
Ovarian features of the wild collared peccary ( Tayassu tajacu) from the northeastern Peruvian...
8
General and Comparative Endocrinology 147 (2006) 268–275 www.elsevier.com/locate/ygcen 0016-6480/$ - see front matter © 2006 Elsevier Inc. All rights reserved. doi:10.1016/j.ygcen.2006.01.010 Ovarian features of the wild collared peccary (Tayassu tajacu) from the northeastern Peruvian Amazon Pedro Mayor a , Mar Fenech a , Richard E. Bodmer b , Manel Lopez-Bejar a,¤ a Department of Animal Health and Anatomy, Faculty of Veterinary, Autonomous University of Barcelona, Bellaterra, E-08193 Barcelona, Spain b Durrell Institute of Conservation and Ecology, University of Kent, Canterbury, Kent CT2 7NS, UK Received 26 October 2005; revised 8 January 2006; accepted 18 January 2006 Available online 3 March 2006 Abstract In this study, the ovaries of 27 wild collared peccaries (Tayassu tajacu) from the Amazonian region of northeastern Peru were examined macroscopically and microscopically, and expression of major steroidogenic enzymes was detected by immunohistochemistry. Our obser- vations suggest a mean ovulation rate of 2.3 § 0.6 follicles and a low rate of reproductive wastage (0.4 § 0.6 oocytes or embryos per preg- nancy). The collared peccary seems to exhibit follicular waves involving the synchronous growth of a cohort of follicles, several of which seem to attain selection. The presence of antral follicles in pregnant females suggests that follicular turnover continues during pregnancy. In cyclic animals, corpora lutea were characterised by the presence of distinct large and small luteal cell populations. The luteal volume in pregnant females was larger than that recorded for non-pregnant females. Through immunohistochemistry, it was observed that luteal cells from active corpora lutea exhibit intensive 3-HSD expression in advanced stages of pregnancy. This suggests that the corpora lutea seems to remain steroidogenically active throughout pregnancy and likely contribute to progesterone production during pregnancy. © 2006 Elsevier Inc. All rights reserved. Keywords: Collared peccary; Tayassu tajacu; Ovary; Steroidogenic enzymes; Reproductive biology 1. Introduction In Amazon regions, wildlife hunting has been a traditional source of food, animal skins, and other essential items for local people, as an important subsistence activity (Fitz Gib- bon, 1998; Lavaigne et al., 1996). Over-hunting, however, has led to a situation in which mammalian species in forested habitats, particularly those of high commercial value, are becoming locally or even widely extinct (Emmons, 1990; Redford, 1993). This threat of extinction has meant that in tropical forest regions there is presently much interest in innovative systems capable of exploiting renewable natural resources, such as captive breeding programmes and commu- nity-based wildlife management programmes. The collared peccary (Tayassu tajacu) is an interesting candidate species for captive breeding, since it has always been considered to have a rapid growth rate similar to other Suiformes. The relatively rapid reproduction growth of the collared peccary as a result of its high fecundity also means that it is less vulnerable to over-hunting than slower repro- ducing species and better suited for sustainable wildlife use management systems (Bodmer et al., 1997). Although col- lared peccaries belong to a separate family within the Suifor- mes, the Tayassuidae, due to general similarities with the domestic pig (Sus scrofa), many data regarding their repro- ductive behaviour in the literature have been extrapolated from this species. The reproductive biology of the collared peccary inhabiting Amazon regions is poorly understood (Gottdenker and Bodmer, 1998). Previous studies have reported on the reproductive fea- tures of captive collared peccaries in southwest North America (Hellgren et al., 1995; Low, 1970; Sowls, 1984, 1997) but the anatomy of the ovaries has been the subject of few investigations. In 1931, Wislocki described two distinct types of luteal cells in the corpora lutea (CL) of a * Corresponding author. Fax: +34 93 581 20 06. E-mail address: [email protected] (M. Lopez-Bejar).
Transcript of Ovarian features of the wild collared peccary ( Tayassu tajacu) from the northeastern Peruvian...

General and Comparative Endocrinology 147 (2006) 268–275
www.elsevier.com/locate/ygcen
Ovarian features of the wild collared peccary (Tayassu tajacu) from the northeastern Peruvian Amazon
Pedro Mayor a, Mar Fenech a, Richard E. Bodmer b, Manel Lopez-Bejar a,¤
a Department of Animal Health and Anatomy, Faculty of Veterinary, Autonomous University of Barcelona, Bellaterra, E-08193 Barcelona, Spainb Durrell Institute of Conservation and Ecology, University of Kent, Canterbury, Kent CT2 7NS, UK
Received 26 October 2005; revised 8 January 2006; accepted 18 January 2006Available online 3 March 2006
Abstract
In this study, the ovaries of 27 wild collared peccaries (Tayassu tajacu) from the Amazonian region of northeastern Peru were examinedmacroscopically and microscopically, and expression of major steroidogenic enzymes was detected by immunohistochemistry. Our obser-vations suggest a mean ovulation rate of 2.3§ 0.6 follicles and a low rate of reproductive wastage (0.4§ 0.6 oocytes or embryos per preg-nancy). The collared peccary seems to exhibit follicular waves involving the synchronous growth of a cohort of follicles, several of whichseem to attain selection. The presence of antral follicles in pregnant females suggests that follicular turnover continues during pregnancy. Incyclic animals, corpora lutea were characterised by the presence of distinct large and small luteal cell populations. The luteal volume inpregnant females was larger than that recorded for non-pregnant females. Through immunohistochemistry, it was observed that luteal cellsfrom active corpora lutea exhibit intensive 3�-HSD expression in advanced stages of pregnancy. This suggests that the corpora lutea seemsto remain steroidogenically active throughout pregnancy and likely contribute to progesterone production during pregnancy.© 2006 Elsevier Inc. All rights reserved.
Keywords: Collared peccary; Tayassu tajacu; Ovary; Steroidogenic enzymes; Reproductive biology
1. Introduction
In Amazon regions, wildlife hunting has been a traditionalsource of food, animal skins, and other essential items forlocal people, as an important subsistence activity (Fitz Gib-bon, 1998; Lavaigne et al., 1996). Over-hunting, however, hasled to a situation in which mammalian species in forestedhabitats, particularly those of high commercial value, arebecoming locally or even widely extinct (Emmons, 1990;Redford, 1993). This threat of extinction has meant that intropical forest regions there is presently much interest ininnovative systems capable of exploiting renewable naturalresources, such as captive breeding programmes and commu-nity-based wildlife management programmes.
