
Phylogenetic relationships in tribe Lepidieae (Brassicaceae) based on chloroplast DNA restriction...
9
Phylogenetic relationships in tribe Lepidieae (Brassicaceae) based on chloroplast DNA restriction site variation Karsten Zunk, Klaus Mummenhoff, and Herbert Hurka Abstract: Tribal and subtribal boundaries within the Brassicaceae are often artificially drawn. Most of the currently recognized tribes and subtribes are separated by only a few morphological characters. Furthermore, these characters are variable even within genera, or they conflict with one another in their distribution patterns, resulting in controversial classification systems. Therefore, tribes and subtribes as traditionally delimited may not reflect natural groups. Up to now no cladistic analysis has been performed in tribe Lepidieae. In the current study, we have analyzed restriction site variation of chloroplast DNA among 41 species representing 19 genera to get insights into phylogenetic relationships within the Lepidieae. Phylogenetic lineages derived from our chloroplast data were critically compared with the traditional concepts. Our chloroplast DNA phylogeny most easily supports the classification of genera in the system of Hayek with some modifications. Key words: chloroplast DNA, restriction site variation, Tribe Lepidieae (Brassicaceae), molecular phylogeny. Résumé : Les frontières entre les tribus et sous-tribus des Brassicaceae sont souvent arbitrairement délimitées. La plupart des tribus et sous-tribus actuellement reconnues ne sont séparées que par quelques caractères morphologiques. De plus, ces caractères sont variables, même à l’intérieur du genre, ou sont en conflit les uns avec les autres dans leurs patrons de distribution, ce qui conduit à des systèmes de classification controversés. Par conséquent, la délimitation traditionnelle des tribus et des sous-tribus peut fort bien ne pas refléter des groupes naturels. Jusqu’à maintenant aucune analyse cladistique n’a été réalisée pour la tribu des Lepidieae. Dans cette étude, les auteurs ont analysé la variation des sites de restriction de l’ADN chloroplastique chez 41 espèces représentant 19 genres, afin de mieux percevoir les relations phylogénétiques chez les Lepidieae. Les auteurs ont comparé de façon critique les lignées phylogénétiques déduites des données sur l’ADN chloroplastique avec les concepts traditionnels. La phylogénie basée sur l’ADN chloroplastique supporte plus facilement, avec quelques modifications, la classification des genres selon le système de Hayek. Mots clés : ADN chloroplastique, variation des sites de restriction, tribu des Lepidieae (Brassicaceae), phylogénie molé- culaire. [Traduit par la Rédaction] Zunk et al. 1512 Introduction Most of the currently recognized tribes of the Brassica- ceae (Cruciferae) are separated by only one or two morpho- logical characters, which are even variable within genera, or they conflict with one another in their distribution patterns among genera and tribes (Al-Shehbaz 1984; Hedge 1976; Price et al. 1994) suggesting high levels of homoplasy for these characters. Apart from the tribes Brassiceae and Thely- podieae, only tribe Lepidieae has much claim to be regarded as natural. The Lepidieae are characterized by angustiseptate silicles (fruits strongly compressed at right angles to the sep- tum) which are generally less than three times longer than they are broad (Al-Shehbaz 1986). With single exceptions, this is contrary to the rest of the family in which the fruits are terete, four-angled, or compressed parallel to the septum, i.e., latiseptate (Hayek 1911). In some genera, such as Co- chlearia and allies, the fruits are inflated or only weakly angustiseptate, and it is difficult to be sure whether the fruits are angustiseptate or latiseptate. Apart from single taxa, fruits are dehiscent and nectar glands are nearly almost small and distinct. For the reasons given above, tribal boundaries are sometimes artificially drawn, and there is controversy among various sub- tribal classifications. Some examples may demonstrate this. The tribe Lepidieae, as described above, was established by Hayek (1911). Later on Schulz (1936) overemphasized the flattening of the fruits in delineating the boundaries of Lepidieae. Schulz maintained Cardaria Desv. in the Lepid- ieae and placed Hymenophysa C.A. Meyer (globose fruits) in tribe Euclidieae. Recent authors included Cardaria and Hymenophysa in the same genus (Al-Shehbaz 1986). The genera Capsella Med., Camelina Crantz, and Neslia Desv. were combined by Hayek (1911) and Janchen (1942) in Lepidieae, subtribe Capsellinae, based on characteristics of the nectar glands, myrosine cells, cotyledons, and the indumentum. Camelina and Neslia, however, are not charac- Can. J. Bot. 77: 1504–1512 (1999) © 1999 NRC Canada 1504 Received February 24, 1999. K. Zunk, K. Mummenhoff, 1 and H. Hurka. Fachbereich Biologie/Chemie, Spezielle Botanik, Universität Osnabrück, Barbarastrasse 11, 49069 Osnabrück, Germany. 1 Author to whom all correspondence should be addressed. e-mail: [email protected]
-
Upload
uni-osnabrueck -
Category
Documents
-
view
6 -
download
0
Transcript of Phylogenetic relationships in tribe Lepidieae (Brassicaceae) based on chloroplast DNA restriction...

Phylogenetic relationships in tribe Lepidieae(Brassicaceae) based on chloroplast DNArestriction site variation
Karsten Zunk, Klaus Mummenhoff, and Herbert Hurka
Abstract: Tribal and subtribal boundaries within the Brassicaceae are often artificially drawn. Most of the currentlyrecognized tribes and subtribes are separated by only a few morphological characters. Furthermore, these characters arevariable even within genera, or they conflict with one another in their distribution patterns, resulting in controversialclassification systems. Therefore, tribes and subtribes as traditionally delimited may not reflect natural groups. Up tonow no cladistic analysis has been performed in tribe Lepidieae. In the current study, we have analyzed restriction sitevariation of chloroplast DNA among 41 species representing 19 genera to get insights into phylogenetic relationshipswithin the Lepidieae. Phylogenetic lineages derived from our chloroplast data were critically compared with thetraditional concepts. Our chloroplast DNA phylogeny most easily supports the classification of genera in the system ofHayek with some modifications.
Key words: chloroplast DNA, restriction site variation, Tribe Lepidieae (Brassicaceae), molecular phylogeny.
Résumé : Les frontières entre les tribus et sous-tribus des Brassicaceae sont souvent arbitrairement délimitées. Laplupart des tribus et sous-tribus actuellement reconnues ne sont séparées que par quelques caractères morphologiques.De plus, ces caractères sont variables, même à l’intérieur du genre, ou sont en conflit les uns avec les autres dans leurspatrons de distribution, ce qui conduit à des systèmes de classification controversés. Par conséquent, la délimitationtraditionnelle des tribus et des sous-tribus peut fort bien ne pas refléter des groupes naturels. Jusqu’à maintenantaucune analyse cladistique n’a été réalisée pour la tribu des Lepidieae. Dans cette étude, les auteurs ont analysé lavariation des sites de restriction de l’ADN chloroplastique chez 41 espèces représentant 19 genres, afin de mieuxpercevoir les relations phylogénétiques chez les Lepidieae. Les auteurs ont comparé de façon critique les lignéesphylogénétiques déduites des données sur l’ADN chloroplastique avec les concepts traditionnels. La phylogénie baséesur l’ADN chloroplastique supporte plus facilement, avec quelques modifications, la classification des genres selon lesystème de Hayek.
