
Pattern of forebrain activation in high novelty-seeking rats following aggressive encounter
13
This article appeared in a journal published by Elsevier. The attached copy is furnished to the author for internal non-commercial research and education use, including for instruction at the authors institution and sharing with colleagues. Other uses, including reproduction and distribution, or selling or licensing copies, or posting to personal, institutional or third party websites are prohibited. In most cases authors are permitted to post their version of the article (e.g. in Word or Tex form) to their personal website or institutional repository. Authors requiring further information regarding Elsevier’s archiving and manuscript policies are encouraged to visit: http://www.elsevier.com/copyright
-
Upload
independent -
Category
Documents
-
view
0 -
download
0
Transcript of Pattern of forebrain activation in high novelty-seeking rats following aggressive encounter

This article appeared in a journal published by Elsevier. The attachedcopy is furnished to the author for internal non-commercial researchand education use, including for instruction at the authors institution
and sharing with colleagues.
Other uses, including reproduction and distribution, or selling orlicensing copies, or posting to personal, institutional or third party
websites are prohibited.
In most cases authors are permitted to post their version of thearticle (e.g. in Word or Tex form) to their personal website orinstitutional repository. Authors requiring further information
regarding Elsevier’s archiving and manuscript policies areencouraged to visit:
http://www.elsevier.com/copyright

Author's personal copy
Research Report
Pattern of forebrain activation in high novelty-seeking ratsfollowing aggressive encounter
Sarah M. Clintona,⁎, 1, Ilan A. Kermana, 1, Hailey R. Orrb, Tracy A. Bedrosianc,Antony D. Abrahamd, Danielle N. Simpsona, Stanley J. Watsonb, Huda Akilb
aDepartment of Psychiatry and Behavioral Neurobiology, University of Alabama-Birmingham, Birmingham, AL, USAbMolecular and Behavioral Neuroscience Institute, University of Michigan, Ann Arbor, MI, USAcNeuroscience Graduate Studies Program, Ohio State University, Columbus, OH, USAdBehavioral Neuroscience Program, Oregon Health and Science University, Portland, OR, USA
A R T I C L E I N F O A B S T R A C T
Article history:Accepted 14 August 2011Available online 19 August 2011
We have previously demonstrated that selectively-bred High (bHR) and Low (bLR) novelty-seeking rats exhibit agonistic differences, with bHRs acting in a highly aggressive mannerwhen facing homecage intrusion. In order to discover the specific neuronal pathways responsi-ble for bHRs' high levels of aggression, the present study compared c-fos mRNA expression inseveral forebrain regions of bHR/bLR males following this experience. bHR/bLR males werehoused with female rats for 2 weeks, and then the females were replaced with a male intruderfor 10min. bHR/bLR residents were subsequently sacrificed by rapid decapitation, and theirbrains were removed and processed for c-fos in situ hybridization. Intrusion elicited robust c-fosmRNAexpression in both phenotypes throughout the forebrain, including the septum, amygda-la, hippocampus, cingulate cortex, and the hypothalamus. However, bHRs and bLRs exhibiteddistinct activation patterns in select areas. Compared to bHR rats, bLRs expressed greater c-fosin the lateral septum andwithinmultiple hypothalamic nuclei, while bHRs showed greater acti-vation in the arcuate hypothalamic nucleus and in the hippocampus. No bHR/bLR differences inc-fos expressionwere detected in the amygdala, cortical regions, and striatum.We also found di-vergent 5-HT1A receptor mRNA expression within some of these same areas, with bLRs havinggreater 5-HT1A, but not 5-HT1B, receptor mRNA levels in the septum, hippocampus and cingu-late cortex. These findings, together with our earlier work, suggest that bHRs exhibit altered se-rotonergic functioning within select circuits during an aggressive encounter.
© 2011 Elsevier B.V. All rights reserved.
Keywords:c-fosBred High Responder (bHR)Bred Low Responder (bLR)SeptumHypothalamusHippocampusAggressionResident–intruder testSerotonin5-HT1A receptor5-HT1B receptor
1. Introduction
Aggression is a common behavior naturally expressed in re-sponse to threat or other environmental challenges. While ag-gression may be appropriate in some instances and importantfor an organism's survival, extreme or uncontrollable aggression
is maladaptive. Impulsive aggression is a common symptom ofseveralmental illnesses, includingattention-deficit hyperactivitydisorder, autism, and some mood disorders (Davidson et al.,2000). Therefore, there is great interest in understanding geneticand biological factors that may predispose certain individuals tobe more aggressive than others.
B R A I N R E S E A R C H 1 4 2 2 ( 2 0 1 1 ) 2 0 – 3 1
⁎ Corresponding author at: 1720 7th Avenue South, Sparks 745, Birmingham, AL 35294, USA.E-mail address: [email protected] (S.M. Clinton).
1 These authors contributed equally.
0006-8993/$ – see front matter © 2011 Elsevier B.V. All rights reserved.doi:10.1016/j.brainres.2011.08.033
Ava i l ab l e on l i ne a t www.sc i enced i r ec t . com
www.e l sev i e r . com/ loca te /b ra i n res

Author's personal copy
Previous studies using laboratory rats andmice have bred an-imals with either naturally high or low levels of aggression, indi-cating that aggressive behavior is strongly influenced by genetics(e.g. Veenema andNeumann, 2007). In recent years, our researchgroup developed two lines of Sprague–Dawley rats based on in-nate differences in emotional or environmental reactivity(Stead et al., 2006). Our selectively-bred High Responder (bHR)rats are hyperactive and vigorously explore novel environments,exhibit elevated impulsivity (Flagel et al., 2010) and enhancedpsychostimulant self-adminstration (Davis et al., 2008) comparedto bred LowResponder (bLR) rats. bLRs, on the other hand, are be-haviorally inhibited, showing much less novelty-induced loco-motor activity and exaggerated anxiety- and depression-likebehavior compared to bHR animals (Clinton et al., 2008; Steadet al., 2006). More recently we found that bHR rats exhibit greaterinter-male aggression compared to bLRs in the resident–intruderparadigm (see companion paper, Kerman, Clinton et al., 2011).When an intruder is introduced into the homecage, bHR animalsreact by aggressively sniffing, attacking and biting the intruderrat, while bLRs aremuchmore passive and sometimes even sub-missive to the intruder (see companion paper, Kerman, Clintonet al., 2011). These divergent behaviors were accompanied byhormonal differences, with bHRs showing enhanced corticoste-rone and testosterone secretion following intrusion comparedto bLRs. Furthermore, these differences appear to be linked to un-derlying bHR/bLR differences in serotonin (5-HT) circuits, sincewe found both baseline and intrusion-evoked gene expressiondifferences within several brainstem 5-HTergic nuclei (see com-panion paper, Kerman, Clinton et al., 2011). Based on these find-ings, we hypothesize activation of specific forebrain circuits inbHR/bLR rats following homecage intrusion, which may contrib-ute to their markedly different behavioral and hormonal re-sponses. In the present study we utilized c-fos mRNAexpression as amarker of neuronal activation to identify the pat-tern of forebrain activation in bHR/bLR males following homec-age intrusion experience (Table 1).
Abundant clinical and preclinical evidence indicates that per-turbed 5-HT neurotransmission plays an important role in regu-lating aggressive behavior (Miczek et al., 2007). Several studiespoint to deficient 5-HTergic transmissionprecipitating anaggres-sive state, including observations of reduced 5-HTmetabolites inthe cerebrospinal fluid of aggressive rodents (Giacalone et al.,1968), monkeys (Higley et al., 1992; Westergaard et al., 1999),and humans (Brown et al., 1979; Kruesi et al., 1990), as well as en-hancement of aggressive behavior following diet-induced trypto-phan depletion (Delgado et al., 1990; Pihl et al., 1995). Ourprevious results suggest that bHRs' aggressive tendencies maystem from a deficiency within certain 5-HTergic cell groups.Thus, a second aim of the present study is to examine 5-HT1Aand 5-HT1B mRNA receptor expression in several forebrain ter-minal regions known to receive 5-HT projections.
