
Phenotypic and genotypic characteristics of Phormidium -like cyanobacteria inhabiting microbial mats...
16
Phenotypic and Genotypic Characteristics of Persistent Methicillin-Resistant Staphylococcus aureus Bacteremia In Vitro and in an Experimental Endocarditis Model Yan Q. Xiong 1,2 , Vance G. Fowler Jr. 4 , Michael R. Yeaman 1,2 , Francoise Perdreau- Remington 3 , Barry N. Kreiswirth 5,6 , and Arnold S. Bayer 1,2 1 Division of Infectious Diseases, Los Angeles Biomedical Research Institute at Harbor-UCLA, Torrance 2 David Geffen School of Medicine at UCLA, Los Angeles 3 Division of Infectious Diseases, Department of Medicine, University of California, San Francisco, California 4 Division of Infectious Diseases, Duke University Medical Center, Durham, North Carolina 5 Public Health Research Institute, Newark, New Jersey 6 University of Medicine and Dentistry of New Jersey, Newark, New Jersey Abstract Background—Persistent MRSA bacteremia (PB) represents an important subset of Staphylococcus aureus infections and correlates with poor clinical outcomes. Methods—We profiled relevant in vitro phenotypic and genotypic characteristics of MRSA isolates from 39 persons with bacteremia (21 had PB and 18 had resolving bacteremia [RB]). We also compared the intrinsic virulence and responsiveness to vancomycin of selected PB and RB strains in an experimental endocarditis model (IE). Results—PB and RB isolates differed significantly with regard to several in vitro characteristics that are believed to impact endovascular infections. PB isolates exhibited significantly more resistance to the cationic defensin hNP-1, enhanced membrane fluidity, and substantially greater adhesion to fibronectin, fibrinogen, and endothelial cells. Genotypically, PB isolates had higher frequency of SCCmec II, CC30, and spa 16; and higher rates of agr type III, cap8, tst-1, and cna carriage. Finally, a prototypic PB strain was more resistant to vancomycin treatment in the infective endocarditis model than a RB comparator strain, despite equivalent virulence profiles. Conclusions—Our findings indicate that PB isolates may have specific virulence signatures that distinguish them from RB isolates. These data suggest that methods might be developed to identify patients at higher risk for PB in real-time, thereby optimizing the effectiveness of anti-MRSA therapeutic strategies. The persistent bacteremia (PB) syndrome is well represented in large clinical series of persons with methicillin-resistant Staphylococcus aureus (MRSA) bloodstream infection (PB © 2008 by the Infectious Diseases Society of America. All rights reserved. Reprints or correspondence: Dr. Yan Q. Xiong, Dept. of Medicine, Div. of Infectious Diseases, Los Angeles Biomedical Research Institute at Harbor-UCLA Medical Center, Torrance, CA 90502 ([email protected]).. Potential conflicts of interest: none reported. NIH Public Access Author Manuscript J Infect Dis. Author manuscript; available in PMC 2010 February 23. Published in final edited form as: J Infect Dis. 2009 January 15; 199(2): 201–208. doi:10.1086/595738. NIH-PA Author Manuscript NIH-PA Author Manuscript NIH-PA Author Manuscript
Transcript of Phenotypic and genotypic characteristics of Phormidium -like cyanobacteria inhabiting microbial mats...

Phenotypic and Genotypic Characteristics of PersistentMethicillin-Resistant Staphylococcus aureus Bacteremia In Vitroand in an Experimental Endocarditis Model
Yan Q. Xiong1,2, Vance G. Fowler Jr.4, Michael R. Yeaman1,2, Francoise Perdreau-Remington3, Barry N. Kreiswirth5,6, and Arnold S. Bayer1,21Division of Infectious Diseases, Los Angeles Biomedical Research Institute at Harbor-UCLA,Torrance2David Geffen School of Medicine at UCLA, Los Angeles3Division of Infectious Diseases, Department of Medicine, University of California, San Francisco,California4Division of Infectious Diseases, Duke University Medical Center, Durham, North Carolina5Public Health Research Institute, Newark, New Jersey6University of Medicine and Dentistry of New Jersey, Newark, New Jersey
AbstractBackground—Persistent MRSA bacteremia (PB) represents an important subset ofStaphylococcus aureus infections and correlates with poor clinical outcomes.
Methods—We profiled relevant in vitro phenotypic and genotypic characteristics of MRSA isolatesfrom 39 persons with bacteremia (21 had PB and 18 had resolving bacteremia [RB]). We alsocompared the intrinsic virulence and responsiveness to vancomycin of selected PB and RB strainsin an experimental endocarditis model (IE).
Results—PB and RB isolates differed significantly with regard to several in vitro characteristicsthat are believed to impact endovascular infections. PB isolates exhibited significantly moreresistance to the cationic defensin hNP-1, enhanced membrane fluidity, and substantially greateradhesion to fibronectin, fibrinogen, and endothelial cells. Genotypically, PB isolates had higherfrequency of SCCmec II, CC30, and spa 16; and higher rates of agr type III, cap8, tst-1, and cnacarriage. Finally, a prototypic PB strain was more resistant to vancomycin treatment in the infectiveendocarditis model than a RB comparator strain, despite equivalent virulence profiles.
Conclusions—Our findings indicate that PB isolates may have specific virulence signatures thatdistinguish them from RB isolates. These data suggest that methods might be developed to identifypatients at higher risk for PB in real-time, thereby optimizing the effectiveness of anti-MRSAtherapeutic strategies.
The persistent bacteremia (PB) syndrome is well represented in large clinical series of personswith methicillin-resistant Staphylococcus aureus (MRSA) bloodstream infection (PB
© 2008 by the Infectious Diseases Society of America. All rights reserved.Reprints or correspondence: Dr. Yan Q. Xiong, Dept. of Medicine, Div. of Infectious Diseases, Los Angeles Biomedical Research Instituteat Harbor-UCLA Medical Center, Torrance, CA 90502 ([email protected])..Potential conflicts of interest: none reported.
NIH Public AccessAuthor ManuscriptJ Infect Dis. Author manuscript; available in PMC 2010 February 23.
