
Süreli Yayın Tasarımı için Temel Tipografi Bilgisi (Ders Notları, 2004)
Upload
independentCategory
view
0download
0
This article was published in the above mentioned Springer issue.The material, including all portions thereof, is protected by copyright;all rights are held exclusively by Springer Science + Business Media.
The material is for personal use only;commercial use is not permitted.
Unauthorized reproduction, transfer and/or usemay be a violation of criminal as well as civil law.
ISSN 0167-6857, Volume 102, Number 3

ORIGINAL PAPER
Production of in vitro haploid plants from in situ induced haploidembryos in winter squash (Cucurbita maxima Duchesne ex Lam.)via irradiated pollen
Ertan Sait Kurtar • Ahmet Balkaya
Received: 18 November 2009 / Accepted: 1 March 2010 / Published online: 23 March 2010
� Springer Science+Business Media B.V. 2010
Abstract The influence of pollen irradiation on the pro-
duction of in vitro haploid plants from in situ induced
haploid embryos was investigated in winter squash
(Cucurbita maxima Duchesne ex Lam.). Pollen were irra-
diated at different gamma-ray doses (50, 100, 200 and 300
Gray) and durations (9, 11, 15, 21, and 28 July). Production
of in vitro haploid plantlets was influenced by irradiation
dose, irradiation duration, genotype, and embryo type and
embryo stage. Embryos were only obtained from lower
irradiation doses (50 Gray and 100 Gray) and earlier irra-
diation durations (9, 11, and 15 July). The greatest embryo
number per fruit was procured from ‘‘G14’’ and ‘‘55SI06’’
genotypes at 50 Gray gamma-ray dose. Necrotic embryos
were higher than normal embryos at delayed harvest times
(5 and 6 weeks after the pollination). The convenient
harvest time for embryo rescue was observed about
4 weeks (between 25 and 30 days) after pollination. All
cotyledon and amorphous embryos had only diploid plants
while late-torpedo, arrow-tip, and pro-cotyledon embryos
produced 33.3, 50.0, and 66.7% haploid plant. The fre-
quency of haploid plantlets was 0.11, 1.17, 10.96 and 0.28
per 100 seeds, 100 embryos, 100 plantlets and a fruit at
50 Gray gamma-ray dose, respectively.
Keywords Irradiated pollen � In vitro haploidization �Winter squash (Cucurbita maxima Duchesne ex Lam.)
Introduction
In a conventional breeding programme, pure lines are
obtained after several generations of selfing and still may
not be 100% homozygous (Germana 2006). Haploidization
is the process of producing haploid plants in a single
generation. Following haploidization, the chromosome
number of haploid plants may be doubled (dihaploidiza-
tion) to obtain complete fertile homozygous lines. These
valuable lines are currently used for breeding programmes
and genetic research.
Haploid plants can be obtained spontaneously (andro-
genesis, gynogenesis, or parthenogenesis, semigamy and
polyembryony), but the frequency of haploid is very low
(Pochard and Dumas de Vaulx 1971; Lacadena 1974).
Recently, haploid plants are produced using in vitro
androgenesis (anther-microspore culture) and gynogenesis
(ovule-ovary culture), and in situ parthenogenesis (pollen
irradiation and treatment with chemicals, etc.).
Pollen irradiation (UV, gamma rays, and X-rays) is the
most widely used technique to induce in situ parthenoge-
netic haploid plants. Gamma rays are commonly used in
haploid programmes because of their simple application,
good penetration, reproducibility, high mutation frequency,
and low disposal (lethal) problems (Chahal and Gosal
2002). This technique was used firstly with embryo culture
on different species of Nicotiana (Pandey and Phung
1982).
Irradiated pollen can germinate on the stigma, grow
within the style and reach the embryo sac, but cannot
fertilize the egg-cell and the polar nuclei (Cuny 1992).
Genetically inactive but germinable pollen can be used to
stimulate the division of the egg cell, and thus induce
parthenogenesis or development of parthenocarpic fruit,
including gynogenic haploid production; overcoming
E. S. Kurtar (&)
High School of Profession of Bafra, Ondokuz Mayis University,
Bafra, Samsun, Turkey
e-mail: [email protected]
A. Balkaya
Horticulture Department of Agriculture Faculty, Ondokuz Mayis
University, Samsun, Turkey
123
Plant Cell Tiss Organ Cult (2010) 102:267–277
DOI 10.1007/s11240-010-9729-1 Author's personal copy

minor cross-incompatibilities, and for physiological studies
of incompatibility (Stairs and Mergen 1964; Savaskan and
Toker 1991; Todorova et al. 2004), gene transformation
(Pandey 1978) and nucleus substitution (Raquin et al.
1989).
The irradiated pollen technique is an effective method
for the induction of haploid embryos in Cucurbit. Induction
of in situ haploid embryos and obtaining in vitro haploid
plants have been achieved using an irradiated pollen
technique in watermelon (Gursoz et al. 1991; Sari 1994),
melon (Sauton and Dumas de Vaulx 1987; Cuny 1992;
Maestro-Tejada 1992; Sari et al. 1992; Abak et al. 1996;
Lotfi et al. 2003), cucumber (Truong-Andre 1988;
Niemirowicz-Szczytt and Dumas de Vaulx 1989; Sauton
1989; Caglar and Abak 1999; Faris et al. 1999; Lotfi et al.
1999; Dolcet-Sanjuan et al. 2006), snake cucumber (Taner
et al. 2000), summer squash (Kurtar et al. 2002), and
pumpkin (Kurtar et al. 2009).
Winter squash is an annual cultivated species of
Cucurbit (Whitaker and Bemis 1964) and genetically dif-
ferent from Cucurbita pepo and Cucurbita moschata
(Athanasios et al. 2009). Cucurbita pepo shares a common
ancestor with C. moschata and C. argyrosperma, but not
with C. maxima (Decker-Walters et al. 1990). Isozyme
study showed high allelic diversity in C. pepo, C. mosch-
ata, and C. maxima (Heikal et al. 2008). To the authors’
knowledge, this is the first report on in vitro haploid pro-
duction in winter squash.
The objective of the present study was to determine the
effects of irradiation dose (Co60), irradiation duration,
genotype, embryo type and embryo stage on production of
in vitro haploid plants from in situ induced haploid
embryos in winter squash.
