
Notes on behaviour, communication and reproduction in captive Hyloxalus toachi (Anura:...
12
Int. Zoo Yb. (2008) 42: 78–89 DOI:10.1111/j.1748-1090.2008.00051.x Notes on behaviour, communication and reproduction in captive Hyloxalus toachi (Anura: Dendrobatidae), an Endangered Ecuadorian frog A. QUIGUANGO-UBILLU ´ S & L. A. COLOMA Museo de Zoologı ´a, Escuela de Biologı ´a, Pontificia Universidad Cato ´lica del Ecuador, Aptdo, 17-01-2184, Quito, Ecuador E-mail: [email protected] We studied, under laboratory conditions, behavioural traits related to social interactions, parental care and commu- nication in Hyloxalus toachi. This species is diurnal. Females spent most of their time maintaining and defend- ing territories and were more territorial than <<. Male vocalizations included long-range advertisement calls, two kinds of close-range courtship calls, aggression calls and sounds produced while attempting to clasp ,,. During body contact, << and ,, produced encounter calls. Preovipositional amplexus was cephalic and lasted 10– 15 minutes. During terrestrial oviposition, the < climbed onto the dorsum of the , in an axillary position, but without clasping the ,. Fertilization occurred as the eggs were deposited. When oviposition was completed, the < left the ,, while the , remained close to the clutch for up to 5hours. The mean clutch size was 11 eggs. Eggs had a mean diameter of 2 4 mm. The incubation time was 13–16 days. During that time, egg attendance and transport of tadpoles to water were performed exclusively by the <. Tadpoles were carried on the <’s dorsum for up to 5 days. The behaviours and features reported here are useful for interpreting trends in the evolution of dendroba- tid behaviour. Key-words: behaviour; Dendrobatidae; Ecuador; Hylox- alus toachi; parental care; reproduction; vocalization. INTRODUCTION Members of the family Dendrobatidae (Anomaloglossinae, Aromobatinae, Allobati- nae, Colostethinae, Hyloxalinae and Dendro- batinae of Grant et al., 2006) have some of the most complex social and reproductive behaviours found among frogs. For example, Hyloxalus species are territorial, attend clutches and transport tadpoles (Myers & Daly, 1983; Weygoldt, 1987). Some species of Dendrobates (Oophaga and Ranitomeya sensu Grant et al., 2006) and Anomaloglossus beebei have larval oophagy, a type of parental care, where ,, provide eggs to feed free-living tadpoles (Grant et al., 2006, and references therein). Workers studying the evolution of these complex behaviours (e.g. Weygoldt, 1987; Zimmermann & Zimmermann, 1988; Grant et al., 2006) have been hampered not only by the limitations of the phylogeny available but also by a lack of knowledge about possible variation in inter-specific behaviour that might occur. The genus Hyloxalus is a large clade (57 species to date, Grant et al., 2006) of non-toxic dendrobatids, which is hypothesized to be more closely related to the derived clade (Dendrobatinae) of aposematic dendrobatids (Myers et al., 1991; Grant et al., 2006). According to Santos et al. (2003), Hyloxalus toachi is the sister taxon of Hyloxalus awa. There have been few behavioural studies carried out for the Hyloxalus genus, with the exception of Hyloxalus sp, H. awa, Hyloxalus azureiventris and Hyloxalus subpunctatus (Mudrack, 1969; Kneller, 1987; Correa 1995; Fandin ˜o et al., 1997; Lo ¨tters et al., 2000). In this study, we describe the behaviour of cap- tive H. toachi (Coloma, 1995). We were parti- cularly interested in documenting aspects of behaviour that might be difficult to examine under natural conditions but could be of critical importance to understanding the evo- lution of complex social and reproductive behaviours. These aspects include actual oviposition and time of fertilization, , voca- lizations, , interclutch interval and </, varia- tion in courtship behaviour. 78 AMPHIBIAN CONSERVATION Int. Zoo Yb. (2008) 42: 78–89. c 2008 The Authors. Journal compilation c 2008 The Zoological Society of London
-
Upload
independent -
Category
Documents
-
view
4 -
download
0
Transcript of Notes on behaviour, communication and reproduction in captive Hyloxalus toachi (Anura:...

Int. Zoo Yb. (2008) 42: 78–89
DOI:10.1111/j.1748-1090.2008.00051.x
Notes on behaviour, communication andreproduction in captive Hyloxalus toachi (Anura:Dendrobatidae), an Endangered Ecuadorian frog
A. QUIGUANGO-UBILLUS & L. A. COLOMAMuseo de Zoologıa, Escuela de Biologıa, Pontificia Universidad Catolica del Ecuador, Aptdo,17-01-2184, Quito, EcuadorE-mail: [email protected]
We studied, under laboratory conditions, behavioural traitsrelated to social interactions, parental care and commu-nication in Hyloxalus toachi. This species is diurnal.Females spent most of their time maintaining and defend-ing territories and were more territorial than <<. Malevocalizations included long-range advertisement calls, twokinds of close-range courtship calls, aggression calls andsounds produced while attempting to clasp ,,. Duringbody contact, << and ,, produced encounter calls.Preovipositional amplexus was cephalic and lasted 10–15minutes. During terrestrial oviposition, the < climbedonto the dorsum of the , in an axillary position, butwithout clasping the ,. Fertilization occurred as the eggswere deposited. When oviposition was completed, the <left the ,, while the , remained close to the clutch for upto 5 hours. The mean clutch size was 11 eggs. Eggs hada mean diameter of 2 � 4mm. The incubation time was13–16days. During that time, egg attendance and transportof tadpoles to water were performed exclusively bythe <. Tadpoles were carried on the <’s dorsum for upto 5 days. The behaviours and features reported here areuseful for interpreting trends in the evolution of dendroba-tid behaviour.
Key-words: behaviour; Dendrobatidae; Ecuador; Hylox-alus toachi; parental care; reproduction; vocalization.
