
оглавление - Институт экспериментальной медицины
173
-
Upload
khangminh22 -
Category
Documents
-
view
2 -
download
0
Transcript of оглавление - Институт экспериментальной медицины


2
ОГЛАВЛЕНИЕ
Введение ..............................................................................................................… 5
Глава 1. Обзор литературы .................................................................................................................. .. 15
1.1. Глобальная и фокальная ишемия головного мозга в клинике ……… 15
1.2. Метаболические нарушения головного мозга, происходящие
вследствие фокального ограничения гемодинамики. Ядро ишемии и
зона «ишемической полутени» …………………………………………….. . 18
1.3. Молекулярные механизмы ишемического повреждения клеток
головного мозга: немедленные и отсроченные ……………………...…… 27
1.4. Основные морфофункциональные особенности клеток мозга …...… 38
1.5. Реактивные изменения различныхтипов клеток головного мозга
в ядре ишемии и в зоне «ишемической полутени» ………….………...…. 47
1.6. Свойства антигипоксантов. Фармакологическая характеристика
метапрота .……………………………...……...……………………….……. 54
1.7. Выбор отделов головного мозга для моделирования последствий
выраженной и слабо выраженной глобальной ишемии ……………..…… 58
Глава 2. Материал и методы исследования .………………………..….……… 62
2.1. Моделирование глобальной ассиметричной ишемии головного мозга .. 63
2.2. Группы исследования …………………..………………………….…... 63
2.3. Морфологическое исследование головного мозга крыс ..……...……. 64
Глава 3. Результаты собственных исследований ……...……………...………. 70
3.1. Особенности строения и расположения разных типов клеток
в некоторых отделах мезокортиколимбической дофаминергической системы
у здоровых крыс ……..…………………………………………… 70
3.1.1. Особенности строения слоев V – VI переднего
цингуцингулярного поля …………………………………….……. 71
3.1.2. Особенности строения прилежащего ядра переднего мозга
………………………………………....…….…… 76 3.1.3.
Особенности строения паранигрального ядра среднего мозга
………………………………………..….....…………. 81

3
3.2. Реактивные изменения разных типов клеток некоторых отделов
мезокортиколимбической дофаминергической системы после
глобальной ишемии .……………………………………...……….………... 90
3.2.1. Изменения клеточного строения в глубоких слоях
передней цингулярной коры (область выраженной ишемии) ...... 90
3.2.2. Изменения клеточного строения прилежащего ядра
переднего мозга (область выраженной ишемии) ………...……....... 96
3.2.3. Изменения клеточного строения паранигрального ядра
среднего мозга (область слабо выраженной ишемии) .……...... 103
3.3. Коррекция метапротом количественных и структурно-топогра-
фических изменений клеточных элементов в фокусе выраженной
ишемии и за его пределами …….……………………………..………… 111
3.3.1. Коррекция метапротом ишемических изменений
клеточных элементов слоев V – VI переднего цингулярного
поля (область выраженной ишемии) …………..………….…… 111
3.3.2. Коррекция метапротом ишемических изменений
клеточных элементов паранигрального ядра среднего мозга
(область слабо выраженной ишемии) …………..…..…………. 118
Глава 4. Обсуждение полученных результатов …………………………….. 127
4.1. Изменения клеток передней цингулярной коры и приле-жащего
ядра, расположенных в области выраженной ишемии . 129 4.2.
Изменения клеток паранигрального ядра, расположенных
в области слабо выраженной ишемии ………………..…….….. 135
4.3. Фармакологическая коррекция метапротом постишемичес-
ких изменений клеток исследованных отделов мезокортико-
лимбической дофаминергической системы ….………….….…. 137
Выводы …………………………….…………….…………………………….. 140
Практические рекомендации …………………………………………………. 142
Список литературы ………………………..…………………………………... 143
Приложение …………………………………………………………………… 164

4
СПИСОК СОКРАЩЕНИЙ
МДoфС – мезокортиколимбическая дофаминергическая
система ПНигЯ – паранигральное ядро среднего мозга
(дофаминергическое)
ПЦинК – передняя цингулярная кора конечного мозга (прегенуальная, поле Cg3 у крыс)
ПЯ – прилежащее ядро конечного мозга
GFAP – глиальный фибриллярный кислый белок астроцитов

5
ВВЕДЕНИЕ
Актуальность исследования.
Проблема ишемического повреждения головного мозга становится все
более важной в последние годы, так как она занимает первое место в
структуре инвалидности и смертности, вытесняя ишемическую болезнь сердца
(Гусев Е.И. и Скворцова В.И., 2001). В связи с этим создание адекватных
экспериментальных моделей и изучение этой патологии в них остается
актуальной проблемой современной медицинской науки и практики.
Модель глобальной церебральной ишемии, вызванная лигированием
обеих общих сонных артерий широко используется для оценки эффективности
коррекции постишемических нарушений фармакологическими препаратами на
основе оценки неврологических и биохимическиих тестов (Пошивалов В.П.,
1978, Шабанов П.Д., Зарубина И.В. и Soultanov Vagif S., 2011). В этом случае
нарушение кровоснабжения более выражено в переднем мозге и меньше – в
стволовой части, где кровообращение поддерживается двумя позвоночными
артериями (Paxinos G., 2004). Животные выживают в течение 2 недель после
операции, если они не подвергаются стрессу. Информативность данной
методологии, однако, может стать намного выше, если ее дополнить данными
реактивных постишемических изменений основных типах клеток головного
мозга – нейроцитов, клеток макро- и микроглии и сосудистого эндотелия,
расположенных в различных гемодинамических условиях, что предже изучено
не было.
Степень разработанности темы исследования
Морфологические исследования глобальной ишемии головного мозга,
не привязаны к определнному его отделу и посвящены только визуальному
описанию общих признаков изменений нейроцитов и развитию защитной
реакции макроглии – сателлитозу. В исследованиях последствий фокальной
ишемии авторы раскрывают изменения разных типов клеток, как в зоне
ишемического инфаркта, так и во внутренней части прилегающей области
(zona penumbra), где начинается формирование капсулы как результат реакции

6
на фокальную циркуляторную гипоксию (Васильев Ю.Г. и Берестов Д.С.,
2011; Szpak G.M. et al., 1999). Совсем не попадаются исследования,
отражающие специфику патологических изменений нейроцитов, клеток
макро- и микроглии, сосудистого эндотелия, обусловленную их расстоянием
от ядра инфаркта и временем, проходящим от начала ишемического
воздействия.
В тех немногочислнных работах, посвященных выявлению реактивных
изменений клеток в фокусе ишмии и ближайшей к ней зоне, все же,
содержится мало сведений о количестве фенотипических форм патологически
модифицированных нейроцитов и других клеток, слабо представлены
количественные и морфометрические данные пространственной пластичности
макроглиальных клеток и самих нейроцитов, (считающихся, в отличие от
микроглиальных, оседлыми) по отношению к телам нейроцитов и стенке
капилляров. Между тем защитно-приспособительная транслокация тел
олигодендроцитов и астроцитов, выявленная при наркотической гипоксии
(Богомолов Д.В., 2001; Должанский О.В., 2001; Дробленков А.В., Карелина
Н.Р. и Шабанов П.Д., 2009), алкогольной абстиненции и хронической
алкогольной интоксикации (Дробленков А.В., 2011), инфекционном процессе
(Voskuhl R.R. et al., 2009), введении суспензии астроцитов, мигрирующих в
направлении сосудистой стенки (Okoye G.S. et al., 1995) может развиваться
также и вследствие ишемии. Имеющиеся сведения о пространственной
пластичности нейронов головного мозга при ишемии, полученные при
электронной микроскопии (Гелеранская О.А. и соавт., 2016) позволяют
убедиться в необходимости более детального изучения таких
приспособительных реакций, как объединения поврежденных нейронов как
между собой, так и со стенкой кровеносных сосудов.
Объектом для изучения особенностей реактивности клеток головного
мозга, расположенных в различных условиях глобальной ишемии мозга, могут
быть основные отделы мезокортиколимбической дофаминергической системы
(МДофС). Интерес к структурно-функциональным особенностям этой

7
системы обычно вызван ее ключевой ролью в формировании зависимости от
употребления психоактивных веществ. В основу зависимости заложен
устойчивый дефицит дофамина в синапсах между аксонами дофаминерги-
ческих нейроцитов среднего мозга и нейроцитами проекционных отделов
МДофС – прилежащего ядра, миндалевидного тела, ядер ложа конечной
полоски, передней цингулярной и префронтальной коры (Анохина И. П.
ссоавт., 2001; McBride W. J. et al.; 1990; Bergstorm H. C. at al., 2008). Большая
протяженность компонентов МДофС (от среднего мозга, где расположены
дофаминергические ядра до конечного мозга, содержащего основные
проекционные центры), а также наличие обширных связей с любым из
отделов мозга (способных повлиять на процесс ишемического повреждения ее
клеток механизмами электрохимической и макромолекулрной передачи)
делают систему пригодной для установления градиента ишемических
изменений. Как правило, МДофС также является структурной мишенью
действия нейропротекторных средств. В качестве последних интересен
препарат метапрот (от слов «МЕТАболический ПРОТектор»), в течение
длительного времени входивший в арсенал медицинского обеспечения
Вооруженных сил РФ в качестве табельного средства, повышающего
работоспособность. В основе действия метапрота лежит его способность
стимулировать энергетические и пластические процессы в головном мозге,
миокарде, печени, почках (Шабанов П. Д., 2009; Шабанов П. Д., 2010).
Разработаны методические основы количественного и морфометричес-
кого изучения тел нейроцитов, тел и отростков астроцитов, нейроцито-
глиальных взаимоотношений в разных отделах МДофС у самок крыс разного
возраста, которые могут быть использованы в ходе настоящего исследования
(Дроблен-ков А. В. и Карелина Н. Р., 2009; Дробленков А. В. и Шабанов П. Д.,
2014).
Таким образом, определение комплекса малоизвестных морфометричес-
ких параметров и ряда визуальных признаков ишемического повреждения
разных типов клеток исследованных отделов МДофС в данной модели

8
глобальной ишемии позволило бы: 1) раскрыть наиболее полно их
реакционные способности, 2) установить стандарт ишемического повреж-
дения этих клеток, 3) верифицировать морфологически эффективность нейро-
протекторных свойств препарата метапрот.
Цель исследования – установление структурных, пространственных и
количественных изменений основных типов клеток некоторых отделов
МДофС, расположенных в разных условиях циркуляторной гипоксии, а также
эффективности метаболического протектора как средства коррекции
постишемических нарушений.
Задачи исследования:
1. Определить основные структурные, пространственные и
количественные характеристики разных типов клеток передней цингулярной
коры и прилежащего ядра конечного мозга (ПЯ и ПЦинК), паранигрального
ядра среднего мозга (ПНигЯ) у здоровых молодых самцов крыс.
2. Установить реактивные изменения основных типов клеток ПЦинК и
ПЯ конечного мозга, расположенных в области выраженного ограничения
кровотока через 7 суток экспериментальной ишемии.
3. Установить особенности реактивных изменений основных типов
клеток ПНигЯ среднего мозга, расположенных в области слабо выраженного
ограничения кровотока через 7 суток экспериментальной ишемии.
4. Выявить морфологические признаки церебропротекторного действия
метапрота на головной мозг по показателям реактивности основных типов
клеток ПЦинК конечного мозга и ПНигЯ среднего мозга через 7 суток после
глобальной ишемии мозга.
Научная новизна исследования. Впервые установлен комплекс
морфометрических параметров для верификации реактивных изменений
различных типов клеток, расположенных в центре частичной ишемии и во
внешней части зоны пенумбры при моделирования ассиметричной глобальной
ишемии головного мозга. Этими параметрами являются: долевое соотношение
различных типов патологически модифицированных нейроцитов – клеток-

9
«теней» (погибших), набухших, сморщенных-гиперхромных и маломодифи-
цированных; площадь тел маломодифицированных и набухших нейроцитов;
доля тел нейроцитов, объединенных в пары; число и площадь тел астроцитов;
глиоцито-нейроцитальный индекс; расстояние между телами нейроцитов,
астроцитов и стенкой капилляров; число микроглиоцитов и площадь их тел;
число эндотелиоцитов капилляров.
Использование данных параметров позволило впервые выявить целый
спектр выраженных патологических изменений клеток различного типа, в том
числе малоизвестных свойств пространственной пластичности тел нейроцитов
и макроглии. В центре частичной ишемии нейроциты склонны к гибели:
завершенной (образование клеток-«теней») и пролонгированной (тип
сморщивания-гиперхромии или отека-набухания), причем тела наименее
поврежденных нейроцитов способны к объединению между собой и клетками
макроглии. Образование нейроцито-макроглиальных кластеров, располагаю-
щихся, главным образом, вблизи стенки разрастающихся капилляров, может
являться для клеток компенсаторно-приспособительным механизмом и
условием их выживания на территории выраженной частичной ишемии мозга.
Белок промежуточных филаментов астроцитов в периферических частях
отростков и перивазальных глиальных мембранах подвергается выраженной
деградации. Между тем, клетки макроглии в фокусе частичной ишемии более
устойчивы к повреждению, чем нейроциты, так как погибшие формы данных
клеток в условиях эксперимента отсутствовали.
На периферии области ишемии нейроциты склонны к переживанию
дистрофических изменений по типу острого набухания и объединению с
клетками макроглии. Образование нейроцито-глио-васкулярных кластеров не
характерно. GFAP в телах, отростках и перикапиллярных глиальных
астроцитарных мембранах повергается изменениям гипертрофического типа.
Впервые определено, что в данной ассиметричной модели глобальной
ишемии мозга параметры изменений микроглиоцитов в исследуемых

10
структурах переднего и среднего мозга значительно не различаются и
выражаются признаками трансформации в клетки амебоидной формы глии.
Определены морфологические признаки выраженного нейроцито(гисто)-
про-текторного эффекта актопротектора и антигипоксанта метапрота. Они
направлены на активацию нейроцито-глиальных взаимоотношений,
протекцию атроцитов и их промежуточных филаментов, в том числе в
периваскулрных глиальных мембранах и сохранение структурных
особенностей нейроцитов, присущих малоизмененным клеткам.
Основные положения, выносимые на защиту
1. Переживание условий выраженной и слабо выраженной
циркуляторной гипоксии в модели глобальной ассиметричной ишемии мозга
индуцирует развитие у разных типов клеток головного мозга целого спектра
выраженных патологических изменений. Нейроциты находятся в процессе
сморщивания-гиперхромии, лизиса и превращения в «клетки-тени». Клетки
ядерной зоны склонны к гибели или сморщиванию-гиперхромии. Нейроциты,
расположенные на периферии области ишемии и испытывающие, главным
образом, дистантное воздействие реакций глютамат-кальциевого каскада,
склонны к острому набуханию.
2. Астроциты, переживающие ишемию в ядерной зоне, отличаются
признаками отека-набухания, деструкции промежуточных филаментов в части
площади их тел, периферических отделах отростков и периваскулярных
глиальных мембранах. На периферии области ишемии происходят изменения
астроцитов гипертрофического типа. Реакция микроглиоцитов в виде слабо
выраженного увеличения их числа и признаков активации является ранним
диффузным проявлением ассиметричной глобальной ишемии мозга.
Пролиферация эндотелиоцитов на ранних сроках ишемического воздействия
происходит лишь в фокусе ишемии.
3. Концентрация жизнеспособных нейроцитов и астроцитов вблизи
стенки кровеносных капилляров, сближение тел нейроцитов с образованием
между собой пар и небольших групп, а также рост числа сателлитных форм

11
глии являются адаптационными механизмами и условием выживания клеток
при различных условиях ишемического воздействия на головной мозг.
4. Применение метапрота значительно усиливает компенсаторно-
приспособительные реакции клеток мозга. Она способствует сдвигу удельного
количества субпопуляций нейроцитов в сторону преобладания
немодифицированных (нормохромных) и существенному сокращению
теневидных и гипохромных форм, не подавляя при этом процесс активации
нейроцито-глиальных взаимоотношений, индуцированный ишемией.
Препарат препятствует формированию контактов клеток со стенкой
капилляров. Усиление защитных реакций клеток мозга, переживающих
циркуляторную гипоксию в сочетании с терапией метапротом, может быть
обусловлено его известными механизмами нейропротекции.
Теоретическая и практическая значимость исследования
Теоретическое значение выполненного диссертационного исследования
состоит в том, что установлен целый ряд морфологических параметров
строения и пространственных взаимоотношений основных типов клеток в
главных отделах МДофС. Величины данных параметров представлют собой
эталон нормы для здоровых самцов крыс молодого возраста и эталон
ишемических изменений, происходящих в условиях выраженного и слабо
выраженного ограничения кровотока при повторном использовании
популярной модели глобальной ишемии мозга с целью установления
эффективности применения различных фармакологических препаратов.
Выявленные параметры пространственной пластичности тел клеток
макроглии, нейроцитов по отношению друг к другу и к сосудистой стенке
представлют собой слабо изученное звено защитно-приспособительного
механизма выживания комплекса клеток головного мозга, которые могут быть
учтены в предстоящих исследованиях генеза различных патологических
процессов, а также в качестве ранних диагностических признаков ишемии.
Практическое значение работы определяется выявлением
морфологических признаков нейропротекторных/нейрореставрационных

12
изменений после курсового назначения актопротектора метапрота.
Полученные данные являются важным доказательством вовлечения разных
типов клеток головного мозга в осуществление системных глиально-
клеточных реакций, вовлекаемых в действие метаболических протекторов.
Методология исследования состояла в изучении последствий
перевязки обеих сонных артерий у молодых самцов крыс методами
морфологического анализа: описательного, иммуноцитохимического,
морфометрического при помощи световой микроскопии гситологических
срезов в проходящем свете. Фармакологическую коррекцию возникающих
вследствие перевязки ишемических нарушений проводили с помощью
метапрота – метаболического протектора, стимулирующего энергетический
обмен, синтез РНК и аминокислот. Исследования выполнены с соблюдением
всех принципов доказательной биологии и медицины и одобрены локальным
комитетом по этике при Федеральном государственном бюджетном научном
учреждении «Институт экспериментальной медицины» (Санкт-Петербург).
Степень достоверности и апробация материалов исследования.
Статистическая обработка результатов
Выборка для каждой группы животных состояла не менее чем из 15
подсчетов в каждой группе животных (состоящей из 4-х крыс). Математико-
статистические методы: было использовано программное обеспечение класса
электронных таблиц Microsoft Excel 7.0. Математико-статистическое описание
объектов исследования осуществлялось с помощью традиционных и давно
утвердившихся в медицинских исследованиях методов (Айвазян С.А., Енюков
И.С. и Мешалкин Л.Д., 1983; Лакин Г.Ф., 1990; Юнкеров В.И., Григорьев С.Г.
и Резванцев М.В., 2011). Устанавливали средние значения (среднего арифме-
тического значения показателей, моды, медианы), характеристики колебле-
мости признаков (дисперсию, среднее квадратическое отклонение, размах
значений), стандартные ошибки средних значений и их доверительных
интервалов. Результаты обрабатывали статистически с использованием t-
критерия Стьюдента на персональном компьютере.

13
Реализация результатов работы. Работа выполнена в соответствии с
планом научно-исследовательских разработок отдела нейрофармакологии им.
С.В. Аничкова Федерального государственного бюджетного научного учреж-
дения «Институт экспериментальной медицины» (Санкт-Петербург). Получен-
ные результаты используются в учебном процессе кафедр морфологии
человека и специализированной терапии Института медицинского образова-
ния Федерального государственного бюджетного образовательного учрежде-
ния высшего профессионального образования «Новгородский государствен-
ный университет имени Ярослава Мудрого» Минобрнауки РФ, кафедры
фармакологии Федерального государственного бюджетного военного образо-
вательного учреждения высшего профессионального образования «Военно-
медицинская академия им. С.М. Кирова» Минобрнауки РФ.
\Апробация и публикация материалов исследования Результаты и
основные положения диссертации доложены и обсуждены
на Всероссийской научной конференции «Актуальные проблемы морфологии,
адаптогенеза и репаративных гистогенезов», посвященной памяти чл.-кор.
АМН СССР проф. Ф.М. Лазаренко (Оренбург, 2013); Всероссийской
молодежной конференции «Нейробиология интегративных функций мозга»
(Санкт-Петербург, 2013), научной конференции молодых ученых
«Перспективы развития медицинской науки и практики» (Санкт-Петербург,
2014), научной конференции, посвященной 100-летию со дня рождения чл.-
кор. АМН СССР проф. А.Г. Кнорре «Актуальные проблемы морфологии,
эмбриональный и репаративный гистогенез, филогистогенез» (Санкт-
Петербург, 2014), Всероссийской научно-практической конференции с
международным участием «Экологические аспекты морфогенеза» (Воронеж,
2015), XIII конгрессе международной ассоциации морфологов (Петрозаврдск,
2016), Всероссийской конференции молодых ученых «Нейробиология
интегративных функций мозга» (Санкт-Петербург, 2016).
Апробация работы прошла на научном заседании отдела нейрофармако-
логии им. С.В. Аничкова ФГБНУ «ИЭМ» (Санкт-Петербург).

14
Публикации
По теме диссертации опубликовано 15 печатных работ, из них 6 статей в
рецензируемых журналах, рекомендованных ВАК, 1 статья в иностранном
журнале, 2 статьи в сборниках научных трудов и 6 тезисов.
Личный вклад автора
Личный вклад автора осуществлялся на всех этапах работы и состоял в
планировании экспериментов (90%), их непосредственном выполнении (95%),
обработке результатов (100%), обсуждении результатов, написании статей и
тезисов (80%), написании диссертации и автореферата (95%).
Объем и структура диссертации
Диссертация состоит из введения, обзора литературы, описания
материалов и методов исследования, главы собственных исследований,
обсуждения результатов, заключения, выводов, научно-практических
рекомендаций и списка литературы. Диссертация изложена на 163 страницах и
содержит следующие разделы: введение, обзор литературы, материал и
методы исследования, результаты собственных исследований, обсуждение
полученных результатов, выводы и список литературы, содержащий 195
источников (60 отечественных и 135 иностранных). Диссертация
иллюстрирована 25 рисунками и 13 таблицами. 5 рисунков составляют
короткое приложение.

15
Глава 1. ОБЗОР ЛИТЕРАТУРЫ
1.1. Глобальная и фокальная ишемия головного мозга в клинике
Общеизвестно, что головной мозг, по сравнению с другими органами и
частями тела, утилизирует кислород (O2) наиболее активно. Во время
ускоренного роста и более высокой активности организма он может
поглощать до 50% O2 от его количества, поглощенного всем организмом.
Митохондрии нейроцитов мозга также вырабатывают до 95% организменного
аденозинтрифосфата (АТФ). Поэтому недостаток O2 в клетках головного
мозга ведет к снижению электрического потенциала их мембран и качеству
синаптической передачи нервного импульса (Ашмарин И.П. и Стукалов П.В.,
1996; П.К. Телушкин, А.Д. Ноздрачев и Потапов П.П., 2008).
Определено, что глюкоза очень быстро усваивается клетками головного
мозга. Его запасы истощаются в мозге, когда приток крови прекращается, в
среднем, через 30 минут (Ашмарин И.П. и Стукалов П.В., 1996; Лукьянова
Л.Д., 1997). Первопричина этого связана с тем, что основной путь
метаболизма глюкозы, основного субстрата окисления в нервной ткани
является аэробный гликолиз, в котором его молекулы расщепляются до
углекислого газа и воды. Интенсивность поглощения глюкозы клетками
головного мозга, особенно нейроцитами, является чрезвычайно высокой, в то
время как его собственные резервы в клетки мозга, чрезвычайно малы и
содержатся в телах астроцитов (Kajihara H., Tsutsumi E., Kinoshita A. et al.,
2001).
Пpи кpитичeском cнижении кoнцeнтpации глюкозы в кpoви нейpоны
начинают использовать гликoген астроцитов для поддержания окислительных
процессов, но из-за его небольших количеств окисление заканчивается через
5-7 минут (Aшмapин И.П. и Cтукaлoв П.В., 1996). Следовательно, функцио-
нирование мозга полностью зависит от регулярного приема глюкозы из крови.
Увеличение скорости окисления глюкозы и связанное с этим увеличение
активности нейроцитов от окислительных процессов как источника энергии

16
происходят по мере развития мозга, пропорционального выполнению задач
(Aшмapин И.П. и Cтукaлв П.В., 1996). Эти авторы также установили
зависимость потребности в церебральных нейроцитах в O2 и глюкозе от их
функциональной активности. В связи с этим очевидна особая важность
проблемы церебральной ишемии, возникающей в результате сокращения
церебрального кровотока и ограничения O2 на нервную ткань (циркуляторная
гипоксия) и глюкоза. Поэтому ишемия является наиболее распространенной
причиной нарушения функции мозга.
Глобальная (общая) церебральная ишемия развивается не только в
результате сердечной недостаточности, нарушений сердечного ритма или
системной артериальной гипотензии. Патогенетическим фактором в
глобальной ишемии мозга может быть патология сосудов головного мозга
(мальформации, атеросклероз), нарушения свертываемости крови,
уменьшение концентрации O2 в атмосфере, а также интоксикация и
органические расстройства клеток нервной ткани, в том числе возраст
(который снижает активность трофического и энергетического обеспечения
клеток и их ферментных систем).
В то же время, ишемия головного мозга встречается реже, чем очаговая
(локальная) ишемия, которая возникает, когда кровоток в отдельном
сосудистом русле нарушается (из-за ишемического инсульта или
транзиторной ишемической атаки).
Увеличение распространенности сосудистых заболеваний, возникающих
в последние годы, вызвал рост частоты острых нарушений мозгового
кровообращения. По эпидемиологическим международным данным (World
Development Report) около 6 миллионов человек земного шара ежегодно
страдают от инсульта, в России это число насчитывает более 450 000 человек.
Каждые 1,5 минуты кто-то из россиян впервые подвергается этой болезни. У
лиц трудоспособного возраста (до 64 лет) отмечается увеличение
распространенности инсульт. Годовая смертность от инсульта зарегистриро-
вана у 4,7 миллиона человек населения мира. В России инсульт занимает

17
второе место в общей структуре смертности, уступая только сердечно-
сосудистой патологии. В 1999 году ранняя 30-дневная смертность после
инсульта составляла 35%, в течение года в среднем умирало около 50%
пациентов.
Инсульт ведет к высокой инвалидизации населения. Более половины
людей, которые переживают до конца третьего года после инсульта, страдают
от неврологических расстройств (Stegmayr V. and Asplund K., 2003). По словам
авторов, не более 20% выживших пациентов могут вернуться к своей
предыдущей работе. Ежегодная потеря полной жизни из-за неадекватной
реабилитации после инсульта составляет 20,3 миллиона человеко-лет в мире
среди мужчин и 22,9 миллиона человек в год среди женщин. Социально-
экономическое значение нарушений мозгового кровообращения усугубляется
тем, что эти нарушения налагают обязательства по уходу за пациентом на
членов его семьи и общества. По данным ВОЗ, которые используют авторы
(Stegmayr V. and Asplund K., 2003), средняя стоимость содержания пациента с
инсультом в США составляет 55000-73000 долларов в год. Исходя из этих
данных, общая стоимость нашего государства, связанная с проблемой
инсульта, должна составлять от 16,5 до 22 млрд. $ США в год.
Таким образом, инсульт представляет собой проблему чрезвычайной
медицинской и социальной значимости. Среди всех типов инсульта
преобладают очаговые ишемические поражения головного мозга. Согласно
международным многоцентровым исследованиям, соотношение ишемичес-
кого и геморрагического инсульта находися в пределах 5,0-5,5.
В результате долгой истории изучения механизмов острых нарушений
кровообращения мозгового кровообращения были установлены четыре
группы основных патогенетических факторов поражения ишемического мозга
(Боголепов Н.К., 1971; Боголепов И.И. и Бурд Г.С., 1981; Бадалян Л.О., 1975;
Гусев Е.И., Бурд Г.С. и Боголепов Н.Н., 1979; Верещагин Н.В., 1980; Hennerici
М. et аll, 1988; Voiculescu V., 1989; Fisher M. and Bogousslavsky J., 1996;
Warlow C.P. et al., 1996). К ним относятся: 1) структурные изменения в стенке

18
сосудов головного мозга; 2) недостаточность церебральной гемодинамики из-
за снижения кровотока до критического уровня и ниже его; 3) изменения
содержания белковых фракций электролитов, недоокисленных продуктов
обмена и других физико-химических свойств крови; 4) индивидуальные и
возрастные особенности метаболизма клеток головного мозга, реализованные
в реакциях на локальные изменения в гемодинамике.
Данные этио-патогенетической структуры ишемического повреждения
головного мозга всего населения (Warlow, C.P. et al., 1996) свидетельствуют о
том, что около 50% острых нарушений мозгового кровообращения являются
тромботическим или эмболическим осложнением атеросклеротического
процесса (атеротромботический инсульт и артериоартериальная эмболия).
Причина многих острых ишемических нарушений связана с патологией
сосудов головного мозга малого калибра (лакунарный инсульт), около 20% - с
кардиогенной эмболией (кардиоэмболический инсульт). Причины около 5%
ишемических поражений встречаются редко (гемодинамический инсульт,
вызванный острым инфарктом миокарда или падением системной
гемодинамики другого генеза, сосудистых нарушений воспалительного генеза,
травмы и расслоения артерий, гематологических расстройств, инфекций, рака ,
излучение и т.д.). Любой из этих патогенетических факторов является
следствием возникновения острой фокальной ишемии.
1.2. Метаболические нарушения головного мозга, происходящие
вследствие фокального ограничения гемодинамики. Ядро ишемии и зона «ишемической полутени»
Нарушения биоэнергетики головного мозга, образующиеся в результате
гипоксии-ишемии, наиболее тщательно изучены при остановке кровотока в
русле середней мозговой артерии. Биохимическими и патофизиологическими
методиками было определено, что снижение мозгового кровотока,
происходящее в фокусе ишемии, инициирует метаболические реакции ткани
головного мозга, интенсивность которых обусловлена прежде всего

19
продолжительностью и степенью ограничения тока крови (Jacewicz M.,
Kiessling M. and Pulsinelli W.A., 1986; Hossmann K.A., 1994a, b; Moghaddasi M.,
Taati M., Asadian P. et al., 2017). Результаты исследований данных авторов
позволили Хоссману К. (1994, а) сформулировать концепцию порогов
жизнеобеспечения клеток в сосудистой гипоксии (Hossmann, K.A., 1994, a).
Эта концепция до настоящего времени наиболее полно раскрывает сущность
повреждения нейроцитов и других клеток головного мозга.
Согласно концепции порогов жизнеобеспчения, было предложено
выделить четыре критических уровня снижения кровотока, каждый из
которых соответствует определенным типам метаболических реакций.
Графическая характеристика этих уровней обобщена в монографии Гусева
Е.И. и Скворцовой В.И. (2001; Приложение, рис. 1).
Первый критический уровень снижения кровотока фиксируется в
результате ограничения гемоциркуляции до 70-80% от неизмененного
(Jacewicz M., Kiessling M. and Pulsinelli W.A., 1986). Авторы обнаружили, что
при скорости кровотока менее 50-55 мл на 100 г ткани головного мозга в
течение 1 минуты клетки реагируют путем ингибирования синтеза белка. По
мере того как время, прошедшее после образования ишемического фокуса или
степень ограничения кровотокаувеличиваются, было определено, что
молекулярная масса синтезируемых белков уменьшается. В последнюю
очередь, как было установлено, прекращается синтез низкомолекулярных
белков теплового шока, которые, как известно, обладают свойством частичной
протекции от аноксии (Андреева Л.И., Маргулис Б.А. и Гужова И.В., 2005).
При выключении тока крови на 50% (которое соответствует скорости 35
мл/100г в 1 мин), регистрируется второй критический уровень биохимических
нарушений и энергообеспечения. Этому уровню соответствует снижение
образования АТФ в области частичной ишемии, что активирует анаэробного
гликолиз и ведет к увеличению производства лактата, Н+, лактат-ацидозу и
цитотоксическому отеку.

20
Метаболический ацидоз в зоне ишемии мозга является одной из первых
чувствительных реакций его клеток, усугубляющейся при нарастании степени
ишемии, которая определяет переход от выборочной гибели клеток к
тотальной – инфаркту мозга (Siesjo B. K. and Bengtsson F. J.,1989). Важнейшим
следствием недостатка АТФ и накопления продуктов анаэробного гликолиза
является накопление свободного Са2+
внутри клеток вследствие конкуренции
Н+ и Са
2+ за места связывания в митохондриях и эндоплазматической сети.
Избыток свободного Са2+
внутри клеток, как известно, усугубляет
оксидантный стресс, способствует выработе токсичного NO и активации ряда
внутриклеточных ферментов (Ooboshi H., Sadoshima S., Yao H. et al., 1992;
Zhang Y., Qiao L., Xu W. et al., 2017). Избыток Н+ при ацидозе, ативируют
аквапорины в области прямых контактов мембран эндотелиоцитов и
астроцитов, что способствует цитотоксическому и перикапиллярному отеку
(Nakada T., 2015), усугубляющему метаболизм клеток.
Третий критический уровень устанавливается при дальнейшем
ограничении кровотока до 25-30% от исходного (Гусев Е.И. и Скворцова В.И.,
2001). Эти условия гемодинамики (скорость кровотока - 20 мл / 100 г в 1 мин)
приводят к дефициту энергии и, как следствие, дефициту высвобождения
значительных количеств токсичных возбуждающих аминоацидергических
нейромедиаторов. Органичение кровотока, соответствующее третьему уровню
пнриводит к уменьшению синтеза аденозинтрифосфата и, как следствие
дисфункции каналов активного переноса ионов, дестабилизации мембраны
нейроцитов, резкому увеличению концентрации медиаторов, особенно
глутамата(Glu) (эксайтотоксичность). Авторы, изучавшие метаболические
нарушения мозга на третьем критическом уровне гемодинамического
ограничения, обратили внимание на ступенчатое и взаимозависимое снижение
концентрации АТФ, синтеза белка и скорости кровотока в 1, 6 и 12 ч после
частичного лигирования средней мозговой артерии (Mies G., Ishimaru S., Xie
Y. et al., 1991).

21
При уменьшении кровотока до скорости 10-15 мл/100 г на 1 минуту (что
соответствует только 15-20% от исходного параметра гемоциркуляции)
наступает четвертый критический уровень. Во время его наступления ионные
градиенты мембран клетки сглаживаются и развивается их аноксическая
деполяризация (Hansen A.J., 1985). Автор обнаружил, что отсутствие O2 в
течении нескольких минут при данном гемодинамическом ограничении
вызывает обратимую потерю функции из-за временного увеличения
проводимости плазмолемы для ионов K+. Это приводит к снижению
резистентности клеточных мембран и их гиперполяризации. После нескольких
минут аноксии четвертого критического уровня происходит неселективное
увеличение проницаемости мембран, приводящее, в свою очередь, к
накоплению внутри клеток Ca2+
и к гибели нейроцитов.
Таким образом, согласно концепции порогов поддержки клеточной
жизни, устанавливающихся с течением времени при различных вариантах
ограничения кровотока в области ишемии, сначала ингибируется синтез белка
(на пороге 0,55 мл/г/мин), затем в ишемизированных клетках устанавливается
анаэробный гликолиз (при 0,35 мл/г/мин), затем происходит высвобождение
из клеток медиаторов и токсических молекул малой величины и развиваются
энергетические расстройства (при скорости кровотока 0,20 мл/г /мин). При
более низкой скорости устанавливается аноксическая деполяризация мембран.
Благодаря другим исследованиям была заложена основа для выделения в
области ишемии-гипоксии подотделов, различающихся интенсивностью
абсорбции O2, глюкозы и степенью тяжести биохимических нарушений. Было
показано, что поглощение O2 и глюкозы в клетках ограничивается
пропорционально уменьшению объема гемоциркуляции: оно больше
нарушается в центральной зоне ишемии и значительно слабее в зоне
демаркации (Scheinberg P., 1991). Область мозга, соответствующая заметному
снижению скорости кровотока до значения менее 0,1 мл/г/мин, повреждается
необратимо быстро. Именно в ней в течение 6-8 мин образуется «сердцевина»
или «ядро» ишемии (Fisher M. and Garcia J.H., 1996).

22
Снаружи области мозга, в которой скорость кровотока снижается до 0,15
мл/г/мин и ниже развиваются биохимико-энергетические нарушения,
соответствующие четвертому критическому уровню, что бысто приводит к
гибели нейроцитов в указанной области. Однако через несколько часов
снаружи нее начинает формироваться область, содержащая ишемические, но
живые клетки. Эта область мозга, расположенная по периферии ядра ишемии,
соответствует зоне «ишемической полутени» или Пенумбры. В ней кровь
перемещается через сосуды со скоростью выше 20 мл / 100 г на 1 мин; Это
значение скорости кровотока было названо «критическим» (прил., рис. 2).
В зоне полутени, согласно Warfow C.P. et al. (1996), энергетический
метаболизм сохраняется, поэтому не происходит патологических изменений
клеток и выражаются только метаболические и функциональные нарушения.
Авторы считают, что функция нервных клеток в области полутени снижается,
потому что метаболические потребности ткани не удовлетворяются в полной
мере; Клетки остаются жизнеспособными, так как ионный баланс по сторонам
плазмолеммы клеток поддерживается на минимально допустимом уровне, при
котором сохраняется электрический потенциал клеток.
Данными авторами также было определено значительное усиение
чувтствительности нейроцитов зоны пенумбры к повреждающим
воздействиям, происходящее с течением времени. Резкое повышаение
чувствительности нейроцитов в этой зоне авторы связывают с падением
количества O2 и глюкозы, которое способны утилизировать клетки пенумбры.
Основной стимул, усугубляющий дальнейшее повреждение нейроцитов в
пенумбре, по мнению авторов, является дальнейшее падение перфузионного
давления крови. Наиболее частыми причинами его дефицита являются
увеличение моторики больного после ишемического инсульта,
постгидратационная гиповолемия, неадекватная гипотензивная терапия.
В связи с этими данными периинфарктная пенумбра должна
рассматриваться как объект терапевтического вмешательства при ишемии: в

23
отсутствие эффективной терапии происходит постепенное увеличение размера
инфаркта. Эту область мозга можно сохранить, поддерживая адекватную
перфузию мозга и используя нейропротективные препараты. Поэтому
основной целью терапии в первые часы и дни после развития невнятного
мозга является именно zona penumbra.
В одном из других исследований была выявлена закономерность
развития нарушений гемоциркуляции при условии восстановления тока крови
после острого церебрального инфаркта спустя 2 минуты от начала окклюзии
артериального сосуда мозга (Tomita M., 1993). Автором было показано, что
при данных условиях начальный этап постишемической гиперемии заменяется
постишемической гипоперфузией, обусловленные первичным началом
метаболических нарушений в ишемической части мозга (прил., рис. 3).
Развитие гипоперфузии после острой мозговой ишемии, как оказалось,
предшествует стадии постишемической гиперемии или «роскошной
перфузии». На этот факт впервые обратил внимание Tanahashi N. (1988).
Дальнейшие исследования позволили установить ее причины. Ими оказались
высвобождение вазоактивных и провоспалительных метаболитов из
ишемической части головного мозга, а также снижение вязкости крови,
развивающиеся при постишемической гиперемии (Hossmann, K.A., 1994 а,б;
Tomita M. and Fukuuchi Y., 1996). Было определено, что в механизмах постишемической гиперемии, вызванной реканализацией ранее
обтурированного просвета артерии, принимает участие циркуляция
значительного объема крови через коллатерали и изменение нейрогенных
вазодилататорных механизмов (Macfarlane R., 1991). Наблюдаемый в данных
условиях избыточный поток крови не соответствует метаболическим
потребностям ишемических, но живых клеток головного мозга, поэтому ограничение их метаболизма сопровождается уменьшением доли
экстрагированного O2.
Приуменьшенииколичестватокакровинижезначения,
предшествующего ишемии, возникающей после стадии постишемической

24
гипоперфузии, наблюдаются метаболические нарушения в ишемических
клетках головного мозга, развивающиеся в нейроцитах через определенный
период времени после начала ишемического воздействия. Токсичные
метаболиты, высвобождающиеся из при этом поврежденных клеток,
вызывают активацию микроглии. Последние в результате активации начинают
продуцировать провоспалительные факторы, которые увеличивают
проницаемость сосудистой стенки. Следствием этого процесса является отек
нейропиля (промежутки между стенкой сосуда, нервные волокна и клеточные
элементы головного мозга), отек клеток астроглии и сжатие просвета сосудов
(Hossmann K.A, 1994 а,б; Hallenbeck J.M., 1996, Iadecola S., 1999, Tomita M.
and Fukuuchi Y., 1996). В результате реакций, вызванных активацией
микроглии, восстановление артериального кровотока является неполным. Это
явление было названо «невосстановленный поток крови» (no-reflow).
При ишемии реагирует и стенка сосудов, особенно наиболее мелких.
Она паретически расширяется в ответ на ишемическую гиперкапнию
(Ladecola S., 1998). Автор приходит к выводу, что в постишемический период
функциональная активация мозга не приводит к адекватному притоку крови.
Это отличает гемоциркуляторно-функциональные связи в ишемически
измененном мозге от пропорционального соотношения между объемом
кровотока и функциями интактного мозга, при котором уровень кровотока
тесно связан с его метаболическими потребностями.
Было также обнаружено, что со временем метаболические и
биоэнергетические нарушения, первоначально сформировавшиеся в ядерной
зоне ишемии, впоследствии распространились на периферию, что ведет к
появлению и расширению зоны полутени (Ginsberg M.D. and Pulsinelli W.A.,
1994). Авторы обнаружили, что продолжительность полутени, ее объем и
степень нарушений биоэнергии являются индивидуальными для каждого
пациента. Эта длительность полутени - это период, в течение которого
терапевтические меры наиболее эффективны (так называемое
«терапевтическое окно»).