The collared peccary (Tayassu tajacu) is an interestingcandidate species for captive breeding, since it has always
* Corresponding author. Fax: +34 93 581 20 06.E-mail address: [email protected] (M. Lopez-Bejar).
0016-6480/$ - see front matter © 2006 Elsevier Inc. All rights reserved.doi:10.1016/j.ygcen.2006.01.010
been considered to have a rapid growth rate similar to otherSuiformes. The relatively rapid reproduction growth of thecollared peccary as a result of its high fecundity also meansthat it is less vulnerable to over-hunting than slower repro-ducing species and better suited for sustainable wildlife usemanagement systems (Bodmer et al., 1997). Although col-lared peccaries belong to a separate family within the Suifor-mes, the Tayassuidae, due to general similarities with thedomestic pig (Sus scrofa), many data regarding their repro-ductive behaviour in the literature have been extrapolatedfrom this species. The reproductive biology of the collaredpeccary inhabiting Amazon regions is poorly understood(Gottdenker and Bodmer, 1998).
Previous studies have reported on the reproductive fea-tures of captive collared peccaries in southwest NorthAmerica (Hellgren et al., 1995; Low, 1970; Sowls, 1984,1997) but the anatomy of the ovaries has been the subjectof few investigations. In 1931, Wislocki described twodistinct types of luteal cells in the corpora lutea (CL) of a

P. Mayor et al. / General and Comparative Endocrinology 147 (2006) 268–275 269
single pregnant collared peccary (Wislocki, 1931). Werecently described the anatomical and histological featuresof the tubular organs of the female genital tract of the col-lared peccary (Mayor et al., 2004a). Field studies in thenortheastern Peruvian Amazon have indicated that the col-lared peccary female can be described as an aseasonallypolyoestrous species (Gottdenker and Bodmer, 1998;Mayor et al., 2004a). Sowls (1997) and Mauget et al. (1997)reported an oestrous cycle length between 22.6 and 27.8days. Other studies have established a mean gestationperiod of 138 days (Mayor et al., 2004b), an ovulation rateof 2.0–2.1 ovulations/cycle (Gottdenker and Bodmer, 1998;Hellgren et al., 1995; Mayor et al., 2004a), and a litter sizeof 1.7–1.9 foetuses or newborns (Lochmiller et al., 1984;Gottdenker and Bodmer, 1998; Mayor et al., 2004a;Nogueira-Filho and Lavorenti, 1997).
EVective wildlife management depends on an under-standing of reproduction, which is the foundation on whicha species survives, thrives or becomes extinct (Pukazhenthiand Wildt, 2004). Reproductive studies beneWt the mainte-nance of viable captive populations (Swanson and Brown,2004) and are essential for understanding the dynamics ofwildlife populations (Greene et al., 1998). Indeed, there is adesperate need for this type of work to improve the numbersof more than 40,000 vertebrate species on the planet thathave gone unstudied (Pukazhenthi and Wildt, 2004). Thepresent study aims to contribute to this gap in the literatureas part of an ongoing project designed to establish severalbiological features of this small artiodactyl. Herein, wedescribe macroscopic and microscopic features and expres-sion of major steroidogenic enzymes detected by immuno-histochemistry in the ovaries of the wild collared peccary.
2. Methods
The study was conducted in the Tamshiyacu–Tahuayo CommunityReserve situated in northeast Peru between the Tamshiyacu, Tahuayo, andYavari Miri rivers. The reserve spans 322,500 ha of continuous forest andis predominantly non-Xooded terra Wrme forest (Bodmer et al., 1990). Thestudy area was in the Blanco river basin region of the reserve, which hasintensive hunting pressures.
2.1. Animals
Reproductive organs of 27 adult wild collared peccary females wereobtained from local hunters of the Blanco river basin between June 2000and August 2001, encompassing the wet and dry seasons. Hunters col-lected the samples as part of an ongoing participatory conservation pro-gramme that involves local hunters in implementing community-basedwildlife management. Whole body and genital organs weights wererecorded, and tissues were maintained in buVered 4% formaldehyde solu-tion (v/v) until analysis.
2.2. Reproductive status
The genital tracts of adult females were examined for evidence ofembryos or foetuses. The uterine tubes and uterine horns of females withovaries bearing corpora lutea but no obvious implantation sites wereXushed with an isotonic saline solution to conWrm the absence of ovulatedoocytes or embryos. Females with at least one embryo or foetus were con-sidered to be pregnant, and the pregnancy stage was deWned as embryonic
or foetal (Nomina Embryologica Veterinaria, 1994). Non-pregnant adultanimals with ovaries bearing large antral follicles and lacking corporalutea (CL) were considered to be in the follicular phase of the oestrouscycle, while those with ovaries containing luteal tissue were described to bein the luteal phase of the oestrous cycle. In the absence of either largeantral follicles or CL, the ovaries were considered inactive.
2.3. Macroscopic and microscopic features of the ovaries
The macroscopic ovarian features were recorded for all animals. Ovar-ian sizes (r1 D Length/2; r2 D Depth/2; and r3 D Width/2) were measuredand the volume estimated using a formula deWning the volume of an ovoidbody: V D 4/3 � (r1 r2 r3).
The ovaries were dehydrated, embedded in paraYn wax and cut. Threeto Wve micron sections were stained with haematoxylin and eosin, PAS–haematoxylin or Masson’s trichrome stain and examined under a lightmicroscope. Measurements of ovarian structures were made using amicrometric ocular device. Diameters were measured as the mean lengthof the two perpendicular axes. Luteal volumes were calculated using theformula: V D 4/3 (D/2)3 as described by Van Aarde and Skinner (1986).The luteal volume per female was calculated as the sum of the volumesfrom all luteal structures that appeared active histologically.
Follicles were classiWed using the optical plane through which theoocyte nucleolus was visible according to a modiWed classiWcation of Joriet al. (2002). Morphometric features of primordial-, primary-, small pre-antral-, large pre-antral-, small antral-, and large antral follicles wererecorded (see Table 2). The presence or absence of the zona pellucida andthecal layers was recorded for each type of follicle.
Corpora lutea were classiWed as cyclic CL (CL from non-pregnantfemales) or CL of pregnancy (CL from pregnant females). CL structureand constituent cell types were described. Corpora albicans were alsodescribed. The ovulation rate was estimated from the mean number ofactive CL per female.
