
Functional segmentation of the brain cortex using high model order group PICA
Upload
kharkov-uaCategory
view
0download
0
Харьковский национальный университетимени В.Н. Каразина
НИИ биологии Харьковского национального университетаимени В.Н. Каразина
Мензбировское орнитологическое общество
Северо-восточное отделение Украинского орнитологического общества имени К.Ф. Кесслера
Рабочая группа по изучению и охране птицбассейна Северского Донца

Somov’s library
BIRD ECOLOGY: SPECIES, COMMUNITIES, INTERRELATIONS
Proceedings of the meeting commemorating the 150th anniversary of the birth of Nikolay N. Somov (1861-1923)
1-4 December 2011, Kharkov, Ukraine
Issue 1
Part 1
Kharkov, 2011

Харьковский национальный университет имени В.Н. Каразина,пл. Свободы, 4, г. Харьков, 61077, Украина
e-mail: [email protected]
О.А. Брезгунова
Ночёвки сорок (Pica pica): индивидуальные стратегии поведения, организация коллективных
ночёвок и биотопические предпочтения
O.A. Bresgunova. Magpie (Pica pica) roosts: individual behavioural strategies, communal roosts’ organization and habitat preference. – The organization of communal roost is poorly understood in most corvids. I investigated the distribution of main communal roosts and sub-roosts of Black-billed Magpie (Pica pica). 15 cluster Magpie roosts, which include main roost sites and several sub-roosts, were discovered in Kharkov city in 2002-2009 years. The general numbers of Magpies’ population within Kharkov city vary in different years from 6500 to 8000 ind. (mean 7360 ind.; n = 5). The Magpie is a sedentary species and maintains breeding territories year-round while travelling long distances (from 1,7 to 6,5 km, mean 3,95 km; n = 15) to roost communally at night in wintertime. The catchment area of communal roosts varied from 3,6 to 50 km² (n = 15). All Magpie communal roosts include main roost with 1-3 sites for sleeping (which Magpies use simultaneously or in different years) and some sub-roosts. The main roosts exist year-round on traditional territories. The Magpies form sub-roosts from April to November and after sub-roost departure birds congregate on main roost sites. The distance between main communal roost site and sub-roosts sites vary from 1,8 to 4,7 km (mean 3,6 km; n = 5). 80,55% of main communal roost sites are confined to floodplain terraces and birds prefer to sleep in willow (Salix cinerea) bushes and reeds (Phragmites australis) (61,11%) (n = 36). The more are the numbers of Magpies on communal roost site in winter the more is the catchment area of the roost (Spearman rank correlation R = 0,956; p < 0,01), the more is the maximum catchment radius of the roost (R = 0,716; p < 0,01), and the more individuals participate in roost formation in breeding period (R = 0,803; p < 0,01). The numbers of Magpies at main roost sites don’t change from early December to the end of February. The numbers of roosting Magpies decline from the end of March – early April. Minimum numbers of Magpies was observed at main roost sites in April and May. In breeding period main roosts were mainly joined by non-breeding Magpies, first-year birds and breeders from breeding territories, which are closer 500 m to main roost sites. From August the numbers of roosting Magpies start to
Серия: Сомовская библиотекаВып. 1. Экология птиц: Виды, сообщества, взаимосвязи

164
О.А. Брезгунова
Материалом для данной работы послужили исследования по распределению мест коллективных ночёвок сорок, проведенные в г. Харькове
Материал и методы
Введение
Коллективные ночёвки птиц – это часть общей социальной структуры, где реализуются социальные связи. Очевидно, что такая форма общения с другими особями популяции может способствовать дисперсии молодых птиц, расселению взрослых птиц, образованию пар и определению места индивида в структуре сообщества (Брезгунова, 2008б). Однако организация таких необхо-димых для птиц социальных образований, как коллективные ночёвки, изучена недостаточно, что обусловлено необходимостью индивидуального распознавания особей, проведением исследований в сумеречное и ночное время, а также не-обходимостью длительных, зачастую круглогодичных наблюдений за птицами в природе. Малоизученными остаются даже такие, на первый взгляд, простые вопросы как сезонная динамика численности птиц на местах ночёвок, характе-ристика ночёвок в репродуктивный период, биотопическое распределение мест коллективного ночлега, особенности ночёвок разных возрастных групп. В работе предпринята попытка обобщить данные по ночёвочному и предночёвочному поведению сороки (Pica pica) в гнездовой и поcлегнездовой периоды, а также дана характеристика мест, используемых сороками для ночлега.
increase. Main roosts include more communal roosting birds in breeding period than sub-roosts (Mann-Whitney U test; U = 17, p < 0,01). Consequently, non-breeding birds prefer to roost at main roost sites. The breeding Magpies have two types of roosting behaviour in breeding period. Females usually don’t join communal roost and often roost at the nest until fledglings leave it. If the breeding territory is situated near communal roost site (up to 500 m) and fledglings already left the nest females may fly to communal roost in the night. If the breeding territory is nearby communal roost site (up to 500 m) breeding males travel to roost communally at night when females incubate and brood and when fledglings already left the nest. Two adult Magpies roosted on breeding territory in incubation and brooding periods but the distance between their breeding sites and communal roost site exceeds 700 m. When the fledglings left the nests on these territories, only one parent bird remains on breeding territory, another adult bird participates in communal roost gathering. Distance between breeding territory and com-munal roost, stage of breeding cycle and roosting behaviour of adjacent breeding pairs play key roles in Magpie roosting strategies in breeding period. The Magpie’ broods roost within parent breeding territory for 20-40 days after leaving the nest. At age of 50-65 days young birds start to participate in communal roost gathering. The Black-billed Magpies form pre-roosting gatherings at breeding territories, near sites of traditional sub-roosts and along the way to roost sites on tree tops (conifers and deciduous trees), in bushes and on roofs of buildings.

165
Ночёвки сорок: их организация и индивидуальные стратегии поведения
в 2002-2008 гг. Всего обнаружены 28 мест ночлега, которые используются сороками из 15 центральных коллективных ночёвочных скоплений (птицы в 8 из 15 ночёвочных скоплений могут перемещаться между двумя-тремя участками для отдыха, в 7-ми – имеют только одно место для ночлега). Зимой 2009 г. сороки стали использовать еще одно место для ночлега, где также проводили наблюдения. При описании мест ночлега использованы данные наблюдений за 6 коллективными ночёвками сорок в Харьковской области (2003-2008 гг.), за одной в г. Киеве (ноябрь 2007 г.) и одной в г. Краснодаре (Российская Федерация; июль и ноябрь 2003 г.; январь 2006 г.).
При описании результатов исследования мы использовали следующую терминологию – предночёвочное скопление, центральная ночёвка, традиционная периферическая ночёвка, временная периферическая ночёвка. В связи с тем, что данные понятия по-разному трактуются в литературе, далее приводятся объяснения терминов, используемых в работе:
Предночёвочное скопление (или клуб) – скопление птиц, формируемое перед полетом на ночёвку, удаленное от места ночлега более чем на 1 км;
Центральная коллективная ночёвка – скопление птиц, поддерживаемое круглый год; птицы ночуют на одном или нескольких традиционных участках, используя их одновременно или в разные зимние сезоны;
Традиционная периферическая коллективная ночёвка – коллективная ночёвка, которую птицы формируют ежегодно в весенний, летний и осенний периоды на традиционных местах;
Временная периферическая коллективная ночёвка – коллективная ночёвка, которую птицы формируют на случайных участках в весенний, летний и осенний периоды.
По материалам наблюдений за центральной коллективной ночёвкой, расположенной в пойме р. Харьков, в 2002-2003 гг. была разработана методика поиска мест коллективных ночёвок и учета собирающихся на них птиц (Коноваленко и др., 2003).
Учет птиц на ночёвках подразумевает поиск всех мест скоплений данного вида на обследуемой территории и непосредственный подсчет всех присутствующих на месте ночлега особей (Бибби и др., 2000). При поиске мест ночлега сорок чаще отслеживают стаи во время перемещения к месту ночёвки (Protocol for censusing.., 2003-2011). В зимний период 2003-2004 гг. в г. Харькове с помощью бинокля определяли направление перелёта птиц в вечернее время в течение одного часа до и 30 минут после захода солнца. Направления перемещений стай и одиночных сорок, летящих на высоте не менее 30 м от земли, наносили на карту. Учетчик следовал за стаями до места ночлега. В пределах города поиск ночёвочных скоплений связан с рядом трудностей – сильное движение транспорта (Protocol for censusing.., 2003-2011), охраняемые и частные

166
О.А. Брезгунова
территории, постройки, заводы. Если наблюдатель не успевал за один вечер обнаружить место коллективной ночёвки, то на следующий день он продолжал отслеживать перемещения отдельных особей и стай, начиная с того участка, где он потерял их из виду в предыдущий день. Поиски велись до тех пор, пока не удавалось обнаружить место ночлега (Protocol for censusing.., 2003-2011). Кроме того, нами были проверены все подходящие для ночёвок биотопы – поймы рек, ручьи, пруды, овраги, пустыри. Следует отметить, что в сумерках сороки всегда находятся недалеко от места ночлега, перемещения в полной темноте нами отмечены не были1. В результате, была обследована вся территория г. Харькова и обнаружены все места коллективных ночёвок сорок.
Подсчет ночующих сорок возможен как после прилёта птиц на ночёвку, например, в свете уличных фонарей (Czechowski et al., 2005), так и во время формирования ночёвочного скопления. После обнаружения места ночёвки необходимо было выбрать наиболее удобную точку для подсчёта птиц (Proto-col for censusing.., 2003-2011). Наблюдатель должен был занять самое высокое место, при этом быть максимально незаметным для сорок (Møller, 1985), чтобы исключить свое влияние на поведение птиц.
Для сорок пути следования на место ночлега вечером и к гнездовым территориям утром полностью совпадают, что объясняется привязанностью птиц к своим гнездовым территориям в течение всего года. Следовательно, выбранное для учета место подходит и для утреннего, и для вечернего подсчёта птиц.
Численность сорок на коллективных ночёвках в зимнее время определяли, преимущественно, в течение полутора часов в вечерние сумерки во время прилёта птиц. Хотя известны случаи учёта сорок на ночёвках за 2-3 ч до захода солнца (Møller, 1985), подсчёт начинали за 1 ч (редко – 1,5 ч) до захода, когда сороки начинают активно перемещаться к месту ночлега. Подсчёт проводил один, реже – два2 наблюдателя, при этом расстояние от учётчика до места ночёвки варьировало от 50 до 150 м, в зависимости от условий конкретного места ночлега: чем крупнее ночёвка, тем на большем удалении от неё находился учетчик. Для тех мест ночёвок, где птицы использовали лишь один участок для ночлега, подсчет проводили по двум секторам – каждый наблюдатель покрывал сектор в 180º. Учёт численности на местах ночёвок, где птицы свободно перемещались между двумя или тремя участками, проводили одновременно или в течение нескольких дней. По возможности, учитывали все перемещения птиц. Учёт прекращали через 30 мин после захода солнца, когда сороки, как правило, уже
1 В темноте возможны перемещения сорок между соседними кустами, деревьями и т.п., то есть смена непосредственного места отдыха.
2 В исследованиях 2002-2004 гг. принимали участие также А.В. Брезгунов, С.В. Коно-валенко.

167
Ночёвки сорок: их организация и индивидуальные стратегии поведения
3 Современные исследования, основанные на изучении последовательности нуклеотидов мДНК, показали, что американские P. nuttalli и P. hudsonia являются подвидами поли-типического вида P. pica (Lee et al., 2003), с этим соглашаются и другие исследователи американских подвидов сорок (Trost, 1999; Koenig, Reynolds, 2009).
не перемещаются. Кроме того, численность птиц на коллективных ночёвках определяли дважды, а в отдельных случаях, трижды в течение зимнего сезона (Коноваленко, Коноваленко, 2004).
При подсчёте численности сорок на ночёвках в утренние часы наблюдатель прибывал на учётный участок за 40-45 мин до восхода солнца, так как известно, что эти птицы просыпаются до восхода (Сметана, 1978а; Møller, 1985). В Америке учёт коллективно ночующих сорок, в частности, Pica pica nuttalli 3, чаще проводят на разлёте с места ночёвки (Protocol for censusing..., 2003-2011). В настоящем исследовании численность сорок определялась, как правило, при сборе на ночёвку. Учёт на утреннем разлете, на наш взгляд, возможен только, если общее число коллективно ночующих птиц не превышает 250-300 особей. Численность птиц в более крупных ночёвочных скоплениях удобнее оценивать в вечернее время, поскольку период формирования ночёвки в два раза продолжительнее утреннего распада. В результате, вечерний учёт позволяет получить более точные данные. В вечерние часы учёт усложнен высокой социальной активностью прибывших на ночёвку сорок, которые часто вовлекаются в погони и преследования. Это особенно затрудняет задачу подсчёта птиц в конце зимы и в период гнездостроения – в марте. Необходимо также учитывать, что на месте ночлега птицы рассредоточены неравномерно – отдельные группы ночуют на расстоянии 50-150 м от основного скопления, что зачастую затрудняет подсчёт птиц.
Методика предполагает получение временной характеристики подлёта птиц к месту ночлега, а также формирования или распада ночёвочного скопления. Поэтому каждые 5-10 мин отмечали всех птиц, подлетающих к месту ночлега или покидающих его (Терновский, Терновская, 1969; Коноваленко, Коноваленко, 2004).
Исследования по поиску мест коллективных ночёвок сорок в 2003-2004 гг. заняли 75 дней. В последующие зимние сезоны 2004-2008 гг. места отдыха всех коллективных ночёвок сорок были проверены, при этом отмечались происходившие изменения.
За двумя центральными ночёвочными скоплениями, каждое из которых использует по два участка для отдыха, вели ежемесячные наблюдения с мая 2003 г. Уделяли внимание динамике прилёта птиц, соотношению особей, прилетающих одиночно, парами или в составе стай. Исследования были направлены на выяснение сезонных различий коллективного ночёвочного поведения сорок.
В апреле мае 2004 г. проводили учет численности сорок на всех местах

168
О.А. Брезгунова
центральных коллективных ночёвок. В июне 2004 г. (в период, когда птенцы начинают покидать гнездовые территории родителей) с целью выявления мест периферических коллективных ночёвок были обследованы поймы рек, ручьи, пруды, овраги, пустыри, районы одноэтажной и многоэтажной застройки в пределах района сбора самой крупной центральной ночёвки (в пойме р. Харьков зимой ночует от 1900 до 2500 сорок). Подсчёт проводил один наблюдатель во время прилета сорок на ночёвку. Расстояние от учётчика до места ночёвки не превышало 70 м. Все места ночёвок посещали по одному разу в течение лета. В мае-июне 2005 г. места периферических ночёвок сорок проверяли с целью выяснения традиционности их использования. В апреле 2006 г. места традиционных периферических ночёвок посещали для выяснения числа птиц, формирующих эти скопления в период гнездостроения и насиживания кладок. На двух местах традиционных периферических ночёвок вели ежемесячные наблюдения с апреля 2006 г.
Для всех мест ночёвочных скоплений сорок отмечали биотоп, который использовался птицами, и определяли особенности положения места ночлега в рельефе местности, например, пойменную зону (прирусловая, центральная или притеррасная). По возможности определяли виды кустарников и деревьев, используемые сороками для отдыха.
Кроме г. Харькова, наблюдения за коллективно ночующими сороками проводили в Харьковской области: в пойме р. Северский Донец от с. Черемушное до с. Мохнач, в пос. Научный, в пойме р. Муром у с. Русские Тишки, на оз. Лиман, в пойме р. Оскол у с. Калиново и др.
Изучение поведения размножающихся сорок в гнездовые периоды 2003-2007 гг. в сумеречное время проводили в пойме р. Харьков (Журавлёвский гидропарк) и в 2003-2005 гг. в Шишковской балке (пос. Жуковского). Анализировали особенности ночёвочного поведения сороки в гнездовой период в зависимости от пола, цикла и стадии размножения, и расстояния между гнездовой территорией и местом коллективного ночлега. Определяли возраст молодых птиц, начинающих посещать места коллективных ночёвок.
В связи с тем, что взрослые птицы реагировали на исследователя, наблюдения вели из укрытий, а гнездовые территории проверяли через 25-40 мин после захода солнца, когда вероятность перемещений сорок была минимальной.
Кроме кольцевания птенцов алюминиевыми кольцами Московского центра кольцевания, мы применяли цветные ножные кольца для индивидуального мечения птенцов. Использовали цветные пластмассовые спирали и пластмассовые кольца (Coulson, 1963). Птенцов метили в 10-20 дневном возрасте (Baeyens, 1981a,b).
В 2003-2007 гг. было окольцовано 74 птенца из 24 выводков первых кладок (2003 г. – 17 из 7 выводков, 2004 г. – 10 из 4, 2005 г. – 14 из 5, 2006 г. – 21 из 5, 2007 г. – 12 из 3) и 10 птенцов из 2 выводков повторных кладок (2004 г.). Удалось

