
Perinatal changes in choroidal 15-hydroxyprostaglandin dehydrogenase: implications for prostaglandin...
11
Developmental Brain Research 121 (2000) 145–155 www.elsevier.com / locate / bres Research report Perinatal changes in choroidal 15-hydroxyprostaglandin dehydrogenase: implications for prostaglandin removal from brain a,c,d c,d,e, b f * Nancy Krunic , S. Lee Adamson , Cameron Ackerley , Richard T. Okita , a,d Flavio Coceani a Integrative Biology Programme, The Hospital for Sick Children, Toronto, Ontario, Canada b Department of Pathology, The Hospital for Sick Children, Toronto, Ontario, Canada c Program in Fetal Health and Development, Samuel Lunenfeld Research Institute, Rm 138P , Mount Sinai Hospital, 600 University Ave., Toronto, Ontario, Canada M5G 1X5 d Department of Physiology, University of Toronto, Toronto, Ontario, Canada e Department of Obstetrics and Gynecology, University of Toronto, Toronto, Ontario, Canada f College of Pharmacy, Washington State University, Pullman, WA, USA Accepted 14 March 2000 Abstract We have previously shown in the sheep fetus at 0.7 and 0.9 gestation that the choroid plexus, unlike brain parenchyma, catabolizes prostaglandins (PGs). Peculiarly, in the choroid plexus, PGE catabolism persists throughout the neonatal period to abate in the adult, 2 while PGF catabolism abates shortly after birth. To explain this differential behavior and elucidate the function of catabolic enzymes, 2a we examined the cellular location and activity of the rate-limiting enzyme for PGE and PGF catabolism, 15-hydroxyprostaglandin 2 2a dehydrogenase (15-PGDH). Immunofluorescence histochemistry and immunogold electronmicroscopy revealed abundant 15-PGDH expression in the epithelial cytosol close to the brush-border membrane at 0.7 and 0.9 gestation. In contrast, at 5 and 15 days postnatal, 15-PGDH was found throughout the cytosol of stromal fibroblasts. No staining was observed at either location in pregnant adults. PGF 2a catabolism was minimal in the total homogenate and 100 0003g supernatant of the fetal choroid plexus at 0.7 and 0.9 gestation, while PGE catabolism was evident at 0.7 gestation only. In contrast, both PGs were catabolized in minced specimens at either age. In 2 conclusion, our study shows immunoreactive 15-PGDH in the choroid plexus from fetal and neonatal, but not pregnant adult, sheep. Results suggest that PGE catabolism is not as critically dependent as that of PGF on tissue integrity and 15-PGDH location. Given the 2 2a key role being assigned to the choroid plexus in PG removal from brain, we speculate that persistence of PGE catabolism into the early 2 postnatal period protects against central respiratory depression caused by the compound during this susceptible stage of development. 2000 Elsevier Science B.V. All rights reserved. Theme: Neurotransmitters, modulators, transporters, and receptors Topic: Uptake and transporters Keywords: Fetus; Newborn; Adult; Choroid plexus; Development; Sheep; Prostaglandin catabolism; Birth 1. Introduction ever, is an exception in that it shows 15-PGDH activity at an early stage of gestation but not in the fetus at term or at 1 NAD -dependent 15-hydroxyprostaglandin dehydrogen- any age after birth [2,28,30,32]. Hence, transport across the ase (15-PGDH), the rate-limiting enzyme for catabolism of choroidal, and possibly extrachoroidal, components of the primary prostaglandins (PGs) such as PGE and PGF , is blood–brain barrier is the primary means for removing 2 2a widely distributed throughout the body [2]. Brain, how- prostaglandins from brain [5,13]. In previous investigations, we have found that the choroid plexus from perinatal sheep is endowed with *Corresponding author. Tel.: 11-416-586-8377; fax: 11-416-586- 15-PGDH activity, and that the enzyme operates in parallel 5993. E-mail address: [email protected] (S. . Adamson) with the prostaglandin carrier system [21,22]. This differs 0165-3806 / 00 / $ – see front matter 2000 Elsevier Science B.V. All rights reserved. PII: S0165-3806(00)00033-X
-
Upload
independent -
Category
Documents
-
view
0 -
download
0
Transcript of Perinatal changes in choroidal 15-hydroxyprostaglandin dehydrogenase: implications for prostaglandin...

Developmental Brain Research 121 (2000) 145–155www.elsevier.com/ locate /bres
Research report
Perinatal changes in choroidal 15-hydroxyprostaglandin dehydrogenase:implications for prostaglandin removal from brain
a,c,d c,d,e , b f*Nancy Krunic , S. Lee Adamson , Cameron Ackerley , Richard T. Okita ,a,dFlavio Coceani
aIntegrative Biology Programme, The Hospital for Sick Children, Toronto, Ontario, CanadabDepartment of Pathology, The Hospital for Sick Children, Toronto, Ontario, Canada
cProgram in Fetal Health and Development, Samuel Lunenfeld Research Institute, Rm 138P, Mount Sinai Hospital, 600 University Ave., Toronto,Ontario, Canada M5G 1X5
dDepartment of Physiology, University of Toronto, Toronto, Ontario, CanadaeDepartment of Obstetrics and Gynecology, University of Toronto, Toronto, Ontario, Canada
fCollege of Pharmacy, Washington State University, Pullman, WA, USA
Accepted 14 March 2000
Abstract
We have previously shown in the sheep fetus at 0.7 and 0.9 gestation that the choroid plexus, unlike brain parenchyma, catabolizesprostaglandins (PGs). Peculiarly, in the choroid plexus, PGE catabolism persists throughout the neonatal period to abate in the adult,2
while PGF catabolism abates shortly after birth. To explain this differential behavior and elucidate the function of catabolic enzymes,2a
we examined the cellular location and activity of the rate-limiting enzyme for PGE and PGF catabolism, 15-hydroxyprostaglandin2 2a
dehydrogenase (15-PGDH). Immunofluorescence histochemistry and immunogold electronmicroscopy revealed abundant 15-PGDHexpression in the epithelial cytosol close to the brush-border membrane at 0.7 and 0.9 gestation. In contrast, at 5 and 15 days postnatal,15-PGDH was found throughout the cytosol of stromal fibroblasts. No staining was observed at either location in pregnant adults. PGF2a
catabolism was minimal in the total homogenate and 100 0003g supernatant of the fetal choroid plexus at 0.7 and 0.9 gestation, whilePGE catabolism was evident at 0.7 gestation only. In contrast, both PGs were catabolized in minced specimens at either age. In2
conclusion, our study shows immunoreactive 15-PGDH in the choroid plexus from fetal and neonatal, but not pregnant adult, sheep.Results suggest that PGE catabolism is not as critically dependent as that of PGF on tissue integrity and 15-PGDH location. Given the2 2a
key role being assigned to the choroid plexus in PG removal from brain, we speculate that persistence of PGE catabolism into the early2
postnatal period protects against central respiratory depression caused by the compound during this susceptible stage of development. 2000 Elsevier Science B.V. All rights reserved.