Published in final edited form as:J Infect Dis. 2009 January 15; 199(2): 201–208. doi:10.1086/595738.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript

prevalence, 20%–30%), and it is especially relevant to patients with endovascular infection[1,2]. However, in up to one-third of cases, no readily identifiable cause of PB is identified,despite extensive clinical evaluation [3]. Therefore, understanding the molecular mechanismsand determinants of PB is essential to optimize prevention and therapy against life-threateningS. aureus infections.
We hypothesize that S. aureus uses specific virulence determinants to persist in the bloodstreamand cause PB in the context of endovascular infection. For example, the organism must avoidimmediate killing by host-defense peptides liberated by platelets at sites of endovascularinfection. Next, the organism must evade or survive phagocytosis and intracellular killing byneutrophil-associated oxidative and nonoxidative killing mechanisms (including thoseassociated with antimicrobial peptides, such as α-defensins). During this phase, the pathogenmust adhere to host cells and ligands to colonize vascular endothelium and avoid clearance bythe reticuloendothelial system. Following adherence, the organism proliferates and invadestissues, creating reservoir foci. Finally, it deploys exotoxins and exoenzymes to reemerge fromthese reservoir sites, reenter the bloodstream, and hematogenously seed metastatic targetorgans (figure 1).
Because distinct infection foci are not synchronized, organisms are continuously reemerginginto the bloodstream, accounting for persistent bacteremia. On the basis of this hypothesizedlife cycle of PB isolates, we investigated a cadre of in vitro phenotypic and genotypiccharacteristics considered to be involved in these phases of endovascular pathogenesis. Inaddition, we examined the in vivo virulence and antibiotic responsiveness of MRSA isolatesassociated with PB (hereafter, “PB isolates”) and those associated with resolving bacteremia(here-after, “RB isolates”) in a rabbit model of infective endocarditis.
PATIENTS, MATERIALS, AND METHODSCollection of strains
All MRSA isolates were recovered from bacteremic patients at Duke University MedicalCenter during 1994–1999. All isolates were susceptible to vancomycin (minimum inhibitoryconcentration [MIC] range, 0.25–1μg/mL), with no significant differences in MICs betweenPB and RB strains.
Two groups of MRSA isolates were studied. The first group was collected from 21 patientswith PB, all of whom had MRSA-positive blood cultures for ≥7 days while receiving anantibiotic to which the isolate was susceptible. The second group was collected from 18 patientswith RB who had an initial blood culture that yielded MRSA and subsequent blood culturesthat yielded no MRSA 2–4 days after therapy initiation. Patients in the 2 cohorts were matchedon the basis of demographic characteristics. Both patient groups had similar initial clinicalcharacteristics and laboratory findings but differed significantly with respect to characteristicsassociated with clinical course and outcome [1,4,5].
In vitro assay for susceptibility to hNP-1In a recent report, we showed that PB isolates were significantly more resistant than RB isolatesto mammalian platelet microbicidal proteins in vitro [1]. To evaluate whether the disparity inthe in vitro susceptibility profiles of PB and RB isolates extended to other important host-defense effectors, we assessed their in vitro responses to hNP-1, a 3.8-kDa cationic α-defensinpeptide from human neutrophils [6-8]. Nonoxidative neutrophil-mediated killing related to thispeptide family is likely important during the initial bacteremic clearance stage of endovascularpathogenesis [6,9]. Purified hNP-1 was purchased from Peptide International. Bacterialsusceptibility to hNP-1 was assayed in vitro, as described in detail elsewhere [7]. Results are
Xiong et al. Page 2
J Infect Dis. Author manuscript; available in PMC 2010 February 23.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript

expressed as the percentage of colony-forming units in the initial inoculum (105 cfu) thatsurvived exposure to hNP-1 after a 2-h incubation period.
In vitro measurement of adherence to host endovascular ligands (fibrinogen and fibronectin)and host cells (endothelial cell and platelets)
Tissue culture plates were coated with purified human fibrinogen (50 μg/mL; Sigma Chemical)or fibronectin (50 μg/mL; Sigma Chemical) and washed with phosphate-buffered saline (PBS;pH 7.2) [10,11]. Plates were then treated with 3% bovine serum albumin (Sigma Chemicals)to prevent nonspecific adhesion and were washed again with PBS before organism seeding.
Human umbilical cord veins were obtained and maintained as previously described [12].Endothelial cell monolayer plates were washed twice with prewarmed Hanks balanced saltsolution before organism seeding.
In assays to detect fibrinogen, fibronectin, and endothelial cell binding, PB or RB isolates thathad been cultured overnight were added at a final inoculum of 5 × 103 cfu/mL to control plateswithout either matrix ligands or endothelial cells, to fibrinogen-coated or fibronectin-coatedplates, or to endothelial cell monolayer-coated plates. The plates were incubated for 1 hunderstatic conditions, as previously described [10-12]. Unbound bacteria were removed by washingwith PBS, and tryptic soy agar was added. Adherence was expressed as the percentage of theinitial inoculum bound.
Bacterial adherence to platelets was tested as previously described [13]. Fresh rabbit platelets(109 platelets/mL) were specifically labeled with CellTracker Red (2.5 mmol/L). Overnight-cultured PB or RB isolates (109 cfu/mL) were labeled with a fluorescent dye, Syto13 (2 mmol/L). Binding studies were done by mixing Syto13-labeled S. aureus cells (108 cfu/mL) and Cell-Tracker Red–labeled platelets (107 platelets/mL). The percentage of S. aureus cells bound toplatelets was calculated as previously described [13].
Measurement of S. aureus cell membrane fluidityWe previously showed that cationic peptide-resistant S. aureus strains (e.g., strains resistantto tPMP-1) exhibited significantly higher degrees of membrane fluidity [14]. Such resistanceis believed to contribute to both initial bloodstream and progressive stages of endovascularinfection [15]. Therefore, we tested the membrane fluidity of the strain sets, using thefluorescent probe 1,6-diphenyl-1,3,5-hexatriene (DPH) [7,14]. Protocols for DPHincorporation into target cell membranes, measurement of fluorescence polarization, andcalculation of the degree of fluorescence polarization index are described in detail elsewhere[14]. As the polarization index decreases, the degree of membrane fluidity increases [14].
E-testMacro E-tests (AB Biodisk) were performed to screen for hetero–vancomycin-intermediatelyresistant S. aureus (hetero-VISA). The assays were done as recommended by the manufacturer.Strains with vancomycin MICs of ≥8 μg/mL were defined as hetero-VISA [16].