169
Ночёвки сорок: их организация и индивидуальные стратегии поведения
Результаты и обсуждение
Организация коллективных ночёвок
Центральные ночёвкиИзвестно, что сорока образует скопления (Коцюруба, 1994), в том числе,
коллективные ночёвки (Linsdale, 1937; Goodwin, 1976; Møller, 1983). В пределах г. Харькова и Харьковской области сорока в зимнее время образует крупные коллективные ночёвки. Одиночные или парные ночёвки сорок в зимний период, описанные другими исследователями (Goodwin, 1976; Birkhead, 1989, 1991; Ка-лякин, Калякина, 1989; Кошелев и др., 2002; Pfister, 2004; Рогачева и др., 2008), в Харьковской области нам не встречались. То, что сороки зимой не ночуют одиночно и в парах, отметил также А. Мёллер (Møller, 1985).
Всего в пределах г. Харькова в зимние периоды 2002-2008 гг. было от-мечено 15 центральных коллективных ночёвочных скоплений сорок (Рис. 1), границы районов сбора которых за указанный период почти не менялись (Брезгу-нова, 2007а,б). Только зимой 2009 г. на границе районов сбора двух коллективных ночёвок образовалось новое ночёвочное скопление (подробнее см. ниже). Птицы семи ночевочных скоплений использовали по одному месту для ночлега (Табл.1), лишь в трёх случаях сороки могли смещаться в пределах 150 м (№№ 3, 6, 7 в Табл. 1). Птицы в остальных ночёвочных скоплениях использовали 2-3 места для ночлега одновременно или в разные зимние сезоны (Табл. 2).
Вероятно, число мест, используемых сороками для коллективного ночлега,
4 Сорока – моноцикличный вид. Под первым циклом размножения подразумевается нормальная первая кладка, под повторным – ещё одна попытка размножения после гибели первой кладки.
выяснить ночёвочное поведение 6 пар сорок с неполными кладками, 22 пар на стадии плотного насиживания и 20 пар на стадии выкармливания птенцов. В период повторного гнездования проводили наблюдения за 3 парами на стадии плотного насиживания и одной парой с птенцами в гнезде. Индивидуальное цветное мечение позволило выяснить особенности поведения в вечернее и ночное время 47 птенцов из 15 выводков первых4 и 10 птенцов из 2 повторных кладок.
Для оценки связи между различными показателями были использованы методы непараметрической статистики: коэффициент ранговой корреляции Спирмена (R), тест Манна-Уитни (U-тест).
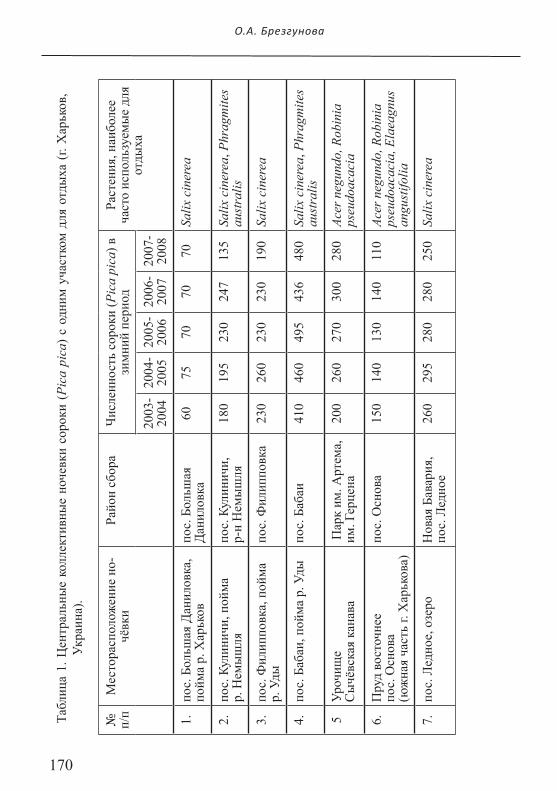
170
О.А. Брезгунова
№
п/п
Мес
тора
спол
ожен
ие н
о-чё
вки
Райо
н сб
ора
Числ
енно
сть
соро
ки (P
ica
pica
) в
зимн
ий п
ерио
дРа
стен
ия, н
аибо
лее
част
о ис
поль
зуем
ые
для
отды
ха20
03-
2004
2004
- 20
0520
05-
2006
2006
-20
0720
07-
2008
1.по
с. Б
ольш
ая Д
анил
овка
, по
йма
р. Х
арьк
овпо
с. Б
ольш
ая
Дан
илов
ка60
7570
7070
Salix
cin
erea
2.по
с. К
улин
ичи,
пой
ма
р. Н
емы
шля
пос.
Кул
инич
и,
р-н
Нем
ыш
ля18
019
523
024
713
5Sa
lix c
iner
ea, P
hrag
mite
s au
stra
lis
3.по
с. Ф
илип
повк
а, п
ойма
р.
Уды
пос.
Фил
иппо
вка
230
260
230
230
190
Salix
cin
erea
4.по
с. Б
абаи
, пой
ма р
. Уды
пос.
Баб
аи41
046
049
543
648
0Sa
lix c
iner
ea, P
hrag
mite
s au
stra
lis
5Ур
очищ
е С
ычё
вска
я ка
нава
П
арк
им. А
ртем
а,
им. Г
ерце
на20
026
027
030
028
0Ac
er n
egun
do, R
obin
ia
pseu
doac
acia
6.П
руд
вост
очне
е по
с. О
снов
а(ю
жна
я ча
сть
г. Х
арьк
ова)
пос.
Осн
ова
150
140
130
140
110
Acer
neg
undo
, Rob
inia
ps
eudo
acac
ia, E
laea
gnus
an
gust
ifolia
7.по
с. Л
едно
е, о
зеро
Нов
ая Б
авар
ия,
пос.
Лед
ное
260
295
280
280
250
Salix
cin
erea
Табл
ица
1. Ц
ентр
альн
ые
колл
екти
вны
е но
чевк
и со
роки
(Pi
ca p
ica)
с о
дним
уча
стко
м дл
я от
дыха
(г.
Хар
ьков
, Ук
раин
а).

171
Ночёвки сорок: их организация и индивидуальные стратегии поведения
№
п/п
Мес
тора
спо-
лож
ение
но-
чёвк
и
Мес
та о
тды
ха
ночё
вочн
ого
скоп
лени
я
Райо
н сб
ора
Раст
ения
, на
ибол
ее ч
асто
ис
поль
зуем
ые
для
отды
ха
Числ
енно
сть
в зи
мний
пер
иод
2003
- 20
0420
04-
2005
2005
- 20
0620
06-
2007
2007
- 20
081.
Шиш
ковс
кая
балк
апо
с. Ж
уков
ског
опо
с. Ж
уков
ског
оШ
ишко
вка
Cra
taeg
us sp
.75
120
6530
30
Пой
ма
р. О
чере
тянк
аSa
lix c
iner
ea,
Phra
gmite
s au
stra
lis
6060
130
140
180
2.П
оймы
р.
Лоз
овен
ька
и р.
Лоп
ань
Овр
аг п
ойма
р.
Лоз
овен
ька
Нов
ая
Але
ксее
вка,
Ф
лори
нка,
Зе
лено
е, С
тара
я А
лекс
еевк
а,
Сор
тиро
вка,
П
авло
во П
оле,
С
арж
ин Я
р
Prun
us sp
inos
a,
Cra
taeg
us sp
.15
00
00
0
Пой
ма
р. Л
озов
еньк
аSa
lix c
iner
ea,
Phra
gmite
s au
stra
lis
095
090
090
078
0
Але
ксее
вски
й лу
гопа
рк
(Але
ксее
вско
е во
дохр
анил
ище)
Salix
cin
erea
550
00
00
Табл
ица 2
. Цен
трал
ьны
е кол
лект
ивны
е ноч
евки
соро
ки (P
ica
pica
) с н
еско
льки
ми у
част
ками
для
отд
ыха
(г. Х
арь-
ков,
Укр
аина
).

172
О.А. Брезгунова
№
п/п
Мес
тора
спо-
лож
ение
но-
чёвк
и
Мес
та о
тды
ха
ночё
вочн
ого
скоп
лени
я
Райо
н сб
ора
Раст
ения
, на
ибол
ее ч
асто
ис
поль
зуем
ые
для
отды
ха
Числ
енно
сть
в зи
мний
пер
иод
2003
- 20
0420
04-
2005
2005
- 20
0620
06-
2007
2007
- 20
083.
Пой
ма
р. Х
арьк
ов
(Жур
авле
в-ск
ий г
идро
-па
рк)
Пус
тырь
по
с. Д
альн
яя
Жур
авле
вка
Стар
ое
Пав
лово
пол
е,
Шат
илов
ка,
Сок
ольн
ики,
Госп
ром,
Пар
к Ш
евче
нко,
Бл
ижня
я Ж
урав
левк
а,
Дал
ьняя
Ж
урав
левк
а,
Шев
ченк
и,
Боль
шая
Д
анил
овка
, С
евер
ная
Сал
товк
а,
Сал
товк
а
Acer
neg
undo
, Ro
bini
a ps
eudo
acac
ia
830*
1600
*0*
2380
0
Пой
ма р
. Хар
ьков
Phra
gmite
s au
stra
lis57
0*30
0*23
00*
00
Ул. Ж
иляр
диAc
er n
egun
do,
Robi
nia
pseu
doac
acia
, Po
pulu
s sp.
00
00*
*22
00**
*
4.П
ойма
р. Н
е-мы
шля
Пой
ма р
. Нем
ыш
-ля
(уч-
к №
1)
Саб
уров
а Д
ача,
Ста
рая
Сал
товк
а,
Раш
кина
Дач
а,
Сел
екци
онна
я,
Нов
озап
адны
й (ч
асти
чно)
Salix
cin
erea
, Ph
ragm
ites
aust
ralis
860
300
600
0
Пой
ма р
. Нем
ыш
-ля
(уч-
к №
2)
Salix
cin
erea
, Ph
ragm
ites
aust
ralis
6020
020
030
010
00
Саб
уров
а Д
ача
(у
ч-к
№ 3
)Ac
er n
egun
do,
Robi
nia
pseu
doac
acia
025
035
00
65

173
Ночёвки сорок: их организация и индивидуальные стратегии поведения
№
п/п
Мес
тора
спо-
лож
ение
но-
чёвк
и
Мес
та о
тды
ха
ночё
вочн
ого
скоп
лени
я
Райо
н сб
ора
Раст
ения
, на
ибол
ее ч
асто
ис
поль
зуем
ые
для
отды
ха
Числ
енно
сть
в зи
мний
пер
иод
2003
- 20
0420
04-
2005
2005
- 20
0620
06-
2007
2007
- 20
085.
Пой
ма р
. Уды
(Г
риго
ровк
а)П
ойма
р. У
ды
(бли
же
к Н
овоб
авар
ском
у во
дохр
анил
ищу)
Лы
сая
гора
, Х
олод
ная
гора
, За
люти
но,
Григ
оров
ка,
Мин
утка
Phra
gmite
s au
stra
lis30
00
075
00
Пой
ма р
. Уды
(б
лиж
е к
окру
жно
й до
роге
)
Elae
agnu
s an
gust
ifolia
400
00
00
Пой
ма р
. Уды
(п
ос. П
есоч
ин)
Salix
cin
erea
3080
067
00
700
6.П
ойма
р.
Лоп
ань
(Мир
ская
ро
ща)
Пой
ма р
. Лоп
ань
(Нов
ожан
ово)
Нов
ожан
ово,
М
ирск
ая р
оща,
М
оска
левк
а,
Заик
овка
, В
ерещ
аков
ка,
Дик
анев
ка,
Дуд
ковк
а
Acer
neg
undo
, Ro
bini
a ps
eudo
acac
ia
625
017
00
0
Пой
ма р
. Лоп
ань
Salix
cin
erea
063
023
042
038
0
7.Ба
лка
Боль
шая
Д
олин
а
Руче
й (п
ос. З
атиш
ье)
Нов
озап
адны
й,
ХТЗ
, Пли
точ-
ный
заво
д
Elae
agnu
s an
gust
ifolia
, Ro
bini
a ps
eudo
acac
ia
150
230
200
300
0
Пру
д на
те
ррит
ории
са
дово
го
това
рищ
еств
а
Phra
gmite
s au
stra
lis50
850
032
0
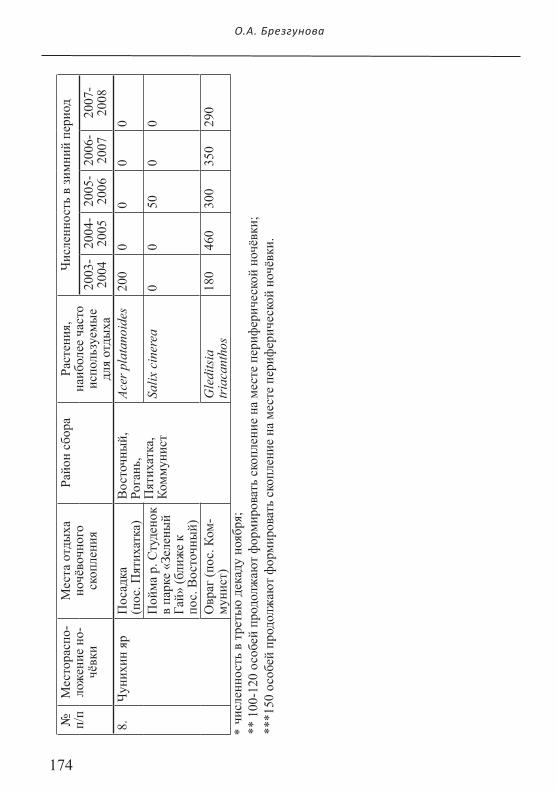
174
О.А. Брезгунова
№
п/п
Мес
тора
спо-
лож
ение
но-
чёвк
и
Мес
та о
тды
ха
ночё
вочн
ого
скоп
лени
я
Райо
н сб
ора
Раст
ения
, на
ибол
ее ч
асто
ис
поль
зуем
ые
для
отды
ха
Числ
енно
сть
в зи
мний
пер
иод
2003
- 20
0420
04-
2005
2005
- 20
0620
06-
2007
2007
- 20
088.
Чуни
хин
ярП
осад
ка
(пос
. Пят
ихат
ка)
Вос
точн
ый,
Ро
гань
, П
ятих
атка
, Ко
ммун
ист
Acer
pla
tano
ides
200
00
00
Пой
ма р
. Сту
дено
к в
парк
е «З
елен
ый
Гай»
(бли
же
к по
с. В
осто
чны
й)
Salix
cin
erea
00
500
0
Овр
аг (п
ос. К
ом-
муни
ст)
Gle
dits
ia
tria
cant
hos
180
460
300
350
290
∗ чи
слен
ност
ь в
трет
ью д
екад
у но
ября
;**
100
-120
осо
бей
прод
олж
ают
форм
иров
ать
скоп
лени
е на
мес
те п
ериф
ерич
еско
й но
чёвк
и;**
*150
осо
бей
прод
олж
ают
форм
иров
ать
скоп
лени
е на
мес
те п
ериф
ерич
еско
й но
чёвк
и.

