
Tác động của chính sách giám sát an toàn vĩ mô tới giá bất ...
Upload
khangminh22Category
view
3download
0
BỘ GIÁO DỤC VÀ ĐÀO TẠO VIỆN HÀN LÂM KHOA HỌC
VÀ CÔNG NGHỆ VIỆT NAM
HỌC VIỆN KHOA HỌC VÀ CÔNG NGHỆ
-------------------------------
HUỲNH THỊ LŨY
NGHIÊN CỨU TẠO, NHÂN PHÔI VÔ TÍNH
VÀ RỄ BẤT ĐỊNH CÂY NGŨ GIA BÌ CHÂN CHIM
(Schefflera octophylla Lour. Harms)
LUẬN ÁN TIẾN SĨ SINH HỌC
THÀNH PHỐ HỒ CHÍ MINH - 2022

BỘ GIÁO DỤC VÀ ĐÀO TẠO VIỆN HÀN LÂM KHOA HỌC
VÀ CÔNG NGHỆ VIỆT NAM
HỌC VIỆN KHOA HỌC VÀ CÔNG NGHỆ
------------------------------
NGHIÊN CỨU TẠO, NHÂN PHÔI VÔ TÍNH
VÀ RỄ BẤT ĐỊNH CÂY NGŨ GIA BÌ CHÂN CHIM
(Schefflera octophylla Lour. Harms)
Chuyên ngành: Sinh lý học thực vật
Mã số: 9 42 01 12
LUẬN ÁN TIẾN SĨ SINH HỌC
NGƯỜI HƯỚNG DẪN KHOA HỌC:
1. TS. Nguyễn Hữu Hổ
2. PGS.TS. Bùi Văn Lệ
THÀNH PHỐ HỒ CHÍ MINH – 2022
HUỲNH THỊ LŨY

i
LỜI CAM ĐOAN
Tôi xin cam đoan luận án: “Nghiên cứu tạo, nhân phôi vô tính và rễ bất
định cây Ngũ gia bì chân chim (Schefflera octophylla Lour. Harms)” là công trình
nghiên cứu của tôi, dưới sự hướng dẫn khoa học của TS. Nguyễn Hữu Hổ và PGS.TS.
Bùi Văn Lệ. Nội dung nghiên cứu và các kết quả trong đề tài này hoàn toàn trung
thực. Toàn bộ số liệu và kết quả nghiên cứu chưa từng được sử dụng để công bố trong
các công trình nghiên cứu để nhận học vị, các thông tin trích dẫn trong luận án này
đều được trích dẫn nguồn gốc rõ ràng. Tôi xin hoàn toàn chịu trách nhiệm về sự cam
đoan này.
Tp. Hồ Chí Minh, ngày 18 tháng 05 năm 2022
Tác giả
Huỳnh Thị Lũy

ii
LỜI CẢM ƠN
Tôi xin trân trọng gửi lời cảm ơn sâu sắc nhất đến PGS.TS. Bùi Văn Lệ và đặc
biệt TS. Nguyễn Hữu Hổ, là những người Thầy đã tận tâm hướng dẫn, giúp đỡ, truyền
đạt những kiến thức quý báu để tôi hoàn thành luận án này.
Tôi xin gửi lời tri ân sâu sắc đến Viện Hàn lâm Khoa học và Công nghệ Việt
Nam, Phòng Đào tạo, Học viện Khoa học và Công nghệ, Viện Sinh học nhiệt đới đã
tạo điều kiện thuận lợi cho tôi học tập và thực hiện nghiên cứu đề tài.
Tôi xin gửi lời cảm ơn chân thành đến Sở Giáo dục và Đào tạo Phú Yên,
Trường THPT chuyên Lương Văn Chánh Phú Yên, đã tạo điều kiện cho tôi học tập,
nghiên cứu.
Xin bày tỏ lòng biết ơn sâu sắc đến các anh chị em của phòng Công nghệ gen
– Viện Sinh học nhiệt đới, đã rất tận tình tạo mọi điều kiện về cơ sở vật chất cũng
như thời gian quý báu để tôi thực hiện nghiên cứu đề tài.
Sự quan tâm và động viên của các Thầy Cô, anh chị em của Viện Sinh học
nhiệt đới là động lực lớn cho tôi thực hiện đề tài nghiên cứu. Chân thành cảm ơn!
Tp. Hồ Chí Minh, ngày 18 tháng 05 năm 2022
Tác giả
Huỳnh Thị Lũy

iii
MỤC LỤC
LỜI CAM ĐOAN ....................................................................................................... i
LỜI CẢM ƠN ............................................................................................................ ii
DANH MỤC CÁC KÝ HIỆU, CÁC CHỮ VIẾT TẮT ........................................... vii
DANH MỤC CÁC BẢNG ........................................................................................ ix
DANH MỤC CÁC HÌNH, ĐỒ THỊ ......................................................................... xi
TÓM TẮT ................................................................................................................. xv
SUMMARY ........................................................................................................... xvii
MỞ ĐẦU ..................................................................................................................... 1
1. Tính cấp thiết của đề tài .......................................................................................... 1
2. Mục tiêu của đề tài .................................................................................................. 2
2.1. Mục tiêu tổng quát .......................................................................................................... 2
2.2. Mục tiêu cụ thể ............................................................................................................... 2
3. Ý nghĩa khoa học và thực tiễn của đề tài ................................................................ 3
3.1. Ý nghĩa khoa học ............................................................................................................ 3
3.2. Ý nghĩa thực tiễn ............................................................................................................. 3
4. Đối tượng và phạm vi nghiên cứu ........................................................................... 3
4.1. Đối tượng nghiên cứu ..................................................................................................... 3
4.2. Phạm vi nghiên cứu ........................................................................................................ 3
5. Tính mới của đề tài .................................................................................................. 4
CHƯƠNG 1. TỔNG QUAN TÀI LIỆU ..................................................................... 5
1.1. Giới thiệu về Ngũ gia bì chân chim (Schefflera octophylla Lour. Harms) .......... 5
1.1.1. Nguồn gốc và phân bố ................................................................................................. 5
1.1.2. Đặc điểm sinh học – sinh thái ...................................................................................... 6
1.1.3. Thành phần hóa học và công dụng .............................................................................. 7
1.1.3.1. Thành phần hóa học .................................................................................................. 7
1.1.3.2. Công dụng ............................................................................................................... 10
1.2. Tình hình nghiên cứu về phôi vô tính, rễ bất định ở một số loài thuộc các chi
quan trọng ở họ Ngũ gia bì (Araliaceae) ................................................................... 11
1.2.1. Chi Panax .................................................................................................................. 11
1.2.1.1. Phôi vô tính ............................................................................................................. 11
1.2.1.2. Rễ bất định .............................................................................................................. 14
1.2.2. Chi Acanthopanax ...................................................................................................... 15
1.2.2.1. Phôi vô tính ............................................................................................................. 15
1.2.2.2. Rễ bất định .............................................................................................................. 16
1.2.3. Chi Polyscias ............................................................................................................. 17
1.2.4. Chi Schefflera ............................................................................................................ 18
1.2.4.1. Schefflera octophylla (Lour.) Harms ...................................................................... 18

iv
1.2.4.2. Schefflera arboricola (Hayata) Merr. ..................................................................... 18
1.2.4.3. Schefflera leucantha Viguier ................................................................................. 19`
1.2.4.4. Didymopanax morototoni (Aublet) Decaisne et Planchon. .................................... 19
1.3. Sự phát sinh phôi vô tính ................................................................................... 20
1.3.1. Cơ sở khoa học của sự phát sinh phôi vô tính ........................................................... 20
1.3.2. Một số nghiên cứu về sự phát sinh phôi vô tính ........................................................ 22
1.4. Sự hình thành rễ bất định .............................................................................................. 29
1.4.1. Cơ sở khoa học của sự hình thành rễ bất định ........................................................... 29
1.4.2. Một số nghiên cứu về sự hình thành rễ bất định ........................................................ 31
CHƯƠNG 2. VẬT LIỆU VÀ PHƯƠNG PHÁP ...................................................... 35
2.1. Nguồn mẫu – Vật liệu nuôi cấy .................................................................................... 35
2.1.1. Nguồn mẫu ...................................................................................................... 35
2.1.2. Tạo vật liệu nuôi cấy ban đầu ......................................................................... 35
2.2. Nội dung nghiên cứu ..................................................................................................... 36
2.2.1. Nội dung 1. Tạo phôi vô tính .......................................................................... 36
2.2.2. Nội dung 2. Nhân phôi vô tính ........................................................................ 36
2.2.3. Nội dung 3. Tạo rễ bất định ............................................................................ 36
2.2.4. Nội dung 4. Nhân rễ bất định .......................................................................... 36
2.3. Nội dung 1. Tạo phôi vô tính ............................................................................. 39
2.3.1. Tạo phôi vô tính trực tiếp từ mô lá ............................................................................ 39
2.3.1.1. Ảnh hưởng của auxin (NAA/IBA) và môi trường khoáng đến sự tạo phôi trực tiếp
từ mô lá ................................................................................................................................ 39
2.3.1.2. Ảnh hưởng của sự kết hợp auxin và BA đến sự tạo phôi trực tiếp từ mô lá ........... 39
2.3.1.3. Ảnh hưởng của nồng độ đường và điều kiện chiếu sáng đến sự tạo phôi trực tiếp từ
mô lá .................................................................................................................................... 40
2.3.1.4. Ảnh hưởng của nước dừa đến sự tạo phôi trực tiếp từ mô lá .................................. 40
2.3.1.5. Tạo cây con in vitro hoàn chỉnh từ phôi vô tính ..................................................... 40
2.3.1.6. Trồng cây con ở chậu đất ........................................................................................ 41
2.3.2. Tạo phôi vô tính gián tiếp qua mô sẹo từ nuôi cấy mô lá .......................................... 41
2.3.2.1. Tạo phôi vô tính gián tiếp qua mô sẹo mảnh lá (10 x 10 mm) ............................... 41
2.3.2.2. Tạo phôi vô tính gián tiếp qua mô sẹo mảnh lá (3 x 10 mm) ................................. 43
2.4. Nội dung 2. Nhân phôi vô tính ........................................................................... 44
2.4.1. Nhân phôi trên môi trường đặc ....................................................................... 44
2.4.2. Nhân phôi trong môi trường lỏng .............................................................................. 44
2.4.2.1. Ảnh hưởng của chất điều hòa sinh trưởng đến sự tạo phôi thứ cấp qua nuôi lỏng lắc
............................................................................................................................................. 44
2.4.2.2. Ảnh hưởng của khối lượng phôi nuôi cấy đến sự tăng trưởng sinh khối phôi ....... 45
2.4.2.3. Ảnh hưởng của kích thước phôi nuôi cấy đến sự tạo phôi thứ cấp ......................... 45

v
2.4.2.4. Ảnh hưởng của nồng độ đường đến sự tăng trưởng sinh khối phôi ....................... 45
2.4.2.5. Ảnh hưởng của cường độ ánh sáng đến sự tăng trưởng sinh khối phôi.................. 45
2.4.2.6. Ảnh hưởng của tỷ lệ nước dừa đến sự tạo phôi thứ cấp ......................................... 46
2.4.2.7. Tạo cây con từ phôi nuôi lỏng lắc ........................................................................... 46
2.4.3. Quan sát cấu trúc giải phẫu phôi sơ cấp và thứ cấp ................................................... 46
2.4.4. Các chỉ tiêu theo dõi và phương pháp xác định ......................................................... 46
2.5. Nội dung 3. Tạo rễ bất định .......................................................................................... 46
2.5.1. Tạo rễ bất định trực tiếp từ mô lá .............................................................................. 46
2.5.1.1. Ảnh hưởng của auxin (NAA/IBA) và môi trường khoáng đến sự tạo rễ bất định
trực tiếp từ mô lá .................................................................................................................. 46
2.5.1.2. Ảnh hưởng của nồng độ đường đến sự tạo rễ bất định trực tiếp từ mô lá .............. 47
2.5.1.3. Ảnh hưởng của điều kiện chiếu sáng đến sự tạo rễ bất định trực tiếp từ mô lá ...... 47
2.5.1.4. Khảo sát hình thái giải phẫu rễ tái sinh trực tiếp từ mô lá ...................................... 48
2.5.2. Tạo rễ bất định từ chồi ............................................................................................... 48
2.5.2.1. Tạo rễ bất định từ chồi có nguồn gốc từ đốt thân cây vườn ươm ........................... 48
2.5.2.2. Tạo rễ bất định từ chồi đốt thân cây in vitro ........................................................... 49
2.5.2.3. Tạo rễ bất định từ chồi có nguồn gốc phôi vô tính ................................................. 49
2.6. Nội dung 4. Nhân rễ bất định trong môi trường lỏng ........................................ 50
2.6.1. Ảnh hưởng của chất điều hòa sinh trưởng đến sự phân nhánh của rễ ....................... 50
2.6.2. Ảnh hưởng của nồng độ đường đến sự tăng trưởng sinh khối rễ .............................. 50
2.6.3. Ảnh hưởng của khối lượng rễ nuôi cấy đến sự tăng trưởng sinh khối rễ .................. 50
2.6.4. Khảo sát diễn biến tăng trưởng sinh khối rễ theo thời gian ....................................... 50
2.7. Điều kiện nuôi cấy in vitro ........................................................................................... 51
2.8. Phương pháp thống kê và xử lý số liệu ......................................................................... 51
CHƯƠNG 3. KẾT QUẢ VÀ THẢO LUẬN ............................................................ 52
3.1. Tạo phôi vô tính ................................................................................................. 52
3.1.1. Tạo phôi vô tính trực tiếp từ mô lá ............................................................................ 52
3.1.1.1. Ảnh hưởng của auxin (NAA/IBA) và môi trường khoáng đến sự tạo phôi vô tính
trực tiếp từ mô lá .................................................................................................................. 52
3.1.1.2. Ảnh hưởng của sự kết hợp NAA và BA đến sự tạo phôi vô tính trực tiếp từ mô lá
............................................................................................................................................. 57
3.1.1.3. Ảnh hưởng của nồng độ đường và điều kiện chiếu sáng đến sự tạo phôi vô tính
trực tiếp từ mô lá .................................................................................................................. 60
3.1.1.4. Ảnh hưởng của tỷ lệ nước dừa đến sự tạo phôi vô tính trực tiếp từ mô lá ............. 62
3.1.1.5. Tạo cây con hoàn chỉnh từ phôi vô tính .................................................................. 64
3.1.1.6. Trồng cây con ở chậu đất ........................................................................................ 66
3.1.2. Tạo phôi vô tính gián tiếp .......................................................................................... 66
3.1.2.1. Tạo phôi vô tính gián tiếp qua mô sẹo mảnh lá (10 x 10 mm) ............................... 66
3.1.2.2. Tạo phôi vô tính gián tiếp qua mô sẹo mảnh lá (3 x 10 mm) ................................. 76

vi
3.2. Nhân phôi vô tính ............................................................................................... 80
3.2.1. Nhân phôi trên môi trường đặc .................................................................................. 80
3.2.2. Nhân phôi trong môi trường lỏng .............................................................................. 82
3.2.2.1. Ảnh hưởng của chất điều hòa sinh trưởng đến sự tạo phôi thứ cấp ........................ 82
3.2.2.2. Ảnh hưởng của khối lượng phôi nuôi cấy đến sự tăng trưởng sinh khối phôi ....... 83
3.2.2.3. Ảnh hưởng của kích thước phôi đến sự tạo phôi thứ cấp ....................................... 86
3.2.2.4. Ảnh hưởng của nồng độ đường đến sự tăng trưởng sinh khối phôi ....................... 88
3.2.2.5. Ảnh hưởng của cường độ ánh sáng đến sự tăng trưởng sinh khối phôi.................. 90
3.2.2.6. Ảnh hưởng của tỷ lệ nước dừa đến sự hình thành phôi thứ cấp ............................. 91
3.2.2.7. Tạo cây con từ phôi nuôi lỏng lắc ........................................................................... 92
3.2.3. Quan sát cấu trúc giải phẫu phôi sơ cấp và phôi thứ cấp ........................................... 93
3.3. Tạo rễ bất định ................................................................................................... 96
3.3.1. Tạo rễ bất định trực tiếp từ mô lá .............................................................................. 96
3.3.1.1. Ảnh hưởng của auxin (NAA/IBA) và môi trường khoáng đến sự tạo rễ bất định
trực tiếp từ mô lá .................................................................................................................. 96
3.1.1.2. Ảnh hưởng của nồng độ đường đến sự tạo rễ bất định trực tiếp từ mô lá ............ 104
3.1.1.3. Ảnh hưởng của điều kiện chiếu sáng đến sự tạo rễ bất định trực tiếp từ mô lá .... 106
3.1.1.4. Minh họa sự tái sinh rễ trực tiếp và khảo sát hình thái giải phẫu rễ tái sinh trực tiếp
........................................................................................................................................... 109
3.3.2. Tạo rễ bất định từ chồi ............................................................................................. 111
3.3.2.1. Tạo rễ bất định từ chồi có nguồn gốc từ đốt thân cây vườn ươm ......................... 111
3.3.2.2. Tạo rễ bất định từ chồi đốt thân cây in vitro ......................................................... 113
3.3.2.3. Tạo rễ bất định từ chồi có nguồn gốc phôi vô tính ............................................... 115
3.4. Nhân rễ bất định trong môi trường lỏng .......................................................... 117
3.4.1. Ảnh hưởng của NAA, IBA đến sự phân nhánh rễ ........................................ 117
3.4.2. Ảnh hưởng của nồng độ đường đến sự tăng trưởng sinh khối rễ ............................ 121
3.4.3. Ảnh hưởng của khối lượng rễ nuôi cấy đến tăng trưởng sinh khối của rễ ............... 123
3.4.4. Khảo sát diễn biến tăng trưởng sinh khối rễ theo thời gian ..................................... 124
KẾT LUẬN VÀ KIẾN NGHỊ................................................................................. 127
KẾT LUẬN ............................................................................................................. 127
KIẾN NGHỊ ............................................................................................................ 128
TÀI LIỆU THAM KHẢO ....................................................................................... 130
PHỤ LỤC

vii
DANH MỤC CÁC KÝ HIỆU, CÁC CHỮ VIẾT TẮT
AUX Auxin
2,4-D 2,4-Dichlorophenoxyacetic acid
2-iP 6-dimethylallylamino purine
ABA Abscisic acid
BA Benzyl adenine
CK Cytokinin
ĐC Đối chứng
ĐHST Điều hòa sinh trưởng
GA3 Gibberellic acid
HPLC High-Performance Liquid Chromatography
IAA Indole-3-acetic acid
IBA Indole-3-butyric acid
IEDC Induced Embryogenic Determined Cell
KNSP Khả năng sinh phôi
KLP Khối lượng phôi
KLTP Khối lượng tươi phôi
LMTB Lát mỏng tế bào
MS Murashige và Skoog
MSSP Mô sẹo sinh phôi
NAA α-Naphthaleneacetic acid
NGBCC Ngũ gia bì chân chim
NSC Ngày sau cấy
PEDC Pre-Embryogenic Determined Cell
RSV Respiratory Syncytial Virus
RN Rễ nhánh
RSC Rễ sơ cấp
SCV Settled Cell Volume
SEM Scanning Electron Microscope
SH Schenk và Hildebrandt
T_TCL transverse_Thin Cell layer

viii
TDZ Thidiazuron
TEM Transmission Electron Microscope
TIBA 2,3,5-triiodobenzoic acid
VDC Vein-Derived Cell
WPM Woody Plant Medium

ix
DANH MỤC CÁC BẢNG
Bảng 3.1. Ảnh hưởng của NAA và môi trường khoáng đến sự tạo phôi vô
tính trực tiếp từ mô lá ở 60 NSC ..................................................
55
Bảng 3.2. Ảnh hưởng của kết hợp NAA và BA đến sự tạo phôi vô tính trực
tiếp từ mảnh lá, ở môi trường SH, 60 NSC .................................
58
Bảng 3.3. Ảnh hưởng của nồng độ đường và điều kiện chiếu sáng đến sự
tạo phôi vô tính trực tiếp từ mô lá, ở môi trường SH, 60 NSC ....
61
Bảng 3.4. Ảnh hưởng của tỷ lệ nước dừa đến sự tạo phôi vô tính trực tiếp
từ mảnh lá (10 x 10 mm), ở môi trường SH, 60 NSC .................
63
Bảng 3.5. Ảnh hưởng của 2,4-D đến sự hình thành mô sẹo có khả năng
sinh phôi, ở môi trường SH, 30NSC.............................................
68
Bảng 3.6. Ảnh hưởng của NAA và BA đến sự tạo phôi từ mô sẹo mảnh lá
(10 x 10 mm), ở môi trường SH, 30 NSC ..................................
70
Bảng 3.7. Ảnh hưởng của NAA và BA đến tạo phôi từ mô sẹo mảnh lá (3
x 10 mm), ở môi trường SH, 30 NSC .........................................
77
Bảng 3.8. Ảnh hưởng của NAA và BA đến sự tạo phôi thứ cấp trên môi
trường đặc SH, 30 NSC ...............................................................
80
Bảng 3.9. Ảnh hưởng của NAA và BA đến sự tạo phôi thứ cấp trong môi
trường lỏng SH, 30 NSC .............................................................
82
Bảng 3.10. Ảnh hưởng của khối lượng phôi nuôi cấy đến tăng trưởng sinh
khối phôi, trong môi trường lỏng SH, 45 NSC ..........................
84
Bảng 3.11. Ảnh hưởng của kích thước phôi đến sự hình thành phôi thứ cấp,
trong môi trường lỏng SH, 30 NSC ............................................
87
Bảng 3.12. Ảnh hưởng của nồng độ đường đến sự tăng trưởng sinh khối
phôi, trong môi trường lỏng SH, 30 NSC ..................................
89
Bảng 3.13. Ảnh hưởng của cường độ ánh sáng đến sự tăng trưởng sinh khối
phôi trong môi trường lỏng SH, 30 NSC ...................................
90
Bảng 3.14. Ảnh hưởng của tỷ lệ nước dừa đến sự tạo phôi thứ cấp, trong
môi trường lỏng SH, 21 NSC ......................................................
91

x
Bảng 3.15. Ảnh hưởng của NAA và môi trường khoáng đến sự tạo rễ bất
định trực tiếp từ mảnh lá (10 x 10 mm), ở môi trường ½MS, 30
NSC ............................................................................................
98
Bảng 3.16. Ảnh hưởng của NAA và môi trường khoáng đến tạo rễ bất định
trực tiếp từ mảnh lá (3 x 10 mm), ở môi trường ½MS, 30 NSC
.....................................................................................................
101
Bảng 3.17. Ảnh hưởng của nồng độ đường đến sự tạo rễ bất định trực tiếp
từ mảnh lá (10 x 10 mm) ở môi trường ½MS, 30 NSC .............
104
Bảng 3.18. Ảnh hưởng của nồng độ đường đến sự tạo rễ bất định trực tiếp
từ mảnh lá (3 x 10 mm), ở môi trường ½MS, 30 NSC ..............
106
Bảng 3.19. Ảnh hưởng của điều kiện chiếu sáng đến tạo rễ bất định trực tiếp
từ mảnh lá (10 x 10 mm), ở môi trường ½MS, 30 NSC ...............
107
Bảng 3.20. Ảnh hưởng của điều kiện chiếu sáng đến sự tạo rễ bất định trực
tiếp từ mảnh lá (3 x 10 mm), ở môi trường ½MS, 30 NSC .........
108
Bảng 3.21. Ảnh hưởng của NAA, IBA đến sự tạo rễ bất định từ chồi có
nguồn gốc từ đốt thân cây vườn ươm, ở môi trường ½MS, 60
NSC ............................................................................................
111
Bảng 3.22. Ảnh hưởng của NAA và IBA đến sự tạo rễ bất định từ chồi đốt
thân cây in vitro, ở môi trường ½MS, 60 NSC ............................
113
Bảng 3.23. Ảnh hưởng của NAA và IBA đến sự tạo rễ bất định từ chồi có
nguồn gốc phôi vô tính, ở môi trường ½MS, 30 NSC ................
115
Bảng 3.24. Ảnh hưởng của NAA, IBA đến sự phân nhánh rễ, ở môi trường
½MS, 21 NSC .............................................................................
118
Bảng 3.25. Ảnh hưởng của nồng độ đường đến sự tăng trưởng sinh khối rễ,
ở môi trường ½MS, 30 NSC ........................................................
121
Bảng 3.26. Ảnh hưởng của khối lượng rễ nuôi cấy đến sự tăng trưởng sinh
khối rễ, ở môi trường ½MS, 45 NSC ..........................................
123

xi
DANH MỤC CÁC HÌNH, ĐỒ THỊ
Hình 1.1. Hình thái một số bộ phận của cây Ngũ gia bì chân chim
(Schefflera octophylla) .............................................................
5
Hình 2.1. Cây Ngũ gia bì chân chim (Schefflera octophylla) ................... 35
Hình 2.2. Lá kép Ngũ gia bì chân chim và vật liệu nuôi cấy ................... 35
Hình 2.3. Vật liệu đốt thân dùng nuôi cấy tạo chồi .................................. 48
Hình 3.1. Các dạng phát triển của phôi vô tính hình thành trực tiếp từ mô
lá ...............................................................................................
52
Hình 3.2. Mảnh lá tạo phôi ở môi trường ½MS, MS, B5, SH có NAA,
60 NSC .....................................................................................
56
Hình 3.3. Các giai đoạn phát triển của phôi vô tính và hình thái giải phẫu
phôi tương ứng .........................................................................
57
Hình 3.4. Mảnh lá tạo phôi trực tiếp ở môi trường SH có NAA kết hợp
với BA, 60 NSC .......................................................................
59
Hình 3.5. Phôi vô tính phát sinh trực tiếp từ mảnh lá ở môi trường SH,
có đường và điều kiện sáng, tối, 60 NSC .................................
62
Hình 3.6. Phôi vô tính phát sinh trực tiếp từ mảnh lá ở môi trường SH,
có bổ sung nước dừa, 60 NSC ..................................................
64
Hình 3.7 Phôi bình thường và một số dạng bất thường của phôi vô tính
...................................................................................................
64
Hình 3.8. Tạo cây con từ phôi vô tính trên môi trường MS, ½MS không
có chất ĐHST ...........................................................................
65
Hình 3.9. Trồng cây từ phôi vô tính ra chậu đất ở vườn ươm ................... 66
Hình 3.10. Ảnh hưởng của cách cấy mảnh lá đến hiệu quả tạo mô sẹo ...... 66
Hình 3.11. Tạo mô sẹo từ mảnh lá (10 x 10 mm)nuôi cấy trên môi trường
SH .............................................................................................
67
Hình 3.12. Ảnh hưởng của 2,4-D đến sự hình thành mô sẹo có khả năng
sinh phôi ở môi trường SH, 30 NSC .........................................
69
Hình 3.13. Tái sinh phôi từ mô sẹo có khả năng sinh phôi trên môi trường
đặc SH, 30 NSC và hình thái giải phẫu phôi tái sinh từ mô sẹo
...................................................................................................
71

xii
Hình 3.14. Các dạng phát triển của phôi tái sinh từ mô sẹo mảnh lá, cây
con hoàn chỉnh từ phôi .............................................................
73
Hình 3.15. Tạo phôi từ mô sẹo có KNSP bằng phương pháp nuôi lỏng lắc,
hình thái cụm tế bào phân chia, cụm mô tạo phôi .....................
74
Hình 3.16. So sánh kết quả tái sinh phôi ở môi trường SH đặc và lỏng, 30
NSC ..........................................................................................
75
Hình 3.17. Mô sẹo hình thành từ mảnh lá (3 x 10 mm) ở môi trường SH
có 2,4-D, 20 – 30 NSC ..............................................................
76
Hình 3.18. Tạo phôi gián tiếp từ mô sẹo mảnh lá (3 x 10 mm) trên môi
trường SH, 30 NSC ..................................................................
78
Hình 3.19. Phôi tái sinh từ mô sẹo mảnh lá (3 x 10 mm) ở môi trường SH,
60 NSC .....................................................................................
79
Hình 3.20. Ảnh hưởng của NAA và BA đến sự nhân phôi trên môi trường
đặc SH, 30 NSC .......................................................................
81
Hình 3.21. Ảnh hưởng của NAA và BA đến sự tạo phôi thứ cấp nuôi lỏng
trong môi trường SH, 30 NSC .................................................
83
Hình 3.22. Ảnh hưởng của khối lượng phôi (w/v) nuôi cấy đến tăng
trưởng sinh khối phôi, trong môi trường lỏng SH, 45 NSC .......
84
Hình 3.23. Nhân phôi qua nuôi cấy lỏng lắc trong môi trường SH, 30 – 60
NSC ..........................................................................................
85
Hình 3.24. Sự đa dạng của kích thước phôi từ quá trình nuôi lỏng lắc trong
môi trường SH ..........................................................................
86
Hình 3.25. Ảnh hưởng của kích thước phôi đến sự tạo phôi thứ cấp trong
môi trường lỏng SH, 30 NSC ....................................................
88
Hình 3.26. Ảnh hưởng của nồng độ đường đến sự tăng sinh khối phôi,
trong môi trường lỏng SH, 30 NSC ...........................................
89
Hình 3.27. Ảnh hưởng của điều kiện chiếu sáng đến sự tăng trưởng sinh
khối phôi, trong môi trường lỏng SH, 30 NSC .........................
91
Hình 3.28. Ảnh hưởng của nước dừa đến sự hình thành phôi thứ cấp, trong
môi trường lỏng SH, 21 NSC ...................................................
92
Hình 3.29. Tạo cây con từ phôi vô tính nuôi lỏng lắc trong môi trường
MS, ½MS .................................................................................
93

xiii
Hình 3.30. Hình thái giải phẫu của phôi sơ cấp và thứ cấp ....................... 94
Hình 3.31. Đĩa cấy mảnh lá trên môi trường ½MS có 3 mg/L NAA, 30
NSC ..........................................................................................
96
Hình 3.32. Tạo rễ trực tiếp từ mảnh lá (10 x 10 mm) ở môi trường MS, có
NAA, 30 NSC .....................................................................
99
Hình 3.33. Tạo rễ trực tiếp từ mảnh lá (10 x 10 mm) ở môi trường ½MS,
có NAA, 30 NSC .....................................................................
99
Hình 3.34. Tạo rễ trực tiếp từ mảnh lá (10 x 10 mm) ở môi trường B5, có
NAA, 30 NSC ...........................................................................
99
Hình 3.35. Tạo rễ trực tiếp từ mảnh lá (10 x 10 mm) ở môi trường SH, có
NAA, 30 NSC ...........................................................................
99
Hình 3.36. Tạo rễ trực tiếp từ mảnh lá (3 x 10 mm) ở môi trường MS, có
NAA, 30 NSC ...........................................................................
102
Hình 3.37. Tạo rễ trực tiếp từ mảnh lá (3 x 10 mm) ở môi trường ½MS,
có NAA, 30 NSC ......................................................................
102
Hình 3.38. Tạo rễ trực tiếp từ mảnh lá (3 x 10 mm) ở môi trường B5, có
NAA, 30 NSC ...........................................................................
102
Hình 3.39. Tạo rễ trực tiếp từ mảnh lá (3 x 10 mm) ở môi trường SH, có
NAA, 30 NSC ...........................................................................
102
Hình 3.40. Ảnh hưởng của nồng độ đường đến sự tạo rễ trực tiếp từ mảnh
lá (10 x 10 mm), ở môi trường ½MS, 30 NSC .........................
105
Hình 3.41. Ảnh hưởng của nồng độ đường đến sự tạo rễ từ mảnh lá (3 x
10 mm), ở môi trường ½MS, 30 NSC .......................................
106
Hình 3.42. Ảnh hưởng của điều kiện chiếu sáng đến sự tạo rễ bất định từ
mảnh lá (10 x 10 mm), ở môi trường ½MS, 30 NSC .................
107
Hình 3.43. Ảnh hưởng của điều kiện chiếu sáng đến sự tạo rễ bất định từ
mảnh lá (3 x 10 mm), ở môi trường ½MS, 30 NSC .................
108
Hình 3.44. Minh họa cận cảnh sự tái sinh trực tiếp rễ bất định từ mảnh lá
nuôi cấy trên môi trường ½MS, 12 – 20 NSC ...........................
109
Hình 3.45. Hình thái giải phẫu sơ khởi rễ bất định hình thành trực tiếp từ
mảnh lá .....................................................................................
110

xiv
Hình 3.46. Tạo rễ bất định từ chồi có nguồn gốc từ đốt thân, ở môi trường
½MS, 60 NSC ..........................................................................
112
Hình 3.47. Tạo rễ bất định từ chồi đốt thân cây in vitro, ở môi trường
½MS, 60 NSC ...........................................................................
114
Hình 3.48. Tạo rễ bất định từ chồi có nguồn gốc phôi vô tính, ở môi trường
½MS, 30 NSC ...........................................................................
116
Hình 3.49. Vật liệu rễ bất định sơ cấp ở 20 NSC ....................................... 117
Hình 3.50. Ảnh hưởng của NAA, IBA đến hình thành rễ nhánh từ rễ đơn,
ở môi trường ½MS ....................................................................
119
Hình 3.51. Rễ tăng trưởng sinh khối liên tục qua quá trình phân nhánh của
rễ đơn ........................................................................................
120
Hình 3.52. Ảnh hưởng của nồng độ đường đến sự tăng trưởng sinh khối
rễ, ở môi trường ½MS, 30 NSC ................................................
122
Hình 3.53. Ảnh hưởng của khối lượng rễ nuôi cấy đến tăng trưởng sinh
khối rễ, ở môi trường ½MS, 45 NSC ........................................
124
Hình 3.54. Diễn biến tăng trưởng sinh khối rễ bất định theo thời gian, 7 –
49 NSC .....................................................................................
126
Biểu đồ 3.1. Tăng trưởng sinh khối rễ bất định theo thời gian ...................... 125

xv
TÓM TẮT
Đề tài: “Nghiên cứu tạo, nhân phôi vô tính và rễ bất định cây Ngũ gia bì
chân chim (Schefflera octophylla Lour. Harms)” được thực hiện tại Viện Sinh học
nhiệt đới –Viện Hàn lâm Khoa học và Công nghệ Việt Nam từ tháng 9 năm 2015 đến
tháng 9 năm 2019.
Mục tiêu tổng quát của đề tài là tạo được phôi vô tính tái sinh trực tiếp, gián
tiếp và nhân được phôi vô tính trên môi trường đặc và trong môi trường lỏng lắc. Tạo
được rễ bất định tái sinh trực tiếp và nhân được rễ bất định trong môi trường lỏng lắc.
Mục tiêu cụ thể của đề tài là: (1) Xác định được một số yếu tố ảnh hưởng (môi
trường khoáng, chất điều hòa sinh trưởng, đường,…) đến tái sinh trực tiếp hoặc/và
gián tiếp phôi vô tính và rễ bất định từ mô lá nuôi cấy in vitro; (2) Xác định được một
số yếu tố ảnh hưởng (môi trường khoáng, chất điều hòa sinh trưởng, số/khối lượng
phôi/rễ, đường,…) đến nhân phôi vô tính và rễ bất định có nguồn gốc từ mô lá nuôi
cấy in vitro; (3) Tạo được cây con in vitro hoàn chỉnh từ phôi vô tính; (4) Tạo được
rễ bất định từ chồi có nguồn gốc đốt thân/phôi vô tính; (5) Xác định được hình thái
giải phẫu của phôi vô tính ở một số giai đoạn phát triển khác nhau, nguồn gốc tế bào
phát sinh phôi vô tính và rễ bất định thông qua kỹ thuật nhuộm hai màu.
Đề tài gồm có các nội dung cơ bản như: (i) Tạo phôi vô tính trực tiếp và gián
tiếp từ mô lá; (ii) Nhân phôi vô tính trên môi trường đặc và trong môi trường lỏng
lắc; (iii) Tạo cây con hoàn chỉnh từ phôi vô tính; (iv) Tạo rễ bất định trực tiếp từ mô
lá, từ chồi có nguồn gốc đốt thân/phôi vô tính; (v) Nhân rễ bất định trong môi trường
lỏng lắc; (vi) Khảo sát hình thái giải phẫu phôi vô tính và rễ bất định.
Qua nghiên cứu, kết quả cho thấy môi trường thích hợp cho sự hình thành phôi
vô tính trực tiếp từ nuôi cấy in vitro mô lá cây Ngũ gia bì chân chim là SH có bổ sung
5 mg/L NAA, 0,25 mg/L BA; 50 g/L sucrose, 10% (v/v) nước dừa kết hợp với nuôi
mô ở điều kiện chiếu sáng 4.000 lux. Môi trường thích hợp cho tạo mô sẹo từ mô lá
là môi trường SH có 2 mg/L 2,4-D. Phôi vô tính tái sinh (gián tiếp) từ mô sẹo trên
môi trường SH có 2 mg/L NAA, 0,25 mg/L BA, 30 g/L sucrose. Đã nhân in vitro
thành công phôi vô tính trên môi trường đặc và trong môi trường lỏng thông qua sự
hình thành phôi thứ cấp theo cơ chế bất định. Nhân phôi trong môi trường lỏng hiệu
quả hơn so với môi trường đặc. Môi trường lỏng thích hợp dùng nhân nhanh phôi vô

xvi
tính là môi trường SH có bổ sung 2 mg/L NAA, 0,25 mg/L BA, 50 g/L sucrose, 10%
(v/v) nước dừa; khối lượng phôi nuôi cấy ban đầu thích hợp là 2% (w/v), kích thước
phôi thích hợp dùng nuôi cấy ~ 10 mm, điều kiện chiếu sáng ~ 4.000 lux. Phôi vô
tính phát triển thành cây con có khả năng sinh trưởng bình thường trong điều kiện in
vitro và ex vitro.
Môi trường thích hợp cho tạo rễ bất định trực tiếp từ mô lá là môi trường đặc
½MS có 3 mg/L NAA, 30 g/L sucrose; cường độ ánh sáng ~ 4.000 lux. Đã nhân thành
công sinh khối rễ bất định thông qua sự hình thành rễ thứ cấp bằng phương pháp nuôi
lỏng lắc dùng môi trường ½MS có bổ sung 2 mg/L NAA, 30 g/L sucrose. Khối lượng
rễ nuôi cấy ban đầu thích hợp là 2% (w/v), sinh khối rễ phát triển rất nhanh ở giai
đoạn 21–35 ngày, đạt mức cao nhất ở giai đoạn 42 ngày sau cấy.
Cũng đã tạo được rễ bất định cho chồi có nguồn gốc đốt thân (cây vườn ươm,
cây in vitro) và phôi vô tính (đã cắt bỏ rễ trụ), ghi nhận được hình thái giải phẫu phôi
vô tính và rễ bất định bằng phương pháp nhuộm hai màu.

xvii
SUMMARY
The study “In vitro induction and multiplication of somatic embryos and
adventitious roots of Schefflera octophylla Lour. Harms” was carried out at the
Institute of Tropical Biology (ITB) – Vietnam Academy of Science and Technology
(VAST) from September of 2015 to September of 2019.
The general objective of this study was to generate directly/indirectly somatic
embryos and multiply somatic embryos on solid and shaking liquid media, to generate
directly adventitious roots and mutiply adventitious roots in shaking liquid medium.
The specific objectives of the study were to identify: (1) The optimal culture
conditions for inducing direct and indirect formation of somatic embryos and
adventitious roots from leaf explants cultured in vitro; (2) The optimal culture
conditions for multiplication of somatic embryos and adventitious roots originated
from leaf explants on solid and/or shaking liquid media; (3) The condition for
generation of in vitro plantlets from somatic embryos; (4) The condition for induction
of adventitious roots from shoots originated from nodal explants/somatic embryos (5)
The morphological and histological properties of somatic embryos at many different
stages of development, cellular origin of somatic embryo and adventitious root
formation through double staining with carmine alum and iodine green.
The study comprised of four contents: (i) To generate directly/indirectly
somatic embryos from leaf explants; (ii) To multiply somatic embryos on the solid
and liquid media; (iii) To generate the plantlets originated from somatic embryos; (iv)
To generate directly adventitious roots from leaf explants, from shoots originated
from nodal explants/somatic embryos; (v) To multiply adventitious roots in liquid
medium; (vi) To examine the morphology and histology of somatic embryo and
adventitious root.
The results showed that the suitable medium for the direct induction of somatic
embryos from the leaf explants cultured in vitro was SH + 5 mg/L NAA + 0.25 mg/L
BA + 10% (v/v) coconut water, 50 g/L sucrose, in light condition ~ 4,000 lux. The
suitable media for callus induction from the leaf explants, for regeneration of somatic
embryos from callus, were SH + 2 mg/L 2,4-D; SH + 2 mg/L NAA + 0.25 mg/L BA,
30 g/L sucrose, respectively. Successfully in vitro multiplied somatic embryos on

xviii
solid and shaking liquid media through secondary embryogenesis by an adventitious
mechanism. Somatic embryo multiplication in liquid medium was more efficient than
on solid media. The suitable liquid medium for multiplication of somatic embryos
was SH + 2 mg/L + 0.25 mg/L BA + 10% (v/v) coconut water, 50 g/L sucrose; the
suitable somatic embryo inoculum density/size for culture was 2% (w/v), the suitable
size of somatic embryos for multiplication in liquid medium was ~ 10 mm, light
condition was 4,000 lux. The somatic embryos developed into plantlets with normal
growth under in vitro/ex vitro conditions.
The suitable medium for direct adventitious roots from leaf explants was ½MS
+ 3 mg/L NAA, 30 g/L sucrose, light condition was ~ 4,000 lux. Through secondary
rhizogenesis by an adventitious mechanism, successfully in vitro multiplied
adventitious roots in liquid medium ½MS + 2 mg/L NAA, 30 g/L sucrose; suitable
root inoculum density for culture was 2% (w/v), root biomass developed quickly at
the period of 21-35 days, reaching the highest level at 42 days after culturing.
The adventitious roots from shoots originated from nodal explants
(greenhouse and in vitro plants) and somatic embryos (with the tap roots removed),
the morphology and anatomy of somatic embryos and adventitious roots via double
staining method were also achieved.