2.4. Immunohistochemistry
Follicles at diVerent developmental stages and CL were immunohisto-chemically labelled to detect the expression of three major steroidogenicenzymes: cytochrome P450 17�-hydroxylase/17–20 lyase (P450c17),involved in androgen synthesis, cytochrome P450 aromatase, whichconverts androgens to oestrogen, and 3�-hydroxysteroid dehydrogenase(3�-HSD), involved in progesterone synthesis.
Immunohistochemistry was performed using a polyclonal rabbit anti-bovine P450c17 antibody (kindly supplied by Dr. Conley, University ofCalifornia, Davis, USA), a monoclonal mouse anti-human aromataseantibody (Serotec, Düsseldorf, Germany), and a polyclonal rabbit anti-human 3�-HSD antibody (kindly supplied by Dr. Van Luu-The,Université Laval, Quebec, Canada) in an avidin-biotin immunoperoxidaseprocedure. Tissue sections were placed on 3-aminopropyltriethoxysilane-coated (APES, Sigma Chemical, St. Louis, MO, USA) slides and, afterdewaxing and rehydration in a graded ethanol series, they were pre-incu-bated in 2% (v/v) hydrogen peroxide for 30 min to inhibit endogenous per-oxidase. After washing in PBS, the sections were immersed in 0.1%protease (Sigma) for 8 min at 37 °C. The sections were then incubated in10% (v/v) normal horse serum for 1 h at room temperature to avoid non-speciWc binding, and incubated with primary antibody used at dilutions of1:5000 (P450c17), 1:500 (aromatase), or 1:400 (3�-HSD) in PBS–BSA (1%w/v BSA, Sigma) overnight at room temperature. Negative control sec-tions were incubated with normal horse serum (1:200) in place of primaryantibody. After three washes in PBS for 5 min each, the sections were incu-bated with biotinylated second antibody (horse anti-mouse IgG andanti-rabbit IgG) for 1 h at room temperature and processed by the biotin–avidin method according to the manufacturer’s instructions (VectastainABC Kit, Vector Laboratories, Peterborough, UK). Peroxidase activitywas visualized by incubating the sections with 3, 3�-diaminobenzidine-4HCl in 0.05 mol Tris–HCl buVer l–1 (pH 7.6) containing 0.01% (v/v) H2O2for 1.5 min. The sections were counterstained with Mayer’s haematoxylinto correlate immunostaining with the morphology of the ovary.

270 P. Mayor et al. / General and Comparative Endocrinology 147 (2006) 268–275
2.5. Statistical analysis
The variables recorded for the 54 ovaries were ovarian size, reproduc-tive state, number and type of antral follicles, number and type of CL,luteal volume, and size of large and small luteal cells. Statistical analyseswere performed using Statistical Analysis System software (SAS, 1988).DiVerences were considered signiWcant when P < 0.05. Values areexpressed as the means § standard deviation (SD).
The eVect of the reproductive state of the animal on the number andtype of antral follicles, number and type of corpora lutea and luteal vol-ume, and size of large and small luteal cells was assessed by analysis of var-iance for unequal numbers. Means were tested for diVerences using theDuncan’s multiple range test.
3. Results
3.1. Reproductive state of the animals
Of the 27 adult females included in this study, 11 were non-pregnant and 16 were found to be at diVerent stages of preg-nancy. Of the 11 non-pregnant females, 5 were in the follicularstage and 6 were in the luteal stage of the oestrous cycle. Ofthe 16 pregnant animals, 15 had two embryos or foetuses yetonly 1 pregnant female had a single foetus. One pregnant pec-cary was described to be at the embryonic stage of pregnancy,with 2 embryos about 5mm in size. Of the 16 pregnantfemales, 10 were found to be pregnant in the wet season and 6in the dry season, including the female at embryonic stage ofpregnancy. No reproductive disorders were detected.
3.2. Ovarian features
The creamy white ovaries of sexually mature collaredpeccary females are ovoid bodies with a smooth surfacepositioned under the fourth lumbar vertebra in non-preg-nant females. They are enveloped by a bursa ovarica, whichis linked to the peritoneal cavity by a Xat 7.4§1.6 mmopening at the place of largest diameter. Table 1 shows thatthe mean ovarian volume was signiWcantly larger in preg-nant than in non-pregnant females at the follicular phase(P < 0.05), but not compared to non-pregnant females inthe luteal phase of the oestrous cycle. Right and left ovariesfailed to diVer signiWcantly in morphometric dimensions.
Growing follicles were visible as transparent bodies thatdid not protrude above the surface. Corpora lutea wereovoid bodies protruding from the surface (Fig. 1). A lowcuboidal surface epithelium covered the ovary. The tunica
albuginea was disrupted by the presence of ovarian folliclesand corpora lutea. Scattered smooth muscle Wbres appearedin the stroma around the follicles.
3.3. Follicular structures and follicle distribution
Table 2 shows the morphometric characteristics of thefollicles of the collared peccary.
Primordial follicles were found in the cortex just beneaththe tunica albuginea. A single layer of Xattened squamousfollicular cells, resting on a basement membrane, envelopedthe spherical oocyte.
Primary follicles were situated in the periphery of thecortex. Proliferation of the Xattened follicular cells sur-rounding the oocyte gave rise to their cuboidal shape.
Small pre-antral follicles were characterised by the pres-ence of 2–4 concentric layers of cuboidal cells. The incipientzona pellucida in the oocyte was 2.9§1.1 �m thick. Thetheca interna was slightly diVerentiated with 1–2 layers offusiform cells. The theca externa was diYcult to diVerenti-ate due its continuity with the stromal tissue.
Large pre-antral follicles had a granulosa membrane ofmore than 4 layers, separated from the oocyte by a zonapellucida 3.5§ 2.1 �m thick. The basement membranebetween the granulosa and theca interna appeared as adistinct boundary separating these layers.
Fig. 1. Ovary of a pregnant collared peccary (Tayassu tajacu) femaleshowing two CL of pregnancy (Bar: 4 cm).
Table 1Size and volume (mean § standard deviation in mm and mm3, respectively) of the ovaries of the collared peccary
r1 D Length/2; r2 D Depth/2; and r3 D Width/2.a,b Values in the same column with diVerent superscripts are signiWcantly diVerent (P < 0.05).
