
Osmotic Bubble: Creative Insight by dint of Synchronized Atmospheres

Physiologia Plantarum 2014 © 2014 Scandinavian Plant Physiology Society, ISSN 0031-9317
Plasticity of vulnerability to leaf hydraulic dysfunctionduring acclimation to drought in grapevines: anosmotic-mediated processSebastian Martorella, Hipolito Medranoa, Magdalena Tomàsa, José M. Escalonaa, Jaume Flexasa
and Antonio Diaz-Espejob,∗
aResearch Group on Plant Biology under Mediterranean Conditions, Departament de Biologia, Universitat de les Illes Balears, Palma de Mallorca07122, SpainbIrrigation and Crop Ecophysiology Group, Instituto de Recursos Naturales y Agrobiología de Sevilla (IRNAS, CSIC), Sevilla 41012, Spain
Correspondence*Corresponding author,e-mail: [email protected]
Received 5 March 2014;revised 28 May 2014
doi:10.1111/ppl.12253
Previous studies have reported correlation of leaf hydraulic vulnerabilitywith pressure–volume parameters related to cell turgor. This link has beenexplained on the basis of the effects of turgor on connectivity among cellsand tissue structural integrity, which affect leaf water transport. In this study,we tested the hypothesis that osmotic adjustment to water stress would shiftthe leaf vulnerability curve toward more negative water potential (Ψleaf) byincreasing turgor at low Ψleaf. We measured leaf hydraulic conductance (Kleaf),Kleaf vulnerability [50 and 80% loss of Kleaf (P50 and P80); |Ψleaf| at 50 and80% loss of Kleaf, respectively), bulk leaf water relations, leaf gas exchangeand sap flow in two Vitis vinifera cultivars (Tempranillo and Grenache), undertwo water treatments. We found that P50, P80 and maximum Kleaf decreasedseasonally by more than 20% in both cultivars and watering treatments.However, Kleaf at −2 MPa increased threefold, while osmotic potential at fullturgor and turgor loss point decreased. Our results indicate that leaf resistanceto hydraulic dysfunction is seasonally plastic, and this plasticity may bemediated by osmotic adjustment.
Introduction
The ability of plants to draw water from soil to leavesdepends on hydraulic conductance, which is thereforean important limitation to leaf gas exchange (Meinzerand Grantz 1990, Hubbard et al. 2001, Sperry et al.2002, Martorell et al. 2014). A significant part ofwhole-plant hydraulic resistance occurs in leaves, com-monly up to 30% (Sack et al. 2003, Sack and Holbrook2006), and perhaps as high as 80% (Sack and Holbrook2006). As a consequence, leaf hydraulic conductance(Kleaf) is considered a hydraulic bottleneck and is nowunderstood to play a pivotal role in the control of
Abbreviations – ABA, abscisic acid; DW, dry weight; FT, full turgor; LA, leaf area; P–V, pressure–volume; PIP, plasmamembrane intrinsic protein; PPFD, photo-synthetic photon flux density; RWC, relative water content; TLP, turgor loss point;WS, water stressed; WW, well watered.
transpiration (Nardini and Salleo 2000, Brodribb et al.2005, Sack and Tyree 2005). This has stimulated muchresearch on acclimation of Kleaf to changing environ-mental conditions, including temperature (Sellin andKupper 2007), light (Sack et al. 2002, Voicu et al. 2008)and dehydration (Brodribb and Holbrook 2003, Guyotet al. 2012, Scoffoni et al. 2012).
One important aspect of Kleaf responses to environ-ment is the decline of Kleaf with leaf water status duringdehydration, which can result from either cavitationor collapse of xylem conduits (Brodribb and Holbrook2003, Johnson et al. 2009, 2012) or a reduction in thepermeability of extra-xylary tissues (Sack and Holbrook
Physiol. Plant. 2014

2006, Pou et al. 2013). Which of these components ismore limiting to water flow is a matter of debate, andthe answer varies among species (Sack et al. 2004, Nar-dini et al. 2005, Charra-Vaskou et al. 2012). There issome consensus that the hydraulic resistance of leafxylem might be similar in magnitude to the resistanceof the extra-xylary pathways (Sack and Holbrook 2006).However, recently Scoffoni et al. (2014) reported thatthe extra-xylary pathway may be more vulnerable thanthe xylem pathway, thereby delaying the onset of xylemembolism or collapse in leaf veins. Whatever the cause,reduced Kleaf under water stress represents an impor-tant limitation to plant productivity worldwide, impact-ing both species distribution (Engelbrecht et al. 2007,Choat et al. 2012) and agriculture (McElrone et al. 2012).Understanding what controls Kleaf vulnerability is partic-ularly important for improving crop water use efficiencyin the face of climate change (Fereres and Soriano 2007,Ruiz-Sánchez et al. 2010).
Grapevine is one of the most extensively culti-vated and economically important crops in the world.However, most studies on hydraulic conductanceof grapevine have focused on stems (Schultz 2003,Alsina et al. 2007, Lovisolo et al. 2008, Brodersenet al. 2010, 2013, Choat et al. 2010), and fewer stud-ies have focused on petioles (Lovisolo et al. 2008,Zufferey et al. 2011) or leaves (Choat et al. 2009,Pou et al. 2012, 2013). As a result, the impact ofwater stress on Kleaf in grapevine – and particularlythe extra-xylary component – is not well understood.Extra-xylar water flow in grapevine may involve eitherof two routes in parallel: an apoplastic route throughcell walls or a symplastic cell-to-cell route acrossone or more cell membranes, involving plasma mem-brane intrinsic protein (PIP) aquaporins (Cochard et al.2007). Both pathways have been reported to dependon turgor pressure. One mechanism for this depen-dence is the loss of hydraulic connectivity betweencells that occurs when cells shrink during dehydra-tion (Sancho-Knapik et al. 2011, Scoffoni et al. 2012,Scoffoni et al. 2014). Another is the putative regulationof aquaporin activity in relation to cell turgor pres-sure (Cochard et al. 2007, Kim and Steudle 2007).Active osmotic adjustment during acclimation todrought could ameliorate either of these effects byreducing the turgor loss point to more negative waterpotentials.
The objective of this study was to test the hypothe-sis that osmotic adjustment during drought reduces Kleafvulnerability in grapevine. We examined Kleaf variationsalong the growing season in well watered (WW) andwater-stressed (WS) vines of two cultivars with contrast-ing drought tolerance.
Materials and methods
Plant material and experimental conditions
The experiment was conducted in the experimental fieldof the University of Balearic Islands during summer 2012on two cultivars of grapevine (Vitis vinifera) var. Tem-pranillo and Grenache (the last reputed to be moredrought tolerant). Climatic conditions were typical of theMediterranean area and consisted of an average temper-ature of 25.2∘C for the experimental period (June 1 to 30August), ranging from a maximum of 37.3∘C in Augustto a minimum 13.4∘C in June. During this period, meanrelative humidity was 58% and average photo-syntheticphoton flux density (PPFD) at noon 1700 μmol m−2 s−1.Soil was a clay loam type 1.5 m deep. Plants were3-year-old grafted on rootstock Richter-110 and plantedin rows (distance between rows was 2.5 m and plants1 m). They were conducted in a bilateral double cor-don having between 10 and 12 canes per plant. Twotreatments were applied: well watered, in which enoughwater was supplied to replenish an average daily waterloss, as described in Medrano et al. (2012), and waterstressed, in which irrigation was withheld for the entiresummer. The total rainfall over the 3 months of the exper-iment was 0.4 mm.