1
MỞ ĐẦU
1. Tính cấp thiết của đề tài
Ngũ gia bì chân chim (Schefflera octophylla Lour. Harms), là loài cây thuộc
họ Ngũ gia bì/Nhân sâm (Araliaceae) – họ các loài thực vật có nguồn hoạt chất sinh
học quan trọng, trong đó có Ngũ gia bì chân chim (NGBCC) [1]. Trong Đông y và y
học hiện đại, có khá nhiều công trình công bố kết quả nghiên cứu về các hợp chất thứ
cấp ở NGBCC với nhiều tác dụng như tăng cường sức khỏe [2], kháng virus [3][4][5]
kháng một số dòng tế bào ung thư [6], làm giảm đau [7] kháng viêm [8], đông máu
[9],... Ở Việt Nam, các vật liệu như lá, vỏ của thân/rễ và rễ nhỏ Ngũ gia bì chân chim
đều được dùng để làm thuốc [10].
Đến nay, một số công trình có liên quan đến NGBCC đã được ghi nhận như
nhân giống bằng phương pháp giâm cành, và bằng phương pháp nuôi cấy mô [3].
Đặng Thị Thanh Tâm (2012) đã nghiên cứu tạo rễ tơ thông qua biến nạp gen dùng vi
khuẩn Agrobacterium rhizogenes và thử nghiệm sản xuất sinh khối loại rễ này bằng
hệ thống bioreactor [11].
Trong những năm gần đây, nghiên cứu ở lĩnh vực sinh học nói chung, sinh lý
học thực vật nói riêng đã đạt được nhiều thành tựu đáng kể, trong đó phương pháp
nuôi cấy mô tế bào luôn được các nhà nghiên cứu quan tâm sử dụng như một công
cụ nhằm tìm hiểu sâu các khía cạnh sinh lý của tính toàn thế của tế bào như vai trò
sinh lý của chất điều hòa sinh trưởng (ĐHST) ngoại sinh, nội sinh và tương tác của
chúng, vai trò của các hệ thống di chuyển và phân bố auxin trong mối quan hệ với hệ
thống gene điều hòa, tuổi sinh lý của mẫu nuôi cấy,.. đối với sự hình thành, phát triển
hai con đường phát sinh hình thái phôi vô tính/rễ bất định thông qua hai kiểu tái sinh
trực tiếp/gián tiếp của hai dạng hình thái phát sinh nói trên [12][13][14][15].
Theo nhiều tài liệu, hệ thống phôi vô tính không chỉ là mục tiêu đối với nghiên
cứu về phát sinh hình thái mà còn là vật liệu được sử dụng trong nhiều nghiên cứu
nhằm các mục đích khác nhau như nhân giống [16], thu hợp chất thứ cấp [17], bảo
quản nguồn gen in vitro [18], tạo hạt nhân tạo [19], biến nạp gen [20], nuôi cấy quang
tự dưỡng [21], chuyển giao công nghệ và thương mại hóa sản phẩm phôi vô tính [22].
Tương tự phôi vô tính, rễ bất định cũng là mục tiêu trong các nghiên cứu về phát sinh
hình thái; ngoài ra, nuôi cấy rễ bất định cũng là một trong các hướng nghiên cứu rất

2
quan trọng nhằm thu nhận các hợp chất thứ cấp. Hệ thống nuôi cấy này khắc phục
được các nhược điểm của hệ thống nuôi cấy tế bào trong một số trường hợp, do các
ưu điểm sau: một là, rễ là một tập hợp có hệ thống các tế bào biệt hóa và như vậy đã
mang một nhiệm vụ nhất định đối với hoạt động sống của cây, do vậy cũng liên kết
với năng lực sinh tổng hợp các hợp chất thứ cấp; hai là, đối với một số cây dược liệu,
rễ là nơi sinh tổng hợp/tích tụ các hợp chất thứ cấp; ba là, có thể nhân nhanh sinh
khối rễ trong môi trường lỏng mà không bị hạn chế vì hiện tượng hóa kính/hóa trong
(vitrification) như trường hợp nuôi thể chồi; bốn là, tương tự như ở một số hệ thống
nuôi cấy khác, có thể nuôi nhân quy mô lớn rễ trong điều kiện khống chế, không phụ
thuộc vào điều kiện môi trường bên ngoài.
Đến nay đã ghi nhận được một số công bố về kết quả nghiên cứu nhân sinh
khối rễ, phôi vô tính phục vụ chiết xuất, thu nhận hợp chất thứ cấp với nhiều quy mô
khác nhau, ví dụ như ở trường hợp Panax ginseng, Panax vietnamensis, Panax
quinquefolius [ 23][24][17]. Tuy nhiên, cho đến nay chưa ghi nhận được công bố nào
trên thế giới và trong nước nghiên cứu về nuôi cấy phôi vô tính và rễ bất định cây
NGBCC.
Dựa vào các cơ sở khoa học và thực tiễn nêu trên và góp phần phát triển nghiên
cứu trên đối tượng NGBCC, luận án được thực hiện với tên đề tài: “Nghiên cứu tạo,
nhân phôi vô tính và rễ bất định cây Ngũ gia bì chân chim (Schefflera octophylla
Lour. Harms)” phục vụ định hướng dài hạn sử dụng nguồn phôi vô tính, rễ bất định
cho các nghiên cứu về nhân giống, thu nhận hợp chất thứ cấp và cho một số nghiên
cứu quan trọng khác trong tương lai.
2. Mục tiêu của đề tài
2.1. Mục tiêu tổng quát
Tạo được phôi vô tính tái sinh trực tiếp, gián tiếp và nhân được phôi vô tính
trên môi trường đặc và trong môi trường lỏng lắc. Tạo được rễ bất định tái sinh trực
tiếp và nhân được rễ bất định trong môi trường lỏng lắc.
2.2. Mục tiêu cụ thể
Xác định được điều kiện môi trường thích hợp cho sự cảm ứng hình thành phôi
vô tính trực tiếp và gián tiếp từ mô lá (môi trường khoáng, loại và nồng độ auxin và
cytokinin, nồng độ đường, tỷ lệ nước dừa, điều kiện chiếu sáng,…).

3
Xác định được điều kiện thích hợp (chất ĐHST, khối lượng/kích thước phôi,
nồng độ đường, tỷ lệ nước dừa) cho sự nhân sinh khối và tăng sinh phôi.
Xác định được điều kiện thích hợp cho sự cảm ứng hình thành và tăng sinh rễ
bất định từ mô lá (môi trường khoáng, loại và nồng độ auxin, nồng độ đường, điều
kiện chiếu sáng,…).
Tạo được rễ bất định cho chồi có nguồn gốc đốt thân/phôi vô tính.
Xác định được hình thái giải phẫu của phôi vô tính ở một số giai đoạn phát
triển khác nhau, nguồn gốc tế bào phát sinh phôi vô tính và rễ bất định thông qua kỹ
thuật nhuộm hai màu.
3. Ý nghĩa khoa học và thực tiễn của đề tài
3.1. Ý nghĩa khoa học
Cung cấp những dẫn liệu khoa học mới có giá trị nghiên cứu về hai con đường
phát sinh hình thái phôi vô tính và rễ bất định, hai kiểu tái sinh trực tiếp/gián tiếp phôi
vô tính và rễ bất định; về nhân phôi vô tính và rễ bất định thông qua cơ chế tạo phôi
và rễ bất định thứ cấp ở NGBCC - một trong các đối tượng thực vật/cây dược liệu
quan trọng thuộc họ Ngũ gia bì. Đồng thời, luận án cũng là tài liệu tham khảo hữu
ích phục vụ cho nghiên cứu và giảng dạy về lĩnh vực sinh lý thực vật, nuôi cấy mô,
tế bào thực vật.
3.2. Ý nghĩa thực tiễn
Kết quả từ nghiên cứu này là tiền đề quan trọng cho nghiên cứu nhân sinh khối
phôi vô tính, rễ bất định trên quy lớn và sản phẩm sinh khối tạo được có thể được sử
dụng trong nghiên cứu nhân giống (phôi vô tính), thu hợp chất thứ cấp (rễ bất định)
phục vụ lĩnh vực y dược và mỹ phẩm.
4. Đối tượng và phạm vi nghiên cứu
4.1. Đối tượng nghiên cứu
Phôi vô tính từ nuôi cấy mảnh lá; rễ bất định từ nuôi cấy mảnh lá/chồi có nguồn
gốc đốt thân/phôi vô tính.
4.2. Phạm vi nghiên cứu
Đề tài thực hiện trên cơ sở ứng dụng phương pháp nuôi cấy mô nhằm tạo và
nhân phôi vô tính, tạo và nhân rễ bất định dưới ảnh hưởng của một số yếu tố như môi
trường khoáng, chất điều hòa sinh trưởng, đường, ánh sáng,…, tạo cây con in vitro

4
hoàn chỉnh từ phôi vô tính và ứng dụng phương pháp khảo sát hình thái/hình thái giải
phẫu mô tế bào nhằm xác định nguồn gốc phát sinh phôi vô tính và rễ bất định.
Do giới hạn về thời gian nên chưa đề cập đến nghiên cứu nhân rễ bất định từ
chồi có nguồn gốc đốt thân/phôi vô tính bằng phương pháp nuôi lỏng lắc, nhân phôi
vô tính/rễ bất định bằng hệ thống bioreactor cũng như không thu thập số liệu các chỉ
tiêu sinh trưởng của cây từ phôi vô tính ở giai đoạn ex vitro.
5. Tính mới của đề tài
Nghiên cứu này đã xác định được môi trường nuôi cấy và điều kiện nuôi cấy
thích hợp cho sự phát sinh phôi vô tính trực tiếp từ mảnh lá NGBCC, đồng thời cũng
xây dựng quy trình tạo phôi gián tiếp thông qua mô sẹo mảnh lá. Nhân in vitro phôi
vô tính trên môi trường đặc và trong môi trường lỏng thông qua sự hình thành phôi
thứ cấp theo cơ chế bất định. Phôi vô tính phát triển thành cây con có khả năng sinh
trưởng bình thường trong điều kiện in vitro và ex vitro. Kết quả này có ý nghĩa rất
lớn trong nhân giống loài dược liệu này.
Xác định được môi trường thích hợp cho tạo rễ bất định trực tiếp từ mảnh lá
nuôi cấy in vitro. Nhân sinh khối rễ bất định thông qua sự hình thành rễ thứ cấp bằng
phương pháp nuôi lỏng lắc. Kết quả này có thể ứng dụng cho hướng nghiên cứu sản
xuất thu các hợp chất thứ cấp.

5
CHƯƠNG 1. TỔNG QUAN TÀI LIỆU
1.1. Giới thiệu về Ngũ gia bì chân chim (Schefflera octophylla Lour. Harms)
1.1.1. Nguồn gốc và phân bố
Giới : Plantae
Ngành: Magnoliopsida
Lớp : Magnoliopsida
Bộ : Apiales
Họ : Araliaceae
Chi : Schefflera
Loài : Schefflera octophylla
`
Họ Ngũ gia bì/họ Nhân sâm (Araliaceae) là họ rất đa dạng về thành phần loài,
có khoảng 70 chi, 900 loài, phân bố chủ yếu ở vùng nhiệt đới và á nhiệt đới, rất ít đại
diện ở vùng ôn đới, phần lớn các chi và loài phân bố ở Đông nam Á, châu Úc, châu
Mỹ nhiệt đới. Ở Việt Nam thực vật thuộc họ Ngũ gia bì cũng khá đa dạng về thành
phần loài, đặc điểm sinh thái – sinh học, hiện thống kê được 141 loài thuộc 19 chi,
phân bố rộng thường mọc rải rác ở vùng rừng núi nhưng mọc tập trung nhiều ở những
vùng núi cao có khí hậu ôn hòa, có nhiều loài sống ở những nơi quang đãng, ẩm ướt,
ven rừng, ven suối…Thực vật họ Ngũ gia bì còn rất đa dạng và phong phú về các hợp
chất thứ cấp, có khả năng tổng hợp và tích lũy các hợp chất triterpenoid saponin,
steroidal saponin, ginsenosides, polysaccharid, các flavonoid, tinh dầu, các hợp chất
Hình 1.1. Hình thái một số bộ phận của cây NGBCC (Schefflera octophylla)
A. Lá; B. Hoa; C. Quả
(Nguồn: A: Wikipedia – Việt Nam; B,C: Wang và cộng sự, 2021[25])

6
hữu cơ khác…[1]. Đây chính là nguồn hợp chất sinh học có hoạt tính rất tốt, vì thế
rất nhiều loài trong họ này đã được sử dụng làm nguồn dược liệu quý có giá trị cao
trong ứng dụng bảo vệ sức khỏe cho con người.
Chi Schefflera là chi lớn nhất trong họ Ngũ gia bì, thành phần loài rất đa dạng
với khoảng 450 loài, phân bố chủ yếu ở vùng nhiệt đới, cận nhiệt đới châu Á như
Nhật Bản, Trung Quốc, Việt Nam, Lào, Campuchia, Thái Lan, Malaysia, Ấn Độ, Sri
Lanka, Philippines, Indonesia, New Zealand, Singapore, Australia và một số đảo ở
Thái Bình Dương [10]. Ở Việt Nam cũng đã thống kê được 56 loài (chiếm 39,7%
tổng số loài của họ Ngũ gia bì), 4 thứ [1]. Nhiều loài trong chi này đã được sử dụng
làm thuốc chữa trị bệnh phong thấp, đau nhức xương khớp ngoài ra còn có tác dụng
điều trị bệnh tiêu hóa, ho, cầm máu…như NGBCC (Schefflera octophylla Lour.
Harms), Chân chim Bắc bộ (Schefflera tonkinensis R. Vig), Chân chim lá nhỏ
(Schefflera pes-avis R. Vig), Ngũ gia bì lá láng (Schefflera nitidifolia Harms), Chân
chim Sapa (Schefflera vietnamensis Grush, N. Skvorts/Schefflera chapana Hamrs), ở
loài Chân chim núi – Chân chim Petelot (Schefflera petelotii Merr.) còn được sử dụng
chữa gãy xương [10].
NGBCC có danh pháp khoa học là Schefflera octophylla (Lour.) Harms [26],
tên khoa học khác Schefflera heptaphylla (L.) Frodin, Aralia octophylla Lour, còn có
tên gọi khác là Sâm nam, cây Đáng, Lá lằng, Kotan (Lào). Đây là loài thực vật thường
xanh, có hoa, có nguồn gốc từ Đông Á là Nhật Bản và miền nam Trung Quốc. Ở Nhật
Bản NGBCC mọc hoang từ miền nam Kyushu đến Okinawa, nó là loài thực vật đại
diện cho vùng Yanbaru của Okinawa Nhật Bản. Ở Trung Quốc NGBCC phân bố chủ
yếu ở Quảng Đông, Quảng Tây, Vân Nam, Phúc Kiến [8]. Ở Việt Nam, NGBCC mọc
rải rác ở vùng núi cao phía bắc, phía nam của dãy Trường Sơn, có nơi mọc tập trung
có diện tích lớn khoảng 5 - 10 hecta, gặp ở các tỉnh như Lạng Sơn, Cao Bằng, Lào
Cai, Vĩnh Phúc, Phú Thọ, Bắc Cạn, Thái Nguyên, Hòa Bình, Hà Tây, Tuyên Quang,
Ninh Bình, Quảng Ngãi, Đắc Lắc, Bình Phước, Lâm Đồng [27][10].
1.1.2. Đặc điểm sinh học – sinh thái
NGBCC thuộc loài cây nhỏ hoặc cây to, cao khoảng 2 - 8 m. Lá chét hình chân
vịt, mọc so le có 6 - 8 lá chét, cuống lá dài 8 - 30 cm, lá chét nguyên, hình trứng, đầu
nhọn hay hơi tù, dài 7 - 17 cm, rộng 3 - 6 cm, cuống lá chét ngắn 1,5 - 2,5 cm, cuống
lá chét giữa dài hơn có thể tới 5 cm, mặt trên lá xanh sẫm bóng, mặt dưới lá nhạt hơn.

7
Cụm hoa mọc thành chùm tán, hoa nhỏ màu trắng thơm, mẫu 5, bao phấn có 2 ngăn,
bầu dưới, trên cuống phụ của cụm hoa đôi khi có hoa riêng lẻ. Quả mọng hình cầu,
đường kính ~ 5 mm, khi chín có màu tím sẫm đen, cây thường ra hoa vào tháng 2 -
3, ra quả vào tháng 4 - 5 [27][10].
NGBCC thuộc cây gỗ trung sinh, ưa ẩm, ưa sáng nhưng ở giai đoạn còn nhỏ
cây chịu bóng, chịu hạn, mọc xen với một số loài cây gỗ, cây bụi ven suối, ven rừng,
ẩm ướt…ngoài ra còn gặp ở rừng non phục hồi hoặc vùng nương rẫy. NGBCC sống
được ở nhiều loại đất khác nhau, thích hợp nhất ở đất feralit vàng đỏ. Đây là loài cây
gỗ phát triển nhanh, nhất là giai đoạn từ 5 - 15 năm tuổi, cây có thể sống đến 45 năm
tuổi, chiều cao khoảng 20 m, đường kính thân khoảng 43 cm, NGBCC có khả năng
tái sinh bằng hạt và bằng chồi rất tốt, những cây tái sinh từ hạt có bộ rễ phát triển tốt
hơn cây tái sinh từ chồi [10].
1.1.3. Thành phần hóa học và công dụng
1.1.3.1. Thành phần hóa học
Đã có nhiều nghiên cứu về thành phần hóa học của NGBCC thông qua chiết
xuất, phân tách và phân lập các chất từ các bộ phận lá, thân, rễ. Kết quả thu được
nhiều hợp chất tự nhiên có hoạt tính tốt được sử dụng trong y học. Chen và cộng sự
(2015) cho rằng thành phần hóa học chính là triterpenoid, đã có hơn 40 loại
triterpenoid được chiết xuất từ các bộ phận của loài thực vật này [7], bao gồm
oleanane [29][30], ursane [31], lupane [2][29][30].
Lá
Phân lập hoạt chất từ lá NGBCC thu được 2 triterpenoid glycoside mới là 3-
epi-betulinic acid 3-O-β-D-glucopyranoside (1); 3α-hydroxylup-20(29)-ene-23,28-
dioic acid 28-O-[α-L-rhamnopyranosyl (1→4)-O-β-D-glucopyranosyl (1→6)]-β-D-
glucopyranoside (2); khác với lup-20(29)-ene-23,28-dioic acid [2].
Nghiên cứu của Sung và cộng sự (1991) phân lập hoạt chất từ lá khô, thu nhận
một glucoside triterpenoid sulfate mới, từ dữ liệu quang phổ và biến đổi hóa học đã
xác định là 3-epi-betulinic axid 3-O-sulfate 28-O-[α-L- rhamnopyranosyl (1→4)-O-
β-D-glucopyranosyl (1→6)]-β-D-glucopyranoside [28]. Tiếp tục phân lập từ lá, Sung
và cộng sự thu nhận được một saponin triterpenoid 3,28-bidesmosidic mới, một
trisaccharid mới và oleanonic acid. Dựa trên dữ liệu quang phổ và sự biến đổi hóa
học, các thành phần mới được xác định là 3-epi-betulinic acid 3-O-β-D-

8
glucopyranoside 28-O-[α-L-rhamnopyranosyl (1→4)-O-β-D-glucopyranosyl
(1→6)]-β-D-glucopyranoside; α-L-rhamnopyranosyl (1→4)-O-D-glucopyranosyl
(1→6)-β-D-glucopyranoside [29]. Phân lập lá, dùng dữ liệu quang phổ và biến đổi
hóa học, Sung và cộng sự thu nhận được 28-O-[α-L-rhamnopyranosyl (1→4)-O-β-
D-glucopyranosyl (1→6)]-β-D-glucopyranoside của 3α-hydroxylup-20(29)-ene-
23,28-dioic acid (1); 3α,11α-dihydroxylup-20(29)-ene-23,28-dioic acid (2); 3-epi-
betulinic acid (3), trong đó hợp chất (2) và (3) được tìm thấy lần đầu tiên trong giới
thực vật [29].
Li và cộng sự (2005) đã phân lập từ cuống lá NGBCC thu được ba dẫn xuất
caffeoylquinic acid là 3,4-di-O-caffeoylquinic axid (1); 3,5-di-O-caffeoylquinic axid
(2); 3-O-caffeoylquinic axid (3). Những hợp chất này có hoạt tính kháng virus - chống
lại virus thể hợp bào gây bệnh hô hấp Respiratory Syncytial Virus (RSV), trong đó
3,4-di-O-caffeoylquinic axid, 3,5-di-O-caffeoylquinic acid có hoạt tính chống RSV
rất mạnh mẽ. Các hợp chất này phát huy tác dụng chống RSV, thông qua việc ức chế
sự dung hợp tế bào-virus trong giai đoạn đầu và ức chế sự dung hợp tế bào-virus ở
giai đoạn cuối [4]. Li và cộng sự (2007) chiết xuất từ lá, tiến hành phân đoạn và phân
lập đã thu được hai triterpenoid tinh khiết có hoạt tính cao, kháng virus hợp bào hô
hấp (RSV) rất mạnh, là 3α-hydroxylup-20(29)-ene-23,28-dioic acid (1); 3-epi-
betulinic acid 3-O-sulfate (2), và một saponin không hoạt động là 3α-hydroxylup-
20(29)-ene-23,28-dioic acid 28-O-[α-L-rhamnopyranosyl (1→4)-O-β-D-
glucopyranosyl-(1→6)]-β-D-glucopyranoside [5]. Tiếp tục phân tích thành phần hóa
học tinh dầu được chiết xuất từ lá, Li và cộng sự (2009) đã thu được 27 hợp chất dễ
bay hơi, trong đó có 17 hợp chất thuộc về monoterpene hoặc sesquiterpene. Tinh dầu
này có hoạt tính kháng sinh với ba dòng tế bào ung thư: tế bào MCF-7, A375 và Hep
G2 [6].
Từ nguyên liệu lá tươi của NGBCC, Liu và cộng sự (2019) đã phân lập và xác
định được 03 lupanine triterpenes có tác dụng đông máu hiệu quả, đó là 3α-hydroxy-
lup-20(29)-ene-23,28-dioic acid (1); betulinic acid 3-0-sulfate (2); 3α-hydroxylup-
20(29)-ene-23,28-dioic acid 28-O-[α-L-rhamnopyranosyl-(1→4)-O-β-D-
glucopyranosyl-(1→6)]-β-D-glucopyranoside (3). Trong đó chất betulinic acid 3-0-
sulfate thúc đẩy sự đông máu rất đáng kể, tác dụng cầm máu hiệu quả cao [9].

9
Thân
Một triterpenoid mới và glycoside được phân lập từ vỏ thân của NGBCC là
asiatic acid; và các asiaticoside được xác định là 3α-hydroxy-urs-12-ene-23,28-dioic
acid; 3α-hydroxy-urs-12-ene-23,28-dioic acid 28-O-[α-L-rhamnopyranosyl (1→4)-
O-β-D-glucopyranosyl (1→6)]-β-D-glucopyranoside. Lần đầu tiên asiaticoside được
phân lập từ một loài thực vật không phải của Centella Asiatica [31].
Wu Chun và cộng sự (2013) đã phân lập từ vỏ thân, trên cơ sở phân tích quang
phổ và phương pháp hóa học đã thu được chín saponin triterpenoid mới gồm bốn
saponin triterpenoid loại ursane là heptursosides AD (1-4), saponin triterpenoid loại
oleanane là heptoleosides AD (5-8), một saponin triterpenoid loại damarane,
heptdamoside A, cùng với hai saponin đã biết là asiaticoside D và scheffoleoside B.
Lần đầu tiên sự hiện diện của saponin triterpenoid tetracyclic từ Schefflera, các
saponin có vai trò chống viêm [32]. Tiếp tục nghiên cứu các hợp chất chiết xuất từ
vỏ thân, Wu Chun và cộng sự (2014) phân lập được năm saponin triterpenoid loại
ursane mới (1-5), các hợp chất được phân lập từ cây này đều được đánh giá về các
hoạt động ức chế sản xuất oxide nitric do lipopolysaccharide gây ra trong tế bào
RAW264.7, và các hợp chất 2 và 5 cho thấy hoạt động chống viêm yếu ở nồng độ
không gây độc tế bào của chúng [33].
Trung tâm Sâm Việt Nam đã hợp tác với Đại học Hiroshima Nhật Bản đã chiết
xuất từ vỏ thân đã xác định được 12 glycozid triterpenoid trong đó có 3 hợp chất đã
biết là asiaticozid, caulozid, 3-α-hydroxylup-ene-23,28-dioic acid 28-α-rhamnosyl
(1→4)-β-D-glucopyranozid (1→6)-β-D-glucopyranozid và chín hợp chất mới. Trong
12 hợp chất được xác định từ vỏ thân có 6 cặp có cấu trúc ursen và oleanen tương
ứng, các cặp glycozid ursen và oleanen tương ứng được đặt tên là scheffursozid A,
B, C, D, E, F và scheffoleozid A, B, C, D, E, F [27].
Rễ
Rễ NGBCC dùng làm thuốc bổ nên còn gọi là Sâm nam [10]. Từ nguồn nguyên
liệu là vỏ rễ của NGBCC được thu thập từ Quảng Châu, tỉnh Quảng Đông, Trung
Quốc, Chen và cộng sự (2015) đã phân lập, dùng kỹ thuật sắc ký, phân tích dữ liệu
quang phổ đã xác định được 6 triterpenoid là taraxerone (1), 3-epi-taraxerol (2),
aleuritolic acid (3), 3-oxofriedelan-28-oic acid (4), 3β,19α-dihydroxy-urs-12-ene-
24,28-dioic acid (5), asiatic aicd (6). Trong đó, các hợp chất từ 1-5 lần đầu tiên thu

10
được từ loài cây này. Những chất hóa học này có vai trò chống viêm, chống ung thư
và chống viêm khớp dạng thấp [7].
1.1.3.2. Công dụng
Trung tâm Sâm Việt Nam hợp tác với Viện nghiên cứu cây thuốc Poznan (Ba
Lan) cho thấy NGBCC có nhiều tác dụng [32].
Tác dụng trên hệ thần kinh trung ương: NGBCC tác động hai hướng trên hệ
thần kinh trung ương liều nhỏ (1/500 của LD50) kích thích nhẹ trên hệ thần kinh trung
ương, liều cao (1/20 của LD50) làm ức chế hệ thần kinh trung ương.
Tác dụng trên nội tiết tố sinh dục: Thực hiện trên hai nhóm súc vật có cơ địa
bình thường, trưởng thành và nhóm súc vật bị suy nhược sinh dục. Nhóm súc vật có
cơ địa bình thường, trưởng thành kết quả cho thấy sự tác động không có biểu hiện rõ
rệt. Ở nhóm súc vật bị suy nhược sinh dục đều có sự gia tăng trọng lượng cơ quan
sinh dục so với đối chứng, đồng thời sự tác động của oestrogen và androgen đều biểu
hiện ở liều thấp. Sự tác động của bộ phận lá mạnh hơn so với tác động của bộ phận
thân NGBCC.
Tác dụng kháng viêm, giảm đau: Nghiên cứu tiến hành trên bột chiết từ lá
NGBCC có tính kháng viêm rõ ở 2 liều thử 0,1 g/kg và 0,5 g/kg trong đó ở liều 0,1
g/kg tác dụng điển hình hơn. Nghiên cứu về tác dụng giảm đau cho kết quả 50% -
60%.
Những nghiên cứu về dược lý những năm gần đây cho thấy, các chất hóa học
được chiết xuất, phân tách, phân lập từ các bộ phận của NGBCC có nhiều hoạt tính
sinh học khác nhau như: kháng virus rất mạnh [3][4][5], tác dụng chống các tế bào
ung thư [6][7], chống viêm [7][32][33], chống viêm khớp [7], giảm đau [7], ngoài ra
NGBCC còn được sử dụng theo cách truyền thống để chữa chảy máu do chấn thương,
và tác dụng tăng đông máu có liên quan đến việc điều hòa mạch hoạt chất nội mô và
các thông số huyết học [8].
Trong nghiên cứu về dược lý saponin triterpenoid loại damarane cũng như các
dẫn xuất của chúng có hoạt tính sinh học chống khối u, chống viêm, kích thích miễn
dịch, tăng sinh tế bào thần kinh, chống lão hóa, chống vi khuẩn, chống tiểu đường,
chống loãng xương.
Từ lâu trong dân gian ở Trung Quốc, Việt Nam các bộ phận của cây NGBCC
(lá, vỏ thân, vỏ rễ, rễ nhỏ) được sử dụng làm thuốc, như rễ dùng làm thuốc bổ, vỏ

11
thân chữa thấp khớp, viêm họng, cảm sốt, vết thương sưng đau, ngoài ra vỏ còn giúp
ăn ngon, ngủ tốt, phục hồi và tăng cường sức khỏe, hạ nhiệt, chống viêm, giảm đau,
trị thấp khớp, đau xương, chữa bệnh gan, tác dụng tốt đến hệ thần kinh trung ương,
chống suy nhược thần kinh, giúp tăng cường trí nhớ [27][10].
1.2. Tình hình nghiên cứu về phôi vô tính, rễ bất định ở một số loài thuộc các
chi quan trọng ở họ Ngũ gia bì (Araliaceae)
1.2.1. Chi Panax
1.2.1.1. Phôi vô tính
Qua sử dụng phôi hợp tử non Panax ginseng, Arya và cộng sự (1993) đã
nghiên cứu sự hình thành phôi vô tính và mô sẹo có khả năng sinh phôi. Các phôi sơ
cấp đã tạo phôi thứ cấp cấp 1, cấp 2 và sự sinh trưởng của chúng phụ thuộc vào chất
điều hòa sinh trưởng (ĐHST). Một số loại auxin được sử dụng như 2,4-D, NAA và
IAA 1 mg/L rất thích hợp cho tạo phôi thứ cấp; ngược lại, kinetin, BA ức chế tạo
phôi thứ cấp [34].
Choi và cộng sự (2003) đã sử dụng các mảnh lá mầm phôi hợp tử sâm Panax
ginseng tạo phôi vô tính trực tiếp trên môi trường đặc MS không bổ sung chất ĐHST,
có 30 g/L đường; ngược lại, môi trường có nồng độ khoáng NH4NO3 cao (60 mM)
chỉ cảm ứng tạo mô sẹo có khả năng sinh phôi. Mô sẹo có khả năng sinh phôi được
cấy chuyển nhiều lần trên môi trường vẫn có khả năng phát triển tiếp tục dù không
bổ sung chất ĐHST. Mô sẹo được nuôi cấy trong bình tam giác (V500 mL) chứa môi
trường MS/½MS có 30 g/L đường để nhân lên phục vụ cho nuôi cấy quy mô lớn hơn
trong bioreactor (V20 lít); khối lượng tươi mô tăng lên 7,1 lần sau 5 tuần nuôi cấy
trong bioreactor chứa môi trường 1/3MS có 30 g/L đường. Hàm lượng ginsenoside
tổng số thấp hơn 6 lần so với rễ cây sâm sinh trưởng ở điều kiện tự nhiên [35].
Cũng từ vật liệu mảnh lá mầm phôi hợp tử, Zhou và Brown (2006) đã xây
dựng được quy trình tạo cây sâm Bắc Mỹ (Panax quinquefolius) có hiệu quả thông
qua nuôi cấy tạo phôi vô tính. Tiền xử lý mảnh lá mầm với 1 M đường ở 4oC đã cải
thiện được số lượng phôi hình thành. Việc kết hợp tiền xử lý với nuôi cấy dùng nồng
độ đường cao (7%) cũng đã nâng cao được tỷ lệ phát sinh phôi từ 40% lên 75%, số
lượng phôi/mẫu cấy từ 10 lên 21. Tỷ lệ hình thành phôi thứ cấp đạt 90% khi mô phôi
vô tính sơ cấp được nuôi cấy trên môi trường MS có 1 mg/L 2,4-D và 1 mg/L NAA.
Phôi phát triển đến giai đoạn trưởng thành (trong đó có ½ lượng phôi nẩy chồi) trên

12
môi trường SH có 1% than hoạt tính và 85% phôi phát triển thành cây con (đầy đủ rễ
thân và lá) khi được nuôi cấy trên môi trường ½SH có 0,5% than hoạt tính. Cây con
có kiểu hình bình thường ở giai đoạn ex vitro [36].
Nghiên cứu của Kim và cộng sự (2012) đã thiết lập hệ thống tái sinh phôi vô
tính thứ cấp Panax ginseng trực tiếp từ phôi vô tính sơ cấp, nuôi cấy trên môi trường
MS không bổ sung chất ĐHST. Môi trường EM (1/3MS, với ½ lượng NH4NO3 và
KNO3, có 20 - 30 g/L sucrose) tạo điều kiện thích hợp cho sự phát triển của phôi thứ
cấp đến giai đoạn cây con. Cây con có rễ cọc hoàn chỉnh khi được nuôi cấy trên môi
trường ½SH có 0,5% than hoạt tính. Tần suất cao của hệ thống tái sinh này thông qua
quá trình phát sinh phôi thứ cấp theo chu kỳ (cyclic manner). Sự tăng sinh liên tục
của phôi vô tính thông qua tạo phôi thứ cấp, nên rất hữu ích cho nhân giống.[37].
Kỹ thuật nuôi cấy lát mỏng tế bào (LMTB) đã được sử dụng trong tạo phôi
trực tiếp từ một số bộ phận khác nhau của cây sâm Ngọc Linh (Panax vietnamensis)
in vitro (3 tháng tuổi) như lá, cuống lá và củ. Mô được nuôi cấy trên môi trường MS
bổ sung NAA và 2,4-D ở các nồng độ 0,1; 0,2; 0,5; 1,0 và 2,0 mg/L. Sau 10 tuần nuôi
cấy, kết quả thu được cho thấy nguồn mẫu từ lá là thích hợp nhất cho sự hình thành
phôi trực tiếp. Mẫu cấy lá được nuôi cấy trên môi trường MS bổ sung 2 mg/L NAA
cho hiệu quả phát sinh phôi trực tiếp cao nhất (29,49 phôi/mẫu). Kết quả khảo sát
hình thái giải phẫu cho thấy phôi phát triển trực tiếp từ các mẫu nuôi cấy [38]. Trong
nghiên cứu của Vũ Thị Hiền và cộng sự (2015) đã nghiên cứu sự phát sinh hình thái
từ LMTB (TCL) mẫu lá, cuống lá và thân rễ sâm Ngọc Linh in vitro. Mẫu được nuôi
cấy trên môi trường đặc MS có bổ sung 30 g/L đường, các chất ĐHST (NAA, 2,4-D,
BA và TDZ riêng lẻ hoặc kết hợp). Sau 10 tuần nuôi cấy, kết quả cho thấy mẫu lá
tTCL_L, mẫu cuống lá lTCL_C, mẫu thân rễ tTCL_R đều cho sự phát sinh phôi, mô
sẹo, rễ, trong khi mẫu cuống lá tTCL_C chỉ cho sự phát sinh mô sẹo và rễ. Trong đó,
tỷ lệ phát sinh phôi cao nhất (89,6%), tỷ lệ phát sinh mô sẹo cao nhất (91 - 98,8%),
tỷ lệ phát sinh rễ cao nhất (98,8%) đã được ghi nhận tương ứng khi mẫu lá tTCL_L
được nuôi cấy trên môi trường bổ sung 2 mg/L NAA và được đặt dưới điều kiện chiếu
sáng 16 h/ngày; lá tTCL_L, cuống lá tTCL_C, lTCL_C, thân rễ tTCL_R được nuôi
cấy trên môi trường có 2,4-D kết hợp với BA dưới điều kiện chiếu sáng 16 h/ngày,
điều kiện tối hoàn toàn; môi trường có bổ sung 1 mg/L NAA và đặt trong điều kiện
tối hoàn toàn. Điều kiện chiếu sáng có tác động đáng kể lên khả năng phát sinh hình

13
thái của mẫu cấy. Việc sử dụng đèn huỳnh quang chiếu sáng 16 h/ngày phù hợp cho
khả năng phát sinh phôi của mẫu cấy, những phôi thu được có dạng hình cầu, hình
tim, hình thủy lôi và cả dạng lá mầm. Ngược lại, điều kiện tối lại kích thích sự hình
thành rễ và mô sẹo tốt hơn. Rễ thu được có màu trắng đục, có phân nhánh, trong khi
mô sẹo lại xốp và có màu vàng nhạt [39]. Ở sâm Ngọc Linh, Mai Trường và cộng sự
(2014) đã nghiên cứu tạo và nhân phôi vô tính trong môi trường lỏng. Lá chét của
cây sâm 6 tháng tuổi ở vườn ươm (sau khử trùng) được nuôi cấy trên môi trường MS
có 1 mg/L 2,4-D và 0,2 mg/L kinetin để tạo mô sẹo. Sau đó, mô sẹo được cấy chuyền
sang môi trường MS lỏng (lắc) có 1 mg/L 2,4-D với 0,2 mg/L kinetin, 500 mg/L
casein hydrolysate để tạo huyền phù tế bào. Sau 2 tháng, huyền phù tế bào được
chuyển sang nuôi cấy trong môi trường B5 lỏng (lắc) có 3 mg/L IBA. Sau nhiều tháng
nuôi, rất nhiều phôi vô tính dạng cầu hình thành và có khả năng nhân ổn định. Mảnh
lá mầm (của cây mầm từ phôi) và phôi vô tính non được nuôi cấy trên môi trường
MS có 10% nước dừa, có hoặc không có 0,2 mg/L IBA để tạo mô sẹo sinh phôi và
phôi thứ cấp. Mô sẹo sinh phôi và phôi thứ cấp này cũng đã được dùng để nuôi nhân
trong môi trường lỏng có và không có chất ĐHST ở quy mô bình tam giác và
bioreactor [40]. Những kết quả đã tạo cơ sở để nhân giống quy mô lớn và thu hợp
chất thứ cấp.
Rễ bất định của cây Panax ginseng hoang dại và cây đột biến được nuôi cấy
tạo mô sẹo trên môi trường MS có 0,5 mg/L 2,4-D và 0,3 mg/L kinetin. Mô sẹo có
khả năng sinh phôi được cấy chuyển sang môi trường MS có 0,5 mg/L 2,4-D nhằm
cảm ứng tạo phôi. Các phôi hình thành trưởng thành trên môi trường MS có 5 mg/L
GA3 và 85% phôi nẩy mầm [41].
Đỗ Mạnh Cường và cộng sự (2020) đã nghiên cứu làm tăng tỷ lệ phát sinh
phôi vô tính sâm Ngọc Linh qua khử trùng lá sâm bằng dung dịch nano bạc và nuôi
cấy dùng môi trường cũng bổ sung nano bạc. Kết quả cho thấy tỷ lệ tạo mô sẹo có
khả năng sinh phôi cao nhất (72,22%), khối lượng tươi cao nhất (0,77 g) khi sử dụng
dung dịch nano bạc xử lý trong 20 phút. Tỷ lệ phát sinh phôi và phôi nẩy mầm cao
nhất trên môi trường MS có 1 mg/L 2,4-D; 0,5 mg/L NAA; 02 mg/L Kin và 1,6 mg/L
nano bạc. Việc bổ sung 1 mg/L NAA và 1,2 mg/L nano bạc cho kết quả tốt nhất về
chiều cao chồi, chiều dài rễ, số rễ, khối lượng tươi/khô cây con [42].

