
PÉCSI TUDOMÁNYEGYETEM TTK BI Általános és Környezeti Mikrobiológiai Tanszék TOXIKOLÓGIA...
88
PÉCSI TUDOMÁNYEGYETEM TTK BI Általános és Környezeti Mikrobiológiai Tanszék TOXIKOLÓGIA GYAKORLATOK Szerkesztette: Vágvölgyi Csaba, Pesti Miklós Az összeállításban résztvettek: Kucsera Judit, Manczinger László, Pesti Miklós, Pfeiffer Ilona, Takács Krisztina, Vágvölgyi Csaba Pécs 2005 A kétciklusú képzés bevezetése a magyar felsőoktatásban a természettudományi szakokon alkalmazkodás a munkaerőpiaci igényeihez. HEFOP-3.3.1-P-2004-06-0016/1.0 Az Európai Szociális Alap támogatásával
-
Upload
pecsimuzeumok -
Category
Documents
-
view
2 -
download
0
Transcript of PÉCSI TUDOMÁNYEGYETEM TTK BI Általános és Környezeti Mikrobiológiai Tanszék TOXIKOLÓGIA...

PÉCSI TUDOMÁNYEGYETEM
TTK BI
Általános és Környezeti Mikrobiológiai Tanszék
TOXIKOLÓGIA
GYAKORLATOK Szerkesztette: Vágvölgyi Csaba, Pesti Miklós
Az összeállításban résztvettek:
Kucsera Judit, Manczinger László, Pesti Miklós, Pfeiffer Ilona, Takács Krisztina, Vágvölgyi Csaba
Pécs
2005
A kétciklusú képzés bevezetése a magyar felsőoktatásban
a természettudományi szakokon
alkalmazkodás a munkaerőpiaci igényeihez.
HEFOP-3.3.1-P-2004-06-0016/1.0
Az Európai Szociális Alap támogatásával

2

BEVEZETÉS
A jegyzet célja, hogy a toxikológia gyakorlatokra való felkészülést elősegítse, a kísérlet menetének leírása segítséget nyújtson a végrehajtáshoz. A gyakorlatos jegyzet egyben jegyzőkönyvként is szolgál, kísérletek kiértékelése rögzíthető benne. A jegyzet elején a laboratóriumi munkavédelmi rendszabályokat az általános anyag- és eszközismeret, majd a mikroszkóp rövid leírása követi. Az egyes gyakorlatokat az elméleti tudnivalók összefoglalásával kezdjük, majd a kísérlet anyag- és eszközigényét adjuk meg (zárójelben feltüntetve az 1 párosra vonatkozó mennyiségeket), ezt követi a gyakorlat menetének leírása. Megadjuk azt, hogy az elvégzett kísérlettel kapcsolatosan milyen kérdésekre kell választ adni, javaslatot teszünk a kísérlet kiértékelésének módjára.
1

I. A LABORATÓRIUM RENDJE
A toxikológia gyakorlatokon a laboratóriumokra vonatkozó általános rendszabályokat be kell tartani (elektromos;- és; gázkészülékek kezelésével, sav, lúg, méreg, robbanásveszélyes vegyszerek kezelésével kapcsolatos előírások). Ha a gyakorlat során mikroorganizmusokkal dolgozunk a fertőzésveszély miatt az általános mikrobiológiai rendszabályokat is szigorúan be kell tartani. A toxikológiai vizsgálatok során számos olyan anyaggal, vegyszerrel dolgozhatunk, amelyek potenciális veszélyt jelenthetnek az emberre, ezért ügyeljünk a megfelelő védőfelszerelés használatára és az anyagok előírásszerű kezelésére. A gyakorlatokon tartsuk be a gyakorlatvezető utasításait!
• Laboratóriumban étkezni, dohányozni, laboratóriumi edényekből inni, laboratóriumi eszközöket egyéb célra felhasználni szigorúan tilos!
• A gyakorlatokon védőköpenyt kell használni. Ezt a munkaköpenyt lehetőleg más célra ne használjuk, vagy egyéb gyakorlatokon való használat esetén gyakran tisztítsuk.
• Csak kifogástalan állapotban levő eszközöket és edényeket használjunk! • A mérgező vegyszerekkel körültekintően bánjunk! • A mérgező anyagokkal szennyezett edényeket, eszközöket a külön erre a célra rendszeresített
kidobóba gyűjtsük! • Élő tenyészetek kiömlése, tenyészeteket tartalmazó edények törése esetén a fertőzésnek kitett
felületeket, a törött edényeket fertőtlenítő folyadékkal (pl. neomagnol) csírátlanítsuk! Az esetről a gyakorlatvezetőt informáljuk.
• Oltóeszközök leégetését előírásszerűen végezzük; nagy mennyiségű oltóanyag rátapadása esetén a láng közepén szárítsuk, utána égessük, különben szétfröccsenhet a mikrobatömeg.
• Tilos a lefolyóba önteni élő mikrobák tenyészetét vagy szuszpenzióját! Elpusztításuk hővel
vagy vegyszerrel történik. Tilos továbbá bármilyen vegyszert a lefolyóba önteni! • Ne öntsünk a lefolyóba megolvasztott agaros táptalajt, mert a csövek eltömődnek! • Mosogatás előtt az edényeket autoklávozással, vagy fertőtlenítő szerekben (formalin,
neomagnol, sterogenol, flóraszept) történő több órás áztatással kell csíramentesíteni. • Munka befejeztével a laboratóriumi asztalt fertőtlenítő folyadékkal (flóraszept vagy
neomagnol) megnedvesített ruhával le kell törölni! • A laboratóriumból való távozás előtt (gyakorlat közben is) kezet kell mosni fertőtlenítő
folyadékban!
2

II. ANYAG ÉS ESZKÖZISMERET
2.1. TÁPTALAJOK
A mikrobák vizsgálata tenyésztés közben vagy tenyésztés után történik. A tenyésztésre táptalajokat használunk. A mikroorganizmusok tápanyagszükségletüket a táptalajból fedezik. A különböző mikróbák tápanyagigénye rendkívül változatos lehet, de vannak olyan táptalaj összetevők amelyek alapvetően szükségesek. A tápközeg komponensei két fő részből állnak:
Esszenciális összetevők Járulékos összetevők: -víz→ oldószer -prekurzorok -energiaforrás→ redukált, nagy energiatartalmú vegyületek -vitamiok -szén-forrás -nitrogén-forrás -ásványi anyagok→ enzimek
A táptalajok osztályozása különböző szempontok alapján történhet.
Halmazállapot szerint 1. Folyékony táptalajok: a tápoldat komponensek összemérése során a szilárdító anyagot kihagyjuk (táplevesek, tápoldatok). 2. Szilárd táptalajok: a folyékony táptalajt szilárdító anyaggal egészítjük ki.
Szilárdító anyagok az alábbiak lehetnek: A) Agar-agar: poliszacharid, 1-3%-ban használatos (1% lágyagar), dermedéspontja
koncentrációtól függően 38°C feletti, tengeri moszatokból készítik, csak igen kevés mikroorganizmus hidrolizálja.
B) Zselatin: polipeptid, 10-30%-ban használatos, dermedéspontja a koncentráció függvénye, számos mikroba bontja.
C) Szilikagél: előnye, hogy szerves anyagokat nem tartalmaz. A táptalaj készítése időigényes. Ritkán alkalmazzák, általában kemolitotróf mikroorganizmusok tenyésztésekor.
Felhasznált anyag szerint 1. Természetes táptalajok: szerves eredetű anyagok, melyeknek pontos összetételét nem ismerjük, pl.
húsfőzet, élesztő kivonat, maláta, melasz, gyümölcslevek, tej. 2. Szintetikus táptalajok: összetételüket pontosan ismerjük. 3. Félszintetikus táptalajok: természetes és szintetikus összetevők keverékei.
3

Az alkalmazott tápanyagkomponensek szerint 1. Alaptáptalajok: a növekedéshez szükséges alapanyagokon kívül (esszenciális összetevők) speciális
adalékanyagokat nem tartalmaznak, pl. T1 – univerzális táptalaj 2. Szelektív táptalajok: alaptáptalaj + szelektív gátlóanyag (pl. antibiotikum). Egyes mikroba
csoportok visszaszorítását célozzák. 3. Differenciáló vagy indikátor táptalajok: alaptáptalaj + indikátor anyag, amely speciális
anyagcsere reakciókat jelez (pl. laktóz bontás, savképzés). 4. Speciális táptalajok: különböző vizsgálatok céljainak megfelelően, vagy különböző speciális
igényű mikrobák számára összeállított táptalajok. Táptalaj készítés: először a szilárd alkotókat mérjük be, majd feloldjuk nem teljes térfogatban. Oldódás után állítjuk be a végleges térfogatot. A pH érték beállítása 5%-os HCl ill. 5→%-os NaOH-dal történik finomskálás indikátor papírral vagy műszerrel történő ellenőrzés mellett, majd autoklávozunk. Autoklávozás után az agar tartalmú táptalajok pH-ja 0,1-0,5 pH értékkel csökken a szilárdító anyag enyhe hidrolízise és változása miatt. Agar tartalmú táptalajok nyomás alatt sterilezhetők, zselatin tartalmúak csak áramló gőzben (nyomás alatt erős hidrolízis következik be). A hőérzékeny táptalaj komponenseket szűréssel való sterilezés után adjuk az autoklávozott és megfelelően lehűtött alaptáptalajhoz.
2.2. TENYÉSZTÉSHEZ HASZNÁLT EDÉNYEK
Minden tenyésztéshez használt laboratóriumi edényt a táptalaj kifejtése előtt vattadugóval kell ellátni (gyapot- vagy papírvatta egyaránt használható).
Kémcsövek: folyékony táptalajból (1.C. ábra) 5 ml mérendő be. Szilárd táptalajból magas agar (1.B. ábra) esetén 10 ml, ferde agarhoz (1.A. ábra) 5 ml szükséges. A ferde táptalaj készítésével célunk a nagyobb felület biztosítása.
A B C 1. ábra: A kémcső mint tenyészedény
4
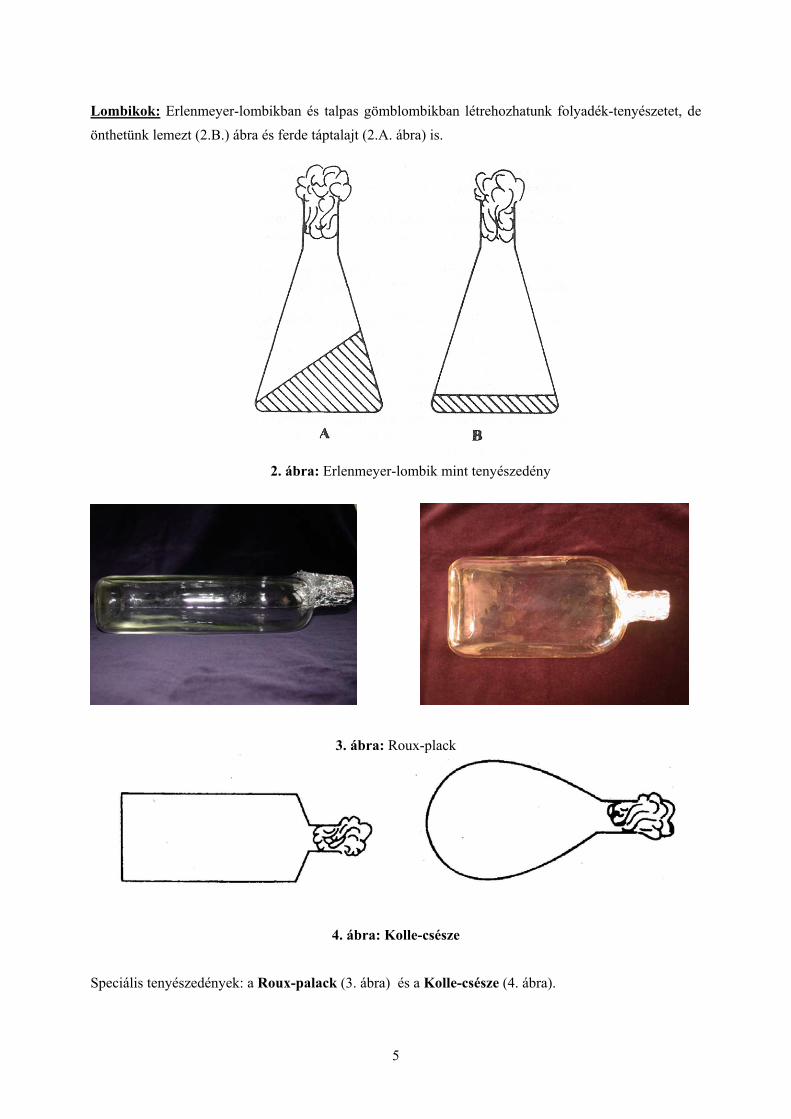
Lombikok: Erlenmeyer-lombikban és talpas gömblombikban létrehozhatunk folyadék-tenyészetet, de önthetünk lemezt (2.B.) ábra és ferde táptalajt (2.A. ábra) is.
2. ábra: Erlenmeyer-lombik mint tenyészedény
3. ábra: Roux-plack
4. ábra: Kolle-csésze Speciális tenyészedények: a Roux-palack (3. ábra) és a Kolle-csésze (4. ábra).
5

A fenti edényféleségek (beleértve a nem speciális Erlenmeyer-lombikot is) mind szilárd, mind nyugvófelszínes folyadékkultúra nevelésére alkalmasak (a táptalaj rétegvastagsága kicsi, felülete nagy). A tenyészedényeket a táptalaj betöltése után vattadugóval zárjuk, majd autoklávozással sterilezzük. Célszerű az edényeket papír, celofán vagy alufólia "sapkával" ellátni sterilezés előtt, nehogy az autokláv gőzterében a vattadugók átnedvesedjenek. Petri-csésze: a leggyakrabban használt laboratóriumi tenyészedény (5. ábra). A táptalaj kiöntése előtt szárazon sterilezzük. A táptalaj kiöntésekor a fedőt csak a szükséges mértékig emeljük. A 9–10 cm átmérőjű csészébe 20-25 ml táptalaj szükséges. A tenyészedényeket az alábbiak szerint jelöljük: 1.) a mikroorganizmus neve és laborkódja, 2.) az oltás dátuma, 3.) a táptalaj típusa, 4.) a gyakorlat száma, 5.) a kivitelező neve.
5. ábra: Petri-csésze
A fermentor olyan változatos méretű tenyészedény, amely gépi segédletekkel a tenyészetek nevelésére használható, ahol a tenyésztés körülményeit megfelelő ellenőrző rendszerekkel tudjuk biztosítani. A tápoldatban szaporodó mikroorganizmusok homogenitását keveréssel, oxigénnel való egyenletes ellátásukat steril levegőbefúvással érjük el.
2.3. OLTÓESZKÖZÖK
Oltótű és az oltókacs: hőszigetelt végű, leégethető fémnyelű (Kolle-nyél, 6. ábra) tűzálló fémtűvel vagy hurokkal (kaccsal) ellátott eszköz. Sterilezése lángban, leégetéssel történik. Mikrotiterkacs: a hurok helyett kalibrált térfogatú fémkosara van, amely ismert térfogatú folyadék (szuszpenzió) átvitelére alkalmas.
6. ábra: Oltótű, oltókacs
6
Pipetták: Folyadéktenyészetek ismert térfogatának átoltására használhatók az üvegpipetták, amelyeket fémdobozban, vagy alufóliába csomagolva száraz hővel sterilezzük. Biopipetták: (automata pipetták) különböző térfogat adagolására alkalmas (rögzített, vagy szabályozható méretű) eszközök, cserélhető sterilezhető műanyag pipetta heggyel.
2.4. STERILEZÉS
A tiszta tenyészetekkel végzett munkák előtt az eszközökön, tenyészedényekben, tápközegben lévő mikrobákat el kell pusztítani. Sterilezés során minden mikróbát elpusztítunk. A dezinficiálás pedig csak a kórokozó mikroorganizmusok elpusztítását jelenti. A sterilezésnek különböző módjai vannak.
I. Fizikai módszerekkel történő sterilezés
A) Sterilezés hővel • Leégetés lánggal: oltótű, oltókacs esetében alkalmazzuk átoltások során. • Száraz hővel: üveg, fémeszközöket, pl. pipettákat, Petri-csészéket sterilezünk így. A
hőlégsterilizálók megadott hőmérsékletre állítható önreguláló elektromos berendezések. A sterilezés időtartama 4 óra 140°C-on vagy 2 óra 160°C-on.
• Nedves hővel Pasteurözés: magas hőmérsékleten (60-96°C) a vegetatív sejtek nagy részének elpusztítása,
időtartama 1-40 perc, pl. 65°C – 30 perc; 85°C – 5 perc. Hátránya, hogy a Clostridium spórák túlélik.
Arnoldozás: sterilizálás áramló gőzben 100°C-on 20-30 percig. Ekkor a vegetatív sejtek elpusztulnak, a spóráknak azonban csak egy része. Kivitelezése le nem zárt autoklávban vagy szelep nélküli kuktafazékban történik.
Tyndallozás: három egymást követő napon arnoldozunk. A kezelések közötti időben a spórák kihajtanak és elpusztíthatók (frakcionált csíramentesítés). Hőérzékeny tápkomponensek esetén (pl. zselatin) alkalmazhatjuk.
Autoklávozás: sterilezés nyomás alatt (autokláv: 7. ábra). A leggyakrabban alkalmazott körülmények: 120°C, 1,0 atmoszféra túlnyomás, 15-30 perc. Egy adott nyomáson elérhető hőmérséklet az autoklávtérben a levegő és gőz arányától függ.
túlnyomás (atm) gőz teljes levegő (°C) gőz 1/2 levegő (°C) gőz levegő nélkül (°C)
0,66 90 105 115
1,00 100 112 121
1,66 115 124 130
2,00 121 128 135
7

A sterilezésre használt berendezésének megfelelő működését ismert hőtűréssel rendelkező mikroorganizmusokkal ellenőrizetjük, pl. autokláv esetén→ B. stearothermophilus; hőlég estetén→ Bacillus speciesek.
Szimplafalú autokláv kezelése: 1. Ellenőrizzük, hogy van-e megfelelő mennyiségű víz az autoklávban. 2. Behelyezzük a sterilizálandó anyagokat (edény, eszköz, tenyészet). 3. Lezárjuk az autokláv fedelét, szorítócsavarokkal egyenletesen leszorítjuk. 4. Ellenőrizzük, hogy a gőz-szelep nyitva van-e. 5. Bekapcsoljuk az autokláv fűtését. 6. Megvárjuk, amíg az autokláv felmelegszik. A gőzszelepet csak az intenzív gőzkiáramlás megindulása
után 2-3 perc múlva zárjuk el. 7. Bekapcsolt autoklávot magára hagyni tilos! A nyomás emelkedésekor ellenőrizzük, hogy a biztonsági
szelep működik-e. 8. A kívánt nyomásérték elérésekor az autoklávot 20 percen keresztül megfelelő hőmérsékleten tartjuk,
majd a fűtést kikapcsoljuk. 9. Megvárjuk amíg a túlnyomás nullára csökken. Nyitjuk a gőz-szelepet, majd az autokláv fedelét.
Vigyázzunk! A felszálló gőz éget! A duplafalú és automata autoklávok kezelése a készülékekhez mellékelt használati utasítás szerint történik.
7. ábra
l. gőz-szelep, 2. nyomásmérő, 3. biztonsági szelep, 4. hőmérő, 5. fedél, 6. vízszintjelző, 7. rakfelület, 8. fűtőtest, 9. vízszint
8

Gyakorlatainkban gyakran alkalmazunk autokláv helyett kuktafazekat sterilezésre. Így is teljes csírátlanítás érhető el. A kuktafazékba desztillált vizet kell tölteni kb. kétujjnyi magasságban, behelyezni az eszközöket, majd lezárjuk és gázlánggal melegítjük a kuktát. A forrástól számítva 20 percig sterilezünk. B) Sterilezés UV-sugárzással: különböző sugárzások (Röntgen-, radioaktív-, UV-sugárzás) csíraölő hatásúak. A gyakorlaton 260-265 nm sugárzási maximumú UV (germicid)-lámpákkal sterilezünk. C) Sterilezés szűrőkkel: folyadék sterilezésére vagy folyadék és szilárd táptalajok hőérzékeny komponenseinek csíramentesítésére szűrőket alkalmazunk. • Abszorpciós szűrők: Chamberland (zománc nélküli porcelán), Berkefeld (kovaföld), Seitz
(azbeszt); ezek egy idő után telítődnek. • Pórus szűrők: leggyakrabban a szinterüveg (zsugorított porózus üveglemez) szűrők és a
membránszűrők használatosak. A bakteriológiai szinterüveg szűrők között legismertebb a G-5 jelzésű szűrő, 0,1µm pórusátmérővel. A membránszűrők anyaga különböző szerves polimer lehet: leggyakrabban a 0,22 vagy a 0,45 µm pórusátmérőjű készítményekkel dolgozunk (Millipore- vagy Sartorius szűrők). A baktériumok eredményes szűréséhez max. 0,3 µm pórusátmérőjű szűrőt használjunk.
A szűrés elősegítésére vízlégszivattyút, olajlégszivattyút (8. ábra), vagy fecskendőhöz hasonló nyomópumpát (9. ábra) alkalmazunk.
8. ábra 9. ábra
l. szűrőtölcsér, 2. szűrőlap l. dugattyú, 2. fecskendőház 3. gumidugó, 4. szívópalack 3. szűrőtartó, 4. szűrőlap, 5. vattazár (membrán)
9