Materials and methods
Plant material
The experiment was carried out with six winter squash
genotypes (57SI06, 57SI21, 55BA02, 55BA03, 55CA06,
and G14) selected from four provinces (Samsun, Amasya,
Sinop and Bolu) of the Black Sea region of Turkey (genetic
material used in the project was funded by TUBITAK-
TOVAG -104O144), except ‘‘G14’’ genotype which was
provided by the Turkish Seed Gene Bank, Menemen, Izmir
(Fig. 1). The seeds were sown in plastic flats (cell volume
150 cm3 and 28 cells per flat) containing mixture of peat-
moss: perlite (2:1 v/v) on 20 April 2008. Seedlings were
raised in unheated glasshouse, and 15 seedlings from each
genotype were planted at 3–4 leaf stage with spacing of
3 9 3 m on 11 May.
Irradiation and pollination
Female flowers were isolated with white cloth bags
(15 9 10 cm) and male flowers were collected around
noon on the day before anthesis. Anthers without filaments
were excised and mixed equally for each genotype, and
placed into small cardboard boxes (5 9 7 9 2 cm). Sam-
ples were irradiated at 50, 100, 200 and 300 Gray doses of
gamma rays by a Co60 source of Theratron 780-C equip-
ment on the same day at 1,482 Ci source activity and
11.96 Gymin-1 dose rate. Irradiation experiments were
performed at different durations (9, 11, 15, 21, and 28 July)
to evaluate the effects of irradiation duration on embryo
induction. Irradiated anthers were incubated at room tem-
perature overnight. Female flowers were pollinated by
irradiated pollen in the morning of the following day at
0700–0900 hours. Pollen of flowers 0 days old (the day
following irradiation) and 1 day old were used for polli-
nation. Female flowers were then isolated with cloth bags
again to avoid undesired pollen contamination. Cloth bags
were removed at 3rd or 4th days of pollination.
Embryo culture
Immature fruits were harvested from 3 to 6 weeks after
pollination to determine the convenient harvest time.
Washed fruits were surface-sterilized in 2% sodium hypo-
chlorite solution for 30 min and then flame-sterilised using
ethanol. Prior to extraction, the laminar-flow hood was
disinfected with UV light for 15 min following 70% etha-
nol. Thereafter, seeds were extracted under axenic condi-
tions in a laminar-flow hood. Liquid medium culture (Lotfi
et al. 2003) and direct extraction methods were investigated
to embryo rescue. Embryos were rescued from 21- to 42-
day-old immature fruits and cultured in magenta boxes and
culture tubes containing solid E20A medium (Sauton and
Dumas de Vaulx 1987) (Table 1). Embryos were classified
according to embryo type and stage of embryo development
(Raghavan 1986; Kurtar et al. 2002), and cultured at
28 ± 1�C with 16-h photoperiod (3,000 lux) thereafter.
Transplantation and acclimatization
After 5–15 days of culture, mini-plantlets (having root and
shoot) were transferred onto fresh E20A medium for fur-
ther development, individually. Well-generated complete
plantlets (27–41 days old) were undergone acclimatization
process. First, the covers of magenta boxes and culture
tubes were gradually opened for 8–10 days. Plantlets were
removed and the roots were washed carefully in running
tap-water. The roots were then soaked for 10 min in 0.2%
solution of fungicide (Maxim XL035FS) to prevent pos-
sible contamination at the beginning of the acclimatization.
268 Plant Cell Tiss Organ Cult (2010) 102:267–277
123
Author's personal copy

The plantlets were transplanted into plastic cups (150 cm3)
containing sterile peat-moss. Each cup was closed a
transparent cup and the plantlets were acclimatized in a
growth cabinet (Nuve TK120) at 28 ± 1�C with 16-h
photoperiod (6,000 lux) and 95% humidity. The transpar-
ent cups were opened gradually and completely removed at
6 days. The humidity of the growth cabinet was regulated
at intervals of 5% over a period of 2 days, until the
greenhouse or open field condition was reached.
Ploidy determination
The ploidy level of plantlets was determined by direct
(chromosome counting in root tip) and indirect (stomata
size, stomata density, and chloroplast number of the guard
cells) methods.
Chromosome counting in root tip was realised by
‘‘Feulgen technique’’ (Darlington and La Cour 1963). In
chromosome counting, root tips of plantlets were excised at
the beginning of acclimatization process. The 4th or 5th
leaves from shoot apex were used to measure stomata size
(width and length), stomata density (number of stoma per
mm2) and chloroplast number (each side of the guard cells)
(Sari 1994). In the measurements, lower epidermal strips of
leaves was placed onto a microscope slide after the addi-
tion of one drop of tap-water, and the cover glass was then
closed (Dore 1986). The sizes of 8 stomata per leaf were
measured for stomata size. 1% AgNO3 solution was used
for chloroplast counting (Rouselle 1992). Stomata density
and chloroplast number were counted in 6 visual areas.
Chromosomes and stomata were observed (40 910 mag-
nification) and photographed (100 910 magnification) by a
light microscope (Nikon, Alphapot, YS-2 model).
Statistical analysis
Responses to induction of in situ haploid embryos and
production of in vitro haploid plants are expressed as
Fig. 1 Representative fruits of investigated genotypes
Plant Cell Tiss Organ Cult (2010) 102:267–277 269
123
Author's personal copy

percentage (%) due to unequal test materials. Statistical
analysis were only carried out in stomata measurements as
completely randomized experimental design including four
replications and the differences between means were
evaluated by using ANOVA test.
Results
Embryo induction
Only 50 Gray and 100 Gray gamma-ray doses and polli-
nation with 0-day-old pollen stimulated embryo induction.
Overall, 50 fruits, 7,071 seeds and 686 embryos were
obtained from the investigated genotypes, and mean
embryo number per fruit was 13.7. The highest fruit (29),
seed (4,981) and embryo number (673) were procured from
50 Gray gamma-ray dose (Table 2). Relatively higher
gamma-ray doses (200 and 300 Gray) were not effective on
embryo induction and gave fruits more or less but fruits
were seedless.
The total seed and embryo number showed varied
responses to different irradiation durations. At earlier
irradiation durations (9, 11, and 15 July) produced the
highest total seed (2,903, 2,979, and 855) and embryo
number (223, 318, and 122) whereas seed (216 and 118)
and embryo number (12 and 11) were the lowest at later
irradiation durations (21 and 28 July), sequentially. The
highest mean embryo number per fruit was recorded as
19.9 on 11 July. Mean seed and embryo number were
varied from genotypes, and the highest mean seed and
embryo number were 381 and 21.2 in ‘‘G14’’ while
55CA06 had the lowest mean seed and embryo number
(133 and 8.2), respectively.
