
Seasonal spatial patterns of two sympatric frogs: California Red-legged Frog and American Bullfrog
Upload
independentCategory
view
1download
0
ORIGINAL ARTICLE
Sympatric amphisbaenids from Manso Dam region, Mato Grosso State, Western Brazil, with thedescription of a new two-pored species of Amphisbaena (Squamata, Amphisbaenidae)
Christine Strussmanna* and Tamı Mottb
aDepartamento de Ciencias Basicas e Producao Animal, Faculdade de Agronomia e Medicina Veterinaria, Universidade Federal
de Mato Grosso, Cuiaba, Mato Grosso, Brazil; bPrograma de Pos Graduacao em Ecologia e Conservacao da Biodiversidade,
Universidade Federal de Mato Grosso, Cuiaba, Bolsista PRODUC CAPES, Brazil
(Received 14 July 2004; accepted 14 November 2008)
A new species in the genus Amphisbaena is described based on a single specimen road killed on MT 351 in theregion under the influence of Manso Dam, at Chapada dos Guimaraes municipality, state of Mato Grosso,Brazil. It is a small amphisbaenid with two precloacal pores, 139 body annuli, 18/14 segments on the midbodyand an immaculate white belly. A checklist of the 25 species of amphisbaenids recorded for the Cerrado biome ispresented. Although ecological and/or historical features might be the reason for the unusual amphisbaenidrichness of eight sympatric species at Manso Dam region, the extensive collecting efforts at this site seem to offera better explanation for finding eight sympatric species.
Uma nova especie de Amphisbaena e descrita com base em um exemplar coletado na rodovia MT 351, regiao deinfluencia da usina hidreletrica de Manso, Chapada dos Guimaraes, estado de Mato Grosso, Brasil. A novaespecie distingue-se de todos os demais anfisbenıdeos que possuem dois poros pre-cloacais pelo menor numero deaneis corporais (139), 18 escamas dorsais e 14 escamas ventrais em um anel no meio do corpo e ventre imaculado.E apresentada uma lista dos anfisbenıdeos registrados no Cerrado, da qual constam 25 especies. Embora fatoresecologicos e/ou historicos possam ser empregados para explicar a elevada riqueza de anfisbenıdeos na regiao doreservatorio de Manso, o intenso esforco amostral naquela area parece melhor justificar o registro de oitoespecies simpatricas.
Keywords: Amphisbaenidae; Amphisbaena brevis sp. n.; new species; sympatry; Cerrado; Mato Grosso
Introduction
Amphisbaenians, or worm lizards, constitute an
array of approximately 190 species of highly specia-
lized fossorial squamates (Gans 2005). They comprise
a monophyletic group (Macey et al. 2004; Kearney &
Stuart 2004; Townsend et al. 2004), distributed over
all continents except Australia and Antarctica. Six
families are recognized in the suborder Amphisbaenia
(Vidal et al. 2008), but only the most speciose family,
Amphisbaenidae, occurs in Brazil, for which at least
57 species in six genera are listed (Gans 2005).
Amphisbaenids are widely distributed in Brazil,
and approximately 50% of the species recorded for
this country are known to occur in the Cerrado. That
region is Brazil’s second largest biome, covering
approximately 2 million km2 in the Central Brazilian
Plateau. Ancientness and the mosaic nature of
ecological conditions in the Cerrado may have
contributed to the development of a diverse herpeto-
fauna, and to a high level of local turnover of species,
at least for lizards (Araujo 1994). Due to biological
richness and high level of menace to this biodiversity,
the whole biome is being treated as a hotspot (see
Myers et al. 2001; Conservation International 2005).
In the last three decades, owing to expanding
agriculture and cattle ranching, the original Cerrado
area was reduced by more than 70% (Alho & Martins1995; Conservation International 2005).
Construction of new hydroelectric power plants in
Cerrado also has been responsible for the loss of huge
extensions of natural habitats, otherwise providing
unique opportunities to locate fossorial fauna,
because underground animals emerge from the soil
following such profound disturbances as soil rooting
and flooding. While conducting herpetofaunal inven-tories in the region of the Manso hydroelectric power
plant in mid-southern Mato Grosso State (western
Brazil), we found an unusual number of sympatric
species of amphisbaenids, one of them still unde-
scribed. Here, we present data on the local amphis-
baenid species composition and provide a description
of the new taxon. Where appropriate, range exten-
sions and meristic data are presented and discussed.
*Corresponding author. Email: [email protected]
Studies on Neotropical Fauna and Environment
Vol. 44, No. 1, April 2009, 37–46
ISSN 0165-0521 print/ISSN 1744-5140 online
# 2009 Taylor & Francis
DOI: 10.1080/01650520802628295
http://www.informaworld.com
Downloaded By: [University of Sao Paulo] At: 17:08 13 May 2009

Materials and methods
Study area
The reservoir behind Manso Dam was created by the
inundation of about 428 km2 of diverse Cerrado
physiognomies from the confluence of Casca and
Manso rivers (approximately at 14u529S, 55u489W)
upstream. The Manso River is the main contributor
to the Cuiaba River, a major tributary of the left
bank of the Paraguay River, which forms the main
axis of the Paraguay River Basin. The climate is
generally hot and semi-humid, with a well-marked
seasonality (Nimer 1979). Mean annual temperature
is around 26uC, with the coldest month being July,
and the hottest, October. The rainy season occurs
during the summer, from October/November to
April/May. Total annual precipitation is, on average,
around 1350 mm. There are 4–5 months of drought
(May to September), and from July to September
relative air humidity may drop to less than 30% (Alho
2000).
The ‘‘region under the influence of the Manso
Dam’’ referred to in this paper is the whole area now
inundated by the reservoir, plus a buffering zone of
10 km along the margins (area of direct influence),
and a few collecting sites situated within 30 km from
those margins (sections of the paved road MT 351
and of the unpaved road to Agua Fria, see Figure 1).
We particularly and repeatedly sampled habitats on
both margins of the Casca River, from its confluence
with the Manso up to the mouth of the Quilombo
River, a major tributary of the Casca (Figure 1).
Fieldwork was mostly done in areas of Fazenda
Morro do Chapeu, Fazenda Jopejo, and in the
neighborhood of Manso facilities, from August
1998 to March 2002.