175
Ночёвки сорок: их организация и индивидуальные стратегии поведения
зависит от численности ночующих птиц и от биотопических условий местности. В условиях спорадичного распределения благоприятных для ночёвок биотопов, коллективно ночующие сороки используют (или даже вынуждены использовать) одно место для ночлега (Коноваленко, Коноваленко, 2005). Крупные коллективные ночёвочные скопления чаще распадаются на более мелкие ночёвочные группы, в отдельные годы вновь объединяясь на одном из традиционных мест ночлега.
Так, в зимний период птицы в крупных ночёвочных скоплениях могут занимать два-три участка сразу. Сороки свободно перемещаются между ними в вечернее время в случае, если расстояние, которое им приходится преодолевать, не превышает 1 км. Такие места ночёвок часто приурочены к поймам рек (Уды, Харьков, Немышля), изобилующих подходящими местообитаниями, – обширными зарослями тростника обыкновенного (Phragmites australis) и ивы пепельной (Salix cinerea).
Известно, что площадь района сбора коллективных ночёвок может
Рис. 1. Распределение основных мест центральных коллективных ночёвок со-роки (Pica pica) в г. Харькове в 2002-2009 гг. (в основе – карта гидрологической сети г. Харькова по Стрижельчик и др., 2003; места отдыха одного ночёвочного скопления обозначены кружками с одинаковой штриховкой).

176
О.А. Брезгунова
составлять 50-75 км² (Brennecke, 1965, цит. по Stiefel, 1979). По нашим дан-ным, площадь района сбора центральных коллективных ночёвок варьирует от 3,6 до 50 км², составляя, в среднем, 17 км² (n = 15). Наблюдается следующая закономерность: чем большее число птиц участвует в образовании коллектив-ных ночёвок в зимний период, тем больше площадь района сбора (R = 0,956; p < 0,01) (Рис. 2).
Общеизвестно, что сороки обитают на своих гнездовых территориях круглогодично (Birkhead, 1991) и при этом привязаны к определённым местам ночлега (Куранов, 1979). Расстояние, с которого сороки собираются на кол-лективные ночёвки в зимний период, может варьировать от 1-1,5 (Ward, 1952; Birkhead et al., 1986; Buitron, 1988; Birkhead, 1991; Коноваленко, Коновален-ко, 2005) до 4-8 км (Utley, 1943; Gyllin, Källander, 1977; Stiefel, 1979; Buitron, 1988; Cramp, Perrins, 1994; Mäck, 1998; Кошелев и др., 2002; Коноваленко, Коноваленко, 2005), реже – до 10 км (Grinnell, 1923 цит. по Linsdale, 1937; Куранов, 1979). Иногда расстояние между местом кормёжки и ночёвки может достигать 40 км (Кричкевич, Смогоржевский, 1977).
По нашим данным, максимальный радиус района сбора, то есть мак-симальное расстояние, преодолеваемое птицами в зимний период при пере-лёте от гнездовых территорий до места коллективного ночлега, составляет от 1,7 до 6,5 км, в среднем, 3,95 км (n = 15)5. Прослеживается зависимость между максимальной численностью птиц на ночёвке и максимальным радиусом района сбора (R = 0,716; p < 0,01) (Рис. 3).
Рис. 2. Корреляция между максимальной (зимней) численностью птиц в ночё-вочном скоплении сорок (Pica pica) и площадью района сбора (S) (n = 15).
5 Данные без учета образовавшейся зимой 2008-2009 гг. коллективной ночёвки в урочище Саржин яр (2,25-5,25, в среднем, 3,3 км; n = 12)

177
Ночёвки сорок: их организация и индивидуальные стратегии поведения
По нашим данным, расстояние между ближай-шими местами отдыха разных центральных коллективных ночёвочных скоплений ва-рьирует от 2,25 до 7 км, в среднем, 3,6 км (n = 12) . По другим источникам, такое расстояние может состав-лять от 1,5-2,5 км (Møller, 1985) до 5 км (Czechowski et al., 2005), а среднее расстоя-
ние между ближайшими местами ночёвки – 1,9 км (Møller, 1985).Сороки формируют центральные коллективные ночёвки круглый год,
включая период размножения (Рис. 4) (Linsdale, 1937; Stiefel, 1979; Коновален-ко, 2006; Брезгунова, 2008а; Georgiev, Iliev, 2009).
Рис. 3. Корреляция между максимальной (зимней) численностью птиц в но-чёвочном скоплении сорок (Pica pica) и максималь-ным радиусом района сбора (n = 15).
Рис. 4. Численность сороки (Pica pica) на центральных ночёвках в зимний пери-од 2003-2004 гг. и гнездовой период (май 2004 г.) (n = 15).

178
О.А. Брезгунова
В гнездовой пери-од центральные ночёвки с одним местом для отдыха посещают от 1,8% до 28,7%, в среднем, 15,5% птиц, фор-мирующих эту ночёвку в зимнее время (подробнее см. ниже). По другим данным, в июле место формирования центральной ночёвки посе-щает до 32,3% птиц, ночую-щих здесь в зимнее время (Georgiev, Iliev, 2009). Для
коллективных ночёвок, где птицы использовали несколько участков для ночлега, численность в период размножения составляет 8,6-13,7%, в среднем, 11,8% от зимней численности. Наблюдается закономерность – чем выше численность птиц на ночёвке в зимнее время, тем большее число особей формируют коллективную ночёвку в гнездовой период (R = 0,803; p < 0,01) (Рис. 5).
Рис. 5. Корреляция между мак-симальной (зимней) численно-стью сорок (Pica pica) на но-чёвках и числом птиц, форми-рующих скопления в гнездовой период (n = 15).
Численность птиц на ночёвкеМногие исследователи отмечают ежедневные перемещения сорок к
местам ночлега (Горбань, Давидович, 1982 цит. по Пасічник, 1995). Заметными становятся даже некрупные ночёвочные скопления численностью до 20 осо-бей (Blake-Knox, 1868; Linsdale, 1937; Абдусалямов, 1973; 1977; Зонов, 1974; Coombs, 1978; Константинов и др., 1982; Хмельков, 1998; Urquhart, 1998), од-нако чаще в литературе описывают ночёвки от 50 до 300 птиц (Linsdale, 1937; Барабаш-Никифоров, Семаго, 1963; Gyllin, Källander, 1977; Кричкевич, Смогоржевский, 1977; Родимцев, Шкарин, 1984; Møller, 1985; Аюпов, 1989; Birkhead, 1989, 1991; Полозов, Бурнашев, 1989; Нечаев, 1990; Cramp, Perrins, 1994; Гержик, 1995; Mäck, 1998; Mäck, Jürgens, 1999; Гуль, Лемко, 2001; Горошко, Ку-сенков, 2003; Кривицкий, Реуцкий, 2003; Емцев и др., 2006; Сотников, 2006; Lee et al., 2008; Мороз, 2009; Сапетина, 2009; Georgiev, Iliev, 2009). Значительно реже встречается информация о ночёвках сорок от 350 до 1600 особей (Linsdale, 1937; Осмоловская, Формозов, 1950; Спангенберг, 1953; Терновский, Терновская, 1969; Stiefel, 1979; Хохлов, 1983; Бокотей, 1991, 1995а,б; Bokotey, 1997; Кошелев и др., 2002; Czechowski, Jerzak, 2003; Жуков, 2006; Georgiev, Iliev, 2009; Мало-

179
Ночёвки сорок: их организация и индивидуальные стратегии поведения
вичко и др., 2010) и даже 1600-2700 особей (Сметана, 1978а; Сеник, 2007; Georgiev, Iliev, 2009). Такие подвиды сорок, как Pica pica sericea, P. p. bottanensis (Linsdale, 1937), P. p. hudsonia (Dice, 1917; Linsdale, 1937; Hall, 1994; Trost, 1999), P. p. nuttalli (Linsdale, 1937; Birkhead, 1991; Koenig, Reynolds, 2009) также фор-мируют крупные коллективные ночёвки.
По нашим данным, максимальная численность птиц (зима), одновре-менно встречающихся на ночлеге, варьирует от 40 до 2500 особей, составляя, в среднем, 374 особи (n = 102)6. В целом, ночёвочные скопления численностью до 300 особей (50%) встречаются чаще, от 300 до 1500 птиц (45%) – несколько реже, а скопления, насчитывающие более 1500 особей (5%), представляют собой и вовсе редкое явление (n = 20)7.
Исследования показали, что общая численность сорок, обитающих в пределах г. Харькова, в разные зимние сезоны варьировала в пределах 6,5-8 тысяч особей и составляла, в среднем, 7360 птиц (n = 5) (Брезгунова, 2009). Для г. Харь-кова численность гнездящихся сорок в 2000 г. оценивалась путём экстраполяции в 2,5-2,8 тысяч пар (Надточий, Зиоменко, 2001). Использованный нами метод учёта позволил определить численность городской популяции сорок, включая неразмножающихся птиц, доля которых в разных частях ареала составляет от 18% до 60% (Baeyens, 1979, 1981a,b; Møller, 1985; Birkhead et al., 1986).
Периферические ночёвкиПри проведении исследований в пределах района сбора центральной
ночёвки, расположенной в пойме р. Харьков (Журавлевский гидропарк), обнару-жено 9 периферических ночёвочных скоплений на участках, где в зимний период птицы формируют предночёвочные скопления, но, как правило, не остаются ночевать (Рис. 6). Численность сорок на этих ночёвках составила от 3 до 32, в среднем, 11 особей8. В большинстве случаев птицы из одного скопления ис-пользовали для ночлега один участок, реже – ночевали на нескольких смежных участках. Удалось установить, что 5 периферических ночёвочных скоплений являются традиционными (55,6%), остальные – временными.
Традиционные периферические ночёвки находились в 1,8-4,7 км (в среднем, 3,6 км; n = 5) от мест отдыха птиц, составлявших центральную но-чёвку. Все традиционные периферические ночёвки сороки, как правило, начи-нают формировать с начала апреля. К ноябрю птицы перемещаются на места центральных ночёвок, при этом места периферических ночёвочных скоплений,
6 Число наблюдений в г. Харькове, Харьковской области, в г. Киеве, в г. Краснодаре (Рос-сийская Федерация) (для г. Киева численность дана для конца ноября).
7 Анализировали данные по коллективным ночёвкам сорок в г. Харькове, в Харьков-ской области, в г. Киеве, в г. Краснодаре (Российская Федерация).
8 На периферических ночёвках из 3-8 птиц учитывали возраст сорок: если среди них было более 2 взрослых птиц, то ночёвка считалась коллективной.

180
О.А. Брезгунова
удаленные на расстояние до 2 км от центральных, распадаются раньше, уже в первой декаде августа. Места временных коллективных ночёвок сорок рас-полагаются на расстоянии 1,7-3,6 км (в среднем, 3,05 км; n = 4) от мест отдыха центральных коллективных ночёвок.
Периферические коллективные ночёвки в ранний послегнездовой период могут представлять собой объединение семейных групп (выводки с родителями) за пределами гнездовых территорий. Скопления после объединения семейных групп описаны и другими исследователями (Сметана, 1978а; Buitron, 1988).
Возможно, часть участков, которые сороки используют на центральных коллективных ночёвках для сна в зимний период, могут быть местами перифе-рических коллективных ночёвок, где по каким-то причинам птицы оставались ночевать и зимой (Табл. 2). Однако постоянные наблюдения в районах сбора центральных колллективных ночёвок не проводили (за исключением места но-чёвки в пойме р. Харьков) и все участки, используемые сороками для ночлега, считали местами отдыха центральных коллективных ночёвок.
Рис. 6. Пространственная организация коллективной ночёвки сорок (Pica pica) (пойма р. Харьков, г. Харьков, Украина).

181
Ночёвки сорок: их организация и индивидуальные стратегии поведения
Динамика формирования и распада ночёвочных скопленийИзвестно, что основными синхронизаторами суточных биоритмов
являются интенсивность освещения, длительность дня и ночи, время захода и восхода солнца (Промптов, 1940; Кузнецова, Правдина, 1977). Время захо-да солнца и освещённость – главные ориентиры для следующих на ночёвку птиц разных видов, которые значительно коррелируют с такими параметрами, как время начала формирования ночёвочного скопления, пик прилёта птиц и время прилёта последних особей (Gill, Dow, 1985; Perlmutter, 1992; Russelli, Gauthreaux, 1999; Peh, 2002). В облачные дни, особенно при наличии дождей, сороки прилетают на место ночлега раньше обычного, а утром покидают его в более позднее время (Gorska, 1998).
Формирование коллективных скоплений сорок в зимнее время начи-нается за 1 час до захода солнца. Однако часть птиц может держаться на месте ночлега и в дневное время (Федосенко, Орлов, 1969; наши данные) или при-летать за 2-3 часа до наступления темноты (Møller, 1985; Birkhead, 1991). По другим данным, формирование ночёвок сорок возможно за более короткое вре-мя – 0,5-1 ч и заканчивается в вечерних сумерках (Терновский, Терновская, 1969). Вечерний прилёт на ночлег начинают одиночные птицы и пары, доля которых в первые 30 мин составляет от 27 до 51% от всех прилетевших на ночёвку со-рок (n = 15). Пик прилёта сорок приходится на сумеречное время, за 20-30 мин до захода солнца. Ближе к ночи возрастает число птиц, прилетевших в группах от 4 до 30-70 особей. После захода солнца птицы, прилетевшие в составе стай, составляют от 49 до 73% от общего числа особей, присоединившихся к скоплению в это время. Весь период формирования ночёвки, в среднем, состав ляет 1 ч 30 мин (n = 15). По другим данным, прилёт занимает 90 мин - 75 мин перед заходом и 15 мин после захода солнца, а пик прилёта приходится на 30-15 мин перед за-ходом солнца, хотя весной большая часть птиц прилетает за 15 мин до и 15 мин после захода солнца (Georgiev, Iliev, 2009). Очевидно, что время формирования ночёвки зависит от освещённости, наличия ветра, осадков и т. д. Так, в пасмур-ную погоду птицы прилетают на ночёвку раньше на 30-40 мин.
В летнее время вечерний прилёт птиц на место ночлега длится около 2 ч и заканчивается до захода солнца, в отличие от зимы, когда птицы вынуждены тратить больше времени на поиски пищи и, следовательно, позже прилетать на место ночлега, что также отметил А. Мёллер (Møller, 1985). Сокращение времени формирования ночёвочного скопления в зимний период характерно и для других видов птиц (Berndt, 1988).
Распад ночёвочного скопления в зимний период занимает 40-45 мин. Общее время, за которое птицы покидают место ночлега, значительно короче времени прилёта. Основная масса сорок покидает место ночёвки за 35-45 мин до восхода солнца. Утренний разлёт начинают группы порядка 10-100 особей, а завершают отлёт одиночки и пары. В летний период сороки начинают покидать

182
О.А. Брезгунова
место ночёвки за 15-25 мин до вос-хода солнца. При этом общее время распада ночёвки остается прежним, около 40-45 мин. Отдельные особи на-ходятся на месте ночлега около 40 мин после восхода.
Общая структура коллек-тивных ночёвок
Известно, что в разных ча-стях ареала в зимнее время сорока формирует коллективные ночёвки на традиционных участках (Родимцев, Шкарин, 1984; Møller, 1985; Хмель-ков, 1998; Czechowski, Jerzak, 2003; Брезгунова, 2005, 2008а, 2010а,в; Коноваленко, Коноваленко, 2005; Czechowski et al., 2005; для P. p. sericea Linsdale, 1937; для P. p. hudsonia Trost , 1999; для P. p. nut tal l i Crosbie et al., 2008 цит. по Koenig, Reynolds, 2009). Так, птицы, состав-ляющие самое крупное в г. Харькове ночёвочное скопление, используют одно и то же место (в пойме р. Харь-ков), по крайней мере, 17 лет (Коно-валенко и др., 2003; А. Реуцкий, личн. сообщ.), а сороки, собирающиеся на ночлег в Шишковской балке, – не менее 20 лет.
Всего на территории г. Харь-кова обнаружено 15 центральных но-чёвочных скоплений, объединяющих несколько участков для ночлега, между которыми, в зависимости от периода года, перемещаются птицы. В весенне-осенний период сороки продолжают посещать места центральных ночёвок, а также формируют небольшие (до 35 особей) периферические ночёвочные скопления на других участках в преде-лах района сбора. Птицы участвуют в формировании периферических кол-
Рис. 7. Переход периферической коллек-тивной ночёвки сороки (Pica pica) в цен-тральную.А – пространственная организация коллек-тивных ночёвок;Б – рост численности птиц, составляющих периферическое ночёвочное скопление;В – место отдыха периферического ночё-вочного скопления сороки начинают ис-пользовать в зимний период.

