
LABORATORIO VIRTUAL REMOTO PARA ROBÓTICA Y EVALUACIÓN DE SU IMPACTO EN LA DOCENCIA
Upload
manchesterCategory
view
2download
0

������
����
���

Opiliones fósilesLos arácnidos actuales de origen más remoto
Russell J. Garwood, Jason A. Dunlop, Gonzalo Giribet y Mark D. Sutton

2
Russell J. Garwood, Jason A. Dunlop, Gonzalo Giribet y Mark D. Sutton (2013). Opiliones fósiles. Los arácnidos actuales de origen más remoto. ¡Fundamental! 23, 1–58.
Este número se ha editado en el marco del 10º Premio Internacional de Investigación en
Paleontología Paleonturología 12, convocado por la Fundación Conjunto Paleontológico
de Teruel–Dinópolis, la Fundación Teruel Siglo XXI y Dinópolis.
Esta publicación forma parte de los proyectos de investigación en Paleontología subvencionados por:
Departamento de Educación, Universidad, Cultura y Deporte del Gobierno de Aragón; Departamento de
Industria e Innovación (Grupo de Investigación Consolidado E–62 FOCONTUR) del Gobierno de Aragón;
Fondo Social Europeo, Instituto Aragonés de Fomento, Dinópolis y Diputación Provincial de Teruel.
EDICIÓN: © Fundación Conjunto Paleontológico de Teruel – Dinópolis
AUTORES: Russell J. Garwood, Jason A. Dunlop, Gonzalo Giribet y Mark D. Sutton
COORDINACIÓN: Luis Alcalá y Francisco Javier Verdú
DISEÑO Y MAQUETA: © EKIX Soluciones Gráficas
DEPÓSITO LEGAL: TE–194–2013
ISBN–13: 978–84–938173–6–7
Queda rigurosamente prohibida, sin la autorización escrita de los autores y del editor, bajo las sanciones establecidas en la ley, la reproducción total o parcial de esta obra por cualquier medio o procedimiento, comprendidos la reprografía y el tratamiento informático. Todos los derechos reservados.

3
10º PREMIO INTERNACIONALDE INVEST IGACIÓN EN PALEONTOLOGÍA
Paleonturología 12
En la ciudad de Teruel, siendo las 22:30 horas del día 11 de diciembre de 2012, se reúnen en calidad de miembros del jurado la Dra. Ana Márquez Aliaga (Profesora de Paleontología de la Universidad de Valencia y Presidenta de la Sociedad Española de Paleontología), el Dr. Enrique Gil Bazán (Profesor de Biología y Geología en el IES Ramón y Cajal de Zaragoza) y el Dr. Heinrich Mallison (Museum für Naturkunde–Leibniz Institute for Research on Evolution and Biodiversity at the Humboldt University Berlin, premiado en Paleonturología 11) y, en calidad de secretario, el Dr. Luis Alcalá (Director Gerente de la Fundación Conjunto Paleontológico de Teruel-Dinópolis/Museo Aragonés de Paleontología),
después de examinar los 19 artículos científicos participantes en la fase final del Décimo Premio Internacional de Investigación en Paleontología Paleonturología 12 (convocado por la Fundación Conjunto Paleontológico de Teruel-Dinópolis, la Fundación Teruel Siglo XXI y Dinópolis) y tras las oportunas deliberaciones,
acuerdan conceder el Premio Paleonturología 12, dotado con 4.500 euros y una edición divulgativa del trabajo premiado, al artículo:
Anatomically modern Carboniferous harvestmen demonstrateearly cladogenesis and stasis in Opiliones
cuyos autores sonRusell J. Garwood, Jason A. Dunlop, Gonzalo Giribet y Mark D. Sutton
(Imperial College London, Humboldt University Berlin, Harvard University Cambridge)
publicado en la revista Nature Communications, 2: 444, 1-7. 2011
Los opiliones o segadores son arácnidos que, superficialmente, se parecen a las arañas; se trata de un grupo de organismos cuyo registro fósil se conoce escasamente. El artículo premiado describe dos nuevas especies de opiliones del Carbonífero de Francia, de unos 305 millones de años de antigüedad, a partir de fósiles excelentemente conservados y utilizando para su estudio microtomografías de rayos X de alta definición.
El artículo presenta, de modo claro y conciso, cómo funciona realmente la evolución, para lo cual utiliza técnicas actuales y combina los análisis tradicionales -morfológicos- con los moleculares para proponer el primer análisis cladístico (es decir, de parentesco) de los opiliones del Paleozoico. A pesar de su gran antigüedad, los dos nuevos opiliones tienen características modernas, demostrando que su configuración anatómica no ha variado en 300 millones de años.
Los autores han desarrollado una aplicación –de descarga gratuita- que permite reproducir resultados e introducen la información de modo muy accesible, de modo que presenta grandes posibilidades para la divulgación de los métodos de investigación en paleontología.
De todo lo cual doy constancia, a las 1:02 horas del día 12 de diciembre de 2012.
El SecretarioLuis Alcalá

Protimesius longipalpis (Stygnidae, Laniatores), Reserva Ducke, Manaos, Amazonas, Brasil. Fotografía: Gonzalo Giribet.

5
Russell Garwood es paleontólogo en la Universidad de
Manchester, Reino Unido. Comenzó su carrera estudiando
geología en la Royal School of Mines, Imperial College,
Londres, antes de completar su doctorado en la misma
institución en 2010. Desde entonces ha trabajado en el Natural
History Museum de Londres, en su laboratorio de Tomografía
Computarizada, y también en la School of Materials en
Manchester. Desde 2012, Russell está siendo financiado,
durante un periodo de tres años, por la Royal Commision for
the Exhibition of 1851, para el estudio del origen y evolución
de los insectos, y por la School of Earth, Atmospheric and
Environmental Sciences. Fundamentalmente utiliza rayos X
para entender mejor a estos animales y a sus ecosistemas y
al mismo tiempo está interesado en una gran variedad de los
primeros animales terrestres. Otros aspectos que le atraen son
el origen y la evolución temprana de la vida, así como el uso de
ordenadores y técnicas de rayos X en geología.
Ru
sse
ll G
arw
oo
d

6
Jason Dunlop nació en Basingstoke, Inglaterra, y creció
en la localidad costera de Cleethorpes. Ya en su infancia le
fascinaban las arañas de su jardín y pronto decidió que le
gustaría convertir esa afición en su carrera profesional. Así,
Jason estudió zoología en la Universidad de Leeds y después
emprendió un doctorado acerca de un grupo extinto de
arácnidos fósiles en el Departamento de Ciencias de la Tierra
de la Universidad de Manchester, bajo la supervisión del Dr.
Paul Senden. Jason permaneció en Manchester con una beca
para investigar el origen de los arácnidos y sus parientes, y
en 1997 obtuvo una plaza de conservador de arácnidos y
miriápodos en el Museum für Naturkunde Berlin, puesto que
aún ocupa en la actualidad. Jason ha publicado numerosos
estudios sobre arácnidos fósiles –con un particular interés en los
opiliones durante los últimos años– y mantiene regularmente
actualizada la lista taxonómica de todos los fósiles de arácnidos
y sus parientes más cercanos. Actualmente es secretario de la
International Society of Arachnology y vicepresidente de la
European Society of Arachnology.
Jaso
nD
un
lop

7
Gonzalo Giribet es un zoólogo interesado en el origen de la
diversidad animal, especialmente en el origen de los grandes filos
animales y su diversidad, especialmente artrópodos y moluscos,
pero también en los llamados “filos menores”, grupos con
animales de pequeño tamaño y poca diversidad de especies. Es
profesor en el departamento de biología evolutiva y de organismos
de la Universidad de Harvard y conservador de la colección de
invertebrados en el Museo de Zoología Comparada de Harvard. Su
trabajo consiste en dar clases de zoología, sistemática, biogeografía
y en dirigir la colección de invertebrados del museo. Su investigación
le ha llevado a recolectar invertebrados en todos los continentes y
océanos. Aunque nació en Burgos, creció en Vilanova i la Geltrú,
en la costa barcelonesa, donde desarrolló su pasión por la zoología
y por los animales marinos. Estudió en la Universitat de Barcelona,
donde también recibió su doctorado. En 1997 inició una estancia
postdoctoral en el American Museum of Natural History de Nueva
York y de ahí pasó a Harvard, primero como profesor ayudante,
luego como profesor asociado y finalmente como catedrático.Go
nza
loG
irib
et

8
Mark Sutton es profesor titular de paleontología en
el Imperial College de Londres. Estudió en Cambridge
(licenciatura) y en Cardiff (doctorado) y ha ocupado cargos
académicos en Lampeter, Oxford y, desde 2005, en el
Imperial College de Londres. Su investigación ha estado
centrada principalmente en la conservación tridimensional
de invertebrados fósiles del ‘Herefordshire Lagerstätte’ del
Silúrico, pero mantiene un gran interés en la filogenia y
evolución de muchos grupos, particularmente braquiópodos,
artrópodos y moluscos. También investiga las aplicaciones de
los ordenadores en paleontología, especialmente técnicas de
reconstrucción tridimensional.
Ma
rkS
utt
on

Opiliones fósilesLos arácnidos actuales de origen más remoto
Russell J. Garwood, Jason A. Dunlop, Gonzalo Giribet y Mark D. Sutton

Phareicranaus manauara (Cranaidae, Laniatores), Reserva Ducke, Manaos, Amazonas, Brasil. Fotografía: Gonzalo Giribet.

11
Puede ser que nunca antes haya oído hablar acerca de un
opilión (también conocido como segador). Y podría pensar que
son completamente insignificantes. Para los aracnófobos que
nos rodean pueden ser causa de una gran aprensión. Lo más
probable es, sin embargo, que usted haya visto a alguno de estos
arácnidos: puede haber alguno en su casa, o dondequiera que
esté leyendo esto ahora mismo, y hay realmente más especies
de opiliones en la actualidad que especies de mamíferos.
Esperamos convencerle en el transcurso de este artículo que los
opiliones son unas criaturas realmente interesantes. No hay que
temerlos sino que, al contrario, merecen un poco de respeto.
Los opiliones han estado por aquí desde hace mucho tiempo,
son biológicamente interesantes, y contribuyen de modo
relevante a los ecosistemas. También son cada vez más útiles
como organismos–modelo (criaturas que los científicos pueden
usar para entender el desarrollo y la evolución de los animales).
Este artículo deriva de una publicación que escribimos en 2011.
Aunque en este artículo presentaremos al grupo en su conjunto,
así como su registro fósil, nos centraremos en los animales de
un tiempo concreto (el Carbonífero; hace 360–299 millones
de años) que hemos estudiado con una técnica muy precisa
(Tomografía Axial Computarizada, TAC, de alta resolución).
Pero antes introduciremos las primeras formas de vida terrestres,
según lo que sabemos a partir del registro fósil. Y no porque
ello sea particularmente importante para este estudio sino
porque es realmente interesante. Luego aportaremos una idea
general de lo que ya conocemos acerca de los opiliones fósiles,
presentaremos nuestro periodo de tiempo (la última parte del
Carbonífero) y explicaremos cómo funciona el escaneado TAC.
Finalmente, mostraremos los nuevos y realmente fascinantes
fósiles que han sido la base de nuestra publicación. Así,
comencemos por echar un vistazo a la vida terrestre.
Terrestrización
Los animales terrestres son aquellos que viven sobre la
tierra y terrestrización es el proceso según el cual evolucionó
este modo de vida (aunque, como sucede con estas cosas,
establecer definiciones precisas no es tan simple como podría
parecer). La inmensa mayoría de especies conocidas se incluye
en dos grupos de artrópodos terrestres; para más información
(incluyendo una definición de artrópodo) consultar el encarte
Artrópodos y arácnidos (pag. 16). Estimaciones acerca de la
diversidad de la vida proponen que los insectos suponen más
del 50% de las especies vivas conocidas, incluyendo plantas,
bacterias y todo el resto de animales. Algunos estudios
sugieren que esta cifra podría alcanzar incluso un 85%,
aunque no conocemos la proporción exacta con certeza. De
las especies restantes, una buena proporción (una quinta
parte o más) son arácnidos: las arañas y sus parientes. Por
tanto, los artrópodos terrestres son grupos numerosos e
importantes. Fueron también los primeros animales terrestres
que conocemos. Pudieron haber sido precedidos por animales
de cuerpos blandos, como gusanos, pero es difícil de decir ya
que raramente se conservan bien como fósiles. Los animales
terrestres más primitivos tienen una antigüedad de unos 427
millones de años y se trata de dos milpiés descubiertos en
Stonehaven, Escocia (Figura 1). Cuando tratamos con fósiles
de esta antigüedad hemos de considerar la posibilidad de
que algo similar a un milpiés podría haber vivido en el agua.
Estamos seguros de que no era el caso de estas dos especies
porque tienen los llamados espiráculos (orificios de su sistema
de respiración que les ayudan a mantenerse húmedos y a
controlar el flujo de oxígeno) que no serían necesarios en un
animal que viviera en el agua. Es probable que los milpiés de
Opiliones fósiles. Los arácnidos actuales de origen más remotoRussell J. Garwood, Jason A. Dunlop, Gonzalo Giribet y Mark D. Sutton

12
entonces fueran detritívoros, como lo son hoy en día, comiendo
materia orgánica en descomposición (probablemente derivada
de plantas). No había bosques en aquellos momentos: de
hecho, las primeras plantas conocidas están datadas en torno
a ese periodo. Por tanto, el registro fósil indica que los primeros
animales terrestres pudieron haber sido milpiés o sus ancestros,
que se alimentaban de materia orgánica en descomposición
derivada de plantas, hongos o lechos bacterianos. Su llegada
al medio terrestre tendría lugar anteriormente a la de los fósiles
que hemos encontrado pues ¡tendrían que haber estado ya allí
para fosilizar! De hecho, las primeras evidencias conocidas de
una criatura similar a un milpiés pueden encontrarse en forma
de rastros fosilizados en una roca decenas de millones de años
más antigua. Los milpiés y otros miembros de los miriápodos
(ciempiés y otros dos grupos más pequeños) preceden en
muy poco tiempo a los arácnidos en el registro fósil. El primer
arácnido conocido es un miembro de un grupo desaparecido,
similar al de las arañas, llamado Trigonotarbida (Figura 2) que,
al contrario de las arañas, careció de capacidad para producir
seda y todavía presentaba un abdomen con segmentos, como
vemos en algunas arañas primitivas que aún viven en el sudeste
asiático. Se encontró disolviendo con ácido rocas de unos 419
millones de años de antigüedad y cribando los restos, pues
los exoesqueletos de los artrópodos no se disuelven ni siquiera
en los ácidos más fuertes. Como casi todos los arácnidos son
depredadores, incluso los miembros de ese grupo, parece que
no pasó demasiado tiempo –en términos geológicos– para que
la depredación comenzara en tierra: un hecho destacado por
la existencia de un fósil de ciempiés de hace 418 millones de
años, también depredador, recuperado de la misma manera.
Hace 411 millones de años ya existían, al menos en
sus formas primitivas, los tres grupos más numerosos de
artrópodos que viven hoy en día en el medio terrestre (insectos,
arañas, miriápodos). Lo sabemos gracias a los yacimientos de
fósiles contenidos en las denominadas Rhynie y Windyfiled
Cherts. Estas rocas, situadas cerca las localidades de Rhynie,
Aberdeenshire, en el norte de Escocia, son muy inusuales:
fueron formadas cuando aguas termales ricas en dióxido de
silicio inundaron el ecosistema terrestre primitivo (de modo
Figura 1 – Fotografía y esquema de un milpiés de hace unos 427 millones de años (Cowiedesmus eroticopodus). Esta figura muestra la parte
frontal del fósil, incluyendo su cabeza, collum (primer segmento detrás de la cabeza, sin extremidades) y varios segmentos corporales tras él.
También se muestra una pata modificada (la undécima), que posiblemente se emplease durante la reproducción. Este fragmento mide 12
mm. Esquema de Garwood y Edgecombe 2011, modificado por Wilson y Anderson 2004, y fotografía de Yong Yi Zhen, Museo Australiano.