Sampling of amphisbaenids
Specimens were collected opportunistically in visual
encounter surveys, pitfall traps (without drift fences)
and while following bulldozers during biological
inventories and monitoring activities in the area to
be inundated by the reservoir of the Manso, and also
during the filling of the reservoir. All the specimens
mentioned in the text are deposited at the Vertebrate
Collection of the Federal University of Mato Grosso,
Cuiaba, Brazil (UFMT collection), except for the
holotype of the new species, which is housed at the
State University of Campinas (UNICAMP, Campinas,
Sao Paulo, Brazil; ZUEC collection). Museum acro-
nyms other than UFMT follow Leviton et al. (1985).
Counts of annuli were made along the left side of
the ventral surface (except when otherwise indicated),
according to Gans & Alexander (1962). We also
followed these authors when describing the pholido-
sis of the new species. Species in Tables 1 and 2 are
arranged as in Vanzolini (2002a), by ascending order
of the number of precloacal pores.
Results
Amphisbaena brevis sp. n.
(Figures 2A–D, 3A–E)
Holotype
Male specimen (ZUEC 2834), road killed near km 63
of the paved road MT 351 (14u579220S, 55u529000W),
70 km north of Cuiaba, municipality of Chapada dos
Guimaraes, state of Mato Grosso, Brazil, collected
on 16 December 2001 by Christine Strussmann.
Etymology
The specific name is derived from the Latin word
brevis, meaning short. It refers to the general
appearance of the holotype, and addresses the most
remarkable trait of this two-pored species: the lowest
number of body annuli among all species presently
known in the genus Amphisbaena.
Diagnosis
Among the two-pored species of Amphisbaena, A.
brevis sp. n. has only 139 body annuli, while A. dubia
Muller, 1924, A. mitchelli Procter, 1923, A. leeseri
Gans, 1964, A. miringoera Vanzolini, 1971, and A.
hiata Montero & Cespedez, 2002 have more than 200.
Amphisbaena brevis sp. n. can be easily distinguished
from A. silvestrii Boulenger, 1902, A. neglecta Dunn
& Piatt, 1936, A. crisae Vanzolini, 1997 and A.
anaemariae Vanzolini, 1997 by the belly that lacks
pigment, in striking contrast with the heavily
pigmented ventral surface of the tail, and the least
number of body annuli (139 instead of more than 150
for the other species). It can be further distinguished
from these four species by a higher number of dorsal
midbody segments (18 instead of 10–12, 12–14, 10,
and 14–16, respectively). The new species differs from
A. trachura Dumeril & Bibron, 1839 and A. hetero-
zonata Burmeister, 1861 by the lower number of body
annuli (139 instead of 178–199 and 190–207, respec-
tively), and lower number of midbody ventral
segments (14 instead of 16–22 and 15–22).
Description
Holotype male: small, robust, with abbreviated,
conical blunt tail. No dorsal or ventral sulci. Faint
38 C. Strussmann and T. Mott
Downloaded By: [University of Sao Paulo] At: 17:08 13 May 2009

lateral sulci. Head not distinct from trunk, with a
relatively wide, rounded snout, without major fusion
of head shields. Paired nasals, prefrontals, frontals
and parietals. Prefrontals are largest scales on top of
the head, being as long as wide, and rhomboidal.
Frontals longer but narrower than prefrontals,
almost trapezoidal in shape. Suture between frontals
is the longest on the top of the head. Parietals
irregular in shape, in point contact in sagittal line.
Anterior tip of each parietal approximately at the
same level of contact point between the third
supralabial and postocular. Parietal in broad contact
with frontal, postocular, and, on the right side, with
three large scales of the second body annulus. On left
side, two of these scales fused, and left parietal in
posterior contact with only two scales of the second
body annulus. Postocular about the same size as
parietal, lance-shaped, and in broad contact with
frontal, parietal, ocular and temporal. Postocular
also in point contact with prefrontal. Three
Figure 1. Region under direct influence of the Manso Dam (gray area) and a portion of Chapada dos Guimaraes plateau(elevations 600–900 m above sea level, stippled). Star: type-locality of Amphisbaena brevis sp. n.; dashed lines: paved roads;dotted line: unpaved road; insert gray area: limits of Cerrado biome.
Studies on Neotropical Fauna and Environment 39
Downloaded By: [University of Sao Paulo] At: 17:08 13 May 2009
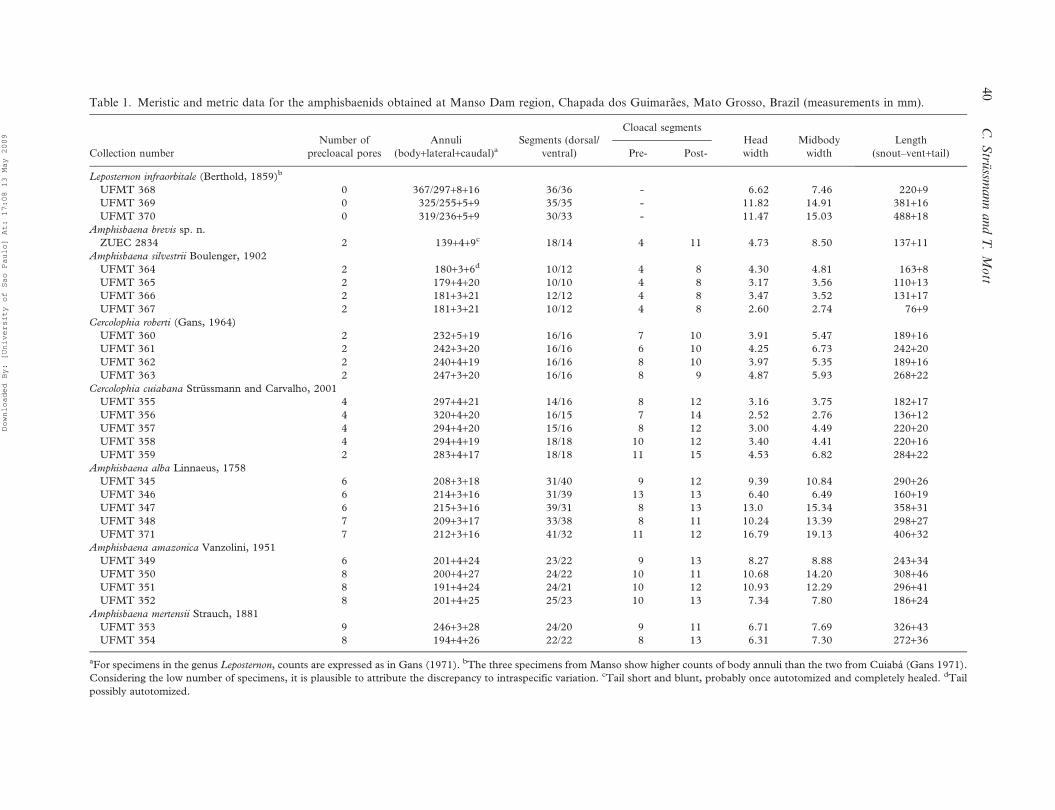
Table 1. Meristic and metric data for the amphisbaenids obtained at Manso Dam region, Chapada dos Guimaraes, Mato Grosso, Brazil (measurements in mm).