Regeneration efficiency
Both normal (white) and necrotic embryos were rescued
and classified according to stages of embryo development
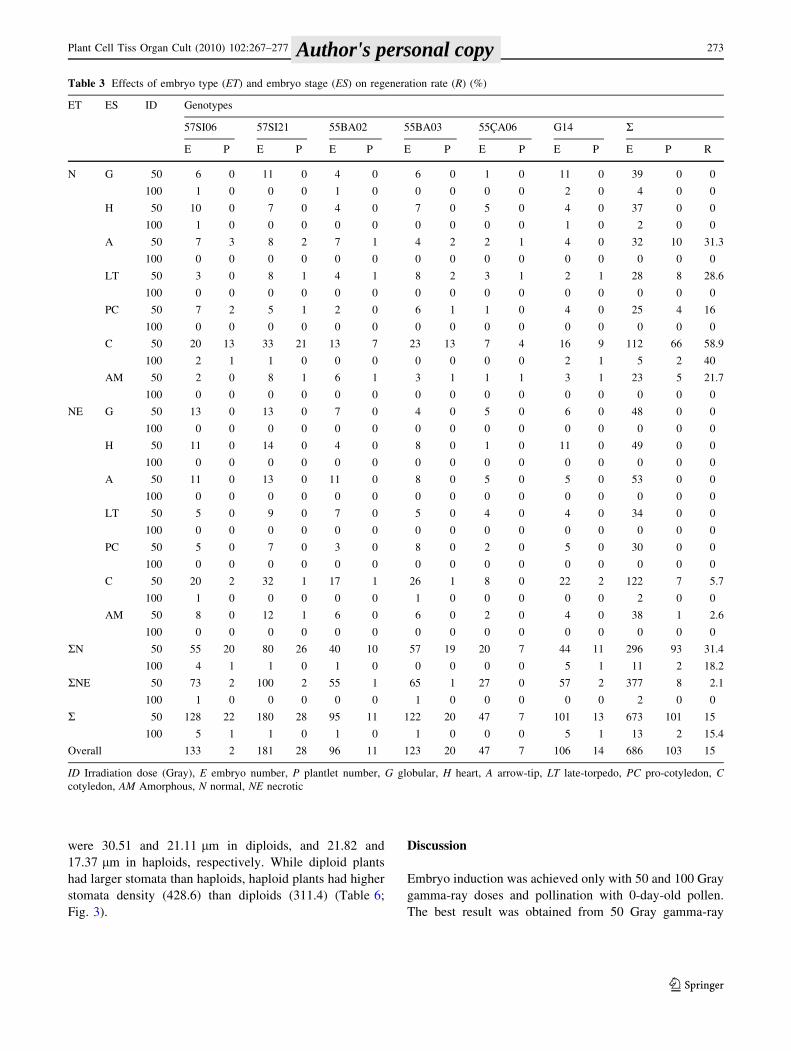
(Raghavan 1986; Kurtar et al. 2002) (Fig. 2). Liquid
medium culture (Lotfi et al. 2003) was found to be inef-
fective on embryo excising, and embryos were not visu-
alized and identified. Moreover, contamination was
observed and caused embryo loss.
Overall, 103 plantlets were regenerated from 686
embryos, and regeneration rate was 15.0%. The number of
necrotic embryos (379) was higher than normal embryos
(307), and regeneration rate were found 31.9% (95 plant-
lets) in normal and 2.1% (8 plantlets) in necrotic embryos.
Well-developed embryos (cotyledon) had higher regener-
ation rate than amorphous and at early developmental
stages embryos (pro-cotyledon, late-torpedo, and arrow-
tip). Despite globular and heart embryos did not regenerate
cotyledon embryos had the highest regeneration rate
(58.9%). Arrow-tip (31.3%) and late-torpedo (28.6%)
embryos also gave noteworthy results compared to pro-
cotyledon embryos (16.0%) (Table 3).
Frequency of haploid plantlets
Frequency of haploid plantlets was influenced from embryo
type and embryo stage, and all haploid plantlets were
obtained from normal embryos. Overall, 8 haploid plantlets
were regenerated from investigated genotypes (3 of
57SI06, 2 of G14, and 3 of 57SI21) at 50 Gray gamma-ray
dose. All cotyledon and amorphous embryos gave only
diploid plantlets, while late-torpedo, arrow-tip, and pro-
cotyledon embryos produced haploid plantlets with values
33.3, 50.0, and 66.7%, respectively (Table 4).
The frequency of haploid in per 100 plantlets was 8.00,
12.2, and 13.3 on 9, 11, and 15 July, respectively. Later
irradiation durations (21 and 28 July) had no effect on
embryo induction. Overall, the frequency of haploid
plantlets in per 100 seeds, 100 embryos, 100 plantlets, and
fruit was determined as 0.11, 1.17, 10.96, and 0.28 at 50
Gray dose, respectively (Table 5).
