
Paleoanthropological research in the Tugen Hills, Kenya. Introduction
10
Introduction Paleoanthropological research in the Tugen Hills, Kenya Andrew Hill Keywords: Tugen Hills, Baringo, Neogene, hominoid, hominid. Department of Anthropology, Yale University, Box 208277, New Haven, Connecticut 06520, U.S.A. E-mail: [email protected] Journal of Human Evolution (2002) 42, 1–10 doi:10.1006/jhev.2001.0520 Available online at http://www.idealibrary.com on The Baringo Paleontological Research Project (BPRP) has been investigating the Tugen Hills succession in the Lake Baringo area of Kenya continuously since 1981. This special issue of Journal of Human Evolution presents some of the more recent paleo- anthropological research carried out in the region by the Project. Papers are included on all significant fossiliferous formations in the succession, from the Middle Miocene to the Middle Pleistocene. This introduction provides a history of work in the Tugen Hills region, placing the research of BPRP in context, particularly the work described in the contributions to this volume. The Tugen Hills are formed by a complex tilted horst block within the Kenya Rift Valley (Figure 1). They extend about 100 km N–S along the Rift west of Lake Baringo and expose some 3000 m of rocks in their fault scarps and eastern foothills, and on the dip-slope to the west. Many of these rocks are fossiliferous, and range in age— without much significant interruption— from over 16 Ma to the present. The first visitor to Baringo who recorded geological observations appears to have been Thomson (1884) who mistakenly described the Tugen Hills as metamorphic in com- position. Gregory (1896, 1921) explored the area in 1892 and 1893, and corrected this notion. Some other relevant early work is described in Prior (1903). Leakey & Solomon (1929) used the area as a basis for their Kamasian Pluvial, in the days when some researchers explained climatic fluctu- ations in terms of the pluvial hypothesis, which ostensibly linked inferred climatic oscillations perceived in East African sedi- ments with European Alpine glacial stages (e.g., Brooks, 1914). Fuchs (1950) reported on some of the results of the Cambridge Expedition to the East African Lakes, which had taken place in 1930 and 1931 (see Worthington & Worthington, 1933). He included a discussion of fossils found in sediments now known to belong to the Kapthurin Formation. Willis (1936) pro- vided information from investigations at about the same time, and Shackleton (1951a,b) worked there during the following decade. Geologists of the Kenya Geological Survey contributed to the basic geological mapping of the southern parts of the region (e.g., McCall et al., 1967; Walsh, 1969). A major phase of research in the area was inaugurated by the formation of the East African Geological Research Unit (EAGRU) in the mid-1960s. Partly funded by the British Government, and working in association with the Geological Survey of Kenya, this project was directed by Basil King at Bedford College, University of London. A number of research students mapped and analyzed the geology of the Hills and areas around the Lake (e.g., Martyn, 1969; Chapman, 1971; Mc Cleneghan, 1971; Carney, 1972). In the course of this work it became apparent that the region was very fossiliferous. Indeed, in the early stages of mapping, one of the first fossil hominids from Kenya was discovered by John Kimengich in 1965, while working 0047–2484/02/010001+10$35.00/0 2002 Academic Press
Transcript of Paleoanthropological research in the Tugen Hills, Kenya. Introduction

Introduction Paleoanthropological research in theTugen Hills, Kenya
Andrew Hill
Keywords: Tugen Hills,Baringo, Neogene,hominoid, hominid.
Department of Anthropology, Yale University, Box 208277,New Haven, Connecticut 06520, U.S.A. E-mail: [email protected]
Journal of Human Evolution (2002) 42, 1–10doi:10.1006/jhev.2001.0520Available online at http://www.idealibrary.com on
The Baringo Paleontological ResearchProject (BPRP) has been investigating theTugen Hills succession in the Lake Baringoarea of Kenya continuously since 1981. Thisspecial issue of Journal of Human Evolutionpresents some of the more recent paleo-anthropological research carried out in theregion by the Project. Papers are includedon all significant fossiliferous formations inthe succession, from the Middle Miocene tothe Middle Pleistocene. This introductionprovides a history of work in the Tugen Hillsregion, placing the research of BPRP incontext, particularly the work describedin the contributions to this volume.
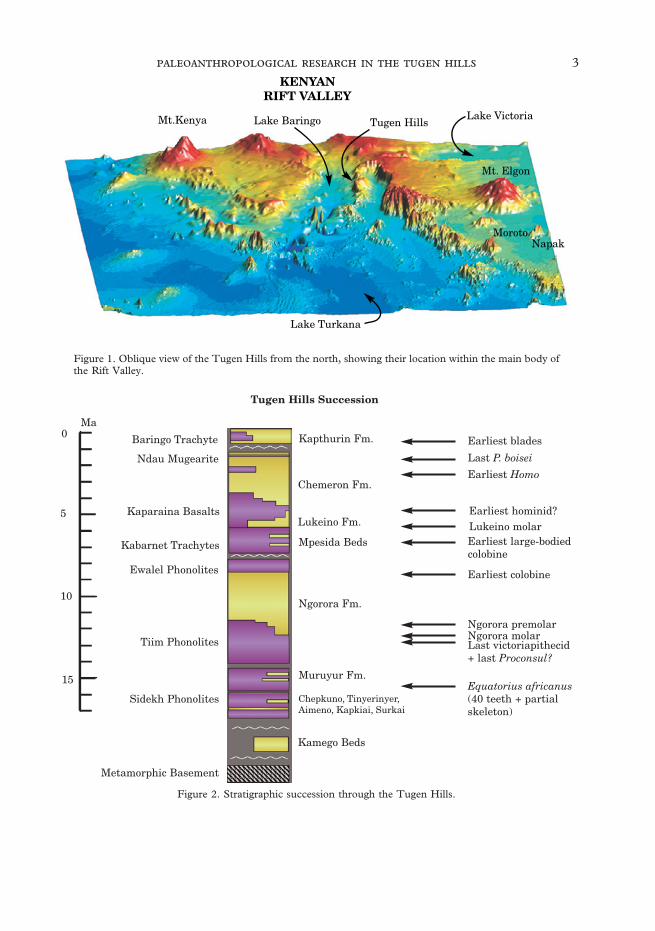
The Tugen Hills are formed by a complextilted horst block within the Kenya RiftValley (Figure 1). They extend about100 km N–S along the Rift west of LakeBaringo and expose some 3000 m of rocks intheir fault scarps and eastern foothills, andon the dip-slope to the west. Many of theserocks are fossiliferous, and range in age—without much significant interruption—from over 16 Ma to the present.
