
Evidence for position-insensitive orthographic priming in the ...
Upload
independentCategory
view
4download
0
International Immunology, Vol. 11, No. 5, pp. 689–698 © 1999 The Japanese Society for Immunology
Paradoxical priming effects of IL-10 oncytokine production
Minou Adib-Conquy, Anne-France Petit, Christelle Marie, Catherine Fitting andJean-Marc Cavaillon
Unite d’Immuno-Allergie, Institut Pasteur, 28 rue Dr Roux, 75015 Paris, France
Keywords: cytokine production, IL-10
Abstract
IL-10 is a well-known immunosuppressive and/or anti-inflammatory cytokine. However, we reportin vitro experimental studies in which IL-10 primed leukocytes and led to an enhanced productionof tumor necrosis factor (TNF) upon further stimulation by lipopolysaccharide (LPS). Monocytesand peripheral blood mononuclear cells (PBMC) prepared from whole blood maintained for 20 h at37°C in the presence of recombinant human IL-10 had an enhanced capacity to produce TNF inresponse to LPS. In addition to TNF, LPS-induced IL-6 and spontaneous IL-1ra production werealso enhanced. When isolated PBMC were first cultured for 20 h in the presence of IL-10 on Teflonto prevent adherence, washed to remove IL-10 and then further cultured in plastic dishes for anadditional 20 h in the presence of LPS or IL-1 β, an enhanced release of TNF was observed. Thiswas not the case when PBMC were pre-cultured in plastic multidishes in the presence of IL-10. TNFmRNA expression induced by LPS was decreased when the pre-treatment of PBMC with IL-10 wasperformed on plastic, whereas this was not the case when cells were pre-cultured with IL-10 onTeflon. Furthermore, NF κB translocation following LPS activation was higher after IL-10 pre-treatment on Teflon than on plastic. Interestingly, an enhanced frequency of CD16 and CD68 F cellsamong the CD14 F cells was observed in the presence of IL-10, independently of the pre-cultureconditions of the PBMC. Altogether, these results indicate that the IL-10-induced up-regulation ofcytokine production depends on the prevention of monocyte adherence by red cells in the wholeblood assays or by cultures of PBMC on Teflon. In contrast, the adherence parameter has no effecton the IL-10-induced modulation of some monocyte surface markers.
Introduction
IL-10 has been identified as a cytokine synthesis inhibitoryfactor capable of suppressing the production of Th1cytokines including IL-2 and IFN-γ (1–4). In addition, IL-10is capable of inhibiting the production of most cytokinesproduced by monocytes/macrophages (5,6) and neutrophils(PMN) (7–9). Thus, IL-10 is known to be an immunosuppres-sive as well as a potent anti-inflammatory mediator. Whilemany in vitro experimental models have established the anti-inflammatory properties of IL-10, several in vivo situationssuggested that in some circumstances, IL-10 could act asa pro-inflammatory and/or an immunostimulatory cytokine.This was observed in graft rejection (10-12), ocular inflam-mation (13), auto-immune diseases (14,15) or anti-tumorimmunity (16). Indeed, a very clear pro-inflammatory activitywas shown following the implant of adenocarcinoma-transfected cells expressing IL-10 (17). Furthermore, during
Correspondence to: J.-M. Cavaillon
Transmitting editor: L. du Pasquier Received 9 September 1998, accepted 19 January 1999
phase I trials of IL-10 in healthy humans, some unexpectedresults emerged including fever and leukocytosis (18,19).
Some reports have shown that IL-10 could also favorcertain aspects of the inflammatory response. Thus, IL-10induces E-selectin expression on small and large bloodvessel endothelial cells (20). IL-10 may behave differentlydepending on the nature of the target cells. For example,a consistent or even enhanced production of IL-8 has beenreported by dendritic cells (21) and endothelial cells (22),while suppression of IL-8 production has been describedin monocytes and PMN (5–9). Similarly, IL-10 inhibits IL-6 release by lipopolysaccharide (LPS)-stimulated humanperipheral blood mononuclear cells (PBMC), whereas itdoes not interfere with cytokine production by activatedhuman endothelial cells (23). Furthermore, IL-10 can repressthe production of NO by macrophages or keratinocytes
by guest on August 26, 2015
http://intimm
.oxfordjournals.org/D
ownloaded from

690 Priming effect of IL-10
(24,25), does not modify its release by mesangial cells(26), and even enhances the production of NO by bonemarrow-derived macrophages and osteoclasts (27,28). Theinhibitory capacity of IL-10 may also depend on the natureof the triggering agent. For example, IL-10 represses theLPS-induced IL-8 production by neutrophils while it doesnot when PMN are activated by tumor necrosis factor (TNF)-α (9). Furthermore, IL-10 could act differently depending onthe state of maturation and/or activation of the target cellsor depending on the sequence of signaling. So far, thishas been essentially demonstrated for IL-4 (29–31), but astudy on blood and sputum neutrophils also showed adifferential responsiveness of these cells to IL-10 (32). Inthe present study we demonstrate that monocytes andPBMC prepared from blood samples maintained overnightin the presence of IL-10 had an enhanced capacity toproduce TNF-α upon stimulation. This finding could bemimicked by pre-incubating PBMC with IL-10 as long asadherence was prevented by pre-cultures on Teflon. Tofurther analyze the underlying mechanisms of the observedphenomenon, TNF mRNA expression and NFκB translocationhave been investigated.
Methods
Whole blood pre-cultureWhole blood samples from healthy donors (Etablissement detransfusion sanguine de l’Assistance Publique, Paris, France),collected on CPD (citrate/phosphate/dextrose), were main-tained in culture flasks (75 cm2; Costar, Cambridge, MA) inthe absence or presence of recombinant human IL-10(10 ng/ml) (DNAX, Palo Alto, CA/Genzyme, Cambridge, MA)for 20 h at 37°C in a 5% CO2 incubator. At the end of thispre-culture period, whole blood samples were diluted 1:3 inRPMI 1640 medium (Glutamax; Gibco Life Technologies,Paisley, UK) and layered on Ficoll-Hypaque (MSL; Eurobio,Les Ulis, France). The ratio was 2 volumes of blood to1 volume of MSL. After centrifugation for 30 min at 15°C and600 g, PBMC were washed twice in RPMI 1640 medium.PBMC (63106 PBMC/ml or adjusted to 106 non-specificesterase1 cells/ml) were suspended in RPMI 1640 mediumsupplemented with antibiotics (100 IU/ml penicillin;100 µg/ml streptomycin), 0.2% heat-inactivated normal humanserum (a pool of sera from healthy volunteers) and1 µg/ml indomethacin, and incubated in 24-well multidishplates (Costar) (0.5 ml/well). In the case of adherent cells,non-adherent cells were removed after a 2 h incubation at37°C and fresh complete medium was added. PBMC andadherent cells were incubated for 20 h in the presence orabsence of various stimuli. Stimuli used were Escherichia coli0111:B4 LPS (2 µg/ml; Sigma, St Louis, MO), Neisseriameningitidis LPS (2 µg/ml; a kind gift of Dr M. Caroff),recombinant human IL-1β (20 ng/ml; Rhone Poulenc, France)and Staphylococcus aureus Cowan I (200 µg/ml; Pansorbin,Calbiochem, La Jolla, CA). At the end of the culture period,supernatants were harvested, centrifuged for 10 min at 15°Cand 300 g, and kept at –30°C until cytokine assessments.
