
Election turnout in '84 reverses 20-year trend Reagan's re ...
Upload
ua-birminghamCategory
view
0download
0
Normalisation of calcium status reverses the phenotype indentin, but not in enamel of VDR-deficient mice
Xueming Zhang a,*, Firoz Rahemtulla a, Ping Zhang b, Xingsheng Li c, Preston Beck a,Huw F. Thomas b
aDepartment of Prosthodontics, School of Dentistry, University of Alabama at Birmingham, Birmingham, AL 35294-2170, USAbDepartment of Pediatric Dentistry, School of Dentistry, University of Alabama at Birmingham, Birmingham, AL, USAcDepartment of Nutrition Science, School of Health Related Professional, University of Alabama at Birmingham, Birmingham, AL, USA
a r c h i v e s o f o r a l b i o l o g y 5 4 ( 2 0 0 9 ) 1 1 0 5 – 1 1 1 0
a r t i c l e i n f o
Article history:
Accepted 17 September 2009
Keywords:
Vitamin D receptor
Enamel
Dentin
Mineralisation
a b s t r a c t
Objective: To determine the effects of vitamin D receptor (VDR) deficiency on mouse dentin
and enamel mineralisation, and how normalisation of serum calcium level affects dentin
and enamel phenotypes in VDR knockout mice.
Materials and methods: Groups of VDR wild-type (VDR+/+), VDR deficient (VDR�/�) and
VDR�/� rescued mice were sacrificed at 70.5 days of life. The rescued group was established
by a high-calcium diet feeding the VDR�/�mice from postnatal 19 days. Micro-CT was used
to compare enamel and dentin mineralisation density (MD) at different levels of mandibular
incisors among the groups. The scanning electron microscope (SEM) was used to examine
the ultrastructure of the enamel and dentin in the corresponding levels and of surface
enamel after acidic treatment.
Results: Micro-CT showed that in VDR�/� rescued group, dentin phenotype was reversed
and dentin MD was reversed to normal; however, enamel mineralisation was not reversible,
and remained as hypermineralisation in molar region and apical region of the incisors. SEM
also revealed enamel hypermineralisation in the VDR�/� rescued group. This early enamel
hypermineralisation was more susceptible to acidic erosion.
Conclusion: Vitamin D affects dentin mineralisation systemically, and it regulates enamel
mineralisation locally.
Published by Elsevier Ltd.
avai lab le at www.sc iencedi rect .com
journal homepage: www.intl.elsevierhealth.com/journals/arob
The vitamin D endocrine system is essential for calcium
homeostasis in mammals. Mineral homeostasis is system-
atically regulated through intestinal calcium absorption and
renal re-absorption into the circulatory system. The classic
pathway of vitamin D function is through binding with the
vitamin D receptor (VDR) in the intestine and the kidney that
govern calcium transport in these organs. The fluctuation of
serum calcium level affects bone mineralisation density (BMD)
through trapping and mobilisation of calcium and phosphorus
in bone.1
Mineralised tissue results from a complex series of well-
coordinated processes that requires optimal function of
* Corresponding author. Tel.: +1 205 934 7823; fax: +1 205 934 6108.E-mail address: [email protected] (X. Zhang).