Multilocus sequence typing (MLST) and clonal complexes (CCs)MLST is a powerful typing tool for defining the molecular epidemiology of S. aureus becauseit can identify specific MLST clones and group strains into unique CCs [17]. It has recentlybeen shown that such bacterial genotypes are associated with distinct clinical outcomes [18].In addition, CC5 and CC30 have been shown to be significantly associated with more-severeS. aureus infection [19]. The protocol for MLST has been published previously [19].
Xiong et al. Page 3
J Infect Dis. Author manuscript; available in PMC 2010 February 23.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript

Staphylococcal protein A (spa) typingSequence analysis of the tandem repeat region in the spa subtypes of MRSA helps furtherdiscriminate the MLST profiles into distinct lineages [20]. We have previously shown thatspa type 16 is associated with more-complicated outcomes in patients with S. aureusbacteremia [19]. The spa typing of the current strain set was performed by means of previouslypublished methods [21].
Staphylococcal cassette chromosome mec (SCCmec) classificationSCCmec typing distinguishes MRSA strains into 5 major classes (types I–V) and providesanother genotypic assessment to further refine strain differentiation on the basis of MLST andspa typing. SCCmec typing was performed for several important reasons: (1) our prior pulsed-field gel electrophoresis profiling suggested that PB isolates were of 2 predominant and relatedlineages (A1 and A2) [1]; (2) because isolates were obtained from a single referral center (DukeUniversity Medical Center), it was likely that clonal bias would be identified; and (3) specificSCCmec types may be associated with specific clinical syndromes (e.g., skin and soft-tissueinfections related to SCCmec type IV) [19]. SCCmec typing was performed on the 39 MRSAstrains, using methods described elsewhere [22].
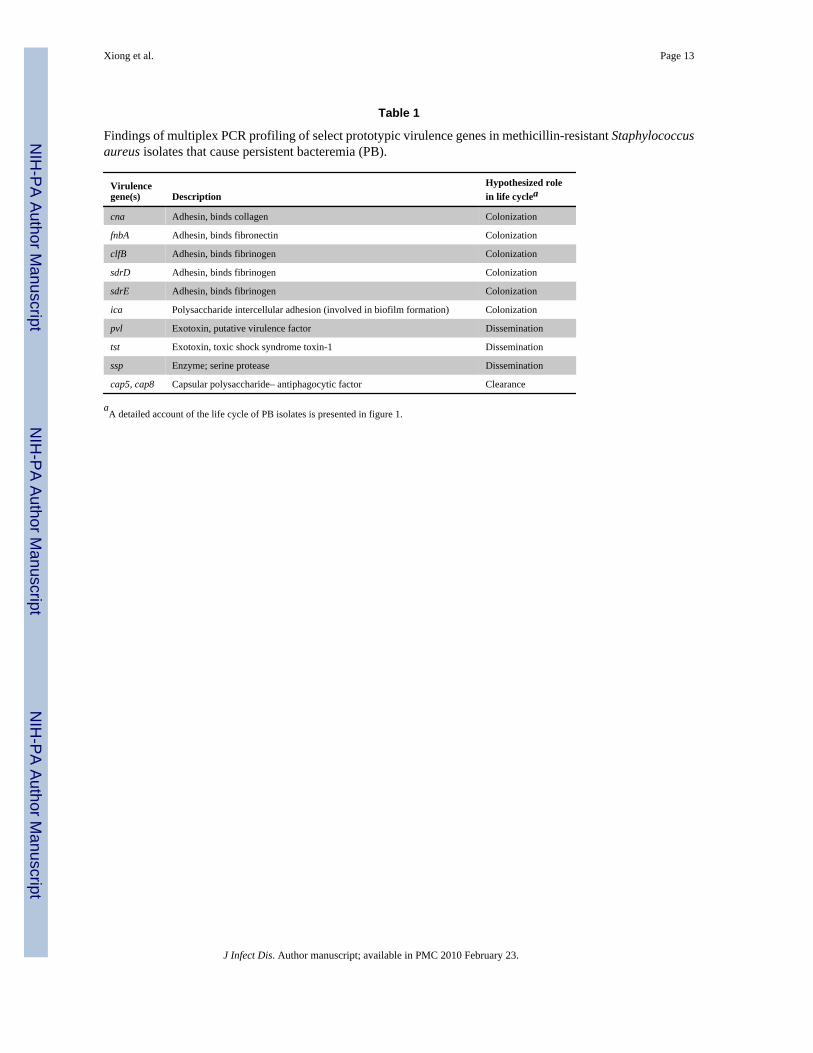
Multiplex PCR for S. aureus virulence genesCoordinated sets of virulence genes or gene networks are believed to be involved in ≥1pathogenic step during MRSA infection (table 1) [23,24]. To compare the endovascular virulonof PB and RB strains, we used multiplex PCR to determine the presence or absence of 33 suchvirulence genes, representing a range of regulatory, adhesin, exotoxin, antiphagocytic, andexoenzyme genes. The PCR-based assays were performed by use of previously publishedtechniques [25].
Rabbit infective endocarditis modelA well-characterized model of catheter-induced infective endocarditis in rabbits was used tostudy the following 4 phasic outcomes of infection due to PB and RB isolates in vivo [15,24]: early bacteremia clearance, initial vegetation colonization, composite virulence, andresponsiveness to vancomycin therapy. Infective endocarditis was produced by intravenousinjection of select PB isolates (from strain 420) or RB isolates (from strain 1507) 24 h aftercatheterization. Strains 420 and 1507 were selected because of their highly distinguishable invitro phenotypic and genotypic profiles (table 2). Phenotypically, the PB strain differed fromthe RB strain in several features likely to impact endovascular pathogenesis: the PB strain hasgreater resistance to killing by a representative platelet antimicrobial peptide, tPMP-1 [4];greater adherence to endothelial cells, fibronectin, and fibrinogen in vitro; and substantiallymore fluidic cell membranes (table 2). Genotypically, the PB strain was CC30 and spa type16 (both of which were associated with more-severe MRSA clinical infections in other studies[19]) and harbored SCCmec type II; in addition, this strain was agr type III and positive forcna and tst-1. In contrast, the RB strain was CC8 and spa type 7, contained SCCmec type IV,and was negative for agr type I, cna, and tst-1 (table 2). Both strains were pvl negative and hadvancomycin MICs of 1 μg/mL. Neither strain exhibited a hetero-VISA phenotype (data notshown).