14
Nguyễn Thị Ngọc Hương và cộng sự (2018) đã nghiên cứu hình thái học và tế
bào học quá trình phát sinh phôi vô tính gián tiếp qua giai đoạn mô sẹo ở Tam thất
hoang (Panax stipuleanatus). Ở nghiên cứu này, mô sẹo được cảm ứng từ mô thân rễ
nuôi cấy trên môi trường MS (½ khoáng đa lượng) có bổ sung 2 mg/L 2,4-D (giai
đoạn nuôi 8 tuần) và 1 mg/L (giai đoạn nuôi 16 tuần). Khối mô sẹo bao gồm các cụm
tế bào rời rạc và cụm tế bào đẳng kính được cấy chuyển sang nuôi cấy trên môi trường
có bổ sung 5 m/L NAA để tạo mô sẹo có khả năng sinh phôi, kết quả cho thấy khối
mô sẹo trở nên mềm hơn với các cụm tế bào có khả năng tái sinh cao: kích thước nhỏ,
đẳng kính, nhân to, nhân rõ và có tế bào chất đậm đặc. Sự hình thành các thể giống
phôi cầu được ghi nhận sau 28 tuần nuôi trên môi trường có NAA nêu trên. Các cấu
trúc giống phôi này tiếp tục phát triển đến giai đoạn hậu phôi cầu, phôi dạng trái tim
và dạng có lá mầm trên môi trường MS như trên có bổ sung 0,5 mg/L BA và 1 mg/L
GA3. Ngoài ra, cũng ghi nhận được một số dạng phát triển bất thường của phôi như
chỉ phát triển rễ/chồi hoặc chỉ có lá mầm [43].
1.2.1.2. Rễ bất định
Gao và cộng sự (2005) đã nghiên cứu sự tái sinh trực tiếp rễ bất định từ đoạn
cuống lá (0,5-0,8 cm) và từ đoạn rễ nhánh (~ 1 cm) cây Panax notoginseng (3 năm
tuổi). Kết quả cho thấy rễ bất định hình thành tốt từ mô cuống lá nuôi cấy trên môi
trường MS có 3 mg/L IBA, có hoặc không bổ sung 0,1 mg/L kinetin. Mô rễ nhánh
mẫn cảm hơn đối với IBA trong quá trình nuôi cấy tạo rễ bất định so với mô cuống
lá. Khối lượng khô rễ bất định tăng 5,25 lần sau quá trình nuôi lỏng lắc trong môi
trường ½MS có 3 mg/L IBA [44].
Ba loại mô dùng tạo rễ bất định sâm Ngọc Linh bao gồm mô sẹo lá chét,
phôi/cụm phôi trưởng thành mang chồi và mô sẹo cứng. Rễ bất định hình thành ngay
trong giai đoạn nuôi cấy mảnh lá (nuôi cấy một giai đoạn, không qua cấy chuyền)
trên môi trường MS bổ sung 2 mg/L 2,4-D và 10% nước dừa; rễ bất định cũng hình
thành từ mô sẹo cấy chuyền (nuôi cấy hai giai đoạn) trên môi trường MS có 3 mg/L
NAA, MS có 3 mg/L NAA, 2 mg/L 2,4-D, 5% (v/v) nước dừa và môi trường B5 có
5 mg/L IBA. Ngoài ra, sự hình thành rễ bất định từ phôi/cụm phôi (đã loại bỏ hoàn
toàn rễ trụ) cũng đã được ghi nhận qua nuôi cấy trên môi trường ½MS bổ sung 2 - 3
mg/L NAA. Rễ bất định từ mô sẹo cứng (từ môi trường B5 có 5 mg/L IBA) đã được
nghiên cứu nuôi cấy tiếp tục trên môi trường đặc/lỏng White có 5 mg/L NAA, B5 có

15
1 - 5 mg/L IBA/NAA nhằm tìm hiểu khả năng tăng sinh khối qua quá trình tạo rễ thứ
cấp. Nghiên cứu khả năng hình thành các sơ khởi rễ và phân nhánh rễ trên các môi
trường White/B5 có 1 - 5 mg/L IBA/NAA từ khúc cắt rễ bất định cũng đã được thực
hiện [45].
Nguyễn Thị Ngọc Hương và cộng sự (2016) đã sử dụng các khúc cắt thân rễ
Panax stipuleanatus có đường kính 1 - 1,5 cm và dày 1 cm được nuôi cấy trên môi
trường MS có bổ sung 30 g/L đường, 6 g/L agar, 0,5 mg/L 2,4-D và đặt trong tối. Mô
sẹo hình thành trên bề mặt thân rễ sau bốn tuần. Sự phân chia đầu tiên trong quá trình
hình thành mô sẹo xảy ra trong hai tuần đầu, ở các tế bào nhu mô vỏ cấp hai và tượng
tầng libe – mộc. Mô sẹo 26 tuần tuổi với các tế bào bên trong cụm chậm tăng trưởng
và các tế bào ở phía ngoài cụm có xu hướng kéo dài được chuyển sang môi trường
hoạt hóa có bổ sung 0,5 mg/L 2,4-D và 0,1 mg/L TDZ trong 6 tuần. Mô sẹo phát triển
trên môi trường này trở nên chặt hơn và hình thành nhiều cụm. Sự hình thành rễ bất
định xảy ra sau 10 tuần khi mô sẹo được chuyển sang môi trường MS có bổ sung 0,5
mg/L NAA. Mô sẹo hình thành các cấu trúc hình cầu giống phôi tuy nhiên chỉ phát
triển một cực rễ. Trong các rễ hình thành từ mô sẹo thân rễ có sự hiện diện của saponin
thuộc nhóm olean [46].
Nghiên cứu cải tiến quy trình nhân nhanh sinh khối rễ thứ cấp sâm Ngọc Linh
từ rễ bất định in vitro thông qua việc tối ưu nguồn mẫu, môi trường nuôi cấy và điều
kiện nuôi cấy đã được thực hiện. Kết quả cho thấy, các rễ thứ cấp sâm Ngọc Linh
được hình thành từ vùng trụ bì tại vị trí của bó mạch dẫn trung tâm. Các mẫu rễ bất
định dài 2 cm tái sinh từ cuống lá với 8 g rễ bất định/1,5 L môi trường nuôi cấy trong
bioreactor hình cầu có sục khí cho kết quả hình thành rễ thứ cấp tốt nhất trên môi
trường MS cải biên với tỷ lệ ½NH4+/NO3
- bổ sung 7 mg/L IBA, 0,5 mg/L BA và 30
g/L đường trong 56 ngày (45 ngày đầu trong tối, 11 ngày sau ngoài sáng) ở 22oC và
pH 5,3 [47].
1.2.2. Chi Acanthopanax
1.2.2.1. Phôi vô tính
Chi Aucanthopanax còn có tên gọi khác là Eleutherococcus, đây là chi đã được
nghiên cứu khá nhiều về phôi vô tính. Các mảnh lá mầm và thân phôi hợp tử
Acanthopanax senticosus tạo phôi vô tính qua nuôi cấy trên môi trường MS có 0,5
mg/L 2,4-D. Phôi thứ cấp hình thành từ phôi vô tính sơ cấp khi được nuôi cấy trên

16
môi trường MS có 2,4-D (0,5 mg/L) hoặc IAA (1-3 mg/L) hoặc Zeatin (0,5 mg/L) và
NAA (0,2 mg/L). Các phôi vô tính nẩy mầm (93%) trên môi trường MS không bổ
sung 2,4-D. Phôi vô tính có nguồn gốc từ cụm tế bào biểu mô và dưới biểu mô [48].
Các đoạn thân cây Acanthopanax koreanum Nakai tạo mô sẹo có khả năng
phát sinh phôi tốt qua nuôi cấy trên môi trường MS chỉ bổ sung 4,5 µM 2,4-D. Mô
sẹo có khả năng phát sinh phôi chỉ tạo phôi cầu khi được nuôi cấy trong môi trường
lỏng có 0,45 µM 2,4-D; ngược lại, ghi nhận phôi phát triển đến giai đoạn trưởng
thành, nẩy mầm khi được nuôi cấy trong môi trường không có 2,4-D. Cytokinin ức
chế sự phát triển bình thường của phôi nhưng kích thích sự phát sinh phôi thứ cấp
trên bề mặt của phôi sơ cấp [49].
Park và cộng sự (2005) đã thực hiện nghiên cứu tạo phôi vô tính
Eleutherococcus koreanum từ nuôi cấy các đoạn rễ bất định trong môi trường lỏng
lắc. Kết quả cho thấy môi trường 1/3MS có 60 g/L đường, không bổ sung chất ĐHST
là thích hợp cho sự hình thành phôi vô tính. Phôi vô tính phát triển đến giai đoạn
trưởng thành và nẩy mầm trong môi trường 1/3MS không bổ sung chất ĐHST. Rễ
bất định, phôi và cây con đã được thu nhận đồng thời sau 12 tuần nuôi cấy rễ bất định
bằng bioreactor trong môi trường như trên [50]. Các kết quả nghiên cứu trên có ý
nghĩa trong nhân giống và thu hợp chất thứ cấp.
Shohael và cộng sự (2007) đã nghiên cứu thu nhận các sản phẩm có giá trị như
eleutherosides và acid chlorogenic từ phôi vô tính có nguồn gốc nuôi cấy huyền phù
tế bào có khả năng sinh phôi Eleutherococcus senticosus. Mô tế bào được xử lý
methyl jasmonate (50 - 400 µM), kết quả cho thấy hàm lượng eleutherosides, acid
chlorogenic, khối lượng tươi/khô, tỷ lệ tăng trường phôi cao nhất khi được xử lý 200
µM methyl jasmonate [51].
1.2.2.2. Rễ bất định
Seo và cộng sự (2003) đã nghiên cứu nhân sinh khối rễ bất định Eleutherococcus
sessiliflorus bằng phương pháp nuôi lỏng lắc và bioreactor. Cảm ứng tạo rễ được thực
hiện bằng nuôi cấy mảnh mô trụ dưới lá mầm, đoạn rễ phôi vô tính trên môi trường đặc
½MS có bổ sung NAA, IBA và IAA. Kết quả cho thấy rễ hình thành từ mô trụ dưới lá
mầm tốt hơn từ đoạn rễ. Trong các auxin được thí nghiệm, nhận thấy tỷ lệ tạo rễ cao nhất
trên môi trường có 0,5 mg/L NAA. Ở nuôi cấy lỏng lắc, hiệu quả tạo rễ ở môi trường
MS (không có NH4NO3) cao hơn trong môi trường MS và ½MS. Rễ tăng khối lượng

17
tươi tốt trong môi trường có 0,5 mg/L IBA. Hệ số nhân (5,5 lần) sau 1 tháng khi rễ được
nuôi trong bioreactor (V10 lít) [52].
Lee và Paek (2012) đã nghiên cứu ảnh hưởng của nguồn đạm đến phát triển
sinh khối rễ bất định Eleutherococcus koreanum Nakai và sự sản sinh các chất có
hoạt tính sinh học từ rễ qua nuôi cấy trong bioreactor sục khí (3 lít). Kết quả cho thấy
tỷ lệ đạm NH4+:NO3
- (5:25 mM và 10:20 mM) cho kết quả sinh khối tươi/khô cao
nhất sau 5 tuần nuôi cấy, hàm lượng eleutherosides B và E đạt cao nhất ở tỷ lệ
NH4+:NO3
- là 5:25. Tăng lượng NH4+ làm giảm sinh khối, đạm nitrate thích hợp hơn
đạm ammonium trong nhân sinh khối rễ và thu hợp chất thứ cấp [53]. Kết quả tạo
tiền đề cho sản xuất sinh khối rễ thu hoạt chất ở quy mô pilot nhằm thương mại hóa
sản phẩm.
1.2.3. Chi Polyscias
Sliwinska và cộng sự (2008) đã tạo thành công phôi vô tính loài đinh lăng lá
to (Polyscias filicifolia). Sau khử trùng, các mảnh lá (5 - 10 mm) của cây 2 năm tuổi
được nuôi cấy để tạo mô sẹo. Mô sẹo loại I hình thành trên môi trường MS có 0,5
mg/L 2,4-D, 1 mg/L BAP và mô sẹo loại II hình thành trên môi trường MS có 2,4-D
2 mg/L, kinetin 0,01 mg/L. Mô sẹo loại I là loại mô sẹo chắc và có màu xanh, mô sẹo
loại II là loại mô sẹo có cấu trúc xốp và màu trắng kem (cả hai loại mô sẹo trên đều
có khả năng tạo phôi). Phôi vô tính sơ cấp và phôi thứ cấp được hình thành trong môi
trường lỏng ½MS không bổ sung chất ĐHST. Phôi tạo cây con tốt nhất trên môi
trường Nitsch và Nitsch cải tiến có bổ sung 0,5 mg/L kinetin, 0,1 mg/L IBA và 10
mg/L adenine sulfate [54].
Phôi vô tính đinh lăng lá nhỏ (Polyscias fruticosa) nuôi lỏng lắc tăng sinh khối
do sự hình thành phôi bất định thứ cấp. Phôi vô tính tăng trưởng sinh khối tốt khi
được nhân in vitro bằng hệ thống nuôi lỏng lắc với khối lượng phôi nuôi cấy ban đầu
là 1 g (1,25% - w/v), tốc độ lắc 100 vòng/phút; khối lượng tăng sinh đạt 7,5 g, hệ số
tăng sinh khối phôi đạt 8,5 lần tại thời điểm 30 ngày sau cấy. Đã xây dựng được
đường biểu diễn tăng trưởng khối lượng, hệ số tăng trưởng sinh khối phôi dưới ảnh
hưởng của khối lượng phôi nuôi cấy ban đầu và tốc độ lắc. Phôi vô tính đinh lăng lá
nhỏ có cấu trúc điển hình, trưởng thành và tạo cây hoàn chỉnh với tỷ lệ cao (95%) khi
được nuôi cấy trên môi trường ½MS không bổ sung chất ĐHST, cây có kiểu hình và
sinh trưởng bình thường trên môi trường nuôi cấy [55].

18
1.2.4. Chi Schefflera
1.2.4.1. Schefflera octophylla (Lour.) Harms
Li và cộng sự (2004) đã xây dựng thành công quy trình nhân giống Schefflera
octophylla bằng phương pháp nuôi cấy in vitro trong đó có giai đoạn tạo rễ bất định.
Các đốt thân (mang chồi ngủ) của cây ở bầu đất, sau khử trùng, được nuôi cấy trên
môi trường MS có bổ sung 0,5 mg/L BA và 2 mg/L kinetin nhằm thích chồi ngủ phát
triển. Kết quả cho thấy, các chồi ngủ phát triển ở 5 ngày sau cấy (NSC) và tạo thành
cụm chồi nhỏ ở 20 NSC với tỷ lệ 91%. Tiếp theo, các chồi nhỏ này phát triển chiều
cao (3 - 4 cm), phát triển số lượng cá thể với hệ số nhân 3,4 lần ở giai đoạn 22 NSC
qua nuôi cấy trên môi trường MS có 0,5 mg/L NAA và 1,5 mg/L BA. Để tạo rễ, các
chồi được nuôi cấy trên môi trường ½MS và 0,2 mg/L NAA có 0,3% than hoạt tính
và kết quả cho thấy, rễ hình thành rất nhiều với chiều dài trung bình ~ 4 cm ở 30
NSC. Các cây con (cao 3 - 4 cm) đã được trồng thành công trong chậu chứa hỗn hợp
perlite, vermiculite và cát mịn (1:1:1) với tỷ lệ sống ~ 75% [3].
1.2.4.2. Schefflera arboricola (Hayata) Merr.
Sau khử trùng, các đốt thân được nuôi cấy trên môi trường nhằm kích thích
chồi ngủ phát triển và tạo cụm chồi; các mảnh lá và trục lá cũng được nuôi cấy nhằm
tạo mô sẹo và tái sinh chồi. Kết quả cho thấy, ở nuôi cấy đốt thân, 0,5 mg/L BA ở
môi trường MS kích thích chồi phát triển tốt trong 8 - 10 ngày, chồi cao 0,5 cm với 4
lá. Thí nghiệm tạo cụm chồi đã cho kết quả âm tính dù đã bố trí thực hiện một số
nghiệm thức khác nhau về tổ hợp chất ĐHST như (NAA và BA); (IAA và kinetin);
(2,4-D và BA) và (BA và kinetin). Mảnh lá tạo mô sẹo trên môi trường MS có 1 - 2
mg/L 2,4-D sau 13 - 16 ngày nuôi cấy; trục lá tạo mô sẹo trên môi trường MS có
NH4NO3 200 mg/L bổ sung 0,5 mg/L 2,4-D và 0,5 mg/L BAP sau 6 ngày. Chỉ thu
được kết quả tái sinh rễ trên môi trường có BA/kinetin/BA và adenine sulphate qua
nuôi cấy tái sinh dùng mô sẹo [56].
Misra (2001) đã nêu một số kết luận như các chồi ngủ được cảm ứng, phát
triển trên môi trường MS có bổ sung 2,21 µM BA; nồng độ BA cao hơn (8,87 µM)
đã cảm ứng sự hình thành cụm chồi; các chồi tạo rễ sau khi được cấy chuyển sang
môi trường tạo rễ MS có bổ sung 2,63 µM NAA. Các cây con được trồng thành công
trong chậu chứa hỗn hợp vermiculite và perlite (tỷ lệ 1:1) và ở chậu đất giai đoạn tiếp
theo [57].

19
Schefflera arboricola (dạng lùn – dwarf schefflera) là một trong các loại cây
cảnh dùng trang trí nội thất, thân và lá cây chứa tinh dầu ức chế đối với nhiều loài vi
khuẩn [58]. Nhân giống vô tính in vitro 03 dòng (‘Luseane’, ‘Charlotte’ và ‘Gold
Capella’) đã được thực hiện [59]. Sau khử trùng, các đốt thân được nuôi cấy trên môi
trường MS có bổ sung cytokinin khác nhau. Kết quả cho thấy số lượng chồi đạt giá
trị cao nhất trên môi trường có bổ sung 0,5 mg/L thidiazuron (TDZ) đối với dòng
‘Luseane’, hoặc 8 mg/L BA đối với 02 dòng ‘Charlotte’ và ‘Gold Capella’. Nuôi cấy
các đốt thân cây in vitro cũng cho kết quả nhân giống tương tự. Hai dòng ‘Luseane’
và ‘Gold Capella’ 100% chồi tạo rễ trên môi trường có bổ sung 2 mg/L NAA trong
khi dòng ‘Charlotte’ tạo rễ với tỷ lệ 93,75% trên môi trường có nồng độ NAA thấp
hơn - 1 mg/L. Các cây hoàn chỉnh của 03 dòng nói trên đã được trồng thành công
trong chậu với các giá loại giá thể khác nhau.
1.2.4.3. Schefflera leucantha Viguier
Schefflera leucantha, tương tự các loài trên, cũng chứa nhiều thành phần hóa
học có khả năng ức chế khá nhiều loài vi khuẩn gây bệnh [60]. Chirakiattikun và
Prakaisrithongkham (2004) đã thực hiện có kết quả nghiên cứu xây dựng môi trường
thích hợp cho vi nhân giống Schefflera leucantha Viguier. – loài cũng hàm chứa nhiều
hợp chất có hoạt tính sinh học. Chồi đã được bố trí nuôi cấy trên môi trường MS
không bổ sung chất ĐHST hoặc có (BA, IAA) với các nồng độ khác nhau, và nước
dừa 15%. Kết quả cho thấy, nghiệm thức ở môi trường có 2 mg/L BA kích thích tạo
chồi tốt nhất so với các nghiệm thức khác với chiều cao chồi 1,4 cm, số lá 3,6, số đốt
6,9, số chồi nách 3,4, chiều cao chồi nách 0,5 cm. Để tạo rễ bất định, chồi được nuôi
cấy trên môi trường có bổ sung 0; 1; 1,5; 2 và 2,5 mg/L IAA và kết quả cho thấy,
chồi trên môi trường MS có 2 mg/L IAA tạo rễ tốt nhất ở các chỉ tiêu như tỷ lệ tạo
rễ, số rễ và chiều dài rễ, lần lượt là 66,7%; 3,2 rễ và 2 cm [61].
Như vậy, đến nay chưa ghi nhận được bất kỳ công bố kết quả nghiên cứu nào
liên quan đến tạo phôi vô tính ở 03 loài Schefflera nêu trên.
1.2.4.4. Didymopanax morototoni (Aublet) Decaisne et Planchon.
Didymopanax morototoni còn có tên đồng nghĩa là Schefflera morototoni
(Aublet) Macguire, Steyerm. et Frodin.
Schefflera morototoni – loài thân gỗ, cây có thân cao, quả có thể sử dụng vì có
giá trị dinh dưỡng; thành phần hóa học trong thân và lá cũng đã được nghiên cứu

20
[62]. Franco và cộng sự (2006) [63] đã sử dụng một số vật liệu từ hạt nẩy mầm như
rễ, chồi, đốt thân và lá mầm để nghiên cứu tạo mô sẹo/phôi vô tính và tạo cây từ phôi.
Kết quả cho thấy mẫu lá mầm, đốt thân có đáp ứng tạo mô sẹo/phôi tốt hơn so với
hai vật liệu còn lại; môi trường WPM [64] có bổ sung 5 mg/L 2,4-D và 0,1 mg/L
kinetin cho tỷ lệ tạo phôi cao nhất (60%), thấp nhất là trên môi trường có 1 mg/L 2,4-
D và 1 mg/L kinetin (20%). Phôi được cấy chuyển sang môi trường WPM không bổ
sung chất ĐHST, có 10 g/L đường (hoặc có bổ sung 0,1 mg/L BAP và 0,5 mg/L GA3)
để tạo cây; cây được trồng ra đất với tỷ lệ sống 33%.
1.3. Sự phát sinh phôi vô tính
1.3.1. Cơ sở khoa học của sự phát sinh phôi vô tính
Phôi vô tính/phôi soma/phôi sinh dưỡng (somatic embryogenis) đều là khái
niệm mô tả một cấu trúc lưỡng cực gồm cực chồi và cực rễ, bắt nguồn từ tế bào sinh
dưỡng hay tế bào soma, dưới những điều kiện thích hợp sẽ phát triển thành một cơ
thể có chức năng hoàn chỉnh. Quá trình hình thành phôi vô tính trải qua các giai đoạn
tương tự như phát sinh phôi hợp tử, trải qua các giai đoạn phôi hình cầu, hình trái tim,
dạng thủy lôi và giai đoạn lá mầm, và sau đó phát triển thành cây con hoàn chỉnh.
Trong quá trình phát triển phôi vô tính, các giai đoạn hình thái không có kết nối với
mô mạch [65]. Phôi vô tính rất giống phôi hữu tính về mặt hình thái, quá trình phát
triển cũng như về mặt sinh lý nhưng không phải là sản phẩm của sự thụ tinh giữa giao
tử đực và giao tử cái, không có quá trình tái tổ hợp di truyền, các phôi vô tính mang
thông tin di truyền giống hệt tế bào soma đã sinh ra nó. Phôi vô tính có thể được hình
thành từ một tế bào đơn hay từ cả một cụm tế bào. Thông qua một quá trình phân chia
có thứ tự phôi diễn ra sự biệt hóa, trưởng thành và phát triển thành cây con [66].
Quá trình phát sinh phôi vô tính đại diện cho mô hình toàn thế hoàn chỉnh, liên
quan đến hoạt động của các tín hiệu phức tạp, cũng như việc lập trình lại sự biểu hiện
gen được điều chỉnh cụ thể. Sự điều hòa này đáp ứng với các kích thích ngoại sinh
được tạo ra do các chất ĐHST thực vật [65]. Cảm ứng phát sinh phôi vô tính có tính
năng kích hoạt lại chu kỳ tế bào trong các tế bào thực vật đã được biệt hóa, dưới tác
động của các kích thích bên ngoài. Từ đó, mở ra con đường chuyển từ loại tế bào
soma sang loại tế bào sinh phôi [67]. Các kích thích này khởi động chương trình di
truyền dẫn đến thiết lập các dòng tế bào có kiểu gen phiên mã bị thay đổi, đầu tiên là
sự hình thành cấu trúc không đối xứng đỉnh – đáy. Phôi vô tính trưởng thành giống

21
phôi hợp tử đều thể hiện đỉnh – đáy và phân cực hướng tâm, có chồi sơ cấp và mô
phân sinh rễ, chứa các cơ quan phôi điển hình, lá mầm, rễ mầm [66] .
Tế bào thực vật có tính toàn thế do vậy có thể tái tạo thành cây hoàn chỉnh
thông qua con đường phát sinh phôi vô tính (nhờ vai trò quan trọng của auxin và
cytokinin, ABA, các stress,..) hoặc/và con đường phát sinh cơ quan. Theo Von Arnold
và cộng sự (2002), sự phát sinh phôi vô tính được xác định là quá trình hình thành
cấu trúc có hai cực chồi và rễ tương tự như phôi hợp tử, mỗi cực đều có mô phân sinh
và hệ thống tiền mạch (provascular) phân biệt với hệ mạch ở thể chồi/cụm chồi tái
sinh theo con đường phát sinh cơ quan. Phôi vô tính hình thành và phát triển từ tế bào
sinh dưỡng, có thể tái sinh trực tiếp từ mô nuôi cấy mà không qua giai đoạn trung
gian là mô sẹo hoặc/và tái sinh từ mô sẹo (tái sinh gián tiếp) [68]. Theo tài liệu, ở tạo
phôi vô tính trực tiếp, phôi có nguồn gốc từ tế bào PEDC (Pre-Embryogenic
Determined Cell) – tế bào được xác định có khả năng tạo phôi; ở tạo phôi gián tiếp,
phôi hình thành từ tế bào IECD (Induced Embryogenic Determined Cell) – tế bào
được xác định có khả năng tạo phôi thông qua cảm ứng.
Ở tạo phôi trực tiếp, theo một số tài liệu, ghi nhận có các bước tạo phôi vô tính
cơ bản như: (1) Khởi tạo loại tế bào có khả năng sinh phôi từ một số loại tế bào khác
nhau ở mô cấy ban đầu như tế bào biểu bì/dưới biểu bì hoặc tế bào ở/gần vùng hệ
mạch dưới tác động của auxin (có hoặc không có cytokinin) ngoại sinh, tác nhân tạo
áp suất thẩm thấu cao,..; (2) Khởi tạo phôi với các giai đoạn phát triển khác nhau, và
(3) Phôi phát triển với hệ mạch độc lập với mô mẹ.
Ở tạo phôi gián tiếp, theo Von Arnold và cộng sự (2002), có 5 bước từ lúc tạo
vật liệu tế bào có khả năng sinh phôi đến giai đoạn nhận được cây hoàn chỉnh, là (1)
Tạo được vật liệu tế bào có khả năng sinh phôi từ nuôi cấy vật liệu ban đầu (như
mảnh lá,..) trên môi trường có auxin (có hoặc không có cytokinin); (2) Nuôi cấy nhân
sinh khối tế bào có khả năng sinh phôi trên môi trường đặc hoặc trong môi trường
lỏng với chất ĐHST như trên; (3) Nuôi cấy ‘tiền trưởng thành’ phôi ở môi trường
không có chất ĐHST, điều này ức chế sự tiếp tục tăng sinh khối và kích thích sự hình
thành và phát triển phôi ở giai đoạn ban đầu; (4) Tạo phôi trưởng thành qua nuôi cấy
trên môi trường có ABA hoặc/và có áp suất thẩm thấu giảm; (5) Tạo cây con qua nuôi
cấy trên môi trường không có chất ĐHST [68].

22
Đến nay, quá trình và cơ chế sinh phôi ở thực vật mô hình Arabidopsis đã được
chỉ rõ. Ở tạo phôi trực tiếp, tế bào xác định đi vào con đường tạo phôi chỉ một thời
gian ngắn sau bước tái lập trình di truyền, không cần trải qua phân bào trước; ngược
lại, ở tạo phôi gián tiếp, khả năng sinh phôi (embryogenic competence) vẫn giữ ở
mức tương đối sau giai đoạn tạo mô sẹo [69].
Phương pháp tạo phôi vô tính trực tiếp có nhiều ưu điểm so với tạo phôi vô
tính gián tiếp:
Do không thông qua giai đoạn nuôi cấy tạo mô sẹo, hơn nữa phôi hình thành
trên bề mặt mô cấy do vậy hiện tượng tái lập trình (reprogramming) về mặt di truyền
ở mức tối thiểu, lại hình thành trong thời gian tương đối ngắn [68], nên phôi tái sinh
ít bị biến dị, do vậy có tiềm năng ứng dụng cao thể phôi này đặc biệt trong trong
nghiên cứu nhân giống, bảo quản lạnh phôi và biến nạp gen,... Tạo phôi trực tiếp đơn
giản hơn trong kỹ thuật nuôi cấy; ngược lại quá trình tái sinh phôi gián tiếp trải qua
các bước cảm ứng và tăng sinh mô sẹo tạo phôi, phải lập trình lại nên đòi hỏi dài về
mặt thời gian, vì vậy, cơ chế điều hòa và cảm ứng phức tạp hơn so với sự hình thành
phôi vô tính trực tiếp. Mức độ phụ thuộc vào chất ĐHST khác nhau giữa tạo phôi
trực tiếp và gián tiếp [70].
Có thể nhân sinh khối phôi thông qua cơ chế tạo phôi thứ cấp bằng phương
pháp nuôi lỏng lắc và phương pháp dùng bioreactor dùng cho nhiều mục đích khác
nhau đặc biệt cho nghiên cứu thu nhận hợp chất thứ cấp.
1.3.2. Một số nghiên cứu về sự phát sinh phôi vô tính
Nói chung, sự phát sinh phôi vô tính (ở các giai đoạn nêu trên) đều chịu ảnh
hưởng bởi một số yếu tố như kiểu gen thực vật, tuổi sinh lý và loại mẫu nuôi cấy,
thành phần môi trường khoáng, chất ĐHST, đường, cường độ ánh sáng,.. Ở
Arabidopsis, khá nhiều loại, kích thước mẫu với tuổi sinh lý khác nhau đã được sử
dụng để tạo phôi vô tính như phôi hợp tử trưởng thành và phôi non, chồi, nụ hoa. Ở
Sapindus mukorosi, lá 6 ngày tuổi là thích hợp cho cảm ứng tạo phôi. Ở Eucalyptus
camaldulensis, lá mầm 10 ngày tuổi cho tỷ lệ tạo mô sẹo sinh phôi cao nhất [70].
Ở nghiên cứu của Kharwanlang và cộng sự (2016a), kết quả cho thấy môi
trường khoáng MS có 5 mg/L 2,4-D thích hợp cho nuôi cấy tạo mô sẹo và mô sẹo có
khả năng sinh phôi (từ vật liệu LMTB thân rễ) Panax pseudoginseng so với môi
trường SH [71]. Trong 3 loại môi trường sử dụng (MS, B5, và SH) để nghiên cứu tạo

23
phôi vô tính Panax ginseng trực tiếp từ mảnh lá mầm phôi hợp tử, kết quả cho thấy
môi trường MS (tỷ lệ NH4+:NO3- là 21:39) là thích hợp nhất (MS có 50 g/L đường,
1% agar) [72].
Đường là yếu tố cung cấp năng lượng cho hoạt động sống của các thể nuôi cấy
mô trong đó có phôi vô tính, nồng độ sử dụng phù hợp với nhu cầu của từng loại mô
nuôi cấy, dao động từ 20 - 50 g/L. Kim và cộng sự (2012) đã sử dụng 20 - 30 g/L
đường cho nuôi cấy phôi thứ cấp Panax ginseng, tùy giai đoạn [73]. Nồng độ đường
50 g/L là thích hợp cho tạo phôi vô tính từ mô sẹo có nguồn gốc từ LMTB cắt ngang
thân rễ Panax vietnamensis [74].
Ánh sáng cũng là yếu tố được quan tâm nghiên cứu đặc biệt đối với thể phôi
vô tính đang trong quá trình chuyển giai đoạn phát triển hay trong quá trình nhân sinh
khối. Thông thường, sử dụng cường độ ánh sáng 4.000 lux để nuôi cấy. Nuôi cấy
phôi vô tính sơ cấp Chrysanthemum nhằm tạo phôi thứ cấp được thực hiện ở điều
kiện chiếu sáng cường độ cao ~ 4.500 - 7.500 lux (16 h sáng : 8 h tối) [75]. Sử dụng
cường độ ánh sáng ~ 3.000 lux đối với nuôi cấy tạo phôi thứ cấp Polyscias filicifolia
[5454]. Tuy nhiên, ở tạo phôi thứ cấp Quercus robur, nuôi cấy được thực hiện trong
điều kiện tối như nuôi cấy vật liệu phôi sơ cấp [76].
Nước dừa (10 - 20% - v/v) cũng được sử dụng trong nghiên cứu liên quan đến
phôi vô tính, vì chứa nhiều chất dinh dưỡng (đường, muối khoáng, acid amin) và chất
ĐHST tự nhiên. Khierallah và Hussein (2013) đã nghiên cứu ảnh hưởng của nước
dừa (% - v/v) với nhiều nồng độ khác nhau trên sự tạo mô sẹo và tái sinh phôi vô tính
Phoenix dactylifera từ nuôi cấy chồi đỉnh. Kết quả cho thấy tỷ lệ nước dừa 20% là
tốt nhất cho tạo mô sẹo (chỉ tiêu khối lượng tươi) và tái sinh phôi vô tính (chỉ tiêu số
phôi) [77]. Tương tự, Tô Thị Nhã Trầm và cộng sự (2020) cũng đã sử dụng nước dừa
trong tạo mô sẹo và tạo phôi đinh lăng lá xẻ nhỏ (Polyscias fruticosa). Kết quả cho
thấy mô sẹo cho tỷ lệ (%) phát sinh phôi vô tính tối ưu trên môi trường MS bổ sung
100 mL/L nước dừa kết hợp với 0,5 mg/L BA sau 6 tuần nuôi cấy. Phôi phát sinh từ
nuôi cấy mô sẹo mẫu lá trên môi trường MS có bổ sung 0,1 mg/L kinetin kết hợp với
150 mL/L nước dừa cho tỷ lệ tái sinh chồi (%) và số chồi/mẫu cao hơn các nghiệm
thức khác sau 8 tuần nuôi cấy. Ngoài ra, cây in vitro được nuôi cấy trên môi trường
có bổ sung 30 g/L đường và 100 mL/L nước dừa cho số rễ/cây cao nhất [78].

24
Chất ĐHST là yếu tố mang tính quyết định đến sự hình thành và phát triển
phôi vô tính tái sinh trực tiếp (từ mảnh lá, mảnh lá mầm,..) và gián tiếp (từ mô sẹo
hoặc cụm tế bào huyền phù có khả năng sinh phôi). Auxin (IAA, NAA, IBA, 2,4-
D,..) và cytokinin (BA, kinetin, TDZ), ngoài tác động ở dạng riêng lẻ, việc kết hợp
chúng cũng có tác động trong kích thích tạo phôi. Việc sử dụng 2,4-D là rất cần thiết
trong khởi tạo phôi vô tính và giúp phôi phát triển ở giai đoạn đầu [79]. Ví dụ sản
lượng cụm tế bào có khả năng sinh phôi chà là tăng 20 lần nhờ bổ sung 2,4-D nồng
độ thấp vào môi trường nuôi cấy [80]. Auxin làm thay đổi biến dưỡng nội sinh theo
cách tích cực, ví dụ ở cà rốt, sử dụng 2,4-D làm tăng khả năng đáp ứng sinh phôi qua
làm tăng hàm lượng IAA nội sinh; 2,4-D tạo điều hòa và cân bằng lượng auxin nội
sinh [81]. Tiền xử lý thực vật với auxin trước khi thực hiện cảm ứng tạo phôi ở C.
canephora cũng đã làm thay đổi biến dưỡng IAA nội sinh; và trong quá trình cảm
ứng tạo phôi C. canephora có sự gia tăng hàm lượng IAA nội sinh và sự biểu hiện
của gen mã hóa enzyme tryptophan aminotransferase (tryptophan aminotransferase
of Arabidopsis 1; CcTAA1) và enzyme flavin mono-oxygenase (YUCCA; CcYUC1 và
CcYUC3). Cả hai enzyme này tạo sinh tổng hợp IAA (Ayil-Gutiérrez và cộng sự,
2013). NAA (2 mg/L) thích hợp cho nuôi cấy tạo phôi trực tiếp từ mảnh lá sâm Ngọc
Linh (Panax vietnamensis) [38]. Chất ĐHST khác, như cytokinin cũng tham gia vào
sự phát triển của thực vật, kích thích sự hình thành nụ, làm chậm sự lão hóa lá và
cùng với auxin kích thích sự phân bào; cả hai auxin và cytokinin có tác động cộng
hưởng [82]. Tỷ lệ cao giữa cytokinin và auxin kích thích sự tạo chồi; ngược lại, tỷ lệ
này thấp cảm ứng tạo rễ và sự hình thành mô phân sinh hoàn chỉnh ở Pisum sativum.
Hai chất ĐHST này hoặc có tác động cộng hưởng hoặc đối kháng trong suốt quá trình
tạo phôi [83]. Ở C. canephora, sự di chuyển phân cực của IAA là cần thiết cho sự
hình thành trục ngọn-gốc [84]. Cytokinin cũng cần thiết để duy trì mức sinh tổng
hợp auxin cơ bản trong quá trình phát triển rễ và chồi, do vậy, có thể có một hệ thống
điều hòa thống nhất nhằm tạo hàm lượng thích hợp giữa auxin và sự phát triển của
thực vật [85].
Quá trình hình thành phôi vô tính có sự tích hợp của các tín hiệu nội sinh và
sự tái lập trình gen nhằm giải phóng tín hiệu khởi tạo quá trình phát sinh phôi. Các
auxin ngoại sinh, ở dạng riêng lẻ hoặc kết hợp với các chất ĐHST khác hoặc stress,
cảm ứng sự biểu hiện của các gen khác dẫn đến sự thay đổi chương trình di truyền

25
của tế bào soma và điều hòa quá trình dịch chuyển qua từng giai đoạn phát triển của
phôi [86]. Hầu hết các gen trên thuộc về một trong 4 nhóm: các yếu tố phiên mã
(transcription factors - TFs), các protein tác động ở chu kỳ tế bào, sinh tổng hợp chất
ĐHST, chủ yếu là auxin, cũng như các protein tham gia vào con đường tín hiệu [87].
Ở cà rốt (Daucus carota), Michalczuk và cộng sự (1992) đã nghiên cứu các
mức auxin nội sinh của tế bào ở các giai đoạn nuôi cấy tạo phôi khác nhau: (1) Nuôi
cấy tế bào huyền phù trong môi trường MS có 1 mg/L 2,4-D; (2) Sau đó chuyển sang
nuôi cấy trong môi trường không có 2,4-D nhằm tạo phôi. Kết quả cho thấy có sự
thay đổi con đường sinh tổng hợp IAA trong suốt quá trình phát sinh phôi vô tính. Ở
nuôi cấy huyền phù tế bào tăng sinh trong môi trường có 2,4-D, nhận thấy IAA nội
sinh hình thành đầu tiên từ cơ chất tryptophan, nhưng ở quá trình phát triển của phôi
vô tính, con đường sinh tổng hợp IAA không từ tryptophan lại có vai trò nổi trội hơn
đối với sinh tổng hợp IAA nội sinh. Như vậy, sự thay đổi về nồng độ auxin ngoại
sinh có ảnh hưởng đến sinh tổng hợp auxin nội sinh - dẫn đến tạo phôi vô tính [88].
Ribnicky và cộng sự (1996) đã nghiên cứu ảnh hưởng của auxin ngoại sinh ở
dạng tự do và phức hợp đến biến dưỡng auxin ở nuôi cấy đoạn trụ dưới lá mầm (1 -
2 cm, của cây 7 ngày tuổi từ hạt Daucus carota L. cv Danvers) trên môi trường MS
có hoặc không có auxin 2,4-D, [2H4]IAA, NAA (10 mM) và có 250 mM [2H5]L-
tryptophan. Kết quả cho thấy 2,4-D kích thích sự hình thành và tăng sinh mô sẹo, 2,4-
D tích lũy ở dạng tự do với lượng lớn và tác động không nhiều lên nồng độ IAA nội
sinh. NAA kích thích sự hình thành và tăng sinh mô sẹo dẫn đến mô ở trạng thái có
thể phát sinh cơ quan – chồi và rễ. NAA được phát hiện chủ yếu ở dạng phức hợp và
cũng có tác động không nhiều lên nồng độ IAA nội sinh. Như vậy, 2,4-D và NAA đã
có tác động trực tiếp lên mô cấy dẫn đến sự hình thành các loại mô tế bào khác nhau,
không do sự thay đổi hàm lượng IAA nội sinh. Không ghi nhận được ảnh hưởng của
[2H4]IAA lên mô cấy có thể do [2H4]IAA bị biến đổi nhanh chóng sang dạng bất
hoạt hoặc có thể [2H4]IAA làm giảm nồng độ IAA nội sinh theo cơ chế phản hồi. Sự
hiện diện của auxin ngoại sinh đã không gây ảnh hưởng đến tryptophan nội sinh hoặc
nguồn (pool) IAA. Như vậy, auxin ngoại sinh không làm thay đổi con đường sinh
tổng hợp IAA nhưng cần thiết cho cảm ứng sự hình thành và tăng sinh mô sẹo cũng
như cảm ứng hình thành tế bào có khả năng sinh phôi [89].