Collection number
Number of
precloacal pores
Annuli
(body+lateral+caudal)a
Segments (dorsal/
ventral)
Cloacal segmentsHead
width
Midbody
width
Length
(snout–vent+tail)Pre- Post-
Leposternon infraorbitale (Berthold, 1859)b
UFMT 368 0 367/297+8+16 36/36 - 6.62 7.46 220+9
UFMT 369 0 325/255+5+9 35/35 - 11.82 14.91 381+16
UFMT 370 0 319/236+5+9 30/33 - 11.47 15.03 488+18
Amphisbaena brevis sp. n.
ZUEC 2834 2 139+4+9c 18/14 4 11 4.73 8.50 137+11
Amphisbaena silvestrii Boulenger, 1902
UFMT 364 2 180+3+6d 10/12 4 8 4.30 4.81 163+8
UFMT 365 2 179+4+20 10/10 4 8 3.17 3.56 110+13
UFMT 366 2 181+3+21 12/12 4 8 3.47 3.52 131+17
UFMT 367 2 181+3+21 10/12 4 8 2.60 2.74 76+9
Cercolophia roberti (Gans, 1964)
UFMT 360 2 232+5+19 16/16 7 10 3.91 5.47 189+16
UFMT 361 2 242+3+20 16/16 6 10 4.25 6.73 242+20
UFMT 362 2 240+4+19 16/16 8 10 3.97 5.35 189+16
UFMT 363 2 247+3+20 16/16 8 9 4.87 5.93 268+22
Cercolophia cuiabana Strussmann and Carvalho, 2001
UFMT 355 4 297+4+21 14/16 8 12 3.16 3.75 182+17
UFMT 356 4 320+4+20 16/15 7 14 2.52 2.76 136+12
UFMT 357 4 294+4+20 15/16 8 12 3.00 4.49 220+20
UFMT 358 4 294+4+19 18/18 10 12 3.40 4.41 220+16
UFMT 359 2 283+4+17 18/18 11 15 4.53 6.82 284+22
Amphisbaena alba Linnaeus, 1758
UFMT 345 6 208+3+18 31/40 9 12 9.39 10.84 290+26
UFMT 346 6 214+3+16 31/39 13 13 6.40 6.49 160+19
UFMT 347 6 215+3+16 39/31 8 13 13.0 15.34 358+31
UFMT 348 7 209+3+17 33/38 8 11 10.24 13.39 298+27
UFMT 371 7 212+3+16 41/32 11 12 16.79 19.13 406+32
Amphisbaena amazonica Vanzolini, 1951
UFMT 349 6 201+4+24 23/22 9 13 8.27 8.88 243+34
UFMT 350 8 200+4+27 24/22 10 11 10.68 14.20 308+46
UFMT 351 8 191+4+24 24/21 10 12 10.93 12.29 296+41
UFMT 352 8 201+4+25 25/23 10 13 7.34 7.80 186+24
Amphisbaena mertensii Strauch, 1881
UFMT 353 9 246+3+28 24/20 9 11 6.71 7.69 326+43
UFMT 354 8 194+4+26 22/22 8 13 6.31 7.30 272+36
aFor specimens in the genus Leposternon, counts are expressed as in Gans (1971). bThe three specimens from Manso show higher counts of body annuli than the two from Cuiaba (Gans 1971).
Considering the low number of specimens, it is plausible to attribute the discrepancy to intraspecific variation. cTail short and blunt, probably once autotomized and completely healed. dTail
possibly autotomized.
40
C.
Stru
ssma
nn
an
dT
.M
ott
Downloaded By: [University of Sao Paulo] At: 17:08 13 May 2009
supralabials, the third one slightly larger than the
other two. First supralabial smallest, in broad contact
with rostral and nasal. Second supralabial in contact
with anterior half of lateral edge of prefrontal.
Second and third supralabials in broad contact with
ocular. Ocular diamond-shaped, almost 50% smaller
than third supralabial. Ocular in point contact with
frontal and temporal, and in broad contact with
prefrontal, postocular, and second and third supra-
labials. Three infralabials, the first smallest, the
second largest. Mental as long as wide. Post-mental
shield-shaped, almost transversally segmented by a
crease in anterior portion of scale. Post-mental in
broad contact with second infralabial and first row of
postgenials. Post-mental also in point contact with
malars. Malars about the same size as second
infralabial, almost trapezoidal in shape, and kept
from medial contact by five postgenials, in two rows.
No postmalar row.
The form has 139 body annuli and 10 caudal
annuli. First body annulus well defined only ven-
trally, formed by 17 segments (including temporals,
postoculars and parietals). Segment counts in the
next seven annuli: 18, 23, 20, 18, 20, 24, 25, the
annulus with fewest segments corresponding to
discreet nuchal constriction. Lateral sulcus beginning
at level of 40th annulus on right side, and 45th on left
side. Middorsal segments 1.5 times longer than wide,
lateral segments as long as wide and midventral
segments two times wider than long. Cloacal region
with four lateral and 11 precloacal and postcloacal
segments, respectively. Two round, small precloacal
pores, situated near posterior suture of the scale
bearing them. Number of segments on tail posterior
to postcloacal segments: 22, 21, 18, 20, 19, 17, 15, 13
and 10. Tail short, cylindrical, without autotomy
constriction, with rounded tip, probably a healed
autotomized tail.
Figure 4 shows some body proportions; numerical
data are summarized in Table 1.