25
«Терапевтическое окно» при острой фокальной церебральной ишемии,
как оказалось, намного короче, чем в глобальной (Ginsberg M.D. and Pulsinelli
W. A., 1994). Авторы показали, что глобальная церебральная ишемия,
возникающая в результате остановки сердца, ведет только к изменениям
нейроцитов в наиболее уязвимых регионах мозга, чувствительных к ишемии
(пирамидальные нейроциты гиппокампа из его области CA3, нейроциты
дорсолатеральной части полосатого тела). Таким образом, патологические
изменения в полосатом теле были зарегистрированы через 4 часа, в
гиппокампе - через 2 дня после развития глобальной ишемии. Следовательно,
терапия при глобальной ишемии может быть эффективной даже через 24-48
чсов после ее начала.
В то же время формирование церебрального инфаркта вследствие
очаговой ишемии заканчивается в течение 3-6 часов с момента появления его
клинических симптомов (Hallenbeck J.M., 1996). Степень выраженности и
длительности формирования морфологического повреждения головного мозга
вследствие фокальной ишемии зависит от чувствительности той или иной его
области (Dereski M.O. et al., 1993).
Исследование динамики формирования очагового ишемического
повреждения головного мозга позволило установить, что «завершение»
инфаркта из-за расширения области полутени продолжается обычно в течение
длительного времени – до 48-72 часов после развития инсульта, возможно
дольше при условии, что существует постоянный отек головного мозга и
другие долгосрочные последствия ишемии. Это подтверждают данные,
полученные различными методами: иммуногистохимических (Matsumoto K. et
al., 1994), использование метаболического красителя (трифенилтетразолин
хлорида), а также современные модификации магнитного резонанса и
позитронно-эмиссионной томографии (Pappata S Et Al., 1993).
Постишемический отек головного мозга, как выяснилось, является
основной причиной, которая препятствует сокращению роста зоны пенумбры

26
в очаге ишемии головного мозга. Отек способствует расширению области
ишемической полутени в течение 1-й недели после инфаркта головного мозга
(Chiamulera C. et al., 1993). Со второй недели, как показали эти авторы,
жидкость в области ишемии начинает резорбироваться, макрофаги удаляют
мертвые клетки и зону инфаркта, а также зону полутени, их размер
уменьшается за счет процессов прогрессирующей клеточной атрофи.
Тот факт, что ишемическая полутень и «терапевтическое окно»
существовуют в течение длительного времени, подтверждется рядом
клинических наблюдений, позволяет надеяться на разработку эффективных
нейропротективных средств, которые уменьшают чувствительность клеток
мозга к ишемии и способствуют их выживанию. Стратегия терапии
ишемического церебрального инфаркта в настоящее время является поиском
различных критериев для определения состояния ишемической полутени и
границ «терапевтического окна», прежде всего клинического и
морфологического. Некоторые исследователи рассматривают клиническую
прогрессию инсульта через 12-24 часа после его развития и ухудшение
состояния пенумбры, вызванное трансформацией переходных функциональ-
ных изменений в необратимый морфологический дефект (инфаркт), как
взаимозависимые клинические вмешательства. Эти явления особенно
выражены при геморрагической трансформации фокуса инфаркта (Fisher M.
and Garcia J.H., 1996). Авторы считают, что такая взаимосвязь возможна с
учетом неблагоприятного влияния гипергликемии, системного ацидоза и
отдаленных влияний ядра ишемии на область пенумбры.
Чтобы определить стратегию и тактику постишемической терапии,
необходимо определить величины пенумбры и временные рамки
«терапевтического окна». Установление данных параметров на оснавание
анализа только клинических признаков неприемлемо, так как начало
клинических проявлений всегда предшествует трансформации изменений
ткани мозга в необратимые (Ginsberg M.D., 1994). Кроме того, даже

27
минимальное повреждение функционально важных зон мозга может
способствовать развитию выраженного неврологического дефицита. С другой
стороны, с образованием ишемического инфаркта возможна частичная или
полная обратимость функционального дефекта вследствие регенеративных
процессов, протекающих в пенумбре, повышения клеточной пластичности и
образования новых синаптических связей (Ginsberg M.D., 1994).
Таким образом, базовые исследования в области процесса развития и
стратегии терапии ишемического повреждения мозга свидетельствуют о
формировании в первые часы и дни после фокальной циркуляторной гипоксии
зоны пенумбры – основной мишени для терапевтических воздействий. Наряду
с этим была создана и доказательно обоснована концепция «терапевтического
окна», значительно изменившая практические подходы к ведению больных в
остром периоде инсульта. Эти фундаментальные события явились мощными
стимулами для разработки новых стратегий нейропротекции клеток в
ишемизированном участке мозга.
1.3. Молекулярные механизмы ишемического повреждения клеток головного мозга: немедленные и отсроченные
Как следствие многолетних многодисциплинарных исследований в
процессе ишемического повреждения головного мозга было определено, что
при развитии очаговой ишемии головного мозга цепь взаимосвязанных
молекулярных реакций происходит в ядерной зоне ишемии и прилегающей
пенумбры в течение первых часов ишемии (Olney J.W.E., McGeer J.W. and
Oiney P., 1978; Choi D.W., 1993; Olney J.W.E., 1994; Choi D.W. and Koh J.Y.,
1998). Цепь этих немедленных реакций обычно называется «каскадом
биохимических реакций» или «глютамат-кальциевым каскадом», который
является объектом терапевтического сдерживания и содержания периода
«терапевтического окна». Результатом или морфологическим выражением
немедленных реакций глютамат-кальциевого каскада является формирование
очага некроза в области ишемии.

28
Патогенетическими механизмами немедленных реакций глютамат-
кальциевого каскада реакций являются: 1) снижение мозгового кровотока, 2)
Gluаматная «эксайтотоксичность», 3) внутриклеточное накопление кальция, 4)
активация внутриклеточных энзимов, 5) повышение синтеза оксида азота и
развитие оксидантного стресса, 6) экспрессия генов раннего реагирования
(Гусев Е.И. и Скворцова В.И., 2001; Choi D.W., 1993; Olney J.W.E., 1994).
Отсроченныи механизмами очагового ишемического повреждения
головного мозга, которые развиваются при ишемии на 3-5-е сутки, являются:
локальная воспалительная реакция, нарушения микроциркуляции крови,
повреждение гемато-энцефалического барьера и апоптоз клеток (Голубев А.М.
и др., 2006; Cohen G.M. еt аl., 1992; Nowak T.S., Nowak J. and Kiessling M.,
1999).
В результате ряда исследований (Гусев Е.И., Скворцова В.И., 2001, Choi
D.W., 1993, Olney J.W.E., 1994) было обосновано выделение трех основных
стадий глютамат-кальциевого каскада: 1) индукция реакций, 2)
амплификация,в процессе котороой накапливается разрушающий потенциал
образовавшихся токсических молекул, синтез которого усиливается, и 3)
экспрессия конечных каскадных реакций, которые непосредственно приводят
к гибели клеток (Приложение 4, рисунки 4 и 5).
Индукция, по мнению авторов, выражается нарушениями переноса
ионов и эксайтотоксичностью Gluамата. Механизм индукции глютамат-
кальциевых каскадных реакций вызван уменьшением аденозинтрифосфата в
ишемической зоне, что ведет к компенсаторной активации анаэробного
гликолиза, что, в свою очередь, вызывает увеличение уровня неорганического
фосфата, увеличение образования Лактата и ионов Н+. Сочетание данных
процессов определяет образование метаболического ацидоза. Кроме того,
расщепление нейроцитального аденозинтрифосфата ведет к «обесточиванию»
ферментной системы Na+
/K+
-ATФ-азной, которая контролирует
энергозависи-мый ионный транспорт (Choi D.W., 1990).

29
Нарушение активного ионного транспорта обусловливает пассивный
отток ионов К+
из клеток и приток ионов Са2+
; ионы Н+
продолжают
накапливаться внутри клеток. Наряду с первичным пассивным притоком
ионов Са2+
в клетки через потенциал-зависимые Сa2+
-каналы в связи с
дефицитом O2 и аденозинтрифосфата нарушается связывания кальция в
митохондриях и эндоплазматическом ретикулуме (Zablocka B. and Domanska-
Janik K., 1996). Эти события вызывают деполяризацию клеточных мембран.
Из-за конкуренции ионов H+ и Ca
2+ на тех же участках связывания и
аккумуляции кислотных валентностей внутри клетки значительное количество
ионов Ca2+
может выделяться из органелл (Kristian T. and Siesjo B.K., 1997,
Khalaf A. and Babiker F., 2016). Накопление ионов Ca2+
внутри клеток при
церебральной ишемии создает перегрузку митохондрий, в результате чего
процесс окислительного фосфорилирования ослабевает, а катаболические
процессы усиливаются (Orrenius S. et al., 1992). По мнению авторов,
диссоциация окислительного фосфорилирования и катаболизма сопровож-
дается переходом ионов Са2+
в активную форму путем взаимодействия с
внутриклеточным рецептором кальмодулина, который индуцирует активацию
кальмодулин-зависимые протеинкиназы, липазы и эндонуклеазы, которые в
конечном итоге приводят к фрагментации ДНК и гибели клеток.
Следовательно, уже в самом начале патобиохимического каскада,
«запущенного» дефицитом макроэргических молекул, начинается процесс
внутриклеточного накопления ионов кальция, что является одним из
ключевых механизмов деструктивных процессов, заложенных в основе
некротической гибели нейроцита (Morley P. et al., 1992).
Наряду с ионным дисбалансом, возникающим на этапе индукции глютамат-
кальциевого каскада, причиной немедленной гибели клеток мозга в течение
первых минут и часов ишемического инсульта, согласно гипотезе
«эксайтотоксической смерти нейроцитов», сформулированной по результатам
эксперимента (Olney J.W.E., 1994), является накопление возбуждающих

30
аминоацидергических нейротрансмиттеров (глютамата и, возможно,
аспартата), которым характерна цитотоксичность. Увеличение количества
высвобождаемых медиаторов из поврежденных нейроцитов и/или уменьшение
скорости, а также интенсивности их повторного захвата нейроцитами и
астроцитами из экстрацеллюлярного пространства, согласно данной гипотезе,
создают условия для накопления в нейропиле больших количеств
цитотоксичных медиаторов. При взаимодействии избыточных количеств
данных медиаторов с перевозбужденными постсинаптическими рецепторами
развиваются повреждения синапсов.
Последующие экспериментальные исследования механизма на
начальном этапе Glu-кальциевого каскада подтвердили избыточный выпуск
Glu и аспартата из окончаний ишемизированных нейроцитов в межклеточное
пространство (Obrenovitch T. P and Zilkha E., 1995; Hegstad E. et al., 1996). В
этих исследованиях было показано, что освобождение этих
нейротрансмиттеров происходит в течение 10-30 минут с момента начала
острой фокальной окклюзии мозгового сосуда. Этот выпуск, по мнению
авторов, является следствием нарушения активного ионного транспорта и
деполяризации пресинаптических мембран. Эти исследования также показали,
что концентрация выше указанных нейротрансмиттеров возвращается к
исходному уровню через 30-40 мин после восстановления кровотока.
Множество доказательств нейронотоксичности избытка возбуждающих
медиаторов привело к трансформации данной гипотезы, практически, в
общепризнанный факт. Эти данные недавно дополнили сведения о подавлении
пофреждающего эффекта Glu через индукцию мутации его гена,
взаимодействующего с каспазой-1, запускающей апоптотическую гибель
клеток (Tang N., Wu J., Zhu H. et al., 2017).
Были также получены уточняющие сведения о том, что высвобождение
значительных количеств возбуждающих медиатров происходит лишь при
достаточно тяжелой церебральной ишемии – в зоне ишемического ядра
(Obrenovitch T.P. and Zilkha E., 1995).

31
Стадия амплификации (усиление потенциала повреждения) связана с
внутриклеточным накоплением ионов кальция, распространением
глутаматной «Экситотоксичности», распространением «депрессии» и
опосредованная цинк «экситотоксичности».
Прежде всего на этом этапе продолжается увеличение концентрации
внутриклеточных ионов Са2+
(Choi D.W., 1993). Частично это можно
объяснить увеличением образования в условиях энергодефицита инозитол-
1,4,5-трифосфата (1Р3), который вызывает высвобождение кальция из депо в
клетках (Mitani A., 1993). Вместе с тем Са2+
поступает внутрь клеток извне
через потенциал-зависимые кальциевые каналы, а также при помощи
мембранных молекул-переносчиков, обменивающих ионы Na+ на ионы Са
2+.
Накопление внутриклеточного Са2+
способствует миграция альбуминов
сыворотки крови через поврежденный гематоэнцефалический барьер(Linde R.,
Laursen H. and Hansen A.J., 1996). Оказалось, что присутствие альбумина в
межклеточном пространстве мозга, способствует длительному раскрытию
агонист-зависимых Са2+
-каналов NMDA -рецепторов, что, по мнению
авторов, увеличивает S ишемического повреждения, так как потенцирует Glu
"эксайтотоксичность".
Избирательная чувствительность разных областей мозга к увеличению
концентрации возбуждающих медиаторов, как оказалась, связана с
плотностью расположения нейроцитов (Siesjo B.K. and Bengtsson F.J., 1989).
Авторы установили, что глютамат, выделяемый в синапсах одной
деполяризованной клетки, индуцирует деполяризацию и высвобождение Glu в
ближайших нервных клетках.
В зоне пенумбры количество Glu увеличивается в меньшей степени, чем в самом центре ишемии (Obrenovitch T. P. and Zilkha E., 1995). Из этого
следует, что в зоне пенумбры механизмом Glu «эксайтотоксичности»
представляет собой процесс метаболических нарушений, ространяющийся
cпонтанно от нейронов в центре ишемии к нейроцитам, расположенным в зоне

32
пенумбры по механизму «домино» (Siesjo B.K. and Bengtsson F.J., 1989;
Obrenovitch T.P. and Zilkha E., 1995). При этом, по данным авторов,
нейроциты, поврежденные сверхвыбросом глютамата, индуцируют
дальнейший выброс нейротрансмиттера ближайшиими нейроцитами.
Другим механизмом немедленной гибели или обратимого повреждения
нейроцитов, развивающимся в зоне пенумбры, является «распространяющаяся
депрессия», или «периинфарктная деполяризация» (Nedergaard M. and Hansen
A.J., 1993). Этот хорошо известный феномен преходящего нарушения ионного
градиента мембран, волна которого медленно движется по связям между
нейроцитами и астроцитами головного мозга (Somjen G.G., Aitken P.G. and
Czeh C.L., 1992). Морфологически при проходжении на периферию волны
депрессии описано сжатие внеклеточного пространства на 50% за счет
набухания астроцитов и дендритов нейроцитов. В момент распространения
волны было определено временное увеличение концентрации ионов K+ во
внеклеточном пространстве, а ионов Na+ и Са
2+ – внутри клеток, что ведет к
открытию анионных каналов для молекул бикарбоната и воды.
Бикарбонатный сдвиг, в свою очередь, ведет к формированию ацидоза в
нейроцитах и внеклеточном пространстве, В астроцитах, напротив,
развивается алкалоз. Данные изменения клеток, по мнению авторов, являются
обратимыми.
Роль «периинфарктной деполяризации» в расширении области инфаркта
головного мозга подтверждается корреляцией между объемом зоны некроза и
числом волн депрессии (Mies G.F., Iljima T. and Hossmann K., 1993). Каждая
волна увеличивает объем ядра церебрального инфаркта примерно на 23%.
Было также определено, что механизмы немедленной нейроцитальной
ишемии в дополнение к Са2+
участвуют и другие катионы. Анализ
селективной гибели нейроцитов в эксперименте с транзиторной глобальной
ишемией, доказал существование цинк-опосредованной «эксайтотоксич-
ности» (см. прил., рис. 5).

33
Ранее была установлена роль внутриклеточного цинка как компонента
многих металлозависимых энзимов и транскрипционных факторов,
нейротрансмиттера и нейромодулятора голвоного мозга (Frederickson C.J.,
1989). Значительное увеличение количества Zn2+
выявлено сразу после
стимуляции нейроцитов в пресинаптических везикулах и синаптической щели;
наряду с этим выявлена способность ионов Zn2+
ингибировать активность
NMDA рецепторов (Choi D.W. and Koh J.Y., 1998).
Цинк-опосредованная нейротоксичность была впервые сформулирована
N. Tonder и соавт. (1990). С временной глобальной ишемии внутри аксонов
нейроцитов СА-3 гиппокампа было выявлено истощение ионов Zi2+
ионы,
тогда как концентрации этого иона в аномально высоких значениях были
зарегистрированы в телах нейроцитов этой области мозга, являющейся
наиболее восприимчивой к ишемии. Такое перемещение ионов Zn2+
и
нейроцитальная дегенерация тормозились при гипотермии головного мозга
(Johansen F.F. et al., 1993).
Нейротоксическая роль ионов Zn2+
подтверждается тем, что их
перемещение из пресинаптических терминалей в постсинаптические
нейроциты непосредственно предшествует явлениям их дегенерации (Choi
D.W. and Koh J.Y., 1998).
В работах S.L. Sensi и соавт. (1997) было показано, что, подобно
кальцию, цинк, проникая в избытке внутрь клеток, может вызывать их некроз
или апоптоз, в зависимости от длительности и интенсивности экспозиции.
Проникновение ионов Zn2+
в клетки происходит через любые кальциевые
потенциал-зависимые каналы (в обмен на внутриклеточные ионы Na+;
агонист-зависимые, сопряженные с NMDA-рецепторами).
Избыток внутриклеточного цинка ведет к снижению уровня
внутриклеточной АТФ, поэтому он может существенно нарушать процессы
гликолиза, опосредуя накопление промежуточных его продуктов и
дигидроксиацетонфосфата (Lee J.M., Zipfel G.J. and Choi D.W., 1999). Темпы

34
цинк-опосредованной гибели клеток сокращаются при введении пирувата,
который в норме образуется в результате метаболических реакций, угнетаю-
щих избыток цинка.
Стадия экспрессии обеспечивается необратимыми механизмами
активации клеточных энзимов, окислительным стрессом и чрезмерным
синтезом оксида азота, накопления других низкомолекулярных
цитотоксических соединений,превращими клетки к некротической смерти.
Избыточное внутриклеточное накопление ионов Ca2+
и переход кальция в
активную форму путем связывания с рецептором кальмодулина и активацией
кальмодулин-зависимых внутриклеточных энзимов (Berridge MJ and Triangi S.,
1985) приводит к диссоциации окислительного фосфорилирования в
митохондриях и усиление катаболизма, вызывающих апоптоз (Orrenius S.,
1992).
"Запуск" каскада ферментативных реакций ведет к множественным
повреждениям биомакромолекул и, в конечном счете, к гибели клеток (Olney
J.W.E., 1994). Особо опасен распад фосфолипидов плазматической мембраны
и мембранах органелл. Через несколько минут после начала ишемии,
уничтожается около 16% мембраного фосфатидилэтаноламина и
высвобождается около 37% свободной арахидоновой кислоты (Yoshida S. еt
al., 1980). Между тем, ишемическое расщепление мембранных фосфолипидов,
таких как арахидоновая кислота, приводит к высвобождению ее в цитозоль.
Метаболизм арахидоновой кислоты сопряжен с образованием простагланди-
нов, тромбоксанов, гидрокси- и гидроперокси жирных кислот, лейкотриенов,
липоперекисей и реактивных свободных радикалов, таким образом
значительно интенсифицируя процессы свободнорадикального окисления и
перекисного окисления липидов (Дамбинова С.А. и Каменская М.А., 1996;
Packer L.L. еt al., 1992; Ugidos I.F., Santos-Galdiano M., Pérez-Rodríguez D. et al.,
2017).
Радикалы O2, образующиеся в результате постишемического
оксидантного стресса, обладают высочайшей реакционной способностью. Они

35
вызывают окисление биомакромолекул, инициируют цепные процессы
перекисного окисления в мембранных липидах (Carney J. M., Starke-Reed P. E.,
Oliver C. N. et al. 1991; Yu K. L. et al., 1994), непосредственное окислительное
повреждение нуклеиновых кислот и белков.
Гидроперекиси, образующиеся при перекисном окислении липидов,
неустойчивы. Их распад ведет к появлению разнообразных вторичных и
конечных продуктов этого окисления, представляющих собой высокотоксич-
ные соединения (диеновые конъюгаты, шиффовы основания и др.), которые
оказывают вторичное повреждающее действие на мембраны клеток. Как
следствие, образуются сшивки биополимеров, определяются набухание
митохондрий и разобщение окислительного фосфорилирования, инактивация
тиоловых энзимов, участвующих в дыхании и гликолизе, дальнейшее разру-
шение липидной основы мембран (Завалишин И.А. и Захарова М.Н., 1996).
Увеличение концентрации свободной арахидоновой кислоты
дополнительно ингибирует митохондриальное дыхание, а также оказывает
непосредственное цитотоксическое действие. Благодаря своим амфофили-
ческим свойствам арахидоновая кислота может соединяться с цитоплаз-
матической мембраной, встраиваясь в мембранные липиды. Жирные кислоты
(в том числе арахидоновая) способны объединяться в мицеллы, имеющие
детергентные свойства, которые позволяют им расплавлять цитоплазмати-
ческие мембраны. Образующиеся при окислении свободных жирных кислот
спирты, кетоны и альдегиды так же повреждающе действуют на мембраны
нейроцитов (Scheinberg P., 1991).
Реакции окислительного стресса тесно связаны с процессами
метаболизма энергии и Glu «эксайтотоксичности», образуя закрытые круги
патологических преобразований. Таким образом промежуточные продукты
перекисного окисления липидов тормозят активность аминотрансферазной
ферментной системы, усугубляя изменения энергозависимого ионного
транспорта (Scheinberg P., 1991), подавляют энзим Glu-аминсинтетазу в

36
астроглии, нарушая обратный захват Glu и способствуя его накоплению в
синаптической щели (Oliver C. N. et al., 1990)..Измененяя структуру белков
рецепторов, свободные радикалы нарушают их функции, замедляя её в одних
условиях и активируя в других. Открытие потенциал зависимых и агонист
зависимых ионных каналов, особенно с Glu рецепторами, ведет к быстрому
входу ионов Са2+
и Na+ в нейроцит, что усугубляет все описанные ранее
реакции Glu-кальциевого каскада, а также обусловливает специфическую
активацию кальмодулина и NO-синтетазы.
Все больше и больше проявляется интерес к участию в механизмах
острой мозговой ишемии свободно-радикального соединения оксида азота
(NO), который является многофункциональным физиологическим
регулятором (Moncada S., Palmer R.M. и Higgs E.A., 1997). Оксид азота обычно
считается первым представителем нового семейства сигнальных молекул,
которые обладают свойствами нейротрансмиттера. В отличие от
традиционных нейротрансмиттеров, NO не сохраняется в синаптических
везикулах нервных окончаний и высвобождается в синаптическую щель путем
свободной диффузиии, а не механизмом экзоцитоза. Молекула синтезируется
с участием энзима NO-синтетазы (NOS) из его предшественника, L-аргинина
аминокислоты. Свойство NO, вызывающее биоэффект, определяется размером
небольшой молекулы и ее самой высокой реакционной способностью,
возможностью диффузии в тканях, включая нервную. Это послужило
основанием назвать NO ретроградным мессенджером (Damon T.M. et al.,
1992).
Кальций-кальмодулин-зависимость тканевой изоформы NOS обусловли-
вает усиление синтеза NO при увеличении значений внутриклеточного Са2+
.
Таким образом, в результате процессов, ведущих к накоплению ионов Са2+
в
клетке (энергетический дефицит, изменения активного ионного транспорта,
Glu «эксайтотоксичность», оксидантный стресс), что сопровождается
повышением уровня NO (Раевский К. С., 1997).

37
Показано, что NO оказывает модулирующее влияние на синаптическую
передачу (Moncada S., Palmer R.M. and Higgs E.A., 1997), на функциональное
состояние рецепторов Glu. Механизм токсичности NO, по мнению авторов,
предполагает ковалентную модификацию белков при взаимодействии с их
тиоловыми группами, и прямое повреждение ДНК.
Активация процессов апоптоза считается конечным компонентом
цепочки событий, приводящих к гипоксической гибели клетки.
Программированную. смерть клетоки способны индуцировать измененные
липиды клеточных мембран. Их действие может быть связано с изменением
содержания цитозольного кальция, опосредующего действие активных форм
O2 на различные сигнальные системы, а также с прямым воздействием на
экспрессию генов, участвующих в запрограммированной гибели клеток
(Nowak TS, Nowak J. and Kiessling M., 1999).
Высказано предположение, что механизмы, связанные с повреждающим
действием активных форм O2, эволюционно записываются и используются
клеткой для реализации программы апоптоза. Уже найдены гены, связанные с
регулированием редокс-состояния клетки , генераторов баланса активных
форм O2 и антиоксидантных механизмов.
Общеизвестно, что центральная роль в развитии апоптоза принадлежит
изменению проницаемости митохондриальных мембран. В митохондриальных
мембранах существует ряд апоптогенных факторов: цитохром C, AIF (фактор,
индуцирующий апоптоз), эндонуклеазой G, SMAC / DIABLO и OMI / HTRA2
белками. Под воздействием AIF открываются митохондриальные поры с
экспрессией и высвобождением апоптозных факторов в цитозоль. Цитохром C
в цитоплазме индуцирует образование апоптосомы, активирующей
высокомолекулярный ансамбль каспазы-9. AIF из митохондрий транслоцирует в ядро, где он индуцирует фрагментацию ДНК и конденсацию
периферического хроматина. Эндонуклеаза G также транслоцируется из
митохондрий в ядро и вызывает деградацию ДНК независимо от каспаз или
дезоксирибонуклеазы. SMAC / DIABLO связывается в белках цитоплазмы

38
рода IAP (ингибитор протеинов апоптоза), которые ингибируют каспазы. OMI
/ HTRA2 представляет собой сериновую протеазу, участвующую в регуляции
как каспаззависимого, так и каспаз-независимого пути апоптоза, а также
блокирует связывание IAP с каспазами (Голубев A.M. и соавт., 2006).
Открытие пор происходит из-за окисления тиольных групп цистеин-
зависимой области белка внутренней митохондриальной мембраны (АТФ /
АДФ-антипортер), которая фактически превращает его в проницаемую
неспецифическую канальную пору. Образование поры канала регулируется
белками рода BCL-2 и изменениями концентрации ионов кальция в
митохондриальном матриксе. Открытие пор превращает митохондрии из
«электростанций» в «печь» окисляющих субстратов без образования АТФ.
Было обнаружено, что нарушение O2 режима тканей, гиперпродукция
экситотоксических аминокислот, уменьшение «нормального» накопления
митохондриями Ca2+
, повреждение митохондриальной мембраны AIF
усиливают открытие пор и высвобождение апоптогенных белков из
поврежденных митохондрий.
Лизосомальные протеазы также повреждают митохондриальные
мембраны и способствуют высвобождению цитохрома С, иных апоптогенных
факторов и активации каспаз. Создание комплекса апоптосомы, состоящего из
цитохрома C, Apaf-1 (Апоптотический протеаз-активирующий фактор-1) и
каспазы-9, играет важную роль в реализации сценария запрограммированной
гибели клеток, вызванного гипоксией (Cohen G.M. et al., 1992).
1.4. Основные морфофункциональные особенности клеток мозга
Нервная ткань головного мозга, по укоренившемуся мнению,
формируется из клеток, которые принадлежат к нескольким линиям
дифференцировки. Клетки основной линии – нейроциты и клетки macroglia
(астроциты, олигодендроциты, эпендимоцитаты), имеющие нейтральное
происхождение. Они обеспечивают ключевым образом, синаптическую
передачу нервных импульсов. Клетки микроглии образуются из моноцитов

39
крови ведущих иммунных клеток. Эндотелиальные клетки и перициты
образуют стенки кровеносных капилляров.
Нейроциты головного мозга, обладающие огромным разнообразием
строения клеточных тел и отростков, связаны между собой при помощи си-
синаптических контактов и образуют в пространстве ансамбли клеток –
нервные центры. Исследования особенностей строения тел и отростков
нейроцитов, их синапсов и органелл, электрофизиологических свойств,
медиаторной передачи, связей с другими клетками, а также особенностей их
развития и патологии охватывают значительный промежуток времени от
конца XIX в. и по ныне (Жаботинский Ю.М., 1965; Поляков Г.И.,1973;
Боголепов Н.Н., 1975; Отеллин В. А., 2003; Marin-Padilla M., 2011 и др.).
Клетки макроглии, расположенные в мозге между сосудами и
нейроцитами,численно значительно доминирует нервными. Основная масса
маленьких нейроцитов настолько тесно окружена макроглиальными клетками,
что часто бывает трудно отличить эти два типа клеток (Васильев Ю.Г. и
Берестов Д.С., 2011).Несмотря на идею о определенной пропорции между
количеством нейроцитов и глиальных клеток в большинстве отделов мозга,
равным соотношению 1:4 (Поляков Г.И., 1973), в некоторых мозговых
центрах, таких как компактная часть черного вещества или глубокие слои
передней цингулярной коры. число нейроцитов значительно превышает
количество макро- и микроглиальных клеток (Дробленков А.В. и Карелина
Н.Р., 2009; Дробленков А.В., Карелина Н.Р. и Шабанов П.Д., 2009).
Нейроциты и глиальные клетки находятся в тесной физиологическом и
патологическом отношении. Итогом их длительного изучения было
формирование концепции существования в центральной нервной системе
динамических нейроцитально-глиальных процессов системы сигнализации. К
ним относятся способность макроглии к регулированию процессов
возбуждения нейроцитов, на которые глиальные клетки в условии нормы
оказывают тормозное влияние (Ginsberg M.D. and Pulsinelli W.A., 1994;
Hallenbeck J.M.,1996). Астроглия и олигодендроглия оказывает на нервные

40
клетки трофическое и пластическое влияние. Эти клетки способствуют
восполнению нейроцитальных глюкозы, лактата, РНК и аминокислот (Певзнер
Л.З., 1972; Богомолов Д.В., Пиголкин Ю.И. и Должанский О.В., 2001; Hyden
H., 1976; Iadecola С., 1999), активируют регенераторные процессы нейронов
благодаря выработке факторов роста (Dai X., Larcher L.D., Clinton P.V. et al.,
2003; Wilkins A., Majet H., Layfield R. et al., 2003).
В последние годы при электронно-микроскопических исследованиях
была подтверждена гипотеза о наличии цитоплазматических связей между
нейроцитами головного мозга позвоночных животных (Парамонова Н.Н. и
Сотников, О.С., 2008; Сотников О.С., Фрумкина Л.Е., Майоров В.Н. и соавт., 2012; Frantseva M.V., Kokarovtseva L. and Perez Velazquez J.L., 2003). По
мнению авторов, эта связь может возникнуть в условиях неполного охвата
нейроцитов глиальными клетками. Между телами нервных клеток. в области
гиппокампа СА-3, выявленны участки соприкосновения наружных клеточных
мембран, плотные и щелевые межнейроцитные мембранные контакты,
мембранные перфорации и цитоплазматическая синцитиальная связь между
нейроцитами (со всеми ее ультраструктурными признаками). Авторы не
исключают адаптационно-приспособительного значения обнаруженного ими
малоизвестного типа межнейроцитального контакта.
Астроциты (волокнистые и протоплазматические) образуют большое
количество отростков, концевые части которых окружают 80% поверхности
объема капилляров, а также тела, область синаптических контактов,
немиелинизированные и миелинизированные фрагменты нейроцитов.
Некоторые из отростков астроцитов расположены вблизи основания
эпендимоцитов, участвующих в лиоэнцефалическом барьере (Balercia G. et al.,
1992). Эти особенности расположения астроцитов указывают на их участие в
метаболизме и естественной защите нейроцитов.
Астроциты имеют относительно крупные светлые ядра, диаметр
которых находился в пределах 5–8 мкм, развитый ядрышковый аппарат. Ядра
были окружены светлым ободком цитоплазмы толщиной 1–4 мкм.

41
В цитоплазме астроцитов содержатся небольшое количество канальцев
гладкой и зернистой эндоплазматической сети, пластинчатый комплекс,
однако содержатся многочисленные мелкие митохондрии. Цитоскелет
умеренно развит в протоплазматических и хорошо – в волокнистых
астроцитах. Специфическим маркером астроцитов является белок их
промежуточных филаментов – глиальный кислый фибриллярный протеин
Отростки астроцитов соединены между собой, а также с плазматической
мембраной тел нейроцитов и миелиновой оболочки на их отростках при
помощи щелевидных и десмосомоподобных контактов. Терминальные
пластинки концевой части отростков соединены при помощи прямых и
десмосомо-подобных контактов с тонкой двухслойной базальной мембраной
капилляра (Сотников О.С., Парамонова Н.М. и Арчакова Л.И., 2010; Nag S.,
2011).
В зависимости от особенностей расположения астроцитов принято
различать: 1) клетки, отростки которых по-разному тяготеют к стенке
ближайших сосудов; 2) астроциты, тела которых расположены вблизи их
стенки – перикапиллярные астроциты и 3) клетки, тела которых расположены
вблизи тел нейроцитов – спутниковая форма астроцитов (Balercia G. et al.,
1992; Васильев Ю.Г. и Берестов Д.С., 2011).
Некоторые авторы полагают, что после рождения астроциты способны к
миграции (Zhou H.F. and Lund R.D., 1992), особенно в зоны повреждения.
Механизм их предполагаемой миграции не изучен. Способность к делению
астроцитов у взрослых рассматривают как слабо выраженную (Васильев Ю.Г.
и Берестов Д.С., 2011).
Астроциты играют важную роль в развитии множества
невропатологических и психопатологические процессы. определено, в
частности, что нейроцит при возбуждении способен к двухстороннему
взаимодействию с астроцитами, во многих случаях вызывая реакции в своих
ионных каналах после собственного возбуждения (Attwell D., 1994; Sontheimer
H. and Richie J.M., 1995). Из-за селективной проницаемости мембран

42
астроцитов для ионов K+ эти клетки регулируют активацию энзимов,
необходимых для поддержания метаболизма нейроцитов, а также для
удаления медиаторов и других агентов, высвобождаемых при активности
нейроцитов (Васильев Ю.Г. и Берестов Д.С., 2011).
Определено, что астроциты принимают участие в синтезе иммунных
медиаторов- цитокинов, и других сигнальных молекул (циклический
гуанозинмонофосфат,Оксид азота), перенесенный впоследствии в нейроциты,
при синтезе глиальных факторов роста (GDNF), участвующих в трофике и
репарации нейроцитов. Способность астроцитов активировать утилизацию
медиаторов, их восстановление и возвращение в нервную клетку, с
увеличением концентрации медиатора в синаптической щели (Hossmann K.A.
(a), 1994, Hallenbeck J.M.,1996). Данная способность астроцитов
подтверждается морфологически, так как разветвления их отростков тесно
переплетены с деревом дендрита нейроцитов, атакже охватывают терминали
отростков нейроцитов. Эта особенность взаимосвязи между астроцитами и
нейроцитами способствует формированию «волны» миграции ионов Ca2 +
между астроцитами, вызывая в некоторых областях мозга колебание в их
концентрации, которое, в свою очередь, может модулировать состояние
многих нейроцитов (Боголепов И.И., Бурд Г.С., 1981). В условиях острой
фокальной ишемии головного мозга механизмы отсроченной смерти
нейроцитов напрямую связаны с агрессивным действием возбужденной
ишемией нейроглии на еще жизнеспособные клетки периинфарктной области
(Garcia J.H. et al., 1996).
Способность астроцитов использовать медиаторы, в частности Glu , и ее
метаболиты из синаптической щели и отдавать их аксону нервной клетки
(Скворцова В.И., 1993; Васильев Ю.Г., Берестов Д.С., 2011). Содержит
значительное адаптационно-приспособительное значение не только и не
столько в физиологических условиях. Особенные условия для реализации
данной адаптационной функции астроцитов формируются, к примеру при
фокальной церебральной ишемии, так как избыточная концентрация

43
глютамата способна вызывать необратимое повреждение нейроцитов (Гусев
Е.И. и Скворцова В.И., 2001).
Олигодендроциты представляют собой наиболее многочисленную
популяцию глиальных клеток центральной нервной системы. Тело
олигодендроцитов меньше, чем тела астроцитов: диаметр клетки составляет 5–
8 мкм, диаметр ядра не превышает 3–5 мкм. Ядра округлые или овальные,
темные. Ядрышки малы. Отростков мало, они тонкие, не ветвятся или слабо
ветвятся, и в белом веществе мозга в основном заканчиваются вокруг аксонов
и дендритов нейроцитов (Miller R. H., 1996). В отличие от астроцитов,
распределение данных клеток в мозге здоровых особей не зависит от
положения сосудов. В цитоплазме олигодендроцитов имеется много
небольших митохондрий, имеется небольшое количество канальцев
эндоплазматического ретикулума, свободных полисом и рибосом, развит
ламеллярный комплекс, но в отличие от нейроцитов тела данных клеток не
имеют развитой системы микрофиламентов. Последние находятся главным
образом в отростках отростках олигодендроцитов (Uehara M. and Ueshima T., 1985).
Олигодендроглиоциты образуют оболочки вокруг тел нервнов,
предоставляя барьер-транспортные функции, формируют миелиновые
оболочки нейроцитных отростков, участвуя в проведении возбуждения. Они
изолируют их, ускоряя проведение возбуждения и предотвращая его затухание и распространение (ирритацию) на соседние отростки (Sontheimer H., 1995).
Функция опоры вызвана как организацией миелина, так и скелетом процессов
клетки. Олигодендроциты механически поддерживают структуру отдельных
нервных волокон и их пучков.
Трофической функции олигодендроцитов связан с транспортом
питательных веществ в телах и отростках нейроцитов. Через них можно
перемещать метаболитов. Олигодендроциты, как нейроциты и астроциты
использовать лактат в качестве энергетического субстрата, и в качестве
прекурсора для синтеза углеводородных полимеров. Активность метаболизма

44
лактата oligodendroglia значительно выше по сравнению с астроцитами и
нейроцитами. Исследование также показало, что глюкоза и ее
метаболитымогут служить источником для синтеза липидов, что, вероятно,
важно для поддержания миелиновых структур (Sánchez-Abarca L. I. et al.,
2001).
Олигодендроциты также участвуют в регенерации поврежденных
отростков нервных клеток путем выделения ряда биологически активных
веществ (Nguyen K.B. and Pender M.P., 1999). Последние предотвращают
гибель поврежденных нейроцитов, инициируют и ориентируют рост их
поврежденных процессов. Кроме того, клетки олигодендролии способны
фагоцитировать остатки поврежденных осевых цилиндров и миелина.
Он выражается путем трансфекции клеток. В плазмолеме отростков
нейроцитов в области их конечной части и межузловых перехватах были
обнаружены рецепторные системы, способные реагировать на эффекты
нейроглии (Ratner N. et al., 1998).
Таким образом, к настоящему времени накапливается достаточное
количество доказательств тому, что целый ряд структурных и биохимических
особенностей организации олигодендроцитов имеют адаптационно-
приспособительное значение при взаимодействии глиоцитов с нейронами,
существенно изменяя структурно-функциональную активность последних,
особенно в патологии (Hajek T. et al., 2005; Walterfang M. et al., 2006).
Эпендимоциты – реснитчатые эпителиоморфные клетки нейрального
происхождения, выстилающии полости ЦНС. На базальной поверхности
содержат инвагинации, и длинные тонкие отростки, идущие к стенкам
микрососудов. Эпендимоциты не экспрессируют нейрофибриллярные и
микротубулин-ассоциированные белки, как нервные клетки. Для их скелета
характерен белок промежуточных филаментов виментин, экспрессируемый
также нейробластами, спонгиобластами и молодыми клетками нейрального
происхождения (Chouaf L. et al., 1989).