Mots clés : ADN chloroplastique, variation des sites de restriction, tribu des Lepidieae (Brassicaceae), phylogénie molé-culaire.
[Traduit par la Rédaction] Zunk et al. 1512
Introduction
Most of the currently recognized tribes of the Brassica-ceae (Cruciferae) are separated by only one or two morpho-logical characters, which are even variable within genera, orthey conflict with one another in their distribution patternsamong genera and tribes (Al-Shehbaz 1984; Hedge 1976;Price et al. 1994) suggesting high levels of homoplasy forthese characters. Apart from the tribes Brassiceae and Thely-podieae, only tribe Lepidieae has much claim to be regardedas natural. The Lepidieae are characterized by angustiseptatesilicles (fruits strongly compressed at right angles to the sep-tum) which are generally less than three times longer thanthey are broad (Al-Shehbaz 1986). With single exceptions,this is contrary to the rest of the family in which the fruits
are terete, four-angled, or compressed parallel to the septum,i.e., latiseptate (Hayek 1911). In some genera, such as Co-chlearia and allies, the fruits are inflated or only weaklyangustiseptate, and it is difficult to be sure whether the fruitsare angustiseptate or latiseptate. Apart from single taxa,fruits are dehiscent and nectar glands are nearly almostsmall and distinct.
For the reasons given above, tribal boundaries are sometimesartificially drawn, and there is controversy among various sub-tribal classifications. Some examples may demonstrate this.
The tribe Lepidieae, as described above, was establishedby Hayek (1911). Later on Schulz (1936) overemphasizedthe flattening of the fruits in delineating the boundaries ofLepidieae. Schulz maintained Cardaria Desv. in the Lepid-ieae and placed Hymenophysa C.A. Meyer (globose fruits)in tribe Euclidieae. Recent authors included Cardaria andHymenophysa in the same genus (Al-Shehbaz 1986).
The genera Capsella Med., Camelina Crantz, and NesliaDesv. were combined by Hayek (1911) and Janchen (1942)in Lepidieae, subtribe Capsellinae, based on characteristicsof the nectar glands, myrosine cells, cotyledons, and theindumentum. Camelina and Neslia, however, are not charac-
Can. J. Bot. 77: 1504–1512 (1999) © 1999 NRC Canada
1504
Received February 24, 1999.
K. Zunk, K. Mummenhoff,1 and H. Hurka. FachbereichBiologie/Chemie, Spezielle Botanik, Universität Osnabrück,Barbarastrasse 11, 49069 Osnabrück, Germany.
1Author to whom all correspondence should be addressed.e-mail: [email protected]
J:\cjb\cjb77\cjb-10\B99-129.vpMonday, January 17, 2000 3:16:57 PM
Color profile: DisabledComposite Default screen

terized by the principal character of the Lepidieae, i.e.,angustiseptate fruits. Therefore, Schulz (1936) placedCamelina in the tribe Sisymbrieae and Neslia, which hasindehiscent and nutlike fruits, in the tribe Euclidieae, an arti-ficial assembly of species sensu Hedge (1976).
As outlined above, there seems to be little agreementamong the various morphologically based classifications asto the limits of the Lepidieae and the number and boundariesof its subtribes. A more natural classification of the Lepidieaewould perhaps be achieved by using an alternative approachto the morphologically based taxonomies and by groupingclosely related genera and working upward.
In the Brassicaceae, molecular studies have already con-tributed to a clarification of phylogenetic relationships in thetribe Brassiceae (Warwick and Black 1997; Francisco-Ortegaet al. 1999), Arabidopsis and related genera (Price et al.1994; Galloway et al. 1998), Lepidium (Mummenhoff et al.1995), Thlaspi and related genera (Mummenhoff and Koch1994; Mummenhoff et al. 1997a, 1997b; Zunk et al. 1996),Cochlearia (Koch et al. 1996), and Cardamine and allies(Franzke et al. 1998). In the current study we used chloro-plast (cp) DNA restriction site analysis to examine phylo-genetic relationships within the Lepidieae and compared ourresults with the traditional concepts of Hayek (1911), Schulz(1936), and Janchen (1942).
We have analysed 18 genera (37 species), of which 15 havebeen placed equivocally into tribe Lepidieae by all relevantauthors (Hayek 1911; Schulz 1936; Janchen 1942; Al-Shehbaz 1986; Table 2). These genera represent four of thefive subtribes of the comprehensive treatment of Hayek(1911), which is generally accepted as being largely natural(Markgraf 1963; Shaw 1965). This system is used here as thestarting point for phylogenetic studies. The monogeneric sub-tribe Subulariinae (two species) was not considered here forlack of material. In addition, we have included three genera,i.e., Camelina, Neslia, and Peltaria Jacq., distributed by rele-vant authors among different tribes of the family (Table 2).
Methods
Plant materialDNA of 70 accessions from 40 Brassicaceae species represent-
ing the main lineages of tribe Lepidieae sensu Hayek (1911) wasexamined (Table 1). Sources of plant material are listed in Table 1.All voucher specimens are deposited in the herbarium of the Uni-versity of Osnabrück (OSBU). A species from the closely relatedfamily Capparaceae (Capparis spinosa L.) served as the outgroup.Because taxa were included in the present study that have beenclassified by some authors within other subtribes of Lepidieae andeven other tribes (Table 2), we decided to use a more remoteoutgroup than perhaps choosing taxa from other tribes of the Bras-sicaceae. The morphological and rbcL analysis of Rodman (1991)and Rodman et al. (1993), respectively, all indicate that the Bras-sicaceae is well nested within the core group of capparalean fami-lies, and that the Brassicaceae is most closely related to thecleomoid subfamily of the Capparaceae, from which it may havebeen derived (Price et al. 1994; Rodman et al. 1993). This providesjustification for the use of a member of the Capparaceae as out-group in phylogenetic studies of Lepidieae.
Molecular methodsYoung, fresh leaves were harvested from plants grown in the
greenhouse at Osnabrück University. In some cases DNA was ex-
tracted from field-collected leaves, dried, and preserved with silicagel (Chase and Hills 1991).
Total DNAs were obtained from 1–2 g of fresh leaf tissue fromsingle plants by the CTAB method, as described by Doyle andDoyle (1987) with additional organic extractions, once with satu-rated phenol, once with phenol–chloroform (1:1), and once withchloroform. DNA samples were digested with 10 restriction en-zymes (BRL) recognizing six base-pair sequences: BglI, EcoRV,HindIII, KpnI, PstI, PvuII, ScaI, SstI, StuI, and XhoI. Restrictionfragments were separated electrophoretically on 0.4–0.6% agarosegels and transferred to reusable nylon filters (Pharmacia-LKB). Fil-ters were subsequently probed with 19 clones and 5 subclones rep-resenting nearly the entire chloroplast genome of Brassica juncea.Clones and subclones were kindly provided by J. Palmer (IndianaUniversity), and S. Warwick and L. Black (Biosystematic ResearchCenter, Ottawa), respectively. Clone number and size are shownin Warwick and Black (1991). Preparation of digoxigenin-labeledprobe DNAs, filter hybridizations, filter decolorization, and proberemoval followed the manufacturer’s instructions (BoehringerMannheim), with the following modifications: hybridizations wereperformed at 68°C with the substitution of 2% powdered milk andherring sperm DNA (200 µg/mL) as a blocking agent. Stringencywashes were performed twice for 2 min each at room temperaturewith 0.3 M NaCl, 30 mM sodium citrate, 0,1% SDS, and twice for15 min each at 50°C with 15 mM NaCl, 1.5 M sodium citrate,0.1% SDS. Immunological detection of hybridized DNA followedthe manufacturer’s instructions (Boehringer Mannheim, DIG DNAlabeling system, detection via NBT/BCIP), with the substitution of1% powdered milk as a blocking agent.