The first visitor to Baringo who recordedgeological observations appears to have beenThomson (1884) who mistakenly describedthe Tugen Hills as metamorphic in com-position. Gregory (1896, 1921) explored thearea in 1892 and 1893, and corrected thisnotion. Some other relevant early workis described in Prior (1903). Leakey &Solomon (1929) used the area as a basis fortheir Kamasian Pluvial, in the days whensome researchers explained climatic fluctu-ations in terms of the pluvial hypothesis,
0047–2484/02/010001+10$35.00/0
which ostensibly linked inferred climaticoscillations perceived in East African sedi-ments with European Alpine glacial stages(e.g., Brooks, 1914). Fuchs (1950) reportedon some of the results of the CambridgeExpedition to the East African Lakes, whichhad taken place in 1930 and 1931 (seeWorthington & Worthington, 1933). Heincluded a discussion of fossils found insediments now known to belong to theKapthurin Formation. Willis (1936) pro-vided information from investigations atabout the same time, and Shackleton(1951a,b) worked there during the followingdecade. Geologists of the Kenya GeologicalSurvey contributed to the basic geologicalmapping of the southern parts of theregion (e.g., McCall et al., 1967; Walsh,1969).
A major phase of research in the area wasinaugurated by the formation of the EastAfrican Geological Research Unit(EAGRU) in the mid-1960s. Partly fundedby the British Government, and working inassociation with the Geological Survey ofKenya, this project was directed by BasilKing at Bedford College, University ofLondon. A number of research studentsmapped and analyzed the geology of theHills and areas around the Lake (e.g.,Martyn, 1969; Chapman, 1971; McCleneghan, 1971; Carney, 1972). In thecourse of this work it became apparent thatthe region was very fossiliferous. Indeed, inthe early stages of mapping, one of the firstfossil hominids from Kenya was discoveredby John Kimengich in 1965, while working
� 2002 Academic Press

2 .
with John Martyn (Martyn, 1967). It is atemporal bone from what is now siteBPRP#2 in the Chemeron Formation, aspecimen initially described by Tobias(1967). Following this discovery, in 1966Louis and Mary Leakey were invited toexcavate the site. At the same time RichardLeakey collected further fossils from thearea, including an entire elephant skeleton(from what is now site BPRP#23 in theChemeron Formation). Margaret Leakeyexcavated sites in the Pleistocene KapthurinFormation that produced archaeologicalremains, and a hominid mandible andpostcranial fossils (Leakey et al., 1969).
Partly in response to such discoveries BillBishop set up a related but separate group atBedford College, to work in association withEAGRU, but focusing on the fossils andtheir sedimentary and paleoenvironmentalcontext. I was the first graduate student onthis project, beginning in 1967, and subse-quent students included Fowler, Tallon andPickford.
The work of EAGRU, with input fromBishop’s group for some sedimentary units,resulted in the documentation of the overallgeological structure of the region and theestablishment of a radiometrically datedstratigraphic framework (Bishop et al., 1971;Chapman & Brook, 1978; Chapman et al.,1978; King & Chapman, 1972). Put simply,although extensive faulting greatly compli-cates the situation, the succession consists ofa number of sedimentary units, mostlyfossiliferous, separated from each other byvolcanic flows. From the bottom to the topthe main sedimentary strata are (withbounding dates refined by our later work):Muruyur Beds (16–13 Ma), NgororaFormation (13–8·5 Ma), Mpesida Beds (7–6·2 Ma), Lukeino Formation (6·2–5·6 Ma),Chemeron Formation (5·6–1·6 Ma) and theKapthurin Formation (700–<200 ka) (seeFigure 2). Most of these sedimentary unitsare exposed over large areas and each con-tains many distinct fossil sites, the Ngorora
and Chemeron Formations in particularbeing of very long duration.
This early work also resulted in thedocumentation of a number of importantlocalities, local sequences, and their fossilfaunas (Bishop, 1971, 1972a,b, 1976;Bishop & Chapman, 1970; Bishop et al.,1971, 1975, 1978; Bishop & Pickford,1975; Carney et al., 1971; Pickford, 1975,1978a,b; Tallon, 1978). It was quickly real-ized that the Tugen Hills sequence wasextensive, contained many sites and gaveinformation about a long period of theAfrican past, from around 16 Ma to quiterecent times. Much of this time period wasotherwise unrepresented in the fossil recordof Africa. For example, it partly filled thelong gap between the Fort Ternan localitydated at about 14 Ma, and much later sitessuch as Lothagam, Kanapoi, Olduvai, andKoobi Fora, that at the time had producedthe earliest hominids.
Expanding knowledge of the Tugen Hillsfaunas, in conjunction with the radiometricdating program, contributed considerablyto the establishment of the sound Africanbiostratigraphy then being developed, andprovided a basis for the correlative datingof sites elsewhere (e.g., Coryndon, 1978;Gentry, 1978; Hill & Walker, 1979;Hooijer, 1969, 1973, 1974; Maglio, 1970,1973; Thomas, 1980, 1981).
The fauna recovered at this time includedprimates. In addition to the Chemeron tem-poral and the material from the KapthurinFormation, two new species of monkey,Paracolobus chemeroni and Theropithecusbaringensis, were described from a site in theChemeron Formation (BPRP#97) (Leakey,1969). Also, an intriguing hominoid toothhad been found from sediments datingto about 12 Ma (BPRP#60) (Bishop &Chapman, 1970), and a cranium and iso-lated teeth of a robust australopithecinewere discovered at Chesowanja on the eastside of Lake Baringo (Carney et al., 1971;Bishop et al., 1975, 1978).