Ploidy determination
Chromosome counting
The results of chromosome counting on root tips of 73 plant-
lets (8 of arrow-tips, 6 of late-torpedo, 3 of pro-cotyledon,
Table 1 Composition of ‘‘E20A’’ medium
Macro and micro elements (mg l-1)
KNO3 1,075 MnSO4�7H2O 11.065
NH4 NO3 619 ZnSO4�7H2O 1.812
MgSO4�7H2O 206 H3 BO3 1.575
CaCl2�2H2O 156.5 KI 0.345
KH2PO4 71 Na2MgO4�2H2O 0.094
Ca(NO3)2�4H2O 25 CuSO4�5H2O 0.008
NaH2PO4�4H2O 19 CoCl2�6H2O 0.008
(NH4)2SO4 17 FeSO4�7H2O 27.8
KCl 3.5 Na2EDTA 37.3
Vitamins and amino acids (mg l-1) Others
Myo-inositol 50.300 Sucrose 20.00 g l-1
Pyridoxine–HCl 5.500 Agar 7.00 g l-1
Nicotinic acid 0.700 IAA 0.01 mg l-1
Thiamine–HCl 0.600 pH 5.90
Calcium pantothenate 0.500
Glycine 0.100
Biotine 0.005
270 Plant Cell Tiss Organ Cult (2010) 102:267–277
123
Author's personal copy

Ta
ble
2F
ruit
nu
mb
er(F
),se
edn
um
ber
(S),
mea
nse
edn
um
ber
(MS),
emb
ryo
nu
mb
er(E
)an
dm
ean
emb
ryo
nu
mb
er(M
E)
ITID
Gen
oty
pes
Ov
eral
l
57
SI0
65
7S
I21
55
BA
02
55
BA
03
55
CA
06
G1
4
FS
MS
EM
EF
SM
SE
ME
FS
MS
EM
EF
SM
SE
ME
FS
MS
EM
EF
SM
SE
ME
FS
MS
EM
E
95
01
25
42
54
29
29
43
69
92
62
16
13
24
32
43
33
31
25
22
52
19
19
12
83
28
32
72
71
65
46
54
46
46
92
,13
62
37
21
62
4.0
10
01
13
81
38
00
31
35
45
10
.31
21
42
14
00
00
00
01
19
61
96
00
18
48
46
67
76
71
10
71
.0
R2
39
21
96
29
14
.57
50
47
26
39
.02
53
82
69
33
16
.51
25
22
52
19
19
.02
47
92
40
27
13
.52
73
83
69
52
26
.01
62
,90
31
81
22
31
3.9
11
50
26
38
31
96
93
4.5
21
46
73
36
18
15
21
52
14
24
22
17
99
09
14
5.5
11
83
18
32
22
21
51
85
18
52
52
92
,18
52
43
31
23
4.7
10
00
00
00
15
25
20
01
17
81
78
00
28
34
24
22
10
61
06
00
13
75
37
52
27
79
41
13
61
.1
R2
63
83
19
69
34
.53
19
89
93
61
2.0
26
99
35
04
22
1.0
42
62
66
95
23
.83
28
91
45
22
7.3
28
93
44
75
42
7.0
16
2,9
79
18
63
18
19
.9
15
50
11
09
10
93
23
22
11
75
86
33
1.5
19
79
71
91
91
10
71
07
88
00
00
00
00
00
54
30
86
12
22
4.4
10
01
97
97
00
12
82
80
00
00
00
00
00
01
27
27
00
12
73
27
30
04
42
51
06
00
R2
20
61
03
32
16
.03
14
54
86
32
1.0
19
79
71
91
9.0
11
07
10
78
8.0
12
72
70
01
27
32
73
00
98
55
95
12
21
3.6
21
50
13
83
82
21
27
27
77
15
95
93
30
00
00
00
00
00
00
00
31
24
43
12
4.0
10
00
00
00
13
93
90
00
00
00
15
35
30
00
00
00
00
00
02
92
46
00
R1
38
38
22
.02
66
33
73
.51
59
59
33
15
35
30
00
00
00
00
00
05
21
64
31
22
.4
28
50
00
00
02
64
32
11
5.5
14
24
20
00
00
00
00
00
00
00
00
31
06
35
11
3.7
10
00
00
00
11
21
20
00
00
00
00
00
00
00
00
00
00
01
12
12
00
R0
00
00
37
62
51
13
.71
42
42
00
00
00
00
00
00
00
00
04
11
83
01
12
.8
Ov
eral
l7
1,2
74
18
21
32
18
.91
89
89
55
18
01
0.0
71
,43
52
05
97
13
.97
67
49
61
22
17
.46
79
51
33
49
8.2
51
,90
43
81
10
62
1.2
50
7,0
71
14
16
86
13
.7
ITIr
rad
iati
on
tim
ein
July
,ID
irra
dia
tio
nd
ose
(Gra
y)
Plant Cell Tiss Organ Cult (2010) 102:267–277 271
123
Author's personal copy

52 of cotyledon, and 4 of amorphous) indicated that ploidy
level changed with embryo stages. Haploid plantlets
(n = 20) were achieved from arrow-tip (4), late-torpedo
(2), and pro-cotyledon (2) embryos. All cotyledon and
amorphous embryos produced only diploid plantlets
(2n = 40).
Stomata observations
Diploid plants had 10 or 12 chloroplast in guard cells
whereas haploid plants had 6 or 8 chloroplast. Average
chloroplast number was counted 11.17 in diploids and 7.21
in haploids. Besides, average stomata length and width
Fig. 2 Some of the different stages of embryos: a,b globular; c,d heart; e arrow-tip; f,g late-torpedo; h,i pro-cotyledon; j,k cotyledon; l,mamorphous; n,o necrotic
272 Plant Cell Tiss Organ Cult (2010) 102:267–277
123
Author's personal copy
were 30.51 and 21.11 lm in diploids, and 21.82 and
17.37 lm in haploids, respectively. While diploid plants
had larger stomata than haploids, haploid plants had higher
stomata density (428.6) than diploids (311.4) (Table 6;
Fig. 3).
Discussion
Embryo induction was achieved only with 50 and 100 Gray
gamma-ray doses and pollination with 0-day-old pollen.
The best result was obtained from 50 Gray gamma-ray
Table 3 Effects of embryo type (ET) and embryo stage (ES) on regeneration rate (R) (%)
ET ES ID Genotypes
57SI06 57SI21 55BA02 55BA03 55CA06 G14 R
E P E P E P E P E P E P E P R
N G 50 6 0 11 0 4 0 6 0 1 0 11 0 39 0 0
100 1 0 0 0 1 0 0 0 0 0 2 0 4 0 0
H 50 10 0 7 0 4 0 7 0 5 0 4 0 37 0 0
100 1 0 0 0 0 0 0 0 0 0 1 0 2 0 0
A 50 7 3 8 2 7 1 4 2 2 1 4 0 32 10 31.3
100 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0
LT 50 3 0 8 1 4 1 8 2 3 1 2 1 28 8 28.6
100 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0
PC 50 7 2 5 1 2 0 6 1 1 0 4 0 25 4 16
100 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0
C 50 20 13 33 21 13 7 23 13 7 4 16 9 112 66 58.9
100 2 1 1 0 0 0 0 0 0 0 2 1 5 2 40
AM 50 2 0 8 1 6 1 3 1 1 1 3 1 23 5 21.7
100 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0
NE G 50 13 0 13 0 7 0 4 0 5 0 6 0 48 0 0
100 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0
H 50 11 0 14 0 4 0 8 0 1 0 11 0 49 0 0
100 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0
A 50 11 0 13 0 11 0 8 0 5 0 5 0 53 0 0
100 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0
LT 50 5 0 9 0 7 0 5 0 4 0 4 0 34 0 0
100 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0
PC 50 5 0 7 0 3 0 8 0 2 0 5 0 30 0 0
100 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0
C 50 20 2 32 1 17 1 26 1 8 0 22 2 122 7 5.7
100 1 0 0 0 0 0 1 0 0 0 0 0 2 0 0
AM 50 8 0 12 1 6 0 6 0 2 0 4 0 38 1 2.6
100 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0
RN 50 55 20 80 26 40 10 57 19 20 7 44 11 296 93 31.4
100 4 1 1 0 1 0 0 0 0 0 5 1 11 2 18.2
RNE 50 73 2 100 2 55 1 65 1 27 0 57 2 377 8 2.1
100 1 0 0 0 0 0 1 0 0 0 0 0 2 0 0
R 50 128 22 180 28 95 11 122 20 47 7 101 13 673 101 15
100 5 1 1 0 1 0 1 0 0 0 5 1 13 2 15.4
Overall 133 2 181 28 96 11 123 20 47 7 106 14 686 103 15
ID Irradiation dose (Gray), E embryo number, P plantlet number, G globular, H heart, A arrow-tip, LT late-torpedo, PC pro-cotyledon, Ccotyledon, AM Amorphous, N normal, NE necrotic
Plant Cell Tiss Organ Cult (2010) 102:267–277 273
123
Author's personal copy

dose, and embryo induction was not performed at 200 and
300 Gray doses. Embryos and haploid plants were also
obtained from lower irradiation doses (25 and 50 Gray) in
summer squash (Kurtar et al. 2002) and in pumpkin (50 and
100 Gray) (Kurtar et al. 2009). On the other hand, haploid
embryo induction was obtained at relatively higher doses
(200–300 Gray) in watermelon (Gursoz et al. 1991; Sari
et al. 1994), melon (Sauton and Dumas de Vaulx 1987;
Cuny 1992; Maestro-Tejada 1992; Sari et al. 1992; Abak
et al. 1996; Lotfi et al. 2003), cucumber (Niemirowicz-
Szczytt and Dumas de Vaulx 1989; Sauton 1989; Caglar
and Abak 1999), and snake cucumber (Taner et al. 2000).