Theme: Neurotransmitters, modulators, transporters, and receptors
Topic: Uptake and transporters
Keywords: Fetus; Newborn; Adult; Choroid plexus; Development; Sheep; Prostaglandin catabolism; Birth
1. Introduction ever, is an exception in that it shows 15-PGDH activity atan early stage of gestation but not in the fetus at term or at
1NAD -dependent 15-hydroxyprostaglandin dehydrogen- any age after birth [2,28,30,32]. Hence, transport across thease (15-PGDH), the rate-limiting enzyme for catabolism of choroidal, and possibly extrachoroidal, components of theprimary prostaglandins (PGs) such as PGE and PGF , is blood–brain barrier is the primary means for removing2 2a
widely distributed throughout the body [2]. Brain, how- prostaglandins from brain [5,13].In previous investigations, we have found that the
choroid plexus from perinatal sheep is endowed with*Corresponding author. Tel.: 11-416-586-8377; fax: 11-416-586-15-PGDH activity, and that the enzyme operates in parallel5993.
E-mail address: [email protected] (S. . Adamson) with the prostaglandin carrier system [21,22]. This differs
0165-3806/00/$ – see front matter 2000 Elsevier Science B.V. All rights reserved.PI I : S0165-3806( 00 )00033-X

146 N. Krunic et al. / Developmental Brain Research 121 (2000) 145 –155
from other organs, such as lung and kidney, where uptake localizing with purified 15-PGDH [30]. In addition, theis rate-limiting for catabolism [3,6]. The latter finding competence of the antibody was confirmed by immuno-implies that the two mechanisms operate in sequence [3]. histochemical analysis of lung samples from the sameIn addition, PGE and PGF differed in their suscep- animals. Lung, unlike brain, is known to be rich in 15-2 2a
tibility to degradation via choroidal 15-PGDH [22]. While PGDH, its expression being particularly high in epithelialPGE catabolism was equally prominent in the fetal and cells [30].2
neonatal choroid plexus, PGF catabolism abated after PGE , PGF , PGA , 15-keto-PGE (15K-PGE ), 13-2a 2 2a 2 2 2
birth [21]. This special pattern may be functionally im- 14-dihydro-15-keto-PGE (15KD-PGE ), 15-keto-PGF2 2 2a
portant since PGE is a potent inhibitor of the central (15K-PGF ), 13,14-dihydro-15-keto-PGF (15KD-2 2a 2a
respiratory drive throughout the perinatal period [14,18]. PGF ), and 13,14-dihydro-PGF (D-PGF ), were a gift2a 2a 2a
Indeed, in support of this concept, brain levels of PGE of The Upjohn Company (Kalamazoo, MI).23fall abruptly at birth and, accordingly, any mechanism [5,6,8,11,12,14,15(N) H]PGE (specific activity, 164 Ci /2
3which keeps these levels low may contribute to the mmol) and [5,6,8,9,11,12,14,15(N) H]PGF (specific ac-2a
maintenance of continuous breathing [1,16,38]. tivity, 180 Ci /mmol) were purchased from DuPont (Bos-The aim of our study was to determine the cellular ton, MA). Their purity on thin-layer radiochromatography
location and activity of 15-PGDH in the sheep choroid was 95 and 96%, respectively. Fluorescein isothiocyanateplexus at different stages of development. For this purpose, (FITC)-labelled anti-guinea pig IgG, nicotinamide adenine
1a combination of immunological and biochemical tech- dinucleotide (NAD ), nicotinamide adenine dinucleotide1niques was used to examine specimens from the fetus, both phosphate (NADP ), bovine serum albumin (BSA, Cohn
preterm and term, the neonate, and the pregnant adult. With fraction V), EDTA, the stabilizing agent, glycerol, and thethis approach, we sought to gain insight into the functional prostaglandin synthase inhibitor, indomethacin, were pur-organization of the enzyme and the reason for the differen- chased from Sigma (St. Louis, MO). Colloidal gold-la-tial catabolism of PGE versus PGF in the early postnat- belled protein A (particle size, 5 and 15 nm) was pur-2 2a
al period. A preliminary account of this work has appeared chased from Amersham (Arlington Heights, IL). Thein abstract form [20]. protease inhibitors, leupeptin and chymostatin, and the
reducing agent, 1,4-dithiothreitol (DTT), were purchasedfrom Boehringer Mannheim (Mannheim, Germany).