45
Микроглиальные, являются клетками моноцитарного происхождения; в
физиологических условиях они неактивны и составляют около 13% от общего
числа нейроглиоцитов (макро- и микроглиоцитов). По расположению
выделяют два типа неактивных микроглиоцитов отличаются положением: 1)
резидентным, распределенным между элементами нервной ткани, а также
вблизи тел нейроцитов (спутниковая форма микроглии) и 2) периваскулярные
макрофаги, расположенные вблизи базальной пластинки капилляров
(Ransohoff RM И Cardona AE, 2010; Quintero OK и Gonzalez-Burgos I., 2012).
Первые обладают малыми размерами, продолговатой формой тела клетки и
плотной сетью тонких ветвящихся отростков; второе, связанное с
гематоэнцефалическим барьером, обладают свойствами дендритных
антигенпредставляющих клеток (Ransohoff R.M. and Cardona A.E., 2010).
Живые микроглиальные клетки обладают слабой фагоцитарной активностью,
но благодаря микродвижениям и восприятию сигналов окружающей среды
они получают информацию, об ее изменениях в результате чего эти
покоящиеся клетки микроглии могут трансформироваться в функциональную
клеточную форму (Imai F., Sawada M., Suzuki H. et al., 1997, Chew L.,
Takanohashi A. and Bell M., Soulet D. and Rivest S., 2008; Jonas RA, Ti-Fei Yuan,
Yu-Xiang Liang et al., 2012).
В развивающемся мозге обнаружена также временная форма клеток
микроглии – амебоидные клетки. Последние ветвящиеся процессы не
имеютоднако, из-за филоподий, складки плазмолеммы и содержание в
цитоплазме большого количества фаголизосом способны мигрировать на
большие расстояния и воплощать активный фагоцитоз. Считают, что
присутствие в развивающемся мозгу амебоидных форм микроглицоитов является особенно необходимым в условиях недостаточности
гематоэнцефалического барьера, в которых вещества из плазмы крови
проникают в ЦНС (Kofler J. and Wiley C.A., 2011). Считается, что надстройка
из амебоидных клеток микроглии не только способствуют удалению
фрагментов нейроцитов гибнущих при развитии мозга в перинатальном

46
периоде механизмом апоптоза, но может и непосредственно обеспечивать эту
гибель. Созревая, амебоидные микроглиальные клетки трансформируются в
неактивные резидентные ветвистые микроглиальные клетки (Imai F., Sawada
M., Suzuki H. et al., 1997; Chew L., Takanohashi A. and Bell M., 2006; Soulet D.
and Rivest S., 2008).
Периваскулярный вид микроглиоцитов в зрелом мозге функционирует
как антиген-представляющие клетки. Активируясь, они выделяют
значительное количество цитокинов, включая и фактор некроза опухоли (TNF-
a) и IL-12 (Bitzer-Quintero О. К. and Gonzalez-Burgos I., 2012). В этих условиях
периваскулярная микроглия передает «сигнал опасности» резидентным
клеткам, приводя к их активации и пролиферации. Активация микроглиоцитов
выражается их трансформацией в крупные неотростчатые клетки амебоидного
фенотипа. Последние, в свою очередь, обеспечивают восстановление
целостности элементов нервной ткани ЦНС.
Эндотелиоциты и перициты. Эндотелиоциты, выстилающие стенку
кровеносных капилляров головного мозга, представляют собой плоские
эпителиальные клетки. Они расположены на очень тонкой двухслойной
базальной мембране, имеют люменальную и базальную поверхности.
Перициты расположены в зонах расслоения базальной мембраны, при
световой микроскопии определяются вокруг сосудов. Эти клетки, являются
сократительными способными регулировать движение крови в просвете
сосуда (Bennet H. et al., 1959). В крупных сосудах эта роль принадлежит
гладким миоцитам.
Базальная мембрана эндотелия капилляров мозга очень тонкая,
расположена в два слоя; ретикулярная пластинка не выражена (Гелеранская О.
А. и соавт., 2016; Bi M., Zhang M., Guo D. et al., 2016). Вдоль стенки
капилляра, обнаруживается плотный футляр, образованный концевой частью
отростков астроцитов .В перикапиллярной глиальной астроцитарной мембране
содержатся пиноцитозные везикулы.

47
Зрелые эндотелиальные клетки характеризуются низким содержанием
мембранных органелл, за исключением митохондрий и транспортных везикул.
Между клеток содержится большое количество мелких десмосомоподобных
соединений. В перицитах много актиновых микрофиламентов.
Эндотелиоциты кровеносных сосудов образуются в зрелом организме из
подобных предшественников. Существует также образование новых сосудов и
во взрослом мозге, но проявления этого процесса, в частности, после
ишемического церебрального инфаркта, становятся более очевидными (Szpak
G.M. et al., 1999).
Процесс неоваскуляризации в этом случае может быть связан с
изменениями пространственного распределения капилляров, возможностью
дегенеративного ангиогенеза, во время которого уменьшается количество
сосудов в областях с низкой энергетической активностью мозга. Поэтому
динамика активности нейроцитов и особенности метаболических процессов в
нервных центрах играют ведущую роль в процессах ангиогенеза, провоцируя
перераспределение сосудов микроциркуляторного русла (Paemeleire K., 2002).
1.5. Реактивные изменения различных клеток головного мозга в ядре ишемии и в зоне «ишемической полутени»
Oстрая церебральная ишемия запускает реакции, протекающие во всех
клеточных пулах нервной ткани. Эти реакции вызывают повреждение
нейроцитов, астроцитов, микроглиальную активации и связанные с ними
изменения нейтрофилов, макрофагов и эндотелиальных клеток, образующих
стенку кровеносных капилляров мозга. Составлено представление о развитии
ранних изменений данных клеток в очаге ишемии при переживании инсульта
(прил., Рис. 6) Динамичность (стадийность) постишемических изменений
олигодендроцитов, эпендимоцитов и перицитов не исследовалась.
Реакция олигодендроцитов в очаге ишемии является выражается
набуханием и гибелью этих клеток путем апоптоза. Способность
активированных микроглиоцитов подавлять апоптоз поврежденных

48
олигодендроцитов в культуре ткани мозга (Brandon A. M., 2007). Вне ядерной
зоны инфаркта после 10-20 дней увеличение числа олигодендроцитов. Одной
из причин перифокального глиоцитоза в коре головного мозга может быть
дифференцировка предшественников олигодендроцитов, мигрирующих из
субвентрикулярной зоны бокового желудочка. В ней у взрослого человека и
грызунов выявлены стволовые нервные клетки, способные дифференци-
роваться в предшественники олигодендроцитов, мигрирующие в мозолистое и
полосатое тела (Zhang R.L., Chopp M. and Zhang Z.G., 2013).
Изменения в эпендимоцитах в фокусе ишемии связаны с нарушенной
перфузией стенки желудочка, выраженной в расширении его просвета и при
набухании клеток. Ультрамикроскопические признаки отека эпендимоцитов,
повреждения органелл, синаптических контактов и смерти нейроцитов были
обнаружены при циркулирующей гипоксии, которая развивается в результате
гравитационной перегрузки (Пащенко П.С. и Сухотерин А.Ф., 2000).
Перициты, локализованные между листками базальной мембраны
капилляра и способные к сокращению, наряду с эндотелиоцитами и
астроцитами, уменьшают кровоток и увеличивают перфузию плазмы крови в
ядерной зоне ишемии (Liu S. et al., 2012). Данные об изменениях количества и
структуры перицитов в условиях ишемии не отражены в доступных
литературных источниках.
Каково же современное представление о последовательности и
выраженности изменений клеток мозга (нейроцитов, астроцитов, микроглио-
цитов, эндотелиоцитов) в ядерной зоне инсульта и в области «ишемической
полутени»?
Размер центрального инсульта и последующая клеточная реакция на
ишемию, определяющая состояние зоны пенумбры, варьируются в
зависимости от тяжести и продолжительности ишемии. При «мягкой»
глобальной или кратковременной фокальной ишемии поражаются только
селективно чувствительные нейроциты, тогда как по мере удлинения
ишемического периода избирательность клеточной гибели теряется, и

49
тканевое повреждение прогрессирует по общим закономерностям (Скворцова
В.И., 1993; Васильев Ю.Г. и Берестов Д.С., 2011; Kogure K., 1996),
обусловливая формирование инфарктного очага по механизмам некроза и
апоптоза.
По данным исследований, проведенных на экспериментальных моделях
инсульта в результате лигирования средней мозговой артерии у крыс были
созданы модели последовательности реакций различных клеточных пулов
мозга при острой фокальной ишемии. Конечно, существуют различия в
развитии процессов повреждения в мозге грызунов и приматов (Tagaya M. et
al., 1997), впрочем рассматриваемые общие особенности дают возможность в
определенной степени экстраполировать экспериментальные данные для
понимания клеточных механизмов острой недостаточности мозгового
кровообращения при инсульте у людей.
В нейроцитах первые изменения наблюдаются через 30 минут после
перевязки сосуда и проявляются признаками сморщивания. Через час гетерохроматиноваягруппировкаамплифицируетсявнейроцитах,
расширяется эндоплазматический ретикулум, вакуолизация, набухание
внутреннего митохондриального матрикса (Garcia J.H. et al., 1996). В течение
первых 24 часов химической гипоксии, вызванной введением хлорида
кобальта,в нейроцитах и астроцитах головного мозга набухание и расширение
просвета трубочек эндоплазматического ретикулума, ансамбля тромбоцитов,
матрикса митохондрий и, вообще, цитоплазматического отека, начиная с
перинуклеарной части клеток (Caltana L. и соавт., 2009).
Эти изменения, по мнению авторов, потенциально обратимы и
развиваются в течение первых 6 часов. Через 10-12 часов в ядерной зоне
ишемии появляются признаки необратимого повреждения клеток обычно
обнаруживаются в форме разрушения цитоплазматических и ядерных
мембран, осаждения богатых кальцием солей во внутренней
митохондриальной мембране. В зоне полутени обратимые изменения
сохраняются в течение более длительного времени. Первые клетки-«тени»

50
(погибшие нейроциты) встречаются в тканях мозга в течение 48-72 часов
после инсульта.
В соответствии с перечисленными признаками некротической смерти
нейроцитов в ишемизированной области мозга, вдоль внутренней границы его
ядра, есть клетки погибающие по механизму программированной гибели –
апоптоза (Charriaut-Marlangue C. et al., 1996; Sadoul R. et al., 1996). Первые из
них появляются в течение 2 ч после инсульта, максимальное количество
апоптотических клеток обнаруживаются после 24-48 ч, затем наблюдается
тенденция к ее снижению (Chopp M.Т. and Li Y., 1996). Однако, нейроциты
гибнущие путем апоптоза, во время ишемического повреждения
обнаруживаются чаще, распространенясь в области "ишемической полутени".
После своего образования апоптотические нейроциты могут быть обнаружены
в течение 4 недель.
Электронная микроскопия клеток новой коры зрелых крыс после
нескольких часов экспериментальной ишемии выявила измененные
нейроциты в виде гиперхромии-сморщивания (Гелеранская, О.А. и соавт.,
2016). В телах гипохромных нейроцитов, отек и расширение просвета
мембранных органелл, выраженный отек и повреждение митохондриального
матрикса отмечались отечные изменения в матриксе цитоплазмы. В
мембранах клеток этого типа не было отмечено никаких заметных
повреждений,в связи с этим авторы должны рассматривать изменения в
клетках по типу гипохромии обратимыми. Матрикс ядра и цитоплазмы
гиперхромных ишемически поврежденных нейроцитов обладали повышенной
электронной плотностью из-за сходой, конденсации органелл. В
плазматической и других мембранах погибающих гиперхромных клеток
отмечались многочисленные перфорации. Авторы наблюдали объединение тел
поврежденных нейроцитных друг с другом и стенкой капилляров,
формирование простых и плотных контактов между телами нейроцитов,
повреждение периваскулярных мембран и прямые нейровазальные контакты.

51
Изменения астроцитов в виде набухания, фрагментации отростков и
дезинтеграции наблюдаются с первых минут ишемии, предшествуя
нейроцитальным изменениям (Garcia J.H. et al. 1993), и сопровождаются
снижением экспрессии астроцитарного маркера – кислого глиального
фибриллярного белка (GFAP). Однако через 4–6 ч после развития ишемии
отмечается активация астроцитов, окружающих ишемическую зону, которые
начинают усиленно синтезировать GFAP (Garcia J.H. et al., 1993). В более
поздние сроки (через 24 ч) вокруг области инфаркта образуется сеть GFAP-
позитивных астроцитов. Астроцитарная реакция становится все более
агрессивной и ведет к формированию глиального рубца в конце 1-й – начале
2-й недели после развития инсульта (Clark R.K. et al., 1993).
В связи с особенностью положения астроцитов относительно
относительно нейронов и стенки капилляров в головном мозге, они
закономерно вовлекаются в процесс ишемического повреждения. Последний
иногда даже рассматривают на уровне условно выделяемой нейроно-глио-
васкулярной единицы (Моргун А.В., Ковачева Н.В. Таранушенко Т.Е. и соавт.,
2013). Между элементами такой единицы в условиях ишемии были выявлены
щелевые соединения, прямые контакты (tight junction) и мембранные
перфорации (Гелеранская, О.А. и соавт., 2016; Frantseva M.V., Kokarovtseva L.
and Perez Velazquez J.L., 2003). Образование дефектов тонкой двухслойной
базальной мембраны эндотелиоцитов капилляров, перфораций мембран между
эндотелиальными клетками и астроцитами, везикул в отростках и телах
астроцитов, прилегающих к эндотелиальным клеткам сосуда, как было
выявлено, способствует повреждению структур гемато-энцефалического барьера, развитию перикапиллярного отека и отека-набухания
ультраструктурных компонетов астроцитов (F. Yang, Z. Wang, X. Wei et al.,
2014). Авторы полагают, что в поврежжение мембран вовлечены рецепторы
адгезии клеток NLRP3, образующих внутри клеток комплекс с активной
каспазой-1 (инфламосома). Последний, в свою очередь, стимулирует секрецию
интерлейкинов и инициирует апоптоз (Eisenbarth S.C. and Flavell R.A., 2012).

52
Микроглиальные клетки в патологических условиях (в том числе
ишемии) включать отростки, увеличиваются в размерах, производят большое
количество лизосом и принимают форму амебоидных, которая соответствует
их выраженной функциональной активации вплоть до полной готовности к
фагоцитозу (Giulian D., 1997). Активированные клетки микроглии способны к
длительному поддержанию структурно-функциональной активности, так что
они синтезируют большое количество соединений, которые являются
токсичными для клеток нервной ткани и клеток сосудистой стенки, тем самым
поддерживая воспалительный ответ (McGeer E. G. and McGeer P. L., 1994). Это
свойство активированных микроглиальных клеток, по мнению авторов, ведет
к задержке потери нейроцитов, микроциркуляторных расстройств, и
изменений проницаемости гематоэнцефалического барьера
В конце первого дня после начала инсульта активированная микроглия
широко представлена во всей области ишемии, особенно в области полутени
(Giulian D. et al., 1993; Giulian D., 1997; Schilling M. et al., 2003). Позже зона ее
локализации продолжает расширяться, и к 5-му дню активированные
микроглиальные клетки обнаруживаются в областях мозга далеко от ишемии,
что, по-видимому, является следствием процессов антеро- и ретроградной
нейроцитной дегенерации (Боголепов И.И. и Бурд Г.С., 1981). Парадокс
транснейроцитальной дегенерации подтверждает открытие авторами на
расстоянии от центра ишемии комплекса гипоксических метаболических
нарушений в нейроцитах в виде изменений в тинкториальных свойств
цитоплазмы повреждения митохондрий, изменений ДНК и РНК ядра и
цитоплазмы, редукции эндоплазматической сети.
Спустя 6-8 ч после инсульта реализуются реактивные изменения
нейтрофилов и гранулоцитов в микроциркуляторном русле, которые вызваны
активацией микроглии и увеличением синтеза провоспалительных факторов,
молекул клеточной адгезии. Наблюдается адгезия нейтрофилов и эндотелия
мелких сосудов, проникновение первых через гематоэнцефалический барьер и
инфильтрация ими ишемизированной ткани мозга (Zhang R.L. et al., 1994;

53
Garcia J.H. et al., 1996). Динамика реакции нейтрофилов во времени варьирует
в зависимости от характеристик ишемического процесса.
Активированая микроглия обладает высокой фагоцитарной
активностью. Она наряду с астроцитами, выполняет представляя функцию
антигенапрезентирования и готова к выведению на свою поверхность главного
комплекса гистосовместимости II класса, который, как Общеизвестно,
необходим для активации иммунокомпетентных клеток лимфоидного ряда
(Giulian D. et al., 1993; Giulian D., 1997; Schilling M. et al., 2003).
Наряду с фагоцитозом, выявлена способность активированных клеток
микроглии в сформированном мозге синтезировать различные цитокины
(Righi M. et al., 1991), блокирующие апоптоз и стимулирующие процессы
регенерации (Stella M.C. et al., 2001).
Цитокины и растворимые факторы микроглии, астроцитов,
нейтрофильных гранулоцитов и лимфоцитов (последние проникают в ткани
ЦНС во время воспаления) играют значительную роль в реакциях центральной
нервной системы. Активированная микроглия оказывает стимулирующее
действие на восстановление поврежденых нейроцитов и усиливает
миелинизацию нервных волокон (Constantinescu C.S. et al., 2000),
производство оксида азота (Betz-Corradin S., 1993), что, в конечном счете
модулирует морфо-функциональное состояние нейроцитов.
Строение эндотелиоцитов микроциркуляторного русла изменяется с
первых минут острой фокальной ишемии мозга. Через 30–40 мин в очаге ост-
острой ишемии наблюдается набухание эндотелиоцитов сосудистой стенки,
через 1 час обнаруживается повышенная проницаемость мембран; через 6
часов в ядерной зоне инфаркта появляются признаки некроза отдельных
клеток, ко 2–3-му дню – пролиферация как эндотелиальных, так и
гладкомышечных клеток сосудистой стенки (Iadecola C., 1999). В то же время,
ишемия головного мозга активизирует процесс ангиогенеза и
неоваскуляризации на 5-7-й день после экспериментального инсульта в районе
периинфарктной зоны, начинается процесс образования новых капилляров

54
(Clark R.K. et al., 1993), которые потенциально вызвают индукцию
ангиогенных факторов роста, молекулами адгезия клетоки,- интегринами.
Работы J. Krupinski с соавт. (1994) показали, что неоваскуляризация является
благоприятным признаком, который может прогнозировать улучшение
процессов восстановления в ткани головного мозга и выживания нейроцитов в
зоне пенумбры.
Известно, что при ишемии клетками мозга увеличивается выработка
фактора роста эндотелия сосудов (VEGF), известого как ключевой медиатор
ангиогенеза (неоваскуляризации). Есть также свидетельства широкого
значения VEGF и цитокинов из его семейства для нейроногенеза (в частности,
синаптогенеза) и нейропротекции при ишемии (Rudko A.S., 2017), благодаря
чему скоординированное взаимодействие ангиогенеза и нейрогенеза
способствует функциональному восстановлению нейронов.
Морфологические события, свидетальствующие о степени повреждения
гематоэнцефалического барьера на уровне элементов нейроно-глио-
васкулярной единицы и механизмы их образования были изложены выше.
1.6. Свойства антигипоксантов. Фармакологическая характеристика метапрота
Общеизвестно, что все виды неврологических расстройств развиваются
в результате гипоксии. Этот унифицированный и универсальный
этиопатогенетический механизм обусловлен тем, что потребность нейроцитов в энергоснабжении во много раз выше, чем для всех других клеток в
организме. Среднее потребление O2 нервной клеткой в состоянии покоя составляет 450 мкл в минуту, тогда как глиальные клетки или гепатоциты
используют до 60 мкл O2 в минуту каждый. Нейроциты, занимающие 5%
объема мозга утилизируют 25% всего церебрального O2 (Морозова О.Г., 2003).
Обычно существуют два основных направления нейропротекции - первичные
и вторичные. Первая включает в себя прерывание быстрых механизмов гибели
некротических клеток в результате реакций глютамат-кальциевого каскада.

55
Она применяется в течение первых минут до первых трех суток ишемии /
гипоксии. Второй используется для предотвращения замедленных механизмов
гибели клеток в результате окислительного стресса, дисбаланса цитокинов,
локального воспаления, вызывающих некроз, апоптоз нейроцитов и
направлена на снижение тяжести «долгосрочных результатов ишемии» (Гусев
Е.И. и Скворцова В.И., 2001).
Основными направлениями нейропротекции считаются: (Шабанов П.Д.,
Зарубина И.В., Цыган В.Н., 2009, Лесиевская Е.Е., 2012) 1). Восстановление
баланса между процессами возбуждения и торможения нейроцитов,
синаптической трансмиссией между ними, с помощью возбуждающих и
тормозных медиаторов. 2) Восстановление и поддержание энергетического
обмена нервных клеток. 3) Активация и поддержание анаболических
процессов в нейроцитах и макроглиях.
Центральное место в нейропротекции занимают антигипоксанты -
препараты, которые предотвращают или устраняют нарушения, вызванные
дефицитом O2.
В направлении действий принято различать антигипоксическое
косвенного и прямого действия. Косвенное стимулирующее воздействие на
клетки головного мозга имеют множество лекарственных препаратов
различных терапевтических групп, например, снотворных, седативных,
наркотических препаратов, способствующих переживанию клеток в условиях
ишемии благодаря адаптации к стрессорным факторам и снижения уровня
метаболизма клетки. Прямые антигипоксаны, в отличие от непрямых,
оказывают непосредственное стимулирующее действие на метаболизм и
функции нервных и глиальных клеток
Методами восстановления и поддержания процессов энергетического
обмена антигипоксантами считаются: 1) восстановление и поддержание
активности энргосинтезирующих процессов; 2) нормализации функциониро-
вания электрон-транспортной системы митохондрий; 3) обеспечение
субстратами компенсаторных метаболических путей; 4) оптимизация

56
процессов перекисного окисления и защита клеточных мембран от
повреждения активными радикалами и метаболитами (Lesiovskaya е. е., 2012).
Метапрот (химическое название препарата – 2- этилтиобензимидазол) –
вторичный нейропротектор группы антиастенических. средств с выражен-
ными психоактивирующими свойствами. Он обладает ноотропной, антигипо-
ксической, антиоксидантной, иммуномодулирующей активностью, способнос-
тью усиливать регенерацию и репарацию (Зарубина И.В., Нурманбетова Ф.Н.
и Шабанов П.Д., 2005; Зарубина И.В. и Шабанов П.Д., 2006).
В 1990-е годы 2-этилтиобензимидазол выпускался в основном как
актопротектор и использовался главным образом в восстановительной,
спортивной и военной медицине. С осени 2009 года метапрот выпускается
отечественным производителем лекарственных препаратов ЗАО НПО
«Антивирал» (Санкт-Петербург).
Метапрот апробирован на кафедре фармакологии ВМедА им. С. М.
Кирова как новый оригинальный отечественный препарат при терапии
гипоксии, инфаркте мозга, энцефалопатии, черепно-мозговой травме, гепатите
А, хронических заболеваниях легких, ишемической болезни сердца, гестозе
беременных и даже при профессиональной тугоухости (Шабанов П. Д., 2009;
Шабанов П. Д., Зарубина И. В. и Цыган В. Н., 2009; Шабанов П. Д., 2010).
Некоторые внемозговые эффекты метапрота позиционируют препарат как
эффективный полиорганный антиоксидант и энергостимулятор.
В тканях плаценты беременных с гестозом на фоне приема метапрота
снижалось содержание продуктов перекисного окисления липидов и
увеличивалась активность антиоксидантных энзимов и содержание
восстановленного глутатиона. Наряду с антиоксидантными эффектами
метапрот нормализовал уровень лактата и пирувата в крови и плаценте
пациенток, что препятствовало развитию лактацидоза и способствовало
стабилизации углеводного обмена (Зарубина И. В. и Шабанов П. Д., 2004). В легких метапрот интенсифицировал регенерацию бронхиального эпителия у
больных различными формами хронического бронхита (Зарубина И. В. и

57
Шабанов П. Д., 2004). В результате этого же исследования были установлены
следующие морфологические эффекты данного препарата: увеличение
количества включений 3
Н-уридина РНК, синтезируемой в эпителиальных
клетках, исчезновение участков эрозивного слущивания реснитчатых клеток,
появление плотной сети канальцев шероховатой эндоплазматической сети в
цитоплазме бокаловидных клеток.
При апробации метапрота у больных, перенесших черепно-мозговую
травму, сделан вывод о высокой эффективности 2-этилтиобензимидазола,
выпускающегося под коммерческим названием метапрот, как в виде
монотерапии, так и в сочетании с антидепрессантом пиразидолом при
рассматриваемой патологии (Шабанов П. Д., 2009).
Черепно-мозговая травма в структуре смертности и инвалидизации
уступает инфаркту мозга вследствие цереброваскулярной болезни, однако
ишемия или гипоксия являются патогенетическим фактором, объединяющим
эти нозологические формы. Так, среди причин черепно-мозговой травмы в
России 50–78% занимает бытовой травматизм, 10–30% – транспортный, 12–
15% – производственный и 1–2% – спортивный (Евсеева М. А. и соавт., 2008;
Новиков В. Е., Понамарева Н. И. и Шабанов П. Д., 2008).
Благодаря работам школы проф. П. Д. Шабанова был уточнен механизм
действия метапрота (Зарубина И. В. и Шабанов П. Д., 2004; Новиков В. Е.,
Понамарева Н. И. и Шабанов П. Д., 2008; Шабанов П. Д., Зарубина И. В. и
Нурманбетова Ф. Н., 2005). Он заключается в активации синтеза РНК и ряда
важнейших белков, в том числе ферментных, имеющих отношение к
иммунной системе. Усиливается синтез энзимов глюконеогенеза, которые
обеспечивают утилизацию лактата (фактора утомляемости, ограничивающего
работоспособность) и синтез углеводов, представляющих собой источник
энергии при интенсивных нагрузках, что ведет к повышению физической
работоспособности. Под влиянием метапрота также усиливается синтез
митохондриальных энзимов и структурных белков митохондрий, что
обеспечивает увеличение энергопродукции и поддерживает высокой уровень

58
сопряженности окисления с фосфорилированием. Воздействие метапрота в
условиях дефицита O2 обеспечивает поддержание синтеза АТФ на высоком
уровне, что обеспечивает его выраженный антигипоксический и
противоишемический эффект. Метапрот усиливает синтез антиоксидантных
энзимов. В целом препарат повышает устойчивость организма к воздействию
экстремальных факторов – физической нагрузке, стрессу, гипоксии и гипер-
термии. Особенностью препарата является его выраженный антиастенический
эффект, так как его применение повышало работоспособность при физической
нагрузке, ускоряло восстановление энергетического обмена и неврологи-
ческих нарушений после экстремальных воздействий.
Коррекция метапротом реактивных изменений клеточных элементов
головного мозга после гипоксии/ишемии прежде не исследовалась. Результаты
многих исследований реактивности клеток мозга убеждают в том, что
коррегирующее действие метапрота на клетки мозга может быть выраженным.
К таким результатам можно отнести установление выраженных продуктивных
изменений клеток эпителиальной выстилки бронхов, обусловленное
воздействием метапрота (Шабанов П. Д., 2010), чувствительных пластических
и структурных изменений нейроцитов, нейроцито-глиальных взаимо-
отношений, о которых шла речь в предыдущем разделе.
1.7. Выбор отделов головного мозга для моделирования последствий выраженной и слабо выраженной глобальной ишемии
Общеизвестно, что фокальная ишемия мозга зачастую не вызывает
выраженных клинических проявлений за счет компенсации утраченных
функций другими отделами мозга. Моделирование ассимметричной
пролонгированной глобальной ишемии мозга в этом смысле имеет
преимущества, так как при использовании этой модели создаются условия для
выраженного ограничения кровотока большого участка мозга, подобные
условиям в периинфарктной зоне (внутренней части зоны пенумбры), и слабо
выраженного ограничения кровотока, вовлекающие в ишемический процесс

59
остальную часть мозга. Нарушения гемодинамики в последней близки тем,
которые развиваются в наружной части зоны пенумбра при инфаркте мозга.
Именно таким условиям отвечает модель с перевязкой обеих общих
сонных артерий, опыт использования которой широко распространен и
включает большой отрезок времени (Пошивалов В.П., 1978; Шабанов П.Д.,
Зарубина И.В. и Soultanov Vagif S., 2011; Ginsberg M.D. and Busto R., 1989; Li
Y. and Liu S., 2017). Ядро ишемии, образующееся при данном способе
моделирования, широкое и захватывает весь передний мозг. Кровоснабжение
среднего мозга нарушается в значительно меньшей степени, так как его
вентромедиальный ядерный комплекс получает кровь по многочисленным
срединным среднемозговым артериям (Paxinos G., 2004). Последние являются
ветвями позвоночной артерии, кровоток в которых и в коллатеральных
сосудах Виллизиева круга частично компенсирует прекращение поступления
крови в передний мозг по ветвям внутренней сонной артерии – передней и
средней мозговым артериям.
Таким образом, данная модель ведет к ассимметричному нарушению
кровотока в переднем и среднем мозге. Оно поддерживается за счет
позвоночных артерий, и крысы выживают в течение 2–4 недель, если их не
подвергать стрессу (Шабанов П.Д., Зарубина И.В. и Soultanov Vagif S., 2011).
Выраженные гемодинамические нарушения в структурах переднего мозга
(ПЦинК и ПЯ) при билатеральной окклюзии обеих общих сонных артерий
ожидаемо будут близки тем, которые развиваются вблизи ядра инфаркта.
Передние лобные септальные артерии, несущие кровь к ПЦинК и ПЯ при этом
способе моделирования ишемии не получают кровь из внутренней сонной
артерии, но могут получать небольшой объем крови из коллатеральных
сосудов большого артериального круга. Слабо выраженное ограничение
кровотока будет охватывать вентральную часть среднего мозга и его ПНигЯ,
что может соответствовать внешней части пенумбры инфаркта.
Некоторые проявления клеточных реакций в зоне пенубры острого
фокального инсульта изучены и обобщены (Гусев Е.И. и Скворцова В.И.,

60
2001). Однако систематические описания изменений клеток различного типа в
зоне пенумбры неизвестны. Также неизвестно, каковы реактивные изменения
клеток мозга в зоне ненарушенного кровотока при моделировании фокальной
ишемии мозга.
Развитие выраженных нарушений клеток среднего мозга в данной
модели ишемии ожидаемо, так как каскадные метаболические реакции,
развивающиеся в фокусе выраженной ишемии, могут оказывать дистантное
действие через поврежденный здесь гематоэнцефалический барьер в системе
«капилляр–астроцит–нейроцит–астроцит–капилляр». Помимо немедленных
реакций глютамат-кальциевого каскада, «волн депрессии», в ПЦинК И ПЯ
предполагаются отсроченные реакции клеток активированной микроглии и
эндотелиоцитов в ответ на образование множества токсичных соединений.
Имеется в виду выработка этими клетками лейкотриенов, тромбоксана,
простациклина, простагландина Е2, вазоконстрикторов, которые индуцируют
дальнейшие цепи реакций (Ernst E., Matrai A.Т. and Paulsen F., 1987).
Увеличение уровня супероксид-радикала, в свою очередь, ведет к активации
перекисного окисления липидов, вторичному повреждению эндотелия с
нарушением его проницаемости. Накопление данных разнообразных
токсических веществ в клетках и нейропиле на больших территориях мозга
могут обусловить дистантный процесс реактивных изменений клеток в
системе «эндотелиоцит–астроцит–нейрон–микроглиоцит–нейрон», оказав
токсическое действие на клетки среднего мозга.
Данная модель ишемии переднего мозга является удобной для оценки
действия фармакологических средств, обладающих нейропротекторными
свойствами (Зарубина И.В. И Шабанов П.Д., 2004; Шабанов П.Д., Зарубина
И.В. и Soultanov Vagif S., 2011). Обычно она используется для установления
функциональных и биохимических постишемических нарушений (Пошивалов
В.П., 1978; Marysheva V.V., Mikheev V.V. and Shabanov P.D., 2013), важных
для оценки эффективности фармакологической коррекции данных нарушений.
Сведения о структурных, пространственных и количественных изменениях

61
клеток различного типа в мозге при таком способе моделирования ишемии в
литературе отсутствуют.
Такие отделы мозга, как глубокие слои ПЦинК, вентромедиальная часть
ПЯ переднего мозга и ПНигЯ среднего мозга были выбраны для настоящего
исследования в связи с тем, что в них прежде уже были установлены многие
чувствительные структурно-пластические параметры клеток после
воздействия на них этанола и морфина (Дробленков А.В., Карелина Н.Р. и
Шабанов П.Д., 2009; Дробленков А.В., 2010; Дробленков А.В. и Шабанов
П.Д., 2014). Этими параметрами, в частности, являлись: площадь тел нейронов
и глиоцитов, долевое соотношение субпопуляций реактивно модифицирован-
ных нейроцитов, глиоцито-нейроцитальный индекс и другие.
Нейроциты дофаминергических ядер и цингулярной коры крупнее
клеток многих остальных центров мозга и расположены компактно, что делает
их удобными для морфологического анализа.
Реактивные изменения клеток в ПНигЯ среднего мозга при данных
условиях моделирования ишемии могут выражать почти исключительно лишь
признаки воздействия токсических молекул каскада биохимических реакций,
передаваемых из переднего мозга через нейроциты, астроциты или меж-
клеточные пространства, так как нарушения гематоэнцефалического барьера в
области мозга, где слабо нарушена гемоциркуляция, не регистрируются.
Изменения клеток ПЦинК и ПЯ переднего мозга при данном способе
ишемизации мозга могут выражать локальные признаки повреждения, так как
находятся в области, где нарушен гематоэнцефалический барьер.
В целом, морфологические исследования последствий глобальной или
фокальной ишемии далеко не приближаются к выявлению целого ряда
лабильных структурных, количественных и пространственных постишемичес-
ких изменений различных клеток мозга, представляющих собой совокупность
критериев, верифицирующих степень повреждения мозга и морфологическую
эффективность ее фармакологической коррекции. Поэтому все изложенное
выше послужило предпосылкой выполнения настоящего исследования.

62
Глава 2. МАТЕРИАЛ И МЕТОДЫ ИССЛЕДОВАНИЯ
Эксперименты проводились на самцах крыс Wistar весом 200-220 г в
возрасте 4 месяцев, полученных из питомника Рапполово РАМН
(Ленинградская область). Изучали три левосторонних формации головного
мозга, которые получали кровь из различных крупных артерий (Paxinos, G.,
2004) - глубокие слои (V-VI) прегенуальная часть передней цингулярной коры
переднего мозга (ПЦинК) вентромедиальная часть прилежащего ядра
переднего мозга (ПЯ) и переднее медиальное подразделение паранигрального
ядра среднего мозга (ПНигЯ). Первые две части переднего мозга получают
кровь из ветвей передней мозговой артерии: передние лобные и восходящие
септальные артерии (соответственно),Третья васкуляризируется из
многочисленных мелких срединных среднемозговых артерий, образующихся
на дистальном конце основной артерии (рис 1).
Рис. 1. Артериальный бассейн общей сонной и позвоночной артерий крыс при
инъекции туши (из: Paxinos G., 2004). Артерии: bas – основная, pcer – задняя мозговая, mmes – срединные среднемозговые, acer – передняя мозговая, aif – передняя лобная, аsa – восходящая септальная, olfa – обонятельная, rsa – передняя септальная, thp – таламоперфорирующие. Структуры: IP – межножковая ямка, MB – сосцевидное тело, ас – передняя спайка, Cg – передняя цингулярная кора, СС – мозолистое тело

63
2.1. Моделирование глобальной ассиметричной ишемии головного
мозга
При воспроизведении кратковременной эфирной анестезии у крыс, их
фиксировали на станке, препарировали обе общие сонные артерии которые
затем плотно лигировали. Рану обработали антисептиком и слоистым.
Отключение обеих общих сонных артерий связано с глубокой гипоксией
переднего мозга. Расположенные в нем ПЦинК и ПЯ подвергаются
выраженной ишемии, так как они могут восполнить дефицит крови только
через коллатеральные сосуды большого артериального круга (Willisii) от
активных позвоночных и основной артерии. ПНигЯ, которые входят в ядера
вентральной области покрышки среднего мозга, испытывает
ишемию,выражается гораздо меньше, чем структура переднего мозга, потому
что он расположен в непосредственной близости от активной основной
артерии. В то же время небольшой диаметр просвета и большое количество
ветвей срединных средних мозговых артерий вносят вклад в формирование
значительно большей устойчивости к кровотоку, чем в широком просвете
основной артерии.
Это означает, что условия гемодинамики формируемые в модели
двусторонней окклюзии обеих общих сонных артерий Пошивалов В. П.
(1978), являются способом моделирования асимметричной глобальной
ишемии головного мозга. Области тяжелой ишемии содержит ПЦинК и ПЯ
переднего мозга, в области легкой ишемии (которая может соответствовать
внешней границе зоны полутени) содержит паранигральное ядро - среднего
мозга. Эти условия моделируются в настоящем эксперименте для выявления
реактивных изменений в разных видов клеток отделов МДОФС,
расположенных в полярных условиях ограничения кровотока.
2.2. Группы исследования
Головной мозг был исследован у четырех групп крыс. Первая группа (5
интактных крыс) представляла собой основной контроль (контроль 1). У крыс

64
экспериментальной группы (5 особей) воспроизводили глобальную
ассиметричную ишемию головного мозга. Контролем для животных с
ишемией служили ложнооперированные животные, у которых
воспроизводили все этапы операции без перевязки сонных артерий (5 особей,
контроль 2). Животным второй контрольной группы ежедневно внутрь
брюшины вводили 1,0 мл физиологического раствора. У крыс 4-й группы (5
животных) устанавливали эффективность медикаментозной коррекции
ишемии мозга препаратом метапрот, стимулирующим энергетический обмен и
биосинтетические процессы во многих органах (Шабанов П. Д. и соавт., 2011).
Препарат, приготовленный на основе 25%-го масляного раствора, в дозе 12,5
мг/кг и объеме 1,0 мл вводили внутрибрюшинно ежедневно 1 раз в сутки в
утренние часы в виде курса в течение 7 дней. Экспериментальных и
ложнооперированных животных умерщвляли через 7 суток опыта.
2.3. Морфологическое исследование головного мозга крыс
Через 3 мин после декапитации крыс, осуществлявшейся в соответствии
с "Правилами лабораторной практики в Российской Федерации" (приказ МЗ
РФ от 2003 г. № 267"), мозг извлекали из полости черепа и фиксировали в 9%
растворе нейтрального формалина, обезвоживали в этаноле возрастающей
концентрации и заливали в парафин. Фронтальные серийные срезы из отделов
мезокортиколимбической дофаминергической системы (МДофС) для
последующей окраски по Нисслю готовили толщиной 3 мкм, с целью
выявления глиального фибриллярного кислого протеина (GFAP) в телах,
главных отростках и перивазальных глиальных мембранах астроцитов – 7
мкм.
Формации МДофС, подлежащие исследованию, вначале идентифициро-
вали ориентировочно при микроскопии неокрашенных парафиновых срезов,
используя стереотаксический атлас головного мозга крыс (Paxinos G. and
Watson C., 1998). Прицельному исследованию подвергали глубокие слои

65
ПЦинК (слои V–VI, с которыми устанавливают синаптический контакт
аксоны нейроцитов ПНигЯ), расположенные в области средней части колена
мозолистого тела (рис. 2). Вентромедиальную часть ПЯ – наиболее широкую
(также являющуюся частью МДофС), определяли между передней спайкой
вблизи основания стриапаллидарного комплекса и обонятельными ядрами,
расположенными вблизи поверхности полушария (рис. 3). Переднюю
медиальную часть ПНИГЯ устанавливали впереди сосцевидных ядер между
черной субстанцией и ножкой мозга латерально, сводом снизу, медиальной
петлей сверху, надсосцевидным ядром и эпендимой 3 желудочка медиально
(рис. 4).
Срезы окрашивали крезиловым фиолетовым по методу Ниссля,
выявляли GFAP астроцитов (с использованием мышиных антител, клон GA-5,
Biocare medical, США, разведение 1:250) и Iba1-антиген мембран клеток
амебоидной формы глии (при помощи козьих поликлональных антител к Iba1,
AbCam, Великобритания, разведение 1:200). Вторичные биотинилированные
антитела применяли из набора VECTASTAIN ABC, США. После проявления
антигенов, связанных диаминобензидином, срезы докрашивали гематоксили-
ном Карацци.
Визуальную и морфометрическую оценку клеточных элементов
исследованных структур мозга осуществляли в трех последовательных
квадратах S 0,01 мм2 у каждого животного в группе (n=15). Для установления
морфометрических параметров клеток в ПЯ, содержавшем две нечетко
разграниченные части – ядерную и скорлуповую (Paxinos G. and Watson C.,
1998), был выбран наиболее широкий вентромедиальный подотдел. В
последнем плотность расположения клеток этих двух частей визуально
различалась в наименьшей степени. Клетки в вентромедиальном подотделе
ПЯ исследовали на площади 0,01 мм2 в семи последовательных квадратах.
В срезах, окрашенных методом Ниссля, выявляли тела нейронов,
астроцитов, олигодендроцитов, микроглиоцитов и эндотелиоциты. Нейроциты
подсчитывали после их идентификации согласно классификации Жаботинс-
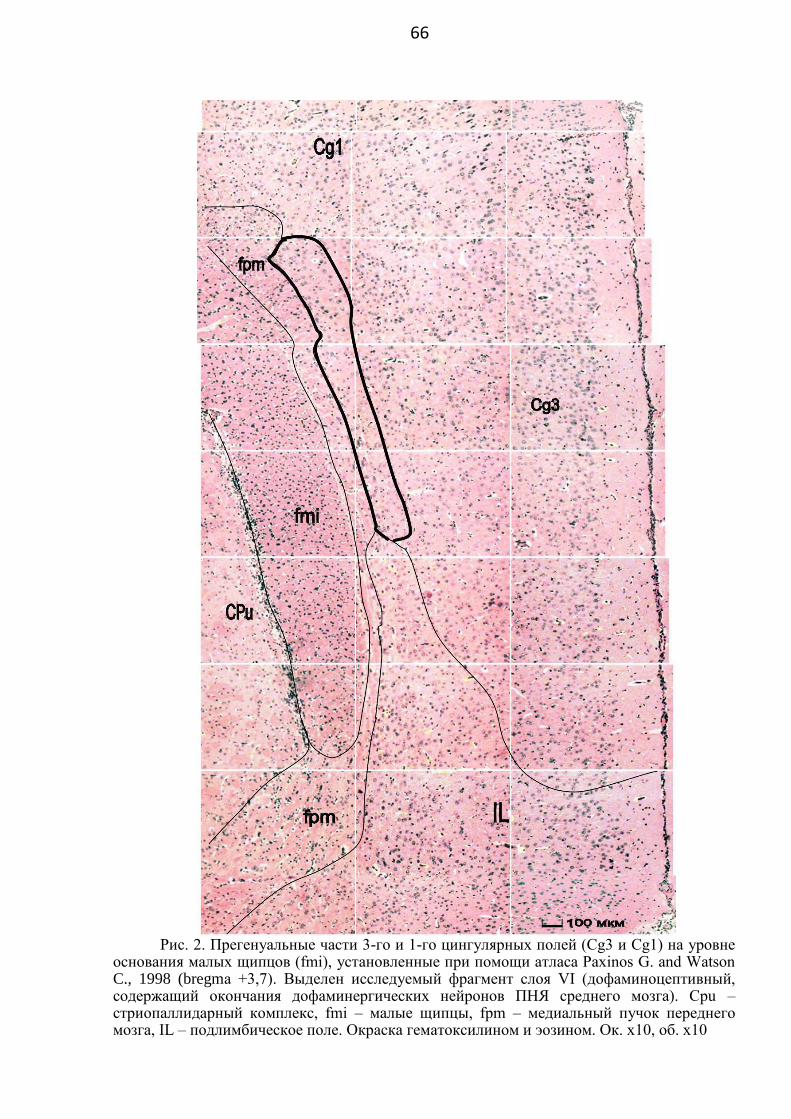
66
Рис. 2. Прегенуальные части 3-го и 1-го цингулярных полей (Cg3 и Cg1) на уровне основания малых щипцов (fmi), установленные при помощи атласа Paxinos G. and Watson C., 1998 (bregma +3,7). Выделен исследуемый фрагмент слоя VI (дофаминоцептивный, содержащий окончания дофаминергических нейронов ПНЯ среднего мозга). Cpu – стриопаллидарный комплекс, fmi – малые щипцы, fpm – медиальный пучок переднего мозга, IL – подлимбическое поле. Окраска гематоксилином и эозином. Ок. х10, об. х10