13
muy parecido a lo que podemos ver actualmente en el
Parque Nacional de Yellowstone en EEUU). El agua petrificó
los animales y plantas reemplazándolos con chert, una roca
compuesta por dióxido de silicio. Aunque esto es muy raro,
cuando sucede puede conservar fósiles con un gran nivel de
detalle: casi hasta a nivel celular. Los Rhynnie y Windyfiled
Cherts han proporcionado: los primeros ancestros de los
insectos; el primer insecto en sentido estricto y, de acuerdo con
partes de una boca que se ha encontrado, posiblemente un
insecto con alas; trigonotárbidos; ácaros y ciempiés. Todo esto
nos lleva hacia el primer fósil de opilión, Eophalangium sheari
(Figura 3) que también procede de Rhyne Chert. Conserva
de su anatomía interna lo suficiente como para para tener la
seguridad de que es un verdadero opilión (de hecho, cuando
se describió el fósil fue situado en un grupo actual, llamado
Eupnoi, del que se hablará más adelante). Conserva elementos
del sistema respiratorio, las extremidades e, incluso, la genitalia.
Así, las Rhynie y Windyfiled Cherts son notables al permitir
atisbar la vida primitiva en el medio terrestre, así como varias de
las primeras apariciones de grupos modernos, incluyendo los
opiliones. Después, se conocen relativamente pocos artrópodos
terrestres fósiles en los siguientes 100 millones de años. Esto
es así porque no se produjeron las condiciones necesarias para
favorecer una conservación extraordinaria. Las excepciones son
algunos maravillosos fósiles terrestres de hace 380 millones
de años recuperados mediante disolución en ácido de rocas
procedentes del estado de Nueva York, así como unos pocos
lugares de Europa donde los animales terrestres se pueden
encontrar aplastados entre capas de esquistos de este periodo
de tiempo. Pero ninguna de ellas contiene opiliones. Hay un
fósil de Opiliones de Escocia, llamado Brigantibunum listoni,
de hace 340 millones de años y –a pesar de una conservación
bastante pobre– parece que puede ser miembro de un grupo
actual. Pero, como se ha indicado al principio, este artículo trata
sobre un grupo particular de animales de un periodo particular
de tiempo en la historia de la Tierra. Ese periodo de tiempo,
llamado Pensilvánico (situado en el Carbonífero tardío) comenzó
hace un poco más de 320 millones de años. De repente, fósiles
Figura 2 – Fotografía y esquema del primer arácnido conocido, un
representante del grupo extinto llamado Trigonotarbida (Palaeotarbus
jerami). El fósil fue recuperado mediante disolución con ácido de la
roca que lo contenía y, dado que le faltan las patas y a su degradada
apariencia, se le dio el apodo de “palomita de maíz quemada”. La
longitud del fósil es de 1,5 mm. Figura modificada de Dunlop, 1996.
14
El Carbonífero
El Periodo Carbonífero abarca desde hace 360 hasta hace
290 millones de años. Se divide en dos partes: el Misisípico
(o Carbonífero Inferior: 359–318 Ma) y el Pensilvánico (o
Carbonífero Superior: 318–299 Ma). La mayoría de las rocas
de su parte más antigua, que es la más extensa, consiste
en calizas (rocas compuestas por carbonato cálcico) de
ambientes marinos someros, y se conocen muy pocos fósiles
de artrópodos (Figura 4, 5) terrestres de este periodo: de
hecho, un único opilión fósil supone la excepción a esta
regla. Sin embargo, la situación cambia por completo en el
Pensilvánico, al menos en términos de rocas que registran vida
terrestre. En Europa y Norteamérica, el Carbonífero tardío
está representado por muchas secuencias repetidas de lutitas,
calizas, areniscas y capas de carbón. Esto es el resultado de
una combinación de factores que nunca hemos vuelto a ver
desde entonces. Durante el Carbonífero aconteció un gran
evento generador de cordilleras, conocido como Orogenia
Varisca o Hercínica. Un gran continente situado en el sur del
planeta, Gondwana, y otro en el norte, Laurasia, colisionaron
y en ese proceso se formó el supercontinente Pangea, que los
dinosaurios poblarían decenas de millones de años después.
Esta colisión también creó un gran cinturón montañoso
ecuatorial (Figura 6). Asociadas a estas montañas variscas, y
tras un descenso del nivel del mar, surgieron muchas áreas
deprimidas que fueron ocupadas por pantanos y bosques.
De hecho, el Carbonífero se describe a menudo como el
primer periodo que tuvo ecosistemas de verdaderas selvas
tropicales. Las plantas de aquellos tiempos nos parecerían
muy sorprendentes. Había organismos relacionados con los
musgos de hoy en día pero con el porte de un árbol; helechos
con gamas de tamaño desde las pequeñas plantas con las que
los asociamos comúnmente hasta el de los grandes árboles; y
parientes de las actuales “colas de caballo” de unos 30 metros
de alto. También caracterizaban estos bosques los helechos
con semillas y parientes primitivas de las coníferas. La mayoría
de esas plantas eran muy ricas en una molécula compleja
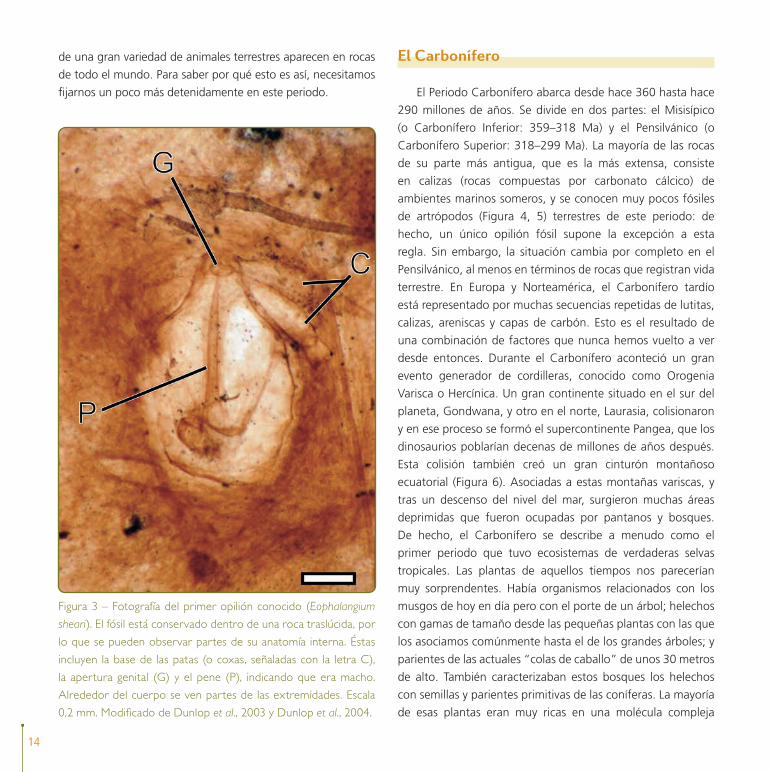
Figura 3 – Fotografía del primer opilión conocido (Eophalangium
sheari). El fósil está conservado dentro de una roca traslúcida, por
lo que se pueden observar partes de su anatomía interna. Éstas
incluyen la base de las patas (o coxas, señaladas con la letra C),
la apertura genital (G) y el pene (P), indicando que era macho.
Alrededor del cuerpo se ven partes de las extremidades. Escala
0,2 mm. Modificado de Dunlop et al., 2003 y Dunlop et al., 2004.
de una gran variedad de animales terrestres aparecen en rocas
de todo el mundo. Para saber por qué esto es así, necesitamos
fijarnos un poco más detenidamente en este periodo.

15
llamada lignina, que contribuye a formar las paredes celulares
de algunas plantas, aportando resistencia a sus tallos. Que
estas plantas fueran ricas en lignina es importante porque la
lignina es muy difícil de descomponer. Incluso actualmente,
después de que muchos animales, como las termitas, han
evolucionado para comer madera, tienen que esforzarse mucho
para digerir la lignina. Esta resistencia a la descomposición
biológica, así como la ausencia de organismos que pudiesen
degradar la lignina, dieron como resultado que los sedimentos
de estos bosques tropicales estuvieran formados por grandes
cantidades de carbón. Especialmente, porque estas regiones
más deprimidas eran propensas a ser rellenadas rápidamente
por sedimentos (por ejemplo, a través del cambio en el
curso de un río o de pequeñas oscilaciones del nivel del
mar). Tales eventos, junto con el bajo nivel de oxígeno, las
condiciones pantanosas y un rápido enterramiento, crearon
las condiciones ideales para producir carbón. De hecho, los
depósitos de carbón del Carbonífero han proporcionado al
periodo su propio nombre y las grandes reservas de carbón
de este periodo de 25 millones de años casi alimentaron por
completo a la revolución industrial en Europa y Norteamérica.
Una enorme cantidad de fósiles de animales terrestres,
las criaturas que vivían en los pantanos y bosques del
Pensilvánico, está asociada con este carbón. Incluyen una serie
de artrópodos y de otros invertebrados, y también algunos
de los primeros vertebrados terrestres (animales que dieron
lugar a los anfibios y a los reptiles, pájaros y mamíferos). Los
ambientes en los que se forma el carbón son también ideales
para el crecimiento de nódulos de carbonato de hierro,
compuestos por siderita (FeCO3), que se desarrollan a partir
de las grandes concentraciones de carbón en el sedimento
y en el agua. Pueden aparecer muy rápidamente después
de la formación y precipitan habitualmente en torno a un
organismo en descomposición que haya sido enterrado en el
sedimento, rodeándolo. Dado que estos nódulos se forman
tan rápidamente, evitan que los organismos sean aplastados,
como sucede con el resto del sedimento. Posteriormente, el
organismo suele descomponerse, dejando un hueco que, a
veces, queda rellenado con otros minerales.
Todo esto significa que tenemos un excelente registro
fósil de la vida terrestre del Pensilvánico. La geografía, el nivel
del mar, las plantas y los sedimentos de este periodo han
generado un gran número de concreciones de carbonato de
hierro, dentro de las cuales encontramos una variedad de los
primeros animales terrestres; por esta razón, el Carbonífero
aporta un conocimiento singular para los paleontólogos que
estudian los primeros ecosistemas terrestres. Hay una ventaja
adicional respecto a que los fósiles estén asociados con las capas
de carbón: con frecuencia son extraídos cuando se explota el
mismo. Otra coincidencia, en este caso histórica, entra aquí en
juego. En Europa, en el siglo XIX, la minería del carbón era una
sólida industria y la paleontología estaba progresando a grandes
zancadas como ciencia. En consecuencia, había un número
elevado de científicos alrededor de las minas en el momento
justo para recolectar estos fósiles y muchos museos todavía
tienen grandes colecciones de tales fósiles de este periodo. Esto
ha llevado a interesarse por depósitos similares de otras partes
del mundo, desde entonces hasta hoy en día. Por lo tanto, no
sólo tenemos fósiles muy bien conservados de esta época –
incluyendo muchos arácnidos– sino que también tenemos una
situación que ha acelerado su descubrimiento. Interesante ¿no?
Sólo hay un problema: los fósiles en 3D son difíciles de gestionar.
La mayoría de los fósiles tiene su origen en la muerte del animal,
que es enterrado en un sedimento y, luego, aplastado dentro del
mismo. La estrategia tradicional de partir una roca en dos mitades
resulta excelente para este tipo de fósiles, puesto que todo lo
que hay que ver de su anatomía se muestra en ese único plano
dividido. La preservación dentro de una concreción, como la
descrita para el Carbonífero, es relativamente rara. Pero cuando
esto sucede, las técnicas tradicionales no son suficientemente
útiles. Si se abre un nódulo, con un fósil 3D dentro de él, en dos
mitades, partes de la anatomía –el extremo de los apéndices, por
ejemplo, o la cabeza– permanecen ocultas en la roca (Figura 7).
Y esto ha limitado tradicionalmente lo que pueden decir los

16
Artrópodos y arácnidos
Los arácnidos constituyen una Clase de animales que
forma parte de un grupo mayor denominado artrópodos.
Los artrópodos son animales definidos por compartir varias
características, como son: un exoesqueleto que se muda
conforme el animal crece; un cuerpo segmentado, con regiones
especializadas llamadas tagmatas; y apéndices articulados.
Otros grupos de artrópodos son los hexápodos (insectos y
unos pocos grupos más pequeños, Figura 4A-C), miriápodos
(ciempiés, milpiés y sus parientes cercanos, Figura 4F), crustáceos
(cangrejos, camarones, percebes, Figura 4G), y los extintos
trilobites. Los arácnidos (Figura 4D-E, 5) son, por su parte,
Figura 4 – Ejemplos de artrópodos actuales. A–C) insectos: A) mariposa, B) abeja melífera, C) un pariente de los saltamontes; D-E)
arácnidos: D) araña, E) ricinúlido, F) milpiés, G) cangrejo de río. Imágenes de: A) Richard Bartz, Munich, Alemania; B-F) Marshal Hedin,
San Diego State University, EEUU.
miembros de un gran subgrupo de artrópodos, los quelicerados.
Este subfilo tiene ese nombre por los apéndices de la boca y
comprende arácnidos, cangrejos cacerola (no emparentados
con los verdaderos cangrejos), probablemente arañas marinas y
varios grupos extintos. La Clase Arachnida consta de 15 grupos
u órdenes más pequeños, incluyendo ácaros y arañas. Otros
órdenes incluyen escorpiones, amblipigios, pseudoescorpiones
y grupos poco conocidos como los ricinúlidos. Los artrópodos
constituyen el linaje de animales más diverso; la mayoría de su
diversidad se debe a insectos y arácnidos pues juntos alcanzan
casi las tres cuartas partes de todos los animales conocidos.

17
Figura 5 – Ilustración de Ernst
Haeckel, erudito alemán del
siglo XIX, que representa la
enorme variedad de formas
de los arácnidos.

18
paleontólogos acerca de los animales. Pero cabe recordar que
estamos hablando de un grupo de animales (opiliones) de un
determinado periodo (el Carbonífero), descubiertos usando una
nueva técnica. Esta técnica es la microtomografía computarizada,
que entra en juego realmente con los fósiles en 3D.
Tomografía computarizada
La tomografía computarizada, o escaneado TAC, es
una técnica no destructiva basada en rayos X. Permite a los
paleontólogos ver dentro de los fósiles y los recrea como
modelos digitales en 3D. Es una herramienta cada vez más
común en el mundo de la paleontología, con la mejora de las
tecnologías de escaneado y de la capacidad de cálculo de los
ordenadores. El escaneado TAC para paleontólogos se basa en
el mismo principio que los escáneres TAC de los hospitales. En
los sistemas médicos una fuente de rayos X y un detector rotan
alrededor de un paciente y toman una serie de radiografías
conforme el paciente se mueve a través de este artilugio
giratorio. Estos escáneres son grandes, y como resultado a
veces son usados en paleontología para fósiles grandes; su
único problema es que estos escáneres están diseñados para
humanos y, por tanto, utilizan fuentes débiles de rayos X para
minimizar la dosis de radiación a los pacientes. A veces, la
fuente no puede penetrar en una roca densa. Una forma más
común de escaneado TAC para fósiles es la microtomografía
de rayos X (TMX o μTM). En este sistema, una muestra (el fósil)
rota aproximadamente 360º entre el emisor de rayos X y un
detector, el cual recibe rayos X de la muestra cada, digamos,
0,1 grados de rotación (Figura 8A). De los más de 3000 rayos
X obtenidos, pueden crearse imágenes de secciones del
fósil mediante la aplicación de algunas ingeniosas fórmulas
matemáticas (Figura 8B). Estas imágenes son, de hecho,
mapas de la atenuación de los rayos X dentro de un objeto,
Figura 6 - Los continentes de la Tierra durante el Pensilvánico. Obsérvese la colisión de los continentes del sur y del norte. En su punto de
encuentro, en el centro, se están formando las cordilleras que se denominan cinturón Varisco. Imagen: Ron Blakey, NAU Geology.

19
es decir, de la cantidad con la que cada parte del fósil reduce
la intensidad de los rayos X. Debido a que a la atenuación es
el resultado de la densidad y de los elementos que componen
cada una de las partes del fósil, estos mapas pueden diferenciar
normalmente al fósil de la roca que lo contiene (y, a veces,
diferentes partes del propio fósil). El resultado consiste en una
serie de imágenes en blanco y negro de secciones transversales
de un fósil. Sin embargo, el proceso de estudiar un fósil en
base a un escaneado TAC no termina ahí: habitualmente se
continúa con el largo y laborioso proceso de hacer un modelo
digital. Hay un buen número de modos diferentes para hacerlo
pero el método usado para crear las figuras que ilustran este
Figura 7 – Un fósil del Pensilvánico en una siderita típica: se trata de un insecto similar a una cucaracha (Archimylacris eggintoni). En este caso,
los apéndices de la boca, las extemidades y el cuerpo permanecen ocultos dentro de la roca. Lo único que podemos observar son las alas
(izquierda) y el escudo de la cabeza-cabeza a la derecha. Modificado de Garwood y Sutton. 2010. Imagen: Natural History Museum, Londres.
20
artículo requiere crear primero imágenes de umbral –una forma
sencilla de convertir píxeles con un cierto rango de niveles de
gris para ser renderizados en 3D, mientras se ignora el resto–.
En el caso de huecos situados en el interior de la roca se deben
eliminar los píxeles más oscuros de los escaneados TAC, dado
que el aire tiene una menor densidad a los rayos X que el
nódulo. Esto creará un modelo 3D del fósil pero también de
cualquier fractura que haya dentro del nódulo y del espacio
que bordea la roca. Para deshacerse de ellos, podemos ir a cada
una de las 2000 imágenes de corte del fósil, coloreando las
diferentes partes de su anatomía –cuerpo, extremidades, piezas
bucales, etc.– y entonces renderizar cada una por separado
en 3D. Mediante este proceso se elimina cualquier artefacto o
las fracturas. Renderizando el modelo en 3D, viendo dónde la
coloración es incorrecta y entonces mejorando cada corte, se
crea finalmente un modelo muy nítido. Al extraer la morfología
del animal completo de la roca –en el caso de nuestro trabajo,
dos opiliones de nódulos del Carbonífero de Francia– la
microtomografía puede decirnos mucho más acerca del modo
de vida del fósil, de su biología y de sus relaciones evolutivas.
Por tanto, en este punto, tiene sentido que nos familiaricemos
con los animales que protagonizarán nuestro artículo.
Opiliones
Los opiliones son arácnidos, miembros de un grupo que
también incluye las arañas, los escorpiones, los ácaros y las
garrapatas (Figura 5). Comparten con otros arácnidos la
presencia de ocho patas y un cuerpo dividido en dos partes
principales: el prosoma (parte anterior) y el opistosoma (parte
posterior). Estas partes de la anatomía diferencian a los
arácnidos de otros animales con los que están relacionados,
como son los insectos (seis pares de patas y cuerpo dividido en
tres partes: cabeza, tórax y abdomen) y de los milpiés/ciempiés
(con muchas patas y cuerpo compuesto por muchos segmentos).
Los miembros del Orden Opiliones se caracterizan por tener las
dos mitades de su cuerpo (es decir, el prosoma y el opistosoma)
unidas por una ancha y, a veces escasamente definida, cintura
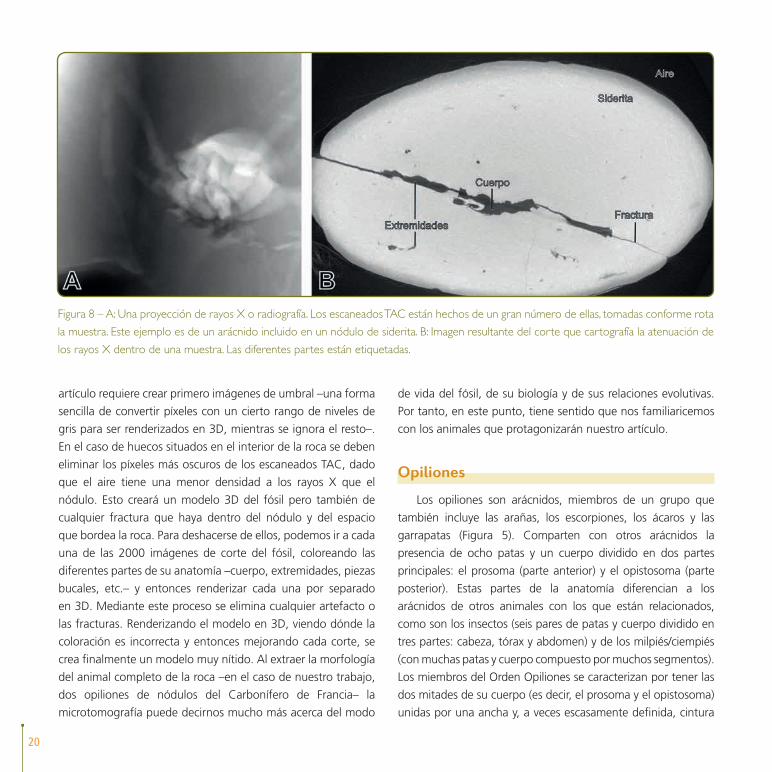
Figura 8 – A: Una proyección de rayos X o radiografía. Los escaneados TAC están hechos de un gran número de ellas, tomadas conforme rota
la muestra. Este ejemplo es de un arácnido incluido en un nódulo de siderita. B: Imagen resultante del corte que cartografía la atenuación de
los rayos X dentro de una muestra. Las diferentes partes están etiquetadas.

