием небольших тоненьких веточек последних порядков. А в остальном так же, как и у особей эпигеогенно-геоксиль- ной биоморфы, средневозрастные генеративные растения трудноотличимы от молодых генеративных растений, так как общий габитус и биометрия остаются почти неизменными.
У старых генеративных особей бересклета бородавчатого отмечаются уменьшение количества плодов и преобладание процессов отмирания над процессами новообразования в побеговой и корневой системах. Растения древовидной формы уменьшают годичные приросты до 5—8 см, в их кроне появляется большое количество усохших скелетных ветвей, а порядок ветвления уменьшается до 5—3. Диаметр кроны 100—140 см.
У аэроксильных вегетативно-неподвижных кустов отмирает большое количество функционирующих побегов 3—4-го порядка, усыхают 1—2 крупных стволика, годичный прирост не превышает 8—12 см. Уменьшается радиус корневой системы (100—150 см).
У эпигеогенно-геоксильных особей бересклета на этом этапе онтогенеза наблюдается отмирание побегов 7—4-го порядка, нарушается связь осей возобновления следующих порядков с материнскими — начинается процесс партикуляции. Годичный прирост 4—5 см, высота 100—ПО (реже более) см.
Сенильные древовидные особи бересклета бородавчатого и аэро- ксильные вегетативно-неподвижные кусты представлены остатками отмерших почти до основания стволиков и системой возникших из спящих почек базальной части растения ювенилоподобных слаборазвитых побегов. Высота их не превышает 50—80 см, годичный прирост 3— 5 см. Радиус корневой системы 40—70 см. Эпигеогенно-геоксильные кусты в сенильном состоянии распадаются на систему сенильных пар- тикул, представляющих собой несколько ослабленных осей с остатками эпигеогенных корневищ.
Р а з в и т и е о с о б е й в е г е т а т и в н о г о п р о и <с х о ж д е н и я
Особи вегетативного происхождения возникают из спящих пазушных почек эпигеогенных корневищ. Осмотр нескольких тысяч экземпляров бересклета бородавчатого не дал возможности обнаружить особи корнеотпрыскового происхождения, отмеченные И. Г. Серебряковым (1954, 1962). Возрастные ряды у особей вегетативного происхождения начинаются с ювенильного состояния.
Ювенильные особи вегетативного происхождения имеют несколько большие размеры, чем семенные растения того же состояния. Высота их достигает 15 см. Корневая система полностью придаточная, поверх
72

ностная. При отсутствии главного корня один из придаточных корней зачастую занимает главенствующее положение.
Среди имматурных особей вегетативного происхождения уже можно выделить древовидные и аэроксильно-кустовидные формы. По биометрическим показателям и дальнейшему развитию они практически ■не отличаются от особей бересклета этого же возрастного состояния семенного происхождения, за исключением следующей специфической особенности. В условиях очень сильного затенения (1—2% от полного света) и достаточной влажности онтогенез становится незавершенным. Здесь имматурные особи после нескольких лет роста вследствие слабого развития механических тканей полегают, укореняются, а из спящих почек, расположенных на плагиотропной части побега, образуются ювенильные оси следующего порядка. Эти побеги либо развиваются до имматурного возрастного состояния, либо полегают и дают начало ювенильным осям новых порядков. Таким образом возникают жизненные формы «стланик» и «полустланик». При изменении условий освещения ювенильные и имматурные особи стлаников могут сформировать любую из описанных выше жизненных форм.
Подобная схема онтогенеза с небольшими отклонениями присуща и бересклету европейскому. Этот вид, по данным Т. Г. Леоновой (1974), отнесен к быстрорастущим и проходит все этапы онтогенеза на открытых местообитаниях быстрее, чем бересклет бородавчатый в тех же условиях. В соответствующей фитоценотической обстановке онтогенез бересклета европейского осуществляется с образованием таких же биоморф, как и у бересклета бородавчатого (табл. 2).
Т а б л и ц а 2Варианты жизненных форм некоторых лесных кустарников
ВидФитоценотип (тип страте
гии)
Древовиднаяформа
(деревце)
Стланик
КуСТОВИД!