Reproductive status n Size (mm) Volume (mm3)
r1 r2 r3
Non-pregnant femalesFollicular stage 10 6.0§ 1.0 5.2 § 0.6a 5.0 § 1.0a 366.9§ 118.7a
Luteal stage 12 6.2§ 1.3 5.8 § 1.6b 5.2 § 1.5a,b 489.1§ 337.2a,b
Pregnant females 32 6.9§ 1.7 6.5 § 1.6b 6.2 § 1.5b 722.8§ 414.5b
P. Mayor et al. / General and Comparative Endocrinology 147 (2006) 268–275 271
Small antral follicles were characterised by a central cav-ity. As the stratum granulosa reached a thickness of 6–10layers (53.1 § 13.6�m in thickness), small Xuid-Wlled cavi-ties coalesced to form a follicular antrum which wasalready visible in follicles measuring 530 �m in diameter.Most of the granulosa cells lining the wall of the antrumwere polyhedral but the basal layer was columnar. Thecumulus oophorus projected into the antrum and sur-rounded the oocyte. The theca, attaining a thickness of56.0§17.1 �m, was clearly divided into the theca interna,deWned and formed by several layers of fusiform cells, andtheca externa, composed of Wbroblast-like cells, and fewlayers of elastic Wbres and vascular plexus. The cumulus celllayer was separated from the oocyte by a zona pellucida6.5§ 1.2�m thick.
Large antral follicles were of mean diameter2252.7§ 558.2�m, signiWcantly larger than the small antralfollicles (P < 0.01). However, mean oocyte plus zona pellu-cida size did not diVer signiWcantly between small and largeantral follicles. The follicular antrum was particularlyprominent. The stratum granulosum appeared to becomethinner as the antrum increased in size, its average thicknessbeing 82.5§ 35.5�m.
The average number of small and large antral folliclesper female at the follicular phase of the oestrous cycle was30.4§1.7 (nD 5). Mean numbers of small antral follicles,large antral follicles smaller than 2.5 mm, and large antralfollicles larger than 2.5 mm were 10.8§4.2, 14.4§3.0, and5.2§ 2.2, respectively. Small antral follicles were observedin ovaries of all the females. Groups of large antral follicleswere also detected in pregnant animals.
In primary and pre-antral atretic follicles, the oocyteappeared to degenerate prior to the follicle wall. In atreticantral follicles, the granulosa and thecal layers both foldinwards, hypertrophy and eventually occupy the antrum.Nuclear pyknosis and chromatolysis of cells provided clearevidence of cellular atresia. During atresia, the basementmembrane thickened and became hyalinised and the zonapellucida turned thicker and folded.
3.4. Interstitial glands
Thecal-type interstitial glands were detected in all thefemales examined and appeared to follow the disappear-ance of the stratum granulosum, hypertrophy of the thecainterna, and remnants of the zona pellucida or basementmembrane centrally.
3.5. Luteal structures
Corpora lutea at diVerent stages of development wereobserved in all the adult females. Table 3 shows the meandiameter and mean luteal volume observed in collared pec-cary females.
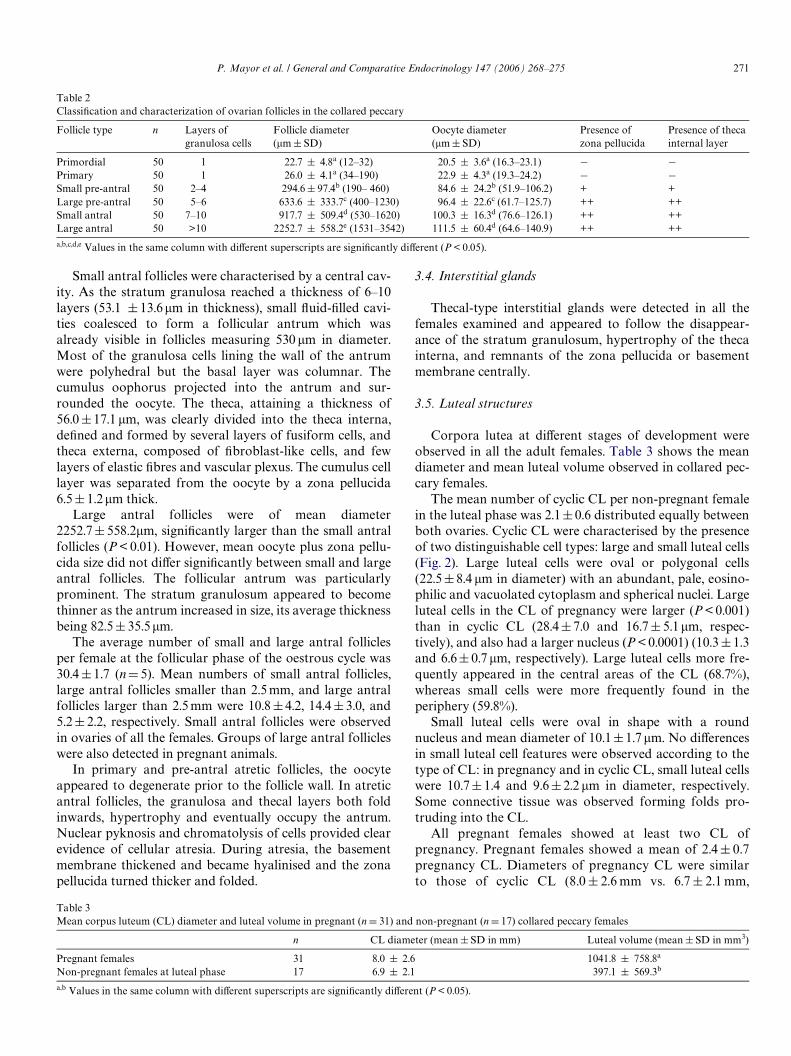
The mean number of cyclic CL per non-pregnant femalein the luteal phase was 2.1§0.6 distributed equally betweenboth ovaries. Cyclic CL were characterised by the presenceof two distinguishable cell types: large and small luteal cells(Fig. 2). Large luteal cells were oval or polygonal cells(22.5§8.4 �m in diameter) with an abundant, pale, eosino-philic and vacuolated cytoplasm and spherical nuclei. Largeluteal cells in the CL of pregnancy were larger (P < 0.001)than in cyclic CL (28.4§7.0 and 16.7§ 5.1�m, respec-tively), and also had a larger nucleus (P < 0.0001) (10.3§1.3and 6.6§ 0.7 �m, respectively). Large luteal cells more fre-quently appeared in the central areas of the CL (68.7%),whereas small cells were more frequently found in theperiphery (59.8%).
Small luteal cells were oval in shape with a roundnucleus and mean diameter of 10.1§1.7 �m. No diVerencesin small luteal cell features were observed according to thetype of CL: in pregnancy and in cyclic CL, small luteal cellswere 10.7§ 1.4 and 9.6§2.2 �m in diameter, respectively.Some connective tissue was observed forming folds pro-truding into the CL.