Data analysisRestriction site maps were constructed for all taxa under study.
Restriction sites of the different taxa were then aligned and pres-ence or absence of restriction sites were scored as a 1/0 datamatrix. The restriction site maps and 1/0 data matrix are availablefrom the authors upon request. Because many restriction sites andfragment sizes among the taxa examined coincided with those fromother taxa of Brassicaceae (Mummenhoff and Koch 1994; Warwickand Black 1991; Zunk et al. 1996), mapping efforts were greatlyfacilitated by scoring our data against these maps.
Parsimony analysis was conducted using PAUP version 3.1.1(Swofford 1993). Heuristic searches for most parsimonious treeswere conducted in PAUP by using simple addition sequence oftaxa followed by TBR branch swapping, and using 100 random ad-dition sequences, followed by the TBR branch swapping algorithm.
Mutations identified were polarized by the outgroup method(Watrous and Wheeler 1981) and the choice of the outgroup is out-lined above.
To test for nonrandom structure in the data, the frequency dis-tribution of the lengths of 10.000 trees randomly selected byPAUP (version 3.1.1; random trees option) from the set of all pos-sible trees was evaluated for left-handed skewness (Hillis andHuelsenbeck 1992). The bootstrap method in PAUP (version 3.1.1)was used to determine statistical support for the groups inferred tobe monophyletic. One hundred bootstrap replications were run inPAUP with heuristic search options. The data were also analyzedby the stepmatrix method of character state weighting of gains overlosses by a factor of 1.3:1 (Albert et al. 1992). This analysis wasperformed with PAUP (version 3.1.1, CONSTRAINTS command)to make the ingroup monophyletic following the steps described byAlbert et al. (1992).
Results
Comparative cpDNA restriction site mapping allowed usto recognize 290 restriction sites in the 38 taxa surveyed.We detected 220 variable sites of which 159 (68.6%) were
© 1999 NRC Canada
Zunk et al. 1505
J:\cjb\cjb77\cjb-10\B99-129.vpMonday, January 17, 2000 3:16:58 PM
Color profile: DisabledComposite Default screen

© 1999 NRC Canada
1506 Can. J. Bot. Vol. 77, 1999
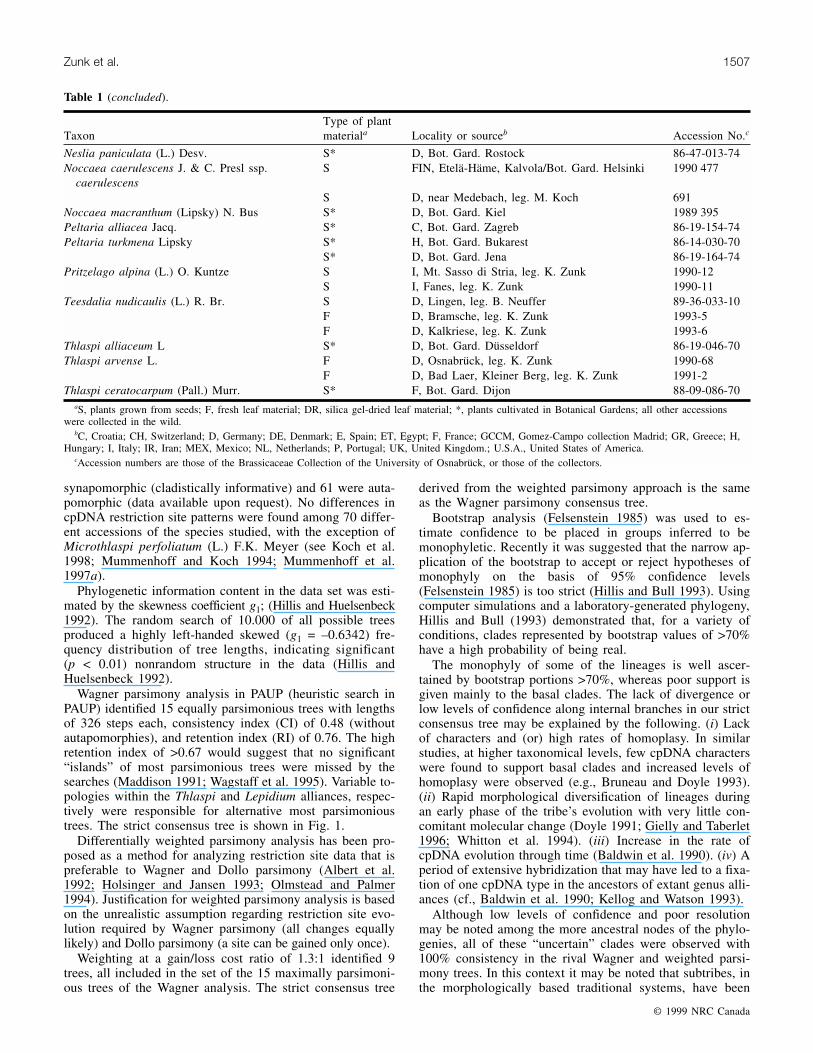
TaxonType of plantmateriala Locality or sourceb Accession No.c
Aethionema grandiflorum Boiss. & Hohenacker S* D, Bot. Gard. Wuppertal DE 92-07-064-70S* Bot. Gard. Kopenhagen 91-31-030-70
Aethionema saxatile (L.) R. Br. S I, Sicily, Quacella/Madonie, leg. K. Bernhardt 86-30-096-14Biscutella cichoriifolia Loisel. S* CH, Bot. Gard. Basel 88-08-062-74Biscutella laevigata L. S D, Mansfelder Hügelland, leg. B. Neuffer 89-46-009-10
S I, near St. Kassian, leg. K. Zunk 1990-7Camelina microcarpa Andrz. ex DC. S U.S.A., Oklahoma, Little Sahara, leg. Neuffer 90-27-008-10
S F, Fontenoy sur Moselle, Conservatoir et JardinBotanique de Nancy
90-17-203-10
Camelina pilosa (DC.) Vassilicz S* D, Bot. Gard. Rostock 86-46-012-74Camelina rumelica Velen. S IR, Quazvin, GCCM 91-50-010-10Capparis spinosa L. F* D, Bot. Gard. Düsseldorf 86-48-062- 80Capsella bursa-pastoris (L.) Med. S U.S.A., Nevada, Reno, leg. H. Hurka 740/4
S ET, Oasis Faijum, leg. H. Hurka 764/2Capsella grandiflora (Fauché & Chaub.) Boiss. S GR, Korfu, leg. H. Hurka 916
S GR, Igouminitza, Votonosi, leg. H. Hurka 926/21S GR, Korfu, leg. H. Hurka 937/28
Capsella rubella Reuter S I, Gargano, Forestra Umbra, leg. S. Bartelheim 774S E, San Millan, leg. H. Hurka 776S GR, Aradena, leg. U. Rösemann 976
Cardaria draba (L.) Desv. F D, Bad Laer, Kleiner Berg, leg. K. Zunk 1991-3F D, Borkum, leg. M. Koch 1992-11F D, Bramsche, leg. K. Zunk 1993-4
Cochlearia danica L. S F, Roscoff, Biol. Gard. 88-20-011-10F D, Melle, leg. M. Koch 92-C-002
Cochlearia glastifolia L. S* NL, Bot. Gard. Amsterdam 88-06-147-70S* NL, Bot. Gard. Amsterdam 86-11-280-70
Coronopus didymus (L.) Sm. S* Bot. Gard. Basel 86-19-148-74S P, Estremadura, Loures, Bot. Gard. Lissabon 93-11-034-10
Coronopus squamatus (Forsk.) Asch. S* D, Bot. Gard. Bonn 86-18-057-74S D, Kleve-Salmorth, leg. P. Gerstberger 89-46-010-10F D, Ziegenberg near Höxter, leg. K. Zunk 1992-7
Hymenolobus procumbens (L.) Nutt. ex Torr.& Gray
S E, Mallorca, Bot. Gard. Kopenhagen 92-25-023-10
S* DE, Bot. Gard. Kopenhagen 92-25-022-70Iberis amara L. S* UK, Bot. Gard. Cambridge 87-04-056-74
S* D, Fritzlar, Bot. Gard. Göttingen 90-08-093-10Iberis pinnata L. S* F, Bot. Gard. Bordeaux 89-27-063-70Ionopsidium abulense (Pau) Rothm. S E, Avila, Piedrahita, GCCM 91-50-045-10Ionopsidium acaule (Desf.) Rchb. S* D, Bot. Gard. Braunschweig 86-11-260-70
S* D, Bot. Gard. Frankfurt 86-11-224-74S GCCM 91-50-046-70