аэроксильныекустарники
(ые формы
геоксильныекустарники
вегета-тквио-непод-вижный
вегета-тивно-
п О ДВИЖИМО
эпигео-генно-
геоксиль-ный
гипогео-генно-
геокснль-ный
Лещина обыкновенная виолент ___ +Бересклет бородавчатый патнент + + + ? + —Бересклет европейский патиент + + + ? + —Жимолость лесная патиент + + + — + —Крушина ломкая патиент + + — + + —Свидина кроваво-красная патиент + + — + + —Бузина красная эксплерент + — + — — —Бузина черная эксплерент + +
Кроме бересклетов бородавчатого и европейского еще несколько видов лесных кустарников характеризуются большим набором жизненных форм. Так, жимолость лесная на опушках, в больших окнах, на вырубках развивается как аэроксильный вегетативно-неподвижный кустарник, реже образует древовидную форму. Под пологом леса в условиях достаточного увлажнения этот вид формирует эпигеогенно-гео- ксильный кустарник, а при очень сильном затенении—стланик и полу- стланик (Мазуренко, 1972, 1977, 1978). Крушина ломкая на свету обычно образует древовидную форму, реже аэроксильный вегетативно- неподвижный кустарник с малым числом осей, а в затенении — эпигео- генно-геоксильный малоосный кустарник или стланик. Корнеотлрыско-4 Бюллетень биологический, выл. 4 73

вый вид свидина кроваво-красная образует те же жизненные формы,, что и бересклеты европейский и бородавчатый. В отличие от этих видов особи вегетативного происхождения возникают у авидины как из спящих почек эпигеогенных корневищ, так и из придаточных почек на горизонтальных корнях, т. е. в последнем случае формируется в е г е т а т и в н о - п о д в и ж н ы й а э р о к с и л ь н ы й кус т .
Несколько иначе проходит онтогенез такого типичного кустарника наших лесов, как лещина обыкновенная. Для нее во взрослом состоянии характерно существование одной жизненной формы — г е о - к с и л ь н о г о г и п о г е о г е н н о г о к у с т а р н и к а . От проростка до имматурного состояния лещина обыкновенная развивается подобно молодому деревцу (Серебряков, 1962). В виргинильном состоянии формируется первичный геоксильный куст в результате образования осей возобновления из подземных (гипокотильных) спящих почек главной оси. Высота растений достигает 2—4 м, а диаметр осей у основания 3—6 см. В корневой системе, постепенно обретающей поверхностный характер, усиливается участие придаточных корней, образующихся в основании осей возобновления, на коротких участках симподиаль- ного ксилоризома, а главный корень и боковые скелетные ослабляют свою деятельность. На этом же этапе онтогенеза из спящих почек в базальной части геоксильного куста развиваются длинные ксилоризо- мы (гипогеогенные корневища до 100 см), образующие из верхушечных почек надземные оси, которые впоследствии дают начало парциальным кустам. Подобные парциальные оси или кусты при механическом отделении от материнского растения начинают развиваться самостоятельно. Они образуют вегетативный ряд особей.
Возникновение и существование короткокор нави щной или длиннокорневищной формы гипогеогенно-геоксильного куста у лещины зависят в основном от эдафических условий (увлажнение, структура почвы, рельеф). Наибольшего развития кусты лещины достигают при переходе в генеративный период. В молодом и средневозрастном генеративном состоянии высота растений 4—6 (и более) м, диаметр стволиков в основании 3—7 (до 12) см. Главная ось начинает суховершинить. Корневая система поверхностная, состоит в основном из придаточных корней, обильно ветвящихся и достигающих 1,5—2,0 м длины. В результате последовательного отмирания осей возобновления первых порядков старый генеративный куст принимает в плане кольцеобразную форму, а затем при переходе в субсенильное и сенильное состояния партикулирует, образуя особи, имеющие 1—2 слабые оси с малым порядком ветвления и небольшими приростами. Подземная часть этих растений представлена остатками отмирающих ксилоризомов с придаточными корнями. В условиях сильного затенения лещина не переходит к образованию стланика. Особи крайне низкой жизненности этого вида также относятся к гипогеогенно-геоксильным кустарникам (Истомина, 1982).
Промежуточное положение по разнообразию жизненных форм занимают такие лесные кустарники, как бузина краоная и черная. В исследованных ценозах под пологом леса и на осветленных местах они чаще всего имеют жизненную форму аэроксильного вегетативно-неподвижного кустарника, реже деревца.