INTRODUCTION
Members of the family Dendrobatidae(Anomaloglossinae, Aromobatinae, Allobati-nae, Colostethinae, Hyloxalinae and Dendro-batinae of Grant et al., 2006) have someof the most complex social and reproductivebehaviours found among frogs. For example,Hyloxalus species are territorial, attend clutchesand transport tadpoles (Myers & Daly, 1983;Weygoldt, 1987). Some species ofDendrobates(Oophaga and Ranitomeya sensu Grant et al.,2006) and Anomaloglossus beebei have larval
oophagy, a type of parental care, where ,,provide eggs to feed free-living tadpoles(Grant et al., 2006, and references therein).Workers studying the evolution of thesecomplex behaviours (e.g. Weygoldt, 1987;Zimmermann & Zimmermann, 1988; Grantet al., 2006) have been hampered not only bythe limitations of the phylogeny available butalso by a lack of knowledge about possiblevariation in inter-specific behaviour that mightoccur. The genusHyloxalus is a large clade (57species to date, Grant et al., 2006) of non-toxicdendrobatids, which is hypothesized to bemore closely related to the derived clade(Dendrobatinae) of aposematic dendrobatids(Myers et al., 1991; Grant et al., 2006).According to Santos et al. (2003), Hyloxalustoachi is the sister taxon of Hyloxalus awa.
There have been few behavioural studiescarried out for the Hyloxalus genus, with theexception ofHyloxalus sp,H. awa,Hyloxalusazureiventris and Hyloxalus subpunctatus(Mudrack, 1969; Kneller, 1987; Correa 1995;Fandino et al., 1997; Lotters et al., 2000). Inthis study, we describe the behaviour of cap-tive H. toachi (Coloma, 1995). We were parti-cularly interested in documenting aspects ofbehaviour that might be difficult to examineunder natural conditions but could be ofcritical importance to understanding the evo-lution of complex social and reproductivebehaviours. These aspects include actualoviposition and time of fertilization, , voca-lizations, , interclutch interval and </, varia-tion in courtship behaviour.
78 AMPHIBIAN CONSERVATION
Int. Zoo Yb. (2008) 42: 78–89. c� 2008 The Authors. Journal compilation c� 2008 The Zoological Society of London

Coloma (1995) provided morphologicaland taxonomic information on H. toachi aswell as a description of the tadpole and dataon its distribution and ecology. Hyloxalustoachi inhabits north-western Ecuador withinthe Tropical Humid Forest and Very HumidPremontane Forest life zones, where it rangesfrom 200 to 1410m above sea level (a.s.l.).At these zones, the mean annual temperatureranges 18–25 1C; the mean annual precipita-tion is 2000–4000mm. Hyloxalus toachi islisted as Endangered in the 2006 IUCN RedList of Threatened Species (IUCN, 2006).
MATERIALS AND METHODS
Collection site and laboratorymaintenance
The seven adult individuals of H. toachiused in this study (QCAZ 9433, 9435–9440)were collected 46 km west of Lita (about7813701000W; 11201000N), 335m a.s.l., provinceof Esmeraldas, Ecuador, in March 1994, and on15 April and 27 August 1995. Adults werefound on stream banks among remains ofnatural vegetation. Tadpoles were found instream pools with slow-moving water.
A < and , were placed in a terrarium85 cm � 28 cm � 30 cm (length � width �depth). A < and two ,, were placed inanother terrarium with the same dimensions.A < and , were placed in a terrarium of61 cm � 36 cm � 35 cm.
Terrariums were situated at a slight incline.The lower portion of the terrarium containedwater covering about 40% of the total length,and rocks were placed in the water to providedry sites. The portion of the terrarium withoutwater contained leaf litter placed in such amanner that animal refugia were available.Following the technique of Zimmermann& Zimmermann (1981), half of a coconut shellwas placed in each terrarium to provide breed-ing and refuge sites. A mirror was placedunder the terrarium, to facilitate observationsunder refugia when frogs occupied these sites.Adults were fed once a week with fruitfliesDrosophila spp, Curculionidae, and larvaeand adult moths (Lepidoptera).
Behaviour observations
Preliminary observations were carried outfrom March to June 1995, to determine timeof maximum activity. Later, 762 hours ofobservations were made on seven individualsfrom October 1995 to April 1996, between0600 and 1900 hours. The description of mostbehavioural traits follows Zimmermann &Zimmermann (1988). In the text, the numbersin parenthesis give the number of times thata behaviour was observed over the total762 hours of observation of 3.4 (<.,). In mostcases, a mean (�x) and standard deviation ( � )are indicated.
Vocalizations
Recordings were made with a Marantz ModelCP 430 tape recorder, a Sennheiser K6 uni-directional microphone and 60minute-longSony SR metallic tape Type IV cassettes.Original tapes are deposited in the tape archiveat Museo de Zoologıa, Pontificia UniversidadCatolica del Ecuador, Quito (QCAZ). Record-ings were made between 1600 and 1800hours.In the terrariums, temperature was main-tained at 21–22 1C and relative humidity was87–88%. Recordings were digitized, editedand analysed using a Macintosh computerequipped with Sound Effects 0.9.1 (Ricci,1994) and Sound Edit 1.0 (Beck et al., 1992)software. Recordings were input in a frequencyrange of 22 kHz and 8 bytes of resolution.Acoustic terminology follows Heyer et al.(1990).
RESULTS
Territoriality
Hyloxalus toachi was most active during theday. Both << and ,, had territories thatthey defended aggressively. This aggressionmanifested as body attacks to intruders orspecific aggressive vocalizations. The terri-tory of the , was a semi-hidden place whereshe spent most of her time. She defended herterritory aggressively against <<. The < usedhis territory when escaping from ,–< aggres-sion during the day, or as a refuge during the
HYLOXALUS TOACHI BEHAVIOUR, COMMUNICATION AND REPRODUCTION 79
Int. Zoo Yb. (2008) 42: 78–89. c� 2008 The Authors. Journal compilation c� 2008 The Zoological Society of London

night. He also used his territory as a callingsite and for courtship and fertilization whenoviposition occurred. Males aggressivelyguarded their clutches during parental care.Thirty-one per cent of the total behaviouralinteractions involved aggression between <<guarding eggs and ,,.