3
Figure 1. Oblique view of the Tugen Hills from the north, showing their location within the main body ofthe Rift Valley.
Figure 2. Stratigraphic succession through the Tugen Hills.

4 .
This phase of research more or less con-cluded with Bishop’s untimely death and theposthumous publication of an edited vol-ume, which included a number of these laterpapers (Bishop, 1978). Pickford had contin-ued to work in the Tugen Hills for a coupleof years in a postdoctoral position withBishop, but abandoned the area to pursueresearch on the Miocene of western Kenya.
The Baringo Paleontological ResearchProject was established by David Pilbeam in1980 to investigate the Tugen Hills succes-sion and neighboring sites, with myself asfield director. It was, and remains, a col-laborative project run jointly with theNational Museums of Kenya. In 1985Pilbeam generously handed over directionto me, and it has been run from YaleUniversity since then. The project is multi-disciplinary and involves the participation ofscientists from a number of institutions andcountries. From the beginning it was con-cerned with the entire succession. Besidesour interest in any hominoids or other pri-mates the sequence might produce, a pri-mary research focus has always been theevolution of the Ethiopian fauna in relationto local environments, global climate, andpaleogeography. Initially we concentratedon the Miocene Ngorora Formation, mainlymotivated by an interest in comparing theevolution of faunas from this time period inAfrica to those in Pakistan (Barry et al.,1985; Hill et al., 1985), but later parts of thesuccession were not neglected (Hill, 1985a).
BPRP’s research has clarified the basicstratigraphy of the Tugen Hills successionand its radiometric and paleomagnetic agecalibration (e.g., Hill et al., 1985, 1986;Deino et al., 1990; Tauxe et al., 1985), andNamwamba (1993) presented preliminaryinternal tephrostratigraphic correlation, andto sites distal to it, such as those in theTurkana Basin.
BPRP’s work on the fauna has resulted innew taxonomic information and has refinedthe biostratigraphic scheme through the
later part of the African Neogene. This hasbeen useful in discerning temporal relation-ships with other sites in Africa, Pakistan(Hill et al., 1985), and Arabia (Bishop &Hill, 1999; Hill, 1999a). In addition tohominoids, the rodents (Winkler, 1990,1992) and pigs (Bishop, 1993, 1994; Bishopet al., 1999) have so far been studied in sometaxonomic detail. This research has resultedin insights into changing environments, bothon a large scale, and more locally, as in thecase of Bishop’s ecomorphological work onsuids (Bishop et al., 1999).
Other paleoenvironmental data derivefrom paleobotanic remains and from iso-topic indicators of environmental andclimate change. Jacobs and colleagues haveexamined macrobotanical fossils at variouspoints in the succession (Jacobs & Kabuye,1987; Jacobs & Winkler, 1992; Jacobs,1994; Jacobs & Deino, 1996; Kabuye &Jacobs, 1986). Kingston has investigated arange of isotopic evidence for vegetation insoil carbonates, and for mammal feedingstrategies in tooth enamel (Bishop et al.,1999; Kingston, 1992, 1999; Kingstonet al., 1992, 1994; Morgan et al., 1994).
Hominoids have been discovered, andearlier discoveries reappraised, since 1981(Hill & Ward, 1988; Hill, 1994, 1998; Hillet al., 1992a). In 1984 a partial mandiblewas found at Tabarin (BPRP#77) (Hill,1985b; Ward & Hill, 1987). It expanded theknown time range of hominids by about25%, and still remains one of the earliesthominids known. Further hominoid teethwere found in the Ngorora Formation (Hillet al., 1985; Hill & Ward, 1988). Ourre-examination of the temporal bonefrom the Chemeron Formation (BPRP#2)convinced us that it belonged to Homo, andradiometric dating of strata at the siteshowed it to be the oldest known example ofthat genus (Hill et al., 1992b,c; Sherwoodet al., 1996). Subsequent work in theMuruyur Formation at the Kipsaramon sitecomplex resulted in a variety of hominoids

5
from the bone bed site (BPRP#89) (Hill,1989; Hill et al., 1991). This was followed,from 1990 onwards, by the discovery of aseries of hominoid teeth from a slightlyyounger Kipsaramon location (BPRP#122)(Brown et al., 1991), and then a partialskeleton at this site (Ward et al., 1996). Thematerial from BPRP#122 was all ultimatelyattributed to the new genus Equatorius(Ward et al., 1999; Kelley et al., 2000).There are a few other isolated hominidspecimens (including ‘‘the Black Cusp’’)from various sites higher in the sequence, sofar unpublished. These include a specimenfound from a new basin of sediments duringour 2001 field season.
This accumulated information is import-ant for understanding the relation of theevolution of the Ethiopian fauna, includinghominoids, to climatic and environmentalchange (Hill, 1986, 1987, 1995, 1999b).Our paleobotanical and isotopic results sofar reveal no evidence at any time from15·5 Ma onwards for a sudden unidirec-tional change in environments that thosewho believe in strong environmental forcingof human evolution have postulated sinceDarwin. Rather the vegetation appears tohave been mixed throughout, with inter-ludes of substantial forest as late as6·5 Ma.
In 1990 an allied project was begun underthe direction of Sally McBrearty to investi-gate the Kapthurin Formation, which hadnot been examined in any detail by BPRP.Although largely independent, the projectscollaborate closely and share some of thesame personnel. The Kapthurin For-mation spans about 500 ka of the MiddlePleistocene, and contains sites recordingevents surrounding the latest Acheulian andearliest Middle Stone Age in Africa. It hasproduced hominid fossils and a variety ofarchaeological occurrences. Among theseare the oldest known sites in Africa attribu-table to the Middle Stone Age, the oldestblades in Africa, perhaps in the world, and
very early pigment in the form of red ochre(McBrearty, 1999; McBrearty et al., 1996,1999, 2001).