Обсуждение результатовВ представлении И. Г. Серебрякова (1964) одной из отличитель
ных черт биологии большинства кустарников является смена побего- вых систем (осей возобновления) в онтогенезе, что не свойственна
74

большинству деревьев. За счет этой биологической особенности у кустарников постоянно обновляются надземные оси, что при ограниченных размерах растений и при недолговечности их побегов позволяет им довольно долго сохранять свое место в растительных сообществах. Однако этот признак не может быть безоговорочно принят как главная отличительная особенность всех кустарников, так как в условиях, неблагоприятных для их вегетативного разрастания и размножения, эти растения могут длительное время развиваться без смены побего- вых систем, т. е. существовать в древовидной или стланиковой форме. Но причислять на этом основании подобные «кустарники» к «деревьям» вряд ли имеет смысл, так же как и не следует причислять к кустарникам многоствольные деревья, способные неоднократно давать порослевые побеги и сменой побеговых систем напоминающие многие кустарники (Чистякова, 1979). Их отличают от кустарников значительная долговечность наземных осей (стволов) и большие размеры стволов и крон взрослых растений (Крылов, 1984).
Используя принципы классификации И. Г. Серебрякова и его последователей (Серебряков и др., 1954, 1962; Лучник, 1960; Мазуренко, Хохряков, 1977, 1978; Соколова, 1982), мы рассматриваем кустарники как тип жизненных форм, включающий в себя деревянистые растения высотой от 0,5 до 6—10 м. Учитывая разнообразие форм роста в этом типе жизненных форм, мы выделили следующие группы и подгруппы жизненных форм кустарников (рис., 2).
Рис. 2. Жизненные формы лесных кустарников: / — древовидная (деревце); II — кустовидные, I — аэрокенльный кустарник (1а — вегетативно-неподвижный, 16 — вегетативно-подвижный), 2 — геоксильный кустарник (2а — эпнгеогенно-корневнщный,
26 — гипогеогенно-корневнщный); III — стланик
I группа — древовидные формы (деревца)— близки к жизненной форме дерева, имеют более или менее выраженный ствол и крону, косовосходящее или ортотролное направление ветвей. Семенные особи довольно долго сохраняют главный корень.
II группа — кустовидные формы (кустарники)— имеют одновременно несколько стволиков (осей возобновления), возникающих из почек в базальной части куста — зоне кущения.
4* 75

1- я подгруппа — аэроксильные кустарники — зона кущения находится над поверхностью почвы на небольшой высоте, продолжительность жизни стволиков 3-5-10 лет и более. В этой подгруппе можно выделить: а) вегетативно-неподвижный аэроксильный кустарник, у которого стволики расположены компактно и почти не образуют придаточных корней; б) вегетативно-подвижный аэроксильный кустарник,, являющийся системой парциальных кустов корнеотпрыскового происхождения; парциальные образования этой системы могут быть представлены как одиночными стволиками (деревцами), так и кустами.
2- я подгруппа—геоксильные кустарники — зона кущения находится на уровне подстилки или под землей, характерны придаточное укоренение стволиков и наличие вегетативного разрастания с помощью специализированных корневищ: а) эпигеогенно-геоксильный кустарник— базальные части стволиков, возникающих надземно, втягиваются в подстилку благодаря сильному придаточному укоренению,, образуя компактный куст (геоксильно-короткокорневищный — Гатцук, 1974). Полегание и укоренение стволиков или отдельных ветвей (возникновение эпигеогенных корневищ) с последующим развитием на них новых побеговых систем приводят к образованию «рыхлых» геоксиль- но-длиннокорневищных кустов; б) гипогеогенно-теоксильный кустарник (настоящий, по И. Г. Серебрякову, 1962) — новые стволики возникают под землей из спящих почек в базальной части куста (компактная форма) или из верхушечных почек гшюгеогенных корневищ (кси- лоризомов), развивающихся из подземных спящих почек в основании осей, — «рыхлая», длиннокорневищная форма куста. В последнем случае каждый стволик обладает значительно большей самостоятельностью, чем у аэроюсильных кустарников, так как он погружен в почву и формирует свою собственную придаточную корневую систему.
III группа — стланик — постоянно стелющаяся форма; побеги нарастают в основном плагиотропно, период ортотрапного роста побегов относительно невелик (1—2 года); вновь образовавшиеся оси растения полегают и укореняются. К этой же группе мы отнесли биоморфу, которую некоторые авторы называют «полустланик», так как посчитали различия между ними (у полустланика побеги формирования растут ортотротшю, а у стланика — плагиотропно) не столь значимыми для характеристики популяционного поведения кустарников.