Animals were quite variable in their ex-pression of behaviours (Table 1), but somegeneralizations are possible. Female–<aggressive encounters showed the followingsequential pattern (Fig. 1): the , looked atthe intruder (usually from a distance of c.4–5 cm); she pulsed her throat (71 occur-rences; n5 4 ,,; 762 hours of observations);and raised her body on all four limbs (12occurrences; n5 4 ,,). If the intruder did not
retreat, the , rammed her snout into the flankof the intruder (30 occurrences; n5 4 ,,) andemitted an encounter call (32 occurrences;n5 4 ,,) (Fig. 1A and B; Table 1). If theintruder persisted, the , jumped onto thedorsum of the < from behind or the side(7 occurrences; n5 4 ,,), and pressedagainst him for 1–2 seconds (7 occurrences;n5 4 ,,); finally, she released him andrammed the distal part of his body with theend of her snout until he left her territory(3 occurrences; n5 4 ,,) (Fig. 1C and D).
The <, while defending a clutch of eggswithin or outside of his territory, continuouslyproduced loud aggression calls (59 occur-rences; n5 3 <<; 762 hours of observations)(Plate 1A; Table 2) when a , approached him.He exuded moisture from his body (visible onthe skin), which also became darker. If the
BEHAVIOUR<<N5 3
,,N5 4
Pulsation of flanks and/or throat 17 71Upright posture 39 325Ramming head against side of other
frog’s body9 30
Body-pushing; jumping at other frog,hitting body against body
3 8
Frontal jumping against the head of theintruder
8 10
Opponents tackling each other withforelimbs, while standing
1 –
Jumping on dorsum of opponent fromback or side
10 7
Clasping the opponent inguinally frombehind
4 3
Body pressing; pushing down rival bysitting on opponent
– 7
Individual struts slowly, with stiff legsin front of rival
– 3
Walking with short and brusquemovements
– 1
Raising body; individual raises bodyon all four extremities
4 12
Subordinate ducks before dominant frog 7 8Encounter call 13 32Loud aggression call 59 –Individual harasses the opponent 3 15Ramming snout against the distal part
of rival3 3
Individual climbs from a frontal positiononto dorsum of the opponent
– 2
Table 1. The total number of occurrences of eachaggressive behaviour observed during 762hours ofobservations for seven Hyloxalus toachi (3.4 <<.,,).Description of behavioural traits follows Zimmer-mann & Zimmermann (1988).
Fig. 1. Sequence of aggression in Hyloxalus toachi,–< (< is shown in grey and , in white): (A) , raisesher body when < approaches her territory; (B) , ramsher snout against <’s body flank; (C) , jumps onto thedorsum of the opponent; (D) , rams <’s distal bodywith her snout and harasses him until he leavesher territory.
80 AMPHIBIAN CONSERVATION
Int. Zoo Yb. (2008) 42: 78–89. c� 2008 The Authors. Journal compilation c� 2008 The Zoological Society of London

Plate 1. Hyloxalus toachi: (A) < emitting an aggressive call; (B) clutch of eggs from below; (C) < moistening aclutch of eggs; (D) < transporting tadpoles. Luis A. Coloma, Pontifica Universidad Catolica del Ecuador.
PARAMETER/KINDOF VOCALIZATION
< ,
LONG-RANGEADVERTISE-MENT
CLOSE-RANGECOURTSHIPTYPE I
CLOSE-RANGECOURTSHIPTYPE II AGGRESSION AMPLEXUS ENCOUNTER
Call length(seconds)
4–21 � 3 12 � 9–26 � 7 0 � 036–1 � 4 22 � 3–60 0 � 229 0 � 035–0 � 057
Calls per minute 2 1 1–6 2 – –Notes per call 10–51 32–73 1–2 31–93 6 1Note length
(seconds)0 � 017–0 � 048 0 � 020–0 � 048 0 � 036–0 � 041 0 � 021–0 � 047 0 � 014–0 � 022 0 � 035–0 � 057
Note interval(seconds)
0 � 35–0 � 38 0 � 20–0 � 85 0 � 72–1 � 08 0 � 50–0 � 68 0 � 022–0 � 024 –
Call interval(seconds)
19 � 4–27 � 6 37–117 5–90 8–12 – –
Fundamentalfrequency (Hz)
2000–2200 2000–2200 2000–2200 2000–2200 500–550 1800–2000
Dominantfrequency (Hz)
4000–4300 4000–4300 4000–4300 4000–4300 – 3800–4300
Sample size (Hz) 25/2 17/2 12/2 18/2 2/1 20/1
Table 2. Parameters of calls ofHyloxalus toachi. Values are given as ranges. Parameters are given as total numberof calls sampled/total number of frogs sampled. Length and time intervals in seconds, frequencies in Hz.
HYLOXALUS TOACHI BEHAVIOUR, COMMUNICATION AND REPRODUCTION 81
Int. Zoo Yb. (2008) 42: 78–89. c� 2008 The Authors. Journal compilation c� 2008 The Zoological Society of London

intruder did not retreat, he performed bodyattacks similar to those described for ,,.During fights, the < also produced encountercalls (13 occurrences; n5 3 <<) similar tothose described for ,, (Table 2). Males didnot defend territories when they were notguarding eggs.