0003–9969/$ – see front matter . Published by Elsevier Ltd.doi:10.1016/j.archoralbio.2009.09.004
numerous genes and proteins. Enamel, dentin and bone are
major tissues in mammals that develop through matrix-
mediated mineralisation processes. However, the origin and
composition, as well as the remodelling potential of these
tissues are different. For example, the mineralisation density
(MD) of the bone relies on a delicate balance between
osteoblast and osteoclast activity, whereas resorption of
enamel and dentin are processes unrelated to their forma-
tion. Furthermore, BMD varies with changes in serum
calcium levels, while the MD in unerupted enamel and
dentin remains at the level attained during their develop-
ment.2,3
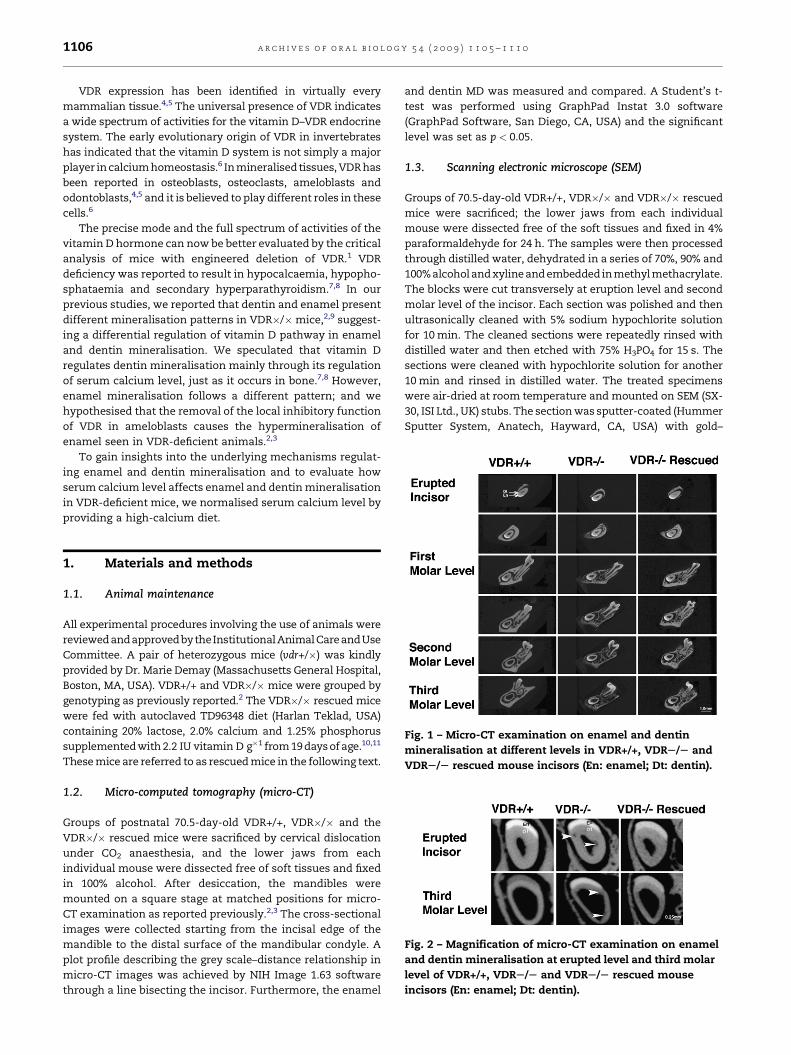
Fig. 2 – Magnification of micro-CT examination on enamel
and dentin mineralisation at erupted level and third molar
level of VDR+/+, VDRS/S and VDRS/S rescued mouse
incisors (En: enamel; Dt: dentin).
Fig. 1 – Micro-CT examination on enamel and dentin
mineralisation at different levels in VDR+/+, VDRS/S and
VDRS/S rescued mouse incisors (En: enamel; Dt: dentin).
a r c h i v e s o f o r a l b i o l o g y 5 4 ( 2 0 0 9 ) 1 1 0 5 – 1 1 1 01106
VDR expression has been identified in virtually every
mammalian tissue.4,5 The universal presence of VDR indicates
a wide spectrum of activities for the vitamin D–VDR endocrine
system. The early evolutionary origin of VDR in invertebrates
has indicated that the vitamin D system is not simply a major
player in calcium homeostasis.6 In mineralised tissues, VDR has
been reported in osteoblasts, osteoclasts, ameloblasts and
odontoblasts,4,5 and it is believed to play different roles in these
cells.6
The precise mode and the full spectrum of activities of the
vitamin D hormone can now be better evaluated by the critical
analysis of mice with engineered deletion of VDR.1 VDR
deficiency was reported to result in hypocalcaemia, hypopho-
sphataemia and secondary hyperparathyroidism.7,8 In our
previous studies, we reported that dentin and enamel present
different mineralisation patterns in VDR�/�mice,2,9 suggest-
ing a differential regulation of vitamin D pathway in enamel
and dentin mineralisation. We speculated that vitamin D
regulates dentin mineralisation mainly through its regulation
of serum calcium level, just as it occurs in bone.7,8 However,
enamel mineralisation follows a different pattern; and we
hypothesised that the removal of the local inhibitory function
of VDR in ameloblasts causes the hypermineralisation of
enamel seen in VDR-deficient animals.2,3
To gain insights into the underlying mechanisms regulat-
ing enamel and dentin mineralisation and to evaluate how
serum calcium level affects enamel and dentin mineralisation
in VDR-deficient mice, we normalised serum calcium level by
providing a high-calcium diet.