This special issue of JHE contains paperson most of the fossiliferous formationsthrough the sequence, particularly asthey relate to fossil primate discoveries.Behrensmeyer et al. (2002) describe thebasic stratigraphy of the Muruyur Beds nearthe base of the sequence, at the Kipsaramonsite complex and in the type area. Fullresults and analytical information on a seriesof age determinations through the wholesuccession are given, including importantdating results relevant to the age of fossilhorizons. These include the bone bed(BPRP#89) from which have come speci-mens of Victoriapithecus, ‘‘Proconsul major’’and Kalepithecus, and BPRP#122, the site ofthe Equatorius africanus partial skeleton andof isolated teeth from other individuals ofthis species. Radiometric correlations withother Muruyur exposures in the type areasome distance away are also provided. Twoadditional papers describe the dentalmaterial of Equatorius from BPRP#122in some detail (Kelley et al., 2002), andprovides a preliminary description of theskeleton (KNM-TH 28860) (Sherwoodet al., 2002).
Hill et al. (2002) provide descriptions ofa collection of cercopithecoid teeth fromBPRP#38 in the succeeding NgororaFormation, dated at 12·5 Ma, and a discus-sion of their context. These teeth belong to aspecies of victoriapithecid 2 million yearslater in time than any other known occur-rence of victoriapithecids. In fact, they arethe only fossil specimens of monkey inAfrica between the age of about 15 Ma and8·5 Ma, when the first colobines arerecorded, also in the Tugen Hills succession(Gundling & Hill, 2000). The Victoria-pithecus assemblage at BPRP#38 occurs justabove one of the finest macro-floral horizonsin Africa, that indicates a forest environ-ment. A hominoid tooth, cf Proconsul (and if

6 .
Acknowledgements
The Baringo Paleontological ResearchProject is based at Yale University, and
so, the youngest occurrence of this genus), isalso described from this site.
Kingston et al. (2002) give informationabout the little known Mpesida Beds. Theselie above the Ngorora Formation, inter-calated within flows of the KabarnetTrachytes. Details are given about a fossilforest at Rormuch (BPRP#85), radiometricdating results, and reference is made to newspecimens of a colobine monkey.
The succeeding sedimentary unit in theTugen Hills succession is the LukeinoFormation, which itself is overlain, abovethe Kaparaina Basalts in this region, by theChemeron Formation. Deino et al. (2002)present a sequence of radiometric dates anda paleomagnetic calibration through theupper part of the Lukeino Formation at thesite of Kapcheberek (BPRP#76), continuingup through the lower part of the ChemeronFormation as represented at the site ofTabarin (BPRP#77) in the same vicinity.The Tabarin site is especially important ashaving produced one of the oldest knownhominids (KNM-TH 13150) (Hill, 1985b;Ward & Hill, 1987; Hill & Ward, 1988) andthe new dates slightly revise previous ageestimates for the specimen. The Tabarinsequence extends up to a volcanic unit thatwe believe may be equivalent to the MoitiTuff in the Turkana Basin.
Later parts of the Chemeron Formationare discussed by Sherwood et al. (2002a)and by Deino & Hill (2002). Sherwood et al.provide new details concerning the taxo-nomic status of the Chemeron Temporal(KNM-BC 1) that reinforce earlier con-clusions that the specimen belongs to genusHomo sensu lato. Deino & Hill (2002) pro-vide additional detailed radiometric datinginformation through the succession in theenvirons of the site (BPRP#2) that confirmsthat the specimen represents, perhaps, theearliest known example of Homo. The latterpaper also includes dates from near the baseof the local Chemeron Formation sequencein the Kapthurin River that are import-
ant for the ages of the type specimens ofParacolobus chemeroni and Theropithecusbaringensis.
Radiometric determinations given byDeino & McBrearty (2002) provide refinedage estimates for hominids from the MiddlePleistocene Kapthurin Formation, andincrease the age for the known Middle StoneAge (MSA) in Africa by about 50 ka. In oneof the following papers, Tryon & McBrearty(2002) present tuff fingerprinting data fromthe Kapthurin Formation, that enhancescorrelation among sites and clarifies thenature of the Acheulian–MSA transition.
Finally, taking a broad perspective,Winkler (2002) discusses the rodents andlagomorphs throughout the whole sequence,and provides valuable information onpaleoenvironments and paleobiogeographyfor the Neogene of Africa.
BPRP intends to continue working in theTugen Hills in the coming years. The succes-sion promises to provide opportunities forbetter understanding the origin of clades andthe pattern of African evolution in the Neo-gene. There is also the strong possibility offossils of interest to paleoanthropology fromtime periods so far unrepresented in therecord elsewhere. One new aspect of ourresearch, so far only published in abstracts(Deino et al., 1999; Kingston et al., 2000),concerns our discovery of clear indicatorsof the 23 ka precessional element of theMilankovitch cycle in the Chemeron For-mation around 2·5 Ma (also involving a moreprecise dating of the Gauss–Matuyamaboundary). At present we are examiningmore closely the fossil specimens, including afew hominids, from the many Tugen Hillssites around this time period in the hope ofdetecting the effects, if any, this climaticcycle has on the evolution of the fauna.

7
carried out in collaboration with theNational Museums of Kenya. We thank theGovernment of the Republic of Kenya forpermission to carry out research in Kenya(Permit OP/13/001/C 1391/), and permis-sion to excavate from the Minister for HomeAffairs and National Heritage. Currently,or in the past, BPRP has been supportedby grants from the National ScienceFoundation (most recently SBR-9208903),the Louise H. and David S. Ingalls Foun-dation, the Louise Brown Foundation, theL. S. B. Leakey Foundation, Mr J. ClaytonStephenson, the Schwartz Family Foun-dation, and Yale University. The Kapthurinwork directed by McBrearty has beenfunded by a grant to her from the NationalScience Foundation (SBR-9601419) and bya University of Connecticut Chancellor’sFellowship. I thank David Pilbeam forinitiating the project, and for generouslypassing the direction of it over to me in1985. Also, I would thank all those whohave worked, and continue to work, on theproject. Sally McBrearty provided valuableeditorial suggestions.