Each animal was challenged intravenously with 109 cfu of PB strain 420 or RB strain 1507 24h after catheterization. Blood samples were obtained for quantitative culture 1 min and 30 minafter challenge. Bacterial blood density was expressed in mean log10 cfu/mL (±SD). Inaddition, animals were sacrificed 30 min after challenge, and all vegetations were removed forquantitative culture. The in vivo adherence of each strain to vegetations was expressed in meanlog10 cfu per g of vegetation (±SD).
Xiong et al. Page 4
J Infect Dis. Author manuscript; available in PMC 2010 February 23.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript

The intrinsic virulence of infective endocarditis strains can be measured as a composite ofinfective endocarditis induction rates over an inoculum challenge range and of target tissuebacterial densities 24 h after receipt of the ID95 inoculum. Thus, 24 h after catheterization,aortic-catheterized animals were challenged intravenously with 104, 105, or 106 cfu of PB orRB strains, the inoculum range that encompasses the ID95 for most S. aureus strains in thismodel [26]. Twenty-four hours after inoculation, all animals were euthanized, and their cardiacvegetations, kidneys, and spleen were removed and quantitatively cultured.
Because PB isolates were obtained from patients with no response to vancomycin therapy, wepostulated that the in vivo response of the PB and RB strains to vancomycin therapy woulddiffer in the infective endocarditis model. Twenty-four hours after receipt of the infectiveendocarditis-inducing inoculation (105 cfu), animals were randomized to receive either notherapy (controls) or vancomycin (15 mg/kg intravenously twice per day for 3 days). Thisvancomycin dose strategy is associated with slow and incomplete clearance of most susceptibleS. aureus strains from vegetations and other target organs over a 3-day treatment period [27].Twenty-four hours after the last antibiotic dose, all animals were sacrificed, and target tissueswere removed and quantitatively cultured as described above.
Statistical analysisData were analyzed by Kruskal-Wallis analysis of variance, with corrections made for multiplecomparisons when appropriate. P values of ≤.05 were considered statistically significant.
RESULTSIn vitro susceptibility to hNP-1
Nearly all strains were resistant to a 20-μg/mL dose of hNP-1. However, at a 40-μg/mL doseof hNP-1, the percentage of isolates that survived was significantly higher in the PB group,compared with the RB group (P = .04) (table 3).
In vitro adherence to host endovascular ligands (fibrinogen and fibronectin) and host cells(endothelial cells and platelets)
In general, PB isolates bound better than RB isolates to matrix ligands and host cells relevantto endovascular infection. For example, the PB group had substantially greater mean adherenceto fibrinogen and fibronectin than the RB group (figure 2; P < .05 for fibrinogen binding).Results also indicated that PB isolates selected for infective endocarditis studies boundsubstantially better to human endothelial cell monolayers than their RB counterparts (figure2). In contrast, there was no difference in the capacity of the PB and RB strain sets overall tobind to platelets (figure 2).
S. aureus cell membrane fluidityThe PB strain set exhibited higher cell membrane fluidity than RB isolates. The meanpolarization indices of membrane fluidity (±SD) for PB and RB isolates were 0.276 ± 0.03 and0.292 ± 0.02, respectively. However, these differences did not reach statistical significance(P = .11).
PB and RB isolates did not differ in other relevant phenotypic characteristicsNo PB or RB isolate had a hetero-VISA phenotype detected by means of a macro E-test.
Xiong et al. Page 5
J Infect Dis. Author manuscript; available in PMC 2010 February 23.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript

MLST and CCsA total of 9 sequence types contained in 5 CCs were represented among the 39 isolates. Themost common CC was CC30, present in 27 (69%) of 39 isolates. PB isolates were significantlymore likely than RB isolates to be CC30 (90% vs. 44%; P < .05) (table 3).
Staphylococcal protein A (spa) typingConsistent with findings in the recent study by Fowler et al. [19], there was an association trendof spa type 16 (W-G-K-A-K-A-O-M-Q-Q-Q-Q) with the PB strain set. Thus, 57% of PBisolates were spa type 16, compared with 39% of RB isolates (P = .16) (table 3).
SCCmec classificationSCCmec II was the most commonly observed type (85% of isolates). In addition, 20 PB isolates(95%) contained the SCCmec II element, compared with 13 RB isolates (72%; P < .05) (table3).
Multiplex PCR for S. aureus virulence genesA total of 81% of PB isolates were agr type III, compared with only 44% of RB isolates (P = .017) (table 3). All isolates possessed the fnbA and clfB surface adhesin genes. Of note, 85% ofPB isolates were cna positive, compared with only 56% of RB isolates (P = .07) (table 3). Also,only 15% of PB isolates carried sdrD and sdrE, compared with 47% of RB isolates (P < .05).Carriage of the tst-1 endotoxin gene was significantly over-represented among PB isolates,compared with RB isolates (86% vs. 44%; P = .03). Panton-Valentine leukocidin locus wasabsent in all strains (table 3). A total of 86% of PB isolates had the cap type 8 genotype(cap8); among RB isolates, 39% had the cap8 genotype, and 61% had the cap5 genotype (table3). Last, all strains carried the biofilm-associated ica gene, as well as the V8 protease genessp. There were no significant differences between the PB and RB strain sets with regard tothe presence or absence of other virulence genes studied (table 3).
Infective endocarditis modelNo significant differences between the rate of early clearance of the PB isolate and the rate ofearly clearance of the RB isolate were observed 1 and 30 min after infection (data not shown).In addition, no significant differences were observed between the PB and RB strains withrespect to the extent of initial colonization of vegetations (data not shown).
At the 104, 105 and 106 cfu inocula, all catheterized animals developed infective endocarditis.At inocula of 104 cfu and 105 cfu (table 4), bacterial densities in the 3 target tissues were notsignificantly different between animals infected with the RB isolate and those infected withthe PB isolate. The majority of animals infected with either the PB or RB strain at an inoculumof 106 cfu died ≤24 h after infection.