Leaf water relations
Midday leaf water potential (Ψmd, measured at noon)and predawn water potential (Ψpd, measured just beforesunrise) were measured monthly (on June 26, July 26and August 27) with a pressure chamber (Soil MoistureEquipment Corp., Santa Barbara, CA). Four replicates pertreatment and cultivar were measured.
Leaf hydraulic conductance
Leaf hydraulic conductance (Kleaf) was measured usingthe leaf rehydration kinetics method described by Bro-dribb and Holbrook (2003):
Kleaf = Cbulk ln[Ψo∕Ψf
]∕t (1)
where Cbulk is the leaf capacitance, Ψo is initial waterpotential, Ψf is final water potential after rehydrationand t is time since rehydration began. Leaf vulnera-bility curves and pressure–volume (P–V) curves weremeasured monthly, in conjunction with water poten-tial measurements described above. Sunlit exposedshoots 1.5–2.5 m long were collected in the after-noon, cut under water and wrapped with parafilm atthe cut surface. Once in the laboratory, samples wererecut under water (Wheeler et al. 2013) and left torehydrate overnight. On the following morning, stemswere cut in portions containing three leaves and were
Physiol. Plant. 2014

allowed to desiccate slowly at light intensities sufficientto ensure light-induced hydraulic function (approxi-mately 50 μmol quanta m−2 s−1). The stem portions weredehydrated to varying degrees, including low enoughΨleaf to cause 100% loss of Kleaf. Stem sections withleaves were equilibrated in sealed plastic bags for 2 hbefore measurement, after which two leaves of thesame branch were measured for initial Ψleaf (Ψo). If thedifference between those two leaves was more than0.1 MPa, the sample was discarded. The remaining leafwas rehydrated for 30 s, and final water potential (Ψf)was measured. A rehydration time of 30 s was chosenbased on preliminary rehydration curves on differentleaves attached at the same plant stem. Measurementswere performed in different ranges of water stress andshowed that 30 s of leaf rehydration was enough. Curveswere constructed with a number of replicates between31 and 59.
P–V curves
P–V curves were measured simultaneously with vulner-ability curves to obtain leaf turgor loss point, osmoticadjustment and leaf capacitance using five replicatesper treatment (Tyree and Hammel 1972). Leaves werecollected the evening before measurements and rehy-drated overnight before P–V determination. During thedehydration of leaves in the laboratory, leaf water poten-tial was periodically measured with a pressure chamber(Soil moisture Equipment Corp.), and leaf weight wasmeasured with an analytical balance (Kern ABT320-4M;KERN & SOHN, Balingen, Germany, 10−4 g resolution).The turgor loss point was identified as the inflection pointof the 1/Ψleaf vs relative water content (RWC) curve.Mean cell modulus of elasticity (𝜖) was estimated as theslope of turgor potential (Ψp) vs RWC through the phasefrom full turgor to turgor loss point. The fitting methodproposed by Sack et al. (2011) was used to fit the P–Vcurves. This method fits lines by the standard major axis(model II regression) so that either variable can be pre-dicted from the other, and the parameter calculation ismore robust.
Bulk leaf capacitance (Cbulk, mol m−2 MPa−1) wasestimated according to Blackman and Brodribb (2011)as follows:
Cbulk = 𝛿RWC∕𝛿Ψleaf × (DW∕LA) × (WWL∕DW) ∕M(2)
where DW is leaf dry weight (g), LA is leaf area (m2),WWL is mass of leaf water at RWC= 1.0 (g) and Mis the molar mass of water. Cbulk was determined fromthe slopes of linear regressions fitted to data on eitherside of the inflection point in 1/Ψleaf vs RWC from P–Vcurves data. These preturgor and postturgor loss slopes
were averaged for each treatment and sampling timeto get a representative value for each treatment andcultivar. Ratios of DW/LA and WWL/DW were measuredfor each cultivar and treatment at the different samplingtimes mentioned above in 10 replicates per cultivar andtreatment. The average value of capacitance on eitherside of the inflection point in 1/Ψleaf vs RWC was used inthe estimation of Kleaf data.
Leaf gas exchange measurements
Leaf gas exchange was measured using a portable openflow gas exchange system (Li-6400; Li-Cor, Inc., Lincoln,NE) equipped with a clear chamber (Li-6400-08). Mea-surements were made at ambient air temperature andPPFD and CO2 concentration of 400 μmol mol−1. Diur-nal time courses of stomatal conductance (gs) were mea-sured in June, July and August in mature leaves. Fivereplicates per treatment were measured hourly.
Sap flow measurements
Sap flow was measured by the stem heat balance methodusing the standard Sap Flow meter P 4.1 from Environ-mental Measuring Systems (EMS, Brno, Czech Republic),with the same protocol as described in Escalona et al.(2002). The mass flow of sap was estimated from the bal-ance of heat fluxes into and out of the heated section ofthe stem (Sakuratani 1981, Baker and Van Bavel 1987).Sensors were insulated with 2 cm thick open-porouspolyurethane foam and protected from radiation withaluminum foil. Four stem gauges were installed for eachtreatment and cultivar. Leaf area of each stem was mea-sured at the end of the experiment using a digital camera(Olympus Corp, Hamburg, Germany; SP800UZ) and theimages analyzed using IMAGE J (http://imagej.nih.gov/ij/).Sap flow measurements were expressed on a LA basis.
Statistical analysis
Statistical differences between treatment means werecalculated using (ANOVA), and significant effects were fol-lowed up using post hoc multiple comparison tests ofDuncan (P< 0.05). Dixon tests were applied to Kleaf datato identify possible outliers. Statistical analyses were per-formed in SPSS 19.0 (IBM Corp., Armonk, NY). To clar-ify the differences between sampling dates, Kleaf datawere grouped into bins by water potential, each with arange of 0.3–0.5 MPa and 3–8 measurements. For sta-tistical prediction, sigmoidal curves (y = y0 +
a
1+e−(x−x0
b ) )
were fitted through nongrouped Kleaf vs Ψleaf data usingSIGMAPLOT 8.0 (Systat Software, Chicago, IL) as has beenreported in other studies (Johnson et al. 2012, Scoffoniet al. 2012).
Physiol. Plant. 2014

Grenache WS
–3–2–100
5
10
15
20
25 JuneJulyAugust
Tempranillo WS
–4–3–2–10
Grenache WW
0
5
10
15
20
25
30Tempranillo WW
leaf (MPa)
Kle
af (
mm
ol m
–2 s
–1 M
Pa–1
)
Fig. 1. Vulnerability curves for both cultivars and water treatments in June, July and August. Lines correspond to the sigmoidal curve: y =y0 + a
1+e−( x−x0b ) fitted to individual measurements of Kleaf. Data were binned in this case to clarify the differences between different months. Each
point represents the average of at least three measurements. Vertical bars represent± SE.