References
Barry, J., Hill, A. & Flynn, L. (1985). Variation de lafaune au Miocene inferieur et moyen de l’Afrique del’est. L’Anthropologie (Paris) 89, 271–273.
Behrensmeyer, A. K., Deino, A. L., Hill, A., Kingston,J. D. & Saunders, J. J. (2002). Geology and geo-chronology of the middle Miocene Kipsaramon sitecomplex, Muruyur Beds, Tugen Hills, Kenya. J.hum. Evol. 1/2, 11–38.
Bishop, L. C. (1993). Hominids of the East African Riftin macroevolutionary context. Am. J. phys. Anthrop.16(Supplement), 57.
Bishop, L. C. (1994). Pigs and the ancestors: hominids,suids and environments during the Plio-Pleistoceneof East Africa. Ph.D. Dissertation, Yale University.
Bishop, L. C. & Hill, A. (1999). Fossil suidae from theBaynunah Formation, Emirate of Abu Dhabi, UnitedArab Emirates. In (P. J. Whybrow & A. Hills, Eds)Fossil Vertebrates of Arabia, pp. 254–270. New Haven:Yale University Press.
Bishop, L. C., Hill, A. & Kingston, J. D. (1999).Palaeoecology of Suidae from the Tugen Hills,Baringo, Kenya. In (P. Andrews & P. Banham, Eds)Late Cenozoic Environments and Hominid Evolution:
a Tribute to Bill Bishop, pp. 99–111. London:Geological Society of London.
Bishop, W. W. (1971). The late Cenozoic historyof East Africa in relation to hominid evolution. In(K. K. Turekian, Ed.) The Late Cenozoic Glacial Ages,pp. 493–527. New Haven: Yale University Press.
Bishop, W. W. (1972a). Stratigraphic succession‘‘versus’’ calibration in East Africa. In (W. W.Bishop, Ed.) Calibration of Hominoid Evolution,pp. 1–28. Edinburgh: Scottish Academic Press.
Bishop, W. W. (1972b). Stratigraphical nomenclaturein the Baringo area of the northern Kenya Rift Valley.In Proceedings of the Pan-African Congress of Prehistory:6th: 1967: Dakar, pp. 332–333. Chambery: Imp.Reunies de Chambery.
Bishop, W. W. (1976). Pliocene problems relating tohuman evolution. In (G. L. Isaac & E. R. McCown,Eds) Human Origins: Louis Leakey and the AfricanEvidence, pp. 139–153. Menlo Park, CA: W. A.Benjamin.
Bishop, W. W. (Ed.) (1978). Geological Backgroundto Fossil Man. Edinburgh: Geological Society ofLondon & Scottish Academic Press.
Bishop, W. W. & Chapman, G. R. (1970). EarlyPliocene sediments and fossils from the northernKenya Rift Valley. Nature 226, 914–918.
Bishop, W. W. & Pickford, M. (1975). Geology, faunaand palaeoenvironments of the Ngorora Formation,Kenya Rift Valley. Nature 254, 185–192.
Bishop, W. W., Chapman, G. R., Hill, A. & Miller,J. A. (1971). Succession of Cainozoic vertebrateassemblages from the northern Kenya Rift Valley.Nature 233, 389–394.
Bishop, W. W., Pickford, M. & Hill, A. (1975).New evidence regarding the Quaternary geology,archaeology, and hominids of Chesowanja, Kenya.Nature 258, 204–208.
Bishop, W. W., Hill, A. & Pickford, M. (1978).Chesowanja: a revised geological interpretation. In(W. W. Bishop, Ed.) Geological Background to FossilMan, pp. 309–327. Edinburgh: Geological Society ofLondon & Scottish Academic Press.
Brooks, C. E. P. (1914). The meterological con-ditions of an ice-sheet and their bearing on thedessication of the globe. R. Meterol. Soc. Quart. J. 40,53–70.
Brown, B., Hill, A. & Ward, S. (1991). New Miocenelarge hominoids from the Tugen Hills, BaringoDistrict, Kenya. Am. J. phys. Anthrop. 12(Supple-ment), 55.
Carney, J. (1972). The geology of the country to theeast of Lake Baringo, Rift Valley Province, Kenya.Ph.D. Dissertation, University of London.
Carney, J., Hill, A., Miller, J. A. & Walker, A. (1971).Late australopithecine from Baringo District, Kenya.Nature 230, 509–514.
Chapman, G. R. (1971). The geological evolution ofthe northern Kamasia Hills, Baringo District, Kenya.Ph.D. Dissertation, University of London.
Chapman, G. R. & Brook, M. (1978). Chronostratig-raphy of the Baringo Basin, Kenya Rift Valley. In(W. W. Bishop, Ed.) Geological Background to Fossil

8 .
Man, pp. 207–223. Edinburgh: Geological Society ofLondon & Scottish Academic Press.
Chapman, G. R., Lippard, S. J. & Martyn, J. E. (1978).The stratigraphy and structure of the KamasiaRange, Kenya Rift Valley. J. Geol. Soc. London 135,265–281.
Coryndon, S. C. (1978). Fossil Hippopotamidae fromthe Baringo Basin and relationships within theGregory Rift, Kenya. In (W. W. Bishop, Ed.)Geological Background to Fossil Man, pp. 279–292.Edinburgh: Geological Society of London & ScottishAcademic Press.
Deino, A. L. & Hill, A. (2002). 40Ar/39Ar dating of theChemeron Formation strata encompassing the site ofhominid KNM-BC 1, Tugen Hills, Kenya. J. hum.Evol. 1/2, 141–15.
Deino, A. L. & McBrearty, S. (2002). 40Ar/39Ar chro-nology for the Kapthurin Formation, Baringo,Kenya. J. hum. Evol. 1/2, 185–210.