As shown in table 4, vancomycin therapy resulted in significant reductions of all target tissueMRSA densities in animals infected with either PB or RB isolates, compared with respectiveuntreated controls (P < .005). Of note, for cardiac vegetations, the residual MRSA density aftervancomycin treatment in animals infected with the PB strain was ~2.0 log10 cfu per g ofvegetation greater than that observed for animals infected with the RB strain (P = .01) (table4).
DISCUSSIONA number of interesting observations emerged from the present investigations. Phenotypically,the current data extend upon past findings regarding antimicrobial peptide resistance; PB
Xiong et al. Page 6
J Infect Dis. Author manuscript; available in PMC 2010 February 23.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript

strains tend to be more resistant to key innate cationic host defense molecules from bothneutrophils (e.g., hNP-1) and platelets (e.g., tPMP-1) [1]. These findings suggest that PBisolates have an intrinsic capacity to survive interactions with 2 predominant host defense cellsearly in the course of bloodstream invasion (figure 1, phase 1 bacteremia). The ability of PBstrains to circumvent such innate immune defenses likely enhance their subsequent pathogenicpotential [15,28]. Furthermore, since these 2 innate defense molecules may be important inmultiple stages of endovascular pathogenesis (e.g., tPMP-1–resistance fostering infectiveendocarditis progression) [28], the ability of a strain to resist their microbicidal actions probablycontributes to PB.
The present findings also demonstrated that PB isolates adhere better than RB isolates to hostcells (i.e., endothelium) and matrix ligands relevant to endovascular pathogenesis (i.e.,fibrinogen and fibronectin). Such capabilities may facilitate the colonization phases of PBinfection (phase 2 colonization) (figure 1). Also, since fibronectin and fibrinogen binding arenow considered integral to endothelial cell and vegetation persistence in experimental infectiveendocarditis [29], increased binding of PB isolates to these ligands, compared with binding ofRB isolates, would theoretically provide an advantage for PB pathogenesis.
Importantly, PB isolates exhibited substantially more fluidic membranes than RB isolates. Afluidic membrane phenotype has previously been linked to cationic antimicrobial peptideresistance in S. aureus laboratory strains [14]. This characteristic may facilitate persistent andprogressive infective endocarditis due to PB isolates [14]. The mechanism (or mechanisms)by which enhanced fluidity causes increased resistance to such peptides is not well understoodbut is postulated to be associated with reduced membrane binding or intramembraneorganization of these cationic molecules.
Genotypically, a greater percentage of PB isolates were associated with SCCmec II (95%,compared with 72% of RB isolates), CC30 (90% vs. 44%), and spa type 16 (57% vs. 39%).This observation is in line with recent findings by Fowler et al. [19], who demonstrated asignificant trend toward more frequent hematogenous complications in strains exhibiting thesegenotypic profiles. Multiplex PCR revealed that PB isolates differed from RB isolates in termsof overrepresentation of capsule type 8 and cna and tst-1 gene carriage. The increased presenceof the adhesin gene, cna, would theoretically allow PB strains to exploit specific anatomictargets, such as bones, joints, or endothelial substrata [30]. This role could potentiallycontribute to enhancement of the colonization and/or persistence phases in the life cycle of PBisolates. In addition, overrepresentation of the tst-1 gene could ostensibly increase the incidenceof “cytokine storm”-associated sepsis syndromes, leading to worse clinical outcomes forpatients with PB [31].
The net intrinsic virulence properties of these 2 strains were not different in the context ofinfective endocarditis induction or progression. In contrast, vancomycin therapy clearlydivulged significant outcome differences between groups. Thus, isolates from the PB strain setwere able to persist within cardiac vegetations to a greater extent than those from the RB strainset during vancomycin therapy. This result occurred despite identical vancomycin MICs andno hetero-VISA subpopulations in the PB and RB strain sets. A similar observation by Fowleret al. [1] and Hawkins et al. [32] demonstrated that vancomycin susceptibility was not decreasedamong PB isolates. However, these findings contrast with those of other studies in which thePB phenotype has been associated with reduced susceptibility to vancomycin [33,34]. Thesein vivo data regarding vancomycin-induced disclosure of the PB outcome suggest severalinteresting possibilities. For example, PB and RB isolates may differ in other features notassessed in our profiling that may impact net responsiveness to vancomycin (e.g., cell wallperturbations, cell surface charge, and global metabolic pathway abnormalities) [7]. Also, it is
Xiong et al. Page 7
J Infect Dis. Author manuscript; available in PMC 2010 February 23.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript

possible that vancomycin may itself differentially impact virulence pathways in PB isolates,compared with RB isolates.
Our study has several potential limitations. First, all strains were obtained from DukeUniversity Medical Center, raising the possibility of single-center and/or geographic bias.Second, these strains emanated from 1994–1999 and therefore do not represent recent shifts.Third, we compared only the initial PB and RB isolates and did not screen for virulencesignatures that may have adaptively evolved during treatment among follow-up blood isolates.Finally, we only examined a single PB-RB strain pair in vivo. Current studies are beingdesigned to address these limitations.
In summary, the present data support our hypothesis that there are significant phenotypic andgenotypic profiles that can distinguish PB isolates from RB isolates. Characterization of PBisolates may afford breakthrough discoveries in the treatment of life-threatening MRSAinfections.
AcknowledgmentsFinancial support:
National Institutes of Health (grants AI-039108 to A.S.B., AI-059111 and AI-068804 to V.G.F., and AI-039001 toM.R.Y.); American Heart Association (grant 0465142Y to Y.Q.X.]).