183
Ночёвки сорок: их организация и индивидуальные стратегии поведения
лективных ночёвок в случае, если их гнездовые территории удалены от места центральной ночёвки более, чем на 1,5 км. Часть таких скоплений существует в гнездовой и ранний послегнездовой периоды на традиционных местах, осталь-ные – не привязаны к определенному участку. К ноябрю большинство сорок присоединяется к традиционным местам отдыха на центральных коллективных ночёвках, покидая периферические ночёвочные скопления (Рис. 7А).
В гнездовой период численность коллективно ночующих сорок значитель-но выше на центральных ночёвках, чем на периферических (тест Манна-Уитни; U = 17, p < 0,01). Следовательно, в гнездовой период неразмножающиеся сороки предпочитают ночевать на местах центральных коллективных ночёвок, что ха-рактерно и для других врановых, например, клушицы Pyrrhocorax pyrrhocorax (Blanco, Tella, 1999).
Численность сорок на местах центральных ночёвок в г. Харькове, как правило, не изменяется с начала декабря до конца февраля (Рис. 8). В других регионах отмечается пик численности в декабре-январе (Gyllin, Källander, 1977; Czechowski, Jerzak, 2003; Georgiev, Iliev, 2009). Наиболее заметными ночёвочные скопления становятся после вылета молодых птиц (Stiefel, 1979), иногда с пер-вой декады августа (Жуков, 2006), но чаще с октября-ноября до начала апреля (Linsdale, 1937; Gyllin, Källander, 1977; Møller, 1985; Mäck, 1998; Сапетина, 2009; Czechowski, Jerzak, 2003; Czechowski et al., 2005). В конце марта – начале апреля
Рис. 8. Динамика формирования коллективной ночёвки сороки (Pica pica) с дву-мя участками для отдыха (Журавлевский гидропарк, пойма р. Харьков, г. Харь-ков, Украина).

184
О.А. Брезгунова
численность сорок в местах центральных ночёвочных скоплений падает в связи с началом периода гнездостроения. Одни птицы в это время начинают формировать периферические коллективные ночёвки, сокращая тем самым расстояние между гнездовой территорией и местом ночлега (Брезгунова, 2006а,б, 2008а), а другие особи могут ночевать на гнездовых территориях с момента начала гнездостроения (Brown, 1924) или сразу после завершения строительства гнезд (Stiefel, 1979). Отдельные пары сорок могут также оставаться на ночлег на путях перелёта с гнездовых территорий к местам центральных коллективных ночёвок, отделяясь от следующих на ночёвку стай при наличии благоприятных для ночлега мест (например, заломов тростника). В апреле, когда самки приступают к откладке яиц, и в мае, в период выкармливания птенцов, численность птиц на местах центральных ночёвок сокращается до минимума (Goodwin, 1976; Coombs, 1978; Брезгунова, 2008а): они поддерживаются неразмножающимися особями (Брез-гунова, 2008а; Georgiev, Iliev, 2009), парами, чьи гнездовые территории располо-жены в пределах места ночлега (Georgiev, Iliev, 2009) и самцами, чьи гнездовые территории удалены не более, чем на 500 м от места ночлега (подробнее см. ниже). Возможно, более опытные размножающиеся пары гнездятся возле мест центральных коллективных ночёвок (Брезгунова, 2008в).
Вероятно, периферические ночёвки поддерживают сороки тех же категорий, что и центральные скопления, но в их составе значительно меньше неразмножающихся птиц.
В июне-августе на местах центральных и периферических ночёвок на-блюдается небольшой рост численности за счет присоединения к скоплениям выводков и самок, участвующих в размножении. Начиная с сентября, численность сорок на местах центральных ночёвок увеличивается за счет присоединения к скоплению особей, формирующих до этого периферические ночёвки. В течение сентября-ноября сороки объединяются на местах центральных ночёвок, покидая участки периферических коллективных ночёвочных скоплений.
Таким образом, размножающиеся пары весь послегнездовой период участвуют в формировании коллективных ночёвок вместе с неразмножающимися птицами, но утром обязательно разлетаются к своим гнездовым территориям (Stiefel, 1979; Deckert, 1980; наши данные; для P. p. hudsonia Buitron, 1988; для P. p. nuttalli Koenig, Reynolds, 2009). Так, например, в сентябре P. p. nuttalli ночуют коллективно, а утром территориальные пары покидают места совмест-ной ночёвки и следуют к гнездовым территориям, где сидят на самых высоких деревьях в течение 30 мин перед перемещением к местам кормёжки (Koenig, Reynolds, 2009). Очевидно, что возможны и другие стратегии ночёвочного поведе-ния (Cramp, Perrins, 1994). Так, многолетние исследования Т. Беркхерда показали, что коллективные ночёвки формируют, в основном, неразмножающиеся особи, а территориальные сороки ночуют одиночно или по две в пределах гнездовой территории в течение всего года (Birkhead, 1989, 1991).
Остается открытым вопрос: чем обусловлено перемещение сорок с

185
Ночёвки сорок: их организация и индивидуальные стратегии поведения
9 р. Карасун – правый приток р. Кубань, ныне – сеть прудов.
мест небольших периферических коллективных ночёвок и объединение их на местах центральных ночёвочных скоплений. На первый взгляд кажется, что формирование центральных коллективных ночёвок не связано с похолоданием. Подтверждением чему является тот факт, что в пойме р. Карасун (г. Краснодар, Российская Федерация)9, где мы проводили наблюдения, в ноябре температура воздуха доходила до +12˚С, а численность птиц на коллективных ночёвках рос-ла, как и в Харькове, где в это время температура опускалась до -2˚С. На наш взгляд, сороки в разных частях ареала имеют различный порог чувствительности к изменениям температуры.
Не менее важен вопрос о том, насколько сороки привязаны к определен-ному месту центральной ночёвки. Данные цветного мечения сорок в г. Томске показали, что птицы не обязательно выбирают место ночёвки, расположенное на минимальном удалении от места гнездования, а зачастую преодолевают значи-тельные расстояния (Куранов, 1979). По нашим данным, за период исследования границы районов сбора центральных коллективных ночёвок практически не изменились. Другими словами, сороки из определённых районов города сле-дуют на одни и те же места ночёвок, что отмечено и другими исследователями (Czechowski et al., 2005).
Пути и причины формирования коллективных ночёвок на новых местах
Распад центрального скопления на две‑три группы с использованием нескольких мест ночлега
Зачастую перемещение птиц одного ночёвочного скопления можно при-нять за формирование новых коллективных ночёвок. Необходимо картировать места ночлега сорок, контролировать перемещения птиц и проводить учёты численности на местах отдыха в течение каждого зимнего сезона. Только в этом случае представится возможность проследить образование новой коллективной ночёвки или установить факт частичного перемещения птиц одного ночёвочного скопления. Вероятно, некоторые данные наблюдений на местах ночёвочных скоплений сорок, показывающие существенное падение численности ночующих птиц в середине зимы (Czechowski, Jerzak, 2003), связаны со сменой традици-онных мест отдыха птицами из одного ночевочного скопления.
Переход периферической коллективной ночёвки в центральнуюНами отмечен случай, когда на месте традиционного периферического
ночёвочного скопления сороки начали оставаться на ночёвку в зимний период (Рис. 7). Этому предшествовало увеличение численности птиц, составлявших периферическое ночёвочное скопление в осеннее время. Зимой 2006-2007 гг. от 100 до 120 сорок не присоединились к центральному ночёвочному скопле-

186
О.А. Брезгунова
нию в пойме р. Харьков (Журавлевский гидропарк) и продолжали участвовать в формировании традиционной периферической ночёвки в пойме р. Очеретянка (до этого птицы зимой здесь не ночевали). В зимний период 2007-2008 гг. число птиц на месте периферической ночёвки увеличилось до 150 особей (Рис. 9). На наш взгляд, причина того, что сороки перестали участвовать в формировании цен-тральной ночёвки и весь зимний период посещали место периферической ночёвки, заключается в том, что на традиционном месте отдыха центрального ночёвочного скопления велись интенсивные строительные работы и, как следствие, площадь доступных для ночлега местообитаний сократилась более чем в три раза.
Формирование ночёвок на местах традиционных предночёвочных скоплений
Есть основания полагать, что в двух случаях сороки ночевали на местах, где они часто формируют предночёвочные скопления в зимний период. К со-жалению, осталось неизвестно, ночевали ли сороки на этих участках в летнее время. Так, после окончания продолжительных морозов зимой 2005-2006 гг. часть птиц, принимавших участие в формировании ночёвки в Чунихином яру, осталась ночевать на месте традиционного предночёвочного скопления в пойме р. Студенок (парк «Зеленый Гай» в пос. Восточный) (Табл. 2). Зимой 2003-2004 гг. 30 сорок, обычно ночевавших в пойме р. Немышля у пос. Кулиничи (Табл. 1) на одну ночь остались на 4 км ниже по течению, приблизившись к соседнему ночёвочному скоплению всего на 1 км.
Рис. 9. Численность сороки (Pica pica) на периферической ночёвке в летне-осенние периоды 2004 г. и 2005 г., а также в зимние периоды 2006 -2007 и 2007-2008 гг. (Шишковская балка, пойма р. Очеретянка, г. Харьков, Украина).

187
Ночёвки сорок: их организация и индивидуальные стратегии поведения
Образование коллективной ночёвки на границе районов сбора централь-ных коллективных ночёвок
Очевидно, что значительная удалённость участков, на которых рас-положены центральные коллективные ночёвки, от гнездовых территорий может способствовать образованию новых ночёвок при наличии подходящих для этого условий (Рис. 10).
Так, зимой 2008-2009 гг. на границе районов сбора двух крупных коллек-
Рис. 10. Образование новой коллективной ночёвки сорок (Pica pica) на границе районов сбора центральных коллективных ночёвок.А – пространственная организация коллективных ночёвок;Б – формирование новой ночёвки на границе районов сбора крупных централь-ных коллективных ночёвок.

188
О.А. Брезгунова
Индивидуальные стратегии ночёвочного поведения в период раз-множения
Ночёвки в гнездахИзвестно, что сороки могут ночевать у своих гнёзд (Северцов, 1950;
Хохлова, Кривцов, 1984) или использовать для ночлега гнездовые постройки, часто старые (Huber, 1944 цит. по Stiefel, 1979; Erpino, 1968 цит. по Birkhead, 1991; Birkhead, 1991). Они могут также ночевать в гнёздах других видов птиц, напри-
тивных ночёвок (№№ 2 и 3 в Табл. 2) образовалось новое ночёвочное скопление, включающее около 100 сорок. Птицы ночевали в пойме р. Саржинки в урочище Саржин яр. Новый участок для ночлега располагался в 3,5 и 4 км от ближайших мест ночёвок. На наш взгляд, причин, по которым птицы остались ночевать на новом месте, несколько. Во-первых, сороки гнездились на периферии районов сбора коллективных ночёвок, а использование нового места ночлега существенно сократило расстояние между гнездовыми территориями и местом коллективного ночлега. Во-вторых, площадь пригодных местообитаний на одном из традици-онных мест отдыха центрального ночёвочного скопления (пойма р. Харьков, Журавлевский гидропарк) уменьшилась в связи со строительными работами.
Возможно, то, что птицы из многих ночёвочных скоплений в зимний период используют для ночлега несколько мест (Табл. 2), также объясняется необходимостью сокращения расстояния перелёта между местом коллективной ночёвки и гнездовой территорией.
Кроме того, возможными причинами, по которым сороки перемещаются между разными местами ночлега, могут служить пожары. Так, скопление сорок в пойме р. Уды (Григоровка) после пожара в пойме использовало для ночлега только одно из традиционных мест отдыха.
Наиболее интересная, на наш взгляд, причина смены мест ночлега связана с понижением температуры воздуха в зимнее время. Находившееся под постоянным наблюдением коллективное ночёвочное скопление сорок в пойме р. Харьков (Журавлевский гидропарк), традиционно использовало для ночлега два участка (Табл. 2). Однако, зимой 2005-2006 гг., после сильных продолжительных морозов (-30°С и ниже), 700 особей (30,5% от общей численности) несколько ночей провели на расстоянии около 1 км от обоих участков. После потепления птицы переместились на традиционное место ночлега.
Описаны также случаи изменения места ночлега под влиянием хищни-ка. Так, сороки P. p. hudsonia могут покидать место отдыха в случае появления виргинского филина Bubo virginianus (Trost, 1999). Принимая во внимание высокую социальность сороки, на наш взгляд, нельзя исключать возможности смены центральных ночёвок отдельными особями в течение жизни без видимых причин.

189
Ночёвки сорок: их организация и индивидуальные стратегии поведения
мер, белого аиста Ciconia ciconia (Linsdale, 1937). Гнёзда часто используют для ночлега зимой (Goodwin, 1976; Cramp, Perrins, 1994; Pfister, 2004). Так, на косе Обиточной (Азовское море) в зимний период до 30% сорок ночуют поодиночке в своих гнёздах (Кошелев и др., 2002). В зимнее время некоторые размножающиеся пары могут использовать старые гнёзда для ночлега, даже находясь недалеко от мест коллективных ночёвок; при этом одна птица ночует в гнезде, а вторая – на ближней к нему ветке (Goodwin, 1976). Интересно, что некоторые пары могут присоединяться к коллективным скоплениям на местах ночлега, но поздним вечером улетать на ночёвку к своим гнёздам (Goodwin, 1976). В Харьковской области нам не удалось отметить аналогичного ночёвочного поведения сорок.
Для североамериканской сороки P. p. hudsonia вопрос об использовании старых гнёзд для ночлега остается открытым (Trost, 1999). Так, одни исследова-тели наблюдали, как сороки с рассветом покидали старые гнёзда (Linsdale, 1946 цит. по Trost, 1999; Erpino, 1968 цит. по Birkhead, 1991), другие такого поведения не отмечали (Verbeek, 1973; Trost, 1999).
Тибетские сороки P. p. bottanensis строят гигантские ночёвочные гнезда, достигающие 2 метров в высоту и примерно столько же – в ширину (Schäfer, 1938). Так, например, одно гнездо располагалось на усохшей пихте (Abies sp.) в 4 м от земли. Это большое гнездо походило на пчелиные соты, так как состояло из нескольких соединенных вместе, небольших по размеру отдельных гнезд и было устроено из неочищенных веток (Schäfer, 1938). В таких гнездах каждый вечер собиралось по 8-10 сорок, хотя днем стаи тибетских сорок могут объеди-нять 15-30 особей. Для размножения (апрель-июль) используется только самое верхнее гнездо (Schäfer, 1938).
Ночёвки в период размножения Известно, что в период гнездостроения некоторые пары ночуют на
деревьях, где расположены гнезда, или в пределах гнездовой территории (Linsdale, 1937; Goodwin, 1976; Møller, 1985; для P. p. nuttalli Verbeek, 197310). В период гнездостроения мы отмечали ночёвки пар сорок в тростнике, на удалении от гнездовых территорий и отдельно от остальных птиц. Интересно, что сороки, приступившие к строительству гнёзд раньше, значительно дольше участвуют в формировании коллективной ночёвки, чем пары, которые позднее начали строить гнёзда (Møller, 1985). Следовательно, более опытные птицы дольше участвуют в формировании коллективных ночёвок.
Нам удалось выяснить, что расстояние между гнездовой территорией и местом коллективной ночёвки влияет на поведение размножающихся сорок в гнездовой период.
10 Самцы P. p. nuttalli ночуют 2-3 дня возле достроенного гнезда, а затем выбирают другие деревья для ночлега в пределах гнездовой территории (Verbeek, 1973).