2. Results
2.1. Homecage intrusion-induced c-fos mRNA expression
Wepreviously found that bHRmales exhibit greater aggressivebehavior than bLRswhen facing homecage intrusion (see com-panion paper, Kerman, Clinton et al., 2011). The present study
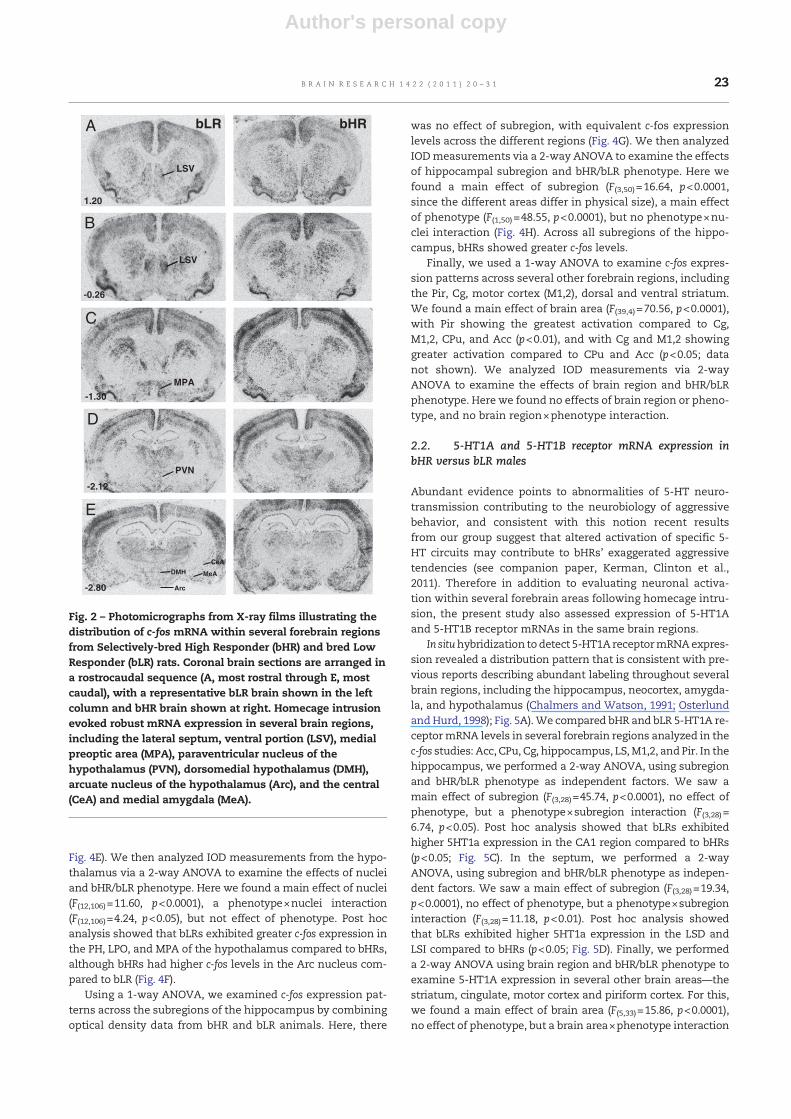
evaluated expression of the immediate early gene c-fos as anindicator of forebrain regional activation following exposureto the resident–intruder paradigm (Fig. 1). Robust c-fos mRNAexpression was detected throughout the forebrain of bHR/bLRresident males at 15 min following initiation of intruder expo-sure, with robust expression within the lateral septum (LS),amygdala, several hypothalamic nuclei, the piriform cortex(Pir), cingulate cortex (Cg), motor cortex (M1,2), caudate puta-men (CPu), nucleus accumbens (Acc), and the hippocampus(Cornu Ammonis fields CA1-CA3, and the dentate gyrus (DG))(Fig. 2). Closer inspection of these data revealed heterogeneousc-fos expression, which was confined to specific subregionswithin several of these brain areas (Fig. 3).
Using a 1-way ANOVA, we examined c-fos expression pat-terns across subregions of the LS by combining optical densitydata from bHR and bLR animals. This analysis revealed amaineffect of subregion in the LS (F(2,24)=6.29, p<0.01), with theventral portion of the LS (LSV) showing greater c-fos levelscompared to the intermediate (LSI; p<0.05) and dorsal (LSD;p<0.01) parts (Fig. 4A). We then analyzed integrated opticaldensity (IOD) measurements via a 2-way ANOVA to examinethe effects of subregion and bHR/bLR phenotype. Here wefound a main effect of phenotype (F(1,21=15.35, p<0.001), sub-region (F(2,21)=10.58, p<0.001), and a significant phenotype×subregion interaction (F(2,21)=3.48, p<0.05). Post hoc analyses
Table 1 – C-fos expression patterns in brain followingaggressive encounter.
Brain region bHR bLR
HypothalamusAH 0.829±0.126 1.139±0.362DMH 0.879±0.106 0.824±0.136LH 0.906±0.229 0.926±0.144PMD 1.367±0.169 1.387±0.557PMV 1.059±0.187 1.002±0.412PVN 1.825±0.271 2.054±0.307SCN 1.083±0.193 1.125±0.084TC 0.616±0.116 0.737±0.252VMH 0.056±0.014 0.101±0.038
AmygdalaBLA 0.898±0.192 0.904±0.292CeA 0.225±0.069 0.136±0.037LaA 0.607±0.125 0.698±0.205MeA 1.988±0.445 2.310±1.390
ForebrainAcc 3.493±0.532 3.153±0.653Cg 11.234±0.806 9.626±1.634CPu 12.252±1.173 10.152±2.291M1,2 9.985±0.752 7.377±1.660Pir 12.114±0.332 7.096±0.819
Values represent average integrated optical density values±standard errors×100.Abbreviations: AH - anterior hypothalamus; DMH - dorsomedialhypothalamus; LH - lateral hypothalamus; PMD - dorsalpremammillary nucleus; PMV - ventral premammillary nucleus;PVN - paraventricular nucleus; SCN - suprachiasmatic nucleus; TC -tuber cinereum; VMH - ventromedial hypothalamus; BLA -basolateral amygdala; CeA - central amygdala; LaA - lateralamygdala; MeA - medial amygdala; Acc - nucleus accumbens; Cg -cingulate cortex; CPu - caudate putamen; M1,2 - motor cortex 1 and2; Pir - piriform cortex.
21B R A I N R E S E A R C H 1 4 2 2 ( 2 0 1 1 ) 2 0 – 3 1

Author's personal copy
showed that bLRs exhibited greater c-fos expression in the LSV(p<0.01) and LSI (p<0.01) compared to bHRs (Fig. 4B).
Using a 1-way ANOVA, we also examined c-fos expressionpatterns across select nuclei of the amygdala by combiningoptical density data from bHR and bLR animals. We found amain effect of nuclei (F(3,36)=5.06, p<0.01), with greater activa-tion in the medial amygdala (MeA) relative to the basolateral,lateral, and central (CeA) nuclei (p<0.05) (Fig. 4C). We then an-alyzed IODmeasurements via a 2-way ANOVA to examine theeffects of nuclei and bHR/bLR phenotype. Here we found amain effect of nuclei (F(3,29)=8.41, p<0.0001), but no effect ofphenotype, and no phenotype×nuclei interaction (Fig. 4D).