Ionopsidium prolongoi (Boiss.) Batt. S E, Cadiz, Frazalema, GCCM 91-50-047-10Lepidium campestre (L.) R. Br. S F, Doubs, Bois de Fourg, Bot. Gard. Besaçnon 822-47
F D, near Minden, leg. M. Koch 1992-13Lepidium sativum L. S D, Bot. Gard. Universität Bochum 342Lepidium virginicum L S MEX, Carrizal Chio, leg.
R. Stockmann / K. Bosbach88-23-025-10
Microthlaspi perfoliatum (L.) S CH, St. Gallen, Bot. Gard. St. Gallen 87-04-059-10F.K. Meyer S D, Bad Laer, leg. K. Zunk 1991-1
S D, near Oberelsungen, leg. K. Zunk 1992-2S CH, Valais, Bot. Gard. Champex 86-19-126-10S D, near Würzburg, leg. K. Zunk 1991-11S I, Monti Sibilli, Mt. Zampa, Bot. Gard. Pesaro 87-04-028-10
Neslia apiculata Fischer, Meyer & Avé-Lall. S I, Sizilien, Selinunte, leg. K. Bernhardt 86-19-233-14Neslia paniculata (L.) Desv. S D, Engen, Bot. Gard. Universität Konstanz 90-14-126-10
DR D, Mt. Staffelstein, leg. K. Zunk 1992-2
Table 1. Collection data and type of plant material examined for chloroplast restriction site variation.
J:\cjb\cjb77\cjb-10\B99-129.vpMonday, January 17, 2000 3:16:59 PM
Color profile: DisabledComposite Default screen
synapomorphic (cladistically informative) and 61 were auta-pomorphic (data available upon request). No differences incpDNA restriction site patterns were found among 70 differ-ent accessions of the species studied, with the exception ofMicrothlaspi perfoliatum (L.) F.K. Meyer (see Koch et al.1998; Mummenhoff and Koch 1994; Mummenhoff et al.1997a).
Phylogenetic information content in the data set was esti-mated by the skewness coefficient g1; (Hillis and Huelsenbeck1992). The random search of 10.000 of all possible treesproduced a highly left-handed skewed (g1 = –0.6342) fre-quency distribution of tree lengths, indicating significant(p < 0.01) nonrandom structure in the data (Hillis andHuelsenbeck 1992).
Wagner parsimony analysis in PAUP (heuristic search inPAUP) identified 15 equally parsimonious trees with lengthsof 326 steps each, consistency index (CI) of 0.48 (withoutautapomorphies), and retention index (RI) of 0.76. The highretention index of >0.67 would suggest that no significant“islands” of most parsimonious trees were missed by thesearches (Maddison 1991; Wagstaff et al. 1995). Variable to-pologies within the Thlaspi and Lepidium alliances, respec-tively were responsible for alternative most parsimonioustrees. The strict consensus tree is shown in Fig. 1.
Differentially weighted parsimony analysis has been pro-posed as a method for analyzing restriction site data that ispreferable to Wagner and Dollo parsimony (Albert et al.1992; Holsinger and Jansen 1993; Olmstead and Palmer1994). Justification for weighted parsimony analysis is basedon the unrealistic assumption regarding restriction site evo-lution required by Wagner parsimony (all changes equallylikely) and Dollo parsimony (a site can be gained only once).
Weighting at a gain/loss cost ratio of 1.3:1 identified 9trees, all included in the set of the 15 maximally parsimoni-ous trees of the Wagner analysis. The strict consensus tree
derived from the weighted parsimony approach is the sameas the Wagner parsimony consensus tree.
Bootstrap analysis (Felsenstein 1985) was used to es-timate confidence to be placed in groups inferred to bemonophyletic. Recently it was suggested that the narrow ap-plication of the bootstrap to accept or reject hypotheses ofmonophyly on the basis of 95% confidence levels(Felsenstein 1985) is too strict (Hillis and Bull 1993). Usingcomputer simulations and a laboratory-generated phylogeny,Hillis and Bull (1993) demonstrated that, for a variety ofconditions, clades represented by bootstrap values of >70%have a high probability of being real.
The monophyly of some of the lineages is well ascer-tained by bootstrap portions >70%, whereas poor support isgiven mainly to the basal clades. The lack of divergence orlow levels of confidence along internal branches in our strictconsensus tree may be explained by the following. (i) Lackof characters and (or) high rates of homoplasy. In similarstudies, at higher taxonomical levels, few cpDNA characterswere found to support basal clades and increased levels ofhomoplasy were observed (e.g., Bruneau and Doyle 1993).(ii) Rapid morphological diversification of lineages duringan early phase of the tribe’s evolution with very little con-comitant molecular change (Doyle 1991; Gielly and Taberlet1996; Whitton et al. 1994). (iii) Increase in the rate ofcpDNA evolution through time (Baldwin et al. 1990). (iv) Aperiod of extensive hybridization that may have led to a fixa-tion of one cpDNA type in the ancestors of extant genus alli-ances (cf., Baldwin et al. 1990; Kellog and Watson 1993).