190
О.А. Брезгунова
Так, в период размножения на стадии откладки яиц (неполная кладка в 1-3 яйца) у сорок наблюдались различные варианты ночёвочного поведения. В одном случае в гнезде ночевала самка11, а самец покинул гнездовую территорию на ночь и присоединился к коллективной ночёвке. Расстояние от гнездового участка до места ночлега не превышало 300 м. В другом гнезде с кладкой из двух яиц сороки не остались ночевать (гнездовая территория прилегала к месту коллективного ночлега), однако, через два дня гнездо было разорено12.
Следующие наблюдения касаются поведения птиц на гнездовых терри-ториях, удаленных более чем на 700 м от места коллективного ночлега. В двух случаях самцы перемещались на соседнюю гнездовую территорию, где ночевали, соответственно, с двумя и пятью взрослыми сороками, предположительно, с самцами со смежных территорий. При этом самки ночевали в гнёздах. В одном случае самец ночевал на границе своей гнездовой территории совместно с че-тырьмя взрослыми птицами. Еще один случай – на гнездовой территории ночью отмечены обе птицы, самец ночевал в 25 м от гнезда. Очевидно, что границы
Рис. 11. Ночёвки взрослых сорок (Pica pica) на гнездовой территории в период размножения.
11 Все взрослые птицы, которые проводили ночь в гнезде, принимались за самок, так как случаи насиживания кладки или обогрева птенцов самцами для сорок неизвестны (Birkhead, 1991; Cramp, Perrins, 1994).
12 Вероятно, редкое явление, так как в остальных случаях самки сороки начинали ночевать в гнезде уже с момента откладки первых яиц (см. также Stiefel, 1979), хотя плотное дневное насиживание чаще начинается перед откладкой предпоследнего яйца.

191
Ночёвки сорок: их организация и индивидуальные стратегии поведения
гнездовых территорий в ночное время не соблюдаются.Из 22 находившихся под наблюдением гнездовых территорий с полны-
ми первыми кладками на 12 самцы ночевали на своей или соседней гнездовой территории. Все без исключения случаи относились к гнездовым территориям, удаленным на расстояние 700-1200 м от места коллективного ночлега. Когда если это расстояние не превышало 500 м, самцы покидали гнездовые территории и присоединялись к коллективным ночёвочным скоплениям (Рис. 11).
В период, когда птенцы уже вылупились, а самка продолжает ночевать в гнезде, самцы в 10 случаях из 20 ночевали на своей или соседней гнездовой территории. Гнездовые территории, в пределах которых (или на незначительном расстоянии от них) в этот период ночевали обе взрослые птицы, были удалены от места коллективной ночёвки на 700-1200 м (Табл. 3).
В 8 из 20 случаев самцы покидали свои гнездовые территории вечером (расстояние между гнездовыми территориями и местом коллективного ночле-га – до 500 м) и присоединялись к коллективным ночёвочным скоплениям, воз-вращаясь к гнездам лишь за 15 минут до восхода солнца. Более того, находясь всего в 150 м от гнезда, самцы не реагировали на сигналы тревоги самки, хотя в дневное время они прилетают на её крик с бóльшего расстояния.
В 2 из 20 случаев обе взрослые птицы стали оставлять на ночь гнез-довые территории и присоединяться к коллективным ночёвкам, когда птенцы достигли 10-14 дневного возраста (Рис. 11). На наш взгляд, такое поведение обусловлено тем, что размножающиеся сороки с соседних гнездовых территорий,
Стадия размножения
Цикл размножения, расстояние до места коллективной ночёвки
Плотное насижива-
ние
Гнездовые птенцы
Слётки
Первый цикл, расстояние до места коллективной ночёвки до 500 м
♀ ♀ или никого
никого
Первый цикл, расстояние до места коллективной ночёвки 700-1200 м
♀♂ ♀♂ 1 взрослая птица (♀ ?)
Повторный цикл, расстояние до места коллективной ночёвки более 700 м
♀ ♀ никого
Таблица 3. Ночёвки сорок (Pica pica) в период размножения при разной удалённости их гнездовых территорий от места коллективно-го ночлега.

192
О.А. Брезгунова
приступившие к гнездованию раньше и, следовательно, раньше покидающие свои территории, могут оказывать влияние на соседние пары, способствуя умень-шению сроков их пребывания на собственной территории. Другими словами, сороки могут значительно раньше вовлекаться в формирование коллективных ночёвок, при этом оставлять птенцов на территории одних, если птицы-соседи уже начинают покидать свои гнездовые территории. В описываемых случаях гнездовые территории находились на расстоянии до 300 м от места ночлега.
В период, когда птенцы достигают возраста 23 суток, в случае, если расстояние между гнездовой территорией и местом коллективного ночлега не превышает 500 м, самки часто ночуют в пределах гнездовой территории (не в гнезде) или покидают её и присоединяются к ночёвочным скоплениям. Возможно, что взрослые птицы ночуют при этом на тех же деревьях, что и ночевали перед этим, что известно для P. p. nuttalli (Verbeek, 1973).
Таким образом, данные наших наблюдений за поведением сорок в ночное время показали, что у размножающихся пар существуют разные стратегии по-ведения (Рис. 11; Табл. 3), и выбор места ночлега между гнездовой территорией и коллективной ночёвкой определяется удаленностью места коллективного ночлега от гнездовой территории и поведением птиц на смежных гнездовых участках (Брезгунова, 2006а,б).
В литературе описаны обе стратегии ночёвочного поведения самцов сороки. Так, по одним данным, в период, когда самки насиживают кладки и обогревают птенцов, самцы держатся на гнездовой территории и ночью (Deckert, 1980; Mäck, 1998; Czechowski et al., 2005; Georgiev, Iliev, 2009; для P. p. hudsonia Buitron, 1988), часто близко к гнезду (Stiefel, 1979; Mäck, 1998; Georgiev, Iliev, 2009). По другим – после откладки первых яиц самец ночует на удалении до 100 м от гнездовой территории или присоединяется к неразмно-жающимся особям, направляющимся на коллективную ночёвку (Brown, 1924; Stiefel, 1979).
В период окончательного вылета птенцов из гнезда, в возрасте 24-30 (в сред-нем, 27) дней, обе птицы покидают гнездовую территорию, оставляя птенцов на ночь, но только при условии, если их гнездовые территории находятся на расстоянии до 500 м от места коллективной ночёвки, что прослежено для 7 пар (Рис. 11). В случае бóльшей удаленности гнездовой территории от места коллективной ночёвки, одна взрослая птица ночует на гнездовом участке вместе с выводком. Такое поведение мы наблюдали у 8 пар. Птенцы P. p. hudsonia, после того как покинут гнёзда, ночуют на гнездовых территориях, а оба родителя возобновляют посещение коллективных ночёвок (Buitron, 1988). Выводки P. p. nuttalli сразу после вылета из гнезда ночуют в пределах гнездовой территории вместе с ро-дителями (Verbeek, 1973). При этом слётки первоначально ночуют отдельно от взрослых птиц; объединение семейной группы на ночёвке зависит от расстояния между деревом, на котором располагалось гнездо, и деревом, выбранным для

193
Ночёвки сорок: их организация и индивидуальные стратегии поведения
ночлега взрослыми птицами (Verbeek, 1973).
Ночёвки во время повторного цикла размноженияВ случае разорения гнезда на стадии кладки, в 52,6% случаев (n = 19) сороки
приступают к строительству гнезда для повторной кладки (Брезгунова, 2008а,б).При повторном размножении, в период насиживания, в трёх случаях
из трёх на территории ночевала только одна птица, хотя расстояние до места ночлега составляло 700 м (Рис. 12).
При повторном размножении, в период, когда птенцы уже покинули гнёзда, но еще ночуют в пределах гнездовой территории, обе размножающиеся пары, находившиеся под наблюдением, покинули гнездовые участки на ночь, хотя до места коллективного ночлега было 700 м (Брезгунова, 2006а,б). По нашим наблюдениям, взрослые птицы вечером покидают свои гнездовые территории со слетками неодновременно.
При повторном размножении наблюдается лишь одна стратегия пове-дения – на стадии насиживания кладок и выкармливания птенцов на гнездовых территориях остаются на ночь только самки, а после вылета птенцов из гнезда и они следуют на коллективные ночёвки (Рис. 12). Объяснением такого поведения может служить то, что в это время птицы на соседних гнездовых территориях начинают покидать свои участки на ночь и присоединяться к коллективным но-чёвкам. Поведение птиц-соседей способствует привлечению размножающихся сорок на коллективные ночёвки.
Рис. 12. Ночёвки взрослых сорок (Pica pica) на гнездовой территории в случае повторного размножения.

194
О.А. Брезгунова
Ночёвки выводковПтенцы, достигшие возраста 20-23 суток, в течение 4-6 дней на не-
сколько часов могут покидать гнездо, но на ночь возвращаются в него. В возрасте 24-30 дней выводок покидает гнездо, но может возвращаться в него на ночь или ночевать на кустарниках и деревьях (Bährmann, 1968 цит. по Goodwin, 1976; Stiefel, 1979). Обычно, птенцы из гнезд, находившихся на вы-соких деревьях, на ночёвку в гнездо не возвращаются (Сметана, 1978б). Зави-симость ночёвочного поведения выводков от вида гнездового дерева отмечена также для P. p. nuttalli: птенцы не возвращаются в гнёзда, если они устроены на деревьях одних видов и посещают их для ночлега, если они построены на деревьях других видов (Verbeek, 1973). В любом случае, выводок после вылета из гнезда возвращается в него на ночлег не более 5 дней (Сметана, 1978б; для P. p. nuttalli Verbeek, 1973).
Молодые птицы из первых и возобновлённых кладок ночуют на гнез-довой территории после вылета из гнезда на протяжении 20-40 дней. Птенцы спят в укромных местах – в тростнике или на нижних ветвях кустарников, деревьев13. Перемещение птенцов в эти укрытия контролируют взрослые пти-цы. Известно, что сороки предпочитают гнездиться на территориях с густой растительностью. Это, возможно, связано и с тем, что в первое время после вылета из гнезда птенцы на таких гнездовых территориях находятся в большей безопасности от хищников.
Интересно, что после вылета из гнезда птенцы P. p. nuttalli ночуют от-дельно от родителей даже в пределах гнездовой территории, что продолжается от 3 до 10 дней (в среднем, 6,4 дня). Затем слётки присоединяются к взрослым птицам и ночуют на облюбованном последними дереве (Verbeek, 1973).
По нашим наблюдениям, в 50-65 дневном возрасте птенцы покидают гнездовые территории на ночь, присоединяясь к коллективным ночёвкам. В дневное время в этот период выводки соседних территорий держатся вместе на одном из гнездовых участков или на их границах. Объединенные выводки могут перемещаться в пределах двух-четырех гнездовых территорий. В формировании этих скоплений участвуют также птицы-родители. Кроме того, к этим группи-ровкам могут присоединяться выводки прошлого года (Baeyens, 1981a,b). Такие объединенные выводки с родителями могут держаться вместе в течение всей зимы (Крушинский, 1986; наши данные). По другим данным, на 2-14 сутки после вылета из гнезда выводок покидает гнездовую территорию, после чего за её пределами происходит объединение семейных групп и формирование стаи (Сметана, 1978а). У P. p. hudsonia также через 9-29 дней после вылета птенцов из гнезда происходит объединение семейных групп (от 2 до 8 выводков с родителями; Buitron, 1988).
13 Очевидно, что выводки сорок, гнездящихся в центральных районах города, сталкива-ются с трудностями при выборе безопасного места ночлега.

195
Ночёвки сорок: их организация и индивидуальные стратегии поведения
Поскольку независимость молодые сороки приобретают на 50-80 день (Birkhead, 1991; Buttermer, 1994), то на места коллективных ночёвок птенцы следуют, как правило, за птицами-родителями. Так, в возрасте около двух ме-сяцев молодые сороки уже участвуют в формировании коллективных ночёвок (Брезгунова, 2007а).
Ночёвки выводков прошлого годаВ ночное время выводки прошлого года держатся группами до 20 особей
(Deckert, 1980) и ночуют коллективно (Birkhead, 1991). По нашим наблюдениям, сороки в возрасте одного года участвуют в формировании коллективных ночёвок на традиционных местах. Эти птицы образуют «ядро» ночёвочного скопления, по крайней мере, на местах центральных коллективных ночёвок. Выводки прошлого года могут держаться на гнездовой территории родителей до 11 месяцев (два наблюдения) (Баник, Брезгунова, 2007). В период откладки яиц они участвуют в активной охране гнездовой территории родителей и прекращают посещать ее после появления в гнезде последнего яйца. Прошлогодние выводки P. p. nuttalli могут даже ночевать на гнездовых территориях родителей (Verbeek, 1973).
Биотопическая характеристика мест коллективных ночёвок Общеизвестно, что места коллективных ночёвок сорок располагаются
в поймах рек, в оврагах, балках, на окраинах лесов, в лесополосах (Формо-зов, 1947; Ward, 1952; Абдусалямов, 1973; Пантелеев, 1974; Кричкевич, Смо-горжевский, 1977; Stiefel, 1979; Бутьев и др., 1983; Ильичев и др., 1987; Cramp, Perrins, 1994; Mäck, 1998; Лысенков и др., 2004; Сапетина, 2009). При этом большинство мест коллективных ночёвок сорок приурочены к берегам рек и озер (Møller, 1985; Birkhead, 1991; Брезгунова, 2010а).
Сороки ночуют в густых кустах и на высоких деревьях, т.е. в наи-более безопасных для них местах (Зарудный, 1888; Goodwin, 1976; Gyllin, Källander, 1977; Reebs, 1986; Нечаев, 1990; Birkhead, 1991; Горошко, Кусен-ков, 2003; для P. p. hudsonia Hall, 1994; для P. p. nuttalli Crosbie et al., 2006; для P. p. sericea Linsdale, 1937). Описаны ночёвки на ветвях ивы Salix sp. (Linsdale, 937; Зонов, 1974; Кричкевич, Смогоржевский, 1977; Мальчевский, Пукинский, 1983; Родимцев, Шкарин, 1984; Møller, 1985; Cramp, Perrins, 1994; Mäck, 1998; для P. p. nuttalli Grinnell, 1923 цит. по Linsdale, 1937), боярышника Crataegus sp. (Brown, 1924; Utley, 1943; Birkhead, 1989; Cramp, Perrins, 1994; Хмельков, 1998; Ponz, Monrós, 2000), лоха Elaeagnus sp. (Кошелев и др., 2002), тёрна Prunus spinosa, шиповника Rosa sp., бузины Sambucus sp., клёна Acer sp. (Mäck, 1998), ежевики Rubus ulmifolius (Ponz, Monrós, 2000), тополя Populus sp. (McGregor, 1906; Крич-кевич, Смогоржевский, 1977; Mäck, 1998; Ponz, Monrós, 2000; для P. p. bactriana Pfister, 2004), ольхи Alnus sp. (Кричкевич, Смогоржевский, 1977), дуба Quercus sp. (Linsdale, 1937; Гержик, 1995; Mäck, 1998; Ponz, Monrós, 2000; для P. p. nuttalli Linsdale, 1937; Verbeek, 1973; Crosbie et al., 2006), берёзы Betula sp. (Møller, 1985;

196
О.А. Брезгунова
Mäck, 1998), яблони Malus sp. (Ponz, Monrós, 2000), ирги овальной Amelanchier ovalis (Ponz, Monrós, 2000), сосны Pinus sp., ели Picea sp., лиственницы Larix sp. и других хвойных (Blake-Knox, 1868; Brown, 1924; Linsdale, 1937; Тернов-ский, Терновская, 1969; Сметана, 1978а; Møller, 1985; Mäck, 1998; Петрович, Редінов, 2007). Обычным ночёвочным биотопом сорок палеарктических подвидов являются также тростниковые заросли (Спангенберг, 1953; Gyllin, Källander, 1977; Stiefel, 1979; Кошелев и др., 1989; Birkhead, 1991; Cramp, Perrins, 1994; Гуль, Лемко, 2001; Кошелев и др., 2002). Очевидно, сорока, для которой свойственно формирование коллективных ночёвок в течение всего года (Брезгунова, 2005, 2008а), использует для ночного отдыха разные типы местности и разные виды растений.
По нашим данным, места центральных коллективных ночёвок сорок находятся, преимущественно, в пределах пойменной террасы (80,55%): в при-русловой пойме (44,44%), в центральной части пойменной террасы в зонах её понижения (8,33%), в приматериковой части пойменной террасы, в области выклинивания грунтовых вод (27,78%) (Рис. 13). Остальные традиционные места ночлега расположены в пределах овражно-балочных систем (16,67%) и у берегов внепойменных озер (2,78%) (n = 36).
Очевидно, что сороки отдают предпочтение ночёвкам в зарослях ивы пепельной и тростника, где расположено большинство мест центральных но-чёвочных скоплений (61,11%; Брезгунова, 2010а,б). Важную роль для ночлега сорок в зимний период играют заросли интродуцентов, таких как клен ясенелист-ный Acer negundo, робиния лжеакация Robinia pseudoacacia, лох узколистный
Рис. 13. Распределение мест центральных коллективных ночёвок сорок (Pica pica) (n = 36).