Courtship
Courtship time lasted up to 2 days. One se-quence (Fig. 2A–E), however, lasted 4 hours.The < began courtship with a long-rangeadvertisement call (Fig. 3B; Table 2). This callwas performed from a semi-hidden place that
was usually the site selected by the < foroviposition. While calling, the < straightenedhis forelimbs and elevated the anterior regionof his body (80 occurrences; n5 3 <<). Oneto 2minutes after initiating vocalization, thedorsum of the < became wet and the entirebody darkened (80 occurrences). The ,showed her receptivity by leaving her terri-tory. If the , did not approach the site wherethe < was calling, he left this area and movedto call from a more visible site. If the , thenapproached the calling <, he led her back tothe initial site (12 occurrences; n5 3 <<).When both animals were at this site, the ,performed short jumps, circling sideways
Fig. 2. Sequence of courtship, oviposition and parental care in Hyloxalus toachi (< is shown in grey and , inwhite): (A) < emits long-range advertisement calls from an oviposition site; (B) , approaches < and , circlesaround him, < emits a close-range Type I courtship call; (C) , touches <’s mid-body at the dorsumwith her snout;(D) < takes position in front of , and emits a close-range courtship call Type II, while , remains bent in front of <;(E) preovipositional cephalic amplexus; (F) < climbs onto the dorsum of , in an axillary position, but withoutgrasping her; (G) during oviposition the pair rotates clockwise each time the , deposits an egg; (H) whenoviposition ends, < leaves the , and the site, while , remains over the clutch; (I) , remains close to the clutch; (J) <returns to the clutch and moistens it; (K) < remains close to the clutch most of the time and defends it fromintruders; (L) < transports tadpoles to water (drawings E and L adapted from Duellman & Trueb, 1986).
82 AMPHIBIAN CONSERVATION
Int. Zoo Yb. (2008) 42: 78–89. c� 2008 The Authors. Journal compilation c� 2008 The Zoological Society of London

around her partner (10 occurrences; n5 4,,). The < changed his call that moment, toa close-range courtship call Type I (17 occur-rences; n5 3 <<) (Table 2), the first beha-
viour associated with a , positive response.After this close-range courtship call Type I,the , touched the dorsum of the < with hersnout (12 occurrences; n5 4 ,,). The <, who
Fig. 3. Calls of Hyloxalus toachi: (A) , encounter call; (B) < long-range advertisement call; (C) < close-range TypeII courtship call; (D)< amplexus call. Oscillogram (above) and audiospectrogram (below). (B) and (C) at same scale.
HYLOXALUS TOACHI BEHAVIOUR, COMMUNICATION AND REPRODUCTION 83
Int. Zoo Yb. (2008) 42: 78–89. c� 2008 The Authors. Journal compilation c� 2008 The Zoological Society of London

was previously at the ,’s side, moved in frontof her (6 occurrences; n5 3 <<) while the ,remained in a bent position (14 occurrences;n5 4 ,,). At this point, the < performed aclose-range courtship call Type II (12 occur-rences; n5 3 <<) (Fig. 3C; Table 2). Afterthe vocal stimulation, the < remained quietfor a few seconds and then jumped onto the,’s dorsum and performed cephalic amplexus(Fig. 2E) (13 occurrences; n5 3 <<), whichlasted 10–15minutes. The < was sometimesunsuccessful in clasping the , on the firstattempt and therefore made several attempts.During attempts at clasping, the < emitted asoft, amplexus call (2 occurrences; n5 3 <<)(Fig. 3D; Table 2). This call differed fromother recorded< vocalizations in having fewernotes per call, a lower fundamental frequencyand a shorter note interval (Table 2).
Oviposition
A typical sequence is indicated in Fig. 2F–I.After cephalic amplexus (sensu Castillo-Trenn & Coloma, in press), the<moved awayand then got on the ,’s dorsum. The , wascompletely bent under the < (5 occurrences).The <’s forelimbs were extended to theground and located just behind the ,’sforelimbs (5 occurrences). At this time noclasping occurred but , peristaltic movementswere evident (5 occurrences). The pair re-mained in this position for 15–30minutes(�x5 23 � 4 � 5 � 9, 5 occurrences), after whichoviposition began. The couple rotated clock-wise each time the , deposited an egg(6 occurrences). The eggs were laid close toeach other under the ,’s ventral region. Ovi-position lasted 10–25minutes (�x517 � 8�6 � 5, 6 occurrences). After oviposition, the <left the , and the site (5 occurrences; n5 3<<), while the , remained over the eggs for45–70minutes (�x5 58 � 3 � 9 � 3, 6 occur-rences; n5 4 ,,). At this point, the , movedslightly away from the eggs but remainedclose to the clutch for about 5 hours (�x5287 � 6minutes � 11 � 2, 6 occurrences; n5 4,,). The < returned to the clutch after the ,had left the oviposition site, and he moistenedthe eggs with liquid from his cloaca.
On one occasion, when the < left the ,after oviposition had occurred, the < wasremoved from the terrarium in order to recordwhether or not the eggs were fertilized. Theeggs continued developing, suggesting thatfertilization had occurred simultaneouslywith oviposition.
In five observed cases, the ovipositionsequence was similar to that described above;however, in one case the < did not get onthe ,’s dorsum after cephalic amplexus; heremained close to the ,. When the , began tooviposit, the < moved to face away fromthe , by putting his cloaca in contact withthe ,’s cloaca.
Parental care
A typical sequence is indicated in Fig. 2J–L.Parental care was performed exclusively bythe <. The < sat near the clutch, defending itfrom ,, (393 occurrences; n5 3 <<;762 hours of observations) and moistening it(40 occurrences; n5 3 <<). He only left theclutch (for short periods of time) in order toeat. The time interval that a < remained overthe clutch was 5–55minutes (�x5 35 � 13 � 8,40 occurrences; n5 3 <<). The < usually satlow to the ground and his venter was incontact with the eggs. He periodically mois-tened the eggs by releasing fluid from hiscloaca (Plate 1B). When the clutch was large,the <’s limbs also covered the eggs; if it wasnot as large as his venter, his limbs borderedthe clutch. The < periodically rotated overthe clutch; intervals between rotations were1–80 seconds (�x5 27 � 6 � 26, 88 occur-rences; n5 3 <<). Several times a < aban-doned a clutch, removed eggs from a clutch(3 occurrences; n5 3 <<) or ate eggs (eitherall or part of the clutch) (3 occurrences; n5 3<<). Fungi invaded a clutch when a <abandoned it.