PBMC pre-culturePBMC were prepared from freshly sampled blood of healthydonors, diluted 1:2 in RPMI 1640 medium and centrifuged
over Ficoll for 20 min at 15°C and 600 g. After washings,PBMC were adjusted to 63106 cells/ml in RPMI 1640 mediumsupplemented with antibiotics and 5% (unless otherwisementioned) heat-inactivated normal human serum or autolog-ous plasma. PBMC were pre-cultured in 24-well multidishplates (0.5 ml/well) in the absence or presence of recombinanthuman IL-10 (10 ng/ml) for 20 h at 37°C in a 5% CO2incubator. At the end of the pre-culture, PBMC were collected,each well was washed twice with 1 ml RPMI 1640 mediumand 1 ml of fresh medium was added to plastic dishes topreserve adherent cell viability. The harvested non-adherentcells were washed once and resuspended in 0.5 ml RPMI1640 medium supplemented with antibiotics, indomethacinand 0.2% normal human serum, and plated in the same wellsas during the pre-culture period after removal of the RPMI1640 medium. PBMC were further cultured for a 20 h periodat 37°C and 5% CO2 in the presence or absence of variouscell activators.
PBMC were also pre-cultured in Teflon containers(2 ml/container) (PolyLabo, Strasbourg, France). At the endof the pre-culture period, PBMC were harvested and Tefloncontainers washed. PBMC were resuspended, plated andcultured in 24-well multidishes as described above. At theend of the second culture period, the supernatants wereharvested, centrifuged for 10 min at 15°C and 300 g, andkept at –30°C until cytokine assays were performed.
Cytokine measurements
Cytokines were measured using specific in-house or commer-cial ELISA: TNF-α (33), IL-6 (34), IL-1ra and IL-10 (R & DSystems, Abingdon, UK).
Apoptosis assay
PBMC apoptosis was assessed according to the percentageof cells containing hypodiploid DNA by using the techniquedescribed by Nicoletti et al. (35). After 20 h of pre-culture ofblood, PBMC were prepared and cells were centrifuged at200 g for 10 min and washed in PBS. The cell pellets weregently resuspended in hypotonic fluorochrome solution(50 µg/ml propidium iodide, 0.1% sodium citrate, 0.1% TritonX-100) and stored at 4°C in the dark overnight before theflow-cytometric analysis using a FACScan flow cytometer(Becton Dickinson Immunocytometry System, San Jose, CA).The red fluorescence of propidium iodide in individual nuclei,the forward scatter and the side scatter were simultaneouslymeasured. Cell debris was excluded from acquisition byraising the forward scatter threshold. Apoptotic nuclei wereeasily distinguishable from residual debris by the high sidescatter value due to the condensation of nuclear chromatin.A total of 10,000 events was collected and analyzed usingthe software Lysys II program. Apoptotic nuclei were distin-guished by their hypodiploid DNA content from the diploidDNA content of normal nuclei.
Electrophoretic mobility shift assay (EMSA)
Nuclear protein extractions were carried out as described(36). PBMC were washed once with PBS, adherent cells wereharvested with a cell scraper, added to non-adherent cellsand suspended in buffer A (10 mM HEPES, pH 7.9, 1.5 mMMgCl2, 10 mM KCl, 0.5 mM DTT and 0.1% NP-40) supple-
by guest on August 26, 2015
http://intimm
.oxfordjournals.org/D
ownloaded from

Priming effect of IL-10 691
mented with protease inhibitors. The protease inhibitorsincluded: 0.5 mM PMSF, 25 µg/ml aprotinin, 10 µg/ml chymo-statin, 2 µg/ml antipain, 8 µg/ml pepstatin, 10 µg/ml leupeptin,0.1 mg/ml α1-antitrypsin and 0.5 mM 3,4-dichloroisocoumarin(all from Sigma). Cells were incubated for 10 min at 4°C andthen centrifuged for 2 min at 10,000 r.p.m. The pellet wassuspended in buffer C and incubated for 20 min at 4°C.Buffer C contained 20 mM HEPES, pH 7.9, 420 mM NaCl,1.5 mM MgCl2, 0.2 mM EDTA, 25% glycerol, 0.5 mM DTTand protease inhibitors. Cells were then centrifuged for 10 minat 14,000 r.p.m., the supernatant corresponding to the nuclearextract was harvested and kept at –80°C. A double-strandedoligonucleotide containing the NFκB motif (promega) wasend-labeled with T4 kinase in the presence of [γ-32P]ATP.Extracts (4 µg) were incubated in the binding buffer for 15 minat room temperature (4% Ficoll, 20 mM HEPES, pH 7, 35 mMNaCl, 60 mM KCl, 0.01% NP-40, 2 mM DTT, 0.1 mg/mlBSA and 1.5 µg/µl salmon sperm DNA). After 15 min, theradiolabeled nucleotide was added (100,000 c.p.m.) and themixture was again incubated for 15 min at room temperature.EMSA was performed in a 5% acrylamide gel in 0.53TBE,and gels were dried and subjected to autoradiography. TheNFκB complexes were quantified using a PhosphorImagerand the ImageQuant software (Molecular Dynamics). Thecomposition of the NFκB complexes was assessed bysupershift assays using specific antibodies (anti-p50 and anti-p65; Santa Cruz Biotechnology, Santa Cruz, CA). The upperEMSA band was identified as a p50–p65 complex, while thesecond one was identified to be a p50 homodimer (datanot shown).
RNA dot-blot
Total RNA from PBMC was extracted using the RNA PLUSreagent (Bioprobe Systems, Montreuil sous bois, France).Several dilutions of each sample were put onto Gene ScreenPlus membranes (Dupont, Boston, MA) using a dot-blotapparatus (Minifold I; Schleicher & Schuell, Dassel, Germany)and hybridized with 32P-labeled TNF-α and β-actin probeskindly provided by Dr N. Haeffner-Cavaillon (INSERM U430,Hopital Broussais). The human TNF-α and the mouse β-actin cDNA probes were 1.18 and 0.97 kb PstI fragmentsrespectively. The transfer and the hybridization conditionswere those recommended by the manufacturer for GeneScreen membranes. After hybridization, the membranes werewashed and the radioactivity was quantified using a Phos-phorImager and the ImageQuant software.
FACS analysis
Cells were cultured in the absence or the presence of IL-10(10 ng/ml) for 20 h, using the whole blood assay, as well asisolated PBMC on plastic or Teflon as previously described.At the end of the pre-incubation, PBMC were isolated fromwhole blood using Ficoll Hypaque, PBMC cultured in theTeflon containers were recovered by centrifugation. For theplastic condition, adherent cells were recovered after a 5 minincubation in PBS/BSA 1%/EDTA 1.3 mM, pooled with corres-ponding non-adherent cells and centrifuged. PBMC werewashed with PBS/BSA 1%, counted and 13106 cells wereused per sample. Double staining was performed using ananti-CD14 mAb (MY4-RD1; Coulter, Miami, FL) coupled to
Fig. 1. TNF-α production by human adherent monocytes isolatedfrom whole blood previously maintained for 20 h in the presence orabsence of IL-10 (10 ng/ml) and further cultured for an additional20 h in the presence of N. meningitidis LPS (2 µg/ml). Each linedefines a single individual experiment (n 5 14). The mean 6 SEM ofTNF-α produced by cells derived from blood in the absence or inthe presence of recombinant human IL-10 was 3214 6 674 and10,517 6 1724 pg/ml respectively (P , 0.001).
phycoerythrin (PE) and an anti-CD16 mAb (3G8; Immunotech,Marseille, France) or an anti-CD68 mAb (KP1; Dako, Glostrup,Denmark) coupled to FITC. Mouse IgG1–FITC (MOPC-21;Sigma) and IgG2b–PE (MOPC-141; Sigma) were used asisotypic controls. Cells were incubated on ice with mAb inPBS/BSA for 30 min, washed with PBS/BSA and data werecollected on 10,000 cells in a FACScan analyzer (BectonDickinson). The monocyte population was analyzed aftergating on side and forward scatters. Results are expressedas percent of positive cells and as mean fluorescence ratio(MFR 5 mean fluorescence of sample/mean fluorescenceof control).
Statistical analysis
Statistical analysis has been performed using the non-para-metric Wilcoxon signed-rank test.