Results
Seasonal changes in Kleaf
Maximum Kleaf (Kleaf,max) declined over the summer, andthe water potential causing 50 and 80% loss of Kleaf(P50 and P80, respectively) became more negative as thesummer progressed, indicating a progressive decrease inhydraulic vulnerability to dysfunction (Fig. 1, Table 1).Kleaf,max declined in both cultivars and watering treat-ments, with the exception of Grenache WW, in whichKleaf,max was similar in July and August. Kleaf,max was sim-ilar between cultivars. As the percentage loss of conduc-tance at a given Ψleaf can be misinterpreted when Kleaf,maxdoes not remain constant, we also analyzed the seasonalprogression of Kleaf at a reference Ψleaf of −2 MPa (Fig. 2).Kleaf at −2 MPa increased as summer progressed in bothcultivars and water treatments, by up to 200% (Fig. 2).This trend in Kleaf at −2 MPa was also found at other ref-erence Ψleaf values like −1.8 or −2.3 MPa. However, onlyat −2 MPa we were able to get enough points of the twocultivars, water treatments and all three dates to make areliable and robust comparison among them.
Kleaf and leaf water relations
Leaf water potential at mid-day (Ψmd) and predawn(Ψpd) declined along the summer (Table 2). Water stress
reduced Ψpd in August to a minimum of −0.85 MPa inGrenache and −0.53 MPa in Tempranillo. A similar pat-tern was observed in Ψmd, which reached −1.88 MPa inGrenache and −1.64 MPa in Tempranillo. As a result, ΔΨ(=Ψpd – Ψmd) was relatively constant along the season forboth cultivars and treatments, with slightly larger valuesin Tempranillo.
Osmotic potential at full turgor (Ψ𝜋,FT) and at the
turgor loss point (Ψ𝜋,TLP) were also lower (more nega-
tive) in July and August than in June (Table 3), whichallowed leaves to maintain turgor at lower Ψleaf laterin summer and in the WS treatments. Cell modulusof elasticity (𝜖) varied between 10 and 16 MPa duringseason and between cultivars (Table 3). Furthermore,the combination of Ψ
𝜋,FT and 𝜖 allowed the plant tokeep RWC at turgor loss point (RWCTLP) at relativelyhigh values (between 88 and 92%) along the season.The minimum Ψleaf (Table 2) reached by each cultivarseasonally was correlated with Ψ
𝜋,TLP, and it was alwayshigher than Ψ
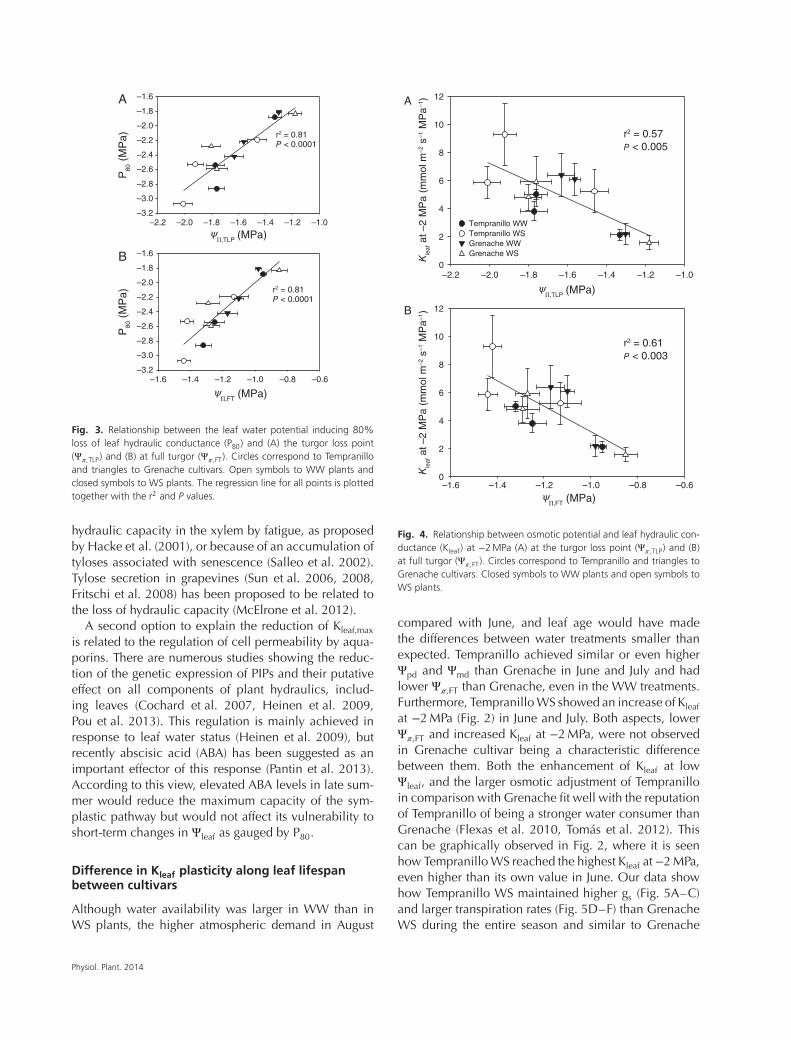
𝜋,TLP (Table 3). We observed strong andpositive linear relationships between P80 and both Ψ
𝜋,TLPand Ψ
𝜋,FT (Fig. 3A, B; P<0.001, r2 = 0.81), indicatinga decrease in hydraulic vulnerability to dysfunctionconcurrent with osmotic adjustment. Kleaf at −2 MPawas also correlated with Ψ
𝜋,TLP and Ψ𝜋,FT (Fig. 4A,
B; P< 0.005, r2 = 0.57; P< 0.003, r2 = 0.61, respec-tively), demonstrating that Kleaf at −2 MPa increased
Physiol. Plant. 2014
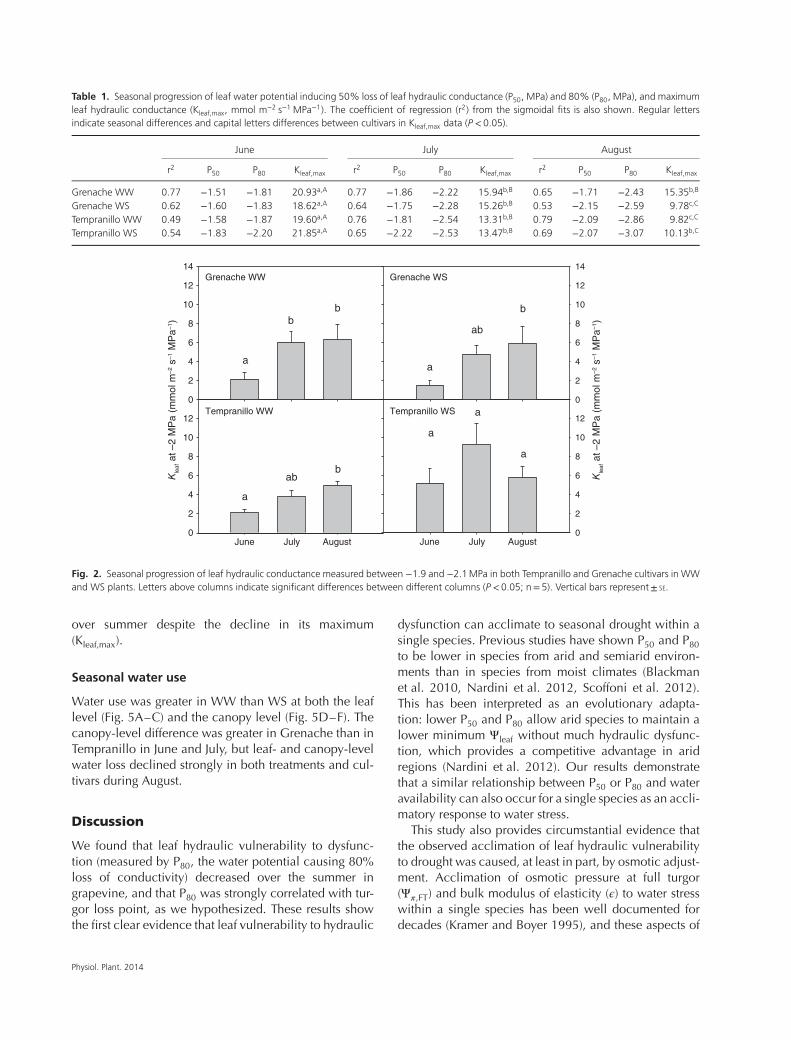
Table 1. Seasonal progression of leaf water potential inducing 50% loss of leaf hydraulic conductance (P50, MPa) and 80% (P80, MPa), and maximumleaf hydraulic conductance (Kleaf,max, mmol m−2 s−1 MPa−1). The coefficient of regression (r2) from the sigmoidal fits is also shown. Regular lettersindicate seasonal differences and capital letters differences between cultivars in Kleaf,max data (P <0.05).