1. Materials and methods
1.1. Animal maintenance
All experimental procedures involving the use of animals were
reviewed andapproved bytheInstitutional Animal CareandUse
Committee. A pair of heterozygous mice (vdr+/�) was kindly
provided by Dr. Marie Demay (Massachusetts General Hospital,
Boston, MA, USA). VDR+/+ and VDR�/�mice were grouped by
genotyping as previously reported.2 The VDR�/� rescued mice
were fed with autoclaved TD96348 diet (Harlan Teklad, USA)
containing 20% lactose, 2.0% calcium and 1.25% phosphorus
supplemented with 2.2 IU vitamin D g�1 from 19 days of age.10,11
These mice are referred to as rescued mice in the following text.
1.2. Micro-computed tomography (micro-CT)
Groups of postnatal 70.5-day-old VDR+/+, VDR�/� and the
VDR�/� rescued mice were sacrificed by cervical dislocation
under CO2 anaesthesia, and the lower jaws from each
individual mouse were dissected free of soft tissues and fixed
in 100% alcohol. After desiccation, the mandibles were
mounted on a square stage at matched positions for micro-
CT examination as reported previously.2,3 The cross-sectional
images were collected starting from the incisal edge of the
mandible to the distal surface of the mandibular condyle. A
plot profile describing the grey scale–distance relationship in
micro-CT images was achieved by NIH Image 1.63 software
through a line bisecting the incisor. Furthermore, the enamel
and dentin MD was measured and compared. A Student’s t-
test was performed using GraphPad Instat 3.0 software
(GraphPad Software, San Diego, CA, USA) and the significant
level was set as p < 0.05.
1.3. Scanning electronic microscope (SEM)
Groups of 70.5-day-old VDR+/+, VDR�/� and VDR�/� rescued
mice were sacrificed; the lower jaws from each individual
mouse were dissected free of the soft tissues and fixed in 4%
paraformaldehyde for 24 h. The samples were then processed
through distilled water, dehydrated in a series of 70%, 90% and
100% alcohol and xyline and embedded inmethyl methacrylate.
The blocks were cut transversely at eruption level and second
molar level of the incisor. Each section was polished and then
ultrasonically cleaned with 5% sodium hypochlorite solution
for 10 min. The cleaned sections were repeatedly rinsed with
distilled water and then etched with 75% H3PO4 for 15 s. The
sections were cleaned with hypochlorite solution for another
10 min and rinsed in distilled water. The treated specimens
were air-dried at room temperature and mounted on SEM (SX-
30, ISI Ltd., UK) stubs. The section was sputter-coated (Hummer
Sputter System, Anatech, Hayward, CA, USA) with gold–
a r c h i v e s o f o r a l b i o l o g y 5 4 ( 2 0 0 9 ) 1 1 0 5 – 1 1 1 0 1107
palladium, and viewed under SEM with an accelerating 15 kV.
The images were taken at 4000� magnification, and then
compared at the matched locations among the groups.
1.4. Surface erosion
Groups of 70.5-day-old VDR+/+, VDR�/� and VDR�/� rescued
mice were sacrificed, the lower jaws from each individual
mouse was dissected free of the soft tissues and fixed in 4%
paraformaldehyde for 24 h. The samples were then ultra-
sonically cleaned in 5% sodium hypochlorite solution. The
cleaned incisors were repeatedly rinsed in distilled water and
then etched with 75% H3PO4 for 15 s. They were cleaned with
hypochlorite solution again and rinsed in distilled water. The
treated specimens were air-dried at room temperature and
mounted for SEM examination as mentioned above.