2. Materials and methods Chemicals were of analytical grade purity and buffers wereprepared with ultrapure water.
2.1. Animals
Experiments were performed on fetal sheep at 0.7 2.3. General proceduresgestation (mean age, 102 days; range, 101–103; term about145 days) and 0.9 gestation (mean age, 136 days; range For light microscopic and biochemical analyses (see134–138), lambs at 5 (range, 5–6) and 15 (range, 15–18) below), choroid plexuses were collected from all cerebraldays, and pregnant ewes (.1 year). Pregnant ewes were ventricles and placed in ice-cold, pre-equilibrated Krebs–anesthetized with intravenous sodium pentobarbital (Nem- Henseleit solution (118 mM NaCl, 4.7 mM KCl, 2.5 mMbutal, 30 mg/kg, Abbott Laboratories, Montreal, Que.) and CaCl ?2H O, 0.9 mM MgSO ?7H O, 1.0 mM KH PO ,2 2 4 2 2 4
ventilated with a mixture of methoxyflurane, nitrous oxide 11 mM glucose, 24.9 mM NaHCO ) for transportation to3
and air. Fetuses were delivered by cesarean section as the laboratory. Tissues were either processed for incuba-previously described [11] and were killed by exsanguina- tion or were flash frozen in OCT and stored at 2708C fortion. Ewes and lambs were killed instead with excess cryosectioning. For electron microscopy (see below), spec-sodium pentobarbital (Euthanyl, 100 mg/kg, M.T.C. Phar- imens were placed in 4% paraformaldehyde immediatelymaceuticals, Cambridge, Ont.). Surgery and experiments after collection. The fixed tissues were transferred to awere approved by our institutional ethics committee and Petri dish and cut into 0.5-mm pieces. The fixative waswere conducted in accordance with the guidelines of the then changed and treatment was continued for 4 h at roomCanadian Council of Animal Care. temperature. Afterwards, tissues were rinsed with 0.1 M
phosphate-buffered saline (PBS, pH 7.4) and stored at 48C2.2. Materials in PBS containing sodium azide (0.02 M). Ultra-thin
cryosections (90 nm) were prepared with a Reichert JungPolyclonal antibodies were raised in guinea pigs against Ultracut E microtome (Vienna, Austria) equipped with a
purified lung 15-PGDH from pregnant rabbits, and were FC-4 cryochamber and, for immunolabelling, were trans-affinity-purified according to a published method [30]. ferred to glow-discharged carbon-Formvar nickel gridsSpecificity of the antibody was verified by demonstrating using a loop of molten sucrose.in the immunoblot of the high-speed supernatant from In some of the term fetuses, lung specimens werepregnant rabbit lung a single cross-reaction product co- collected under the same conditions and were used as a

N. Krunic et al. / Developmental Brain Research 121 (2000) 145 –155 147
positive control for histochemical [30] and biochemical 1:20) or the 100 0003g supernatant (ratio, 1:10) was[31] analyses of 15-PGDH. prepared for incubation. In certain experiments with the
total homogenate, the homogenization buffer was sup-2.4. Morphological analysis plemented with one of the following compounds: in-
1domethacin (2.8 or 28 mM), NAD (5 mM), DTT (12.4.1. Light microscopy immunofluorescence mM), EDTA (1 mM), glycerol (20% by volume), leupep-histochemistry tin (1 or 10 mM), or chymostatin (10 or 100 mM). To
Cryosections (5 mm) were fixed in 2% paraformal- prepare the high-speed supernatant, the homogenate wasdehyde for 30 min, washed thoroughly with BSA/glycine centrifuged first at 10 0003g (15 min, 48C) and the(0.5%/0.15%) in PBS and blocked with BSA (0.5% in resulting supernatant was re-centrifuged at 100 0003g (1PBS). The sections were then incubated with a 1:75 h, 48C). Protein was measured by a standard proceduredilution (in 5% milk stock) of the primary antibody for 60 [24] and its average concentration was 1.34 and 1.06min. Following a thorough wash in BSA (0.5% in PBS), mg/ml for samples of choroid plexus and lung, respective-the sections were incubated with a 1:75 dilution (in PBS) ly.of FITC-conjugated anti-guinea pig IgG for 60 min in the The total homogenate (choroid plexus, lung) anddark. Afterwards, they were washed with PBS and 100 0003g supernatant fraction (choroid plexus) were
3mounted in Elvanol. As a negative control, incubations incubated (60 min, 378C) with either [ H]PGF or2a3 1were carried out with 0.5% BSA alone or with serum from [ H]PGE (0.25 mCi/ml) and the cofactor NAD (5 mM)2
a non-immunized guinea pig in place of the primary in ambient air. The incubation volume was 1 ml. In thoseantibody. The positive control consisted of lung tissue instances where the homogenization buffer had already
1from the term fetus which had been processed in the same been supplemented with NAD (see above), no cofactormanner. Immunohistochemical staining was carried out in was added to the incubation proper. Incubation wasa humid chamber, and images were acquired on a confocal terminated by adding 1 M citric acid and the incubate wasmicroscope (Leica, Model DM 1RB). then extracted three times with 2 vol ethyl acetate.
Preliminary work did not reveal any non-enzymatic break-2.4.2. Electron microscopy immunogold histochemistry down of PGE metabolites. Consequently, unlike previous2
Immunolabelling was carried out as described above, studies in the intact tissue [22], homogenized tissues wereexcept for the secondary antibody, which consisted of not treated with alkali prior to extraction to ensurecolloid gold-conjugated protein A (1:20 in PBS). After- complete conversion of 15KD-PGE to the bicyclo-deriva-2
wards, sections were washed thoroughly with PBS and tive. Following extraction, organic phases were washeddouble-distilled water in sequence and then incubated in twice with distilled water, pooled, dried under a stream ofneutral uranyl acetate for 10 min. Following a final wash nitrogen, and the residue was re-suspended in a smallwith double-distilled water, sections were stabilized in volume (20 ml) of ethyl acetate. In certain cases, incuba-0.2% uranyl acetate in methyl cellulose for 10 min. tion was terminated by rapid freezing in a solid CO –2
Specimens were viewed and photographed in a JEOL acetone bath and the frozen samples were stored at 2208C.model 1200 ExII transmission electron microscope (Pea- When required, samples were thawed and processed asbody, MA). Particles were counted (25 fields /cell) at a described above. Tissue extracts were spotted on silica gelmagnification of 25 000 using the morphometric program G60 plates alongside reference standards and developedNIH Image. For epithelial cells, fields were randomly once at room temperature in either diethyl ether /acetic
3chosen within a 0.3-mm band at the apical surface. For acid (99:1, by vol, [ H]PGE incubation) or chloroform/2
stromal cells, fields were randomly chosen from perinu- methanol /acetic acid /water (90:9:1:0.65, by vol,3clear and peripheral regions of the cytosol. Regardless of [ H]PGF incubation). Distribution of radioactivity on the2a
cell type and age, the number of cells sampled in each plates was ascertained by thin layer radiochromatographyspecimen ranged between 15 and 24. Gold particle density (TLC) [21,22].