Results
Reactivity of adherent mononuclear cells isolated from IL-10pretreated blood samples
In a first set of experiments, LPS-induced TNF-α productionby human monocytes was investigated with adherent cellsprepared from blood samples maintained in culture for 20 heither in the absence or in the presence of recombinanthuman IL-10. The results depicted in Fig. 1 indicate thatincreased amounts of TNF were detected in the supernatantsof monocytes prepared from IL-10 pre-treated blood samplesas compared to adherent cells derived from blood maintainedfor 20 h in the absence of IL-10. Fourteen different donorswere studied and the observation was highly reproducible(p , 0.001). The minimum timing of IL-10 pre-incubation inthe whole blood samples leading to the enhanced TNF
by guest on August 26, 2015
http://intimm
.oxfordjournals.org/D
ownloaded from

692 Priming effect of IL-10
production was 14 h (data not shown). While this long periodsuggests that protein synthesis might be involved in theobserved phenomenon, preliminary experiments to demon-strate it with cycloheximide failed due to a long-lasting effectof the drug which later on interfered with the cytokine synthesisinduced upon activation (data not shown). A wide range ofTNF levels was observed consistent with previous findingsdue to the wide genetic diversity seen with human cells (37).The mean TNF increase of 3.3-fold was not due to a differencein the number of apoptotic cells since a similar total numberof PBMC and a similar number of apoptotic PBMC (11 6 4%)were recovered from blood samples maintained for 20 h at37°C in the absence or presence of IL-10. However, wenoticed an enhanced proportion (42 6 13%) of adheringmonocytes recovered after incubation onto plastic of PBMCprepared from the blood samples supplemented with IL-10.
Reactivity of PBMC isolated from IL-10 pretreated bloodsamplesAlthough the observed enhancement of adherent cells wasfar below the enhancement of the TNF production, it couldreflect a selection of a highly reactive subpopulation. So, wedecided to pursue the analysis without selecting the adherentcells. As shown in Fig. 2A, similar enhancement of TNFproduction was observed when whole PBMC were culturedand activated for an additional 20 h after being preparedfrom blood in which increasing amounts of IL-10 had beenadded. The enhancement was independent of the bacterialorigin of the LPS as well as when IL-1 was employed as atriggering signal. The presence of low (0.2%) or high (2%)concentrations of normal human serum or the addition ofindomethacin influenced the amounts of produced TNFα butdid not modify the IL-10 induced effect (Fig. 2B). Fourteennew different experiments have been performed with PBMCand a fixed amount of recombinant human IL-10 (10 ng/ml).The mean enhancement of Escherichia coli LPS-induced TNFproduction by PBMC derived from blood maintained for 20 hin the presence of IL-10 was 3.4 6 0.9-fold (P 5 0.016).Although high LPS concentrations (1–2 µg/ml) have beenused throughout the study, the amplificatory activity of IL-10was also observed with lower concentrations of LPS (1–100ng/ml) (Table 1). The mean enhancements were 2.1 6 0.4-,2.6 6 0.5- and 3.4 6 1.1-fold for 1, 10 and 100 ng/mlrespectively. A weaker amplificatory activity was found whenS. aureus Cowan was used as TNF-α inducer (1.3 6 0.2-fold).
To assess whether the enhancing effect of the IL-10 pre-treatment was specific for TNF, we also investigated its roleon IL-6, IL-1ra and IL-10 production. As shown in Table 2,the pre-culture of whole blood in the presence of IL-10significantly increased the E. coli LPS-induced IL-6 productionby PBMC (P 5 0.03) and to a lesser extent that induced byN. meningitidis LPS (P 5 0.07). Such IL-10 pre-treatmentgreatly increased the spontaneous IL-1ra production as wellas the LPS-induced IL-1ra. The S. aureus Cowan-induced IL-6 and IL-1ra productions were not significantly enhanced. IL-10 production was not affected.
Reactivity of PBMC following an IL-10 pretreatment in TefloncontainersWe and others have demonstrated that pre-incubation ofmonocytes with IL-10 led to a reduction of TNF production
Fig. 2. (A) TNF-α production by human PBMC cultured for 20 h inthe absence or in the presence of either E. coli (E.c.) LPS (2 µg/ml),N. meningitidis (N.m.) LPS (2 µg/ml) or IL-1β (20 ng/ml). PBMC wereisolated from blood samples maintained at 37°C for 20 h in thepresence of increasing amounts of recombinant human IL-10. Thedose–response curve is representative of two experiments. (B) TNF-α production by N.m. LPS (2 µg/ml)-activated PBMC in the presenceof 0.2% or 2% normal human serum (NHS) and in the presence orabsence of indomethacin. PBMC were isolated from blood samplesmaintained at 37°C for 20 h in the presence of increasing amountsof IL-10.
upon LPS stimulation (33,38). The converse obtained withpre-incubation of whole blood with IL-10 suggested thateither compounds present in whole blood or experimentalparameters may later on affect the responsiveness of the cells.Autologous plasma, presence of platelets and adherence ofthe cells were considered. Further experiments were per-formed with freshly isolated PBMC which were first maintainedin the presence or absence of IL-10 in the presence of variousamounts of autologous plasma. To prevent adherence, PBMCwere pre-cultured for 20 h in Teflon containers before beingtransferred in the classical plastic 24-well multidishes andfurther cultured for 20 h in the presence of various stimuli. Asshown in Fig. 3, the presence of IL-10 in such pre-culturesresulted in cells which were capable of producing higher
by guest on August 26, 2015
http://intimm
.oxfordjournals.org/D
ownloaded from

Priming effect of IL-10 693
Fig. 3. TNF-α production by human PBMC pre-cultured for 20 h in Teflon containers in the absence (light shaded) or in the presence (heavyshaded) of recombinant human IL-10 (10 ng/ml) and in the presence of 5% (left panel) or 50% (right panel) autologous plasma. PBMC werefurther cultured in plastic 24-well multidish plates in the presence or absence of either E. coli (E.c.) LPS (2 µg/ml), N. meningitidis (N.m.) LPS(2 µg/ml), S. aureus Cowan (200 µg/ml) or IL-1β (20 ng/ml). The experiment is representative of three.
Table 1. TNF-α production (pg/ml), upon activation by increasing amounts of E. coli LPS, by PBMC isolated from bloodsamples maintained for 20 h at 37°C in the presence or absence of IL-10 (10 ng/ml)
E. coli LPS 1 ng/ml 10 ng/ml 100 ng/ml
IL-10 No Yes No Yes No Yes
Experiment 1 ND ND 856 2993 (33.5) 1284 7085 (35.3)Experiment 2 2966 7360 (32.5) 3135 8352 (32.7) 2884 7495 (32.6)Experiment 3 800 1358 (31.7) 812 1396 (31.7) 946 2000 (32.1)
Table 2. IL-6, IL-10 and IL-1ra production (pg/ml) by PBMC derived from blood maintained for 20 h in the absence or presenceof IL-10 (10 ng/ml)a
Pre-culture 0–20 h Activators 20–40 h
None E. coli LPS N. meningitidis LPS S. aureus Cowan
IL-6 whole blood 572 6 115 44,687 6 6497 59,131 6 7143 64,707 6 5904whole blood 1 IL-10 660 6 190 79,455 6 13,553 84,158 6 15,379 88,968 6 13,186P NS 0.03 NS NS
IL-1ra whole blood 13,794 6 4772 28,693 6 8604 30,172 6 9096 42,285 6 10,691whole blood 1 IL-10 30,028 6 5796 43,477 6 10,066 41,786 6 9390 50,679 6 9542P 0.01 0.01 0.05 NS
IL-10 whole blood 31 6 10 1059 6 426 1222 6 469 2283 6 880whole blood 1 IL-10 64 6 26 1014 6 266 1169 6 251 2455 6 723P NS NS NS NS
aThe results are the mean 6 SEM of six different experiments.