At the time of egg hatching the < bent overthe clutch, with his dorsum curved convexly.The tadpoles climbed onto the dorsum of the< at his flanks, inguinal region or legs. At thesame time, the < moved over the clutch,intervals between rotations were 3–80 seconds(�x5 29 � 3 � 25 � 7, 73 occurrences; n5 3
84 AMPHIBIAN CONSERVATION
Int. Zoo Yb. (2008) 42: 78–89. c� 2008 The Authors. Journal compilation c� 2008 The Zoological Society of London
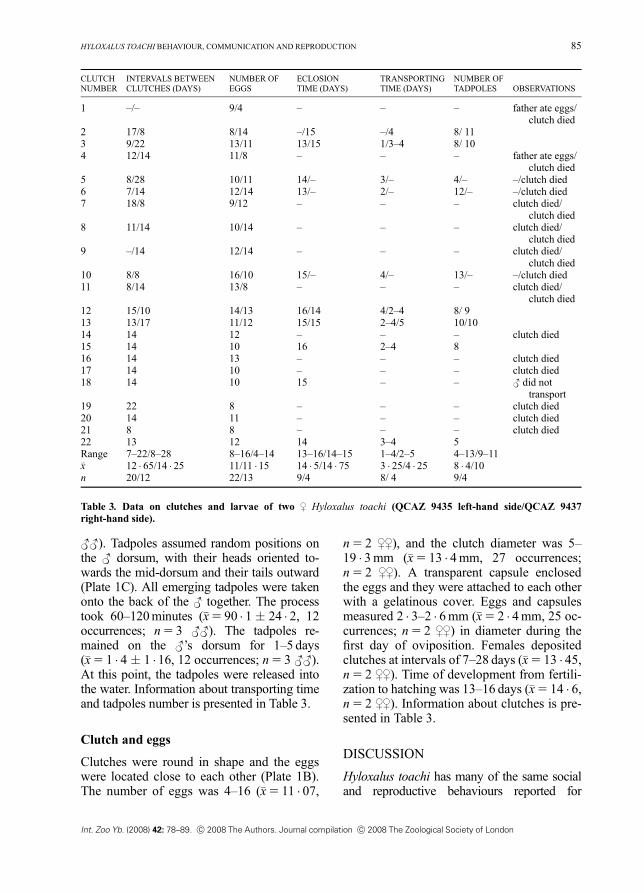
<<). Tadpoles assumed random positions onthe < dorsum, with their heads oriented to-wards the mid-dorsum and their tails outward(Plate 1C). All emerging tadpoles were takenonto the back of the < together. The processtook 60–120minutes (�x5 90 � 1 � 24 � 2, 12occurrences; n5 3 <<). The tadpoles re-mained on the <’s dorsum for 1–5 days(�x5 1 � 4 � 1 �16, 12 occurrences; n5 3 <<).At this point, the tadpoles were released intothe water. Information about transporting timeand tadpoles number is presented in Table 3.
Clutch and eggs
Clutches were round in shape and the eggswere located close to each other (Plate 1B).The number of eggs was 4–16 (�x5 11 � 07,
n5 2 ,,), and the clutch diameter was 5–19 � 3mm (�x5 13 � 4mm, 27 occurrences;n5 2 ,,). A transparent capsule enclosedthe eggs and they were attached to each otherwith a gelatinous cover. Eggs and capsulesmeasured 2 � 3–2 � 6mm (�x5 2 � 4mm, 25 oc-currences; n5 2 ,,) in diameter during thefirst day of oviposition. Females depositedclutches at intervals of 7–28 days (�x5 13 � 45,n5 2 ,,). Time of development from fertili-zation to hatching was 13–16 days (�x5 14 � 6,n5 2 ,,). Information about clutches is pre-sented in Table 3.
DISCUSSION
Hyloxalus toachi has many of the same socialand reproductive behaviours reported for
CLUTCHNUMBER
INTERVALS BETWEENCLUTCHES (DAYS)
NUMBER OFEGGS
ECLOSIONTIME (DAYS)
TRANSPORTINGTIME (DAYS)
NUMBER OFTADPOLES OBSERVATIONS
1 –/– 9/4 – – – father ate eggs/clutch died
2 17/8 8/14 –/15 –/4 8/ 113 9/22 13/11 13/15 1/3–4 8/ 104 12/14 11/8 – – – father ate eggs/
clutch died5 8/28 10/11 14/– 3/– 4/– –/clutch died6 7/14 12/14 13/– 2/– 12/– –/clutch died7 18/8 9/12 – – – clutch died/
clutch died8 11/14 10/14 – – – clutch died/
clutch died9 –/14 12/14 – – – clutch died/
clutch died10 8/8 16/10 15/– 4/– 13/– –/clutch died11 8/14 13/8 – – – clutch died/
clutch died12 15/10 14/13 16/14 4/2–4 8/ 913 13/17 11/12 15/15 2–4/5 10/1014 14 12 – – – clutch died15 14 10 16 2–4 816 14 13 – – – clutch died17 14 10 – – – clutch died18 14 10 15 – – < did not
transport19 22 8 – – – clutch died20 14 11 – – – clutch died21 8 8 – – – clutch died22 13 12 14 3–4 5Range 7–22/8–28 8–16/4–14 13–16/14–15 1–4/2–5 4–13/9–11�x 12 � 65/14 � 25 11/11 �15 14 � 5/14 � 75 3 � 25/4 � 25 8 � 4/10n 20/12 22/13 9/4 8/ 4 9/4
Table 3. Data on clutches and larvae of two , Hyloxalus toachi (QCAZ 9435 left-hand side/QCAZ 9437right-hand side).