67
Рис. 3. Вентромедиальная часть прилежащего ядра на уровне перехода колена мозолистого тела в ствол, установленная при помощи атласа Paxinos G. and Watson C.,
1998 (bregma +0,2). Выделен исследуемый фрагмент ядра. IG – обонятельные ядра, Cpu – стриопаллидарный комплекс, Pir – грушевидная кора, SepN – ядра перегородки, VL – боковой желудочек, ca – передняя спайка мозга. Окраска гематоксилином и эозином. Ок. х10, об. х10

68
Рис. 4. Карта левосторонней передней медиальной части среднего мозга на уровне переднего полюса паранигрального ядра и компактной части черной субстанции (установлена согласно Paxinos G. and Watson C., 1998: bregma –4,8). Рамкой выделена
исследованная часть паранигрального ядра, равная трем квадратам площадью 0,01 мкм2
каждый. Обозначения: 3v – третий желудочек, APR – предкрасноядерное поле, f – свод, fr – fasciculus retroflexus, GD – зубчатая извилина, lm – медиальная петля, MNC – комплекс сосцевидных ядер, mp – сосцевидная ножка, mtg – сосцевидно-покрышечный путь, NIlm – интерстициальное ядро медиальной петли, NPN – паранигральное ядро, pc – ножка мозга, SNC – компактная часть черной субстанции, ZI – zona incerta. Окраска гематоксилином и эозином. Ок. х10, об. х10
кого Ю.М. (1965), широко используемой в современных нейроморфологи-ческих
исследованиях (Дробленков А.В. и соавт., 2009; Дробленков А.В., 2011;
Литвинцев Б.С. и соавт., 2011; Парамонова Н.М. и соавт., 2016), как
неизмененные (маломодифицированные), гипохромные, в том числе набухшие,
сморщенные гиперхромные (пикноморфные) и теневидные. Определяли
абсолютное количество и долю данных форм, долю нейроцитов, объединен-ных в
пары, площадь тел жизнеспособных нейроцитов (маломодифициро-ванных и
гипохромных), расстояние между телами нейроцитов данных разновидностей и
стенкой капилляра, а также между телами астроцитов и

69
стенкой капилляра в пределах окружности радиусом 20 мкм. Астроциты при
окрашивании по Нисслю идентифицировали по светлым ядрам овальной и
округлой формы, диаметр которых находился в пределах 5–8 мкм, и имевшим
на срезе отчетливо различимое ядрышко. Ядро астроцитов было окружено
светлым ободком цитоплазмы толщиной 1–4 мкм. Определяли глиоцито-
нейроцитальный индекс (как отношение количества клеток-сателлитов:
олигодендроцитов и астроцитов к количеству маломодифицированных и
гипохромных нейроцитов), количество эндотелиоцитов и площадь их ядер.
Олигодендроциты идентифицировали по гиперхромному округлому ядру,
диаметр которого составлял 3–5 мкм, снаружи которых часто был различим
узкий светлый ободок цитоплазмы толщиной не более 2 мкм. Эндотелиоциты
идентифицировали по самым небольшим гиперхромным ядрам вытянутой
формы, обращенным в просвет капилляра, часто содержащего эритроциты.
В срезах, окрашенных на GFAP, устанавливали количество астроцитов,
расстояние между их телами и стенкой капилляра в пределах окружности
радиусом 20 мкм (контроль результата, полученного аналогичным способом в
срезах, окрашенных по Нисслю) и площадь тел астроцитов. При окраске Iba1
устанавливали количество и площадь тел микроглиоцитов.
Морфометрию проводили с помощью программы Imagescope,
«Электронный анализ», Россия (рис. 5). Среднее арифметическое ( X S X ) ,
Рис. 5. Морфометрическое изучение астроцитов при помощи программы Imagescope («Электронный анализ», Россия). На экране монитора производили выделение и автоматическое установление площади тел астроцитов (а) и расстояния между их телом и стенкой капилляра (б). Выявление GFAP астроцитов и окрашивание ядер гематоксилином. Ок. х10, об. х100

70
Всего было исследовано 155 срезов МДофС (табл. 1). В этих срезах
Таблица 1
Схема эксперимента, количество исследованных срезов МДофС и подсчетов
Число Исследованные m на 1 m на 5
Количество подсчетов, n числа расстояния,
Группа крыс в формации крысу крыс клеток площади тел животных группе мозга каждого клеток, числа
типа пар нейроцитов
Переднее цин- 3 45
Интактные 5 гулярное поле,
(1, 1, 1) 15
(15х3) 10–53
слои V–VI
Прилежащее 3 105
ядро переднего 15 10–53 (1, 1, 1) (15х7) мозга
Параниграль- 3 45
ное ядро сред- 15 10–53 (1, 1, 1) (15х3) него мозга
После Переднее цин- 3 45
ложной 5 гулярное поле, (1, 1, 1)
15 (15х3)
10–53
слои V–VI
операции
Прилежащее 3
105
ядро переднего 15 10–53 (1, 1, 1) (15х7) мозга
Параниграль- 3 45
ное ядро сред- 15 10–53 (1, 1, 1) (15х3) него мозга
После Переднее цин- 3
15 45
5 гулярное поле, 10–53 (1, 1, 1) (15х3) ишемии слои V–VI
Прилежащее 3 105
ядро переднего 15 10–53 (1, 1, 1) (15х7) мозга
Параниграль- 3 45
ное ядро сред- 15 10–53 (1, 1, 1) (15х3) него мозга
После Переднее цин- 2 30
ишемии и 5 гулярное поле, (1, 1)
10 (10х3)
10–53
слои V–VI
терапии
Параниграль- 2
30
ное ядро сред- 10 10–53 (1, 1) (10х3) него мозга
Всего 20 — 31 155 645 110–583
Примечание: m – количество исследованных срезов; n – количество подсчетов в группе животных. В скобках через запятую приведено число срезов при окрашивании по Нисслю, выявлении GFAP астроцитов, Iba1 микроглиоцитов

69
после окрашивания методом Ниссля, выявления GFAP астроцитов Iba1
микроглиоцитов было произведено 645 подсчетов (в каждом из 645
квадратов площадь равна 0,01 мм2) числа тел фенотипических форм
нейроцитов, астроцитов, микроглиоцитов и эндотелиоцитов. Также было
выполнено 110–583 подсчета расстояния, 550–2915 подсчетов площади тел
маломодифицированных и гипохромных нейроцитов, астроцитов,
миокроглиоцитов, ядер эндотелиоцитов, 110–583 подсчета нейроцитов,
объединенных в пары.

70
Глава 3. РЕЗУЛЬТАТЫ СОБСТВЕННЫХ ИССЛЕДОВАНИЙ
3.1. Особенности строения и расположения разных типов клеток в некоторых отделах мезокортиколимбической дофаминергической
системы у здоровых крыс
При изучении обзорных срезов головного мозга крыс, выполненных во
фронтальной плоскости между основанием малых щипцов мозолистого тела
и медиальной поверхностью полушария, было идентифицировано
прегенуальное цингулярное поле (ПЦинП, Cg3 по атласу Paxinos, G., Watson,
C., 1998; см. рис. 2). Оно представляло собой наибольшую часть медиальной
прегенуальной коры, имевшую треугольную форму и простирающуюся от
небольшого подлимбического поля (расположенного под клювом
мозолистого тела) до широкого первого цингулярного поля Cg1
(занимающего верхнее-медиальную часть передней лобной коры). Основание
глубокого слоя (VI) в Cg3 располагалось на поверхности пояска (сingulum), в
составе передних щипцов мозолистого тела. Слой VI, содержащий
дофаминергические волокна медиального пучка переднего мозга,
заканчивающихся на нейронах слоев VI и V этого поля (Bjorklund A. and
Lindvall O., 1984), был представлен телами крупных пирамидных нейронов,
при окраске по Нисслю выглядевших овальными, плотность расположения
которых визуально была значительно больше, чем в V, III и II слоях.
Прилежащее ядро переднего мозга было исследовано во фронтальной
плоскости на уровне перехода колена мозолистого тела в его ствол и области
дна переднего рога бокового желудочка мозга – в его средней, наиболее
крупной части (см. рис. 3). Снизу и медиально контуры прилежащего ядра
охватывал медиальный пучок переднего мозга. Он был представлен
прерывистыми гнездами оксифильных волокнистых структур и цепочками
олигодендроцитов. В наиболее широкой медиальной части ядра нейроны
были расположены наиболее равномерно. Подсчет количества и изучение
строения клеточных элементов прилежащего ядра были осуществлены в

71
каждом из семи последовательных квадратов данной части ядра (ограничена
прямоугольником).
Передний медиальный подотдел паранигрального ядра среднего мозга
(ПНигЯ) во фронтальной плоскости занимал обширную территорию
вентральной части покрышки среднего мозга на уровне стыка заднего края
сосцевидных ядер гипоталамуса спереди и переднего края ядер и
проводниковых структур среднего мозга сзади (см. рис. 4). Немногочис-
ленные тела нейронов визуально обладали наибольшими размерами,
наибольшими компактностью и равномерностью расположения в средней
медиальной части этого подотдела ПНигЯ, которое и подлежало
исследованию.
3.1.1. Особенности строения слоев V–VI переднего цингулярного поля
В глубоких слоях ПЦинК тела нейронов неправильной, округлой и овальной
формы были распределены в слоях равномерно и располагались компактно,
причем их плотность расположения вблизи пояска (слой VI) визуально была
выше, чем во внутренней части слоя V (рис. 6). Многие тела нейронов слоя
VI, расположенные вблизи пояска, обладали вытянутой формой и
ориентировались относительно пояска параллельно. У части клеток было
различимо основание главного дендрита, направленного в направлении
поверхности коры.
Большинство нейронов содержало хорошо развитую цитоплазму, в
которой отмечалось преимущественно обильное скопление мелких
базофильных нисслевских глыбок. Последние в теле нейрона занимали почти
всю площадь цитоплазмы, обычно за исключением ее части, переходящей в
главный отросток. Крупное светлое ядро нейронов, обладавшее правильной
округлой или овальной формой, содержало преимущественно эухроматин и
крупное ядрышко, обычно занимавшее его центральную часть. Рельеф
поверхности ядра был четким на всем протяжении, контуры большей части
поверхности тела клетки – ровными и слегка выпуклыми.
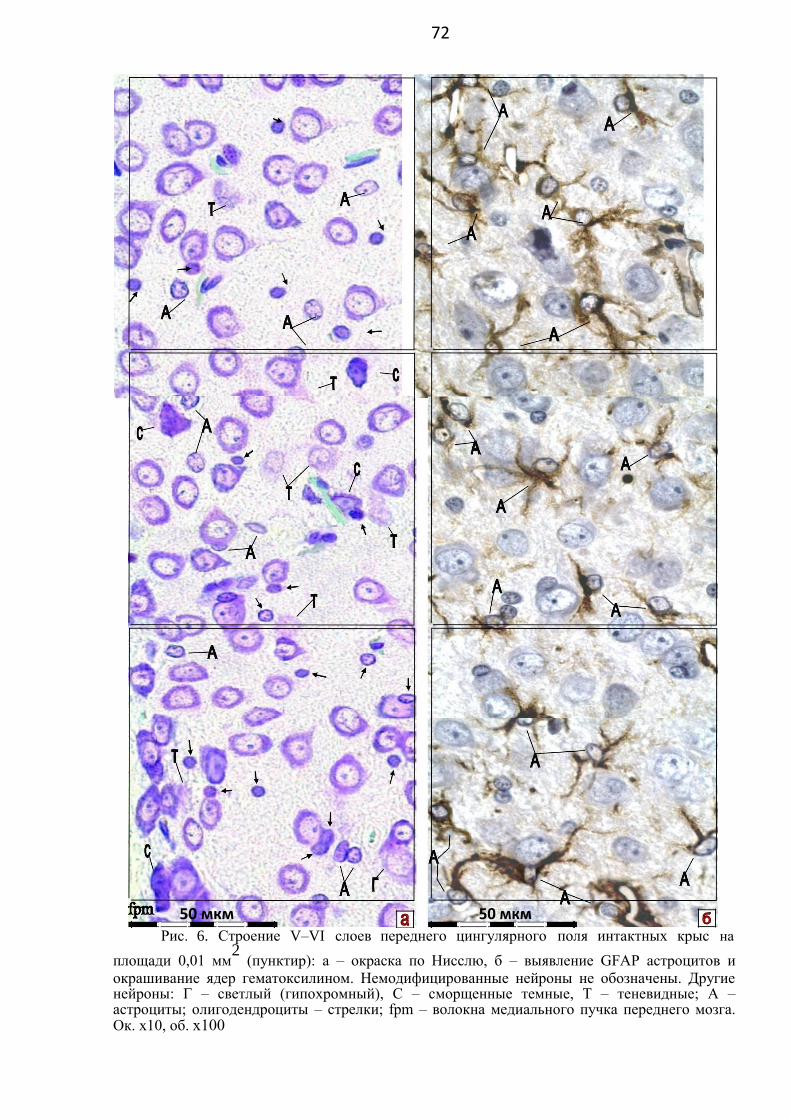
72
50 мкм 50 мкм
Рис. 6. Строение V–VI слоев переднего цингулярного поля интактных крыс на
площади 0,01 мм2
(пунктир): а – окраска по Нисслю, б – выявление GFAP астроцитов и
окрашивание ядер гематоксилином. Немодифицированные нейроны не обозначены. Другие нейроны: Г – светлый (гипохромный), С – сморщенные темные, Т – теневидные; А – астроциты; олигодендроциты – стрелки; fpm – волокна медиального пучка переднего мозга. Ок. х10, об. х100

73
Нейроны данного вида определялись как немодифицированные или
маломодифицированные (нормохромные). Их удельное число составило
83,6±2% от процента остальных видов нейронов ПЦинК (табл. 2). Площадь
тел нейронов этого типа составила 85,4±2,9 мкм2 (табл. 3). Некоторые тела
нейронов были объединены в пары, они примыкали друг к другу на
значительной части своей поверхности. Доля тел нейронов, объединенных в
пары, составила 9,3±1,4%. Относительно стенки капилляров немодифициро-
ванные клетки находились на разном расстоянии, параметр расстояния
между телом нейронов этого вида и стенкой капилляра составил 8,7±0,8 мкм.
Вблизи тел некоторых немодифицированных клеток располагались клетки-
сателлиты – светлые и темные формы олигодендроцитов. Реже клетками-
сателлитами являлись астроциты, тела которых были идентифицированы при
окраске по Нисслю. Глиоцито-нейрональный индекс в ПЦинК составил
0,13±0,03.
Единичные нейроны выглядели светлыми, они были лишены вещества
Ниссля, некоторые из них выглядели сморщенными, другие – набухшими.
Ядерный хроматин этих клеток, как и немодифицированных, был
конденсирован, преимущественно на периферии ядерной оболочки; ядрыш-
ко, более светлое и мелкое, чем у немодифицированных нейронов, было
обычно смещено ко внутренней поверхности кариотеки. Данные нейроны
обозначались как светлые (гипохромные) клетки. Доля светлых нейронов
была самой небольшой в исследованной части ПЦинК (см. табл. 2). Площадь
сечения тел клеток этого вида была в 1,2 раза меньше, чем этот параметр у
немодифицированных клеток (р<0,05; см. табл. 3), между тем среднее
расстояние между телами светлых нейронов и стенкой капилляра
значительно не отличалось от этого параметра у неизмененных клеток
(р<0,05). Среди гипохромных нейронов встречались клетки, объединенные в
пары с немодифицированными.

74
В ПЦинК также содержались единичные сморщенные тмные
(гиперхромные) нейроны (4,0±1,1%). Они обладали сморщенными контурами
поверхности ядра и цитоплазмы. Ядро содержало однородный гиперхромный
материал, у некоторых клеток этого типа было слабо различимо ядрышко.
Тела темных сморщенных клеток также могли быть объединены в пары с
немодифицмрованными нейронами. Некоторые клетки этого вида прилежали
к стенке капилляра и граничили с клеткой сателлитной формой макрогии.
Среди нейронов ПЦинК также были идентифицированы клетки-«тени».
Ониобладали стертыми контурами поверхности, были слабо окрашены,
сморщены, иногда имели вид слабо окрашенных базофильных пятен или
деформированных контуров кариотеки. Клетки-«тени» были малочисленны,
но в среднем они встречались в 2 раза чаще, чем светлые и сморщнные
темные клетки (8,4±1,0%). Клетки-«тени» могли образовывать пары с
немодифицированными клетками и располагались, визуально, на разном
расстоянии от стенки капилляров, однако с сателлитоцитами они не
граничили.
Астроциты глубоких слоев ПЦинК при окраске по Нисслю отличались
от остальных клеточных элементов строением ядра. Светлые ядра астроцитов
обладали округлой и овальной формой, диаметром 5–9 мкм, отчетливым
ядрышком, часто занимающим часто центральную часть ядра и содержали
преимущественно эухроматин. Вокруг ядра был различим светлый
однородный ободок цитоплазмы толщиной 1–4 мкм. Контур наружной
поверхности у некоторых клеток образовывал выступы – основания главных
отростков, придававшие клетке звездчатую форму. Астроциты, как и
нейроны, были распределены относительно равномерно и на разном
расстоянии от стенки кровеносных капилляров. Некоторые астроциты
представляли собой сателлитную форму глии.
GFAP в телах астроцитов имел вид рыхлой тонкопетлистой сети, во
многих клетках оплетавшей ядро со всех сторон, и часто образовывал
сгущение в виде глыбки вытянутой формы (см. рис. 6б). С материалом

75
глыбки часто была связана рыхлая сеть промежуточного филамента,
отростки. Главные отростки некоторых астроцитов, оказавшиеся в плоскости
среза, были ориентированы в направлении стенки капилляров и тел
нейронов. GFAP в концевой части отростков формировали тонкие
непрерывные периваскулярные глиальные мембраны.
Количество астроцитов в ПЦинК, а также расстояние между их телами
и стенкой ближайших кровеносных капилляров, установленные при окраске
по Нисслю, значительно не отличались от данных параметров, выявленных
при окрашивании GFAP (табл. 4). Средняя площадь тел астроцитов составила
41,5±3,3 мкм2
.
Глиальные макрофаги – покоящиеся микроглиоциты – были
идентифицированы как при окраске по Нисслю, так и их Iba-антигену.
Клетки обладали вытянутой формой гиперхромного ядра и гиперхромной
иммунопозитивной цитоплазмой (рис. 7). Отростки почти не
экспрессировали Iba-антиген. Микроглиоциты чаще располоагались вдоль
стенки кровеносных капилляров, однако встречался вариант их
перинейронального расположения (см. рис. 1б). Количество этих клеток было
самым малым, а площадь их тел – более чем в 1,6 раза меньшей, чем площадь
тел астроцитов (табл. 5).
Рис. 7. Количество, строение и расположение микроглиоцитов в слоях V–VI передней цингулярной коры у интактных крыс. Пунктир – площадь исследования,
равная 0,01 мм2
. Вставки – увеличенные фрагменты среза; fpm – поясок, содержащий волокна медиального пучка переднего мозга. Выявление Iba микроглиоцитов и окрашивание ядер гематоксилином. Ок. х10, об. х63

76
Стенка кровеносных капилляров в ПЦинК была идентифицирована при
окраске по Нисслю как граница слабо базофильного нейропиля с полым
просветом сосуда или как тонкая базофильная полоса на границе с
просветом. Стенка микрососуда выглядела значительно рельефнее при
окраске GFAP периваскулярных глиальных мембран. Эндотелиоциты
капилляров были идентифицированы по мелким гиперхромным ядрам
вытянутой формы, выступающим в просвет сосуда. Их количество было
меньше, чем клеток макроглии, но больше числа покоящихся макрофагов
(см. табл. 5). Площадь ядер эндотелицоитов составила всего 11,0±1,1 мкм2
.
3.1.2. Особенности строения прилежащего ядра переднего мозга
Тела нейронов исследуемой медиальной части прилежащего ядра (ПЯ)
были представлены фракцией компактно расположенных небольших клеток
округлой и неправильной формы. Большинство их, содержавших ободок
плотно расположенных мелких базофильных глыбок в перикарионе,
имевшие ровные, слегка выпуклые контуры клеточной поверхности, светлые
ядра округлой формы с одним или двумя мелкими ядрышками, были
расценены как немодифицированные клетки (рис. 8). Светлые, темные
сморщенные и теневидные нейроны характеризовались структурными
особенностями, аналогичными особенностям этих типов клеток в ПЦинК.
Доля немодифицированнх нейронов в ПЯ была значительно больше, а доля
клеток-«теней» – значительно меньше, чем в ПЦинК (см. табл. 2). Доли
светлых и темных сморщенных нейронов в ПЯ и ПЦинК значительно не
различались (р>0,05).
Все типы нейронов располагались относительно равномерно. Так же,
как и в исследованных слоях ПЦинК, они были удалены от стенки
кровеносных капилляров и эндотелиоцитов на разное расстояние. Среднее
расстояние немодифицированных и светлых нейронов от стенки капилляра в
ПЯ значительно не различалось с этим параметром в ПЦинК (р>0,05).
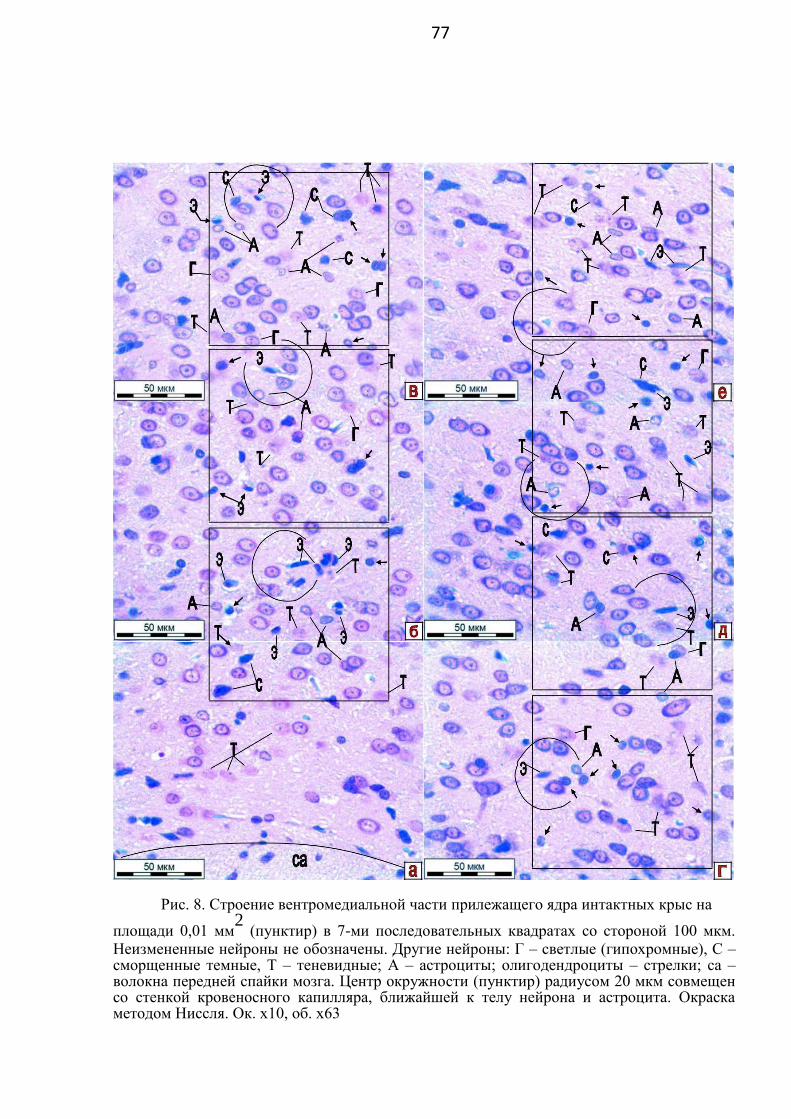
77
Рис. 8. Строение вентромедиальной части прилежащего ядра интактных крыс на
площади 0,01 мм2
(пунктир) в 7-ми последовательных квадратах со стороной 100 мкм.
Неизмененные нейроны не обозначены. Другие нейроны: Г – светлые (гипохромные), С – сморщенные темные, Т – теневидные; А – астроциты; олигодендроциты – стрелки; са – волокна передней спайки мозга. Центр окружности (пунктир) радиусом 20 мкм совмещен со стенкой кровеносного капилляра, ближайшей к телу нейрона и астроцита. Окраска методом Ниссля. Ок. х10, об. х63

78
Тела некоторых нейронов ПЯ были объединены в пары. В состав этих
пар входили, наряду с неизмененными, все описанные типы нейронов, в том
числе клетки-«тени». Тела некоторых нейронов были окружены клетками-
сателлитами. Доля нейронов, объединенных в пары, и глиоцито-
нейрональный индекс в ПЯ значительно не различались с данными
параметрами в ПЦинК (см. табл. 3; р>0,05).
Астроциты исследованной части прилежащего ядра при окраске
методом Ниссля так же отличались по строению от остальных клеточных
элементов, как и в ПЦинК. Наружный контур ядерной поверхности окружал
различимый у некоторых клеток светлый ободок цитоплазмы. Тела
астроцитов располагались относительно равномерно и на разном расстоянии
от ближайших кровеносных капилляров. Некоторые астроциты представляли
собой клетки-сателлиты.
При окраске GFAP астроциты имели те же особенности строения, что и
в глубоких слоях ПЦинК (рис. 9). Отростки некоторых астроцитов,
оказавшиеся в части своей длины срезанными продольно, были направлены в
сторону ближайших к ним тел нейронов или кровеносных капилляров и
образовывали снаружи эндотелиоцитов астроцитарные периваскулярные
мембраны.
Количество астроцитов, их расстояние от стенки ближайших
кровеносных капилляров, установленные при окраске по Нисслю и их GFAP
в ПЯ, площадь тел астроцитов значительно не различались с
соответствую-щими параметрами в глубоких слоях ПЦинК (см. табл. 4).
Микроглиоциты ПЯ, идентифицируемые при окрашивании Iba-1
антигена, располагались преимущественно вблизи стенки капилляров. Они
обладали преимущественно вытянутой формой тела и небольшими
размерами (рис. 10). Периферические части их отростков иммунный маркер
не экспрессировали. Число и площадь тел покоящихся микроглиоцитов, а
также количество и площадь ядер эндотелицоитов в ПЯ также значительно
не различались с соответствующими параметрами в исследованной части
ПЦинК (см. табл. 5).
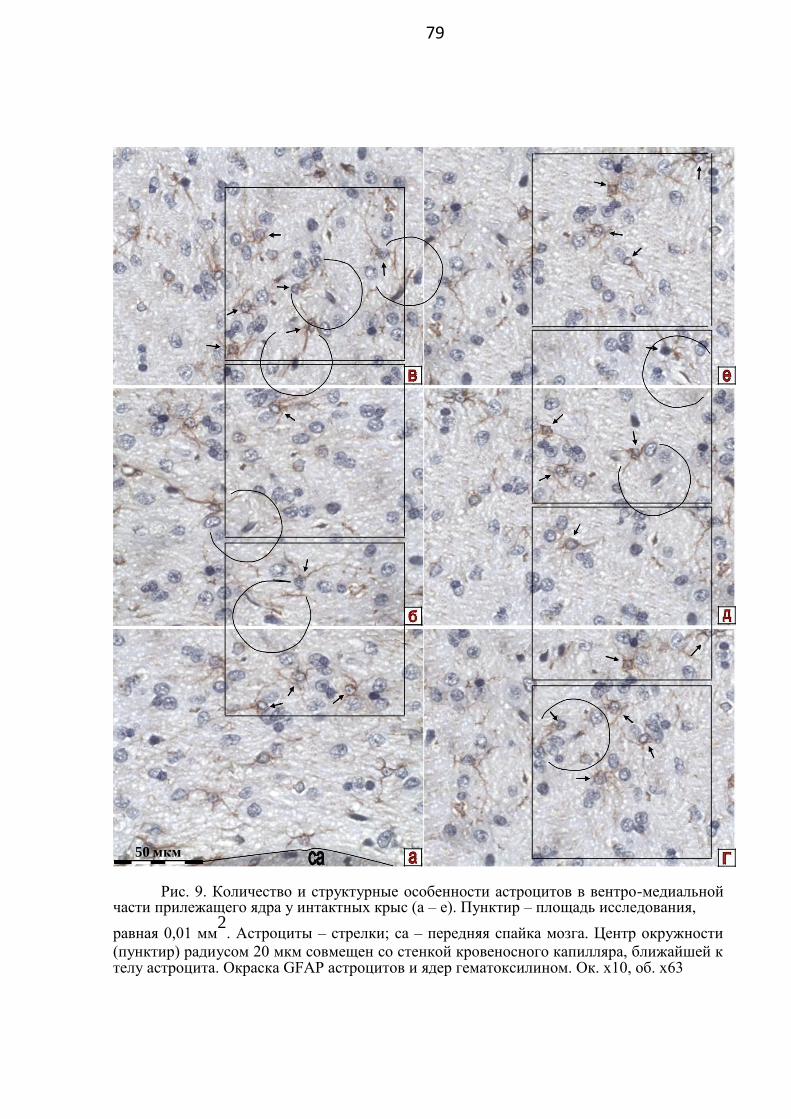
79
50 мкм
Рис. 9. Количество и структурные особенности астроцитов в вентро-медиальной части прилежащего ядра у интактных крыс (а – е). Пунктир – площадь исследования,
равная 0,01 мм2
. Астроциты – стрелки; са – передняя спайка мозга. Центр окружности
(пунктир) радиусом 20 мкм совмещен со стенкой кровеносного капилляра, ближайшей к телу астроцита. Окраска GFAP астроцитов и ядер гематоксилином. Ок. х10, об. х63

80
50 мкм
Рис. 10. Количество, расположение и структурные особенности микроглио-цитов в вентромедиальной части прилежащего ядра у интактных крыс (а – е). Пунктир – площадь
исследования, равная 0,01 мм2
. Вставки – увеличенные фрагменты среза; са – передняя спайка мозга. Окраска Iba микроглиоцитов и ядер гематоксилином. Ок. х10, об. х63

81
3.1.3. Особенности строения паранигрального ядра среднего мозга
Большинство нейронов в исследованной части паранигрального ядра
(ПНигЯ) характеризовалось крупными размерами тел, имевшими слегка
удлиненную полигональную форму, отчетливые, ровные, слегка выпуклые
контуры клеточной и ядерной поверхности. Нисслевская субстанция имела
вид плотно упакованных глыбок, занимала большую часть цитоплазмы, в
которой была распределена преимущественно вблизи ядра. Основание
отростков и небольшая территория вблизи клеточной мембраны были
просветлены (рис. 11). Эти нейроны были расценены как неизмененные. Эти
клетки составили 70,4±4,2% от общего количества. Площадь их тел 2
Значительно меньшую часть нейронов составили светлые, сморщенные
темные и теневидные клетки. Гипохромные нейроны имели вид
просветленных, лишенных хроматофильной субстанции, некоторые из них
выглядели уменьшенными в размерах, сморщенными, иные – набухшими,
имели участки стертости клеточной и ядерной поверхности. Некоторые
гипохромные нейроны, обладавшие слегка сморщенной формой цитоплазмы
и ядра, отличались от неизмененных клеток ослабленным окрашиванием
ядерного хроматина, ядрышка и цитоплазмы. Клетки данного типа возникли,
по-видимому, вследствие нарушения водно-солевого обмена или апоптоза;
они рассматривались как находящиеся на пути к гибели и превращению в
клетку-«тень». В целом площадь гипохромных нейронов оказалась немного
меньше, чем неизмененных (р<0,05).
Темные сморщенные нейроны были расценены как утрачивающие
влагу и склерозирующиеся. Клетки-«тени» обладали стертыми контурами
поверхности, были слабо окрашены, сморщены, иногда имели вид слабо
окрашенных базофильных комочков или деформированных контуров
ядерной оболочки. Более трети всех нейронов ПНигЯ были измененными,
главным образом, по типу гипохро мии (во-вилимому вследствие нарушения

82
50 мкм 50 мкм
Рис. 11. Строение передней медиальной части паранигрального ядра среднего
мозга у интактных крыс на площади 0,01 мм2
(пунктир): а – окраска методом Ниссля, б – выявление GFAP астроцитов и докрашивание ядер гематоксилином. Нейроны: Н – неизмененные, Г – светлый (гипохромный), С – сморщенный темный, Т – клетки-«тени». А – астроциты; олигодендроциты – стрелки. Ок. х10, об. х100

83
водно-солевого обмена) или гиперхромии-сморщивания (вероятно,
вследствие начала программированной гибели (см. табл. 2).
Рассматриваемые типы нейронов визуально располагались в
исследованных областях ядра относительно равномерно. Они были удалены
от стенки кровеносных капилляров и эндотелиоцитов на разное расстояние:
клетки могли располагаться вплотную к сосуду или находиться на
расстоянии от ближайшего сосуда, превышающем R=20 мкм. Среднее
расстояние немодифицированных и светлых нейронов от стенки капилляра в
паранигральном ядре составило, соответственно, 7,0±2,0 и 5,8±2,5 мкм.
Некоторые нейроны были окружены клетками-сателлитами, единичные
– объединены в пары. Доля пар нейронов, глиоцито-нейрональный индекс и
расстояние между телами нейронов и стенкой капилляра в ПНЯ значительно
не отличались от соответствующих параметров в ПЦинК и ПЯ.
Клетки астроцитарной глии при окраске методом Ниссля были
идентифицированы по строению ядер. Они выглядели светлыми, овальными,
реже округлыми, имевшими ровный выпуклый контур ядерной поверхности.
Их хроматин был представлен преимущественно рыхлой сетью тонких
нитей; другая часть хроматина была конденсирована и образовывала тонкую
линию вблизи внутренней поверхности ядра. Продольный диаметр ядер
астроцитов в срезах на уровне одного-двух мелких ядрышек составлял 6–9
мкм (см. рис. 1б). Наружный контур ядерной поверхности окружал
различимый у некоторых клеток светлый ободок цитоплазмы.
Астроциты в исследованной части паранигрального ядра, как и
нейроны, располагались относительно равномерно и на разном расстоянии от
ближайших кровеносных капилляров.
При окраске глиального GFAP вокруг ядер астроцитов был различим
плотный иммунопозитивный материал, охватывающий ядро полностью или,
частично, в виде полумесяца (см. рис. 11б). От тел астроцитов отходили 2–5
наиболее широких главных отростков, некоторые из них на периферии
ствола и на конце выглядели разволокненными (пушистыми). Отростки
располагались прямолинейно, реже имели слабо извилистый вид, многие из