26
Ayil-Gutiérrez và cộng sự (2013) cho rằng hầu hết quá trình phát sinh phôi vô
tính đòi hỏi sự hiện diện của ít nhất của một loại auxin ngoại sinh, hoặc hiện diện
trước đó hoặc hiện diện trong suốt quá trình phát sinh phôi; ngoài ra, còn kể đến vai
trò của auxin nội sinh. Auxin ngoại sinh cảm ứng sự hình thành auxin nội sinh – cũng
cần thiết cho sự phát sinh phôi. Các tác giả trên đã nghiên cứu động học của nồng độ
IAA và một số phức hợp của IAA (IAA-Ala, IAA-Glu) trong suốt thời kỳ cảm ứng
phát sinh phôi vô tính ở cà phê (Coffea canephora) qua nuôi cấy các mảnh lá (0,25
cm2, dùng môi trường lỏng lắc có 5 µM BA) của cây có nguồn gốc phôi vô tính (40
tuần tuổi, có 6 cặp lá, được tiền nuôi cấy trong môi trường MS có 0,54 µM NAA và
2,32 µM kinetin trong 14 ngày). Trong suốt thời gian tiền nuôi cấy cây trong môi
trường nuôi cấy có auxin NAA nói trên, nhận thấy có sự gia tăng mạnh của cả IAA ở
dạng tự do, IAA ở dạng phức hợp và IBA. Sự gia tăng này đi kèm với sự gia tăng
biểu hiện của các gen có liên quan. Sự cân bằng giữa IAA tự do và IAA ở dạng phức
hợp là rất cần thiết cho sự biểu hiện của các gen liên quan đến quá trình tạo phôi. Như
vậy, ở nghiên cứu này, quá trình phát sinh phôi vô tính bao gồm 2 pha cơ bản: Pha 1
cần có auxin NAA và kinetin (2 tuần), pha 2 chỉ cần có sự hiện diện của BA [90].
Nghiên cứu của Verma và cộng sự (2018) đã cho thấy vai trò của auxin và sự
di chuyển phân cực của auxin đã ảnh hưởng đến sự tạo phôi Digitalis trojana qua
nuôi cấy các đoạn trụ dưới lá mầm (5 mm) phôi hợp tử trên môi trường MS có IAA
(0,1 - 1 mg/l), TIBA (chất ức chế sự di chuyển auxin) (0,1 - 1 mg/l), IAA và TIBA.
Kết quả cho thấy ở nghiệm thức chỉ có IAA (0,5 mg/L), tỷ lệ tạo phôi cao nhất -
52%, số phôi 10 phôi/mẫu; ngược lại, ở NT có 0,5 mg/L IAA, 1 mg/L TIBA, tỷ lệ
tạo phôi chỉ đạt 24%; 3,6 phôi/mẫu, và có nhiều phôi bất thường. TIBA đã gây đáp
ứng chậm đối với tạo phôi và tái sinh nhiều phôi bất thường. Như vậy, auxin và sự di
chuyển phân cực của auxin có vai trò quan trọng đối với tạo phôi vô tính ở Digitalis
trojana [14].
Gaoyin và cộng sự (2020) đã nghiên cứu sự thay đổi các đặc tính hóa sinh
trong quá trình phát sinh phôi vô tính ở Ormosia henryi Prain qua nuôi cấy phôi hợp
tử trưởng thành trên các môi trường theo thứ tự: (1) Môi trường tạo mô sẹo: B5 có 2
mg/L 2,4-D (auxin mạnh), 0,2 mg/L BA, 30 g/L đường, 500 mg/L glutamine, thời
gian nuôi 6 tuần; (2) Môi trường tăng sinh mô sẹo đồng thời thu nhận mô sẹo có khả
năng sinh phôi: B5 có 1 mg/L 2,4-D (giảm nồng độ), 0,5 mg/L kinetin, thời gian nuôi

27
4 tuần; (3) Môi trường nuôi mô sẹo có khả năng sinh phôi nhằm tạo phôi cầu: như
trên, nuôi 2 tuần trong tối ở 25 ± 2oC; (4) Môi trường nuôi phôi cầu tạo phôi có lá
mầm: B5 có 0,2 mg/L NAA (auxin yếu), 0,5 mg/L thidiazuron, thời gian nuôi 60
ngày. Kết quả nghiên cứu cho thấy: (1) Các hợp chất cung cấp năng lượng: Hàm
lượng đường hòa tan và tinh bột giảm dần trong suốt quá trình sinh phôi do nhu cầu
tiêu thụ lượng lớn các hợp chất này; ngược lại, hàm lượng protein lại tăng lên. Ở mô
sẹo có khả năng sinh phôi, sự tích lũy đường hòa tan, tinh bột và protein cao – cơ sở
quan trọng về năng lượng cho quá trình tạo phôi so với mô sẹo không có khả năng
sinh phôi; (2) chất ĐHST nội sinh: Ở các giai đoạn phát triển khác nhau của phôi,
hàm lượng IAA tăng có ý nghĩa ở giai đoạn phôi có lá mầm – có liên quan đến sự
hình thành cực phôi. Tương tự, hàm lượng cytokinin cũng tăng ở phôi giai đoạn này
– có liên quan đến sự trưởng thành và nẩy mầm của phôi. IAA cũng cao ở mô sẹo có
khả năng sinh phôi so với mô sẹo không có khả năng sinh phôi; (3) Về tỷ lệ giữa các
chất ĐHST nội sinh: Tương tác giữa các chất ĐHST nội sinh ở các giai đoạn phát
triển khác nhau của quá trình phát sinh phôi vô tính có thể được thể hiện qua tỷ lệ
giữa chúng và được xem như chỉ số sinh lý để tìm hiểu sự phản biệt hóa và tái biệt
hóa của tế bào. Tỷ lệ IAA/ABA, IAA/GAs, AUX/GAs và AUX/ABA giảm dần, tỷ lệ
IAA/CKs, AUX/CKs, ABA/CKs and GAs/CKs lúc đầu tăng và sau đó giảm. Tỷ lệ
IAA/CKs và AUX/CKs thấp nhất ở phôi có lá mầm [12].
Huyền phù tế bào đơn của dòng Medicago falcata 47/1/150 có khả năng sinh
phôi cao đã được thực hiện. Huyền phù tế bào này bao gồm nhiều loại tế bào đơn
khác nhau. Hầu hết các tế bào đều có khả năng tạo phôi sau quá trình phân chia không
đối xứng. Nhằm khảo sát rõ quá trình phân chia này, huyền phù tế bào mang gen gusA
(mã hóa enzyme β-glucoronidase) cũng đã được thực hiện và nuôi cấy theo hướng
phân bào tạo phôi trực tiếp. Từ đó, các bước phát triển của phôi đã được khảo sát chi
tiết bắt đầu từ sự phân chia không đối xứng – tín hiệu đầu tiên của quá trình hình
thành phôi vô tính [67].
Quy trình tạo phôi trực tiếp, tạo phôi thứ cấp và tái sinh cây từ phôi ở
Dendrobium cv. Chiengmai Pink đã được thực hiện. Kết quả cho thấy phôi tái sinh
tốt nhất trên môi trường có bổ sung 1 mg/L TDZ. Phôi tái sinh trực tiếp nhiều ở vị trí
gần vết cắt mẫu và ít hơn ở vị trí đầu lá. Quá trình cấy chuyền cho thấy có hiện tượng

28
tạo phôi thứ cấp trực tiếp từ phôi sơ cấp. Khảo sát hình thái giải phẫu cho thấy phôi
thứ cấp hình thành từ lớp tế bào bên ngoài của phôi sơ cấp [91].
Gow và cộng sự (2009) đã ghi nhận được phôi vô tính Phalaenopsis amabilis
Shimadzu var. formosa và Phalaenopsis ‘Nebula’ tái sinh tái sinh trực tiếp từ mảnh
lá (1-2 cm, cấy úp) trên môi trường ½MS có bổ sung 3 mg/L TDZ hoặc 3 mg/L BA.
Qua quan sát dưới kính hiển vi điện tử quét, nhận thấy phôi vô tính với nhiều dạng
khác nhau hình thành trực tiếp từ lớp tế bào bề mặt của mảnh lá ở cả 2 giống [92].
Phôi Curcuma amada Roxb. hình thành trực tiếp qua nuôi cấy 2 giai đoạn (1)
Nuôi cấy 2 tuần trên môi trường có bổ sung 2,24 μM và 1,11 μM BA, sau đó (2) Nuôi
cấy tiếp tục trên môi trường có 9,10 μM TDZ và 1,33 μM NAA. Qua khảo sát hóa
mô tế bào và quan sát dưới kính hiển vi điện tử, kết quả cho thấy phôi có nguồn gốc
từ quá trình phân chia tế bào ở biểu mô và tế bào dưới biểu mô [93].
Cũng qua khảo sát mô tế bào, Wang và cộng sự (2014) cho rằng ngoài tái sinh
phôi theo cách thông qua mô sẹo, các tiền phôi đa bào cũng được hình thành trực tiếp
từ lớp tế bào bề mặt của phôi non Tapiscia sinensis qua nuôi cấy trên môi trường MS
có 1 mg/L 2,4-D và 0,5% than hoạt tính [94].
Các mẫu cấy lá sâm Ngọc Linh có kích thước 0,5 x 0,5 cm được nuôi cấy trên
môi trường MS có 1 mg/L 2,4-D và 1 mg/L NAA để cảm ứng tạo mô sẹo. Các mẫu
mô sẹo này được cắt thành mảnh nhỏ có đường kính 0,5 cm và nuôi cấy trên môi
trường MS bổ sung 1 mg/L 2,4-D kết hợp với NAA và/hoặc kinetin ở các nồng độ
khác nhau để cảm ứng quá trình phát sinh phôi vô tính. Sau 8 tuần nuôi cấy, các cấu
trúc hình cầu bắt đầu được ghi nhận. Tần suất phát sinh phôi đạt được cao nhất trên
môi trường MS có bổ sung 1 mg/L 2,4-D, 0,2 mg/L kinetin và 0,5 mg/L NAA. Sau
12 tuần nuôi cấy, tất cả các giai đoạn phát triển của phôi vô tính, bao gồm phôi hình
cầu, hình tim, hình thủy lôi và phôi giai đoạn lá mầm đều được phát hiện trong các
cụm phôi. Các quan sát hình thái giải phẫu và phân tích dưới kính hiển vi điện tử quét
cho thấy phôi có cấu trúc lưỡng cực với cực chồi và cực rễ phát triển rõ ràng [95].
Trong phát sinh phôi vô tính từ mô sẹo Tam thất hoang (Panax stipuleanatus)
sau 24 tuần nuôi cấy trên môi trường có 2,4-D. Mô sẹo có các cụm tế bào đẳng kính
rời rạc được chuyển sang môi trường có bổ sung 0,5 mg/L NAA để thu nhận tế bào
có khả năng sinh phôi. Mô sẹo phát triển trên môi trường này trở nên nhão hơn với
các cụm tế bào có đặc tính của tế bào sinh phôi: kích thước nhỏ, đẳng kính, nhân to,

29
thấy rõ hạch nhân và tế bào chất đậm đặc. Sự hình thành những cụm gồm các cấu
trúc hình cầu, kích thước đồng đều xảy ra tại thời điểm 28 tuần trên môi trường có bổ
sung NAA. Các cấu trúc giống phôi này trải qua các giai đoạn phát triển: phôi hình
cầu muộn, hình tim và tử diệp trên môi trường có bổ sung 0,5 mg/L BA, 1 mg/L GA3.
Bên cạnh đó, có sự xuất hiện của phôi bất thường chỉ chỉ tạo rễ, tạo chồi hoặc lá [43].
Mảnh lá (~ 10 mm) cây in vitro Tolumnia Louise Elmore ‘Elsa’ được nuôi cấy
trên môi trường ½MS có bổ sung 5 loại cytokinin khác nhau (2-iP, BA, kinetin, TDZ
và zeatin) với các nồng độ 0,3; 1; 3 mg/L và nuôi trong tối. Kết quả cho thấy, sau 90
ngày nuôi, nồng độ 3 mg/L TDZ cho kết quả tái sinh trực tiếp phôi cầu tốt nhất. Khảo
sát hình thái giải phẫu cho thấy phôi có nguồn gốc từ các tế bào mô phân sinh (kích
thước nhỏ, tế bào chất đậm đặc) có nguồn gốc từ lớp tế bào biểu mô và dưới biểu mô
(tế bào diệp lục) ở mảnh lá [96].
Liang và cộng sự (2020) đã nghiên cứu tạo mô sẹo và tái sinh chồi bất định và
phôi vô tính Scaevola sericea qua nuôi cấy mảnh lá (1 x 1 cm, từ cây in vitro) và đoạn
rễ (dài 0,5 cm, bao gồm đầu rễ, cũng từ cây in vitro) trên môi trường MS có 0,5 - 2,5
mg/L NAA, MS có 2,5 mg/L NAA, theo thứ tự. Đã xác định được môi trường tối ưu
cho tái sinh gián tiếp chồi bất định và phôi vô tính từ mô sẹo lá, rễ là MS có bổ sung
2,5 mg/L BA, MS có 2,5 mg/L TDZ, theo thứ tự. Phôi vô tính tái sinh có nguồn gốc
từ nhu mô nằm bên dưới lớp tế bào biểu mô của lá [97].
1.4. Sự hình thành rễ bất định
1.4.1. Cơ sở khoa học của sự hình thành rễ bất định
Rễ bất định là rễ được phát sinh từ những vùng khác nhau trên cơ thể thực vật
như thân, cành, lá; nhưng không phát sinh từ rễ.
Tương tự trường hợp tạo phôi vô tính, tế bào với đặc điểm có tính toàn thế có
thể tái sinh rễ bất định theo cách trực tiếp và gián tiếp. Rễ có thể được tái sinh trực
tiếp/gián tiếp từ một số loại mô nuôi cấy ban đầu khác nhau như mảnh lá/lá mầm,
đoạn cuống lá, đoạn rễ, LMTB thân rễ,..
Quá trình tạo rễ bất định trực tiếp và gián tiếp bao gồm 3 giai đoạn: (1) Cảm
ứng; (2) Khởi tạo sơ khởi rễ từ tế bào liên kết với mô mạch hoặc thuộc bó mạch hoặc
từ tế bào nhu mô khuyết (đối với vật liệu nuôi cấy là mô lá), hoặc từ vòng tế bào
tượng tầng bên ngoài bó mạch/trụ bì (đối với vật liệu nuôi cấy là mô thân/rễ); (3)
Biểu hiện/hình thành rễ. Mỗi giai đoạn có sự tương tác giữa các yếu tố như tín hiệu

30
bên ngoài, sự thay đổi hàm lượng các chất ĐHST nội sinh, và sự biểu hện của một số
gen. Trong những năm gần đây, nhiều nghiên cứu đã được thực hiện nhằm xác định
cơ chế phân tử của sự tạo rễ bất định, có nhiều gen liên quan trực tiếp đến cảm ứng
và khởi tạo rễ bất định đã được xác định ở nhiều loài thực vật khác nhau như
Arabidopsis thaliana, Solanum lycopersicon, Medicago sativa,.. Auxin được biết như
yếu tố trung tâm điều hòa các giai đoạn của quá trình tạo rễ bất định như đã nêu trên;
có 3 gen tín hiệu auxin (auxin signaling) tham gia vào quá trình này bao gồm gen
sinh tổng hợp auxin và điều hòa cân bằng nội sinh (homeostasis), gen vận chuyển
auxin và gen đáp ứng với auxin, trong đó gen sinh tổng hợp auxin và điều hòa cân
bằng nội sinh có tác động trực tiếp, ví dụ như gen YUCCA mã hóa flavin
monooxygenase – tham gia trong sinh tổng hợp IAA (từ tryptophan) nhằm duy trì
mức IAA ở mô phân sinh đầu rễ [98].
Yu và cộng sự (2017) đã thực hiện nuôi cấy mảnh lá Arabidopsis thaliana trên
môi trường B5 không có auxin nhằm tạo rễ bất định trực tiếp và nuôi cấy mảnh lá
trên môi trường có auxin nồng độ cao để tạo mô sẹo và sau đó cấy chuyển mô sẹo
sang môi trường có nồng độ auxin thấp/không có auxin để tái sinh rễ (gián tiếp). Từ
kết quả thu được, các tác giả trên cho rằng ở tái sinh rễ trực tiếp có 4 hiện tượng sinh
lý tuần tự xảy ra: (1) Bước ‘tạo mồi’: auxin nội sinh được vận chuyển đến các tế bào
có khả năng tái sinh (procambium và tế bào nhu mô mạch), kích chuyển các tế bào
này thành các tế bào ‘nguồn’; (2) Bước ‘khởi tạo’: các tế bào ‘nguồn’ phát triển thành
sơ khởi rễ - nơi tích tụ auxin cao; (3) Bước ‘tạo hình’: các tế bào sơ khởi rễ tiếp tục
phân chia tạo mô phân sinh đầu rễ - nơi auxin giảm tích tụ; (4) Bước ‘hiện hình’: đầu
rễ đạt mức trưởng thành, tiếp tục phát triển ra khỏi mảnh lá. Ở trường hợp tái sinh
gián tiếp, cũng có các bước tương tự như trên: (1) Bước ‘tạo mồi’ tạo tế bào ‘nguồn’;
(2) Bước ‘khởi tạo’: tạo mô sẹo thay vì sơ khởi rễ; (3) Bước tạo rễ khi mô sẹo được
cấy chuyển sang môi trường có nồng độ auxin giảm/không có auxin [99]
Tương tự trường hợp phôi vô tính, tạo rễ bất định cũng là một quá trình phức
tạp, chịu ảnh hưởng bởi một số yếu tố cơ bản như vai trò sinh lý của auxin (loại và
nồng độ sử dụng), sự di chuyển phân cực của auxin, sự tích tụ auxin ở vị trí nhất
định,… và ở mức độ phân tử có sự biểu hiện của các gen có liên quan. Như vậy, tạo
rễ bất định là đặc tính di truyền số lượng, được điều hòa bởi cả các yếu tố môi trường
bên ngoài và yếu tố nội sinh. Auxin đóng vai trò quyết định trong điều hòa phát triển

31
rễ, quá trình này có liên quan đến các gen cảm ứng bởi auxin và con đường tín hiệu
auxin.
1.4.2. Một số nghiên cứu về sự hình thành rễ bất định
Một số yếu tố chính ảnh hưởng đến các giai đoạn hình thành rễ bất định bao
gồm chất ĐHST, thành phần môi trường khoáng, nguồn carbon, mật độ mô/tế bào
nuôi cấy, độ pH,.. Các loại môi trường nuôi cấy khác nhau có tác dụng khác nhau đối
với cảm ứng phân nhánh và gia tăng sinh khối rễ. MS, ½MS là môi trường thích hợp
trên nhiều đối tượng cây trồng như Boesenbergia rotunda, Vernonia amygdalina,
Camellia sinensis và Echinacea purpurea.
Nguồn carbon cần thiết cho sự hình thành rễ cũng khác nhau do khả năng hấp
thụ của từng loài thực vật và quá trình chuyển hóa nguồn carbon có khác nhau. Nồng
độ đường 50 g/L là rất thích hợp cho Boesenbergia rotunda và Eurycoma longifolia,
trong khi nồng độ đường 30 g/L thích hợp đối với Camellia sinensis... Ngoài ra, nồng
độ đường thích hợp cũng ảnh hưởng đến việc tăng sản xuất các hợp chất thứ cấp.
Nhằm tối ưu hóa các điều kiện nuôi cấy rễ Gynura procumbens, các môi trường MS
(½ - 2), nồng độ đường (1 – 10%), độ pH (5 - 6,5), và điều kiện tối/sáng đã được
nghiên cứu. Kết quả cho thấy đường 2% là thích hợp nhất cho sự tăng trưởng rễ. Dựa
trên nghiên cứu của Cui và cộng sự (2010), nồng độ cao của đường (5%, 7%, 9%) đã
ảnh hưởng đến sinh khối do ảnh hưởng của áp suất thẩm thấu - là rất cao. Hơn nữa,
trong nghiên cứu nuôi rễ Psammosilene tunicoide với các điều kiện nuôi cấy như
nồng độ đường (0 - 50 g/L); các chu kỳ sáng (0 - 12 h/ngày); pH ban đầu (5 - 6,5); và
cường độ ánh sáng 2.000 lux, kết quả cho thấy các yếu tố ảnh hưởng thích hợp là 30
g/L đường, chu kỳ sáng 10 h/ngày, và pH ban đầu là 5,8 [100].
Ahkami và cộng sự (2013) đã nghiên cứu vai trò của sự di chuyển phân cực
của auxin đối với sự tích tụ auxin và sự hình thành rễ bất định ở đoạn thân mang chồi
của Petunia hybrida. Sau cắt và trồng đoạn thân trong khay nhựa, nhận thấy IAA tăng
lên ở gốc đoạn thân và đạt đỉnh cao ở thời điểm 2 h và 24 h, giảm mạnh ở ở 48 h và
giảm tối đa ở thời điểm 192 h. Xử lý Naphthylphthalamic acid (NPA) (chất ức chế
vận chuyển auxin) đã gây ức chế tạo đỉnh IAA ở 24 h dẫn đến ức chế hoàn toàn sự
tạo rễ. Các tác giả trên kết luận sự phát sinh rễ bất định ở Petunia phụ thuộc vào (1)
Sự vận chuyển phân cực của auxin, và (2) Đỉnh IAA ở thời điểm 24 h ở vùng tạo rễ
- tạo bể và cảm ứng sự phân chia đầu tiên của tế bào mô phân sinh rễ. IAA giảm sau

32
thời điểm 24 h tạo thuận lợi cho quá trình phát triển tiếp theo của rễ. Sự phát triển
tiếp theo của rễ kích thích sự đường phân và hoạt động của con đường pentose
phosphate – giúp tạo năng lượng, khung carbon dùng sinh tổng hợp acid amin, protein
và acid nucleic ở các tế bào trong quá trình tăng sinh, biệt hóa và tăng trưởng [101].
Guan và cộng sự (2019) đã nghiên cứu vai trò điều hòa của auxin đối với quá
trình tạo rễ bất định từ đoạn thân mang chồi (~ 4 cm) dòng cà chua Alisa Craig (AC)
và dòng chuyển gen DR5pro:YFP trans nuôi trong dung dịch dinh dưỡng Hoagland.
Kết quả cho thấy rễ bất định bắt nguồn từ tế bào nguồn ở lớp tế bào trụ bì của đoạn
thân. Sau đó rễ bất định kéo dài sau quá trình chết của tế bào biểu bì. Mức auxin và
ethylene tăng lên ở phần gốc đoạn thân trong vòng 1 h, có sự phân bố auxin gia tăng
ở pha khởi tạo rễ, tập trung ở mô phân sinh đang phát triển; ngược lại, zeatin, acid
salicylic và acid abscisic chỉ đóng vai trò nhỏ trong quá trình tạo rễ. Các tác gỉả trên
còn ghi nhận sự biểu hiện của gen vận chuyển auxin đặc hiệu tăng lên trong suốt pha
phát triển của rễ [102].
Guan và cộng sự (2020) đã nghiên cứu sự di chuyển phân cực của auxin thông
qua sự tạo rễ bất định từ đoạn thân của giống táo lùn (Malus domestica) M.9. Các tác
giả cho rằng sự hình thành rễ bất định bắt đầu bằng sự phân chia và kéo dài của các
tế bào nguồn – là tế bào tượng tầng đã phản biệt hóa, ở vị trí gần kề hai bó mạch, quá
trình này xảy ra ở điều kiện có mức tương đối cao của IAA trong mô và có sự phân
giải các hạt tinh bột dùng cung cấp năng lượng. Auxin ngoại sinh đã kích thích sự
phân chia tế bào, sự tăng sinh và tái cấu trúc hệ võng nội chất và thể golgi. Ngược
lại, ở nghiệm thức xử lý N-1-naphthylphthalamic acid (NPA) - tác nhân gây ức chế
sự di chuyển auxin, nhận thấy có hiện tượng ức chế sự phân chia tế bào ở vùng gốc
của đoạn thân dẫn đến sự phân bào bất thường ở suốt giai đoạn sớm của quá trình
hình thành rễ [13].
Auxin là chất ĐHST có tính quyết định đối với nuôi cấy rễ bất định, một số
auxin thường được sử dụng là IAA, IBA, NAA, kể cả 2,4-D. Hiệu quả đến tăng
trưởng rễ khá phụ thuộc vào loài thực vật. Dựa trên các báo cáo khác nhau cho thấy
IBA là auxin thích hợp để cảm ứng và phát triển nuôi cấy rễ của một số cây thuốc/cây
trồng như Centella asiatica, Costus igneus, Couroupita guianensis Aubl., Labisia
pumila, Psoralea coryfolia, Silyabum marianum. Nồng độ IBA 7 m/L; 0,5 mg/L là
nồng độ tối ưu cho sự hình thành rễ bất định so với NAA/IAA ở Centella asiatica,

33
Costus igneus, theo thứ tự. Bên cạnh IBA, NAA cũng có ảnh hưởng mạnh đến cảm
ứng và gia tăng sinh khối rễ. NAA là auxin tốt nhất đối với nuôi cấy rễ nhiều loài như
Andrographis paniculata, Boesenbergia rotunda, Eurycoma longifolia, Fagonia
indica, Mondia whitei, và Rumex crispus. Trong nghiên cứu nuôi cấy rễ cây thuốc
Andrographis paniculata, NAA ở nồng độ 2,7 µM tạo sinh khối rễ khô/tươi và hàm
lượng andrographolide cao sau 4 tuần nuôi cấy so với IAA và IBA. Đối với Rumex
crispus, 5 µM NAA là nồng độ rất thích hợp để tạo và tăng sinh rễ. Trong khi đó,
nồng độ cao hơn, 10 µM NAA - tối ưu cho nuôi cấy rễ của Mondia whitei. So với
IBA và NAA, IAA có hiệu quả thấp trong thúc đẩy phát triển rễ cây thuốc, trừ ở
trường hợp Orthosiphon stamineus (nồng độ 3 mg/L là tốt nhất để thúc đẩy sự ra rễ
từ mẫu cấy lá). Ngoài ra, một số nghiên cứu cũng cho rằng IBA và NAA có thể được
kết hợp với nhau để tạo ra các điều kiện tăng trưởng tối ưu cho nuôi cấy rễ cây thuốc.
Ví dụ, sự kết hợp 1 mg/L IBA và 1 mg/L NAA rất thích hợp cho rễ Luffa acutangula
(L.) Roxb. và sự kết hợp của 0,05 mg/L IBA và 0,1 mg/L NAA là điều kiện tối ưu
cho rễ Psammosilene tunicoides [98]. Ở họ Ngũ gia bì, cũng nhận thấy có sự đa dạng
về loại môi trường, loại và nồng độ chất ĐHST,.. sử dụng trong nghiên cứu tạo rễ. Ví
dụ rễ bất định Panax notoginseng tái sinh tốt từ mô cuống lá, đoạn rễ nhánh nuôi cấy
trên môi trường MS có 3 mg/L IBA, có hoặc không bổ sung kinetin; nuôi lỏng lắc rễ
bất định được thực hiện trong môi trường ½MS có 3 mg/L IBA [44]. Đối với tạo rễ
bất định thứ cấp từ rễ bất định sơ cấp Panax vietnamensis, rễ thứ cấp phát triển tốt
nhất trên môi trường MS cải biên (½NH4+/NO3
-) có IBA và BA, 30 g/L đường, ở
22oC và pH 5,3 [47]. Li và cộng sự (2004) đã tạo rễ cho các chồi (có nguồn gốc từ
đốt thân) Schefflera octophylla qua sử dụng môi trường ½MS có bổ sung chất ĐHST
thích hợp là NAA (0,2 mg/L). Môi trường MS có bổ sung 0,5 mg/L NAA sử dụng để
tái sinh rễ bất định Panax stipuleanatus từ mô sẹo có nguồn gốc từ thân rễ [43].
Lo (1997) cũng nghiên cứu sự tái sinh rễ bất định Saintpaulia ionantha
(African violet) đồng thời với nghiên cứu tái sinh chồi từ mảnh lá theo cơ chế phát
sinh cơ quan dưới ảnh hưởng của một số yếu tố như tuổi sinh lý, tạo vết thương, cách
đặt mẫu cấy vào môi trường và sự trao đổi khí. Kết quả thu được cho thấy rễ bất định
có nguồn gốc từ tế bào nhu mô diệp lục khuyết [103].
Mảnh lá Medicago truncatula được sử dụng để nghiên cứu sự cảm ứng tạo rễ
bởi auxin từ nuôi cấy mảnh lá trên môi trường P4 có 10 µM NAA. Rễ tái sinh sau

34
giai đoạn mô sẹo chỉ mới bắt đầu hình thành. Khảo sát mô tế bào cho thấy, sau nuôi
cấy, có các lớp tế bào khác biệt hình thành từ bó mạch của mảnh lá và từ đây sơ khởi
rễ và tế bào mô mạch mới hình thành. Như vậy, các tế bào có nguồn gốc mô mạch
(VDC) lá tương tự như tế bào tiền tượng tầng và có chức năng của tế bào gốc, qua
đáp ứng với auxin ngoại sinh sẽ có xu hướng tạo tế bào mô phân sinh sơ khởi rễ và
mô mạch của rễ mới hình thành (kết quả đã được chứng minh qua sử dụng đột biến
sickle ‘skl’ – bị hư hại ở khả năng truyền tín hiệu ethylene trong khi dòng hoang dại
và dòng đột biến ‘sunn’ - khiếm khuyết ở chức năng di chuyển phân cực của auxin).
VDC có thể có nguồn gốc từ mô libe hoặc gần mô libe [104].
Qua các thông tin cho thấy, phương pháp nuôi cấy mô tế bào đã và đang có
nhiều đóng góp trong lĩnh vực nghiên cứu thực vật ở khía cạnh sinh lý. Cơ sở khoa
học của phương pháp này là tính toàn thế của tế bào. Qua nuôi cấy trong điều kiện
thích hợp, tế bào có thể phát triển qua nhiều bước tiếp theo thành các cấu trúc mới
như phôi vô tính/chồi/rễ và thành cây hoàn chỉnh theo hình thức tái sinh trực tiếp
hoặc gián tiếp. Có một số yếu tố ảnh hưởng đến quá trình nuôi cấy tạo các sản phẩm
mục tiêu như phôi/chồi/rễ như vai trò sinh lý của chất ĐHST, tuổi sinh lý mẫu cấy,
điều kiện nuôi cấy, kiểu gene thực vật,… Theo nhiều tài liệu, trong các nhóm chất
ĐHST, auxin là tác nhân đóng vai trò quan trọng của quá trình tái lập trình để tế bào
sinh dưỡng (ở trạng thái biệt hóa hoặc phản biệt hóa) chuyển sang trạng thái mang
tính toàn thế (founder cell) - hướng đích tạo phôi vô tính hoặc rễ bất định. Phương
pháp khảo sát hình thái/hình thái giải phẫu mô tế bào với đối tượng nghiên cứu là các
loại mô/tế bào, hình thức tổ chức và phát triển của chúng cũng đã góp phần quan
trọng trong tìm hiểu về tính toàn thế của tế bào.
Tóm lại, các thông tin tổng quan nêu trên đã khái quát tầm quan trọng của một
số loài thực vật quan trọng thuộc họ Ngũ gia bì nói chung và NGBCC nói riêng, trong
lĩnh vực nghiên cứu khoa học và thực tiễn cuộc sống. Như đã trình bày, điểm tồn
tại/hạn chế hiện nay là các nghiên cứu liên quan đến xây dựng hai hệ thống nuôi cấy
là phôi vô tính và rễ bất định - vốn có rất nhiều ưu điểm còn rất hạn chế trên đối tượng
thực vật này. Do vậy, đề tài này với nội dung triển khai thực nghiệm nghiên cứu tạo
và nhân sinh khối phôi vô tính và rễ bất định hy vọng sẽ góp phần phát triển hướng
nghiên cứu quan trọng này, đồng thời cung cấp một số dẫn liệu khoa học mới về hai
hệ thống nêu trên.

35
CHƯƠNG 2. VẬT LIỆU VÀ PHƯƠNG PHÁP
2.1. Nguồn mẫu – Vật liệu nuôi cấy
2.1.1. Nguồn mẫu
Cây NGBCC khoảng 8 năm tuổi được thu thập từ nhà vườn, trồng trong chậu
(Hình 2.1), được lưu trữ ở nhà lưới Phòng thí nghiệm trọng điểm phía nam về Công
nghệ tế bào thực vật (Viện Sinh học nhiệt đới) để tạo vật liệu nuôi cấy cho các thí
nghiệm.
Hình 2.1. Cây Ngũ gia bì chân chim (Schefflera octophylla)
2.1.2. Tạo vật liệu nuôi cấy ban đầu
Sử dụng tất cả các lá chét (lá đơn) (Hình 2.2A) được cắt ra từ lá kép thứ 2 (tính
từ ngọn) cây NGBCC khoảng 8 năm tuổi (Hình 2.1). Các lá chét được rửa bằng nước
xà phòng, rửa lại dưới vòi nước chảy trong 10 phút, khử trùng tiếp bằng cồn 70%
trong 10 phút, bằng nước Javel (NaOCl, hàm lượng chlor hoạt chất 38 g/L) 30% (v/v)
trong 20 phút, rửa lại bằng nước cất vô trùng 3 lần. Sau khử trùng, cắt phiến lá chét
thành các mảnh lá kích thước 10 x 10 mm (mảnh lá I), 3 x 10 mm (mảnh lá II) làm
vật liệu nuôi cấy (Hình 2.2B,C,D).
Hình 2.2. Lá kép Ngũ gia bì chân chim và vật liệu nuôi cấy
A. Lá kép, LC: Lá chét, B. Cách cắt mẫu; Mảnh lá 10 x 10 mm (C), 3 x 10 mm (D)

36
2.2. Nội dung nghiên cứu
2.2.1. Nội dung 1. Tạo phôi vô tính
Nghiên cứu sự cảm ứng phát sinh phôi vô tính trực tiếp từ mảnh lá thông qua
khảo sát ảnh hưởng bởi loại và nồng độ chất ĐHST auxin (NAA/IBA), nồng độ của
cytokinin (BA), loại môi trường khoáng (½MS, MS, B5, SH), nồng độ sucrose, tỷ lệ
% nước dừa, điều kiện chiếu sáng. Tạo cây con hoàn chỉnh từ phôi vô tính, trồng cây
con ra chậu đất ở vườn ươm. Tạo phôi vô tính gián tiếp qua mô sẹo mảnh lá nuôi cấy
ở môi trường đặc và lỏng, mô sẹo hình thành từ mảnh lá ở môi trường SH có bổ sung
2,4-D.
Quan sát, theo dõi các giai đoạn phát sinh phôi: phôi cầu, phôi trái tim, phôi
thủy lôi, phôi trưởng thành có dạng hai lá mầm. Quan sát cấu trúc giải phẫu phôi;
hình thái cụm tế bào/cụm mô trong quá trình nuôi cấy lỏng.
2.2.2. Nội dung 2. Nhân phôi vô tính
Sử dụng nguồn phôi vô tính được hình thành từ thí nghiệm tạo phôi vô tính
làm vật liệu cho nuôi cấy nhân phôi. Nhân phôi ở môi trường lỏng: khảo sát chất
ĐHST, khối lượng và kích thước phôi nuôi cấy, nồng độ đường, tỷ lệ % nước dừa,
cường độ ánh sáng ảnh hưởng đến tạo phôi thứ cấp.
Giải phẫu phôi thứ cấp và xác định nguồn gốc hình thành của phôi thứ cấp.
2.2.3. Nội dung 3. Tạo rễ bất định
Khảo sát các yếu tố như: loại và nồng độ auxin thích hợp (NAA/IBA), loại
môi trường khoáng, nồng độ đường, điều kiện chiếu sáng ảnh hưởng đến cảm ứng
tạo rễ bất định trực tiếp từ mô lá NGBCC. Tiến hành khảo sát giải phẫu sơ khởi rễ.
Ngoài ra, khảo sát ảnh hưởng NAA/IBA đến cảm ứng tạo rễ bất định từ chồi có nguồn
gốc từ đốt thân vườn ươm/đốt thân in vitro, chồi có nguồn gốc phôi vô tính.
2.2.4. Nội dung 4. Nhân rễ bất định
Sử dụng các rễ đơn 20 ngày tuổi làm vật liệu nuôi cấy cho nhân rễ bất định.
Tiến hành khảo sát chất ĐHST (NAA/IBA) ảnh hưởng đến sự phân nhánh rễ;
ảnh hưởng của nồng độ đường, khối lượng rễ nuôi cấy đến tăng trưởng sinh khối rễ.
Khảo sát diễn biến tăng trưởng sinh khối rễ theo thời gian.