All pregnant females showed at least two CL ofpregnancy. Pregnant females showed a mean of 2.4§ 0.7pregnancy CL. Diameters of pregnancy CL were similarto those of cyclic CL (8.0§ 2.6 mm vs. 6.7§ 2.1 mm,
Table 2ClassiWcation and characterization of ovarian follicles in the collared peccary
a,b,c,d,e Values in the same column with diVerent superscripts are signiWcantly diVerent (P < 0.05).
Follicle type n Layers of granulosa cells
Follicle diameter (�m § SD)
Oocyte diameter (�m § SD)
Presence of zona pellucida
Presence of theca internal layer
Primordial 50 1 22.7 § 4.8a (12–32) 20.5 § 3.6a (16.3–23.1) ¡ ¡Primary 50 1 26.0 § 4.1a (34–190) 22.9 § 4.3a (19.3–24.2) ¡ ¡Small pre-antral 50 2–4 294.6§ 97.4b (190– 460) 84.6 § 24.2b (51.9–106.2) + +Large pre-antral 50 5–6 633.6 § 333.7c (400–1230) 96.4 § 22.6c (61.7–125.7) ++ ++Small antral 50 7–10 917.7 § 509.4d (530–1620) 100.3 § 16.3d (76.6–126.1) ++ ++Large antral 50 >10 2252.7 § 558.2e (1531–3542) 111.5 § 60.4d (64.6–140.9) ++ ++
Table 3Mean corpus luteum (CL) diameter and luteal volume in pregnant (n D 31) and non-pregnant (n D 17) collared peccary females
a,b Values in the same column with diVerent superscripts are signiWcantly diVerent (P < 0.05).
n CL diameter (mean § SD in mm) Luteal volume (mean § SD in mm3)
Pregnant females 31 8.0 § 2.6 1041.8 § 758.8a
Non-pregnant females at luteal phase 17 6.9 § 2.1 397.1 § 569.3b
272 P. Mayor et al. / General and Comparative Endocrinology 147 (2006) 268–275
respectively). The CL of females in advanced stages ofpregnancy showed a more intense, yellow-brownpigmentation.
Pregnant females had a signiWcantly greater averageluteal volume than non-pregnant cyclic females (P < 0.01;Table 3).
Corpora albicans were occasionally found in thefemales. Only one pregnant female showed two corporaalbicans besides pregnancy CL. Corpora albicans exhibitedextensive Wbrosis, markedly reduced vascularization com-pared to active CL and nuclear pyknosis.
Fig. 2. Detail of a pregnancy CL in a collared peccary showing large andsmall (arrows) luteal cells. Haematoxylin and eosin staining. Scale barrepresents 25 �m.
3.6. Ovulation rate
According to the number of CL observed in pregnantand cyclic females, the mean ovulation rate was estimatedto be 2.3§0.6 corpora lutea/female (nD16). Of the 16 preg-nant and cyclic females, 7 females ovulated unilaterally (4females on the right ovary and 3 females on the left ovary).There was no signiWcant diVerence in the mean number ofCL between the left and right ovary (1.1§ 0.7 vs. 1.2§ 0.7corpora lutea/female, respectively). The numbers of CLfound in pregnant and non-pregnant females did not varysigniWcantly. The average litter size for pregnant femaleswas 1.9§0.3 embryos or foetuses/pregnant female (nD 16).
3.7. Expression of steroidogenic enzymes
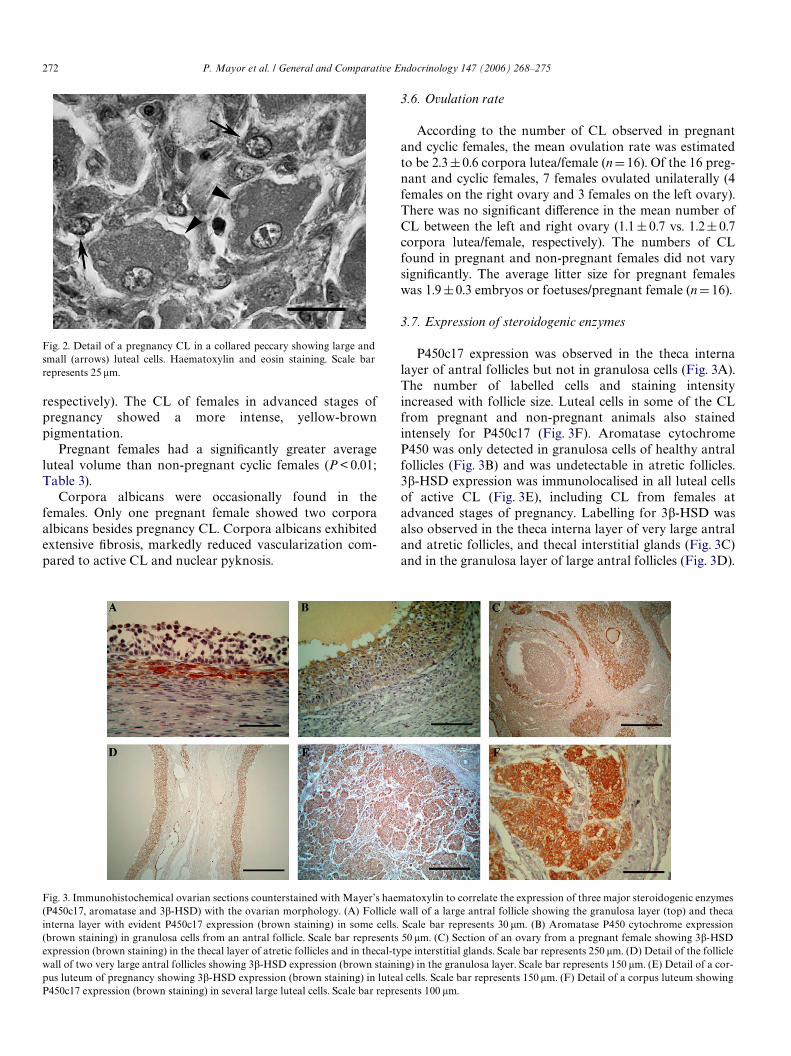
P450c17 expression was observed in the theca internalayer of antral follicles but not in granulosa cells (Fig. 3A).The number of labelled cells and staining intensityincreased with follicle size. Luteal cells in some of the CLfrom pregnant and non-pregnant animals also stainedintensely for P450c17 (Fig. 3F). Aromatase cytochromeP450 was only detected in granulosa cells of healthy antralfollicles (Fig. 3B) and was undetectable in atretic follicles.3�-HSD expression was immunolocalised in all luteal cellsof active CL (Fig. 3E), including CL from females atadvanced stages of pregnancy. Labelling for 3�-HSD wasalso observed in the theca interna layer of very large antraland atretic follicles, and thecal interstitial glands (Fig. 3C)and in the granulosa layer of large antral follicles (Fig. 3D).