June July August
r2 P50 P80 Kleaf,max r2 P50 P80 Kleaf,max r2 P50 P80 Kleaf,max
Grenache WW 0.77 −1.51 −1.81 20.93a,A 0.77 −1.86 −2.22 15.94b,B 0.65 −1.71 −2.43 15.35b,B
Grenache WS 0.62 −1.60 −1.83 18.62a,A 0.64 −1.75 −2.28 15.26b,B 0.53 −2.15 −2.59 9.78c,C
Tempranillo WW 0.49 −1.58 −1.87 19.60a,A 0.76 −1.81 −2.54 13.31b,B 0.79 −2.09 −2.86 9.82c,C
Tempranillo WS 0.54 −1.83 −2.20 21.85a,A 0.65 −2.22 −2.53 13.47b,B 0.69 −2.07 −3.07 10.13b,C
Tempranillo WW
June July August0
2
4
6
8
10
12
a
abb
Tempranillo WS
June July August0
2
4
6
8
10
12
a
a
a
Grenache WW
0
2
4
6
8
10
12
14Grenache WS
0
2
4
6
8
10
12
14
b
ab
a
bb
a
Kle
af a
t –2
MP
a (m
mol
m–2
s–1
MP
a–1)
Kle
af a
t –2
MP
a (m
mol
m–2
s–1
MP
a–1)
Fig. 2. Seasonal progression of leaf hydraulic conductance measured between −1.9 and −2.1 MPa in both Tempranillo and Grenache cultivars in WWand WS plants. Letters above columns indicate significant differences between different columns (P <0.05; n=5). Vertical bars represent± SE.
over summer despite the decline in its maximum(Kleaf,max).
Seasonal water use
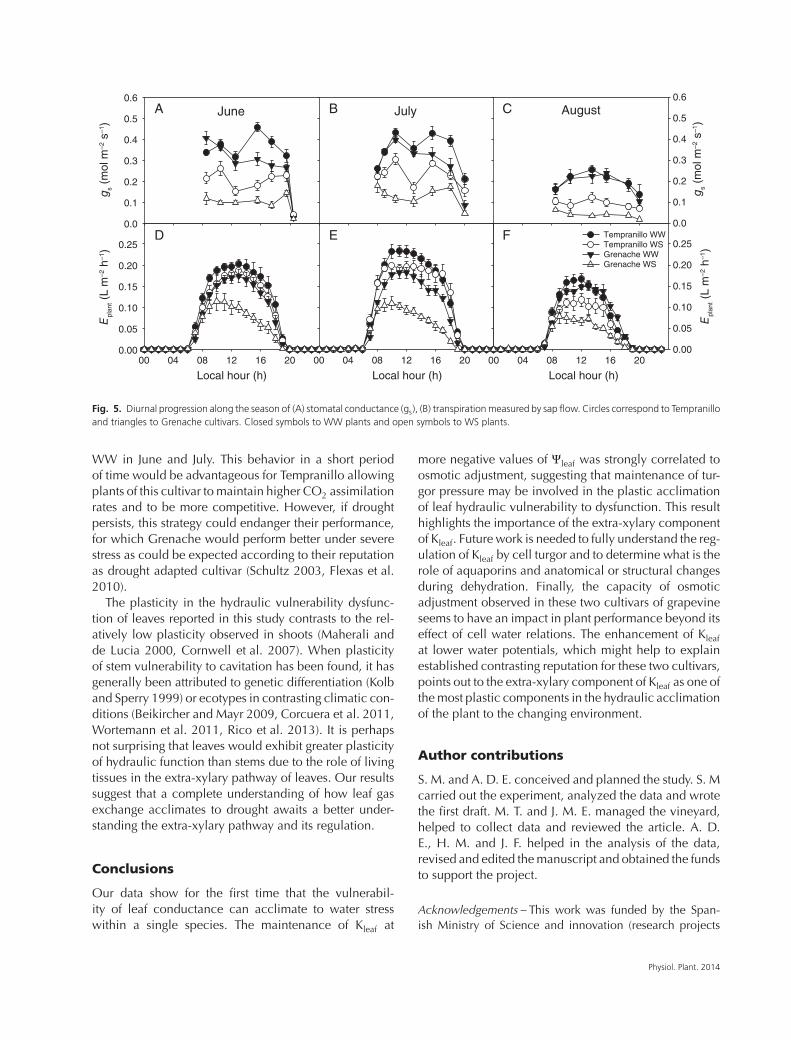
Water use was greater in WW than WS at both the leaflevel (Fig. 5A–C) and the canopy level (Fig. 5D–F). Thecanopy-level difference was greater in Grenache than inTempranillo in June and July, but leaf- and canopy-levelwater loss declined strongly in both treatments and cul-tivars during August.
Discussion
We found that leaf hydraulic vulnerability to dysfunc-tion (measured by P80, the water potential causing 80%loss of conductivity) decreased over the summer ingrapevine, and that P80 was strongly correlated with tur-gor loss point, as we hypothesized. These results showthe first clear evidence that leaf vulnerability to hydraulic
dysfunction can acclimate to seasonal drought within asingle species. Previous studies have shown P50 and P80to be lower in species from arid and semiarid environ-ments than in species from moist climates (Blackmanet al. 2010, Nardini et al. 2012, Scoffoni et al. 2012).This has been interpreted as an evolutionary adapta-tion: lower P50 and P80 allow arid species to maintain alower minimum Ψleaf without much hydraulic dysfunc-tion, which provides a competitive advantage in aridregions (Nardini et al. 2012). Our results demonstratethat a similar relationship between P50 or P80 and wateravailability can also occur for a single species as an accli-matory response to water stress.
This study also provides circumstantial evidence thatthe observed acclimation of leaf hydraulic vulnerabilityto drought was caused, at least in part, by osmotic adjust-ment. Acclimation of osmotic pressure at full turgor(Ψ
𝜋,FT) and bulk modulus of elasticity (𝜖) to water stresswithin a single species has been well documented fordecades (Kramer and Boyer 1995), and these aspects of
Physiol. Plant. 2014

Table 2. Predawn leaf water potential (Ψpd, MPa), midday water poten-tial (Ψmd, MPa) and soil to leaf water potential gradient (ΔΨ=Ψmd −Ψpd,MPa) measured at both treatments in June, July and August. Letters indi-cate for significant differences (P <0.05) among the different dates onthe same treatment.