HYLOXALUS TOACHI BEHAVIOUR, COMMUNICATION AND REPRODUCTION 85
Int. Zoo Yb. (2008) 42: 78–89. c� 2008 The Authors. Journal compilation c� 2008 The Zoological Society of London

Hyloxalus, Allobates, Colostethus, Epipedo-bates, Mannophryne and Rheobates (seeCrump, 1995, Grant et al., 2006, and Lotterset al., 2007, for general reviews). Hyloxalustoachi is particularly similar to six species ofdendrobatids: H. awa, Hyloxalus sp (as Phyllo-bates sp), H. azureiventris, Mannophryne col-laris, Mannophryne trinitatis and Rheobatespalmatus (Mudrack, 1969; Durant & Dole,1975; Wells, 1980a; Correa, 1995; Luddecke,1999; Lotters et al., 2000). In all these species,,, appear to defend territories and act aggres-sively. The << initially court ,, from a dis-tance and do not use close-range tactilestimulation until at the actual oviposition site(Bourne et al., 2001, and references therein).
Parental-care behaviours recorded inH. toachi differ from some other dendrobatids,with regard to which gender cares for the eggs,frequency of < attendance, duration of tadpolesstaying on the parent’s dorsum and other fea-tures [see also Grant et al. (2006) for a discus-sion on sex of nurse frogs and Lotters et al.(2007) for parental care]. In H. toachi, the <<carry the tadpoles. The latter behaviour con-trasts with the sister species H. awa andHyloxalus sp that receive biparental care (Mu-drack, 1969; Correa, 1995). The dark colourchange of the calling << reported for H. toachialso occurs inM. trinitatis and R. palmatus.
Observations of captive H. toachi haverevealed interesting additional details regard-ing amplexus and fertilization, for example,the uncoupling of amplexus from oviposit-ing that occurs in some dendrobatid frogs(Bourne et al., 2001; Grant et al., 2006;Castillo-Trenn & Coloma, 2008). AmongHyloxalus, the most remarkable differenceoccurs between H. toachi and H. azureiven-tris, because the latter lacks amplexus. Inmost anurans, << grasp the ,, in either anaxillary or an inguinal position while ovipo-siting (Weygoldt, 1987). In contrast, inH. toachi amplexus takes place before egglaying and is in a cephalic position. Duringoviposition, the < is either situated above the, with his pads on the ground or abuts to the, so that both cloacal openings are in contactwith each other. In both cases, the , laysher eggs without being grasped by the <. In
H. toachi, behaviours related to the time offertilization are described herein. Crump(1972), Weygoldt (1987), Jungfer (1985)and Jungfer et al. (1996) indicated the timewhen fertilization occurred in Dendrobatesgranuliferus, Dendrobates tinctorius, Den-drobates speciosus and Dendrobates vicentei.Spermatoposition occurs before the , hasstarted laying and right before the < leavesthe oviposition site. The , only starts layingafter that. In other studies, some vague datahave been presented (Silverstone, 1976; Les-cure, 1984; Weygoldt, 1987; Zimmermann,1989; Summers, 1992; Lotters et al., 2000).
Cephalic amplexus may serve two roles inH. toachi: stimulatory and informational.Cephalic amplexus always occurs beforesuccessful oviposition, suggesting that itmay have some function in stimulating the ,to release eggs. A stimulatory role is alsosupported by observations of multiple boutsof cephalic amplexus before oviposition inspecies of Epipedobates and Anomaloglossus(Correa, 1995; Junca, 1998; this study), andobservations in other anurans. For example, ,Osteocephalus oophagus will not lay eggswithout being clasped by a <, although theeggs only serve as food for the larvae andfertilization is unnecessary (Jungfer & Schie-sari, 1995). Female H. toachi vary consider-ably in the size of clutch they produce (4–16eggs) and another possibility is that someaspect of cephalic amplexus, such as intensityor stimulus, may influence the number ofeggs a , might lay. Prolonged courtship hasbeen suggested to be a means by which <<determine that a , is receptive (Wells, 1977).Also, in this study, a , sometimes left aftercephalic amplexus but before oviposition.This raises the possibility that the amplexusmay also convey additional informationabout < quality. Observations on H. toachiof egg laying without simultaneous cephalicamplexus are intriguing but, clearly, moreobservations are necessary to determine howwidespread the uncoupling of amplexus andoviposition is among species of Hyloxalus,as well as to determine how it evolvedamong Dendrobatidae (see discussion inCastillo-Trenn & Coloma, 2008).
86 AMPHIBIAN CONSERVATION
Int. Zoo Yb. (2008) 42: 78–89. c� 2008 The Authors. Journal compilation c� 2008 The Zoological Society of London

Information on , interclutch intervalsis rare (Tsuji & Lue, 2000). An importantresult of this study is data on the interclutchintervals of , H. toachi. Captive ,, of thisspecies laid eggs as frequently as once aweek. While this is not the shortest inter-clutch interval recorded for any frog species(see Tsuji & Lue, 2000), it is quite differentfrom other published data on a species ofDendrobatidae. The shortest known inter-clutch interval for field-studied , Colostethuspanamensis (as Colostethus inguinalis) is61 days comparing observations betweencaptive and field animals and, clearly, thedata from this study need to be supplementedby field observations. Nonetheless, a 1weekegg-laying interval is now known to bebiologically possible for a species of Hylox-alus and could be used in broader, moretheoretical studies that attempt to modelthe evolution of complex reproductive beha-viour in dendrobatid frogs (e.g. Summers &Earn, 1999).
Female encounter calls have been reportedin Allobates femoralis (Weygoldt, 1980),C. panamensis (Wells, 1980b), H. awa andEpipedobates machalilla (Correa, 1995), andherein, but , vocalization may turn out bemore common, when more detailed studiesare carried out.