21
Figura 9 – Arácnido opilión (Leiobunum aldrichi) con su anatomía
indicada. Obsérvese la división del cuerpo en dos mitades (el
prosoma y el opistosoma) con una unión ancha, las piezas bucales
(pedipalpos y quelíceros) y los apéndices locomotores 1-4 (L1-L4).
Imagen: Marshal Hedin, San Diego Sate University, EEUU.
(Figura 9). Esto contrasta con las arañas, por ejemplo, que
presentan un estrechamiento entre estas dos partes (o tagmas)
del cuerpo. De estos tagmas, el prosoma tiene los cuatro pares
de patas para caminar y los apéndices bucales: los quelíceros
(que son equivalentes a los de las arañas) y los pedipalpos,
que son los mismos apéndices que en los escorpiones han
evolucionado para usarse como pinzas (Figura 10). El opistosoma
no tiene verdaderos apéndices en los arácnidos, y los opiliones
no son una excepción. La mitad posterior tiene las aperturas del
sistema respiratorio, pequeños orificios llamados espiráculos, y
también la abertura genital, o gonostomo, que está cubierta a
menudo por una solapa llamada opérculo. Los machos tienen
pene, mientras que las hembras tienen un ovopositor, que es un
órgano especializado para poner huevos. En muchos grupos, el
segundo par de apéndices locomotores no se usa realmente para
desplazarse; en cambio, es más largo que las otras extremidades
y actúa a modo de antena, palpando para poder percibir el
entorno del animal.
Origen de los nombres
Los opiliones reciben nombres variados en las diferentes
culturas. En inglés el nombre de segadores -también
llamados arañas de la cosecha- puede derivar de que los
opiliones tienden a ser abundantes durante la temporada
de la cosecha. Alternativamente, también puede deberse
a que cuando sus patas están separadas, lo que sucede
comúnmente en algunos grupos como una forma de
defensa, los apéndices describen un movimiento similar
al de la siega. De modo que en varios idiomas se conocen
también como segadores. El nombre en español de agostero
tiene un origen similar, mientras que en algunas provincias
los opiliones son conocidos como pedros, en referencia al
día de San Pedro, que se celebra durante la temporada de la
siega. El nombre en latin del grupo, Opiliones, se estableció
en 1833. El zoólogo Suizo Karl J. Sundevall propuso ese
nombre del latín opilio, que significa maestro ovejero (usado
por el dramaturgo romano Plauto, 254-184 AC) o pastor,
según Virgilio en sus Bucólicas (37 AC). Se ha sugerido
que este origen pudo deberse a las largas patas y al cuerpo
elevado de algunas de las especies europeas, recordando
los opiliones a los antiguos pastores europeos que usaban
zancos para vigilar sus rebaños.
Los opiliones son un grupo grande: dentro de los arácnidos
sólo hay dos grupos más grandes –las arañas y los ácaros– y sólo
hay un grupo mayor y más diverso que los arácnidos, el de los
insectos. Una medida de la diversidad de los opiliones es el número
de especies que han sido descritas hasta la fecha: hay alrededor
de 6400 y las dos primeras ya fueron descritas en 1758 por
Linneo (el inventor del sistema de definición de especies que los
científicos todavía usan hoy en día). Como dato de comparación,
hay aproximadamente 5400 especies conocidas de mamíferos. A
pesar de ser un gran grupo, la biología de los opiliones es todavía
un poco desconocida y quedan muchas cosas por investigar. Los

22
aracnólogos (científicos que estudian los arácnidos) piensan que
tal vez pueden ser de los miembros más primitivos del grupo.
Muchos creen que son los más estrechamente relacionados con
escorpiones, pseudoescorpiones y solífugos. Esta agrupación de
cuatro órdenes se llama Dromopoda, aunque los aracnólogos
todavía debaten acerca de si esto es correcto y, si lo fuera, qué
relaciones puede haber entre los cuatro órdenes. También existe
bastante debate con respecto a las relaciones evolutivas dentro de
los propios opiliones. Los científicos se muestran de acuerdo en
que los opiliones comprenden cuatro linajes distintos (cada uno
de los cuales es técnicamente un suborden). Los Cyphophthalmi
(Figura 11A) probablemente representan el primer suborden,
la primera divergencia con respecto a los otros. Es pequeño (6
familias y 130 especies) pero tiene una distribución global. Los
miembros de este grupo normalmente son pequeños (1–5 mm),
tienen patas pequeñas y habitualmente son ciegos. Tienen un
exoesqueleto grueso y suelen vivir en la hojarasca, aunque
Apodos y mitos de los opiliones
En muchas partes de Estados Unidos, los opiliones se
llaman “papá piernas largas”, o “abuelito piernas largas”
por lo que, a veces, son confundidos en Europa con los
tipúlidos, que tienen el mismo apodo en las costas europeas.
Algunos apodos para los opiliones en diferentes países
reflejan el hecho de que se parecen a las arañas pero sin ser
verdaderos miembros del grupo (por ejemplo, Afterspinne
en Alemania, que significa falsa araña). En Estados Unidos
y Australia, una leyenda urbana muy común insinúa que el
grupo es extremadamente venenoso pero que sus piezas
bucales son incapaces de atravesar la piel humana. Como
muchas leyendas urbanas, es completamente falsa: los
opiliones no tienen veneno.
Figura 10 – Los pedipalpos (en azul) y los quelíceros (en rojo) indicados en dos arácnidos que no son opiliones: una araña y uno de los
primeros escorpiones fósiles. Obsérvese también que ambos tienen cuerpos que pueden dividirse en prosoma y opistosoma (uno de los
rasgos que define a un arácnido).

23
algunos viven en cuevas. Los otros tres linajes restantes de
opiliones se agrupan frecuentemente en un linaje llamado
Phalangida. El suborden Eupnoi (6 familias, aproximadamente
1800 especies, Figura 11C) incluye los opiliones más fácilmente
reconocibles: criaturas con pequeños cuerpos blandos
colgando bajo unas patas muy largas. Este grupo se encuentra
también en la mayor parte del mundo. Otro suborden,
Dyspnoi (7 familias, 290 especies, Figura 11B) muestra una
gran variedad de formas que están unificadas por tener un
par de dientes pequeños en los quelíceros. Este suborden se
encuentra ampliamente extendido en el Hemisferio Norte. El
último grupo es el de los Laniatores, el suborden más grande
(27 familias, aproximadamente 3750 especies, Figura 11D)
que, a menudo, son de colores vivos y tienen muchas espinas
pero están principalmente restringidos a las regiones tropicales
y templadas del Hemisferio Sur. No tenemos completamente
establecidas las relaciones entre estos tres subórdenes y dos
posibles ordenaciones se muestran en la Figura 12.
Figura 11 – Ejemplos de cada uno de los cuatro linajes de opiliones. A: Cyphophthalmi. B: Dyspnoi. C: Eupnoi. D: Laniatores. Imagen: Marshal
Hedin, San Diego State University, EEUU.

24
Estas encantadoras criaturas son bastante variadas en su
ecología y modos de vida. Muchas viven en suelos y hojarasca, en
lugares oscuros y húmedos tales como bajo las rocas y en cuevas.
Algunas, sin embargo, viven en paredes y troncos de árboles y
varias lo hacen entre la vegetación. Una consideración particular
de los opiliones es la humedad del lugar donde están viviendo,
ya que muchas especies son poco eficaces en la reducción de
la pérdida de agua. Los ambientes húmedos y oscuros ayudan
a evitar cualquier contratiempo que, de otra forma, surgiría
al perder demasiada agua. Encontramos la mayor diversidad
de los opiliones en ambientes tropicales, especialmente en
las pluviselvas. Siendo omnívoros, los opiliones tienen una
alimentación muy diversificada: pequeños invertebrados, plantas,
carroña y hongos. ¡Incluso han llegado a sobrevivir en la espuma
de un cappuccino cuando se escaparon en el laboratorio! Tener tal
variedad en su dieta es inusual ya que los otros arácnidos son casi
todos puramente depredadores y tienden a digerir su alimento
por vómito de enzimas digestivas sobre él, antes de filtrarlo y
succionarlo usando un estómago bombeador. Los opiliones
son depredadores que tienden a emboscar a sus presas u,
ocasionalmente, dejar que se agoten. Su sistema de alimentación,
sin embargo, tiene problemas: debido a que los opiliones utilizan
sus apéndices bucales –que terminan en pinzas– para romper la
comida en pequeños pedazos antes de comérsela (en lugar de
succionarla como un líquido, lo que permitiría filtrar el alimento),
son muy propensos a ingerir parásitos. Además, los opiliones
a menudo son presas de pájaros, anfibios, pequeños reptiles
e incluso arañas. Pero no están completamente indefensos:
algunas especies segregan una sustancia pegajosa y adhieren
partículas sobre su superficie externa para camuflarse. Otros
fingen estar muertos si son atacados y algunos se mueven
arriba y abajo para confundir a los potenciales depredadores.
Todos los opiliones tienen orificios para glándulas que emiten
olores desagradables en la mitad anterior de su cuerpo (otro
mecanismo de defensa). Finalmente, muchos pueden perder
sus patas; por ejemplo, si han quedado atrapadas en cualquier
lugar. Esta estrategia defensiva se conoce como apendoctomía.
Los apéndices perdidos no vuelven a crecer, pero la ruptura tiene
lugar a lo largo de unas líneas de debilidad. El resultado, aparte
de la pérdida de una pata (lo que no parece tener demasiado
impacto en la criatura), no ocasiona muchos problemas ya que
los músculos de la pata cierran la herida.
Figura 12 – Dos posibles relaciones entre los diferentes subórdenes de opiliones, ambas con Cyphophthalmi como suborden basal, pero que
difieren en la disposición de los otros tres grupos. Fotografías: Marshal Hedin, San Diego State University, EEUU.