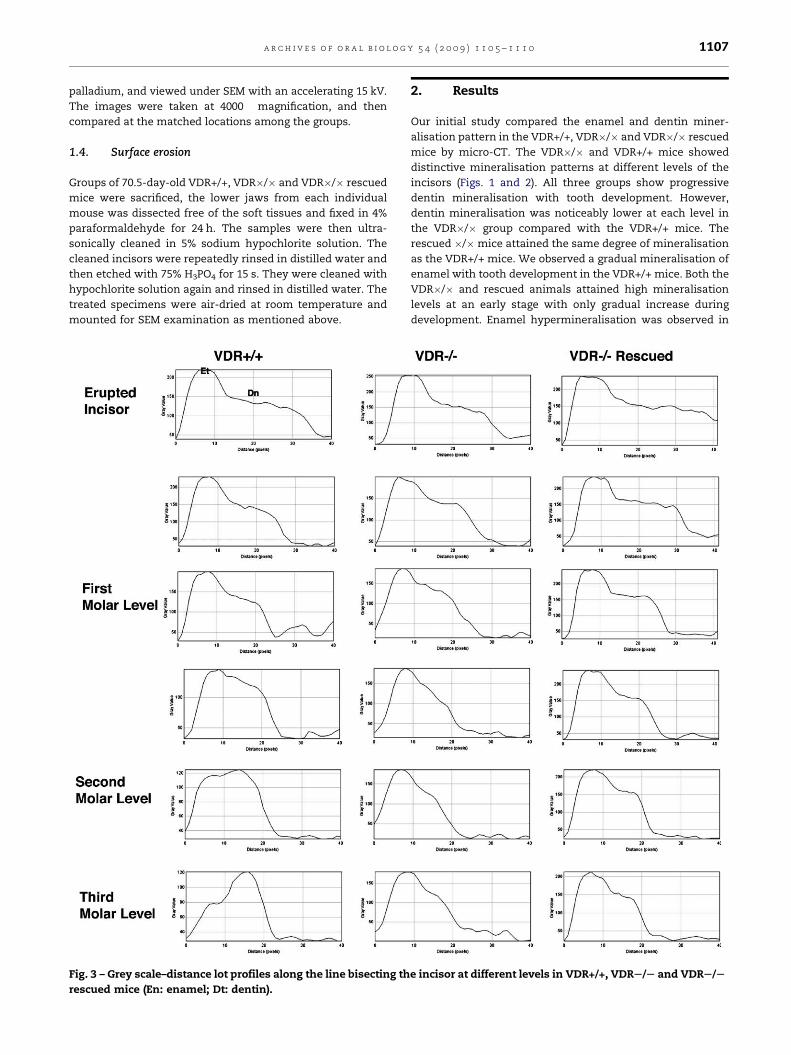
Fig. 3 – Grey scale–distance lot profiles along the line bisecting th
rescued mice (En: enamel; Dt: dentin).
2. Results
Our initial study compared the enamel and dentin miner-
alisation pattern in the VDR+/+, VDR�/� and VDR�/� rescued
mice by micro-CT. The VDR�/� and VDR+/+ mice showed
distinctive mineralisation patterns at different levels of the
incisors (Figs. 1 and 2). All three groups show progressive
dentin mineralisation with tooth development. However,
dentin mineralisation was noticeably lower at each level in
the VDR�/� group compared with the VDR+/+ mice. The
rescued �/�mice attained the same degree of mineralisation
as the VDR+/+ mice. We observed a gradual mineralisation of
enamel with tooth development in the VDR+/+ mice. Both the
VDR�/� and rescued animals attained high mineralisation
levels at an early stage with only gradual increase during
development. Enamel hypermineralisation was observed in
e incisor at different levels in VDR+/+, VDRS/S and VDRS/S
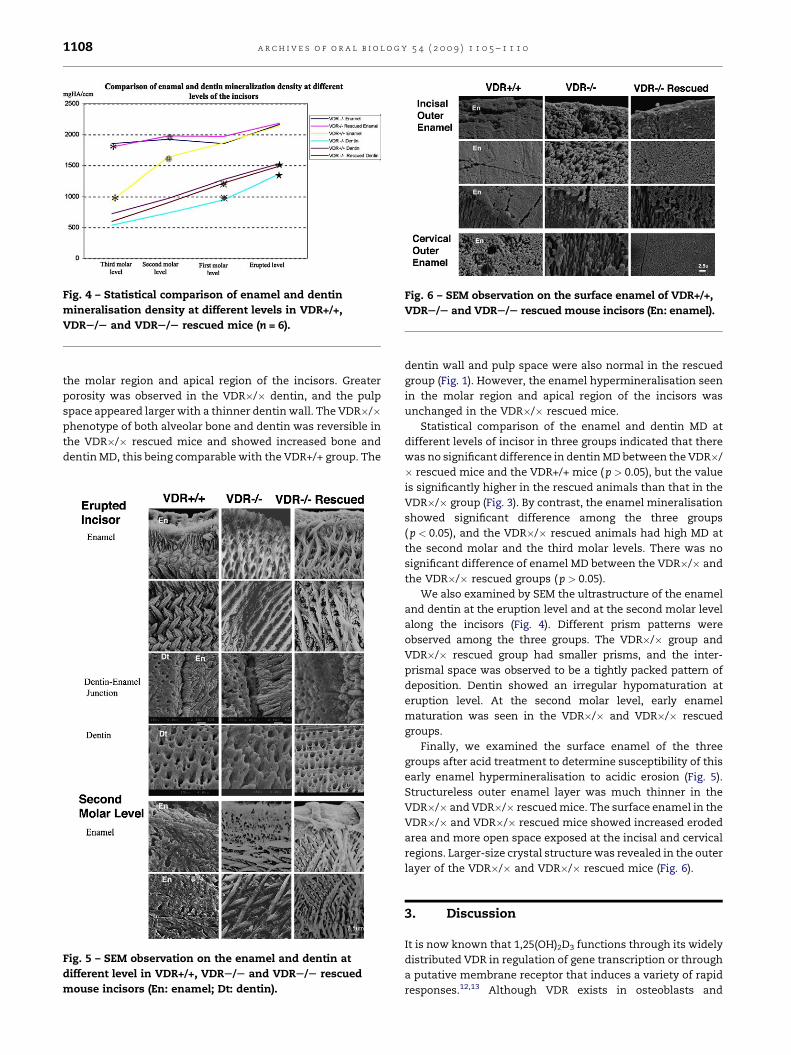
Fig. 4 – Statistical comparison of enamel and dentin
mineralisation density at different levels in VDR+/+,
VDRS/S and VDRS/S rescued mice (n = 6).
Fig. 6 – SEM observation on the surface enamel of VDR+/+,
VDRS/S and VDRS/S rescued mouse incisors (En: enamel).
a r c h i v e s o f o r a l b i o l o g y 5 4 ( 2 0 0 9 ) 1 1 0 5 – 1 1 1 01108
the molar region and apical region of the incisors. Greater
porosity was observed in the VDR�/� dentin, and the pulp
space appeared larger with a thinner dentin wall. The VDR�/�phenotype of both alveolar bone and dentin was reversible in