Although low levels of confidence and poor resolutionmay be noted among the more ancestral nodes of the phylo-genies, all of these “uncertain” clades were observed with100% consistency in the rival Wagner and weighted parsi-mony trees. In this context it may be noted that subtribes, inthe morphologically based traditional systems, have been
© 1999 NRC Canada
Zunk et al. 1507
TaxonType of plantmateriala Locality or sourceb Accession No.c
Neslia paniculata (L.) Desv. S* D, Bot. Gard. Rostock 86-47-013-74Noccaea caerulescens J. & C. Presl ssp.
caerulescensS FIN, Etelä-Häme, Kalvola/Bot. Gard. Helsinki 1990 477
S D, near Medebach, leg. M. Koch 691Noccaea macranthum (Lipsky) N. Bus S* D, Bot. Gard. Kiel 1989 395Peltaria alliacea Jacq. S* C, Bot. Gard. Zagreb 86-19-154-74Peltaria turkmena Lipsky S* H, Bot. Gard. Bukarest 86-14-030-70
S* D, Bot. Gard. Jena 86-19-164-74Pritzelago alpina (L.) O. Kuntze S I, Mt. Sasso di Stria, leg. K. Zunk 1990-12
S I, Fanes, leg. K. Zunk 1990-11Teesdalia nudicaulis (L.) R. Br. S D, Lingen, leg. B. Neuffer 89-36-033-10
F D, Bramsche, leg. K. Zunk 1993-5F D, Kalkriese, leg. K. Zunk 1993-6
Thlaspi alliaceum L S* D, Bot. Gard. Düsseldorf 86-19-046-70Thlaspi arvense L. F D, Osnabrück, leg. K. Zunk 1990-68
F D, Bad Laer, Kleiner Berg, leg. K. Zunk 1991-2Thlaspi ceratocarpum (Pall.) Murr. S* F, Bot. Gard. Dijon 88-09-086-70
aS, plants grown from seeds; F, fresh leaf material; DR, silica gel-dried leaf material; *, plants cultivated in Botanical Gardens; all other accessionswere collected in the wild.
bC, Croatia; CH, Switzerland; D, Germany; DE, Denmark; E, Spain; ET, Egypt; F, France; GCCM, Gomez-Campo collection Madrid; GR, Greece; H,Hungary; I, Italy; IR, Iran; MEX, Mexico; NL, Netherlands; P, Portugal; UK, United Kingdom.; U.S.A., United States of America.
cAccession numbers are those of the Brassicaceae Collection of the University of Osnabrück, or those of the collectors.
Table 1 (concluded).
J:\cjb\cjb77\cjb-10\B99-129.vpMonday, January 17, 2000 3:16:59 PM
Color profile: DisabledComposite Default screen

characterized by only few consistent character, at best (Al-Shehbaz 1984; Hedge 1976).
The topology of the Wagner parsimony strict consensustree retained considerable resolution even under a differentcharacter state weighting scheme (Albert et al. 1992).Grouping of plastomes found here fits best to the taxonomictreatment of Hayek (1911).
To avoid confusion with traditional subtribal terms (seeTable 2), cpDNA-groups are designated in our tree as “genusalliances.” Our genus alliances correlate in part with tradi-tionally described subtribes: namely the Capsella alliance(subtribe Capsellinae), the Lepidium alliance (subtribeLepidiinae), and the Thlaspi alliance (subtribe Thlaspidinae).Other cpDNA groups differ from previous treatments (Co-chlearia alliance, Iberis alliance). The genera Aethionema
and Biscutella cannot be associated with any of the coregroups of tribe Lepidieae.
Discussion
In the absence of a comprehensive cladistic analysis of al-ternative characters, our phylogenetic tree can only be dis-cussed in the light of the morphologically based treatmentsof previous authors (i.e., Hayek 1911; Janchen 1942; Schulz1936).
Lepidium alliance (Lepidium, Cardaria, Coronopus)These genera share a number of characters: (i) embryo
notorrhizal and cotyledons folded above the insertion,(ii) fruits with one seed per locule, (iii) a special pattern of
© 1999 NRC Canada
1508 Can. J. Bot. Vol. 77, 1999
Genusa,b Current analysis Hayek (1911) Schulz (1936) Janchen (1942)
Genus alliance Tribe / subtribe Tribe / subtribe Tribe / subtribe
Lepidium
Cardaria Lepidium Lepidieae / Lepidiinae Lepidieae / Lepidiinae Lepidieae / Lepidiinae
Coronopus
Iberis
Lepidieae / Iberidinae Lepidieae / Iberidinae
Hymenolobus Iberis Lepidieae / Iberidinaec
Pritzelago Lepidieae / Capsellinae
Capsella
Lepidieae / Capsellinae
Camelina Capsella Lepidieae / Capsellinae Sisymbrieae / Camelininae
Neslia Euclidieae
Cochlearia
Lepidieae / Cochleariinae Lepidieae / CochleariinaeCochlearia
Ionopsidium
Noccaea
MicrothlaspiLepidieae / Thlaspidine
Lepidieae / Thlaspidinae Lepidieae / Thlaspidinae
Teesdalia Thlaspi
Thlaspi
Peltaria Lunarieae Alysseae / Lunariinae
Biscutella
Lepidieae / Iberidinae Lepidieae / IberidinaeLepidieae / Lepidiinae
Aethionema Lepidieae / Thlaspidinae Lepidieae / ThlaspidinaeaGenus alliances of the cpDNA phylogeny.bNomenclature of Thlaspi, Noccaea, and Microthlaspi follows Meyer (1973, 1979).cHymenolobus has no generic rank (see text).
Table. 2. Diagrammic presentation of alternative classification systems for the Lepidieae relative to the genus alliances identified in ourcpDNA phylogeny (see Fig. 1).
J:\cjb\cjb77\cjb-10\B99-129.vpMonday, January 17, 2000 3:17:00 PM
Color profile: DisabledComposite Default screen

© 1999 NRC Canada
Zunk et al. 1509
Fig. 1. Strict consensus of 15 equally most parsimonious Wagner trees (tree length = 326, CI = 0.48). Bootstrap values >50% aregiven above the branches, and genus alliances are indicated in brackets. The tree shows relationships among taxa of tribe Lepidieaebased upon 154 chloroplast DNA restriction site mutations. This tree is identical to the strict consensus of 9 most parsimonious treesderived from a weighted parsimony analysis with a gain/loss cost ratio of 1.3:1.
J:\cjb\cjb77\cjb-10\B99-129.vpMonday, January 17, 2000 3:17:03 PM
Color profile: DisabledComposite Default screen

aromatic glucosinolates (Al-Shehbaz and Al-Shammary1987; Daxenbichler et al. 1964, 1991; Kjær 1960), and (iv) aseed oil composition that is characterized by dominance ofC18 fatty acids and similar amounts of mono- and triple-unsaturated fatty acids (Mikolajczak et al. 1961; Miller et al.1965). Based on the morphological characters, the generamentioned above are traditionally classified in subtribeLepidiinae (Table 2). Our cpDNA restriction site datastrongly confirm this grouping with a bootstrap value of100%. Although we have studied only three species of thegenus Lepidium and closely related genera, our data wouldindicate that Lepidium is not monophyletic (Fig. 1). A com-prehensive sequence analysis of 100 species of Lepidium,Cardaria, and Coronopus (ITS regions of nrDNA, noncodingcpDNA (K. Mummenhoff and H. Brüggemann, unpublisheddata) clearly suggest that Lepidium and Coronopus are para-phyletic and polyphyletic, respectively, as currently delim-ited. Furthermore, Cardaria is closely related to Lepidiumcampestre and related species (section Lepia), and should beincluded in Lepidium. (K. Mummenhoff and H. Brüggemann,unpublished data).