197
Ночёвки сорок: их организация и индивидуальные стратегии поведения
(Elaeagnus angustifolia, гледичия Gleditsia triacanthos (27,78%; Рис. 14). Реже сороки в зимний период ночуют в кустах боярышника и тёрна (8,33%), ино-гда остаются на ночь на ветвях клена остролистного Acer platanoides (2,78%) (n = 36).
Места периферических коллективных ночёвок располагались в поймах рек и по берегам озер (69,23%), часто находились у дорог, в парковой зоне, во дворах многоэтажной и частной застроек (30,77%) (n = 13). Птицы, прилетаю-щие на места периферических коллективных ночёвок, устраиваются на ночлег в зарослях ивы пепельной и тростника (53,85%), на ветвях клена ясенелистного, робинии лжеакации, лоха узколистного (38,46%), реже – на ветвях плодовых деревьев (7,69%) (n = 13) (Брезгунова, 2010а,б).
Очевидно, что для сорок наиболее характерно устраиваться на ночлег в пределах пойменной террасы, особенно в прирусловой пойме и притеррасье, где в понижениях произрастают тростник и ива пепельная – наиболее типичные виды растений для мест коллективных ночёвок сорок. Немаловажное значение играют интродуценты из Азии и Северной Америки, заросли которых создают благоприятные места для ночлега сорок как в поймах рек, так и в пределах овражно-балочной системы. В Северной Америке места зимних ночёвок P. p. nuttalli также приурочены к прибрежным коридорам и удалены от гнездовых местообитаний (Crosbie et al., 2006; Koenig, Reynolds, 2009).
Нехарактерные типы ночлегаВ Северной Америке описаны регулярные ночёвки P. p. hudsonia груп-
пами по 6-7 особей на спинах крупного рогатого скота (Criddle, 1923).
Рис. 14. Растения, используемые сороками (Pica pica) для отдыха на местах цен-тральных коллективных ночёвок (n = 36).

198
О.А. Брезгунова
Предночёвочное поведение
Предночёвочные скопления и перемещение к месту ночлегаОбразование предночёвочных скоплений – характерная черта поведения
европейских (Goodwin, 1976; Birkhead, 1991) и американских (P.p.hudsonia; Trost, 1999) сорок. Скопления птицы формируют на удалении от мест ночёвок, часто возле мест кормёжки (Goodwin, 1976). Особи, участвующие в образовании предночёвочных скоплений, следуют на одну и ту же коллективную ночёвку.
В зимний период большинство традиционных предночёвочных скоплений сорока формирует вблизи мест периферических ночёвок14. Однако, для этого социального вида характерно образование массовых предночёвочных скоплений и на других участках. По нашим наблюдениям, сорока также образует клубы на гнездовых территориях и по пути следования к местам ночлега на верхушках деревьев (включая хвойные), в гуще кустов и на крышах зданий. Число птиц, формирующих предночёвочные скопления, обычно составляет от 15 до 70, в среднем, 35 особей (n = 11).
Образование предночёвочных скоплений на гнездовых территориях связано с объединением выводков с соседних участков. Так, в течение зимы 2004-2005 гг. на одной из находившихся под наблюдением гнездовых территорий сорок держался выводок текущего года. К этому скоплению присоединились птицы с соседних гнездовых территорий, и общая численность предночёвочного скопления достигла 60 особей.
Мы наблюдали формирование крупного скопления в условиях обильного снегопада 7 января 2004 г. в Залютинском яру (южная часть г. Харькова). Птицы за 1 ч 45 мин до захода солнца прятались в кустах ивы пепельной, залетая туда группами по 2-3 особи. За 15 мин до захода солнца собралось около 200 сорок. Затем птицы по одной или в составе стай покидали место скопления и следовали на ночёвку в пойму р. Уды (Григоровка). Последние сороки покинули место предночёвочного скопления через 10 мин поле захода солнца.
В случае, если температура воздуха опускается ни же -100С, можно наблюдать формирование крупных предночёвочных скоплений по пути следо-вания птиц на ночёвку. Сороки периодически останавливаются и отдыхают на верхушках деревьев, к ним присоединяются единичные особи и целые стаи. В такие скопления может вовлекаться до 50% следующих на ночёвку птиц.
Численность птиц в отдельных традиционных предночёвочных ско-плениях постоянна. Сороки следуют на ночлег традиционными маршрутами, что также отметили З.А. Горошко и А.Н. Кусенков (2003). Так, предночёвочное
14 Сороки формируют предночёвочные скопления возле мест периферических коллек-тивных ночёвок в зимний период, когда эти участки уже не используются для ночного отдыха.

199
Ночёвки сорок: их организация и индивидуальные стратегии поведения
скопление из 15 сорок в пойме р. Жихорец регулярно перемещалось на место коллективной ночёвки в пойму р. Уды (пос. Бабаи), причем птицы следовали по одной и той же трассе, над сосновым бором (Рис. 15).
Вероятно, наблюдения Н.Н. Сомова (1897) за перемещением сорок в зимний период в поймах рек Харьковской области относятся к птицам, следовавшим на коллективную ночёвку. Так, на место ночлега сороки летят разреженными (Bree, 1862; Hadfield, 1874; Georgiev, Iliev, 2009) или скучен-
Рис.15. Пути перемещения сорок (Pica pica) из поймы р. Жихорец на место цен-тральной коллективной ночёвки в пойму р. Уды (пос. Бабаи).

200
О.А. Брезгунова
ными (Спангенберг, 1953; Гержик, 1995) стаями по 20-25 особей (Utley, 1943; Georgiev, Iliev, 2009), группами по 3-7 особей и отдельными парами (Ward, 1952; Goodwin, 1976; Birkhead, 1991). По нашим данным, число птиц в одной стае может достигать 70 особей. Размер стай возрастает при приближении к месту ночлега, что связано с объединением следующих на ночёвку птиц.
Следуя на место ночлега, «сороки летят высоко, не привлекая лишнего внимания, и, долетев до места, отвесно пикируют» (Семаго, 1975, Стр. 248). Поведение птиц, занимающих места для отдыха, описано Е.П. Спангенбергом: «каждая сорока, то складывая, то распуская свои короткие крылья, ныряла в воздухе и в несколько приемов пикировала почти до самой воды…» (Спан-генберг, 1953, Стр. 165). Такое поведение становится особенно заметно уже в сентябре (Utley, 1943).
Поведение на месте ночлегаНа месте коллективного ночлега сороки могут чистить оперение, кор-
миться, формировать скопления. Одни птицы прилетают на место коллективной ночёвки и сразу устраиваются на отдых, другие – кормятся вплоть до насту-пления сумерек (могут прилетать на место ночлега с кормом или разыскивать его рядом с местом ночлега). Летом стаи сорок кормятся совместно вплоть до ухода на общую ночёвку. Интересно, что к скоплениям сорок на кормёжке в вечернее время могут присоединяться насиживающие самки, покидая гнезда на 15-20 мин.
Известно, что взрослые сороки и птицы в возрасте одного года группами от 2 до 6 особей участвуют на местах коллективных ночёвок в погонях и пре-следованиях (Ward, 1952; наши данные). По мнению многих исследователей, «погони» – это проявление демонстративного поведения, играющего роль в об-разовании пар (Linsdale, 1937; Erpino, 1969). Нами такая активность на местах коллективных ночёвок отмечалась в течение всего года. С середины ноября до середины февраля вовлечение птиц в погони наблюдалось значительно реже, а с конца февраля – начала марта в «преследования» включаются десятки птиц. Сороки принимают участие в погонях даже после захода солнца в полной тем-ноте. Кроме того, вблизи мест коллективных ночёвок сороки часто образуют церемониальные скопления15, в которые вовлекаются от 8 до 35 особей (ред-ко – до 60-70 птиц).
Птицы, прилетевшие на ночлег до захода солнца, могут собираться вокруг места коллективной ночёвки. Такие скопления формируются на расстоя-
15 Церемониальные или игровые скопления – скопления, формируемые размножающимися парами и холостыми сороками с февраля до апреля; птицы в скоплениях используют разные формы поведения – погони, «пересаживания» с ветки на ветку и т.п. Собираются птицы чаще на деревьях, лишённых листьев, на земле.

201
Ночёвки сорок: их организация и индивидуальные стратегии поведения
нии от 10 (Møller, 1985) до 200 м от места ночлега (Birkhead, 1989, 1991). Для P. p. nuttalli отмечены подобные скопления на деревьях и на земле (Grinnell, 1923 цит. по Linsdale, 1937). Прилетевшие на место ночлега раньше других часто за-нимают самые высокие деревья (Georgiev, Iliev, 2009), в основном, лишённые листьев, реже – ветви кустарников (Utley, 1943; наши данные). На некоторых местах ночёвок сороки могут ежедневно соблюдать определенный «ритуал» перемещения на места ночлега с высоких «присад» (Горошко, Кусенков, 2003). Одни птицы могут занимать такие «посадочные места» («alighting spot») перед тем, как переместиться на место ночного отдыха (Ward, 1952), другие – сразу «пикируют» к нему. В случае если большинство птиц после прилёта заняли верхушки деревьев или крыши зданий, они могут синхронно перемещаться на места отдыха за 20 мин до или через 15-20 мин после захода солнца. Иногда со-роки собираются на одних кустах, а затем, после захода солнца, перемещаются на ночлег на ветви соседних. Если места коллективных ночёвок расположены возле линий электропередачи, то сороки рассаживаются на проводах. Числен-ность таких скоплений достигает 50-200 особей, расстояние между сидящими рядом птицами колеблется от 10 см до 1 м. Некоторые особи, занявшие высокие присады, могут перекрикиваться с вновь прилетевшими птицами, не покидая свои «посты» вплоть до завершения формирования ночёвки.
Поведение сорок после прилёта на место коллективного ночлега в течение года меняется несущественно. Однако весной птицы ведут себя более оживленно, могут целыми стаями включаться в преследования, или даже по-кидать место ночлега (вероятно, затем возвращаются обратно).
Если, на месте ночлега находится клуб серых ворон (Corvus cornix) или грачей (C. frugilegus), то после их отлёта, сороки сначала поднимаются вверх (на высоту более 100 м), а затем перемещаются над местом ночёвки сотенными стаями. Интересно, что такой массовый «подъем» в воздух, может быть вызван также пожаром возле места ночлега или появлением тетеревятника (Accipiter gentilis). Сороки могут «отводить» тетеревятника (или человека) с места ночёвки, «ныряя» перед ним в тростник на участке, где обычно не ночуют, но чаще появ-ляющихся на местах коллективных ночёвок ястребов они просто преследуют.
Сороки выбирают для ночлега центральную часть зарослей, площадь которых может достигать 800-1000 м² (Utley, 1943; Brennecke, 1965 цит. по Stiefel, 1979; Mäck, 1998). В разные годы птицы могут ночевать на разных участ-ках места ночлега (Терновский, Терновская, 1969; для P. p. nuttalli Protocol for censusing.., 2003-2011), что может зависеть от погодных условий (Møller, 1985). Часто выбирают место для сна соответственно условиям погоды: в ветреную погоду – подветренную сторону, а в морозы – нижние ветви (Møller, 1985). Ве-роятно, ветер не только влияет на выбор места ночлега, но и может определять численность птиц на коллективной ночёвке (Czechowski et al., 2005). Степень закрытия клюва и плюсен также зависит от погоды: в самые холодные ночи птицы

202
О.А. Брезгунова
прячут клюв под крыло, а перьями живота закрывают плюсны (Reebs, 1986).Непосредственно тростник сороки используют для ночлега в зимний
период крайне редко, чаще спят на ветвях ивы пепельной. Ночуя в зарослях боя-рышника, сороки предпочитают устраиваться на ветвях в средней части кроны (Birkhead, 1991), хотя небольшая часть птиц спит на верхних и нижних ветвях. В хвойных деревьях располагаются близко к стволу на нижних ветвях (Reebs, 1987 цит. по Birkhead, 1991). Спят на высоте 0,5-6 м (наши данные), реже – до 12 м; чаще ночуют в 2-5 м от земли (Møller, 1985).
В кустарнике ночуют группами от 5-8 (Зонов, 1974) до 40-50 особей. Расстояние между птицами составляет от 20 до 70 см, в среднем, 50 см (n = 20) (0,3-1 м для P. p. hudsonia Splendoria, 1971 цит. по Trost, 1999). По одним данным могут спать, плотно прижавшись друг к другу (Кривицкий, 1999; Lee et al., 2008), по другим – не скучиваются даже в холодные ночи (для P. р. hudsonia Trost, 1999).
На ночёвках сороки пугливы (Терновский, Терновская, 1969), предва-рительно осматривают место отдыха (Холодковский, Силантьев, 1901), на месте ночлега замолкают (Utley, 1943; Спангенберг, 1953). Могут менять непосредствен-ное место отдыха ночью – пересаживаться (Utley, 1943; наши наблюдения).
Наблюдателя подпускают на расстояние до 10 м, а затем очень тихо слетают, при этом издают тихое «ча», при сокращении расстояния издают очень резкое «ча-ча-ча-ча-ча». Подобные звуки часто можно услышать в сумерках, когда сороки устраиваются на ночлег (Linsdale, 1937). Чувствительность к шуму у сорок понижается по мере снижения температуры воздуха (Reebs, 1986).
Пробуждение сорок сопровождается характерными для этого вида сиг-налами «мааг» или «вэк-вэк-вэк»; подобные сигналы описаны и для P. p. nuttalli (Protocol for censusing.., 2003-2011).
Выводы
1. Сороки образуют коллективные ночёвки круглый год; большинство мест отдыха расположены на традиционных участках.
2. Центральные коллективные ночёвки существуют в течение всего года и могут включать от одного до трех участков для отдыха; в весенне-осенний период в пределах района сбора центральных коллективных ночёвок птицы также образуют периферические ночёвочные скопления.
3. Небольшие (до 35 особей) периферические ночёвки сороки формируют в случае удаленности их гнездовых территорий от места центральной ночёвки на расстояние, превышающее 1,5 км. Часть периферических ночёвок существует в гнездовой и ранний послегнездовой периоды на традиционных местах, осталь-ные – не привязаны к определённому участку.
4. Площадь района сбора центральных коллективных ночёвок варьирует

203
Ночёвки сорок: их организация и индивидуальные стратегии поведения
от 3,6 до 50 км², составляя, в среднем, 17 км² (n = 15). Максимальный радиус района сбора, то есть максимальное расстояние, преодолеваемое птицами в зимний период при перелёте от гнездовых территорий до места коллективного ночлега, составляет от 1,7 до 6,5 км, в среднем, 3,95 км (n = 15).
5. Чем большее число птиц участвует в образовании коллективных ночёвок в зимний период, тем больше площадь района сбора (R = 0,956; p < 0,01) и мак-симальный радиус сбора (R = 0,716; p < 0,01), а также тем большее число особей формируют коллективную ночёвку в гнездовой период (R = 0,803; p < 0,01).
6. Общая численность популяции сороки г. Харькова в разные зимние сезо-ны варьировала в пределах 6,5-8 тысяч особей, составляя, в среднем, 7360 птиц (n = 5).
7. Места традиционных периферических ночёвок расположены на рас-стоянии 1,8-4,7 км (в среднем, 3,6 км; n = 5) от мест отдыха птиц центральной ночёвки. Сороки формируют периферические ночёвки с начала апреля до августа-ноября, что зависит от удаленности участков, на которых они образуются, от мест отдыха центральной коллективной ночёвки. Места временных коллективных ночёвок могут располагаться на расстоянии 1,7-3,6 км (в среднем, 3,05 км; n = 4) от мест отдыха центральных коллективных ночёвок.
8. В гнездовой период численность коллективно ночующих сорок значитель-но выше на центральных ночёвках, чем на периферических (тест Манна-Уитни; U = 17, p < 0,01). Следовательно, в гнездовой период неразмножающиеся сороки предпочитают ночевать на местах центральных коллективных ночёвок.
9. Численность сорок на центральных ночёвках не изменяется с начала декабря до конца февраля, падает в конце марта – начале апреля, достигая ми-нимального значения в апреле-мае. Затем число птиц начинает увеличиваться, и заметными центральные ночёвочные скопления становятся с первой декады августа, но чаще – с октября-ноября.
10. В период размножения коллективные ночёвки поддерживается, в основном, за счёт неразмножающихся особей, пар, чьи гнездовые территории расположены в пределах места ночлега, и самцов, чьи гнездовые территории удалены не более чем на 500 м от места ночлега.
11. Выводки прошлого года участвуют в формировании коллективных ночёвок на традиционных местах и являются «ядром» скоплений птиц на центральных коллективных ночёвках. При этом в дневное время они могут держаться на гнездовой территории родителей до 11-месячного возраста.
12. У размножающихся пар существуют разные стратегии ночёвочного пове-дения. Выбор места ночлега между гнездовой территории и местом коллективной ночёвки определяется циклом (первый или повторный), стадией размножения и удаленностью места коллективной ночёвки от гнездовой территории. На выбор места ночлега оказывает также влияние ночёвочное поведение пар со смежных гнездовых территорий.