D) Sterilezés ultrahanggal: Ritkán alkalmazott eljárás.
II. Sterilezés kémiai úton
A kemikáliák fehérjékkel és egyéb sejtkomponensekkel való reakciók révén fejtik ki sterilező hatásukat. A hatás a koncentráció és az idő függvénye. Alkalmazhatók nehézfémsók (HgCl2, AgNO3), oxidálószerek (KMnO4, H2O2), halogének (klór, jód), szerves oldószerek (pl. 70%-os etanol,
dimetilszulfoxid), a fenol 3%-os vizes oldata (karbol), a formalin 3%-os vizes oldata; gáz (etilénoxid). Felületaktív, zsíremulgeáló kation szappan, membránkárosító, spórákat nem öl. A neomagnol (benzol-szulfonkloramid-nátrium) 0,1%-os vizes oldatában a keletkező NaOCl a fehérjék aminocsoportjait klórozza. Megfelelő koncentrációban sporocid. Hatástalanítja a toxinokat is.
Steril oltófülke
Használata a táptalajoknak steril edényekbe való kifejtésekor, Petri-csészékbe történő lemez készítésekor, ezek leszárításakor, valamint oltási műveletek végzése esetén ajánlatos. Különösen Petri-csészék, illetve folyadéktenyészetek, hosszú tenyészidőre beállított kultúrák inokulálását célszerű a steril fülkében elvégezni. Az ún. "biohazard" típusú oltófülke lényege, hogy a levegő a beépített szívó és nyomóhatást kifejtő elektromotorok hatására a fülke hasznos belső terének tetején elhelyezett szűrőn keresztül csíramentesen kerül a munkatérbe, ott lefelé áramlik, majd a lyuggatott munkalapon keresztül elhagyja a munkateret. Az ismételt steril szűrőn való átjutás előtt egy előszűrőn halad át a levegőáram. Ez a mozgásirány, valamint a munkatér és a külvilág határfelületén egyéb segédberendezések (kiküszöbölendő az örvénymozgást) megakadályozzák, hogy a fülkébe a külső térből bárminemű szennyezés, fertőzésforrás bekerülhessen, egyben megakadályozza a munka során a belső térben használt mikroorganizmusok kikerülését is. A steril fülkék ezen típusa alkalmas a patogén
mikroorganizmusokkal való munkára is.
2.5. A MIKROORGANIZMUSOK TENYÉSZTÉSE
A mikrobákat kontrollált körülmények között kell tenyészteni. Fontos az optimális hőmérsékleten való tartás a gyors növekedés biztosítása érdekében. Erre a célra termosztátokat használunk (duplafalú, vízköpennyel, önreguláló elektromos fűtéssel ellátott szekrények). A mikróbák különböző levegőztetést igényelnek. Az aerob szervezetek nevelésekor – ha azok nem szilárd táptalaj felületén nőnek – a megfelelő levegőztetésről körkörös rázatással vagy fermentorban való tenyésztéssel gondoskodhatunk. A rázógépek termosztálását vízfürdő segítségével vagy száraz levegő befúvással biztosítjuk. Anaerobok esetén oxigén elvonást, illetve reduktív körülményeket kell a tápközegben biztosítani. Az egyes mikroorganizmusok tápnygszükséglete eltérő, ezért tenyésztéskor a megfelelő táptalajt kell alkalmazni. Végül, pedig a tenyésztési időt is figyelembe kell venni, mivel ez is más-más
10

az egyes mikróbáknál (baktériumoknál 1-2 nap, élesztőgombáknál 2-3 nap, penész- ill. fonalasgombáknál 1 hét).
2.6. MIKROSZKÓP-ISMERET
Ebben a fejezetben röviden áttekintjük a mikroszkóp használatával kapcsolatos tudnivalókat. Mikroszkópunk binokuláris tubussal, keresztasztallal, négy revolverváltóval, cserélhető achromát objektívvel, két pár okulárral rendelkező készülék. A fényforrás beépített alacsonyfeszültségű lámpa (10. ábra). Az objektíveken néhány jellemző érték van feltüntetve, pl.:
HI 100/1,25
160/0,17 (W
Glyc ) fedőlemez vastagsága
numerikus apertura
Az első szá(160) egyensorban a tönumerikus a A leszavak röviakkor érvénúgynevezetaz üveg fénszámot (lásdMűszerünk A ffelismerhetüobjektív nuszerint függ
olajimmerzió
I - vízimmerzió
- glicerin immerzió
m az objektív nagyítóképességét lő. Az első sorban a törtvonal urtvonal utáni szám azt a fedőlempertura érték érvényes. gnagyobb nagyítású objektíven a
dítése. Azt jelzi, hogy ennél az obyesek, ha az objektív frontlencséj
t száraz-objektívnél), hanem immeytörésével azonos. Az okulárokon később).
teljesítőképességét a fenti adatok aeloldóképesség annak a legkisebbnk. A feloldóképesség (F) a m
merikus aperturájától (NA) legked:
F
nagyítás
tubushosszjelzi, ha a tubus hossza mm-ben az alatta lévő számmal táni szám az objektív numerikus aperturája. A második
ez vastagságot jelzi mm-ben, melynél a felette lévő
nagyítást jelző szám előtt álló HI a "homogén immerzió" jektívnél a jelzett numerikus apertura és nagyítás értékek e és a preparátum között nem levegő van (mint a többi, rziós olaj (cédrus olaj vagy anizol), melynek fénytörése jelzik a nagyítóképességet és (kis körbe írva) az ún. mező
lapján egyszerűen kiszámíthatjuk. tárgynak az átmérője, amelyet a műszerrel még éppen egvilágításra használt fény hullámhosszától (λ) és az vezőbb megvilágítási viszonyok esetén az alábbi képlet
λ = ––––––– 2 x NA
11

A numerikus apertura a tárgy és az objektív frontlencséje között lévő közeg törésmutatója (n) valamint a frontlencse félnyílásszöge (α) sinusának a szorzata:
NA = n x sinα
frontlencse A nyílásszöget a tárgy egy pontjából a frontlencse széleihez húzott két egyenes adja (élesre állított kép esetén!). A sinα maximális értéke 1; gyakorlatban 0,95 az elérhető érték, ekkor a teljes nyílásszög kb. 140°. Száraz objektíveknél n = 1 (levegő) homogén immerziós objektíveknél n = 1,51. (cédrusolaj)
tárgy Számításainknál a fény hullámhosszát 500 nm-nek vegyük; ez az emberi szem által legjobban érzékelhető, világoszöld színű fény hullámhossza. A mikroszkóp nagyítása az objektív nagyításának és az okulár nagyításának szorzata. Az objektíven jelölt nagyítás 160 mm tubushosszra vonatkozik. Egyes mikroszkópoknál a binokuláris tubus 160 mm-nél hosszabb; ebben az esetben a nagyítás kiszámításánál a binokuláris tubuson látható faktorral (1,6x) is szorozni kell. Azt, hogy a mikroszkóp látómezejében megfigyelt terület mekkora a valóságban, kiszámíthatjuk, ha az okuláron jelzett mezőszámot elosztjuk az objektív nagyításával, így mm-ben kapjuk a kérdéses kör átmérőjét. Természetesen a tubushosszat itt is figyelembe kell vennünk. A binokuláris tubust mindenki beállítja a saját pupillatávolságára, a tubuson lévő 55-től 75-ig számozott skála a pupillatávolságot jelzi mm-ben. A binokuláris tubuson korrigálható a két szem közötti esetleges dioptriakülönbség. A bal szemet behunyva a képet a mikrométer-gombbal élesre állítjuk, majd a jobb szemet behunyva a képet a tubuson lévő gyűrűvel állítjuk élesre. Akiknél a két szem egyforma, a 0-jel vonása egybeesik a tubus oldalán lévő vonással. Az élesre állítást úgy végezzük, hogy oldalról ellenőrizve először a tárgyasztalt a durvabeállító gombbal az objektív felé közelítjük, míg az objektív a preparátumot csaknem érinti, majd a mikroszkópba nézve a tárgyasztal süllyesztésével (előbb a durvabeállítóval, majd amikor a kép már látszik, a mikrométer gombjával) állítunk élesre. A mikroszkópos kép jó minőségét és egyenletes kivilágítottságát a lámpa égőjének és blendéjének, a kondenzor magasságának és blendéjének megfelelő beállításával, továbbá a kondenzorba homályos üveg illetve színszűrő behelyezésével igyekszünk elérni. A kondenzor alsó részén található, kihajlítható lencsét csak a 6,3x-os nagyítású objektívvel kapcsolatban iktassuk be. A mikroszkóphoz csatlakoztatható fáziskontraszt berendezés, sötétlátóteres kondenzor és UV-fényforrás is (lásd a megfelelő gyakorlatokat).
12

Mikroszkópos fotózás Amikor a mikroszkópozás során látott képet rögzíteni akarjuk, akkor szükségessé válik a mikroszkóp és a fényképezőgép (kamera) összekapcsolása. Ez történhet egyszerű mechanikai úton, de az összekapcsoláskor közbeiktathatunk különböző típusú automatikus rendszereket elsősorban a vizsgált tárgyon áthaladó fény mérésére, és az ehhez szükséges expozíciós idő automatikus mérésére. Gyakorlatunkban a BA2 típusú fénymérő és exponáló automatát használjuk. Ezen keresztül csatlakoztatjuk a kamerát a fáziskontraszt mikroszkóphoz. (Többféle típusú kamera használható mikroszkópos fotózáshoz, ha a normál fényképezőgép optikáját eltávolítva illeszteni tudjuk a mikroszkóphoz.) Az exponáló automata segítségével különböző fedettségű, sötét illetve világos, jól megvilágított és gyenge világítottságú preparátumról készíthető jól exponált film. Az automata fénymérő üvegszál optikán beméri az adott preparátum megvilágítási viszonyait és ennek megfelelően vezérli a felvétel expozíciós idejét (ez a másodperc tört részeitől 5-6 perc hosszúságú lehet). A fényképezésre szánt mikroszkópos preparátummal szemben magas minőségi követelményeket támasztunk:
l.) A tárgylemezek, fedőlemezek teljesen tiszták, karcmentesek és optikailag is megfelelőek legyenek.
2.) A vizsgálandó anyag, élő sejt, vagy rögzített preparátum mindig egyrétegben (egy síkban) helyezkedjen el. Ez lényeges, mert az átvilágítás során egyik réteg zavarja a másikat. Az egy síkban elhelyezkedő mikrobák mélységélessége jelentősen javul, így morfológiájuk jól tanulmányozható és a képi látvány is tökéletesebb.
3.) Beszáradt, vagy zsugorodott preparátum nem fényképezhető.
4.) A tárgylemezre felvitt anyag (intakt sejtek, fonalak, konidiumok) mindig folyadékba legyen ágyazva. Ez a fénytörésből eredő zavaró problémákat csökkenti, a fényképezéshez elengedhetetlen kontúrélességet fokozza.
A tárgylemezre vitt anyagra cseppentsünk az alkalmazandó folyadékból, vagy eleve ebbe a folyadékcseppbe készítsük el a fényképezendő preparátumot, tiszta fedőlemezzel fedjük le, a felesleges folyadékot itassuk fel. (Ne ússzon a fedőlemez!)
5.) Az expozíciós idő alatt állítsuk le, vagy fékezzük le a preparátum mozgását. A pillanatnyi mozdulatlanság előfeltétele az éles felvételnek. A mozgás leállítása, vagy lassítása történhet zselatinba, glicerinbe történő ágyazással, tárgylemezhez történő fixálással, ragasztással, vagy pl. konidiumok cellux szalagra történő ragasztásával.
6.) Válasszunk ki jellemző, nem zsúfolt látómezőt a fényképezésre.
7.) A felvételeket viszonylag gyorsan készítsük el, mert a fényforrás melegít, a preparátum száradása megindul, és beindul a Brown-féle mozgás is.
13

8.) A kép "megszerkesztésekor" síkban gondolkodjunk (a binokuláris mikroszkópok térben adják a képet). A fénykép "kevesebbet" ad vissza a binokuláris feltétben látottaknál.
9.) A mikroszkópos felvétel készítése előtt ügyeljünk a preparátum élesre állítására. Itt a képkeresőben látható szálkereszt élesre állításához igazodjunk.
A mikroszkópos fekete-fehér felvételek készítéséhez leggyakrabban a közepes érzékenységű (15/10, vagy 17/10 din) filmek alkalmazhatók. Ezek szemcsézettsége finom, jól nagyíthatók, kontrasztosak, a részleteket jól visszaadják. Színes felvételek készítésekor műfény diafilm, fluorescens-mikroszkóp esetében napfény diafilm használható 18/10 din alsó érzékenység határral. Mikroszkópunk vázlata (10. ábra): l: talp; 2: durva beállítógomb; 3: mikrométer beállítógomb; 4: kondenzor emelő-süllyesztő gomb; 5: kondenzor, kihajlítható alsó lencse, kihajlítható tartógyűrű színszűrő vagy opálüveg számára, blende; 6: tárgyasztal keresztasztallal; 7: keresztasztal mozgató gombjai; 8: objektívek a revolverváltóban; 9: binokuláris tubus okulárokkal; 10: mikroszkóp megvilágító rendszere (talpba épített transzformátor, izzó).
10. ábra: Laboratóriumi mikroszkóp vázlata
14

III. GYAKORLATOK
A gyakorlatokon párokban dolgozunk, ezért az “Anyagok és eszközök” pontban az egy párra
vonatkozó mennyiségeket zárójelben tüntettük föl.
15

1. gyakorlat: ÁTOLTÁSOK GYAKORLÁSA
Az elkészített táptalajokat (T-1—T-5a) különböző steril tenyészedényekbe szétosztjuk, majd a táptalajokat hagyjuk lehűlni, illetve megdermedni. A kémcsöveket megdöntve helyezzük el a ferdeagar, és függőlegesen a magas agar készítéséhez. A lehűlt tápoldatba, illetve a megdermedt táptalajra oltunk.
a) Oltás ferde agarra
Mikroorganizmusok:Escherichia coli, Serratia marcescens, Bacillus cereus var. mycoides, B. megaterium, Micrococcus luteus, (T-1); Saccharomyces cerevisiae, Candida utilis, Rhodotorula rubra, Syncephalastrum racemosum, Aspergillus niger 48 órás tenyészetei T-5 ferde agaron (a baktériumok T-1, a gombák T-5 táptalajon növekednek jól).
Anyagok és eszközök: • T-1 ferde agar (4 db) • T-5 ferde agar (4 db)
A leoltás menete: 1) Bal kézbe fogjuk a leoltandó tenyészetet tartalmazó és az "üres " kémcsövet (11. ábra). A csövek
helyzete közel vízszintes legyen. Hüvelykujjunkkal a tenyerünkhöz szorítjuk a csöveket. (Rögzíthetjük a csöveket az ujjak közé szorítással is.) Az agar oltandó felülete mindig felénk legyen fordítva.
2) Az üres cső vattadugóját megforgatjuk, hogy könnyen kiemelhető legyen, ha a belefejtett táptalajtól beragadt volna.
3) Az oltóeszközt úgy fogjuk jobb kézzel, mintha íróeszközt fognánk. Közel függőleges helyzetben a gázlángba tartva, felizzítjuk az oltókacsot, majd a nyél fém részét lángoljuk le.
4) Jobb kezünk gyűrűs- és kisujjával kiemeljük a tenyészetes cső dugóját úgy, hogy a kiemelés után a dugó szabad része ne érintkezzen semmivel. Lángoljuk le a kémcső száját.
5) A kaccsal benyúlunk a kémcsőbe, lehűtjük üres agarfelületen. A tenyészetből inokulumot veszünk ki. (Inokulum = az a mikrobamennyiség, amellyel be/le/oltjuk az új táptalajt.) A kacs nem érhet a cső oldalához. Lelángoljuk a kémcső száját, a dugót visszahelyezzük a tenyészetes csőbe.
6) A beoltandó kémcső dugóját kivesszük, lelángoljuk a kémcső száját (vigyázzunk, az inokulum ne érjen a lángba!). Beoltjuk a táptalaj felületét zegzugos vonal mentén (12.A. ábra) lángoljuk a frissen leoltott cső száját, visszahelyezzük a vattadugót. Leégetjük az oltókacsot (először szárítva, majd izzítva), állványba állítjuk, majd állványba helyezzük a kémcsöveket is.
16

11. ábra: Ferde agarra történő oltás egy lehetséges módja
Figyelem! Az egész műveletet a gázláng közvetlen közelében végezzük, ügyelve arra, hogy a beoltás folyamata kellő ütemű legyen, ugyanakkor nagy levegőmozgást széles mozdulatokkal, kapkodással ne keltsünk. A tenyészedényeket oltás előtt lássuk el jelöléssel. Átoltunk 4 baktérium, 2 élesztőgomba és 2 fonalasgomba törzset.
b) Oltás magas agarba (szúrt tenyészet)
Mikroorganizmusok: Serratia marcescens, Bacillus cereus var. mycoides, Escherichia coli, Saccharomyces cerevisiae, Rhodotorula rubra 48 órás tenyészetei T-1 illetve T-5 ferde agaron.
Anyagok és eszközök: • T-1 magas agar (2 db) • T-5 magas agar (2 db)
A gyakorlat menete: Oltás menete ugyanaz mint előbb, csak tűvel és zegzugos vonalhúzás helyett egyetlen szúró mozdulattal oltunk be 2 baktérium és 2 élesztőgomba törzset (12.B. ábra).
A B
12. ábra: Ferde- illetve magas agar leoltása
17

c) Oltás kémcsőből folyékony táptalajba
Mikroorganizmusok: Serratia marcescens, Escherichia coli, Bacillus megaterium, Micrococcus luteus, Saccharomyces cerevisiae, Syncephalastrum racemosum, Aspergillus niger 48 órás tenyészetei T-1 illetve T-5 ferde agaron
Anyagok és eszközök:• 15 ml T-1a tápoldat kémcsőben (4 db) A beoltás menete: az a) gyakorlatnál leírtak szerint történik. A kémcsöveket egyenként beoltjuk 2 baktérium, 1 élesztőgomgba és 1 fonalasgomba törzzsel A mikroorganizmusokat szuszpendáljuk a folyadékban.
1. gyakorlat: Eredmények és értékelés
1. Rajzoljuk és írjuk le, hogyan szaporodnak a különböző mikrobák a szúrt tenyészetekben (gázterelés, milyen mélyen nő).
18

1. gyakorlat: Eredmények és értékelés 2. Ellenőrizzük az átoltás eredményességét. 3. Figyeljük meg az átoltott mikroorganizmusok színét, telepmorfológiai (fénye-matt felszín) tulajdonságait. Ábrázoljuk megjelenési formájukat. 4. Rögzítsük hogy folyadéktenyészetben milyen a különböző mikrobák tenyészete (egyformán zavarosít, felületen úszik, kiülepszik, milyen a színük)?
Folyékony tenyészet Mikroba Ferde agar Magas agar
keverés előtt keverés után
19