Fig. 3. Immunohistochemical ovarian sections counterstained with Mayer’s haematoxylin to correlate the expression of three major steroidogenic enzymes(P450c17, aromatase and 3�-HSD) with the ovarian morphology. (A) Follicle wall of a large antral follicle showing the granulosa layer (top) and thecainterna layer with evident P450c17 expression (brown staining) in some cells. Scale bar represents 30 �m. (B) Aromatase P450 cytochrome expression(brown staining) in granulosa cells from an antral follicle. Scale bar represents 50 �m. (C) Section of an ovary from a pregnant female showing 3�-HSDexpression (brown staining) in the thecal layer of atretic follicles and in thecal-type interstitial glands. Scale bar represents 250 �m. (D) Detail of the folliclewall of two very large antral follicles showing 3�-HSD expression (brown staining) in the granulosa layer. Scale bar represents 150 �m. (E) Detail of a cor-pus luteum of pregnancy showing 3�-HSD expression (brown staining) in luteal cells. Scale bar represents 150 �m. (F) Detail of a corpus luteum showing
P450c17 expression (brown staining) in several large luteal cells. Scale bar represents 100 �m.
P. Mayor et al. / General and Comparative Endocrinology 147 (2006) 268–275 273
Control sections were negatively stained except for thecounterstaining of nuclei with haematoxylin.
4. Discussion
In this study, we characterise the reproductive physiol-ogy of the wild collared peccary through the description ofmacroscopic and microscopic features of the ovaries. Theovaries of the collared peccary female are enclosed by alarge bursa ovarica that completely surrounds the ovary.Contrary to observations in the sow (Dyce et al., 1987;Getty, 1982; Schwarze, 1984), growing follicles did not pro-trude above the ovary surface, but were visible as transpar-ent bodies. The constituent cell types of the ovary of thecollared peccary were found to be similar to those describedfor other Suidae (Mossman and Duke, 1973). The cortexshows many follicles at various stages of development andcorpora lutea.
Collared peccary females at the follicular phase of theoestrous cycle had average of 30.4 antral follicles. Amongthe large antral follicles, 14.4 and 5.2 follicles were smallerand larger than 2.5 mm, respectively, while the mean num-ber of CL observed per female indicates a mean ovulationrate of 2.3§0.6 follicles. This suggests there are follicularwaves involving the synchronous growth of a cohort of fol-licles, from which several seem to be selected to continuedevelopment. The low mean ovulation rate of the collaredpeccary could act as a biological limit for the proliWcacy ofthis Suiform. The domestic pig has a mean ovulation rate of10–24 follicles (Gordon, 1997) while the rate in the wild pigis 4–6 follicles (Mauget, 1982). On the other hand, the preg-nant collared peccary female had a low rate of reproductivewastage, averaging 0.4§ 0.6 oocytes or embryos per preg-nancy, based on the diVerence between the number of trueCL, and observed embryos or foetuses (Van Aarde andSkinner, 1986). This eYciency could be considered an opti-mal reproductive feature, compared to the 40% wastage perpregnancy reported for the sow (Pomeroy, 1992) and the12.5% for the wild boar (Mauget, 1982).
Corpora lutea were larger in the pregnant than in thecyclic collared peccary female. As in the domestic pig (Mur-phy, 2000), there are two distinct populations of luteal cells:small and large luteal cells. Also similar to the situation inthe pig (Priedkalns, 1987), small luteal cells comprise asmall proportion of the luteal tissue and mainly occupy tra-becular and periphery of the corpus luteum. The meanluteal volume calculated for the collared peccary wasgreater in pregnant than non-pregnant animals. Due to thelarge volume occupied, CL protruded from the smoothouter surface of the ovary, giving rise to a higher meanovarian volume in pregnant females. This increased volumeof CL in pregnancy could in part be the result of hypertro-phy of large luteal cells. During gestation, large luteal cellsprobably increase their production of steroid hormones,mainly progesterone. In mammals, progesterone is respon-sible for the development and secretion of uterine glands,such that the endometrium becomes receptive to the
implantation of the embryo, and promotes behaviourappropriate of pregnancy (Priedkalns, 1987).
Corpora albicans were found to occur alongside func-tional CL in the ovaries of adult females. As reported byMossman and Duke (1973), this could suggest that regres-sion of the functional CL is slow and that CL can persistinto the following gestation.
Large antral follicles were found regularly in the ovariesof pregnant females. This Wnding suggests that, as in the pigand cow (Guilbault et al., 1986; Pierson and Ginther, 1986;Rexroad and Casida, 1975; Taylor and Rajamahendran,1991), follicular growth continues throughout pregnancy incollared peccaries.
The pregnancies detected in our study population wereevenly distributed across wet and dry seasons. In agreementwith the Wndings of Gottdenker and Bodmer (1998), thissuggests that the collared peccary apparently breeds yearround in the NE Peruvian Amazon. In contrast, Sowls(1984) and Hellgren et al. (1995) reported that the collaredpeccary usually gives birth during the rainy season inTexas. This diVerent behaviour could be attributable to thelack of huge seasonal diVerences in resources in the tropicalenvironment, such that there is a suYcient food supply forpeccaries to sustain a pregnancy regardless of the month ofthe year (Gottdenker and Bodmer, 1998).
Our immunohistochemical data revealed that in antralfollicles, the theca interna layer expresses cytochromeP450c17, the intensity of the expression pattern increasingwith follicle size. Thus, the theca cells provide granulosacells with androgen substrate, as described for the domesticpig (Shores and Hunter, 1999; Tonetta and Hernandez,1989). In the peccary, P450 aromatase expression could notbe detected in theca interna cells, although it has beenreported in pig antral follicles (Conley et al., 1994; Corbinet al., 2003; Lautincik et al., 1994; Shores and Hunter,1999). The thecal cells of species other than the pig, such ascattle or sheep, do not express P450 aromatase (Lautinciket al., 1994). On the other hand, as in the pig (Conley et al.,1994; Garrett and Guthrie, 1997), 3�-HSD expression wasdetected in the theca interna of large antral follicles, con-Wrming the regulatory role of theca cells in steroidogenesisin the collared peccary.