Capsella alliance (Capsella, Camelina, Neslia)Our cpDNA data clearly demonstrate that these genera
form a monophyletic lineage with a bootstrap proportion of96%. This corresponds with the concepts of most authors.The classification systems of de Candolle (1821), Hayek(1911), Jaretzky (1928), and Hedge (1976), based on differ-ences in life form, the structure of nectar glands (mediannectaries absent and lateral nectaries half-moon like with asmall process towards the median stamens), are in agreementwith our data. Schulz (1936), however, distributed the threegenera into three different tribes (see Table 2) based onflower and fruit characters (Capsella: dehiscent angustisep-tate fruits; Camelina: dehiscent and slightly latiseptate fruit;Neslia: fruit nutlike and indehiscent). Even Al-Shehbaz(1986, 1987) stated that the genera have only superficialsimilarity and he transferred Camelina to tribe Alysseaewhile leaving Capsella and Neslia in tribe Lepidieae.
Indeed the three genera are very similar in a number ofmorphological characters, and our phylogenetic analysissuggests that this similarity is due to common ancestry.
The fruits of Neslia and Camelina, although very differentwhen ripened, are nearly identical in early stages of de-velopment. Like those of Camelina, the ovaries of Nesliaare characterized by two locules with two (sometimes four)ovules per locule. In later phases of development, only oneovule is becoming a seed (Bayer 1905, p. 167).
In addition, the fatty acid spectrum (monosaturated fattyacids larger than C18 as main seed oil component;Mikolajczak et al. 1961; Miller et al. 1965; Mukherjee et al.1984) and glucosinolate composition of these three generaso far as investigated (long-chained ω-methyl sulphinyl glu-cosinolates; Daxenbichler et al. 1991; Kjær 1960) is verysimilar and different from other Brassicaceae. Furthermore,the structure of the seed coat (Vaughan and Whitehouse1971) would support our cpDNA grouping. The seed coat ofCapsella, Camelina, and Neslia is identical, except for thestructure of the epidermis, which is mucilaginous in Cap-sella and Camelina, but not in Neslia.
Nonmucilaginous seeds are typical of Brassicaceae taxawith indehiscent fruits (Al-Shehbaz 1986). Summing up, theunification of Camelina, Capsella, and Neslia into the Cap-sella alliance (= subtribe Capsellinae) seems to be justifiedbased on cpDNA data, morphological characters (habit, nec-tar, glands) and biochemical data.
Iberis alliance (Iberis, Pritzelago, Hymenolobus)The treatment of Iberis, Pritzelago, and Hymenolobus, as
one lineage, is preliminary and needs special discussion, be-cause (i) taxa of this alliance are traditionally placed in dif-ferent subtribes along with various other genera (compareTable 2), and (ii) this alliance is not strongly supported bybootstrap analysis. Consequently the question arises if thereis a justification for this grouping by looking at additionalcharacters.
A compilation of traditionally used characters shows thatmembers of this alliance share very small lateral nectar glandsand fruit locules containing one or few seeds. The system-atic position of Hymenolobus is not easy to clarify, becausethe genus was previously included in Capsella, Hornungia,and Prizelago. Hayek (1911) includes Hymenolobus pro-cumbens in Hornungia (Hornungia procumbens (L.) Hayek)but this genus does not appear in his phylogenetic treatmentof the whole family (Hayek 1911). Bayer (1905) and Hayek(1911) both argue for close relationships of Pritzelago andIberis, because lateral nectar glands are small and the myro-sin cells are associated with the phloem of vascular bundles,respectively. With regard to the commonly used morphologi-cal characters described above, the three genera are placedbest near the Lepidium alliance, a view favoured by Hayek(1911). However, in addition to lateral nectar glands, smallmedian glands are present in Lepidium and Coronopus, thecore group of the Lepidium alliance. Furthermore, the mostcharacteristic feature of the Lepidium alliance, i.e., cotyle-dons folded at right angles to the leaf axis, is not found intaxa of the Iberis alliance.
In contrast with this interpretation, Schulz (1936) andJanchen (1942) maintained Iberis in sutribe Iberidinae buttransferred Pritzelago and Hymenolobus to Capsella and al-lies (subtribe Capsellinae) based on presence of branchedtrichomes in the latter group (but note the different composi-tion of subtribe Capsellinae sensu Schulz and Janchen (Ta-ble 2).
But as outlined above, concurrence in the structure andform of nectar glands and in the number of seeds per fruitlocule in the Capsella alliance, in Pritzelago and Hymeno-lobus, respectively, is apparently not in agreement withSchulz’s and Janchen’s treatment.
Although not strongly supported by high bootstrap pro-portions in our cpDNA tree, there is the general impressionthat genera of the Iberis alliance are somewhat intermediatebetween the Capsella and Lepidium alliance.
The systematic position of Biscutella is a matter of con-troversy. It was classified close to Lepidium by Hayek, how-ever next to Iberis by Schulz and Janchen (Table 2). Theclassification of Biscutella is difficult because the fruit type(a strongly compressed, didymous silicula, indehiscent butwith the one-seeded locules eventually breaking away fromthe axis, Ball et al. 1993) is uncommon within the Bras-sicaceae, but similar fruits occur in the Asian Megacarpaea
© 1999 NRC Canada
1510 Can. J. Bot. Vol. 77, 1999
J:\cjb\cjb77\cjb-10\B99-129.vpMonday, January 17, 2000 3:17:04 PM
Color profile: DisabledComposite Default screen

DC., North American Dithyrea Harv., and DimorphocarpaRollins, and South American Cremolobus DC. Also the nec-tar glands are very variable between the species. This wasalready pointed out by Bayer (1905), and for these reasonshe even excluded Biscutella from his system of Brassica-ceae. The cpDNA data do not support any traditional treat-ment of Biscutella, but would suggest an isolated positionwithin tribe Lepidieae. Al-Shehbaz (1986) and de Candolle(1821) both suggested affinities of Biscutella to the SouthAmerican tribe Cremolobeae. Certainly the classification ofBiscutella needs further studies.
Thlaspi alliance (Thlaspi, Microthlaspi, Noccaea,Peltaria, Teesdalia)
With the exception of Peltaria (see Table 2) these taxawere traditionally classified in subtribe Thlaspidinae. A de-tailed systematic analysis of this group was recently pre-sented by Zunk et al. (1996). The molecular data wouldsuggest that subtribe Thlaspidinae, as traditionally described,is not monophyletic and should be reduced to a core groupconsisting of Thlaspi s.str., Peltaria, Teesdalia, Microthlaspi,and Noccaea including Raparia. Cochlearia, Ionopsidium,and Aethionema, which have been included in various com-binations by previous authors in the Thlaspidinae (seeTable 2), apparently do not belong to this lineage. RbcL se-quencing studies (Price et al. 1994) as well as sequencingstudies of nuclear ITS (K. Mummenhoff, unpublished data)and arginine decarboxylase (Galloway et al. 1998) also dem-onstrate Aethionema as a sister group to the other Bras-sicaceae studied, using Capparaceae as outgroup. This issurprising since the angustiseptate fruits of Aethionema aredifferent from typical fruits of the outgroup in the Cap-paraceae. The systematic position of Aethionema and theidentification of phylogenetically basal taxa within the Bras-sicaceae needs closer inspection.