По способности иметь тот или иной набор жизненных форм можно выделить следующие группы кустарников. К I группе мы отнесли доминант кустарникового яруса лещину обыкновенную, которая во- взрослом состоянии как в благоприятных, так и в неблагоприятных условиях не выходит за пределы гипогеогенно-геоксильной биоморфы. Стабильность жизненной формы, высокая скорость роста и конкурентная мощность осей, большая продолжительность жизни куста, господство взрослых особей в популяциях — все эти признаки позволяют причислить лещину к конкурентно-сильным видам, виолентам кустарникового яруса.
II группу образуют такие виды, как жимолость лесная, берескле- ты бородавчатый и европейский, крушина ломкая, свидина кроваво- красная. Они отличаются широким набором жизненных форм, т. е. высокой степенью лабильности структуры особи, что помогает им не только выносить давление древесного и доминантов кустарникового ярусов, но и адекватно реагировать на изменение эколого-ценотиче- ских условий. Толерантность (фитоценотическая) этих видов проявляется в том, что в благоприятной ценотичеокой обстановке (вдоль просек, на опушках) они существуют в виде аэроксильного вегетативна-
76

неподвижного кустарника (жимолость, бересклеты, крушина, свиди- на) или в древовидной форме (жимолость, крушина, свидина). Под пологом леса эти виды образуют более «рыхлые» формы — эпигеоген- но-геоксильный кустарник (жимолость, бересклет), аэроксильный вегетативно-подвижный кустарник (крушина, свидина), реже встречаются «деревца» вегетативного происхождения (бересклет). При сильном затенении эти варианты жизненных форм заменяются формой стланика. Многообразие жизненных форм, вегетативная подвижность, способность максимально снижать приросты в условиях стресса — эти признаки характеризуют перечисленные виды как патиенты кустарникового яруса.
В III группу попадают бузина красная и черная. Они отличаются небольшим разнообразием вариантов жизненных форм (табл. 2). Эти кустарники, по нашим наблюдениям, занимают наиболее нарушенные места в фитоценозах (вывалы, вырубки) и относятся по своим биологическим особенностям (большая скорость роста в молодости, мощные, но с коротким циклом побеги формирования, раннее плодоношение, лабильность возрастного состава популяций) к эксплерентным видам.
Заключение
Исследования некоторых черт популяционной жизни лесных кустарников (лещины обыкновенной, бересклета бородавчатого и европейского, жимолости лесной, крушины ломкой, свидины кроваво-красной, бузины красной и черной) показали, что существенное разнообразие жизненных форм этих видов, возникающее в результате поливариантности онтогенеза, не выходит за пределы трех основных групп биоморф: древовидной, кустовидной и стланиковой.
Конвергентное сходство внешнего облика разных видов кустарникового яруса в однородной ценотичеекой обстановке позволяет распространить вышеприведенную классификацию жизненных форм на более широкий крут объектов (на виды родов: калина, черемуха, кара- гана, смородина).
Изучение поливариантности онтогенеза широко распространенных видов кустарников широколиственных и хвойно-широколиственных лесов европейской части СССР показало, что она может рассматриваться как один из механизмов устойчивого существования популяций и синузии в целом. Степень выраженности поливариантности онтогенеза можно оценивать как одно из проявлений типа стратегии вида.