25
mediante cinta adhesiva. Después del escaneado, y tras una
hora de procesamiento para convertir las simples radiografías en
secciones, obtuvimos 2000 imágenes mostrando las secciones
trasversales de los fósiles. En uno de los conjuntos de datos cada
píxel representó un cuadrado de 26,5µm x 26,5µm y, en el otro,
de 14,2µm x 14,2µm. En cada una de las imágenes de secciones
trasversales, los fósiles aparecían como formas oscuras, debido a
que eran menos densos a los rayos X que la roca que los contenía.
Utilizamos un software personalizado para reconstruir los fósiles
en 3D, como describimos en la sección de microtomografías.
Esto fue un proceso un poco lento: requería hacer conjeturas
acerca de lo que estábamos viendo en las imágenes de las
secciones, renderizar el modelo en 3D y entonces mejorar la
coloración. Sin embargo, finalmente creamos los modelos que se
presentan en este artículo: dos modelos de opiliones fósiles con
muy buen aspecto y notablemente completos. Para confeccionar
las imágenes de este trabajo, exportamos estos modelos como
un archivo que registra la forma de las superficies en 3D y lo
importamos en un programa CGI de código libre llamado
Blender, lo que permitió obtener imágenes y animaciones con
sombras e iluminación realistas.
Otra gran cuestión que se plantea cuando tratamos la biología
de criaturas tales como los opiliones es cómo se reproducen. La
mayoría de ellos lo hacen sexualmente: se necesita un macho
y una hembra de la misma especie para generar crías fértiles
(aunque hay unas pocas especies que se ha observado que se
multiplican asexualmente). Los opiliones emplean fertilización
interna; en la mayoría de los casos (todos los grupos excepto
los Cyphophthalmi) esto significa que los machos tienen un
pene y que una pareja copula para criar. Después de la cópula,
las hembras pueden poner huevos inmediatamente o tras un
periodo de tiempo. Usan su ovopositor para hacerlo y los huevos
eclosionarán después en un periodo de tiempo que varía entre
unos pocos días y varios meses. La mayoría de las especies
muestra diversos tipos de cuidado parental para los huevos/crías.
Podemos concluir de esta rápida introducción sobre los
opiliones que se trata de un grupo de criaturas realmente
interesante. Hasta ahora hemos introducido el periodo de
tiempo del que proceden estos fósiles (el Pensilvánico), el
método usado para estudiarlos (microtomografía de rayos X)
y los animales que han fosilizado (los opiliones). Así que ahora
presentaremos nuestra investigación.
La investigación
Fuimos bastante afortunados al encontrar dos nuevos fósiles de
opiliones que permitieron llevar a cabo esta investigación. Ambos
estaban en el interior de nódulos de siderita, provenientes de un
yacimiento llamado Montceau-Les-Mines, en el Macizo Central
de Francia. Sus cuerpos apenas se apreciaban en la superficie
visible tras romper el nódulo para abrirlo; incluso uno de ellos
no se identificó que era un opilión hasta que no lo escaneamos.
Ambos estaban depositados en el Muséum d’Histoire Naturelle
d’Autun y nos fueron prestados para estudiarlos. Para conseguir
revelar la anatomía oculta en la roca, llevamos a cabo escaneados
de ambos fósiles en un escáner TAC en el Natural History
Museum de Londres (Figura 13). Para ello juntamos de nuevo
las mitades de los nódulos partidos y las mantuvimos unidas
Figura 13 – El escáner TAC usado para este estudio, que puede
escanear muestras con tamaños desde unos pocos milímetros hasta un
cuarto de metro. La habitación donde se encuentra el equipamiento
está forrada de plomo para proteger del escape de radiación.
26
Los fósiles
Los resultados de nuestro trabajo –o, al menos, de su
mayor parte– son los modelos 3D que se pueden ver en las
figuras. Son bastante diferentes de cualquier otro opilión
que conozcamos de una edad similar, por lo que estábamos
capacitados para describirlos como especies nuevas. Esto
significaba que los dos necesitaban un nombre; dos, de
hecho, ya que también necesitaban un nombre para el género.
Tradicionalmente, estos nombres tienen su origen en el latín o
el griego, ajustándose al sistema que fue implantado a partir
de los trabajos de Linneo en el siglo XVIII.
Ameticos
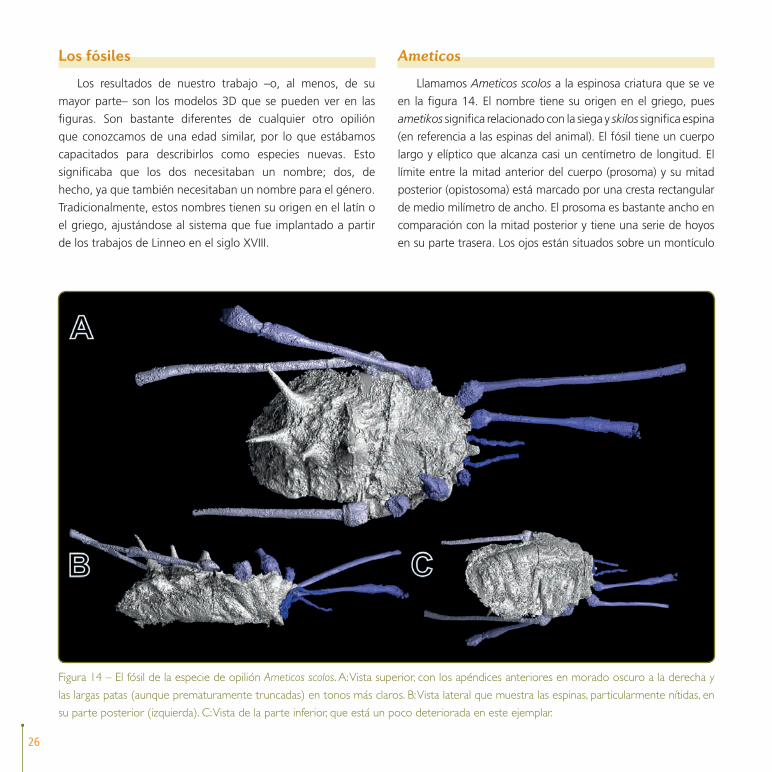
Llamamos Ameticos scolos a la espinosa criatura que se ve
en la figura 14. El nombre tiene su origen en el griego, pues
ametikos significa relacionado con la siega y skilos significa espina
(en referencia a las espinas del animal). El fósil tiene un cuerpo
largo y elíptico que alcanza casi un centímetro de longitud. El
límite entre la mitad anterior del cuerpo (prosoma) y su mitad
posterior (opistosoma) está marcado por una cresta rectangular
de medio milímetro de ancho. El prosoma es bastante ancho en
comparación con la mitad posterior y tiene una serie de hoyos
en su parte trasera. Los ojos están situados sobre un montículo
Figura 14 – El fósil de la especie de opilión Ameticos scolos. A: Vista superior, con los apéndices anteriores en morado oscuro a la derecha y
las largas patas (aunque prematuramente truncadas) en tonos más claros. B: Vista lateral que muestra las espinas, particularmente nítidas, en
su parte posterior (izquierda). C: Vista de la parte inferior, que está un poco deteriorada en este ejemplar.
27
(el oculario) que se sitúa en la parte más frontal del animal,
donde hay una pequeña proyección (la estructura que se parece
un poco a una pequeña nariz). La parte inferior de la criatura
está muy fracturada pero podemos ver la base de sus patas en
uno de los lados. Los apéndices bucales –los quelíceros– son
pequeños (0,8 mm de longitud) y tienen tres segmentos pero,
por otra parte, no se ven bien. Probablemente, se debe a que no
están bien conservados y no a que no hayamos podido registrar
sus detalles en nuestro escaneado TAC. El segundo par de
apéndices (pedipalpos) son también bastante pequeños y han
perdido la uña de su extremo final; podemos distinguir los seis
segmentos que componen un apéndice de 2,3 mm de longitud.
La pata más completa del fósil es la tercera izquierda, que tiene
cuatro segmentos conservados y casi 9 mm de longitud. Las otras
extremidades parecen haberse desprendido entre el segundo y
el tercer segmento (el trocánter y el fémur, respectivamente).
Desplazándonos hacia el dorso de la criatura, lo más obvio son
los tres pares de espinas prominentes (cada una de 2,4 mm de
longitud). Esto hace que la criatura completa tenga alrededor de
medio centímetro de altura. Podemos ver cinco segmentos en el
opistosoma y las espinas se encuentran sobre los tres primeros.
Toda la superficie superior está recubierta por hoyos profundos
y esféricos de alrededor de un cuarto de milímetro de diámetro.
En la parte inferior también podemos distinguir cinco segmentos
en el opistosoma, cada vez más cortos hacia la parte posterior.
Macrogyion
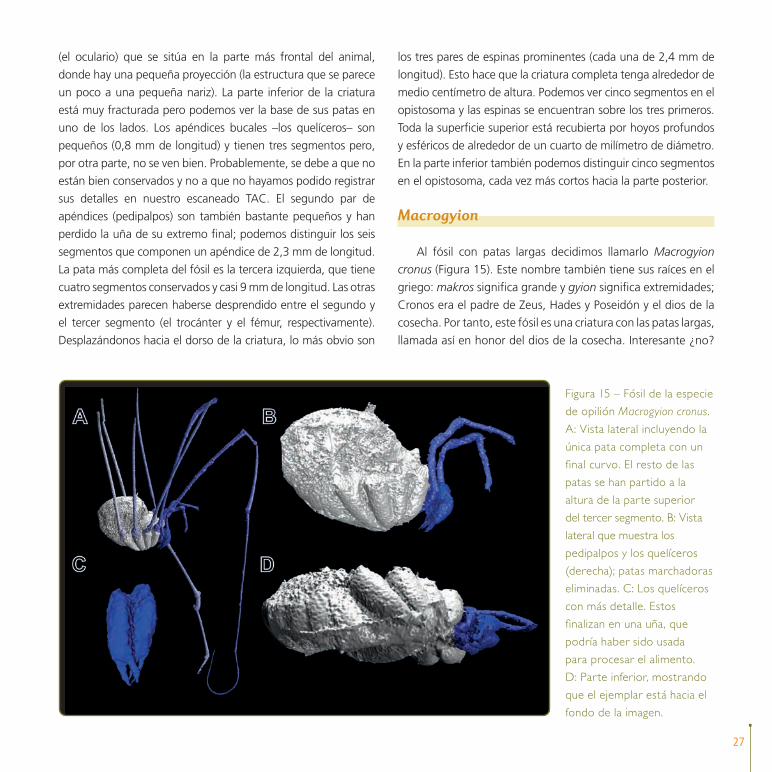
Al fósil con patas largas decidimos llamarlo Macrogyion
cronus (Figura 15). Este nombre también tiene sus raíces en el
griego: makros significa grande y gyion significa extremidades;
Cronos era el padre de Zeus, Hades y Poseidón y el dios de la
cosecha. Por tanto, este fósil es una criatura con las patas largas,
llamada así en honor del dios de la cosecha. Interesante ¿no?
Figura 15 – Fósil de la especie
de opilión Macrogyion cronus.
A: Vista lateral incluyendo la
única pata completa con un
final curvo. El resto de las
patas se han partido a la
altura de la parte superior
del tercer segmento. B: Vista
lateral que muestra los
pedipalpos y los quelíceros
(derecha); patas marchadoras
eliminadas. C: Los quelíceros
con más detalle. Estos
finalizan en una uña, que
podría haber sido usada
para procesar el alimento.
D: Parte inferior, mostrando
que el ejemplar está hacia el
fondo de la imagen.

28
Evaluando las relaciones evolutivas
Cuando se tiene un fósil, una de las primeras cosas
que se desea resolver es con qué otros animales está
relacionado. Los organismos vivos tienen una estructura
jerarquizada: organismos con un parentesco cercano
se parecen más entre sí que a aquellos otros con los que
están menos estrechamente relacionados. Sobre esta
base, los paleontólogos han situado tradicionalmente a
los fósiles dentro de grupos que abarcan diferentes niveles
de semejanza. Por ejemplo, los miembros de una Familia
(tales como Hominidae: los grandes simios, incluyendo a
los humanos, chimpancés, bonobos, gorilas y orangutanes)
compartirán un número mayor de características que las
familias dentro de un Orden (como Primates, que incluye
también monos y lémures, entre otros). Sin embargo, esas
familias todavía compartirán un número de características
que definen al Orden. A su vez, los órdenes son los miembros
de una Clase, un grupo definido por otro conjunto de
caracteres compartidos (probablemente menos específico).
Este camino continúa hacia el nivel de Filo o Reino, que son
las divisiones verdaderamente gruesas de la vida.
Los paleontólogos modernos todavía usan este sistema;
los fósiles que hemos descrito aquí, por ejemplo, pertenecen
al Orden Opiliones y a la Clase Arachnida. Sin embargo,
resolver las relaciones evolutivas sólo en base al parecido
anatómico puede ser algo subjetivo. Por esta razón, muchos
paleontólogos usan ahora la cladística para entender las
relaciones evolutivas, o filogenia, de un grupo. Esto implica
crear una lista de aspectos diferentes de la anatomía de un
animal. Por ejemplo, se podría usar el número de patas, la
presencia (o ausencia) de ojos, si las patas tienen un número
determinado de subsegmentos, y un enorme número de
otros aspectos de la naturaleza de un organismo, como
caracteres. Una selección de los animales en cuestión son
entonces codificados con estos caracteres; por tanto, por
ejemplo, el número de patas podría ser introducido en una
lista para todos los animales, como se podría incluir también la
presencia (codificado como 1) o la ausencia (codificado como 0)
de ojos. Esto crea una matriz de un gran número de caracteres
para un gran número de especies, que registra en esencia las
similitudes y diferencias entre los animales codificados. Con
esta matriz, se puede usar un algoritmo informático para
resolver qué árbol evolutivo (o árboles, en su caso) es el que
implica el menor número de cambios en los caracteres. Por
tanto, en el caso de los ojos, si a 5 de las 20 especies en el
análisis le faltan los ojos y forman un único grupo, los ojos
solamente sería necesario perderlos en una ocasión. Otro
ejemplo se muestra en la Figura 16, con exactamente la misma
comparación pero usando otro carácter. De este modo, un
gran número de caracteres diferentes pueden ser usados para
tratar de elaborar un árbol evolutivo factible: el que implica el
menor número de cambios. Hacer esto, por supuesto, exige
asumir que el menor número de cambios proporcionará el más
plausible de los árboles evolutivos. Pero el uso de esta técnica
permite proporcionar hipótesis que se pueden contrastar y
verificar. Esto nos permite demostrar cómo de bien –o de mal–
está soportada cualquier agrupación y, a menudo, cuáles son
las alternativas. Si hay un número pequeño de árboles de igual
longitud, podemos elaborar un árbol de consenso (un árbol que
sólo tiene los pedacitos en los cuales todos los otros árboles
más cortos están de acuerdo).
En los últimos años se ha comenzado a utilizar la misma
técnica con DNA de una serie de organismos para tratar de
evaluar cuán estrechamente relacionados pueden estar. Cuanto
más tiempo hace que dos organismos comparten un ancestro

29
Figura 16 – Ejemplo de un solo carácter cuyos cambios están marcados en dos árboles posibles. Los grupos de opiliones Laniatores y
Dyspnoi tienen hembras que carecen de articulaciones en su ovopositor (el órgano que usan para poner los huevos). Si asumimos que
ésta es la condición primitiva para los opiliones, estas articulaciones deberían de haberse perdido en algún lugar en el camino entre
Laniatores y Dyspnoi. Si los dos no están estrechamente relacionados, como se ve a la izquierda, esto debería haber pasado una vez
en cada linaje. Sin embargo, si están más estrechamente relacionados entre sí que con el resto de opiliones, esta pérdida podría haber
sucedido sólo una vez, en el ancestro de ambos grupos. Así, al menos para este carácter, el árbol de la derecha tiene menos pasos. Si se
tiene un gran número de caracteres, se pueden usar comparaciones como éstas para construir un árbol con el menor número de pasos,
y proporcionar una hipótesis contrastable para las relaciones evolutivas. Fotografías: Marshal Hedin, San Diego State University, EEUU.
común (es decir, cuanto más tiempo hace que sus líneas se
dividieron) mayor tiempo han tenido para que mutaciones
se incorporen a su DNA. De acuerdo con esto, si se pueden
comparar dos de las mismas cadenas de DNA, los animales más
estrechamente relacionados tendrán menos diferencias que los
menos estrechamente relacionados y, de nuevo, esto puede
usarse para elaborar un árbol. También se pueden combinar
estas dos aproximaciones, permitiendo codificar la morfología
tanto de fósiles como de organismos vivos y entonces usar el
DNA de las especies actuales para ayudar a construir el árbol.
Hemos usado esta aproximación aquí para intentar resolver las
relaciones de nuestros dos fósiles.

30
El fósil es más pequeño que Ameticos pues tiene un cuerpo
ovalado de unos 6,5 mm de longitud. Es difícil decir dónde
está el límite prosoma/opistosoma y, de nuevo, partes de
las bases de las patas y la zona inferior del opistosma están
parcialmente rotas. El cuerpo tiene también aproximadamente
medio centímetro de altura, por lo que es más esférico que
el de Ameticos, que es más plano. En este caso los quelíceros
están muy bien conservados. Tienen unos 3,3 mm de largo y
se componen de tres segmentos que terminan en una pinza.
Están situados entre unos pedipalpos, bien conservados,
situados en alto y al frente del prosoma, y que finalizan en
una uña simple. Los seis segmentos de los pedipalpos están
bastante claros y la extremidad completa está cerca de los 8
mm de longitud. Las patas están generalmente rotas una vez
más pero la primera izquierda está conservada en sus 2,6 cm
completos; sus primeros tres segmentos apuntan hacia arriba,
luego hay un segmento horizontal tras una curva y luego otra
curva más; el resto del apéndice se orienta hacia abajo desde
aquí. La última parte de la pata está curvada, sugiriendo que
está compuesta por un gran número de sub–segmentos, algo
que vemos en varios grupos de opiliones actuales. El resto de las
patas se han roto en diferentes lugares y están mucho menos
completas. El prosoma está un poco aplastado también pero la
parte superior no ha cedido como la inferior. Tiene oculario, un
montículo que, presumiblemente, sostenía los ojos en vida. Los
primeros segmentos del opistosoma están fusionados y uno de
ellos tiene una pequeña cresta sobre él, aunque tres de ellos se
distinguen claramente en el dorso del animal. La segmentación
está bien conservada en la parte inferior: hay cinco segmentos
claros, el primero mucho más largo que los que le siguen.
Métodos cladísticos
Para resolver las relaciones evolutivas de estos dos fósiles,
hemos desarrollado un análisis cladístico (ver Evaluando las
relaciones evolutivas; pág. 28). Para ello usamos 201 caracteres
morfológicos, codificando todas las partes diferentes de la
anatomía de los opiliones. Después usamos un software
llamado TNT para calcular el árbol que requería el menor
número de cambios anatómicos. Tuvimos en cuenta varios
aspectos diferentes de la técnica para evaluar las relaciones.
Estos incluyen dar a todos los caracteres la misma importancia
(“mismos pesos”) y dar menos importancia (“pesos implícitos”)
a los que parecen ser menos fiables (es decir, que se invierten
más en muchos de los árboles). También demostramos si el
árbol estaba bien soportado, usando una medida llamada
remuestreado simétrico. Además de esta matriz morfológica,
usamos el DNA de las especies actuales para ayudar a construir
el árbol.
Resultados cladísticos
Todo este análisis nos dio algunos resultados interesantes
y novedosos, que perfilaremos aquí pero cuya importancia
discutiremos después. El análisis incluyó arácnidos no opiliones
que fueron usados para enraizar el árbol; es decir, para resolver
qué caracteres eran más similares a los de otros arácnidos y,
por lo tanto, probablemente los grupos con esos caracteres
se separaron antes. Usando solo la morfología de las especies
actuales y de los fósiles, analizándola con los mismos pesos,
obtuvimos una serie de agrupaciones. Éstas incluían a todos los
opiliones –los primitivos Cyphophthalmi y Phalangida (todos
los opiliones excepto los Cyphophthalmi)– compartiendo un
único ancestro común que definía cada grupo (Figura 17).
Los fósiles, a pesar de ser situados en Phalangida, no
tenían una posición clara: si el análisis se ejecutaba sin
los fósiles, tanto Eupnoi como Dyspnoi aparecían como
verdaderos grupos. Pero al añadir nuestros fósiles se cambiaba
el resultado cladístico: con M. cronus los Eupnoi ya no se
obtenían como un grupo verdadero y A. scolos tenía el
mismo efecto en los Dyspnoi cuando hacíamos un árbol de
consenso. En otras palabras, en todos los árboles que tenían
la misma longitud, los fósiles fueron encontrados en tantas
posiciones que relativamente pocas de las relaciones evolutivas
obtenidas eran compartidas entre los árboles. Los fósiles han
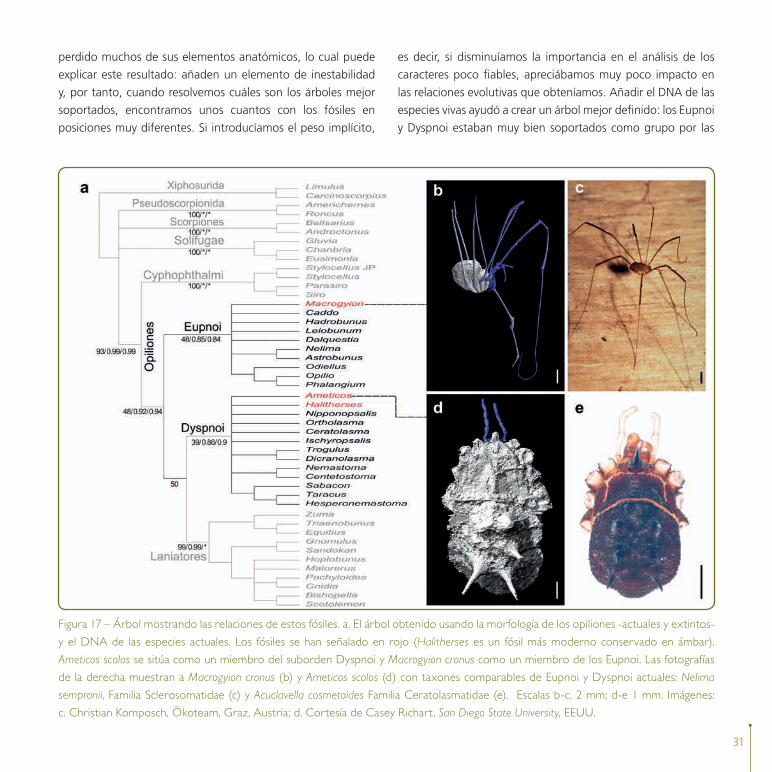
31
perdido muchos de sus elementos anatómicos, lo cual puede
explicar este resultado: añaden un elemento de inestabilidad
y, por tanto, cuando resolvemos cuáles son los árboles mejor
soportados, encontramos unos cuantos con los fósiles en
posiciones muy diferentes. Si introducíamos el peso implícito,
es decir, si disminuíamos la importancia en el análisis de los
caracteres poco fiables, apreciábamos muy poco impacto en
las relaciones evolutivas que obteníamos. Añadir el DNA de las
especies vivas ayudó a crear un árbol mejor definido: los Eupnoi
y Dyspnoi estaban muy bien soportados como grupo por las
Figura 17 – Árbol mostrando las relaciones de estos fósiles. a. El árbol obtenido usando la morfología de los opiliones -actuales y extintos-
y el DNA de las especies actuales. Los fósiles se han señalado en rojo (Halitherses es un fósil más moderno conservado en ámbar).
Ameticos scolos se sitúa como un miembro del suborden Dyspnoi y Macrogyion cronus como un miembro de los Eupnoi. Las fotografías
de la derecha muestran a Macrogyion cronus (b) y Ameticos scolos (d) con taxones comparables de Eupnoi y Dyspnoi actuales: Nelima
sempronii, Familia Sclerosomatidae (c) y Acuclavella cosmetoides Familia Ceratolasmatidae (e). Escalas b-c, 2 mm; d-e 1 mm. Imágenes:
c. Christian Komposch, Ökoteam, Graz, Austria; d. Cortesía de Casey Richart, San Diego State University, EEUU.