the VDR�/� rescued mice and showed increased bone and
dentin MD, this being comparable with the VDR+/+ group. The
Fig. 5 – SEM observation on the enamel and dentin at
different level in VDR+/+, VDRS/S and VDRS/S rescued
mouse incisors (En: enamel; Dt: dentin).
dentin wall and pulp space were also normal in the rescued
group (Fig. 1). However, the enamel hypermineralisation seen
in the molar region and apical region of the incisors was
unchanged in the VDR�/� rescued mice.
Statistical comparison of the enamel and dentin MD at
different levels of incisor in three groups indicated that there
was no significant difference in dentin MD between the VDR�/
� rescued mice and the VDR+/+ mice ( p > 0.05), but the value
is significantly higher in the rescued animals than that in the
VDR�/� group (Fig. 3). By contrast, the enamel mineralisation
showed significant difference among the three groups
( p < 0.05), and the VDR�/� rescued animals had high MD at
the second molar and the third molar levels. There was no
significant difference of enamel MD between the VDR�/� and
the VDR�/� rescued groups (p > 0.05).
We also examined by SEM the ultrastructure of the enamel
and dentin at the eruption level and at the second molar level
along the incisors (Fig. 4). Different prism patterns were
observed among the three groups. The VDR�/� group and
VDR�/� rescued group had smaller prisms, and the inter-
prismal space was observed to be a tightly packed pattern of
deposition. Dentin showed an irregular hypomaturation at
eruption level. At the second molar level, early enamel
maturation was seen in the VDR�/� and VDR�/� rescued
groups.