Deino, A. L., Tauxe, L., Monaghan, M. & Drake, R.(1990). 40Ar/39Ar age calibration of the litho- andpaleomagnetic stratigraphies of the Ngorora For-mation, Kenya. J. Geol. 98, 567–587.
Deino, A. L., Glen, J. M., Kingston, J. D. & Hill, A.(1999). 40Ar/39Ar age calibration of the Gauss/Matuyama boundary and recognition of precessionalclimatic cycles in the Chemeron Formation, centralKenya Rift. Am. Geophys. Union Meetings: Abstracts.
Deino, A. L., Tauxe, L., Monaghan, M. & Hill, A.(2002). 40Ar/39Ar geochronology and paleomagneticstratigraphy of the Lukeino and lower Chemeronsuccession at Tabarin and Kapcheberek, TugenHills, Kenya. J. hum. Evol. 1/2, 117–140.
Fuchs, V. E. (1950). Pleistocene events in the BaringoBasin. Geol. Mag. 87, 149–174.
Gentry, A. W. (1978). The fossil Bovidae of theBaringo Area, Kenya. In (W. W. Bishop, Ed.)Geological Background to Fossil Man, pp. 293–308.Edinburgh: Geological Society of London & ScottishAcademic Press.
Gregory, J. W. (1896). The Great Rift Valley. London:John Murray.
Gregory, J. W. (1921). The Rift Valley and Geology ofEast Africa. London: Seeley Service.
Gundling, T. & Hill, A. (2000). Geological context offossil Cercopithecoidea from eastern Africa. In (P. F.Whitehead & C. J. Jolly, Eds) Old World Monkeys,pp. 180–213. Cambridge: Cambridge UniversityPress.
Hill, A. (1985a). Les variations de la faune du Miocenerecent et du Pliocene d’Afrique de l’est.L’Anthropologie (Paris) 89, 275–279.
Hill, A. (1985b). Early hominid from Baringo, Kenya.Nature 315, 222–224.
Hill, A. (1986). The Tugen Hills sequence: environ-mental change in the late Miocene and Pliocene.The Longest Record: The Human Career in Africa. AConference in Honour of J. Desmond Clark. Abstracts,pp. 45–46.
Hill, A. (1987). Causes of perceived faunal change inthe later Neogene of East Africa. J. hum. Evol. 16,583–596.
Hill, A. (1989). Kipsaramon: a Miocene hominoid sitein Kenya. Am. J. phys. Anthrop. 78, 241.
Hill, A. (1994). Late Miocene and early PlioceneHominoids from Africa. In (R. S. Corruccini &R. L. Ciochon, Eds) Integrative Paths to the Past:Paleoanthropological Advances in Honor of F. ClarkHowell, pp. 123–145. Englewood Cliffs, NJ: PrenticeHall.
Hill, A. (1995). Faunal and environmental change inthe Neogene of east Africa: evidence from the TugenHills sequence, Baringo District, Kenya. In (E. S.Vrba, G. H. Denton, T. C. Partridge & L. H.Burckle, Eds) Paleoclimate and Evolution, WithEmphasis on Human Origins, pp. 178–193. NewHaven: Yale University Press.
Hill, A. (1998). Very early plausible hominids from theTugen Hills, Kenya. In (M. A. Raath, H. Soodyall,D. Barkhan, K. L. Kuykendall & P. V. Tobias, Eds)Abstracts of Contributions to the Dual Congress 1998,p. 20. Sun City/Johannesburg, South Africa:International Association for the Study of HumanPalaeontology & International Association of HumanBiologists.
Hill, A. (1999a). Late Miocene sub-Saharan verte-brates, and their relation to the Baynunah fauna, AbuDhabi, United Arab Emirates. In (P. J. Whybrow &A. Hill, Eds) Fossil Vertebrates of Arabia, pp. 420–429.New Haven: Yale University Press.
Hill, A. (1999b). The Baringo Basin, Kenya: from BillBishop to BPRP. In (P. Andrews & P. Banham, Eds)Late Cenozoic Environments and Hominid Evolution: aTribute to Bill Bishop, pp. 85–97. London: GeologicalSociety of London.
Hill, A. & Walker, A. (1979). A fossil marabou (Aves:Ciconiidae) from the Miocene Ngorora Formation,Baringo District, Kenya. Netherlands J. Zool. 29,217–224.
Hill, A. & Ward, S. (1988). Origin of the Hominidae:the record of African large hominoid evolutionbetween 14 my and 4 my. Yearb. phys. Anthrop. 31,49–83.
Hill, A., Drake, R., Tauxe, L., Monaghan, M., Barry,J. C., Behrensmeyer, A. K., Curtis, G., Jacobs, B. F.,Jacobs, L. L., Johnson, N. & Pilbeam, D. (1985).Neogene Palaeontology and geochronology of theBaringo Basin, Kenya. J. hum. Evol. 14, 759–773.
Hill, A., Curtis, G. & Drake, R. (1986). Sedimentarystratigraphy of the Tugen Hills, Baringo Disjtrict,Kenya. In (L. E. Frostick, R. W. Renaut, I. Reid &J.-J. Tiercelin, Eds) Sedimentation in the African Rifts,pp. 285–295. Oxford: Blackwell & GeologicalSociety of London Special Publication 25.
Hill, A., Behrensmeyer, A. K., Brown, B., Deino, A. L.,Rose, M., Saunders, J. J., Ward, S. & Winkler, A. J.(1991). Kipsaramon: a lower Miocene hominoid sitein the Tugen Hills, Baringo District, Kenya. J. hum.Evol. 20, 67–75.
Hill, A., Ward, S. & Brown, B. (1992a). Anatomy andage of the Lothagam mandible. J. hum. Evol. 22,439–451.
Hill, A., Ward, S., Deino, A. L., Curtis, G. & Drake, R.(1992b). Earliest Homo. Nature 355, 719–722.

9
Hill, A., Ward, S., Deino, A. L., Curtis, G. & Drake, R.(1992c). Scientific correspondence: earliest Homodebate. Nature 358, 289–290.