32
evidencias de su DNA. Con el DNA, y con mismos pesos para
los caracteres, M. cronus fue identificado como un miembro
de los Eupnoi y A. scolos como Dyspnoi. Comprobamos este
resultado usando unas pocas técnicas más para evaluar las
relaciones evolutivas de los fósiles, incluyendo una técnica
estadística conocida como inferencia bayesiana. Todos los
análisis que hicimos confirmaban esa misma disposición
de los fósiles. Para todos estos métodos de situación de los
fósiles, aportamos un indicador del grado de soporte del árbol;
cuando se incluían los fósiles, éste era habitualmente bajo,
pues muchos rasgos anatómicos de estos animales cuando
estaban vivos, tales como los espiráculos (orificios del sistema
traqueal), la genitalia, o los ojos no pueden ser determinados
con certeza en cada fósil, ya que son demasiado pequeños o
bien son internos y no pueden encontrarse con este tipo de
conservación. Esto incluye la mayoría de los caracteres usados
para definir cada suborden de los opiliones. No obstante, el
hecho de que en todos los análisis obtuviésemos esencialmente
el mismo resultado sugiere que la posición para los fósiles es
bastante fiable dentro del árbol de la vida de los opiliones.
¿Qué significa esto?
Por tanto tenemos dos nuevos fósiles, reconstruidos en 3D,
y hemos resuelto sus relaciones probables con otros opiliones.
Esto nos permite crear unas reconstrucciones precisas de su
aspecto en vida (Figura 18) y decir algo sobre la paleobiología
de ambas especies fósiles (como, por ejemplo, cómo vivieron).
Podemos usar también su descubrimiento para decir algo
sobre la historia de los opiliones en conjunto. Dado que estos
ejemplares están tan bien conservados (y esto es inusual
en fósiles de opiliones), se trata de la primera vez que unos
fósiles con más de unas pocas decenas de millones de años
han sido situados en el árbol de los opiliones en base a un
análisis cladístico. Como anteriormente se ha esbozado, los
opiliones pueden dividirse en cuatro subórdenes (Eupnoi,
Dyspnoi, Laniatores y Cyphophthalmi) y nuestro trabajo ha
situado a Ameticos en Dyspnoi y a Macrogyion en Eupnoi. Esto
significa que ambos subórdenes, Dyspnoi y Eupnoi, deben de
haber existido desde hace 305 millones de años, cuando estos
fósiles fueron depositados en una delgada lámina de agua
en uno de los bosques de carbón del Carbonífero. Ya se ha
sugerido que los Eupnoi existen desde hace 411 millones de
años en base a los opiliones mencionados anteriormente de
las Rhynie Chert, como fue inferido esencialmente a partir de
su morfología genital, aunque datos actuales basados en un
nuevo fósil contradicen la posición del opilión de Rhynie Chert.
La confirmación, usando una gran variedad de observaciones
anatómicas, de que existió un Eupnoi (Macrogyion) en
el Carbonífero añade peso a la propuesta de que Eupnoi
constituye un suborden antiguo. Por el contrario, el fósil más
antiguo de Dyspnoi situado mediante un análisis cladístico
antes de nuestra publicación sólo tiene pocas decenas de
millones de años de antigüedad. Así, Ameticos no sólo es el
miembro más antiguo conocido, sino que mueve un buen
trecho hacia atrás a los primeros fósiles conocidos del grupo. De
igual manera que se usan las similitudes en DNA para resolver
las relaciones entre grupos, el DNA puede también usarse
para estimar cuándo se dividen grupos diferentes, el uno con
respecto al otro, basándose en la frecuencia con la que ocurren
las mutaciones. Varios biólogos han usado esta técnica en la
última década para estimar la edad de diferentes grupos de
opiliones y nuestros dos fósiles corroboran tales estimaciones.
Las relaciones que descubrimos también aclaran que los dos
subórdenes de opiliones –ambos con representantes actuales
todavía– ya estaban presentes en los bosques de Europa hace
305 millones de años. En contraste, en aquel tiempo, otros
arácnidos como las arañas y los escorpiones no estaban
presentes (al menos en la forma que los vemos hoy en día).
Más bien, tienden a ser reconocidos sólo en sus formas
más primitivas: linajes que ahora están extintos. Los ácaros
son los únicos arácnidos que sabemos que eran similares
anatómicamente en aquel momento a las formas modernas.
Por tanto, nuestro trabajo proporciona evidencias convincentes
acerca de un origen temprano de los opiliones, pues parece

33
probable que estén entre las primeras ramas de los arácnidos.
Más allá de estos orígenes tan tempranos, nuestros dos fósiles
son esencialmente modernos en apariencia; esto demuestra
que el plan corporal de los opiliones fue establecido en el
Carbonífero tardío y que han permanecido mucho tiempo sin
cambiar desde hace 305 millones de años. Son un excelente
ejemplo de estasis evolutiva.
Podemos usar esta similitud para hacer conjeturas
fundamentadas acerca de cómo nuestros fósiles pudieron
haber vivido. Ameticos scolos presenta un parecido
llamativo con algunos miembros actuales de los Dyspnoi
(particularmente del género Acuclavella –miembros de una
familia llamada Ceratolasmatidae–; Figura 17e). El segundo
par de patas de nuestro fósil espinoso era probablemente
alargado, rasgo en común con Acuclavella. Sugerimos que
este es el caso porque incluso aunque el segundo par de
patas estén rotas por la mitad a través del tercer segmento,
el fémur, ese fragmento ya es más largo que los fémures
de todas las otras patas. Conocemos que en las formas
modernas de opiliones –como Acuclavella– estas patas son
usadas para palpar delante del animal cuando camina, un
poco a modo de antena. El hecho de que Ameticos tenga
unos hoyos profundos en su superficie superior sugiere un
Figura 18 – Reconstrucción idealizada de Ameticos scolos (arriba) y Macrogyion cronus (abajo), mostrando el aspecto en vida de ambas
especies. Escala 5 mm.

34
exoesqueleto robusto y bien construido, lo que implica que
la criatura es más probable que haya vivido sobre el suelo del
bosque que en los árboles. Acuclavella, el Dyspnoi actual, vive
en los márgenes de aguas perennes poco extensas, en restos
de madera húmedos, y pensamos que sus espinas pueden
proporcionarle defensa frente a depredadores anfibios. Del
mismo modo, nuestros fósiles podrían haber habitado zonas
similares de los bosques húmedos de carbón ricos en detritos
del Carbonífero y, probablemente, tenían espinas por las
mismas razones. Podemos hacer el mismo tipo de inferencias
para Macrogyion cronus, que es similar en forma a los Eupnoi
actuales de las familias Phalangiidae y Sclerosomatidae
(Figura 17c). Por ejemplo, las proporciones que vemos en
los segmentos de las extremidades, la forma y el tamaño del
cuerpo y los apéndices bucales son muy similares. El hecho
de que el cuerpo de los fósiles esté parcialmente roto y peor
conservado que las extremidades sugiere que –como en
los miembros modernos de estas familias– el exoesqueleto
de estos fósiles tenía probablemente una cutícula blanda y
curtida en vida. La pata completa muestra que el animal poseía
muchos subsegmentos al final de las patas locomotoras, que
le habrían permitido enrollarlos cuando fuera necesario. Esto
es una condición típica de los miembros actuales de Eupnoi
y una posible adaptación hacia la vida entre la vegetación,
permitiendo al animal agarrarse mejor a los tallos de las
plantas. Esta sugerencia está respaldada por sus extremidades
muy alargadas, que en las especies modernas son indicativas
de una vida por encima del nivel del suelo. De acuerdo con
esto, Macrogyion podría haber tenido su morada en la
vegetación o en los árboles.
Conclusiones
Esperamos haber aportado un trabajo interesante y que
nuestra introducción a elementos de paleobiología, cladística,
microtomografías, opiliones y la historia de la vida terrestre
tenga sentido… ¡Hemos abarcado mucho terreno! Querríamos
finalizar resumiendo las cosas que hemos mencionado.
Hemos aprendido que los primeros animales terrestres fueron
artrópodos. Los artrópodos incluyen a los arácnidos y los
opiliones son sólo un grupo de estos. Son bastante raros
como fósiles. Sin embargo, debido a un conjunto especial
de condiciones en el Periodo Carbonífero, muchos animales
terrestres de este tiempo –incluyendo nuestros dos nuevos
fósiles– están conservados en tres dimensiones. Los fósiles
conservados en esas condiciones pueden ser problemáticos:
usando técnicas tradicionales, los paleontólogos se están
dejando partes del organismo enterradas dentro de la roca.
La microtomografía puede superar esto utilizando rayos X
para obtener imágenes de secciones que pueden, a su vez,
ser usadas para crear un modelo en 3D de los fósiles. Esto
nos ha permitido reconstruir dos opiliones fósiles de hace 305
millones de años y usar los modelos resultantes para intentar
evaluar sus relaciones evolutivas. El presente trabajo sugiere
que los opiliones son uno de los primeros grupos de arácnidos
en diversificarse y que también son un excelente ejemplo
de estasis evolutiva. Los modelos y las comparaciones con
opiliones vivos nos han permitido proponer que una especie
tenía su morada en el suelo y la otra vivió en la vegetación
o en los árboles. Todo eso, a partir de dos pequeños fósiles
escondidos en una roca. ¡No está mal! ¿eh?

35
Dunlop, J.A. (1996) A trigonotarbid arachnid from the Silurian of Ludford Lane, Shropshire.
Palaeontology. 39, 605–614.
Dunlop, J.A. (2010) Geological history and phylogeny of Chelicerata. Arthropod Structure
and Development. 39 (2-3), 124–142. doi:10.1016/j.asd.2010.01.003.
Dunlop, J.A. (2011) Fossil Focus: Arachnida. Palaeontology Online. Volume 1, Article 7.
http://www.palaeontologyonline.com/articles/2011/fossil-focus-arachnida/
Dunlop, J.A. (2011) Fossil Focus: Chelicerata. Palaeontology Online. Volume 1, Article 1.
http://www.palaeontologyonline.com/articles/2011/fossil-focus-chelicerata/
Dunlop, J.A., Anderson, L.I., Kerp, H. & Hass, H. (2003) Preserved organs of Devonian
harvestmen. Nature. 425, 916. doi:10.1038/425916a
Dunlop, J.A., Anderson, L.I., Kerp, H. & Hass, H. (2004) A harvestman (Arachnida:
Opiliones) from the Early Devonian Rhynie cherts, Aberdeenshire, Scotland.
Transactions of the Royal Society of Edinburgh, Earth Sciences. 94 (04), 341–354.
doi:10.1017/S0263593300000730.
Garwood, R.J. & Edgecombe, G.D. (2011) Early Terrestrial Animals, Evolution,
and Uncertainty. Evolution: Education and Outreach. 4 (3), 489–501.
doi:10.1007/s12052-011-0357-y.
Garwood, R.J., Rahman, I.A. & Sutton, M.D. (2010) From clergymen to computers—the
advent of virtual palaeontology. Geology Today. 26 (3), 96–100.
Garwood, R.J. & Spencer, A.R. (2011) The geology and terrestrial life of the Carboniferous.
Deposits Magazine. 28, 38–47.
ReferenciasMás información en:

36
Garwood, R.J. & Sutton, M.D. (2010) X-ray micro-tomography of Carboniferous
stem-Dictyoptera: new insights into early insects. Biology Letters. 6, 699–702.
doi:10.1098/rsbl.2010.0199.
Giribet, G., Vogt, L., Pérez González, A., Sharma, P. & Kury, A.B. (2010) A multilocus
approach to harvestman (Arachnida: Opiliones) phylogeny with emphasis
on biogeography and the systematics of Laniatores. Cladistics. 26, 408–437.
doi: 10.1111/j.1096-0031.2009.00296.x.
Hedin, M.C., Starrett, J., Akhter, S., Schönhofer, A.L. & Shultz, J.W. (2012) Phylogenomic
resolution of Paleozoic divergences in harvestmen (Arachnida, Opiliones) via
analysis of next-generation transcriptome data. PLoS ONE. 7 (8), e42888.
doi:10.1371/journal.pone.0042888.
Pinto-da-Rocha, R., Machado, G. & Giribet, G. (eds.) (2007) Harvestmen: The Biology of
Opiliones. Harvard University Press, Cambridge, MA. 597 pp.
Slater, B. (2011) Fossil Focus: Coal swamps. Palaeontology Online. Volume 1, Article 4.
http://www.palaeontologyonline.com/articles/2011/fossil-focus-coal-swamps/
Wilson, H.M. & Anderson, L.I. (2004) Morphology and taxonomy of Paleozoic millipedes
(Diplopoda: Chilognatha: Archipolypoda) from Scotland. Journal of Paleontology.
78 (1), 169–184.doi:10.1666/0022-3360(2004)078<0169:MATOPM>2.0.CO;2.

Fossil harvestmenThe oldest surviving arachnids
Russell J. Garwood, Jason A. Dunlop, Gonzalo Giribet & Mark D. Sutton

38
Jason Dunlop was born in Basingstoke, England and grew up in the
coastal town of Cleethorpes. From childhood he was fascinated by
the spiders in his garden and decided early on he would like to turn
his hobby into a career. Jason thus studied zoology at the University
of Leeds and then undertook a PhD on an extinct group of fossil
arachnids in the Earth Sciences department of the University of
Manchester under the supervision of Dr Paul Selden. Jason remained
in Manchester on a research fellowship to investigate the origins of
arachnids and their relatives, and then in 1997 was appointed to the
position of curator for arachnids and myriapods at the Museum für
Naturkunde Berlin; a post which he has held until today. Jason has
published numerous studies about fossil arachnids –with a particular
focus on harvestmen in the last few years– and maintains a regularly
updated taxonomic list of all fossil arachnids and their relatives. He is
currently Secretary of the International Society of Arachnology, and
Vice-President of the European Society of Arachnology.
Russell Garwood is a palaeontologist based at the University of
Manchester, UK. He started his career studying geology as an
undergraduate at the Royal School of Mines, Imperial College,
London, before completing a PhD at the same institution in 2010.
Since then he has worked at the Natural History Museum, London,
running the CT lab there, and also in the School of Materials in
Manchester. For three years from 2012 Russell is being funded by
the Royal Commission for the Exhibition of 1851 to study the origin
and evolution of the insects, split between materials and the School
of Earth, Atmospheric and Environmental Sciences. Much of his
work uses X-ray techniques to better understand these animals and
their ecosystems, but he maintains an interest in a wide range of
early terrestrial animals. Other topics he is interested in include the
origin and early evolution of life, and the use of computers and
X-ray techniques in geology.
Russell J. Garwood Jason A. Dunlop

39
Gonzalo Giribet is an invertebrate zoologist interested in the origins
of animal diversity, especially the diversification of animal phyla
and the disparity of the large phyla, arthropods and molluscs. He
is also interested in the so-called “minor phyla”, often composed
of animals of small size and containing just a few species. He is a
Professor at the Department of Organismic and Evolutionary Biology
at Harvard and curator of the Department of Invertebrate Zoology
in the Museum of Comparative Zoology. His job therefore combines
research and teaching zoology, systematics, biogeography, and in
directing the invertebrate collections at the MCZ. His research has
taken him to collect invertebrates in every continent and ocean.
Born in Burgos, he grew up in Vilanova i la Geltrú, on the south
coast of Barcelona, where he developed his passion for zoology by
studying marine creatures. He went to school at the Universitat de
Barcelona, where he also received his PhD. In 1997 he moved to
New York City to conduct postdoctoral research at the American
Museum of Natural History, and from there he got an Assistant
Professorship at Harvard, where he went through the ranks until
becoming a Full Professor in 2007.
Mark Sutton is a Senior Lecturer in Palaeontology at Imperial
College, London. He studied at Cambridge (undergraduate) and
Cardiff (PhD), and has held academic appointments at Lampeter,
Oxford and, since 2005, Imperial College, London. His research
has primarily centered around the three-dimensionally preserved
invertebrate fossils of the Silurian ‘Herefordshire Lagerstätte’,
but he maintains broad interests in the phylogeny and evolution
of many groups, particularly the brachiopods, arthropods and
molluscs. His research interests also include computer-applications
in Palaeontology, especially three-dimensional reconstruction
techniques.
Gonzalo Giribet Mark D. Sutton

40 Speleosiro argasiformis (Pettalidae, Cyphophthalmi), un opilión cavernícola de Weynberg caves, Table Mountain, Ciudad del Cabo, Sudáfrica. Fotografía: Gonzalo Giribet.