Cochlearia alliance (Cochlearia, Ionopsidium)Systematic relationships of this group relative to the
Thlaspi alliance has recently been discussed in Zunk et al.(1996). Our present data support common ancestry of Co-chlearia and Ionopsidium. This conclusion is not supportedby previous authors (Table 2). Solely, de Candolle (1821)emphazised close relationships among both genera in treat-ing Ionopsidium as a section of Cochlearia.
OutlookComprehensive molecular studies in the Brassicaceae are
needed to obtain a robust phylogenetic framework that re-flects a more natural classification, which allows to deter-mine phylogenetically basal genera and helps to unravelevolutionary history of traditional morphological characters.
This may be achieved by grouping closely related generaand working upward. Current analyses in our lab includegenera of other tribes that are clearly unrelated to theLepidieae to see whether this tribe is a natural lineage.
Acknowledgments
We thank those who provided plant material for this pro-ject and U. Coja for technical assistance, A. Franzke, I.A.Al-Shehbaz, and one anonymous referee for valuable com-
ments on the manuscript. This work was supported by theGerman Research Foundation (Deutsche Forschungsgemein-schaft).
References
Albert, V.A., Mishler, B.D., and Chase, M.W. 1992. Character stateweighting for restriction site data in phylogenetic reconstruc-tion, with examples from chloroplast DNA. In Molecular sys-tematics of plants. Edited by P.S. Soltis, D.E. Soltis, and J.J.Doyle. Chapman & Hall, New York. pp. 369–401.
Al-Shehbaz, I.A. 1984. The tribes of Cruciferae (Brassicaceae) inthe southeastern United States. J. Arnold Arbor. Harv. Univ. 65:343–373.
Al-Shehbaz, I.A. 1986. The genera of Lepidieae (Cruciferae; Bras-sicaceae) in the southeastern United States. J. Arnold Arbor.Harv. Univ. 67: 265–311.
Al-Shehbaz, I.A. 1987. The genera of Alysseae (Cruciferae; Bras-sicaceae) in the southeastern United States. J. Arnold Arbor.Harv. Univ. 68: 185–240.
Al-Shehbaz, I.A., and Al-Shammary, K.I. 1987. Distribution andchemosystematic significance of glucosinolates in certain mid-dle-eastern Cruciferae. Biochem. Syst. Ecol. 15: 559–569.
Baldwin, B.G., Kyhos, D.W., and Dvorak, J. 1990. ChloroplastDNA evolution and adaptive radiation in the Hawaiian silvers-word alliance (Asteraceae). Ann. Mo. Bot. Gard. 77: 96–109.
Ball, P.W., Heywood, V.H., and Akeroyd, J.R. 1993. Cruciferae. InFlora Europaea. Edited by T.G. Tutin, N.A. Burges, O.A. Chater,J.R. Edmondson, V.H. Heywood, D.M. Moore, D.H. Valentine,S.M. Walters, and D.A. Webb. 2nd ed., Vol. 1. Cambridge Uni-versity Press, Cambridge, U.K. pp. 313–417.
Bayer, A. 1905. Beiträge zur systematischen Gliederung der Cruci-feren. Beih. Bot. Centralbl. 18: 119–180.
Bruneau, A., and Doyle, J.J. 1993. Cladistic analysis of cpDNA re-striction site characters in Erythrina (Leguminosae: Phaseoleae).Syst. Bot. 18: 229–247.
Chase, M.W., and Hills, H.H. 1991. Silica gel: An ideal materialfor field preservation of leaf samples for DNA studies. Taxo, 40:215–220.
Daxenbichler, M.E., van Etten, C.H., Brown, F.S., and Jones, Q.1964. Oxaolidinethiones and volatile isothiocyanates in enzyme-treated seed meals from 65 species of Cruciferae. Agric. FoodChem. 12: 127–130.
Daxenbichler, M.E., Spencer, G.F., Carlson, D.G., Rose, G.B.,Brinker, A.M., and Powell, R.G. 1991. Glucosinolate composi-tion of seeds from 297 species of wild plants. Phytochemistry,38: 2623–2638.
de Candolle, A.P. 1821. Regni vegetabilis systema naturale, vol 2.Treuttel et Würz, Paris, France.
Doyle, J.J. 1991. The pros and cons of DNA systematic data: stud-ies of the wild perennial relatives of soybean. Evol. TrendsPlants, 5: 99–104.
Doyle, J.J., and Doyle, J.L. 1987. A rapid DNA isolation procedurefor small quantities of fresh leaf tissue. Phytochem. Bull. 19:11–15.
Felsenstein, J. 1985. Confidence limits on phylogenies: An ap-proach using the bootstrap. Evolution, 93: 783–791.
Francisco-Ortega, J., Fuertes-Aguilar, J., Gomez-Campo, C.,Santos-Guerra, A., and Jansen, R.K. 1999. ITS sequence phy-logeny of Crambe L. (Brassicaceae): Molecular data reveal twoOld Worl disjunctions. Mol. Phylogenet. Evol. 11: 361–380.
Franzke, A., Pollmann, K., Kohrt, R., and Hurka, H. 1998. Molec-ular systematics of Cardamine and allied genera (Brassicaceae):
© 1999 NRC Canada
Zunk et al. 1511
J:\cjb\cjb77\cjb-10\B99-129.vpMonday, January 17, 2000 3:17:05 PM
Color profile: DisabledComposite Default screen

© 1999 NRC Canada
1512 Can. J. Bot. Vol. 77, 1999
ITS and non-coding chloroplast DNA. Folia Geobot. Phytotax.33: 225–240.
Galloway, G.L., Malmberg, R.L., and Price, R.A. 1998. Phylogen-etic utility of the nuclear gene arginine decarboxylase: an exam-ple from Brassicaceae. Mol. Biol. Evol. 15: 1312–1320.
Gielly, L., and Taberlet, P. 1996. A phylogeny of the Europeangentians inferred from chloroplast trnL (UAA) intron sequences.Bot. J. Linn. Soc. 120: 57–75.
Hayek, A. 1911. Entwurf eines Cruciferensystems auf phylogene-tischer Grundlage. Beih. Bot. Centralbl. 27: 127–335.
Hedge, I.C. 1976. A systematic and geographical survey of the OldWorld Cruciferae. In The biology and chemistry of the Cruci-ferae. Edited by J.G. Vaughan, A.J. MacLeod, and M.B. Jones.Academic Press, London, England. pp. 1–45.
Hillis, D.M., and Bull, J.J. 1993. An empirical test of bootstrap-ping as a method for assessing confidence in phylogenetic anal-ysis. Syst. Biol. 42: 182–192.