References1. Fowler VG Jr, Sakoulas G, McIntyre LM, et al. Persistent bacteremia due to methicillin-resistant
Staphylococcus aureus infection is associated with agr dysfunction and low-level in vitro resistanceto thrombin-induced platelet microbicidal protein. J Infect Dis 2004;190:1140–9. [PubMed:15319865]
2. El-Ahdab F, Benjamin DK Jr, Wang A, et al. Risk of endocarditis among patients with prosthetic valvesand Staphylococcus aureus bacteremia. Am J Med 2005;118:225–9. [PubMed: 15745719]
3. Khatib R, Johnson LB, Fakih MG, et al. Persistence in Staphylococcus aureus bacteremia: incidence,characteristics of patients and outcome. Scand J Infect Dis 2006;38:7–14. [PubMed: 16338832]
4. Fowler VG Jr, McIntyre LM, Yeaman MR, et al. In vitro resistance to thrombin-induced plateletmicrobicidal protein in isolates of Staphylococcus aureus from endocarditis patients correlates withan intravascular device source. J Infect Dis 2000;182:1251–4. [PubMed: 10979928]
5. Fowler VG Jr, Kong LK, Corey GR, et al. Recurrent Staphylococcus aureus bacteremia: pulsed-fieldgel electrophoresis findings in 29 patients. J Infect Dis 1999;179:1157–61. [PubMed: 10191218]
6. Ganz T. Defensins: antimicrobial peptides of innate immunity. Nat Rev Immunol 2003;3:710–20.[PubMed: 12949495]
7. Xiong YQ, Mukhopadhyay K, Yeaman MR, Adler-Moore J, Bayer AS. Functional interrelationshipsbetween cell membrane and cell wall in antimicrobial peptide-mediated killing of Staphylococcusaureus. Anti-microb Agents Chemother 2005;49:3114–21.
8. White SH, Wimley WC, Selsted ME. Structure, function, and membrane integration of defensins. CurrOpin Struct Biol 1995;5:521–7. [PubMed: 8528769]
9. Nathan C. Neutrophils and immunity: challenges and opportunities. Nat Rev Immunol 2006;6:173–82. [PubMed: 16498448]
10. Kupferwasser LI, Yeaman MR, Nast CC, et al. Salicylic acid attenuates virulence in endovascularinfections by targeting global regulatory pathways in Staphylococcus aureus. J Clin Invest2003;112:222–33. [PubMed: 12865410]
11. Xiong YQ, Bayer AS, Yeaman MR, Van Wamel W, Manna AC, Cheung AL. Impacts of sarA andagr in Staphylococcus aureus strain Newman on fibronectin-binding protein A gene expression andfibronectin adherence capacity in vitro and in experimental infective endocarditis. Infect Immun2004;72:1832–6. [PubMed: 14977998]
Xiong et al. Page 8
J Infect Dis. Author manuscript; available in PMC 2010 February 23.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript

12. Fu Y, Ibrahim AS, Sheppard DC, et al. Candida albicans Als1p: an adhesin that is a downstreameffector of the EFG1 filamentation pathway. Mol Microbiol 2002;44:61–72. [PubMed: 11967069]
13. Yeaman MR, Sullam PM, Dazin PF, Norman DC, Bayer AS. Characterization of Staphylococcusaureus-platelet binding by quantitative flow cytometric analysis. J Infect Dis 1992;166:65–73.[PubMed: 1318911]
14. Bayer AS, Prasad R, Chandra J, et al. In vitro resistance of Staphylococcus aureus to thrombin-inducedplatelet microbicidal protein is associated with alterations in cytoplasmic membrane fluidity. InfectImmun 2000;68:3548–53. [PubMed: 10816510]
15. Dhawan VK, Bayer AS, Yeaman MR. In vitro resistance to thrombin-induced platelet microbicidalprotein is associated with enhanced progression and hematogenous dissemination in experimentalStaphylococcus aureus infective endocarditis. Infect Immun 1998;66:3476–9. [PubMed: 9632628]
16. Tenover FC, Moellering RC Jr. The rationale for revising the Clinical and Laboratory StandardsInstitute vancomycin minimal inhibitory concentration interpretive criteria for Staphylococcusaureus. Clin Infect Dis 2007;44:1208–15. [PubMed: 17407040]
17. Feil EJ, Li BC, Aanensen DM, Hanage WP, Spratt BG. eBURST: inferring patterns of evolutionarydescent among clusters of related bacterial genotypes from multilocus sequence typing data. JBacteriol 2004;186:1518–30. [PubMed: 14973027]
18. Melles DC, Gorkink RF, Boelens HA, et al. Natural population dynamics and expansion of pathogenicclones of Staphylococcus aureus. J Clin Invest 2004;114:1732–40. [PubMed: 15599398]
19. Fowler VG Jr, Nelson CL, McIntyre LM, et al. Potential associations between hematogenouscomplications and bacterial genotype in Staphylococcus aureus infection. J Infect Dis 2007;196:738–47. [PubMed: 17674317]
20. Harmsen D, Claus H, Witte W, et al. Typing of methicillin-resistant Staphylococcus aureus in auniversity hospital setting by using novel software for spa repeat determination and databasemanagement. J Clin Microbiol 2003;41:5442–8. [PubMed: 14662923]
21. Koreen L, Ramaswamy SV, Graviss EA, Naidich S, Musser JM, Kreiswirth BN. spa typing methodfor discriminating among Staphylococcus aureus isolates: implications for use of a single marker todetect genetic micro- and macrovariation. J Clin Microbiol 2004;42:792–9. [PubMed: 14766855]