2. gyakorlat: MIKROORGANIZMUSOK SZAPORODÁSA KÜLÖNBÖZŐ TÁPTALAJOKON
Jelen gyakorlat a címben megjelölt célkítűzés mellett a kémcsőből Petri-csészére történő oltás technikájának elsajátításának célját is szolgálja. A mikroorganizmusok szaporodás intenzitása függ a törzs jellegétől és a tenyésztési körülményektől. Ugyanazon mikroorganizmus különböző táptalajokon különböző gyorsasággal szaporodhat és eltérő morfológiai sajátságokat mutathat. Ez a jelenség a mikroszervezetek gyors adaptív tulajdonságával magyarázható, nem örökletes, ún. modifikatív sajátság.
Mikroorganizmusok: Escherichia coli, Serratia marcescens, Bacillus megaterium, Bacillus cereus var. mycoides, Saccharomyces cerevisiae, Rhodotorula rubra, Trichoderma viridae, Aspergillus niger (vad típusú, prototrof), Aspergillus niger 341 ade- (adenin auxotrof mutáns törzse) és 48 órás tenyészetei T-1, illetve T-5 ferde agaron.
Anyagok és eszközök: • T-1, T-2, T-3, T-4 és T-5 táptalajt tartalmazó Petri-csésze (1-1 db) • félkémcső 2 ml steril fiziológiás sóoldattal (3 db)
A gyakorlat menete: Az egyes táptalajokból 3-3 Petri-csészét öntünk ki. A táptalajok megdermedése után a táplemezeket le kell szárítani! Amennyiben steril fülke nem áll rendelkezésre, a csészéket óvatosan nyitjuk, a fedelet kézben lefelé tartva steril vattával páramentesítjük, ezután a 13. ábra szerint az asztallapra helyezzük, majd a táplemezt tartalmazó csésze felet tetőcserépszerűen rátesszük. (E művelet előtt célszerű az asztallapot fertőtlenítőszeres ruhával letörölni.) Steril fülkében a csészék normál helyzetben nyitott fedővel száríthatók. A leoltani kívánt mikrobákból a steril fiziológiás sóoldatban szuszpenziót készítünk (úgy mintha kémcsőből folyadék tápoldatba oltanánk). Fonalasgombák esetében célszerű a konidium szuszpenziót sterilvizes lemosással készíteni. Az agar dermedése, majd a csészék szárítása után a szuszpenziókból oltókaccsal 3-3 mikrobát oltunk le Petri-csészénként egymástól 2-2,5 cm távolságra, egyenes vagy zegzugos vonal mentén. A leoltás menete: leégetett oltókaccsal inokulumot veszünk ki a szuszpenziót tartalmazó félkémcsőből, majd azt állványba helyezzük, ezután a Petri-csésze fedőt felemeljük annyira, hogy az oltóeszköz alá férjen. A Petri-csészéket termosztátba helyezzük, 48 órán át 30°C-on inkubálunk.
13. ábra: Táptalajjal frissen kiöntött Petri-csészék szárítása
20
2. gyakorlat: Eredmények és értékelés l. Táblázatban tüntessük fel az egyes fajok szaporodáserősségét a különböző táptalajokon.
Táptalajok Mikroorganizmus
T-1 T-2 T-3 T-4 T-5
+++ erős, ++ közepes,+ gyenge szaporulat, – nem szaporodik
2. Írjuk le a morfológiai jellegzetességeket, eltéréseket a különböző táptalajokon.
21

3. gyakorlat: TISZTATENYÉSZETEK KÉSZÍTÉSE
Kevert tenyészetek, a természetben található többféle fajból álló úgynevezett nyers tenyészetek, ezek tisztítása, egyes fajok tiszta (ún. egysejt) tenyészetének kinyerése szilárd táptalaj alkalmazásával szélesztés, illetve lemezöntés módszerével történhet.
A) Szélesztés Mikroorganizmusok: Escherichia coli, Serratia marcescens, Micrcoccus luteus 48 órás tenyészete T-1 ferde agaron. Anyagok és eszközök:
• T-1 Petri csésze (2 db)
• 4 ml steril fiz.só félkémcsőben (1 db)
A gyakorlat menete: A táptalaj kiöntése után kevert szuszpenziót készítünk steril fiziológiás sóoldatban a három baktérium fajjal. (Micrococcus luteus-ból nagy oltókacsnyi, Escherichia coli-ból kis oltókacsnyi, Serratia marcescens-ből pedig csak oltótűnyi mennyiséget vigyünk a sóoldatba!) Ezután a szuszpenziót kémcsőkeverővel homogenizáljuk. Petri-csészére egy-egy kacsnyit kiviszünk a felületre és a 14. ábra szerint szélesztjük: (1) zegzugos vonalat húzunk, a kacsot leégetjük (2) innen újabb csíkozást végzünk, ismét leégetünk, majd (3) harmadszor is így szélesztünk. A másik csészét a 15. ábra szerint szélesztjük: (1) csíkot húzunk a csésze közepére, hagyjuk a csíkot beszáradni, majd a száradás után (2) zegzugos vonal mentén szétkenjük a beszáradt szuszpenziót.
14. ábra: 15. ábra:
A szélesztés különböző módjai
A csíkhúzás során a baktériumok száma egyre fogy, végül különálló telepeket kapunk. 48 órás inkubáció után a szín alapján elkülönítjük a telepeket (természetesen kevert szuszpenzióknál egymás után többször szélesztünk a már különálló telepekből, így győződünk meg arról, hogy hasad-e tovább több fajra az izolált telep).
22

3. gyakorlat: Eredmények és értékelés l. Rajzoljuk le, milyen eloszlásban jelennek meg a telepek szélesztésnél.
23

B) Lemezöntés Mikroorganizmusok: A szélesztésnél használt kevert szuszpenzió. Anyagok és eszközök:
• T-1 1% agarral Petri csésze (4 db) (frissen készítendő!)
• 4,5 ml fiz.só félkémcsőben (7 db)
• 100 ml főzőpohár
• automata pipetta (100 ul)
• bakterológiai pipetta
A gyakorlat menete: A kevert szuszpenzióból 0,5 ml-t kivéve steril pipettával a 16. ábra szerint 10x hígítási sorozatot készítünk 7 lépcsőben. Hígítás során az egyre híguló szuszpenzióval mindig "átöblítjük" (felszívjuk, kifolyatjuk) a pipettát, mielőtt a 0,5 ml szuszpenziót a következő csőbe átvisszük. A megolvasztott 1% agartartalmú T-1 táptalajt eril kémcsövekbe adagoljuk 15 ml-ként és 40°C-os vízfürdőben tartjuk a beoltásig. A hígítási sorozat utolsó öt hígítási lépcsőjéből (IV.-V.-VI.-VII.) steril pipettaheggyel egy-egy steril folyékony1% T-1 táptalajt tartalmazó kémcsőbe 0,1ml-nyi szuszpenziót viszünk át. (A leghígabb szuszpenzióval kezdjük, így egy pipettaheggyel dolgozhatunk végig.) Kémcsőkeverővel homogenizálunk, majd steril Petri-csészékbe öntjük a kémcsövek tartalmát. (lemezöntés) és hagyjuk megszilárdulni. Az agarba dermedt sejtek egymástól elválva képeznek telepeket.
16. ábra: 10x-es léptékű hígítási sorozat készítése
Megjegyzés: Mindkét módszer kvantitatívvá tehető és így sejtszámolásra alkalmas. Dermedés után 48 órán keresztül 30°C-on inkubálunk.
24

3. gyakorlat: Eredmények és értékelés 1. Írjuk le, melyik hígításban különülnek el egymástól a baktérium telepek lemezöntésnél. 2. Rajzoljunk le olyan lemezeket, ahol a telepek jól számolhatók.
25

4. gyakorlat: MIKROORGANIZMUSOK MORFOLÓGIAI VIZSGÁLATA RÖGZÍTETT
FESTETT PREPARÁTUMOKKAL, EGYSZERŰ FESTÉSEK
Festéssel jobban megfigyelhetővé tudjuk tenni a mikroorganizmusokat, nagyobb sejteken belül sejtalkotókat ismerhetünk fel és egyéb, a sejtek beltartalmára vonatkozó adatokat nyerhetünk. Az 1-2 µm nagyságú baktérium sejtek esetében a festés magát a sejtalak, a mikroszkópos kép felismerését teszi lehetővé, hisz méretük kicsinysége miatt önmaguk csekély interferencia jelenséget idéznek elő. A festett preparátumon áthaladó nem monochromatikus fénynyaláb a festékre jellemző hullámhossz tartományban elnyelést szenved, így a kép vizuális érzékelhetősége növekszik.
Rögzített preparátumok készítésével kapcsolatos általános tudnivalók A tárgylemezeket detergenssel zsírtalanítjuk, desztillált vízzel alaposan leöblítjük és törlés nélkül szárítjuk. Kaccsal vagy tűvel kis csepp vízbe szuszpendáljuk a vizsgálandó mikroba kis sejttömegét, a cseppet szétkenjük és hagyjuk beszáradni. A szárítás melegítéssel való siettetése zsugorodást eredményez! A baktériumokat (1-2 µm) a tárgylemezre felvitt szuszpenzió teljes beszáradása után hővel rögzítjük úgy, hogy a beszáradt kenetet 4–5-ször áthúzzuk gázláng felett. A gombák rögzítésére etanol : éter = 1 : 1 keverékét használjuk, mert hő hatására - a gombasejt (4-5 µm) nagyobb mérete miatt - alakja megválozhat. Az etanol-éter keveréket rácsepegtetjük a beszáradt kenetre, majd hagyjuk bepárlódni. A rögzítést minden festés előtt végezzük el! Festés után a preparátumot először 40x-es nagyítású objektívvel ellenőrizzük, majd immerziós olaj (vagy anizol) alkalmazásával l00x-os nagyítású immerziós lencsével vizsgáljuk. A mikroszkóppal kapcsolatos tudnivalókat lásd a mikroszkóp leírásánál!
A) Festés metilénkékkel
Mikroorganizmusok: Micrococcus luteus, Staphylococcus aureus, Bacillus subtilis 24 órás tenyészete T-1, Saccharomyces cerevisiae, Geotrichum candidum 24 órás tenyészete T-5 ferde agaron.
Anyagok és eszközök: Metilénkék festékoldat (R-1), tárgylemezek, fedőlemezek, festőkád, mikroszkóp.
A gyakorlat menete:A megfelelő rögzítés után 5 percig festjük a preparátumot, utána csapvízzel mossuk, amíg a mosófolyadék színtelenül folyik le. A preparátum szárítása siettethető szűrőpapírral való leitatással.
26

B) Festés karbolos fukszinnal Mikroorganizmusok: Staphylococcus aureus, Bacillus megaterium, Candida utilis 24 órás tenyészetei T-1 illetve T-5 ferde agaron.
Anyagok és eszközök: Karbolos fukszin (R-2), tárgylemezek, fedőlemezek, festőkád, mikroszkóp.
A gyakorlat menete: A rögzített kenetet 30 mp-ig festjük, majd csapvízzel mossuk, szárítjuk. Megjegyzés: Immerziós olaj alkalmazása után az objektívlencsét benzolos vagy xilolos vattával tisztítsuk meg. Alkoholt e célra ne használjunk, mert a lencsék ragasztottak és az alkohol a ragasztóanyagot kioldja!
4. gyakorlat: Eredmények és értékelés 1. Rajzoljuk le a mikroszkópos képeket, ábrázoljunk egy-egy látómezőt a mikrobákkal, vigyázva a méretarányok megtartására.
27

5. gyakorlat: MIKROORGANIZMUSOK MORFOLÓGIAI VIZSGÁLATA RÖGZÍTETT
FESTETT PREPARÁTUMOKKAL, ÖSSZETETT FESTÉSEK
Az összetett festések esetén több egyszerű festékkel egymás után festünk, vagy festékkomplexeket viszünk a kenetre. E technikával azt kívánjuk elérni, hogy különböző sejttípusokat, csoportokat elkülönítsünk, illetve sejten belül tudjunk elkülöníteni egyes elemeket.
A) Gram-festés (Hucker szerint) Ha kristályibolyával festett baktériumokra jódoldatot öntünk, a sejtekben sötétlila színű jód-festék komplex képződik. Ez a komplex a Gram-negatív baktériumokból etanollal könnyen kimosható, a Gram-pozitívakból nem. Az elszíntelenített Gram-negatív baktériumokat szafraninnal pirosra festjük, hogy jól felismerhetők legyenek. Az eltérő festődés alapja az, hogy a Gram-pozitív és Gram-negatív baktériumok között lényeges strukturális és kémiai különbségek vannak, elsősorban a sejtfalban.
Mikroorganizmusok: Micrococcus luteus, Staphylococcus aureus, Bacillus cereus var. mycoides, Escherichia coli, Serratia marcescens 24 órás tenyészete T-1 ferde agaron.
Anyagok és eszközök:R-3, R-4, R-5 festékoldatok, etanol, tárgylemezek, festőkád, mikroszkóp, immerziós olaj.
A gyakorlat menete: Kenetet készítünk az egyes baktériumokból külön-külön, majd egy Gram-pozitív és egy Gram-negatív baktérium keverékéből. Száradás után hővel rögzítünk. 1 percig festünk kristályibolyával (R-3), ezt pár másodperces csapvizes mosás követi. 1 percig Lugol-oldattal kezelünk (R-4), ezután csapvizes öblítés, majd szárítás következik. A tárgylemezt csipesszel megfogva festőkád felett pipettából etanolt csepegtetve a kenetre 30 másodpercig mossuk. Szárítjuk a preparátumot. Szafraninnal (R-5) kontrasztfestést alkalmazunk 10-20 másodpercig, majd csapvizes öblítés és szárítás következik. Megjegyzés: Feltétlenül fiatal, 24 órásnál nem idősebb tenyészetekkel dolgozzunk, idős sejteknél a különbségek elmosódnak!
28

5. gyakorlat: Eredmények és értékelés 1. Rajzoljuk le a mikroszkópos képeket a festődésnek megfelelő színnel vigyázva a méretarányok megtartására.
29

B) Spórafestés A baktériumok és gombák spórái egyszerű festéssel nem festődnek, mert a többrétegű spórafalba a festék nehezen épül be, ezért a festék bevitelére ún. agresszív festést alkalmazunk (fenol és melegítés). Az így megfestett spóra etanolos mosáskor nem színtelenedik el, míg a vegetatív sejtből a festék kimosódik. A vegetatív sejtek láthatóvá tételére ún. kontrasztfestést alkalmazhatunk. Az alábbi spórafestési eljárások baktériumok és gombák esetében egyaránt alkalmazhatók. Spórafestés előtt a hővel való rögzítés szükségtelen.
a) Módosított Dorner-féle módszer: Mikroorganizmus:Bacillus megaterium 48 órás tenyészete T-1 ferde agaron.
Anyagok és eszközök:Karbolos fukszin (R-2) festékoldat, etanol, tárgylemez, fedőlemezek, szűrőpapír, üvegkád, mikroszkóp, immerziós olaj.
A gyakorlat menete: A tenyészetből kenetet készítünk, rögzítés nélkül szárítjuk. Szűrőpapírt helyezünk a kenetre, erre karbolos fukszint csepegtetünk. 10-15 percig forró vízfürdő fölé helyezzük a preparátumot. A festéket folyamatosan pótoljuk. A szűrőpapírt eltávolítjuk, a preparátumot etanollal mossuk a lecsepegő folyadék elszíntelenedéséig, majd csapvízzel öblítjük, leitatással szárítjuk. A spórák vörösek, a vegetatív sejtek fehérek.
b) Schaeffer-Fulton módszer:
Mikroorganizmus:
Saccharomyces kluyveri CBS 3082 törzse egy hetes tenyészete T-5 ferde agaron.
Anyagok és eszközök:
Malachit-zöld (R-6) és szafranin (R-5) festékoldat, tárgylemezek, fedőlemezek, festőkád, mikroszkóp.
A gyakorlat menete: A Saccharomyces kluyveri tenyészetből tárgylemezen szuszpenziót készítünk, szárítjuk rögzítés nélkül. Szűrőpapírt helyezünk a kenetre rácsepegtetünk malachit-zöld oldatot (R-6) és 10 percig forró vízfürdő felett melegítjük. Közben az elpárolgott folyadékot a festékoldat állandó adagolásával pótoljuk. Ezután 30 mp-ig folyóvízben mossuk a preparátumot, majd 30 mp-ig kontrasztfestést alkalmazunk szafraninnal (R-5). Végül csapvizes öblítés, majd szárítás következik. A spórák zöldek, a vegetatív alakok pirosra festődnek.
30

5. gyakorlat: Eredmények és értékelés 2. Rajzoljuk le fiatal és idős tenyészet mikroszkópos képét a festődésnek megfelelő színnel. 3. Rajzoljuk le a mikroszkópos képet méretarányosan és a festődésnek megfelelő színnel.
31

6. gyakorlat: MIKROORGANIZMUSOK NAGYSÁGÁNAK MÉRÉSE
Különböző vizsgálatoknál gyakran szükséges a szervezetek méreteinek megállapítása. A méreteket az okulár- és az objektívmikrométer segítségével határozzuk meg. Az okulármikrométer az okulárba helyezhető kerek üveglemez, általában 0-100 beosztású skálával. Ez az okulárral forgatható, a tárgy pedig a tárgyasztallal mozgatható, így a skálát rá tudjuk állítani a látómezőben található bármely objektumra. Leolvashatjuk, hogy a vizsgált mikroba méretei hány okulár beosztásnak felelnek meg. Az okulármikrométer kalibrálása az objektívmikrométerrel történik. Az objektívmikrométer egy speciális tárgylemez, amelynek közepén 1 mm-nyi távolság 100 egységre van osztva. Az objektívmikrométer osztásközeinek mérete 10 µm.
Mikroorganizmusok: Schizosaccharomyces pombe, Geotrichum candidum 24 órás 25°C-on rázatott tenyészete T-5a tápoldatban. (A Geotrichum inokulum készítéshez maximum 48 órás ferde agaros tenyészet használható).
Anyagok és eszközök: R-2 festékoldat, okulár- és objektívmikrométer, tárgylemezek, fedőlemezek, mikroszkóp.
A gyakorlat menete: Az objektívmikrométert tárgyként használjuk, élesre állítjuk a skáláját. Az okulár skálát párhuzamos állásba állítjuk, a kezdő pontokat fedésbe hozzuk (17. ábra).
17. ábra: 1. okulármikrométer skála, 2. objektívmikrométer skála
1) Olvassuk le, hogy a 16x-os és a 40x-es nagyítású lencse alkalmazásakor hány objektívmikrométer
beosztásnak felel meg az okulár 0-100 beosztása!
32

2) Számítsuk ki a 16x-os és a 40x-es nagyítású objektíveknél az okulár mikrométer egy osztásközének értékét µm-ben! A leolvasott objektívmikrométer osztások számát osztjuk az okulárosztások számával (jelen esetben ez 100) és szorozzuk 10-zel (az objektívmikrométer egy osztásközének nagysága µm-ben).
3) festünk, (lásd 4.
4) eg az átlagértéket
l. Soszt2. Marth3. S
Okulármikrométer köz = leolvasott objktívmikrométer osztások száma X 10
okulárosztások száma (100)
A gombatenyészetekből készítsünk kenetet karbolos fukszinnal (R-2) 30 mp-iggyakorlat B rész). Mérjük le 10-10 vegetatív sejt, illetve arthrospóra hosszát, szélességét és adjuk mµm-ben!
6. gyakorlat: Eredmények és értékelés zámítsuk ki a l6x-os és a 40x-es nagyítású objektívek esetében az okulármikrométer egy ásközének értékét µm-ben.
érjük meg tíz Schizosaccharomyces pombe sejt és tíz Geotrichum candidum vegetatív sejt és rospóra hosszát és szélességét. zámoljuk ki az átlagméreteket.
33

7. gyakorlat: BAKTÉRIUMOK ÉS ÉLESZTŐK SEJTSZÁMMEGHATÁROZÁS MÓDSZEREI
A legtöbb baktérium és az élesztőgombák esetében a sejtek szaporodása során az utódsejtek elválnak egymástól. Ezeknél az élőlényeknél a sejtek szaporodása jól nyomonkövethető mind közvetlen, mind közvetett módszerekkel. A közvetlen sejtszámmeghatározás módszerei közül a mikroszkópos számolást, és a turbidimetriát alkalmazzuk leggyakrabban, de pl. a nedvessúlymeghatározás is ilyen módszer.
I. Közvetlen sejtszámolási módszerek: 1. Bürker-kamrás sejtszámolás: Nagyobb méretű sejteknél közvetlen sejtszámolás valósítható meg Bürker-kamrával. Ez a módszer az összsejtszám megállapítására ad lehetőséget. Gyakran alkalmazott, gyors és egyszerű sejtszámlálási módszer. 2. Turbidimetria: A meghatározás folyadéktenyészetek "zavarosságának" mérésén alapszik. A turbidimetriás mérésnél a műszeren mért "sűrűség"-értékeket kalibrálnunk kell. Ez közvetlen sejtszámolással vagy közvetve, élőcsíraszám megállapítással történik. Gyakorlatunkon a közvetlen sejtszámolás módszerét alkalmazzuk. Kalibrációs görbét veszünk fel, ahol egy-egy turbiditás értéknek meghatározott sejtszám felel meg. A vizsgálandó szuszpenzióból egyrészt felező hígítást készítünk és műszeresen megmérjük az egyes fokozatok zavarosságát, optikai denzitását (OD), másrészt Bürker-kamrás számolással meghatározzuk az eredeti sejtszámot. Az összsejtszámméréssel történő kalibrálás különböző tenyészeteknél alkalmazható, az élőcsíraszám meghatározással történő kalibrálás csak a logaritmikus szaporodási fázisban (ahol az összsejtszám és az élőcsíraszám gyakorlatilag azonos) ad pontos eredményt. A kalibrálást minden mikroorganizmus törzzsel külön kell elvégezni a mikroorganizmusok eltérő nagysága miatt, s a kalibrációs görbe mindig csak egy meghatározott korú tenyészetre vonatkoztatható. Megjegyzés: Mivel a mérés fotométerrel történik, az eredményeket befolyásolhatja a táptalaj színe vagy színes mikrobáknál a mikroorganizmus pigmentáltsága. Sárga színű tápoldatok esetén célszerű a zöld tartomány (500-520 nm) helyett vörösben (630-700 nm) elvégezni a méréseket. Ha spektrofotométerrel dolgozunk, az intervallumokon belül ott mérjünk, ahol maximális az elnyelés.
II. Közvetett sejtszámolási módszerek: A közvetett sejtszámolási eljárások esetében nem a tényleges sejtszámot kapjuk meg, hanem az "élőcsíraszámot", azaz a vizsgált tenyészetek azon sejtjeinek (egységeinek) számát, amelyek különálló telepeket képesek létrehozni. Bürker-kamrás méréssel összehasonlítva különösen idősebb tenyészetek esetében jelentős sejtszámkülönbséget kaphatunk a közvetlen számolás és a tenyésztési
34