37
Sơ đồ tóm tắt các nội dung nghiên cứu
TN 1. Ảnh hưởng của auxin (NAA/IBA) và môi trường
khoáng đến sự tạo phôi trực tiếp từ mô lá
TN 2. Ảnh hưởng của sự kết hợp auxin và BA đến sự
tạo phôi trực tiếp từ mô lá
TN 3. Ảnh hưởng của nồng độ đường và điều kiện chiếu
sáng đến sự tạo phôi trực tiếp từ mô lá
TN 4. Ảnh hưởng của nước dừa đến sự tạo phôi trực tiếp
từ mô lá
TN 6. Trồng cây con ở chậu đất
Quan sát cấu trúc giải phẫu phôi
TN 7. Tạo mô sẹo từ nuôi cấy mảnh lá (10 x 10 mm)
TN 5. Tạo cây con in vitro hoàn chỉnh từ phôi vô tính
TN 8. Tạo phôi từ mô sẹo nuôi cấy mảnh lá (10 x 10 mm)
trên môi trường đặc
Quan sát hình thái cụm tế bào/cụm mô nuôi lỏng lắc
TN 10. Tạo mô sẹo từ nuôi cấy mảnh lá (3 x 10 mm)
TN 11. Tạo phôi từ mô sẹo mảnh lá (3 x 10 mm)
NỘI DUNG 1
Tạo phôi vô tính
TN 13. Ảnh hưởng của chất ĐHST đến sự tạo phôi thứ cấp
Vậ
t liệu
TN 9. Tạo phôi từ mô sẹo nuôi trong môi trường lỏng
Quan sát cấu trúc giải phẫu phôi tái sinh trên môi
trường đặc
TN 14. Ảnh hưởng của khối lượng phôi nuôi cấy đến sự
tăng trưởng sinh khối phôi
TN 17. Ảnh hưởng của cường độ ánh sáng đến sự tăng
trưởng sinh khối phôi
NỘI DUNG 2
Nhân phôi vô tính
TN 15. Ảnh hưởng của kích thước phôi nuôi cấy đến sự
tạo phôi thứ cấp
TN 18. Ảnh hưởng của nước dừa đến sự tạo phôi thứ cấp
Quan sát cấu trúc giải phẫu phôi sơ cấp và thứ cấp.
TN 16. Ảnh hưởng của nồng độ đường đến sự tăng trưởng
sinh khối phôi
TN 19. Tạo cây con từ phôi nuôi lỏng lắc
Nh
ân
ph
ôi tro
ng
mô
i trườ
ng lỏ
ng
TN 12. Nhân phôi trên môi trường đặc
Tạ
o p
hô
i vô
tính
trự
c tiếp từ
mô
lá
Tạ
o p
hô
i vô
tính
giá
n tiếp
qu
a m
ô sẹo
mả
nh
lá

38
Tạ
o rễ b
ất đ
ịnh
trự
c tiếp
từ m
ô lá
TN 20. Ảnh hưởng của auxin (NAA/IBA) và môi trường
khoáng đến sự tạo rễ bất định trực tiếp từ mô lá
TN 22. Ảnh hưởng của điều kiện chiếu sáng đến sự tạo rễ
bất định trực tiếp từ mô lá
Khảo sát hình thái giải phẫu rễ tái sinh trực tiếp từ mô
lá
TN 23. Ảnh hưởng của NAA, IBA đến sự tạo rễ bất định
từ chồi in vitro của đốt thân cây vườn ươm
TN 24. Ảnh hưởng của NAA, IBA đến sự tạo rễ bất định
từ chồi của đốt thân cây in vitro
TN 25. Ảnh hưởng của NAA, IBA đến sự tạo rễ bất định
từ chồi có nguồn gốc phôi vô tính
NỘI DUNG 4
Nhân rễ bất định
TN 26. Ảnh hưởng của NAA, IBA đến sự phân nhánh rễ
TN 27. Ảnh hưởng của nồng độ đường đến sự tăng trưởng
sinh khối rễ
TN 28. Ảnh hưởng của khối lượng rễ nuôi cấy đến sự tăng
trưởng sinh khối rễ
TN 29. Khảo sát diễn biến tăng trưởng sinh khối rễ theo
thời gian
Tạo rễ b
ất đ
ịnh
từ
chồi
Vậ
t liệu
TN 21. Ảnh hưởng của nồng độ đường đến sự tạo rễ bất
định trực tiếp từ mô lá
NỘI DUNG 3
Tạo rễ bất định

39
2.3. Nội dung 1. Tạo phôi vô tính
2.3.1. Tạo phôi vô tính trực tiếp từ mô lá
2.3.1.1. Ảnh hưởng của auxin (NAA/IBA) và môi trường khoáng đến sự tạo phôi
trực tiếp từ mô lá
Để khảo sát ảnh hưởng riêng rẽ của loại auxin với nồng độ thích hợp và môi
trường khoáng đến sự phát sinh phôi trực tiếp từ mảnh lá, tiến hành bố trí thí nghiệm
theo kiểu hai yếu tố: yếu tố chất ĐHST với các mức nồng độ khác nhau; yếu tố về
môi trường khoáng với 4 loại môi trường.
Vật liệu nuôi cấy: mảnh lá kích thước 10 x 10 mm.
Môi trường nuôi cấy là ½MS, MS [105], B5 [106] và SH [107] bổ sung loại
auxin riêng lẻ (NAA hoặc IBA) ở các nồng độ khác nhau (0, 1, 2, 3, 4, 5, 6 mg/L), có
chứa 30 g/L đường (sucrose). Thí nghiệm được bố trí theo kiểu hoàn toàn ngẫu nhiên,
tiến hành cấy 5 mẫu/đĩa, 6 đĩa/lần lặp lại, mỗi nghiệm thức lặp lại 3 lần. Ghi nhận các
chỉ tiêu về tỷ lệ mẫu tạo phôi (%); số phôi/mẫu tạo phôi tại thời điểm 60 NSC.
Quan sát cấu trúc giải phẫu phôi
Cắt dọc phôi (cầu, trái tim, thủy lôi) thành lát mỏng bằng dao lam, ngâm vi
phẫu vào nước javel đến khi mẫu trắng (loại bỏ nội chất tế bào), rửa sạch mẫu bằng
nước cất. Sau đó, ngâm vi phẫu trong dung dịch acid acetic 10% trong 10 phút (loại
bỏ javel trước khi nhuộm). Thực hiện nhuộm kép vi phẫu bằng thuốc nhuộm son phèn
- lục iod trong 15 phút, rửa sạch mẫu [43]. Quan sát tiêu bản bằng kính hiển vi soi
nổi Leica (vật kính 20X) và chụp hình.
Chỉ tiêu theo dõi: Các dạng hình thái của phôi vô tính (dạng phôi cầu, phôi
tim, phôi thủy lôi, phôi dạng hai lá mầm)
2.3.1.2. Ảnh hưởng của sự kết hợp auxin và BA đến sự tạo phôi trực tiếp từ mô lá
Nhằm xác định nồng độ BA tối ưu kết hợp với loại auxin có nồng độ thích hợp
đến cảm ứng tạo phôi vô tính trực tiếp từ mô lá.
Vật liệu nuôi cấy: mảnh lá kích thước 10 x 10 mm
Môi trường nuôi cấy là môi trường khoáng có loại auxin với các nồng độ có
tác động tích cực đến sự hình thành phôi (kế thừa kết quả từ thí nghiệm 2.3.1.1) và
có bổ sung BA với các nồng độ khác nhau (0; 0,25; 0,5; 1mg/L), 30 g/Lsucrose. Thí
nghiệm bố trí theo kiểu hai yếu tố: yếu tố về nồng độ của loại auxin với các mức nồng
độ tác động tích cực đến tạo phôi (kết quả thí nghiệm trên), yếu tố về nồng độ của

40
BA (có 4 mức nồng độ). Cấy 5 mẫu/đĩa, 6 đĩa/lần lặp lại, mỗi nghiệm thức lặp lại 3
lần. Ghi nhận các chỉ tiêu tại thời điểm 60 NSC: tỷ lệ mẫu tạo phôi (%); số phôi/mẫu.
2.3.1.3. Ảnh hưởng của nồng độ đường và điều kiện chiếu sáng đến sự tạo phôi
trực tiếp từ mô lá
Khảo sát nồng độ đường và điều kiện chiếu sáng thích hợp đến cảm ứng tạo
phôi. Môi trường nuôi cấy được sử dụng là môi trường khoáng có nồng độ của loại
auxin (NAA/IBA); nồng độ BA (kế thừa kết quả từ hai thí nghiệm trên) có bổ sung
sucrose với các nồng độ khác nhau (10, 30, 50, 70 g/L) và nuôi mẫu ở điều kiện chiếu
sáng/tối. Ở điều kiện chiếu sáng, mẫu đặt dưới ánh sáng đèn huỳnh quang, chiếu sáng
12 giờ chiếu sáng/ngày, cường độ sáng ~ 4.000 lux; ở điều kiện tối mẫu đặt trong
điều kiện tối hoàn toàn.
Vật liệu nuôi cấy: mảnh lá kích thước 10 x 10 mm.
Thí nghiệm được bố trí theo kiểu hai yếu tố: yếu tố sucrose với 4 mức nồng độ
khác nhau (10, 30, 50, 70 g/L), yếu tố điều kiện chiếu sáng với 2 mức sáng (4.000
lux)/tối. Cấy 5 mẫu/đĩa, 6 đĩa/lần lặp lại, mỗi nghiệm thức lặp lại 3 lần. Theo dõi, ghi
nhận các chỉ tiêu tại thời điểm 60 NSC: tỷ lệ mẫu tạo phôi (%); số phôi/mẫu.
2.3.1.4. Ảnh hưởng của nước dừa đến sự tạo phôi trực tiếp từ mô lá
Các điều kiện tối ưu của thí nghiệm 2.3.1.3 được áp dụng làm môi trường nuôi
cấy có bổ sung nước dừa với các tỷ lệ khác nhau 0, 5, 10, 15, 20% (v/v); để khảo sát
ảnh hưởng của tỷ lệ nước dừa đến cảm ứng tạo phôi trực tiếp từ mô lá.
Vật liệu nuôi cấy: mảnh lá kích thước 10 x 10 mm.
Cấy 5 mẫu/đĩa, 6 đĩa/lần lặp lại, mỗi nghiệm thức lặp lại 3 lần. Các chỉ tiêu
theo dõi được ghi nhận tại thời điểm 60 NSC gồm tỷ lệ mẫu tạo phôi (%); số
phôi/mẫu. Thí nghiệm được bố trí hoàn toàn ngẫu nhiên, theo kiểu đơn yếu tố.
2.3.1.5. Tạo cây con in vitro hoàn chỉnh từ phôi vô tính
Nhằm xác định môi trường nuôi cấy thích hợp để tạo cây con in vitro hoàn
chỉnh, từ đó tạo cây con có thân, bộ lá, rễ tốt phục vụ việc trồng cây ra chậu đất.
Vật liệu nuôi cấy là các cây con có thân cao ~ 1 cm, đã có lá thật 3 – 4 mm
(nguồn gốc từ phôi vô tính).
Tách các cây con, nuôi cấy trên các đĩa petri có chứa 30 mL môi trường nuôi
cấy là MS hoặc ½MS, có chứa 20 g/L sucrose, không bổ sung chất ĐHST. Cấy 5 cây
con/đĩa với 6 đĩa/lần lặp lại, mỗi nghiệm thức lặp lại 3 lần. Quan sát hình thái cơ bản,

41
thu thập số liệu về các chỉ tiêu theo dõi sau cấy 90 ngày. Tại thời điểm 90 NSC tiến
hành đo đếm các chỉ tiêu theo dõi: Chiều cao cây (cm), số lá/cây, số rễ/cây; chiều dài
rễ (mm): được xác định từ gốc rễ đến chóp rễ; quan sát hình thái cơ bản.
Sử dụng môi trường tốt nhất của thí nghiệm làm môi trường nuôi cấy tạo nguồn
cây con in vitro khỏe mạnh. Sử dụng các cây con có nguồn gốc từ phôi vô tính như
trên cấy vào bình tam giác chứa 50 mL môi trường nuôi cấy, cấy 3 cây/bình, 10
bình/lần lặp lại, lặp lại 3 lần. Sau 3 tháng thu các cây con để trồng ra chậu đất ở vườn
ươm. Thí nghiệm được bố trí hoàn toàn ngẫu nhiên theo kiểu đơn yếu tố.
2.3.1.6. Trồng cây con ở chậu đất
Các cây con 3 tháng tuổi được nuôi cấy trong bình tam giác (từ kết quả thí
nghiệm trên), lấy ra khỏi bình tam giác, rửa sạch agar, bảo quản và đảm bảo độ ẩm
tránh mất nước cho cây, sau đó được trồng ra chậu (có Ø ~ 20 cm) chứa đất sạch
thương hiệu Tribat (Cty TNHH Công nghệ sinh học Sài Gòn Xanh, TP. HCM). Các
chậu cây được đặt trong vườn ươm, trồng 1 cây con/chậu, 10 chậu/lần lặp lại, 3 lần
lặp lại, ghi nhận tỷ lệ sống (%) và hình thái cơ bản của cây con ở thời điểm 3 tuần
sau trồng.
Tỷ lệ sống (%) = (Số cây sống/số cây trồng) x 100%
Trồng cây ở vườn ươm với điều kiện che sáng ~ 50% bằng lưới đen (cường độ
sáng ~ 8.000 lux), tưới phun 2 ngày/lần, nhiệt độ 30 - 32oC, độ ẩm 60 - 70%. Theo
dõi, ghi nhận, đánh giá cảm quan hình thái cây đến giai đoạn 3 - 12 tháng sau trồng.
2.3.2. Tạo phôi vô tính gián tiếp qua mô sẹo từ nuôi cấy mô lá
2.3.2.1. Tạo phôi vô tính gián tiếp qua mô sẹo mảnh lá (10 x 10 mm)
Tạo mô sẹo
Vật liệu nuôi cấy: các mảnh lá có kích thước 10 x 10 mm
Mẫu được cấy vào môi trường tạo mô sẹo SH có bổ sung 2,4-D với các nồng
độ (0; 1; 2; 3 mg/L), 30 g/L sucrose. Sau cấy 40 ngày, cấy chuyền các mẫu và nuôi
tiếp trong 20 ngày nhằm để mô sẹo hình thành trên toàn bộ bề mặt mảnh lá. Thí
nghiệm được thực hiện trên các đĩa petri, cấy 5 mảnh lá/đĩa, 3 đĩa/lần lặp lại. Quan
sát, theo dõi sự hình thành mô sẹo ở 20, 40, 60 NSC.
Cắt các mảnh lá mang mô sẹo (sau đây gọi tắt là mảnh mô sẹo) ở 60 NSC nói
trên thành các mảnh có kích thước (~ 10 x 10 mm/~ 1 cm2), nuôi cấy tiếp trong thời

42
gian 30 ngày để tạo mô sẹo có khả năng sinh phôi (KNSP). Tiến hành cấy 5 mảnh
mô sẹo/đĩa, cấy 6 đĩa/lần lặp lại, mỗi nghiệm thức lặp lại 3 lần.
Theo dõi, ghi nhận các chỉ tiêu ở thời điểm 30 NSC: tỷ lệ mẫu tạo mô sẹo có
KNSP (%), số cụm mô sẹo có KNSP/mẫu mô sẹo.
Thí nghiệm được bố trí hoàn toàn ngẫu nhiên theo kiểu đơn yếu tố.
Xác định mô sẹo có KNSP dựa vào đặc điểm hình thái, màu sắc, cấu trúc, mô
sẹo có KNSP: có dạng cứng, trắng hơi vàng, vàng, cấu trúc chắc, dạng cụm hoặc hạt,
phát triển chậm. Mô sẹo không có KNSP có cấu trúc xốp, phát triển nhanh, màu trắng
hoặc xám, mềm.
Tạo phôi từ mô sẹo nuôi trên môi trường đặc
Vật liệu nuôi cấy: các cụm mô sẹo có KNSP
Các cụm mô sẹo có KNSP thu từ thí nghiệm trên được cắt thành các cụm nhỏ
có kích thước ~ 5 mm, cấy vào môi trường tạo phôi là môi trường nuôi cấy có bổ
sung loại auxin và nồng độ thích hợp của BA (kế thừa kết quả thí nghiệm tạo phôi
trực tiếp). Loại auxin này được bố trí với các nồng độ 1; 2; 3; 4; 5 mg/L; có 30 g/L
sucrose.
Tiến hành cấy 5 cụm mô sẹo có KNSP/đĩa, 6 đĩa/lần lặp lại, mỗi nghiệm thức
được lặp lại 3 lần, thí nghiệm bố trí hoàn toàn ngẫu nhiên theo kiểu đơn yếu tố . Theo
dõi, ghi nhận số liệu ở thời điểm 30 NSC với các chỉ tiêu theo dõi: số cụm mô sẹo có
KNSP tạo phôi, số phôi/cụm mô sẹo có KNSP, đặc điểm hình thái phôi.
Quan sát cấu trúc giải phẫu phôi tái sinh trên môi trường đặc
Sử dụng dao lam cắt ngang phôi tái sinh từ cụm mô sẹo có KNSP nuôi trên
môi trường đặc để tạo các lát mỏng tế bào, tiếp theo các mẫu cắt được nhuộm bằng
phương pháp nhuộm màu kép như trên [43]. Quan sát tiêu bản bằng kính hiển vi soi
nổi Leica (vật kính 20X) và chụp hình.
Tạo phôi từ mô sẹo nuôi trên môi trường lỏng
Thí nghiệm được tiến hành với nguồn vật liệu là các cụm mô sẹo có KNSP có
đường kính khoảng 5 mm, cấy 30 cụm mô sẹo có KNSP vào bình tam giác V250 mL
chứa 60 mL môi trường kế thừa từ kết quả thí nghiệm trên, mỗi nghiệm thức lặp lại
3 lần. Theo dõi, ghi nhận sự hình thành phôi ở 10, 14, 30 NSC.
Quan sát hình thái cụm tế bào/cụm mô nuôi lỏng lắc

43
Quan sát, ghi nhận hình thái cụm tế bào sống (không nhuộm màu), cụm đa bào
sinh phôi (qua nhuộm màu) (ở giai đoạn ~ 10 ngày nuôi lỏng lắc mô sẹo có khả năng
sinh phôi) dưới kính hiển vi soi ngược với độ phóng đại 50X, 20X, theo thứ tự.
2.3.2.2. Tạo phôi vô tính gián tiếp qua mô sẹo mảnh lá (3 x 10 mm)
Tạo mô sẹo từ nuôi cấy mảnh lá (3 x 10 mm)
Thí nghiệm này nhằm tạo nguồn vật liệu mô sẹo cho sự khảo sát tạo phôi sau
đó. Vật liệu nuôi cấy được sử dụng là mảnh lá có kích thước 3 x 10 mm. Thí nghiệm
được bố trí hoàn toàn ngẫu nhiên theo kiểu đơn yếu tố.
Mẫu được cấy trên môi trường tạo mô sẹo là SH có bổ sung 2,4-D với các
nồng độ 0; 1; 2; 3 mg/L, 30 g/L sucrose. Tiến hành cấy 10 mẫu/đĩa, 3 đĩa/lần lặp lại,
mỗi nghiệm thức lặp lại 3 lần. Theo dõi, ghi nhận sự hình thành mô sẹo, mô sẹo có
KNSP ở thời điểm 30 NSC.
Tạo phôi
Vật liệu nuôi cấy: mô sẹo hình thành từ thí nghiệm 2.3.2.2
Mẫu mô sẹo được cấy vào môi trường tạo phôi giống môi trường nuôi cấy của
thí nghiệm tạo phôi ở môi trường đặc. Tiến hành cấy 5 mẫu mô sẹo/đĩa, cấy 6 đĩa/lần
lặp lại, mỗi nghiệm thức lặp lại 3 lần. Thí nghiệm được bố trí theo kiểu đơn yếu tố và
hoàn toàn ngẫu nhiên. Theo dõi, quan sát sự hình thành phôi; thu thập số liệu ở thời
điểm 30 NSC với các chỉ tiêu: số mẫu mô sẹo tạo phôi, số phôi/mẫu mô sẹo. Tiếp tục
quan sát, ghi nhận sự phát triển của phôi ở giai đoạn 60 NSC (không thu thập số liệu).
2.3.3. Các chỉ tiêu và phương pháp theo dõi
Phôi vô tính được xác định là một trong bốn dạng hình thái phát triển của phôi:
dạng phôi cầu, phôi tim, phôi thủy lôi, dạng phôi trưởng thành (dạng phân cực có tiền
lá mầm và tiền rễ). Đếm phôi bằng cách quan sát dưới kính lúp và ghi nhận số phôi.
Tiến hành lấy chỉ tiêu theo dõi với 30 mẫu cho mỗi nghiệm thức.
Các chỉ tiêu ở thí nghiệm tạo phôi trực tiếp, theo dõi và thu thập số liệu ở thời
điểm ngày thứ 60 sau cấy:
Tỷ lệ mẫu tạo phôi (%) = (Số mẫu tạo phôi/số mẫu cấy) x 100%
Số phôi/mẫu = Tổng số phôi/số mẫu tạo phôi
Các chỉ tiêu ở thí nghiệm tạo phôi gián tiếp, theo dõi và thu thập số liệu ở thời
điểm ngày thứ 30 sau cấy:
Tỷ lệ mẫu tạo mô sẹo có KNSP (%) = (𝑆ố 𝑚ẫ𝑢 𝑡ạ𝑜 𝑚ô 𝑠ẹ𝑜 𝑐ó 𝐾𝑁𝑆𝑃
𝑆ố 𝑚ẫ𝑢 𝑚ô 𝑠ẹ𝑜 𝑐ấ𝑦) x 100%

44
Số cụm mô sẹo có KNSP/mẫu = 𝑆ố 𝑐ụ𝑚 𝑚ô 𝑠ẹ𝑜 𝑐ó 𝐾𝑁𝑆𝑃
𝑠ố 𝑚ẫ𝑢 𝑚ô 𝑠ẹ𝑜 𝑐ó 𝐾𝑁𝑆𝑃
Số phôi/cụm mô sẹo = 𝑇ổ𝑛𝑔 𝑠ố 𝑝ℎô𝑖
𝑆ố 𝑐ụ𝑚 𝑚ô 𝑠ẹ𝑜 𝑡ạ𝑜 𝑝ℎô𝑖
2.4. Nội dung 2. Nhân phôi vô tính
2.4.1. Nhân phôi trên môi trường đặc
Vật liệu nuôi cấy là các cụm phôi vô tính sơ cấp (hình thành từ mảnh lá).
Môi trường nhân phôi được sử dụng từ kết quả của thí nghiệm tạo phôi trực
tiếp từ mảnh lá về môi trường khoáng, loại auxin, nồng độ BA. Loại auxin này được
sử dụng với các nồng độ khác nhau (0; 1; 2; 3; 4; 5 mg/L), 30 g/L sucrose. Thí nghiệm
được bố trí trên các đĩa petri, theo kiểu đơn yếu tố và hoàn toàn ngẫu nhiên. Cấy 5
cụm phôi/đĩa (3 phôi/cụm), 6 đĩa/lần lặp lại, mỗi nghiệm thức lặp lại 3 lần. Thu thập
số liệu về số phôi/cụm phôi, hệ số nhân phôi, quan sát đặc điểm hình thái phôi, cụm
phôi ở 30 NSC. Chú ý quan sát sự hình thành phôi thứ cấp ở các bộ phận (từ thân, lá,
rễ) của phôi sơ cấp.
2.4.2. Nhân phôi trong môi trường lỏng
2.4.2.1. Ảnh hưởng của chất điều hòa sinh trưởng đến sự tạo phôi thứ cấp qua
nuôi lỏng lắc
Sử dụng kết quả của thí nghiệm trên làm cơ sở cho thí nghiệm khảo sát tác
động của các chất ĐHST đến sự phát sinh phôi thứ cấp trong môi trường lỏng. Do
đó, môi trường nuôi cấy được kế thừa về môi trường khoáng, các mức nồng độ của
loại auxin có phát sinh phôi tích cực và nồng độ BA; nuôi cấy lỏng lắc. Sử dụng máy
lắc hiệu SARTORIUS của Đức, thực hiện nuôi lỏng lắc với 80 vòng/phút.
Vật liệu nuôi cấy là các phôi đơn 60 ngày tuổi (từ thí nghiệm tạo phôi vô tính
trực tiếp từ mô lá).
Thí nghiệm được tiến hành trong bình tam giác V250 mL, chứa 60 mL môi
trường nuôi cấy, nuôi lỏng lắc 80 vòng/phút, mỗi nghiệm thức lặp lại 3 lần. Thí
nghiệm được bố trí theo kiểu đơn yếu tố, cấy 30 phôi/bình. Quan sát, theo dõi thí
nghiệm, thu thập số liệu về số phôi thứ cấp hình thành, đặc điểm hình thái và màu sắc
phôi ở 30 NSC.

45
2.4.2.2. Ảnh hưởng của khối lượng phôi nuôi cấy đến sự tăng trưởng sinh khối
phôi
Vật liệu nuôi cấy: sử dụng phôi qua nuôi cấy lỏng lắc sau 60 ngày nuôi cấy từ
phôi 60 ngày tuổi của các thí nghiệm tạo phôi vô tính trực tiếp từ mô lá.
Để khảo sát ảnh hưởng khối lượng phôi nuôi cấy đến sự tăng trưởng sinh khối
phôi, sử dụng khối lượng phôi ban đầu như sau: 0,3; 0,6; 1,2; 1,8 g tương ứng với
0,5%, 1%, 2% và 3% (w/v) vào bình tam giác V250 mL chứa 60 mL môi trường nuôi
cấy kế thừa thí nghiệm ảnh hưởng chất ĐHST, 30 g/L đường. Thu thập số liệu về
khối lượng tươi phôi (g), hệ số nhân phôi (lần) và đặc điểm hình thái phôi/cụm phôi
ở 30 NSC.
Thí nghiệm bố trí theo kiểu đơn yếu tố, cấy 3 bình/lần lặp lại, mỗi nghiệm thức
lặp lại 3 lần. Quan sát, theo dõi và thu thập số liệu về số phôi
2.4.2.3. Ảnh hưởng của kích thước phôi nuôi cấy đến sự tạo phôi thứ cấp
Sử dụng vật liệu là các phôi vô tính với các kích thước khác nhau: kích thước
nhỏ (7 - 8 mm), trung bình (~ 10 mm), to (~ 15 mm). Tiến hành cấy 30 phôi nhỏ/trung
bình/to vào bình tam giác V250 mL chứa 60 mL môi trường nuôi cấy giống thí
nghiệm ảnh hưởng khối lượng phôi nuôi cấy, có 30 g/L đường.
Thí nghiệm bố trí theo kiểu đơn yếu tố, cấy 3 bình/lần lặp lại, mỗi nghiệm thức
lặp lại 3 lần. Quan sát, theo dõi và thu thập số liệu về số phôi thứ cấp hình thành, đặc
điểm hình thái, màu sắc phôi/ cụm phôi hình thành ở 30 NSC.
2.4.2.4. Ảnh hưởng của nồng độ đường đến sự tăng trưởng sinh khối phôi
Cấy khối lượng phôi 0,3 g (0,5% - w/v) vào bình tam giác V250 mL chứa 60
mL môi trường như thí nghiệm trên, sucrose được khảo sát với các mức nồng độ (20,
30, 40, 50 g/L); mỗi nghiệm thức được lặp lại 3 lần. Thu thập số liệu về khối lượng
tươi phôi (g), hệ số nhân phôi (lần) và đặc điểm hình thái, màu sắc phôi/cụm phôi ở
45 NSC. Thí nghiệm bố trí hoàn toàn ngẫu nhiên theo kiểu đơn yếu tố
2.4.2.5. Ảnh hưởng của cường độ ánh sáng đến sự tăng trưởng sinh khối phôi
Cấy khối lượng phôi 0,30 g (0,5% - w/v) vào bình tam giác V250 mL chứa 60
mL môi trường như trên, 30 g/L sucrose, nuôi ở các điều kiện chiếu sáng khác nhau:
tối (0 lux), chiếu sáng với cường độ 2.000 lux (~ 27 μmol m-2 s-1), 4.000 lux (~ 54
μmol m-2 s-1), mỗi nghiệm thức lặp lại 3 lần. Quan sát, theo dõi, thu thập số liệu về

46
khối lượng tươi phôi (g), hệ số nhân phôi (lần), đặc điểm hình thái, màu sắc phôi/cụm
phôi ở 30 NSC. Thí nghiệm bố trí hoàn toàn ngẫu nhiên theo kiểu đơn yếu tố.
2.4.2.6. Ảnh hưởng của tỷ lệ nước dừa đến sự tạo phôi thứ cấp
Các phôi đơn có kích thước trung bình (~ 10 mm) được cấy vào bình tam giác
V250 mL chứa 60 mL môi trường thí nghiệm ảnh hưởng khối lượng phôi nuôi cấy
trên, 30 g/L sucrose, nước dừa được khảo sát với các mức tỷ lệ (0%, 5%, 10%). Cấy
30 phôi đơn/bình, mỗi nghiệm thức lặp lại 3 lần. Thu thập số liệu về số phôi hình
thành, đặc điểm hình thái, màu sắc phôi/cụm phôi ở 21 NSC.
Thí nghiệm bố trí hoàn toàn ngẫu nhiên theo kiểu đơn yếu tố
2.4.2.7. Tạo cây con từ phôi nuôi lỏng lắc
Sử dụng các phôi đơn có kích thước trung bình (~ 10 mm), cấy vào bình tam
giác V250 mL chứa 60 mL môi trường MS hoặc ½MS (1/2 khoáng đa lượng) [105]
không bổ sung chất điều hòa sinh trưởng, 20 g/L sucrose, cấy 30 phôi đơn/bình, mỗi
nghiệm thức lặp lại 3 lần; quan sát, ghi nhận sự phát triển phôi đến giai đoạn trưởng
thành ở 14 NSC. Sau đó, cấy chuyển phôi sang môi trường đặc thích hợp (kế thừa kết
quả thí nghiệm) để phôi phát triển thành cây con trong 60 ngày.
2.4.3. Quan sát cấu trúc giải phẫu phôi sơ cấp và thứ cấp
Cắt tạo các LMTB dọc thân phôi/rễ, ngang phiến lá mầm (của phôi sơ cấp)
mang phôi thứ cấp bằng dao lam, mẫu cắt được nhuộm bằng phương pháp nhuộm
kép của Bộ môn Thực vật - Khoa Dược, ĐH. Y Dược TP. HCM [43]. Quan sát tiêu
bản bằng kính hiển vi soi nổi Leica (vật kính 20X) và chụp hình.
2.4.4. Các chỉ tiêu theo dõi và phương pháp xác định
Số phôi/cụm phôi = Tổng số phôi/số cụm phôi
Hệ số nhân phôi = Số phôi/cụm phôi
𝑆ố 𝑝ℎô𝑖 𝑐ấ𝑦/𝑐ụ𝑚
Khối lượng tươi phôi thu nhận (g): Thu và cân sinh khối lượng phôi ở mỗi
nghiệm thức sau 30 ngày nuôi.
Hệ số nhân sinh khối phôi (lần) = 𝐾ℎố𝑖 𝑙ượ𝑛𝑔 𝑡ươ𝑖 𝑝ℎô𝑖 𝑡ℎ𝑢 𝑛ℎậ𝑛(𝑔)
𝐾ℎố𝑖 𝑙ượ𝑛𝑔 𝑝ℎô𝑖 𝑛𝑢ô𝑖 𝑐ấ𝑦 (𝑔)
2.5. Nội dung 3. Tạo rễ bất định
2.5.1. Tạo rễ bất định trực tiếp từ mô lá
2.5.1.1. Ảnh hưởng của auxin (NAA/IBA) và môi trường khoáng đến sự tạo rễ bất
định trực tiếp từ mô lá

47
Khảo sát tác động của riêng lẻ của auxin và môi trường khoáng lên sự cảm
ứng tạo rễ bất định của mô lá, nhằm xác định loại và nồng độ auxin thích hợp, trên
môi trường khoáng thích hợp.
Sử dụng vật liệu là các mảnh lá I (10 x 10 mm), mảnh lá II (3 x 10 mm), cấy
úp mặt trên lá tiếp xúc vào môi trường MS, ½MS, B5 và SH có bổ sung các chất
ĐHST riêng lẻ NAA/IBA có cùng nồng độ (0; 0,5; 1; 2; 3; 4; 5) mg/L, 30 g/L sucrose,
chiếu sáng 4.000 lux, 12 h chiếu sáng/ngày. Cấy 5 mảnh lá I/đĩa, 6 đĩa/lần lặp lại; 10
mảnh lá II/đĩa, 3 đĩa/lần lặp lại; mỗi nghiệm thức lặp lại 3 lần.
Thí nghiệm bố trí hoàn toàn ngẫu nhiên theo kiểu hai yếu tố: yếu tố về các
mức nồng độ của NAA/IBA và yếu tố về môi trường khoáng với 4 môi trường khác
nhau (MS, ½MS, B5, SH). Theo dõi thí nghiệm, thu thập số liệu về tỷ lệ mẫu tạo rễ
(%), số rễ/mẫu, chiều dài rễ (mm) ở thời điểm 30 NSC.
2.5.1.2. Ảnh hưởng của nồng độ đường đến sự tạo rễ bất định trực tiếp từ mô lá
Nhằm khảo sát sự tác động của nồng độ đường đến sự phát sinh rễ từ các mẫu
cấy mảnh lá, sử dụng môi trường khoáng với chất ĐHST (NAA/IBA) có nồng độ tạo
rễ tích cực nhất (kế thừa từ thí nghiệm trên), có đường với các nồng độ khác nhau
(20; 30; 40; 50 g/L) làm môi trường nuôi cấy.
Vật liệu nuôi cấy: mảnh lá I (10 x 10 mm), mảnh lá II (3 x 10 mm). Mẫu được
cấy vào môi trường nuôi cấy trên với 5 mảnh lá I/đĩa, 6 đĩa/lần lặp lại; 10 mảnh lá
II/đĩa, 3 đĩa/lần lặp lại; mỗi nghiệm thức lặp lại 3 lần. Thí nghiệm được bố trí hoàn
toàn ngẫu nhiên, theo kiểu đơn yếu tố. Theo dõi thí nghiệm, số liệu về tỷ lệ mẫu tạo
rễ (%), số rễ/mẫu, chiều dài rễ (mm) được thu thập ở thời điểm 30 NSC.
2.5.1.3. Ảnh hưởng của điều kiện chiếu sáng đến sự tạo rễ bất định trực tiếp từ mô
lá
Để khảo sát điều kiện chiếu sáng có tác động đến sự phát sinh rễ bất định từ
mảnh lá, tiến hành cấy mẫu trên môi trường khoáng, có loại và nồng độ auxin thích
hợp, nồng độ sucrose có tác động hiệu quả nhất đến sự phát sinh rễ bất định từ mảnh
lá (kế thừa kết quả từ hai thí nghiệm trên). Các đĩa mẫu cấy được đặt ở điều kiện
chiếu sáng khác nhau 4.000 lux, 2.000 lux, tối hoàn toàn.
Sử dụng vật liệu nuôi cấy là các mảnh lá I (10 x 10 mm) và mảnh lá II (3 x 10
mm). Với mảnh lá I cấy 5 mẫu/đĩa, 6 đĩa/lần lặp lại; mảnh lá II cấy 10 mẫu/đĩa, 3
đĩa/lần lặp lại; mỗi nghiệm thức lặp lại 3 lần. Thí nghiệm được bố trí hoàn toàn ngẫu

48
nhiên, theo kiểu đơn yếu tố. Quan sát, theo dõi thí nghiệm, ghi nhận số liệu về tỷ lệ
mẫu tạo rễ (%), số rễ/mẫu (tạo rễ), chiều dài rễ (mm) thu thập ở thời điểm 30 NSC.
2.5.1.4. Khảo sát hình thái giải phẫu rễ tái sinh trực tiếp từ mô lá
Cắt dọc các sơ khởi rễ 10 ngày tuổi, dài ~ 4 mm (hình thành trực tiếp ở mặt
trên của mảnh lá nuôi cấy trên môi trường ½MS có 3 mg/L NAA) tạo LMTB bằng
dao lam, vi phẫu được nhuộm bằng phương pháp nhuộm kép [43]. Quan sát tiêu bản
bằng kính hiển vi soi nổi Leica (độ phóng đại 20X) và chụp hình.
2.5.2. Tạo rễ bất định từ chồi
2.5.2.1. Tạo rễ bất định từ chồi có nguồn gốc từ đốt thân cây vườn ươm
Khử trùng mẫu.
Hình 2.3. Vật liệu đốt thân dùng nuôi cấy tạo chồi.
A. Nhánh cây (vạch ngang là vị trí cắt tạo các đốt thân; CL: cuống lá); B, C. Các đốt
thân dùng nuôi cấy tạo chồi (mũi tên chỉ vị trí chồi ở trạng thái ngủ) (thanh ngang 10
mm).
Sử dụng nhánh non (8 - 10 cm) cây NGBCC (Hình 2.3 A). Trước hết cắt bỏ
lá, rửa nhánh dưới vòi nước chảy trong 10 phút, rửa tiếp bằng nước xà phòng. Cắt
nhánh thành các đoạn mang chồi ngủ - đốt nhánh (sau đây gọi là đốt thân) dài ~ 1,5
cm, không sử dụng chồi ngọn, tiếp theo khử trùng bằng cồn 70% trong 1 phút, bằng
nước Javel 30% (v/v) trong 30 phút, rửa lại 3 lần bằng nước cất vô trùng. Các đốt
thân, sau khử trùng, cắt bỏ phần mô chết ở hai đầu (Hình 2.3 B, C) được dùng làm
vật liệu nuôi cấy trong thí nghiệm tạo chồi.
Tạo chồi từ đốt thân cây vườn ươm
Cấy các đốt thân sau khử trùng vào môi trường MS có bổ sung 0,5 mg/L BA
và 2 mg/L kinetin, 30 g/L sucrose [3]. Ghi nhận sự tạo chồi ở 60 NSC.
Tạo rễ bất định từ chồi
Tách các chồi (cao ~ 1 cm) và cấy vào môi trường tạo rễ bất định ½MS có bổ
sung 0,2 mg/L NAA [3], 20 g/L sucrose, 10 g/L agar, pH 5,8; IBA cũng được sử dụng

49
với nồng độ 0,2 mg/L. Cấy 2 chồi nách/bình, cấy 15 bình/lần lặp lại, mỗi nghiệm thức
lặp lại 3 lần. Theo dõi, ghi nhận số liệu về các chỉ tiêu: tỷ lệ chồi tạo rễ (%), số rễ/chồi,
chiều dài rễ (mm) ở 60 NSC.
Thí nghiệm được bố trí hoàn toàn ngẫu nhiên, theo kiểu đơn yếu tố
2.5.2.2. Tạo rễ bất định từ chồi đốt thân cây in vitro
Tạo chồi từ đốt thân cây in vitro
Cây in vitro ~ 5 tháng tuổi (có nguồn gốc từ phôi vô tính được nuôi trên môi
trường ½MS không bổ sung chất ĐHST, có 30 g/L sucrose) được cắt bỏ toàn bộ lá,
sau đó cắt thân thành các đoạn dài ~ 8 mm mang chồi nách và cấy vào môi trường
tạo chồi như trên. Theo dõi sự tạo chồi ở 60 NSC.
Tạo rễ bất định từ chồi
Tách các chồi nách 60 ngày tuổi (cao 0,7 - 1 cm), cấy vào môi trường tạo rễ
bất định như trường hợp trên. Cấy 2 chồi nách/bình, cấy 15 bình/lần lặp lại, mỗi
nghiệm thức lặp lại 3 lần. Theo dõi, ghi nhận số liệu về các chỉ tiêu: tỷ lệ chồi tạo rễ
(%), số rễ/chồi, chiều dài rễ (mm) ở 60 NSC.
2.5.2.3. Tạo rễ bất định từ chồi có nguồn gốc phôi vô tính
Sử dụng vật liệu cây con có nguồn gốc từ phôi vô tính có chiều cao thân ~ 15
mm, cắt bỏ phần rễ trụ, giữ phần thân (trụ dưới lá mầm) và các lá phía trên, dùng như
vật liệu tạo rễ bất định. Mẫu được cấy vào môi trường ½MS có bổ sung NAA hoặc
IBA cùng nồng độ 0,1 mg/L, 20 g/L sucrose. Cấy 2 chồi/bình, cấy 15 bình/lần lặp lại,
mỗi nghiệm thức lặp lại 3 lần. Chỉ tiêu theo dõi: Tỷ lệ mẫu tạo rễ (%); số rễ/mẫu;
chiều dài rễ (mm) và đặc điểm hình thái, màu sắc rễ tại thời điểm 30 NSC
2.5.3. Các chỉ tiêu và phương pháp theo dõi
Tỷ lệ mẫu tạo rễ (%) = (Số mẫu tạo rễ/số mẫu cấy) x 100%
Số rễ/mẫu = Tổng số rễ/số mẫu tạo rễ
Chiều dài rễ (mm) = Tổng chiều dài các rễ/số rễ được đo
Các chỉ tiêu của thí nghiệm tạo rễ từ chồi đốt thân cây in vitro được thu thập
ở 60 NSC:
Tỷ lệ chồi tạo rễ (%) = (Số chồi tạo rễ/số chồi cấy) x 100%
Số rễ/chồi = Số rễ hình thành/số chồi tạo rễ
Chiều dài rễ (mm) = Tổng chiều dài các rễ/số rễ được đo

50
2.6. Nội dung 4. Nhân rễ bất định trong môi trường lỏng
2.6.1. Ảnh hưởng của chất điều hòa sinh trưởng đến sự phân nhánh của rễ
Sử dụng các rễ đơn (rễ sơ cấp, dài ~ 1 cm) tái sinh trực tiếp từ mảnh lá trên
môi trường đặc ½MS có 3 mg/L NAA ở thời điểm 20 NSC làm vật liệu nuôi cấy.
Cấy ~ 120 rễ đơn vào bình tam giác V250 mL chứa 60 mL (~ 0,5% - w/v) môi
trường lỏng ½MS có bổ sung NAA hoặc IBA với các nồng độ khác nhau 0, 1, 2, 3
mg/L. Sau 20 ngày nuôi cấy lấy ngẫu nhiên 30 rễ/lần, 3 lần đo đếm, thu thập số liệu
về chiều dài rễ sơ cấp (mm), số rễ thứ cấp cấp 1, chiều dài rễ thứ cấp cấp 1 (mm), số
rễ thứ cấp (sau đây gọi là rễ nhánh) cấp 2 hình thành và đặc điểm hình thái rễ.
2.6.2. Ảnh hưởng của nồng độ đường đến sự tăng trưởng sinh khối rễ
Cấy khối lượng rễ 0,3 g (0,5% - w/v) vào bình tam giác V250 mL chứa 60 mL
môi trường ½MS bổ sung NAA/IBA với nồng độ kế thừa kết quả thí nghiệm (2.6.1);
sucrose với các nồng độ (20, 30 và 40 g/L). Thu thập số liệu về khối lượng tươi rễ
(g), hệ số nhân rễ (lần) và đặc điểm hình thái, màu sắc rễ, cụm rễ ở thời điểm 30 NSC,
mỗi nghiệm thức lặp lại 3 lần. Thí nghiệm bố trí theo kiểu hai yếu tố: yếu tố chất
ĐHST (NAA, IBA) và yếu tố nồng độ sucrose với ba mức nồng độ (20, 30, 40 g/L).
2.6.3. Ảnh hưởng của khối lượng rễ nuôi cấy đến sự tăng trưởng sinh khối rễ
Để khảo sát ảnh hưởng của khối lượng rễ nuôi cấy đến tăng sinh khối rễ, sử
dụng khối lượng rễ 0,35 g; 0,70 g; 1,4 g; 2,1 g tương ứng với 0,5%, 1%, 2% và 3%
(w/v) vào bình tam giác V250 mL chứa 70 mL môi trường ½MS có bổ sung NAA
hoặc IBA có nồng độ kế thừa thí nghiệm (2.6.1), nồng độ sucrose được sử dụng từ
thí nghiệm (2.6.2). Thu thập số liệu về khối lượng tươi rễ (g), hệ số nhân rễ (lần) và
đặc điểm hình thái, màu sắc rễ, cụm rễ ở 30 NSC, mỗi nghiệm thức lặp lại 3 lần. Thí
nghiệm bố trí theo kiểu hai yếu tố: yếu tố chất ĐHST (NAA, IBA) và yếu tố khối
lượng rễ nuôi cấy với bốn mức nồng độ (0,35; 0,7; 1,4; 2,1%).
2.6.4. Khảo sát diễn biến tăng trưởng sinh khối rễ theo thời gian
Cấy khối lượng rễ thích hợp nhất (từ kết quả của thí nghiệm 2.6.3) vào bình
tam giác V250 mL chứa 70 mL môi trường nuôi cấy giống với thí nghiệm (2.6.3).
Sau thời gian nuôi cấy 28 ngày, tiến hành thay 70 mL môi trường nuôi cấy có thành
phần giống môi trường ban đầu. Thu thập số liệu về khối lượng tươi rễ (g), hệ số nhân
rễ (lần) ở 7, 14, 21, 28, 35, 42 và 49 NSC, mỗi nghiệm thức lặp lại 3 lần. Thí nghiệm

51
bố trí theo kiểu hai yếu tố: yếu tố chất ĐHST (NAA, IBA) và yếu tố thời gian với các
mức thời gian (7, 14, 21, 28, 35, 42, 49 ngày).
2.6.5. Các chỉ tiêu và phương pháp theo dõi
Chiều dài rễ sơ cấp (mm) = Chiều dài các rễ/số rễ được đo
Số rễ nhánh cấp 1 = Số rễ hình thành từ rễ sơ cấp/số rễ sơ cấp
Chiều dài rễ nhánh cấp 1 (mm) = Chiều dài của các rễ nhánh/số rễ nhánh
Số rễ nhánh cấp 2 = 𝑇ổ𝑛𝑔 𝑠ố 𝑟ễ 𝑛ℎá𝑛ℎ ℎì𝑛ℎ 𝑡ℎà𝑛ℎ 𝑡ừ 𝑟ễ 𝑐ấ𝑝 1
𝑆ố 𝑟ễ 𝑛ℎá𝑛ℎ 𝑐ấ𝑝 1
Khối lượng tươi rễ (g): khối lượng rễ tươi thu nhận được 30 NSC
Hệ số nhân (lần) = 𝐾ℎố𝑖 𝑙ượ𝑛𝑔 𝑟ễ 𝑡ℎ𝑢 𝑛ℎậ𝑛 (𝑔)
𝐾ℎố𝑖 𝑙ượ𝑛𝑔 𝑟ễ 𝑛𝑢ô𝑖 𝑐ấ𝑦 (𝑔)
2.7. Điều kiện nuôi cấy in vitro
Các thí nghiệm được thực hiện ở nhiệt độ phòng 25 - 28oC, độ ẩm trung bình
50 – 60%, các thí nghiệm đều được đặt ở ngoài sáng để dưới ánh sáng đèn huỳnh
quang, chiếu sáng 12 giờ chiếu sáng/ngày, cường độ sáng ~ 4.000 lux (trừ thí nghiệm
tạo phôi trực tiếp ở điều kiện tối đặt trong điều kiện tối hoàn toàn; thí nghiệm ảnh
hưởng của cường độ ánh sáng đến tăng trưởng sinh khối phôi; thí nghiệm ảnh hưởng
của điều kiện chiếu sáng đến tạo rễ bất định ở điều kiện sáng 2.000 lux, ở điều kiện
tối mẫu đặt ở điều kiện tối hoàn toàn). Sử dụng môi trường đặc (agar 10 g/L, pH 5,8;
hấp khử trùng ở 1 atm/121oC trong 20 phút); thí nghiệm trong môi trường lỏng lắc
với tốc độ lắc 80 vòng/phút.
2.8. Phương pháp thống kê và xử lý số liệu
Thí nghiệm được bố trí hoàn toàn ngẫu nhiên, mỗi nghiệm thức lặp lại 3 lần.
Các kết quả trung bình của 3 lần lặp lại được xử lý và phân tích bằng ANOVA một
yếu tố, kiểm tra mức độ khác biệt giữa các nghiệm thức bằng LSD hoặc Duncan. Các
thí nghiệm 2 yếu tố được xử lý theo phân tích phương sai đa biến một chiều (One-
way MANOVA) của phép thử Duncan với p ≤ 0,05 bằng phần mềm SPSS 25.0. (Số
liệu tỷ lệ % biến động từ 0 – 100% chuyển đổi sang arcsin √𝑥 khi xử lý thống kê,
hoặc từ 70 – 100% được chuyển đổi sang (x + 0,5)1/2.