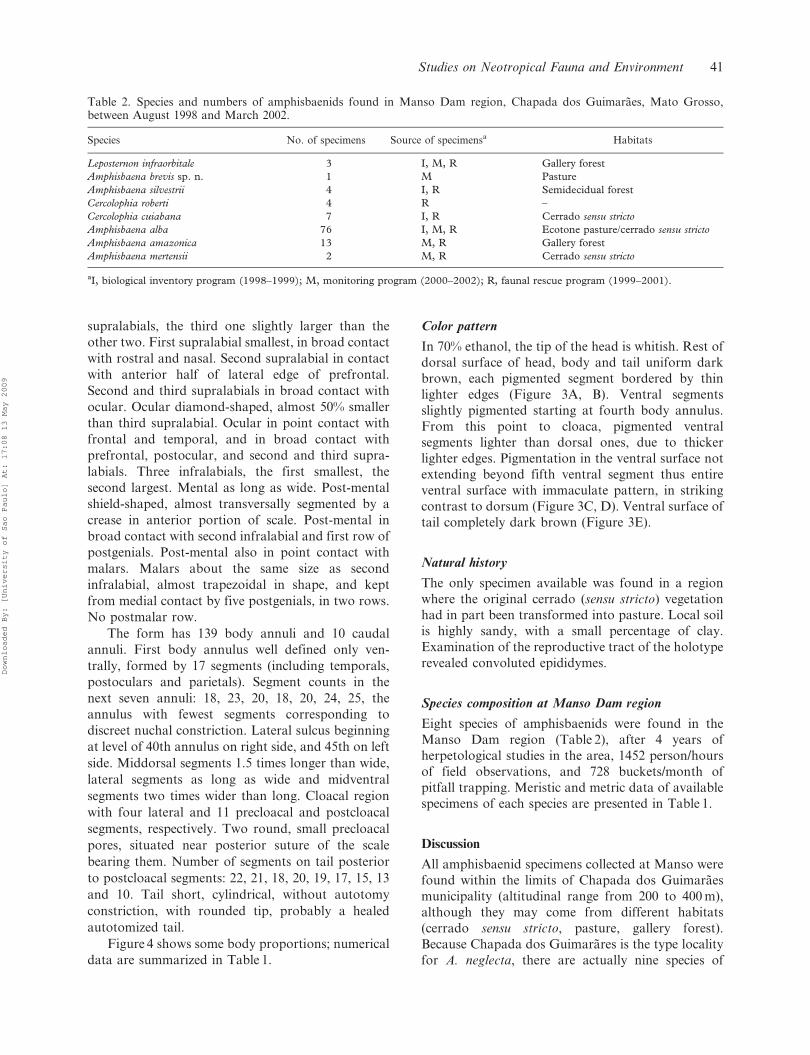
Color pattern
In 70% ethanol, the tip of the head is whitish. Rest of
dorsal surface of head, body and tail uniform dark
brown, each pigmented segment bordered by thin
lighter edges (Figure 3A, B). Ventral segments
slightly pigmented starting at fourth body annulus.
From this point to cloaca, pigmented ventral
segments lighter than dorsal ones, due to thicker
lighter edges. Pigmentation in the ventral surface not
extending beyond fifth ventral segment thus entire
ventral surface with immaculate pattern, in striking
contrast to dorsum (Figure 3C, D). Ventral surface of
tail completely dark brown (Figure 3E).
Natural history
The only specimen available was found in a region
where the original cerrado (sensu stricto) vegetation
had in part been transformed into pasture. Local soil
is highly sandy, with a small percentage of clay.
Examination of the reproductive tract of the holotype
revealed convoluted epididymes.
Species composition at Manso Dam region
Eight species of amphisbaenids were found in the
Manso Dam region (Table 2), after 4 years of
herpetological studies in the area, 1452 person/hours
of field observations, and 728 buckets/month of
pitfall trapping. Meristic and metric data of available
specimens of each species are presented in Table 1.
Discussion
All amphisbaenid specimens collected at Manso were
found within the limits of Chapada dos Guimaraes
municipality (altitudinal range from 200 to 400 m),
although they may come from different habitats
(cerrado sensu stricto, pasture, gallery forest).
Because Chapada dos Guimarares is the type locality
for A. neglecta, there are actually nine species of
Table 2. Species and numbers of amphisbaenids found in Manso Dam region, Chapada dos Guimaraes, Mato Grosso,between August 1998 and March 2002.
Species No. of specimens Source of specimensa Habitats
Leposternon infraorbitale 3 I, M, R Gallery forest
Amphisbaena brevis sp. n. 1 M Pasture
Amphisbaena silvestrii 4 I, R Semidecidual forest
Cercolophia roberti 4 R –
Cercolophia cuiabana 7 I, R Cerrado sensu stricto
Amphisbaena alba 76 I, M, R Ecotone pasture/cerrado sensu stricto
Amphisbaena amazonica 13 M, R Gallery forest
Amphisbaena mertensii 2 M, R Cerrado sensu stricto
aI, biological inventory program (1998–1999); M, monitoring program (2000–2002); R, faunal rescue program (1999–2001).
Studies on Neotropical Fauna and Environment 41
Downloaded By: [University of Sao Paulo] At: 17:08 13 May 2009
amphisbaenids known to occur in the region. This
number of sympatric (but not necessarily syntopic)
species of amphisbaenids is presently unparalleled in
any other locality in Brazil.
Following an analysis of the herpetological
material housed in the Universidade de Brasılia
collection (CHUNB), Araujo & Colli (2003) stated
that for localities in the better sampled East–West
main axis of the Cerrado, evidence suggests the
coexistence of only two sympatric species of amphis-
baenids, in a radius of 20 km. This suggestion clearly
needs to be reexamined, in the light of recent findings,
at least for Amazon Forest–Cerrado contact zones.
Five species of amphisbaenids were found in an
area of 1784 km2 flooded by the Serra da Mesa
hydroeletric power plant (Castro-Mello 1999). In
Figure 2. Holotype of Amphisbaena brevis sp. n. (ZUEC2834). (A–C) Dorsal, lateral (right side), and ventral view ofthe head; (D) ventral view of the cloacal region and tail.Scale bars: 1.5 mm.
Figure 3. Holotype of Amphisbaena brevis sp. n. (ZUEC2834); pigmentation pattern and relative segmental propor-tions. (A) Dorsal view of head; (B) dorsal view of midbody;(C) lateral view of midbody; (D) ventral view of midbody;(E) ventral view of cloacal region and tail.
42 C. Strussmann and T. Mott
Downloaded By: [University of Sao Paulo] At: 17:08 13 May 2009
another open formation, the Brazilian Caatinga,
Rodrigues (2002) reported the presence of seven
species of amphisbaenids on Quaternary sand dunes
of the Middle Sao Francisco River (9–11uS, 41–
43uW), in the state of Bahia. Although this appears
an impressive number of sympatric amphisbaenids, it
must be considered that sand dunes represent an
extensive area (approximately 5000 km2, according to
Rodrigues 1996). Additionally, the Sao Francisco
River represents a geographical barrier between
herpetofaunal assemblages from opposite riverbanks,
and no more than four sympatric amphisbaenids
were recorded on the same bank.