by guest on August 26, 2015
http://intimm
.oxfordjournals.org/D
ownloaded from
694 Priming effect of IL-10
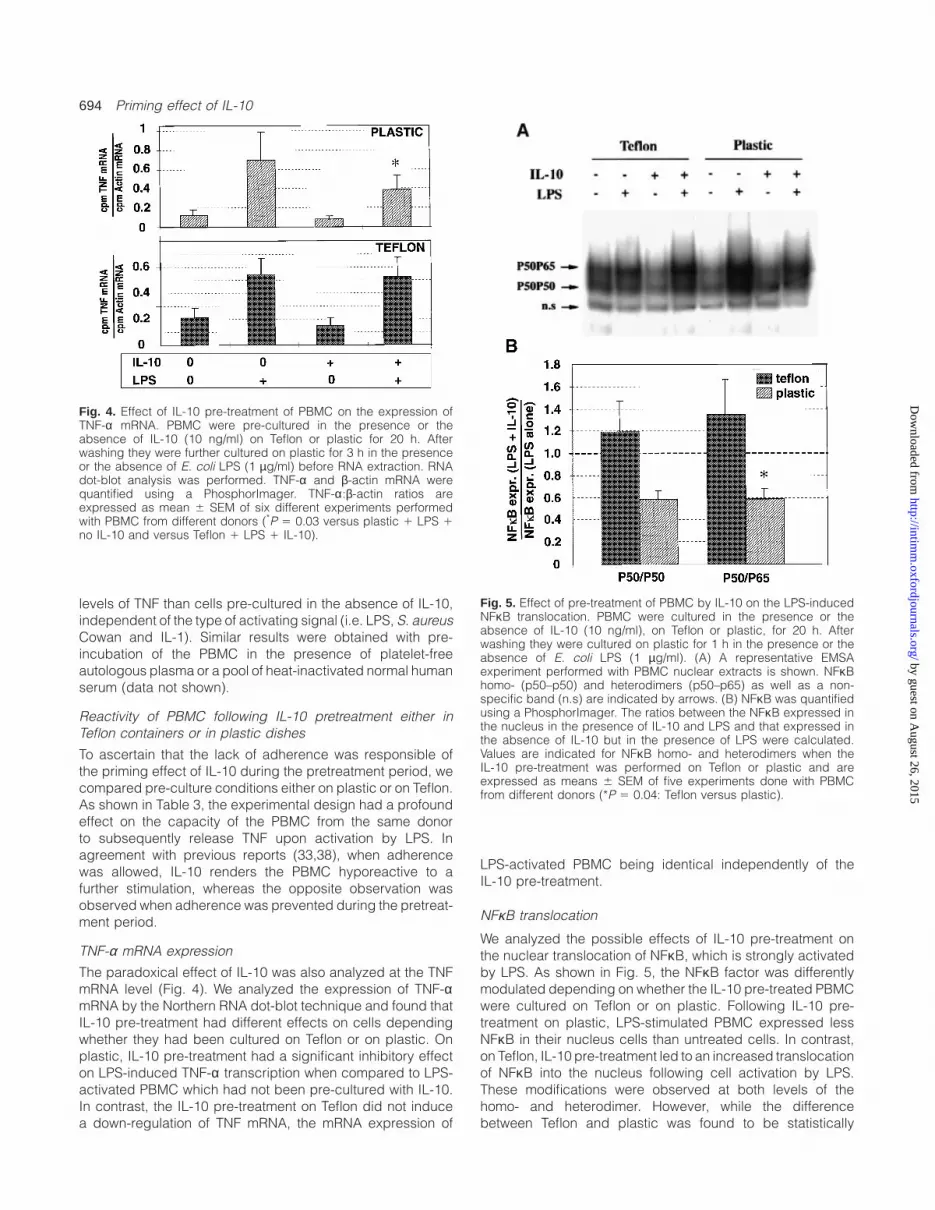
Fig. 4. Effect of IL-10 pre-treatment of PBMC on the expression ofTNF-α mRNA. PBMC were pre-cultured in the presence or theabsence of IL-10 (10 ng/ml) on Teflon or plastic for 20 h. Afterwashing they were further cultured on plastic for 3 h in the presenceor the absence of E. coli LPS (1 µg/ml) before RNA extraction. RNAdot-blot analysis was performed. TNF-α and β-actin mRNA werequantified using a PhosphorImager. TNF-α:β-actin ratios areexpressed as mean 6 SEM of six different experiments performedwith PBMC from different donors (*P 5 0.03 versus plastic 1 LPS 1no IL-10 and versus Teflon 1 LPS 1 IL-10).
levels of TNF than cells pre-cultured in the absence of IL-10,independent of the type of activating signal (i.e. LPS, S. aureusCowan and IL-1). Similar results were obtained with pre-incubation of the PBMC in the presence of platelet-freeautologous plasma or a pool of heat-inactivated normal humanserum (data not shown).
Reactivity of PBMC following IL-10 pretreatment either inTeflon containers or in plastic dishes
To ascertain that the lack of adherence was responsible ofthe priming effect of IL-10 during the pretreatment period, wecompared pre-culture conditions either on plastic or on Teflon.As shown in Table 3, the experimental design had a profoundeffect on the capacity of the PBMC from the same donorto subsequently release TNF upon activation by LPS. Inagreement with previous reports (33,38), when adherencewas allowed, IL-10 renders the PBMC hyporeactive to afurther stimulation, whereas the opposite observation wasobserved when adherence was prevented during the pretreat-ment period.