Contrary to these reports, the best irradiation dose was
found 100 Gray (Faris et al. 1999; Lotfi et al. 1999), 150
Gray (Xie et al. 2005), and 500 Gray (Dolcet-Sanjuan et al.
2006) in cucumber, and 750 Gray in melon (Sun et al.
2006).
These results may be attributed to the radio-resistance of
pollen and also to biologic efficiency of irradiation. A
linear relationship between radio-resistance and pollen
size, which is also a function of the amount of DNA in the
nucleus has been reported (Brewbaker and Emery 1962;
Alison and Casareft 1968; Shridhar 1992; Jain et al. 1996).
Pollen of winter squash are one of the largest pollen (as in
squash and pumpkin) in vegetables (average width
180 lm). Melon, watermelon, and cucumber pollen are
smaller than winter squash (average 50, 60 and 65 lm,
respectively) (Sensoy et al. 2003). Moreover, melon,
watermelon and cucumber have 3 apertures, whereas
winter squash has 12 apertures. Hence, winter squash
pollen are more sensitive to dehydration and rapid loss of
their viability as reported in squash (Nepi and Pacini 1993).
Pollen viability, germination ability and fruit and seed-
set also decreased along with increasing of irradiation dose,
irradiation duration and pollen age in pumpkin and winter
squash (Kurtar 2009).
Biologic efficiency of irradiation relates to the source
activity (Ci) and dose rate (Gymin-1) (Ozalpan 2001).
Irradiation efficiency influences from oxygen and water
contents of tissue (Bernstein et al. 1993), division volume
of cell (Tokarek et al. 1994) and also type of radiation
(Goldschmidt et al. 1994). Although the same irradiation
procedure was followed in our previous work, the best
result was obtained from 25 to 50 Gray gamma ray doses in
summer squash (Kurtar et al. 2002). Irradiation treatments
were realised at 6,313 Ci source activity and 44.64
Gymin-1 dose rate in summer squash while these values
were 1,482 Ci and 11.96 Gymin-1 in the present study,
respectively. Based on this concept, source activity (Ci),
dose rate (Gymin-1), pollen characteristics (size, sensitiv-
ity condition of dehydration) and irradiation and
Table 4 Effects of embryo type (ET) and embryo stage (ES) on
haploid plantlet number (HN) and haploid plantlet rate (HR) (%)
ET ES E P IP HN HR (%)
N G 43 0 0 0 0.0
H 39 0 0 0 0.0
A 32 10 8 4 50.0
LT 28 8 6 2 33.3
PC 25 4 3 2 66.7
C 117 68 49 0 0.0
AM 23 5 4 0 0.0
NE G 48 0 0 0 0.0
H 49 0 0 0 0.0
A 53 0 0 0 0.0
LT 34 0 0 0 0.0
PC 30 0 0 0 0.0
C 124 7 3 0 0.0
AM 38 1 0 0 0.0
RN 307 95 73 8 10.96
RNE 379 8 0 0 0.0
E Embryo number, P plant number, IP investigated plant number,
G globular, H heart, A arrow-tip, LT late-torpedo, PC pro-cotyledon,
C cotyledon, AM amorphous, N normal, NE necrotic
Table 5 Frequency of haploid production
IT ID HP S E IP F 100
seeds
100
embryos
100
plantlets
Per
fruit
9 50 2 2,136 216 25 9 0.09 0.93 8 0.22
100 0 767 7 0 7 0 0 0 0
11 50 4 2,185 312 33 9 0.18 1.28 12.2 0.44
100 0 794 6 0 7 0 0 0 0
15 50 2 430 122 15 5 0.47 1.64 13.3 0.4
100 0 425 0 0 4 0 0 0 0
21 50 0 124 12 0 3 0 0 0 0
100 0 92 0 0 2 0 0 0 0
28 50 0 106 11 0 3 0 0 0 0
100 0 12 0 0 1 0 0 0 0
R 50 8 4,981 673 73 29 0.16 1.19 10.96 0.28
100 0 2,090 13 0 21 0 0 0 0
Overall 8 7,071 686 73 50 0.11 1.17 10.96 0.28
IT Irradiation time in July, ID irradiation dose (Gray), HP haploid
plant number, S seed number, E embryo number, IP investigated plant
number, F fruit number
Table 6 Stomata dimension and density of haploid and diploid plants
SL SW SD CN
Haploid 21.82 b 17.37 b 428.6 a 7.21 b
Diploid 30.51 a 21.11 a 311.4 b 11.17 a
Means followed by the same letter in the same column are not sig-
nificantly different
SL (lm) Stomata length, SW (lm) stomata width, SD (number/per
mm2) stomata density, CN chloroplast number
274 Plant Cell Tiss Organ Cult (2010) 102:267–277
123
Author's personal copy

pollination details (time from collection to irradiation and
incubation and storage conditions of anthers) must be
clarified in irradiation experiments like this.