Using a 1-way ANOVA, we examined c-fos expression pat-terns across select nuclei of the hypothalamus by combining
optical density data from bHR and bLR animals. We found amain effect of nuclei (F(12,119)=18.42, p<0.0001), with the para-ventricular nucleus (PVN) showing the greatest activation(p<0.0001 compared to all other nuclei). The ventromedial hy-pothalamus (VMH) showed the least c-fos expression (p<0.05compared to all nuclei except the lateral hypothalamus (LH)and lateral preoptic area (LPO)). The remaining nuclei can beroughly divided into two groups, with one group showing an in-termediate to high level of c-fos (dorsal premammillary nucleus(PMD), arcuate nucleus (Arc), dorsomedial hypothalamus(DMH), and ventral premammillary nucleus (PMV)), and theother group showing a comparatively lower activation level(tuber cinereum (TC), medial preoptic area (MPA), posterior hy-pothalamus (PH), anterior hypothalamus (AH), LH, and LPO;
Bregma 1.20 mm
Cg
M2
CPu
Pir
M1
Bregma -0.26 mm
ccLSD
LSI
ox
LPOMPA
LSV
Bregma -1.30 mm
ox
cc
PVN
cc
AH
TC
Ar c
LHf
BLA
LaACeA
MeA
DGCA3CA2CA1cc
f
opt
PMV
PMD
f
PH
cc
Bregma -2.12 mm
Bregma -2.80 mm
Bregma -4.16 mm
Acc
VMH
DMH
opt
Fig. 1 – Location of templates used to sample integrated optical density measurements within specific brain regions.Abbreviations: Acc—nucleus accumbens; AH—anterior hypothalamus; Arc—arcuate nucleus; BLA—basolateral amygdala;cc—corpus callosum; CeA—central amygdala; Cg—cingulate cortex; CPu—caudate-putamen; DG—dentate gyrus;DMH—dorsomedial hypothalamus; f—fornix; LaA—lateral amygdala; LH—lateral hypothalamus; LPO—lateral preoptic area;LSD—dorsal lateral septal nucleus; LSI—intermediate lateral septal nucleus; LSV—ventral lateral septal nucleus; M1,2—motorcortex 1 and 2; MeA—medial amygdala; MPA—medial preoptic area; opt—optic tract; ox—optic chiasm;PH—posterior hypothalamus; Pir—piriform cortex; PMD—dorsal premammillary nucleus; PMV—ventral premammillary nucleus;PVN—paraventricular nucleus; TC—tuber cinereum; VMH—ventromedial hypothalamus.
22 B R A I N R E S E A R C H 1 4 2 2 ( 2 0 1 1 ) 2 0 – 3 1
Author's personal copy
Fig. 4E). We then analyzed IOD measurements from the hypo-thalamus via a 2-way ANOVA to examine the effects of nucleiand bHR/bLR phenotype. Here we found a main effect of nuclei(F(12,106)=11.60, p<0.0001), a phenotype×nuclei interaction(F(12,106)=4.24, p<0.05), but not effect of phenotype. Post hocanalysis showed that bLRs exhibited greater c-fos expression inthe PH, LPO, and MPA of the hypothalamus compared to bHRs,although bHRs had higher c-fos levels in the Arc nucleus com-pared to bLR (Fig. 4F).
Using a 1-way ANOVA, we examined c-fos expression pat-terns across the subregions of the hippocampus by combiningoptical density data from bHR and bLR animals. Here, there
was no effect of subregion, with equivalent c-fos expressionlevels across the different regions (Fig. 4G). We then analyzedIODmeasurements via a 2-way ANOVA to examine the effectsof hippocampal subregion and bHR/bLR phenotype. Here wefound a main effect of subregion (F(3,50)=16.64, p<0.0001,since the different areas differ in physical size), a main effectof phenotype (F(1,50)=48.55, p<0.0001), but no phenotype×nu-clei interaction (Fig. 4H). Across all subregions of the hippo-campus, bHRs showed greater c-fos levels.
Finally, we used a 1-way ANOVA to examine c-fos expres-sion patterns across several other forebrain regions, includingthe Pir, Cg, motor cortex (M1,2), dorsal and ventral striatum.We found a main effect of brain area (F(39,4)=70.56, p<0.0001),with Pir showing the greatest activation compared to Cg,M1,2, CPu, and Acc (p<0.01), and with Cg and M1,2 showinggreater activation compared to CPu and Acc (p<0.05; datanot shown). We analyzed IOD measurements via 2-wayANOVA to examine the effects of brain region and bHR/bLRphenotype. Here we found no effects of brain region or pheno-type, and no brain region×phenotype interaction.
2.2. 5-HT1A and 5-HT1B receptor mRNA expression inbHR versus bLR males
Abundant evidence points to abnormalities of 5-HT neuro-transmission contributing to the neurobiology of aggressivebehavior, and consistent with this notion recent resultsfrom our group suggest that altered activation of specific 5-HT circuits may contribute to bHRs' exaggerated aggressivetendencies (see companion paper, Kerman, Clinton et al.,2011). Therefore in addition to evaluating neuronal activa-tion within several forebrain areas following homecage intru-sion, the present study also assessed expression of 5-HT1Aand 5-HT1B receptor mRNAs in the same brain regions.
In situhybridization todetect 5-HT1AreceptormRNAexpres-sion revealed a distribution pattern that is consistent with pre-vious reports describing abundant labeling throughout severalbrain regions, including the hippocampus, neocortex, amygda-la, and hypothalamus (Chalmers and Watson, 1991; OsterlundandHurd, 1998); Fig. 5A).We compared bHR and bLR 5-HT1A re-ceptormRNA levels in several forebrain regions analyzed in thec-fos studies: Acc, CPu, Cg, hippocampus, LS,M1,2, andPir. In thehippocampus, we performed a 2-way ANOVA, using subregionand bHR/bLR phenotype as independent factors. We saw amain effect of subregion (F(3,28)=45.74, p<0.0001), no effect ofphenotype, but a phenotype×subregion interaction (F(3,28)=6.74, p<0.05). Post hoc analysis showed that bLRs exhibitedhigher 5HT1a expression in the CA1 region compared to bHRs(p<0.05; Fig. 5C). In the septum, we performed a 2-wayANOVA, using subregion and bHR/bLR phenotype as indepen-dent factors. We saw a main effect of subregion (F(3,28)=19.34,p<0.0001), no effect of phenotype, but a phenotype×subregioninteraction (F(3,28)=11.18, p<0.01). Post hoc analysis showedthat bLRs exhibited higher 5HT1a expression in the LSD andLSI compared to bHRs (p<0.05; Fig. 5D). Finally, we performeda 2-way ANOVA using brain region and bHR/bLR phenotype toexamine 5-HT1A expression in several other brain areas—thestriatum, cingulate, motor cortex and piriform cortex. For this,we found a main effect of brain area (F(5,33)=15.86, p<0.0001),no effect of phenotype, but a brain area×phenotype interaction
A bLR bHR
1.20
B
-0.26
-1.30
C
LSV
MPA
LSV
D
-2.12
PVN
-2.80
E
Arc
DMH
CeA
MeA
Fig. 2 – Photomicrographs from X-ray films illustrating thedistribution of c-fos mRNA within several forebrain regionsfrom Selectively-bred High Responder (bHR) and bred LowResponder (bLR) rats. Coronal brain sections are arranged ina rostrocaudal sequence (A, most rostral through E, mostcaudal), with a representative bLR brain shown in the leftcolumn and bHR brain shown at right. Homecage intrusionevoked robust mRNA expression in several brain regions,including the lateral septum, ventral portion (LSV), medialpreoptic area (MPA), paraventricular nucleus of thehypothalamus (PVN), dorsomedial hypothalamus (DMH),arcuate nucleus of the hypothalamus (Arc), and the central(CeA) and medial amygdala (MeA).