СПИСОК ЛИТЕРАТУРЫ
Г а т ц у к Л. Е. 1974. К методам описания и определения жизненных форм в сезонном климате//Бюл. МОИП. Отд. биол. Т. 79, вып. 3. Ж у к о в а Л. А. 1986. Поливариантность онтогенеза луговых растений//Жизненные формы в экологии и систематике растений. М. 3 а у г о л ь н о в а Л. Б. и др. 1988а. Ценопопуляции растений (очерки популяционной биологии растений). М. З а у г о л ь н о в а Л. Б. и др. 19886. Жизненные формы и популяционное поведение многолетних травянистых растенийу/Экология популяций. Тез. докл. Всесоюз. совещ. Ч. 1. М. И с т о м и н а И. И. 1982. Явление квази- сеннльности и его роль в жизни ценопопуляций на примере лещины обыкновенной/! //Биология, экология и взаимоотношения ценопопуляций растений. М. К р ы л о в А. Г. 1984. Жизненные формы лесных фитоценозов. Л. Л е о н о в а Т. Г. 1974. Бересклеты СССР и сопредельных стран. Л. Л у ч н и к 3. И. 1960. Обрезка кустарников. М. М а з у р е н к о М. Т. 1972. Некоторые особенности морфогенеза трех видов жимолости// ]//Бюл. Глав. бот. сада АН СССР. Вып. 83. М а з у р е н к о М. Т. 1977. Онтогенез жимолости камчатской в оптимальных условиях произрастания//Бюл. МОИП. Отд. биол. Т. 82, вып. 2. М а з у р е н к о М. Т. 1978. Онтогенез жимолости камчатской в условиях глубокого затенения/Дам же. Т. 83, вып. 1. М а з у р е н к о М. Т., Х о х р я к о в А. П.
77

1977. Структура и морфогенез кустарников. М. М а з у р е н к о М . Т., Х о х р я к о в А .П .1978. Экологические аспекты онтогенеза древовидных жизненных форм//Тез. докл. IV делегат, съезда Всесоюз. бот. о-ва. Л. Н и к о л а е в а М. Г. 1956. Биологическое прорастание семян бересклета в связи с его видовым географическим происхождением// //Бот. журн. Т. 41, № 3. Р а б о т н о в Т. А. 1964. Определение возрастного состава по- шуляций видов в сообществе//Полевая геоботаника. Т. 3. М.; Л. С е р е б р я к о в И. Г. 1962. Экологическая морфология растений. М. С е р е б р я к о в И. Г. 1964. Жизненные формы высших растений и их изучение//Полевая геоботаника. Т. 3. М.; Л. С е р е б р я к о в И. Г., Д о м а н с к а я М. Г., Р о д м а н Л. С. 1954. О морфогенезе жизненных форм кустарника на примере орешннка//Бюл. МОИП. Отд. биол. Т. 59, вып. 2. С е р е б р я к о в И. Г., С е р е б р я к о в а Т. И. 1965. О двух типах формирования корневищ у травянистых многолетников//Там же. Т. 70, вып. 2. С м и р н о в а О. В. 1980. Поведение видов и функциональная организация травяного покрова широколиственных лесов европейской части СССР//Там же. Т. 85, вып. 5. С м и р н о в а О. В., Ч и с т я к о в а А. А. 1980. Анализ фитоценотических потенций некоторых древесных видов широколиственных лесов европейской части СССР//Жури. общ. биол. Т. 41, № 3. С о к о л о в а ( Д е р - в и з - С о к о л о в а ) Т. Г. 1982. Морфология нз Северо-Востока СССР в связи С проблемами жизненной формы покрытосеменных растений: Автореф. докт. дис. М. Ч и с т я к о в А. Р. 1935. Биологические особенности бересклета бородавчатого//Сов. бот. № 4. Ч и с т я к о в а А. А. 1979. Большой жизненный цикл Tilia cordata МШ.//)Бюл. МОИП. Отд. бнол. Т. 84, вып. 1. Ч и с т я к о в а А. А. 1986. Жизненные формы деревьев и их эколого-ценотическая обусловленность//Жнзненные формы в экологии и систематике растений. М. Ч и с т я к о в а А. А. 1988. Жизненные формы и их спектры как показа- Уели состояния вида в ценозе (на примере широколиственных деревьев)//Бюл. МОИП. Отд. бнол. Т. 93, вып. 6. G r i m e I. Р. 1979. Plant strategies and vegetation processes. N. Y. etc.
МГПИ им. В. И. Ленина Поступила в редакцию16.05.90
ONTOGENESIS POLYVARIABILITY AND LIFE FORMS OF FOREST SHRUBS
1.I. Istomina, N. N. Bogomolova
Summary
The detailed description of the ontogenesis polyvariability in shrubs is given, being revealed within the platyphyllous and conifer-platyphyllous forests of the European part of the USSR, a comprehensive list of life form varieties in different eco-cenotic conditions being worked out for eight shrub species. Developmental polyvariability is shown to be related with some other features of populational life in the studied species. Polyvariability of ontogenesis is supposed to be one of the main mechanisms of the shrub populational tolerance, as well as of the shrub synusia tolerance in general.




