7. gyakorlat: Eredmények és értékelés 1. Bürker-kamrás sejtszámolás: Számoljuk ki a Bürker-kamrás mérés alapján az eredeti szuszpenziók sejtszámát. Tíz Bürker-kamra egység átlagsejtszámából kiszámoljuk 1 mm3 szuszpenzió sejtszámát (egy kamraegységre jutó átlagértéket szorozzuk a kamratérfogat nevezőjével), majd kiszámítjuk a ml-kénti sejtszámot.
Sejtszám
Eredeti 10x 102x 103x 104x 105x 106x 107x
Lemezöntés (1 ml)
Szélesztés (50µl)
35
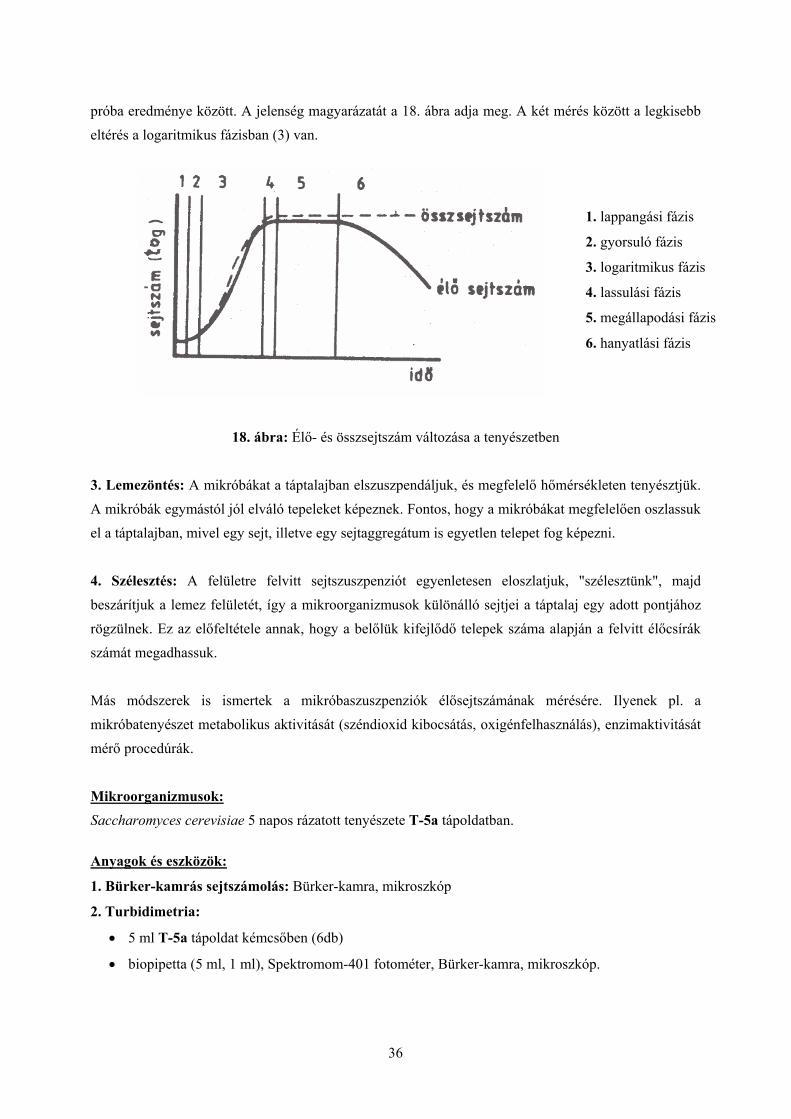
próba eredménye között. A jelenség magyarázatát a 18. ábra adja meg. A két mérés között a legkisebb eltérés a logaritmikus fázisban (3) van.
1. lappangási fázis
2. gyorsuló fázis
3. logaritmikus fázis
4. lassulási fázis
5. megállapodási fázis
6. hanyatlási fázis
18. ábra: Élő- és összsejtszám változása a tenyészetben
3. Lemezöntés: A mikróbákat a táptalajban elszuszpendáljuk, és megfelelő hőmérsékleten tenyésztjük. A mikróbák egymástól jól elváló tepeleket képeznek. Fontos, hogy a mikróbákat megfelelően oszlassuk el a táptalajban, mivel egy sejt, illetve egy sejtaggregátum is egyetlen telepet fog képezni.
4. Szélesztés: A felületre felvitt sejtszuszpenziót egyenletesen eloszlatjuk, "szélesztünk", majd beszárítjuk a lemez felületét, így a mikroorganizmusok különálló sejtjei a táptalaj egy adott pontjához rögzülnek. Ez az előfeltétele annak, hogy a belőlük kifejlődő telepek száma alapján a felvitt élőcsírák számát megadhassuk. Más módszerek is ismertek a mikróbaszuszpenziók élősejtszámának mérésére. Ilyenek pl. a mikróbatenyészet metabolikus aktivitását (széndioxid kibocsátás, oxigénfelhasználás), enzimaktivitását mérő procedúrák.
Mikroorganizmusok: Saccharomyces cerevisiae 5 napos rázatott tenyészete T-5a tápoldatban. Anyagok és eszközök:
1. Bürker-kamrás sejtszámolás: Bürker-kamra, mikroszkóp
2. Turbidimetria:
• 5 ml T-5a tápoldat kémcsőben (6db)
• biopipetta (5 ml, 1 ml), Spektromom-401 fotométer, Bürker-kamra, mikroszkóp.
36
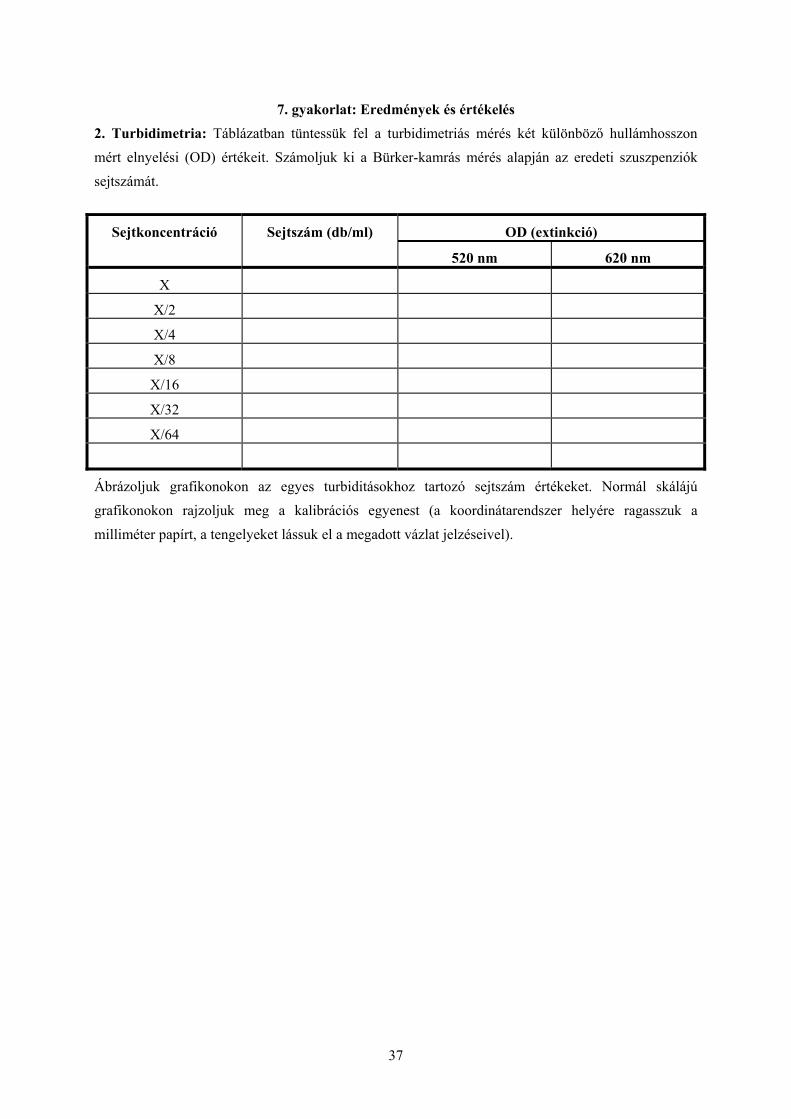
7. gyakorlat: Eredmények és értékelés 2. Turbidimetria: Táblázatban tüntessük fel a turbidimetriás mérés két különböző hullámhosszon mért elnyelési (OD) értékeit. Számoljuk ki a Bürker-kamrás mérés alapján az eredeti szuszpenziók sejtszámát.
OD (extinkció) Sejtkoncentráció Sejtszám (db/ml)
520 nm 620 nm
X
X/2
X/4
X/8
X/16
X/32
X/64
Ábrázoljuk grafikonokon az egyes turbiditásokhoz tartozó sejtszám értékeket. Normál skálájú grafikonokon rajzoljuk meg a kalibrációs egyenest (a koordinátarendszer helyére ragasszuk a milliméter papírt, a tengelyeket lássuk el a megadott vázlat jelzéseivel).
37

3. Lemezöntés:
• 4,5 ml fiz.só félkémcsőben (7 db)
• 3x25 ml T-5 1% agarral (frissen készítendő!)
• 1 ml automata pipetta
• 2x 100 ml-es főzőpohár
• steril Petri csésze (3 db)
4. Szélesztés:
• T-5 Petri csésze (3 db)
• 50 µl automata pipetta
• szélesztőbot, alkohol
A gyakorlat menete: 1. A Bürker-kamrát detergenssel zsírtalanítjuk, desztillált vízzel való alapos öblítés után törlés nélkül szárítjuk. Rászorítjuk a kamra fedőlemezét a "Newton-gyűrűk" megjelenéséig. Az élesztő tenyészetből 10x-es és 100x-os hígítást készítünk (lásd 3. gyakorlat 16. ábra). A szuszpenziókból üvegbottal egy cseppet rácseppentünk a Bürker-kamrára a fedőlemez mellé (19. ábra). Ha az üveg zsírmentes, a szuszpenzió egyenletesen "szívódik" a fedőlemez alá. 16x-os vagy 40x-es nagyítású objektívvel megnézzük, melyik szuszpenzió számolható könnyen és megbízhatóan (kamra alapegységenként 5-10 sejt legyen).
1. szorító fémből, 2. fedőlemez, 3. négyzetrácsos karcolat, 4. felcseppentés helye
19. ábra: Bürker-kamra felülnézetben
A számolást elvégezhetjük a nagy négyzet alapú egységben (20. ábra) 1/5 x l/5 = 1/25 mm2, vagy a téglalapalapú egységben 1/5 x l/20 = 1/100 mm2. A kamra magassága 1/10 mm, így a nagy négyzet alapú kamra 1/250 mm3, a téglalap alapú kamra 1/1000 mm3 térfogatú.
1. 1/20 mm-es karcolattávolság
2. 1/5 mm-es karcolattávolság
20. ábra: Bürker-kamra cellaméretei Tíz egységet leszámolunk, kiszámoljuk az egy egységre jutó átlagsejtszámot, kiszámoljuk 1 mm3 szuszpenzió sejtszámát (az egy kamraegységre jutó átlagértéket szorozzuk a kamratérfogat nevezőjével), majd kiszámítjuk a ml-kénti sejtszámot. Amennyiben nem az eredeti tenyészetet
38

számoltuk, a hígítási értékkel beszorozva megkapjuk az 5 napos tenyészet ml-kénti sejtszámát. A sejtkoncentráció megjelölésének módja pl. 3,6 x 107 sejt/ml.
2. Turbidimetria: A Saccharomyces cerevisiae tenyészetből felező hígítási sorozatokat készítünk 6 lépcsőben, ahol az első koncentráció maga az eredeti szuszpenzió: a felező hígításokhoz 5-5 ml szuszpenziót 5-5 ml T-5a tápoldattal sorozatban hígítunk. Az összehasonlító oldat a T-5a tápoldat. A hígítási sorozatok fényelnyelését 520 nm-nél és 620 nm-nél mérjük. A mérés 1 cm-es szögletes küvettában történik.
3. Lemezöntés: 7 tagból álló, 10-es léptékű hígítási sorozatot készítünk a Saccharomyces cerevisiae tenyészetből. Felolvasztjuk az 1% agartartalmú steril T-5 táptalajt, majd felhasználásig 40°C-os vízfürdőben tartjuk.. A táptalaj hőmérsékletének 40°C-ra történő beállítása után a hígítási sorozat megfelelő 3 tagjából (sejtszámolás alapjákiválasztott hígítások) 1-1 ml szuszpenzióval beoltunk 3 db steril Petri-csészét. A beoltás után a Petri-csészébe beleöntjük a 25ml meleg 1%-os T-5 táptalajt és óvatosan ∞ alakban mozgatva keverjük a csészéket. Dermedés után 48 órán át 30°C-on inkubálunk. A számolandó szuszpenzió hígításakor, a táptalajba vitelekor ügyelnünk kell az egyenletes eloszlásra és arra, hogy a mikroorganizmusok sejtjei egymástól elváljanak (ne legyenek aggregátumok), mivel egy sejt vagy 5-10 sejtből álló "egység" is egyaránt egyetlen telepet hoz létre.
4. Szélesztés: 3 db steril T-5 táptalajt tatalmazó Petri-csészébe Saccharomyces cerevisiae hígítási sorozatainak megfelelő hígítási fokozataiból 3-at kiválasztunk és a lemezek felületére oltjuk. A hígítási lépcsőkből biopipettával csészénként 0,05 ml-nyi mennyiségeket oltunk. A felületre vitt szuszpenziót üvegbotból hajlított oltóeszközzel egyenletesen szétszélesztjük. Steril oltófülkében szárítjuk le a lemezeket. 30°C-on 48 órán keresztül inkubáljuk a csészéket.
7. gyakorlat: Eredmények és értékelés
3. Lemezötés és szélesztés: Értékeléskor kiválasztjuk azt a hígítást, amely jól számolható telepmennyiséget ad. Megszámoljuk a telepeket a megfelelő hígításhoz tartozó 3 párhuzamos leoltásban, majd átlagot számolunk. Hasonlítsuk össze a közvetlen sejtszámolás adatait az élőcsíraszám értékekkel!
Összsejtszám Élőcsíraszám
Hígítás foka
Sejtszám (db/ml)
Sejtszám átlag
(db/ml)
Hígítás foka
Sejtszám (db/ml)
Sejtszám átlag
(db/ml)
% különbség
Lemezöntés
Lemezöntés
Lemezöntés
Szélesztés
Szélesztés
Szélesztés
39

8. gyakorlat: MIKROORGANIZMUSOK ANTAGONIZMUSÁNAK VIZSGÁLATA
Az élő szervezetek egymásra gyakorolt gátló hatása nem csak abban nyilvánul meg, hogy a környezeti feltételeket egymás számára aspecifikus anyagcsere termékekkel kedvezőtlenné teszik, hanem abban is, hogy specifikus gátló anyagokat termelnek. A gátlóanyagok egy másik fajhoz vagy törzshöz tartozó mikrobát megölik ("cidikus" hatás pl. baktericid, fungicid anyagok) vagy csak szaporodásukban gátolják ("sztatikus" hatás pl. bakteriosztatikus, fungisztatikus anyagok). Az antagonizmus ezen formája az antibiózis, a gátlóanyag az antibiotikum. Az antibiotikum specifikus hatású anyagcseretermék, amely s koncentrációban hatékony. Szilárd felületre oltva a gátlóanyagot termelő mikroorganizmust, a keletkezett antibiotikum a táptalajba diffundál és gátolja a termelő törzs mellé oltott érzékeny mikrobákat. Két mikroba faj között a kapcsolat lehet:
A faj B faj
neutralizmus Ø Ø
kommenzalizmus + Ø
protokooperáció + +
szimbiózis + +
antagonizmus - -
→kompetíció - -
→amenzalizmus - Ø
→parazitizmus + -
→predáció + -
→antibiózis + -
Mikroorganizmusok: Pseudomonas aeruginosa, Escherichia coli, Serratia marcescens, Staphylococcus aureus, Micrococcus luteus, Bacillus subtilis, Bacillus cereus var. mycoides, Candida utilis 3 napos, valamint Streptomyces griseus és Penicillium chrysogenum 5 napos tenyészete T-1 illetve T-5 táptalajon, ferde agaron.
Anyagok és eszközök: • 3 ml fiz.só félkémcsőben (3 db)
A gyakorlat menete: A csészék átmérőjének vonalába (21. ábra l.) 2-2 csészére a következő törzseket oltjuk le (gyakorlatra már előre leoltva), mint termelő mikrobákat: Pseudomonas aeruginosa, Escherichia coli, Bacillus subtilis, Streptomyces griseus, Penicillium chrysogenum. A leoltás szuszpenzióból történik, egyetlen
40

csíkhúzással (a három baktérium esetében a sejtmennyiség egy kacs/ml szuszpenzióként, a Streptomyces és Penicillium törzseknél az egész ferde agart lemossuk, 3 ml fiziológiás sóoldattal). Baktériumok esetén 48 órás, a Streptomyces griseus és Penicillium chrysogenum törzsek alkalmazásakor 5 napos inkubálás után "a termelő mikrobák" mellé a 21. ábra 2. szerint szuszpenzióból csíkhúzással leoltjuk a tesztmikrobákat majd újabb 48 órás inkubálás után értékeljük a lemezeket. (Célszerű a 48 órás tenyésztés után a "termelő " baktériumokkal leoltott csészéket hidegen tárolni, amíg a lassabban növő "termelők" is elszaporodnak majd ezután keresztbe oltani a tesztorganizmusokat!) Vigyázat: a kísérlet fokozottan fertőzésveszélyes! Megjegyzés: Vigyázzunk, hogy a tesztorganizmusok leoltásakor a már kifejlődött kultúrát ne érintsük oltókacsunkkal.
21. ábra
1. az antibiotikumtermelő mikroorganizmus 2. a tesztmikroorganizmusok
8. gyakorlat: Eredmények és értékelés 1. Táblázatba írjuk be, mely mikroorganizmusok között van antagonizmus, számmal jelöljük, hány mm a gátlási zóna nagysága.
“Teszt mikrobák” “Termelő mikrobák”
41

9. gyakorlat: OXIDO-REDUKCIÓS ENZIMEK VIZSGÁLATA
A.) Kataláz teszt A kataláz jelenléte vagy hiánya a mikroorganizmus anyagcseréjének fontos jellemzője. Az aerobok kataláz termelők (pozitívok), az anaerobok kataláz hiányosak (negatívok). A légköri oxigén jelenlétében az obligát anaerob szervezetekben a kataláz hiánya miatt toxikus hidrogénperoxid keletkezik. Kivételt képeznek a Lactobacillaceae család tagjai, amelyek fakultatív anaerob szervezetek de kataláz negatívok. Ezekben a baktériumokban az aerob körülmények között keletkező hidrogénperoxidot peroxidáz enzimek bontják le. (A nem kataláz típusú hidrogénperoxid bontó aktivitást egyes szerzők "pszeudokataláznak" is nevezik.)
Mikroorganizmusok: Escherichia coli, Staphylococcus aureus, Bacillus subtilis 48 órás tenyészete T-1 ferde agaron. Bacillus subtilis 48 órás rázatott tenyészete 15 ml T-1a táplevesben Erlenmeyer-lombikban. Lactobacillus sp. 48 órás tenyészete T-7a táplevesben anaerob körülmények között előnevelve (15 ml álló tenyészet kémcsőben).
Anyagok és eszközök: 100 ml T-1 táptalaj 3% agarral, 100 ml 3%-os H2O2 oldat (R-15), 3 db steril Petri-csésze, T-1 ferde
agarral, 2 db centrifugacső, centrifuga.
A gyakorlat menete: a) A folyadéktenyészetben elszaporodott baktériumok (Bacillus subtilis és Lactobacillus sp.) sejtjeit
centrifugálással összegyűjtjük (10 perc 8000 x g), majd a centrifugacsőben 3%-os H2O2-ot
rétegzünk rá, megfigyeljük b) 3 db Petri-csészébe T-1 táptalajból lemezt készítünk. Csészénként 4-4 foltba oltjuk a ferde agaron
tenyésztett mikrobákat. 48 órás 30°C-os inkubálás után 3%-os H2O2 -ot rétegzünk közvetlenül a
tenyészetek felületére. Vigyázat: a kísérlet fokozottan fertőzésveszélyes! .
42