The total number of amphisbaenid individuals
obtained at Manso Dam region is less than the
amount recorded for other hydroelectric power
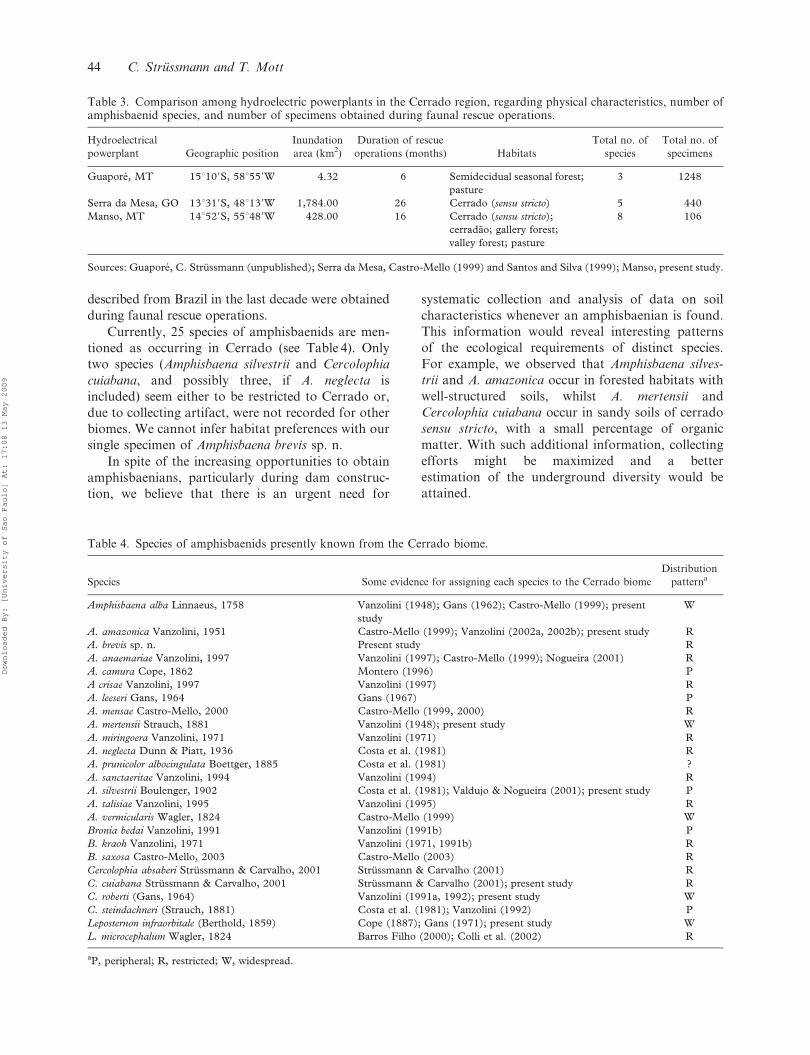
plants in the Cerrado (Table 3). Data in Table 3
indicate that the number of specimens is not
proportional to the size of the reservoir. Therefore,
differential faunal rescue efforts, timing of reservoir
filling, and topography, among other factors, may
have influenced the numbers of amphisbaenids
obtained in the hydroelectric power plants. This
point needs further investigation.
Although ecological and/or historical features
might be the reason for the unusual amphisbaenid
richness at Manso Dam region, the extensive collecting
efforts at this site seem to offer a better explanation for
finding eight sympatric species. Despite the dearth of
appropriate methodologies for capturing fossorial
herpetofauna and scant zoological knowledge of the
Cerrado, the Manso Dam site has become a unique
opportunity for researchers. The reservoir of Manso
not only flooded a huge area, but also demanded the
longest zoological inventory, faunal rescue and long-
term monitoring programs ever done in Mato Grosso
state. Field efforts spent in this area were likely to be
decisive for finding several species of sympatric
amphisbaenids at Chapada dos Guimaraes. In fact,
the high rate at which new species of amphisbaenids are
being described in recent years is clearly related to the
increasing number of rescue operations during the
filling of reservoirs from hydroelectric power plants.
The types of six out of the 12 species of Amphisbaena
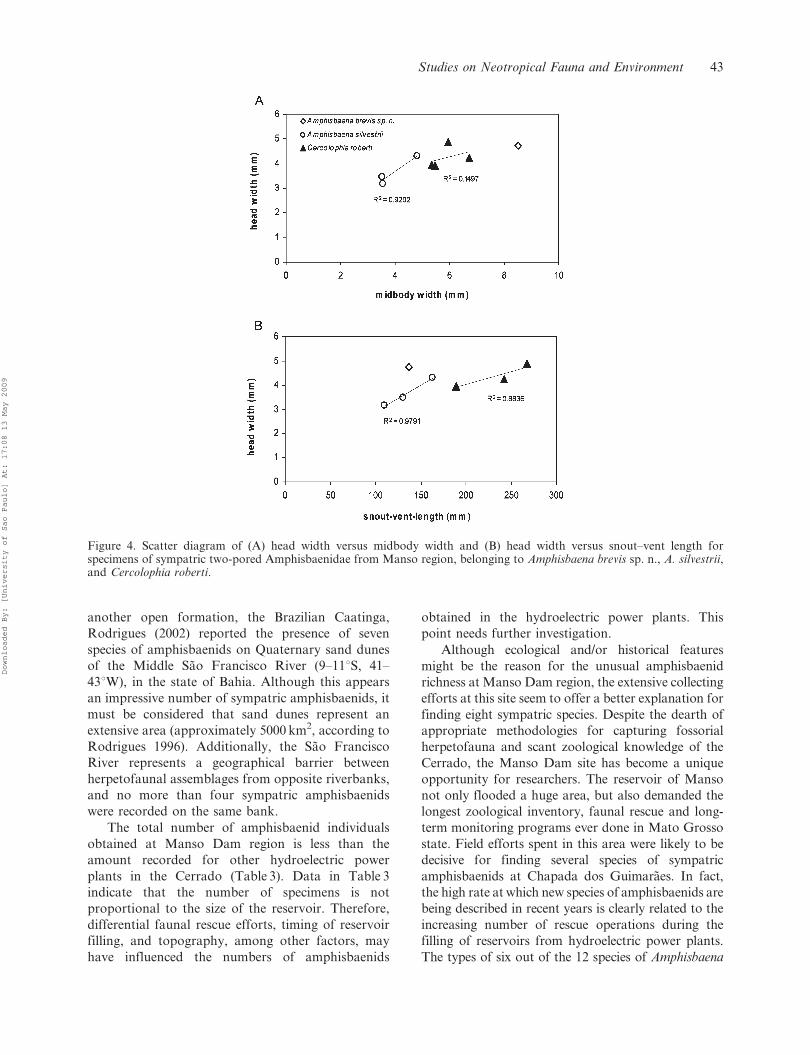
Figure 4. Scatter diagram of (A) head width versus midbody width and (B) head width versus snout–vent length forspecimens of sympatric two-pored Amphisbaenidae from Manso region, belonging to Amphisbaena brevis sp. n., A. silvestrii,and Cercolophia roberti.