TNF-α mRNA expression
The paradoxical effect of IL-10 was also analyzed at the TNFmRNA level (Fig. 4). We analyzed the expression of TNF-αmRNA by the Northern RNA dot-blot technique and found thatIL-10 pre-treatment had different effects on cells dependingwhether they had been cultured on Teflon or on plastic. Onplastic, IL-10 pre-treatment had a significant inhibitory effecton LPS-induced TNF-α transcription when compared to LPS-activated PBMC which had not been pre-cultured with IL-10.In contrast, the IL-10 pre-treatment on Teflon did not inducea down-regulation of TNF mRNA, the mRNA expression of
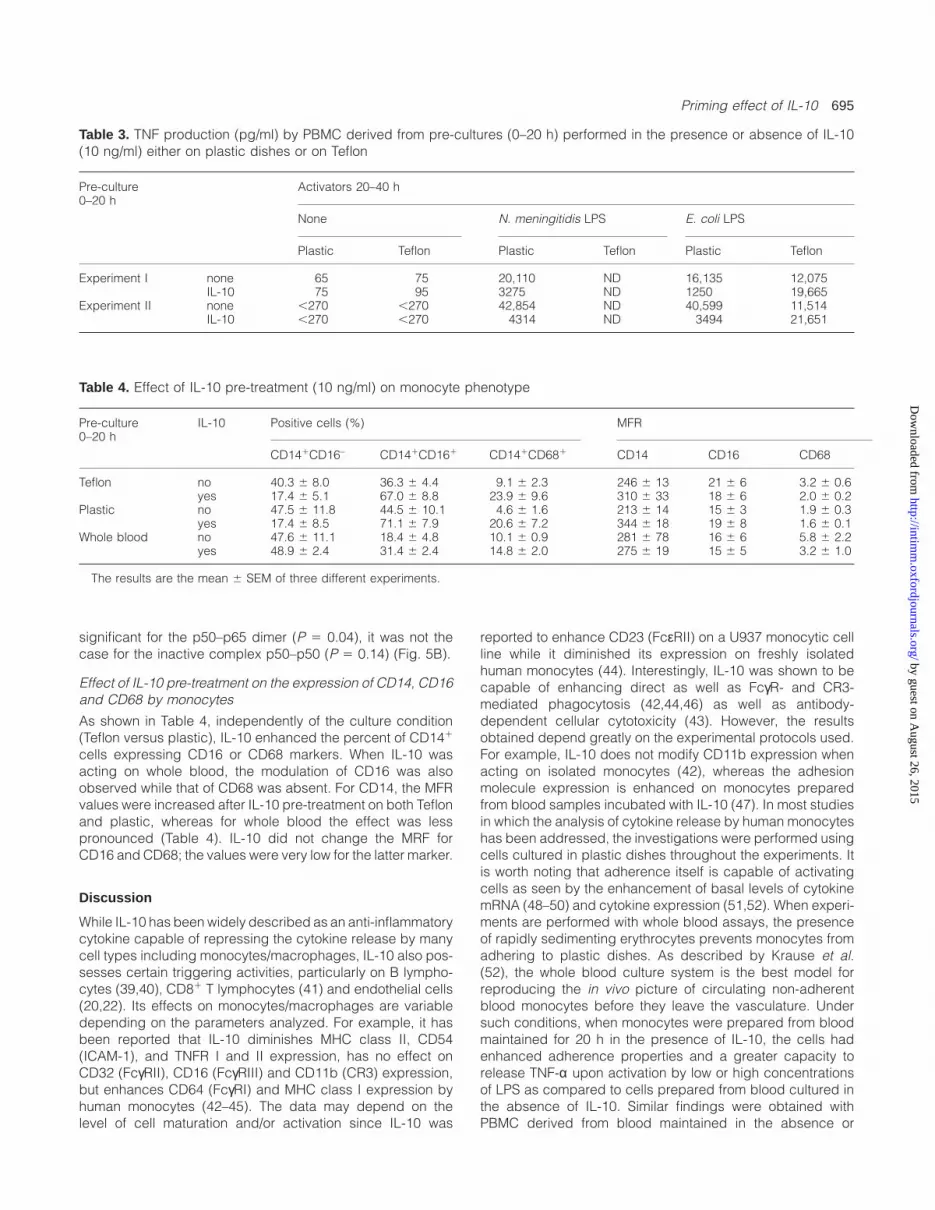
Fig. 5. Effect of pre-treatment of PBMC by IL-10 on the LPS-inducedNFκB translocation. PBMC were cultured in the presence or theabsence of IL-10 (10 ng/ml), on Teflon or plastic, for 20 h. Afterwashing they were cultured on plastic for 1 h in the presence or theabsence of E. coli LPS (1 µg/ml). (A) A representative EMSAexperiment performed with PBMC nuclear extracts is shown. NFκBhomo- (p50–p50) and heterodimers (p50–p65) as well as a non-specific band (n.s) are indicated by arrows. (B) NFκB was quantifiedusing a PhosphorImager. The ratios between the NFκB expressed inthe nucleus in the presence of IL-10 and LPS and that expressed inthe absence of IL-10 but in the presence of LPS were calculated.Values are indicated for NFκB homo- and heterodimers when theIL-10 pre-treatment was performed on Teflon or plastic and areexpressed as means 6 SEM of five experiments done with PBMCfrom different donors (*P 5 0.04: Teflon versus plastic).
LPS-activated PBMC being identical independently of theIL-10 pre-treatment.
NFκB translocation
We analyzed the possible effects of IL-10 pre-treatment onthe nuclear translocation of NFκB, which is strongly activatedby LPS. As shown in Fig. 5, the NFκB factor was differentlymodulated depending on whether the IL-10 pre-treated PBMCwere cultured on Teflon or on plastic. Following IL-10 pre-treatment on plastic, LPS-stimulated PBMC expressed lessNFκB in their nucleus cells than untreated cells. In contrast,on Teflon, IL-10 pre-treatment led to an increased translocationof NFκB into the nucleus following cell activation by LPS.These modifications were observed at both levels of thehomo- and heterodimer. However, while the differencebetween Teflon and plastic was found to be statistically
by guest on August 26, 2015
http://intimm
.oxfordjournals.org/D
ownloaded from
Priming effect of IL-10 695
Table 3. TNF production (pg/ml) by PBMC derived from pre-cultures (0–20 h) performed in the presence or absence of IL-10(10 ng/ml) either on plastic dishes or on Teflon
Pre-culture Activators 20–40 h0–20 h
None N. meningitidis LPS E. coli LPS
Plastic Teflon Plastic Teflon Plastic Teflon
Experiment I none 65 75 20,110 ND 16,135 12,075IL-10 75 95 3275 ND 1250 19,665
Experiment II none ,270 ,270 42,854 ND 40,599 11,514IL-10 ,270 ,270 4314 ND 3494 21,651
Table 4. Effect of IL-10 pre-treatment (10 ng/ml) on monocyte phenotype
Pre-culture IL-10 Positive cells (%) MFR0–20 h
CD141CD16– CD141CD161 CD141CD681 CD14 CD16 CD68
Teflon no 40.3 6 8.0 36.3 6 4.4 9.1 6 2.3 246 6 13 21 6 6 3.2 6 0.6yes 17.4 6 5.1 67.0 6 8.8 23.9 6 9.6 310 6 33 18 6 6 2.0 6 0.2
Plastic no 47.5 6 11.8 44.5 6 10.1 4.6 6 1.6 213 6 14 15 6 3 1.9 6 0.3yes 17.4 6 8.5 71.1 6 7.9 20.6 6 7.2 344 6 18 19 6 8 1.6 6 0.1
Whole blood no 47.6 6 11.1 18.4 6 4.8 10.1 6 0.9 281 6 78 16 6 6 5.8 6 2.2yes 48.9 6 2.4 31.4 6 2.4 14.8 6 2.0 275 6 19 15 6 5 3.2 6 1.0
The results are the mean 6 SEM of three different experiments.
significant for the p50–p65 dimer (P 5 0.04), it was not thecase for the inactive complex p50–p50 (P 5 0.14) (Fig. 5B).
Effect of IL-10 pre-treatment on the expression of CD14, CD16and CD68 by monocytes
As shown in Table 4, independently of the culture condition(Teflon versus plastic), IL-10 enhanced the percent of CD141
cells expressing CD16 or CD68 markers. When IL-10 wasacting on whole blood, the modulation of CD16 was alsoobserved while that of CD68 was absent. For CD14, the MFRvalues were increased after IL-10 pre-treatment on both Teflonand plastic, whereas for whole blood the effect was lesspronounced (Table 4). IL-10 did not change the MRF forCD16 and CD68; the values were very low for the latter marker.