2was quantified as the number of particles per mm and wasexpressed as the average for three animals in each age 2.5.2. Minced tissuegroup. Choroid plexus samples collected from fetuses (see
above) were kept in Krebs–Henseleit medium and equili-2.5. 15-PGDH assays brated for 1 h on ice with a gas mixture (2.5% O /5%2
CO in N ) approximating fetal oxygen levels in vivo2 2
2.5.1. Homogenized tissue (P(O ), 13–21 mmHg) [21]. Using a razor blade, they2
Specimens of fetal choroid plexus and lung were were then chopped into small pieces over a chilled glasstransferred from chilled Krebs–Henseleit medium to phos- plate and the resulting tissue mince was placed in Krebs–phate buffer (50 mM KH PO /NaOH, pH 7.4) and Henseleit medium (250 ml) to be centrifuged at 20003g2 4
homogenized (Polytron) on ice. The weight:volume ratio for 5 min. A sealed tube which had been flushed with thevaried depending on whether the total homogenate (ratio, equilibration gas mixture was used for centrifugation.

148 N. Krunic et al. / Developmental Brain Research 121 (2000) 145 –155
Afterwards, fluid was separated from tissue and the two medium supplemented with the 100 0003g supernatantfractions were incubated separately (60 min, 378C) with fraction (10%, by vol) from the choroid plexus of the other
3 3either [ H]PGE or [ H]PGF (0.25 mCi/ml). Following lateral ventricle (final incubation vol, 2 ml). Tissues were2 2a
incubation, samples were processed and analyzed by TLC dried overnight (608C), homogenized (Polytron) in phos-as described above for homogenized tissues. phate buffer and processed for TLC analysis [21].
In all cases, catabolism is expressed as the percentage of2.5.3. Intact tissue total radioactivity migrating with metabolites of PGE or2
Some control incubations were carried out in the intact PGF (PGM). Only metabolite peaks exceeding back-2a
choroid plexus. In this case, the choroid plexus from one ground (i.e., .8% of total radioactivity) were counted.lateral ventricle was equilibrated in Krebs–Henseleit Any radioactivity corresponding to non-enzymatic prod-medium as described above, and was incubated (60 min, ucts of the parent compound (e.g., PGA ) was not consid-2
3378C) with [ H]PGF (0.25 mCi/ml) in Krebs–Henseleit ered in this computation.2a
Fig. 1. Sheep choroid plexus. 15-PGDH immunofluorescence in (a) fetus, 0.7 gestation; (b) fetus, 0.9 gestation; (c) lamb, 5 days; (d) lamb, 15 days.Epithelium and stroma are indicated, respectively, with an arrow and asterisk. Note that the epithelium is not evident in (c) and (d). Scale bar is 25 mm.
N. Krunic et al. / Developmental Brain Research 121 (2000) 145 –155 149
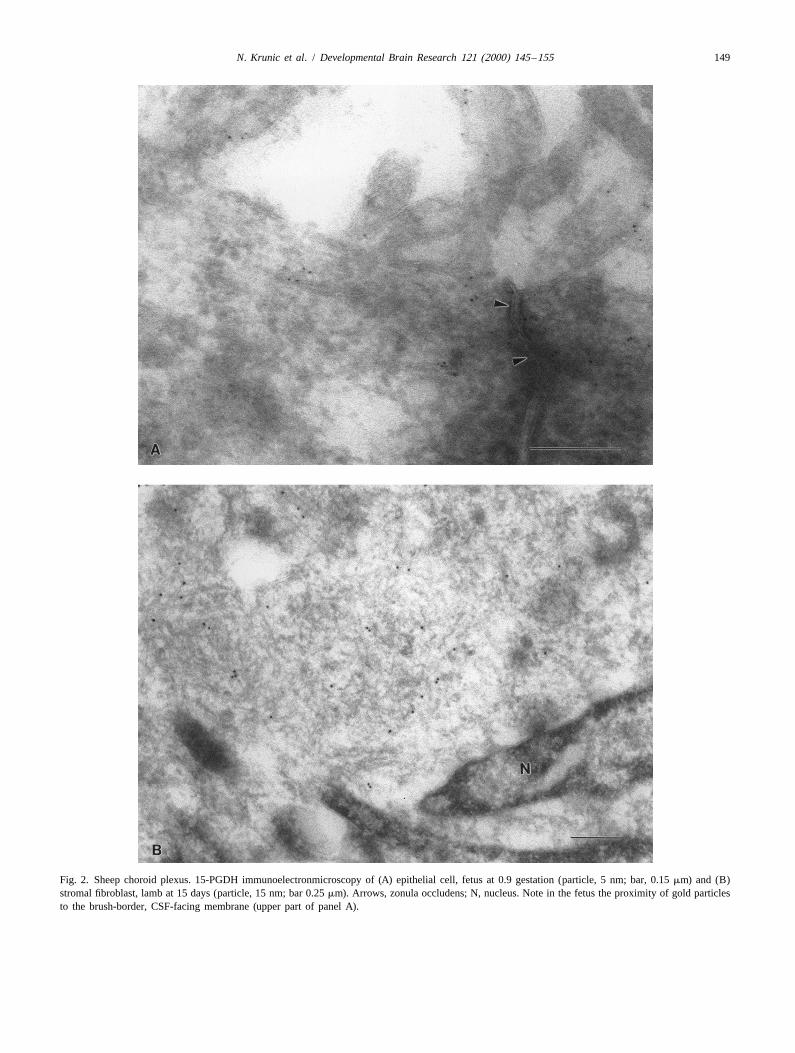
Fig. 2. Sheep choroid plexus. 15-PGDH immunoelectronmicroscopy of (A) epithelial cell, fetus at 0.9 gestation (particle, 5 nm; bar, 0.15 mm) and (B)stromal fibroblast, lamb at 15 days (particle, 15 nm; bar 0.25 mm). Arrows, zonula occludens; N, nucleus. Note in the fetus the proximity of gold particlesto the brush-border, CSF-facing membrane (upper part of panel A).