Studies on Neotropical Fauna and Environment 43
Downloaded By: [University of Sao Paulo] At: 17:08 13 May 2009
described from Brazil in the last decade were obtained
during faunal rescue operations.
Currently, 25 species of amphisbaenids are men-
tioned as occurring in Cerrado (see Table 4). Only
two species (Amphisbaena silvestrii and Cercolophia
cuiabana, and possibly three, if A. neglecta is
included) seem either to be restricted to Cerrado or,
due to collecting artifact, were not recorded for other
biomes. We cannot infer habitat preferences with our
single specimen of Amphisbaena brevis sp. n.
In spite of the increasing opportunities to obtain
amphisbaenians, particularly during dam construc-
tion, we believe that there is an urgent need for
systematic collection and analysis of data on soil
characteristics whenever an amphisbaenian is found.
This information would reveal interesting patterns
of the ecological requirements of distinct species.
For example, we observed that Amphisbaena silves-
trii and A. amazonica occur in forested habitats with
well-structured soils, whilst A. mertensii and
Cercolophia cuiabana occur in sandy soils of cerrado
sensu stricto, with a small percentage of organic
matter. With such additional information, collecting
efforts might be maximized and a better
estimation of the underground diversity would be
attained.
Table 3. Comparison among hydroelectric powerplants in the Cerrado region, regarding physical characteristics, number ofamphisbaenid species, and number of specimens obtained during faunal rescue operations.
Hydroelectrical
powerplant Geographic position
Inundation
area (km2)
Duration of rescue
operations (months) Habitats
Total no. of
species
Total no. of
specimens
Guapore, MT 15u109S, 58u559W 4.32 6 Semidecidual seasonal forest;
pasture
3 1248
Serra da Mesa, GO 13u319S, 48u139W 1,784.00 26 Cerrado (sensu stricto) 5 440
Manso, MT 14u529S, 55u489W 428.00 16 Cerrado (sensu stricto);
cerradao; gallery forest;
valley forest; pasture
8 106
Sources: Guapore, C. Strussmann (unpublished); Serra da Mesa, Castro-Mello (1999) and Santos and Silva (1999); Manso, present study.
Table 4. Species of amphisbaenids presently known from the Cerrado biome.
Species Some evidence for assigning each species to the Cerrado biome
Distribution
patterna
Amphisbaena alba Linnaeus, 1758 Vanzolini (1948); Gans (1962); Castro-Mello (1999); present
study
W
A. amazonica Vanzolini, 1951 Castro-Mello (1999); Vanzolini (2002a, 2002b); present study R
A. brevis sp. n. Present study R
A. anaemariae Vanzolini, 1997 Vanzolini (1997); Castro-Mello (1999); Nogueira (2001) R
A. camura Cope, 1862 Montero (1996) P
A crisae Vanzolini, 1997 Vanzolini (1997) R
A. leeseri Gans, 1964 Gans (1967) P
A. mensae Castro-Mello, 2000 Castro-Mello (1999, 2000) R
A. mertensii Strauch, 1881 Vanzolini (1948); present study W
A. miringoera Vanzolini, 1971 Vanzolini (1971) R
A. neglecta Dunn & Piatt, 1936 Costa et al. (1981) R
A. prunicolor albocingulata Boettger, 1885 Costa et al. (1981) ?
A. sanctaeritae Vanzolini, 1994 Vanzolini (1994) R
A. silvestrii Boulenger, 1902 Costa et al. (1981); Valdujo & Nogueira (2001); present study P
A. talisiae Vanzolini, 1995 Vanzolini (1995) R
A. vermicularis Wagler, 1824 Castro-Mello (1999) W
Bronia bedai Vanzolini, 1991 Vanzolini (1991b) P
B. kraoh Vanzolini, 1971 Vanzolini (1971, 1991b) R
B. saxosa Castro-Mello, 2003 Castro-Mello (2003) R
Cercolophia absaberi Strussmann & Carvalho, 2001 Strussmann & Carvalho (2001) R
C. cuiabana Strussmann & Carvalho, 2001 Strussmann & Carvalho (2001); present study R
C. roberti (Gans, 1964) Vanzolini (1991a, 1992); present study W
C. steindachneri (Strauch, 1881) Costa et al. (1981); Vanzolini (1992) P
Leposternon infraorbitale (Berthold, 1859) Cope (1887); Gans (1971); present study W
L. microcephalum Wagler, 1824 Barros Filho (2000); Colli et al. (2002) R
aP, peripheral; R, restricted; W, widespread.
44 C. Strussmann and T. Mott
Downloaded By: [University of Sao Paulo] At: 17:08 13 May 2009

Range extensions
Amphisbaena amazonica
Known distribution of the species is based on reports
by Vanzolini (1951), Castro-Mello (1999), Chalkidis
(2000), Chalkidis et al. (2002), and Vanzolini (2002b).
The only records for the Cerrado region are those of
Castro-Mello (1999): Minacu (13u319S, 48u139W),
and of Vanzolini (2002b): Piracanjuba (17u189S,
49u019W), both in the state of Goias. All other
specimens mentioned in the literature are from
localities in Amazonian or Atlantic rainforest. The
presence of A. amazonica at Manso is the first record
for the Paraguay River Basin (see Brasil 1997), and
the westernmost record in the Cerrado biome,
extending its known distribution for approximately
750 km northwestwards of Piracanjuba.
Amphisbaena mertensii
In his redescription of the species, Gans (1966)
provided a distribution map depicting Igarapava
(state of Sao Paulo, near 20uS) as the northernmost
known record for A. mertensii. Actually, the north-
ernmost locality cited by Gans in the text is Colonia
Teresa Cristina, the type locality of A. mattogrossen-
sis Peracca, 1898, a junior synonym for A. mertensii.