41
You may never have heard of a harvestman before.
And you might think they are entirely unimportant. To the
arachnophobes amongst you, they may be a cause of much
fear. The odds are, though, that you have seen one these
arachnids in the past: there may well be some in your home, or
wherever you’re reading this right now, and there are actually
more species of harvestmen alive than there are extant species
of mammals. We hope to convince you in the course of this
article that they are really cool creatures. They are nothing to be
afraid of, but they do deserve a little respect. Harvestmen have
been around for a very long time, are biologically interesting,
and make important contributions to ecosystems. They’re also
increasingly useful as model organisms–creatures scientists
can use to understand animal development and evolution.
This article stems from a paper we wrote in 2011. While we’ll
introduce the group as a whole, and their fossil record, some
this article will focus on animals from a particular time (the
Carboniferous; 360–299 million years ago) which we have
studied with a neat technique (high resolution CT scanning).
But we’ll first introduce early life on land, based on what
we know from the fossil record. Not because it’s particularly
important to this study, but because it’s really interesting. We’ll
then: outline the fossil harvestmen we already know about;
introduce our time period, the last part of the Carboniferous;
and explain CT scanning. Finally, we’ll introduce the really
exciting new fossils which were the basis of our paper. So, let’s
start by having a look at life on land.
Figure 1 – Photograph and sketch of a ~427 million year old
millipede (species name: Cowiedesmus eroticopodus). This
figure shows the front of the fossil, including its head, collum
(first segment behind the head, with no limbs), and a number
of body segments behind this. A modified leg (the eleventh)
is also shown. It is possible this was used for reproduction.
Shown portion of fossil 12 mm in length. (Figure from
Garwood and Edgecombe 2011, modified after Wilson and
Anderson 2004, and photograph courtesy of Yong Yi Zhen,
Australian Museum).
Terrestrialisation
Terrestrial animals are those which live on land, and
terrestrialisation is the process by which this way of living
evolved (although, as with all these things, if you look in detail
at either, creating a proper definition of either isn’t as simple
as it might appear). The vast majority of known living species
is found within two groups of terrestrial arthropods –for more
information (including a definition of the word arthropod) see
the Arthropods and Arachnids box–. Estimates of the diversity
of life suggest insects account for more than 50% of known
living species, including plants, bacteria, and all other animals.
Some studies suggest this figure may be as high as 85%,
although we don’t know the exact proportion for sure. Of the
leftover species, a good proportion (about a fifth or more) are
arachnids: spiders and their kin. So, terrestrial arthropods are
big and important groups. They were also the first animals
that we know of which came onto the land. They may have
been preceded by soft bodied animals like worms, but it is
hard to tell, as these rarely preserve well as fossils. The earliest
land animals are 427 or so million years old, and are found in
the form of two millipedes: fossils discovered in Stonehaven,
Scotland (Figure 1). When dealing with fossils this old we
have to consider the possibility that something millipede-like
in appearance could have lived in water. We’re sure that is
Fossil harvestmen. The oldest surviving arachnidsRussell J. Garwood, Jason A. Dunlop, Gonzalo Giribet & Mark D. Sutton
Page12

42
not the case with these two species because they have things
called spiracles - openings to their breathing system, which
help keep it moist and control oxygen flow. This wouldn’t be
necessary in an animal living in water. Millipedes then, as now,
were probably detritivores - eating decaying organic matter
(probably plant–derived). There were no forests around at this
point: in fact, the earliest known plants date from around this
time period. So it appears from the fossil record that the first
land animals may have been millipedes or their ancestors, who
ate decaying organic matter from plants, fungi or bacterial
mats. Their arrival on land must have come before the first
fossils we find - after all, they have to be there already to be
fossilised! In fact, the earliest known traces of a millipede-
like creature can be found in the form of tracks fossilised in
a rock that date from tens of millions of years before this.
Millipedes and other members of the myriapods (centipedes
and two smaller groups) only shortly precede the arachnids in
the fossil record. The earliest known arachnid is a member of
an extinct, spider–like group called the Trigonotarbida (Figure
2) - one which, unlike spiders, lacked the ability to make silk
and still had an abdomen with segments (as we see in some
primitive spiders that still live in southeast Asia). This was found
by dissolving away rocks ~419 million years old with strong
acid and sifting the remains (arthropods’ exoskeletons don’t
dissolve in even the strongest acids). As almost all arachnids are
predators, including members of this group, it suggests that it
didn’t take all that long –in geological terms– for predation
to begin on land: a fact highlighted by the existence of a 418
million year old fossil centipede, also a predator, recovered the
same way.
Figure 2 – Photograph and sketch of the first known arachnid,
a member of an extinct group called the Trigonotarbida (species
name: Palaeotarbus jerami). The fossil was recovered by
dissolving away the host rock with acid – hence the fossils lack
of legs, and less-than-pristine appearance which has earned it
the nickname ‘the burnt corn flake’. Fossil 1.5 mm in length.
(Modified after Dunlop 1996).
By 411 million years ago the three largest arthropod
groups that live on land today (insects, spider, myriapods)
were present - at least in their primitive forms. We know
this from fossil deposits called the Rhynie and Windyfield
Cherts. These two rocks - situated near the village of Rhynie,
Aberdeenshire, in Northern Scotland - are very unusual: they
were formed when water rich in silicon dioxide from a hot
spring inundated this early terrestrial ecosystem (much like we
can see at Yellowstone National Park in the USA today). The
water petrified the animals and plants by replacing them with
chert - a rock made of silicon dioxide. While this is very rare,
when it does occur, it can preserve fossils in huge amounts
of detail: almost down to a cellular level. The Rhynie and
Windyfield cherts provide: the earliest relatives of the insects;
the first true insect, and possibly one with wings on the basis of
some mouthparts found in the deposit; trigonotarbids; mites;
and centipedes. That neatly brings us to the earliest fossil
harvestman, Eophalangium sheari (Figure 3), which is also from
the Rhynie Chert. This preserves enough internal anatomy to
make us sure that it is a true harvestman - in fact, when the
fossil was described it was placed in a living group, called the
Eupnoi (which we introduce later). It preserves elements of the
breathing system, the limbs, and even the genitalia. Hence the
Rhynie and Windyfield Cherts are notable in providing an early
glimpse of life on land and a number of earliest occurrences
of modern groups, including the Opiliones. After this there are
relatively few terrestrial arthropod fossil for the next 100 million
years. This is because the conditions needed for exceptional
preservation did not occur. Exceptions can be found in some
wonderful land fossils from 380 million years ago recovered
by dissolving rocks from New York state in acid, and a few
sites in Europe exist during this time where land animals can
be found flattened between the layers of shale rocks. None of
these contain harvestmen. There is one fossil Opiliones called
Brigantibunum listoni from Scotland, which is 340 million years
old, and –despite fairly poor preservation– also looks like it may
be a member of a still living group. But you’ll remember at the
start, that we mentioned that this article is about a particular
Page13

43
group of animals, from a particular time period in the Earth’s
history. That time period, late in the Carboniferous, called the
Pennsylvanian, kicked off a little over 320 million years ago.
Suddenly fossils of a huge range of land animals appear in
rocks all over the world. To know why this is we need to look a
little more closely at this period.
Figure 3 – Photograph of the earliest known harvestman (species
name: Eophalangium sheari). The fossil is preserved within a
translucent rock, allowing parts of the internal anatomy to be
seen. These include the base of the legs (or coxae, labelled C),
genital opening (G), and penis (P), showing that this was a male.
Around the body you can see parts of the legs. Scale bar 0.2
mm. (Modified after Dunlop et al. 2003 and Dunlop et al. 2004).
Arthropods and Arachnids
Arachnids are class of animals, member of a bigger
group called the arthropods. Arthropods are animals
which are defined by having a number of shared
features including: an exoskeleton, which is moulted as
the animal grows; a segmented body, with specialised
regions called tagmata; and jointed appendages. Other
groups of arthropods include the hexapods (insects
and a few smaller groups, Figure 4A–C), myriapods
(centipedes and millipedes and close relatives, Figure
4F), crustaceans (including crabs, shrimp, barnacles,
Figure 4G), and the extinct trilobites. Arachnids (Figure
4D–E, 5) are themselves member of a larger subgroup
of the arthropods, the Chelicerates. This subphylum is
name after the mouthparts, and comprises arachnids,
horseshoe crabs (not related to the true crabs),
probably sea spiders, and a number of extinct groups.
The class Arachnida consists of 15 smaller groups or
orders, including the mites and spiders. Other orders
include scorpions, whip spiders, whip scorpions,
pseudoscorpions, and poorly known groups like the
ricinuleids. Arthropods are the most diverse lineage of
animals around - the majority of their diversity is found
in the insect and arachnids, which together account for
around ¾ of all known living organisms.
Figure 4 – Examples of extant arthropods. A-C insects:
A- a butterfly, B- a honey bee, C- a relative of the
grasshoppers; D-E arachnids: D- a spider, E- a ricinuleid;
F- a millipede; G- a crayfish. Image credit: A) Richard
Bartz, Munich, Germany; B-F) Marshal Hedin, San
Diego State University, USA.
Figure 5 – An illustration by 19th century German
polymath Ernst Haeckel showing the huge range of
different forms found within arachnids.
The Carboniferous
The Carboniferous Period spanned from 360 to 290
million years ago. It’s split into two parts - the earlier
Mississippian (or Lower Carboniferous: 359–318 Ma) and
later Pennsylvanian (or Upper Carboniferous: 318–299 Ma).
Most of the rocks we have from the longer, earlier part are
shallow marine limestones (calcium carbonate rocks), and
very few fossils of land arthropods are even known from this
period: in fact, a single harvestman fossil is the exception to
this rule. In the Pennsylvanian, however –at least in terms
of rocks which record life on land– everything kicks off.
In Europe and North America, the later Carboniferous is
represented by lots of repeated sequences of mudrocks,
limestones, sandstones and coal layers. This results from a
combination of factors which we haven’t really seen since.
During the Carboniferous, a mountain building event was
happening, called the Variscan Orogeny (or Hercynian, or in
fact Alleghenian depending on where in the world you are).
Page14
Page16
Page17

44
A Southern continent, Gondwana, and a northern continent,
Laurussia (which eventually went on to make up many of the
rocks of Europe and North America) collided. In the process
the supercontinent Pangaea was formed, upon which the
dinosaurs lived tens of millions of years later. This collision also
created a large equatorial mountain belt (Figure 6). Associated
with these Variscan mountains, and a drop in sea level, were
lots of low–lying regions, which were host to swamps and
forests. In fact the Carboniferous is sometimes described as
having the first true rain forest ecosystems. The plants of
the time would have looked very unusual to our eyes. There
were: tree–like organisms related to extant club mosses; ferns
running the gamut from the small plants with which we are
familiar, to large trees; and thirty metre tall relatives of living
horsetails. Seed–bearing ferns and early relatives of conifers
also featured in these forests. Most of these plants were very
rich in a complex molecule called lignin which helps make up
some plant cell walls, giving wood its strength in the process.
The fact that the plants were so rich in this lignin is important.
Lignin is very hard to break down. Even today when lots of
animals such as termites, have evolved to eat wood, they
struggle to digest lignin. This resistance to biological decay,
and a lack of organisms which could degrade the lignin,
resulted in lots of carbon building up in the sediments of these
rainforests. This especially true because these low lying areas
were prone to rapid inundations of sediment (for example,
through a river changing course, or minor shifts in sea level).
Such events, coupled with low–oxygen, swampy conditions,
and rapid burial, created the ideal conditions to make coal.
Indeed, it is coal deposits from the Carboniferous that provide
the period with its name, and the vast coal reserves from this
25 million year period almost single–handedly fuelled the
industrial revolution in the Europe and North America.
Figure 6 – The Earth’s continents during the Pennsylvanian –
note the southern and northern continents colliding. Where
they meet, in the middle, mountains called the Variscan belt are
forming. (Image credit: Ron Blakey, NAU Geology).
Associated with this coal, are a huge number of fossils of
land animals - the creatures living in the Pennsylvanian swamps
and forests. These include a range of arthropods and other
invertebrates, and also some of the earliest land vertebrates
- animals which gave rise to the amphibians, and the reptiles,
birds and mammals. Environments in which coal forms are
also ideal for the growth of iron carbonate nodules. These are
made of the mineral siderite (FeCO3) and develop because of
high carbon concentrations in the sediment and water. They
can occur very rapidly after formation and will often precipitate
out around a decaying organism that has been buried with the
sediment, encasing it. Because these nodules form so quickly,
they prevent the organism from being squashed, as happens
to the rest of the sediment. The organism will then often rot
away leaving a void, which is sometimes then in–filled with
other minerals.
What this all means in that we have an excellent fossil
record of life on land from the Pennsylvanian. The geography,
sea level, plants and sediments of the time have created a
huge number of iron carbonate concretions, inside of which
we find a range of early land animals - for this reason the
Carboniferous is a unique insight for palaeontologists who
look at early terrestrial ecosystems. There’s an added bonus
that the fossils are associated with coal seams: they are often
dug up when people mine for coal. Another coincidence, this
one historical, comes into play here. In Europe, in the 19th
century, coal mining was a massive industry, and palaeontology
as a science was just hitting its stride. Accordingly there were
a large number of scientists around at just the right time to
collect these fossils, and many museums still have very large
collections of such fossils from this period. This has led to a
focus on similar deposits in other parts of the world, from this
time until today. So not only do we have very nicely preserved
fossils from this time - including lots of arachnids - but we
have a situation which has hastened their discovery. Neat huh?
There’s only one problem: 3D fossils are difficult to deal with.
Most fossils come about by an animal dying, being buried in
Page18

45
a sediment, and then being flattened within it. The traditional
approach of splitting open a rock is excellent for these
because everything there is to see of its anatomy is shown
on that single split plane. Preservation within a concretion, as
described for the Carboniferous, is relatively rare. But when
it does happen, traditional techniques aren’t so useful. If you
split open a nodule with a 3D fossil in it, parts of the anatomy
- the ends of the limbs, for example, or the head - remain
buried in the rock (Figure 7). And this has traditionally limited
what palaeontologists can say about the animals. But you’ll
recall, we’re talking about one group of animals (harvestmen)
from one time (the Carboniferous), discovered using a new
technique. That technique is computed tomography, and it’s
with 3D fossils such as these that it really comes into play.
Figure 7 – A typical siderite hosted Pennsylvanian fossil,
this example being a cockroach-like insect (species name:
Archimylacris eggintoni). In this case the mouthparts, limbs,
and body all remain hidden within the rock - all we see are
the wings (left), and head shield/head on the right. (Modified
after Garwood & Sutton 2010, Image credit: Natural History
Museum, London).
Computed tomography
Computed tomography, or CT scanning, is a non–
destructive X–ray based technique. It allows palaeontologists
to see inside fossils and recreate them as 3D digital models. It’s
an increasingly common tool in the world of palaeontology
as scanning technology and computing power has improved.
CT scanning for palaeontologists comes in a number of
flavours, all following the same principle as CT scanners in
hospitals. In medical systems an X–ray source and detector
rotate around a patient, and take a series of radiographs
as the patient moves through this rotating contraption.
These scanners are big, and as a result are sometimes
used in palaeontology for large fossils - the only problem
being that the scanners are designed for humans, and so
have weak X–ray sources to minimise the radiation dose to
patients. Sometimes the source can’t penetrate dense rock.
A more common form of CT scanning for fossils is X–ray
microtomography (XMT or μCT). In this setup a sample (the
fossil) is rotated around 360 between an X–ray source and a
detector, which takes an X–ray of the sample every, say, 0.1
degrees of rotation (Figure 8A). From the resulting 3000+
X–rays, some clever maths can be used to create slice images
of the fossil (Figure 8B). These images are actually maps of
the X–ray attenuation within an object - the amount that any
given part of the fossil reduces the intensity of the X–rays.
Because attenuation is the result of both density and the
elements that make up any part of the fossil, these maps
can usually differentiate the fossil from the host rock (and
sometimes different parts of the fossil from each other). The
result is a series of black and white images of the cross–
section of a fossil. The process of studying a fossil on the
basis of CT scanning doesn’t end there, however - normally
it is followed by the long and laborious process of making a
digital model. There are quite a few different ways to do this,
but the method used to create the figures later in this article
involves first creating threshold images - a simple means of
turning pixels within a certain range of grey levels ‘on’ to
be rendered in 3D, and ignoring the rest. With CT scans of
voids within rock, this involves picking out the darkest pixels,
because air has a lower X–ray density that the host nodule.
This will create a 3D model of the fossil, but also any cracks
within the nodule, and airspace outside the rock. To get rid
of these, we can go through all 2000 slice images of the
fossil, colouring in the different parts of the anatomy - body,
legs, mouthparts etc., and then rendering each separately
in 3D. In the process this removes any artefacts, or cracks.
By rendering the model in 3D, seeing where the colouring is
wrong, and then making improvements on each slice, a very
neat model is finally created. By extracting the morphology
of the animal in full from the host rock - in the case or this
article, two harvestmen from Carboniferous nodules of
France - μCT can tell us a lot more about a fossil’s mode of
Page19