Discussion
While IL-10 has been widely described as an anti-inflammatorycytokine capable of repressing the cytokine release by manycell types including monocytes/macrophages, IL-10 also pos-sesses certain triggering activities, particularly on B lympho-cytes (39,40), CD81 T lymphocytes (41) and endothelial cells(20,22). Its effects on monocytes/macrophages are variabledepending on the parameters analyzed. For example, it hasbeen reported that IL-10 diminishes MHC class II, CD54(ICAM-1), and TNFR I and II expression, has no effect onCD32 (FcγRII), CD16 (FcγRIII) and CD11b (CR3) expression,but enhances CD64 (FcγRI) and MHC class I expression byhuman monocytes (42–45). The data may depend on thelevel of cell maturation and/or activation since IL-10 was
reported to enhance CD23 (FcεRII) on a U937 monocytic cellline while it diminished its expression on freshly isolatedhuman monocytes (44). Interestingly, IL-10 was shown to becapable of enhancing direct as well as FcγR- and CR3-mediated phagocytosis (42,44,46) as well as antibody-dependent cellular cytotoxicity (43). However, the resultsobtained depend greatly on the experimental protocols used.For example, IL-10 does not modify CD11b expression whenacting on isolated monocytes (42), whereas the adhesionmolecule expression is enhanced on monocytes preparedfrom blood samples incubated with IL-10 (47). In most studiesin which the analysis of cytokine release by human monocyteshas been addressed, the investigations were performed usingcells cultured in plastic dishes throughout the experiments. Itis worth noting that adherence itself is capable of activatingcells as seen by the enhancement of basal levels of cytokinemRNA (48–50) and cytokine expression (51,52). When experi-ments are performed with whole blood assays, the presenceof rapidly sedimenting erythrocytes prevents monocytes fromadhering to plastic dishes. As described by Krause et al.(52), the whole blood culture system is the best model forreproducing the in vivo picture of circulating non-adherentblood monocytes before they leave the vasculature. Undersuch conditions, when monocytes were prepared from bloodmaintained for 20 h in the presence of IL-10, the cells hadenhanced adherence properties and a greater capacity torelease TNF-α upon activation by low or high concentrationsof LPS as compared to cells prepared from blood cultured inthe absence of IL-10. Similar findings were obtained withPBMC derived from blood maintained in the absence or
by guest on August 26, 2015
http://intimm
.oxfordjournals.org/D
ownloaded from

696 Priming effect of IL-10
presence of IL-10. While the effects of plasma and plateletswere shown not to contribute to the observed phenomenon,we investigated the effects of adherence. When PBMC main-tained during the pre-culture period for 20 h in the presenceof IL-10 were first cultured on plastic or on Teflon, a significantdifference was observed. In the presence of IL-10, a reducedTNF-α production was found among cells pre-cultured onplastic as we previously reported (33), whereas enhancedTNF-α production was detected when cells were derived fromTeflon containers. The phenomenon was not specific to LPSstimulation, suggesting that the modulation of CD14 expres-sion by IL-10 (44) is not sufficient to explain the observation.
Indeed, our results suggest that the up-regulation of CD14expression by IL-10 does not explain its paradoxical effecton TNF-α production since a similar modulation was observedon plastic and Teflon. The analysis of the phenotype of PBMCin terms of CD16 and CD68 revealed that IL-10 increasedthe frequency of CD141 CD161 monocytes, in accordancewith another report (53). The important point was that IL-10acted on the expression of the tested molecules in the sameway on Teflon and plastic. The IL-10 pre-treatment alsoincreased the percent of CD141 cells expressing the CD68marker. Indeed, in vitro experiments on monocytes haveshown that IL-10 in the presence of granulocyte macrophagecolony stimulating factor and IL-13 induced their maturationto macrophages from day 4 of culture (53). Thus, there wasa discrepancy between the capacity of IL-10 to modulatesome cell surface markers which are not influenced byadherence and the capacity of IL-10 to modulate a latercytokine production which is influenced by adherence.
To further understand the mechanisms involved in thisobservation performed at the protein level, we investigatedwhether the various conditions of pre-cultures with IL-10 mayalso differently affect the TNF mRNA expression. This wasthe case and the IL-10 pre-treatment on Teflon did not modifythe LPS-induced mRNA expression as compared to untreatedcells, whereas similar IL-10 pre-treatment on plastic led to afurther decreased expression of TNF mRNA. The observationsat the transcriptional level did not totally parallel those obtainedat the protein level, and analysis of TNF mRNA half-life andkinetics would require further studies. Nevertheless, the LPS-induced mRNA expression following IL-10 pre-culture onTeflon was significantly higher than that obtained followingIL-10 pre-treatment of plastic (P 5 0.03). Thus we wanted toknow whether the observed different modulations of the cellreactivity by IL-10 were upstream the mRNA transduction.For that purpose we analyzed the translocation of NFκB, oneof the main nuclear factor that regulates the transcription ofnumerous genes of cytokines, including TNF-α, IL-1β, IL-6and IL-8 (54). The NFκB family is composed of variousmembers, p50 (NFκB1), p52 (NFκB2), p65 (RelA), RelB andc-Rel, which can form homo- and heterodimers. In mostcells, the complex that is commonly found is the p50–p65 heterodimer which is a potent transactivator, while it isgenerally admitted that the p50–p50 homodimer is not (55).In plastic culture conditions, IL-10 showed an inhibitory activitytowards NFκB translocation. However, when IL-10 pre-treat-ment of PBMC was performed in the absence of adherence,i.e. on Teflon, the nuclear expression of NFκB was moderatelyenhanced. The p50–p65 NFκB heterodimer expression
induced by LPS was significantly lower in the cells culturedin plastic than in Teflon. There are few in vitro studies in theliterature dealing with the effects of IL-10 on NFκB activationin human PBMC and monocytes showing contradictory results(56,57). Both experiments were carried out in classical cultureconditions on plastic; Wang et al. (56) found that IL-10inhibited the LPS-induced NFκB activity, while Dokter et al.(57) found that it did not. Our results for PBMC cultured onplastic are similar to those of Wang et al. and in addition weshow that in the absence of adherence the effect of IL-10can be completely reversed.
An in vivo model in which an enhanced TNF-α productionwas observed following IL-10 treatment of mice in a model ofmultiple organ dysfunction syndrome induced by i.p. injectionof zymosan further supports our findings (58). The sequenceof the signals delivered to the cells as well as the nature ofthe responding cells are crucial for the quality of the cellreactivity. This has been well illustrated when studying themodulatory properties of IL-4 (29–31) or of cortisol (59).
Our experiments clearly indicate that adherence is animportant co-factor which greatly influences the furtherbehavior of the monocytes/macrophages in response toIL-10. These data suggest that depending on the compart-ment, circulating blood versus tissues, IL-10 may differentiallyinfluence the responsiveness of monocytes/macrophages.Finally, these results should be kept in mind when IL-10 isconsidered as a therapeutic agent in human inflammatoryconditions.
Acknowledgements
The authors thank Dr P. Poncet for his valuable help for the FACSanalysis and K. W. Moore (DNAX) for his kind gift of IL-10. This workwas presented in part at the 37th ICAAC Meeting, Toronto, September1997, abstract symposium S13.
Abbreviations
EMSA electrophoretic mobility shift assayLPS lipopolysaccharideMRF mean fluorescence ratioPBMC peripheral blood mononuclear cellsPE phycoerythrinPMN polymorphoneutrophilsTNF tumor necrosis factor
References
1 Fiorentino, D. F., Bond, M. W. and Mosmann, T. R. 1989. Twotypes of mouse T helper cell. IV. Th2 clones secrete a factor thatinhibits cytokine production by Th1 clones. J. Exp. Med. 170:2081.
2 Vieira, P., de Waal-Malefyt, R., Dang, M. N., Johnson, K. E.,Kastelein, R., Fiorentino, D. F., de Vries, J. E., Roncarolo, M. G.,Mosmann, T. R. and Moore, K. W. 1991. Isolation and expressionof human cytokine synthesis inhibitory factor cDNA clones:homology to Epstein–Barr virus open reading frame BCRFI. Proc.Natl Acad. Sci. USA 88:1172.
3 Fiorentino, D. F., Zlotnik, A., Vieira, P., Mosmann, T. R., Howard, M.,Moore, K. W. and O’Garra, A. 1991. IL-10 acts on the antigen-presenting cell to inhibit cytokine production by Th1 cells.J Immunol. 146:3444.
4 Taga, K. and Tosato, G. 1992. IL-10 inhibits human T cellproliferation and IL-2 production. J Immunol. 148:1143.
5 de Waal Malefyt, R., Abrams, J., Bennet, B., Figdor, C. G. andde Vries, J. E. 1991. Interleukin 10 (IL-10) inhibits cytokine
by guest on August 26, 2015
http://intimm
.oxfordjournals.org/D
ownloaded from

Priming effect of IL-10 697
synthesis by human monocytes: an autoregulatory role of IL-10produced by monocytes. J. Exp. Med. 174:1209.
6 Fiorentino, D. F., Zlotnik, A., Mosmann, T. R., Howard, M. andO’Garra, A. 1991. IL-10 inhibits cytokine production by activatedmacrophages. J. Immunol. 147:3815.