This locality was misplaced in Gans’ map, beyond
Igarapava and far from its actual geographic position
(approximately 16u389S, 55u149W), in the state of
Mato Grosso instead of Mato Grosso do Sul, as one
can infer from the map. Presence of A. mertensii at
the Manso Dam region extends the known range of
the species ca. 200 km northwards of Colonia Teresa
Cristina (most probably, an indigenous reserve
mentioned as ‘‘Aldeia Teresa Cristina’’ in recent
maps.
Cercolophia cuiabana
This species has been recorded only from the type
locality Cuiaba (Strussmann & Carvalho 2001). The
occurrence in the neighborhood of the Manso Dam
extends its known range for about 60 km to the
northeast.
Cercolophia roberti
At the time of its description as Amphisbaena roberti,
the species was known from 11 localities in the
Brazilian states of Sao Paulo, Minas Gerais, and
Goias (Gans 1964). Examination of additional
material allowed the range of the species to be
expanded to localities in the states of Espirito Santo
and Parana (Vanzolini 1991a; Moura-Leite et al.
1996). Montero and Terol (1999) referred to
Paraguayan material from Amambay (20u179S,
58u179W), this being the westernmost record for the
species. The presence of C. roberti at the Manso Dam
region adds another Brazilian state to the list: Mato
Grosso.
Acknowledgements
Jair Serratel offered permission and facilities for the
fieldwork at Fazenda Morro do Chapeu; Cleber J.R.
Alho led the biological programs conducted at
Manso; many volunteers and workers involved in
these programs offered field companionship and/or
specimens; Wolf Dieter Eberhard helped with the
map; Catia Nunes da Cunha (Project Ecologia do
Gran Pantanal/Federal University of Mato Grosso)
offered us the laboratory support which made it
possible to examine and draw the type specimen.
Sheda Morshed reviewed our English on a first draft
of the manuscript, and Marvalee Wake gave valuable
suggestions to improve the text. We express our
gratitude to all these people.
References
Alho CJR. 2000. Fauna Silvestre da regiao do rio Manso, MT.
Brasılia: Ministerio do Meio Ambiente, Edicoes IBAMA and
Centrais Eletricas do Norte do Brasil. Chapter I, Paisagens e
enfoque. p. 19–29.
Alho CJR, Martins ES. 1995. De grao em grao o Cerrado perde
espaco. Brasılia: WWF and Sociedade de Pesquisas Ecologicas
do Cerrado.
Araujo AFB. 1994. Herpetologia no Brasil. Belo Horizonte (MG):
PUC-MG, Fundacao Biodiversitas and Fundacao Ezequiel
Dias. p. 58–67 (Comunidades de lagartos brasileiros).
Araujo AFB, Colli GR. 2003. Biodiversidade do Cerrado:
Herpetofauna [Internet]. Belo Horizonte (MG): Biodiversitas;
[cited 2003 Dec 21]. Available from: http://www.bdt.fat.org/pdf/
workcerrado/REPTEIS.PDF.
Barros Filho JD. 2000. Osteologia craniana comparada de especies
do genero Leposternon Wagler, 1824 (Reptilia, Amphisbaenia)
[PhD dissertation]. Sao Paulo (SP): University of Sao Paulo.
Brasil. 1997. Programa Nacional do Meio Ambiente/Projeto
Pantanal. Plano de Conservacao da Bacia do Alto Paraguai
(Pantanal) – PCBAP: diagnostico dos meios fısico e biotico.
Meio Biotico. Brasılia: MMA and PNMA.
Castro-Mello C. 1999. Estudo de uma comunidade de Amphisbaena
(Squamata: Amphisbaenidae). Paper presented at: Workshop A
Fauna do AHE Serra da Mesa, GO; 2003; Goiania, Brazil.
Castro-Mello C. 2000. A new species of Amphisbaena from Central
Brasil (Squamata: Amphisbaenidae). Pap Avulsos Zool.
41(16):243–246.
Castro-Mello C. 2003. Nova especie de Bronia Gray, 1845, do
estado do Tocantins, Brasil (Squamata: Amphisbaenidae). Pap
Avulsos Zool. 43(7):139–143.
Chalkidis HM. 2000. Amphisbaena fuliginosa varia. Geographic
distribution. Herpetol Rev. 31(4):253.
Chalkidis HM, Cordeiro-Duarte AC, Vogt RC. 2002. Amphisbaena
fuliginosa fuliginosa. Geographic distribution. Herpetol Rev.
33(2):148.
Studies on Neotropical Fauna and Environment 45
Downloaded By: [University of Sao Paulo] At: 17:08 13 May 2009

Colli GR, Bastos RP, Araujo AFB. 2002. The Cerrados of Brazil.
Ecology and natural history of a Neotropical savanna. Columbia
University Press (NY): Oliveira PS, Marquis RJ. Chapter 12,
The character and dynamics of the Cerrado herpetofauna;
223–239.
Conservation International, Biodiversity Hotspots, Brazilian
Cerrado [Internet]. Washington (DC); [cited 2005 Jan 2].
Available from: http://www.biodiversityhotspots.org/xp/Hotspots/
cerrado.
Cope ED. 1887. Synopsis of the Batrachia and Reptilia obtained by
H.H. Smith in the Province of Mato Grosso, Brazil. Proc Am
Philos Soc. 24(12):44–60.
Costa CCC, Lima JP, Cardoso LD, Henriques VQ. 1981. Fauna do
Cerrado: lista preliminar de aves, mamıferos e repteis. Rio de
Janeiro: IBGE.
Gans C. 1962. Notes on amphisbaenids (Amphisbaenia, Reptilia).
5. A redefinition and biogeography of Amphisbaena alba
Linnaeus. Am Mus Novit. 2105(24):1–31.
Gans C. 1964. Notes on amphisbaenids 15. The South American
species of Amphisbaena with a vertically keeled tail (Reptilia,
Amphisbaenidae). Senck Biol. 45(3–5):387–416.
Gans C. 1966. Redescription of Amphisbaena mertensii Strauch,
with comments on its geographic variation and synonymy
(Amphisbaenia: Reptilia). Copeia. 1966(3):534–548.
Gans C. 1967. A new record of Amphisbaena leeseri from Mato
Grosso. Herpetologica. 23(3):247–248.
Gans C. 1971. Studies on Amphisbaenians (Amphisbaenia,
Reptilia) 4. A review of the Amphisbaenid genus Leposternon.
Bull Am Mus Nat Hist. 144(6):383–464.