46
life, its biology, and its evolutionary relationships. So, at this
point, it makes sense for us to acquaint ourselves with the
animals that provide the subject of our article.
Figure 8 – A: An X–ray projection, or radiograph. CT scans are
made of a large number of these, acquired as a sample rotates.
This example is an arachnid within a siderite nodule. B: The
resulting slice image which maps the X–ray attenuation within a
sample. Different features are labelled.
Harvestmen
The harvestmen are arachnids - members of a group that
also includes the spiders, scorpions, mites and ticks (Figure 5). In
common with other arachnids they possess eight legs, and a body
split into two major parts - the prosoma (front) and opisthosoma
(back). These parts of the anatomy set arachnids apart from
related animals like insects (six pairs of legs and three body parts:
a head, thorax and abdomen), and millipedes/centipedes (many
legs, and lots of segments). Harvestmen arachnids - members
of the order Opiliones - are defined by having the two halves
of their body (that is, the prosoma and opisthosoma), joined by
a broad and sometimes poorly–defined juncture (Figure 9). This
is in contrast to spiders, for example, which have a narrowing
between these two parts (or tagma) of the body. Of these
tagma, the prosoma has the four pairs of walking legs, and the
mouthparts - the chelicerae, which are the equivalent of a spider’s
fangs, and the pedipalps, which are the same limb that scorpions
have evolved to use as claws (Figure 10). The opisthosoma has
no real limbs in arachnids, and the harvestmen are no exception.
Rather, the back half has openings to the breathing system,
small holes called spiracles, and also the genital opening, which
is often covered by a flap called the operculum. Males have a
penis, whilst females have an ovipositor - a specialised organ for
laying eggs. In many groups the second pair of walking legs isn’t
actually used for moving around. Instead, it is longer than the
other limbs, and acts a bit an antenna, waving around and letting
the animal feel its surroundings.
Figure 9 – A harvestman arachnid (species name: Leiobunum
aldrichi) with the anatomy labelled. Note the division into two
halves of the body (the prosoma and opisthosoma), with a
broad junction, the mouthparts (pedipalps and chelicerae), and
walking legs 1 – 4 (L1 – L4). (Image credit: Marshal Hedin, San
Diego State University, USA).
Name origins
Harvestmen have a variety of different names in
different cultures. In English the names harvestmen
- or alternatively harvest spiders - may well stem from
the fact that Opiliones tend to be abundant during the
harvest season. Alternatively this may be because when
their legs are detached, which happens commonly in
some groups as a form of defence, the limb makes a
reaping motion. Hence they are known in a number of
languages as reaper too. The Spanish name agostero
has similar origins, whilst in some provinces harvestmen
are known as pedros. This is in reference to Saint Peter’s
Day which falls during harvest season. The group’s Latin
name, Opiliones, dates from 1833. Swedish zoologist
Karl J. Sundevall proposed the name, after the Latin
opilio, meaning sheep–master (used by a Roman
dramatist Plautus, 254–184 BC) or shepherd, as featured
in Virgil’s Eclogues (37 BC). It is suggested the origin of
this could be due to the long legs and high body of some
European species making harvestmen reminiscent of
ancient European shepherds who used stilts to oversee
their flocks.
Figure 10 – The pedipalps (blue) and chelicerae (red) marked
on two non–harvestman arachnids: A spider and an early fossil
scorpion. Note also that both have bodies which can be divided
into prosoma and opisthosoma - one of the things that defines
an arachnid.
Page20
Page21
Page22

47
Harvestman nicknames and myths
In many parts of the US, harvestmen are called daddy
longlegs, or grandaddy longlegs - this means they are
sometimes confused in Europe with crane flies, which
have the same nickname on European shores. Some
nicknames for the harvestmen in different countries
reflect the fact that they resemble spiders, but are not
true members of the group - for example Afterspinne
in German, which means false spider. In the US and
Australia, a common urban legend suggests the group
is extremely poisonous, but that their mouthparts are
incapable of breaking human skin. In common with
many urban legends, this is entirely untrue - harvestmen
have no venom.
Harvestmen are a big group - in terms of arachnids there
are only two bigger: the spiders, and the mites, and there’s only
one bigger group which is more diverse than the arachnids,
which is the insects. If we measure the diversity of harvestmen
by the number of harvestman species that have been described
to date, there are around 6400 - the first two being described in
1758 by Linnaeus, who invented the system of defining species
which scientists still use today. By comparison there are ~5400
known species of mammals. Despite being a big group, the
biology of harvestmen is still a little murky, and many things
remain unknown. Arachnologists - scientists that work on
arachnids - believe they may represent fairly primitive members
of the group. Many think they are most closely related to the
scorpions, pseudoscorpions, and solifuges (a small arachnid
order otherwise known as camel/sun spiders or wind scorpions).
This grouping of four orders is called the Dromopoda, although
arachnologists still debate both whether this is correct, and if
so, what the relationships between the four orders may be.
Quite a lot of debate regarding evolutionary relationships within
the harvestmen also remains. Scientists agree the harvestmen
comprise four distinct lineages (each is technically a suborder).
The Cyphophthalmi (Figure 11A) is likely to represent the first
of these to branch from the others - it is small (6 families, 130
species) but has a worldwide distribution. Members of the group
are typically small (1–5mm), with short legs, and are usually blind.
They have a thick exoskeleton, and tend to live in the leaf litter on
forest floors, although some live in caves. The remaining three
lines of harvestmen are often grouped together as a lineage
called the Phalangida. The suborder Eupnoi (6 families, ~1800
species, Figure 11C) includes arguably the most easily recognised
harvestmen - creatures with small soft bodies, hanging down
between very long legs. This group is also found in most parts
of the world. Another suborder, the Dyspnoi (7 families, 290
species, Figure 11B) shows a great variety of form, but is united
by having small, regular teeth on the chelicerae. Interestingly,
this suborder is found largely in the Northern Hemisphere. The
final group is the Laniatores - the biggest suborder (27 families,
~3750 species, Figure 11D), which are often colourful and
spined, but are largely restricted to the tropical regions, and the
temperate Southern Hemisphere. We’re not 100% settled on
the relationships between these three suborders - two possible
arrangements are shown in Figure 12.
Figure 11 – Examples of each of the four lineages of harvestmen.
A: Cyphophthalmi. B: Dyspnoi. C: Eupnoi. D: Laniatores. (Image
credit: Marshal Hedin, San Diego State University, USA).
These lovely creatures are rather varied in their ecology and
way of life. Many live in soils and leaf litter, and in dark damp
places such as under rocks and in caves. Some, however, live on
walls and tree trunks and a number spend their time amongst
vegetation. A particular consideration with harvestmen is
the humidity of the place they’re living, as many species are
poor at minimising water loss. Dark, and humid environments
help avoid any issues which may otherwise arise from losing
too much water. We see the greatest diversity amongst
harvestmen in tropical, especially rainforest, environments.
Page23

48
Being omnivorous, Opiliones eat a wide range of things - small
invertebrates, plants, carrion and fungi. They have even been
known to survive on cappuccino mousse when raised in the
laboratory! Having such variety in their diet is unusual because
other arachnids are almost all purely predatory, and tend to
digest their food by vomiting digestive enzymes on it, prior to
filtering and sucking up food using a pumping stomach. Where
they are predators, harvestmen tend to ambush their prey,
or occasionally run it down. Their feeding system has issues,
however - because the harvestmen use their mouthparts,
which end in pincers, to break food into small chunks before
eating it rather than sucking up a liquid (which would allow
the food to be filtered), they are very prone to ingesting
parasites. Also, Opiliones often fall prey to birds, amphibians,
small reptiles, and even spiders. They are not completely
defenceless, however; some species secrete a sticky substance,
gluing debris onto their outside to provide camouflage. Others
feign death if they are attacked, and some bob up and down to
confuse potential predators. All harvestmen have openings to
glands which emit nasty smells on the front half of their body
- another defence mechanism. Finally, many can detach their
legs if these get stuck somewhere, for example. This defensive
strategy is known as autospasy or appendotomy. The lost limbs
don’t grow back, but the break occurs along special lines of
weakness. As a result - other than the loss of a leg (which
doesn’t seem to impact on the creature too much) - it doesn’t
cause many issues as the muscles of the leg close the wound.
Figure 12 – Two possible relationships between the different
Opiliones orders - both have the Cyphophthalmi as the earliest
branching suborder, but differ in the arrangement of the other
three groups. (Photograph credits: Marshal Hedin, San Diego
State University, USA).
Another big question when dealing with the biology of
creatures such as harvestmen is how they reproduce. The
majority of harvestmen do this sexually - a male and a female
member of the same species are required for fertile young
to be created (although there are a few species which have
been seen to multiply asexually). Harvestmen employ internal
fertilisation - in the majority of cases (all groups except the
Cyphophthalmi), this means males have a penis, and a pair
copulate in order to breed. After copulation, females can lay
eggs immediately or after a period of time. They use their
ovipositor to do this, and eggs will hatch somewhere between
a few days and several months later. Most species show a range
of forms of parental care for the eggs/young. That brings to a
conclusion our quick introduction to the harvestmen - a really
interesting group of creatures.
We’ve now introduced the time period from which these
fossils come (the Pennsylvanian), the method used to study
them (X–ray microtomography), and the animals which
have been fossilised (the harvestmen). So, we’ll move on to
introducing our study.
This study
We were lucky enough to find two new harvestman fossils
for this investigation. Both were found within siderite nodules,
from a deposit called Montceau-Les-Mines, in the Massif
Central, France. Relatively little of their body was showing on
the crack sued to break the nodule open - for one of them,
we didn’t even realise the fossil was a harvestman until we
had scanned it. Both were housed in the Muséum d’histoire
naturelle d’Autun, and we borrowed the fossils from there
to study. In order to reveal the anatomy hidden in the rock,
we conducted scans of both on a CT scanner in the Natural
History Museum, London - to do so we put the bits of the
cracked nodules back together, and held them in place with
cling film. After scanning, and then an hour’s processing to
turn the plain radiographs into slices, we had 2000 images
showing the fossils’ cross sections. In one dataset each pixel
represented a 26.5µm x 26.5µm square, and in the other each
was 14.2µm x 14.2µm. In each cross sectional image, because
the fossils were less dense to X-rays than the surrounding rock,
Page24

49
they appeared as dark shapes in the slices. We used custom-
built software to reconstruct the fossils in 3D as we described
in the micro-CT section. This was a bit of a slow process - it
required us to make a guess at what we were looking at in
slice images, render the model in 3D, and then improve the
colouring. However, eventually we created the models that
you see in this article - two very nice looking, and remarkably
complete models of fossil harvestmen. To make the images you
see here, we took these models, and exported them as a file
recording the shape of their 3D surfaces and imported them
into an open source CGI program called Blender. This allowed
the images and movies to have shadows and realistic lighting.
The fossils
The results of our work - or at least a major part of them
- are the 3D models which you can see in the figure. They are
both quite unlike any other harvestmen we know of a remotely
similar age, so we were able to name both of them as new
species. This meant that both needed a name - two, in fact, as
both needed a genus name too. Traditionally these are Latin
or Greek in origin, conforming to a system which has been in
place since the work of Linneaus in the 18th century.
Figure 13 – The CT scanner used for this study. This can scan
samples from a few millimetres to a quarter of a metre in size.
The cabinet housing the equipment is lead lined to prevent
radiation escaping.
Ameticos
The spiny creature you see in our figures we called Ameticos
scolos. This is Greek in origin, ametikos meaning relating to the
harvest, and skolos meaning thorn, in reference to the animal’s
spines. The fossil has a large, elliptical body which was almost
one centimetre in length. The boundary between the front half
(prosoma) and back half (opisthosoma) of the body is marked
by a half-a-millimetre wide rectangular ridge. The prosoma is
quite wide compared to the back half, and has a series of pits
towards the posterior. The eyes are situated on a mound (the
ocularium) found at the very front of the animal, where there is
a small projection (the structure which looks a little like a nose).
The underside of the creature is heavily crushed but we can see
the base of the walking legs on one side. The “mouthparts”
- the chelicerae - are small (0.8mm in length) and have three
segments, but are otherwise a little unclear. This is probably
because they are not well preserved, rather than because we
couldn’t get the details out in our CT scan. The second pair of
appendages (pedipalps) are also rather small. They are missing
a claw at the end, and we can see all six segments which make
up the 2.3mm long limb. The most complete leg on the fossil
is the third left leg, which has four preserved segments, and is
almost 9 mm in length. The other limbs appear to have been
snapped off between the second and third segments (the
trochanter and femur respectively). Moving to the back of the
creature, what is most obvious is the three pairs of prominent
spines (each 2.4mm long). This makes the entire creature
around half a centimetre deep. We can see five segments in
the opisthosoma - the spines being found on the first three.
The entire top surface is covered with deep, spherical pits
about a quarter of a millimetre across. On the underside we
can also make out five segments in the opisthosoma - they get
shorter towards the back.
Figure 14 – The fossil harvestman species Ameticos scolos. A:
View from above, with the anterior appendages in the darkest
purple to the right, and then longer (but prematurely truncated)
legs in lighter shades. B: View from the side. This makes the
spines particularly clear, shown towards the posterior (left). C: A
view of the underside, which in this specimen is crushed a little,
especially towards the bottom of the image.
Macrogyion
The fossil with long legs we decided to call Macrogyion
cronus. This also stems from Greek - makros means long and
Page25
Page26

50
gyion means limb. Cronus/Kronos was the father of Zeus, Hades
and Poseidon, and the god of the harvest. So this fossil is a long
legged creature, named in honour of the harvest god. Cool,
huh? The fossil is smaller than Ameticos - it has an oval body
around 6.5 mm in length. It’s hard to tell where the prosoma/
opisthosoma boundary is, and again, parts of the leg bases and
underside of the opisthosoma have been partially crushed. The
body is around half a centimetre deep again, so this is more
spherical than the flatter Ameticos. This time the chelicerae
are very well preserved. They are 3.3 mm long, and made of
three segments, ending in a pincer. They are tucked in between
well preserved pedipalps - which are held aloft and in front of
the prosoma, and end in a simple claw. All six segments of the
pedipalps are quite clear, and the entire limb is nearly 8mm
long. The legs are generally snapped off again, but the first left
is preserved in its 2.6cm entirety - the first three segments point
upwards, before a bend and a horizontal segment, then another
bend. The rest of the limb points downwards from there. The last
part of the leg is curved, suggesting it is made from a large number
of sub-segments - something we see in a number of groups which
are still around today. The rest of the limbs have snapped off in
various places, and all are much less complete. The prosoma is a
little squashed too, but the top has not been caved in like the
underside. It has an ocularium - a mound, which presumably
held the eyes in life. The first segments of the opisthosoma are
fused together, and one of them has a small ridge on it, but three
segments are visible differentiated at the back of the animal. The
segmentation on the underside is well preserved - there are five
clear segments, the first being a lot longer than those behind it.
Figure 15 – The fossil harvestman species Macrogyion cronus.
A: View from the side, including the single complete leg with
a curved termination. The rest of the legs have snapped off
towards the top of the third segment. B: A side view showing
pedipalps and chelicerae (right), walking legs removed. C: The
chelicerae in more detail. These end in a claw, which would
have been used to process food. D: The underside, showing
that the specimen is towards the bottom of the image.
Assessing evolutionary relationships.
When you have a fossil, one of the first things
you want to be able to say is what other animals it
is related to. Living organisms have a hierarchical
structure: closely related organisms look more similar
to each other than they do to those less closely related.
On this basis, palaeontologists have traditionally placed
fossils into groups encompassing different levels of
similarity - for example, members of a family (such as
Hominidae: the great apes, including humans, chimps
and bonobos, gorillas and orang-utans) will share
a number more features than do the families within
an order (e.g. Primates, which also includes monkeys
and lemurs, amongst others). However, those families
will still share a number of features which defines the
order. In turn, orders are the members of a class - a
group defined by another, probably less specific set of
shared characters. This goes all the way to a phylum
or kingdom level, which are the truly coarse divisions
of life.
Modern palaeontologists still use this system - the
fossils we have described here, for example, belong to
the Opiliones, and class Arachnida. However, working
out evolutionary relationships just on the basis of
anatomical similarity can be somewhat subjective. For
this reason most palaeontologists now use something
called cladistics to understand the evolutionary
relationships, or phylogeny, of a group. This involves
creating a list of different aspects of the anatomy of an
animal. For example, you could use the number of legs,
the presence (or absence) of eyes, whether the leg ends
in a number of sub–segments, and a huge number of
other aspects of an organisms nature, as characters. A
selection of the animals in question are then coded for
these characters - so, for example, the number of legs
could be entered in a list for all the animals, as could the
Page27