Hill, A., Leakey, M., Kingston, J. D. & Ward, S. C.(2002). New cercopithecoids and a hominoid from12·5 Ma in the Tugen Hills succession, Kenya.J. hum. Evol. 1/2, 75–93.
Hooijer, D. A. (1969). Pleistocene East African rhi-noceroses. In (L. S. B. Leakey, Ed.) Fossil Vertebratesof Africa, Vol. 1, pp. 71–98. New York: AcademicPress.
Hooijer, D. A. (1973). Additional Miocene toPleistocene rhinoceroses of Africa. Zool. Meded. 46,149–180.
Hooijer, D. A. (1974). The hipparions of the BaringoBasin sequence. Nature 254, 211–212.
Jacobs, B. F. (1994). Paleoclimate reconstructionsusing middle to late Miocene paleofloras from centralKenya. Am. J. Botany 81(Supplement), 94.
Jacobs, B. F. & Deino, A. L. (1996). Test ofclimate-leaf physiognomy regression models, theirapplication to two Miocene floras from Kenya,and 40Ar/39Ar dating of the Late Miocene Kapturosite. Palaeogeogr., Palaeoclimatol., Palaeoecol. 123,259–271.
Jacobs, B. F. & Kabuye, C. H. S. (1987). A middleMiocene (12·2 my old) forest in the East African RiftValley, Kenya. J. hum. Evol. 16, 147–155.
Jacobs, B. F. & Winkler, D. A. (1992). Taphonomy ofa middle Miocene autochthonous forest assemblage,Ngorora Formation, central Kenya. Palaeogeogr.,Palaeoclimatol., Palaeoecol. 99, 31–40.
Kabuye, C. H. S. & Jacobs, B. F. (1986). An interestingrecord of the genus Leptaspis, Bambusoideae fromMiddle Miocene flora deposits in Kenya, East Africa.Abstracts: International Symposium on Grass Sys-tematics and Evolution, Washington DC: SmithsonianInstitution, 32.
Kelley, J., Ward, S., Brown, B., Hill, A. & Downs, W.(2000). Middle Miocene Hominoid Origins. Science287. http://www.sciencemag.org/cgi/content/full/287-5462/2375a.
Kelley, J., Ward, S., Brown, B., Hill, A. & Duren, D.(2002). Dental remains of Equatorius africanus fromKipsaramon, Tugen Hills, Baringo District, Kenya.J. hum. Evol. 1/2, 39–62.
King, B. C. & Chapman, G. R. (1972). Volcanism ofthe Kenya rift valley. Phil. Trans. R. Soc. London 271,185–208.
Kingston, J. D. (1992). Stable isotopic evidence forhominid paleoenvironments in East Africa. Ph.D.Dissertation, Harvard University.
Kingston, J. D. (1999). Environmental determinants inearly hominid evolution; issues and evidencefrom the Tugen Hills, Kenya. In (P. Andrews & P.Banham, Eds) Late Cenozoic Environments and Homi-nid Evolution: a Tribute to Bill Bishop, pp. 69–84.London: Geological Society of London.
Kingston, J. D., Hill, A. & Marino, B. D. (1992).Isotopic evidence of late Miocene/Pliocene veg-etation in the East African Rift Valley. Am. J. phys.Anthrop. Supplement 13, 100.
Kingston, J. D., Marino, B. D. & Hill, A. (1994).Isotopic evidence of late Miocene/Pliocene veg-etation in the East African Rift Valley. Science 264,955–959.
Kingston, J. D., Hill, A. & Deino, A. L. (2000).Evolution of East African hominid communitiesand high resolution Pliocene climatic change in thecentral Kenyan Rift. J. hum. Evol. 38, A15.
Kingston, J. D., Jacobs, B. F., Hill, A. & Deino, A. L.(2002). Stratigraphy, age and environments of thelate Miocene Mpesida Beds, Tugen Hills, Kenya.J. hum. Evol. 1/2, 95–116.
Leakey, L. S. B. & Solomon, J. D. (1929). East Africanarchaeology. Nature 124, 9.
Leakey, M., Tobias, P. V., Martyn, J. E. & Leakey,R. E. F. (1969). An Acheulian industry with pre-pared core technique and the discovery of a contem-porary hominid mandible at lake Baringo, Kenya.Proc. Prehist. Soc. 3, 48–76.
Leakey, R. E. F. (1969). New Cercopithecoideafrom the Chemeron Beds of Lake Baringo, Kenya.In (L. S. B. Leakey, Ed.) Fossil Vertebrates ofAfrica, Vol. 1, pp. 53–70. New York: AcademicPress.
Maglio, V. J. (1970). Early Elephantidae of Africa anda tentative correlation of African Plio-Pleistocenedeposits. Nature 225, 328–332.
Maglio, V. J. (1973). Origin and evolution of theElephantidae. Trans. Am. Phil. Soc. 63, 1–149.
Martyn, J. (1967). Pleistocene deposits and new fossillocalities in Kenya. Nature 215, 476–478.
Martyn, J. (1969). The geological history of the countrybetween Lake Baringo and the Kerio River, BaringoDistrict, Kenya. Ph.D. Dissertation, University ofLondon.
McBrearty, S. (1999). The archaeology of theKapthurin Formation. In (P. Andrews & P. Banham,Eds) Late Cenozoic Environments and Hominid Evol-ution: a Tribute to Bill Bishop, pp. 143–156. London:Geological Society of London.
McBrearty, S., Bishop, L. C. & Kingston, J. D. (1996).Variability in traces of middle Pleistocene hominidbehavior in the Kapthurin Formation, Baringo,Kenya. J. hum. Evol. 30, 563–580.
McBrearty, S., Brown, B., Deino, A. L., Kingston,J. D. & Ward, S. (1999). Anatomy, context, age andaffinities of hominids from the Kapthurin Formation,Kenya. J. hum. Evol. 36, A12.
McBrearty, S., Deino, A. L. & Tryon, C. (2001).Chronology of Middle Pleistocene events in theKapthurin Formation, Baringo, Kenya. J. hum. Evol.40, A14.