June July August
Ψpd
Grenache WW −0.16±0.02a −0.27±0.02a −0.24±0.06a
Grenache WS −0.31±0.02a −0.55±0.02b −0.85±0.07c
Tempranillo WW −0.16±0.02a −0.33±0.02b −0.30±0.04b
Tempranillo WS −0.28±0.01a −0.48±0.02b −0.53±0.02c
Ψmd
Grenache WW −1.07±0.03a −1.05±0.06a −1.00±0.04a
Grenache WS −1.13±0.05a −1.60±0.01b −1.88±0.04c
Tempranillo WW −1.09±0.01a −1.28±0.05b −1.11±0.06ab
Tempranillo WS −1.27±0.04a −1.59±0.03b −1.64±0.11b
ΔΨGrenache WW 0.91±0.05a 0.78±0.08a 0.76±0.08a
Grenache WS 0.82±0.04a 1.06± 0.03b 1.04± 0.04b
Tempranillo WW 0.94±0.02a 0.95±0.06a 0.80±0.08a
Tempranillo WS 0.99±0.04a 1.12±0.04a 1.11±0.11a
leaf acclimation to water stress may help to maintain theconductivity of the extra-xylary pathway for water flow intwo ways. First, maintaining turgor may prevent exces-sive cell shrinkage during water stress (Sancho-Knapiket al. 2011, Scoffoni et al. 2012, Scoffoni et al. 2014),thereby maintaining cell-to-cell pathways (symplasticand transcellular paths) and preventing a reduction inevaporative surface in the intercellular spaces (Scof-foni et al. 2014). Second, turgor may directly modulateaquaporin function, so maintenance of turgor may berequired to sustain high conductivity of the symplasticpathway (Cochard et al. 2007, Kim and Steudle 2007).This hypothesis is supported by the strong relationshipthat we found between Ψ
𝜋,TLP and P80. Furthermore,these changes in P80 were not caused by changes in max-imum Kleaf (which also declined over the summer), asdemonstrated by the fact that Kleaf at a reference waterpotential of −2.0 MPa actually increased over the sum-mer, and still maintained the correlation with Ψ
𝜋,TLP andΨ
𝜋,FT (Fig. 4).Although it is widely interpreted that concomitant
changes in 𝜖 have an effect on Ψ𝜋,TLP, Bartlett et al.
(2012) recently demonstrated that 𝜖 changes vary widelybetween studies of drought stress and play a secondaryrole in driving drought tolerance as some times val-ues can increment or decrease in water stress condi-tions. This reinforce the hypothesis that changes in Ψ
𝜋,TLPare driven primarily by osmotic adjustment (which isreflected by changes in Ψ
𝜋,FT; Bartlett et al. 2012). How-ever, 𝜖 could play a role in preventing loss of Kleaf at lowΨleaf, because the decline in Kleaf caused by shrinkageduring dehydration (Scoffoni et al. 2014) is likely caused
Table 3. Leaf water relation parameters obtained by P–V curves atJune, July and August in both cultivars and water treatments. Osmoticpotential at full turgor (Ψ
𝜋,FT, MPa), turgor loss point (Ψ𝜋,TLP, MPa),
relative water content at turgor loss point (RWCTLP, %) and mean bulkmodulus of elasticity (𝜖, MPa). Values are means of five samples± SE.Letters indicate for differences between dates in the same treatment andcultivar (P < 0.05).
June July August
Grenache WWΨ
𝜋,FT −0.98±0.02b −1.10±0.03ab −1.17±0.06a
Ψ𝜋,TLP −1.30±0.02b −1.56±0.03a −1.63±0.07a
RWCTLP 92.52±0.33a 89.85±0.56b 90.28±0.66ab
𝜖 12.46±0.60ab 10.85±0.67b 13.59± 0.50a
Grenache WSΨ
𝜋,FT −0.85±0.05b −1.29±0.07a −1.27±0.05a
Ψ𝜋,TLP −1.18±0.05b −1.80±0.07a −1.76±0.08a
RWCTLP 92.56±0.59b 88.53± 0.92a 88.26± 0.91a
𝜖 11.23±0.73a 12.14± 0.96a 9.98± 0.50a
Tempranillo WWΨ
𝜋,FT −0.95±0.02b −1.25±0.06a −1.32±0.05a
Ψ𝜋,TLP −1.33±0.05b −1.77±0.07a −1.76±0.06a
RWCTLP 91.37±0.90a 89.99±0.74a 91.74± 0.64a
𝜖 10.53±1.35a 12.74±1.45ab 16.45±1.05b
Tempranillo WSΨ
𝜋,FT −1.13±0.09b −1.42±0.04a −1.44±0.04a
Ψ𝜋,TLP −1.46±0.07b −1.92±0.06a −2.01±0.07a
RWCTLP 90.57±0.73a 88.79±1.25a 89.16± 0.70a
𝜖 12.95±0.99a 13.49±1.16a 13.09± 0.54a
by reduced cell volume rather than turgor pressure perse. This secondary role of 𝜖 in the response to drought ofleaves, i.e. facilitation of Kleaf at lower Ψleaf by the main-tenance of higher RWC and preventing cell dehydration,might have gone unnoticed by Bartlett et al. (2012) wheninterpreting their results on the acclimation to drought. Inany case, the role of cell volume on Kleaf deserves furtherexperimental support.
Although we did not partition Kleaf into xylem andextra-xylary components in this study, changes in thexylem of petioles or minor leaf veins are unlikely toexplain the changes in P80 and in Kleaf at −2.0 MPaobserved between June and August. All leaves were ofsimilar age, and acclimation during ontogeny, whichcould have allowed for the construction of a veinnetwork of different vulnerability, cannot be involved.Although it has been reported that changes in the ioniccomposition of the sap might enhance the hydraulicconductivity of xylem vessels (Nardini et al. 2011), theincrease as high as the one observed in this study hasnever been reported (Fig. 2). The most parsimoniousexplanation to both the decrease of P80 and the increaseof Kleaf at −2 MPa is the shift of the vulnerability curvetoward more negative Ψleaf values, in line with ourinitial hypothesis. On the other hand, the decline inKleaf,max over the summer could be because of the loss of
Physiol. Plant. 2014
–2.2 –2.0 –1.8 –1.6 –1.4 –1.2 –1.0–3.2
–3.0
–2.8
–2.6
–2.4
–2.2
–2.0
–1.8
–1.6
–1.6 –1.4 –1.2 –1.0 –0.8 –0.6–3.2
–3.0
–2.8
–2.6
–2.4
–2.2
–2.0
–1.8
–1.6
r2 = 0.81P < 0.0001
r2 = 0.81P < 0.0001
A
B
P80
(M
Pa)
P80
(M
Pa)
�,FT (MPa)
�,TLP (MPa)
Fig. 3. Relationship between the leaf water potential inducing 80%loss of leaf hydraulic conductance (P80) and (A) the turgor loss point(Ψ
𝜋,TLP) and (B) at full turgor (Ψ𝜋,FT). Circles correspond to Tempranillo
and triangles to Grenache cultivars. Open symbols to WW plants andclosed symbols to WS plants. The regression line for all points is plottedtogether with the r2 and P values.
hydraulic capacity in the xylem by fatigue, as proposedby Hacke et al. (2001), or because of an accumulation oftyloses associated with senescence (Salleo et al. 2002).Tylose secretion in grapevines (Sun et al. 2006, 2008,Fritschi et al. 2008) has been proposed to be related tothe loss of hydraulic capacity (McElrone et al. 2012).