Finally, we examined the surface enamel of the three
groups after acid treatment to determine susceptibility of this
early enamel hypermineralisation to acidic erosion (Fig. 5).
Structureless outer enamel layer was much thinner in the
VDR�/� and VDR�/� rescued mice. The surface enamel in the
VDR�/� and VDR�/� rescued mice showed increased eroded
area and more open space exposed at the incisal and cervical
regions. Larger-size crystal structure was revealed in the outer
layer of the VDR�/� and VDR�/� rescued mice (Fig. 6).
3. Discussion
It is now known that 1,25(OH)2D3 functions through its widely
distributed VDR in regulation of gene transcription or through
a putative membrane receptor that induces a variety of rapid
responses.12,13 Although VDR exists in osteoblasts and

a r c h i v e s o f o r a l b i o l o g y 5 4 ( 2 0 0 9 ) 1 1 0 5 – 1 1 1 0 1109
osteoclasts, bone mineralisation and bone remodelling are
believed to be independent of VDR local expression; rather, it is
regulated by the serum calcium level.14 Several studies have
shown that serum calcium level in the VDR�/� mice can be
reversed by feeding with a high-calcium diet.10,11 It has been
reported that 1,25(OH)2D3 down-regulates cadherin-17 and
aquaporin-8. Thus, the accumulation of 1,25(OH)2D3 can route
calcium absorption through a paracellular path in the intestine,
which contributes to normalisation of serum calcium level.15,16
Our previous studies found that both bone and dentin
exhibit a hypomineralisation phenotype in VDR�/� mice,
suggesting that dentin shares a similar mechanism to bone in
the mineralisation process.2,9 The present study has demon-
strated that dentin hypomineralisation phenotype in VDR�/�mice can be reversed by feeding with a high-calcium diet early
in life. This indicates that dentin mineralisation is indeed
regulated by serum calcium level and that VDR is not required
for normal dentin mineralisation.
In contrast to dentin, our study found hypermineralisation
patterns in VDR�/� enamel. More importantly, we found that
the enamel hypermineralisation phenotype could not be
reversed by a high-calcium diet. Enamel differs from bone
and dentin in its unique epithelial origin.17 In epithelium,
calcium ions are carried through the capillaries into cells. Such
is the case with enamel. During enamel mineralisation,
enamel organ is required to supply calcium in bulk and to
quickly move the calcium and phosphate ions from blood
stream into the enamel layer. This process takes place despite
the presence of many ionic barriers possibly created by
intervening capillary endothelial cells, papillary layer cells and
ameloblasts. Furthermore, ameloblasts maintain low free
cytosolic calcium concentrations to avoid toxicity and to
retain cell responsiveness to intracellular calcium signalling
cascades that use calcium transiently.17–19 Transcellular cal-
cium transfer across the ameloblast barrier consists of a passive
concentration gradient-dependent transportationandanactive
ATP-dependent transportation.20 Transcellular calcium is
responsible for transferring calcium ions through the tightly
sealed ruffle-ended ameloblasts whereas paracellular calcium
transportation is believed to occur between leaky smooth-
ended ameloblasts.19 Disruption of calcium transportation
could contribute to a variety of developmental defects in the
enamel. Removal of the VDR may open the tight junction in the
ruffle-ended ameloblasts and enhance paracellular calcium
transport as observed in the intestines.15,16 Further studies need
to be pursued in future to determine the relative change
between the transcellular and the paracellular calcium trans-
portation in VDR-deficient mouse ameloblasts. Understanding
underlying cellular mechanisms in VDR-mediated disruption of
calcium transportation will provide potential targets for drug
development to improve enamel quality.
The surface enamel serves as a barrier to block bacterial
penetration and resist abrasion and attrition. In our study, we
found that the surface enamel layer is much thinner in VDR�/
� mice and is more susceptible to acid erosion. This reduced
surface enamel layer is not reversible by a high-calcium diet.