22. Okuma K, Iwakawa K, Turnidge JD, et al. Dissemination of new methicillin-resistant Staphylococcusaureus clones in the community. J Clin Microbiol 2002;40:4289–94. [PubMed: 12409412]
23. Entenza JM, Foster TJ, Ni Eidhin D, Vaudaux P, Francioli P, Moreillon P. Contribution of clumpingfactor B to pathogenesis of experimental endocarditis due to Staphylococcus aureus. Infect Immun2000;68:5443–6. [PubMed: 10948180]
24. Xiong YQ, Willard J, Yeaman MR, Cheung AL, Bayer AS. Regulation of Staphylococcus aureusalpha-toxin gene (hla) expression by agr, sarA, and sae in vitro and in experimental infectiveendocarditis. J Infect Dis 2006;194:1267–75. [PubMed: 17041853]
25. Diep BA, Carleton HA, Chang RF, Sensabaugh GF, Perdreau-Remington F. Roles of 34 virulencegenes in the evolution of hospital-and community-associated strains of methicillin-resistantStaphylococcus aureus. J Infect Dis 2006;193:1495–503. [PubMed: 16652276]
26. Bayer AS, Coulter SN, Stover CK, Schwan WR. Impact of the high-affinity proline permease gene(putP) on the virulence of Staphylococcus aureus in experimental endocarditis. Infect Immun1999;67:740–4. [PubMed: 9916085]
27. Dhawan VK, Yeaman MR, Bayer AS. Influence of in vitro susceptibility phenotype against thrombin-induced platelet microbicidal protein on treatment and prophylaxis outcomes of experimentalStaphylococcus aureus endocarditis. J Infect Dis 1999;180:1561–8. [PubMed: 10515816]
28. Dhawan VK, Yeaman MR, Cheung AL, Kim E, Sullam PM, Bayer AS. Phenotypic resistance tothrombin-induced platelet microbicidal protein in vitro is correlated with enhanced virulence inexperimental endocarditis due to Staphylococcus aureus. Infect Immun 1997;65:3293–9. [PubMed:9234789]
29. Que YA, Haefliger JA, Piroth L, et al. Fibrinogen and fibronectin binding cooperate for valve infectionand invasion in Staphylococcus aureus experimental endocarditis. J Exp Med 2005;201:1627–35.[PubMed: 15897276]
30. Elasri MO, Thomas JR, Skinner RA, et al. Staphylococcus aureus collagen adhesin contributes to thepathogenesis of osteomyelitis. Bone 2002;30:275–80. [PubMed: 11792597]
Xiong et al. Page 9
J Infect Dis. Author manuscript; available in PMC 2010 February 23.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript

31. Galperine T, Neau D, Lina G, et al. Menstrual staphylococcal toxic shock is still a reality [in French].Presse Med 2003;32:1121–2. [PubMed: 12947741]
32. Hawkins C, Huang J, Jin N, Noskin GA, Zembower TR, Bolon M. Persistent Staphylococcusaureus bacteremia: an analysis of risk factors and outcomes. Arch Intern Med 2007;167:1861–7.[PubMed: 17893307]
33. Charles PG, Ward PB, Johnson PD, Howden BP, Grayson ML. Clinical features associated withbacteremia due to heterogeneous vancomycin-intermediate Staphylococcus aureus. Clin Infect Dis2004;38:448–51. [PubMed: 14727222]
34. Howden BP, Ward PB, Charles PG, et al. Treatment outcomes for serious infections caused bymethicillin-resistant Staphylococcus aureus with reduced vancomycin susceptibility. Clin Infect Dis2004;38:521–8. [PubMed: 14765345]
Xiong et al. Page 10
J Infect Dis. Author manuscript; available in PMC 2010 February 23.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript

Figure 1.Hypothesized life cycle of methicillin-resistant Staphylococcus aureus (SA) isolates that causepersistent bacteremia in the context of endovascular infection. AP, antimicrobial peptides; α-Tox, α-toxin; CM, cell membrane; FBG, fibrinogen; FBN, fibronectin; O-P, PMNopsonophagocytosis; PLT, platelets; PMN, neutrophils; SAK, staphylokinase.
Xiong et al. Page 11
J Infect Dis. Author manuscript; available in PMC 2010 February 23.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
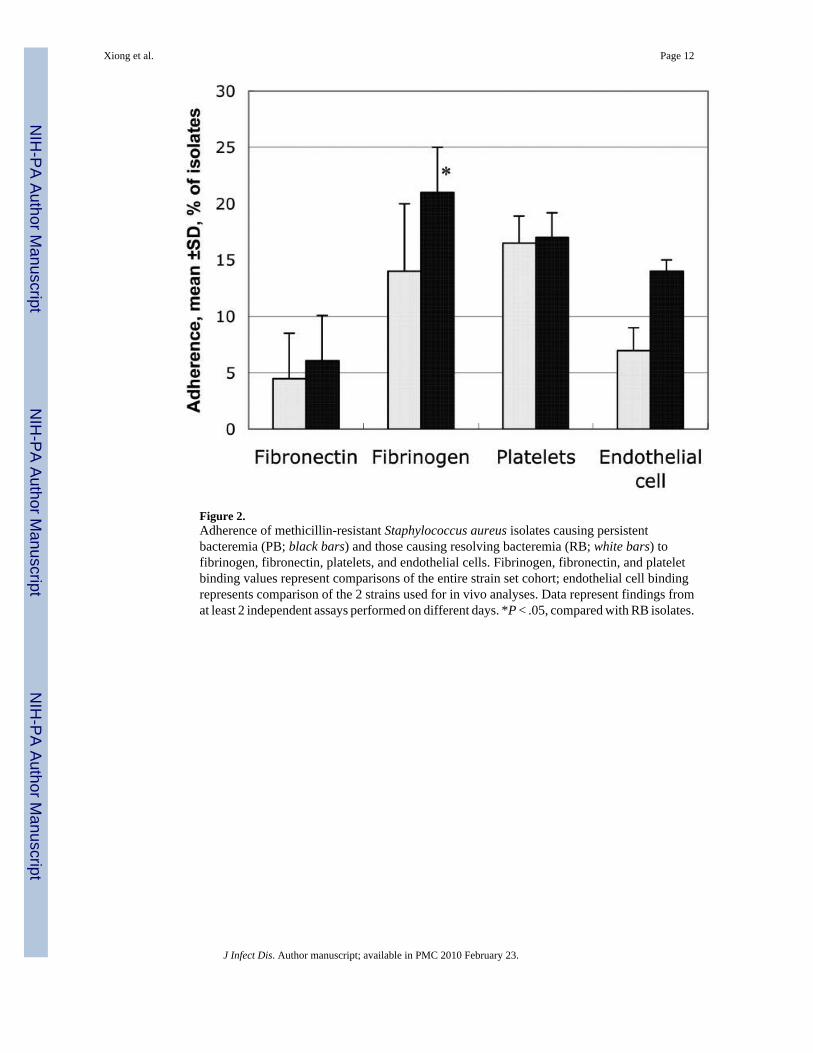
Figure 2.Adherence of methicillin-resistant Staphylococcus aureus isolates causing persistentbacteremia (PB; black bars) and those causing resolving bacteremia (RB; white bars) tofibrinogen, fibronectin, platelets, and endothelial cells. Fibrinogen, fibronectin, and plateletbinding values represent comparisons of the entire strain set cohort; endothelial cell bindingrepresents comparison of the 2 strains used for in vivo analyses. Data represent findings fromat least 2 independent assays performed on different days. *P < .05, compared with RB isolates.
Xiong et al. Page 12
J Infect Dis. Author manuscript; available in PMC 2010 February 23.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
Xiong et al. Page 13
Table 1
Findings of multiplex PCR profiling of select prototypic virulence genes in methicillin-resistant Staphylococcusaureus isolates that cause persistent bacteremia (PB).