A second option to explain the reduction of Kleaf,maxis related to the regulation of cell permeability by aqua-porins. There are numerous studies showing the reduc-tion of the genetic expression of PIPs and their putativeeffect on all components of plant hydraulics, includ-ing leaves (Cochard et al. 2007, Heinen et al. 2009,Pou et al. 2013). This regulation is mainly achieved inresponse to leaf water status (Heinen et al. 2009), butrecently abscisic acid (ABA) has been suggested as animportant effector of this response (Pantin et al. 2013).According to this view, elevated ABA levels in late sum-mer would reduce the maximum capacity of the sym-plastic pathway but would not affect its vulnerability toshort-term changes in Ψleaf as gauged by P80.
Difference in Kleaf plasticity along leaf lifespanbetween cultivars
Although water availability was larger in WW than inWS plants, the higher atmospheric demand in August
A
–2.2 –2.0 –1.8 –1.6 –1.4 –1.2 –1.00
2
4
6
8
10
12
B
–1.6 –1.4 –1.2 –1.0 –0.8 –0.60
2
4
6
8
10
12
Tempranillo WWTempranillo WSGrenache WWGrenache WS
P < 0.005r2 = 0.57
P < 0.003r2 = 0.61
Kle
af a
t –2
MP
a (m
mol
m–2
s–1
MP
a–1)
Kle
af a
t –2
MP
a (m
mol
m–2
s–1
MP
a–1)
�,FT (MPa)
�,TLP (MPa)
Fig. 4. Relationship between osmotic potential and leaf hydraulic con-ductance (Kleaf) at −2 MPa (A) at the turgor loss point (Ψ
𝜋,TLP) and (B)at full turgor (Ψ
𝜋,FT). Circles correspond to Tempranillo and triangles toGrenache cultivars. Closed symbols to WW plants and open symbols toWS plants.
compared with June, and leaf age would have madethe differences between water treatments smaller thanexpected. Tempranillo achieved similar or even higherΨpd and Ψmd than Grenache in June and July and hadlower Ψ
𝜋,FT than Grenache, even in the WW treatments.Furthermore, Tempranillo WS showed an increase of Kleafat −2 MPa (Fig. 2) in June and July. Both aspects, lowerΨ
𝜋,FT and increased Kleaf at −2 MPa, were not observedin Grenache cultivar being a characteristic differencebetween them. Both the enhancement of Kleaf at lowΨleaf, and the larger osmotic adjustment of Tempranilloin comparison with Grenache fit well with the reputationof Tempranillo of being a stronger water consumer thanGrenache (Flexas et al. 2010, Tomás et al. 2012). Thiscan be graphically observed in Fig. 2, where it is seenhow Tempranillo WS reached the highest Kleaf at −2 MPa,even higher than its own value in June. Our data showhow Tempranillo WS maintained higher gs (Fig. 5A–C)and larger transpiration rates (Fig. 5D–F) than GrenacheWS during the entire season and similar to Grenache
Physiol. Plant. 2014
D
Local hour (h) Local hour (h) Local hour (h)00 04 08 12 16 20
0.00
0.05
0.10
0.15
0.20
0.25E
00 04 08 12 16 20
Tempranillo WWTempranillo WSGrenache WWGrenache WS
F
00 04 08 12 16 20
A
0.0
0.1
0.2
0.3
0.4
0.5
0.6B C
Tempranillo WWTempranillo WSGrenache WWGrenache WS
June July August
0.00
0.05
0.10
0.15
0.20
0.25
0.0
0.1
0.2
0.3
0.4
0.5
0.6g s
(m
ol m
–2 s
–1)
Epl
ant (
L m
–2 h
–1)
g s (
mol
m–2
s–1
)E
plan
t (L
m–2
h–1
)
Fig. 5. Diurnal progression along the season of (A) stomatal conductance (gs), (B) transpiration measured by sap flow. Circles correspond to Tempranilloand triangles to Grenache cultivars. Closed symbols to WW plants and open symbols to WS plants.
WW in June and July. This behavior in a short periodof time would be advantageous for Tempranillo allowingplants of this cultivar to maintain higher CO2 assimilationrates and to be more competitive. However, if droughtpersists, this strategy could endanger their performance,for which Grenache would perform better under severestress as could be expected according to their reputationas drought adapted cultivar (Schultz 2003, Flexas et al.2010).
The plasticity in the hydraulic vulnerability dysfunc-tion of leaves reported in this study contrasts to the rel-atively low plasticity observed in shoots (Maherali andde Lucia 2000, Cornwell et al. 2007). When plasticityof stem vulnerability to cavitation has been found, it hasgenerally been attributed to genetic differentiation (Kolband Sperry 1999) or ecotypes in contrasting climatic con-ditions (Beikircher and Mayr 2009, Corcuera et al. 2011,Wortemann et al. 2011, Rico et al. 2013). It is perhapsnot surprising that leaves would exhibit greater plasticityof hydraulic function than stems due to the role of livingtissues in the extra-xylary pathway of leaves. Our resultssuggest that a complete understanding of how leaf gasexchange acclimates to drought awaits a better under-standing the extra-xylary pathway and its regulation.
Conclusions
Our data show for the first time that the vulnerabil-ity of leaf conductance can acclimate to water stresswithin a single species. The maintenance of Kleaf at
more negative values of Ψleaf was strongly correlated toosmotic adjustment, suggesting that maintenance of tur-gor pressure may be involved in the plastic acclimationof leaf hydraulic vulnerability to dysfunction. This resulthighlights the importance of the extra-xylary componentof Kleaf. Future work is needed to fully understand the reg-ulation of Kleaf by cell turgor and to determine what is therole of aquaporins and anatomical or structural changesduring dehydration. Finally, the capacity of osmoticadjustment observed in these two cultivars of grapevineseems to have an impact in plant performance beyond itseffect of cell water relations. The enhancement of Kleafat lower water potentials, which might help to explainestablished contrasting reputation for these two cultivars,points out to the extra-xylary component of Kleaf as one ofthe most plastic components in the hydraulic acclimationof the plant to the changing environment.
Author contributions
S. M. and A. D. E. conceived and planned the study. S. Mcarried out the experiment, analyzed the data and wrotethe first draft. M. T. and J. M. E. managed the vineyard,helped to collect data and reviewed the article. A. D.E., H. M. and J. F. helped in the analysis of the data,revised and edited the manuscript and obtained the fundsto support the project.
Acknowledgements – This work was funded by the Span-ish Ministry of Science and innovation (research projects
Physiol. Plant. 2014

AGL2008-04525-C02-01, AGL2009-11310/AGR andAGL2011-30408-C04-01). S. M. benefited from a Forma-ción personal investigador (FPI) grant BES-2009-016906from the Spanish Ministry of Science and Innovation.We thank Tim Brodribb for stimulating discussions andintroducing us to the measurements of leaf hydraulics. Wealso thank Dr Pep Cifre for his help on statistics and TomBuckley for helpful comments on the manuscript and forassistance with English editing.