Hillis, D.M., and Huelsenbeck, J.P. 1992. Signal, noise, and reliabil-ity in molecular phylogenetic analyses. J. Hered. 83: 189–195.
Holsinger, K.E., and Jansen, R.K. 1993. Phylogenetic analysis ofrestriction site data. Methods Enzymol. 224: 439–455.
Janchen, E. 1942. Das System der Cruciferen. Oesterr. Bot. Z. 91:1–28.
Jaretzky, R. 1928. Untersuchungen über Chromosomen und Phylo-genie bei einigen Cruciferen. Jahrb. Wiss. Bot. 68: 1–45.
Kellog, E.A., and Watson, L. 1993. Phylogenetic studies of a largedata set. I. Bambusoideae, Andropogonaceae, and Pooidae (Gra-mineae). Bot. Rev. 59: 273–343.
Kjær, A. 1960. Naturally derived iso-thiocyanates (mustard oils)and their parent glucosinolates. Fortschr. Chem. Org. Naturst.18: 122–176.
Koch, M., Hurka, H., and Mummenhoff, K. 1996. ChloroplastDNA restriction site variation and RAPD-analyses in Cochlearia(Brassicaceae): Biosystematics and speciation. Nord. J. Bot. 16:585–605.
Koch, M., Mummenhoff, K., and Hurka, H. 1998. Molecular bio-geography and evolution of the Micothlaspi perfoliatum s.l.polyploidy complex (Brassicaceae): chloroplast DNA and nu-clear ribosomal DNA restriction site variation. Can. J. Bot. 76:382–396.
Maddison, D.R. 1991. The discovery and importance of multipleislands of most parsimonious trees. Syst. Zool. 40: 315–328.
Markgraf, F. 1963. Cruciferae. In G. Hegi, Illustrierte Flora vonMitteleuropa. 2nd ed., vol 4 (1). Edited by F. Markgraf. Hauser,Munich. pp. 73–514.
Meyer, F.K. 1973. Conspectus der “Thlaspi” Arten Europas,Afrikas und Vorderasiens. Feddes Repert. 84: 449–470.
Meyer, F.K. 1979. Kritische Revision der “Thlaspi” Arten Europas,Afrikas und Vorderasiens. Feddes Repert. 90: 129–154.
Mikolajczak, K.L., Miwa, T.K., Earle, F.R., and Wolff, I.A. 1961.Search for new industrial oils. V. Oils of Cruciferae. J. Am. OilChem. Soc. 38: 678–691.
Miller, R.W., Earle, F.R., and Wolff, I.A. 1965. Search for new in-dustrial oils. XIII. Oils from 102 species of Cruciferae. J. Am.Oil Chem. Soc. 42: 817–821.
Mukherjee, K.D., Kiewitt, I., and Hurka, H. 1984. Lipid contentand fatty acid composition of seeds of Capsella species fromdifferent geographical localities. Phytochemistry, 23: 117–119.
Mummenhoff, K., and Koch, M. 1994. Chloroplast DNA restric-tion site variation and phylogenetic relationships in the genusThlaspi sensu lato (Brassicaceae). Syst. Bot. 19: 73–88.
Mummenhoff, K., Kuhnt, E., Koch, M., and Zunk, K. 1995. Sys-tematic implications of chloroplast DNA variation in Lepidiumsections Cardamon, Lepiocardamon and Lepia (Brassicaceae).Plant. Syst. Evol. 196: 75–88.
Mummenhoff, K., Franzke, A., and Koch, M. 1997a. Moleculardata reveal convergence in fruit characters used in the classifica-tion of Thlaspi s.l. (Brassicaceae). Bot. J. Linn. Soc. 125: 183–199.
Mummenhoff, K., Franzke, A., and Koch, M. 1997b. Molecularphylogenetics of Thlaspi s.l. (Brassicaceae) based on sequencesof the internal transcribed spacers of the nuclear ribosomalDNA. Can. J. Bot. 75: 469–482.
Olmstead, R.G., and Palmer, J.D. 1994. Chloroplast DNA system-atics: A review of methods and data analysis. Am. J. Bot. 81:1205–1224.
Price, R.A., Palmer, J.D., and Al-Shehbaz, I.A. 1994. Systematicrelationships of Arabidopsis: A molecular and morphologicalperspective. In Arabidopsis. Edited by E.M. Meyerowitz and C.Somerville. Cold Spring Harbor Press, Cold Spring Harbor, N.Y.pp. 7–19.
Rodman, J.E. 1991. A taxonomic analysis of glucosinolate-producing plants, part 2: cladistics. Syst. Bot. 16: 619–629.
Rodman, J.E., Price, R.A., Karo, L.K., Conti, E., Sytsma, K.J., andPalmer, J.D. 1993. Nucleotide sequences of the rbcL gene indi-cate monophyly of mustard oil plants. Ann. Mo. Bot. Gard. 80:686–699.
Schulz, O.E. 1936. Cruciferae. In Die natürlichen Pflanzenfami-lien, 17B. Edited by A. Engler and K. Prantl. Engelmann, Leip-zig, Germany. pp. 227–658.
Shaw, E.A. 1965. Taxonomic revision of some Australian endemicgenera of Cruciferae. Trans. R. Soc. S. Aust. 89: 145–253.
Swofford, D.L. 1993. PAUP: Phylogenetic analysis using parsi-mony, version 3.1.1 for Macintosh. Illinois Nat. Hist. Survey,Champaign, Ill.
Vaughan, J.G., and Whitehouse, J.M. 1971. Seed structure and thetaxonomy of the Cruciferae. Bot. J. Linn. Soc. 64: 383–409.
Wagstaff, S.J., Olmstead, R.G., and Cantino, P.D. 1995. Parsimonyanalysis of cpDNA restriction site variation in subfamilyNepetoideae (Labiatae). Am. J. Bot. 82: 886–892.
Warwick, S.I., and Black, L.D. 1991. Molecular systematics ofBrassica and allied genera (subtribe Brassicinae, Brassiceae) —chloroplast genome and cytodeme congruence. Theor. Appl.Genet. 82: 81–92.
Warwick, S.I., and Black, L.D. 1997. Molecular phylogenies fromtheory to application in Brassica and allies (tribe Brassiceae,Cruciferae). Opera Bot. 132: 149–168.
Watrous, L.E., and Wheeler, Q.D. 1981. The outgroup comparisonmethod of character analysis. Syst. Zool. 30: 1–11.
Whitton, J., Wallace, R.S., and Jansen, R.K. 1995. Phylogenetic re-lationships and patterns of character change in the tribe Lac-tuceae (Asteraceae) based on chloroplast DNA restriction sitevariation. Can. J. Bot. 73: 1058–1073.
Zunk, K., Mummenhoff, K., Koch, M., and Hurka, H. 1996. Phylo-genetic relationships of Thlaspi s.l. (subtribe Thlaspidinae,Lepidieae) and allied genera based on chloroplast DNA restric-tion-site variation. Theor. Appl. Genet. 92: 375–381.
J:\cjb\cjb77\cjb-10\B99-129.vpMonday, January 17, 2000 3:17:05 PM
Color profile: DisabledComposite Default screen




