84
них образовывали тонкие вторичные ветви, отходящие от основного ствола
под углом. Некоторые отростки астроцитов были направлены в сторону
ближайших к ним тел нейронов или кровеносных капилляров и образовывали
снаружи ядер эндотелиоцитов астроцитарные периваскулярные мембраны.
Значительная часть пространств между телами клеточных элементов
содержала тонкопетлистую сеть GFAP-позитивных филаментов.
Количество астроцитов, их расстояние до стенки ближайших
кровеносных капилляров, установленные при окраске как при окраске по
Нисслю, так и при выявлении их GFAP, значительно не различались с
соответствующими параметрами в исследованных структурах переднего
мозга (см. табл. 4). Площадь тел астроцитов, более чем в 2,5 раза меньшая
площади неизмененных нейронов ПНигЯ, значительно не различалась с
соответствующим параметром в ПЦинК и ПЯ. Тела некоторых астроцитов,
темных и светлых видов олигодендро-цитов являлись клетками-сателлитами.
Глиальные макрофаги – покоящиеся микроглиоциты были иденти-
фицированы как при окраске по Нисслю, так и их Iba-антигену. Клетки
обладали гиперхромным ядром вытянутой формы и гиперхромной
иммунопозитивной цитоплазмой (рис. 12). Отростки почти не
экспрессировали Iba-антиген. Микроглиоциты чаще располагались вдоль
стенки кровеносных капилляров. Количество этих клеток было самым
низким в ПНЯ, а площадь их тел – более чем в 1,5 раза меньшей, чем
площадь тел астроцитов (см. табл. 5). Морфометрические параметры
микроглиоцитов различались незначительно с таковыми в структурах
переднего мозга (р>0,05).
Стенка кровеносных капилляров в ПНигЯ была идентифицирована при
окраске по Нисслю как граница слабо базофильного нейропиля с полым
просветом сосуда или как тонкая базофильная полоса на границе с ним.
Стенка микрососуда выделялась значительно рельефнее при окраске GFAP
периваскулярных глиальных мембран. Эндотелиоциты капилляров были
идентифицированы по мелким гиперхромным ядрам вытянутой формы,
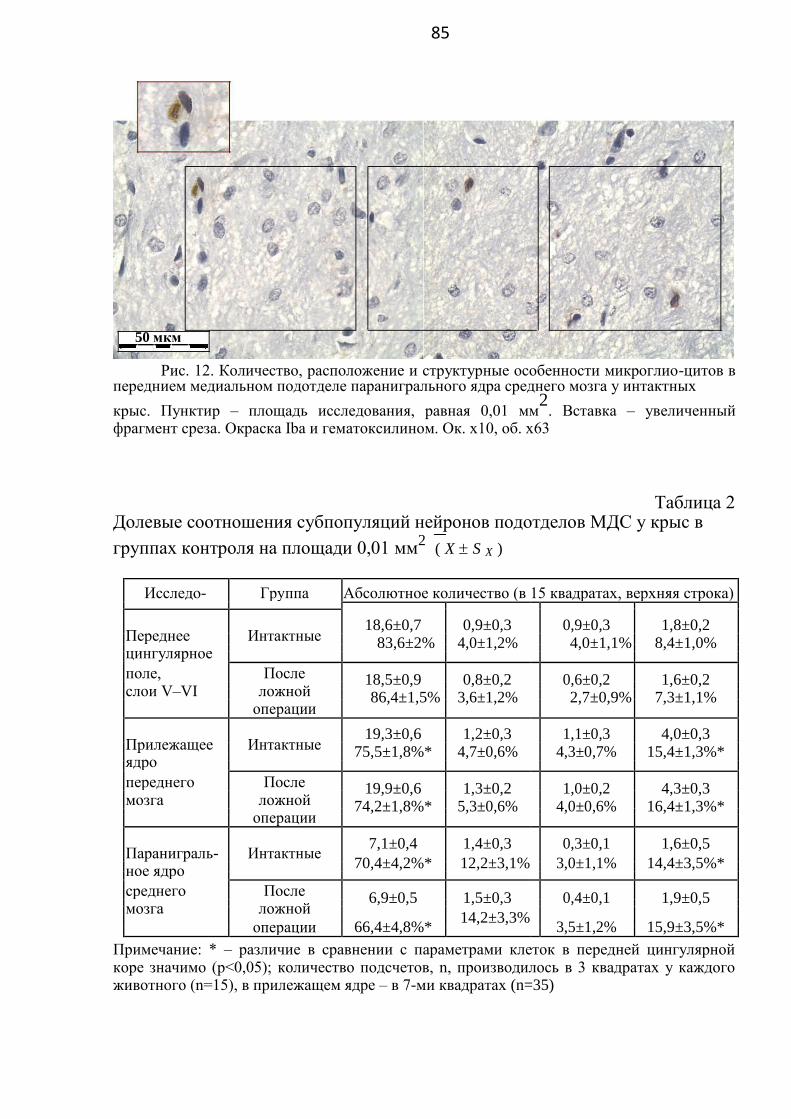
85
50 мкм
Рис. 12. Количество, расположение и структурные особенности микроглио-цитов в переднием медиальном подотделе паранигрального ядра среднего мозга у интактных
крыс. Пунктир – площадь исследования, равная 0,01 мм2
. Вставка – увеличенный фрагмент среза. Окраска Iba и гематоксилином. Ок. х10, об. х63
Таблица 2 Долевые соотношения субпопуляций нейронов подотделов МДС у крыс в
группах контроля на площади 0,01 мм2 ( X S X )
Исследо- Группа Абсолютное количество (в 15 квадратах, верхняя строка)
18,6±0,7 0,9±0,3
0,9±0,3 1,8±0,2 Переднее Интактные
83,6±2% 4,0±1,2% 4,0±1,1% 8,4±1,0%
цингулярное
поле, После 18,5±0,9 0,8±0,2 0,6±0,2 1,6±0,2 слои V–VI ложной
86,4±1,5% 3,6±1,2% 2,7±0,9% 7,3±1,1%
операции
Прилежащее Интактные 19,3±0,6 1,2±0,3 1,1±0,3 4,0±0,3
75,5±1,8%* 4,7±0,6% 4,3±0,7% 15,4±1,3%* ядро
переднего После 19,9±0,6 1,3±0,2 1,0±0,2 4,3±0,3 мозга ложной
74,2±1,8%* 5,3±0,6% 4,0±0,6% 16,4±1,3%*
операции
Параниграль- Интактные 7,1±0,4 1,4±0,3 0,3±0,1 1,6±0,5
70,4±4,2%* 12,2±3,1%
3,0±1,1% 14,4±3,5%* ное ядро
среднего После 6,9±0,5 1,5±0,3 0,4±0,1 1,9±0,5 мозга ложной
14,2±3,3%
операции 66,4±4,8%*
3,5±1,2% 15,9±3,5%*
Примечание: * – различие в сравнении с параметрами клеток в передней цингулярной коре значимо (р<0,05); количество подсчетов, n, производилось в 3 квадратах у каждого животного (n=15), в прилежащем ядре – в 7-ми квадратах (n=35)
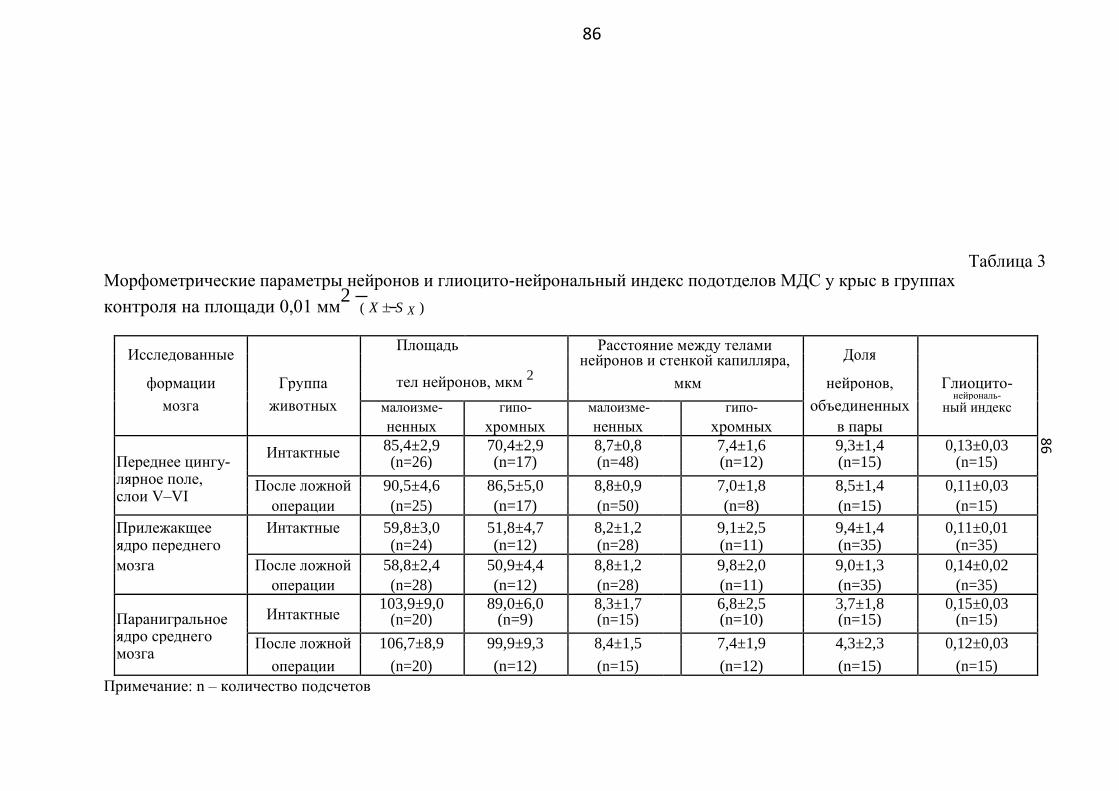
86
Таблица 3 Морфометрические параметры нейронов и глиоцито-нейрональный индекс подотделов МДС у крыс в группах
контроля на площади 0,01 мм2
( X S X )
Исследованные Площадь Расстояние между телами
Доля
нейронов и стенкой капилляра,
Глиоцито-
формации Группа тел нейронов, мкм 2
мкм нейронов,
мозга животных
объединенных нейрональ-
малоизме- гипо- малоизме- гипо- ный индекс
ненных хромных ненных хромных в пары 86
Переднее цингу- Интактные 85,4±2,9 70,4±2,9 8,7±0,8 7,4±1,6 9,3±1,4 0,13±0,03
(n=26) (n=17) (n=48) (n=12) (n=15) (n=15)
лярное поле,
После ложной 90,5±4,6 86,5±5,0 8,8±0,9 7,0±1,8 8,5±1,4 0,11±0,03
слои V–VI
операции (n=25) (n=17) (n=50)
(n=8) (n=15) (n=15)
Прилежакщее Интактные 59,8±3,0 51,8±4,7 8,2±1,2 9,1±2,5 9,4±1,4 0,11±0,01 ядро переднего (n=24) (n=12) (n=28) (n=11) (n=35) (n=35)
мозга После ложной 58,8±2,4 50,9±4,4 8,8±1,2 9,8±2,0 9,0±1,3 0,14±0,02
операции (n=28) (n=12) (n=28) (n=11) (n=35) (n=35)
Паранигральное Интактные 103,9±9,0 89,0±6,0 8,3±1,7 6,8±2,5 3,7±1,8 0,15±0,03
(n=20) (n=9) (n=15) (n=10) (n=15) (n=15)
ядро среднего
После ложной 106,7±8,9 99,9±9,3 8,4±1,5 7,4±1,9 4,3±2,3 0,12±0,03
мозга
операции (n=20) (n=12) (n=15)
(n=12) (n=15) (n=15)
Примечание: n – количество подсчетов

87
Таблица 4 Количество и морфометрические параметры астроцитов подотделов МДС у
крыс в группах контроля на площади 0,01 мм2 (
S
X
)
X
Исследо -
Количество при
Расстояние между
Группа телами астроцитов Площадь
ванные окрашивании и стенкой капилляров, тел,
формации животных
мкм, при окраске
мкм
мозга
методом
GFAP по Нисслю
GFAP
Ниссля
Переднее Интактные
4,6±0,6 4,7±0,5 9,4±1,9 9,7±1,7 41,5±3,3 (n=15) (n=15) (n=15) (n=15) (n=19)
цингуляр-
После
4,8±0,4
43,4±2,2
ное поле, 4,7±0,5 10,3±1,6 8,9±1,5
слои V–VI ложной
(n=15)
(n=15) (n=15)
(n=15) (n=20)
операции
Прилежащее Интактные 3,5±0,3 3,6±0,3 10,4±1,1 10,9±1,1 38,0±1,5
(n=35)
(n=35) (n=35)
(n=35) (n=35)
ядро
переднего После 3,4±0,3 3,3±0,3 10,8±1,2 11,0±1,2 38,0±1,5
мозга ложной (n=35)
(n=35) (n=35)
(n=35) (n=35)
операции
Пара- Интактные 3,7±0,4 3,7±0,4 9,7±1,5 10,4±1,6 38,7±3,2 нигральное (n=15) (n=15) (n=15) (n=15) (n=30)
ядро
После 3,7±0,3
3,9±0,5 9,8±1,5
10,4±1,3 39,3±2,9
среднего
ложной
мозга (n=15)
(n=15) (n=15)
(n=15) (n=29)
операции
Примечание: n – количество подсчетов
Таблица 5 Количество, морфометрические параметры микроглиоцитов и эндотелиоцитов капилляров подотделов МДС у крыс в группах контроля на
площади 0,01 мм2 (
S
X
)
X
Исследованные Группа
Микроглиоциты Эндотелиоциты
формации
площадь площадь животных
мозга
количество тел, мкм2
количество ядер, мкм2
Переднее цингу- Интактные 0,9±0,2 28,2±2,2 2,2±0,3 11,9±1,1 лярное поле,
После ложной
слои V–VI 0,9±0,3 28,6±2,3 2,3±0,4 10,5±0,7
операции
Прилежащее Интактные 1,0±0,1 28,7±1,3 2,5±0,3 11,5±0,5
переднего
После ложной 1,1±0,2 27,1±1,2 2,3±0,2 12,3±0,6
мозга
операции
Паранигральное Интактные 1,1±0,2 26,3±1,7 2,6±0,4 10,9±0,6 ядро
среднего После ложной 1,2±0,3 26,8±1,3 2,3±0,4 10,4±0,8 мозга операции
Примечание: количество подсчетов, n, производилось в 3-х квадратах у каждого животного (n=15), в прилежащем ядре – в 7-ми квадратах (n=35)

88
выступающим в просвет сосуда. Ядрышко этих клеток обычно не
выявлялось. Морфометрические параметры эндотелиоцитов в ПНигЯ
значительно не различались с таковыми в ПЦинК и ПЯ (см. табл. 5).
Морфометрические параметры разных типов клеток в исследованных
отделах дофаминергической системы мозга у ложно оперированных и
интактных животных значительно не различались (см. табл. 2–5).
Таким образом, левосторонние глубокие слои передней цингулярной
коры, вентромедиальная часть прилежащего ядра переднего мозга и
передний медиальный подотдел паранигрального ядра среднего мозга у
молодых взрослых крыс-самцов образованы скоплениями высокодифферен-
цированных клеток, прежде всего нейронов и протоплазматических
астроцитов, обладающих рядом существенных морфолого-топографических
параметров. Их изменения могут нести важную информацию для оценки
степени повреждения клеточных систем исследованных структур мозга,
вызванного рядом внешних факторов.
Среди нейронов, помимо преобладающих немодифицированных
клеток, встречаются субпопуляции единичных гипохромных, темных
сморщенных и немногочисленных клеток-«теней». Удельная плотность
последних наименьшая в глубоких слоях передней цингулярной коры и
наибольшая, соответственно, в дофаминергических ядрах. Наибольшая
компактность расположения тел нейронов и наименьший их размер из всех
исследованных дофаминергических центров был присущ вентромедиальной
части прилежащего ядра переднего мозга.
Нейроны разных фенотипических форм в исследованных отделах
дофаминергической системы способны образовывать пары, количество
которых невелико, они находятся на разном расстоянии от сосудистой стенки
и некоторые из них окружены клетками сателлитной формы глии.
Для установления числа астроцитов и площади их тела
информативными могут быть и срезы, окрашенные методом Ниссля. Число и
площадь тел астроцитов в исследованных дофаминергических центрах

89
значительно не различаются. Тела астроцитов в исследованных формациях
мозга находятся на разном расстоянии от стенки капилляров, величина
которого в этих структурах мозга значительно не различается. Концевые
части главных отростков астроцитов в исследованных формациях мозга
образуют тонкие непрерывные периваскулярные глиальные мембраны,
рельефно ограничивающие просвет кровеносных капилляров.
При окрашивании Iba-1 антигена в исследованных дофаминергических
центрах выявляются немногочисленные неактивные тела микроглиоцитов,
расположенные главным образом вблизи стенки капилляров.
Эндотелиальные клетки капилляров, обладавшие мелкими гиперхромными
ядрами, в которых ядрышко обычно не выявлялось, были
немногочисленными и численно в исследованных структурах мозга
значительно не различались.
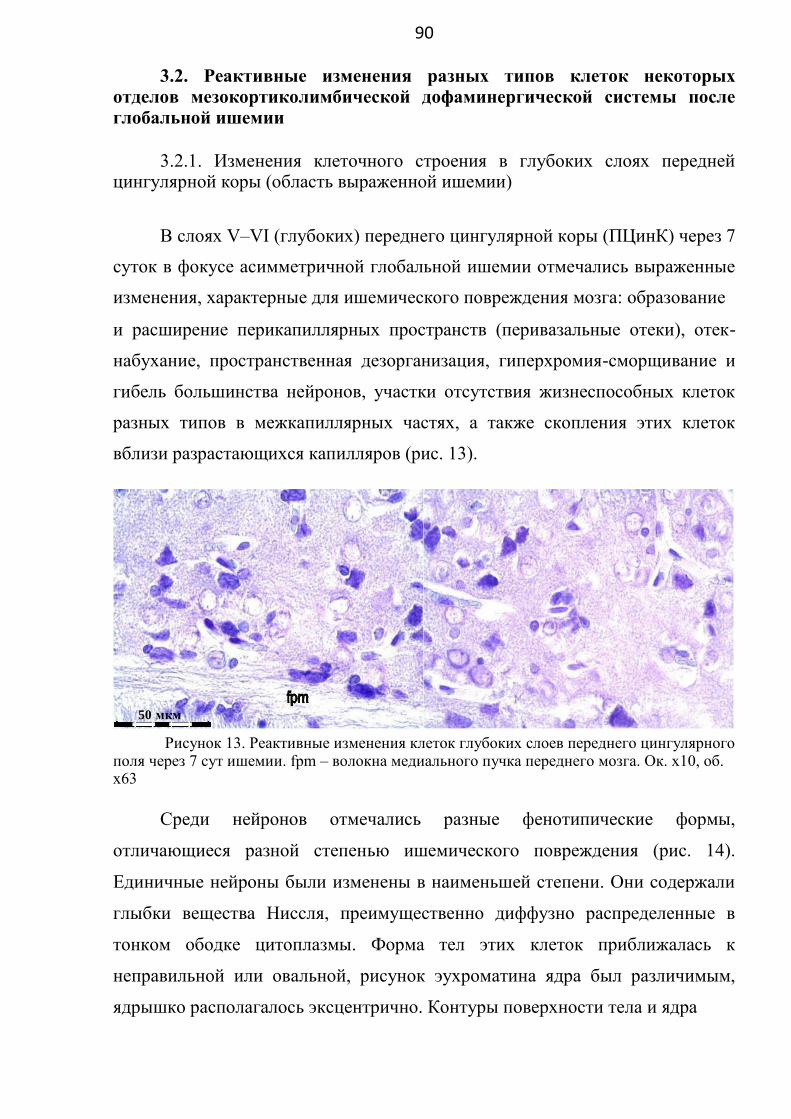
90
3.2. Реактивные изменения разных типов клеток некоторых
отделов мезокортиколимбической дофаминергической системы после
глобальной ишемии
3.2.1. Изменения клеточного строения в глубоких слоях передней цингулярной коры (область выраженной ишемии)
В слоях V–VI (глубоких) переднего цингулярной коры (ПЦинК) через 7
суток в фокусе асимметричной глобальной ишемии отмечались выраженные
изменения, характерные для ишемического повреждения мозга: образование
и расширение перикапиллярных пространств (перивазальные отеки), отек-
набухание, пространственная дезорганизация, гиперхромия-сморщивание и
гибель большинства нейронов, участки отсутствия жизнеспособных клеток
разных типов в межкапиллярных частях, а также скопления этих клеток
вблизи разрастающихся капилляров (рис. 13).
50 мкм
Рисунок 13. Реактивные изменения клеток глубоких слоев переднего цингулярного поля через 7 сут ишемии. fpm – волокна медиального пучка переднего мозга. Ок. х10, об. х63
Среди нейронов отмечались разные фенотипические формы,
отличающиеся разной степенью ишемического повреждения (рис. 14).
Единичные нейроны были изменены в наименьшей степени. Они содержали
глыбки вещества Ниссля, преимущественно диффузно распределенные в
тонком ободке цитоплазмы. Форма тел этих клеток приближалась к
неправильной или овальной, рисунок эухроматина ядра был различимым,
ядрышко располагалось эксцентрично. Контуры поверхности тела и ядра

91
были отчетливыми. Маломодифицированные клетки визуально были
несколько крупнее клеток этого типа в контролях, располагались вблизи
стенки кровеносных капилляров и были окружены клетками-сателлитами.
Статистической оценки площади тел клеток этого типа не производилось в
связи с их небольшим количеством.
Тела светлых нейронов выглядели более крупными, чем остальные
виды нейронов, обладали округлой или овальной формой, не содержали
нисслевских глыбок. Округлое ядро этих клеток, визуально было
значительно крупнее ядра нейронов других видов; пространства между
нитями хроматина в нем были шире, за счет чего ядро представлялось более
светлым. Ко внутренней поверхности контура ядра, более тонкого, чем в
контролях, но отчетливого на всем протяжении, обычно прилежало ядрышко
(см. рис. 14). Доля светлых нейронов в ПЦинК после ишемии, по сравнению
с долей этих клеток в группах контроля, увеличилась в 3,6–4 раза (до
14,5±1,8; табл. 6). Площадь тел светлых нейронов после ишемии оказалась
больше этого значения в группах контроля в 2–2,4 раза (р<0,05).
Нейроны при ишемии приобретали светлый фенотип вследствие
набухания, поскольку они, в отличие от контролей, становились более
крупными, утрачивали неправильную форму и становились округлыми,
содержали истонченный контур кариотеки; в их ядрах увеличивались
межхроматинные пространства и становилась выраженной эксцентричное
положение ядрышек. Большинство тел светлых набухших нейронов
располагалось вблизи стенки капилляров, а также они были объединены в
пары и группы, состоявшие их нескольких тел нейронов: таких же набухших
клеток, темных сморщенных, маломодифицированных или клеток-«теней».
Расстояние между телами светлых, единичных маломодифицирован-
ных нейронов и стенкой капилляра сократилось в 3,3–4 раза (табл. 7). Вблизи
тел большинства набухших нейронов располагались клетки-сателлиты.
Среди нейронов ПЦинК после ишемии выявлялись также тела
нейронов, обладавшие гиперхромными ядром и цитоплазмой, находившиеся

92
50 мкм
Рисунок 14. Изменения клеток в V–VI слоях передней цингулярной коры через 7 сут
ишемии на площади 0,01 мм2 (пунктир): а – окраска методом Ниссля, б – выявление GFAP
астроцитов и окрашивание ядер гематоксилином. Нейроны: Н – маломодифициро-ванные (в паре), Г – светлые набухшие (гипохромные), С – темные сморщенные, Т – клетки-«тени»; А – астроциты, Э – эндотелиоциты; олигодендроциты – стрелки, микроглиоциты – звездочки; fpm – волокна медиального пучка переднего мозга. Ок. х10, об. х100

93
на разных этапах сморщивания. Деформированный контур поверхности этих
темных нейронов в ряде участков был нечетким; в цитоплазме многих клеток
этого вида слабо выделялись грубые темные базофильные комочки. В теле
этих клеток также слабо выделялся деформированный контур поверхности
ядра и ядрышко. Многие темные сморщенные нейроны были объединены в
пары с другими нейронами, были окружены клетками-сателлитами и часто
располагались вблизи стенки кровеносного капилляра.
Доля темных сморщенных клеток после ишемии увеличилась 9,1–13,4
раза по сравнению с их долей в контролях (до величины 36,3±2,4; р<0,05) и,
таким образом, клетки данного типа в ПЦинК при ишемии стали преобла-
дающей формой потенциально жизнеспособных нейронов (см. табл. 6).
Наружный контур тела клетки-«тени» обычно был нечетким,
«размытым», деформированным по типу набухания или сморщивания, у
части клеток не определялся. Теневидные нейроны часто имели вид
деформированных фрагментов ядра, содержащих просветленные остатки
нуклеоплазмы. Иногда клетки-«тени», находясь в начале гибели, содержали
набухшие ядра, отличались от ядер набухших гипохромных клеток
размытыми контурами ядерной оболочки, повышенной прозрачностью ядер
и участками отсутствия эухроматина. Клетки-«тени» могли входить в состав
пар и групп нейронов наряду с жизнеспособными клетками, располагаться
вблизи стенки капилляров, однако визуально чаще на значительно большем
расстоянии от стенки капилляров, чем остальные формы поврежденных
нейронов. Последние могли располагаться между ними и стенкой сосуда.
Вблизи теневидных нейронов клетки-сателлиты обычно отсутствовали,
однако нередко располагались микроглиоциты, что подтверждается при
выявлении их Iba-1 антигена (рис. 15). Доля теневидных клеток ПЦинК при
ишемии возросла в 5,8–6,6 раза по сравнению с их долей в группах контроля
(р<0,05), достигнув величины, почти равной половине потенциально
жизнеспособных и жизнеспособных клеток (48,5±2,0).

94
Клетки-сателлиты в ПЦинК после ишемии располагались вблизи тел
малоизмененных, большинства гипохромных, сморщенных гиперхромных
нейронов и их групп. Среди последних были различимы светлые и темные
разновидности тел олигодендроцитов, так и тела астроцитов, выявленные в
том числе при окрашивании GFAP (см. рис. 14б).
Расстояние между телами маломодифицированных, набухших нейронов
и стенкой капилляра после ишемии сократилось, по сравнению с параметром
в группах контроля, в 3,3–4,1 раза (р<0,05; см. табл. 7). Доля нейронов
разных фенотипов, объединенных в пары, при ишемии, а также глиоцито-
нейрональный индекс, по сравнению с этими величинами в контрольных
группах значительно возросли (р<0,05).
При окрашивании по Нисслю были различимы тела астроцитов,
обычно обладавшие звездчатой формой, содержавшие округлое или овальное
ядро и светлый ободок цитоплазмы, переходящий на периферии в основание
главных отростков. Тела астроцитов визуально были в 1,5–4 раза мельче тел
нейронов. Ядро астроцитов было светлее, чем у клеток в группах контроля,
обычно почти прозрачным и однородным, поскольку эухроматин не
обнаруживался. Контуры ядерной оболочки были четкими и ровными,
ядрышко располагалось вблизи внутренней поверхности кариотеки.
Многие астроциты являлись клетками-сателлитами для набухших,
темных сморщенных и маломодифицированных клеток. Большинство
астроцитов одновременно являлись также клетками перивазальной формы
глии, поскольку располагались вблизи стенки кровеносных капилляров,
ориентируясь вдоль этой стенки. Друг с другом тела астроцитов обычно пары
не образовывали.
В срезах ПЦинК, окрашенных для выявления промежуточного
филамента астроцитов, тела последних также характеризовались отеком
цитоплазмы и ядра, разреженностью, фрагментацией и смещением GFAP+
материала к одной из сторон клеточного тела (см. рис. 14б). Протеин во
многих главных отростках выглядел более грубым, чем в группах контроля,

95
комковатым и прерывистым, смещенным к телу клетки. Вследствие этого
отростки, оказавшиеся в плоскости среза, выглядели более короткими.
Участки разрыхленности или тонкопетлистого строения протеина в
отростках астроцитов, характерные клеткам в группах контроля,
отсутствовали. Уровень экспрессии GFAP в концевой части отростков
астроцитов и периваскулярных глиальных мембранах был снижен, протеин в
этих частях клеток выглядел истонченным, прерывистым и в наибольшей
части данных сруктур отсутствовал.
GFAP многих отростков клеток, был вытянут в направлении стенки
ближайшего капилляра или прилежал к ней, формируя перикапиллярную
глиальную мембрану. Тела астроцитов, расположенные вблизи сосудов,
также были ориентированы параллельно их стенке.
Количество астроцитов в ПЦинК при ишемии не различалось с их
числом в группах контроля (табл. 8). Площадь тел астроцитов, по сравнению
с контролями, возросла в 1,4–1,5 раза (р<0,05). Расстояние между телами
астроцитов и стенкой капилляра, установленное как при обзорном
окрашивании клеток, так и при выявлении их GFAP, сократилось в 4,1–14,6
раза (р<0,05).
Микроглиоциты в ПЦинК при переживании ишемии обладали
признаками трансформации в клетки амебоидной формы. Их тела были
увеличены в размерах и изменены в форме. Клетки приобрели округлую,
овальную, веретеновидную, звездчатую, треугольную формы (см. рис. 14а и
15). При окраске их Iba-1 антигена маркер был выявлен в составе их
утолщенных главных отростках, расположенных вдоль оси миграции клеток.
Многие из этих клеток были удалены от стенки кровеносных капилляров и
окружали тела измененных нейронов (теневидных, гипохромных и
пикноморфных).
Активированные микроглиоциты, по сравнению с группами контроля,
были увеличены в количестве (в 2,2 раза; р<0,05). Была значительно
увеличена также и площадь их тел (в 1,3–1,5 раза; табл. 9).

96
Капилляры в срезах ПЦинК после ишемии определялись чаще, чем в
препаратах групп контроля, были гораздо протяженнее, часто содержали
бифуркационные разделения. Наиболее часто капилляры в срезах были
расположены продольно и косо-продольно, причем их длина в плоскости
среза визуально была значительно больше, чем в контролях (см. рис. 14).
Рисунок 15. Структурно-топографические и количественные особенности микроглиоцитов в глубоких слоях передней цингулярной коры через 7 сут ишемии.
Пунктир – площадь исследования, равная 0,01 мм2 . fpm – волокна медиального пучка
переднего мозга. Окраска Iba микроглиоцитов и ядер гематоксилином. Ок. х10, об. х63
Ядра эндотелиоцитов визуально были многочисленнее, значительно
крупнее и светлее, чем в группах контроля. Число эндотелиоцитов
капилляров при переживании ишемии было больше их количества в
контролях в 1,7 раз; площадь ядер – в 1,9–2 раза (р<0,05; см. табл. 9).
3.2.2. Изменения клеточного строения прилежащего ядра переднего мозга (область выраженной ишемии)
Среди нейронов прилежащего ядра (ПЯ) при переживании глобальной
ишемии мозга преобладали клетки-«тени» и набухшие гипохромные формы
клеток. Сморщенные-гиперхромные нейроны были малочисленными, мало-
измененные нейроны определялись в единичном количестве (рис. 16; см.
табл. 6). Строение тел этих видов патологически измененных нейронов в ПЯ
и ПЦинК после ишемии было аналогичным. Доля гипохромных клеток (по
сравнению со значением неизмененных нейронов в группах контроля) увели-
чилась в 7,8–8,8 раза, а площадь – в 2,4 раза, достигнув величины 143,4±4,5

97
мкм2
(см. табл. 7). Наиболее многочисленными клетками после ишемии в ПЯ
оказались клетки-«тени». Их доля, по сравнению с контролями, значительно
возросла и достигла 49,8±2,4%.
50 мкм
Рисунок 16. Изменения клеток прилежащего ядра переднего мозга через 7 сут
ишемии на площади 0,01 мм2 (в пределах каждого из 7 квадратов). Нейроны: Н –
мамодифицированные, Г – набухшие (гипохромные), С – темные сморщенные, Т – клетки-«тени»; А – астроциты, Э – эндотелиоциты; олигодендроциты – стрелки , микроглиоциты – звездочки ; са – передняя спайка мозга. Центр окружности (пунктир) радиусом 20 мкм совмещен со стенкой кровеносного капилляра, ближайшей к телу нейрона. Окраска методом Ниссля. Ок. х10, об. х63

98
5 0 мк м
Рисунок 16в. Увеличенный фрагмент «в» из рис. 16. Обозначения те же. Окраска методом Ниссля. Ок. х10, об. х63
Тела многих нейронов были между собой объединены в пары,
небольшие группы и располагались преимущественно вблизи стенки
капилляров. Положение между капиллярами занимали, главным образом,
клетки-«тени». Большинство нейронов – набухших гипохромных, сморщен-
ных темных и маломодифицированных клеток были окружены клетками-
сателлитами, среди которых идентифицировались тела астроцитов и
олигодендроцитов.
Расстояние между телами жизнеспособных нейронов (маломодифи-
цированных, гипохромных) и стенкой капилляров сократилось (по
сравнению с основным контролем) в 2,5–2,8 раза (р<0,05). Доля нейронов,
образующих пары в ПЯ, возросла в 2,7–2,8 раз (р<0,05). Глиоцито-
нейрональный индекс возрос в 2,1 раза по сравнению с индексом в ПЯ групп
контроля, тогда как количество астроцитов, по сравнению с их количеством у
контрольных крыс, осталось прежним (см. табл. 8).

99
Через 7 суток ишемии строение астроцитов ПЯ, в сравнении с этими
клетками в ПЦинК, изменилось аналогичным образом. Астроциты
отличались от клеток в группах контроля признаками отека цитоплазмы и
ядра, разреженностью, фрагментацией и смещением GFAP+ материала к
одной из сторон клеточного тела (рис. 17; см. также рис. 16). Площадь тел
астроцитов, по сравнению с этим параметром группах контроля, увеличилась
в 1,3 раза (р<0,05; см. табл. 8). Уровень экспрессии GFAP в отростках
астроцитов был снижен, GFAP-материал на некоторых участках
периваскулярных глиальных мембран капилляров был неразличим. Между
GFAP-материалом в составе некоторых перикапиллярных глиальных
мембран, и прилежащим к нему снаружи веществом мозга были различимы
щелевидные оптически прозрачные пространства (см. рис. 17 а, е).
Расстояние между телами астроцитов и стенкой капилляра, измененное как
при окраске по Нисслю, так и на GFAP, сократилось (в 2,6–3,4 раза).
Тела микроглиоцитов прилежащего ядра при переживании гипоксии
равномерно и интенсивно экспрессировала Iba-1 антиген (рис. 18). Форма тел
клеток характеризовалась наличием у них закругленных наружных контуров
и коротких немногочисленных выступов. Она была округлой, овальной,
треугольной, трапециевидной, листовидной, звездчатой, но в целом
приближена к округлой. Большинство микроглиоцитов определялись на
значительном расстоянии от стенки капилляров. Клетки, выступая в качестве
нейронофагов, располагались вблизи патологически нейронов, в том числе
теневидных, реже – одиночно или вблизи других глиальных клеток.
По сравнению с группами контроля микроглиоциты были увеличены в
количестве (в 1,7 раза; р<0,05); также была увеличена и площадь их тел (в
1,2–1,7 раза; р<0,05; см. табл. 9).

100
5 0 мк м
Рисунок 17. Структурно-топографические изменения астроцитов прилежащего
ядра переднего мозга крыс через 7 сут ишемии на площади 0,01 мм2 в 7-ми
последовательных квадратах. Вставки – увеличенные фрагменты среза. Пунктир –
площадь исследования, равная 0,01 мм2. Астроциты – стрелки; са – передняя спайка
мозга. Центр окружности (пунктир) радиусом 20 мкм совмещен со стенкой кровеносного капилляра, ближайшей к телу астроцита. Окрашивание GFAP астроцитов и ядер клеток гематоксилином. Ок. х10, об. х63
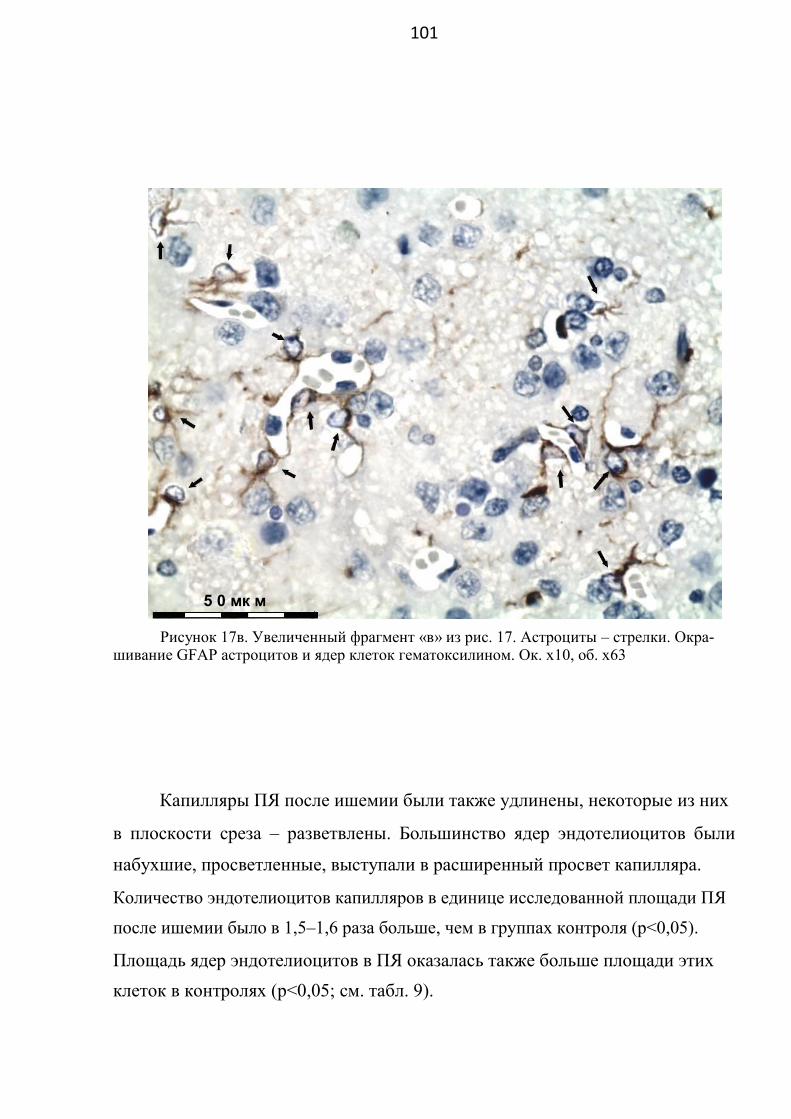
101
5 0 мк м
Рисунок 17в. Увеличенный фрагмент «в» из рис. 17. Астроциты – стрелки. Окра-шивание GFAP астроцитов и ядер клеток гематоксилином. Ок. х10, об. х63
Капилляры ПЯ после ишемии были также удлинены, некоторые из них
в плоскости среза – разветвлены. Большинство ядер эндотелиоцитов были
набухшие, просветленные, выступали в расширенный просвет капилляра.
Количество эндотелиоцитов капилляров в единице исследованной площади ПЯ
после ишемии было в 1,5–1,6 раза больше, чем в группах контроля (р<0,05).
Площадь ядер эндотелиоцитов в ПЯ оказалась также больше площади этих
клеток в контролях (р<0,05; см. табл. 9).

102
5 0 м к м
Рисунок 18. Изменения расположения и строения микроглиоцитов прилежащего ядра через 7 сут экспериментальной ишемии (а – е). Окраска Iba-1 микроглиоцитов и
гематоксилином. Ок. х10, об. х63. Пунктир – площадь исследования, равная 0,01 мм2
. Вставки – увеличенные фрагменты среза; са – передняя спайка мозга

103
3.2.3. Изменения клеточного строения паранигрального ядра среднего мозга (область слабо выраженной ишемии)
Через 7 суток глобальной ишемии мозга в препаратах паранигрального
ядра среднего мозга (ПНигЯ) были выражены перикапиллярный отек и
спектр патологических изменений тел нейронов, в котором превалировало
отек-набухание клеток. Тела многих нейронов ПНигЯ при ишемии, как и в
группах контроля, характеризовавшиеся удлиненной формой, различимым (у
некоторых из них) основанием главного отростка, располагались
параллельно друг другу и их общей оси, соответствующей направлению
волокон медиального пучка переднего мозга (рис. 19).
Некоторые нейроны были изменены в наименьшей степени и, как и в
группах контроля, содержали многочисленные мелкие глыбки вещества
Ниссля. Последние были распределены в цитоплазме диффузно или, реже,
занимали ее наибольшую часть. В основании главных отростков нейронов
хроматофильные глыбки отсутствовали. Между глыбками были различимы
тонкие светлые пространства, визуально более широкие, чем в группах
контроля. Ядро было более просветленным, чем в группах контроля, а размер
тела визуально – более крупным.
Доля малоизмененных клеток при ишемии в ПНигЯ (4,7±1,6%)
оказалась значительно больше, чем в ПЦинК и ПЯ (в 2,1 и 6,7 раза
соответственно; см. табл.6). Маломодифицированные клетки были окружены
сателлитами, располагались преимущественно вблизи стенки капилляров,
пар с другими нейронами обычно не образовывали (см. табл. 7). Площадь тел
малоизме-ненных клеток была в 1,2 раза больше, чем в группах контроля
(р<0,05).
Тела набухших клеток были лишены нисслевских глыбок или
содержали отдельные глыбки; цитоплазма и ядро этих клеток выглядели
набухшими и просветленными. Многие набухшие нейроны обладали слегка
выпуклыми четкими контурами клеточной и ядерной оболочки.

104
50 мкм
50 мкм
Рисунок 19. Изменения строения и расположения клеток паранигрального ядра
среднего мозга через 7 сут ишемии на площади 0,01 мм2 (пунктир). Нейроны: Н –
маломодифицированные, Г – набухшие (гипохромные), С – сморщенные темные, Т – клетки-«тени»; А – астроциты, Э – эндотелиоциты; олигодендроциты – стрелки, микроглиоциты – звездочки. Центр окружности (пунктир) радиусом 20 мкм совмещен со стенкой кровеносного капилляра, ближайшей к телу нейрона. Окраска методом Ниссля. Ок. х10, об. х63
Доля этих клеток в исследованных формациях возросла в 4,1–4,8 раза
(по сравнению со значением у крыс в группах контроля) и превысила в них
50% от общего числа нейронов (см. табл. 6). Площадь тел набухших
нейронов в исследованных ядрах увеличилась (по сравнению с контролями) в
1,8–2,1 раза и достигла 186,1±13,5 мкм2. Тела нейронов этого типа в ПНигЯ
обычно были окружены сателлитоцитами, пар не образовывали и
располагались как вблизи стенки капилляров, так и вдали от последней.
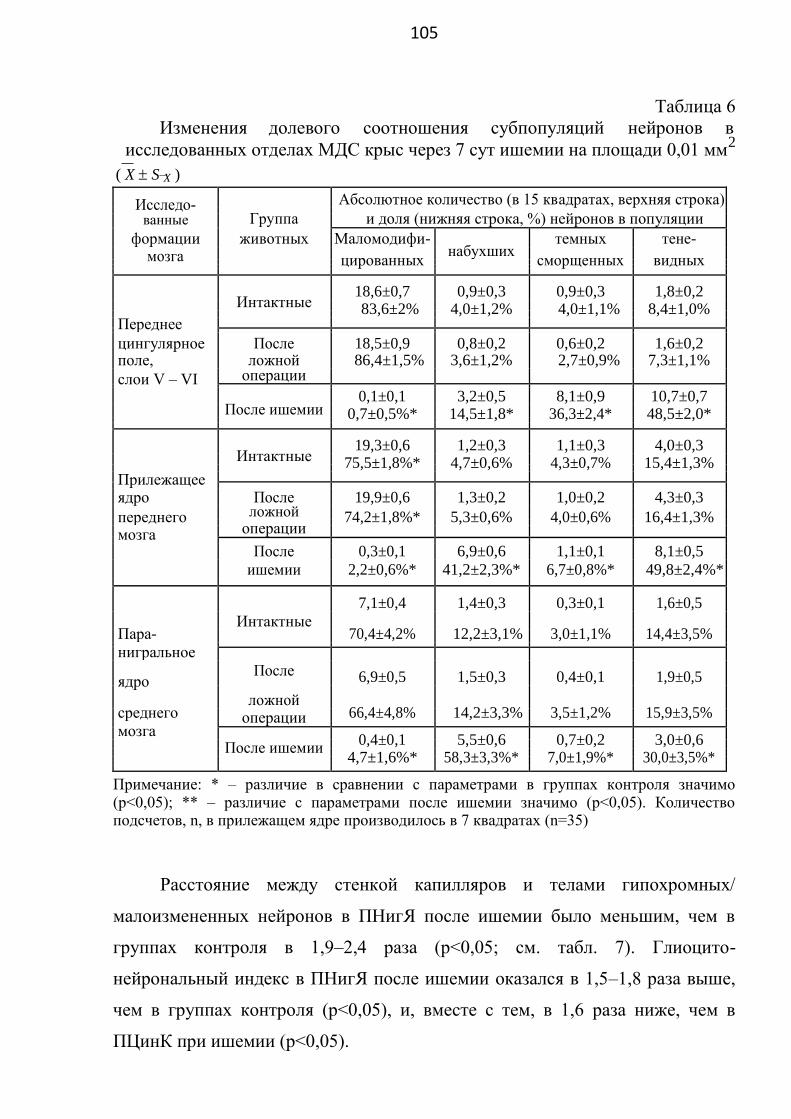
105
Таблица 6
Изменения долевого соотношения субпопуляций нейронов в
исследованных отделах МДС крыс через 7 сут ишемии на площади 0,01 мм2
( X S X )
Исследо- Абсолютное количество (в 15 квадратах, верхняя строка)
Группа и доля (нижняя строка, %) нейронов в популяции ванные
формации животных Маломодифи- набухших
темных тене- мозга
цированных сморщенных видных
Интактные
18,6±0,7 0,9±0,3 0,9±0,3 1,8±0,2 83,6±2% 4,0±1,2% 4,0±1,1% 8,4±1,0% Переднее
После
цингулярное 18,5±0,9 0,8±0,2 0,6±0,2 1,6±0,2 поле, ложной 86,4±1,5% 3,6±1,2% 2,7±0,9% 7,3±1,1%
слои V – VI операции
После ишемии 0,1±0,1 3,2±0,5 8,1±0,9 10,7±0,7
0,7±0,5%* 14,5±1,8* 36,3±2,4* 48,5±2,0*
Интактные
19,3±0,6 1,2±0,3 1,1±0,3 4,0±0,3 75,5±1,8%* 4,7±0,6% 4,3±0,7% 15,4±1,3% Прилежащее
После
ядро 19,9±0,6 1,3±0,2 1,0±0,2 4,3±0,3
переднего ложной 74,2±1,8%* 5,3±0,6% 4,0±0,6% 16,4±1,3% операции мозга
После 0,3±0,1 6,9±0,6 1,1±0,1 8,1±0,5
ишемии 2,2±0,6%* 41,2±2,3%* 6,7±0,8%* 49,8±2,4%*
Интактные
7,1±0,4 1,4±0,3 0,3±0,1 1,6±0,5
Пара- 70,4±4,2% 12,2±3,1% 3,0±1,1% 14,4±3,5%
нигральное
После 6,9±0,5 1,5±0,3 0,4±0,1 1,9±0,5 ядро
ложной 66,4±4,8% 14,2±3,3% 3,5±1,2% 15,9±3,5% среднего операции
мозга
После ишемии 0,4±0,1 5,5±0,6 0,7±0,2 3,0±0,6
4,7±1,6%* 58,3±3,3%* 7,0±1,9%* 30,0±3,5%*
Примечание: * – различие в сравнении с параметрами в группах контроля значимо (р<0,05); ** – различие с параметрами после ишемии значимо (р<0,05). Количество подсчетов, n, в прилежащем ядре производилось в 7 квадратах (n=35)
Расстояние между стенкой капилляров и телами гипохромных/
малоизмененных нейронов в ПНигЯ после ишемии было меньшим, чем в
группах контроля в 1,9–2,4 раза (р<0,05; см. табл. 7). Глиоцито-
нейрональный индекс в ПНигЯ после ишемии оказался в 1,5–1,8 раза выше,
чем в группах контроля (р<0,05), и, вместе с тем, в 1,6 раза ниже, чем в
ПЦинК при ишемии (р<0,05).