52
CHƯƠNG 3. KẾT QUẢ VÀ THẢO LUẬN
3.1. Tạo phôi vô tính
Trong nghiên cứu này, phôi vô tính hình thành trực tiếp được định nghĩa là
phôi tái sinh ngay ở lần nuôi cấy đầu tiên, khác với phôi tái sinh gián tiếp qua hai giai
đoạn nuôi cấy là giai đoạn tạo mô sẹo và phát sinh phôi từ mô sẹo [108][109].
3.1.1. Tạo phôi vô tính trực tiếp từ mô lá
3.1.1.1. Ảnh hưởng của auxin (NAA/IBA) và môi trường khoáng đến sự tạo phôi
vô tính trực tiếp từ mô lá
Hình 3.1. Các dạng phát triển của phôi vô tính hình thành trực tiếp từ mô lá.
A,B. Phôi cầu đơn, cụm phôi cầu; C. Phôi dạng tim (1) và dạng cầu (2); D. Phôi dạng
thủy lôi; E,F,G,H. Phôi có lá mầm và rễ mầm phát triển; I. Cây con hoàn chỉnh lá
mầm, lá thật (đơn, kép), rễ cọc điển hình. Mũi tên chỉ lá mầm (trắng), rễ mầm (đen).
Thanh ngang 2 mm.
Sau 5 tuần nuôi cấy, phôi vô tính xuất hiện ở nhiều nghiệm thức, ở môi trường
SH có bổ sung 5 mg/L NAA phôi xuất hiện sớm và nhiều nhất, môi trường ½MS phôi
phát sinh chậm, ít hơn. Sự phân hóa cấu trúc trong quá trình phát sinh phôi từ mảnh
lá NGBCC bắt đầu từ dạng phôi cầu, xuất hiện khoảng 32 – 35 NSC, có màu vàng
nhạt (Hình 3.1A), giai đoạn phôi hình tim có màu xanh, xuất hiện sau đó khoảng 10
ngày, đây là dấu hiệu tiền hai lá mầm, dạng đặc trưng của sự hình thành phôi ở cây
hai lá mầm (Hình 3.1C). Dạng phôi thủy lôi (Hình 3.1D) phát triển tiếp đến giai đoạn
có hai lá mầm khoảng 15 – 20 ngày tiếp theo (Hình 3.1E,F,G,H). Phôi phát sinh trực

53
tiếp từ mảnh lá NGBCC đã trải qua tuần tự 04 dạng phát triển của phôi.
Quan sát sự phát sinh phôi từ mảnh lá NGBCC nhận thấy, các hình dạng khác
nhau và các giai đoạn phát triển của phôi cùng xuất hiện trên cùng mẫu cấy (Hình
3.1C1,C2), điều đó chứng tỏ quá trình phát sinh phôi không đồng bộ và các tế bào
khi đủ điều kiện sẽ phát sinh phôi.
Sau 60 ngày nuôi cấy, ở môi trường không bổ sung NAA/IBA, các mẫu cấy
đều không phát sinh hình thái, không xuất hiện phôi, mẫu cấy hóa nâu và chết sau đó,
điều đó chứng tỏ lượng chất ĐHST nội sinh ít không đủ cảm ứng phát sinh phôi. Tất
cả các mẫu cấy trên môi trường ½MS, MS, B5, SH có bổ sung IBA (1; 2; 3; 4; 5; 6
mg/L), đều không ghi nhận được cấu trúc phôi xuất hiện trong suốt 60 ngày nuôi cấy
(không trình bày số liệu). Ngược lại, các nghiệm thức bổ sung NAA (1; 2; 3; 4; 5; 6
mg/L) vào môi trường nuôi cấy đều có cảm ứng tạo phôi vô tính theo hướng tích cực
(Hình 3.2). Điều đó chứng tỏ, sự cảm ứng tạo phôi của mảnh lá NGBCC đối với mỗi
loại auxin trong môi trường nuôi cấy là khác nhau và cần có loại auxin ngoại sinh
thích hợp là NAA, còn IBA hoàn toàn không thích hợp cho sự cảm ứng tạo phôi.
Kết quả (Bảng 3.1) cho thấy, tỷ lệ mẫu tạo phôi và số phôi/mẫu tăng khi tăng
nồng độ NAA (1 – 5 mg/L) trong môi trường nuôi cấy, khi tăng nồng độ 6 mg/L NAA
thì tỷ lệ mẫu tạo phôi và số phôi/mẫu đều giảm. Khi xét các mức nồng độ NAA và
các loại môi trường khoáng, nhận thấy môi trường ½MS có bổ sung 1 mg/L NAA có
tỷ lệ mẫu tạo phôi (%) và số phôi/mẫu thấp nhất (24,44% và 6,18 phôi/mẫu). Các chỉ
tiêu này đạt giá trị cao nhất khi môi trường SH có bổ sung 5 mg/L NAA với tỷ lệ mẫu
tạo phôi 88,89% và 19,95 phôi/mẫu (Hình 3.2L). Kết quả này có ý nghĩa khác biệt về
thống kê so với tất cả các nghiệm thức còn lại. Điều đó chứng tỏ, tỷ lệ mẫu tạo phôi
(%) và số phôi/mẫu chịu ảnh hưởng bởi các mức nồng độ của loại auxin thích hợp
(NAA) và loại môi trường khoáng phù hợp. Như vậy, nồng độ 5 mg/L NAA là tối ưu
để kích thích tạo phôi đạt hiệu quả cao nhất từ mảnh lá NGBCC, trong môi trường
nuôi cấy có hàm lượng và thành phần khoáng thích hợp là SH.
Sự cảm ứng phát sinh phôi từ mô lá NGBCC chỉ ở những nghiệm thức có bổ
sung NAA, số phôi phát sinh ít hoặc nhiều tùy thuộc nồng độ NAA sử dụng, điều này
cho thấy vai trò của auxin ngoại sinh là mang tính quyết định. Qua vết thương của
mẫu cấy làm tăng sự hấp thụ auxin ngoại sinh, sự hiện diện của auxin ngoại sinh
(NAA) trong tế bào làm tăng IAA nội sinh thông qua quá trình tổng hợp IAA nội

54
sinh, sự gia tăng của IAA nội sinh không thể thiếu để thay đổi chương trình di truyền
dẫn đến phát sinh phôi, sự cân bằng nội môi của các auxin rất quan trọng cho cảm
ứng tạo phôi [90]. Sự khác nhau về loại và nồng độ auxin ngoại sinh áp dụng trong
mối tương tác với sinh tổng hợp auxin nội sinh có vai trò quan trọng trong phát sinh
phôi vô tính [88][89]. Vai trò của auxin ngoại sinh và sự vận chuyển phân cực của
auxin, đóng vai trò quan trọng trong sự phát sinh phôi vô tính, điều này được Verma
và cộng sự (2018) chứng minh trong nghiên cứu tác động của auxin ngoại sinh đến
phát sinh phôi vô tính trực tiếp của Digitalis trojana [14].
Như vậy, auxin trong đó có NAA là yếu tố truyền tín hiệu cảm ứng phát sinh
phôi, có vai trò quan trọng kênh thông tin tế bào - làm tế bào biến đổi đến trạng thái
cuối cùng đã được chương trình hóa là phát sinh phôi vô tính [65]. Tuy nhiên khả
năng đáp ứng tạo phôi của mô thực vật khác nhau đối với các loại auxin trong môi
trường là khác nhau [70].
Môi trường nuôi cấy là một trong những yếu tố quan trọng nhất trong nuôi cấy
in vitro, chứa thành phần và hàm lượng các chất khoáng cần thiết cho sự phát sinh
phôi. Trong thí nghiệm này, sử dụng các môi trường khoáng thường sử dụng trong
nghiên cứu phát sinh phôi vô tính ở họ Ngũ gia bì là MS, B5, SH [72][71][38] và
½MS. Kết quả sau 60 ngày nuôi cấy, phôi phát sinh cao nhất ở môi trường SH và
thấp nhất ở môi trường ½MS, điều đó chứng tỏ thành phần và hàm lượng chất khoáng
có tác động đến sự phát sinh phôi và môi trường khoáng SH thích hợp nhất cho cảm
ứng tạo phôi trực tiếp từ mô lá NGBCC.
Như vậy, hiệu quả phát sinh phôi chịu ảnh hưởng bởi loại, nồng độ auxin thích
hợp, hàm lượng và thành phần khoáng của môi trường nuôi cấy, trong đó auxin có
vai trò quyết định. Trong nghiên cứu này, tỷ lệ % mẫu tạo phôi và số phôi/mẫu nhiều
nhất ở môi trường nuôi cấy SH có 5 mg/L NAA. Đây là nghiên cứu đầu tiên thực
hiện thành công tạo phôi vô tính trực tiếp từ mảnh lá NGBCC.
Thí nghiệm này, 2,4-D cũng đã được bố trí thực hiện trên NGBCC nhưng với
kết quả âm tính trong tạo phôi, chỉ ghi nhận được hiện tượng hình thành mô sẹo.
Theo một số tác giả, kiểu gen thực vật [109], loại môi trường khoáng cơ bản
[110], một số thành phần khoáng đơn lẻ [111], công thức môi trường nuôi cấy bao
gồm chất ĐHST,..[112][70] có ảnh hưởng quan trọng đến sự hình thành phôi vô tính
trực tiếp/gián tiếp.

55
Bảng 3.1. Ảnh hưởng của NAA và môi trường khoáng đến sự tạo phôi vô tính
trực tiếp từ mô lá ở 60 NSC.
NAA
(mg/L)
Môi trường
khoáng
Tỷ lệ mẫu tạo
phôi (%)
Số phôi/mẫu
0 ½MS 0.00q* 0,00r
1 ½MS 24,44p 6,18v
2 ½MS 33,33o 6,29u
3 ½MS 42,22n 7,02s
4 ½MS 52,22l 7,74n
5 ½MS 56,67k 8,71k
6 ½MS 46,67m 7,16p
0 MS 0.00q 0,00r
1 MS 57,78k 6,58t
2 MS 61,11j 7,29p
3 MS 66,67gh 8,67k
4 MS 75,56c 10,00h
5 MS 78,89b 11,69e
6 MS 68,89f 9,07j
0 B5 0.00q 0,00r
1 B5 47,78m 7,11st
2 B5 51,11l 7,47p
3 B5 56,67k 8,36l
4 B5 65,56hi 9,19i
5 B5 73,33d 12,48d
6 B5 64,44i 10,23g
0 SH 0.00q 0,00r
1 SH 60.00j 7,89m
2 SH 67,78fg 9,11ij
3 SH 71,11e 11,27f
4 SH 78,89b 15,24b
5 SH 88,89a 19,95a
6 SH 74,44cd 13,57c
*Các chữ cái khác nhau trong một cột thể hiện sự khác biệt có ý nghĩa ở mức
p ≤ 0,05 trong phép thử Duncan. Số liệu (%) được chuyển đổi sang dạng arcsin √𝑥
khi phân tích thống kê.

56
Hình 3.2. Mảnh lá tạo phôi ở môi trường ½MS, MS, B5, SH có NAA, 60 NSC.
A,B,C,D. Môi trường ½MS, MS, B5, SH có 3 mg/L NAA; E,F,G,H. Môi trường
½MS, MS, B5, SH có 4 mg/L NAA; I,J,K,L. Môi trường ½MS, MS, B5, SH có 5
mg/L NAA; thanh ngang 10 mm.
Khác với cảm ứng tạo phôi trực tiếp, quá trình tạo phôi gián tiếp có thể gây ra
biến dị dòng soma ở chuỗi DNA do trải qua giai đoạn mô sẹo [113]. Do vậy, theo
chúng tôi tạo phôi trực tiếp rất có ý nghĩa trong nhân giống cây trồng – nghiên cứu
dựa trên nguyên tắc đồng nhất về mặt di truyền.
Quan sát cấu trúc giải phẫu phôi
Khảo sát cấu trúc giải phẫu phôi bằng phương pháp nhuộm hai màu đối với 03
thể phôi (cầu, trái tim, thủy lôi) đã được thực hiện. Quan sát dưới kính hiển vi, đã ghi
nhận được lớp tế bào biểu bì (epidermis) đặc trưng ở cả 03 dạng phôi, và sự hiện diện
của mạch dẫn ở cấu trúc phôi dạng thủy lôi (Hình 3.3).

57
Hình 3.3. Các giai đoạn phát triển của phôi vô tính và hình thái giải phẫu phôi
tương ứng.
A,B,C. Dạng phôi cầu, phôi tim, phôi thủy lôi; D,E,F. Hình thái giải phẫu tương ứng
dạng phôi cầu, phôi tim, phôi thủy lôi, (LM: Lá mầm, BB: Biểu bì, M: Mạch; thanh
ngang 1 mm).
3.1.1.2. Ảnh hưởng của sự kết hợp NAA và BA đến sự tạo phôi vô tính trực tiếp từ
mô lá
Phân tích số liệu (Bảng 3.2) cho thấy, môi trường nuôi cấy có NAA được bổ
sung BA với các mức nồng độ (0,25; 0,5; 1 mg/L) thì kết quả tạo phôi có khác biệt
về thống kê giữa các nghiệm thức. Điều đó chứng tỏ, sự kết hợp của NAA và BA có
ảnh hưởng đến sự phát sinh phôi. Nhận thấy, khi sử dụng BA (0,25; 0,5; mg/L) kết
hợp với NAA (3, 4, 5 mg/L) mô lá có biểu hiện đáp ứng tích cực, các chỉ tiêu theo
dõi đều cao hơn khi môi trường nuôi cấy bổ sung auxin (NAA) riêng lẻ, không kết
hợp với BA, cụ thể tỷ lệ mẫu cấy tạo phôi và số phôi/mẫu tăng, cao nhất ở nghiệm
thức có 5 mg/L NAA kết hợp với 0,25 mg/L BA (tỷ lệ mẫu tạo phôi là 94,44% và
28,56 phôi/mẫu), cao hơn nghiệm thức đối chứng 5 mg/L NAA (tỷ lệ mẫu tạo phôi
88,89% và 19,95 phôi/mẫu). Nhưng các nghiệm thức có NAA (3, 4, 5 mg/L) khi bổ
sung 1 mg/L BA, thì mẫu cấy tạo phôi chậm, tỷ lệ tạo phôi và số phôi/mẫu đều giảm,
thấp nhất ở nghiệm thức có 5 mg/L NAA và 1 mg/L BA (tỷ lệ mẫu tạo phôi 57,78%
và 9,67 phôi/mẫu). Như vậy, khi bổ sung BA với nồng độ tối ưu đã phát huy vai trò
của NAA trong quá trình phân bào và phát sinh phôi vô tính trực tiếp với hiệu quả

58
cao nhất. Từ kết quả trên cho thấy, sự hiện diện của auxin (NAA) kết hợp với
cytokinin (BA) là cần thiết cho quá trình phát sinh phôi NGBCC, sự tương tác này có
tác động tích cực và hiệu quả khi nồng độ của chất ĐHST thích hợp (Hình 3.4D,E).
Bảng 3.2. Ảnh hưởng của kết hợp NAA và BA đến sự tạo phôi vô tính trực tiếp từ
mảnh lá, ở môi trường SH, 60 NSC.
NAA
(mg/L)
BA
(mg/L)
Tỷ lệ mẫu
tạo phôi (%)
Số phôi/mẫu
3 0 71,11g* 11,27h
4 0 78,89e 15,24e
5 0 88,89c 19,95b
3 0,25 75,56f 15,78e
4 0,25 82,22d 18,83c
5 0,25 94,44a 28,56a
3 0,5 72,22g 12,67g
4 0,5 80,00e 18,00d
5 0,5 91,11b 20,67b
3 1 67,78h 10,67i
4 1 65,56i 14,33f
5 1 57,78j 9,67j
*Các chữ cái khác nhau trong một cột thể hiện sự khác biệt có ý nghĩa ở mức
p ≤ 0,05 trong phép thử Duncan. Số liệu (%) được chuyển đổi sang dạng arcsin √𝑥
khi phân tích thống kê.
Trong nuôi cấy in vitro, đa số các thực vật (có khuynh hướng tái sinh phôi)
đều có nhu cầu auxin và cytokinin nhằm đáp ứng tạo phôi vô tính. Auxin ngoại
sinh/bổ sung giúp điều hòa và cân bằng lượng auxin nội sinh [70], cytokinin cùng với
auxin kích thích sự phân bào, và cả hai có tác động cộng hưởng đối với nhiều quá
trình sinh học ở thực vật và mô nuôi cấy, do vậy cũng có vai trò quan trọng trong tạo
phôi [82]. Ở NGBCC, tổ hợp NAA và BA ở nồng độ thích hợp đã cho kết quả tạo
phôi tích cực - phù hợp với nhận định chung về vai trò kết hợp của auxin và cytokinin
đối với tạo phôi như đã nêu trên.

59
Hình 3.4. Mảnh lá tạo phôi trực tiếp ở môi trường SH có NAA và BA, 60 NSC.
A,B,C. Môi trường SH có 4 mg/L NAA và BA 0,25; 0,5; 1 mg/L; D,E,F. Môi trường
SH có 5 mg/L NAA và BA 0,25; 0,5; 1 mg/L. Thanh ngang 5 mm.
Kết quả tác động tích cực của BA đối với sự hình thành phôi NGBCC ở nghiên
cứu này phù hợp với kết quả nghiên cứu của Kharwanlang và cộng sự (2016a) khi bổ
sung thêm kinetin/BA (0,25 – 5 mg/L) vào môi trường dùng tạo phôi Panax
pseudoginseng [71]. Trong nuôi cấy in vitro, đa số các thực vật (có khuynh hướng tái
sinh phôi) đều có nhu cầu auxin và cytokinin nhằm đáp ứng tạo phôi vô tính. Auxin
ngoại sinh/bổ sung giúp điều hòa và cân bằng lượng auxin nội sinh [70], cytokinin
cùng với auxin kích thích sự phân bào và cả hai có tác động cộng hưởng đối với nhiều
quá trình sinh học ở thực vật và mô nuôi cấy và do vậy cũng có vai trò quan trọng
trong tạo phôi [82]. Từ nuôi cấy mảnh lá mầm phôi hợp tử Cucumis sativa trên môi
trường có BA ở điều kiện chiếu sáng, Li và cộng sự (2007) nhận thấy BA, ngoài tác
động làm tăng kích thước mảnh lá mầm cũng đã làm tăng hàm lượng cytokinin nội
sinh [5]. Qua nghiên cứu mối liên quan giữa hàm lượng chất ĐHST nội sinh với các
giai đoạn phát triển khác nhau của phôi vô tính từ nuôi cấy mô phôi hợp tử Ormosia
henryi, Gaoyin và cộng sự (2020) [12] nhận thấy hàm lượng auxin đạt cao ở giai
đoạn phát triển và sự phân cực của phôi (tỷ lệ IAA/CKs, AUX/CKs cao), và hàm

60
lượng cytokinin cũng cao ở giai đoạn trưởng thành và nẩy mầm của phôi (tỷ lệ
IAA/CKs, AUX/CKs giảm).
Như vậy, chất ĐHST ngoại sinh có vai trò không thể thay thế trong cảm ứng
tạo phôi vô tính và nồng độ của chúng ảnh hưởng đến sự cân bằng chất ĐHST nội
sinh trong đó có cytokinin, giúp điều hòa sự tăng trưởng và phát sinh hình thái. Ở
NGBCC, tổ hợp NAA và BA ở nồng độ thích hợp đã cho kết quả tạo phôi tích cực -
phù hợp với nhận định chung về vai trò kết hợp của auxin và cytokinin đối với tạo
phôi như đã nêu trên.
3.1.1.3. Ảnh hưởng của nồng độ đường và điều kiện chiếu sáng đến sự tạo phôi vô
tính trực tiếp từ mô lá
Kết quả sau 60 ngày nuôi cấy (Bảng 3.3, Hình 3.5) cho thấy, nồng độ sucrose
50 g/L ở trường hợp chiếu sáng đã có ảnh hưởng tích cực nhất đến sự phát sinh phôi
vô tính (Hình 3.5C) so với các trường hợp nồng độ sucrose còn lại và so với trường
hợp để mẫu trong tối, tỷ lệ mẫu tạo phôi và số phôi/mẫu lần lượt là 98,89% và 35,95
phôi/mẫu. Khi nồng độ sucrose cao đã làm tăng khả năng giữ nước của môi trường,
tế bào nhận nước từ môi trường kém thuận lợi - mô khô hơn do vậy kích thích sinh
lý tế bào biến đổi mạnh, tạo thuận lợi cho quá trình phát sinh phôi. Khi nồng độ
sucrose ở môi trường nuôi cấy tăng cao 70 g/L làm tỷ lệ mẫu tạo phôi và số phôi/mẫu
giảm (tương ứng là 87,78% và 18,72 phôi/mẫu), do áp suất thẩm thấu tăng cao nên tế
bào bị ‘sốc’ thiếu nước, sinh lý tế bào bị ức chế, trao đổi chất giảm. Khi nồng độ
sucrose giảm còn 10 g/L thì quá trình tạo phôi giảm rõ rệt, phát sinh hình thái chậm
dù vẫn ở điều kiện sáng. Nghiệm thức có 50 g/L sucrose ở điều kiện tối không khác
biệt về mặt thống kê so với nghiệm thức 10 g/L sucrose ở điều kiện sáng khi xét về
tỷ lệ % mẫu tạo phôi, nhưng số phôi/mẫu có khác biệt rõ. Như vậy, với các mức nồng
độ sucrose khác nhau trong điều kiện chiếu sáng khác nhau đã có ảnh hưởng khác
biệt lên hiệu quả phát sinh phôi vô tính trực tiếp từ mô lá NGBCC, ở điều kiện chiếu
sáng phát sinh phôi hiệu quả hơn trong điều kiện tối hoàn toàn, vì ở điều kiện chiếu
sáng có ảnh hưởng đến quang hợp của mẫu cấy.
Trong điều kiện nuôi cấy in vitro, mô và tế bào thực vật sống chủ yếu theo
phương thức dị dưỡng hoặc bán dị dưỡng nhờ điều kiện ánh sáng nhân tạo và lục lạp
có khả năng quang hợp, mẫu cấy chủ yếu sử dụng dinh dưỡng từ môi trường nuôi
cấy. Vì vậy, việc bổ sung đường vào môi trường nuôi cấy và bố trí mẫu cấy ở những

61
điều kiện chiếu sáng khác nhau có ý nghĩa rất lớn cho sự phát sinh hình thái, phát
sinh phôi. Đường đối với cơ thể thực vật là yếu tố không thể thiếu trong cung cấp
năng lượng và khung carbon cho sự phát triển của tế bào. Đường còn giúp điều hòa
áp suất thẩm thấu, bảo vệ tính toàn vẹn của màng tế bào, quyết định hình dạng và sự
phát triển của tế bào [12].
Bảng 3.3. Ảnh hưởng của nồng độ đường và điều kiện chiếu sáng đến sự tạo phôi vô
tính trực tiếp từ mô lá, ở môi trường SH, 60 NSC
Điều kiện
nuôi cấy
Nồng độ
sucrose (g/L)
Tỷ lệ mẫu tạo
phôi (%)
Số phôi/mẫu
Sáng 10 91,11c* 23,19c
30 94,44b 28,56b
50 98,89a 35,95a
70 87,78d 18,72d
Tối 10 78,89f 10,08g
30 82,22e 11,28f
50 90,00cd 15,93e
70 71,11g 9,72g
*Các chữ cái khác nhau trong một cột thể hiện sự khác biệt có ý nghĩa ở
mức p ≤ 0, 05 trong phép thử Duncan. Số liệu (%) được chuyển đổi sang dạng
(x+0,5)1/2 khi xử lý thống kê.
Đã ghi nhận được công trình công bố đường ở nồng độ 50 g/L thúc đẩy khả
năng phát sinh phôi vô tính từ mô sẹo phôi hợp tử cây dược liệu quan trọng
Terminalia chebula [114]. Nồng độ đường 50 g/L là nồng độ tối ưu cho sự phát sinh
phôi vô tính sâm Ngọc Linh (Panax vietnamensis) [74]. Ở họ Ngũ gia bì, nhiều kết
quả nghiên cứu cho thấy đường được sử dụng như vừa là tác nhân tạo áp suất thẩm
thấu cao và chất dinh dưỡng đã tạo ảnh hưởng rất tích cực lên quá trình tạo phôi, ví
dụ [36] đã cải thiện được chất lượng và tần số tạo phôi Panax quinquefolius trực tiếp
qua kết hợp tiền xử lý (pretreatment) mẫu lá mầm với tác nhân đường (342,3 g/L, ở
4oC) với nuôi cấy mẫu sau đó trên môi trường cảm ứng tạo phôi có nồng độ đường
cao (70 g/L); ngoài ra, Kim và cộng sự (2010) cũng đã ghi nhận tần số tạo phôi đơn
Panax ginseng trực tiếp (ở 2 giống Yun-Poong và Chun-Poong) cao nhất khi mảnh
lá mầm phôi non được nuôi cấy trên môi trường MS có 70 g/L đường và tần số tạo

62
phôi đơn tăng cao khi xử lý gây co nguyên sinh (plasmolysis) mẫu bằng đường (0,1
– 0,5 – 1 M, trong 24 h) trước khi cấy mẫu vào môi trường [115].
Hình 3.5. Phôi vô tính phát sinh trực tiếp từ mảnh lá ở môi trường SH, có đường và
điều kiện sáng, tối, 60 NSC
A,B,C,D. Phôi hình thành ở môi trường SH có sucrose 10, 30, 50 và 70 g/L, điều kiện
sáng. E,F,G,H. Phôi hình thành ở môi trường SH có sucrose 10, 30, 50 và 70 g/L,
điều kiện tối. Thanh ngang 10 mm.
3.1.1.4. Ảnh hưởng của tỷ lệ nước dừa đến sự tạo phôi vô tính trực tiếp từ mô lá
Qua tối ưu hóa các yếu tố đã khảo sát, môi trường SH có 5 mg/L NAA và 0,25
mg/L BA, 50 g/L đường được sử dụng làm môi trường nuôi cấy để xác định nồng độ
thích hợp của nước dừa (%) cho sự cảm ứng tạo phôi vô tính từ mảnh lá nuôi ở điều
kiện chiếu sáng ở 4000 lux.
Kết quả cho thấy ở 60 NSC, nghiệm thức có tỷ lệ nước dừa 10% đã có ảnh
hưởng tích cực nhất so với các nồng độ còn lại, tất cả mẫu cấy đều tạo phôi, với 39,50
phôi/mẫu (Bảng 3.4; Hình 3.6), nghiệm thức có tỷ lệ nước dừa 5% và 10% không có
khác biệt thống kê về chỉ tiêu số mẫu tạo phôi nhưng có sự khác biệt về số phôi/mẫu.
Các mẫu cấy ở môi trường có tỷ lệ nước dừa cao 15%, 20% làm giảm khả năng tạo
phôi vô tính (tỷ lệ mẫu tạo phôi là 95,56%, 94,44%), số phôi/mẫu giảm rõ rệt ở
nghiệm thức có 20% nước dừa (29,78 phôi/mẫu). Ở nghiệm thức đối chứng không bổ
sung nước dừa và nghiệm thức có tỷ lệ nước dừa 15% không khác biệt về thống kê
về số phôi/mẫu nhưng khác biệt rõ về tỷ lệ mẫu tạo phôi. Như vậy, khi bổ sung nước
dừa với các tỷ lệ khác nhau đã có tác động rõ rệt đến kết quả tạo phôi vô tính.

63
Bảng 3.4. Ảnh hưởng của tỷ lệ nước dừa đến sự tạo phôi vô tính trực tiếp từ mảnh lá,
ở môi trường SH, 60 NSC.
Tỷ lệ nước
dừa (%)
Tỷ lệ mẫu tạo
phôi (%)
Số phôi/mẫu
0 98,89b* 35,95c
5 100,00a 38,19b
10 100,00a 39,50a
15 95,56c 35,19c
20 94,44d 29,78d
*Các chữ cái khác nhau trong một cột thể hiện sự khác biệt có ý nghĩa ở
mức p ≤ 0, 05 trong phép thử LSD. Số liệu (%) được chuyển đổi sang dạng
(x+0,5)1/2 khi xử lý thống kê.
Theo Prades và cộng sự (2012) [116], trong nước dừa xanh có chứa đường,
vitamin, muối khoáng, acid amin và một số phytohormone quan trọng nhất là
cytokinin. Cũng theo các tác giả trên, IAA tham gia vào nhiều quá trình liên quan đến
tăng trưởng và phát triển của thực vật, sự di chuyển phân cực của auxin tạo lượng
auxin cực đại (auxin maximum)/gradient về nồng độ nhằm điều hòa nhiều quá trình
sinh lý dẫn đến sự phát sinh hình thái trong đó có phát sinh phôi vô tính. Trong nước
dừa cũng có nhiều loại cytokinin khác nhau như trans-zeatin, trans-zeatin riboside-
5’- monophosphate, kinetin riboside, kinetin, N6-isopentenyladenine,...- có tác dụng
trong phân chia tế bào, làm chậm sự lão hóa, tăng khả năng biến dưỡng,.. và như vậy
cũng kích thích tăng trưởng thực vật. Do đó, khi bổ sung nước dừa vào môi trường
với lượng phù hợp sẽ làm tăng khả năng tạo và phát triển phôi, chồi/cây. Khierallah
và Hussein (2013) đã sử dụng nước dừa trong nghiên cứu và kết luận nước dừa 20%
(v/v) rất thích hợp cho tạo phôi vô tính Phoenix dactylifera (date palm) [77]. Ở họ
Ngũ gia bì trong đó có loài NGBCC đã ghi nhận được ảnh hưởng tích cực của nước
dừa (10%) lên quá trình hình thành phôi từ mô sẹo/mô sẹo có khả năng sinh phôi sâm
Ngọc Linh - Panax vietnamensis [117].