204
О.А. Брезгунова
13. Места центральных коллективных ночёвок сорок находятся, преимуще-ственно, в пределах пойменной террасы (80,55%): в прирусловой пойме (44,44%), в центральной части пойменной террасы в зонах её понижения (8,33%), на при-материковой части пойменной террасы, в области выклинивания грунтовых вод (27,78%), а также в пределах овражно-балочных систем (16,67%) и у берегов внепойменных озер (2,78%) (n = 36). При этом сороки ночуют в зарослях ивы пепельной и тростника (61,11%), клена ясенелистного, робинии лжеакации, лоха узколистного, гледичии обыкновенной (27,78%), реже – в кустах боярыш-ника и тёрна (8,33%) или на ветвях клёна остролистного (2,78%) (n = 36; места центральных коллективных ночёвок).
14. В течение 20-40 дней после вылета из гнезда выводки сорок из пер-вых и повторных кладок ночуют в пределах гнездовой территории родителей. В 50-65-дневном возрасте птенцы присоединяются к птицам на коллективных ночёвках. Индивидуальная жизнь у 2-месячных сорок полностью социализи-рована.
15. Сорока формирует предночёвочные скопления на гнездовых территори-ях, возле мест традиционных периферических ночёвок и по пути следования к местам ночлега на верхушках деревьев (включая хвойные), в гуще кустарников и на крышах зданий.
Абдусалямов И.А. 1973. Фауна Таджикской ССР. Том XIX. Ч. 2. Птицы. Душанбе: Изд-во «Дониш». 396 с.
Абдусалямов И.А. 1977. Фауна Таджикской ССР. Том XIX. Ч. 3. Птицы. Душанбе: Изд-во «Дониш». 275 с.
Аюпов А.Н. 1989. Зимующие врановые птицы города Ташкента // Врановые птицы в естественных и антропогенных ландшафтах: Материалы II Всесоюзн. совещ. Ч. I. Липецк. С. 142-143.
Баник М.В., Брезгунова О.А. 2007. Дериваты образа жизни предковых популяций в со-временных особенностях экологии и поведении некоторых видов птиц // Совре-менные проблемы биологической эволюции: Материалы конф. к 100 летию Гос. Дарвиновского музея (17-20 сент. 2007 г., г. Москва). М.: Изд-во ГДМ. С. 223-225.
Барабаш-Никифоров И.И., Семаго Л.Л. 1963. Птицы юго-востока Черноземного цен-тра. Воронеж. 211 с.
Бибби К., Джонс М., Марсден С. 2000. Методы полевых экспедиционных исследова-ний. Исследования и учеты птиц. М. 186 c.
Бокотей А.А. 1991. Орнитофауна Львова // Материалы X Всесоюзн. орнитол. конф. (17-20 сент. 1991 г., г. Витебск). Ч. 2. Кн. 1. Минск: Навука і тэхніка. С. 66-67.
Литература

205
Ночёвки сорок: их организация и индивидуальные стратегии поведения
Бокотей А.А. 1995а. Огляд орнітофауни міста Львова // Беркут. т. 4. Вип. 1-2. С. 3-13.
Бокотей А.А. 1995б. Атлас птиц города Львова: Основные принципы и результаты пер-вого года работы // Проблеми вивчення та охорони птахів: Матеріали VI наради орнітологів Західної України (м. Дрогобич, 1-3 лютого 1995 р.). Львів-Чернівці. С. 14-16.
Брезгунова О.А. 2005. Различия в организации коммунальных ночёвок врановых родов Сorvus и Pica // Животный мир: охрана и рациональное использование: Материа-лы научн.-практ. конф. (г. Харьков, с. Гайдары, 20-22 окт. 2005 г.). С. 62-64.
Брезгунова О.А. 2006а. Индивидуальные стратегии поведения размножающихся сорок: перемещение на коммунальную ночёвку или охрана гнездовой территории // Ор-нитологические исследования в Северной Евразии: Тез. XII Междунар. орнитол. конф. Сев. Евразии (Ставрополь, 31 янв.- 5 февр. 2006 г.). Ставрополь. С. 93-94.
Брезгунова О.А. 2006б. Ночёвки взрослых сорок и их выводков в период размножения // Бранта. Сб. научн. тр. Азово-Черноморской орнитол. станции. Вып. 9. С. 184-189.
Брезгунова О.А. 2007а. Коммунальные ночёвки сороки обыкновенной (Pica pica (L.)) в гнездовой и послегнездовой периоды // IV Всероссийская конференция по поведе-нию животных: Сб. тез. М.: Тов-во научн. изд. КМК. С. 347-348.
Брезгунова О.А. 2007б. Многовидовые ночёвочные скопления с участием сороки обык-новенной // Птицы бассейна Северского Донца. Вып. 10: Материалы 13-14 совещ. Рабочей группы «Изучение и охрана птиц бассейна Северского Донца». Харьков. С. 111-125.
Брезгунова О.О. 2008а. Колективні ночівлі воронових птахів: розподіл, типи організації та стратегії поведінки (на прикладі м. Харкова) // автореферат дис. на здобуття наук. ст. канд. біолог. наук. 03.00.08 /зоологія/. Київ. 20 с.
Брезгунова О.А. 2008б. О причинах формирования коллективных ночёвок у птиц и явлениях, связанных с общественным ночёвочным поведением // Орнитология. Вып. 35. С. 14-30.
Брезгунова О.А. 2008в. Особенности гнездовой биологии сороки в Харьковской обла-сти // Беркут. т. 17. № 1-2. С. 67-78.
Брезгунова О.А. 2009. Результаты учета численности врановых птиц на коллективных ночёвках в Харькове // Биоразнообразие и роль животных в экосистемах: Мате-риалы V Междунар. научн. конф. Днепропетровск: Лира. С. 286-287.
Брезгунова О.А. 2010а. Биотопическая характеристика мест коллективных ночё-вок сорок (Pica pica (L.)) // Врановые птицы Северной Евразии: Материалы IХ Mеждунар. конф. по изучению врановых птиц Сев. Евразии (23-26 сент. 2010 г.). Омск. С. 28-31.
Брезгунова О.А. 2010б. К вопросу о путях заселения г. Харькова сорокой (Pica pica (L.) // Птицы бассейна Северского Донца. Вып. 11. Материалы 15 научн. конф. Рабочей

206
О.А. Брезгунова
группы по птицам басейна Северского Донца, посвящ. памяти И.А. Кривицкого. Донецк: ДонНУ. С. 142-149.
Брезгунова О.А. 2010в. Совместные коллективные ночёвки сороки Pica pica (L.) и дру-гих видов птиц // Орнитология в Северной Евразии: Материалы XIII Междунар. орнитол. конф. Сев. Евразии / Тез. докл. (Оренбург, 30 апр. - 6 мая 2010 г.). Орен-бург: Изд-во Оренбургского гос. пед. ун-та, ИПК ГОУ ОГУ. С. 72-73.
Бутьев В.Т., Константинов В.М., Бабенко В.Г., Барышева И.К., Самойлов Б.Л. 1983. Зимняя авифауна г. Москвы // Влияние антропогенных факторов на структуру и функционирование биоценозов: Межвуз. сб. научн. тр. М. С. 3-36.
Гержик И.П. 1995. Массовое скопление сорок на ночёвку // Экосистемы дикой приро-ды: охрана, природопользование, мониторинг. Вып. 2. Одесса. С. 33.
Горбань И., Давидович Л. 1982. Суточные миграции сорок в г. Львове // Экологические исследования и охрана птиц Прибалтийских республик. Каунас. С. 168-169.
Горошко З.А., Кусенков А.Н. 2003. Особенности послегнездовых перемещений неко-торых воробьинообразных птиц // Современные проблемы орнитологии Сибири и Центральной Азии: Материалы II Междунар. орнитол. конф. (Россия, Улан-Удэ, 16-19 мая 2003 г.). Ч. I. Улан-Удэ: Изд-во Бурятского гос. ун-та. С. 166-168.
Гуль И.Р., Лемко Т.М. 2001. Врановые Покутья // Актуальные проблемы изучения и охраны птиц Восточной Европы и Северной Азии: Материалы XI Междунар. конф. Казань. С. 199-200.
Емцев А.А., Попов С.В., Сесин А.В. 2006. К фауне птиц севера Ханты-Мансийского автономного округа // Материалы к распространению птиц на Урале, в Приуралье и Зап. Сибири. Екатеринбург: Изд-во Уральск. ун-та. С. 75-101.
Жуков В.С. 2006. О некоторых птицах Новосибирска и его окрестностей в холодный период года, в особенности в связи с суровой зимой 2005-2006 годов // Материа-лы к распространению птиц на Урале, в Приуралье и Зап. Сибири. Екатеринбург: Изд-во Уральск. ун-та. С. 110-120.
Зарудный Н.А. 1888. Орнитологическая фауна Оренбургского края. Под ред. Ф.Д. Пле-ске // Зап. Импер. Акад. наук. т. 57. Прил. № 1. С. 1-338.
Зонов Г.Б. 1974. Географические особенности зимнего обитания млекопитающих и птиц в некоторых горных системах Восточной Сибири // Вопросы зоогеографии Сибири. Иркутск. С. 21-32.
Ильичев В.Д., Бутьев В.Т., Константинов В.М. 1987. Птицы Москвы и Подмосковья. М. 272 с. (Серия «Человек и окружающая среда»).
Калякин В.Н., Калякина Н.М. 1989. О некоторых видовых особенностях серой вороны и сороки // Врановые птицы в естественных и антропогенных ландшафтах: Мате-риалы II Всесоюзн. совещ. Ч. I. Липецк. С. 27-29.
Коноваленко (Брезгунова) О.А. 2006. Коммунальные ночёвки сорок в период размноже-

207
Ночёвки сорок: их организация и индивидуальные стратегии поведения
ния // Пiдвищення продуктивностi сiльскогосподарських тварин: Зб. наук. праць. т. 16. Харків: Харк. держ. зовет. акад. С. 435-438.
Коноваленко (Брезгунова) О.А., Коноваленко С.В. 2004. К методике учета численно-сти коммунально ночующих врановых птиц (на примере сороки обыкновенной) // Облік птахів: підходи, методики, результати (Зб. наук. ст. другої міжнар. наук.-практ. конф.). Житомир. С. 49-51.
Коноваленко (Брезгунова) О.А., Коноваленко С.В. 2005. Распределение территорий коммунальных ночёвок сороки обыкновенной (Pica pica (L.)) в г. Харькове // Пти-цы бассейна Северского Донца. Вып. 9: Материалы 11 и 12 конф. «Изучение и охрана птиц бассейна Северского Донца». Донецк. С. 47-52.
Коноваленко (Брезгунова) О.А., Кривицкий И.А., Коноваленко С.В. 2003. О групповых зимних ночёвках сорок Pica pica (Passeriformes, Corvidae) в г. Харькове // Вісн. Дніпропетр. ун-ту. Вип. 11. т. 1. С. 174-177.
Константинов В.М., Бабенко В.Г., Барышева И.К. 1982. К характеристике зимней ави-фауны и населения птиц долины р. Сумбар (Западный Копетдаг) // Орнитология. Вып. 17. С. 171-172.
Кошелев А.И., Пересадько Л.В., Калякин М.В. 1989. Особенности гнездования сороки в тростниковых зарослях на юге Украины // Врановые птицы в естественных и ан-тропогенных ландшафтах: Материалы II Всесоюзн. совещ. Ч. III. Липецк. С. 3-5.
Кошелев А.И., Покуса Р.В., Кошелев В.А. 2002. К экологии сороки на Косе Обиточной (Азовское море) // Бранта. Сб. научн. тр. Азово-Черноморской орнитол. станции. Вып. 5. С. 39-57.
Коцюруба В. 1994. Зимовки врановых в Криворожье // Матеріали 1-ї конф. молодих орнітологів України. Чернівці. С. 117-119.
Кривицкий И.А. 1999. Казахстанская «целина» и птицы. Харьков. 97 с.
Кривицкий И., Реуцкий А. 2003. Птицы в нашем городе. Харьков. 120 с.
Кричкевич Н.Г., Смогоржевский Л.А. 1977. О ночёвках сорок в лесостепной части Сумской области УССР // VII Всесоюзн. орнитол. конф.: Тез. докл. (Черкас-сы, 27-30 сент. 1977 г.). Киев: Наукова думка. С. 264.
Крушинский Л.В. 1986. Биологические основы рассудочной деятельности. Эволюцион-ный и физиолого-генетический аспекты поведения. Изд. 2-е., перераб. и доп. М.: Изд-во МГУ. 272 с.
Кузнецова С.С., Правдина Г.М. 1977. Сезонная вариабельность суточных физиологиче-ских ритмов // Эколого-физиологические исследования в природе и эксперименте: Тез.V Всесоюзн. конф. по экологич. физиологии, биохимии и морфологии. Фрун-зе. С. 39.
Куранов Б.Д. 1979. Изучение распределения сороки в гнездовой период с помощью ме-чения // Экология гнездования птиц и методы её изучения: Тез. Всесоюзн. конф.

208
О.А. Брезгунова
молодых ученых (Самарканд, 23-25 мая 1979 г.). Самарканд. С. 122-123.
Лысенков Е.В., Спиридонов С.Н., Константинов В.М., Лапшин А.С. 2004. Экология и биоценотическое значение врановых птиц Мордовии. Саранск. 230 с.
Маловичко Л.В., Гаврилов А.И., Зубалий А.М. 2010. Значение степных рек в распреде-лении врановых птиц в Ставропольском крае // Врановые птицы Северной Евра-зии: Материалы IX Междун. научн.-практ. конф. (Омск, 23-26 сент. 2010 г.). Омск. C. 83-85.
Мальчевский А.С., Пукинский Ю.Б. 1983. Птицы Ленинградской области и сопредель-ных территорий: История, биология, охрана. В 2 х томах. т. 2. Л.: Изд-во Ленингр. ун-та. 504 с.
Мороз В.А. 2009. К экологии сороки в Провальской степи (Луганская область) // При-родничий альманах: зб. наук. праць. № 13. С. 114-123. (Сер.: Біологічні науки).
Надточий А.С., Зиоменко С.К. 2001. Врановые птицы города Харькова // Актуальные проблемы изучения и охраны птиц Восточной Европы и Северной Азии: Материа-лы XI Междунар. конф. Казань. С. 461-462.
Нечаев В.А. 1990. Материалы к биологии сороки – Pica pica (L.) в Южном Приморье // Экология и распространение птиц юга Дальнего Востока. С. 49-54.
Осмоловская В.И., Формозов А.Н. 1950. Очерки экологии некоторых полезных птиц леса. // А.Н. Формозов, В.И. Осмоловская, К.Н. Благосклонов. Птицы и вредители леса. Сер. «Среди природы». Вып. 19. М.: Изд-во МОИП. С. 34-142.
Пантелеев П.А. 1974. Об изучении зимовок врановых в городах // Материалы VI Всесо-юзн. орнитол. конф. (1-5 февр. 1974 г., г. Москва). Ч. II. М. С. 348-349.
Пасічник А.О. 1995. Гніздове розміщення сороки у Львові // Беркут. т. 4. Вип. 1-2. С. 47-49.
Петрович З.О., Редінов К.О. 2007. Значення Кінбурнської коси в збереженні орлана-білохвоста в зимовий період // Бранта. Сб. научн. тр. Азово-Черноморской орни-тол. станции. Вып. 10. С. 156-164.
Полозов С.А., Бурнашев С.И. 1989. Особенности размещения и поведения врановых птиц Западного Копетдага // Врановые птицы в естественных и антропогенных ландшафтах: Материалы II Всесоюз. совещ. Липецк. Ч. II. С. 53-55.
Промптов А.Н. 1940. Изучение суточной активности птиц в гнездовой период // Зоол. журн. т. 19. Вып. 1. С. 143-159.
Рогачева Э.В., Сыроечковский Е.Е., Черников О.А. 2008. Птицы Эвенкии и сопредель-ных территорий. М.: Тов-во науч. изд. КМК. 754 с.
Родимцев А.С., Шкарин В.С. 1984. К размножению сороки в Кузбассе // Гнездовая жизнь птиц (Межвуз. сб. научн. трудов). Пермь. С. 27-35.
Сапетина И.М. 2009. Птицы Окского заповедника и сопредельных территорий (био-логия, численность, охрана). т. 2. Воробьиные птицы. М.: Изд-во научн. изд.