150 N. Krunic et al. / Developmental Brain Research 121 (2000) 145 –155
Fig. 3. Age-related change in gold particle density in choroid plexus epithelial cells (left) and stromal fibroblasts (right). Values are the means6S.E.M. forthree animals in each age group. For details, see Section 2. Different letters denote a significant difference among ages. Note that no gold particle weredetected in the epithelium from lambs at 15 days of age.
3 3Fig. 4. Sheep choroid plexus. Catabolism of [ H]PGE (open bar) and [ H]PGF (stipled bar) (0.25 mCi/ml for both) by the total homogenate and2 2a1100 0003g supernatant fraction from the fetus at 0.7 (left) and 0.9 (right) gestation. Incubations lasted 60 min and NAD (5 mM) was used as cofactor.
Values are the means6S.E.M. for the number of animals given above each column. A significant difference between the two prostaglandins whenincubated in the same tissue fraction is indicated with an asterisk, while a difference between the two tissue fractions (identical age) when incubated withthe same prostaglandin is indicated with a spade. In this and the following figures, catabolism is expressed as the percent of total radioactivity migratingwith metabolites of either prostaglandin (for details, see Section 2).

N. Krunic et al. / Developmental Brain Research 121 (2000) 145 –155 151
2.6. Analysis of data particularly its apical portion, and little of the enzyme wasfound in the stroma (Fig. 1a,b). Conversely, in the lamb at
All data are given as the mean6S.E.M. where n is the 5 and 15 days, 15-PGDH was virtually absent in thenumber of animals. Comparisons were made with either a epithelium and became abundant in the stroma (Fig. 1c,d).one-way or a two-way analysis of variance (ANOVA) Neither site, on the other hand, showed staining exceedingfollowed by the Student–Newman–Keuls test for indi- background levels in the pregnant adult (data not shown).vidual comparisons. Differences were considered signifi-cant when P,0.05. 3.1.2. Immunogold electronmicroscopy
The distribution of immunogold particles accorded withhistochemical findings. In the fetus, particles were found
3. Results predominantly near the brush-border, CSF-facing mem-brane of choroid plexus epithelial cells (Fig. 2A). This
3.1. 15-PGDH localization epithelial staining was cytosolic and was not linked to anyparticular organelle (Fig. 2A). In the lamb, immuno-
3.1.1. Immunofluorescence histochemistry reactivity was instead distributed diffusely throughout theImmunoreactive 15-PGDH was abundant in the choroid cytosol of stromal fibroblasts (Fig. 2B). Fig. 3 summarizes
plexus of both fetus and lamb. Its location, however, these findings and documents the reciprocal expression ofdiffered between the two ages. In the fetus at either 0.7 or gold particles in epithelial versus stromal cells, depending0.9 gestation, 15-PGDH was expressed in the epithelium, on the age of the animal.
3Fig. 5. Sheep fetus at 0.9 gestation. Catabolism of [ H]PGF (0.25 mCi/ml) by the total homogenate of the choroid plexus (stippled bar) and lung (open2a1bar). Incubations lasted 60 min and NAD (5 mM) was used as cofactor. One of the following compounds was added to the buffer prior to tissue
1homogenization: indomethacin (indo), NAD , DTT, EDTA, glycerol, leupeptin (leup), or chymostatin (chym). In those instances in which the1homogenization buffer was supplemented with NAD , the same cofactor was omitted from the incubation proper. Each bar represents the mean6S.E.M.
for three animals. With either tissue, different letters (bold, lung; italics, choroid plexus) denote a significant difference among treatment groups. Asignificant difference between lung and choroid plexus is indicated with an asterisk.

152 N. Krunic et al. / Developmental Brain Research 121 (2000) 145 –155
3.2. 15-PGDH activity
3.2.1. Tissue fractionsWhen using the total homogenate of the choroid plexus,
15-PGDH-mediated catabolic activity varied depending onthe prostaglandin and gestational age. At 0.7 gestation,
3catabolism of [ H]PGE was marked and exceeded that of23[ H]PGF , which was barely measurable (Fig. 4).2a3[ H]PGE catabolism also tended to be greater than23[ H]PGF catabolism at 0.9 gestation; however, this2a
difference did not reach significance (Fig. 4). 15-PGDHactivity was also present in the 100 0003g supernatantfrom the fetal choroid plexus at 0.7 gestation and, as in the
3total homogenate, it was more effective against [ H]PGE23than [ H]PGF (Fig. 4). However, by 0.9 gestation,2a
catabolism of both prostaglandins in the supernatantbecame marginal (Fig. 4).