Embryo induction (number/per fruit) was achieved from
all investigated genotypes and varied in the range of 3.5–
52.0. Genotype specificity was observed, but the general
reaction of the genotypes was low. When comparing the
average values, ‘‘G14’’ (21.2) demonstrated good response.
Data presented in this paper confirm the relationship
between embryo induction and genotype that has been
previously demonstrated for cucumber (Sauton 1989; Faris
et al. 1999), summer squash (Kurtar et al. 2002) and
pumpkin (Kurtar et al. 2009). Genotypic effect can be
explained by the responsiveness of parthenogenesis
(Pandey and Phung 1982) or lower parthenocarpic response
(Sauton 1989; Faris et al. 1999). Moreover, embryo yield
was highly influenced by different factors such as irradia-
tion dose, irradiation duration, and genotype (Brewbaker
and Emery 1962; Niemirowicz-Szczytt and Dumas de
Vaulx 1989; Sauton 1989; Sari et al. 1992, 1994;
Ficcadenti et al. 1995; Abak et al. 1996; Caglar and Abak
1999; Kurtar et al. 2002, 2009).
Overall, 686 embryos were rescued and the number of
necrotic embryos (379) was higher than normal embryos
(307) due to delayed harvest times (5 and 6 weeks). A total
of 103 plantlets were regenerated from 686 embryos, and
the regeneration rate was 15.0%. The greatest number of
plantlets (95 plantlets) were regenerated from normal
embryos. Regeneration rate changed with embryo type and
embryo stage. Globular and heart embryos were not
regenerated, and the greatest regeneration rate was
observed in cotyledon embryos (58.9%). Regeneration rate
was determined 31.3% in arrow-tip, 28.6% in late-torpedo,
and 16.0% in pro-cotyledon embryos. Regeneration rate
was reported 12.0% in arrow-tip, 31.3% in torpedo, 44.4%
in heart, and 31.3% in cotyledon embryos in summer
squash (Kurtar et al. 2002). On the other hand, cotyledon
embryos had the highest regeneration rate (66.7%), and
torpedo (45.5%), heart (44.0%) and arrow-tip (41.7%)
embryos also gave better results in pumpkin (Kurtar et al.
2009). Regeneration rate was 80% in heart embryos and
20% in globular embryos in melon (Sari et al. 1999b).
28.1% of globular, 55.0% of heart, 80% of torpedo, and
81.8% of cotyledon type embryos were regenerated in
cucumber (Caglar and Abak 1999). However, Faris et al.
(1999) found relatively lower regeneration efficiency (3.3–
7.7%) in cucumber. The success of embryo rescue depends
on the stage of embryo development and composition of
culture media (Jaskani et al. 2005). Different culture media
and several subcultures may be used for increasing
regeneration efficiency (Ondrej et al. 2002).
Embryo rescue is a very laborious and tedious process in
winter squash. Opening 100 seeds within a fruit takes
approximately 70–80 min. Therefore, liquid medium cul-
ture (Lotfi et al. 2003) was investigated to facilitate embryo
rescue. This procedure appeared simple and convenient,
but it was found to be ineffective in winter squash.
Embryos were not visualized and identified because of
having the thick seed-coat. Moreover, contamination was
occurred in magenta boxes excessively and led to embryo
loss. In a solution, X-ray technique can also be used for
identification of embryos (Savin et al. 1988; Sauton 1989).
But this technique requires specific equipment which is not
available in many laboratories.
The frequency of haploid plantlets was the highest at
earlier harvest times (3rd and 4th week), because of later
harvest times (5th and 6th week) led to a greater number of
necrotic embryos (Renata and Visser 1987; De Witte 2000;
Taner et al. 2000). Overall, 8 haploid plants were regen-
erated from arrow-tip (4), late-torpedo (2) and pro-cotyle-
don (2) embryos. The frequency of haploid plantlets was
0.11, 1.17, 10.96 and 0.28 per 100 seeds, 100 embryos, 100
plantlets and fruit, respectively. Frequency of haploid
plantlets was found per 100 seeds, 100 embryos and fruit
1.2, 10.4 and 0.7 in summer squash (Kurtar 1999), and
0.24, 0.94 and 0.33 in pumpkin (Kurtar et al. 2009),
respectively. Low frequency of haploid (range 0.11–0.36)
was also reported in cucumber (Caglar and Abak 1999).
Fig. 3 Stomata dimension and chloroplast number in guard cells: haploid (a), diploid (b)
Plant Cell Tiss Organ Cult (2010) 102:267–277 275
123
Author's personal copy

Stomata size, stomata density, chloroplast number and
chromosome number in root tip were different from hap-
loid to diploid plants. Measurement of stoma and chloro-
plast counting was simple and more practical than
chromosome counting. These results indicated that stomata
observations can be used successfully to determine the
ploidy in winter squash. Abak et al. (1996) in melon, Sari
et al. (1999a) in watermelon, Kurtar et al. (2002) in sum-
mer squash and Kurtar et al. (2009) in pumpkin reported
similar results. Flow-cytometry can be used a potential
method to determine the ploidy level, but it is expensive,
labour-intensive and requires specific equipment (Sari et al.
1999a). Besides, no convenient method for flow-cytometry
has been found by many researchers working on haploid/
dihaploid production (Lim and Earle 2009) because many
plants scored as diploid by flow-cytometry do not produce
pollen or fruit-set in vivo (Lim and Earle 2008). This may
be explained by genetic modification of plants (especially
polyploidy, aneuploidy, euploidy and mixopoidy) in tissue
culture (Berlyn et al. 1986). Aneuploid plants were also
reported in cucumber (Truong-Andre 1988; Niemirowicz-
Szczytt and Dumas de Vaulx 1989) and in melon (Ezura
and Oosawa 1994). Considering these findings, in addition
to chromosome counting and stomata observations, mor-
phological observation (checking of flowers and pollen,
fruit-set and growth characteristics, etc.) seems to be nec-
essary to determine the ploidy level in winter squash.
However, morphological observations are time consuming,
as it requires plant development to reach an appropriate
stage.
Although our results indicated low frequency of haploid
plantlets, this is the first report on induction of partheno-
genetic haploid embryo via irradiated pollen technique in
winter squash. Low frequency of haploid may be explained
by the greatest number of necrotic embryos (due to delayed
harvest times), absence of regeneration (globular and heart
embryos were not regenerate) and spontaneous diploidi-
zation. Spontaneous diploidization has also been reported
for root meristems of cucumber (Sauton 1989) and melon
(Sauton 1988; Lotfi et al. 2003).