23B R A I N R E S E A R C H 1 4 2 2 ( 2 0 1 1 ) 2 0 – 3 1

Author's personal copy
(F(4,33)=3.80, p<0.05). Post hoc analysis showed that 5-HT1AmRNA levels were increased in the Cg of the bLR rats comparedto bHRs (p<0.05; Fig. 5E).
5-HT1B receptor mRNA expression was also consistentwith previous reports, with the greatest levels apparent inAcc, CPu, CA1, and hypothalamus, and moderate expressionin the septum and several cortical regions (Neumaier et al.,1996). We evaluated 5-HT1B receptor mRNA expression in thesame brain areas analyzed for 5-HT1A receptor and c-fos.
In the hippocampus, there was a main effect of subregion(F(3,28)=165.24, p<0.0001), but no effect of phenotype, and nosubregion×phenotype interaction. In the septum, there was amain effect of subregion (F(2,21)=14.63, p<0.0001), but no effectof phenotype, and no subregion×phenotype interaction. Forthe remaining areas (cingulate, piriform cortex, motor cortexand striatum) there was a main effect of subregion (F(4,32)=45.65, p<0.0001), but no effect of phenotype, and no subregion×phenotype interaction (data not shown).
Cresyl VioletCresyl Violet bLR bHR
CeAMeA
LaA
BLA
A
-2.80
Arc
B
-2.80
-0.26
C
LSILSD
LSV
-2.56
D
CA2CA1
CA3DG
Fig. 3 – High-power photomicrographs of cresyl violet-stained sections (left column) and similar sections processed for in situhybridization to detect c-fos mRNA expression in a representative bLR and bHR rat (middle and right columns, respectively).C-fos levels were assessed within multiple amygdalar nuclei, including: lateral (LaA), basolateral (BLA), central (CeA), andmedial (MeA; A). The arcuate nucleus of the hypothalamus (Arc) was one hypothalamic regionwhere we observed greater c-fosexpression in bHR rats compared to bLRs (B). Three subdivisions of the lateral septum (LS) were analyzed: the dorsal (LSD),ventral (LSV), and intermediate (LSI; C). C-fos mRNA levels were also assessed in 4 subregions of the hippocampus: CornuAmmonis fields CA1-CA3 and the dentate gyrus (DG) (D).
Fig. 4 – Homecage intrusion elicited robust region-specific patterns of c-fos mRNA expression in multiple forebrain regions ofbHR and bLR animals. The homecage intrusion experience differentially activated subdivisions of the lateral septum (LS), withthe greatest activation within the ventral subdivision (LSV) and the least activation within the dorsal subdivision (LSD) (datacollapsed across bHR and bLR groups; A). Compared to bHRs, bLR males exhibited greater c-fos mRNA expression in theintermediate and ventral portions of the lateral septum (LSI and LSV, respectively) following the homecage intrusionexperience (B). Homecage intrusion also produced a nuclei-specific response in the amygdala, with the greatest activationoccurring in the medial nucleus of the amygdala compared to the other nuclei (data collapsed across groups; C). Surprisingly,there were no bHR/bLR differences in c-fos expression within these nuclei (D). Of the thirteen hypothalamic nuclei examined,the paraventricular nucleus of the hypothalamus (PVN) showed the greatest activation compared to the other nuclei (datacollapsed across groups; E). Abbreviations for the other hypothalamic nuclei are the same as those defined in Fig. 1C. bLRsshowed greater intrusion-evoked c-fos activation within the posterior hypothalamus (PH), lateral preoptic area (LPO), andmedial preoptic area (MPA) compared to bHRs, although bHRs showed greater activation in the arcuate nucleus (Arc). Therewere no phenotypic differences in the paraventricular nucleus (PVN) or ventromedial hypothalamus (VMH) (F). Finally, whilethe intrusion experience produced robust activation of the hippocampus, all subregions showed similarly high levels of c-fosexpression (data collapsed across groups; G). bHR animals exhibited greater overall c-fos activation across all hippocampalsubfields compared to bLRs (H). ***p<0.001; **p<0.01; *p<0.05.
24 B R A I N R E S E A R C H 1 4 2 2 ( 2 0 1 1 ) 2 0 – 3 1

Author's personal copy
3. Discussion
We previously showed that exposure to homecage intrusionelicits markedly divergent behavioral and hormonal re-sponses in bHR and bLR rats, with bHRs exhibiting exaggerat-ed aggressive behavior, testosterone and corticosteronerelease, and diminished activation of select 5-HTergic brain-stem nuclei (see companion paper, Kerman, Clinton et al.,2011). The present study revealed a distinct pattern of fore-brain c-fos mRNA expression in bHR versus bLR animals
following the homecage intrusion experience, suggestingthat their behavioral and hormonal differences are accompa-nied, or perhaps driven by, activation of specific forebrain cir-cuits. Several brain regions were activated in response to thehomecage intrusion experience, with distinct septal, amygda-lar, hypothalamic, and cortical subregions demonstrating in-creased c-fos mRNA levels. A number of these subregionsalso showed bHR/bLR differences in c-fos expression withbLR rats exhibiting greater activation of the LS and multiplehypothalamic nuclei in response to intrusion, whereas bHRsshowed greater activation in the Arc and in the hippocampus.
25B R A I N R E S E A R C H 1 4 2 2 ( 2 0 1 1 ) 2 0 – 3 1

Author's personal copy
We also found divergent 5-HT1A receptor mRNA expressionwithin some of these same areas, with bLRs having greatermRNA levels in the LS, hippocampus, and Cg. Taken togetherwith our earlier work, these findings suggest that bHRs havealtered 5-HTergic activation within select circuits during anaggressive encounter, which is consistent with reports ofhyposerotonergic states contributing to aggressive behavior.
3.1. Neuroanatomy of aggression
Our c-fos expression data indicate a distinct forebrain activationpattern in response to intruder exposure (Fig. 4). These data re-veal a gradient so that LSV, PVN, MeA, and Pir exhibited particu-larly pronounced levels of c-fos. This activation pattern likelyreflects complexity of the resident exposure experience. For ex-ample, social interaction is inherently stressful rodents (DeVries,2002), and it reliably activates the PVN, a key stress-regulatorybrain region (Sawchenko et al., 1996). Resident exposure also im-pacts the general level of arousal and anxiety, and activation ofthe LS has been implicated in these processes (Trent andMenard, 2010). Rats are also highly sensitive to olfactory cues,and intruder-elicited activation of Pir and MeA, both of whichprocess olfactory stimuli (Gottfried, 2010; Price, 2003), likely rep-resents strong olfactory stimulation by intruder presentation.
Interestingly, all of the brain regions that we examinedshowed a distinct pattern of c-fos expression within their subre-gions. For example, within the LS its ventral, intermediate, anddorsal subdivisions showed progressive decreases in c-fos. Like-wise within the hypothalamus, PVN exhibited the greatest levelof activation, while PMD, Arc, DMH, and PMV exhibited interme-diate levels, with TC, MPA, PH, AH, LH, LPO, and VMH showingthe least amount of c-fos. In the amygdala, its basolateral andlateral subdivisions exhibited lesser c-fos levels than the medialsubdivision, but greater levels than CeA. Among the other fore-brain regions, Cg and M1,2 showed equivalent activation,which was less than Pir, but greater than that of the striatum.Together these data suggest that exposure to an intruder elicitsa specific pattern of activation within the limbic system. It islikely that this pattern is specific to social interaction and maybe different in response to other emotionally-salient stimuli,such as exposure to a novel environment or to chronic stress.Future studies will be required to test this notion.