9. gyakorlat: Eredmények és értékelés 1. Írjuk le, mit tapasztaltunk az A/a gyakorlat esetében. 2. Írjuk le az A/b gyakorlattal kapcsolatban látottakat.
43

B.) A metilénkék redukciója A citokróm rendszer rövidre zárását redox festékekkel megvalósíthatjuk. Anaerob viszonyok között a metilénkék elektronakceptorként viselkedik. A redukált metilénkék színtelen (leukometilénkék). Az elszíntelenedés mértéke arányos a redukció intenzitásával. Ha a rendszerbe oxigén jut, a metilénkék visszakékül.
Mikroorganizmus: Escherichia coli és Staphylococcus aureus 48 órás tenyészete T-1 ferde agaron.
Anyagok és eszközök: 100 ml T-8a metilénkék tápleves, 6 db kémcső, kémcsőkeverő.
A gyakorlat menete: 6 db kémcsőbe 10-10 ml T-8a tápoldatot öntünk, kuktafazékban sterilezzük. 2 csövet Escherichia coli-val, 2 csövet Staphylococcus aureus-szal beoltunk, 2 cső kontrollként sterilen marad. 48 órán keresztül 30°C-on inkubálunk. (A csövek belsejében közel anaerob viszonyok uralkodnak.) Vigyázat: a kísérlet fokozottan fertőzésveszélyes!
C.) Oxidáz reakció A citokróm oxidáz a terminális oxidációban a citokrómnak a molekuláris oxigén általi oxidációját katalizálja. A reakció könnyen kimutatható a dietil-p-feniléndiamin-HCl színtelen oldatának enzim hatására történő elszíneződésével. A reakció diagnosztikailag fontos az Enterobacteriaceae család tagjainak a Pseudomonas és Vibrio fajoktól való differenciálásában.
Mikroorganizmus: Escherichia coli és Pseudomonas aeruginosa 24 órás tenyészete T-1 ferde agaron.
Anyagok és eszközök: Oxidáz reagens (R-16), Petri-csésze, szűrőpapírcsíkok.
A gyakorlat menete: Petri-csészébe helyezett szűrőpapír darabkára néhány cseppet teszünk a reagensből, majd rákenjük a vizsgálandó tenyészet kacsnyi mennyiségét. Pozitív reakció esetén a papír 10 másodperc alatt sötétbíbor színű lesz.
44

Vigyázat: a kísérlet fokozottan fertőzésveszélyes! A kísérlet során használt, mikroorganizmusokat tartalmazó papírdarabokat neomagnol oldatban csíramentesítjük.
9. gyakorlat: Eredmények és értékelés 3. Figyeljük meg a beoltott csövek színváltozását. 4. Rázzuk fel erősen a csöveket, figyeljük meg a színváltozást a kontrollhoz képest. 5. Írjuk le a kísérlet során tapasztaltakat.
45

10. gyakorlat: MAKROMOLEKULÁK EXOENZlMES BONTÁSA
Az egyes fajok identifikálásában jelentős szereppel bírnak a különböző biokémiai sajátosságokat feltáró tesztek. Ezen tesztek elsősorban az orvosi mikrobiológia területén elterjedtek. A legfontosabb biokémiai tulajdonságokat vizsgáló eljárásokat ma már kitek formájában árusítját a különböző forgalmazók, amelyek segítségével a fajok azonosítása gyorsan és pontosan elvégezhető.
A.) Amiláz kimutatása. Számos mikroorganizmus termel és bocsát ki a környezetbe amilázt. A keménytőt tartalmazó tápfelületre oltott mikroba által termelt enzim a táptalajba diffundál. Az enzim mikrobatelep körül a szénforrásként táptalajba bevitt keményítőt dextrinig illetve glükózig bontja. A folyamat végterméke befolyásolhatja az enzim indukcióját. Megjegyzés: A diffúziós zóna nagyságát több tényező befolyásolja, pl. a diffundáló vegyület molsúlya, polárossága, mennyisége, a diffúziós közeg (táptalaj) milyensége (ionerősség, pH). A diffúziós zóna nagysága alapján mennyiségi viszonyokra csak azonos anyagok és azonos körülmények esetén következtethetünk.
Mikroorganizmusok: Escherichia coli, Bacillus subtilis, Streptomyces albus, Streptomyces griseus, Saccharomyces diastaticus, Aspergillus niger 65 jelzésű törzse, Penicillium ramigena, Penicillium purpurogenum 48 órás tenyészetei T-1 illetve T-5 ferde agaron. Anyagok és eszközök:
• T9 és T9+ 2%glükóz Petri-csésze (1-1 db)
• 2ml fiz.só félkémcsőben (2 db)
• steril szemcseppentő
A gyakorlat menete: A mikroorganizmusokból fiziológiás sóoldatban tömény szuszpenziót készítünk, majd a T-9 illetve T-9+glükózt tartalmazó Petri-csészék felületére, csészénként 2 foltba 2 mikroorganizmus szuszpenzióját szemcseppentővel felvisszük. (Beszárítás nélkül óvatosan helyezzük a Petri-csészéket a termosztátba!) 30°C-on 72 órán keresztül inkubálunk. Az inkubációs idő után a lemezeket Lugol-oldattal (R-17) öntjük le. A keményítő kék színreakciót ad; a telepek körül színtelen "oldási udvar" látható.
46

B.) Zselatinbontó exoenzim ("zselatináz") kimutatása. A zselatinbontó képesség határozó bélyeg. Szénhidrátmentes táptalajon a mikroorganizmusok tekintélyes hányada bontja a zselatint, míg asszimilálható szénhidrát jelenlétében a bontás elmarad. A zselatináz kimutatását két módon végezzük el.
a) Zselatin elfolyósítás vizsgálata Mikroorganizmusok: Escherichia coli, Serratia marcescens, Pseudomonas aeruginosa, Proteus vulgaris, Staphylococcus aureus, Micrococcus luteus, Bacillus subtilis, Bacillus cereus var. mycoides, Streptomyces aureus, Streptomyces griseus 48 órás tenyészete T-1 ferde agaron.
Anyagok és eszközök: • T-10 magas táptalaj, tyndallozással sterilezve (2 db)
A gyakorlat menete: Oltótűvel szúrt tenyészetet készítünk a magas táptalajba, egy cső kontrollként beoltás nélkül marad. Szobahőmérsékleten 1 hétig inkubálunk.
b) Zselatináz kimutatása zselatin-agar táptalajon Mikroorganizmusok: Serratia marcescens, Proteus vulgaris, Micrococcus luteus, Bacillus cereus var. mycoides, Bacillus subtilis, Streptomyces griseus 48 órás tenyészete T-1 ferde agaron.
Anyagok és eszközök: 100 ml T-11 táptalaj, R-18 reagens, 3 db steril Petri-csésze.
A gyakorlat menete: A Frazier-táptalajt (T-11) felolvasztjuk és 3 db Petri-csészébe kiöntjük. Dermedés után felületre oltjuk a mikrobákat egyenes vonalú csíkhúzás mentén (csészénként 2-2 leoltás). 30°C-on 40 órán keresztül inkubálunk. Az inkubációs idő után "előhívjuk" a lemezeket. Sósavas HgCl2 oldatot (R-18) öntünk a csészékbe és pár percig állni hagyjuk rajta. A HgCl2 zselatinnal opálos csapadékot ad, az elbontott
zselatin helyén világos udvar látszik. Vigyázat: a kísérlet fokozottan fertőzésveszélyes! A HgCl2-ot tartalmazó Petri-csészéket alumínium kidobó edénybe NE tegyünk!
47

C.) Kazein hidrolízis A mikroorganizmusok a zselatin bontásához hasonlóan egyéb fehérjék bontására is képesek exoenzimjeik segítségével. Számos mikroorganizmus képes a tej fehérjéinek pl. a kazeinnek a bontására.
Mikroorganizmusok: Escherichia coli, Serratia marcescens, Pseudomonas aeruginosa, Proteus vulgaris, Staphylococcus aureus, Micrococcus luteus, Bacillus megaterium, Bacillus cereus var. mycoides 24 órás tenyészete T-1 ferde agaron.
Anyagok és eszközök: • T-12 Petri-csésze (2 db)
A gyakorlat menete: 2-2 mikroorganizmust oltsunk egy-egy T-12 csészébe csíkhúzással. 48 órás 30°C-os inkubálás után figyeljük az oldási zónákat, illetve öntsük le a felületet R-18 reagenssel. Vigyázat: a kísérlet fokozottan fertőzésveszélyes! A HgCl2-ot tartalmazó Petri-csészéket aluminium kidobó edénybe NE tegyünk!
48

10. gyakorlat: Eredmények és értékelés 1. Írjuk le, mely mikrobáknál tapasztaltunk keményítőbontást, melyik törzs esetében nem represszál a glükóz?
Bontás mértéke Tesztorganizmus
glükóz jelenlétében glükóz nélkül
2. Rajzoljuk le a zselatináz elfolyósítási formákat. 3. Írjuk le, mely mikrobák termelnek zselatinázt. 4. Foglaljuk össze táblázatba a kazeinbontásra vonatkozó adatokat!
Oldási zóna Mikroorganizmus
Előhívás nélkül reagenssel
49

11. gyakorlat: ÉLESZTŐGOMBÁK CUKORASSZIMILÁCIÓJÁNAK ÉS ERJESZTÉSÉNEK
VIZSGÁLATA
Élesztőgombák esetében az asszimiláció oxidatív körülmények közötti cukorfelhasználást, az erjesztés anaerob körülmények közötti, gázfejlődéssel járó cukorhasznosítást jelent. Az asszimilációs és erjesztési próbák során különböző cukrok hasznosítását vizsgáljuk és így jellemezzük a fajokat és törzseket. Az asszimilációt auxonográfiás módszerrel határozzuk meg (szénforrásmentes táptalajba lemezöntéssel rögzített sejtek a táptalaj felületére vitt cukrok körül szaporodni kezdenek, a pozitív próbát "opálosodás" jelzi), az erjesztési képességet a közeg pH-jának jelentős csökkenésével (indikátor színváltozása jelzi) és gázképződéssel bizonyítjuk. Az élesztőgombáknál redukált morfológiai bélyegeik miatt a rendszertani besorolást a biokémiai jellegek meghatározása nélkül nem tudjuk elvégezni.
Mikroorganizmusok: Saccharomyces cerevisiae ( "pékélesztő" jelzésű törzse), és Candida utilis 48 órás tenyészete T-5 ferde agaron. Anyagok és eszközök:
• 2 ml fiz.só félkémcsőben (2 db)
• szemcseppentő
• automata pipetta
A) Asszimiláció:
• T-17 táptalaj (frissen készítendő!) (2 db)
• diagnosztikus cukrok→ glükóz (D), galaktóz (G), szacharóz (S), maltóz (M), laktóz (L), raffinóz
(R), 1-arabinóz (Ar) és d-xilóz (X)
B) Erjesztés:
• 2,5ml steril T-17a tápoldat + cukrok Wassermann-csőben→ 0,5% glükóz (D), galaktóz (G),
szacharóz (S), maltóz (M), laktóz (L), raffinóz (R), 1-arabinóz (Ar) és d-xilóz (X)
• vaspar
A gyakorlat menete: A.) Asszimiláció: Az asszimilációs táptalajt (T-17) felolvasztjuk, majd 40°C-on vízfürdőzzük. A két élesztőgombából steril vízben szuszpenziót készítünk (1 kacs/2 ml), ebből steril pipettával 1-1 ml-t oltunk Petri-csészébe, majd a 40°C-os táptalajt ráöntjük és óvatosan jól elkeverjük. A táptalaj megdermedése után a lemezre 1-2 mg-nyi cukrot szórunk a 22. ábra szerint sorrendben a csésze széléhez közel. A glükózt mind a két csészébe bevisszük kontrollként. Az asszimilációs vizsgálatot a diagnosztikus cukrokon kívül 1-arabinóz és d-xilóz pentózokkal is elvégezzük.
50
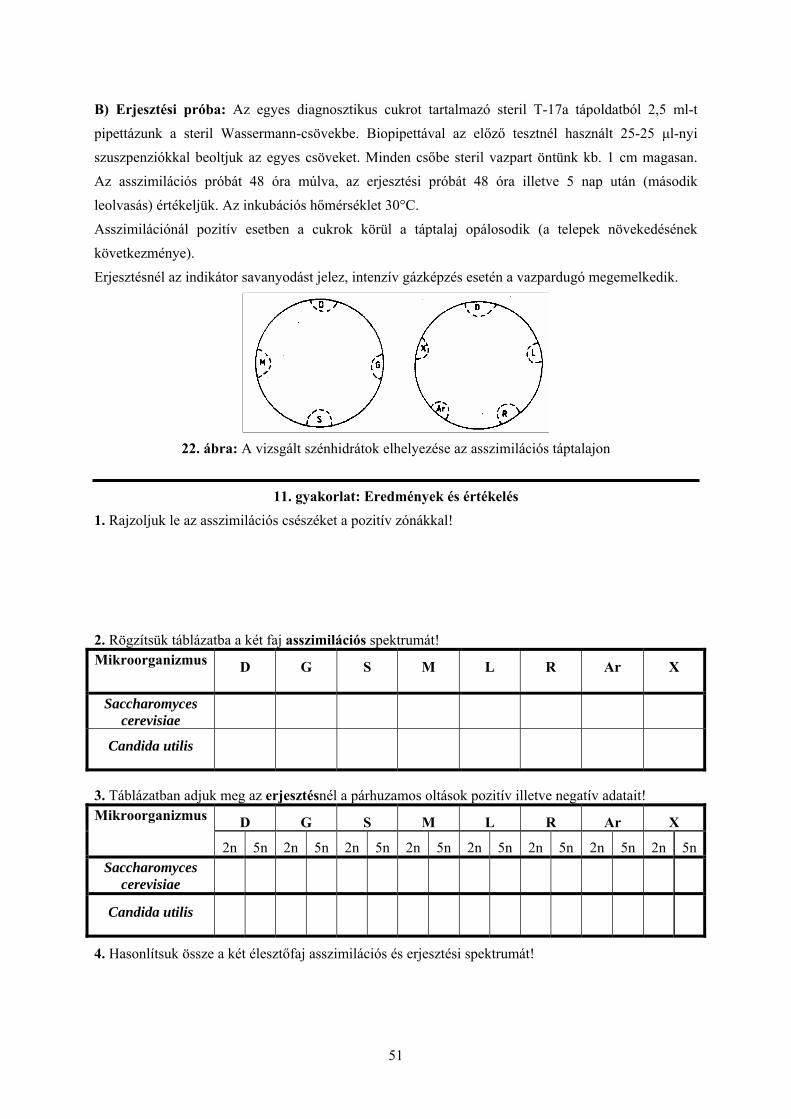
B) Erjesztési próba: Az egyes diagnosztikus cukrot tartalmazó steril T-17a tápoldatból 2,5 ml-t pipettázunk a steril Wassermann-csövekbe. Biopipettával az előző tesztnél használt 25-25 µl-nyi szuszpenziókkal beoltjuk az egyes csöveket. Minden csőbe steril vazpart öntünk kb. 1 cm magasan. Az asszimilációs próbát 48 óra múlva, az erjesztési próbát 48 óra illetve 5 nap után (második leolvasás) értékeljük. Az inkubációs hőmérséklet 30°C. Asszimilációnál pozitív esetben a cukrok körül a táptalaj opálosodik (a telepek növekedésének következménye). Erjesztésnél az indikátor savanyodást jelez, intenzív gázképzés esetén a vazpardugó megemelkedik.
22. ábra: A vizsgált szénhidrátok elhelyezése az asszimilációs táptalajon
11. gyakorlat: Eredmények és értékelés
1. Rajzoljuk le az asszimilációs csészéket a pozitív zónákkal! 2. Rögzítsük táblázatba a két faj asszimilációs spektrumát! Mikroorganizmus
D G S M L R Ar X
Saccharomyces cerevisiae
Candida utilis
3. Táblázatban adjuk meg az erjesztésnél a párhuzamos oltások pozitív illetve negatív adatait!
D G S M L R Ar X Mikroorganizmus
2n 5n 2n 5n 2n 5n 2n 5n 2n 5n 2n 5n 2n 5n 2n 5nSaccharomyces
cerevisiae
Candida utilis
4. Hasonlítsuk össze a két élesztőfaj asszimilációs és erjesztési spektrumát!
51

12. gyakorlat: BAKTÉRIUMOK ANTIBIOTIKUM ÉRZÉKENYSÉGÉNEK VIZSGÁLATA
Az antibiotikumok használatának széleskörű elterjedésével az antibiotikumokkal szembeni rezisztencia gyakoriságának növekedése figyelhető meg a baktériumok között. A kórokozó mikroorganizmusok különböző törzsei különböző mértékben gátolhatók egyes antibiotikumokkal, illetve gyakran megjelennek úgynevezett polirezisztens törzsek, amelyek egyszerre több antibiotikummal szemben rezisztensek. Ezen okoknál fogva a sikeres terápia szempontjából feltétlenül szükséges a humán megbetegedésekből izolált baktériumok rezisztenciájának meghatározása. A különböző módszerek közül a leggyakrabban a szemikvantitatív korongdiffúziós eljárást ("bioteszt-korong" módszer) alkalmazzák. A szűrőpapírra felvitt antibiotikum a táptalajba diffundál és a diffúzió gyorsaságától, a hatóanyag mennyiségétől és a mikroba érzékenységétől függően kisebb-nagyobb átmérőjű gátlási zónát hoz létre. Pontosabb eredményt kapunk, ha meghatározzuk hígításos módszerrel azt a legkisebb antibiotikum mennyiséget, amely 1 ml térfogatú oldatban (táptalajban) megakadályozza valamely baktérium szaporodását. Ezt az értéket minimális gátló koncentrációnak (minimum inhibitory concentration = MIC) nevezzük. Hasonlóan informatív lehet az antibiotikumok minimális baktériumölő koncentrációja (minimum bactericid concentration = MBC), az a mennyiség, amely az inokulumként bevitt baktériumok 99,9%-át elpusztítja.
Mikroorganizmusok: 1.) Pseudomonas aeruginosa, Escherichia coli, Serratia marcescens, Staphylococcus aureus, Streptococcus faecalis, Bacillus subtilis 48 órás tenyészete T-1 ferde agaron; 2.) Escherichia coli 48 órás tenyészete T-1 táptalajon Anyagok és eszközök:
1.) Bioteszt korongsorozattal:
• T-1 táptalaj Petri-csészében (2 db)
• 3 ml fiz.só félkémcsőben (2 db)
• antibiotikum tesztkorongok
• szűrőpapír, kidobóedény
2.) 1,28 mg/ml-es gentamycin törzsoldat (desztillált vízben), 50 ml steril fiziológiás sóoldat, 11 db steril kémcső, automata pipetták, 25 ml Mueller-Hinton-tápleves (T-40a).
A gyakorlat menete: 1.) Bioteszt korongsorozattal: a vizsgálandó baktériumokból 1-2 kacsnyi mennyiséget 3 ml fiziológiás sóoldatban szuszpendálunk. A mikroorganizmusok szuszpenziójával egy-egy csészét masszív oltással beoltunk, majd rövid ideig (5 perc) steril fülkében szárítjuk. A csészesorozatra
52
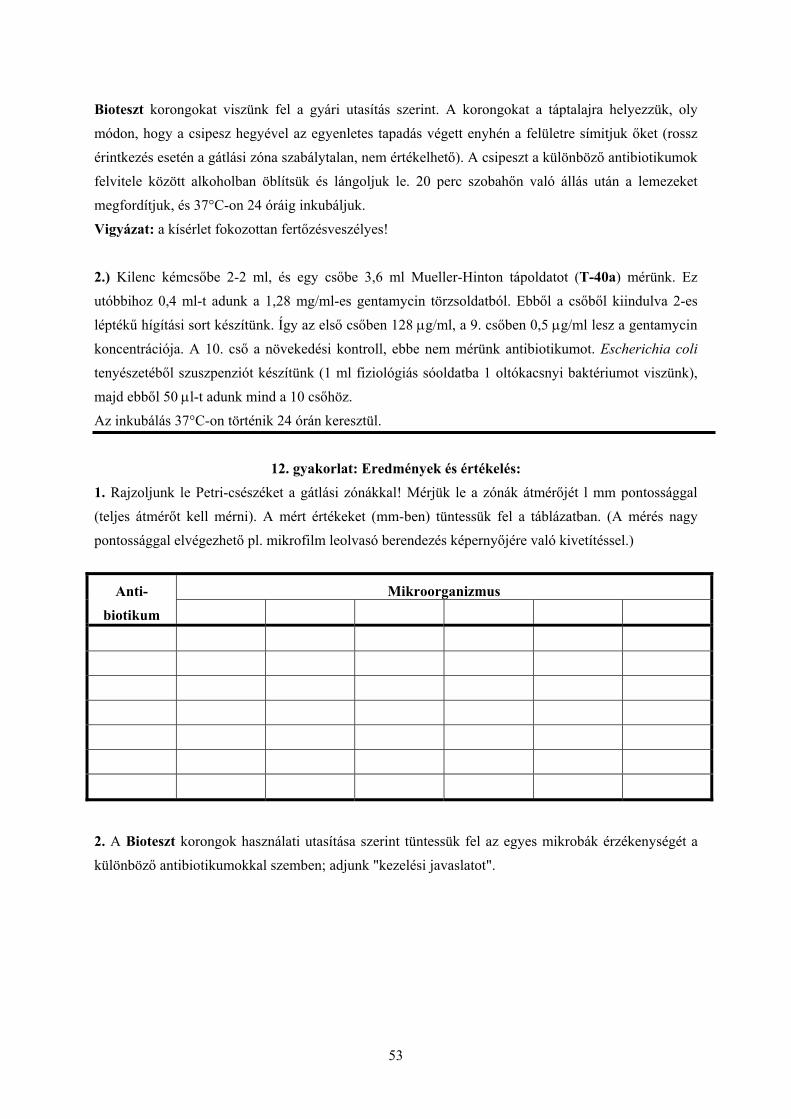
Bioteszt korongokat viszünk fel a gyári utasítás szerint. A korongokat a táptalajra helyezzük, oly módon, hogy a csipesz hegyével az egyenletes tapadás végett enyhén a felületre símitjuk őket (rossz érintkezés esetén a gátlási zóna szabálytalan, nem értékelhető). A csipeszt a különböző antibiotikumok felvitele között alkoholban öblítsük és lángoljuk le. 20 perc szobahőn való állás után a lemezeket megfordítjuk, és 37°C-on 24 óráig inkubáljuk. Vigyázat: a kísérlet fokozottan fertőzésveszélyes!
2.) Kilenc kémcsőbe 2-2 ml, és egy csőbe 3,6 ml Mueller-Hinton tápoldatot (T-40a) mérünk. Ez utóbbihoz 0,4 ml-t adunk a 1,28 mg/ml-es gentamycin törzsoldatból. Ebből a csőből kiindulva 2-es léptékű hígítási sort készítünk. Így az első csőben 128 µg/ml, a 9. csőben 0,5 µg/ml lesz a gentamycin koncentrációja. A 10. cső a növekedési kontroll, ebbe nem mérünk antibiotikumot. Escherichia coli tenyészetéből szuszpenziót készítünk (1 ml fiziológiás sóoldatba 1 oltókacsnyi baktériumot viszünk), majd ebből 50 µl-t adunk mind a 10 csőhöz. Az inkubálás 37°C-on történik 24 órán keresztül.
12. gyakorlat: Eredmények és értékelés: 1. Rajzoljunk le Petri-csészéket a gátlási zónákkal! Mérjük le a zónák átmérőjét l mm pontossággal (teljes átmérőt kell mérni). A mért értékeket (mm-ben) tüntessük fel a táblázatban. (A mérés nagy pontossággal elvégezhető pl. mikrofilm leolvasó berendezés képernyőjére való kivetítéssel.)
Mikroorganizmus Anti-biotikum
2. A Bioteszt korongok használati utasítása szerint tüntessük fel az egyes mikrobák érzékenységét a különböző antibiotikumokkal szemben; adjunk "kezelési javaslatot".
53