Virulencegene(s) Description
Hypothesized rolein life cyclea
cna Adhesin, binds collagen Colonization
fnbA Adhesin, binds fibronectin Colonization
clfB Adhesin, binds fibrinogen Colonization
sdrD Adhesin, binds fibrinogen Colonization
sdrE Adhesin, binds fibrinogen Colonization
ica Polysaccharide intercellular adhesion (involved in biofilm formation) Colonization
pvl Exotoxin, putative virulence factor Dissemination
tst Exotoxin, toxic shock syndrome toxin-1 Dissemination
ssp Enzyme; serine protease Dissemination
cap5, cap8 Capsular polysaccharide– antiphagocytic factor Clearance
aA detailed account of the life cycle of PB isolates is presented in figure 1.
J Infect Dis. Author manuscript; available in PMC 2010 February 23.

NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
Xiong et al. Page 14
Table 2
Phenotypic and genotypic characteristics of methicillin-resistant Staphylococcus aureus strains used to inducepersistent bacteremia (PB) or resolving bacteremia (RB) in an infective endocarditis model.
Characteristic PB strain 420 RB strain 1507
Phenotypica
tPMP-1 susceptibilityb 61 ± 11 9 ± 3.5
Fibrinogen binding 15.2 ± 2.3 7.4 ± 1.6
Fibronectin binding 17.0 ± 0.2 1.3 ± 0.1
Platelet binding 16.6 ± 3.7 17.7 ± 2.2
Endothelial cell binding 12.7 ± 0.4 7.3 ± 0.2
Membrane fluidity, polarization index ± SD 0.319 ± 0.005 0.333 ± 0.016
Genotypic
MLST CC30 CC8
spa type 16 7
SCCmec type II IV
agr type III I
pvl Absent Absent
cna Present Absent
cap type 8 5
tst Present Absent
NOTE. MLST, multilocus sequence type; SCCmec, staphylococcal cassette chromosome mec.
aData are percentage of isolates (±SD), unless otherwise indicated.
bData are percentage of isolates that survived exposure to 1 μg/mL of tPMP-1.
J Infect Dis. Author manuscript; available in PMC 2010 February 23.

NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
Xiong et al. Page 15
Table 3
Phenotypic and genotypic characteristics of methicillin-resistant Staphylococcus aureus strains used to inducepersistent bacteremia (PB) or resolving bacteremia (RB).
CharacteristicPB isolates
(n = 21)RB isolates
(n = 18) P
Phenotype
hNP-1 susceptibilitya 52 ± 5 36 ± 10 <.05
Fibrinogen bindingb 21 ± 4 14 ± 6 <.05
Membrane fluidity, polarization index 0.276 (more fluid) 0.290 (less fluid) .11
Genotype
MLST CC30 19 (90) 8 (44) <.05
spa16 12 (57) 7 (39) .16
SCCmec type II 20 (95) 13 (72) <.05
agr type III 17 (81) 8 (44) <.05
cna 18 (85) 10 (56) .07
cap5 3 (14) 11 (61) <.05
cap8 18 (86) 7 (39) <.05
pvl 0 (0) 0 (0) NS
tst 18 (86) 8 (44) <.05
NOTE. Data are no. (%) of isolates, unless otherwise indicated. The following genes were also tested, but there were no significant differences betweenPB and RB isolates: bsaA1, sea-e, sej, sem, sei, seg, seo, sen, sdrC-E, bbp, ebpS, icaA, efb, sbl, ssp, clfB, splB, fnbA, lukDE, lukM, and EDIN. CC30,clonal complex 30; MLST, multilocus sequence type; NS, not signficant; SCCmec, staphylococcal cassette chromosome mec.
aData are percentage of isolates (±SD) that survived exposure to 40 μg/mL of hNP-1.
bData are percentage of isolates (±SD) that bound to fibrinogen.
J Infect Dis. Author manuscript; available in PMC 2010 February 23.

NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
Xiong et al. Page 16
Tabl
e 4
Den
sitie
s of S
taph
yloc
occu
s aur
eus s
train
s tha
t ind
uced
per
sist
ent b
acte
rem
ia (P
B) o
r res
olvi
ng b
acte
rem
ia (R
B) i
n ta
rget
tiss
ues o
f an
infe
ctiv
e end
ocar
ditis
mod
el, w
ith o
r with
out v
anco
myc
in th
erap
y.
Gro
up, i
sola
tety
peA
nim
als,
no.
Inoc
ulum
, cfu
Rec
eive
d15
mg/
kgva
ncom
ycin
MR
SA d
ensi
ty, l
og10
cfu
/g o
f spe
cim
en, m
ean
± SD
Veg
etat
ions
Kid
neys
Sple
en
Gro
up 1
PB
710
4N
o7.
21 ±
0.7
25.
31 ±
0.1
14.
76 ±
0.5
1
R
B6
104
No
6.28
± 1
.18
5.41
± 1
.83
4.88
± 1
.68
Gro
up 2
PB
910
5N
o7.
46 ±
1.5
35.
23 ±
0.9
14.
83 ±
1.5
4
R
B9
105
No
7.61
± 1
.34
5.53
± 0
.73
5.12
± 0
.50
Gro
up 3
PB
810
5Y
es4.
23 ±
1.4
1a2.
19 ±
0.2
3a2.
38 ±
0.5
0a
R
B7
105
Yes
2.31
± 0
.92b
1.81
± 0
.63c
1.96
± 0
.38c
NO
TE
. The
PB
isol
ate
was
stra
in 4
20, a
nd th
e R
B is
olat
e w
as st
rain
150
7.
a P <
.005
, com
pare
d w
ith th
e de
nsity
in c
orre
spon
ding
spec
imen
s fro
m P
B c
ontro
ls fr
om g
roup
2.
b P <
.01,
com
pare
d w
ith th
e de
nsity
in c
orre
spon
ding
spec
imen
s fro
m R
B c
ontro
ls fr
om g
roup
2.
c P <
.005
, com
pare
d w
ith th
e de
nsity
in c
orre
spon
ding
spec
imen
s fro
m R
B c
ontro
ls in
gro
up 2
.
J Infect Dis. Author manuscript; available in PMC 2010 February 23.




