In conclusion, the irradiated pollen technique is pro-
posed to induce embryos and obtain haploid plantlets in
winter squash. However, in order to evaluate the irradiated
pollen approach for the recovery of haploid plants in winter
squash, further research should be based on appropriate
irradiation doses, irradiation durations, genotypes, culture
media and culture conditions, and also acclimatization
processes.
Acknowledgments We gratefully acknowledge the support of the
Scientific and Technical Research Council of Turkey (TUB_ITAK
Project No: TOVAG 108 O 390) and partial funding by the Voca-
tional School of Bafra, Ondokuz Mayis University in Turkey.
References
Abak K, Sari N, Paksoy M, Yılmaz H, Aktas H, Tunalı C (1996)
Genotype response to haploid embryo induction with pollination
by irradiated pollen in melon, obtaining of dihaploid lines,
determination of haploid plants by different techniques. Turk J
Agric For 20:425–430
Alison A, Casareft P (1968) Radiation biology. United States Atomic
Energy Commission, Washington, DC
Athanasios LT, Koutita O, Anastasiadou A (2009) Description and
analysis of genetic diversity among squash accessions. Braz
Arch Biol Technol 52:271–283
Berlyn GP, Beck RC, Renfroe MH (1986) Tissue culture and the
propagation and genetic improvement of conifers: problems and
possibilities. Tree Physiol 1:227–240
Bernstein EF, Sullivan FJ, Mitchel JB (1993) Biology of chronic
radiation effect on tissues and wound healing. Clin Plast Surg
20:435–453
Brewbaker JL, Emery GC (1962) Pollen radiobotany. Radiat Bot
1:101–154
Caglar G, Abak K (1999) Obtention of in vitro haploid plants from in
situ induced haploid embryos in cucumber (Cucumis sativus L.).
Turk J Agric For 23:283–290
Chahal GS, Gosal SS (2002) Principles and procedures of plant
breeding. Alpha Science, Oxford
Cuny F (1992) Processus d’induction d’embryons haploides par du
pollen irradie chez le melon (Cucumis melo L.) responses du
pollen a l’irradiation gamma. These de Docteur, Universite
d’Avignon et des Pays de Vaucluse, Avignon
Darlington CD, La Cour LF (1963) Methoden der chromosome-
nuntersuchungten. Keller, Stuttgart
De Witte K (2000) Review of research at fruitteeltcentrum on the
production of homozygous plants through androgenesis in vitro
and parthenogenesis in situ. Acta Hort 520:135–148
Decker-Walters DS, Walters TW, Posluszny U (1990) Genealogy and
gene flow among annual and domesticated species of Cucurbita.
Can J Bot 68:782–789
Dolcet-Sanjuan R, Claveria E, Garcia-Mas J (2006) Cucumber
(Cucumis sativus L.) dihaploid line production using in vitro
rescue of in vivo induced parthenogenic embryos. Acta Hort
725:837–844
Dore C (1986) Evaluation du niveau de ploidie des plantes d’une
population de choux de Bruxelles (Brassica oleracea L. ssp.
gemmifera) d’origine pollinique. Agronomie 6:797–801
Ezura H, Oosawa K (1994) Ploidy of somatic embryos and the ability
to regenerate plantlets in melon (Cucumis melo L.). Plant Cell
Rep 14:107–111
Faris NM, Nikolova V, Niemirowicz-Szczytt K (1999) The effect of
gamma irradiation dose on cucumber (Cucumis sativus L.)
haploid embryo production. Acta Phys Plant 21:301–396
Ficcadenti N, Veronese P, Sestili S, Crino P, Lucretti S, Schiavi M,
Saccardo F (1995) Influence of genotype on the induction of
haploidy in Cucumis melo L. by using irradiated pollen. J Genet
Breed 49:359–364
Germana MA (2006) Doubled haploid production in fruit crops. Plant
Cell Tiss Organ Cult 86:131–146
Goldschmidt H, Breneman JC, Breneman DL (1994) Ionizing
radiation therapy in dermatology. J Am Acad Dermatol
30:157–182
Gursoz N, Abak K, Pitrat M, Rode JC, Dumas de Vaulx R (1991) Obten-
tion of haploid plants induced by irradiated pollen in watermelon
(Citrullus lanatus). Cucurbit Genetic Coop 14:109–110
Heikal AH, Abdel-Razzak HS, Hafez EE (2008) Assessment of
genetic relationships among and within cucurbita species using
RAPD and ISSR markers. J App Sci Res 4:515–525
276 Plant Cell Tiss Organ Cult (2010) 102:267–277
123
Author's personal copy

Jain SM, Sopory SK, Veilleux RE (1996) In vitro haploid production
in higher plants. Kluwer, Dordrecht
Jaskani MJ, Khan IA, Khan MM (2005) Fruit set, seed development
and embryo germination in interploid crosses of citrus. Scientia
Hort 107:51–57
Kurtar ES (1999) Research on the effects of genotypes and growing
seasons on in situ haploid embryo induction and in vitro plant
obtention via irradiated pollen in squash. PhD Thesis, University
of Cukurova
Kurtar ES (2009) Influence of gamma irradiation on pollen viability,
germinability and fruit and seed-set of pumpkin and winter
squash. Afr J Bio 8:6918–6926
Kurtar ES, Sari N, Abak K (2002) Obtention of haploid embryos and
plants through irradiated pollen technique in squash (Cucurbitapepo L.). Euphytica 127:335–344
Kurtar ES, Balkaya A, Ozbakır M, Ofluoglu T (2009) Induction of
haploid embryo and plant regeneration via irradiated pollen
technique in pumpkin (Cucurbita moschata Duchesne ex. Poir).