References
Alsina MM, De Herralde F, Aranda X, Save R, Biel CC(2007) Water relations and vulnerability to embolism arenot related: experiments with eight grapevine cultivars.Vitis 46: 1–7
Baker JM, Van Bavel CHM (1987) Measurement of massflow of water in the stems of herbaceous plants. PlantCell Environ 10: 777–782
Bartlett MK, Scoffoni C, Sack L (2012) The determinants ofleaf turgor loss point and prediction of drought toleranceof species and biomes: a global meta-analysis. Ecol Lett15: 393–405
Beikircher B, Mayr S (2009) Intraspecific differences indrought tolerance and acclimation in hydraulics ofLigustrum vulgare and Viburnum lantana. Tree Physiol29: 765–775
Blackman CJ, Brodribb TJ (2011) Two measures of leafcapacitance: insights into the water transport pathwayand hydraulic conductance in leaves. Funct Plant Biol38: 118–126
Blackman CJ, Brodribb TJ, Jordan GJ (2010) Leaf hydraulicvulnerability is related to conduit dimensions anddrought resistance across a diverse range of woodyangiosperms. New Phytol 188: 1113–1123
Brodersen CR, McElrone AJ, Choat B, Matthews MA,Shackel KA (2010) The dynamics of embolism repair inxylem: in vivo visualizations using high-resolutioncomputed tomography. Plant Physiol 154: 1088–1095
Brodersen CR, Choat B, Chatelet DS, Shackel KA,Matthews MA, McElrone AJ (2013) Xylem vessel relayscontribute to radial connectivity in grapevine stems (Vitisvinifera and V. arizonica; Vitaceae). Am J Bot 100:314–332
Brodribb TJ, Holbrook NM (2003) Stomatal closure duringleaf dehydration, correlation with other leafphysiological traits. Plant Physiol 132: 2166–2173
Brodribb TJ, Holbrook NM, Zwieniecki MA, Palma B(2005) Leaf hydraulic capacity in ferns, conifers andangiosperms: impacts on photosynthetic maxima. NewPhytol 165: 839–846
Charra-Vaskou K, Badel E, Burlett R, Cochard H, Delzon S,Mayr S (2012) Hydraulic efficiency and safety ofvascular and non-vascular components in Pinus pinasterleaves. Tree Physiol 32: 1161–1170
Choat B, Gambetta GA, Wada H, Shackel KA, MatthewsMA (2009) The effects of Pierce’s disease on leaf andpetiole hydraulic conductance in Vitis vinifera cv.Chardonnay. Physiol Plant 136: 384–394
Choat B, Drayton WM, Brodersen C, Matthews MA,Shackel KA, Wada H, McElrone AJ (2010) Measurementof vulnerability to water stress-induced cavitation ingrapevine: a comparison of four techniques applied to along-vesseled species. Plant Cell Environ 33:1502–1512
Choat B, Jansen S, Brodribb TJ, Cochard H, Delzon S,Bhaskar R, Bucci SJ, Field TS, Gleason SM, Hacke UG,Jacobsen AL, Lens F, Maherali H, Martínez-Vilalta J,Mayr S, Mencuccini M, Mitchell PJ, Nardini A,Pittermann J, Pratt RB, Sperry JS, Westoby M, Wright IJ,Zanne AE (2012) Global convergence in thevulnerability of forests to drought. Nature 491: 752–756
Cochard H, Venisse JS, Barigah TS, Brunel N, Herbette S,Guilliot A, Tyree MT, Sakr S (2007) Putative role ofaquaporins in variable hydraulic conductance of leavesin response to light. Plant Physiol 143: 122–133
Corcuera L, Cochard H, Gil-Pelegrin E, Notivol E (2011)Phenotypic plasticity in mesic populations of Pinuspinaster improves resistance to xylem embolism (P50)under severe drought. Trees 25: 1033–1042
Cornwell WK, Bhaskar R, Sack L, Cordell S (2007)Adjustment of structure and function of HawaiianMetrosideros polymorpha at high vs. low precipitation.Funct Ecol 21: 1063–1071
Engelbrecht BM, Comita LS, Condit R, Kursar TA, TyreeMT, Turner BL, Hubbell SP (2007) Drought sensitivityshapes species distribution patterns in tropical forests.Nature 447: 80–82
Escalona J, Flexas J, Medrano H (2002) Drought effects onwater flow, photosynthesis and growth of pottedgrapevines. Vitis 41: 57–62
Fereres E, Soriano MA (2007) Deficit irrigation for reducingagricultural water use. J Exp Bot 58: 147–159
Flexas J, Galmés J, Gallé A, Gulias J, Pou A, Ribas-CarbóM, Tomàs M, Medrano H (2010) Improving water useefficiency in grapevines: potential physiological targetsfor biotechnological improvement. Aust J Grape WineRes 16: 106–121
Fritschi FB, Lin H, Walker MA (2008) Scanning electronmicroscopy reveals different response pattern of fourVitis genotypes to Xylella fastidiosa infection. Plant Dis92: 276–286
Guyot G, Scoffoni C, Sack L (2012) Combined impacts ofirradiance and dehydration on leaf hydraulicconductance: insights into vulnerability and stomatalcontrol. Plant Cell Environ 35: 857–871
Hacke UG, Stiller V, Sperry JS, Pittermann J, McCulloh KA(2001) Cavitation fatigue. Embolism and cycles canweaken cavitation resistance of xylem. Plant Physiol125: 779–786
Physiol. Plant. 2014

Heinen RB, Ye Q, Chaumont F (2009) Role of aquaporinsin leaf physiology. J Exp Bot 60: 2971–2985
Hubbard RM, Ryan MG, Stiller V, Sperry JS (2001)Stomatal conductance and photosynthesis vary linearlywith plant hydraulic conductance in Ponderosa pine.Plant Cell Environ 24: 113–121
Johnson DM, Meinzer FC, Woodruff DR, McCulloh KA(2009) Leaf xylem embolism, detected acoustically andby cryo-SEM, corresponds to decreases in leaf hydraulicconductance in four evergreen species. Plant CellEnviron 32: 828–836
Johnson DM, McCulloh KA, Woodruff DR, Meinzer FC(2012) Evidence for xylem embolism as a primary factorin dehydration-induced declines in leaf hydraulicconductance. Plant Cell Environ 35: 760–769
Kim YX, Steudle E (2007) Light and turgor affect the waterpermeability (aquaporins) of parenchyma cells in themidrib of leaves of Zea mays. J Exp Bot 58: 4119–4129
Kolb KJ, Sperry JS (1999) Differences in drought adaptationbetween subspecies of sagebrush (Artemisia tridentata).Ecology 80: 2373–2384
Kramer PJ, Boyer JS (1995) Water Relations of Plants andSoils. Elsevier Academic Press, London
Lovisolo C, Perrone I, Hartung W, Schubert A (2008) Anabscisic acid-related reduced transpiration promotesgradual embolism repair when grapevines arerehydrated after drought. New Phytol 180: 642–651
Maherali H, De Lucia EH (2000) Xylem conductivity andvulnerability to cavitation of ponderosa pine growing incontrasting climates. Tree Physiol 20: 859–867
Martorell S, Diaz-Espejo A, Medrano H, Ball MC, Choat B(2014) Rapid hydraulic recovery in Eucalyptus paucifloraafter drought: linkages between stem hydraulics and leafgas exchange. Plant Cell Environ 37: 617–626