After enamel maturation, the thickness of surface enamel is
maintained by the sum of two distinct and opposing
processes, mineralisation and demineralisation.21 The pH,
titratable acidity, phosphate and calcium concentrations as
well as fluoride content of enamel determine the degree of
tooth mineral saturation, and this degree of saturation is the
driving force of its demineralisation and remineralisation.22
Continuously erupting mouse incisors share some char-
acteristic features with human primary teeth, and acidic
erosion on the mouse incisors could be viewed as a good
simulator of the development of early childhood caries (ECC)
in American children. Despite efforts to intervene in the
transmission of the microbes and practices to correct
unhealthy eating habits, the average number of decayed
and filled teeth among 2- to 4-year-olds has remained
unchanged over the past 25 years.23 Some individuals appear
to be more susceptible to caries, regardless of environmental
risk factors to which they are exposed, indicating that genetic
factor may play an essential role in the development of ECC.24
The VDR gene polymorphisms were first reported to be
associated with variation in BMD in 1994.25 Since then, much
attention has been focussed on this relationship to resolve
controversies regarding the association of such polymorph-
isms and BMD.26–28 The conflicting results may result from
some compensation in calcium homeostasis since the bone
mineralisation is mainly regulated by systemic serum calcium
level. However, enamel mineralisation is associated with VDR
local expression and function. VDR could be a good candidate
gene to be investigated in the relationship between genetic
factors and caries susceptibility.
4. Conclusion
In this study, we used a high-calcium diet administration to
normalise serum calcium level and found that high calcium
supplementation can reverse dentin hypomineralisation, but
not enamel hypermineralisation. The result demonstrated
that dentin mineralisation is regulated systemically by serum
calcium level; however, VDR affects enamel mineralisation
locally.
Acknowledgements
We thank Dr. Marie Demay of Massachusetts General Hospital,
Harvard Medical School, for providing us with breeding pairs
of VDR heterozygous mice.
Funding: UABSOD Dean’s Research Fund.
Competing interests: None declared.
Ethical approval: Not required.
r e f e r e n c e s
1. Bouillon R, Carmeliet G, Verlinden L, van Etten E, Verstuyf A,Luderer HF, et al. Vitamin D and human health: lessonsfrom vitamin D receptor null mice. Endocr Rev2008;29(6):726–76.
2. Zhang X, Rahemtulla FG, MacDougall MJ, Thomas HF.Vitamin D receptor deficiency affects dentin maturation inmice. Arch Oral Biol 2007;52(12):1172–9.
3. Zhang X, Rahemtulla F, Zhang P, Beck P, Thomas HF.Different enamel and dentin mineralization observed in

a r c h i v e s o f o r a l b i o l o g y 5 4 ( 2 0 0 9 ) 1 1 0 5 – 1 1 1 01110
VDR deficient mouse model. Arch Oral Biol 2009;54(4):299–305.
4. Berdal A, Hotton D, Pike JW, Mathieu H, Dupret JM. Cell- andstage-specific expression of vitamin D receptor andcalbindin genes in rat incisor: regulation by 1,25-dihydroxyvitamin D3. Dev Biol 1993;155(1):172–9.
5. Demay M, Sabbagh Y, Carpenter T. Calcium and vitamin D:what is known about the effects on growing bone. Pediatrics2007;119(Suppl. 2):S141–4.
6. Whitfield GK, Dang HT, Schluter SF, Bernstein RM, Bunag T,Manzon LA, et al. Cloning of a functional vitamin D receptorfrom the lamprey (Petromyzon marinus), an ancientvertebrate lacking a calcified skeleton and teeth.Endocrinology 2003;144(6):2704–16.
7. Li Y, Pirro A, Amling M, Delling G, Baron R, Bronson R, et al.Targeted ablation of the vitamin D receptor: an animalmodel of vitamin D-dependent rickets type II with alopecia.Proc Natl Acad Sci USA 1997;94(18):9831–5.
8. Yoshizawa T, Handa Y, Uematsu Y, Takeda S, Sekine K,Yoshihara Y, et al. Mice lacking the vitamin D receptorexhibit impaired bone formation, uterine hypoplasia andgrowth retardation after weaning. Nat Genet 1997;16(4):391–6.
9. Lezot F, Descroix V, Mesbah M, Hotton D, Blin C, PapagerakisP, et al. Cross-talk between Msx/Dlx homeobox genes andvitamin D during tooth mineralization. Connect Tissue Res2002;43(2–3):509–14.