Afr J Bio 8:5944–5951
Lacadena JR (1974) Spontaneous and induced parthenogenesis and
androgenesis. In: Kasha KJ (ed) Haploids in higher plants—
advances and potential. University of Guelph, Guelph, Canada,
pp 13–32
Lim W, Earle ED (2008) Effect of in vitro and in vivo colchicine
treatments on pollen production and fruit recovery on melon
plants obtained after pollination with irradiated pollen. Plant Cell
Rep 95:115–124
Lim W, Earle ED (2009) Enhanced recovery of doubled haploid lines
from parthenogenetic plants of melon (Cucumis melo L.). Plant
Cell Rep 98:351–356
Lotfi M, Kashi A, Onsinejad R (1999) Induction of parthenogenetic
embryos by irradiated pollen in cucumber. Acta Hort 492:323–
328
Lotfi M, Alan AR, Henning MJ, Jahn MM, Earle ED (2003)
Production of haploid and doubled haploid plants of melon
(Cucumis melo L.) for use in breeding for multiple virus
resistance. Plant Cell Rep 21:1121–1128
Maestro-Tejada MC (1992) Resistance du melon aux virus. Interac-
tion avec les pucerons vecteurs. Analyse genetique sur les
lignees haplodiploides. These de Docteur, Universite de Droit,
d’Economie et des Sciences d’Aix-Marseille
Nepi M, Pacini E (1993) Pollination, pollen viability and pistil
receptivity in Cucurbita pepo. Ann Bot 72:527–536
Niemirowicz-Szczytt K, Dumas de Vaulx R (1989) Preliminary data
on haploid cucumber (Cucumis sativus L.) induction. Cucurbit
Genetics Coop 12:24–25
Ondrej V, Navratilova B, Lebeda A (2002) In vitro cultivation of
Cucumis sativus ovules after fertilization. Acta Hort 588:339–343
Ozalpan A (2001) Temel Radyobiyoloji. Halic University Press,
Cambridge
Pandey KK (1978) Gametic gene transfer in Nicotiana by means of
irradiated pollen. Genetica 49:53–69
Pandey KK, Phung M (1982) Hertwig effect in plants: induced
parthenogenesis through the use of irradiated pollen. Theor Appl
Genet 62:295–300
Pochard E, Dumas de Vaulx R (1971) La monoploidie chez le piment
(Capsicum annuum L.). Z Pflanzenzuchtg 65:23–46
Raghavan V (1986) Embryogenesis in angiosperms: a developmental
and experimental study. Cambridge University Press, Cambridge
Raquin C, Cornu A, Farcy E, Maizonnier D, Pelletier G, Vedel F
(1989) Nucleus substitution between Petunia species using
gamma ray-induced androgenesis. Theor Appl Genet 78:337–
341
Renata S, Visser T (1987) Embryo development and fruit set in pear
induced by untreated and irradiated polen. Euphytica 36:287–
294
Rouselle F (1992) Techniques d’estimation nombre de chloroplastes.
In: Jahier J et al (eds) Techniques de Cytogenetique Vegetale.
INRA , Paris
Sari N (1994) Effect of genotype and season on the obtention of
haploid plants by irradiated pollen in watermelon and alterna-
tives to the irradiation. PhD Thesis, University of Cukurova
Sari N, Abak K, Pitrat M, Dumas de Vaulx R (1992) Induction of
parthenogenetic haploid embryos and plant obtention in melon
(Cucumis melo L. var. inodorus Naud ve C. melo L. var.
reticulatus Naud). Trans J Agric For 16:302–314
Sari N, Abak K, Pitrat M, Rode JC, Dumas de Vaulx R (1994)
Induction of parthenogenetic haploid embryos after pollination
by irradiated pollen in watermelon. HortScience 29:1189–1190
Sari N, Abak K, Pitrat M (1999a) Comparison of ploidy level
screening methods in watermelon (Citrullus lanatus (Thunb.)
Matsum. and Nakai). Scientia Hort 82:265–277
Sari N, Ekiz H, Yucel S, Yetisir H, Ekbic H, Abak K (1999b)
Investigation of new protected cultivation melon lines resistant
to Fusarium oxysporium f.sp. melonis using dihaplodization.
Turkey IIIrd National Horticultural Congress, 14–17 September
1999, Ankara, pp 498–503
Sauton A (1988) Effect of season and genotype on gynogenetic haploid
production in muskmelon, Cucumis melo L. Sci Hort 35:71–75
Sauton A (1989) Haploid gynogenesis in Cucumis sativus induced by
irradiated pollen. Cucurbit Genetics Coop 12:22–23
Sauton A, Dumas De Vaulx R (1987) Obtention de plantes haploides
chez le melon (Cucumis melo L.) par gynogenese induite par du
pollen irradie. Agronomie 7:141–148
Savaskan C, Toker MC (1991) The effects of various doses of gamma
irradiation on the seed germination and root tips chromosomes of
rye (Secale cereale L.). Trans J Bot 15:349–359
Savin F, Decomble V, Le Couviour M, Hallard J (1988) The X-ray
detection of haploid embryos arisen in muskmelon (Cucumis meloL.) seeds, and resulting from a parthenogenetic development
induced by irradiated pollen. Cucurbit Genetics Coop 11:39–42
Sensoy AS, Ercan N, Ayar F, Temirkaynak M (2003) Determination
of some morphologic characteristics and viability of some
vegetable crops in Cucurbitaceae family. J Agric Fac Akdeniz
Univ 16:1–6
Shridhar (1992) Pollen grains of cultivated Cucurbits. In: Proceedings
of the 5th Eucarpia Cucurbitaceae Symp, July 27–31, Warsaw,
Poland, 28–33
Stairs GR, Mergen F (1964) Potential uses of irradiated pollen in
forest genetics. In: Proceedings of the 11th northeastern forest
tree improved conference, pp 38–41
Sun Y, Mei S, Peng J, Zhang L, Nie Q, Zeng H, Du N (2006) Induced
haploid plants after pollination by irradiated pollen in Cucumismelo L. Hubei Agr Sci 4:98–100
Taner KY, Yanmaz R, Kunter B (2000) The effects of irradiation dose
and harvest period on haploid plant formation via irradiated
pollen in snake cucumber (Cucumis melo var. flexuosus Naud.).
IIIrd National Vegetable Culture Symp, Isparta-Turkey, 177–181
Todorova M, Ivanov P, Ninova N, Encheva J (2004) Effect of female
genotype on the efficiency of induced perthenogenesis in
sunflower (Helianthus annuus L.). Hella 27:67–74
Tokarek R, Bernstein EF, Sullivan F (1994) Effect of therapeutic
radiation on wound healing. Clin Dermatol 12:57–70
Truong-Andre I (1988) In vitro haploid plants derived from pollin-
isation by irradiated pollen on cucumber. In: Proceedings of the
Eucarpia meeting on cucurbit genetics and breeding. May 31–
June 2, Avignon-Montfavet, pp 143–144
Whitaker TW, Bemis WP (1964) Evolution in the genus. Cucurbita.
Evolution 18:553–559
Xie M, Zhao J, Pan J, He H, Wu A, Cai R (2005) Induced haploid
plants after pollination by irradiated pollen in Cucumis sativus L.
J Shanghai Jiaotong Univ (Agr Sci) 2:45–49
Plant Cell Tiss Organ Cult (2010) 102:267–277 277
123
Author's personal copy
Copyright © 2022 FDOKUMEN