McElrone AJ, Brodersen CR, Alsina MM, Drayton WM,Matthews MA, Shackel KA, Wada H, Zufferey V, ChoatB (2012) Centrifuge technique consistently overestimatesvulnerability to water stress-induced cavitation ingrapevines as confirmed with high-resolution computedtomography. New Phytol 196: 661–665
Medrano H, Pou A, Tomás M, Martorell S, Gulias J, FlexasJ, Escalona JM (2012) Average daily light interceptiondetermines leaf water use efficiency among differentcanopy locations in grapevine. Agric Water Manag 114:4–10
Meinzer FC, Grantz DA (1990) Stomatal and hydraulicconductance in growing sugarcane: stomatal adjustmentto water transport capacity. Plant Cell Environ 13:383–388
Nardini A, Salleo S (2000) Limitation of stomatalconductance by hydraulic traits: sensing or preventingxylem cavitation? Trees 15: 14–24
Nardini A, Salleo S, Andri S (2005) Circadian regulation ofleaf hydraulic conductance in sunflower (Helianthusannuus L. cv Margot). Plant Cell Environ 28: 750–759
Nardini A, Salleo S, Jansen S (2011) More than just avulnerable pipeline: xylem physiology in the light ofion-mediated regulation of plant water transport. J ExpBot 62: 4701–4718
Nardini A, Pedà G, Rocca NL (2012) Trade-offs betweenleaf hydraulic capacity and drought vulnerability:morpho-anatomical bases, carbon costs and ecologicalconsequences. New Phytol 196: 788–798
Pantin F, Monnet F, Jannaud D, Costa JM, Renaud J, MullerB, Genty B (2013) The dual effect of abscisic acid onstomata. New Phytol 197: 65–72
Pou A, Medrano H, Tomàs M, Martorell S, Ribas-Carbó M,Flexas J (2012) Anisohydric behaviour in grapevinesresults in better performance under moderate waterstress and recovery than isohydric behaviour. Plant Soil359: 335–349
Pou A, Medrano H, Flexas J, Tyerman SD (2013) A putativerole for TIP and PIP aquaporins in dynamics of leafhydraulic and stomatal conductances in grapevineunder water stress and re-watering. Plant Cell Environ36: 828–843
Rico C, Pittermann J, Polley HW, Aspinwall MJ, Fay PA(2013) The effect of subambient to elevated atmosphericCO2 concentration on vascular function in Helianthusannuus: implications for plant response to climatechange. New Phytol 199: 956–965
Ruiz-Sánchez MC, Domingo-Miguel R, Castel-Sánchez JR(2010) Deficit irrigation in fruit trees and vines in Spain.Span J Agric Res 8: S5–S20
Sack L, Holbrook NM (2006) Leaf hydraulics. Annu RevPlant Physiol Plant Mol Biol 57: 361–381
Sack L, Tyree MT (2005) Leaf hydraulics and itsimplications in plant structure and function. In:Holbrook NM, Zwieniecki MA (eds) Vascular Transportin Plants. Elsevier Academic Press, London
Sack L, Melcher PJ, Zwieniecki MA, Holbrook NM (2002)The hydraulic conductance of the angiosperm leaflamina: a comparison of three measurement methods. JExp Bot 53: 2177–2184
Sack L, Cowan PD, Jaikumar N, Holbrook NM (2003) The‘hydrology’ of leaves: co-ordination of structure andfunction in temperate woody species. Plant Cell Environ26: 1343–1356
Sack L, Streeter CM, Holbrook NM (2004) Hydraulicanalysis of water flow through leaves of sugar maple andred oak. Plant Physiol 134: 1824–1833
Sack L, Pasquet-Kok J, PrometheusWiki contributors (2011)Leaf pressure-volume curve parameters. Available athttp://prometheuswiki.publish.csiro.au/tiki-index.php?page=Leaf+pressure-volume+curve+parameters(accessed 9 October 2014)
Sakuratani T (1981) A heat balance method for measuringwater flux in the stem of intact plants. J Agric Meteor 37:9–17
Physiol. Plant. 2014

Salleo S, Nardini A, Lo Gullo MA, Ghirardelli LA (2002)Changes in stem and leaf hydraulics preceding leafshedding in Castanea sativa L. Biol Plant 45: 227–234
Sancho-Knapik D, Álvarez-Arenas TG, Peguero-Pina JJ,Fernández V, Gil-Pelegrín E (2011) Relationshipbetween ultrasonic properties and structural changes inthe mesophyll during leaf dehydration. J Exp Bot 62:3637–3645
Schultz HR (2003) Differences in hydraulic architectureaccount for near-isohydric and anisohydric behaviour oftwo field-grown Vitis vinifera L. cultivars during drought.Plant Cell Environ 26: 1393–1405
Scoffoni C, McKown AD, Rawls M, Sack L (2012)Dynamics of leaf hydraulic conductance with waterstatus: quantification and analysis of species differencesunder steady state. J Exp Bot 63: 643–658
Scoffoni C, Vuong C, Diep S, Cochard H, Sack L (2014)Leaf shrinkage with dehydration: coordination withhydraulic vulnerability and drought tolerance. PlantPhysiol 164: 1772–1788. DOI: 10.1104/pp.113.221424
Sellin A, Kupper P (2007) Temperature, light and leafhydraulic conductance of little-leaf linden (Tilia cordata)in a mixed forest canopy. Tree Physiol 27: 679–688
Sperry JS, Hacke UG, Oren R, Comstock JP (2002) Waterdeficit and hydraulic limits to leaf water supply. PlantCell Environ 25: 251–263
Sun Q, Rost TL, Matthews MA (2006) Pruning-inducedtylose development in stems of current-year shoots ofVitis vinifera (Vitaceae). Am J Bot 93: 1567–1576
Sun Q, Rost TL, Matthews MA (2008) Wound-inducedvascular occlusions in Vitis vinifera (Vitaceae): tyloses insummer and gels in winter. Am J Bot 95: 1498–1505
Tomás M, Medrano H, Pou A, Escalona JM, Martorell S,Ribas-Carbó M, Flexas J (2012) Water-use efficiency ingrapevine cultivars grown under controlled conditions:effects of water stress at the leaf and whole-plant level.Aust J Grape Wine Res 18: 164–172
Tyree MT, Hammel HT (1972) Measurement of turgorpressure and water relations of plants by pressure bombtechnique. J Exp Bot 23: 267–282
Voicu MC, Zwiazek JJ, Tyree MT (2008) Light response ofhydraulic conductance in bur oak (Quercusmacrocarpa) leaves. Tree Physiol 28: 1007–1015
Wheeler JK, Huggett BA, Tofte AN, Rockwell FE, HolbrookNM (2013) Cutting xylem under tension orsupersaturated with gas can generate PLC and theappearance of rapid recovery from embolism. Plant CellEnviron 36: 1938–1949
Wortemann R, Herbette S, Barigah TS, Fumanal B, Alia R,Ducousso A, Comory D, Roeckel-Drevet P, Cochard H(2011) Genotypic variability and phenotypic plasticity ofcavitation resistance in Fagus sylvatica L. across Europe.Tree Physiol 31: 1175–1182
Zufferey V, Cochard H, Ameglio T, Spring JL, Viret O(2011) Diurnal cycles of embolism formation and repairin petioles of grapevine (Vitis vinifera cv. Chasselas). JExp Bot 62: 3885–3894
Edited by M. Oliver
Physiol. Plant. 2014
Copyright © 2022 FDOKUMEN




