51
presence (e.g. coded as 1) or absence (coded as 0) of
eyes. This creates a matrix - a large number of characters
for a large number of species, which in essence records
the similarities and differences between the coded
animals. With this matrix, a computer algorithm can be
used to calculate work out which evolutionary tree(s)
which involve require the smallest number of character
changes. So in the case of eyes, if 5 of the 20 species
in an analysis lack eyes, and they form a unique group,
the eyes would only need to be lost once. Another
example is shown in Figure 16, of exactly the same
comparison but with another character. In this way,
a large number of different characters can be used to
try and work out a likely evolutionary tree: that with
the smallest number of changes. Doing this, of course,
requires the assumption that the smallest number of
changes will provide the most likely evolutionary tree.
But using this technique does allow us to provide
a testable, and verifiable hypothesis. It allows us to
demonstrate how well - or how poorly - any given
grouping is supported, and often what the alternatives
are. If there are a number of shortest trees of equal
length, we can produce a consensus tree - this is a tree
which has only the bits that all the different shortest
trees agree on.
Figure 16 – An example of a single character, whose
changes are marked onto two possible trees. The
harvestman groups Laniatores and Dyspnoi both have
females who lack joints in their ovipositor (the organ
they use to lay eggs). If we assume this is the primitive
condition for the harvestmen, such joints must have
been lost somewhere on the way to the Laniatores and
Dyspnoi. If the two are not closely related, as shown
on the left, this must have happened once in each
lineage. However, if they are more closely related to
each other than the rest of the harvestmen, this loss
could have occurred just once, in the ancestor of both
groups. Thus - for this character at least - the tree on
the right has less steps. If you have a large number of
characters, you can use comparisons like these to build
a tree with the smallest number of steps, and provide
a testable hypothesis for evolutionary relationships.
(Photograph credits: Marshal Hedin, San Diego State
University, USA).
In recent years people have started using the same
technique with the DNA of a range of organisms to try
and assess how closely related they may be. The longer
ago two organisms shared a common ancestor (i.e. the
longer ago their lines split), the more time there has
been for mutations to occur in their DNA. Accordingly,
if you can compare two of the same stretches of DNA,
more closely related animals will have less differences
than less closely related animals, and again, you can
use this to work out a tree. You can also combine these
two approaches, allowing you to code the morphology
of both fossils and living organisms, and then use the
DNA from the living species to help build the tree. We
have used this approach here to try and work out the
relationships of our two fossils.
Cladistics methods
To work out the evolutionary relationships of these two
fossils, we performed a cladistic analysis (see box Assessing
evolutionary relationships). For this we used 201 morphological
characters, all coding different parts of harvestmen anatomy.
We then used a piece of software called TNT to calculate
the tree which required the smallest number of anatomical
changes. We used a number of different aspects of the
technique to assess the relationships. These included giving
all the characters the same importance (“equal weights”),
and giving those which appear to be less reliable (i.e. they
Page29

52
reverse more in many of the trees) less importance (“implied
weights”). We showed how well supported the tree was as
well, using a measure called symmetric resampling. As well
as this morphological matrix, we used the DNA of the living
species to help build the tree.
Cladistics results
All of this analysis gave us some interesting and exciting
results, which we’ll outline here, but the importance of which
we’ll discuss later. The analysis included non–harvestman
arachnids were used to root the tree - i.e. to work out
which harvestmen are most similar to the other arachnids,
and therefore probably split first. Using just the morphology
of the extant species and the fossils, analysing with equal
weights, we recovered a number of groupings. These included
the Opiliones as a whole, the primitive Cyphophthalmi, and
Phalangida (all harvestmen minus the Cyphophthalmi) - all
came out as sharing a single common ancestor which defined
each group (Figure 17).
Figure 17 – A tree showing the relationships of these fossils.
A: The tree recovered using the morphology of living and
extinct harvestmen, and the DNA of non–fossil species. Fossils
are shown in red (Halitherses is a younger fossil preserved
in amber). Ameticos scolos is recovered as a member of the
suborder Dyspnoi, and Macrogyion cronus as a member of the
Eupnoi. The images on the right show B: Macrogyion cronus and
D: Ameticos scolos with comparable taxa of the modern Eupnoi
and Dyspnoi; C: Nelima sempronii (family Sclerosomatidae)
E: Acuclavella cosmetoides, (family Ceratolasmatidae). Scale bar
B–C, 2mm; D–E 1mm. (Image credits: C: Christian Komposch,
Ökoteam, Graz, Austria; D: courtesy of Casey Richart, San Diego
State University, USA).
The fossils - despite being placed in the Phalangida - had no
clear position: if the analysis was run without the fossils, Eupnoi
and Dyspnoi both also came out as true groupings. Hence adding
our fossils changed the cladistic result: adding M. cronus stopped
the Eupnoi being recovered as a true group, and A. scolos had
the same effect on the Dyspnoi when we made a consensus tree.
In other words, in all of the trees which were of the same length,
the fossils were found in so many positions that relatively few
of the evolutionary relationships recovered were shared between
trees. The fossils have lots of missing parts of the anatomy, which
may explain this result: they add an element of instability, so
when we work out which are the best supported trees, there are
a number with the fossils in very different places. If we added
the implied weighting - i.e. lowered the importance of seemingly
unreliable characters in the analysis - it had relatively little impact
on the evolutionary relationships we recovered. Adding the
DNA of extant species helped create a better defined tree - the
Eupnoi and Dyspnoi were strongly supported as a group by their
DNA evidence. With DNA, and equal weights, M. cronus was
identified as a member of the Eupnoi and A. scolos the Dyspnoi.
We tested this result by using a few more techniques to assess
the evolutionary relationships of the fossils, including a statistical
technique known as Bayesian inference. All the analyses we did
supported this same placement of the fossils. For all of these
methods of placing the fossils, we provided an indication of
how well supported the tree was. When including the fossils,
this was usually low - a lot of the anatomy we may use to place
living fossils, such as spiracles (openings to the tracheal system),
genitalia, or eyes, can’t be resolved for certain on either fossil,
as they are too small, or are internal and thus aren’t found with
this type of preservation. This includes most of the characters
which are used to define each suborder of the harvestmen.
Nevertheless, the fact that in all analyses we obtained essentially
the same result suggests that this is a fairly reliable position for
the fossils in the Opiliones family tree.
What this means
So we have two new fossils, reconstructed in 3D, and
we have worked out how they are probably related to other
harvestmen. This allows us to create accurate reconstructions
Page31

53
of how they would have looked in life (Figure 18), and say
something about both the palaeobiology of these fossil species
(such as how they lived their lives).
Figure 18 – Idealised reconstructions of Ameticos scolos (above)
and Macrogyion cronus (below) showing how the two species
may have looked in life. Scale bar 5mm.
We can also use their discovery to say something about the
history of harvestmen as a whole. Because these specimens are
so well preserved, and this is unusual for harvestmen fossils,
this actually represents the first time any fossils older than a
few tens of millions of years have been placed in a harvestman
tree on the basis of a cladistic analysis. As previously outlined,
the Opiliones can be divided into four suborders, the Eupnoi,
Dyspnoi, Laniatores and Cyphophthalmi, and our work has
placed Ameticos with Dyspnoi, and Macrogyion with Eupnoi.
This means that both of the suborders Dyspnoi and Eupnoi
must have been around 305 million years ago, when these
fossils were deposited in a shallow body of water in one of the
Carboniferous coal forests. Now, it has already been suggested
that the Eupnoi existed 411 million years ago on the basis of
the harvestman mentioned earlier from the Rhynie Chert - this
was largely inferred on the basis of its genital morphology
– although new discoveries may suggest that the Rhynie
specimen is not a Eupnoi. The confirmation using a broader
range of anatomical observations that a Carboniferous eupnoid
(Macrogyion) existed adds weight to the suggestion that the
Eupnoi is an ancient suborder. In contrast, the oldest fossil
Dyspnoi placed with a cladistic analysis before this paper is a
few tens of millions of years old. Hence Ameticos is not only
the earliest known member, but it pushes the group’s earliest
known fossil back by a long way. As well as using the similarities
in DNA to work out the relationships between groups, the
DNA can also be used to estimate when different groups split
from each other based on the frequency with which mutations
occur. A number of biologists have used this technique in
the last decade to estimate the age of different harvestman
groups, and both these fossils corroborate such estimates. The
relationships we discovered also makes it very clear that two
harvestman suborders - both of which are still around today
- were present in the forests of Europe 305 million years ago.
In contrast, at this time, other arachnids such as spiders and
scorpions are not present - at least in the form we see them
today. Rather, they tend to be known in just their earliest
forms: lineages which are now extinct. The only arachnids we
know were similarly modern in anatomy by this point are the
mites. Hence this work provides compelling evidence for an
early origin of the harvestmen - it seems likely that they are
amongst the earliest branching arachnids. Beyond these early
origins, both fossils are essentially modern in appearance - this
demonstrates that Opiliones’ body plans were established by
the Late Carboniferous, and have remained largely unchanged
in the 305 million years since. They are an excellent example of
evolutionary stasis.
We can use this similarity to make educated guesses about
how our fossils may have lived their lives. Ameticos scolos bears
a striking resemblance to some living members of the Dyspnoi
- particularly members of the genus Acuclavella (members of a
family called Ceratolasmatidae - Figure 17E). The second pair
of walking legs on our spiny fossil was probably elongate - in
common with Acuclavella. We suggest this is the case because
even though the second legs are broken off midway through
the third segment, the femur, this part is already longer than
the femora on all the other legs. We know from modern
harvestmen - such as Acuclavella - that these legs are used
to feel ahead of the animal as it walks, a bit like an antenna.
The fact Ameticos has deep pits on its upper surface suggests
a robust, well–built exoskeleton, that implies the creature is
more likely to have lived on the forest floor, rather than in the
trees. Acuclavella, the modern dyspnoid, lives at the margins of
small perennial water features in moist, woody debris, and we
think that its spines may provide defence against amphibian
predators. As such, our fossil could have inhabited similar
regions in the moist, detritus–rich Carboniferous coal forests,
Page33

54
and likely bore spines for the same reasons. We can make the
same sort of inferences for Macrogyion cronus, which is similar
in form to modern eupnoids in the families Phalangiidae and
Sclerosomatidae (Figure 17C). For example, the proportions we
see in the segments of the limb, the body shape and size, and
the mouthparts are very similar. Hence, the fact that the body
of the fossil is partly crushed and less well preserved than the
limbs suggests that - like modern members of these families
- the exoskeleton of this fossil was probably soft and leathery
cuticle in life. The complete leg shows the animal possessed
lots of sub–segments at the end of walking legs, and would
have allows it to coil these where required. This is a condition
typical of living members of the Eupnoi, and a possible
adaptation towards life in vegetation, allowing the animal to
better grip plant stems. This suggestion is supported by the
highly–elongate limbs, which in modern species is indicative of
life above ground–level. Accordingly, Macrogyion could have
had a vegetation or tree dwelling mode of life.
Conclusions
We hope this has proved an interesting article. We hope
also that our introduction to elements of palaeobiology,
cladistics, CT scanning, harvestmen and the history of life
and land all made sense: we have covered a lot of ground!
We wanted to finish by summing up the things we have
mentioned. We have learned that the first animals on land
were arthropods. Arthropods include arachnids, and the
harvestmen are just one group of these. They are fairly rare
as fossils. However, due to a special set of conditions in the
Carboniferous period, a lot of land animals from this time are
preserved in three dimensions - including these two new fossils.
Three–dimensionally preserved fossils can be problematic:
using traditional techniques, palaeontologists are left with
unrecovered parts of the organism buried within the rock. CT
scanning can overcome this by using X–rays to recover slice
images, which can in turn be used to create a 3D model of the
fossils. Here it allowed us to reconstruct two 305 million year
old harvestman fossils, and use the resulting models to try and
assess their evolutionary relationships. This work suggests that
harvestmen are one of the earliest branching arachnid groups,
and are also an excellent example of evolutionary stasis. The
models and comparison with living harvestmen have allowed
us to suggest that one species was a ground–dwelling creature,
and the other lived in vegetation or trees. All that, from two
little fossils buried in a rock. Not bad, huh?

55Caddella sp. (Acropsopilionidae, Dyspnoi), cerca de Knysna, Western Cape Province, Sudáfrica. Fotografía: Gonzalo Giribet.

56
Terrestrización Página 11
El Carbonífero Página 14
Tomografía computarizada Página 18
Opiliones Página 20
La investigación Página 25
Los fósiles Página 26
Ameticos Página 26
Macrogyion Página 27
Métodos cladísticos Página 30
Resultados cladísticos Página 30
¿Qué significa esto? Página 32
Conclusiones Página 34
Índice

57Eucynortella duapunctata (Cosmetidae, Laniatores), Reserva Ducke, Manaos, Amazonas, Brasil. Fotografía: Gonzalo Giribet.

58
Terrestrialisation Página 41
The Carboniferous Página 43
Computed tomography Página 45
Harvestmen Página 46
This study Página 48
The fossils Página 49
Ameticos Página 49
Macrogyion Página 49
Cladistics methods Página 51
Cladistics results Página 52
What this means Página 52
Conclusions Página 54
Contents

59
¡Fundamental! es una serie de publicaciones de difusión paleontológica de la Fundación Conjunto Paleontológico de Teruel–Dinópolis que consta de subseries temáticas para cuentos (Fundacuentos paleontológicos) o para versiones divulgativas de artículos científicos (Paleonturología) y, en función de la complejidad de los contenidos, tiene previsto dirigirse a públicos de tres niveles, especificados en el lomo de cada número mediante un pictograma.
Dirección de la serie: Luis Alcalá.
El tercer número, Evolución humana en el valle del río Omo (Etiopía), se edita como consecuencia de la adjudicación del Primer Premio Internacional de Investigación en Paleontología Paleonturología 03.
03
El segundo número, Dinosaurios de Teruel, ha sido editado en el marco del III Seminario sobre Paleontología y Desarrollo de la Universidad de Verano de Teruel (Universidad de Zaragoza) y con la colaboración de ésta.
02
El Gigante Europeo: excavando un dinosaurio. Documental y juego interactivo de la excavación del gran dinosaurio de Riodeva (Teruel), Turiasaurus riodevensis. Programa Nacional de Fomento de la Cultura Científica y Tecnológica del Ministerio de Educación y Ciencia (492839C1).
05
Teoría del Homosaurus (Paleontología imaginaria de Dino y Saura) es un cuento del que es autor Elifio Feliz de Vargas (Teruel, 1964), veterinario. En 1990 obtuvo la “Ayuda a la Creación Literaria” concedida por el Instituto de Estudios Turolenses y en 1991 el Premio “Teruel” de relatos.
04
07 –3.200.000 + 2005 Tejidos (óseos, arquitectónicos, pictóricos) es el catálogo de la exposición artística presentada en el Museo de Teruel desde el 4 de noviembre al 11 de diciembre de 2005.
06 XVI Reunión Bienal de la Real Sociedad Española de Historia Natural contiene los trabajos de investigación presentados en dicha reunión, celebrada en Teruel (septiembre-octubre de 2005).
01 El primer número, El apasionante mundo de la Paleontología, se editó en el marco del proyecto La paleontología al alcance de todos, desarrollado por la Fundación y financiado por el Ministerio de Ciencia y Tecnología (DIF2003–10062–E).
08 El octavo número, Modalidades de ornamentación en bivalvos, se edita como consecuencia de la adjudicación del Segundo Premio Internacional de Investigación en Paleontología Paleonturología 04.
09 El noveno número, Los huesos gastrales de los terópodos: insuflando vida a los dinosaurios, se edita como consecuencia de la adjudicación del Tercer Premio Internacional de Investigación en Paleontología Paleonturología 05.
10 Laboratorios de Paleontología (IV Seminario sobre Paleontología y Desarrollo de la Universidad de Verano de Teruel), ha sido editado en el marco del Año de la Ciencia 2007 con una ayuda de la FECYT y del Ministerio de Educación y Ciencia (proyecto Paleontología en Teruel: dos libros mejor que uno CCT005–07–00629).

60
Los ojos de los trilobites: el sistema visual más antiguo conservado se edita como consecuencia de la adjudicación del Quinto Premio de Investigación en Paleontología Paleonturología 07.
13
12 Aligerando a los gigantes: los huesos neumáticos de los dinosaurios saurópodos y sus implicaciones para la estimación de la masa corporal se edita durante el Año de la Ciencia 2007 como consecuencia de la adjudicación del Cuarto Premio de Investigación en Paleontología Paleonturología 06.
El dinosaurio que excavó su madriguera se edita como consecuencia de la adjudicación del Sexto Premio de Investigación en Paleontología Paleonturología 08.
15
Mesozoic Terrestrial Ecosystems in Eastern Spain es una síntesis en inglés de ecosistemas mesozoicos continentales –Pirineos orientales y centrales, Teruel y Cuenca– preparada con motivo de su visita durante el 10th MTE Symposium, celebrado en Teruel (septiembre de 2009).
14
En la Tierra como en el Cielo: Río Tinto como análogo terrestre de Marte se edita como consecuencia de la adjudicación del Séptimo Premio de Investigación en Paleontología Paleonturología 09.
17
Myotragus: la economía energética en la evolución se edita como consecuencia de la adjudicación del Octavo Premio de Investigación en Paleontología Paleonturología 10.
19
El Plateosaurus virtual se edita como consecuencia de la adjudicación del Noveno Premio de Investigación en Paleontología Paleonturología 11.
21
Dinojuegos (serie Fundajuegos paleontológicos, de nivel infantil) se ha realizado en colaboración con el Museo Nacional de Ciencias Naturales (CSIC) y en el marco del proyecto e-dino 10, financiado por la Fundación Española para la Ciencia y la Tecnología – Ministerio de Ciencia e Innovación (FCT-10-980). Incluye un DVD.
18
10th Annual Meeting of the European Association of Vertebrate Palaeontologists incluye las comunicaciones presentadas en la reunión anual de la EAVP celebrada en Teruel (junio de 2012).
20
Escuela Taller de Restauración Paleontológica IV describe trabajos de laboratorio y de campo promovidos para la recuperación de patrimonio y para la formación profesional.
22
XVI Simposio sobre Enseñanza de la Geología incluye las 34 comunicaciones presentadas en la reunión conmemorativa del vigésimo aniversario de la Asociación Española para la Enseñanza de las Ciencias de la Tierra (Teruel, julio de 2010).
16
11 Teruel: territorio paleontológico (Seminario sobre Paleontología y Desarrollo de la Universidad de Verano de Teruel), ha sido editado en el marco del Año de la Ciencia 2007 con ayuda de la FECYT y del Ministerio de Educación y Ciencia (proyecto Paleontología en Teruel: dos libros mejor que uno CCT005–07–00629).

������
����
���

Copyright © 2022 FDOKUMEN




