The granulosa cells of the antral follicles were found toexpress cytochrome P450 aromatase with an intensityincreasing with follicle size, as occurs in the pig (Conleyet al., 1994). In addition, the granulosa layer in very largeantral follicles begin to express 3�-HSD, as reported for thecow (Conley et al., 1995) but not the pig (Conley et al.,1994, 1995).
All luteal cells in active CL expressed 3�-HSD intensely,and this expression persisted in CL obtained from femalesat advanced stages of pregnancy, suggesting that the CL ofthe collared peccary continue to be active during a largepart of pregnancy, consistent with Wndings in the pig(Moeljono et al., 1977; Perry et al., 1976). Luteal cells in thepeccary showed no aromatase expression, as described forpig CL by mRNA analysis (Guthrie et al., 1994;

274 P. Mayor et al. / General and Comparative Endocrinology 147 (2006) 268–275
Slomczynska et al., 2001). Contrary to the pig (Guthrieet al., 1994), luteal cells from some CL of the collared pec-cary expressed P450c17 intensely, suggesting the produc-tion of androgen substrate by CL in the collared peccary.
Thecal layers in atretic follicles and thecal-type intersti-tial glands showed 3�-HSD expression, pointing to steroi-dogenic activity in these ovarian structures.
The expression of major steroidogenic enzymes in folli-cles, corpora lutea, and interstitial structures of the collaredpeccary showed similarities but also subtle diVerences withrespect to the domestic pig. Further work is needed toestablish the expression proWles of steroidogenic enzymesduring pregnancy and the diVerent phases of the oestrouscycle.
In summary, the present study describes ovarian featuresof the wild collared peccary that suggest that folliculargrowth involves the synchronous growth of groups of folli-cles in waves, which continues in pregnant females. In thefollicle, theca cells have a steroidogenic regulating role.Given their mean ovulation rate and proliWcacy, the rate ofreproductive wastage is extremely low. Our observationsalso suggest that in this small mammal, corpora lutearemain active during much of pregnancy.
Acknowledgments
The authors thank Dr. Alan J. Conley for his helpfulcomments regarding the manuscript, Pablo Puertas andEtersit Pezo for help with the sample collection and theI.V.I.T.A- Iquitos, Peru, for data collection. This study wassupported by the European Union (Research for the Devel-opment, INCO-DEV; Fifth Framework Programme; con-tract ICA4-CT-2001-10045) and Spanish Ministry ofScience and Technology (MCYT; AGL2001-4961-E).
References
Bodmer, R.E., Eisenberg, J.F., Redford, K.H., 1997. Hunting and the likeli-hood of extinction of Amazonian mammals. Conserv. Biol. 11, 460–466.
Bodmer, R.E., Penn, J.W., Fang, T.G., Moya, L., 1990. Management pro-grammes and protected areas—the case of the Reserva Comunal Tam-shiyacu–Tahuayo, Peru. Parks 1, 21–25.
Conley, A.J., Howard, H.J., Slanger, W.D., Ford, J.J., 1994. Steroidogenesisin the preovulatory porcine follicle. Biol. Reprod. 51, 655–661.
Conley, A.J., Kaminski, M.A., Dubowsky, S.A., Jablonka-ShariV, A., Red-mer, D.A., Reynolds, L.P., 1995. Immunohistochemical localization of3 beta-hydroxysteroid dehydrogenase and P450 17 alpha-hydroxylaseduring follicular and luteal development in pigs, sheep, and cows. Biol.Reprod. 52, 1081–1094.
Corbin, C.J., Moran, F.M., Vidal, J.D., Ford, J.J., Wise, T., Mapes, S.M.,Njar, V.C., Brodie, A.M., Conley, A.J., 2003. Biochemical assessment oflimits to estrogen synthesis in porcine follicles. Biol. Reprod. 69, 390–397.
Dyce, K.M., Sack, W.O., Wensing, C.J.G., 1987. The pelvis and reproduc-tive organs of the pig. In: Dyce, K.M., Sack, W.O., Wensing, C.J.G.(Eds.), Textbook of Veterinary Anatomy. W.B. Saunders Company,Philadelphia, pp. 759–767.
Emmons, L.H., 1990. Neotropical Rainforest Mammals: A Field Guide.University of Chicago Press, Chicago IL.
Fitz Gibbon, C., 1998. The management of subsistence harvesting: behav-ioural ecology of hunter and their mammalian prey. In: Caro, T. (Ed.),
Behavioural Ecology and Conservation Biology. Oxford UniversityPress, Oxford, pp. 449–474.
Garrett, W.M., Guthrie, H.D., 1997. Steroidogenic enzyme expressionduring preovulatory follicle maturation in pigs. Biol. Reprod. 56,1424–1431.
Getty, R., 1982. The Anatomy of the Domestic Animals. W.B. SaundersCompany, Philadelphia PA.
Gordon, I., 1997. Controlled Reproduction in Pigs. CAB International,New York NY.
Gottdenker, N., Bodmer, R., 1998. Reproduction and productivity ofwhite-lipped and collared peccaries in the Peruvian Amazon. J. Zool.245, 423–430.
Greene, C., Umbanhowar, J., Mangel, M., Caro, T., 1998. Animal breedingsystems, hunter selectivity, and consumptive use in wildlife conserva-tion. In: Caro, T. (Ed.), Behavioural Ecology and Conservation Biol-ogy. Oxford University Press, Oxford, pp. 271–274.
Guilbault, L.A., Dufor, J.J., Thatcher, W.W., Drost, M., Haibel, G.K., 1986.Ovarian follicular development during early pregnancy in cattle. J.Reprod. Fertil. 78, 127–135.
Guthrie, H.D., Barber, J.A., Leighton, J.K., Hammond, J.M., 1994. Steroi-dogenic cytochrome P450 enzyme messenger ribonucleic acids and fol-licular Xuid steroids in individual follicles during preovulatorymaturation in the pig. Biol. Reprod. 51, 465–471.
Hellgren, E.C., Syntzke, D.R., Oldelberg, P.W., Guthery, F.S., 1995.Demography of a collared peccary population in South Texas. J. Wild-life Manag. 59, 153–163.
Jori, F., López-Béjar, M., Mayor, P., López, C., 2002. Functional anatomyof the ovaries of the wild brush-tailed porcupine (Atherurus africanus,Gray 1842) from Gabon. J. Zool. 256, 35–43.