Among examined brain regions several nuclei showed signif-icantly greater c-fos levels in bLR rats, including: PH, LPO, MPA,LSI, and LSD,while Arc and the hippocampal subfields containedincreased c-fos levels in bHRs. These data are consistentwith pre-vious reports implicating these brain regions in mediating ag-gressive behaviors. For example, diminished activation withinthe LS has been reported in aggressive rodents (Haller et al.,2006; Veenema and Neumann, 2007; Veenema et al., 2007). Thisbrain region regulates diverse behaviors, including autonomic,sexual, and emotional responses, and it does so via its reciprocalprojections to other limbic regions, including the olfactory bulb,hippocampus, amygdala, hypothalamus, and Cg (Mizumoriet al., 2000; Risold andSwanson, 1997; Sheehanet al., 2000). Septallesions produce the so-called septal rage, an effect that is drivenby exaggerated fear and defensive behavior (Blanchard et al.,1979; Sheehan et al., 2000). Considering the role of the septumin modulating emotional behavior, it has been suggested that asuppressionof its activity contributes to enhanced impulsiveness
and aggressiveness (Siegel et al., 1999); thus, our current findingsof reduced LS activation in bHRs may well underlie, at least inpart, their impulsive and aggressive tendencies akin to whathas been reported in other aggressive rodent models (Haller etal., 2006; Veenema and Neumann, 2007; Veenema et al., 2007).
In addition to the LS, multiple subregions of the hypothala-mus have also been implicated in mediating aggressive behav-ior. Classic studies have delineated the hypothalamic attackarea, which spans across multiple hypothalamic nuclei, includ-ing: AH, LH, VMH, and Arc (Hrabovszky et al., 2005). Electricalstimulation of this region elicits aggressive behavior in ratsand other species, although specific types of aggressive behav-iors seem to be linked to activation of its discreet subregions(Siegel et al., 1999). In contrast, activation of other hypothalamicsubregions, such asmedial hypothalamic areas, has been linkedwith defensive behavior (Bandler et al., 1985). Because of thebHR/bLR differences in aggressive behavior and in defensivefreezing (see companion paper, Kerman, Clinton et al., 2011),the increased activation of Arc and decreased activation ofMPA in bHR animals fits well with these previous observations.
Additionally,we also observedmuch greater c-fos expressionacross all hippocampal subfields (CA1-3 and DG) following theresident–intruder test. This finding fits well previous studies,which have reported increased hippocampal activation in high-ly aggressive mice (Haller et al., 2006) and have linked aberranthippocampal activity with heightened aggression (Adamec,1991; Adamec and Stark-Adamec, 1983; Ibi et al., 2008; Mellanbyet al., 1981). Alternatively, the strong c-fos expression in the hip-pocampusmay not have been dependent on aggression per se,and insteadmay have beenmore related to other aspects of be-havioral arousal that accompany homecage intrusion, such asinvestigation of novel odor of the intruder male.
In addition to these differences in expression, bHR/bLRrats showed similar activationwithin several other forebrain re-gions, including the PVN, VMH, MeA, and CeA, which manifestenhanced activation following aggressive encounter in ratsbred for low anxiety behavior (Veenema et al., 2007). ThoughbHR animals also manifest diminished anxiety-like behaviorsand show similarly heightened aggressive responses, their dif-ferent pattern of forebrain activation suggests that distinct neu-robiological mechanisms may underlie aggression in animalsselected for high novelty-seeking (i.e. bHR) as opposed to thoseselected for low anxiety. This notion is consistent with previousstudies, which have found that a number of brain regions regu-late aggressive behavior, and since aggression itself canmanifestin numerous ways (i.e. defensive aggression, territorial aggres-sion, etc.) (Miczek et al., 2007), it seems feasible that the natureof bHR/bLR aggression differences may due to differences in theactivation of a unique set of forebrain limbic regions.
Although the resident–intruder paradigm is commonly usedto test for aggressive tendencies, it is important to note that thehome intrusion experience potentially elicits a wide range of be-havioral responses, including fear and anxiety. One interpreta-tion of our c-fos expression patterns in bHR versus bLR animalsis that the activation of certain brain areas (i.e. hippocampusand Arc) and failure to activate other regions (i.e. LS) producesan exaggerated aggressive state, a finding that is generally con-sistent with other studies in aggressive rats and mice (Halleret al., 2006; VeenemaandNeumann, 2007). However, analternateexplanation is that the observed c-fos expression differencesmay
26 B R A I N R E S E A R C H 1 4 2 2 ( 2 0 1 1 ) 2 0 – 3 1

Author's personal copy
reflect increased behavioral inhibition (e.g. fear, anxiety) in bLRsfacing intrusion. Our earlier work has demonstrated bLRs'heightened anxiety in novel situations (Stead et al., 2006), so theobserved c-fos expression differences may be related to bLRs' in-creased anxiety or fear when facing the novel social encounter,as evidenced by their increased defensive freezing (see compan-ionpaper, Kerman, Clinton et al., 2011). Future effortswill explorethese different possibilities.
3.2. Serotoninergic transmission and aggression
Abundant evidence supports the hypothesis of dysregulated5-HT neurotransmission playing an important role in regulat-ing aggressive behavior (Miczek et al., 2007). Findings of di-minished 5-HT metabolites in cerebral spinal fluid ofaggressive individuals (Brown et al., 1979; Giacalone et al.,1968; Kruesi et al., 1990) and of tryptophan-depleted diets
**
*
**
CA3 DG LSV
A) 5HT1amRNA expression B) 5HT1b mRNA expression
0.00
0.02
0.04
0.06
IOD
0.00
0.05
0.10
0.15
0.20C) 5HT1a,Hippocampus D) 5HT1a, Septum
CA1 CA2
IOD
0.00
0.05
0.10
0.15
0.20
IOD
LSD LSI MS
bHR
bLR
E) 5HT1a,Cingulate
Fig. 5 – bHR and bLR males exhibit differences in 5-HT1A receptor mRNA levels within select forebrain regions. Panels A and Bdepict photomicrographs from X-ray films illustrating the distribution of 5-HT1A receptor mRNA (A) and 5-HT1B receptormRNA (B) within several forebrain regions. 5-HT1A receptor mRNA levels are particularly enriched in the septum andhippocampus (A), while 5-HT1B receptor mRNA is highly expressed in the caudate-putamen and the CA1 subfield of thehippocampus (B). We found increased 5-HT1A receptor mRNA expression in the CA1 region of the hippocampus (C), dorsal andintermediate portions of the lateral septum (LSD and LSI, respectively; D), and the cingulate cortex (E) in bLR animals comparedto bHRs. There were no bHR/bLR differences in the expression of 5-HT1B receptor mRNA (data not shown). **p<0.001; *p<0.05.