13. gyakorlat: AZ OZMOTIKUS NYOMÁS HATÁSA A MIKROORGANIZMUSOKRA
A táptalajok és tápoldatok különböző sókat, oldott szerves anyagokat tartalmaznak. Az oldott anyagok mennyisége alapján a mikroorganizmusok szempontjából a közeg lehet izotóniás, hypotóniás és hypertóniás. Az izotóniás értéktől való mindkét irányú eltérés károsan befolyásolja az életfolyamatokat, a sejtek lecsökkentik anyagcsere-folyamataikat, sőt el is pusztulhatnak. Ismeretesek konzervipari eljárások, melyekben az ozmotikus érték emelésével tartósítunk. A hipertóniás közeg meggátolhatja egyes készítmények mikroorganizmusok okozta romlását. A közeg ozmotikus értékének növekedésére a mikroorganizmusok különböző mértékben érzékenyek.
Mikroorganizmusok: Escherichia coli és Bacillus subtilis 24 órás tenyészete, T-l, Saccharomyces cerevisiae, Hansenula anomala és Penicillium frequentans 48 órás tenyészete T-5 ferde agaron. Anyagok és eszközök:
• kémcsőben 5 ml T-5a táptalaj 1, 10, 20, 40, 60% glükózzal (1-1 db)
• kémcsőben 5 ml T-5a táptalaj 1, 5, 10, 20, 30% NaCl-al (1-1 db)
• 2 ml fiz.só félkémcsőben (1 db)
• 100 ul automata pipetta
A gyakorlat menete: A mikroorganizmusok tenyészeteiből szuszpenziókat készítünk a steril fiziológiás sóoldatban (2 ml-ben). A koncentrációsorozatok minden tagjából 1-1 csövet beoltunk a vizsgálandó mikroorganizmusok szuszpenziójából. A beoltás biopipettával történik (0,1 ml csövenként). 30°C-on egy héten keresztül inkubálunk. A mikroorganizmusok szaporodását 48 óra és 7 nap múlva értékeljük.
54
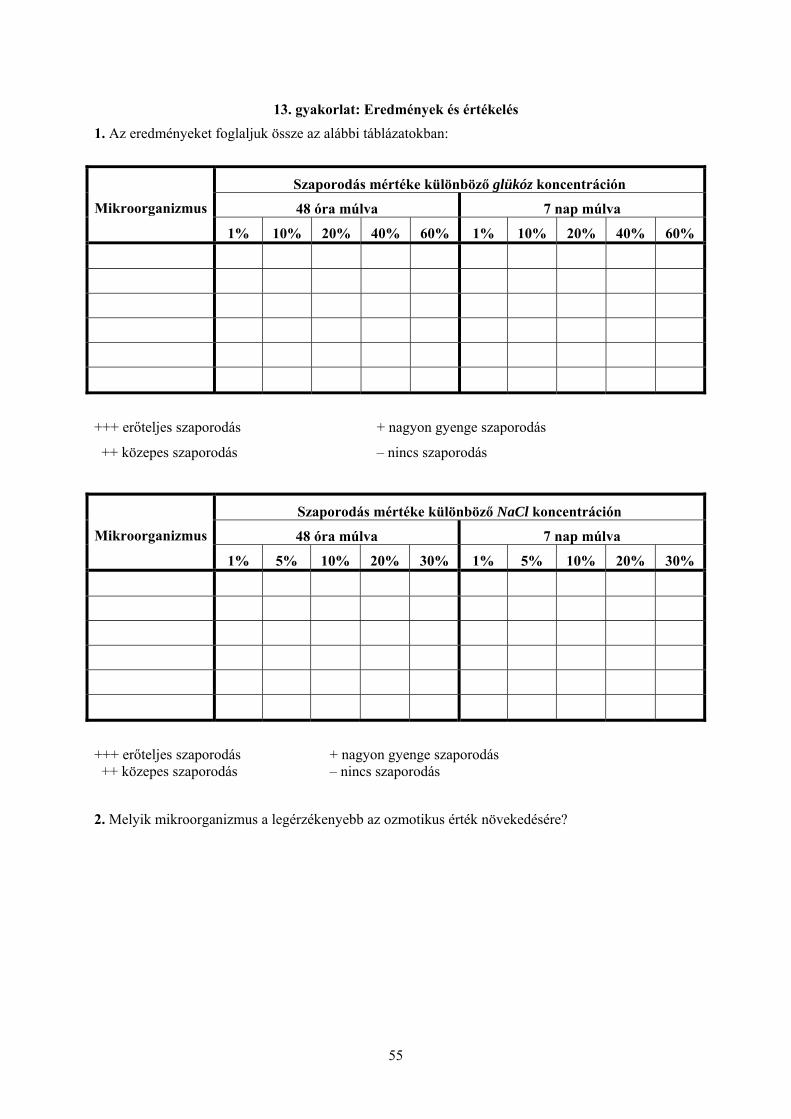
13. gyakorlat: Eredmények és értékelés 1. Az eredményeket foglaljuk össze az alábbi táblázatokban:
Szaporodás mértéke különböző glükóz koncentráción
48 óra múlva 7 nap múlva Mikroorganizmus
1% 10% 20% 40% 60% 1% 10% 20% 40% 60%
+++ erőteljes szaporodás + nagyon gyenge szaporodás
++ közepes szaporodás – nincs szaporodás
Szaporodás mértéke különböző NaCl koncentráción
48 óra múlva 7 nap múlva Mikroorganizmus
1% 5% 10% 20% 30% 1% 5% 10% 20% 30%
+++ erőteljes szaporodás + nagyon gyenge szaporodás ++ közepes szaporodás – nincs szaporodás
2. Melyik mikroorganizmus a legérzékenyebb az ozmotikus érték növekedésére?
55

14. gyakorlat: FÉMIONOK SZAPORODÁSGÁTLÓ HATÁSA
A nehézfémek pl. ezüst, réz, nikkel ionjai már igen kis koncentrációban toxikus hatásúak a mikrobákra. Egyes fémek enzimekben megtalálható fémekkel lép interakcióba, megakadályozva ezzel az enzim normális működését. Más fémek a DNS-hez kötődnek, s mutációk jönnek létre hatásukra. Jelen gyakorlatunkban az ionok a szilárd táptalajban diffundálnak és így gátlózónát hoznak létre.
Mikroorganizmusok: Micrococcus luteus, Bacillus cereus var. mycoides és Saccharomyces cerevisiae 48 órás tenyészete T-1 illetev T-5 ferde agaron. Anyagok és eszközök:
• T-1 Petri-csésze (2 db)
• 3ml fiz.só félkémcsőben (2 db)
• fémsó oldatok
• papírkorongok
• szűrőpapír, kidobóedény
A gyakorlat menete: A mikróbákból szabad szemmel már zavarosnak látszó szuszpenziót készítünk a 3 ml fiziológiás sóoldatot tartalmazó félkémcsövekbe, majd az egész szuszpenziót a T-1 táptalajt tartalmazó Petri-csésze felületére öntjük úgy, hogy az egész lemezt befedje. A felesleges, csészékről lecsurgatott szuszpenziót fertőtlenítő folyadékba öntjük. A csészéket steril oltófülkébe helyezzük. Nyitott tetővel ferde állásban a lecsurgó szuszpenziót szűrőpapír csíkokkal felszívatjuk (23. ábra), a lemezek felületét beszárítjuk. Ez az ún. "masszív oltás", mellyel egy összefüggő mikrobapázsitot képezünk a csésze teljes felszínén. Beszáradás után a fémsókat tartalmazó oldatokba belehelyezzük a papírkorongokat, majd csipesszel óvatosan a beoltott csészékre helyezzük őket. Egy csészére maximum 4 korongot helyezzünk egymástól egyenlő távolságba. 24 órán keresztül 30°C-on inkubálunk.
23. ábra: Masszív oltás után felület leszárítása
1. Petri-csésze fedő, 2. táptalaj, 3. szűrőpapírcsík
56

14. gyakorlat: Eredmények és értékelés 1. Táblázatba írjuk be hány mm a gátlási zóna nagysága. 2. Rajzoljuk le a Petri-csészéket, tüntessük fel a látottakat. 3. Írjuk le, melyik mikroorganizmus az érzékenyebb és melyik fém hoz létre nagyobb gátlási zónát.
Nehézfém oldat Mikroba
57

15. gyakorlat: NÖVÉNYI ANTIMIKROBIÁLIS VEGYÜLETEK HATÁSÁNAK
VIZSGÁLATA
A magasabbrendű növények által termelt, mikroorganizmusokra kis koncentrációban toxikus vegyületeket nevezzük fitoncidoknak. A magasabbrendű növények által termelt antimikrobiális anyagok, hasonlóan a szűkebb értelemben vett antibiotikumokhoz, a mikrobák kisebb-nagyobb csoportjaival szemben toxikusak, hatásuk szelektív. Gyakorlatunk során több növényi anyag hatását vizsgáljuk különböző mikroba csoporthoz tartozó fajokkal szemben.
Mikroorganizmusok:
Escherichia coli, Bacillus cereus var. mycoides, Hansenula anomala, Aspergillus niger 48 órás tenyészete T-1 illetve T-5 ferde agaron.
Anyagok és eszközök: 400 ml T-1 táptalaj (pH 7) 3% agartartalommal, Solanum laciniatum (levél), Cannabis sativa (nővirágzat pálhalevelei), Cynanchum vincetoxicum (gyökér), Cnicus benedictus (levél) őrlemény, vöröshagyma, dimetilszulfoxid (DMSO), 6 db kémcső, 4 db kémcső 5 ml steril vízzel, 12 db steril Petri-csésze, biopipetta (200 µl) steril hegyekkel, dörzscsésze, táptalaj lyukasztó eszköz. A gyakorlat menete:
Kémcsőben friss, szárított növényi őrlemények 0,5 g-jához 5 ml DMSO-t mérünk, a hagymából 0,5 g-ot eldörzsölünk, rövid ideig állni hagyjuk (prekurzor anyagból enzimatikusan keletkezik a hatóanyag, az aktiváló enzim a szövetek roncsolódásakor fejti ki hatását), ezután adunk hozzá 5 ml DMSO-t, 0,5 g hagymára pedig közvetlenül, eldörzsölés nélkül mérjük rá az oldószert. A növényi anyagokat forrásban levő vízfürdőben 10 percig extraháljuk, utána hagyjuk lehűlni a kivonatokat. A T-1 táptalajt felolvasztjuk, a pH-t ellenőrizzük, ha szükséges ismét 7-re állítjuk, (a hatóanyagok egy részének hatása erősen függ a pH-tól). 12 db steril csészébe lemezt öntünk. Masszív oltással a felületre visszük a 4 mikroorganizmus steril vizes szuszpenzióját (1 nagy kacs/5 ml). Minden mikrobával 3-3 lemezt oltunk. A lemezek felületének beszáradása után a lyukasztó eszközzel lemezenként 3 lyukat vágunk. A lyukakba a vizsgálandó növénykivonatok 0,1 ml mennyiségeit, illetve kontrollként a harmadik lyukba tiszta DMSO-t viszünk be. 48 órán keresztül 30°C-on inkubálunk.
58

15. gyakorlat: Eredmények és értékelés 1. Rajzoljuk le a csészéket a gátlási zónákkal! 2. Mérjük le a gátlási zónákat (a teljes átmérőt mérjük mm-ben)! Az adatokat (mindkét párhuzamost) táblázatban tüntessük fel! ––––––––––––––––––––––––––––––––––––––––––––––––––––––––––––––––– Mikroba E. coli B. cereus v. H. anomala A. niger Növény mycoides ––––––––––––––––––––––––––––––––––––––––––––––––––––––––––––––––– Solanum laciniatum ––––––––––––––––––––––––––––––––––––––––––––––––––––––––––––––––– Cannabis sativa ––––––––––––––––––––––––––––––––––––––––––––––––––––––––––––––––– Cynanchum vincetoxicum ––––––––––––––––––––––––––––––––––––––––––––––––––––––––––––––––– Cnicus benedictus ––––––––––––––––––––––––––––––––––––––––––––––––––––––––––––––––– Allium cepa eldörzsölés nélkül ––––––––––––––––––––––––––––––––––––––––––––––––––––––––––––––––– Allium cepa eldözsölve ––––––––––––––––––––––––––––––––––––––––––––––––––––––––––––––––– DMSO (kontrol)
59

16. gyakorlat: AZ UV-SUGÁRZÁS HATÁSA A MIKROORGANIZMUSOKRA
A bevezetőben szóltunk az UV-sugárzás csírátlanító hatásáról. A hatás maximuma 260-265 nm, egybeesik a nukleinsavak, pontosabban a pirimidin bázisok elnyelési maximumával. UV hatására a DNS kettős spirál azonos szálán lévő szomszédos pirimidinek közötti kovalens kötések kialakulásával dimerek (citozin-citozin, citozin-timin, timin-timin) jönnek létre. Legnagyobb jelentőségűek a timin-timin dimerek. Keletkezésük során a timin és a szemben levő adenin közti H-híd kötések gyengülnek. Ezt követően a timin C5-C6 atomjai közötti kettőskötés felszakad és a
szomszédos timin bázis azonos pozíciójú (5-6) szénatomjaival kovalens kötődéssel dimer jön létre. A dimerek a nukleinsavak normális szintézisének, replikálódásának menetét zavarják, amelynek következménye mutánsok létrejötte lehet. A mutáció következménye lehet letális, közönbös és előnyös (nagyon ritka). Esszenciális gének mutációja a sejt pusztulását eredményezi, ha a károsodást nem követi enzimatikus javítás. Az egyik lehetséges javító mechanizmus a 1) fotoreaktiváció, melynek során látható fény (300-600 nm) hatására a sejtben olyan enzim aktiválódik ("fotoliáz"), ami a timin dimereket összetartó ciklobutil gyűrű elhasításával a DNS helyes szerkezetét visszaállítja. Ez a javítómechanizmus baktériumokban fordul elő. Az ún. 2) kimetszéses javítás során először egy endonukleáz kihasítja a dimert, az így keletkezett rést egy exonukleáz kitágítja, majd a DNS polimeráz megszintetizálja a hiányzó szakaszt és ligáz összeköti a végeket. Az UV sugárzás károsító hatása függ:
• fajtól→ az egyes faj más-más érzékenységgel bírnak • a besugárzott tenyészet korától→ a fiatal, log fázisban lévő tenyészetek esetében nagyobb az
UV sugárzás károsító hatása, mivel ebben a fázisban a legintenzívebb a sejtek szaporodása, így a DNS állandó működésben van, lazább szerkezetű és ezáltal könnyebben támadható
• az UV lámpa teljesítményétől→ gyakorlatunkon ST 131 germicid lámpa (250 nm), 30 Wattos • az UV lámpa távolsága a tenyészettől→ gyakorlatunkon 45 cm • a besugárzás időtartamától
A gyakorlat során azért használjuk a Serratia marcescens-t, mert rendelkezik egy pigment (prodigiozin) termelésért felelős génnel, amelyet az UV sugárzással könnyen “elronthatunk”. A mutációt szenvedett génnel rendelkező sejtek pigmenttermelése hiányzik, a fotoliáz enzim működésének hatásra azonban a pigmenttermelés helyreállhat.
Mikroorganizmus: Serratia marcescens 24 órás tenyészete (620 nm-nél OD = 0,1 értékkel) T-1a tápoldat.
Anyagok és eszközök:
• T-1 Petri-csésze (5 db)
• 4,5 ml fiz.só félkémcsőben (4 db)
• 50 ul automata pipetta
• szélesztőbot, alkohol
60

16. gyakorlat: Eredmények és értékelés 1. Számoljuk meg az egyes kezelési idők utáni telepszámot és tüntessük fel a táblázatban. 2. Hasonlítsuk össze a sötétben és a fényben tartott sorozatot, tapasztalunk-e a telepszámokban eltérést? 3. Figyeljük meg, megjelennek-e, pigmentet nem termelő színtelen mutánsok?
Besugárzási idő (sec) Telepszám
Sötétben inkubált Fényen inkubált
0
1
2
5
10
61

A gyakorlat menete: A 24 órás Serratia marcescens tenyészetből négy lépcsőben 10-es léptékű hígítási sort készítünk. A IV. Hígítási lépcsőből 50 µl mennyiségeket viszünk fel a T-1 Petri-csésze felületére és üvegbottal szélesztünk. Steril oltófülkében beszárítjuk a beoltott táptalaj felületét. A Petri-csészék fedelét levesszük és UV-lámpa alá helyezzük őket és az egyes csészéket 0, 1, 2, 5, 10 másodpercig besugározzuk. Két sorozatot kezelünk. Egyik sorozatot kezelés után 30°C-os sötét termosztátba helyezzük, a másikat DNS-reparálódás céljából 6000 lux erősségú F-csöves megvilágításnak tesszük ki. 30°C-on 5 napon keresztül inkubálunk.
62