Gans C. 2005. Checklist and bibliography of the Amphisbaenia of
the world. Bull Am Mus Nat Hist. 289:1–130.
Gans C, Alexander A. 1962. Studies on Amphisbaenids
(Amphisbaenia, Reptilia). 2. On the Amphisbaenids of the
Antilles. Bull Mus Comp Zool. 128(3):65–158.
Kearney M, Stuart L. 2004. Repeated evolution of limblessness and
digging heads in worm lizards revealed by DNA from old bones.
Proc Roy Soc Lond B Biol Sci. 271(1549):1677–1683.
Leviton AE, Gibbs Jr RH, Heal E, Dawson CE. 1985. Standards in
herpetology and ichthyology: Part I. Standard symbolic code for
institutional resource collections in herpetology and ichthyology.
Copeia. 1985(3):802–832.
Macey JR, Papenfuss TJ, Kuel JV, Fourcade HM, Boore JL. 2004.
Phylogenetic relationships among amphisbaenian reptiles based
on complete mitochondrial genome sequences. Mol Phylogenet
Evol. 33(1):22–31.
Montero R. 1996. Amphisbaena bolivica Mertens 1929, nueva
combinacion (Squamata: Amphisbaenia). Cuad Herp.
9(2):75–84.
Montero R, Cespedez J. 2002. A new two pored Amphisbaena
(Squamata: Amphisbaenidae) from Argentina. Copeia.
2002(3):792–797.
Montero R, Terol GJ. 1999. Los Amphisbaenidae en Paraguay,
listado geografico. Cuad Herp. 13(1–2):89–95.
Moura-Leite JC, Morato SAA, Bernils RS. 1996. New records of
reptiles from the state of Parana, Brazil. Herpetol Rev.
27(4):216–217.
Myers N, Mittermeier RA, Mittermeier CG, Fonseca GAB, Kent
J. 2001. Biodiversity hotspots for conservation priorities. Nature.
403(6772):853–858.
Nimer E. 1979. Climatologia do Brasil. Rio de Janeiro: IBGE,
Superintendencia de Recursos Naturais e Meio Ambiente.
Nogueira CC. 2001. New records of Squamate reptiles in Central
Brazilian Cerrado. II. Herpetol Rev. 32(4):285–287.
Rodrigues MT. 1996. Lizards, snakes and amphisbaenians from
the Quaternary sand dunes of the Middle Rio Sao Francisco,
Bahia, Brazil. J Herpetol. 30(4):513–523.
Rodrigues MT. 2002. Herpetofauna of the Quaternary sand dunes
of the middle Rio Sao Francisco, Bahia, Brazil. VIII.
Amphisbaena arda sp. nov., a fuliginosa-like checkered patterned
Amphisbaena (Squamata, Amphisbaenidae). Phyllomedusa.
1(2):51–56.
Santos HGP, Silva Jr NJ. 1999. Resgate da Fauna do
Aproveitamento Hidreletrico (AHE) Serra da Mesa, Goias.
Paper presented at: Resumos Seminario Nacional Producao e
Transmissao Energia Eletrica (XV); Foz do Iguacu, Brazil.
Strussmann C, Carvalho MA. 2001. Two new species of
Cercolophia Vanzolini, 1992 from the state of Mato Grosso,
western Brazil (Reptilia, Amphisbaenia, Amphisbaenidae). Boll
Mus Reg Sci Nat (Torino). 18(2):487–505.
Townsend TM, Larson A, Louis E, Macey JR. 2004. Molecular
phylogenetics of Squamata: the position of snakes, amphisbae-
nians, and dibamids, and the root of the squamate tree. Syst
Biol. 53(5):735–757.
Valdujo PH, Nogueira C. 2001. New records of Squamate reptiles
in Central Brazilian Cerrado: Emas National Park Region.
Herpetol Rev. 32(3):128–130.
Vanzolini PE. 1948. Notas sobre os ofıdios e lagartos da Cachoeira
de Emas, no municıpio de Pirassununga, Estado de Sao Paulo.
Rev Bras Biol. 8(3):377–400.
Vanzolini PE. 1951. Contributions to the knowledge of the
Brazilian lizards of the family Amphisbaenidae Gray, 1925. 6.
On the geographical distribution and differentiation of
Amphisbaena fuliginosa Linne. Bull Mus Comp Zool.
106(1):1–67.
Vanzolini PE. 1971. New Amphisbaenidae from Brazil (Sauria).
Pap Avulsos Zool. 24(14):191–195.
Vanzolini PE. 1991a. Biometry and geographical differentiation of
Amphisbaena roberti Gans, 1964 (Reptilia, Amphisbaenia). Pap
Avulsos Zool. 37(24):363–377.
Vanzolini PE. 1991b. A third species of Bronia Gray, 1865
(Reptilia, Amphisbaenia). Pap Avulsos Zool. 37(25):379–388.
Vanzolini PE. 1992. Cercolophia, a new genus for the species of
Amphisbaena with a terminal vertical keel on the tail (Reptilia,
Amphisbaenia). Pap Avulsos Zool. 37(27):401–412.
Vanzolini PE. 1994. A new species of Amphisbaena from the state
of Sao Paulo (Reptilia, Amphisbaenia, Amphisbaenidae). Pap
Avulsos Zool. 39(3):29–32.
Vanzolini PE. 1995. A new species of Amphisbaena from the state
of Mato Grosso, Brazil (Squamata, Amphisbaenidae). Pap
Avulsos Zool. 39(10):217–221.
Vanzolini PE. 1997. The silvestrii species group of Amphisbaena,
with the description of two new Brazilian species (Reptilia:
Amphisbaenia). Pap Avulsos Zool. 40(3):65–85.
Vanzolini PE. 2002a. An aid to the identification of the South
American species of Amphisbaena (Squamata, Amphisbaenidae).
Pap Avulsos Zool. 42(15):351–362.
Vanzolini PE. 2002b. A second note on the geographical
differentiation of Amphisbaena fuliginosa L., 1758 (Squamata,
Amphisbaenidae), with a consideration of the forest refuge
model of speciation. An Acad Bras Cienc. 74(4):609–648.
Vidal N, Azvolinsky A, Cruaud C, Hedges SH. 2008. Origin of
tropical American burrowing reptiles by transatlantic rafting.
Biol Lett. 4(1):115–118.
46 C. Strussmann and T. Mott
Downloaded By: [University of Sao Paulo] At: 17:08 13 May 2009
Copyright © 2022 FDOKUMEN




