Studies of captive animals offer a uniqueopportunity for documenting intraspecificbehavioural variation because of the ease ofobservation and the possibility of obtaininglarge behavioural sample sizes. Aspects ofsocial behaviour are often divided into quali-tative, discrete character states for inter-specific phylogenetic analysis or for mappingthe evolution of behaviour on an indepen-dently derived phylogenetic hypothesis. Theintraspecific variation in behaviour documen-ted in this study (Table 1) suggests caution inconstructing such qualitative, discrete inter-specific character categories. For example,H. toachi was observed on different occa-sions in varying postures during oviposition.Those different postures are usually thoughtto represent interspecific differences in beha-viour rather than intraspecific variation owingto polymorphisms.
Sex roles in dendrobatid frogs appear to bemore variable than among most other anuranspecies. In a number of species, includingH. toachi, , frogs are aggressive and haveterritories that they actively defend. Usinginterspecific field comparisons, Summers(1992) found support for a causal relationshipbetween < parental care and , aggression intwo species of Dendrobatinae. An importantnext step in understanding the behaviourobserved in captive H. toachi would be fieldstudies examining whether , H. toachi suffera cost, such as reduced < parental care, iftheir mate also mates with other ,,.
ACKNOWLEDGEMENTSThanks to Gloria M. Correa, who shared her knowledgewith us and made valuable suggestions during thisresearch. She and Nestor Acosta-Buenano helpedwith fieldwork. Cristina Novoa-Felix helped with layoutof figures. Eugenia M. del Pino, Violeta Rafael, membersof the AQ thesis committee, Michele Auber-Thomay, andMartha L. Crump and two anonymous reviewers madehelpful comments. This manuscript also benefited froma critical review by Sharon B. Emerson. We are indebtedto Sharon B. Emerson and Giovanni Onore for theirencouragement to publish this study.
PRODUCTS MENTIONED IN THE TEXTMarantz Model CP 430: portable cassette recorder,manufactured by Marantz, Kanagawa, Japan. http://www.marantz.comSennheiser K6: microphone, manufactured by Sennhei-ser electronic GmbH & Co. KG, 30900 Wedemark,Germany. http://www.sennheiser.comSony SRmetallic tape Type IV: cassettes, manufacturedby Sony Corporation, Minato-ku, Tokyo, Japan. http://www.sony.net
REFERENCESBECK, A., PETERSON, A., SENSENDORF, J., CHAPLIN, M.,POMPA, M. & KONAR, M. (1992): SoundEdit pro 1.0. SanFrancisco, CA: Macromind–Paracomp.BOURNE, G. R., COLLINS, A. C., HOLDER A. M. &
MCCARTHY C. L. (2001): Vocal communication andreproductive behavior of the frog Colostethus beebei inGuyana. Journal of Herpetology 35: 272–281.CASTILLO-TRENN, P. & COLOMA, L. A. (In press): Noteson behaviour and reproduction in captive Allobateskingsburyi (Anura: Dendrobatidae), with comments onevolution of reproductive amplexus. International ZooYearbook 42. DOI:10.1111/j.1748-1090.2007.00039.x.COLOMA, L. A. (1995): Ecuadorian frogs of the genusColostethus (Anura: Dendrobatidae). University ofKansas, Natural History Museum, Miscellaneous Pub-lications 87: 1–72.
HYLOXALUS TOACHI BEHAVIOUR, COMMUNICATION AND REPRODUCTION 87
Int. Zoo Yb. (2008) 42: 78–89. c� 2008 The Authors. Journal compilation c� 2008 The Zoological Society of London

CORREA, G. M. (1995): Comportamiento, vocalizacion ynotas biologicas de Colostethus awa y C. machalilla(Anura: Dendrobatidae) en cautiverio. Tesis de Licen-ciatura, Pontificia Universidad Catolica del Ecuador,Quito.CRUMP, M. L. (1972): Territoriality and mating behaviorin Dendrobates granuliferus (Anura: Dendrobatidae).Herpetologica 28: 195–198.CRUMP, M. L. (1995): Parental care. In Amphibianbiology 2. Social behaviour: 518–567. Heatwole, H. &Sullivan, B. K. (Eds). Chipping Norton, NSW: Surrey,Beatty and Sons.DUELLMAN, W. E. & TRUEB, L. (1986): Biology of amphi-bians. New York, NY: McGraw-Hill Book Co.DURANT, P. & DOLE, J. W. (1975): Aggressive behavior inColostethus (Prostherapis) collaris (Anura: Dendrobati-dae). Herpetologica 31: 23–36.FANDINO, M. C., LUDDECKE, H. & AMEZQUITA, A. (1997):Vocalization and larval transportation of male Colos-tethus subpunctatus (Anura: Dendrobatidae). Amphibia–Reptilia 18: 39–48.GRANT, T., FROST, D. R., CALDWELL, J. P., GAGLIARDO, R.,HADDAD, C. F. B., KOK, P. J. R., MEANS, D. B., NOONAN,B. P., SCHARGEL, W. E. & WHEELER, W. C. (2006):Phylogenetic systematics of dart-poison frogs and theirrelatives (Amphibia: Athesphatanura: Dendrobatidae).Bulletin of the American Museum of Natural History299: 262.HEYER, W. R., RAND, A. S., GONCALVES DA CRUZ, C. A.,PEIXOTO O. L. & NELSON C. E. (1990): Frogs of Boraceia.Arquivos de Zoologia 31: 237–358.IUCN (2006): 2006 IUCN red list of threatenedspecies. Gland and Cambridge: IUCN http://www.iucnredlist.org/.JUNCA, F. A. (1998): Reproductive biology of Colostethusstepheni and Colostethus marchesianus (Dendrobatidae),with the description of a new anuran mating behavior.Herpetologica 54: 377–387.JUNGFER, K.-H. (1985): Beitrag zur Kenntnis vonDendrobates speciosus O. Schmidt, 1857. Salamandra21: 263–280.JUNGFER, K.-H. & SCHIESARI, L. C. (1995): Description ofa Central Amazonian and Guianan tree frog, genusOsteocephalus (Anura, Hylidae), with oophagustadpoles. Alytes 13(1): 1–13.JUNGFER, K.-H., WEYGOLDT, P. & JURASKE, N. (1996):Dendrobates vicentei, ein neuer Pfeilgiftfrosch ausZentral-Panama. Herpetofauna 18(103): 17–26.KNELLER, M. (1987): Beobachtungen an Phyllobatesazureiventris im naturlichen Lebensraum und imTerrarium. Herpetofauna 9(50): 6–8.LESCURE, J. (1984): Las larvas de dendrobatidae. ReunionIberoamericana de Conservacion y Zoologıa de Vertebrados Actas II: 37–45.LOTTERS, S., JUNGFER, K.-H. & WIDMER, A. (2000): A newgenus of aposematic poison frog (Amphibia: Anura:Dendrobatidae) from the upper Amazon Basin with noteson its reproductive behaviour and tadpole morphology.Jahreshefte der Gesellschaft fur Naturkunde in Wurttem-berg 156: 233–243.