106
Таблица 7 Изменения морфометрических параметров нейронов и глиоцито-нейронального индекса в исследованных
отделах МДС крыс через 7 сут ишемии на площади 0,01 мм2 (
S
X
)
X
Исследо-
Площадь Расстояние между телами Доля Глиоцито-
тел нейронов, мкм 2 нейронов и стенкой капилляра, мкм
ванные Группа нейронов,
нейрональ-
формации животных
объединенных
маломодифи- набухших
маломодифи-
набухших ный индекс
мозга
цированных цированных
в пары
Интактные
85,4±2,9 70,4±2,9 8,7±0,8 7,4±1,6 9,3±1,4 0,13±0,03
Переднее (n=26) (n=17)
(n=48)
(n=12) (n=15) (n=15)
цингуляр- После ложной 90,5±4,6 86,5±5,0 8,8±0,9 7,0±1,8 8,5±1,4 0,11±0,03
ное поле, операции (n=25) (n=17) (n=50) (n=8) (n=15) (n=15)
слои V – VI После ишемии — 172,3±9,1* 2,1±1,0* 23,5±2,0* 0,36±0,03*
(n=20) (n=20) (n=15) (n=15)
Интактные 59,8±3,0 51,8±4,7 8,2±1,2 9,1±2,5 9,4±1,4 0,11±0,01 1
06
Прилежащее (n=24) (n=12) (n=28) (n=11) (n=35) (n=35)
После ложной 58,8±2,4 50,9±4,4 8,8±1,2
9,8±2,0 9,0±1,3 0,14±0,02
ядро
переднего операции (n=28) (n=12) (n=28) (n=11) (n=35) (n=35) мозга
После ишемии 79,5±7,4* 143,4±4,5* 3,1±1,1* 3,2±0,9* 25,3±1,9* 0,29±0,02*
(n=12) (n=28)
(n=10)
(n=28) (n=35) (n=35)
Пара- Интактные
103,9±9,0 89,0±6,0 8,3±1,7 6,8±2,5 3,7±1,8 0,15±0,03 (n=20) (n=9) (n=15) (n=10) (n=15) (n=15)
нигральное
После ложной 106,7±8,9 99,9±9,3 8,4±1,5 7,4±1,9 4,3±2,3 0,12±0,03
ядро
операции (n=20) (n=12)
(n=15)
(n=12) (n=15) (n=15)
среднего
125,1±6,2* 183,1±11,6*
3,5±1,2* 3,6±1,5 0,22±0,04*
мозга После ишемии
(n=11) (n=15) (n=20) (n=15) (n=15) Примечание: n – количество подсчетов; * – различие в сравнении с параметрами в группах контроля значимо (р<0,05); ** – различие с параметрами после ишемии значимо (р<0,05)

107
Доля темных сморщенных и теневидных нейронов ядер среднего мозга
при переживании ишемии увеличилась, по сравнению с их удельным
количеством в группах контроля, в 2–2,2 раза (р<0,05). Пикноморфные
клетки наряду с маломодифицированными являлись при ишемии самыми
малочисленными субпопуляциями нейронов в ПНигЯ: доля первых
составила 7,0±1,9%, вторых – 4,7±1,6%. Удельное количество клеток этих
двух субпопуляций между собой значительно не различалось (р<0,05). Доля
клеток-«теней» в ПНигЯ при гипоксии составила треть от общего числа
нервных клеток (30,0±3,5%). Сморщенные темные нейроны и клетки-«тени»
в ПНигЯ при ишемии обычно не были окружены клетками-сателлитами и
располагались вдали от стенки сосудов.
Астроциты ПНигЯ после ишемии были значительно изменены. Ядро
астроцитов отчетливо окрашивалось методом Ниссля, по интенсивности
окрашивания и строению не различалось с контролями; в нем был различим
эухроматин и ядрышко, обычно занимавшее центральную часть ядра. Ядро
было окружено светлым ободком цитоплазмы, значительная часть которой у
многих клеток была сдвинута к одной из сторон клетки (рис. 19).
При окрашивании GFAP астроцитов было заметно значительное
увеличение размеров тел клеток за счет их GFAP-материала, смещение GFAP
к одной из сторон клеточного тела, оттесняющего ядро в противоположную
сторону, и расширение оснований главных отростков по сравнению с
контролями, в которых концентрировалась основная часть протеина (рис. 20).
Главные отростки астроцитов, срезанные на части своей длины продольно,
визуально были, по сравнению с контролями, утолщены. Вблизи поверхности
тел и отростков астроцитов обычно определялась периферическая
тонкопетлистая сеть протеина. Перикапиллярые глиальные мембраны
содержали широкую непрерывную сеть промежуточного филамента. Между
перикапиллярной глиальной мембраной и прилежащим к ней снаружи
веществом мозга были видны щелевидные прозрачные пространства.

108
50 мкм
Рисунок 20. Структурно-топографические изменения астроцитов паранигрального ядра среднего мозга крыс через 7 сут ишемии. Пунктир – площадь исследования, равная
0,01 мм2
. Окрашивание GFAP астроцитов и ядер клеток гематоксилином. Ок. х10, об. х63
Астроциты располагались преимущественно вблизи стенки кровеносных
капилляров и являлись сателлитами.
Количество астроцитов в ПНигЯ после ишемии, по сравнению с
группами контроля, значительно не изменилось (р>0,05). Площадь их тел
увеличилась в 1,7–1,8 раза. Расстояние между телами астроцитов и стенкой
капилляра, установленное как при окраске по Нисслю, так и в срезах,
окрашенных на GFAP, сократилось в 4,3–6,1 раза (табл. 8).
Микроглиоциты в ПНигЯ после ишемии приобрели фенотип
активированных амебоидных клеток (рис. 21, см. также рис. 19). Они
обладали значительно более крупными размерами, чем в группах контроля,
округлой или овальной формой тел и располагались обычно на удалении от
стенки капилляров. Отдельные микроглиоциты располагались вблизи клеток-
«теней», являясь нейронофагами; иные определялись по ходу сосудов. Среди
последних встречались элементы, связанные со стенкой капилляра короткой
цитоплазматической ножкой.
Число клеток микроглии в ПНнигЯ, в сравнении с их количеством в
контролях, было увеличено в 1,6–1,7 раза (р<0,05). Также была увеличена и
площадь их тел (в 1,5 раза; р<0,05; см. табл. 9).
Капилляры ПНигЯ после ишемии были малочисленными, срезаны
под разным углом, просвет их содержал эритроциты, плазму крови, либо был

109
оптически прозрачный. Снаружи стенки капилляров обычно определялись
оптически прозрачные щели. Ядра эндотелиоцитов были мелкими,
гиперхромными, ядрышко обычно различимо не было (рис. 19–21).
Таблица 8
Изменения количества и морфометрических параметров астроцитов в
исследованных отделах МДС через 7 сут ишемии ( X S X )
Исследо- Коли- Расстояние между
Площадь Группа чество телами астроцитов ванные при и стенкой капилляров, тел, формации животных
окраске мкм, при окраске мкм2
мозга
GFAP по Нисслю GFAP
Интактные
4,7±0,5 9,4±1,9 9,7±1,7 41,5±3,3
(n=15) (n=15) (n=15) (n=19) Переднее
После
цингуляр- 4,8±0,4 10,3±1,6 8,9±1,5 43,4±2,2 ное поле, ложной
(n=15) (n=15) (n=15) (n=20) слои V – VI операции
После 4,3±0,4 2,0±0,9* 1,5±0,8* 60,1±5,8*
ишемии (n=15) (n=15) (n=15) (n=15)
Интактные
3,6±0,3 10,4±1,1 10,9±1,1 38,0±1,5
(n=35) (n=35) (n=35) (n=35)
Прилежащее После 3,3±0,3 10,8±1,2 11,0±1,2 38,0±1,5
ядро ложной (n=35) (n=35) (n=35) (n=35) операции переднего
мозга После 3,7±0,3 2,9±0,7* 3,4±0,8* 49,5±2,4*
ишемии (n=35) (n=35) (n=35) (n=35)
Интактные
3,7±0,4 9,7±1,5 10,4±1,6 38,7±3,2
(n=15) (n=15) (n=15) (n=30)
Пара- После
3,9±0,5 9,8±1,5 10,4±1,3 39,3±2,9
ложной нигральное (n=15) (n=15) (n=15) (n=29) операции ядро
После 3,5±0,4 1,6±0,9* 2,4±1,3* 68,0±4,9* среднего
ишемии (n=15) (n=15) (n=15) (n=18) мозга
Примечание: n – количество подсчетов астроцитов; * – различие в сравнении с параметрами в группах контроля значимо (р<0,05)
Число эндотелиоцитов (на единице исследованной площади) в ПНЯ
после ишемии значительно не различалось с числом этих клеток в контролях

(р>0,05); площадь ядер этих клеток при ишемии была немного больше (в 1,7
раза), чем в группах контроля (р<0,05; см. табл. 9).

110
50 мкм
Рисунок 21. Структурно-топографические изменения микроглиоцитов параниграль-ного ядра среднего мозга крыс через 7 сут ишемии . Пунктир – площадь исследования,
равная 0,01 мм2. Окрашивание Iba микроглиоцитов и ядер клеток гематоксилином. Ок.
х10, об. х63
Таблица 9
Изменения количества и морфометрических параметров микроглио-
цитов и эндотелиоцитов в исследованных отделах МДС крыс через 7 сут
ишемии ( X S X )
Исследо- Микроглиоциты Эндотелиоциты ванные
Группа
площадь
площадь формации
количество количество животных тел, мкм2
ядер, мкм2
мозга
Интактные 0,9 ± 0,2 28,2 ± 2,2 2,2 ± 0,3 11,9 ± 1,1
Переднее
После
цингуляр- 0,9 ± 0,3 28,6 ± 2,3 2,3 ± 0,4 10,5 ± 0,7 ложной ное поле, операции слои V – VI
После 2,0 ± 0,4* 39,6 ± 2,2* 3,9 ± 0,3* 23,0 ± 1,5*
ишемии
Интактные 1,0 ± 0,1 28,7 ± 1,3 2,5 ± 0,3 11,5 ± 0,5
Прилежащее
После
ядро 1,1 ± 0,2 27,1 ± 1,2 2,3 ± 0,2 12,3 ± 0,6 ложной переднего операции
мозга
После 2,1 ± 0,3* 37,1 ± 1,2* 3,7 ± 0,4* 22,5± 1,9*
ишемии
Пара- Интактные 1,1 ± 0,2 26,3 ± 1,7 2,6 ± 0,4 10,9 ± 0,6
нигральное После 1,2 ± 0,3 26,8 ± 1,3 2,3 ± 0,4
ядро ложной 10,4± 0,8
среднего операции
мозга После 1,9 ± 0,4* 40,2 ± 3,6* 2,8 ± 0,4 18,1± 1,7* ишемии

Примечание: * – различие в сравнении с параметрами в группах контроля значимо
(р<0,05). Количество подсчетов, n, в прилежащем ядре производилось в 7 квадратах (n=35)

111
3.3. Коррекция метапротом количественных и структурно-топографических изменений клеточных элементов в фокусе выраженной ишемии и за его пределами
3.3.1. Коррекция метапротом ишемических изменений клеточных элементов слоев V–VI переднего цингулярного поля (область выраженной ишемии)
Через 7 суток после лигирования обеих сонных артерий и ежедневного
внутрибрюшинного введения препарата метапрот в передней цингулярной коре
(ПЦинК), расположенной в области выраженного ограничения кровотока, тела
большинства нейронов визуально были малоизмененными, поскольку
сохранили признаки, характерные для этих клеток у интактных крыс (рис. 22).
Они содержали светлое округлое ядро, базофильную цитоплазму и обладали
полигональной, слегка удлиненной и выпуклой формой. Многие
малоизмененные нейроны были объединены в группы, пары и располагались на
разном расстоянии от сосудистой стенки. Значительная, но меньшая часть
нейронов была представлена гиперхромными клетками, уменьшенными в
размерах и сморщенными, которые располагались в пределах глубоких слоев.
Рисунок 22. Передняя цингулярная кора после ишемии и воздействия метапрота. Выделены слой VI и внутренняя часть слоя V, которые были исследованы. Fmi – передние щипцы мозолистого тела. Окраска методом Ниссля. Ок. х10, об. х10

112
При исследовании глубоких слоев ПЦинК с помощью иммерсионного
объектива в цитоплазме маломодифицированных нейронов были различимы
многочисленные глыбки субстанции Ниссля, имеющие вид мелких зерен,
обильно и преимущественно равномерно рассеянных в перикарионе.
Исключение составляли чаще – основание главных отростков, реже – какая-
либо другая часть цитоплазмы, в которых глыбки отсуствовали (рис. 23).
Иногда глыбки Ниссля образовывали скопления в виде одного-трех темных
хлопьев (комков). В светлом ядре, часто имевшем правильную округлую или
овальную форму и четкие контуры поверхности, был различим нитчатый
рисунок эухроматина и крупное ядрышко. Доля малоизмененных нейронов
после ишемии и терапии метапротом возросла, по сравнению с их долей при
ишемии без лечения (0,7±0,5%), в 90,7 раз (до 63,5±4,1%), и оказалась
меньше, чем в группах контроля всего в 1,3–1,4 раза (р<0,05; табл. 10).
Площадь сечения тел малоизмененных нейронов ПЦинК после ишемии и
терапии значительно не различалась с соответствующей величиной в группах
контроля (табл. 11).
Гипохромные нейроны, почти лишенные хроматофильных глыбок,
обладали менее четкими контурами клеточной и ядерной поверхности на
одном из участков или на всем протяжении поверхности, содержали
эксцентрично расположенное ядрышко, которое было меньшей величины и
окрашено слабее, чем у маломодифицированных клеток. Рельеф поверхности
оболочки ядра этих нейронов в некоторых участках был деформирован. Доля
гипохромных нейронов после ишемии и лечения, по сравнению с долей этих
клеток после ишемии без лечения, снизилась в 2,9 раза до 5,0±1,4% (р<0,05).
Площадь сечения перикарионов гипохромных нейронов после ишемии и
лечения метапротом оказалась меньше площади тел нейронов этого типа
после ишемии без лечения в 1,5 раза, но больше в 1,3 раза, чем в группах
контроля (р<0,05, см. табл. 11).
Среди нейронов после ишемии и лечения метапротом выявлялись
также тела нейронов, обладавшие гиперхромными ядром и цитоплазмой,

113
находившиеся на разных этапах сморщивания. В ПЦинК животных, состав-
лявших экспериментальную группу с лечением был выражен разброс числа
сморщенных гиперхромных клеток: от их преобладания над числом мало-
измененных нейронов на исследованной площади до отсутствия (см. рис. 23
а, б). Доля темных сморщенных нейронов после ишемии и лечения в ПЦинК
по сравнению с их долей при ишемии уменьшилась в 2,2 раза (р<0,05).
Теневидные нейроны часто имели вид деформированных фрагментов
ядра, содержащих просветленные остатки нуклеоплазмы и скопления
базофильного материала вблизи его оболочки; иногда клетки-«тени» были
представлены слабо контурируемыми фрагментами ядра или, находясь в
начале гибели, выглядели сморщенными и отличались от жизнеспособных
клеток размытыми контурами клеточной и ядерной поверхности. Доля
теневидных клеток после ишемии и терапии метапротом сократилось в 3,0
раза по сравнению с их долей после ишемии без лечения (р<0,05). Доля
клеток-«теней» в ПЦинК после ишемии и терапии все же оказалась выше
удельного количества этих клеток в группах контроля (в 3,2 раза; р<0,05).
Все типы жизнеспособных нейронов располагались на разном расстоянии
от сосудистой стенки и редко прилежали к последней. Среднее расстояние
между телами малоизмененных, гипохромных нейронов и стенкой капилляра
после ишемии и терапии метапротом значительно не отличалось от
соответствующей величины в группах контроля (р>0,05). Вместе с тем
расстояние между телами гипохромных и малоизмененных клеток, составивших
группу жизнеспособных нервных клеток ПЦК, после ишемии и лечения
оказалось значительно больше, чем после ишемии без лечения (р<0,05).
Доля нейронов разных фенотипов, объединенных в пары, при ишемии
и терапии метапротом, по сравнению с их долей при ишемии без лечения,
значительно не изменилась, вместе с тем она оставалась в 2,5 раза больше,
чем в группах контроля (р<0,05; см. табл. 10). Визуально в пару (группу)
перикарионов редко входили клетки-«тени»; последние чаще располагались
по-отдельности или в составе группы таких же теневидных клеток.

114
50 мкм
Рисунок 23. Строение глубоких слоев передней цингулярной коры после ишемии и
воздействия метапрота на площади 0,01 мм2 (пунктир): а, б – окраска по Нисслю, в –
окрашивание GFAP астроцитов и ядер гематоксилином. Нейроны: маломодифицированные – Н, темные сморщенные – С, светлые (гипохромные) – Г, теневидные – Т; астроциты – А, олигодендроциты – стрелки; эндотелиоциты – Э; микроглиоциты – звездочки. Центр окружности (R=20 мкм) совмещен со стенкой капилляра, ближайшей к телу нейрона и астроцита. Ок. х10, об. х100

115
50 мкм

116
Вблизи тел малоизмененных, гипохромных, сморщенных гиперхромных
нейронов и их групп в ПЦинК после ишемии и лечении метапротом часто
располагались клетки-сателлиты. Среди последних были различимы как
светлые и темные формы тел олигодендроцитов, так и тела астроцитов,
выявленные в том числе и при окрашивании GFAP (см. рис. 23 а,б,в; табл.11).
Глиоцито-нейрональный индекс после ишемии и терапии метапротом
был в 1,3 раза меньше, чем после ишемии (р<0,05), но в 2,5 раза выше, чем в
группах контроля (р<0,05).
В срезах ПЦинК, окрашенных методом Ниссля, обнаруживались тела
астроцитов звездчатой формы, которые выглядели в 1,5–2,5 раза мельче, чем
тела нейронов. Светлое ядро астроцитов обладало чаще правильной округлой
или овальной формой, четкими контурами ядерной оболочки; в нем был
различим хроматин, преимущественно в виде тонкой сеточки, а также одно,
реже два отчетливых ядрышка, расположенных как эксцентрично, так и в
центральной части. Цитоплазма имела вид светлого ободка, расширяющегося
вблизи основания главных отростков.
Тела некоторых астроцитов располагались попарно, причем между
наружной поверхностью оболочки ядер этой пары клеток цитоплазма
различима не была. При окрашивании на GFAP между ядрами многих
объединенных клеток промежуточный филамент также не определялся.
Экспрессия GFAP в срезах ПЦинК была интенсивной и непрерывной в
перинуклеарной части цитоплазмы астроцитов, основании главных отростков
и перикапиллярой глиальной мембране. В областях основания главных
отростков и прилежащей части клеточного тела астроцитов наблюдались
разрастание, сгущение и слияние GFAP этих частей клеток, в результате
которых они выглядели более крупными, чем в контролях, и
деформированными.
Главные отростки астроцитов, срезанные продольно и косо, содержали
интенсивно окрашенную непрерывную сеть промежуточного филамента и
были ориентированы в направлении тел нейронов и стенки капилляра.

117
Вблизи тел нейронов многие отростки астроцитов разветвлялись и
охватывали поверхность перикариона. GFAP-материал, составляющий пери-
капиллярную глиальную мембрану, был визуально утолщен; в концевой
части отростков клеток волокна филамента образовывали участки сгущений.
Число астроцитов в ПЦинК после ишемии и терапии метапротом было
незначительно увеличено, по сравнению с контролями (табл. 12). Расстояние
между стенкой капилляра и телами астроцитов, установленное как при
окраске по Нисслю, так и при выявлении GFAP, по сравнению с этой
величиной при ишемии, было увеличено в 5–6,4 раза и значительно не
различалось с этой величиной у крыс в группах контроля (р>0,05). Площадь
тел астроцитов после ишемии и терапии была в 1,4 больше, чем площадь
клеток у крыс в контроле (р<0,05), но значительно не различалась с этим
параметром у крыс после ишемии.
Микроглиоциты ПЦинК, выявляемые в этой части эксперимента при
окрашивании по Нисслю, располагались преимущественно вблизи тел
патологически измененных нейронов, главным образом теневидных и,
следовательно, являлись нейронофагами. Визуально они обладали более
крупными размерами, чем в контроле, содержали гиперхромное округлое или
бобовидное ядро, были окружены ободком просветленной цитоплазмы, часто
образующей выступы в направлении поврежденных клеток.
Эндотелиоциты капилляров ПЦинК после ишемии и терапии метапротом
по своему количеству и строению ядра визуально не различались с клетками в
контролях. Их ядро было мелким и гиперхромным, в нем часто были различимы
тонкий темный контур ядерной оболочки и ядрышко. Число эндотелиоцитов
после ишемии и лечения, по сравнению с их количеством при ишемии без
лечения, было уменьшено в 1,4 раза (р<0,05; табл. 13). Вместе с тем,
относительно контролей эта величина значительно изменена не была (р<0,05).
Площадь ядер этих клеток после ишемии и лечения также была значительно
уменьшена по сравнению с параметром при ишемии (в 2,3 раза; р<0,05) и
была близка параметрам клеток в группах контроля (р>0,05).

118
3.3.2. Коррекция метапротом ишемических изменений клеточных элементов паранигрального ядра среднего мозга (область слабо выраженной ишемии)
Через 7 суток после лигирования обеих сонных артерий и ежедневного
внутрибрюшинного введения метапрота в ядерных центрах вентральной
части среднего мозга, расположенной за пределами области выраженного
ограничения кровотока, тела большинства нейронов, цитоплазма которых
интенсивно базофильно окрашивалась методом Ниссля (рис. 24), визуально
были малоизмененными.
Рисунок 24. Передняя медиальная часть среднего мозга после ишемии и воздействия метапрота. Выделена исследованная площадь паранигрального ядра, равная
3×0,01 мм2
. Обозначения: 3v – третий желудочек, APR – предкрасноядерное поле, f –
свод, fr – fasciculus retroflexus, lm – медиальная петля, MNC – комплекс сосцевидных ядер, mp – сосцевидная ножка, mtg – сосцевидно-покрышечный путь, NIlm – интерстициальное ядро медиальной петли, NPN – паранигральное ядро, pc – ножка мозга, SNC – компактная часть черной субстанции, ZI – zona incerta, V – полнокровный просвет сосуда венозного типа. Окраска методом Ниссля. Ок. х10, об. х10
При исследовании передней медиальной части паранигрального ядра
(ПНигЯ) с помощью иммерсионного объектива в цитоплазме тел
малоизмененных нейронов обнаруживались многочисленные глыбки Ниссля,

119
имеющие вид мелких зерен, распределенных преимущественно равномерно
(рис. 25). Различий в строении ядра нейронов у клеток этого типа при
лечении ишемии и в группах контроля не отмечалось. Доля малоизмененных
нейронов в ПНигЯ после ишемии и терапии метапротом возросла, по
сравнению с их долей при ишемии без лечения (4,7±1,6%), в 90,7 раз (до
56,9±4,1%), и оказалась меньше, чем в группах контроля всего в 1,2 раза
(р<0,05; табл. 10). Площадь сечения тел нейронов этого типа после ишемии и
терапии значительно не различалась с соответствующей величиной в группах
контроля (р>0,05; табл. 11).
Светлые (гипохромные) нейроны, не содержавшие базофильных
глыбок, отличались от малоизмененных клеток более слабым окрашиванием
ядра, менее четкими контурами клеточной и ядерной поверхности в одном из ее
участков или на всем протяжении. Ядрышко светлых нейронов располагалось
вблизи ядерной оболочки, было слабо окрашено и было меньше, чем у
малоизмененных клеток. Рельеф поверхности ядерной оболочки некоторых
гипохромных нейронов был деформирован. Доля светлых клеток после ишемии
и лечения, по сравнению с их долей после ишемии без лечения, снизилась в 9,7
раза (от 58,3±3,3% до 6,0±2,5%; р<0,05). Площадь сечения тел светлых нейронов
после ишемии и лечения метапротом оказалась меньше площади тел нейронов
этого типа после ишемии без лечения в 1,9 раза; причем она значительно не
различалась с параметрами площади тел гипохромных клеток в группах
контроля (р>0,05; см. табл. 11).
Сморщенные темные нейроны после ишемии и лечения метапротом
были единичными (всего 3,5±1,3% от всех фенотипических форм клеток).
Доля нейронов этого типа после ишемии и лечения метапротом сократилась
по сравнению с их долей при ишемии без лечения в 2,0 раза (р<0,05).
Теневидные нейроны в препаратах были представлены, главным
образом, ядрами и их фрагментами, рельеф оболочки которых был
деформирован, ослаблен и размыт; нуклеоплазма и ее остатки во фрагментах

120
50 мкм 50 мкм
Рисунок 25. Строение паранигрального ядра среднего мозга через 7 сут ишемии и
терапии метапротом на площади 0,01 мм2 (пунктир): а – окраска методом Ниссля, б –
выявление GFAP астроцитов и окрашивание ядер гематокси -лином. Нейроны: маломодифицированные – Н, светлые (гипохромные) – Г, теневидные – Т; астроциты – А, олигодендроциты – стрелки; эндотелиоциты – Э; микроглиоциты – звездочки. Центр окружности (R=20 мкм) совмещен со стенкой капилляра, ближайшей к телу нейрона и астроцита. Ок. х10, об. х100
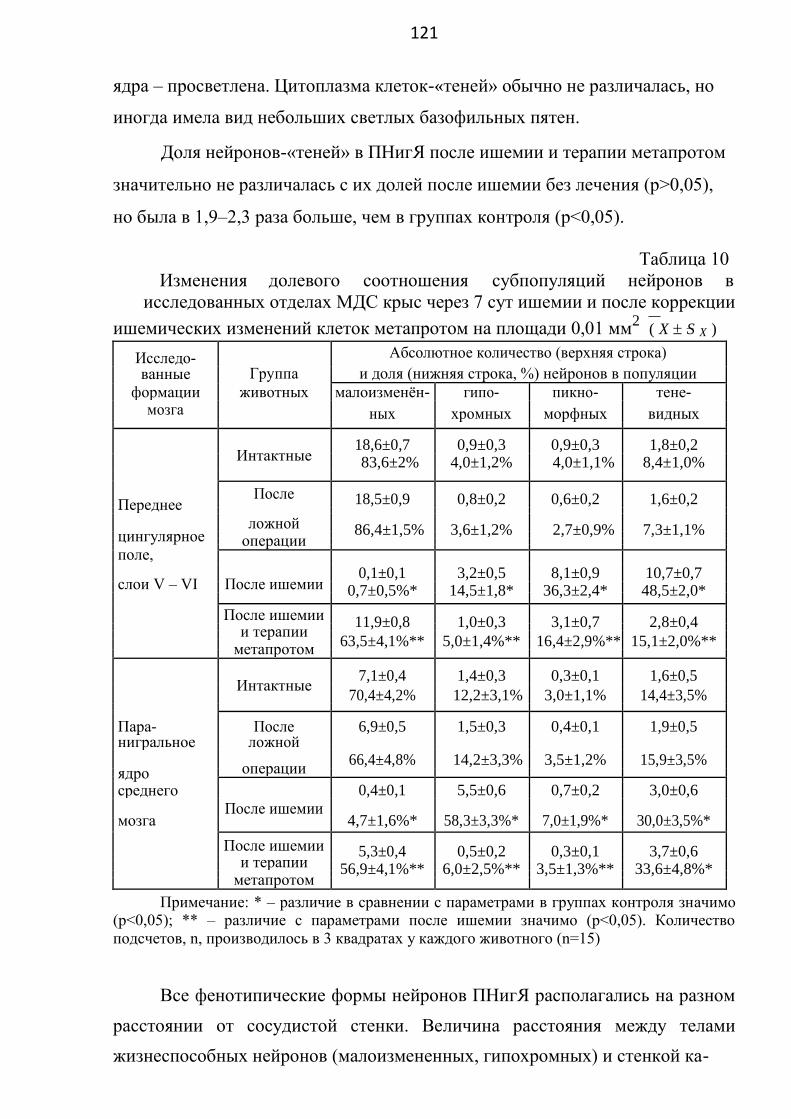
121
ядра – просветлена. Цитоплазма клеток-«теней» обычно не различалась, но
иногда имела вид небольших светлых базофильных пятен.
Доля нейронов-«теней» в ПНигЯ после ишемии и терапии метапротом
значительно не различалась с их долей после ишемии без лечения (р>0,05),
но была в 1,9–2,3 раза больше, чем в группах контроля (р<0,05).
Таблица 10
Изменения долевого соотношения субпопуляций нейронов в
исследованных отделах МДС крыс через 7 сут ишемии и после коррекции
ишемических изменений клеток метапротом на площади 0,01 мм2 ( X S X )
Исследо- Абсолютное количество (верхняя строка)
Группа и доля (нижняя строка, %) нейронов в популяции ванные
формации животных малоизменён- гипо- пикно- тене- мозга ных хромных морфных видных
Интактные
18,6±0,7 0,9±0,3 0,9±0,3 1,8±0,2 83,6±2% 4,0±1,2% 4,0±1,1% 8,4±1,0%
Переднее После 18,5±0,9 0,8±0,2 0,6±0,2 1,6±0,2
ложной 86,4±1,5% 3,6±1,2% 2,7±0,9% 7,3±1,1% цингулярное операции
поле,
0,1±0,1 3,2±0,5 8,1±0,9 10,7±0,7 слои V – VI После ишемии 0,7±0,5%* 14,5±1,8* 36,3±2,4* 48,5±2,0*
После ишемии 11,9±0,8 1,0±0,3 3,1±0,7 2,8±0,4
и терапии 63,5±4,1%** 5,0±1,4%** 16,4±2,9%** 15,1±2,0%** метапротом
Интактные
7,1±0,4 1,4±0,3 0,3±0,1 1,6±0,5
70,4±4,2% 12,2±3,1% 3,0±1,1% 14,4±3,5%
Пара- После 6,9±0,5 1,5±0,3 0,4±0,1 1,9±0,5 нигральное ложной
66,4±4,8% 14,2±3,3% 3,5±1,2% 15,9±3,5% ядро операции
0,4±0,1 5,5±0,6 0,7±0,2 3,0±0,6 среднего
После ишемии мозга 4,7±1,6%* 58,3±3,3%* 7,0±1,9%* 30,0±3,5%*
После ишемии 5,3±0,4 0,5±0,2 0,3±0,1 3,7±0,6
и терапии 56,9±4,1%** 6,0±2,5%** 3,5±1,3%** 33,6±4,8%*
метапротом
Примечание: * – различие в сравнении с параметрами в группах контроля значимо (р<0,05); ** – различие с параметрами после ишемии значимо (р<0,05). Количество
подсчетов, n, производилось в 3 квадратах у каждого животного (n=15)
Все фенотипические формы нейронов ПНигЯ располагались на разном
расстоянии от сосудистой стенки. Величина расстояния между телами
жизнеспособных нейронов (малоизмененных, гипохромных) и стенкой ка-

122
пилляра после ишемии и терапии метапротом значительно не отличалось от
соответствующей величины в группах контроля (р>0,05). Вместе с тем
расстояние между телами жизнеспособных клеток ПНигЯ после ишемии и
лечения оказалось значительно больше, чем после ишемии без лечения
(р<0,05).
Нейроны разных фенотипов ПНигЯ, объединенные в пары, при
ишемии и терапии метапротом, были единичными. Их доля, по сравнению с
удельным количеством пар нейронов при ишемии без лечения, а также в
группах контроля значительно не изменилась (р<0,05; см. табл. 11).
Клетки-сателлиты в ПНигЯ после ишемии и лечении метапротом –
светлые и темные формы олигодендроцитов, астроциты, тела которых
обнаруживались как при окрашивании по Нисслю, так и при выявлении
GFAP, часто сопровождали тела малоизмененных, гипохромных, сморщенных
гиперхромных нейронов. Глиоцито-нейрональный индекс после ишемии и
терапии метапротом был таким же высоким, как и при ишемии без лечения
(р>0,05), но в 1,7–2,1 раза выше, чем в группах контроля (р<0,05).
При окраске срезов ПНигЯ методом Ниссля тела астроцитов
обнаруживались как небольшие клетки звездчатой формы, немного меньшей
величины, чем тела нейронов. Светлое ядро астроцитов обладало правильной
округлой или овальной формой, четкими контурами ядерной оболочки; в нем
был различим эухроматин и одно-два отчетливых ядрышка, расположенных
как эксцентрично, так и в центральной части ядра. Цитоплазма имела вид
светлого ободка, расширяющегося вблизи основания главных отростков.
Тела астроцитов, объединенных в пары, были единичными. Между
ядрами сдвоенных астроцитов обычно был различим GFAP-материал.
Экспрессия GFAP была интенсивной и непрерывной в перинуклеарной
части цитоплазмы астроцитов, отростках, расположенных в плоскости среза
продольно (или косо-продольно), и перикапиллярой глиальной мембране. В
области основания главных отростков некоторых астроцитов и прилежащей
части их клеточного тела было различимо сгущение и слияние GFAP этих

123
Таблица 11 Изменения морфометрических параметров нейронов и глиоцито-нейронального индекса в исследованных отделах МДС крыс
через 7 сут ишемии и после коррекции ишемических изменений клеток метапротом на площади 0,01 мм2 (
S
)
X
X
Исследо-
Площадь Расстояние между телами Доля Глиоцито-
тел нейронов, мкм 2 нейронов и стенкой капилляра, мкм
ванные Группа нейронов,
нейрональ-
формации животных
объединенных
малоизме- гипо- малоизме- гипо- ный индекс
мозга ненных хромных ненных хромных в пары (%)
Интактные
85,4±2,9 70,4±2,9 8,7±0,8 7,4±1,6 9,3±1,4 0,13±0,03
(n=26) (n=17) (n=48)
(n=12)
(n=15) (n=15)
После ложной 90,5±4,6 86,5±5,0 8,8±0,9 7,0±1,8 8,5±1,4 0,11±0,03
Переднее операции (n=25) (n=17) (n=50) (n=8) (n=15) (n=15) цингуляр-
После ишемии — 170,7±9,1*
2,1±1* 23,5±2,0* 0,36±0,03*
12
3
ное поле,
(n=20) (n=20) (n=15) (n=15)
слои V – VI
После ишемии 95,6±3,4 114,8±5,7** 8,4±1,4** 8,3±1,7** 23,0±2,6* 0,27±0,04**
и терапии (n=34) (n=22) (n=20) (n=12) (n=15) (n=15)
Интактные
103,9±9,0 89,0±6,0 8,3±1,7 6,8±2,5 3,7±1,8 0,15±0,03
(n=20) (n=9) (n=15)
(n=10)
(n=15) (n=15)
Пара- После ложной 106,7±8,9 99,9±9,3 8,4±1,5 7,4±1,9 4,3±2,3 0,12±0,03
нигральное операции (n=20) (n=12) (n=15) (n=12) (n=15) (n=15)
ядро После ишемии 125,1±6,2* 183,1±11,6* 3,5±1,2* 3,6±1,5 0,22±0,04*
среднего (n=11) (n=15) (n=20) (n=15) (n=15)
мозга
После ишемии 111,4±7,8** 95,6±7,4** 8,1±1,5** 7,8 ±1,8** 4,2±1,9 0,25±0,04*
(n=20) (n=15) (n=15)
(n=12)
(n=15) (n=15)
и терапии
Примечание: n – количество подсчетов; * – различие в сравнении с параметрами в группах контроля значимо (р<0,05); ** – различие с параметрами после ишемии значимо (р<0,05)

124
частей клеток. Однако, в отличие от строения GFAP при воспроизведении
выраженной ишемии переднего мозга и терапии метапротом, в ПНигЯ
визуально тела астроцитов оставались такими же небольшими, как в группах
контроля.
Главные отростки астроцитов были ориентированы в направлении стенки
капилляра и тел нейронов, ближайших к ним. Концевые части некоторых главных
отростков огибали поверхность тел нейронов или были дихотомически
разветвлены. GFAP-материал, составляющий перикапиллярную глиальную
мембрану, визуально был тонким и непрерывным.
Количество астроцитов ПНигЯ после ишемии и терапии метапротом с их
количеством в контролях и экспериментальных группах значительно не
различалось (см. табл. 12). Расстояние между стенкой капилляра и телами
астроцитов, по сравнению с этой величиной при ишемии, было увеличено в
4,3–6,6 раза и значительно не различалось с этой величиной у крыс в группах
контроля (р>0,05). Площадь тел астроцитов после ишемии и терапии была
значительно меньше площади этих клеток при ишемии без лечения (р<0,05), но
со значениями в группах контроля значительно не различалась (р>0,05).
Микроглиоциты, располагавшиеся вблизи тел теневидных нейронов,
обладали крупными размерами и округлой формой ядра, были окружены
ободком просветленной цитоплазмы, образующей выступы в направлении
поврежденных клеток (псевдоподии). Отдельные микроглиоциты, более
мелкие и уплощенные, не имевшие различимых всевдоподий, располагались
вблизи стенки капилляра или тел жизнеспособных нейронов.
Эндотелиоциты капилляров ПНигЯ после ишемии и терапии метапротом
по своему количеству и строению ядра визуально не различались с клетками в
контролях. Число эндотелиоцитов после ишемии и лечения, по сравнению с их
количеством при ишемии без лечения и группами контроля, значительно не
различалось (р>0,05; см. табл. 13). Площадь ядер этих клеток после ишемии и
лечения была значительно меньше, чем при ишемии без лечения (р<0,05) и со
значением площади ядер в группах контроля значимо не различалась (р>0,05).