7 Cassatella, M. A., Meda, L., bonora, S., Ceska, M. andConstantin, G. 1993. Interleukin 10 (IL-10) inhibits the releaseof proinflammatory cytokines from human polymorphonuclearleukocytes. Evidence for an autocrine role of tumor necrosis factorand IL-1β in mediating the production of IL-8 triggered bylipopolysaccharide. J. Exp. Med. 178:2207.
8 Kasama, T., Strieter, R. M., Lukacs, N. W., Burdick, M. D. andKunkel, S. L. 1994. Regulation of neutrophil derived chemokineexpression by IL-10. J. Immunol. 1994:3559.
9 Marie, C., Pitton, C., Fitting, C. and Cavaillon, J.-M. 1996.Regulation by anti-inflammatory cytokines (IL-4, IL-10, IL-13,TGFβ) of interleukin-8 production by LPS- and /or TNF-α-activatedhuman polymorphonuclear cells. Mediators Inflam. 5:334.
10 Zheng, X. X., Steele, A. W., Nickerson, P. W., Steurer, W., Steiger, J.and Strom, T. B. 1995. Administration of noncytolytic IL-10/Fc inmurine models of lipopolysaccharide-induced septic shock andallogeneic islet transplantation. J. Immunol. 154:5590.
11 Qian, S. G., Li, W., Li, Y. P., Fu, F. M., Lu, L. N., Fung, J. J.and Thomson, A. W. 1996. Systemic administration of cellularinterleukin-10 can exacerbate cardiac allograft rejection in mice.Transplantation 62:1709.
12 Li, W., Lu, L., Li, Y., Fu, F., Fung, J. J., Thomson, A. W. andQian, S. 1997. High-dose cellular IL-10 exacerbates rejection andreverses effects of cyclosporine and tacrolimus in mouse cardiactransplantation. Transplant. Proc. 29:1081.
13 Rosenbaum, J. T. and Angell, E. 1995. Paradoxical effects of IL-10 in endotoxin-induced uveitis. J. Immunol. 155:4090.
14 Wogensen, L., Lee, M. S. and Sarvetnick, N. 1994. Productionof interleukin-10 by islet cells accelerates immune-mediateddestruction of β cells in nonobese diabetic mice. J. Exp. Med.179:1379.
15 Moritani, M., Yoshimoto, K., Tashiro, F., Hashimoto, C., Miyazaki, J.,Li, S., Kudo, E., Iwahana, H., Hayashi, Y., Sano, T. and Itakura, M.1994. Transgenic expression of IL-10 in pancreatic islet A cellsaccelerates autoimmune insulitis and diabetes in non-obesediabetic mice. Int. Immunol. 6:1927.
16 Berman, R. M., Suzuki, T., Tahara, H., Robbins, P. D., Narula,S. K. and Lotze, M. T. 1996. Systemic administration of cellular IL-10 induces an effective specific and long-lived immune responseagainst established tumor in mice. J. Immunol. 157:231.
17 Di Carlo, E., Coletti, A., Modesti, A., Giovarelli, M., Forni, G. andMusiani, P. 1998. Local release of interleukin-10 by transfectedmouse adenocarcinoma cells exhibits pro- and anti-inflammatoryactivity and results in a delayed tumor rejection. Eur. CytokineNetw. 9:61.
18 Chernoff, A., Granowitz, E., Shapiro, L., Vannier, E.,Lonnemann, G., Angel, J., Kennedy, J., Rabson, A., Wolff, S. andDinarello, C. 1995. A randomized, controlled trial of IL-10 inhumans. Inhibition of inflammatory cytokine production andimmune responses. J. Immunol. 154:5492.
19 Huhn, R. D., Radwanski, E., O’Connell, S. M., Sturgill, M. G.,Clarke, L., Cody, R. P., Affrime, M. B. and Cutler, D. L. 1996.Pharmacokinetics and immunomodulatory properties ofintravenously administered recombinant human interleukin-10 inhealthy volunteers. Blood 87:699.
20 Vora, M., Romero, L. I. and Karasek, M. A. 1996. Interleukin-10induces E-selectin on small and large blood vessel endothelialcells. J. Exp. Med. 184:821.
21 Buelens, C., De Groote, D., Goldman, M. and Willems, F. 1996.Differential effects of interleukin-10 on the production ofinterleukin-12 and interleukin-8 by human dendritic cellsgenerated from peripheral blood. Tranplant. Proc. 28:3255.
22 De Beaux, A. C., Maingay, J. P., Ross, J. A., Fearon, K. C. andCarter, D. C. 1995. Interleukin-4 and interleukin-10 increaseendotoxin-stimulated human umbilical vein endothelial cellinterleukin-8 release. J. Interferon Cytokine Res. 15:441.
23 Sironi, M., Munoz, C., Pollicino, T., Siboni, A., Sciacca, F. L.,Bernasconi, S., Vecchi, A., Colotta, F. and Mantovani, A. 1993.
Divergent effects of interleukin-10 on cytokine production bymononuclear phagocytes and endothelial cells. Eur. J. Immunol.23:2692.
24 Becherel, P. A., Le Goff, L., Ktorza, S., Ouaaz, F., Mencia-Huerta,J. M., Dugas, B., Debre, P., Mossalayi, M. D. and Arock, M.1995. Interleukin-10 inhibits IgE-mediated nitric oxide synthaseinduction and cytokine synthesis in normal human keratinocytes.Eur. J. Immunol. 25:2992.
25 Cunha, F. Q., Moncada, S. and Liew, F. Y. 1992. Interleukin-10(IL-10) inhibits the induction of nitric oxide synthase by interferon-gamma in murine macrophages. Biochem. Biophys. Res.Commun. 182:1155.
26 Fouqueray, B., Boutard, V., Philippe, C., Kornreich, A.,Marchant, A., Perez, J., Goldman, M. and Baud, L. 1995.Mesangial cell-derived interleukin-10 modulates mesangial cellresponse to lipopolysaccharide. Am. J. Pathol. 147:176.
27 Betz-Corradin, S. B., Fasel, N., Buchmuller-Rouiller, Y., Ransijn, A.,Smith, J. and Mauel, J. 1993. Induction of macrophage nitricoxide production by interferon-gamma and tumor necrosis factor-alpha is enhanced by interleukin-10. Eur. J. Immunol. 23:2045.
28 Sunyer, T., Rothe, L., Jiang, X., Osdoby, P. and Collin-Osdoby, P.1996. Proinflammatory agents, IL-8 and IL-10, upregulateinducible nitric oxide synthase expression and nitric oxideproduction in avian osteoclast-like cells. J. Cell Biochem. 60:469.
29 Hart, P. H., Jones, C. A. and Finlay-Jones, J. J. 1995. Monocytescultured in cytokine-defined environments differ from freshlyisolated monocytes in their responses to IL-4 and IL-10. J. Leuk.Biol. 57:909.
30 D’Andrea, A., Ma, X., Aste-Amezaga, M., Paganin, C. andTrinchieri, G. 1995. Stimulatory and inhibitory effects of interleukin(IL)-4 and IL-13 on the production of cytokines by humanperipheral blood mononuclear cells: priming for IL-12 and tumornecrosis factor alpha production. J. Exp. Med. 181:537.
31 Kambayashi, T., Jacob, C. O. and Strassmann, G. 1996. IL-4 andIL-13 modulate IL-10 release in endotoxin-stimulated murineperitoneal mononuclear phagocytes. Cell. Immunol. 171:153.
32 Pang, G., Ortega, M., Zighang, R., Reeves, G. and Clancy,R. 1997. Autocrine modulation of IL-8 production by sputumneutrophils in chronic bronchial sepsis. Am. J. Respir. Crit. CareMed. 155:726.
33 Cavaillon, J. M., Pitton, C. and Fitting, C. 1994. Endotoxin toleranceis not a LPS-specific phenomenon: partial mimicry with IL-1,IL-10 and TGFβ. J. Endotoxin Res. 1:21.