27B R A I N R E S E A R C H 1 4 2 2 ( 2 0 1 1 ) 2 0 – 3 1

Author's personal copy
leading to exaggerated aggressive behavior (Delgado et al.,1990; Pihl et al., 1995) suggest that a general 5-HT deficiencyunderlies enhanced aggression. Moreover, microdialysis stud-ies have shed considerable light on the dynamic fluctuationsof 5-HT during the course of aggressive encounters, revealinghow levels change before, during and after an encounter, andhow these patterns markedly vary across brain regions. Forexample, 5-HT levels in the prefrontal cortex, but not Acc, de-cline as an animal is actively engaged in an aggressive display(van Erp and Miczek, 2000), although 5-HT levels decrease inthe Acc in anticipation of an impending fight (Ferrari et al.,2003). Our earlier study revealed diminished c-fos expressionwithin specific 5-HT cell groups in bHR animals exposed tohomecage intrusion, suggesting that deficient activation ofthis subset of 5-HT circuitry may contribute to their excessive-ly aggressive behavior (see companion paper, Kerman, Clin-ton et al., 2011). Our present findings extend this work,showing regional and 5-HT receptor subtype-specific bHR/bLR differences that appear to involve 5-HT1A, but not 5-HT1B, receptors (at least in the forebrain regions assessed inthis study). “Previous work has implicated both pre- andpost-synaptic 5-HT1a and 5-HT1b receptors in themodulationof aggressive behavior, and each receptor subtype may con-tribute to unique aspects of aggression and/ormediate specifictypes of aggression (de Boer and Koolhaas, 2005). The presentstudy focuses only on post-synaptic 5-TH1a and 5-HT1b recep-tors in the forebrain.We found significant bHR/bLR differenceswithin select brain regions—the hippocampus, septum, andcingulate, and these differences were restricted to 5-HT1amRNA. Other studies have demonstrated a role for both 5-HT1a and 5-HT1b in testosterone-mediated aggression(Simon et al., 1998), and 5-HT1b receptors in food restriction-induced aggressive behavior (Nakamura et al., 2008). Our find-ings suggest that bHRs' aggression is solely mediated by 5-HT1a receptors within a subset of brain regions where this re-ceptor subtype is particularly highly expressed. Importantly,the major source of 5-HT in these areas are the median anddorsal raphe (Lowry et al., 2008), brainstem regions thatshowed a very precise pattern of bHR/bLR gene expression dif-ferences and activation by intruder exposure (see companionpaper, Kerman, Clinton, et al., 2011). Future efforts will exam-ine serotonergic levels in bHR versus bLR brains both at base-line and at different points during the resident–intruder test.Additional pharmacological experiments will examine the ef-fects of 5-HT1A receptor agonists (applied either systemically,or delivered locally into specific brain regions), to determinewhether this can attenuate bHRs' aggressive tendencies.
3.3. Conclusions
Aggressive behaviors can manifest in multiple ways, and it islikely that they are mediated by distinct neurobiological pro-cesses. In humans aggression frequently coexists with hyper-activity, impulsivity, and increased propensity for drug abuse,a behavioral profile that represents externalizing disorders(Brown and Linnoila, 1990; Brown et al., 1989; Kruesi et al.,1990; Westergaard et al., 1999, 2003). In these companion pa-pers we demonstrate that bHR animals, which wereselectively-bred for increased novelty-induced locomotionand that also manifest increased impulsivity together with
an increased propensity for drug self-administration, exhibitheightened inter-male aggression. This behavioral responseis accompanied by hormonal changes that are typical of ag-gression, as well as a distinct pattern of brain activationalong with alterations in the expression of key genes that reg-ulate 5-HTergic neurotransmission. These differences in brainactivation and gene expression were restricted to specificbrainstem and forebrain subregions, suggesting that function-al alterations of specific 5-HT circuits mediate aggressive dis-plays in animals with an inborn predisposition to heightenednovelty-seeking, impulsivity, and drug self-administration.
4. Experimental procedures
4.1. Animals
Adult male bHR and bLR males (n=10 per group) were ac-quired from our in-house breeding colony where we havemaintained these lines for several generations (Stead et al.,2006). bHR and bLR rats used in these studies were from the22nd generation of our colony. In addition, a group of adultSprague–Dawley male rats (n=20) was purchased fromCharles River (Wilmington, MA) and allowed to acclimate toour housing facilities for at least 1 week prior to any behavior-al testing. All rats were housed in a dedicated animal facilitywith 12:12 light–dark cycle (lights on at 6 a.m.). Rat chow andwater were available ad libitum. All experiments were ap-proved by the University of Michigan University Committeeon Use and Care of Animals, and were conducted in accor-dance with the National Institute of Health (NIH) guidelineson laboratory animal use and care dictated by the National Re-search Council in 1996. All work was carried out in accordancewith European Commission (EC) Directive 86/609/EEC for ani-mal experiments and the Horizontal Legislation on the Protec-tion of Animals Used for Scientific Purposes (http://ec.europa.eu/environment/chemicals/lab_animals/legislation_en.htm).
4.2. Resident–intruder paradigm
bHR/bLR males were housed with a female mating partner ofthe same selectively-bred line (i.e. male bHR with femalebHR, and male bLR with female bLR) for 14 days. On the dayof testing (day 15), the female partner was removed andeach resident was presented for 10 min with a size-matchedcommercially-purchased male Sprague–Dawley rat classifiedas the intruder. Each intruder rat was only used once in theresident–intruder paradigm. This social encounter was video-taped, and subsequently scored for aggressive, defensive, andother non-antagonistic behaviors in resident and intruderrats, although those behavioral data were reported elsewhere(see companion paper, Kerman, Clinton et al., 2011).
4.3. Tissue collection and in situ hybridization
Five minutes following resident–intruder testing (15 min afterbeginning of testing) bHR/bLR resident animals were sacrificedby rapid decapitation. Brains were removed, snap frozen andstored at −80 °C. They were cryostat sectioned at 12 μm, imme-diately thaw-mounted onto Fisherbrand Superfrost Plus
28 B R A I N R E S E A R C H 1 4 2 2 ( 2 0 1 1 ) 2 0 – 3 1

Author's personal copy
Microscope Slides (Fisher Scientific, http://www.fishersci.com/),and subsequently prepared for in situhybridization. The expres-sion of c-fos, 5-HT1A receptor and 5-HT1B receptor mRNAs wasassessed at 240 μm intervals throughout several forebrain re-gions. At each anatomical level, adjacent slides were processedfor each of the three transcripts. An additional adjacent slidewas stainedwith cresyl violet andwas used to guide anatomicalidentification of the regions of interest.
Our in situ hybridization protocol has been previously de-scribed (Clinton et al., 2008). Briefly, sections were fixed in4% paraformaldehyde at room temperature for 1 hour. Theslides were then washed 5×3 min in 2× SSC (300 mMNaCl/30 mM sodium citrate, pH 7.2). Next, the slides wereplaced in a solution containing acetic anhydride (0.25%) intriethanolamine (0.1 M), pH 8.0, for 10 min at room tempera-ture, rinsed in distilled water, and dehydrated through gradedethanol washes (50%, 75%, 85%, 95%, and 100%). After air dry-ing, adjacent tissue sections were hybridized overnight at55 °C with a 35S-labeled cRNA probes (see below). Followinghybridization, coverslips were removed and the slides werewashed with 2× SSC and incubated for 1 hour in RNaseA at37 °C, followed by multiple washes in increasingly stringentSSC solutions. Slides were then rinsed in distilled H2O, dehy-drated through graded ethanol washes, air-dried, and ap-posed to Kodak XAR film (Eastman Kodak, Rochester, NY).Slides processed for c-fos, 5HT1A and 5HT1B were exposed tofilm for 7 days; this time was selected because it produced sig-nal within the linear portion of the development curve foreach probe for the brain regions of interest.