64
Hình 3.6. Phôi vô tính phát sinh trực tiếp từ mảnh lá ở môi trường SH, có bổ sung
nước dừa, 60 NSC.
A,B,C,D. Mảnh lá tạo phôi ở môi trường SH có nước dừa 5%, 10%, 15%, 20%; E.
Đĩa cấy mảnh lá tạo phôi trên môi trường SH có 10% nước dừa. Thanh ngang 10 mm.
3.1.1.5. Tạo cây con hoàn chỉnh từ phôi vô tính
Trong quá trình nuôi cấy tái sinh, trước và sau giai đoạn trưởng thành hầu hết
các phôi tiếp tục sinh trưởng dẫn đến phát triển thân, lá mầm, rễ mầm, chồi/lá bình
thường (Hình 3.7A). Ngược lại, bên cạnh dạng phôi vô tính hình thái tiêu biểu cũng
đã ghi nhận được trường hợp phôi tái sinh (với số lượng không đáng kể) có hình dạng
bất thường như phôi dính nhau ở phần lá mầm/thân, không có rễ mầm, chồi không
phát triển (Hình 3.7B,C).
Hình 3.7. Phôi bình thường và một số dạng bất thường của phôi vô tính
A. Hình thái phôi bình thường; B. Hình thái phôi có lá mầm dị dạng, rễ mầm phát
triển bình thường; C. Các phôi dính nhau với lá mầm dị dạng, có chung phần thân và
không có rễ mầm. Thanh ngang 5 mm.
Sự phân chia tế bào ở vùng mô phân sinh xảy ra trước khi có sự biệt hóa của
cực chồi và lá mầm, nếu sự phân bào vẫn tiếp tục sau khi hình thành sơ khởi lá mầm,

65
dẫn đến hình thành phôi bất thường có hai hay nhiều phôi dính lại với nhau (Hình
3.7C). Miekle và cộng sự, (1995) cho rằng, các lá mầm hợp nhất như vậy cũng là đặc
điểm của phôi soma dạng sừng thường được phục hồi từ nuôi cấy tế bào, có thể là
auxin ngoại sinh hiện diện trong môi trường có thể đủ làm nhiễu loạn sự vận chuyển
bình thường của auxin trong phôi ở giai đoạn hình cầu, làm cho sự phát triển lá mầm
bình thường bị ngăn cản [108].
Phôi phát triển với 2 lá thật nhỏ ban đầu (Hình 3.8A) được nuôi cấy trên môi
trường MS và ½MS [54] không bổ sung chất ĐHST để so sánh khả năng tạo cây hoàn
chỉnh. Kết quả cho thấy, cây con ở hai môi trường này phát triển khá khác biệt ở 90
NSC. Chiều cao cây, số lá, số rễ và chiều dài rễ trung bình của cây ở môi trường
½MS, MS lần lượt là 2,63 cm; 5,2; 7,3; 9,6 cm; là 2,58 cm; 4,9; 6,5 và 6,9 cm, theo
thứ tự (không trình bày bảng số liệu). Như vậy, môi trường ½MS là thích hợp cho
việc tạo cây con hoàn chỉnh từ phôi vô tính (Hình 3.8C). Điểm đặc biệt là các lá thật
hình thành ban đầu ở cây con đều là các lá đơn, sau đó mới tạo các lá kép có thùy (lá
đơn) đặc trưng (Hình 3.8B,C).
Hình 3.8. Tạo cây con từ phôi vô tính trên môi trường MS, ½MS không có chất
ĐHST.
A. Vật liệu phôi dùng thí nghiệm tạo cây con; B,C. Cây con từ phôi nuôi cấy trên
môi trường MS, ½MS.
Điểm đặc biệt ở NGBCC là phôi có thể tạo chồi khá thuận lợi trên môi trường
không bổ sung chất ĐHST; theo chúng tôi, có thể do phôi không ở trạng thái ngủ/hưu
miên (dormancy) như ở một số trường hợp khác như phôi các dòng Panax ginseng
đột biến [41] và dòng lai khác loài giữa Panax ginseng và Panax quinquefolius [118]
- đều cần được nuôi cấy trên môi trường có GA3 (5 mg/L); và cũng cần được nuôi
trên môi trường có bổ sung BA (1 mg/L), NAA (0,2 mg/L) đối với Coriandrum
sativum [119]. Qua thử nghiệm nuôi cấy ban đầu 200 phôi, ghi nhận được số phôi tạo
chồi (sau ~ 3 tháng nuôi) là 196 (tỷ lệ 98%). Theo Sliwinska và cộng sự (2008), môi

66
trường ½MS không bổ sung chất ĐHST là thích hợp cho sự tạo cây từ phôi chỉ trong
một lần nuôi cấy đối với Đinh lăng Polyscias filicifolia [54].
3.1.1.6. Trồng cây con ở chậu đất
Hình 3.9. Trồng cây từ phôi vô tính ra chậu đất ở vườn ươm
A. Bình cây chuẩn bị đưa ra trồng ở vườn ươm, ở 90 NSC; B,C. Cây từ phôi ở chậu
đất vườn ươm sau 3 - 6 tháng trồng. D. Cây sau 12 tháng trồng.
Các cây con (nuôi trong bình tam giác chứa môi trường ½MS) (Hình 3.9A)
được đem trồng ra chậu chậu đất, kết quả cho thấy các lá mầm dần thoái hóa, cây có
tỷ lệ sống rất cao (≥ 95%) tính đến thời điểm 3 tuần sau trồng và cây có sinh
trưởng/kiểu hình bình thường trong điều kiện vườn ươm; số lá chét ban đầu thường
thấp hơn so với cây mẹ, tăng dần ở thời gian sau đó (Hình 3.9B,C).
3.1.2. Tạo phôi vô tính gián tiếp
3.1.2.1. Tạo phôi vô tính gián tiếp qua mô sẹo mảnh lá (10 x 10 mm)
Tạo mô sẹo
Hình 3.10. Ảnh hưởng của cách cấy mảnh lá đến hiệu quả tạo mô sẹo.
A. Cấy mặt trên mảnh lá tiếp xúc với môi trường (a,c), cấy mặt dưới mảnh lá tiếp xúc
với môi trường (b); B. Mảnh lá nuôi cấy trên môi trường SH không có 2,4-D.
Trước khi bố trí thí nghiệm tạo mô sẹo, tiến hành thử nghiệm ảnh hưởng của
02 cách cấy mảnh lá (trên môi trường SH có 2 mg/L 2,4-D) đến hiệu quả tạo mô sẹo
đã được thực hiện; một là, cấy mặt trên mảnh lá (có màu xanh đậm, sáng bóng) tiếp

67
xúc với môi trường; hai là, cấy mặt dưới mảnh lá (màu xanh nhạt, mờ) trên mặt môi
trường.
Kết quả cho thấy cách cấy thứ nhất cho kết quả tạo nhiều mô sẹo hơn ở mép
cắt (Hình 3.10Aa,c) so với cách cấy thứ hai (Hình 3.10Ab), có thể do mặt trên lá tiếp
xúc với môi trường nên sự hấp thu chất dinh dưỡng ở môi trường tốt hơn, đồng thời
mặt dưới lá có nhiều khí khổng, sự trao đổi khí thuận lợi hơn. Không ghi nhận sự
hình thành mô sẹo trường hợp nuôi cấy mảnh lá trên môi trường không bổ sung 2,4-
D (Hình 3.10B). Do vậy, cách đặt mẫu mặt trên lá tiếp xúc với môi trường được sử
dụng cho nghiên cứu tiếp theo.
Hình 3.11. Tạo mô sẹo từ mảnh lá (10 x 10 mm) nuôi cấy trên môi trường SH.
A,B,C. Mảnh lá tạo mô sẹo ở môi trường có 2,4-D 1, 2, 3 mg/L, 20 NSC; D,E,F.
Mảnh lá tạo mô sẹo (và rễ) ở môi trường có 2,4-D 1, 2, 3 mg/L, 40 NSC; G,H,I. Cụm
mô sẹo (sau bỏ rễ, cấy chuyền) trên môi trường có 2,4-D 1, 2, 3 mg/L, 60 NSC (thanh
ngang 10 mm).
Đã ghi nhận nhiều công bố kết quả nghiên cứu như tạo mô sẹo Coffea arabica
[120], tạo phôi Coffea arabica và Coffea canephora [121], và kể cả tạo chồi Petunia
hybrida [122] và Sinningia speciosa [123] từ mảnh lá sử dụng thao tác cấy mẫu theo
cách này. Ngược lại, cấy đặt mặt dưới mảnh lá Malus sieversii tiếp xúc với môi trường
lại cho kết quả tạo mô sẹo và tái sinh chồi tốt hơn [124]. Như vậy, cách cấy phụ thuộc
vào loài thực vật nghiên cứu và ở NGBCC, cấy mặt trên mảnh lá tiếp xúc với môi

68
trường đã tạo ảnh hưởng tích cực đến quá trình tạo mô sẹo và nhìn chung phù hợp
với các kết quả công bố chung như đã nêu trên.
Kết quả thí nghiệm tạo mô sẹo cho thấy, ở 20 ngày sau cấy, mô cấy đã tăng
kích thước, mô sẹo hình thành ở xung quanh mép cắt của tất cả các mẫu cấy mảnh lá
trên môi trường có bổ sung 2,4-D (tỷ lệ 100%); hiệu quả cao nhất ở 3 mg/L 2,4-D, ít
nhất ở nghiệm thức có 1 mg/L 2,4-D (Hình 3.11A,B,C). Ngày thứ 40 sau nuôi cấy,
nhận thấy các mẫu cấy hình thành khá nhiều rễ ngắn có nhiều lông hút, đầu rễ tròn,
nhiều nhất ở 1 mg/L 2,4-D (Hình 3.11D,E,F). Mẫu cấy sau khi cắt bỏ rễ, cấy chuyền
vào môi trường tạo mô sẹo trong 20 ngày tiếp theo, kết quả cho thấy mô sẹo hình
thành trên khắp bề mặt của mảnh lá, hình thành nhiều ở các mẫu cấy trên môi trường
có 2,4-D ở nồng độ 2 và 3 mg/L), mô sẹo phát triển kém ở 1 mg/L 2,4-D. Các mẫu
cấy ở 2 mg/L 2,4-D, ghi nhận có sự hình thành một số cụm mô hơi cứng, màu hơi
vàng/trắng tương tự mô sẹo có KNSP, (Hình 3.12H); mô sẹo xốp hơn ở 3 mg/L 2,4-
D (Hình 3.12I). Như vậy, việc cắt bỏ rễ đã tạo điều kiện thuận lợi cho sự hình thành
mô sẹo.
Bảng 3.5. Ảnh hưởng của 2,4-D đến sự hình thành mô sẹo có KNSP, ở môi trường
SH, 30NSC.
2,4-D
(mg/L)
Tỷ lệ (%) mẫu tạo
mô sẹo có KNSP
Số cụm mô sẹo có
KNSP/mẫu
1 100 2,20b*
2 100 3,43a
3 100 1,87c
*Các chữ cái khác nhau trong một cột thể hiện sự khác biệt có ý nghĩa
ở mức p ≤ 0,05 trong phép thử LSD.
Theo tài liệu, đối với cây một và hai lá mầm, 2,4-D có tác dụng rất hiệu quả
đến sự hình thành mô sẹo từ nhiều loại vật liệu nuôi cấy khác nhau như mảnh lá/lá
mầm, đoạn thân/trụ mầm/rễ, phôi non,… ở nhiều đối tượng thực vật khác nhau
[125][126]. Tương tự, đối với một số loài thuộc họ Ngũ gia bì [127], mô sẹo hình
thành có nhiều dạng khác nhau như xốp/cứng và màu sắc khác nhau như trắng
đục/trắng trong, hơi vàng/vàng. Ở NGBCC khoảng dao động nồng độ 2,4-D để tạo
mô sẹo, nhận được với các dạng mô sẹo và màu sắc khác nhau phù hợp với kết quả
nghiên cứu chung ở lĩnh vực này.

69
Sau 30 ngày cấy các mảnh mô sẹo để tạo mô sẹo có KNSP, kết quả cho thấy
có sự hình thành các cụm mô sẹo hơi cứng, màu vàng sáng/hơi đục hình thành trên
bề mặt cụm mô sẹo nuôi cấy, đây là các cụm mô sẹo có KNSP, số lượng cụm mô sẹo
hình thành khác nhau tùy thuộc nồng độ 2,4-D nhiều nhất ở 2 mg/L 2,4-D (3,43), thấp
nhất ở 3 mg/L 2,4-D (1,87) (Bảng 3.5, Hình 3.12A,B,C).
Hình 3.12. Ảnh hưởng của 2,4-D đến sự hình thành mô sẹo có KNSP, ở môi trường
SH, 30 NSC.
A,B,C. Cụm mô sẹo có KNSP (vị trí mũi tên) hình thành trên môi trường có 2,4-D
1; 2; 3 mg/L (thanh ngang 10 mm)
Trong nghiên cứu tạo/duy trì/phát triển sinh khối mô sẹo có khả năng sinh phôi
ở nhiều loài thực vật thì auxin ở dạng riêng lẻ hoặc kết hợp với cytokinin là yếu tố
mang tính quyết định, và loại mô có khả năng tái sinh này có vai trò rất quan trọng
đối với nghiên cứu tái sinh cây theo con đường tạo phôi vô tính (somatic
embryogenesis) [68][128][129], kể cả theo con đường tạo chồi. Mô sẹo có KNSP có
thể hình thành ngay ở lần nuôi cấy đầu tiên hoặc/và ở một số lần cấy chuyền nuôi cấy
tiếp theo. Theo Von Arnold và cộng sự (2002), sự khác biệt giữa mô sẹo có KNSP và
mô sẹo không có KNSP dễ ghi nhận, dựa trên hình thái, màu sắc; mô sẹo có KNSP
là thể mô mang các cụm tế bào có khả năng sinh tiền phôi (proembryogenic masses -
PEMs) [68]. Mô sẹo có KNSP (embryogenic) là thể mô cứng, trắng hơi vàng, ở dạng
cụm/hạt và phát triển chậm – khác với mô sẹo không có KNSP thường có cấu trúc
xốp, phát triển nhanh, màu trắng/xám [130]. Theo Gaoyin và cộng sự (2020), ở mô
sẹo có khả năng sinh phôi có sự tích lũy lượng IAA nội sinh, đường hòa tan, tinh bột
và protein cao – cơ sở quan trọng về năng lượng cho quá trình tạo phôi so với mô sẹo
không có khả năng sinh phôi [12]. Như đã trình bày, ở NGBCC, sự phát sinh mô sẹo
có KNSP được ghi nhận sau giai đoạn cấy chuyền mô sẹo có thể do ở lần nuôi cấy
đầu tiên sự hiện diện của rễ tái sinh đã gây hạn chế sự hình thành loại mô này. Từ các

70
kết quả trên đây, mảnh lá được nuôi cấy trên môi trường có 2 mg/L 2,4-D để thu mô
sẹo có KNSP phục vụ nghiên cứu tái sinh phôi ở giai đoạn tiếp theo.
Tạo phôi từ mô sẹo mảnh lá (10 x10 mm) trên môi trường đặc
Từ kết quả thí nghiệm trên thu được các cụm mô sẹo có KNSP (Hình 3.13A)
sử dụng làm vật liệu tạo phôi, được nuôi cấy trên môi trường SH có 0,25 mg/L BA
kết hợp với NAA (1, 2, 3, 4, 5 mg/L). Quan sát mẫu sau 30 ngày cấy, kết quả cho
thấy ở tất cả các cụm mô sẹo có KNSP trên môi trường nuôi cấy SH đều có hiện
tượng tái sinh phôi (tỷ lệ 100%), số phôi hình thành nhiều nhất ở môi trường có 2 –
3mg/L NAA tương ứng số phôi tái sinh/cụm là 15,5; 15,8 ít nhất ở môi trường có
NAA 1 mg/L với số phôi/cụm mô sẹo là 7,2. Ở nghiệm thức có 2, 3 mg/L NAA không
có sự khác biệt về thống kê cả hai chỉ tiêu theo dõi, nhưng khác biệt so với các nghiệm
thức còn lại (Bảng 3.6, Hình 3.13).
Bảng 3.6. Ảnh hưởng của NAA và BA đến sự tạo phôi từ mô sẹo mảnh lá (10 x 10
mm), ở môi trường SH, 30 NSC.
NAA
(mg/L)
BA
(mg/L)
Số cụm mô
sẹo có KNSP
dùng nuôi cấy
Số phôi
tái
sinh/cụm
Đánh giá định tính về phôi tái sinh
1 0,25 30 7,20d* Phôi dài, rễ phôi ngắn
2 0,25 30 15,50a Phôi dài, rễ phôi ngắn
3 0,25 30 15,80a Phôi ngắn, rễ phôi ngắn có lông hút
4 0,25 30 10,80b Phôi ngắn; một số phôi có rễ dài,
nhiều lông hút
5 0,25 30 9,50c Phôi rất ngắn, nhiều rễ có lông hút
trên cụm phôi; cụm có nhiều mô
sẹo
*Các chữ cái khác nhau trong một cột thể hiện sự khác biệt có ý nghĩa ở mức
p ≤ 0,05 trong phép thử LSD.
Tương tự trường hợp tạo mô sẹo có KNSP, cũng sử dụng auxin (2,4-D/NAA)
kết hợp với cytokinin (BA/kinetin) trong tạo phôi vì cytokinin thường giúp kích thích
phôi hình thành và phát triển; trong một số trường hợp, sự tạo phôi và sự trưởng thành
của phôi được ghi nhận trong điều kiện sử dụng cùng nồng độ chất điều hòa sinh
trưởng [131]. Tuy nhiên, ở đa số trường hợp tạo phôi, mô sẹo có khả năng sinh phôi

71
cần được nuôi cấy trên môi trường có nồng độ auxin giảm hoặc không bổ sung auxin
[132]. Theo hướng này, auxin NAA (có hoạt tính sinh lý thấp hơn 2,4-D) thường
được sử dụng nhằm kích thích tái sinh phôi và kích thích phát triển phôi tiếp tục sau
khi đã hình thành và ở nghiên cứu này, auxin NAA được lựa chọn sử dụng phục vụ
nghiên cứu tái sinh phôi. Tương tự như ở nghiên cứu của Gaoyin và cộng sự (2020)
[12]. Theo Yu và cộng sự (2017) [99], tuy ở điều kiện nuôi cấy có nồng độ auxin cao
nhưng ở khối mô sẹo luôn có sự phân bố không đồng đều của auxin theo không gian
(spatial) do sự di chuyển phân cực của auxin có khác nhau ở các vị trí khác nhau, do
vậy có thể xảy ra hiện tượng biệt hóa từng phần/toàn phần khi ở điều kiện thuận lợi
(ví dụ chuyển sang nuôi cấy sử dụng nồng độ auxin thấp, hoặc dùng loại auxin nuôi
cấy phù hợp) – các cấu trúc như phôi/rễ bất định có thể được tái sinh.
Hình 3.13. Tái sinh phôi từ mô sẹo có khả năng sinh phôi trên môi trường đặc SH,
30 NSC và hình thái giải phẫu phôi tái sinh từ mô sẹo.
A. Vật liệu mô sẹo có KNSP (vị trí mũi tên); B,C,D,E,F. Phôi tái sinh trên môi trường
có NAA 1; 2; 3; 4; 5 mg/L; G,H. Phôi bước đầu hình thành (vị trí mũi tên) từ cụm
mô sẹo có KNSP và hình thái giải phẫu phôi tương ứng (vạch chéo đen chỉ vị trí cắt
thu lát mỏng tế bào phục vụ nhuộm màu kép); I. Cụm mô sẹo xốp không có KNSP.

72
Đến nay đã ghi nhận nhiều nghiên cứu tạo phôi hiệu quả khi kết hợp 2,4-D và
BA/kinetin, như trường hợp tạo phôi Passiflora mollissima từ mô sẹo lá mầm trên
môi trường MS có 4,5 μM 2,4-D và 4,5 μM BA [133]. Với Psoralea corylifolia, sự
kết hợp NAA và BA rất quan trọng đối với tạo mô sẹo có KNSP (từ vật liệu trụ dưới
lá mầm) với NAA (2,7 - 10,8 µM), BA 2,2 µM và tái sinh phôi từ mô sẹo với NAA
1,4 µM, BA 2,2 µM [134]. Sự kết hợp của NAA (0,5 - 5 mg/L) với BA 0,5 mg/L luôn
cần thiết đối với tạo phôi vô tính trực tiếp từ mảnh lá/lá mầm Solanum nigrum [135].
Tương tự, tổ hợp NAA và BA cũng rất thích hợp để tạo phôi từ mô sẹo lá Citrullus
colocynthis [136], mô sẹo phôi hạt non Tapiscia sinensis [94], mô sẹo phôi hợp tử
non Picea abies và P. omorika [137]. Kết quả thí nghiệm cho thấy, lần nữa NAA đã
chứng tỏ rất thích hợp cho tái sinh phôi gián tiếp từ mô sẹo với nồng độ 2 mg/L và
cytokinin BA cũng rất cần thiết cho quá trình tái sinh phôi với nồng độ sử dụng là
0,25 mg/L – tương tự như ở nghiên cứu tạo phôi trực tiếp từ mảnh lá.
Quan sát cấu trúc giải phẫu phôi
Khảo sát hình thái giải phẫu phôi tái sinh từ cụm mô sẹo có khả năng sinh phôi
(Hình 3.13G) bằng phương pháp nhuộm hai màu và quan sát mẫu vật dưới kính hiển
vi đã được thực hiện. Kết quả cho thấy phôi cầu hình thành trên bề mặt khối mô sẹo
với lớp biểu mô đặc trưng (Hình 3.13H). Qua khảo sát tế bào học, Sakr và Sayed
(2018) kết luận phôi Oryza sativa có nguồn gốc từ một số tế bào phôi ở phần ngoài
của các cụm mô sẹo phôi hợp tử đang phát triển sinh khối trên môi trường MS có (2
- 2,5 mg/L) 2,4-D [138]. Ở trường hợp tạo phôi Carica papaya từ mô sẹo lá mầm
phôi hạt, phôi tái sinh bắt nguồn từ các cấu trúc dạng cầu có có khả năng sinh tiền
phôi (pre-embryogenic globular structures) hình thành từ các tế bào vùng bề mặt và
dưới bề mặt của cụm mô sẹo [139]. Tương tự, các tiền phôi Tapiscia sinensis hình
thành từ lớp tế bào bề mặt (surface layer) và dưới bề mặt hoặc bên trong (internally)
của cụm mô sẹo có KNSP [94]. Như vậy, theo chúng tôi, phôi NGBCC tạo từ mô sẹo
có KNSP cũng bắt nguồn từ hoạt động phân chia của loại mô tế bào đặc trưng tương
tự như ở kết quả nghiên cứu của các tác giả nêu trên và cơ sở sinh lý của quá trình
tạo phôi là do sự di chuyển phân cực của auxin, làm thay đổi lập trình di truyền của
tế bào mô sẹo dẫn đến sự hình thành tế bào mang tính toàn thế - có khả năng tái sinh
cơ quan [15].

73
Hình 3.14. Các dạng phát triển của phôi tái sinh từ mô sẹo mảnh lá, cây con hoàn
chỉnh từ phôi.
A. Phôi cầu đơn; B. Phôi dạng tim; C. Giai đoạn đầu của phôi dạng thủy lôi; D,E.
Phôi thủy lôi đơn và cụm 2 phôi thủy lôi phát triển; F,G,H. Phôi có lá mầm và rễ
mầm phát triển đến giai đoạn trưởng thành; I. Cây con hoàn chỉnh từ phôi với lá mầm,
lá thật (đơn và kép), rễ cọc điển hình và nhiều rễ phụ
Tương tự như ở quá trình nghiên cứu tạo phôi trực tiếp từ mảnh lá, trong quá
trình tái sinh phôi từ mô sẹo, cũng đã ghi nhận được đầy đủ các dạng phát triển khác
nhau của phôi như phôi dạng cầu (Hình 3.14A), dạng tim (Hình 3.14B), dạng thuỷ
lôi (Hình 3.14C,D,E), dạng bước đầu có lá mầm, rễ mầm (Hình 3.14F,G) và dạng
phôi trưởng thành (Hình 3.14H). Phôi trưởng thành phát triển bình thường thành cây
con qua nuôi cấy tiếp tục trên môi trường ½MS không bổ sung chất điều hòa sinh
trưởng (Hình 3.14I).
Tạo phôi từ mô sẹo mảnh lá (10 x10 mm) trong môi trường lỏng
Từ vật liệu là các cụm mô sẹo có KNSP (Hình 3.15A), nuôi cấy tái sinh phôi
trong môi trường lỏng lắc cũng đã được thực hiện dùng môi trường SH có 2 mg/L
NAA và 0,25 mg/L BA (kế thừa từ thí nghiệm trên). Kết quả cho thấy, phôi hình
thành dần theo thời gian ở 7, 10, 14 và 30 NSC (Hình 3.15B,C,D,E,F,G).

74
Hình 3.15. Tạo phôi từ mô sẹo có KNSP bằng phương pháp nuôi lỏng lắc, hình thái
cụm tế bào phân chia, cụm mô tạo phôi.
A. Các cụm mô sẹo có KNSP làm vật liệu tạo phôi (vị trí mũi tên); B,C,D. Bình nuôi
các cụm mô sẹo có KNSP giai đoạn mới cấy, 7 NSC, 30 NSC; E,F,G. Cận cảnh tạo
phôi từ cụm mô sẹo có KNSP ở 10, 14, 30 NSC; H. Hình thái cụm tế bào phân chia
theo hướng tạo phôi (vòng tròn (a) chỉ vị trí tế bào phân chia không cân xứng và vòng
tròn (b) chỉ vị trí cụm tế bào nhỏ đẳng kính); I. Phôi dạng cầu hình thành ở giai đoạn
đầu ở cụm mô có KNSP (vị trí mũi tên); J. Phôi dạng cầu phát triển (các hình H,I,J
được chụp dưới kính hiển vi với độ phóng đại 50X, 20X, 20X, theo thứ tự)
Quan sát hình thái cụm tế bào/cụm mô (có nguồn gốc mô sẹo có khả năng sinh
phôi) nuôi lỏng lắc
Trong quá trình theo dõi, quan sát mô sẹo nuôi lỏng lắc, ghi nhận được hai
hiện tượng xảy ra có thể liên quan đến sự tạo phôi, một là, sự phân chia tế bào không
đối xứng (asymmetric division/ unequal division pattern) dẫn đến sự hình thành hai
tế bào không cùng kích cỡ (Hình 3.15Ha) – tương tự như ở trường hợp hình thành
phôi hợp tử và nhiều trường hợp hình thành phôi vô tính [68]; hai là, sự hình thành
cụm tế bào tròn nhỏ, đẳng kính (isometric), có vách dày (Hình 3.15Hb). Grzebelus
và cộng sự (2012) đã ghi nhận được hiện tượng phân chia tế bào không đối xứng ở

75
nuôi cấy tế bào trần có nguồn gốc từ mô lá, trụ dưới lá mầm Daucus carota dẫn đến
sự hình thành cụm đa bào (multi-cell colonies) có kết cấu chặt (tightly packed), tế
bào dạng tròn, tế bào chất đậm đặc ở 5 – 10 NSC, với kích cỡ nhỏ hơn tế bào ban
đầu; sau đó các cụm này phát triển thành cụm tiền phôi (proembryonic mass), tạo
phôi cầu ở 21 – 28 NSC và phôi có lá mầm ở 30 – 60 NSC [140]. Iantcheva và cộng
sự (2004) đã nghiên cứu chi tiết sự phân chia không đối xứng của quần thể tế bào đơn
(100 - 200 µm) Medicago falcata đối chứng và chuyển gen (gusA) và kết luận hầu
hết các tế bào có sự phân chia không đối xứng (~ 80%, 50 – 75% tế bào phân chia
không đối xứng ở đối chứng và chuyển gen, theo thứ tự, ở ngày thứ 15 sau cấy) đều
dẫn đến tạo phôi cầu ở giai đoạn sau cấy 20 ngày, tạo phôi trưởng thành và tạo cây
con ở ngày thứ 15 – 20 sau cấy sau khi được cấy chuyển sang môi trường thích hợp
[67]. Trong quần thể tế bào huyền phù Coffea arabica cv. Catimor, các tế bào dạng
tròn, kích thước nhỏ, vách dày, tế bào chất đậm đặc có phân sự chia tế bào theo kiểu
không đối xứng cũng dẫn đến tạo phôi [109].
Theo Friml và cộng sự (2003), ở Arabidopsis thaliana, sau sự phân chia lần
thứ nhất không đối xứng thành hai tế bào gốc (basal) to và tế bào ngọn (apical) nhỏ,
auxin di chuyển phân cực từ tế bào gốc đến tế bào ngọn - kích hoạt chương trình di
truyền chuyển dịch tế bào ngọn sang quá trình tạo phôi [141].
Hình 3.16. So sánh kết quả tái sinh phôi ở môi trường SH đặc và lỏng, 30 NSC.
A,B. Cụm phôi tái sinh trên môi trường đặc, lỏng (thanh ngang 10 mm).
Tuy không bố trí thí nghiệm so sánh hiệu quả phương pháp nuôi cấy tái sinh
nhưng bước đầu so sánh kết quả tái sinh phôi ở môi trường SH có 2 mg/L NAA và
0,25 mg/L BA đã được thực hiện. Kết quả cho thấy phôi tái sinh từ mô sẹo nuôi lỏng
lắc đạt số lượng gần gấp đôi (21,4) số phôi tái sinh trên môi trường đặc (12,6) (Hình

76
3.16A,B), có thể do sự hấp thu nước, chất dinh dưỡng, chất điều hòa sinh trưởng của
mô tế bào trong môi trường lỏng tốt hơn [142]. Kết quả tái sinh phôi tích cực trong
môi trường lỏng tạo điều kiện cho thí nghiệm nhân phôi quy mô lớn phục vụ nhân
giống, nghiên cứu sản xuất hợp chất thứ cấp,...
3.1.2.2. Tạo phôi vô tính gián tiếp qua mô sẹo mảnh lá (3 x 10 mm)
Tạo mô sẹo
Tương tự trường hợp tạo mô sẹo từ mảnh lá (10 x 10 mm), các mảnh lá kích thước
nhỏ (3 x 10 mm) cũng được nuôi cấy tạo mô sẹo trên môi trường SH có 2,4-D với
các nồng độ khác nhau (0, 1, 2, 3 mg/L).
Hình 3.17. Mô sẹo hình thành từ mảnh lá (3 x 10 mm) ở môi trường SH có 2,4-D,
20 – 30 NSC.
A,B,C,D. Môi trường ĐC, có 2 mg/L 2,4-D ở 20 NSC, có 1 mg/L 2,4-D 30 NSC, có
2 mg/L 2,4-D 30 NSC; E. Cận cảnh sự hình thành các cụm mô sẹo nhỏ có KNSP/phôi
cầu (ô vuông được phóng to thành hình E); F. Môi trường có 3 mg/L 2,4-D. Cụm mô
sẹo nhỏ có KNSP, MSSP (mô sẹo sinh phôi - mũi tên đen); vị trí phôi cầu (mũi tên
trắng).
Sau 20 ngày nuôi cấy, mô sẹo hình thành nhiều ở vị trí vết cắt (Hình 3.17B),
đặc biệt không có hiện tượng tái sinh rễ như ở nuôi cấy mảnh lá. Ở ngày 30 sau nuôi

77
cấy, mô sẹo hình thành trên khắp bề mặt mẫu cấy mảnh lá; không ghi nhận có sự hình
thành mô sẹo ở nghiệm thức đối chứng (Hình 3.17A). Qua quan sát dưới kính hiển vi
nhận thấy trên bề mặt mô sẹo ở tất cả các nghiệm thức có 2,4-D đều có các cụm mô
nhỏ có bề mặt khô, màu hơi trắng/vàng, xuất hiện rải rác trên bề mặt mô sẹo, đây là
các cụm mô sẹo nhỏ có KNSP (không thu thập số liệu). Điểm đặc biệt các mẫu cấy ở
2 mg/L 2,4-D, ngoài sự hình thành các cụm mô sẹo nhỏ nêu trên còn ghi nhận có sự
hình thành một số phôi dạng cầu có màu hơi xanh ở một số mẫu (Hình 3.17D,E). Do
vậy, 2 mg/L 2,4-D được sử dụng để tạo vật liệu mô sẹo phục vụ nghiên cứu tạo phôi.
Tạo phôi
Mô sẹo hình thành từ mảnh lá (3 x 10 mm) được nuôi cấy tái sinh trên môi
trường SH có sự kết hợp của NAA với các nồng độ khác nhau và 0,25 mg/L BA. Kết
quả thu được, ở ngày thứ 30 sau nuôi cấy, sự tái sinh phôi rất hiệu quả, phôi xuất hiện
trên tất cả các mẫu cấy mô sẹo, số lượng phôi nhiều ở môi trường có NAA từ 2 - 5
mg/L (Bảng 3.7) (Hình 3.18B,C,D,E,F).
Bảng 3.7. Ảnh hưởng của NAA và BA đến tạo phôi từ mô sẹo mảnh lá (3 x 10 mm),
ở môi trường SH, 30 NSC.
NAA
(mg/L)
BA
(mg/L)
Số mẫu mô
sẹo tạo phôi
Số phôi/mẫu
mô sẹo
Đặc điểm hình thái về phôi tái
sinh
1 0,25 30 4,47d* Phôi chưa có rễ, mô sẹo kém
phát triển
2 0,25 30 15,10a Một số phôi có rễ, phôi dài,
nhiều phôi nhỏ đang trong quá
trình phát sinh/phát triển
3 0,25 30 14,84a Một số phôi có rễ, phôi dài, lá
mầm to; mô sẹo phát triển to
4 0,25 30 9,37b Một số phôi có rễ, lá mầm to;
mô sẹo phát triển mạnh
5 0,25 30 7,80c Một số phôi có rễ, lá mầm to;
mô sẹo phát triển mạnh
* Các chữ cái khác nhau trong một cột thể hiện sự khác biệt có ý nghĩa
ở mức p ≤ 0,05 trong phép thử LSD.

78
Tương tự trường hợp nuôi cấy tạo phôi từ mô sẹo mảnh lá, ở thí nghiệm này,
cũng đã ghi nhận được đầy đủ các dạng phát triển khác nhau của phôi như dạng cầu,
dạng tim, dạng thủy lôi, dạng có lá mầm và rễ mầm (Hình 3.18G,H).
Hình 3.18. Tạo phôi gián tiếp từ mô sẹo mảnh lá (3 x 10 mm) trên môi trường SH,
30 NSC.
A. Mẫu mô sẹo làm vật liệu nuôi cấy; B,C,D,E,F. Phôi tái sinh trên môi trường SH
có NAA 1, 2, 3, 4, 5 mg/L (thanh ngang 5 mm); G. Sự hình thành phôi dạng cầu (C),
dạng có lá mầm và rễ mầm (vị trí mũi tên); H. Phôi dạng tim (T) và dạng thủy lôi
(TL) (vị trí mũi tên).
Phôi tái sinh tạo cây con hoàn chỉnh qua nuôi cấy trên môi trường đặc ½MS
không bổ sung chất ĐHST (Hình 3.19F).
Trong nghiên cứu nuôi cấy mô, vật liệu LMTB cũng được sử dụng khá phổ
biến với ưu điểm là khả năng phát sinh hình thái cao hơn vì có sự tương tác tốt giữa
tế bào với môi trường so với các mẫu cấy có kích thước to/dày [143].
Sự phát triển tiếp tục của phôi tái sinh cũng đã được theo dõi đến giai đoạn
sau nuôi cấy 60 ngày (không thu thập số liệu) (Hình 3.19). Ở cuối giai đoạn này, nhận
thấy phôi phủ kín bề mặt khối mô sẹo ở môi trường có NAA 2 mg/L (Hình 3.19B).
Nhìn chung, các cụm phôi hình thành đều có màu xanh rõ, ở môi trường có NAA (3
- 5 mg/L) phôi có rễ mang nhiều lông hút (Hình 3.19C,D,E).

79
Hình 3.19. Phôi tái sinh từ mô sẹo mảnh lá (3 x 10 mm) ở môi trường SH, 60 NSC.
A,B,C,D,E. Phôi phát triển trên môi trường SH có 0,25 mg/L BA và NAA (1, 2, 3,
4, 5 mg/L), thanh ngang 6 mm; F. Cây con từ phôi qua nuôi cấy ở môi trường ½MS,
20 NSC.
Auxin (hoặc kết hợp với cytokinin) thường được sử dụng để tạo mô sẹo/phôi
từ LMTB. Vu Thi Hien và cộng sự (2016) đã sử dụng 2,4-D 1 mg/L kết hợp với BA
0,5 mg/L hoặc TDZ 0,1 mg/L để tạo mô sẹo sâm Ngọc Linh (Panax vietnamensis) từ
nuôi cấy lát mỏng tế bào cắt ngang (t-TCL) lá cây in vitro trên môi trường MS [144];
tương tự, mô sẹo rễ cây Panax vietnamensis in vitro 3 tháng tuổi cũng đã được tạo
thành công qua nuôi cấy t-TCL (dày ~ 1 mm) trên môi trường MS có 2,4-D 1 mg/L
kết hợp với NAA 0,5 mg/L hoặc/và kết hợp với kinetin 0,2 mg/L [74]. Trong nghiên
cứu tạo phôi từ mô sẹo t-TCL chồi in vitro 02 giống Blackberry (Rubus sanctus và
Rubus hirtus), Sabooni và Shekafandeh, (2017) đã sử dụng NAA 10,84 μM và BA
8,88 μM [145]. Scherwinski-Pereira và cộng sự (2010) đã sử dụng môi trường MS có
bổ sung 0 – 450 µM Picloram, 2,4-D, 3% sucrose, 500 mg/L glutamine, 0,6 µM NAA
và 12,30 µM 2iP để duy trì mô sẹo có KNSP cây cọ dầu Elaeis guineensis [146].
Từ kết quả của các thí nghiệm tạo phôi gián tiếp qua mô sẹo mảnh lá NGBCC
ngoài vật liệu mảnh lá (10 x10 mm), thì mảnh lá kích thước nhỏ (3 x 10 mm) đã
chứng tỏ là vật liệu tốt đối với nghiên cứu tạo mô sẹo có khả năng phát sinh hình thái
phôi/tái sinh phôi qua các bước phát triển khác nhau.

80
3.2. Nhân phôi vô tính
3.2.1. Nhân phôi trên môi trường đặc
Ở 30 NSC, kết quả (Bảng 3.8, Hình 3.20) cho thấy số phôi/cụm cao ở các
nghiệm thức có 2; 3; 4; và 5 mg/L NAA, lần lượt là 20,8; 22,6; 25,3 và 26,8; nhiều ở
nghiệm thức 4 mg/L và 5 mg/L NAA và ít nhất ở nghiệm thức 1 mg/L NAA (16,20).
Ở các nồng độ khác nhau của NAA khi kết hợp với 0,25 mg/L BA kết quả tạo phôi
có khác biệt về thống kê. Kết quả này, một lần nữa chứng minh NAA, không chỉ có
tác động hiệu quả đến tạo phôi trực tiếp từ mô lá mà còn có ảnh hưởng tích cực đến
tạo phôi thứ cấp từ phôi sơ cấp. Nhận thấy kích thước phôi tăng với lá mầm to, số rễ
và kích thước rễ tăng khi nồng độ NAA tăng. Ở nghiệm thức đối chứng (0 mg/L
NAA), phôi phát triển chiều cao, không ghi nhận tạo phôi thứ cấp (Hình 3.20C).
Bảng 3.8. Ảnh hưởng của NAA và BA đến sự tạo phôi thứ cấp, ở môi trường đặc SH,
30 NSC.
NAA
(mg/L)
BA
(mg/L)
Số
phôi/cụm
Hệ số
nhân
Đặc điểm hình thái cụm phôi/phôi
0 0,25 3,00f* 1,00f Phôi phát triển chiều cao, không ghi
nhận hiện tượng tạo phôi thứ cấp; lá
mầm phôi to, rễ ít, ngắn
1 0,25 16,20e 5,40e Kích thước cụm phôi nhỏ; thân phôi
thấp, lá mầm phôi nhỏ, rễ ít, nhỏ và
ngắn
2 0,25 20,80d 6,93d Kích thước cụm phôi to; thân phôi
thấp, lá mầm phôi trung bình, rễ ít,
nhỏ và ngắn
3 0,25 22,60c 7,50c Kích thước cụm phôi to; lá mầm
phôi to, rễ nhiều, to và dài
4 0,25 25,30b 8,40b Kích thước cụm phôi rất to; thân
phôi cao, lá mầm phôi to, rễ rất
nhiều, to và dài
5 0,25 26,80a 8,93a Kích thước cụm phôi rất to; thân
phôi cao, lá mầm phôi rất to, rễ rất
nhiều, rất to và rất dài
* Các chữ cái khác nhau trong một cột thể hiện sự khác biệt có ý nghĩa ở mức
p ≤ 0,05 trong phép thử Duncan.