16. gyakorlat: Eredmények és értékelés 4. Ábrázoljuk grafikonon a sejtpusztulás mértékét. (A koordináta rendszer helyére ragasszunk be milliméter papírt, a tengelyeket lássuk el a megadott vázlat jelölésével.)
63

17. gyakorlat: MUTAGENITÁSI VIZSGÁLAT AMES-TESZTTEL
A tesztet eredetileg Ames dolgozta ki 1973-ban. Ma már a módosított változatát az ún. Ames-tesztet
világszerte alkalmazzák, hogy meghatározzák bizonyos új kemikáliák mutagén potenciálját. A
módszer végrehajtására nemzetközi szabványt dolgoztak ki, így új gyógyszerek és vegyszerek
tesztelésére is általánosan használt eljárás.
A tesztben alkalmazott Salmonella typhimurium törzsben a mutagenitás vizsgálatára specifikus,
mesterséges genetikai módosításokat hoztak létre: a) exciziós repair mechanizmus inaktivált, b)
permeábilis membrán c) hisztidin auxotrófia. Az excíziós repair mechanizmus inaktívlása
megakadályozza a bekövetkező mutációs változásoijavását, a permeábilis membrán lehetővé teszi az
anyagok bejutását a sejtbe.
Az eljárás lényege, hogy a hisztidin auxotróf törzs nem képes növekedni a minimál táptalajon. Ha a
kísérletben vizsgált anyag mutagén, akkor a hisztidin szintéziséért felelős génben létrejöhet mutáció és
a mutációt szenvedett sejtek prototrófok lesznek, azaz képesek növekedni minimál táptalajon. Minél
több a visszamutálódott sejt annál erősebb mutagén a tesztelt anyag.
Mikroorganizmusok:
Salmonella typhimurium TA 98 (∆ bio chlD uvrB gal; rfa; pKM101) törzsének 48 órás ferde agaros
tenyészetéből készített overnight kultúra.
Anyagok és eszközök:
• Oxoid nurient broth No.2 tápoldat
• Ames-puffer oldat
• frissen készített meleg top agar kiegészítve 0,05 mM hisztidinnel és biotinnal
• GM agar tartalmú Petri-csésze
• steril félkémcsövek
• 4-nitro-o-feniléndiamin törzsoldat (kezelés koncentráció: 2,5 µg/csésze);
• 0,1 mM CdCl2/csésze, 0,5 mg etídium-bromid/csésze, 0,025mM kálium-dikromát/csésze
A gyakorlat menete:
A 2 napos ferde agaros tenyészetről 2 kaccsal beoltunk 20 ml Oxoid nutrient broth No.2 tápoldatot,
majd 4 órát szobahőn tartjuk és ezután 11-14 órát (overnight) rázatjuk (100 rpm) 37°C-on.
Félkémcsövekbe a következő anyagokat mérjük össze:
• 2 ml felolvasztott 43-45ºC-os top agar táptalaj kiegészítve 0,05 mM hisztidinnel és biotinnal
• 500 µl Ames-puffer
• 50 µl vizsgált anyagminta
• 100 µl Salmonella typhimurium egyéjszakás előkultúra
64

Az összekevert elegyet GM agar tartalmú Petri-csészékre öntjük, majd dermedés után a csészéket 2
napig inkubáljuk 37°C-on.
Megjegyzés: célszerű a vizsgált anyag törzsoldatából 10x hígítási sort készíteni.
17. gyakorlat: Eredmények és értékelés
1. Számoljuk meg a revertáns telepek számát a kontrol és a vizsgált anyagot tartalmazó csészéken!
65

18. gyakorlat: PETIT-MUTÁNS INDUKCIÓ
A Saccharomyces cerevisiae kitűnő modell a mitokondriumok biogenezisének
tanulmányozásához, a légzési lánc funkcionális komplexeinek elemzéséhez. Ez főként annak
köszönhető, hogy a gomba életben marad akkor is, ha mitokondriumai a mtDNS-en
bekövetkező mutáció vagy a kedvezőtlen külső környezeti feltételek mellett nem működnek.
A mitokondriális genetikai tanulmányok során háromféle mutáns kategóriát állapítottak meg:
• pont mutációk vagy kisméretű deléciók következtében fellépő, fenotípikusan légzés
deficiens mutációk (mit-, syn-)
• pont mutációk következtében kialakuló, normális légzésű, de antibiotikumokra vagy
más gátló anyagokra rezisztens mutációk (antR)
• nagy deléciók és a megmaradó szentek amplifikációjával keletkező légzés deficiens
ún. petit (ρ-) mutációk
• extrém esetben a teljes mitokondriális genom elveszhet (ρ0).
A kísérlet során a vizsgált anyag jelenlétében inkubáljuk 24 órán át Saccharomyces cerevisiae
(a ura3 ade1) tenyészetét. Ha az anyag képes mutációt indukálni a mikroorganizmus
mitokondriális DNS-ben, légzésdeficiens ún. petit-mutáns telepek jönnek létre. Ha a kezelt
tenyészetből kioltjuk a sejteket komplett (2xYPD) táptalajra, majd utána átreplikázzuk a
légzésdeficiens mutánsok számára nem fermentálható szénforrást (glicerin) tartalmazó YPG
táptalajra, összehasonlítva a növekedést a csészék között, megkapjuk a petit-mutánsokat. A
petit-mutáns sejtek a 2xYPD táptalajon képesek növekedni, a YPG táptalajon azonban nem.
Mikroorganizmusok:
Saccharomyces cerevisiae (a ura3 ade1)
Anyagok és eszközök:
• 2xYPD Petri-csésze
• YPG Petri-csésze
• 4x YPD tápoldat
• fiziológiás só
• 10 mg/ml etídium-bromid oldat
• 12% DMSO, 14 % DMSO oldat
• alkohol, üvegbot, 50 ml-es lombik, félkémcsövek
66

A gyakorlat menete:
2 napos 2xYPD ferde agaros táptalajon tenyésztett törzsből 2 kaccsal beoltunk 5 ml folyékony
2xYPD oldatot és rázatjuk 30°C -on, 14 órán át, majd centrifugáljuk 3000 rpm-el, 5 percig. A
sejteket felvesszük 2 ml fiziológiás sóoldatban. Bürker-kamrával sejtszámolást végzünk és a
sejtszámot 107/ml-re állítjuk be. Összemérjük a következőket 50 ml lombikba: 2,5 ml 4xYPD
és 2,5 ml a vizsgálni kívánt anyag oldatából (amiből késztünk eredeti, 10x, 100x hígítást is).
Etídium-bromid esetén 5 ml 2xYPG tápoldatba oltsunk 50 µl 10 mg/ml etídium bromid
oldatot. A tenyészeteket 24 órán át rázatjuk, ismét centrifugálunk, 3000 rpm-el, 5 percig.
Felvesszük a sejteket 5 ml fiziológiás sóoldatba és Bürker-kamrával sejtszámolást végzünk. A
sejtszámot 103/ml-re állítjuk. 100 µl kiszélesztünk 2xYPD táptalajra és 3-5 napig 30 °C –on
inkubáljuk.
A kinőtt telepeket átreplikázzuk YPG táptalajra és ismét 3-5 napig 30 °C –on inkubáljuk.
18. gyakorlat: Eredmények és értékelés
1. Állapítsuk meg, hogy hány százalékos a petit mutáns telepek elfordulása a különböző indukáló
ágensek hatására.
Vizsgált anyag 2xYPG csésze
(telepszám)
YPG csésze
(telepszám)
Petit-mutáns
előfordulás (%)
6% DMSO
7% DMSO
Etídium-bromid
67

19. gyakorlat: AZOTOBACTER AGILE TESZT
Az Azotobacter agile laboratóriumi tenyészete alkalmas a hulladékban lévő mérgező szerves és
szervetlen komponense talajbaktériumokra kifejtett hatásának jelzésére. A hulladék ökotoxikus hatását
a tesztbaktérium 28ºC hőmérsékleten meghatározott inkubációs idő (48 óra) alatti szaporodás, illetve
szaporodás gátlása alapján értékeljük. A szaporodást vörös szín megjelenése jelzi, amely a táptalajhoz
adott trifenil-tetrazóum-klorid dehidrogenáz enzim jelenlétében redukálódott formájától, trifenil-
formazántól származik.
Mikroorganizmusok:
Azotobacter agile két hétnél nem idősebb tenyészete
Anyagok és eszközök:
• mikrotitrátorlemez, 18 cm átmérőjű Petri-csésze
• félkémcsövek
• Fjodorov-féle táptalaj (teszt), frissen készített
• 2, 3, 5,-trifenil-2H tetrazolium-orid (TTC) 1% vizes oldata
• 0,85% NaCl oldat
• 1 mg/ml koncentrációjú CuSO4 oldat, 0,005 mg/ml HgCl2, 0,01 mg/ml NiSO4, 0,04 mg/ml ZnSO4,
0,05 mg/ml CdCl2, 0,1 mg/ml KCr2O4, 0,2 mg/ml Pb(NO3)2
A gyakorlat menete:
A vizsgált anyagokból hígítási sort készítünk a 24 ábra alapján, desztillált vízzel. A mikrotitrátor
lemez mélyedéseibe bemérjük (50 µl) a hígításokat (mindegyiket 3 párhuzamosban), utolsónak
bemérve a negatív kontrollként szolgáló CuSO4 oldatot s végül pozitív kontrollt a desztillált vizet. A
vizsgálandó anyagokat váltott pipettavéggel vagy hígabbtól a töményebb felé egy heggyel mérjük be.
A negatív kontrollt mindig külön heggyel vigyük fel.
Sorszám Hígítófolyadék (ml) Minta Hígítási fok
1. 4 4 ml eredetiből 2x
2. 8 2 ml eredetiből 5x
3. 9 1 ml eredetiből 10x
4. 9 1 ml a 2x hígításból 20x
5. 9 1 ml az 5x hígításból 50x
24. ábra: Vizsgált kísérleti anyagok hígítása.
Azotobaer agile ferde agaros tenyészetére 5 ml 0,85% NaCl oldatot öntünk, kaccsal lesodorjuk a
sejteket, vortexeljük pár másodpercig. A 100 ml Fjodorov-féle (teszt) táptalajhoz hozzáadunk 1 ml
1%-os TTC oldatot, majd beoltjuk az Azotobacteres szuszpenzióval (5 ml). Az így elkészített
68
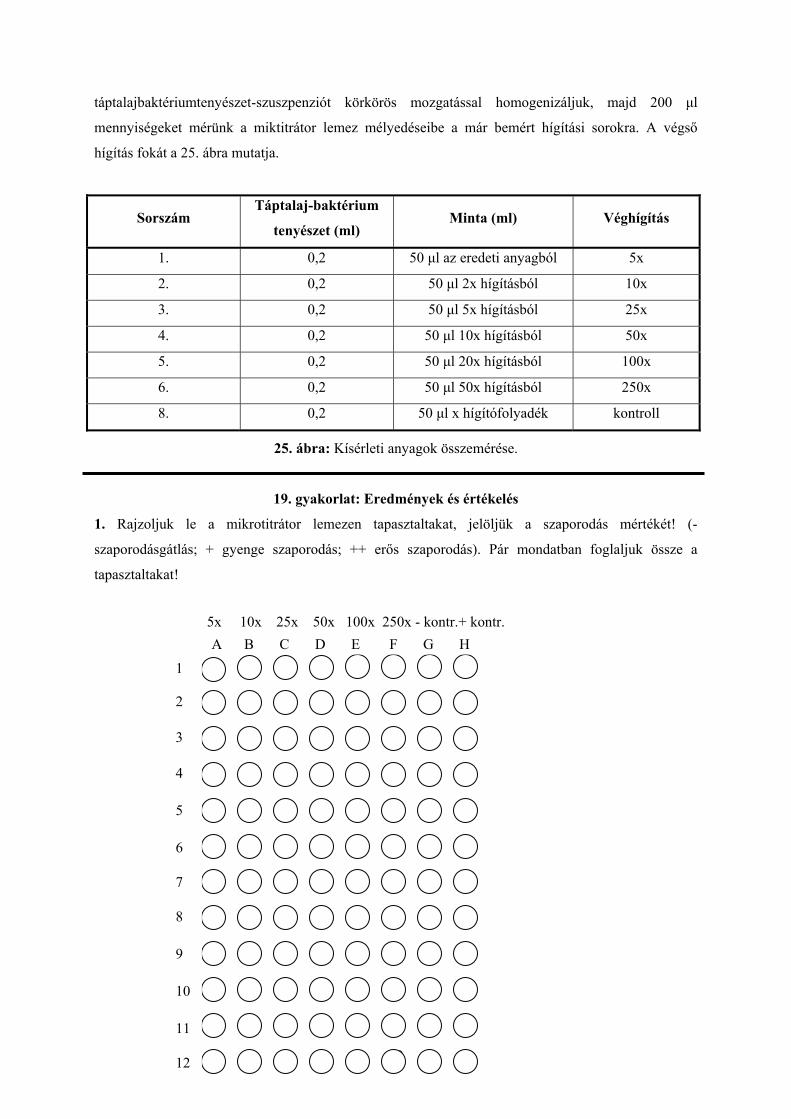
táptalajbaktériumtenyészet-szuszpenziót körkörös mozgatással homogenizáljuk, majd 200 µl
mennyiségeket mérünk a miktitrátor lemez mélyedéseibe a már bemért hígítási sorokra. A végső
hígítás fokát a 25. ábra mutatja.
Sorszám Táptalaj-baktérium
tenyészet (ml) Minta (ml) Véghígítás
1. 0,2 50 µl az eredeti anyagból 5x
2. 0,2 50 µl 2x hígításból 10x
3. 0,2 50 µl 5x hígításból 25x
4. 0,2 50 µl 10x hígításból 50x
5. 0,2 50 µl 20x hígításból 100x
6. 0,2 50 µl 50x hígításból 250x
8. 0,2 50 µl x hígítófolyadék kontroll
25. ábra: Kísérleti anyagok összemérése.
19. gyakorlat: Eredmények és értékelés
1. Rajzoljuk le a mikrotitrátor lemezen tapasztaltakat, jelöljük a szaporodás mértékét! (-
szaporodásgátlás; + gyenge szaporodás; ++ erős szaporodás). Pár mondatban foglaljuk össze a
tapasztaltakat!
A B C D E F G H69
1
2
3
4
5
6
7
8
9
10
11
12
5x 10x 25x 50x 100x 250x - kontr.+ kontr.

20. gyakorlat: TALAJTOXICITÁS VIZSGÁLAT AZOTOBAKTERES TALAJBLOKK
MÓDSZERREL
A talajokban általánosan elterjedt és toxikus anyagokkal szemben magas fokban érzékeny,
Azotobacter chroococcum teszt-mikroorganizmussal beoltott nitrogénmentes tápagarlemez felületére
nedves talajból préselt talajblokkokat helyezünk. A talajblokkból a toxikus anyagok a tápagarba
diffundálnak é abban a zónában, ahol a gátló hatású anyagok toxikus koncentrációban vannak jelen, a
tesztmikroorganizmus nem növekedik. A toxikusságot a gátlási gyűrű mm-ekben mért átmérőjével
jellemezzük.
Mikroorganizmusok:
Azotobacter chroococcum egy hétnél nem idősebb tenyészete
Anyagok és eszközök:
• Fjodorov-féle táptalaj (kálcium-karbonátból az eredeti recept 1/5 tartalmazza), frissen készítve
• fiziológiás sóoldat (3 ml)
• steril Petri-csésze
• talajminták (a talajmintákat a felhasználásig 4ºC-on tároljuk)
• talajblokk készítő présszerszám
A gyakorlat menete:
Ferde agarra oltott Azotobacter chroococcum kultúrából steril fiziológiás sóoldattal készítünk
sejtszuszpenziót, úgy hogy ferde agaros tenyészetére 3 ml 0,85% NaCl oldatot öntünk, kaccsal
lesodorjuk a sejteket, vortexeljük pár másodpercig. Laminár box alatt lemezöntést végzünk: bemérünk
a sejtszuszpenzióból 1 ml-t Petri-csészébe és a steril még meleg Fjodorov-féle táptalajt (kálcium-
karbonátból az eredeti recept 1/5 tartalmazza) ráöntjük a szuszpenzióra, majd finoman mozgatva a
csészét elkeverjük a sejteket. Az agar dermedése után közvetlenül megkezdjük a talajblokkok
elhelyezését. A talajminták nedvességtartalmát steril desztillált víz hozzáadásával a kapilláris
vízkapacitás 60%-nak megfelelő értékre állítjuk. A talajminták egy részét közvetlenül felhasználjuk a
vizsgálatra, a másik részét autoklávozzuk. Ezek a steril minták kontrollként szolgálnak a toxikus,
illetve serkentő hatás megítéléséhez és részben információt adnak arról, hogy a gátló anyagok
hőstabilak-e. A talajblokkokat présszerszám (26. ábra) segítségével elhelyezzük a táptalaj felszínén. A
csészéket 16 órára 4ºC-os hűtőbe helyezzük, itt a baktérium nem növekedik, de a talajból
kioldódhatnak a toxikus anyagok és a táptalajba diffundálnak. Inkubálásra a csészéket 28ºC-os
termosztátba helyezzük 40-48 órára.
70

26. ábra: Présszerszám.
Minden vizsgálat során készítsünk kontroll csészéket is: 1) Azotobacter tenyészet nélkül talajblokk
ráhelyezésével (táptalaj steril kontrollja) és 2) Azotobacter szuszpenziójával talajblokk nélkül
(Azotobacter növekedésének ellenőrzése).
A talajtoxicitást a gátlási gyűrűátmérőjével jellemezzük. A teljes gátló zóna és a talajblokk
átmérőjének ismeretében a gátlási gyűrű átmérőjét az alábbi számítás alapján határozzuk meg:
ahol
d: gátlási gyűrű átmérője
a: teljes gátlózóna átmérője
b: a talajblokk átmérője
Toxikusnak tekintjük azokat a talajokat, amel
20. gyakorlat:
1. Számoljuk ki a gátlási zónákat és vonjuk le
d = a-b
2
yeknél a gátlási gyűrű átlagos átmérője eléri az 1mm-t.
Eredmények és értékelés
a következtetéseket az eredményekből.
71

21. gyakorlat: PSEUDOMONAS PUTIDA NÖVEKEDÉSGÁTLÁSI TESZTJE
A módszer a felszíni, a talaj- és a szennyvíz Pseudomonas putida (talajban és elszíni vizekben
mindenütt előfordul) baktériumra gyakorolt gátló hatásának meghatározását teszi lehetővé. A
sejtnövekedés mérésével meghatározzuk a teszt minta különböző hígításainál és összehasonlítjuk egy
olyan kultúra sejtnövekedési adataival, amelyet azonos körülmények között vizsgáltunk a tesztanyag
nélkül. A sejtkoncentrációt optikai sűrűségként határozzuk meg a 16 órás tenyészidőszak után.
Mikroorganizmusok:
Pseudomonas putida legfeljebb 7 napos ferde agaros tenyészete
Anyagok és eszközök:
• I., II., III., IV: jelű oldat
• 3, 5 diklór-fenol
• vízminta
• WTW nefelométer
A gyakorlat menete:
Pseudomonas putida ferde agaros tenyészetére öntsünk rá 5 ml steril előkultúra tápoldatot, majd
oltókaccsal sodorjuk le a sejteket az agar felszínéről. Pipettázzuk át a szuszpenziót egy 50 ml
lombikba és hígítsuk addig előkultúra tápoldattal 100 FNU zavarosságértékig. FNU-mérés helyett
optikai denzitást is mérhetünk: A610=0,02 → 10 FNU, A610=0,1 → 50 FNU, A610=0,2 → 100 FNU.
Oltsunk be steril 250 ml lombikban 90ml előkultúra tápközeget 10 ml sejtszuszpenzióval, majd 5 órán
keresztül rázassuk 23ºC-on rázassuk. Az inkubálási idő lejárta után hígítsuk a szuszpenziót 250 ml
lomiban a tesztkultúra táptalajjal a 27. ábra szerint.
Törzsolatok Hígítóvíz Tesztanyag
II. III. IV.Előkultúra Végső térfogat
Hígítási fok
ml ml ml ml ml ml ml
kontroll 80 0 2,5 2,5 5 10 100
2 30 50 2,5 2,5 5 10 100
4 55 25 2,5 2,5 5 10 100
8 67,5 12,5 2,5 2,5 5 10 100
16 73,7 6,3 2,5 2,5 5 10 100
32 76,9 3,1 2,5 2,5 5 10 100
64 78,5 1,5 2,5 2,5 5 10 100
27. ábra: Kísérleti anyagok összemérése.
Inkubáljuk 16 órát rázatva 23ºC-on. Végezzük el a zavarosságmérést.
72