The following cRNA riboprobes were used: c-fos (alias Fos;NCBI reference sequence—X06769, pos.—585–1,368); 5-HT1A(alias Htr1a; NCBI reference sequence—J05276, pos.—333–1,241); 5-HT1B (alias Htr1b; NCBI reference sequence—X62944,pos.—210–1,070). Specificity of labeling was confirmed by theabsence of signal following hybridization with sense ribop-robes generated for the same positions of the target mRNAs(data not shown). After the 7-day exposure period, autoradio-grams were developed and digitized using a ScanMaker1000XL Pro flatbed scanner (Microtek, Carson, CA) with Laser-Soft Imaging software (AG, Kiel, Germany) at 1600 dpi. Digi-tized images were analyzed using ImageJ Analysis Softwarefor PC from NIH. Adjacent cresyl violet-stained sections weredigitized at 1600 dpi using Super CoolScan 4000 ED slide scan-ner (Nikon, http://www.nikon.com/). The cresyl violet-stainedsections provided cytoarchitectonic information to guide thedelineation of brain regions and sub-nuclei, particularly forsubregions of the amygdala and hypothalamus.
4.4. Data analysis
In an effort to standardize measurements of in situ hybridiza-tion signal and optical density, we utilized a template for eachbrain region or sub-nucleus based on the shape and size of theregion. Fig. 1 indicates the location and relative size of the tem-plates; illustrations have beenmodified from the rat brain atlasof Paxinos and Watson (1998). Using these templates, opticaldensity measurements were collected for each brain regionfrom the left and right sides of the brain (where possible) and/or from rostral–caudal sections spaced 240 μm apart. Opticaldensity values were corrected for background, multiplied by
the area sampled to produce the Integrated Optical Density(IOD)measurement, whichwas then averaged to produce a sin-gle data point for each brain region per animal. These datapoints were averaged per group and compared statistically.
We first evaluated general patterns of homecage intrusion-evoked c-fos expression (collapsing across bHR/bLR groups); forthiswe used a one-factor ANOVA and compared c-fos levels (op-tical density values) across subregions of LS, amygdala, hypo-thalamus, and hippocampus. The effect of bHR/bLR phenotypewas determined by comparing the mean level of c-fos, 5-HT1A,and 5-HT1B mRNA within the same brain regions betweenintrusion-exposed bHR and bLR males using a two-factorANOVA (bHR/bLR phenotype and sub-nucleus/subregion as in-dependent factors and IOD as dependent factorwhen analyzingbrain regions composed of multiple nuclei, such as the hypo-thalamus, septum, amygdala and hippocampus). Post-hoccomparisons were performed using Fisher's Exact Test. Datawere analyzed using SPSS 17.0 for Windows (SPSS Institute,http://www.spss.com/) and are presented as mean±SEM. Forall tests α=0.05.
Acknowledgments
This study was supported by NIMH 1K99MH085859 (SMC), NIMH5K99MH081927 (IAK), NARSAD Young Investigator award (IAK),Office of Naval Research N00014-09-1-0598 (HA and SJW), andNIDA PPG 5P01DA021633-02 (HA and SJW). The authors wouldlike to thankMs. Sharon Burke andMs. SueMiller for their excel-lent technical assistance and Ms. Rebecca Simmons for helpfuldiscussions of an earlier version of this manuscript. None ofthe funding sources had roles in study design; data collection,analysis and interpretation,manuscript preparation, or the deci-sion to submit thework for publication. There are no biomedicalfinancial interests or conflicts of interest for any of the authors.
R E F E R E N C E S
Adamec, R.E., 1991. Partial kindling of the ventral hippocampus:identification of changes in limbic physiology whichaccompany changes in feline aggression and defense. Physiol.Behav. 49, 443–453.
Adamec, R.E., Stark-Adamec, C., 1983. Partial kindling andemotional bias in the cat: lasting aftereffects of partial kindlingof the ventral hippocampus. I. Behavioral changes. Behav.Neural Biol. 38, 205–222.
Bandler, R., Depaulis, A., Vergnes, M., 1985. Identification ofmidbrain neurones mediating defensive behaviour in the ratby microinjections of excitatory amino acids. Behav. Brain Res.15, 107–119.
Blanchard, D.C., Blanchard, R.J., Lee, E.M., Nakamura, S., 1979.Defensive behaviors in rats following septal andseptal–amygdala lesions. J. Comp. Physiol. Psychol. 93, 378–390.
Brown, G.L., Linnoila, M.I., 1990. CSF serotonin metabolite(5-HIAA) studies in depression, impulsivity, and violence.J. Clin. Psychiatry 51, 31–41 Suppl, discussion 42–3.
Brown, G.L., Goodwin, F.K., Ballenger, J.C., Goyer, P.F., Major, L.F.,1979. Aggression in humans correlates with cerebrospinal fluidamine metabolites. Psychiatry Res. 1, 131–139.
Brown, C.S., Kent, T.A., Bryant, S.G., Gevedon, R.M., Campbell, J.L.,Felthous, A.R., Barratt, E.S., Rose, R.M., 1989. Blood platelet
29B R A I N R E S E A R C H 1 4 2 2 ( 2 0 1 1 ) 2 0 – 3 1

Author's personal copy
uptake of serotonin in episodic aggression. Psychiatry Res. 27,5–12.
Chalmers, D.T., Watson, S.J., 1991. Comparative anatomicaldistribution of 5-HT1A receptor mRNA and 5-HT1A binding inrat brain—a combined in situ hybridisation/in vitro receptorautoradiographic study. Brain Res. 561, 51–60.
Clinton, S., Miller, S., Watson, S.J., Akil, H., 2008. Prenatal stressdoes not alter innate novelty-seeking behavioral traits, butdifferentially affects individual differences in neuroendocrinestress responsivity. Psychoneuroendocrinology 33, 162–177.
Davidson, R.J., Putnam, K.M., Larson, C.L., 2000. Dysfunction in theneural circuitry of emotion regulation—a possible prelude toviolence. Science 289, 591–594.
Davis, B.A., Clinton, S.M., Akil, H., Becker, J.B., 2008. The effects ofnovelty-seeking phenotypes and sex differences onacquisition of cocaine self-administration in selectivelybred High-Responder and Low-Responder rats. Pharmacol.Biochem. Behav. 90, 331–338.
de Boer, S.F., Koolhaas, J.M., 2005. 5-HT1A and 5-HT1B receptoragonists and aggression: a pharmacological challenge of theserotonin deficiency hypothesis. Eur. J. Pharmacol. 526,125–139.
Delgado, P.L., Charney, D.S., Price, L.H., Aghajanian, G.K., Landis,H., Heninger, G.R., 1990. Serotonin function and themechanism of antidepressant action. Reversal ofantidepressant-induced remission by rapid depletion ofplasma tryptophan. Arch. Gen. Psychiatry 47, 411–418.
DeVries, A.C., 2002. Interaction among social environment, thehypothalamic–pituitary–adrenal axis, and behavior. Horm.Behav. 41, 405–413.
Ferrari, P.F., van Erp, A.M., Tornatzky, W., Miczek, K.A., 2003.Accumbal dopamine and serotonin in anticipation of the nextaggressive episode in rats. Eur. J. Neurosci. 17, 371–378.
Flagel, S.B., Robinson, T.E., Clark, J.J., Clinton, S.M., Watson, S.J.,Seeman, P., Phillips, P.E., Akil, H., 2010. An animal model ofgenetic vulnerability to behavioral disinhibition andresponsiveness to reward-related cues: implications foraddiction. Neuropsychopharmacology 35, 388–400.
Giacalone, E., Tansella, M., Valzelli, L., Garattini, S., 1968. Brainserotonin metabolism in isolated aggressive mice. Biochem.Pharmacol. 17, 1315–1327.
Gottfried, J.A., 2010. Central mechanisms of odour objectperception. Nat. Rev. Neurosci. 11, 628–641.