125
Таблица 12
Изменения количества и морфометрических параметров астроцитов в
исследованных отделах МДС через 7 сут ишемии и после коррекции
ишемических изменений клеток метапротом на площади 0,01 мм2 ( X S X )
Исследо- Коли- Расстояние между
Площадь Группа чество телами астроцитов ванные при и стенкой капилляров, тел, формации животных
окраске мкм, при окраске мкм2
мозга
GFAP по Нисслю GFAP
Интактные
4,7±0,5 9,4±1,9 9,7±1,7 41,5±3,3*
(n=15) (n=15) (n=15) (n=19)
Переднее После 4,8±0,4 10,3±1,6 8,9±1,5 43,4±2,2
цингуляр- ложной
(n=15) (n=15) (n=15) (n=20) ное поле, операции
слои V – VI После 4,3±0,4 2,0±0,9* 1,5±0,8* 60,1±5,8* ишемии (n=15) (n=15) (n=15) (n=15)
После 4,9±0,6 9,5±1,5** 9,6±1,5** 58,8±5,8* ишемии и
(n=15) (n=15) (n=15) (n=15) терапии
Интактные
3,7±0,4 9,7±1,5 10,4±1,6 38,7±3,2 (n=15) (n=15) (n=15) (n=30)
Пара- После 3,9±0,5 9,8±1,5 10,4±1,3 39,3±2,9
ложной (n=15) (n=15) (n=15) (n=29) нигральное операции
ядро
После 3,5±0,4 1,6±0,8* 2,4±1,3* 68,0±4,9* среднего
ишемии (n=15) (n=15) (n=15) (n=18) мозга
После 3,4±0,4 10,5±1,4** 10,4 ±1,3** 42,7±2,1**
ишемии и (n=15) (n=15) (n=15) (n=31) терапии
Примечание: n – количество подсчетов астроцитов; * – различие в сравнении с параметрами в группах контроля значимо (р<0,05); ** – различие с параметрами после ишемии значимо (р<0,05)
Таким образом, использование препарата метапрот в течение 7 суток
экспериментальной глобальной ишемии мозга способствует нивелированию
признаков токсического отека астроцитов, нейронов и эндотелиоцитов
капилляров, сохраняя при этом многие нейроны маломодифицированными, а

также инициирует укрепление каркаса промежуточного филамента в
перикапиллярных глиальных мембранах, отростках и телах астроцитов.
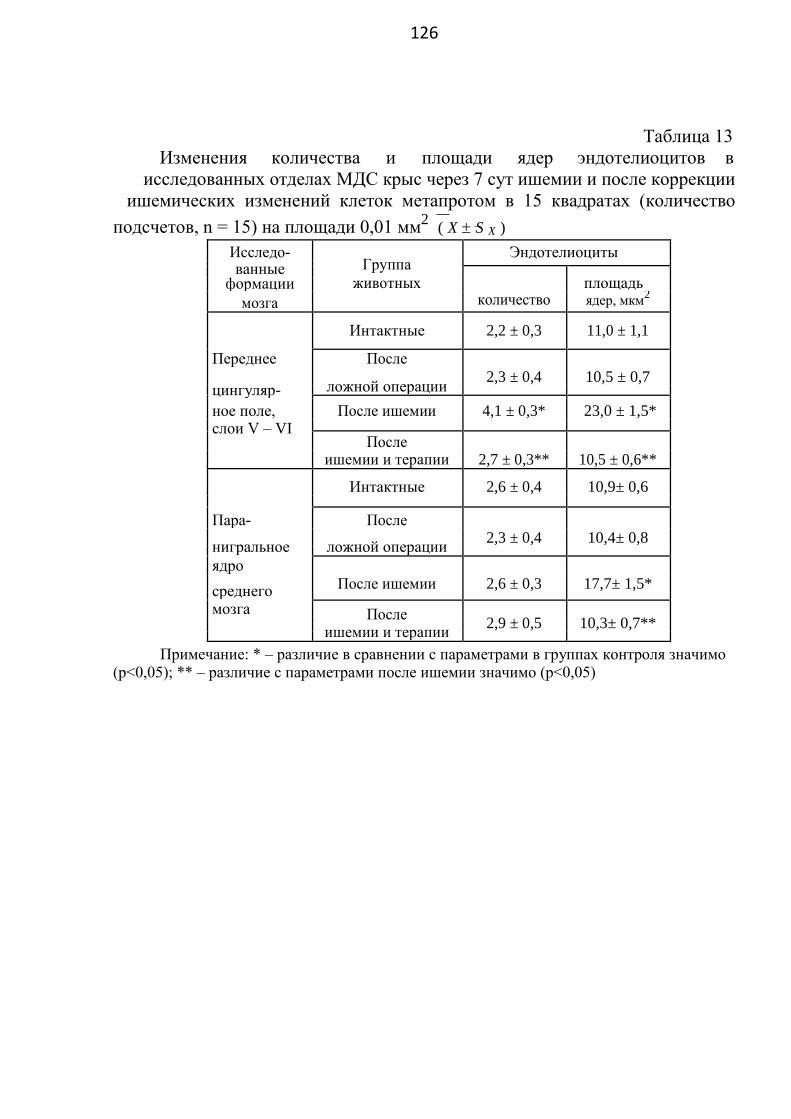
126
Таблица 13
Изменения количества и площади ядер эндотелиоцитов в
исследованных отделах МДС крыс через 7 сут ишемии и после коррекции
ишемических изменений клеток метапротом в 15 квадратах (количество
подсчетов, n = 15) на площади 0,01 мм2 ( X S X )
Исследо- Группа
Эндотелиоциты ванные
площадь животных
формации количество мозга ядер, мкм
2
Интактные 2,2 ± 0,3 11,0 ± 1,1
Переднее После
2,3 ± 0,4 10,5 ± 0,7 цингуляр- ложной операции
ное поле, После ишемии 4,1 ± 0,3* 23,0 ± 1,5* слои V – VI
После 2,7 ± 0,3** 10,5 ± 0,6**
ишемии и терапии
Интактные 2,6 ± 0,4 10,9± 0,6
Пара-
После 2,3 ± 0,4 10,4± 0,8
нигральное ложной операции
ядро После ишемии 2,6 ± 0,3 17,7± 1,5*
среднего
мозга После 2,9 ± 0,5 10,3± 0,7**
ишемии и терапии
Примечание: * – различие в сравнении с параметрами в группах контроля значимо (р<0,05); ** – различие с параметрами после ишемии значимо (р<0,05)

127
Глава 4. ОБСУЖДЕНИЕ ПОЛУЧЕННЫХ РЕЗУЛЬТАТОВ
Полученные данные о строении тел нейронов, в частности, параметры
долей неизмененных, гипохромных, сморщенных гиперхромных нейронов и
клеток-«теней», площади тел нейронов, количества астроцитов, глиоцито-
нейрональный индекс в исследованных частях передней цингулярной коры
(ПЦинК), прилежащего ядра переднего мозга (ПЯ) ипаранигрального ядра
срденего мозга (ПНигЯ) у молодых самцов крыс существенно не различались
с соответствующими парамерами, установленными ранее у интактных
взрослых крыс-самок (Дробленков А.В., 2010; Дробленков А.В. и Шабанов
П.Д.,2014). Основное гендерное различие заключалось в плотности
расположения нейронов глубоких слоев ПЦинК: у самоцов крыс оно
оказалась в 1,7 раза меньше, чем у самок.
Интересными явдяются сведения, подтверждающие более высокие
показатели удельного количества измененных форм нейронов (клеток-
«теней», гипохромных и сморщенных гиперхромных форм) в дофамин-
ергическом ПНигЯ взрослых крыс-самцов (через которое проходят волокна
медиального пучка переднего мозга) по сравнению с проекционными центрами
МДофС (ПЯ и ПЦинК). Это обстоятельство может быть следствием
незавершенности физиологического процесса программированной гибели
нейронов у крыс после рождения и подтверждает наличие «градиента
прочности» мезокортикальной дофаминергической системы, установленного
у здоровых взрослых самок крыс (Дробленков А.В., 2010): он ниже в
дофаминергических ядрах и выше – в проекционных зонах.
Распределение тел нейронов и дифференцированных протоплазматичес-
ких астроцитов по отношению к сосудистой стенке и между собой было
равномерным. Обнаружение значительного количества клеток-сателлитов из
числа клеток макроглии в исследованных частях МДофС подтверждает и

128
дополняет численной характеристикой данные других авторов (Дробленков
А.В. и Карелина Н.Р., 2009; Дробленков А.В., 2011; Васильев Ю.Г. и
Берестов Д.С., 2011; Дробленков А.В. и Шабанов П.Д., 2014). Сведения о
том, что белки промежуточных филаментов в телах, отростках и периваску-
лярных глиальных мембранах отделов МДофС образуют непрерывнную сеть,
также согласуются с результатами исследований этих же авторов.
Высокий парамтр отношения числа клеток-сателлитов из числа
макроглии к числу жизнеспособных нейронов (глиоцито-нейрональный
индекс) и большое количество клеток-сателлитов в МДофС взрослых
здоровых крыс свидетельствуют о хорошо развитом глиальном покрытии
нервных клеток. Это, в свою очередь, может являться объяснением
невысокой способности нейронов ПНигЯ, ПЯ и ПЦинК образовывать пары
(от 3,7 до 9,4% тел нейронов в этих центрах были сгруппированы).
Способность тел нейронов образовывать мембранные перфорации и
синцитиальные связи в норме, в ходе онтогенетического развития и при
патологии связана именно с выраженностью глиального покрытия в
пирамидном слое СА-1 и СА-2 частей гиппокампа, в зернистом слое коры
мозжечка (Парамонова Н.М. и Сотников О.С., 2008; Сотников О.С. и
Парамонова Н.М., 2010; Сотников О.С., Парамонова Н.М. и Арчакова Л.И.,
2010; Сотников О.С., Фрумкина Л.Е. и Майоров В.Н., 2012) и медильном
аркутном ядре гипоталамуса (Ходулева Ю.Н. и соавт., 2015). По данным
авторов, изучавших ультраструктуру контактов спаренных нейронов
(Сотников О.С. и Парамонова Н.М., 2010; Сотников О.С., Фрумкина Л.Е. и
Майоров В.Н., 2012), между телами клеток образуются перфорации
плазматическтих мембран и синцитиальная связь, со всеми ее
ультраструктурными признаками.
Плотность расположения нейронов в глубоких слоях ПЦинК много
ниже, чем в обозначенных отделах гиппокампа и зернистом слое коры
мозжечка, но в ПЦинК и ПЯ значительно выше, чем в ПНигЯ (р<0,05); см.
табл. 3). Вместе с тем плотность расположения астроцитов, олигодендро-

129
цитов и значение глиоцито-нейронального индекса в исследованных частях
МДофС значительно не различаются (см. рис. 6, 8, 10; табл. 3 и 4).
Следовательно, данные настоящего исследования подтверждают концепцию,
сформулированную перечисленными авторами о вероятном
происпособительном значении объединения нейронов в кластеры и связи
числа таких кластеров нейронов с количеством клеток их глиального
окружения, дополнив эту концепцию численными параметрами в структурах
МДофС.
4.1. Изменения клеток передней цингулрной коры и прилежащего ядра, расположенных в области выраженной ишемии
Выявленные выраженные реактивные изменения тел нейронов в
ПЦинК и ПЯ через 7 суток ишмии, безусловно, являются признаками
отсроченных проявлениий глютамат-кальциевого каскада, наблюдавшихся у
нейронов в фокусе ишемии (Гусев Е. И. и Скворцова В. И., 2001). Здесь,
вследствие острой ишемии, по мнению авторов монографии, развиваются и
нарастают ваимообусловленные молекулярные реакции, приводящие клетки
к гибели. В частности, речь идет об активации кальмодулин-зависимых
клеточных ферментов, оксидантном стрессе, избыточной выработке оксида
азота, самого по себе токсичного для клеток, и накопление
низкомолекулярныхцитотоксичныхсоединений.Отек-набухание,
гиперхромию и сморщивание тел нейронов во многом объясняют
ультраструктурные изменения нервных и макроглиальных клеток,
вывленные через несколько часов ишемии, индуцированной инъекцией
хлорида кобальта (Caltana L. еt al., 2009). Авторы имеют в виду процесс
набухания и расширения просвета канальцев эндоплазматической сети,
пластинчатого комплекса, матрикса митохондрий и, в целом, отек
цитоплазмы, начинающийся в перинуклеарной части клеток, что
сопровождалось сморщиванием оболочки ядра и клетки.

130
Обстоятельство, что как в фокусе частичной ишемии, так и во внешней
части зоны пенумбры большую долю нейронов составляют сморщенные
гиперхромные формы, может быть следствием вступления клеток данного
фенотипа в процесс гибели путем апоптоза. Именно такому способу
умирания клетки в самом начале характерно ее сжатие и сгущение органелл,
выявляемому часто в зоне пенумбры (Chopp M.Т. and Li Y., 1996), которое на
светооптическом уровне проявляется гиперхромией цитоплазмы.
Возможно, присутствие двух фенотипов поврежденных клеток –
светлых набухших и темных сморщенных объяснет начало детерминации
двух различных путей их умирания: некроза и апоптоза. Поскольку контуры
поверхности ядра и тела нейронов на светооптическом уровне дефектов не
обнаруживают, а ядрышко остается отчетливо выявляемым, данные измене-
ния представляются обратимыми. Вероятно, гиперхромия-сморщивание
нейронов развивается в исходе ишемического отека и предшествует
образованию клетки-«тени».
Что касается состояния астроцитов в фокусе частичной ишемии,
данные клетки были гораздо менее повреждены, чем нейроны. Во-первых,
среди астроцитов не было клеток-«теней» или сморщенных форм, во-вторых
каждая астроцитарная клетка экспрессировла промежуточный филамент
(хотя и в мньшей степени, чем в контролях). Более выраженная устойчивость
астроцитов, чем нейронов, в условиях ишемии-гипоксии, на первый взгляд,
является непонятной, если учесть особеннсоти ее положения в системе
«нейрон-астроцит-капилляр». Однако, с другой стороны, эта же особенность
положения астроцитов, вероятно, формирует более выгодный тип питания
клеток этого типа в условиях ишемии, при котором астроцит забирает
значительную чать важных нутриентов себе.
Значительная устойчивость астроцитов в условиях выраженной
ишемии также можно объяснить и с позиций увеличения интенсивности
взаимоотношений этих клеток со стенкой капилляров и телами нейронов.

131
Вполне вероятно, что выявленное усиление обмена жизненно важными
молекулами и ростовыми факторами при неблагоприятном воздействии
происходит не только в направлении от глиальной клетки к нейрону (Певзнер
Л.З., 1972; Hyden H., 1967; Dai Х., Larcher L.D., Clinton P.V. et al.,2003;
Wilkins А., Majed H., Layfield R. et al., 2003), но и в обратном направлениии
или между люмыми клетками в этой коопрации. Данное умозаключение
подтверждает тот выявленный факт, что большинство тел астроцитов ПЦинК
и ПЯ располагались периваскулярно и одновременно являлись клетками-
сателлитами. Расстояние же между телами астроцитов и стенкой капилляра
значительно сократилось, причем количество астроцитов спустя неделю
ишемического воздействия значительно не изменилось.
То обстоятельство, что выраженное нарушение гемодинамики в ПЦинК
и ПЯ вызывало гибель лишь некоторой части клеток, тогда как значительная
часть нейронов этих формаций оказывалась в состоянии, близком к
обратимому повреждению, и даже определялись единичные малоизмененные
клетки, может быть связано с компенсационным влиянием на выживающие
клетки переднего мозга нейронов среднего мозга. Так, ПНигЯ, ПЯ и
глубокие слои ПЦинК являются важнейшими взаимосвязанными частями
МДофС: их объединяют между собой дофаминергические волокна
медиального пучка переднего мозга (Bjorklund A. and Lindvall O., 1984).
Следовательно, значительная часть аксонов нейронов дофаминергических
ядер среднего мозга, расположенных в области слабо выраженной ишемии,
являются транспортерами для метаболитов, необходимых для выживания
клеток ПЯ и ПЦинК, расположенных в фокусе выраженной ишемии.
Реактивные изменения астроцитов в ПЯ и глубоких слоях ПЦинК в
условиях данного эксперимента принципиально не различались: клетки
находились в состоянии цитотоксического отека, признаки которого у
астроцитов наблюдали также через 6–7 суток в полосатом теле при
пролонгированной гипоксии на стороне окклюзии основания средней

132
мозговой артерии (Garcia J. H. еt al., 1993). Авторы полагают, что эти
изменения происходят в зоне, прилегающей к ядру инфаркта мозга
(супраоптическая область в их модели), так называемой области
ишемической «полутени» (зона пенумбры), то есть области частичной
ишемии. Грубые нарушения экспрессии GFAP в телах, отростках и
периваскулярных глиальных мембранах, установленные при использовании
нашей модели ишемии как в ПЯ, так и в ПЦинК свидетельствуют о том, что
эти клетки находятся именно в ядре выраженной ишемии и испытывают
альтернативное воздействие концентрирующихся здесь токсичных молекул
глютамат-кальциевого каскада (Гусев Е.И. и Скворцова В.И., 2001).
Воздействие токсичных молекул, образующихся в ходе немедленных и
отсроченных реакций этого каскада на нейроны и атроциты, усугубляется
признаками повреждения их промжуточного филамента в основном
компоненте гемато-энцефалического барьера – перикапиллярной глиальной
астроцитарной мембране. Они, по данным настоящего исследования,
выражаются истончением, фрагментацией и могли произойти в нашей
модели ишемии прежде, чем GFAP стал частично восстановленным. Для
уточнения механизмов процесса перемещения клеток макроглии в мозге у
взрослых особей, уже ставшего очевидным в отношении олигодендроцитов
(Богомолов Д. В. и соавт., 2001; Дробленков А.В. и Шабанов П.Д., 2014), но
малоизвестного в отношении астроцитов, установленного при ишемии мозга
и терапии полипренолами (Дробленков А.В., Монид М.В. И Валькович Э.И.,
2014), а также при хронической алкогольной интоксикации (Дробленков А.В.
и Шабанов П.Д., 2014) требуются дальнейшие исследования с большим
количеством промежуточных сроков эксперимента.
Защитно-приспособительный процесс направленного перемещения тел
нейронов и макроглиоцитов, считающихся оседлыми клетками, малоизучен,
однако наши данные подтверждают способность астроцитов к изменению
положения их тел после частичного разрушения GFAP. Косвенно

133
подтвердить способность к перемещению тел астроцитов в головном мозге
взрослых организмов при определенных условиях может результат
эксперимента Okoye G.S., Powel E.M. and Geller H.M. (1995). После введения
суспензии астроцитов в стриатум взрослого мозга авторы наблюдали
миграцию этих клеток в направлении сосудистой стенки со скоростью 31
мкм в день.
Объединение тел поврежденных нейронов между собой и стенкой
капилляров в условиях ишемии наблюдали также при электронной
микроскопии (Гелеранская О.А. и соавт., 2016). Авторы описывали простые и
плотные контакты между телами нейронов, образование между ними
трансмембранных перфораций, прямые нейровазальные контакты; они не
исключали защитно-приспособительной роли подобных коммуникативных
процессов.
Тенденцию к образованию кластеров клеток различного типа
(нейронов, клеток макроглии, эндотелиоцитов капилляров, возможно также и
активированных клеток амебоидной формы глии) на молекулярном уровне
можно объяснить увеличением экспрессии и активации в их плазмолемме
молекул межклеточной адгезии. Так, активацию последних выявили при
ишемии в плазмолемме эндотелиоцитов (Siren A.L., McCarron R.M., Lu Y. et
al.) и в плазмолемме нейронов гиппокампа, что морфологически сочеталось
со сближением поверхности тел пирамидных клеток (Li Y. and Liu S., 2017).
Увеличение числа периваскулярных форм астроцитов, олигодендро-
цитов и нейроцитов в сочетании с увеличением числа сателлитных клеток
может иметь компенсаторно-адаптивное значение для всех клеток этого
комплекса и, в частности, нейронов, поскольку во многих исследованиях
приводятся убелительные доказательства передачи РНК, аминокислот и
факторов роста клетками макроглии в тела нейронов при функциональном
истощении последних (Певзнер Л.З., 1972; Богомолов Д.В. и соавт., 2001; Dai
X. at al., 2003).

134
Увеличение числа сателлитных клеток и рост индекса глиоцит/нейрон,
наблюдаемого в условиях выраженной пролонгированной ишемии, следует
рассматривать как чувствительную компенсаторную неспецифическую
глиальную реакцию, также выявленную при абстиненции после поглощения
алкоголя и опиатов (Богомолов Д.В. и соавт., 2001; Дробленков А.В., 2011;
Васильев Ю.Г. и Берестов Д.С., 2011) и наркотической гипоксии (Богомолов
Д.В. и соавт., 2001). Увеличение индекса глиоцит/нейрон в 9 раз может
указывать на выраженное увеличение передачи глиальных питательных
веществ в тела нервных клеток.
Декструктивные изменения нейронов и астроцитов в фокусе
выраженной ишемии может быть обусловлено также влиянием клеток
активированной микроглии. На это обстоятельство указывает то факт, что в
обеих структурах МДофС, расположенных области выраженной ишемии,
микроглиоциты обладали признаками активированных амебоидных клеток.
Такое положение подтверждается новыми сведениями о токсичности
активированных микроглиоцитов (Рыбакина E.Г. и Корнева E.А., 2005; Kim
S.U. and Vellis J., 2005; Smith J.A., Das A., Ray S.K. and Banik N.L., 2011).
Активированная форма клеток микроглии, по данным авторов, синтезирует
потенциально нейротоксичные вещества, включая NO, активные формы
кислорода, протеазы, возбуждающие аминокислоты, провоспалительные
цитокины и хемокины.
Сосудистая реакция через 7 сут в области выраженной ишемии
выражалась увеличением длины, петлистым ходом ноообразованных частей
кровеносных капилляров, увеличением числа эндотелиоцитов и отеком-
набуханием ядер. Обнаружение начальных признаков гиперплазии
эндотелиоцитов к концу 7-х суток опыта не может быть неожиданным,
поскольку в ядре полной ишемии эти явления наблюдали через 3–4 суток
(Васильев Ю. Г. и Берестов Д. С., 2011). Перечисленные изменения стенки
капилляров являются ответом на воздействие токсичных продуктов
глютамат-кальциевого каскада (Гусев Е. И. и Скворцова В. И., 2001).

135
4.2. Изменения клеток паранигрального ядра, расположенных в области слабо выраженной ишемии
Полученные данные свидетельствуют о том, что в фокусе
ишемии,который в нашей модели включает ПЦинК и ПЯ, деструктивные
изменения нейронов и астроцитов выражены больше, чем за ее пределами
(ПНигЯ). Эти изменения, конечно же, являются следствием распространения
токсических продуктов глютамат-кальциевого каскада (Гусев Е. И. и
Скворцова В. И., 2001). Допустим, что в ПЦинК и ПЯ создается область
повышенной концентрации кальмодулин-зависимых клеточных ферментов,
оксида азота и других низкомолекулярных цитотоксинов, которая
соответствует зоне пенумбры, близкой к фокусу полной ишемии. Клеточные
элементы в ней могут испытывать влияние токсикантов не только через
пространства между клетками, но и через гемато-энцефалический барьер
(систему «нейрон– атроцит–капилляр–нейрон»). Такой двойной механизм
действия токсикантов в фокусе частичной ишемии подтверждается
вывленной деструкцией GFAP в периваскулярных глиальных мембранах и
признаками токсического отека астроцитов.
В ПНигЯ, испытывающем в течение 7 суток ишемию, выраженную
гораздо в меньшей степени, деструктивным изменениям подвергаются лишь
нейроны, причем эти изменения клеток выражены значительно слабее, чем в
структурах переднего мозга.
Феномен меньшей выраженности постишемических изменений
нейронов ПНигЯ, чем в ПЦинК и ПЯ, можно объаснить следующими
механизмами влиния альте-рирующих факторов, запускаемых глютамат-
кальциевым каскадом в очаге ишемии. 1) Образовавшись в переднем мозге,
альтеранты достигают нейронов ПНигЯ через нейропиль, 2) либо по свзям
отростков нервных клеток ПЦинК и ПЯ с нейронами ПНигЯ, выявленными
благодаря изучению пути перемещения изотопов (Bjorklund A., Lindvall O.,
1984). При этом на пути распространения их концентраци снижается. 3)
Токсиканты могут образовываться локально (в ПНигЯ) в малом количестве.

136
Передача их соседним нейронам через гемато-энцефалический барьер
представляется маловеротной, поскольку астроцитарное звено этого барьера
укреплено благодаря защитному усилению выработки клетками GFAP.
Выявленные изменения астроцитов ПНигЯ гипертрофического типа,
такие как увеличение площади тел, увеличение количества GFAP-материала
в телах, отростках и перикапиллярных мембранах, по-видимому, аналогичны
тем,которые происходят в зоне пенумбры после острой фокальной ишемии и
связаны с накоплением в клетках большого количества включений гликогена
(Kajihara H. at al., 2001).
Можно предположить, что астроциты в ПНигЯ приобрели перикапил-
лярное положение в начале ишемического воздействия, поскольку через 7
суток многие из них уже представляли собой периваскулярную форму
клеток, отростки которых были укорочены, окружали стенку капилляров,
обладали признаками гипертрофии и увеличенной экспрессией GFAP.
Также логично предположить, что миграция астроцитов и связанных с
ними нейронов ПЦинК и ПЯ, находящихся в фокусе частичной ишемии,
приобрели периваскулярную локализацию в более ранние сроки опыта,
поскольку на 7-е сутки астроциты уже обладали выраженными признаками
дистрофии.
Таким образом, клеточные элементы головного мозга при переживании
разных условий циркуляторной гипоксии характеризуются спектром
выраженных патологических изменений. Нейроны находятся в процессе
клеточной гибели, который, вероятно, для многих клеток из-за частичности
ишемии стал пролонгированным во времени, в результате чего нейроны
приобрели фенотип сморщивания-гиперхромии, лизиса и превращения в
клетки-«тени». Клетки центральной зоны ишемии склонны к гибели или
сморщиванию-гиперхромии. Нейроны, расположенные за пределами области
выраженной гипоксии и испытывающие воздействие токсикантов в малой
концентрации, склонны к острому набуханию.

137
Реакция микроглиоцитов в виде слабо выраженного увеличения их
количества и признаков активации является ранним диффузным проявлением
глобальной гипоксии.
Пролиферация эндотелиоцитов на ранних сроках ишемического
возвоздействия наблюдается лишь в фокусе гипоксии.
Концентрация жизнеспособных нейронов и астроцитов вблизи стенки
кровеносных капилляров, а также рост числа сателлитных форм глии
являются адаптационными механизмами и условием выживания клеток при
разных условиях ишемического воздействия на головной мозг.
4.3. Фармакологическая коррекция метапротом постишемических изменений клеток исследованных отделов МДС
Снижение числа погибших нейронов, поддержание структуры многих
нейронов и астроцитов на уровне, близком этим клеткам, у здоровых
животных являются основными морфологическими коррелятами
компенсаторно-приспособительного влияния метаболического протектора на
головной мозг при моделировании его глобальной ассиметричной ишемии.
Существенное коррекционное влияние метапрота в дозе 12,5 мг/кг на
разные типы клеток мозга при глобальной ишемии, сдерживающее процесс
их альтеративных изменений, может быть объснено следующими
биохимичес-кими механизмами действия препарата.
1). Препарат усиливает синтез РНК и белка, поскольку выявлены
включение 75Se-метионина в клетки различных органов, а также активация
препаратом действия экзогенно вводимых РНК, угнетенных
предварительным введением избирательного ингибитора синтеза РНК
актиномицина Д (Зарубина И. В. и Шабанов П. Д., 2004).
2). Метапрот интенсифицирует процессы глюконеогенеза из недоокислен-
ных продуктов распада (лактата, пирувата) и аминокислот, что вывлено при
изучении биохимии коркового вещества почек и печени (Зарубина И. В. И
Шабанов П. Д. 2004; Шабанов П. Д., 2010; Шабанов П. Д. и соавт., 2010).

138
3). Данное вещество стимулирует митохондриальное окисление при
экстремальных и повреждающих воздействиях на клетку, в том числе при
гипоксии и ишемии. Сохранение высокого уровня синтеза АТФ при
дефиците кислорода способствует выраженной антигипоксической и
противоишемической активности метапрота (Зарубина И. В. и Шабанов П. Д.,
2004; Шабанов П. Д. и соавт., 2010). Влияние на митохондрии заключается в
ослаблении торможения НАД-зависимого дыхания и активности сукцинат-
дегидрогеназы, в уменьшении разобщения окисления с фосфорилированием,
предотвращении глубокого низкоэнергетического сдвига. Этот эффект также
связан с оптимизирующим действием метапрота на функции митохондрий и
активацией синтеза митохондриальных белков (Зарубина И. В. и Шабанов П.
Д. 2004; Шабанов П. Д. и соавт., 2005).
4). Метапрот оказывает экономизирующее и восстановительное
влияние на углеводный и энергетический обмен. При стандартной
физической нагрузке под его влиянием наблюдается меньшее снижение
содержания гликогена, АТФ и креатинфосфата в организме, меньший
прирост теплопродукции и потребления кислорода, чем при нагрузке без
влини препарата (Шабанов П. Д. и соавт., 2005; Зарубина И. В. и соавт., 2006;
Евсеева М. А. и соавт., 2008).
Функциональное влияние метапрота изучено на организменном уровне
(Шабанов П.Д. и Ганапольский В.П., 2009; Шабанов П.Д., 2010; Шабанов П.
Д., 2011). Препарат существенно повышает физическую работоспособность
животных и человека и ускоряет ее восстановление после предельных
нагрузок. Этот эффект особенно ярко проявился в осложненных условиях
(при высотной гипоксии, перегревании и др.).
Таким образом, применение метапрота в постишемическом периоде
значительно увеличивает долю малоизмененных нейронов, соответственно
сокращает удельное количество клеток-«теней» и других измененных
нейронов как в области ограниченного кровотока, так и за ее пределами.

139
Коррегирующее действие метапрота на астроциты в постишемический
период как в фокусе ишемии, так и в ядре среднего мозга выражается
сохранением GFAP-материала в телах, главных отростках и периваскулярных
мембранах.
Терапия метапротом в постишемическом периоде усиливает нейроно-
глиальные взаимоотношения, подавляет изменение положения тел
астроцитов и нейронов относительно сосудистой стенки.
Действие метапрота на нейроны и макроглию в постгипоксическом
периоде направлено на усиление компенсаторно-приспособительных реакций
этих клеток. Его эффекты можно объяснить такими свойствами препарата,
как усиление энергетического обмена (увеличение концентрации АТФ,
креатинфосфата), увеличение белкового синтеза и углеводного обмена.

140
ВЫВОДЫ
1. В основных отделах мезокортиколимбической дофаминергической
системы интактных молодых самцов крыс содержится некоторое количество
измененных нейронов: клеток-«теней», гипохромных и сморщенных
гиперхромных форм, что может быть следствием незавершенности
физиологического процесса программированной гибели нейронов у крыс
после рождения. Глиальный фибриллярный кислый белок астроцитов
формирует непрерывный каркас в их телах, отростках и периваскулярных
мембранах. Тела клеток разных типов (нейронов, макроглиоцитов, в том
числе астроцитов) распределены равномерно по отношению к стенке
капилляров и друг к другу: среднее расстояние между ними и стенкой
капилляров превышает 5–9 мкм, глиоцито-нейрональный индекс – не более
0,14. Покоящиеся микроглиоциты представлены единичными периваскуляр-
ными формами клеток, площадь тел которых не превышает 29,9±2,2 мкм2.
Количество эндотелиоцитов в единице площади исследованных отделов
мозга постоянно и не превышает 2,7±0,6 клеток.
2. Выраженное ограничение кровотока через 7 суток индуцирует
гибель большей части нейронов по темному и светлому типу (состояние,
близкое к тому, которое устанавливается во внутренней части zona penumbra
при формировании ишемического инфаркта мозга). Астроциты более устойчивы
к процессу ишемического повреждения, чем нейроны. Они испытывают отек-
набухание, деструкцию промежуточных филаментов в части площади их тел,
периферических отделах отростков и периваскулярных глиальных мембранах.
Наблюдаются признаки капиллярной реакции.
3. При моделировании выраженного ограничения кровотока в головном
мозге изменяются пространственные взаимоотношения клеток. Тела повреж-
денных нейронов объединяются между собой, клетками макроглии и со
стенкой кровеносных капилляров. Формирование нейроно-глио-васкулярных
комплексов представляет собой защитный механизм и условие выживания
клеток в фокусе частичной ишемии.

141
4. В области слабо выраженной ишемии нейроны более склонны к
отеку-набуханию, нежели к гибели; объединение тел нейронов в группы не
происходит. Астроциты, представленные, главным образом, околососудистой
формой клеток, испытывают изменения гипертрофического типа
(аналогичные происходящим на внешней границе zona penumbra). Признаки
капиллярной реакции отсутствуют.
5. Микроглиоциты при моделировании разных условий глобальной
ишемии мозга отличаются от клеток в контролях признаками трансформации
в активированную амебоидную форму. Параметры изменений клеток
микроглии в исследуемых структурах переднего и среднего мозга
значительно не различаются.
6. Выраженный нейроно(гисто)протекторный эффект актопротектора и
антигипоксанта метапрота направлен на защиту структуры атроцитов и их
промежуточных филаментов, в том числе в периваскулярных глиальных
мембранах, сохранение нейронами структурных особенностей малоизменен-
ных, поддержание нейроно-глиальных взаимоотношений на высоком уровне.

142
ПРАКТИЧЕСКИЕ РЕКОМЕНДАЦИИ
Особенности реактивных изменений клеток разных типов, выявленные
в исследованных отделах МДофС при часто используемом моделировании
глобальной ишемии мозга методом окклюзии обеих общих сонных артерий,
могут быть использованы в учебном процессе на кафедрах гистологии и
эмбриологии, патололгической анатомии, судебной медицины для обучения
основам клеточных реакций, устанавливающихся в головном мозге после
частичной выраженной и частичной слабо выраженной ишемии.
Реактивные изменения клеток разных типов в исследованных отделах
МДС, выявленные после частичной выраженной и частичной слабо
выраженной ишемии, могут являться основой для сравнения с результатами
повторных гистологиеских исследований мозга в данной модели ишемии, а
также для установления эффективности коррекции процесса ишемии мозга
различными фармакологическими препаратами. Результаты настощей работы
убеждают в том, что использование при моделировании глобальной ишемии
мозга гистологических методик значительно повышает информативность
данных, полученных другими традиционными методами, при оценке
эффективности фармакологического влияния на ишемизированный мозг.

143
СПИСОК ЛИТЕРАТУРЫ
1. Айвазян, С. А. Прикладная статистика: Основы моделирования и
первичная обработка данных / С. А. Айвазян, И. С. Енюков, Л. Д. Мешалкин.
– М. : Финансы и статистика, 1983. – 471 с.
2. Андреева, Л. И. Центральные эффекты белка теплового шока с
молекулярной массой 70 кДа / Л. И. Андреева, Б. А. Маргулис, И. В. Гужова
// Психофармакол. и биол. наркол. – 2005. – Т. 5. – № 1. – С. 794–803.
3. Анохина, И. П. Диагностика генетической предрасположенности к
зависимости от психоактивных веществ / И. П. Анохина, Ю. Л. Арзуманов,
А. Г. Веретинская и др. // Проблемы диагностики и лечения алкоголизма и
наркоманий. – М. : Анахарсис, 2001. – С. 6–29.
4. Ашмарин, И. П. Нейрохимия / И. П. Ашмарин, П. В. Стукалов. – М. :
Изд-во Ин-та био-мед. химии РАМН, 1996.
5. Бадалян, Л. О. Неврологические синдромы при болезнях сердца / Л.
О. Бадалян. – М. : Медицина, 1975. – 416 с.
6. Боголепов, Н. К. Церебральные кризы и инсульты / Н. К. Боголепов.
– М. : Медицина, 1971. – 392 с.
7. Боголепов, Н. Н. Ультраструктура синапсов в норме и патологии /
Н. Н. Боголепов. – М. : Медицина, 1975. – 214 с.
8. Боголепов, И. И., Бурд, Г. С. Материалы VII Всесоюз. съезда
невропатол. и психиатров / И. И. Боголепов, Г. С. Бурд. – М., 1981. – Т. 2. –
С. 32–35.
9. Богомолов, Д. В. Судебно-медицинская диагностика наркотической
интоксикации по морфологическим данным : автореф. …. д-ра мед. наук / Д.
В. Богомолов. – М., 2001. – 29 с.
10. Богомолов, Д. В. Морфометрическое исследование нейроглиальных
комплексов головного мозга при судебно-медицинской диагностике
наркоманий / Д. В. Богомолов, Ю. И. Пиголкин, О. В. Должанский // Суд.-
мед. эксп. – 2001. – Т. 44. – № 4. – С. 18–19.

144
11. Васильев, Ю. Г. Гомеостаз и пластичность мозга / Ю. Г. Васильев,
Д. С. Берестов. – Ижевск : ФГБОУ ВПО Ижевская ГСХА, 2011. – 216 с.
12. Верещагин, Н. В. Патология вертебробазилярной системы и
нарушения мозгового кровообращения / Н. В. Верещагин. – М. : Медицина,
1980. – 310 с.
13. Гелеранская О. А. Влияние физической нагрузки на
ультраструктурные изменения в коре головного мозга крыс / О. А.
Гелеранская, Н. В. Гончаров, Н. М. Маслова, Н. М. Парамонова, Т. В.
Тавровская // Морфология. – 2016. – Т. 149. – Вып. 3. – С. 62.
14. Голубев, А. М. Апоптоз при критических состояниях / А. М.
Голубев, Е. Ю. Москалева, С. Е. Северин // Общая реаниматология. – 2006. –
Т. 2. – № 5 –6. – С. 184–190.
15. Гусев, Е. И. Ишемия головного мозга / И. И. Гусев, В. И. Скворцова.
– М. : Медицина, 2001. – 328 с.
16. Гусев, Е. И. Сосудистые заболевания головного мозга / Е. И. Гусев,
Г. С. Бурд, Н. Н. Боголепов. – М. : Медицина, 1979. – 142 с.
17. Дамбинова, С. А. Молекулярные механизмы передачи импульса в
мембранах нейронов. Ионные каналы, рецепторы / С. А. Дамбинова, М. А.
Каменская // Нейрохимия. (Ред. Ашмарин И. П., Стукалов П. В.). – М. :
Медицина, 1996. – С. 246–295.
18. Дегенеративные изменения нейронов медиального аркуатного
гипоталамического ядра в модели мужского гипогонадизма / Ю. Н.
Ходулева, З. П. Асауленко, А. А. Байрамов, И. Л. Никитина, А. В.
Дробленков // Педиатр. – 2015. – Т. 6. – № 3. – С. 62–68.
19. Должанский, О. В. Судебно-медицинская оценка морфологических
изменений головного мозга при хронических опийных наркоманиях :
автореф. дис. … канд. мед. наук / О. В. Должанский. – М., 2001. – 26 с.
20. Дробленков, А. В. Краткий микроскопический атлас ядерных и
корковых центров мезокортиколимбической и некоторых других
дофаминергических систем головного мозга крысы / Под ред. проф.

145
Карелиной Н. Р. / А. В. Дробленков. – СПб. : СПбГПМА, 2010. – С. 16, 33–35.
21. Дробленков, А. В. Патологические изменения нейронов
мезокортиколимбической дофаминергической системы у здоровых людей и
крыс / А. В. Дробленков // Морфология. – 2010. – Т. 137. – Вып. 3. – С . 11–17.
22. Дробленков, А. В. Морфологические признаки отравления этанолом,
алкогольной абстиненции и хронической алкогольной интоксикации в
мезокортиколимбической дофминергической системе / А. В. Дробленков //
Суд.-мед. экспертиза. – 2011. – Т. 54. – № 5. – С. 11–17.
23. Дробленков, А. В. Структурные особенности нейронов и
макроглиоцитов взаимосвязанных отделов мезоаккумбоцингулярной
дофаминергической системы крыс / А. В. Дробленков, Н. Р. Карелина //
Морфология. – 2009. – Т. 136. – № 5. – С. 11–17.
24. Дробленков, А. В. Изменения нейронов и глиоцитов
мезоаккумбоцингулярной системы при перинатальном воздействии морфина
у крыс / А. В. Дробленков, Н. Р. Карелина, П. Д. Шабанов // Морфология. –
2009. – Т. 136. – № 6. – С. 35–37.
25. Дробленков, А. В. Дофаминергические механизмы алкогольной
зависимости / А. В. Дробленков, П. Д. Шабанов. – СПб. : Art-Xpress, 2014. –
256 с.
26. Евсеева, М. А., Евсеев, А. В., Правдивцев В. А., Шабанов П. Д.
Механизмы развития острой гипоксии и пути ее фармакологической
коррекции // Обзоры клин. фармакол. лекарств. терап. – 2008. – Т. 6. – № 1. –
С. 3–25.
27. Жаботинский, Ю.М. Нормальная и патологическая морфология
нейрона / Ю. М. Жаботинский. – Л. : Медицина,1965. – 323 с.
28. Завалишин, И. А. Оксидантный стресс: общий механизм
повреждения при заболеваниях нервной системы / И. А. Завалишин, М. Н.
Захарова // Журн. невропатол. и психиатр. – 1996. – Т. 2. – С. 111–114.
29. Зарубина, И. В. Молекулярная фармакология антигипоксантов / И. В.
Зарубина, П. Д. Шабанов. – СПб. : Элби-СПб, 2004. – 368 с.