LOTTERS, S., JUNGFER, K.-H., HENKEL, F. W. & SCHMIDT, W.(2007): Poison frogs. Biology, species captive husban-dry. Frankfurt am Main: Edition Chimaira.LUDDECKE, H. (1999): Behavioral aspects of the repro-ductive biology of the Andean frog Colostethus palma-tus. Revista de la Academia Colombiana de CienciasExactas, Fısicas y Naturales 23: 303–316.MUDRACK, W. (1969): Pflege und Zucht eines Blattstei-gerfrosches der Gattung Phyllobates aus Ecuador. Sala-mandra 5: 81–84.MYERS, C. W. & DALY, J. W. (1983): Dart-poison frogs.Scientific American 248: 120–133.MYERS, C. W., PAOLILLO, A. & DALY, J. W. (1991):Discovery of a defensively malodorous and nocturnalfrog in the family Dendrobatidae: phylogenetic signifi-cance of a new genus and species from the VenezuelanAndes. American Museum Novitates 3002: 1–33.RICCI, A. (1994): Sound effects V. 0.9.1. Torino, Italy:Corso De Gasperi.SANTOS, J. C., COLOMA, L. A. & CANNATELLA, D. C.(2003): Multiple, recurring origins of aposematism anddiet specialization in poison frogs. Proceedings of theNational Academy of Sciences United States of America100: 12792–12797.SILVERSTONE, P. A. (1976): A revision of the poison-arrowfrogs of the genus Phyllobates Bibron in Sagra (FamilyDendrobatidae). Natural History Museum of Los AngelesCounty Science Bulletin 27: 1–53.SUMMERS, K. (1992): Mating strategies in two species ofdart-poison frogs: a comparative study. Animal Beha-viour 43: 1–13.SUMMERS, K. & EARN, D. (1999): The cost of polygynyand the evolution of female care in poison frogs.Biological Journal of the Linnean Society 66:515–538.TSUJI, H. & LUE, K. Y. (2000): The reproductive ecologyof female Rana (Limnonectes) kuhlii, a fanged frog ofTaiwan, with particular emphasis on multiple clutches.Herpetologica 56(2): 153–165.WELLS, K. D. (1977): The courtship of frogs. In Thereproductive biology of amphibians: 233–262. Taylor, D.H. & Guttman, S. I. (Eds). New York, NY: Plenum Press.WELLS, K. D. (1980a): Social behavior and communica-tion of a dendrobatid frog (Colostethus trinitatis). Herpe-tologica 26: 189–199.WELLS, K. D. (1980b): Behavioral ecology and socialorganization of a dendrobatid frog (Colostethus inguina-lis). Behavior, Ecology and Sociobiology 6: 199–209.WEYGOLDT, P. (1980): Zur Fortpflanzungsbiologie vonPhyllobates femoralis (Boulenger) im Terrarium. Sala-mandra 16: 215–226.WEYGOLDT, P. (1987): Evolution of parental care in dartpoison frogs (Amphibia: Anura: Dendrobatidae). Zeits-chrift fuer Zoologische Systematik und Evolutions-forschung 25: 51–67.ZIMMERMANN, H. (1989): Conservation studies on the‘‘dart poison’’ frogs Dendrobatidae in the field and incaptivity. International Zoo Yearbook 28: 31–44.ZIMMERMANN, H. & ZIMMERMANN, E. (1981): Sozialver-halten, Fortpflanzungsverhalten und Zucht der
88 AMPHIBIAN CONSERVATION
Int. Zoo Yb. (2008) 42: 78–89. c� 2008 The Authors. Journal compilation c� 2008 The Zoological Society of London

Farberfrosche Dendrobates histrionicus und D. lehmannisowie einiger anderer Dendrobatiden. Zeitschrift desKolner Zoo 24(3): 83–99.ZIMMERMANN, H. & ZIMMERMANN, E. (1988): Etho-Tax-onomie und zoogeographische Artengruppenbildung beiPfeilgiftfroschen (Anura: Dendrobatidae). Salamandra24: 125–160.
Manuscript submitted 18 June 2007; revised 11December 2007; accepted 30 January 2008
Supplementary material
The following supplementary material isavailable for this article online:
Abstract S1. Spanish abstract.This material is available as part of the onlinearticle from: http://www.blackwell-synergy.com/doi/abs/10.1111/j.1748-1090.2007.00051.x(This link will take you to the article abstract).Please note: Blackwell Publishing are notresponsible for the content or functionalityof any supplementary materials supplied bythe authors. Any queries (other than missingmaterial) should be directed to the corre-sponding author of the article.
HYLOXALUS TOACHI BEHAVIOUR, COMMUNICATION AND REPRODUCTION 89
Int. Zoo Yb. (2008) 42: 78–89. c� 2008 The Authors. Journal compilation c� 2008 The Zoological Society of London




