A teszt érvényesnek tekinthető, ha a kontrollhoz használt inokulum (5FNU) a teszt alatt legalább 60x-
ra szaporodott és ha a referenciavegyületként használt 3,5-diklór-fenol EC50 értéke 10 mg/l és 30 mg/l
közé esik.
A sejtszaporodás-gátlás (I) százalékos értékét a következőképp számolhatjuk ki:
I = Bc - Bn
Bc – B00
ahol
I: a sejtszaporodás-gátlás, százalékban kifejezve
Bn: biomassza mért zavarosság a tesztidőszak végén a tesz
Bc: a biomassza mért zavarossága a tesztidőszak végé a ko
B0: a biomassza kezdeti zavarosságmérésekor kapott érték
21. gyakorlat: Eredménye
1. Számoljuk ki a fenti képlet alapján a szaporodás-gátlás
Hígítási érték Mérési adatok
Kontroll
2
4
8
16
32
64
2. Ábrázoljuk koordináta rendszerben mindegyik hígítá
hígítási faktorral szemben. Az EC10 és EC50 értékek
valamint az 50% ordinátaértékeknél az abszcisszával pá
való metszéspontjánál található).
73
x 10
tminta n-edik koncentrációjára
ntrollban
, t0 időpontban, a kontrollban.
k és értékelés
t és vezessük z eredményeket a táblázatba.
Átlag Növekedés-gátlás (%)
sra kiszámolt gátlási értéket (I) a megfelelő
et állapítsuk meg (az egyenesnek a 10%,
rhuzamosan szerkesztett egyenes vonalakkal

IV. TÁPTALAJOK
T-1 : Kiegészített hústáptalaj 1000 ml-re
• 4 g húskivonat (beef extract) • 4 g pepton • g glükóz • 1 g élesztőkivonat (yeast extract) • 20 g agar (2%)
→illetve más koncentrációknál az agar megfelelő %-ban (az agartartalom az agar minőségének
függvénye, a jegyzetben megadott viszonylag magas értékek (3%) szálas agarra vonatkoznak).
T-1a : Kiegészített hústápleves
Mint T-1, agar nélkül.
T-2 : Hústáptalaj 1000 ml-re
• 4 g húskivonat • 4 g pepton • 1 g élesztőkivonat • 3 % agar
T-3 : Czapek-Dox minimál táptalaj 1000 ml-re
• 2 g NaNO3 • 0,5 g MgSO4 x 7 H2O • 0,5 g KCl • 0,5 g K2HPO4 • 30 g szacharóz • 10 mg ZnSO4 x 7H2O • 10 mg FeSO4 x 7H2O • 3 mg CuSO4 • 3% agar
T-4 : Vitaminos szintetikus táptalaj 1000 ml-re
• 5 g (NH4)2SO4 • 1 g KH2PO4 • 0,5 g MgSO4 x 7 H2O • 10 g glükóz • 3 % agar • kiegészítve 1 ml Wickerham-féle vitaminoldattal.
74

Wickerham-féle vitaminkeverék: 100 ml desztillált vízre
0,2 mg folsav 20,0 mg p-aminobenzoesav
0,2 mg biotin 20,0 mg riboflavin
40,0 mg Ca-pantotenát 40,0 mg aneurin.HCl (tiamin)
200,0 mg inozitol 40,0 mg piridoxin.HCl
40,0 mg niacin
T-4a : Vitaminos szintetikus tápoldat
Mint a T-4, agar nélkül.
T-5 : Élesztőkivonat táptalaj 1000 ml desztillált vízre
• 5 g élesztőkivonat • 10 g glükóz • 3 % agar
T-5a : Élesztőkivonat tápoldat
Mint T-5, agar nélkül.
T-7a : Tápoldat Lactobacillus-ok tenyésztéséhez 1000 ml deszt. vízre
• 3,0 g glükóz • 2,27 g Na-acetát x 3H2O • 1,36 g KH2PO4 • 0,23 g MgSO4 x 7H2O • 0,05 g MnSO4 x 2H2O • 0,02 g FeSO4 x 7H2O • 0,02 g (NH4)2SO4 x 6H2O • 7,5 g Trypton • 2,5 g élesztőkivonat
pH = 6,5
T-8a : Metilénkék tápleves 1000 ml-re
• 4 g húskivonat • 4 g pepton • 1 g élesztőkivonat • 100 mg metilénkék
T-9 : Keményítős táptalaj 1000 ml-re
• 10 g vízoldékony keményítő • 5 g élesztőkivonat • 3 % agar
75

T-10 : Tápzselatin 1000 ml-re
• 4 g húskivonat • 4 g pepton • 1 g élesztőkivonat • 200 g zselatin
A zselatinos táptalajokat áramló gőztérben, nyomás nélkül sterilezzük!
T-11 : Frazier-féle zselatin-agar 1000 ml-re
• 4 g húskivonat • 4 g pepton • 4 g zselatin • 1 g élesztőkivonat • 3 % agar
T-12 : Tejtáptalaj 1000 ml-re
• 2 g pepton • 1 g glükóz • 2 g Na2HPO4 • 10 g NaCl • 3 % agar
Autoklávozás után 90 ml alaptáptalajhoz 10 ml 10-15 percig 5000 x g-n centrifugált majd nyomás
alatt hősterilezett tejet adunk.
T-17 : Asszimilációs (auxonográfiás) táptalaj 1000 ml-re
• 5,0 g (NH4)2SO4 • 1,0 g KH2PO4 • 0,5 g MgSO4 x 7H2O • 1 ml Wickerham-féle vitaminoldat • 1,5 % mosott, vagy Bacto agar
(Agar mosása: kimérjük az agart abba a lombikba, amelyikben a táptalajt meg akarjuk autoklávozni.
12 óráig csapvízben mossuk, majd friss desztillált vízzel többször átöblítjük.)
T-17a : Tápoldat erjesztési próbához 1000 ml-re
• 5,0 g (NH4)2SO4 • 1,0 g KH2PO4 • 0,5 g MgSO4 x 7H2O • 1ml Wickerham-féle vitaminoldat • 0,5% a megfelelő diagnosztikus cukorból • 10 mg brómkrezolbíbor indikátorfesték
A pH-t 5%-os NaOH-dal semlegesre állítjuk, hogy a tápoldat éppen lilaszínű legyen!
76

T-40 : Mueller-Hinton agar 1000 ml-re
• 5 g marhahúskivonat • 17,5 g savval emésztett kazein (pl. Casamino Acid) • 2 g glükóz • 15 g agar
pH 7,4
T-40a : Mueller-Hinton tápleves
Ugyanaz mint T-40 agar nélkül.
Top agar kiegészítve hisztidinnel és biotinnal (Ames-teszt), 1000 ml-re
• 6 g NaCl
• 100 ml hisztidin/biotin oldat (0,5 mM)
• 6 g agar
Hisztidin/biotin oldat (0,5 mM) ), 1000 ml-re (Ames-teszt)
• 124mg D-biotin
• 96 mg L-hisztidin·HCl
Nutrient broth 1000 ml-re(Ames-teszt)
• 25 g Oxoid nutrient broth
Ames-puffer (nátrium-foszfát puffer) 1000 ml-re (Ames-teszt)
• 13,8 g NaH2PO4·H2O
• 14,2 g Na2HPO4·H2O
pH: 7,4 0,1M Na2HPO4·H2O oldattal beállítva
GM agar 1000 ml-re (Ames-teszt)
• 20 ml 50xVB oldat
• 50 ml 10% glükóz oldat
• 15 g agar
Vogel-Bonner (VB) oldat 1000 ml-re (Ames-teszt)
• 10 g MgSO4·H2O
• 100 g citromsav, monohidrát
• 500 g KH2PO4
• 175g Na2NH2PO4·4H2O
77

2xYPD táptalaj 1000 ml-re
• 10 g élesztőkivonat
• 20 g pepton
• 20 g glükóz
• 20 g agar
2xYPD tápoldat 1000 ml-re
• ua. mint a 2xYPD táptalaj agar nékül
4xYPD tápoldat 1000 ml-re
• 20 g élesztőkivonat
• 40 g pepton
• 40 g glükóz
YPG táptalaj 1000 ml-re
• 1% élesztőkivonat
• 2% g pepton
• 2,5% g glicerin
Fjodorov-féle táptalaj 1000 ml-re (törzsfenntartáshoz)
• 20 g glükóz
• 0,3 g dikálium-hidrogén-foszfát
• 0,2 g kalcium-hidrogén-foszfát, kritályvizes
• 0,2 g dikáum-foszfát
• 0,3 g magnézium-szulfát, kristályvizes
• 0,5 g nátrium-klorid
• 0,01 g vas(III)klorid, kristályvizes
• 5 g kálcium-karbonát
• 20 g agar
• 1 ml nyomelemkeverék
pH: 7,2-7,4
Fjodorov-féle táptalaj 1000 ml-re (teszthez)
• 20 g glükóz
• 0,3 g dikálium-hidrogén-foszfát
• 0,2 g kalcium-hidrogén-foszfát, kritályvizes
78

• 0,2 g dikálium-foszfát
• 0,3 g magnézium-szulfát, kristályvizes
• 0,5 g nátrium-klorid
• 0,01 g vas(III)klorid, kristályvizes
• 5 g kálcium-karbonát
• 1,5 g agar
• 1 ml nyomelemkeverék
pH: 7,2-7,4
Nyomelemkeverék Fjodorov-féle táptalajhoz 1000 ml-re
• 5 g bórsav
• 5 g ammónium-molibdenát, kristályvizes
• 0,5 g kálium-jodid
• 05 g nátrium-bromid
• 0,2 g cink-szulfát, kristályvizes
• 0,3 g alumínium-szulfát
I. jelű oldat (P. putida teszt) 500 ml-re
• 10 g NaNO3
• 2,4 g K2HPO4
• 1,2 g KH2PO4
• 1 g élesztőkivonat
II. jelű oldat (P. putida teszt) 500 ml-re
• 10 g NaNO3
• 2,4 g K2HPO4
• 1,2 g KH2PO4
III. jelű oldat (P. putida teszt) 500 ml-re
• 40 g glükóz
IV. jelű oldat (P. putida teszt) 1000 ml-re
• 4 g MgSO4·7H2O
• 0,01 g vas(III)-citrát
79

Fenntaró kultúra táptalaj (P. putida teszt) 1000 ml
• 50 ml I. jelű oldat
• 125 ml III. jelű oldat
• 100 ml IV. jelű oldat
• 18 g agar
Előkultúra táptalaj (P. putida teszt) 1000 ml
• 900 ml steril desztillált víz
• 25 ml I. jelű oldat
• 25 ml III. jelű oldat
• 50 ml IV. jelű oldat
80

V. REAGENSEK, FESTÉKEK
R-1 : Metilénkék festékoldat
• 50 mg metilénkék • 5 ml etanol (96%-os) • 95 ml desztillált víz
R-2 : Karbolos fukszin oldat
A oldat: 300 mg bázikus fukszin
10 ml etanol (96%-os)
B oldat: 5 g fenol
95 ml desztillált víz
Az "A" oldatot keverés közben a "B" oldatba öntjük, majd leszűrjük!
R-3 : Oxalátos kristályibolya (Gram-festéshez)
A oldat: 1 g kristályibolya 10 ml (96%-os) etanolba oldva
B oldat: 1 g ammóniumoxalát 100 ml desztillált vízben oldva
Az "A" oldatot keverés közben a "B" oldatba öntjük!
R-4 : Lugol-oldat (Gram-festéshez)
• 1 g jód • 2 g KI • 300 ml desztillált víz
R-5 : Szafranin oldat
• 250 mg szafranin • 10 ml etanol (96%-os) • 100 ml desztillált víz
R-6 : Malachitzöld festékoldat
• 1 g malachitzöld • 1 g fenol • 100 ml desztillált víz
R-15 : Hidrogénperoxid oldat kataláz próbához
• 9 ml H2O2 (33%-os) • 91 ml desztillált víz
(Csak a frissen bontott H2O2 esetében elegendő a 3%-os mennyiség.)
Használat előtt frissen készítendő!
81

R-16 : Oxidáz reagens dietil-p-feniléndiamin-HCl
• 1%-os vizes oldata Célszerű mindig friss készítményt használni (légmentesen lezárva hűtőszekrényben 1 hétig tárolható,
ha kékre színeződik, ne használjuk)!
R-17 : Lugol-oldat (keményítőbomlás kimutatásához)
Törzsoldat:
• 5 g jód • 10 g KI • 100 ml desztillált víz
(Hidegen tartjuk, használat előtt desztillált vízben 50x-re hígítjuk!)
R-18 : HgCl2, oldat
• g HgCl2 • ml cc.HCl • ml desztillált víz
82

VI. A GYAKORLATOKON ALKALMAZOTT MIKROORGANIZMUSOK JEGYZÉKE
Gram-negatív baktériumok: Élesztőgombák:
Pseudomonas aeruginosa Schizosaccharomyces pombe
Escherichia coli Saccharomyces cerevisiae
Serratia marcescens Saccharomyces kluyveri
Proteus vulgaris Saccharomyces diastaticus
Saccharomycodes ludwigii
Gram-pozitív baktériumok: Geotrichum candidum
Micrococcus luteus Candida utilis
Staphylococcus aureus Rhodotorula rubra
Streptococcus faecalis Hansenula anomala
Streptococcus pyogenes
Bacillus cereus var. mycoides Fonalasgombák:
Bacillus megaterium Aspergillus nidulans
Bacillus subtilis Aspergillus niger 65
Clostridium sp. Aspergillus niger ade- 341
Lactobacillus sp. Aspergillus violaceus
Streptomyces griseu Aspergillus albertensis
Streptomyces albus Neurospora crassa
Streptomyces aureus Penicillium chrysogenum
Penicillium claviforme
Penicillium frequentans
Penicillium purpurogenum
Penicillium ramigena
Sordaria fimicola
Trichoderma viridae
Actinomucor repens
Mucor hiemalis
Mucor genevensis
Mucor rouxii
Mycotypha pyriphaera
Syncephalastrum racemosum
Thamnostylum piriforme
83

VII. FORRÁSMUNKÁK ÉS AJÁNLOTT IRODALOM
Bálint, P. (szerk.) (1974) Klinikai laboratóriumi diagnosztika, Medicina Könyvkiadó, Budapest,
Brock, T.D., Madigan, M.T., Martinko, J.M, Parker, J. (1997) Biology of Microorganisms, Eighth Edition, Prentice Hall Inter. Ltd., London
Capuccino, J.G., Sherman, N. (1983) Microbiology a Laboratory Manual, Addison-Wesley Publ.
Co., Reading
Case C.L., Johnson T.R. (1984.) Laboratory experiments in Microbiology, Benjamin/Cummings
Publ. Co. Inc., Menlo Park,
Clark, G. (1981) Staining Procedures. (Fourth edition.) Williams and Wilkins, Baltimore,
Collins, C.H.( 1967). Microbiological Methods. ButterWorths, London,
Davis, B.D., Dulbecco,R., Eisen, H.N., Ginsberg, H.S. (1990) Microbiology Fourth Edition, J. B.
Lipincott Company, Philadelphia, London, Sydney, Tokyo
Ferenczy, L., Zsolt, J. (1979) Mikrobiológia, Tankönyvkiadó, Budapest
Gerhardt, P. (szerk.) (1993) Manual of Methods for General Bacteriology, ASM, Washington
Horváth, S. (1981) Mikrobiológiai praktikum, Tankönyvkiadó, Budapest
Kavanagh, F. (1963) Analytical Microbiology, Academic Press, New York, London
Lányi, B. (szerk.) (1980) Járványügyi és klinikai bakteriológia. OKI, Budapest
Norris, J.R., Ribbons, D.W. (szerk.) (1969-1992) Methods in microbiology. (1-24 kötet) Academic
Press, London and New York
Primrose, S.B., Wardlaw, A.C. (1982) Source Book of Experiments for the Teaching of
Microbiology, Academic Press, London
Schlegel H.G. (1993) General Microbiology, Seventh Edition, Cambridge Univ.Press, Cambridge
Sharp, M.S., Lyles, S.T. (1969) Laboratory Instructions in Biology of Microorganisms, Moshby
Co., Saint Louis
Sós, J. (szerk.) (1974) Laboratóriumi diagnosztika, Medicina Könyvkiadó, Budapest
Szegi, J., (1979) Talajmikrobiológiai vizsgálati módszerek, Mezőgazdasági Könyvkiadó, Budapest
Kristen Mortelmans, Errol Zeiger: The Ames Salmonella/microsome mutagenicity assay. (2000),
Mutation Resaerch 455, 29-60.
Kucsera Judit: Mikrobiológiai Gyakorlatok II. (JATE Press, Szeged, 2003.)
Országos Szabvány, MSZ 21978/30-1988
Országos Szabvány, MSZ-08-1721/1-86
Európai Szabvány, EN ISO 10712
84

TARTALOMJEGYZÉK
BEVEZETÉS .......................................................................................................................................... 1
I. A LABORATÓRIUM RENDJE.......................................................................................................... 2
II. ANYAG ÉS ESZKÖZISMERET....................................................................................................... 3 2.1. TÁPTALAJOK ................................................................................................................................ 3 2.2. TENYÉSZTÉSHEZ HASZNÁLT EDÉNYEK ............................................................................... 4 2.3. OLTÓESZKÖZÖK .......................................................................................................................... 6 2.4. STERILEZÉS................................................................................................................................... 7 2.5. A MIKROORGANIZMUSOK TENYÉSZTÉSE .......................................................................... 10 2.6. MIKROSZKÓP-ISMERET............................................................................................................ 11
III. GYAKORLATOK .......................................................................................................................... 15 1. gyakorlat: ÁTOLTÁSOK GYAKORLÁSA ..................................................................................... 16 2. gyakorlat: MIKROORGANIZMUSOK SZAPORODÁSA KÜLÖNBÖZŐ TÁPTALAJOKON .... 20 3. gyakorlat: TISZTATENYÉSZETEK KÉSZÍTÉSE .......................................................................... 22 4. gyakorlat: MIKROORGANIZMUSOK MORFOLÓGIAI VIZSGÁLATA RÖGZÍTETT FESTETT
PREPARÁTUMOKKAL, EGYSZERŰ FESTÉSEK........................................................................ 26 5. gyakorlat: MIKROORGANIZMUSOK MORFOLÓGIAI VIZSGÁLATA RÖGZÍTETT FESTETT
PREPARÁTUMOKKAL, ÖSSZETETT FESTÉSEK....................................................................... 28 6. gyakorlat: MIKROORGANIZMUSOK NAGYSÁGÁNAK MÉRÉSE ........................................... 32 7. gyakorlat: BAKTÉRIUMOK ÉS ÉLESZTŐK SEJTSZÁMMEGHATÁROZÁS MÓDSZEREI .... 34 8. gyakorlat: MIKROORGANIZMUSOK ANTAGONIZMUSÁNAK VIZSGÁLATA ..................... 40 9. gyakorlat: OXIDO-REDUKCIÓS ENZIMEK VIZSGÁLATA........................................................ 42 10. gyakorlat: MAKROMOLEKULÁK EXOENZlMES BONTÁSA.................................................. 46 11. gyakorlat: ÉLESZTŐGOMBÁK CUKORASSZIMILÁCIÓJÁNAK ÉS ERJESZTÉSÉNEK
VIZSGÁLATA .................................................................................................................................. 50 12. gyakorlat: BAKTÉRIUMOK ANTIBIOTIKUM ÉRZÉKENYSÉGÉNEK VIZSGÁLATA ......... 52 13. gyakorlat: AZ OZMOTIKUS NYOMÁS HATÁSA A MIKROORGANIZMUSOKRA .............. 54 14. gyakorlat: FÉMIONOK SZAPORODÁSGÁTLÓ HATÁSA......................................................... 56 15. gyakorlat: NÖVÉNYI ANTIMIKROBIÁLIS VEGYÜLETEK HATÁSÁNAK VIZSGÁLATA . 58 16. gyakorlat: AZ UV-SUGÁRZÁS HATÁSA A MIKROORGANIZMUSOKRA ............................ 60 17. gyakorlat: MUTAGENITÁSI VIZSGÁLAT AMES-TESZTTEL.................................................. 64 18. gyakorlat: PETIT-MUTÁNS INDUKCIÓ ...................................................................................... 66 19. gyakorlat: AZOTOBACTER AGILE TESZT................................................................................. 68
85

20. gyakorlat: TALAJTOXICITÁS VIZSGÁLAT AZOTOBAKTERES TALAJBLOKK
MÓDSZERREL ................................................................................................................................. 70 21. gyakorlat: PSEUDOMONAS PUTIDA NÖVEKEÉSGÁTLÁSI TESZTJE.................................... 72
IV. TÁPTALAJOK ............................................................................................................................... 74
V. REAGENSEK, FESTÉKEK ............................................................................................................ 81
VI. A GYAKORLATOKON ALKALMAZOTT MIKROORGANIZMUSOK JEGYZÉKE.............. 83
VII. FORRÁSMUNKÁK ÉS AJÁNLOTT IRODALOM .................................................................... 84
86
