Experiments were carried out to determine whether thepeculiar loss of catabolic activity upon tissue homogeniza-tion had a methodological cause. Fetal lung was used as areference since it is known to express 15-PGDH activity
3[21,31]. As shown in Fig. 5, [ H]PGF catabolism was2a
not restored by adding indomethacin to the buffer prior tochoroid plexus homogenization. This argues against satura-tion of 15-PGDH by endogenously formed prostaglandins.Similarly ineffective were stabilizing agents (DTT, gly-cerol) and protease inhibitors (EDTA, leupeptin, chymos-
Fig. 6. Sheep fetus at 0.9 gestation. Thin-layer radiochromatogram of the1 1tatin) (Fig. 5). Substitution of NAD with NADP during product of incubation of the intact lateral ventricle choroid plexus3incubation was also unable to restore catabolic activity incubated with [ H]PGF (0.25 mCi/ml) in the absence (top) and2a
presence (bottom) of the 100 0003g supernatant fraction from the(data not shown). Furthermore, catabolism was still margi-1 contralateral choroid plexus. For details, see Section 2. Markers are (i)nal when NAD was added to the buffer prior to homoge-
PGF , (ii) D-PGF , (iii) 15K-PGF , (iv) 15KD-PGF , and (v) 15KD-2a 2a 2a 2anization rather than at the start of the incubation (Fig. 5).PGE . Origin is at arrow. Note that in the lower panel, unlike the upper2In contrast, 15-PGDH-mediated catabolism was high in the panel, some radioactivity migrated with unreacted PGF .2a
lung homogenate and this activity remained unabated withall but the treatments with indomethacin and chymostatin
tissue homogenization results from the lack of structural(Fig. 5). This reduction was statistically significant withintegrity, incubations were carried out in minced speci-high doses of chymostatin (Fig. 5). While the inhibitorymens of the choroid plexus from the 0.7 and 0.9 gestationtrend observed with indomethacin accords with earlier datafetus. Contrary to homogenization, mincing did not inter-[36], no precedent is available for the chymostatin effect.fere with 15-PGDH-mediated catabolism. As shown in Fig.Separate controls were also done to determine whether7, catabolism was consistently high, regardless of gesta-an inhibitor of 15-PGDH was being released duringtional age or the prostaglandin used in the incubation.homogenization. This possibility is supported by a report
in the literature [26]. For this purpose, the intact choroidplexus from one lateral ventricle was incubated with
3 4. Discussion[ H]PGF after adding an aliquot of the 100 0003g2a
supernatant (10% by vol) of the choroid plexus from the3 Using an immunological approach, the present study hasother lateral ventricle. [ H]PGF was extensively catabol-2a
demonstrated 15-PGDH in the choroid plexus of theized in this preparation, its overall magnitude beingperinatal sheep. In addition, it has shown that the enzymecomparable to that observed in the intact choroid plexus
3 relocates from epithelial cells to stromal fibroblasts duringincubated with [ H]PGF alone (Fig. 6). Nevertheless, the2a
the transitional period at birth. 15-PGDH immunoreactivitysupernatant may cause some inhibition, as indicated by thewas cytosolic in both cell types, which is consistent withappearance of a small peak migrating with unreacted
3 the expression pattern of the enzyme elsewhere [10,19,30].[ H]PGF (Fig. 6, lower panel).2a
However, in stromal fibroblasts 15-PGDH was distributeddiffusely throughout the cytosol, while in the epithelium it3.2.2. Tissue mincewas clustered in proximity to the brush-border, CSF-To determine whether the loss in catabolic activity after

N. Krunic et al. / Developmental Brain Research 121 (2000) 145 –155 153
3 3Fig. 7. Sheep choroid plexus. Catabolism of [ H]PGE and [ H]PGF by minced preparations from the fetus at 0.7 and 0.9 gestation. Tissue (open bar);2 2a
supernatant (stipled bar). Values are the mean6S.E.M. for the number of animals given above each column. A significant difference between tissue andsupernatant is indicated with an asterisk.
facing, membrane. Contrary to the perinatal animal, 15- sion and would, consequently, appear to be good candi-PGDH immunoreactivity was not detectable in the choroid dates for this role. Specifically, one may speculate that theplexus of the pregnant adult. The latter finding accords normal rise in plasma cortisol during the last third ofwith the absence of catabolic activity in the choroid plexus gestation [23] upregulates 15-PGDH in the choroid plexusof non-pregnant rabbits and pregnant and non-pregnant epithelium. This upregulation could be mediated by spe-sheep [4,4,21,22]. The distribution pattern of 15-PGDH cific receptors in the epithelium [17] or by a glucocorticoidwithin epithelial cells of the fetal choroid plexus suggests response element on the gene encoding 15-PGDH [25].that the enzyme forms a functional unit with the apical Downregulation of the epithelial enzyme in the neonatemembrane. Consistent with this idea is the fact that, at 0.9 would then follow the normal fall in plasma cortisolgestation, enzyme activity was high in the minced tissue, occurring after birth [7,37]. Extending this speculationwhere structural integrity of membranes may still be further, upregulation of stromal 15-PGDH could be linkedrelatively preserved, but was low in the homogenized to the transient decrease in plasma thyroid hormonestissue, where cell disruption is nearly complete. Neverthe- during the neonatal period [7,8]. As a corollary, theless, any such structural requirement for the function of the progressive increase in plasma thyroid hormones duringenzyme does not appear to be as strict at 0.7 as at 0.9 the first postnatal month [8] might ultimately lead togestation, particularly for PGE catabolism. downregulation of stromal 15-PGDH, thereby explaining2
Rapid relocation of 15-PGDH from the epithelium to the the very low levels of 15-PGDH in the pregnant adultstroma during perinatal development is an unexpected choroid plexus. Thyroid hormones not only reduce 15-observation. Generally, 15-PGDH is found in the cytosol PGDH expression but also interfere with the activity of theof epithelial cells, and this applies specifically to the lung enzyme [27] and, by being transported by a specific carrier[30], gastric mucosa [19] and placenta [10]. In none of in the choroid plexus [33], they may exert their effectthese organs has evidence been obtained for the presence locally. Verification of our scheme, which implies not onlyof the enzyme in stromal fibroblasts [10,19,30]. Consider- a role for these hormones but also their differentialing the linkage of this phenomenon to birth, it is reason- influence on epithelial versus stromal cells, would entailable to assume the involvement of hormonal influences. further work.Glucocorticoids [39] and thyroid hormones [27] are known The presence of 15-PGDH in the choroid plexus of theto increase and decrease, respectively, 15-PGDH expres- fetus and lamb is consistent with extensive PGE catabo-2