McCall, G. J. H., Baker, B. H. & Walsh, J. (1967). LateTertiary and Quaternary sediments of the Kenya RiftValley. In (W. W. Bishop & J. D. Clark, Eds)Background to Evolution in Africa, pp. 191–220.Chicago: Chicago University Press.
McClenaghan, M. P. (1971). Geology of the Ribkwoarea, Baringo District, Kenya. Ph.D. Dissertation,University of London.
Morgan, M. E., Kingston, J. D. & Marino, B. D.(1994). Carbon isotopic evidence for the emergence

10 .
of C4 plants in the Neogene from Pakistan andKenya. Nature 367, 162–165.
Namwamba, F. L. (1993). Tephrostratigraphy of theChemeron Formation, Baringo Basin, Kenya. M.Sc.Dissertation, University of Utah.
Pickford, M. (1975). Stratigraphy and paleoecology offive Late Cainozoic Formations in the Kenya RiftValley. Ph.D. Dissertation, University of London.
Pickford, M. (1978a). Geology, palaeoenvironmentsand vertebrate faunas of the mid-Miocene NgororaFormation. In (W. W. Bishop, Ed.) Geological Back-ground to Fossil Man, pp. 237–262. Edinburgh: Geo-logical Society of London & Scottish AcademicPress.
Pickford, M. (1978b). Stratigraphy and mammalianpaleontology of the late-Miocene Lukeino For-mation, Kenya. In (W. W. Bishop, Ed.) GeologicalBackground to Fossil Man, pp. 263–278. Edinburgh:Geological Society of London & Scottish AcademicPress.
Prior, G. T. (1903). Contributions to the petrology ofBritish East Africa. Mineralog. Mag. 13, 228–262.
Shackleton, R. M. (1951a). A review of some recentwork in the rift valleys of Kenya. 18th InternationalGeological Congress 14, 213.
Shackleton, R. M. (1951b). A contribution to thegeology of the Kavirondo Rift Valley. Q. J. Geol. Soc.London 106, 345–392.
Sherwood, R. J., Ward, S. & Hill, A. (1996). Mandibu-lar fossa anatomy of the Chemeron temporal bone(KNM-BC 1). Am. J. phys. Anthrop. Supplement 22,214–215.
Sherwood, R. J., Ward, S. & Hill, A. (2002a). Thetaxonomic status of the Chemeron temporal(KNM-BC 1). J. hum. Evol. 1/2, 153–184.
Sherwood, R. J., Ward, S., Hill, A., Duren, D., Brown,B. & Downs, W. (2002b). Preliminary description ofthe Equatorius africanus partial skeleton (KNM-TH28860) from Kipsaramon, Tugen Hills, BaringoDistrict, Kenya. J. hum. Evol. 1/2, 63–73.
Tallon, P. W. J. (1978). Geological setting of thehominid fossils and Acheulian artifacts from theKapthurin Formation, Baringo District, Kenya. In(W. W. Bishop, Ed.) Geological Background to FossilMan, pp. 361–373. Edinburgh: Geological Society ofLondon & Scottish Academic Press.
Tauxe, L., Monaghan, M., Drake, R., Curtis, G. &Staudigel, H. (1985). Paleomagnetism of MioceneEast African Rift sediments and the calibration of theGeomagnetic Reversal Time Scale. J. Geophys. Res.90, 4639–4646.
Thomas, H. (1980). Les bovides du Miocene superieurdes couches de Mpesida et de la formation deLukeino (district de Baringo, Kenya). In (R. Leakey& B. A. Ogot, Eds) Proceedings of the 8th Pan-AfricanCongress of Prehistory and Quaternary Studies,pp. 82–91. Nairobi: The International Louis LeakeyMemorial Institute for African Prehistory.
Thomas, H. (1981). Les Bovides Miocenes de laformation de Ngorora du Bassin de Baringo (RiftValley, Kenya). Proc. Kon. Ned. Akad. Wet. 84,335–410.
Thomson, J. (1884). Through Masailand. London:Samson Low.
Tobias, P. V. (1967). Pleistocene deposits and newfossil localities in Kenya. Nature 215, 478–480.
Tryon, C. & McBrearty, S. (2002). Tephrostratigraphyand the Acheulian to Middle Stone Age transition inthe Kapthurin Formation, Baringo, Kenya. J. hum.Evol. 1/2, 211–235.
Walsh, J. (1969). Geology of the EldamaRavine—Kabarnet area. Rep. Geol. Surv. Kenya 83.
Ward, S. & Hill, A. (1987). Pliocene hominid partialmandible from Tabarin, Baringo, Kenya. Am. J.phys. Anthrop. 72, 21–37.
Ward, S., Brown, B. & Hill, A. (1996). Forelimb ofKenyapithecus africanus from the Tugen Hills,Baringo District, Kenya. Am. J. phys. Anthrop.22(Supplement), 240.
Ward, S., Brown, B., Hill, A., Kelley, J. & Downs, W.(1999). Equatorius: a new hominoid genus from theMiddle Miocene of Kenya. Science 285, 1382–1386.
Willis, B. (1936). East African Plateaus and Rift Valleys.Washington: Carnegie Institution.
Winkler, A. J. (1990). Systematics and biostratigraphyof Neogene rodents from the Baringo District,Kenya. Ph.D. Dissertation, Southern MethodistUniversity.
Winkler, A. J. (1992). Systematics and biogeographyof middle Miocene rodents from the MuruyurBeds, Baringo District, Kenya. J. Vert. Paleontol. 12,236–249.
Winkler, A. J. (2002). Neogene paleobiogeography andeast African paleoenvironments: contributions fromthe Tugen Hills rodents and lagomorphs. J. hum.Evol. 1/2, 237–256.
Worthington, S. & Worthington, E. B. (1933). InlandWaters of Africa: the Result of Two Expeditions to theGreat Lakes of Kenya and Uganda, With Accountsof Their Biology, Native Tribes and Development.London: Macmillan.




