34 Cavaillon, J.-M., Marie, C., Caroff, M., Ledur, A., Godard, I.,Poulain, D., Fitting, C. and Haeffner-Cavaillon, N. 1996. CD14/LPSreceptor exhibits lectin-like properties. J. Endotoxin Res. 3:471.
35 Nicoletti, I., Migliorati, G., Pagliacci, M. C., Grignani, F. andRiccardi, C. 1991. A rapid and simple method for measuringthymocyte apoptosis by propidium iodide staining and flowcytometry. J. Immunol Methods. 139:271.
36 Israe¨l, N., Gougerot-Pocidalo, M. A., Aillet, F. and Virelizier, J. L.
1992. Redox status of cells influences constitutive or inducedNFkB translocation and HIV long terminal repeat activity in humanT and monocytic cell lines. J. Immunol. 149:3386.
37 Pociot, F., Briant, L., Jongeneel, C. V., Molvig, J., Worsaae, H.,Abbal, M., Thomsen, M. J. N. and Cambon-Thomsen, A. 1993.Association of tumor necrosis factor (TNF) and class II majorhistocompatibility complex alleles with the secretion of TNF-α andTNF-β by human mononuclear cells : a possible link to insulin-dependent diabetes mellitus. Eur. J. Immunol. 23:224.
38 Randow, F., Syrbe, U., Meisel, C., Krausch, D., Zuckermann, H.,Platzer, C. and Volk, H. 1995. Mechanism of endotoxindesensitization: involvement of interleukin 10 and transforminggrowth factor β. J. Exp. Med. 181:1887.
39 Rousset, F., Garcia, E., DeFrance, T., Peronne, C., Vezzio, N.,Hsu, D. H., Kastelein, R., Moore, K. W. and Banchereau, J. 1992.Interleukin-10 is a potent growth and differentiation factor foractivated human B lymphocytes. Proc. Natl Acad. Sci. USA89:1890.
40 Itoh, K. and Hirohata, S. 1995. The role of IL-10 in human B cellactivation, proliferation and differentiation. J. Immunol. 154:4341.
41 Chen, W. F. and Zlotnik, A. 1991. IL-10: a novel cytotoxic T celldifferentiation factor. J. Immunol. 147:528.
by guest on August 26, 2015
http://intimm
.oxfordjournals.org/D
ownloaded from

698 Priming effect of IL-10
42 Capsoni, F., Minonzio, F., Ongari, A. M., Carbonelli, V., Galli, A.and Zanussi, C. 1995. IL-10 up-regulates human monocytepahogocytosis in the presence of IL-4 and IFNγ. J. Leuk. Biol.58:351.
43 te Velde, A. A., de Waal Malefijt, R., Huijbens, J. F., de Vries, J. E.and Figdor, C. G. 1992. IL-10 stimulates monocytes FcγR surfaceexpression and cytotoxic activity. Distinct regulation of antibody-dependent cellular cytotoxicity by IFNγ, IL-4 and IL-10. J.Immunol. 149:4048.
44 Spittler, A., Schiller, C., Willheim, M., Tempfer, C., Winkler, S. andBoltz-Nitulescu, G. 1995. IL-10 augments CD23 expression onU937 cells and down-regulates IL-4 driven CD23 expression oncultured human blood monocytes: effects of Il-10 and theircytokines on cell phenotype and phagocytosis. Immunology85:311.
45 Joyce, D. A., Gibbons, D. P., Green, P., Steer, J. H., Feldmann, M.and Brennan, F. M. 1994. Two inhibitors of pro-inflammatorycytokine release, interleukin-10 and interleukin-4, have contrastingeffects on release of soluble p75 tumor necrosis factor receptorby cultured monocytes. Eur. J. Immunol. 24:2699.
46 Roilides, E., Dimitriadou, A., Kadiltsoglou, I., Sein, T.,Karpouzas, J., Pizzo, P. A. and Walsh, T. J. 1997. IL-10 exertssuppressive and enhancing effects on antifungal activity ofmononuclear phagocytes against Aspergillus fumigatus. J.Immunol. 158:322.
47 Buchwald, U. K., Geerdes-Fenges, H. F., Vockler, J., Ziege, S.and Lode, H. 1997. Effects of interleukin-10 and prednisolone onthe phagocytosis and adhesion molecule expression of humanpolymorphonuclear neutrophils and monocytes. ICAAC Meeting(Toronto) abstr G23.
48 Fuhlbridgge, R. C., Chaplin, D. D., Kiely, J. M. and Unanue, E. R.1987. regulation of interleukin-1 expression by adherence andlipopolysaccharide. J. Immunol. 138:3799.
49 Darville, T., Tabor, D. R., Theus, S. A. and Jacobs, R. F. 1992.Surface matrix binding alters murine peritoneal mononuclearphagocyte TNF-α and IL-6 induction. Immunol. Invest. 21:539.
50 Osipovich, O. A., Fegeding, K. V., Misuno, N. I., Kolesnikova,
T. S., Savostatin, I. K., Sudarikov, A. B. and Voitenok, N. N. 1993.Differential action of cycloheximide and activation stimuli ontranscription of tumor necrosis factor-α, IL-1β, IL-8 and p53 genesin human monocytes. J. Immunol. 150:4958.
51 Standiford, T. J., Kunkel, S. L., Kasahara, K., Milia, M. J., Rolfe,M. W. and Strieter, R. M. 1991. Interleukin-8 gene expression fromhuman alveolar macrophages: the role of adherence. Am. J.Respir. Cell Mol. Biol. 5:579.
52 Krause, S. W., Kreutz, M. and Andreesen, R. 1997. Differentialeffects of cell adherence on LPS-stimulated cytokine productionby human monocytes and macrophages. Immunobiology196:522.
53 Allavena, P., Piemonti, L., Longoni, D., Bernasconi, S.,Stoppacciao, A., Ruco, L. and Mantovani, A. 1998. IL-10 preventsthe differentiation of monocytes to dendritic cells but promotestheir maturation to macrophages. Eur. J. Immunol. 28:359.
54 Wulczyn, F. G., Krappmann, D. and Scheidereit, C. 1996. TheNFkB/Rel and IkB families: mediators of immune response andinflammation. J. Mol. Med. 74:749.
55 Schmitz, M. L. and Baeuerle, P. A. 1991. The p65 subunit isresponsible for the strong transcription activating potential ofNFkB. EMBO J. 10:3805.
56 Wang, P., Wu, P., Siegel, M. I., Egan, R. W. and Billah, M. M. 1995.Interleukin (IL)-10 inhibits nuclear factor κB (NFκB) activation inhuman monocytes. IL-10 and IL-4 suppress cytokine synthesisby different mechanisms. J. Biol. Chem. 270:9558.
57 Dokter, W. H. A., Koopmans, S. B. and Vellenga, E. 1996. Effectsof IL-10 and IL-4 on LPS-induced transcription factors (AP-1,NFIL6 and NFκB) which are involved in IL-6 regulation.Leukemia 10:1308.
58 Jansen, M. J. J. M., Hendriks, T., de Man, B., Knapen, M., vander Meer, J. W. M. and Goris, R. J. A. 1997. Interleukin-10 andchloropromazine mitigate the development of multiple organdysfunction syndrome in mice. Shock 7 (Suppl.):97.
59 Barber, A. E., Coyle, S. M., Marano, M. A., Fischer, E., Calvano,S. E., Fong, Y., Moldawer, L. L. and Lowry, S. F. 1993.Glucocorticoid therapy alters hormonal and cytokine responsesto endotoxin in man. J. Immunol. 150:1999.
by guest on August 26, 2015
http://intimm
.oxfordjournals.org/D
ownloaded from
Copyright © 2022 FDOKUMEN




