154 N. Krunic et al. / Developmental Brain Research 121 (2000) 145 –155
movements in fetal sheep, Can. J. Physiol. Pharmacol. 75 (1997)lism in intact [22] and minced (this study) specimens at135–142.this stage of development. Proximity of 15-PGDH to the
¨ ˚[2] E. Anggard, C. Larsson, B. Samuelsson, The distribution of 15-brush-border membrane of the epithelium in the fetus 13hydroxy prostaglandin dehydrogenase and prostaglandin-D -reduc-suggests that any PGE , present either in the CSF in vivo tase in tissues of the swine, Acta Physiol. Scand. 81 (1971) 396–2
or in the incubation medium in vitro, is readily accessible 404.[3] L.Z. Bito, Inhibition of renal prostaglandin metabolism and excre-to the enzyme. At the same time, persistence of catabolism
tion by probenecid, bromcresol green and indomethacin, Prosta-in the lamb implies that PGE can cross the epithelium to2 glandins 12 (1976) 639–646.reach the stromal enzyme. No such accessibility to the[4] L.Z. Bito, R. Baroody, Concentrative accumulation of
3inner region of the choroid plexus is evident in the case of [ H]prostaglandins by some rabbit tissues in vitro: the chemical3PGF [21,22], and this peculiar difference from PGE has nature of the accumulated [ H]labelled substances, Prostaglandins 72a 2
(1974) 131–140.been tentatively ascribed to an accessory carrier system[5] L.Z. Bito, H. Davson, J.R. Hollingsworth, Facilitated transport ofspecifically directing PGF away from the enzyme [22].2a prostaglandins across the blood–cerebrospinal fluid and blood–brainAn organic acid transporter expressed in the developing
barriers, J. Physiol. 256 (1976) 273–285.choroid plexus [15] is a possible candidate for this [6] L.Z. Bito, R.A. Baroody, M.E. Reitz, Dependence of pulmonaryaccessory uptake. Whether PGF is actually a substrate prostaglandin metabolism on carrier-mediated transport processes,2a
Am. J. Physiol. 232 (1977) E382–E387.for this carrier remains to be verified.[7] G. Cabello, D. Levieux, Hormonal status in the newborn lambThe existence of a functional 15-PGDH in the choroid
(cortisol, T3, T4). Relationships to the birth weight and the length ofplexus for only a limited period of development raises thegestation: effect of the litter size, Biol. Neonate 39 (1981) 208–216.
possibility of a special role of catabolism in prostaglandin [8] G. Cabello, C. Wrutniak, Thyroid function in the newborn lamb.homeostasis. Significantly, the period in question coincides Physiological approach of the mechanism inducing the changes in
plasma thyroxine, free thyroxine and triiodothyronine concentra-with the stage when PGE inhibits breathing [9,14,18].2tions, J. Dev. Physiol. 13 (1990) 25–32.Hence, the enzyme, in concert with the transport mecha-
¨[9] L.A. Carlson, L.-G. Ekelund, L. Oro, Circulatory and respiratorynism(s) [21,22], may prevent PGE from attaining levels in2 effects of different doses of prostaglandin E in man, Acta Physiol.1the CSF effective on PGE receptors located in brainstem2 Scand. 75 (1969) 161–169.respiratory centers [35]. By operating at the interface [10] P.Y.C. Cheung, J.C. Walton, H.H. Tai, S.C. Riley, J.R.G. Challis,
Localization of 15-hydroxyprostaglandin dehydrogenase in humanbetween blood and CSF, choroidal 15-PGDH lends itselffetal membranes, decidua, and placenta during pregnancy, Gynecol.well to the degradation of PGE originating from the2Obstet. Invest. 33 (1992) 142–146.periphery [16], brain [12,34], or in the choroid plexus itself
[11] F. Coceani, I. Bishai, E. White, E. Bodach, P.M. Olley, Action of[29]. prostaglandins, endoperoxides, and thromboxanes in the lamb ductus
In conclusion, the present study reports for the first time arteriosus, Am. J. Physiol. 234 (1978) H117–H122.[12] F. Coceani, I. Bishai, D. Engelberts, R.V. House, S.L. Adamson,the presence of 15-PGDH immunoreactivity in the choroid
Response of newborn and adult sheep to pyrogens: relation betweenplexus. It also demonstrates a singular, developmentallyfever and brain eicosanoid changes, Brain Res. 700 (1995) 191–linked change in the location of the enzyme, resulting in204.
preferential catabolism of PGE . This selectivity in the2 [13] F.E. DiBenedetto, L.Z. Bito, Transport of prostaglandins and otheroperation of 15-PGDH during the early period after birth eicosanoids by the choroid plexus: its characterization and physio-
logical significance, J. Neurochem. 46 (1986) 1725–1731.may be important not only for the maintenance of continu-[14] F.A. Guerra, R.D. Savich, L.D. Wallen, C.H. Lee, R.I. Clyman, F.E.ous breathing but also as a protection against any disrup-
Mauray, J.A. Kitterman, Prostaglandin E causes hypoventilation2tion caused by PGE formed in response to infectious or2 and apnea in newborn lambs, J. Appl. Physiol. 64 (1988) 2160–inflammatory noxae. 2166.
[15] R. Hogue Angeletti, A.J. Bergwerk, P.M. Novikoff, A.W. Wolkoff,Dichotomous development of the organic anion transport protein inliver and choroid plexus, Am. J. Physiol. 275 (1998) C882–C887.Acknowledgements
[16] S.A. Jones, S.L. Adamson, I. Bishai, J. Lees, D. Engelberts, F.Coceani, Eicosanoids in third ventricular cerebrospinal fluid of fetal
This work was supported by the Medical Research and newborn sheep, Am. J. Physiol. 264 (1993) R135–R142.Council of Canada. The technical assistance of Eric [17] E. Kitraki, M.N. Alexis, M. Papalopoulou, F. Stylianopoulou,
Glucocorticoid receptor gene expression in the embryonic rat brain,Seidlitz and Aina Tilups is gratefully acknowledged. N.K.Neuroendocrinology 63 (1996) 305–317.is supported by the Canadian Foundation for the Study of
[18] J.A. Kitterman, G.C. Liggins, J.E. Fewell, W.H. Tooley, InhibitionSudden Infant Death and the Genesis Research Founda-of breathing movements in fetal sheep by prostaglandins, J. Appl.
tion. S.L.A. is a Career Investigator of the Heart and Physiol. 54 (1983) 687–692.Stroke Foundation of Ontario. [19] K. Kobayashi, K. Higuchi, T. Arakawa, T. Matsumoto, H. Nagura,
Effect of sofalcone on localization of 15-hydroxyprostaglandindehydrogenase, an enzyme that metabolizes prostaglandin E , in rat2
gastric mucosa: an immunohistochemical study, J. Clin. Gastroen-References terol. 14 (1992) S39–S42.