Haller, J., Toth, M., Halasz, J., De Boer, S.F., 2006. Patterns of violentaggression-induced brain c-fos expression in male miceselected for aggressiveness. Physiol. Behav. 88, 173–182.
Higley, J.D., Mehlman, P.T., Taub, D.M., Higley, S.B., Suomi, S.J.,Vickers, J.H., Linnoila, M., 1992. Cerebrospinal fluidmonoamine and adrenal correlates of aggression in free-ranging rhesus monkeys. Arch. Gen. Psychiatry 49, 436–441.
Hrabovszky, E., Halasz, J., Meelis, W., Kruk, M.R., Liposits, Z., Haller,J., 2005. Neurochemical characterization of hypothalamicneurons involved in attack behavior: glutamatergic dominanceand co-expression of thyrotropin-releasing hormone in a subsetof glutamatergic neurons. Neuroscience 133, 657–666.
Ibi, D., Takuma, K., Koike, H., Mizoguchi, H., Tsuritani, K., Kuwahara,Y., Kamei, H., Nagai, T., Yoneda, Y., Nabeshima, T., Yamada, K.,2008. Social isolation rearing-induced impairment of thehippocampal neurogenesis is associated with deficits in spatialmemory and emotion-related behaviors in juvenile mice.J. Neurochem. 105, 921–932.
Kruesi, M.J., Rapoport, J.L., Hamburger, S., Hibbs, E., Potter, W.Z.,Lenane, M., Brown, G.L., 1990. Cerebrospinal fluid monoaminemetabolites, aggression, and impulsivity in disruptivebehavior disorders of children and adolescents. Arch. Gen.Psychiatry 47, 419–426.
Lowry, C.A., Evans, A.K., Gasser, P.J., Hale,M.W., Staub, D.R., Shekhar,A., 2008. Topographic organization and chemoarchitecture of thedorsal raphe nucleus and themedian raphe nucleus. In: Monti, J.
M., Pandi-Perumal, S.R., Jacobs, B.L., Nutt, D.J. (Eds.), Serotoninand Sleep: Molecular, Functional and Clinical Aspects.Birkhäuser Verlag, Switzerland. Vol.
Mellanby, J., Strawbridge, P., Collingridge, G.I., George, G., Rands,G., Stroud, C., Thompson, P., 1981. Behavioural correlates of anexperimental hippocampal epileptiform syndrome in rats.J. Neurol. Neurosurg. Psychiatry 44, 1084–1093.
Miczek, K.A., Faccidomo, S.P., Fish, E.W., DeBold, J.F., 2007.Neurochemistry and Molecular Neurobiology of AggressiveBehavior. In: Lajtha, A. (Ed.), Handbook of Neurochemistry andMolecular Neurobiology. Vol. Behavioral Neurochemistry,Neuroendocrinology and Molecular Neurobiology. Springer, US.
Mizumori, S.J., Cooper, B.G., Leutgeb, S., Pratt, W.E., 2000. A neuralsystems analysis of adaptive navigation. Mol. Neurobiol. 21,57–82.
Nakamura, K., Kikusui, T., Takeuchi, Y., Mori, Y., 2008. Changes insocial instigation- and food restriction-induced aggressivebehaviors and hippocampal 5HT1B mRNA receptor expressioninmalemice from earlyweaning. Behav. Brain Res. 187, 442–448.
Neumaier, J.F., Szot, P., Peskind, E.R., Dorsa, D.M., Hamblin, M.W.,1996. Serotonergic lesioning differentially affects presynapticand postsynaptic 5-HT1B receptor mRNA levels in rat brain.Brain Res. 722, 50–58.
Osterlund, M.K., Hurd, Y.L., 1998. Acute 17 beta-estradioltreatment down-regulates serotonin 5HT1A receptor mRNAexpression in the limbic system of female rats. Brain Res. Mol.Brain Res. 55, 169–172.
Paxinos, G., Watson, C., 1998. The Rat Brain in StereotaxicCoordinates. Academic Press, San Diego. Vol.
Pihl, R.O., Young, S.N., Harden, P., Plotnick, S., Chamberlain, B.,Ervin, F.R., 1995. Acute effect of altered tryptophan levels andalcohol on aggression in normal human males.Psychopharmacology (Berl) 119, 353–360.
Price, J.L., 2003. Comparative aspects of amygdala connectivity.Ann. N. Y. Acad. Sci. 985, 50–58.
Risold, P.Y., Swanson, L.W., 1997. Connections of the rat lateralseptal complex. Brain Res. Brain Res. Rev. 24, 115–195.
Sawchenko, P.E., Brown, E.R., Chan, R.K., Ericsson, A., Li, H.Y.,Roland, B.L., Kovacs, K.J., 1996. The paraventricular nucleus ofthe hypothalamus and the functional neuroanatomy ofvisceromotor responses to stress. Prog. Brain Res. 107,201–222.
Sheehan, T.P., Cirrito, J., Numan, M.J., Numan, M., 2000. Usingc-Fos immunocytochemistry to identify forebrain regions thatmay inhibit maternal behavior in rats. Behav. Neurosci. 114,337–352.
Siegel, A., Roeling, T.A., Gregg, T.R., Kruk, M.R., 1999.Neuropharmacology of brain-stimulation-evoked aggression.Neurosci. Biobehav. Rev. 23, 359–389.
Simon, N.G., Cologer-Clifford, A., Lu, S.F., McKenna, S.E., Hu, S.,1998. Testosterone and its metabolites modulate 5HT1Aand 5HT1B agonist effects on intermale aggression.Neurosci. Biobehav. Rev. 23, 325–336.
Stead, J.D., Clinton, S., Neal, C., Schneider, J., Jama, A., Miller, S.,Vazquez, D.M., Watson, S.J., Akil, H., 2006. Selective breedingfor divergence in novelty-seeking traits: heritability andenrichment in spontaneous anxiety-related behaviors. Behav.Genet. 36, 697–712.
Trent, N.L., Menard, J.L., 2010. The ventral hippocampus and thelateral septum work in tandem to regulate rats' open-armexploration in the elevated plus-maze. Physiol. Behav. 101,141–152.
van Erp, A.M., Miczek, K.A., 2000. Aggressive behavior, increasedaccumbal dopamine, and decreased cortical serotonin in rats.J. Neurosci. 20, 9320–9325.
Veenema, A.H., Neumann, I.D., 2007. Neurobiologicalmechanisms of aggression and stress coping: a comparativestudy in mouse and rat selection lines. Brain Behav. Evol. 70,274–285.
30 B R A I N R E S E A R C H 1 4 2 2 ( 2 0 1 1 ) 2 0 – 3 1

Author's personal copy
Veenema, A.H., Torner, L., Blume, A., Beiderbeck, D.I., Neumann, I.D.,2007. Low inborn anxiety correlates with high intermaleaggression: link to ACTH response and neuronal activation of thehypothalamic paraventricular nucleus. Horm. Behav. 51, 11–19.
Westergaard, G.C., Suomi, S.J., Higley, J.D., Mehlman, P.T., 1999.CSF 5-HIAA and aggression in female macaque monkeys:
species and interindividual differences. Psychopharmacology(Berl) 146, 440–446.
Westergaard, G.C., Suomi, S.J., Chavanne, T.J., Houser, L., Hurley,A., Cleveland, A., Snoy, P.J., Higley, J.D., 2003. Physiologicalcorrelates of aggression and impulsivity in free-ranging femaleprimates. Neuropsychopharmacology 28, 1045–1055.
31B R A I N R E S E A R C H 1 4 2 2 ( 2 0 1 1 ) 2 0 – 3 1




