Lautincik, J., Kolodzieyski, L., Elias, V., Hyttel, P., Osawa, Y., Sirotkin, A.,1994. Immunocytochemical localization of aromatase in the ovary ofsuperovulated cattle, pigs and sheep. Acta Vet. Scand. 35, 185–191.
Lavaigne, D.M., Callaghan, C.J., Smith, R.J., 1996. Sustainable utilization:the lessons of history. In: Taylor, V.J., Dunstone, N. (Eds.), TheExploitation of Mammals Populations. Chapman and Hall, London,pp. 250–265.
Lochmiller, R.L., Hellgren, E.C., Grant, W.E., 1984. Selected aspects of col-lared peccary Dycotyles tajacu reproductive biology in a captive Texasherd. Zoo. Biol. 3, 145–149.
Low, W.A., 1970. The InXuence of Aridity on Reproduction of the Col-lared Peccary (Dicotyles tajacu (Linn)) in Texas. Ph.D. Thesis. Univer-sity of British Columbia, BC.
Mauget, R., 1982. Seasonality of reproduction in the wild boar. In: Cole,D.J.A., Foxcroft, G.R. (Eds.), Control of Pig Reproduction. Butter-worth ScientiWc, London.
Mauget, R., Feer, F., Henry, O., Dubost, G., 1997. Hormonal and behav-ioural monitoring of ovarian cycles in peccaries. Proceedings of theFirst International Symposium on Physiology and Ethology of Wildand Zoo Animals II, 145–149.
Mayor, P., Jori, F., Lopez-Bejar, M., 2004a. Anatomicohistological charac-teristics of the tubular genital organs of the female collared peccary(Tayassu tajacu) from Northeastern Amazon. Anat. Histol. Embryol.33, 65–74.
Mayor, P., López-Gatius, F., López-Béjar, M., 2004b. Integrating ultraso-nography within the reproductive management of the collared peccary(Tayassu tajacu). Theriogenology 63, 1832–1843.
Moeljono, M.P.E., Thatcher, W.W., Bazer, F.W., 1977. A study of prosta-glandin F2� as the luteolysin in swine. II. Characterization and com-parison of prostaglandin F2�, estrogens and progestin concentrationsin utero-ovarian vein plasma of nonpregnant and pregnant gilts. Pros-taglandins 14, 543–555.
Mossman, H.W., Duke, K.L., 1973. Comparative Morphology of theMammalian Ovary. University of Wisconsin Press, Madison.
Murphy, B.D., 2000. Models of luteinization. Biol. Reprod. 63, 2–11.Nogueira-Filho, S.L., Lavorenti, A., 1997. O Manejo do caititu (Tayassu
Tajacu) e do queixada (Tayassu Pecari) em cativeiro. In: Manejo eConservação de Vida Silvestre no BrasilMamirauá-CNPq, Belém/PA-Brasília/DF, pp. 2851.

P. Mayor et al. / General and Comparative Endocrinology 147 (2006) 268–275 275
Nomina Embryologica Veterinaria, 1994. International Committee onVeterinary Embryological Nomenclature, Ithaca, NY.
Perry, J.S., Heap, R.D.B., Burton, R.D., 1976. Endocrinology of the blasto-cyst and its role in the establishment of pregnancy. J. Reprod. Fertil.25, 85–104.
Pierson, R.A., Ginther, O.J., 1986. Ovarian follicular populations duringearly pregnancy in heifers. Theriogenology 26, 649.
Pomeroy, R.W., 1992. Embryogenesis recapitulates oogenesis in swine.Proc. Soc. Exp. Biol. Med. 199, 273–281.
Priedkalns, J., 1987. Female reproductive system. In: Dellmann, H.-D.,Brown, E.M. (Eds.), Textbook of Veterinary Histology. Lea and Febi-ger, Philadelphia, pp. 313–339.
Pukazhenthi, B.S., Wildt, D.E., 2004. Which reproductive technologies aremost relevant to studying, managing, and conserving wildlife? Reprod.Fertil. Dev. 16, 1–14.
Rexroad, C.E., Casida, L.E., 1975. Ovarian follicular development in cows,sows, and ewes in diVerent stages of pregnancy as aVected by numberof corpora lutea in the same ovary. J. Anim. Sci. 41, 1090–1097.
Redford, K.H., 1993. Hunting in neotropical forests: a subsidy fromnature. In: Hladik, C.M., Hladik, A., Linares, O.F., Pagezy, H., Semple,A., Hadley, M. (Eds.), Tropical Forests, People, and Food: BioculturalInteractions and Applications to Development. The Parthenon PubGroup, Paris, pp. 227–246.
Schwarze, C., 1984. Compendio de Anatomía Veterinaria. Acribia, Zara-goza. pp. 277–289.
Shores, E.M., Hunter, M.G., 1999. Immunohistochemical localization ofsteroidogenic enzymes and comparison with hormone production dur-ing follicle development in the pig. Reprod. Fertil. Dev. 11, 337–344.
Slomczynska, M., Duda, M., Slzak, K., 2001. The expression of androgenreceptor, cytochrome P450 aromatase and FSH receptor mRNA in theporcine ovary. Folia Histochem. Cytobiol. 39, 9–13.
Sowls, L.K., 1984. The Peccaries. The University of Arizona Press, Tucson.Sowls, L.K., 1997. Javelines and Other Peccaries: The Biology, Manage-
ment and Use. College Station, Texas A. and M. University Press,Texas. pp. 84–106.
Swanson, W.F., Brown, J.L., 2004. International training programs inreproductive sciences for conservation of Latin America Welds. Anim.Reprod. Sci. 82-83, 21–34.
Taylor, C., Rajamahendran, R., 1991. Follicular dynamics and corpus lut-eum growth and function in pregnant versus nonpregnant dairy cows.J. Dairy Sci. 74, 115–123.
Tonetta, S.A., Hernandez, M., 1989. Modulation of 17 alpha-hydroxylase/C17,20-lyase activity in porcine theca cells. J. Steroid Biochem. Mol.Biol. 33, 263–270.
Van Aarde, R.J., Skinner, J.D., 1986. Functional anatomy of the ovaries ofpregnant and lactating cap porcupines Hystrix africaustralis. J.Reprod. Fertil. 76, 553–559.
Wislocki, G.B., 1931. Notes on the female reproductive tract (ovaries,uterus and placenta) of the collared peccary (Pecari angulatus bangsiGoldman). J. Mammal. 121, 143–149.