[20] N. Krunic, S.L. Adamson, I. Bishai, F. Coceani, Prostaglandin[1] S.L. Adamson, D. Engelberts, K.J. Whiteley, Low dose in- catabolism in the fetal choroid plexus: a special arrangement, Soc.
domethacin via fetal vein or cerebral ventricle stimulates breathing Neurosci. Abstr. 22 (1996) 773.

N. Krunic et al. / Developmental Brain Research 121 (2000) 145 –155 155
[21] N. Krunic, S.L. Adamson, I. Bishai, F. Coceani, Prostaglandin [31] C.R. Pace-Asciak, Prostaglandin biosynthesis and catabolism in theuptake and catabolism by the choroid plexus during development in developing fetal sheep lung, Prostaglandins 13 (1977) 649–660.sheep, Dev. Brain Res. 100 (1997) 82–89. [32] C.R. Pace-Asciak, G. Rangaraj, Prostaglandin biosynthesis and
[22] N. Krunic, S.L. Adamson, F. Coceani, Differential uptake and catabolism in the developing fetal sheep brain, J. Biol. Chem. 251catabolism of prostaglandin (PG)E versus PGF in the sheep (1976) 3381–3385.2 2a
choroid plexus during development, Dev. Brain Res. 119 (2000) [33] G. Schreiber, A.R. Aldred, A. Jaworowski, C. Nilsson, M.G. Achen,11–19. M.B. Segal, Thyroxine transport from blood to brain via trans-
[23] G.C. Liggins, Adrenocortical-related maturational events in the thyretin synthesis in choroid plexus, Am. J. Physiol. 258 (1990)fetus, Am. J. Obstet. Gynecol. 126 (1976) 931–941. R338–R345.
[24] O.H. Lowry, N.J. Rosebrough, A.L. Farr, R.J. Randall, Protein [34] K.R. Sinervo, G.N. Smith, A.D. Bocking, J. Patrick, J.F. Brien,measurement with the Folin phenol reagent, J. Biol. Chem. 193 Effect of ethanol on the release of prostaglandins from ovine fetal(1951) 265–275. brain stem during gestation, Alcohol Clin. Exp. Res. 16 (1992)
[25] M. Matsuo, C.M. Ensor, H.H. Tai, Characterization of the genomic 443–448.1structure and promoter of the mouse NAD -dependent 15-hydroxy- [35] T.C. Tai, S.J. Lye, S.L. Adamson, Expression of prostaglandin E2
prostaglandin dehydrogenase gene, Biochem. Biophys. Res. Com- receptor subtypes in the developing sheep brainstem, Mol. Brainmun. 235 (1997) 582–586. Res. 57 (1998) 161–166.
[26] M. Mibe, K. Nagai, T. Oshige, N. Mori, Endogenous inhibitors of [36] T. Takizawa, Y. Ikeda, H. Togashi, M. Yamamoto, K. Arishima, F.human placental prostaglandin dehydrogenase, Prostaglandins Akahori, T. Masaoka, Inhibitory effect of indomethacin on neonatalLeukotrienes Essent. Fatty Acids 46 (1992) 241–245. lung catabolism of prostaglandin E : possible mechanism of the2
[27] P.K. Moore, J.R.S. Hoult, Experimental hyperthyroidism in rats re-opening of the ductus arteriosus after indomethacin therapy, J.suppresses in vitro prostaglandin metabolism in lung and kidney, Tox. Sci. 21 (1996) 243–248.Prostaglandins 16 (1978) 335–349. [37] S.J. Thomas, D.W. Wilson, C.G. Pierrepoint, E.H.D. Cameron, K.
[28] J. Nakano, A.V. Prancan, S.E. Moore, Metabolism of prostaglandin Griffiths, Measurement of cortisol, cortisone, 11-deoxy-cortisol andE in the cerebral cortex and cerebellum of the dog and rat, Brain corticosterone in foetal sheep plasma during the perinatal period, J.1
Res. 39 (1972) 545–548. Endocrinol. 68 (1976) 181–189.[29] J.L. Norton, S.L. Adamson, A.D. Bocking, V.K.M. Han, Prosta- [38] D.W. Walker, N. Pratt, Effect of probenecid on breathing movements
glandin-H synthase-1 (PGHS-1) gene is expressed in specific and cerebral clearance of prostaglandin E in fetal sheep, J. Physiol.2
neurons of the brain of the late gestation ovine fetus, Dev. Brain 506 (1998) 253–262.Res. 95 (1996) 79–96. [39] C.Q. Xun, C.M. Ensor, H.H. Tai, Regulation of synthesis and
1[30] R.T. Okita, A.R. Sinning, J.R. Okita, H.H. Tai, R.R. Markwald, J.M. activity of NAD -dependent 15-hydroxyprostaglandin dehydrogen-1Bergholte, NAD -dependent 15-hydroxyprostaglandin dehydrogen- ase (15-PGDH) by dexamethasone and phorbol ester in human
ase: immunochemical characterization of the lung enzyme from erythroleukemia (HEL) cells, Biochem. Biophys. Res. Commun.pregnant rabbits, Arch. Biochem. Biophys. 279 (1990) 242–248. 177 (1991) 1258–1265.




