146
30. Зарубина, И. В. Антигипоксанты при черепно-мозговой травме /
И. В. Зарубина, П. Д. Шабанов. – СПб. : Элби-СПб., 2006. – 208 с.
31. Зарубина, И. В. Эффективность бемитила и пиразидола у больных с
церебрастенией вследствие черепно-мозговой травмы / И. В. Зарубина, Ф. Н.
Нурманбетова, П. Д. Шабанов // Клин. мед. – 2005. – Т. 83. – № 11. – С. 59–62.
32. Лакин, Г. Ф. Биометрия : учеб. пособие для биол. спец. вузов. 4-е
изд., перераб. и доп. / Г. Ф. Лакин. – М. : Высш. шк., 1990. – 352 с.
33. Лесиовская, Е. Е. Антигипоксанты прямого действия –
перспективные нейропротекторы / Е. Е. Лесиовская // TERRA MEDICA
34. Литвинцев, Б.С., Одинак, М.М., Гайкова, О.Н., Онищенко, Л.С.,
Трофимова, А.В. Клинико-морфологическая характеристика неврологических
проявлений наркомании / Б. С. Литвинцев, М. М. Одинак, О. Н. Гайкова, Л. С.
Онищенко, А. В. Трофимова. // Профилактич. и клин. медицина. – 2011. – Т.
39. – № 2. – С. 99–104.
35. Лукьянова, Л.Д. Биоэнергетическая гипоксия: понятие, механизмы и
способы коррекции / Л.Д. Лукьянова // Бюллетень экспериментальной
биологии и медицины. – 1997. – Т. 124. – № 9. – С. 244–254.
36. Моргун, А.В. Современные представления о патогенезе
перинатального ишемического повреждения клеток нейроваскулярной
единицы головного мозга: молекулы-мишени для нейропротекции / Моргун
А.В., Кувачева Н.В., Таранушенко Т.Е. и др. // Вестник Рос. акад. мед. наук –
2013. – № 12. – С. 26–35.
37. Морозова, О.Г. Применение нейропротекторов для лечения
осложнений сахарного диабета / О. Г. Морозова // Здоровье Украины. – 2003.
– № 4. – С. 10–11.
38. Новиков, В.Е. Аминотиоловые антигипоксанты при травматическом
отеке мозга / В.Е. Новиков, Н.И. Понамарева, П.Д. Шабанов. – Смоленск-
СПб. : Элби-СПб., 2008. – 176 с.

147
39. Отеллин, В. А. Структурное развитие головного мозга и
формирование его патологии в пренатальном онтогенезе млекопитающих /
В. А. Отеллин // Мозг. Теоретические и клинические аспекты / Гл. ред.
И. В. Покровский. – М. : Медицина, 2003. – С. 117–138.
40. Парамонова, Н. М. Цитоплазматическая синцитиальная связь между
телами нейронов ЦНС взрослых животных / Н. М. Парамонова, О. С.
Сотников // Морфология. – 2008. – Т. 134. – № 6. – С. 13–18.
41. Парамонова, Н. М. Межнейрональные мембранные контакты и
синцитиальные перфорации в поле СА2 гиппокампа при травматическом
повреждении мозга / Н. М. Парамонова, О. С. Сотников, Т. В. Краснова //
Бюлл. эксп. биол. мед. – 2010. – Т. 150. – № 7. – С. 113–116.
42. Пашченко, П. С., Сухотерин, А. Ф. Структурная организация области
postrema здоровых крыс и в условиях хронического воздействия перегрузок /
П. С. Пашченко, А. Ф. Сухотерин // Морфология. – 2000. –117(2) – С. 36–41.
43. Певзнер, Л. З. Функциональная биохимия нейроглии / Л. З. Певзнер.
– Л. : Наука, 1972. – 198 с.
44. Поляков, Г. И. Основы систематики нейронов коры головного мозга
человека / Г. И. Поляков. – М. : Медицина, 1973. – 216 с.
45. Пошивалов, В. П. Этологический атлас для фармакологических
исследований на лабораторных грызунах / В. П. Пошивалов. – М., 1978. Деп.
в ВИНИТИ, № 3164 – 3178.
46. Раевский, К. С. Оксид азота: избыточный синтез в механизмах
острой ишемии мозга / К. С. Раевский // Бюлл. экспер. биол. и мед. – 1997. –
Т. 123. – № 4. – С. 484–490.
47. Рыбакина, E. Г. Трансдукция сигнала интерлейкина-1 в процессах
взаимодействия нервной и иммунной систем организма / Е. Г. Рыбакина, Е.
А. Корнева // Вестник Рос. акад. мед. наук. – 2005. – № 7. – С. 3–8.
48. Скворцова, В. И. Клинический и нейрофизиологический мониторинг,
метаболическая терапия в остром периоде церебрального ишемического

148
инсульта : дис. … д-ра мед. наук / В. И. Скворцова. – М., 1993. – 379 с.
49. Сотников, О. С. Цитоплазматическая синцитиальная связь – одна из
трех форм межнейрональной связи / О. С. Сотников, Н. М. Парамонова //
Усп. физиол. наук. – 2010. – Т. 41. – № 1. – С. 45–57.
50. Сотников, О. С. Динамика ультраструктурных превращений
спаренных биологических мембран в нервной системе / Сотников О. С.,
Парамонова Н. М. и Арчакова Л. И. // Журн. мембр. клет. биол. – 2010. –
Т. 27. – № 6. – С. 489–497.
51. Сотников, О. С. Реабилитация межнейрональной синцитиальной
связи в нервной системе / О. С. Сотников, Л. Е. Фрумкина, В. Н. Майоров //
Тихоокеан. мед. журн. – 2012. – № 2. – С. 75–83.
52. Телушкин, П. К. Показатели энергетического и азотистого обмена у
крыс при инсулиновой гипогликемии / П. К. Телушкин, А. Д. Ноздрачев, П.
П. Потапов // Известия АН. Сер. Биол. – 2008. – № 3. – С. 324–332.
53. Шабанов, П. Д. Применение метапрота в неврологии / П. Д. Шабанов
// TERRA MEDICA NOVA. – 2009. – № 3. – С. 34–38.
54. Шабанов, П. Д. Клиническая фармакология метапрота, нового
противоастенического препарата с психоактивирующими свойствами. – URL:
http://medi.ru/doc/a40g790405j.htm. – 2010.
55. Шабанов, П. Д. Метеоадаптогенные свойства антигипоксантов / П. Д.
Шабанов, В. П. Ганапольский // Эксп. клин. фарм. – 2009. – № 7. – С. 36–41.
56. Шабанов, П. Д. Фармакологические корректоры гипоксии / П. Д.
Шабанов, И. В. Зарубина, В. Н. Цыган. Под ред. А. Б. Белевитина. – СПб. : Н-
Л, 2009. – 916 с.
57. Шабанов, П. Д. К механизму действия полипренолов при ишемии
головного мозга / П. Д. Шабанов, И. В. Зарубина, Vagif S. Soultanov // Мед.
акад. журн. – 2011. – Т. 11. – № 2. – С. 24–31.
58. Шабанов, П. Д. Лечение посттравматической церебрастении

149
антигипоксантами / П. Д. Шабанов, И. В. Зарубина, Ф. Н. Нурманбетова //
Вестн. Рос. воен.-мед. акад. – 2005. – Т. 14. – № 2. – С. 38–41.
59. Юнкеров, В. И. Математико-статистическая обработка данных
медицинских исследований. 3-е изд., доп. / В. И. Юнкеров, С. Г. Григорьев,
М. В. Резванцев. – СПб. : ВМедА, 2011. – 318 с.
60. Activated astrocytes with glycogen accumulation in ischemic penumbra
during the early stage of brain infarction: immunohistochemical and electron
microscopic studies / Kajihara H., Tsutsumi E., Kinoshita A., Nakano J., Takagi
K., Takeo S. // Brain Res. – 2001. – V. 909. – P. 92–101.
61. Attwell, D. Glia and neurons in dialogue / D. Attwell // Nature. – 1994. –
62. Balercia, G. Fine structural organization of the ependymal region of the
paraventricular nucleus of the rat thalamus and its relation with projection neurons
/ G. Balercia, M. Bentivoglio, L. Kruger // Journal of Neurocytology. – 1992. –
63. Bennett, H. Morphological classification of vertebrate blood capillaries /
H. Bennett, J. Luft, J. Hampton // American Journal of Physiology. – 1959. –
64. Bergstorm, H. C. Continuous nicotine administration produces selective,
age-dependent structural alteration of pyramidal neurons from prelimbic cortex /
H. C. Bergstorm, C. D. McDonald, H. T. French, R. F. Smith // Synapse. – 2008. –
65. Berridge, M. J. Relationship between secretagogue-induced Ca2+ release
and inositol polyphosphate production in permeabilized pancreatic acinar cells / M.
J. Berridge, S. Triangi // J. Biol. Chem. – 1985. – V. 260. – № 12. – Р. 7309–
7315.
66. Betz-Corradin, S. Inducible nitric oxide synthase activity of cloned
murine microglial сells / S. Betz-Corradin et al. // Glia. – 1993. – N 7. – P. 255–
262.

150
67. Bi, M. N-Butylphthalide Alleviates Blood-Brain Barrier Impairment in
Rats Exposed to Carbon Monoxide / M. Bi, M. Zhang, D. Guo , W. et al. // Front
Pharmacol. – 2016. –V. 7. – P. 394.
68. Bitzer-Quintero, О. К., Gonzalez-Burgos I. Immune System in the Brain:
A Modulatory Role on Dendritic Spine Morphophysiology? // Neural Plasticity. –
2012. – Article ID. 348642. – 7 p.
69. Bjorklund, A. Dopamine-containing systems in the CNS / А. Bjorklund,
О. Lindvall. // In: Handbook of neuroanatomy. –V. 2. Classical neurotransmitters
in the CNS. Part 1. – Amsterdam, New-York, Oxford : Elsevier Sci. Publ. – 1984.
– P. 55–122.
70. Brandon, A. М. Developmental stage of oligodendrocytes determines
their response to activated microglia in vitro / A. М. Brandon et al. // Journal of
Neuroinflammation. – 2007. – V. 4. – P. 28.
71. Caltana, L. Neuronal and glial alterations due to focal cortical hypoxia
induced by direct cobalt chloride (CoCl2) brain injection / L. Caltana, А. Merelli,
А. Lazarowski, А. Brusco // Neurotox. Res. – 2009. – V. 15, № 4. – P. 348–358.
72. Carney, J. M. Reversal of age-related increase in brain protein oxidation,
decrease in enzyme activity, and loss in temporal and spatial memory by chronic
administration of the spin-trapping compound N-tert-butyl-alpha-phenylnitrone / J.
M. Carney, P. E. Starke-Reed, C. N. Oliver et al. // Proc. Natl. Acad. Sci. USA. –
1991. – V. 88. – № 9. – P. 3633–3636.
73. Charriaut-Marlangue, C. Apoptosis and necrosis after reversible focal
ischemia: an in situ DNA fragmentation analysis / C. Charriaut-Marlangue, I.
Margaill, A. Represa. et al. // J. Cereb. Blood Flow Metab. – 1996. – V. 16. –
P. 186–194.
74. Chew, L. J. Microglia and inflammation: impact on developmental brain
injuries // L. J. Chew, A. Takanohashi, M. // Bell Ment. Retard. Dev. Disabil. Res.
Rev. – 2006. – V. 12. – P. 105–112.
75. Chiamulera, С. Qualitative and quantitative analysis of the progressive

151
cerebral damage after middle cerebral artery occlusion in mice / С. Chiamulera, А.
Terron, А. Reggiani et al. // Brain Res. – 1993. – V. 606. – № 2. – Р. 251–258.
76. Chouaf, L. Comparative marker analysis of the ependymocytes of the
subcommissural organ in four different mammalian species / L. Chouaf et al. //
Cell and tissue research. – 1989. – V. 257. – № 2. – P. 55–62.
77. Chopp, M. Т. Apoptosis in focal cerebral ischemia / M. Т. Chopp, Y. Li.
// Acta Neurochir. Suppl. – 1996. – V. 66. – P. 21–26.
78. Choi, D. W. Cerebral hypoxia: some new approaches and unanswered
question / D. W. Choi // J. Neurosci. – 1990. – V. 10. – Р. 2493–2501.
79. Choi, D. W. Nitric oxide: foe or friend to the injured brain? / D. W. Choi
// Proc. Natl. Acad. Sci. USA. – 1993. – V. 90. – P. 9741–9743.
80. Choi, D. W. Zinc and brain injury / D. W. Choi, J. Y. Koh. // Annu. Rev.
Neurosci. – 1998. – V. 21. – P. 347–375.
81. Clark, R. K. Development of tissue damage, inflammation and resolution
following stroke: an immunohistochemical and quantitative planimetric study / R.
K. Clark, E. V. Lee, C. J. Fish et al. // Brain Res. Bull. – 1993. – V. 31. – P. 565–
572.
82. Cohen, G. M. Key morphological features of apoptosis may occur in the
absence of internucleosomal fragmentation / G. M. Cohen, X. M. Sun, R. T.
Snowden. at al. // Biochem. J. – 1992. – V. 286 (pt2). – P. 331–334.
83. Constantinescu, C. S. Murine macrophages stimulated with central and
peripheral nervous system myelin or purified myelin proteins release inflammatory
products / C. S. Constantinescu et al. // Neuroscience Letters. – 2000. – V. 287. –
P. 171–174.
84. Damon, T. M. A novel neuronal messenger molecule in brain: the free
radical, nitric oxide / T. M. Damon, V. L. Dawson, S. H. Snyder // Ann. Neurol. –
1992. – V. 32. – № 3. – P. 297–311.
85. Dai, X. The trophic role of oligodendrocytes in the basal forebrain /

152
X. Dai, L. D. Larcher, P. V. Clinton et al. // J. Neurosci. – 2003. – V. 23. – № 13б.
– Р. 5846–5853.
86. Dereski, M. O. The heterogeneous temporal evolution of focal ischemic
neuronal damage in the rat. / M. O. Dereski, M. Chopp, R. A. Knight, L. C.
Rodolosi, J. H. Garcia // Acta Neuropathol. – 1993. – V. 85. – № 3. – P. 327–333.
87. Ernst, E. Leukocyte rheology in recent stroke / Е. Ernst, А. Т. Matrai, F.
Paulsen // Stroke. – 1987. – V. 18. – №. 1. – Р. 59 – 62.
88. Eisenbarth, SC. Innate instruction of adaptive immunity revisited: the
inflammasome / S.C. Eisenbarth, R.A. Flavell. // EMBO Mol. Med. – 2009. – V. 1.
–P. 92 – 98.
89. Fisher, M. Current Review of Cerebrovascular Disease / М. Fisher, J.
Bogousslavsky // Philadelphia: CM. – 1996. – 237 р.
90. Fisher, M. Evolving stroke and the ischemic penumbra / M. Fisher, J. H.
Garcia // Neurology. – 1996. – V. 47. – № 4. – P. 884–888.
91. Frantseva, M.V. Ischemia-induced brain damage depends on specific gap-
junctional coupling / M.V. Frantseva, L. Kokarovtseva, J.L. Perez Velazquez // J.
Cereb. Blood Flow Metab. – 2003. – V. 23. – № 2. –Р. 261 – 270.
92. Frederickson, C. J. Neurobiology of zinc and zinc-containing neurons / C.
J. Frederickson // Rev. Neurobiol. – 1989. – V. 31. – P. 145–238.
93. Garcia, J. H. Progression from ischemic injury to infarct following middle
cerebral artery occlusion in the rat / J. H. Garcia, Y. Yoshida, H. Chen et al. // Am.
J. Pathol. – 1993. – V. 142. – P. 623–635.
94. Garcia, J. H. Ischemic stroke and incomplete infarction / J. H. Garcia, N.
A. Lassen, C. Weiller, B. Sperling, J. Nakagawara. // Stroke. – 1996. – V. 27. – №
4. – Р. 761–765.
95. Ginsberg, M. D. Rodent models of cerebral ischemia / M. D. Ginsberg, R.
Busto // Stroke. – 1989. – V. 20. – №. 12. – С. 1627–1642.
96. Ginsberg, M. D. New strategies to prevent neural damage from ischemic

153
stroke / M. D. Ginsberg. – New York : Acad. Press, 1994. – 251 р.
97. Ginsberg, M. D. The ischemic penumbra, injury thresholds, and the
therapeutic window for acute stroke / M. D. Ginsberg, W. A. Pulsinelli // Ann.
Neurol. – 1994. – V. 36. – № 4. – Р. 553–554.
98. Giulian, D. Reactive microglia and ischemic injury. In: Primer on
Cerebrovascular Diseases (Welsh M., Caplan L. Siesjo B. Weir B. Reis D. J. eds.).
/ D. Giulian. – San Diego : Acad. Press, 1997. – Р. 117–124.
99. Giulian, D. Reactive mononuclear phagocytes release neurotoxins after
ischemic and traumatic injury to the central nervous system / D. Giulian, M.
Corpuz, S. Chapman, M. Mansouri, C. Robertson. // J. Neurosci. Res. – 1993. – V.
36. – №. 6. – Р. 681–693.
100. Hajek
101. , T. Neuroanatomical abnormalities as risk factors for bipolar disorder /
T. Hajek, N. Carrey, M. Alda // Bipolar disorder. – 2005. – V. 7. – P. 393–403.
102. Hallenbeck, J. M. Mechanisms of Secondary Brain Damage in Cerebral
Ischemia and Trauma / J. M. Hallenbeck // New York: Acad. Press, 1996. – P. 27–
31.
103. Hansen, A. Effect of anoxia on ion distribution in the brain / A. J.
Hansen // Physiol. Rev. – 1985. – V. 65. – Р. 101–148.
104. Hegstad, E. Amino-acid release from human cerebral cortex during
simulated ischaemia in vitro / E. Hegstad, J. Berg-Johnsen, T. S. Haugstad et al. //
Acta neurochir. – 1996. – V. 138 – № 2. – Р. 234–241.
105. Hennerici, M. The subclavian steal phenomenon: a common vascular
disorder with rare neurologic deficits / М. Hennerici, С. Klemm, W. Rautenberg //
Neurology – 1988. – V. 38. – P. 669–673.
106. Hossmann, K. A. (а) Glutamate-mediated injury in focal cerebral
ischemia: the excitotoxin hypothesis revised / К. А. Hossmann // Brain Pathol. –
1994. – V. 4. – № 1. – Р. 23–36.
107. Hossmann, K. A. (в) Viability thresholds and the penumbra of focal
ischemia // Ann Neurol. / К. А. Hossmann. – 1994. – V. 36. – № 4. – Р. 557–565.

154
108. Hyden, H. Behavior, neural function and RNA / H. Hyden // Prog.
Nucleic Acid Res. Mol. Biol. – 1967. – V. 6. – Р. 187–218.
109. Iadecola, С. Mechanisms of cerebral Ischemic damage. In: Cerebral
Ischemia (Wolfgang Walz ed.) / С. Iadecola. – New Jersey, Totowa: Humana Press.
– 1999. – Р. 3–33.
110. Imai, F. Migration activity of microglia and macrophages into rat brain / F.
Imai, M. Sawada, H. Suzuki et al. // Neurosci. Lett. – 1997. – V. 237. – P. 49–52.
111. Ischemic thresholds of cerebral protein synthesis and energy state
following middle cerebral artery occlusion in rat / G. Mies, S. Ishimaru, Y. Xie, K.
Seo, K. A. Hossmann // J. Cereb. Blood Flow Metab. – 1991. – V. 11. – № 5. – Р.
753–61.
112. Jacewicz, M. Selective gene expression in focal cerebral ischemia /
М. Jacewicz, М. Kiessling, W. A. Pulsinelli // J. Cereb. Blood Flow Metab. – 1986.
– V. 6. – № 3. – Р. 263–272.
113. Johansen, F. F. Hypothermia protects somatostatinergic neurons in rat
dentate hilus from zinc accumulation and cell death after cerebral ischemia / F. F.
Johansen, N. Tonder, M. Berg et al. // Mol. Chem. Neuropathol. – 1993. – V. 18. –
Р. 161–172.
114. Jonas R. A., Ti-Fei Yuan, Yu-Xiang Liang et al. The Spider Effect:
Morphological and Orienting Сlassification of Microglia in Response to Stimuli in
Vivo // PLoS ONE. – 2012. – V. 7.
115. Khalaf, A. Discrepancy in calcium release from the sarcoplasmic
reticulum and intracellular acidic stores for the protection of the heart against
ischemia/reperfusion injury / A. Khalaf, F. Babiker // J. Physiol. Biochem. – 2016.
– V. 72. № 3. – Р. 495 – 508.
116. Kim, S. U. Microglia in health and disease / S. U. Kim, J. Vellis //
Neurobiol. Dis. – 2005. – V. 81. – № 3. – P. 302–313.
117. Kofler, J. Microglia: key innate immune cells of the brain / J. Kofler, C.
A. Wiley // Toxicol. Pathol. – 2011. – V. 39. – P. 103 –114.
118. Kogure, K. Inflammation of the brain after ischemia / K. Kogure,

155
Y. Yamasaki, Y. Matsuo, H. Kato, H. Onodera // Acta Neurochir. Suppl. – 1996. –
V. 66. – Р. 40–43.
119. Kristian, Т. Changes in ionic fluxes during cerebral ischemia. In:
Neuroprotective Agents and Cerebral Ischemia / Т. Kristian, B. K. Siesjo // New
York: Academic press, 1997. – Р. 27–45.
120. Krupinski, J. Role of angiogenesis in patients with cerebral ischemic
stroke / J. Krupinski, J. Kaluza, P. Kumar. et al. // Stroke. – 1994. – V. 25.
– Р. 1794–1798.
121. Ladecola, C. Cerebral circulatory dysregulation in ischemia. In:
Cerebrovascular Diseases / С. Ladecola // Blackwell Med. Cambridge, 1998. – Р.
319–336.
122. Lee J.-M. The changing landscape of ischaemic brain injury mechanisms
/ J.-M. Lee, G. J. Zipfel, D. W. Choi // Nature. – 1999. – V. 24. – Р. 7–14.
123. Li, Y. The Effect of Dexmedetomidine on Oxidative Stress Response
Following Cerebral Ischemia-Reperfusion in Rats and the Expression of
Intracellular Adhesion Molecule-1 (ICAM-1) and S100B / Y. Li, S.Liu // Med. Sci.
Monit. – 2017. – V. 23. –Р. 867–873.
124. Linde, R. Calcium Accumulation Post-Injury an Indicator of Cell
Damage / R. Linde, H. Laursen, A. J. Hansen. In.: Mechanisms of Secondary Brain
Gamage in Cerebral Ischemia and Trauma. – Wien, New-York: Springer. 1996. –
Р. 15–20.
125. Liu, S. The role of pericytes in blood-brain barrier function and stroke /
S. Liu, D. Agalliu, C. Yu, M. Fisher // Curr. Pharm. Des. – 2012. – V. 18. – Р.
3653–3662.
126. Macfarlane, R. Chronic trigeminal ganglionectomy or topical capsaicin
application to pial vessels attenuates postocclusive cortical hyperemia but does not
influence postischemic hypoperfusion / R. Macfarlane, E. Tasdemiroglu, M. A.
Moskowitz, Y. Uemura, E. P. Wei, H. A. Kontos // J Cereb Blood Flow Metab. –
1991. – V. 11. – Р. 261–271.
127. McBridge, W. J. Localization of brain reinforcement mechanisms:

156
intracranial self-administration and intracranial place-conditioning studies / W. J.
McBridge, J. M. Murphy, S. Ikemoto // Beh. Brain Res. – 1990. – V. 101. – P.
129– 152.
128. McGeer, E. G. Neurodegeneration and the immune system. In:
Neurodegenerative Diseases (Calne D. B. ed.) / E. G. McGeer, P. L. McGeer. –
Philadelphia: Brace & World, 1994. – Р. 277–299.
129. Marin-Padilla, M. The human brain: prenatal development and structure
/ M. Marin-Padilla – Berlin: Springer. – 2011. – 145 p.
130. Marysheva, V. V. Effect of amtizol on resistance of SHR mice to acute
hypoxia with hypercapnia under conditions of isolated functioning of one cerebral
hemisphere / V. V. Marysheva, V. V. Mikheev, P. D. Shabanov // Bull. Exp. Biol.
Med. – 2013. – V. 154. – № 4. – Р. 453–456.
131. Matsumoto, K. Role of pyruvate in ischaemia-like conditions on cultured
neurons / K. Matsumoto, K. Yamada, E. Kohmura, A. Kinoshita, T. Hayakawa //
Neurol Res. – 1994. – V. 16. – № 6. – Р. 460–464.
132. Mies, G. Correlation between peri-infarct DC shifts and ischaemic
neuronal damage in rat / G. Mies, T. Iijima, K. A . Hossmann. // Neuroreport. –
1993. – V. 4. – Р. 709–711.
133. Miller R. H. Oligodendrocyte origins / R. H. Miller // Trends in
Neuroscience. – 1996. – V. 19. – P. 92–96.
134. Mitani, A. Origin of intracellular Ca2+ elevation induced by in vitro
ischemia-like condition in hippocampal slices / A. Mitani, S. Yanase, K. Sakai. et
al. // Brain Res. – 1993. – V. 601. – Р. 103–110.
135. Moghaddasi, M. The effects of two-stage carotid occlusion on spatial
memory and pro-inflammatory markers in the hippocampus of rats / M.
Moghaddasi, M.Taati, P. Asadian et al. // J. Physiol. Sci. – 2017. – V. 67. № 3. – Р.
415–423.
136. Moncada, S. Nitric oxide: physiology, pathophysiology and
pharmacology / S. Moncada, R. M. Palmer, E. A. Higgs // Pharmacol. Rev. – 1991.

157
– V. 43. – № 2. – Р. 109–142.
137. Morley, P. Cerebral Ischemia / P. Morley, J. S. Tauskela, A. M. Hakim. –
138. Nag, S. Morphology and properties of astrocytes / S. Nag // Methods
Mol. Biol. – 2011. – V. 686. – P. 69–100.
139. Nedergaard, M. Characterization of cortical depolarizations evoked in focal
cerebral ischemia / M. Nedergaard, A. J. Hansen // Cereb. Blood Flow Metab.
– 1993. – V. 13. – Р. 568–593.
140. Nguyen, K. B. Survival And Mitosis Of Myelinating Oligodendrocytes
In Experimental Autoimmune Encephalomyelitis: An Immunocytochemical Study
With Rip Antibody / K. B.Nguyen et al. // Acta neuropathologica. – 1999. – V. 98.
– № 1. – P. 39–47.
141. Nowak, T. S. Reprogramming of Gene Expression after Ischemia. In:
Cerebral Ischemia / T. S. Nowak , J. Nowak, M. Kiessling. – New Jersey: Humana
Press, 1999. – P. 145–217.
142. Obrenovitch, T. P. Changes in extracellular glutamate concentration
associated with propagating cortical spreading depression. In: Experimental
headache models in animal and man / T. P. Obrenovitch, E. Zilkha. – New York:
Raven press, 1995. – Р. 113–117.
143. Okoye, G. S. Migration of A7 immortalized astrocyte cells grafted into
the adult rat striatum / G. S. Okoye, E. M. Powel, H. M. Geller // J. Comp. Phys. –
1995. – V. 262. – Р. 524–534.
144. Oliver, C. N. Oxidative damage to brain proteins, loss of glutamine
synthetase activity, and production of free radicals during ischemia/reperfusion-
induced injury to gerbil brain / C. N. Oliver, P. E. Starke-Reed, E. R. Stadtman et
al. // Proc. Nat. Acad Sci USA. – 1990. – V. 87. – № 13. – Р. 5144–5147.
145. Olney, J. W. E. New mechanisms of excitatory transmitter neurotoxicity
/ J. W. E. Olney // J. Neural Transm. Suppl. – 1994. – V. 43. – Р. 47–51.

158
146. Olney, J. W. E. Neurotoxicity of excitatory aminoacids: Kainic Acid as
tool in Neurobiology / J. W. E. Olney, J. W. McGeer, P. Oiney. – New York:
McGeer J.W. ed., 1978. – P. 95–121.
147. Ooboshi, H. Inhibition of ischemia-induced dopamine release by omega-
conotoxin, a calcium channel blocker, in the striatum of spontaneously
hypertensive rats: in vivo brain dialysis study / Ooboshi H., Sadoshima S., et al. //
J. Neurochem. – 1992. – V. 58. – №. 1. –Р. 298 – 303.
148. Orrenius, S. Ca(2+)-dependent mechanisms of cytotoxicity and
programmed cell death / S. Orrenius, M. J. J. McCabe, P. Nicotera // Toxicol. Lett.
– 1992. – V. 64. – Р. 357–364.
149. Packer, L. L. Free radicals in the brain. / L. L. Packer, L. Packer, Y.
Prilipko, Y. Cyirsten. – Berlin : Springer, 1992. – 120 р.
150. Paemeleire, K. The cellular basis of neurovascular metabolic coupling /
K. Paemeleire // Acta. Neurol. Belg. – 2002. – V. 102. – P. 153–157.
151. Pappata, S. PET study of changes in local brain hemodynamics and
oxygen metabolism after unilateral middle cerebral artery occlusion in baboons / S.
Pappata, M. Fiorelli, T. Rommel et al // J. Cereb. Blood Flow Metab. – 1993. –
V. 13. – Р. 31–34.
152. Paxinos G. The Rat Nervous System. 3-d ed. / G. Paxinos. – Amsterdam,
Boston, Heidelberg, London, New York, Oxford, Paris, San Diego, San Francisco,
Singapore, Sydney, Tokyo: Elsevier Acad. Press, 2004. – 1780 р.
153. Paxinos, G., Watson, C. The rat brain in stereotaxic coordinate. 4-th ed. /
G. Paxinos, C. Watson. –Sydney, Orlando, San Diego, New-York, Austin, London,
Montreal, Toronto : Academic press, 1998. – Pр. 16–24, 414, 429.
154. Racchetti, G. Astrocyte stellation, a process dependent on Rac1 is
sustained by the regulated exocytosis of enlargeosomes / G. Racchetti, R.
D'Alessandro, J. Meldolesi // Glia. – 2012. – V. 60, issue 3. – P. 465–475.
155. Ransohoff, R. M. The myeloid cells of the central nervous system
parenchyma / R. M. Ransohoff, A. E. Cardona // Nature. – 2010. – V. 11. – Р. 253–

159
262.
156. Ratner, N. A. Role for Cdk5 Kinase in Fast Anterograde Axonal
Transport: Novel Effects of Olomoucine and the APC Tumor Suppressor Protein /
N. Ratner, G. S., Sadoul R., Dubois-Dauphin M., Fernandel P. A. et al. //
Mechanisms of neuronal cell death // Adv. Neurol. – 1996. – V. 71. – Р. 419–424.
157. Righi, M. The microglial cell: a cytokine source in the CNS / M. Righi et
al. // J. Сhemother. – 1991. – V. 3. – P. 41 – 43.
158. Rudko, A.S. Influence of vascular endothelial growth factor on
angiogenesis and neurogenesis / A.S. Rudko, M.K. Efendieva, M.V. Budzinskaya,
M.A. Karpilova // Vestn. Oftalmol. – 2017. – V. 133. – №. 3. –Р. 75 – 81.
159. Sánchez-Abarca, L. I. Oligodendrocytes use lactate as a source of energy
and as a precursor of lipids / Sánchez-Abarca L. I., Tabernero A., Medina J. M. //
Glia. – 2001. – V. 36. – № 3. – Р. 321–329.
160. Sadoul, R. Mechanisms of neuronal cell death / R. Sadoul, M. Dubois-
Dauphin, P. A. Fernandez et al. // Adv. Neurol. – 1996. – V. 71. – Р. 419–424.
161. Scheinberg P. The biologic basis for the treatment of acute stroke / P.
Scheinberg // Neurology – 1991. – V. 41. – Р. 1867–1873.
162. Schilling, M. Microglial activation precedes and predominates over
macrophage infiltration in transient focal cerebral ischemia: a study in green
fluorescent protein transgenic bone marrow chimeric mice / M. Schilling et al. //
Experimental Neurology. – 2003. – V. 183. – P. 25–33.
163. Scheinberg, P. The biologic basis for the treatment of acute stroke / P.
Scheinberg // Neurology. – 1991. – V. 41. – № 12. – Р. 1867–1873.
164. Sensi, S. L. Measurement of intracellular free zinc in living cortical
neurons: routes of entry / S. L. Sensi, L. M. Canzoniero, S. P. Yu. et al. // J.
Neurosci. – 1997. – V. 17. – № 24. – Р. 9554–9564.
165. Siesjo, B. K. Calcium fluxes, calcium antagonists, and calcium-related
pathology in brain ischemia, hypoglycemia, and spreading depression: a unifying
hypothesis / B. K. Siesjo, F. J. Bengtsson // Cereb. Blood Flow Metab. – 1989. –

160
V. 9. – № 2. – Р. 127–140.
166. Siren, A.L. Microcirculatory stasis in the brain / A.L. Siren, R.M.
McCarron, Y. Lu et al. // Amsterdam: Experta Medica, 1993. –P. 169 – 175.
167. Smith, J. A. Role of pro-inflammatory cytokines released from microglia
in neurodegenerative diseases / J. A. Smith, A. Das, S. K. Ray, N. L. Banik // Brain
Res. Bull. – 2011. – V. 87. – P. 10–20.
168. Snchez-Abarca, L. I. Oligodendrocytes use lactate as a source of energy
and as a precursor of lipids / L. I. Sаnchez-Abarca, A. Tabernero, J. M. Medina //
Glia. –2001. – Vol. 36(3). – P. 1–9.
169. Somjen, G. G. Mechanism of spreading depression: a review of recent
findings and a hypothesis / G. G. Somjen, P. G. Aitken, C. L. Czeh // Can. J.
Physiol. Pharmacol. – 1992. – V.70. Suppl. – P. 248–254.
170. Sontheimer, H. Voltage-gated sodium and calcium channels / H.
Sontheimer, J. M. Richie // Neuroglia. – New York: Oxford University Press,
1995. – P. 246–258.
171. Soulet, D. Microglia / D. Soulet, S. Rivest // Curr. Biol. – 2008. – V. 18.
– P. 506–508.
172. Stegmayr, B. Stroke in Northern Sweden / В. Stegmayr, К. Asplund //
Scand. J. Public Health. Suppl. – 2003. – V. 61. – Р. 60–69.
173. Stella M. C. Macrophage Stimulating Protein Is a Novel Neurotrophic
Factor / M. C. Stella et al. // Molecular biology of the cell. – 2001. – V. 12. – P.
341–352.
174. Stroke. A practical guide to management / Warfow C. P., Dennis M. S.,
van Gijn J. et al. – Oxford, Blackwell: Science, 1996. – 160 p.
175. Szpak, G. M. Border zone neovascularization in cerebral ischemic
infarct / G. M. Szpak et al. // Folia Neuropathol. – 1999. – V. 37. – P. 264–268.
176. Tagaya, M. DNA scission after focal brain ischemia. Temporal
differences in two species / M. Tagaya, K.F. Liu, B. Copeland, D. Seiffert,

161
R. Engler, J. H. Garcia, G. J. del Zoppo // Stroke. – 1997. – V. 28. – № 6. – Р.
1245–1254.
177. Tanahashi, N. Cerebral microvascular reserve for hyperemia. In:
Cerebral hyperemia and ischemia / M. Tomita, Т. Sawada, H. Naritomi, W.-D.
Heiss. eds. – Amsterdam : Excerpta Medica. – 1988. – P. 173–182.
178. Tang, N. Genetic Mutation of GluN2B Protects Brain Cells Against
Stroke Damages / N. Tang, J. Wu, H. Zhu et al. // PLoS ONE. Mol. Neurobiol. –
2017. – V. 54.
179. Tomita, M. Microcirculatory stasis in the brain. In: Microcirculatory
stasis in the brain / M. Tomita, G. Mchedlishvili, W.I. Rosenblum, W-D. Heiss, Y.
Fukuuchi eds.// Amsterdam: Excerpta Medica. – 1993. – P.1 – 7.
180. Tomita, M. Leukocytes, macrophages and secondary brain damage
following cerebral ischemia / M. Tomita, Y. Fukuuchi // Acta Neurochir Suppl. –
1996 – V. 66. – P. 32–39.
181. Tonder, N. Possible role of zinc in the selective degeneration of dentate
hilar neurons after cerebral ischemia in the adult rat / N. Tonder, F. F. Johansen, C.
J. Frederickson et al. // Neurosci. Lett. – 1990. – V. 109. – № 3. – Р. 247–252.
182. Ueharа, M. Morphological features and frequencies of various types of
glial cells in the ventral horn of the chicken spinal cord / M. Uehara, T. Ueshima //
Jap. J. Veterinary Sci. – 1985. – V. 47. – P. 791–798.
183. Ugidos, I.F. Neuroprotective effect of 2-hydroxy arachidonic acid in a
rat model of transient middle cerebral artery occlusion / I.F. Ugidos, M. Santos-
Galdiano, D. Pérez-Rodríguez et al. // Biochim. Biophys. Acta. – 2017. – V. 1859.
– № 9, рt B. –Р. 1648 – 1656.
184. Voiculescu, V. Current concepts in brain ischemia. A review / V.
Voiculescu // Neurol. Psychiatr. Med. – 1989. – V. 27. – № 3. – P. 175–186.
185. Voskuhl R. R. Reactive astrocytes form scar-like perivascular barriers to
leukocytes during adaptive immune inflammation of the CNS / R. R. Voskuhl, R.
S. Peterson, B. Song, Y. Ao, L. B. Morales, S. Tiwari-Woodruff, M. V.

162
Sofroniew // J. Neurosci. – 2009. – V. 29. – P. 11511–11522.
186. Walterfang, M. Neuropathological, neurogenetic and neuroimaging
evidence for white matter pathology in schizophrenia / M. Walterfang et al. //
Neurosci. Biobehav. reviews. – 2006. – V. 30. – P. 918–948.
187. Wilkins, А. Oligodendrocytes promote neuronal survival and axonal
length by distinct intracellular mechanisms: a novel role for oligodendrocyte-
derived glial cell line-derived neurotrophic factor / А. Wilkins, H. Majed, R.
Layfield et al. // J. Neurosci. – 2003. – V. 23. – № 12. – P. 4967–4974.
188. Yang, F. NLRP3 deficiency ameliorates neurovascular damage in
experimental ischemic stroke / F. Yang, Z. Wang, X. Wei et al. // J. Cereb. Blood
Flow Metab. – 2014. – V. 34. – № 4. –Р. 660 – 667.
189. Yoshida, S. Effect of transient ischemia on free fatty acids and
phospholipids in the gerbil brain. Lipid peroxidation as a possible cause of
postischemic injury / S. Yoshida, S. Inoh, T. Asano et al. // J. Neurosurg. – 1980. –
V. 53. – № 3. – Р. 323–331.
190. Yu, K. L. Second messenger regulation of mouse gonadotropin-releasing
hormone gene expression in immortalized mouse hypothalamic GT1-3 cells / K. L.
Yu, T. T. Yeo, K. W. Dong et al. // Mol. Cell. Endocrinol. – 1994. – V. 102. – Р 85–
92.
191. Zablocka, B. Enhancement of 3[H]D-aspartate release during ischemia
like conditions in rat hippocampal slices: source of excitatory amino acids /
В. Zablocka, К. Domanska-Janik // Acta Neurobiol. Exp. Warsz. – 1996. – V. 56. –
№ 1. – Р. 63–70.
192. Zhang, R. L. Temporal profile of ischemic tissue damage, neutrophil
response, and vascular plugging following permanent and transient (2H) middle
cerebral artery occlusion in the rat / R. L. Zhang, M. Chopp, H. Chen, J. H. Garcia.
// J. Neural. Sci. – 1994. – V. 125. – Р. 3–10.
193. Zhang, R. L. Oligodendrogenesis after cerebral ischemia / R. L. Zhang,
M. Chopp, Z. G. Zhang // Front. Cell. Neurosci. – 2013. – V. 29. – № 7. – С. 201.

163
194. Zhang, Y. Paeoniflorin Attenuates Cerebral Ischemia-Induced Injury by
Regulating Ca2+/CaMKII/CREB Signaling Pathway / Zhang Y., Qiao L., Xu W. et
al. // Molecules. – 2017. – V. 22. – № 3. –Р. 359 – 371.
195. Zhou, H. F. Migration of astrocytes transplanted to the midbrain of
neonatal rats / H. F. Zhou, R. D. Lund // Journal of Compar. Neurol. – 1992. – V.
317. – P. 145–155.
П Р И Л О Ж Е Н И Е

164
Рисунок 1. Биохимико-энергетические реакции мозга на разных фокальных уровнях снижения мозгового кровотока. Обобщено Гусевым Е. И. и Скворцовой В. И. (2001) на
основе данных циркуляторно-биохимических исследований (Jacewicz M., Kiessling М. and Pulsinelli W. A., 1986; Mies G. et al., 1991; Hossmann, K. A. 1994 а, б)
Рисунок 2. Формирование инфаркта мозга на фоне снижения церебрального кровотока (по: Fisher M., Takano К., 1995)

165
Мозговой кровоток,
мл/100 г в мин
Рисунок 3. Гемодинамические события, происходящие после церебральной ишемии
и реперфузии (по: Tomita M., 1993)

166
Рисунок 4. Этапы глютамат-кальциевого каскада (по: Choi D. W., 1993; Olney J. W. E.,
1994; цитата из: Гусев Е. И. и Скворцова В. И., 2001)

167
Рисунок 5. Основные механизмы глютамат-кальциевого каскада (по: Choi D. W.,
1993; Olney J. W. E., 1994; цитата из: Гусев Е. И. и Скворцова В. И., 2001)

168




