10. Li Y, Amling M, Pirro A, Priemel M, Meuse J, Baron R, et al.Normalization of mineral ion homeostasis by dietary meansprevents hyperparathyroidism, rickets, and osteomalacia,but not alopecia in vitamin D receptor-ablated mice.Endocrinology 1998;139(10):4391–6.
11. Amling M, Priemel M, Holzmann T, Chapin K, Rueger J,Baron R, et al. Rescue of the skeletal phenotype of vitamin Dreceptor-ablated mice in the setting of normal mineral ionhomeostasis: formal histomorphometric and biomechanicalanalyses. Endocrinology 1999;140(11):4982–7.
12. Agadir A, Lazzaro G, Zheng Y, Zhang XK, Mehta R.Resistance of HBL100 human breast epithelial cellsto vitamin D action. Carcinogenesis 1999;20(4):577–82.
13. Norman AW, Bishop JE, Bula CM, Olivera CJ, Mizwicki MT,Zanello LP, et al. Molecular tools for study of genomicand rapid signal transduction responses initiated by 1alpha,25(OH)(2)-vitamin D(3). Steroids 2002;67(6):457–66.
14. Anderson P, May B, Morris H. Vitamin d metabolism: newconcepts and clinical implications. Clin Biochem Rev2003;24(1):13–26.
15. Kutuzova GD, Sundersingh F, Vaughan J, Tadi BP, Ansay SE,Christakos S, et al. TRPV6 is not required for 1alpha,25-dihydroxyvitamin D3-induced intestinal calcium absorptionin vivo. Proc Natl Acad Sci USA 2008;105(50):19655–9.
16. Benn BS, Ajibade D, Porta A, Dhawan P, Hediger M, Peng JB,et al. Active intestinal calcium transport in the absence oftransient receptor potential vanilloid type 6 and calbindin-D9k. Endocrinology 2008;149(6):3196–205.
17. Hubbard MJ. Calcium transport across the dental enamelepithelium. Crit Rev Oral Biol Med 2000;11(4):437–66.
18. Simmer JP, Fincham AG. Molecular mechanisms of dentalenamel formation. Crit Rev Oral Biol Med 1995;6(2):84–108.
19. Smith CE. Cellular and chemical events during enamelmaturation. Crit Rev Oral Biol Med 1998;9(2):128–61.
20. Hoenderop JG, Bindels RJ. Is vitamin D indispensable forCa2+ homeostasis: lessons from knockout mouse models?Nephrol Dial Transplant 2005;20(5):864–7.
21. Briner WW, Francis MD, Widder JS. Factors affecting the rateof post-eruptive maturation of dental enamel. Calcif TissueRes 1971;7(3):249–56.
22. Fejerskov O, Larsen MJ, Richards A, Baelum V. Dental tissueeffects of fluoride. Adv Dent Res 1994;8(1):15–31.
23. American Academy of Pediatric Dentistry Council onClinical Affairs. Policy on early childhood caries (ECC):unique challenges and treatment options. Pediatr Dent2005;27(7 Suppl.):34–5.
24. Slayton RL, Cooper ME, Marazita ML. Tuftelin, mutansstreptococci, and dental caries susceptibility. J Dent Res2005;84(8):711–4.
25. Morrison NA, Qi JC, Tokita A, Kelly PJ, Crofts L, Nguyen TV,et al. Prediction of bone density from vitamin D receptoralleles. Nature 1994;367(6460):284–7.
26. Uitterlinden AG, Fang Y, van Meurs JB, van Leeuwen H, PolsHA. Vitamin D receptor gene polymorphisms in relation toVitamin D related disease states. J Steroid Biochem Mol Biol2004;89–90(1–5):187–93.
27. Liu YZ, Liu YJ, Recker RR, Deng HW. Molecular studies ofidentification of genes for osteoporosis: the 2002 update. JEndocrinol 2003;177(2):147–96.
28. Thakkinstian A, D’Este C, Eisman J, Nguyen T, Attia J. Meta-analysis of molecular association studies: vitamin Dreceptor gene polymorphisms and BMD as a case study. JBone Miner Res 2004;19(3):419–28.
Copyright © 2022 FDOKUMEN




