209
Ночёвки сорок: их организация и индивидуальные стратегии поведения
КМК. 172 с.
Северцов Н.А. 1950. Периодические явления в жизни зверей, птиц и гад Воронежской губернии. М.: Изд-во АН СССР. 308 с. (по изданию 1855 года).
Семаго Л.Л. 1975. Сто свиданий с природой. Воронеж: Изд-во Воронежского ун-та. 262 с.
Сеник М.А. 2007. Особенности зимовок грача и других врановых в г. Львове // Эко-логия врановых в естественных и антропогенных ландшафтах: Материалы VIII Междунар. конф. по врановым птицам. С. 143-146.
Сметана Н.М. 1978а. Сезонные и суточные миграции сорок в Наурзумском заповеднике // Вторая Всесоюзн. конф. по миграциям птиц: Тез. сообщ. М. С. 142-144.
Сметана Н.М. 1978б. Экология сороки в Наурзумском заповеднике (Северный Казах-стан) в период инкубации яиц и выкармливания птенцов // Биология птиц в Казах-стане: Тр. Ин-та зоол. т. 38. Алма-Ата. С. 20-28.
Сомов Н.Н. 1897. Орнитологическая фауна Харьковской губернии. Харьков: тип. А. Дарре. IX, 194, 680 с.
Сотников В.Н. 2006. Птицы Кировской области и сопредельных территорий. т. 2. Во-робьинообразные. Ч. 1. Киров: ООО «Триада плюс». 448 с.
Спангенберг Е.П. 1953. Из жизни натуралиста. М.: Мол. гвардия. 526 с.
Стрижельчик Г.Г., Соколов Ю.П., Гольдфельд И.А., Чебанов А.Ю., Николенко Н.С. 2003. Подтопление в населенных пунктах Харьковской области. Харьков. 160 с.
Терновский Д.В., Терновская Ю.Г. 1969. Учёт численности сорок и обыкновенных скворцов на ночёвках // Орнитология в СССР: Материалы V Всесоюзн. орнитол. конф. Кн. 2. Ашхабад. С. 646-649.
Федосенко А.К., Орлов Г.И. 1969. Зимнее питание сороки в предгорьях Заилийского Алатау (Тянь-Шань) // Орнитология в СССР: Материалы V Всесоюзн. орнитол. конф. Кн. 2. Ашхабад. С. 658-661.
Формозов А.Н. 1947. Фауна // Природа города Москвы и Подмосковья. М.-Л.: Изд-во АН СССР. С. 287-370.
Хмельков С.А. 1998. К экологии воробьиных птиц на ночёвках // Мордовск. орнитол. вестн.: Сб. научн. трудов. Вып. 1. Саранск. С. 105-107.
Холодковский Н.А., Силантьев А.П. 1901. Птицы Европы. Ч. II. Специальная орнитоло-гия. СПб.: Изд. А.Ф. Девриена. 636 с.
Хохлов А.Н. 1983. Сравнительная экология и практическое значение массовых видов врановых птиц в антропогенных ландшафтах Ставропольского края // Авторефе-рат дисс. на соиск. уч. ст. канд. биол. наук. М. 16 с.
Хохлова Н.А., Кривцов С.К. 1984. Изменчивость биологии сороки при гнездовании в населенных пунктах // Наземные и водные экосистемы. Вып. 7. С. 22-29.

210
О.А. Брезгунова
Baeyens G. 1979. Description of the social behaviour of the Magpie (Pica pica) // Ardea. vol. 67. P. 28-41.
Baeyens G. 1981a. Functional aspects of serial monogamy: The magpie pair-bond in relation to it’s territorial system // Ardea. vol. 69. P. 145-166.
Baeyens G. 1981b. The role of the sexes in territory defense in the Magpie (Pica pica) // Ardea. vol. 69. P. 69-82.
Bährmann U.1968. Die Elster. Die Brehm-Bücherei. Nr. 393. Wittenberg Lutherstadt: A. Ziemsen Verlag. 72 S.
Berndt R.K. 1988. Zum Schlafplatzflug der Amsel (Turdus merula) am Kieler Stadtrand // Corax. Bd. 13. S. 82-90.
Birkhead T.R. 1989. Studies of West Palearctic birds 189. The Magpie // British Birds. vol. 2. № 12. P. 583-600.
Birkhead T.R. 1991. The Magpies. The Ecology and Behavior of Black-billed and Yellow-billed Magpies. New York: Academic Press. 270 р.
Birkhead T.R., Eden S.F., Clarkson K., Goodburn S.F., Pellatt J. 1986. Social organization of a population of Magpies Pica pica // Ardea. vol. 74. P. 59-68.
Blake-Knox H. 1868. Ornithological notes from the County Dublin for 1867 // Zoologist. P. 1401-1411.
Blanco G., Tella J.L. 1999. Temporal, spatial and social segregation of red-billed choughs between two types of communal roost: a role for mating and territory acquisition // Anim. Behav. vol. 57. № 6. P. 1219-1227.
Bokotey A.A. 1997. Numbers and distribution of Magpie Pica pica in Lvov (Ukraine) // Acta ornithol. vol. 32. № 1. P. 5-7.
Bree W.T. 1862. Gathering of magpies // Zoologist. vol. 20. P. 7846.
Brennecke H.-E. 1965. Notizen an einem schleswig-holsteinischen Elsternschlafplatz // Corax. Bd. 1. S. 82-87.
Brown R.H. 1924. Field-notes on the Magpie, as observed in Cumberland // British Birds. vol. 18. № 5. P. 122-128.
Buitron D. 1988. Female and male specialization in parental care and its consequences in Black-billed Magpies // Condor. vol. 90. P. 29-39.
Buttermer W.A. 1994. Size and energy considerations in avian winter roosting habits // J. Orn. Bd. 135. Sonderheft. Research Notes on Avian Biology 1994: selected contributions from the 21-st International Ornithological Congress. P. 429.
Coombs F. 1978. The Crows. A study of the Corvids of Europe. London: B.T. Batsford Ltd. 255 p.
Coulson J.C. 1963. Improved coloured rings // Bird study. vol. 10. Iss. 2. P. 109-111.
Cramp S., Perrins C.M. (eds.). 1994. The Birds of the Western Palearctic: Handbook of the

211
Ночёвки сорок: их организация и индивидуальные стратегии поведения
Birds of Europe, the Middle East, and North Africa. Vol. VIII. Crows to Finches. Oxford: Oxford University Press. 899 p.
Criddle M. 1923. The American magpie in Manitoba // Can. Field-Natur. vol. 37. № 2. P. 25-26.
Crosbie S.P., Bell D.A., Bolen G.M. 2006. Vegetative and thermal aspects of roost-site selection in urban Yellow-billed Magpies // Wilson J. Orn. vol. 118. № 4. P. 532-536.
Crosbie S.P., Koenig W.D., Reisem W.K., Kramer V.L., Marcus L., Carney R., Pandolfino E., Bolen G.M., Crosbie L.R., Bell D.A., Ernest H.B. 2008. Early impact of West Nile virus on the Yellow-billed Magpie (Pica nuttalli) // Auk. vol. 125. № 3. P. 542-550.
Czechowski P., Jerzak L. 2003. The roosting behaviour of the magpie Pica pica in an urban environment // Vogelwarte. vol. 42. Abstr. 4th Conference of the European Ornithologists’ Union (Chemnitz, Germany, 16-21 august 2003). Links and perspectives in European ornithology. № 1-2. S. 14-15.
Czechowski P., Jerzak L., Zduniak P., Bochenski M. 2005. Is the communal roosting behaviour of the Magpie (Pica pica) wind dependent? An example from an isolated population in W. Poland // Ardeola. vol. 52. № 2. P. 333-339.
Deckert G. 1980. Siedlungsdichte und Nahrungssuche bei Elster Pica p. pica (L.), und Nebelkrähe, Corvus corone cornix (L.) // Beitz. Vogelk. Bd. 26. № 6. S. 305-334.
Dice L.R. 1917. Habits of the Magpie in Southeastern Washington // Condor. vol. 19. P. 121-124.
Erpino M.J. 1968. Nest-related activities of Black-billed Magpies // Condor. vol. 70. P. 154-165.
Erpino M.J. 1969. Seasonal cycle of reproductive physiology in the Black-billed Magpie // Condor. vol. 71. P. 267-279.
Georgiev K., Iliev M. 2009. Numbers and arrival dynamics of communally roosting Magpies (Pica pica) in the dendrological park of the University of forestry – Sofia, Bulgaria // Biotechnol. & Biotechnol. EQ. Vol. 23. Special Edition On-line. XI Anniversary scientific conference 120 years of academic education in biology 45 years faculty of biology. P. 250-253.
Goodwin D. 1976. Crows of the World. Ithaca, New York: Comstock publ. assoc., the British Museum (Natural History). 354 p.
Gorska E. 1998. The influence of weather factors on the daily activity of urban populations of birds at their common roosts in Slupsk // Acta Zool. Cracov. vol. 41. № 1. Р. 35-49.
Gill B.J., Dow D.D. 1985. Waking and roosting of Grey-crowned Babblers Pomatostomus temporalis in South-east Queensland during spring // Emu. vol. 85. №. 2. P. 97-105.
Grinnell J. 1923. Notes on some birds observed in the vicinity of Colusa, California // Condor. vol. 25. P. 172-176.
Gyllin R., Källander H. 1977. Sovsträck och övernattning hos skaten Pica pica // Fauna och

212
О.А. Брезгунова
Flora. vol. 72. P. 18-24.
Hadfield H.1874. Remarks on birds seen during a three weeks’ tour in Brittany // Zoologist. P. 3945-3947.
Hall T.C. 1994. Magpies // Hygnstrom S., Timm R., Larson G. (eds.). Prevention and control of wildlife damage. Lincoln: University of Nebraska. P. E79-E85.
Huber J. 1944. Aus dem Leben der Elster im Sempacherseegebiet // Orn. Beob. Bd. 41. S. 1-7.
Koenig W., Reynolds M.D. 2009. Yellow-billed Magpie (Pica nuttalli) // The Birds of North America Online (A. Poole, Ed.). Ithaca: Cornell Lab of Ornithology; Retrieved from the Birds of North America Online: http://bna.birds.cornell.edu/bna/species/180.
Lee S., Parr C.S., Hwang Y., Mindell D.P., Choe. J.C. 2003. Phylogeny of magpies (genus Pica) inferred from mtDNA data // Mol. Phylogen. Evol. vol. 29. № 2. Р. 250-257.
Lee W.Y., Lee S., Jablonski P.G., Choe. C.J. 2008. Genetic Relationships within a Communal winter roost of Magpies using microsatellite markers // SNU J. Stud. Res. vol. 2. 6 p.
Linsdale J.M. 1937. The Natural History of Magpies // Pacific coast avifauna. vol. 25. P. 1-134.
Linsdale J.M. 1946. American Magpie (Pica pica hudsonia) // Bent A.C. (Ed.). Life Histories of North American jays, crows, and titmice. U.S. Nat. Mus. Bull. vol. 191. P. 133-155.
Mäck U. 1998. Populationsbiologie und Raumnutzung der Elster (Pica pica Linnaeus 1758) in einem urbanen Ökosystem – Untersuchungen im Großraum Ulm // Ökol. Vögel. Bd. 20. Ht. 1. S. 1 215.
Mäck U., Jürgens M.-E. 1999. Aaskrähe, Elster und Eichelhäher in Deutschland. Bericht über den Kenntnisstand und die Diskussionen zur Rolle von Aaskrähe (Corvus corone), Elster (Pica pica), und Eichelhäher (Garrulus glandarius) im Naturhaushalt sowie die Notwendigkeit eines Bestandsmanagements. Bundesamt für Naturschutz. 252 S.
McGregor P.J.C. 1906. Notes on birds observed at Monastir, Turkey in Europe // Ibis. Ser. 8. vol. 6. P. 285 307.
Møller A.P. 1983. Habitat selection and feeding activity in the Magpie Pica pica // J. Orn. vol. 124. № 2. P. 147-161.
Møller A. P. 1985. Communal roosting in the Magpie (Pica pica) // J. Orn. vol. 126. № 4. P. 405-419.
Peh K. S.-H. 2002. Roosting behaviour of House Crow (Corvus splendens) in relation to environmental variables // The Raffles Bull. vol. 50. Iss. 1. P. 257-262.
Perlmutter G.B. 1992. Environmental factors influencing roost arrival of Black-crowned Night-Herons // J. Field Orn. vol. 63. № 4. P. 462-465.
Pfister O. 2004. Birds and Mammals of Ladakh. New Delhi: Oxford University Press. 361 p.
Ponz M.A., Monrós G. J.S. 2000. Dos factores que influyen en el establecimiento de

213
Ночёвки сорок: их организация и индивидуальные стратегии поведения
los dormideros comunales en la urraca (Pica pica): las actividades humanas y las temperaturas mínimas // Ardeola. vol. 47. P. 85-87.
Protocol for censusing Yellow-billed Magpies (Pica nuttalli) at communal roosts. 2003-2011. (www.prbo.org/calpif/pdfs/YBMA.pdf )
Reebs S.G. 1986. Sleeping behavior of Black-billed Magpies under a wide range of temperatures // Condor. vol. 88. P. 524-526.
Reebs S.G. 1987. Roost characteristics and roosting behavior of Black-billed Magpies Pica pica, in Edmonton, Alberta // Can. Field-Nat. vol. 101. P. 519-525.
Russelli K.R., Gauthreaux J.R. 1999. Spatial and temporal dynamics of a Purple Martin pre-migratory roost // Wilson Bull. vol. 111. Iss. 3. P. 354-362.
Schäfer E. 1938. Ornithologische Ergebnisse zweier Forschungsreisen nach Tibet // J. Orn. Bd. 86. Sonderheft May 1938. S. 261-279.
Splendoria F. 1971. Roost ecology of the Black-billed Magpie (Pica pica hudsonia). Master’s Thesis. Pocatello: Idaho State Univ.
Stiefel A. 1979. Ruhe und Schlaf bei Vögeln. Wittenberg Lutherstadt: A. Ziemsen Verlag. 216 S.
Trost C.H. 1999. Black-billed Magpie (Pica hudsonia), The Birds of North America Online (A. Poole, Ed.). Ithaca: Cornell Lab of Ornithology; Retrieved from the Birds of North America Online: http://bna.birds.cornell.edu/bna/species/389
Urquhart D. 1998. East Mersea in winter // The Essex bird report 1997. P. 141-158.
Utley J.P. 1943. A North Yorkshire Magpie roost // British Birds. vol. 36. №. 8. P. 159-160.
Verbeek N.A.M. 1973. The exploitation system of the Yellow-billed Magpie. Berkeley-Los-Angeles-London: University of California press. 58 p.
Ward E. 1952. Some observations at a Magpie roost // British Birds. vol. 45. № 11. P. 403-405.
Copyright © 2022 FDOKUMEN




