81
Hình 3.20. Ảnh hưởng của NAA và BA đến sự nhân phôi trên môi trường đặc SH,
30 NSC.
A. Đĩa phôi nuôi cấy ban đầu; B. Cụm phôi nuôi cấy ban đầu; C. Cụm phôi ĐC
(không bổ sung NAA); D,E,F,G,H. Môi trường SH có bổ sung 1, 2, 3, 4, 5 mg/L
NAA, (thanh ngang 1 cm); I,J,K. Vị trí phát sinh phôi thứ cấp (mũi tên) trên thân
phôi; L. Vị trí phát sinh phôi thứ cấp từ rễ phôi (mũi tên). Hình chụp dưới kính hiển
vi soi nổi với độ phóng đại 10X (I,J,K); 20 X (L).
Tương tự trường hợp tạo mô sẹo có KNSP và tạo phôi sơ cấp từ mô sẹo,
auxin (2,4-D/NAA,…) ở dạng riêng lẻ hoặc kết hợp với cytokinin (BA/kinetin) cũng
có vai trò rất quan trọng trong kích thích sự hình thành phôi thứ cấp từ phôi sơ cấp
ở nhiều loài thực vật [129]. Môi trường tạo phôi thứ cấp Castanea sativa [136],
Solanum nigrum [135] thích hợp là MS có (0,1 mg/L NAA; 0,1 mg/L BA), (2 mg/L
NAA và 0,5 mg/L BA), theo thứ tự. Tương tự, tổ hợp (NAA và BA) cũng được xác
định là thích hợp trong một số trường hợp khác, ví dụ (2 µM NAA và 2 µM BA)
dùng cho tạo phôi thứ cấp Angelica glauca Edgew [147]; môi trường SH chứa (0,5
mg/L NAA và 2 mg/L BA) rất thích hợp đối với tạo phôi thứ cấp Crocus vernus
[148]. Arya và cộng sự (1993) kết luận 2,4-D, NAA, và IAA ở nồng độ 1 mg/L có

82
tác động rất hiệu quả đến sự hình thành phôi thứ cấp cấp 1, cấp 2 ở Panax ginseng
[34]. Và ở NGBCC, NAA và BA cũng đóng vai trò quan trọng trong tạo phôi thứ
cấp tương tự như ở các trường hợp trên.
3.2.2. Nhân phôi trong môi trường lỏng
3.2.2.1. Ảnh hưởng của chất điều hòa sinh trưởng đến sự tạo phôi thứ cấp
Bảng 3.9. Ảnh hưởng của NAA và BA đến sự tạo phôi thứ cấp, ở môi trường lỏng
SH, 30 NSC.
NAA
(mg/L)
BA
(mg/L)
Số phôi
thứ cấp
Đặc điểm hình thái, màu sắc phôi/ cụm phôi
0 0,25 0,00e* Không có hiện tượng tạo phôi thứ cấp; phôi sơ cấp
phát triển với hình thái rễ, thân và lá mầm rõ rệt
1 0,25 15,23d Phôi thứ cấp nhỏ, dạng nốt tròn, màu hơi xanh; cụm
phôi ít rễ
2 0,25 17,30c Phôi thứ cấp nhỏ, dạng nốt tròn/thon dài, màu hơi
xanh; cụm phôi ít rễ
3 0,25 19,37a Phôi thứ cấp to, màu hơi xanh; cụm phôi có nhiều
rễ. Có hiện tượng sùi mô sẹo ở viền lá mầm phôi
cấy ban đầu.
4 0,25 18,56b Phôi thứ cấp to, màu hơi xanh; cụm phôi có nhiều
rễ. Có hiện tượng sùi mô sẹo ở viền lá mầm phôi
cấy ban đầu và ở rễ
*Các chữ cái khác nhau trong một cột thể hiện sự khác biệt có ý nghĩa ở
mức p ≤ 0,05 trong phép thử LSD.
Từ kết quả trên, 04 nghiệm thức NAA (1; 2; 3; 4 mg/L) được tiếp tục sử dụng
cho nghiên cứu tạo phôi thứ cấp trong môi trường lỏng vì ở nồng độ 5 mg/L NAA,
phôi có lá mầm rất to, rễ rất to và rất dài có thể gây hạn chế quá trình nhân phôi. Kết
quả cho thấy số phôi thứ cấp phát sinh từ phôi sơ cấp đơn nuôi cấy trong môi trường
có 1; 2; 3; 4 mg/L NAA lần lượt là 15,23; 17,30; 19,37; 18,56 (Bảng 3.9) và phôi thứ
cấp hình thành ở phần thân phôi sơ cấp (Hình 3.21D,E,F,G). Ở phôi nuôi cấy trong
môi trường không bổ sung NAA, không ghi nhận hiện tượng tạo phôi thứ cấp (Hình
3.21C). Tuy số phôi ở nghiệm thức 2 mg/L NAA không nhiều như ở 02 nghiệm thức
3 mg/L và 4 mg/L NAA nhưng phôi thứ cấp hình thành có dạng nốt tròn/thon dài với

83
số lượng khá nhiều, rễ phôi ngắn - tạo điều kiện thuận lợi cho nghiên cứu nhân phôi
sau này.
Như đã nêu trên, vai trò của auxin và cytokinin là rất quan trọng đối với sự
hình thành/phát triển phôi nói chung và nói riêng đối với phôi nuôi lỏng [22][149].
Môi trường lỏng MS có bổ sung 0,5 μM NAA và 5 μM BA tạo hiệu quả cao đối với
sự hình thành phôi Narcissus từ nuôi cấy mô sẹo có KNSP có nguồn gốc mô bầu
noãn [150]. Tương tự, môi trường lỏng MM có 2 mg/L NAA kích thích sự tạo phôi
từ mô sẹo có KNSP (từ nuôi cấy mảnh lá non) và môi trường có 2 mg/L BA tạo
phôi Coffea arabica với lá mầm phát triển [151].
Hình 3.21. Ảnh hưởng của NAA và BA đến sự tạo phôi thứ cấp trong môi trường
lỏng SH, 30 NSC.
A. Bình phôi đơn thu thập dùng thí nghiệm; B. Cận cảnh phôi đơn dùng nuôi cấy; C.
Cận cảnh phôi nuôi trong môi trường không bổ sung NAA (ĐC); D,E,F,G. Phôi thứ
cấp hình thành ở môi trường SH có 1, 2, 3, 4 mg/L NAA.
3.2.2.2. Ảnh hưởng của khối lượng phôi nuôi cấy đến sự tăng trưởng sinh khối
phôi
Sau 45 ngày nuôi cấy, kết quả thí nghiệm thu được ở (Bảng 3.10) cho thấy, KLP
nuôi cấy là 0,3 g (0,5%) có hệ số nhân sinh khối có giá trị thấp nhất là 10,40 và KLTP
thu nhận được là 3,12 g (Hình 3.22A); khi KLP nuôi cấy là 1,2 g (2%) thu được KLTP
là 17,85 g có hệ số nhân sinh khối cao nhất 14,88 (Hình 3.22C); KLP thu nhận được

84
đạt giá trị cao nhất là 23,45 g nhưng hệ số nhân sinh khối chỉ đạt 13,03 ở trường hợp
KLP nuôi cấy là 1,8 g (3%) (Hình 3.22D). Như vậy, khối lượng phôi nuôi cấy ban
đầu có ảnh hưởng đến sinh khối phôi thu nhận được, do liên quan đến mật độ trong
bình nuôi cấy.
Trong nuôi cấy mô sinh phôi/phôi nói chung, mật độ mẫu cấy (khối lượng mẫu
cấy - g hoặc số phôi)/đơn vị thể tích môi trường có vai trò quan trọng. Khi mật độ
nuôi cấy cao, làm hạn chế tiếp xúc của phôi cấy với chất dinh dưỡng và oxygen của
môi trường nuôi cấy, gây ra sự cạnh tranh, do đó làm giảm sự hấp thu chất dinh dưỡng
và oxygen, nên ảnh hưởng không tốt đến sinh trưởng của phôi [152]. Vì vậy, xác định
khối lượng phôi nuôi cấy ban đầu rất quan trọng cho sự tăng trưởng sinh khối phôi.
Như vậy, trong thí nghiệm này, sử dụng khối lượng phôi nuôi cấy ban đầu là 1,2 g
(tương ứng 2% w/v) thì sự tăng trưởng sinh khối phôi NGBCC đáp ứng sinh trưởng
tốt với hệ số nhân cao nhất.
Bảng 3.10. Ảnh hưởng của khối lượng phôi nuôi cấy đến tăng trưởng sinh khối phôi,
trong môi trường lỏng SH, 45 NSC
KLP nuôi
cấy (g)
KLTP thu
nhận (g)
Hệ số nhân
sinh khối (lần)
Đặc điểm hình thái cụm phôi
0,30 (0,5%) 3,12d* 10,40d Cụm phôi nhỏ, lá mầm to
0,60 (1%) 6,75c 11,25c Cụm phôi to, lá mầm to
1,20 (2%) 17,85b 14,88a Cụm phôi rất to, lá mầm nhỏ
1,80 (3%) 23,45a 13,03b Cụm phôi rất to, lá mầm nhỏ
*Các chữ cái khác nhau trong một cột thể hiện sự khác biệt có ý nghĩa ở mức p ≤
0,05 trong phép thử LSD.
Hình 3.22. Ảnh hưởng của khối lượng phôi (w/v) nuôi cấy đến tăng trưởng sinh
khối phôi trong môi trường lỏng SH, 45 NSC.
A,B,C,D. Môi trường nuôi cấy SH có 2 mg/L NAA và 0,25 mg/L BA, với khối lượng
phôi nuôi cấy 0,3 g; 0,6 g; 1,2 g; 1,8 g.

85
Đã ghi nhận một số nghiên cứu về khối lượng mẫu cấy ban đầu ảnh hưởng đến
hiệu quả của sự phát sinh phôi, như C. Guillou và cộng sự (2018) nghiên cứu về phát
sinh phôi của Theobroma cacao khi nuôi cấy trong bình tam giác đã sử dụng 1 – 2%
(w/v) mô sinh phôi PEMs (pro-embryogenic masses) là phù hợp; ngược lại, 5% gây
ức chế sinh trưởng [153]. Tương tự, B.B. Misra và S. Dey (2013), khi nuôi cấy mô
sinh phôi Santalum album (trong bioreactor 1,5 – 7,5 L) với khối lượng cấy ban đầu
là 1 – 2% là thích hợp nhất [154]. Đối với nuôi cấy phôi chuối lá [plantain FHIA-21
AAAB)] trong bình tam giác, García-Águila và cộng sự (2016) kết luận khối lượng
cấy ban đầu thích hợp là 2% (qua so sánh các khối lượng phôi nuôi cấy từ 0,7% đến
2,7%) [155].
Hình 3.23. Nhân phôi qua nuôi cấy lỏng lắc trong môi trường SH, 30 – 60 NSC.
A. Cụm phôi dùng nuôi cấy; B,C. Bình nuôi phôi ở 30 NSC, 60 NSC; D,E. Phôi với
các dạng phát triển và kích thước khác nhau; F. Tập hợp phôi thu được ở 60 NSC; G,
H. Các dạng phôi điển hình (PC: phôi cầu, PTL: Phôi thủy lôi, PT: Phôi tim); I. Các
dạng phát triển phôi từ dạng cầu đến dạng trưởng thành; J. Phôi trưởng thành sau cấy
chuyển sang môi trường đặc. Hình được chụp dưới kính hiển vi soi nổi với độ phóng
đại 10X (I,J), 20X (G,H).

86
Trong quá trình nuôi lỏng lắc, nhận thấy có hiện tượng tạo phôi thứ cấp liên
tục theo chu kỳ để tạo các phôi/cụm phôi mới - phù hợp với kết quả nghiên cứu của
[156][157] về sự hình thành cụm phôi theo chu kỳ (cyclic manner) từ cực rễ của phôi
Piper nigrum và từ cực chồi của phôi Elaeis guineensis, theo thứ tự. Theo dõi trong
thời gian dài (đến 60 ngày) cho thấy, các phôi do tác động cơ học của quá trình lắc
và do tương tác với nhau đã treo vào môi trường với nhiều hình dạng (dạng cầu, tim,
thủy lôi, hai lá mầm) và kích cỡ khác nhau (nhỏ, to, trung bình) (qua quan sát đáy
bình nuôi) (Hình 3.23D,E,F,G,H).
Qua sàng lọc, tập hợp các phôi có kích thước khác nhau được dùng cho các
nghiên cứu tiếp theo (Hình 3.24). Sự hình thành phôi/cụm phôi với các kích cỡ và
hình dạng khác nhau đã được ghi nhận trong quá trình tạo/nhân phôi trên môi trường
đặc [158] và trong môi trường lỏng [22].
Hình 3.24. Sự đa dạng của kích thước phôi từ quá trình nuôi lỏng lắc trong môi
trường SH.
A. Tập hợp các loại phôi có kích thước khác nhau; B,C,D. Tập hợp các phôi có kích
thước nhỏ, trung bình, to, theo thứ tự.
3.2.2.3. Ảnh hưởng của kích thước phôi đến sự tạo phôi thứ cấp
Sau 30 ngày nuôi cấy, kết quả (Bảng 3.11) cho thấy, trong 03 loại kích thước
phôi, với phôi có kích thước trung bình (~ 10 mm) tạo số phôi thứ cấp cao nhất
(17,84) ở 30 NSC; ở phôi nhỏ và phôi to có số phôi thứ cấp là 7,51 và 13,92. Quan
sát về hình thái của phôi thứ cấp nhận thấy phôi cấy ban đầu có kích thước nhỏ (~ 7
mm, Hình 3.25A), hầu như tạo phôi có dạng cầu (Hình 3.25B,C,D,E), còn phôi cấy

87
có kích thước trung bình (~ 10 mm, Hình 3.25F) phôi hình thành rất đa dạng, từ dạng
cầu đến dạng có lá mầm (Hình 3.25G,H,I), với phôi cấy có kích thước lớn (~ 15 mm,
Hình 3.25J) phôi hình thành có dạng có lá mầm (Hình 3.25K,L,M). Điều đó chứng
tỏ, sự khác nhau về kích thước phôi cấy ban đầu có ảnh hưởng đến số phôi thứ cấp
hình thành và sự phát sinh hình thái phôi thứ cấp. Như vậy, ở NGBCC vật liệu sử
dụng có kích thước trung bình (bước đầu có lá mầm) là thích hợp cho nhân phôi.
Bảng 3.11. Ảnh hưởng của kích thước phôi đến sự tạo phôi thứ cấp, trong môi trường
lỏng SH, 30 NSC.
Kích thước
phôi (mm)
Số phôi thứ
cấp
Đặc điểm hình thái, màu sắc phôi/cụm phôi
~ 7 7,51c* Phôi nhỏ, thường có dạng cầu hình thành
trên các bộ phận của phôi
~ 10 17,84a Phôi to, nhiều dạng phôi khác nhau: dạng
nốt tròn, dạng thon dài và dạng có lá mầm
và hình thành trên các bộ phận của phôi
~ 15 13,92b Phôi to, thường ở dạng có lá mầm, hình
thành trên các bộ phận của phôi
*Các chữ cái khác nhau trong một cột thể hiện sự khác biệt có ý nghĩa
ở mức p ≤ 0,05 trong phép thử LSD.
Ảnh hưởng của kích thước phôi đến sự tạo phôi thứ cấp đã được nghiên cứu
trên một số đối tượng cây trồng. Phôi hợp tử Brassica oleracea (var. gongylodes) ở
giai đoạn mới có lá mầm là vật liệu nuôi cấy cho tỷ lệ tạo phôi thứ cấp cao (80,7%)
khi được nuôi cấy trên môi trường không có chất điều hòa sinh trưởng [159]. Phôi vô
tính giai đoạn thủy lôi của dòng cà phê Arabica Arabica (AS2K) tạo phôi thứ cấp tốt
trên môi trường MS (đặc và bán rắn) có bổ sung Thidiazuron ở nồng độ 9,08 μM
[160]. Corredoira và cộng sự (2015) sử dụng phôi vô tính giai đoạn có lá mầm và
cụm phôi cầu và thủy lôi Eucalyptus globulus và E. saligna × E. maidenii nuôi cấy
trên môi trường có 16,11 µM NAA nhằm phát triển sinh khối [161]. Tuy nhiên, đối
với Malus x domestica Borkh. (cv. 'Gloster 69'), phôi vô tính kích thước to lại cho tỷ
lệ tạo phôi thứ cấp cao (> 73%) khi được nuôi cấy trên môi trường có tổ hợp
NAA/BAP/KIN hoặc chỉ có TDZ (10 μM) [162].

88
Hình 3.25. Ảnh hưởng của kích thước phôi đến sự tạo phôi thứ cấp trong môi
trường lỏng SH, 30 NSC.
A. Phôi kích thước nhỏ dùng nuôi cấy; phôi thứ cấp hình thành trên thân phôi (B,C),
rễ (D), lá (E) (vị trí mũi tên). F. Phôi kích thước trung bình dùng nuôi cấy; phôi thứ
cấp hình thành trên thân phôi (G), rễ (H), lá (I). J. Phôi kích thước to dùng nuôi cấy;
phôi thứ cấp hình thành trên thân phôi (K), rễ (L), lá (M).
3.2.2.4. Ảnh hưởng của nồng độ đường đến sự tăng trưởng sinh khối phôi
Ở 30 NSC, kết quả (Bảng 3.12) cho thấy, KLT phôi đạt giá trị cao nhất (4,67
g) và hệ số nhân phôi cũng đạt cao nhất 15,57 ở môi trường nuôi cấy SH có bổ sung
50 g/L sucrose. Nghiệm thức có nồng độ sucrose thấp (20 g/L) KLT phôi thu nhận
được là thấp nhất (2,14 g) phôi có màu xanh, lá mầm to, rễ dài so với phôi trong môi
trường có nồng độ sucrose cao (40 – 50 g/L) - phôi màu hơi vàng, lá mầm nhỏ, và rễ
ngắn (Hình 3.26).

89
Bảng 3.12. Ảnh hưởng của nồng độ đường đến sự tăng trưởng sinh khối phôi trong
môi trường lỏng SH, 30 NSC.
Nồng độ
sucrose (g/L)
KLT phôi
thu nhận (g)
Hệ số
nhân phôi
Đặc điểm hình thái, màu sắc
cụm phôi
20 2,14d* 7,29d Cụm phôi màu xanh, phôi có lá
mầm to, rễ dài
30 2,87c 9,57c Cụm phôi màu xanh, phôi có lá
mầm to, rễ dài
40 3,56b 11,87b Cụm phôi màu hơi vàng, phôi có
lá mầm nhỏ, rễ ngắn
50 4,67a 15,57a Cụm phôi màu hơi vàng, phôi có
lá mầm nhỏ, rễ ngắn
*Các chữ cái khác nhau trong một cột thể hiện sự khác biệt có ý nghĩa
ở mức p ≤ 0,05 trong phép thử LSD.
Hình 3.26. Ảnh hưởng của nồng độ đường đến sự tăng sinh khối phôi, trong môi
trường lỏng SH, 30 NSC.
A,B,C,D. Sự phát sinh phôi thứ cấp trong môi trường có nồng độ sucrose 20, 30, 40,
50 g/L
Naing và cộng sự (2013) kết luận cảm ứng tạo phôi thứ cấp Chrysanthemum
cv. Euro tốt hơn trên môi trường (MS có 2 mg/L 2,4-D, 2 mg/L kinetin) có 60 g/L
đường so với 30 g/L [75]. Ngược lại, phôi thứ cấp Panax ginseng phát sinh trên môi
trường MS, không có chất điều hòa sinh trưởng, với nồng độ đường tương đối thấp
(20 - 30 g/L); và nồng độ 20 g/L kích thích sự phát triển tiếp tục của phôi nhưng nồng
độ 30 g/L gây hạn chế dù kích thích tăng sinh khối [73]. Tương tự, Al Shamari và
cộng sự (2018) kết luận phôi thứ cấp Brassica oleraceae var. Botrytis phát sinh tốt
từ trụ dưới lá mầm phôi sơ cấp cũng trên môi trường bán rắn MS (không bổ sung chất

90
điều hòa sinh trưởng) có 30 g/L đường [163]. Nhìn chung, sự phát triển sinh khối
phôi NGBCC có nhu cầu về đường khá cao – tương tự một số trường hợp nêu trên.
3.2.2.5. Ảnh hưởng của cường độ ánh sáng đến sự tăng trưởng sinh khối phôi
Kết quả cho thấy (Bảng 3.13, Hình 3.27), cường độ ánh sáng 2.000 - 4.000 lux
đã có ảnh hưởng tích cực đến sự tăng trưởng sinh khối phôi so với nuôi phôi trong
điều kiện tối. Hệ số nhân phôi ở nghiệm thức tối, 2.000 lux và 4,000 lux lần lượt là
8,42; 9,37 và 10,40. Nói chung, phôi nuôi ngoài sáng xanh hơn phôi nuôi trong tối.
Phôi là cấu trúc mang nhiều lục lạp nên thường được nuôi cấy trong điều kiện có
chiếu sáng. Naing và cộng sự (2013) đã thực hiện nuôi cấy phôi sơ cấp
Chrysanthemum cv. Euro (môi trường MS có 2 mg/L 2,4-D và 2 mg/L kinetin) nhằm
tạo phôi thứ cấp ở điều kiện chiếu sáng cao 60 - 100 µmol m-2 s-1 (~ 4.500 – 7.500
lux) (16 h sáng : 8 h tối) [75]. Sử dụng cường độ ánh sáng 80 µmol m-2 s-1 (~ 6.000
lux), 40 µmol m-2 s-1 (~ 3.000 lux) đối với nuôi cấy tạo phôi thứ cấp Brassica oleracea
var. Botrytis [163], Polyscias filicifolia [54], theo thứ tự, cũng đã được ghi nhận.
Ngược lại, ở tạo phôi thứ cấp Quercus robur, nuôi cấy được thực hiện trong cùng
điều kiện tối [76] như nuôi cấy vật liệu phôi sơ cấp. Ở NGBCC, nuôi nhân phôi ở
điều kiện ánh sáng cao đã thúc đẩy sự hình thành phôi thứ cấp dẫn đến tăng sinh khối
tốt – phù hợp với tình hình chung về bố trí điều kiện sáng khi dùng vật liệu chứa
nhiều lục lạp.
Bảng 3.13. Ảnh hưởng của cường độ ánh sáng đến sự tăng trưởng sinh khối phôi,
trong môi trường lỏng SH, 30 NSC.
Cường độ ánh
sáng (lux)
KLT phôi
thu nhận (g)
Hệ số
nhân phôi
Đặc điểm hình thái, màu sắc
phôi
0 (tối) 2,53c* 8,42c Phôi màu hơi vàng, lá mầm
nhỏ, rễ phôi nhiều, dài
2.000 2,81b 9,37b Phôi màu xanh, lá mầm to, rễ
phôi trung bình
4.000 3,12a 10,40a Phôi màu xanh, lá mầm to, rễ
phôi ít, ngắn
*Các chữ cái khác nhau trong một cột thể hiện sự khác biệt có ý nghĩa ở
mức p ≤ 0,05 trong phép thử LSD.

91
Hình 3.27. Ảnh hưởng của điều kiện chiếu sáng đến sự tăng trưởng sinh khối phôi,
trong môi trường lỏng SH, 30 NSC
A. Phôi nuôi trong tối (ĐC); B,C. Phôi nuôi ngoài sáng cường độ 2.000 lux, 4.000
lux; D,E. Cận cảnh phôi nuôi trong tối, ánh sáng 4.000 lux.
3.2.2.6. Ảnh hưởng của tỷ lệ nước dừa đến sự hình thành phôi thứ cấp
Ở 21 NSC, kết quả cho thấy nước dừa đã ảnh hưởng tích cực đến quá trình
hình thành và phát triển phôi thứ cấp. Ở nghiệm thức 10% nước dừa, số phôi trung
bình đạt giá trị cao nhất (17,20) so với nghiệm thức không bổ sung nước dừa (13,40);
ngoài ra, cũng ghi nhận được lá mầm to và rễ dài (Bảng 3.14, Hình 3.28). Nước dừa
chứa nhiều thành phần dinh dưỡng tự nhiên như đường, vitamin, muối khoáng, acid
amin, các chất điều hòa sinh trưởng [164]. Vì vậy, bổ sung 10% nước dừa vào môi
trường nhân phôi ảnh hưởng tích cực đến sự phát sinh và phát triển phôi thứ cấp.
Bảng 3.14. Ảnh hưởng của tỷ lệ nước dừa đến sự tạo phôi thứ cấp, trong môi trường
lỏng SH, 21 NSC.
Nước
dừa (%)
Số phôi
thứ cấp
Đặc điểm hình thái, màu sắc phôi/cụm phôi
0 13,40c* Cụm phôi nhỏ, lá mầm phôi nhỏ, ngắn; rễ phôi ít, ngắn
5 14,27b Cụm phôi trung bình, lá mầm phôi trung bình; rễ phôi
nhiều, dài
10 17,20a Cụm phôi to, lá mầm phôi to, dài; rễ phôi nhiều, dài
*Các chữ cái khác nhau trong một cột thể hiện sự khác biệt có ý nghĩa
ở mức p ≤ 0,05 trong phép thử LSD.

92
Hình 3.28. Ảnh hưởng của tỷ lệ nước dừa đến sự tạo phôi thứ cấp, trong môi trường
lỏng SH, 21 NSC.
A. Bình vật liệu phôi nuôi cấy. B,C,D. Phôi nuôi trong môi trường SH có nước dừa
0%, 5%, 10%.
Qua nuôi cấy mô sẹo có KNSP mang phôi sơ cấp (từ mô hoa đực non chuối
Musa spp. AAB cv. Dwarf Brazilian) trên môi trường MS chỉ có 10% nước dừa, kết
quả cho thấy có sự hình thành phôi thứ cấp với tỷ lệ 39,8%, ở các nghiệm thức không
có nước dừa đều không có sự hình thành phôi thứ cấp [165]. Nước dừa (2 – 10%) có
ảnh hưởng tích cực đến cảm ứng tạo phôi từ mô sẹo có KNSP (từ nuôi cấy mảnh lá)
Kalopanax septemlobus [166]; nước dừa ngoài làm tăng sự phát sinh phôi từ mô sẹo
(ở nồng độ 20%), còn làm tăng sự tạo chồi từ phôi trưởng thành (ở nồng độ 10%) đối
với chà là Phoenix dactylifera [77]. Trong tạo phôi Phoenix dactylifera, nước dừa (10
– 15%) có tác động rất quan trọng [167]. Đối với loài Schefflera leucantha cùng chi
Ngũ gia bì, với tỷ lệ nước dừa 15% đã ảnh hưởng rất tốt đến chiều cao chồi, số lá, số
đốt thân,.. [168]. Như vậy, ở NGBCC nước dừa cũng là yếu tố quan trọng ảnh hưởng
đến sự phát sinh, phát triển phôi thứ cấp, với tỷ lệ thích hợp là 10%.
3.2.2.7. Tạo cây con từ phôi nuôi lỏng lắc
Các phôi đơn (Hình 3.29A) được nuôi cấy trong môi trường lỏng MS hoặc
½MS (không bổ sung chất điều hòa sinh trưởng, 20 g/L sucrose) nhằm thử nghiệm
tạo cây con. Kết quả cho thấy, tương tự như tạo cây đối với phôi nuôi trên môi trường
đặc, phôi phát triển trong môi trường khoáng lỏng ½MS tốt hơn so với môi trường
MS ở các chỉ tiêu tạo rễ, tạo lá (Hình 3.29B,C,D,E) (không thu thập số liệu). Ở 14
NSC, các phôi được cấy chuyển sang môi trường đặc ½MS để tạo chồi và phát triển
thành cây con (Hình 3.29F,G,H,I ). Các cây (đã có lá thật) sau đó được trồng ra chậu
đất ở vườn ươm (Hình 3.29J,K,L).

93
Hình 3.29. Tạo cây con từ phôi vô tính nuôi lỏng lắc trong môi trường MS, ½MS
A. Vật liệu phôi đơn nuôi cấy tạo cây; B,C. Phôi nuôi trong môi trường MS, ½MS ở
14 NSC; D,E. Phôi đơn trong môi trường MS, ½MS ở 14 NSC; F. Cụm phôi mới cấy
chuyển sang môi trường đặc ½MS. G,H,I. Phôi nuôi cấy trên môi trường đặc ½MS
ở 15, 30 và 60 NSC; J,K,L. Cây từ phôi trồng ra đất ở 7, 30, 45 ngày sau trồng.
Việc sử dụng môi trường có hàm lượng khoáng thấp (½MS), không hoặc có
bổ sung chất ĐHST với nồng độ thấp và lượng đường thấp (10 – 20 g/L) nhằm nuôi
phôi nhằm mục đích tạo cây được ghi nhận là mang tính quyết định.. Môi trường
½MS cũng đã được kết luận là phù hợp đối với phôi Brassica oleracea L. var. Italica
(từ nuôi cấy trong môi trường lỏng) nhằm tạo chồi (½MS, 1 mg/L BA) và tạo cây
trên môi trường đặc (½MS không có chất điều hòa sinh trưởng) [169]. Phôi mang
chồi Ranunculus sceleratus (chưa có rễ, 1 - 2 cm) cũng được cấy chuyển sang môi
trường ½MS, ½vit B5, 10 g/L đường để tạo rễ và phát triển thành cây con [170]. Nói
chung, việc sử dụng môi trường có hàm lượng khoáng thấp, giảm thiểu hàm
lượng/không sử dụng chất điều hòa sinh trưởng và đường là nguyên tắc nuôi cấy phổ
biến ở giai đoạn này [68][149], và nuôi cấy phôi nhằm tạo cây ở NGBCC là phù hợp
với các nghiên cứu.
3.2.3. Quan sát cấu trúc giải phẫu phôi sơ cấp và phôi thứ cấp
Khảo sát hình thái giải phẫu mô tế bào phôi bắt đầu có lá mầm phát sinh từ
thân (trụ dưới lá mầm) phôi sơ cấp nuôi cấy trên môi trường đặc (Hình 3.30A), phôi
cầu tạo từ thân phôi sơ cấp nuôi cấy trong môi trường lỏng (Hình 3.30D), phôi cầu
hình thành từ lá mầm (Hình 3.30H) và rễ phôi sơ cấp (Hình 3.30J) đã được thực hiện.
Từ kết quả thu được, theo chúng tôi, phôi thứ cấp phát sinh từ sự phân chia của
một/vài/nhiều tế bào/lớp tế bào bên ngoài thuộc vùng vỏ (cortex) của thân phôi sơ
cấp (Hình 3.30B,C,E,F), cũng không ghi nhận được sự thay đổi nhiều của mô tế bào

94
ở gần/vị trí mạch dẫn. Tương tự, phôi thứ cấp phát sinh từ lá mầm có nguồn gốc từ
lớp biểu mô của lá mầm phôi sơ cấp (Hình 3.30I) và phôi thứ cấp từ rễ có nguồn gốc
từ biểu mô vùng vỏ rễ phôi sơ cấp (Hình 3.30K).
Hình 3.30. Hình thái giải phẫu của phôi sơ cấp và thứ cấp.
A,B,C. Phôi thứ cấp hình thành trên thân phôi sơ cấp ở môi trường đặc, hình thái giải
phẫu tương ứng. D,E,F. Phôi thứ cấp hình thành trên thân phôi sơ cấp ở môi trường
lỏng, hình thái giải phẫu tương ứng. G,H,I. Phôi thứ cấp hình thành từ lá mầm phôi
sơ cấp ở mặt dưới, mặt trên, hình thái giải phẫu tương ứng, (vạch đen chỉ vị trí cắt
thu lát mỏng tế bào); J,K. Phôi thứ cấp hình thành ở phần rễ phôi sơ cấp, hình thái
giải phẫu tương ứng, hình vuông chỉ vị trí vùng rễ khảo sát; mũi tên đen/trắng chỉ vị
trí phôi thứ cấp; M: mạch dẫn ở thân phôi, ở lá mầm và rễ phôi; hình chụp dưới kính
hiển vi soi nổi với độ phóng đại 20X)
Sukhada và cộng sự (2010) đã sử dụng kỹ thuật hóa mô để khảo sát sự hình
thành phôi thứ cấp từ phôi sơ cấp chuối (cv. Nanjangud Rasbale) (tái sinh từ mô sẹo
có khả năng sinh phôi từ hoa đực non nuôi cấy trên môi trường MS có 18,10 μM 2,4-
D, 5,37 μM NAA và 5,71 μM IAA) với kết quả đã ghi nhận được phôi thứ cấp hình
thành trực tiếp từ lớp biểu bì (epidermis) của phôi sơ cấp, không qua trung gian giai
đoạn mô sẹo theo hai cách, một là, phôi thứ cấp có nguồn gốc từ tế bào đơn biểu mô

95
của phôi cầu sơ cấp non; hai là, phôi thứ cấp có nguồn gốc từ nhiều tế bào biểu mô
lá mầm của phôi sơ cấp trưởng thành [171]. Trong nghiên cứu tạo phôi NGBCC, đã
ghi nhận được sự hình thành phôi cầu thứ cấp cấp 2 (tertiary) ở mặt dưới của lá (Hình
3.30G, vị trí mũi tên trắng) từ phôi cầu thứ cấp cấp 1; theo chúng tôi, phôi này cũng
có thể có nguồn gốc từ tế bào biểu mô phôi cầu tương tự kết luận của các tác giả vừa
nêu trên.
Phôi thứ cấp Arabidopsis thaliana (L.) Heynh hình thành trực tiếp từ lớp tế
bào bề mặt và dưới bề mặt, mặt trên lá mầm phôi hợp tử non nuôi cấy trên môi trường
B5 có 5 µM 2,4-D [172]; tương tự, ở Dendrobium cv. Chiengmai Pink, phôi thứ cấp
phát sinh từ các lớp tế bào biểu bì mặt trên, ở vị trí đầu lá và gần vị trí cắt của đoạn
lá mầm phôi sơ cấp (1 tháng tuổi, nuôi cấy trên môi trường ½MS có 1 mg/L TDZ)
[91]. Và ở Pistacia vera, phôi hình thành có nguồn gốc tế bào biểu mô hoặc dưới biểu
mô của lá [173].
Cũng qua khảo sát mô tế bào, Wang và cộng sự (2014) cho rằng ngoài tái sinh
phôi theo cách thông qua mô sẹo, các tiền phôi đa bào (multicellular proembryos)
cũng được hình thành trực tiếp từ lớp tế bào bề mặt (surface layer) của phôi non
Tapiscia sinensis qua nuôi cấy trên môi trường MS có bổ sung 1 mg/L 2,4-D, 0,5%
than hoạt tính [94].
Xu và cộng sự (2019) đã ghi nhận được sự hình thành trực tiếp phôi thứ cấp
Ranunculus sceleratus từ bề mặt các loại mẫu cấy nh mảnh lá (1 – 1,5 cm2), đoạn
thân và đoạn rễ (1 - 2 cm) của cây mầm (2 tuần tuổi, cao 4 – 5 cm, có nguồn gốc phôi
hợp tử) nuôi cấy trên môi trường MS có nồng độ NAA cao (10 mg/L) [170]. Phôi thứ
cấp hình thành trực tiếp từ đoạn rễ (~ 8 mm) cây mầm (7 ngày tuổi, từ phôi hợp tử
Brassica oleracea L. var. Italica) nuôi cấy trong môi trường MS có 1 mg/L 2,4-D,
tuy nhiên phôi thứ cấp lại có nguồn gốc từ các lớp tế bào trụ bì (pericycle cell layers)
gần hệ mạch [169]. Ở Quercus robur L., Zegzouti và cộng sự (2001) đã ghi nhận
được hai kiểu sinh phôi thứ cấp khác nhau từ vật liệu nuôi cấy là thân và rễ phôi sơ
cấp (đã bỏ lá mầm): một là, sinh phôi gián tiếp, ở trường hợp này có sự phân chia tế
bào ở mô vùng vỏ dẫn đến sự hình thành mô sẹo, sau đó tạo các khối phôi thứ cấp
tròn từ vùng mô có khả năng phân bào mạnh ở mặt ngoài) mô sẹo; hai là, sinh phôi
trực tiếp, có hai trường hợp xảy ra: (1) sự phân chia tế bào biểu bì trên diện rộng, và

96
(2) sự phân chia của một số tế bào biểu bì (3 – 4 tế bào) dẫn đến kết quả tạo phôi thứ
cấp [76].
Như vậy, sự sinh phôi thứ cấp từ thân, lá và rễ phôi sơ cấp ở NGBCC cũng
theo hướng chung của sự sinh phôi thứ cấp nói chung ở thực vật/cây trồng – tương tự
như kết quả của một số tác giả nêu trên.
3.3. Tạo rễ bất định
3.3.1. Tạo rễ bất định trực tiếp từ mô lá
3.3.1.1. Ảnh hưởng của auxin (NAA/IBA) và môi trường khoáng đến sự tạo rễ bất
định trực tiếp từ mô lá
Ảnh hưởng của auxin (NAA/IBA) và môi trường khoáng đến sự tạo rễ bất
định trực tiếp từ mảnh lá (10 x10 mm)
Ở nghiệm thức đối chứng, môi trường nuôi cấy không bổ sung chất ĐHST
(NAA/IBA), các mẫu cấy đều không có biểu hiện đáp ứng tạo rễ 30 NSC.
Trên môi trường nuôi cấy MS, ½MS, B5, SH có bổ sung IBA ở nồng độ từ
(0,5 – 5mg/L), kết quả cho thấy sau 30 ngày nuôi cấy các mẫu cấy mảnh lá hoàn toàn
không có biểu hiện cảm ứng tạo rễ ở tất cả các nghiệm thức (không trình bày số liệu,
Hình 3.31A,B).
Hình 3.31. Đĩa cấy mảnh lá trên môi trường ½MS có 3 mg/L NAA, 30 NSC.
A. Mặt trên của đĩa cấy mảnh lá, B. Mặt dưới của đĩa cấy mảnh lá
Sau 30 ngày nuôi cấy mẫu trên môi trường MS, ½MS, B5 và SH có NAA ở
tất cả các nghiệm thức, kết quả cho thấy có sự tác động rõ rệt của NAA đến khả năng
tạo rễ xét ở tất cả các chỉ tiêu như tỷ lệ (%) mẫu tạo rễ, số rễ/mẫu, chiều dài rễ (mm)
thể hiện ở (Bảng 3.15, Hình 3.32, Hình 3.33, Hình 3.34, Hình 3.35).
Quan sát mẫu cấy ở nghiệm thức có bổ sung NAA sau 7 ngày cho thấy, mô
xung quanh vết cắt có biểu hiện trương to và rễ xuất hiện ở ngày thứ 12 – 15. Tỷ lệ

97
mẫu ra rễ, số rễ/mẫu, chiều dài rễ tăng theo nồng độ NAA (0,5 - 3 mg/L), hiệu quả
cao nhất ở 3 mg/L NAA. Khi nồng độ NAA tăng trong khoảng 4, 5 mg/L, giá trị các
chỉ tiêu theo dõi đều giảm. Mặt khác, trên các môi trường nuôi cấy khác nhau khi bổ
sung NAA có cùng nồng độ, kết quả về các chỉ tiêu theo dõi khác nhau, chứng tỏ sự
phát sinh rễ bất định chịu ảnh hưởng bởi các mức nồng độ khác nhau của NAA và
các môi trường khoáng khác nhau. Trong nghiên cứu này cho thấy, hiệu quả tạo rễ
trên môi trường ½MS là cao nhất với tỷ lệ mẫu tạo rễ đạt 100%, số rễ/mẫu 69,78;
chiều dài rễ (mm) 16,96, kết quả này khác biệt với các nghiệm thức còn lại, rễ hình
thành sớm hơn ở ngày thứ 12 sau cấy, so với các môi trường còn lại. Rễ hình thành
nhiều ở xung quanh vết cắt, có màu trắng, nhiều lông hút trên tất cả các loại môi
trường.
Trên môi trường B5 có 3 mg/L NAA, rễ có kích thước dài nhất (18,68 mm).
Ở nghiệm thức có bổ sung 3 mg/L NAA, rễ ngắn nhất ghi nhận được khi mẫu nuôi
cấy ở môi trường MS (11,33 mm).

98
Bảng 3.15. Ảnh hưởng của NAA và môi trường khoáng đến sự tạo rễ bất định trực
tiếp từ mảnh lá (10 x 10 mm), ở môi trường ½MS, 30 NSC.
NAA
(mg/L)
Môi trường
khoáng
Tỷ lệ mẫu
tạo rễ (%)
Số rễ/mẫu Chiều dài rễ
(mm)
0 MS 00,0r* 00,0p 0,00o
0,5 MS 62,22q 11,61n 8,63l
1 MS 64,45o 15,92l 9,22k
2 MS 70,00lm 23,71g 10,44j
3 MS 84,45h 30,32e 11,33i
4 MS 81,11ij 26,60f 8,25m
5 MS 66,67n 20,82j 7,54n
0 ½MS 00,0o 00,0p 0,00o
0,5 ½MS 80,00j 18,49k 12,28h
1 ½MS 84,44h 26,76f 13,50f
2 ½MS 91,11cd 43,56c 15,54d
3 ½MS 100,00a 69,78a 16,96b
4 ½MS 94,45b 56,41b 10,56j
5 ½MS 88,89ef 36,05d 9,54k
0 B5 00,0o 00,0p 0,00o
0,5 B5 68,89m 9,46o 13,71f
1 B5 75,56k 12,72mn 14,50e
2 B5 86,67g 18,68k 16,57c
3 B5 92,22c 26,53f 18,68a
4 B5 90,00de 23,15gh 11,52i
5 B5 82,22i 16,33l 10,66j
0 SH 00,0o 00,0p 0,00o
0,5 SH 63,33po 11,67n 9,56k
1 SH 66,67n 13,98m 10,81j
2 SH 74,45k 21,89hi 11,38i
3 SH 87,78fg 29,08e 12,69g
4 SH 82,22i 26,15f 9,22k
5 SH 71,11l 16,89l 8,69l
*Các chữ cái khác nhau trong một cột thể hiện sự khác biệt có ý nghĩa ở mức
p ≤ 0, 05 trong phép thử Duncan. Số liệu (%) được chuyển đổi sang dạng arcsin √𝑥
khi phân tích thống kê.

99
Hình 3.32. Tạo rễ trực tiếp từ mảnh lá
(10 x 10 mm) ở môi trường MS, có
NAA, 30 NSC.
A. 0,5 mg/L, B. 1 mg/L, C. 2 mg/L,
D. 3 mg/L, E. 4 mg/L, F. 5 mg/L.
Hình 3.33. Tạo rễ trực tiếp từ mảnh lá (10
x 10 mm) ở môi trường ½MS, có NAA,
30 NSC.
A. 0,5 mg/L, B. 1 mg/L, C. 2 mg/L,
D. 3 mg/L, E. 4 mg/L, F. 5 mg/L.
Hình 3.34. Tạo rễ trực tiếp từ mảnh lá
(10 x 10 mm) ở môi trường B5, có
NAA, 30 NSC.
A. 0,5 mg/L, B. 1 mg/L, C. 2 mg/L,
D. 3 mg/L, E. 4 mg/L, F. 5 mg/L.
Hình 3.35. Tạo rễ trực tiếp từ mảnh lá
(10 x 10 mm) ở môi trường SH, có
NAA, 30 NSC.
A. 0,5 mg/L, B. 1 mg/L, C. 2 mg/L,
D. 3 mg/L, E. 4 mg/L, F. 5 mg/L.
Ảnh hưởng của auxin (NAA/IBA) và môi trường khoáng đến sự tạo rễ bất
định trực tiếp từ mảnh lá ( 3 x 10 mm)
Tương tự như trường hợp mẫu cấy mảnh lá (10 x10 mm), sau 30 ngày cấy ở
nghiệm thức đối chứng (không có NAA) và tất cả các nghiệm thức có IBA (0,5 – 5

100
mg/L) hoàn toàn không ghi nhận được hiện tượng tạo rễ. Như vậy, lần nữa cho thấy
mô lá hoàn toàn không có biểu hiện cảm ứng phát sinh rễ bất định đối với IBA.
NAA có tác động tích cực đến cảm ứng tạo rễ, mẫu cũng có đáp ứng tạo mô
sẹo tuy không nhiều sau 5 ngày sau nuôi cấy trên môi trường MS và rễ xuất hiện sớm
ở ngày thứ 12 nuôi cấy. Các nồng độ khác nhau của NAA đều có tác động kích thích
quá trình tạo rễ, giá trị các chỉ tiêu theo dõi tăng tỷ lệ thuận với nồng độ NAA (0,5 -
3 mg/L), giảm khi nồng độ NAA cao hơn 3 mg/L (Bảng 3.16). Môi trường tạo rễ bất
định hiệu quả nhất vẫn là ½MS có 3 mg/L NAA cụ thể tỷ lệ mẫu tạo rễ 100%; số
rễ/mẫu 19,67; chiều dài rễ (6,29 mm), rễ hình thành sớm sau 10 ngày cấy. Rễ hình
thành trên môi trường ½MS và B5 (Hình 3.37, 3.38) đều dài hơn rễ ở môi trường MS,
SH (Hình 3.36, 3.39). Các giá trị trung bình của các chỉ tiêu theo dõi đều có sự khác
biệt về mặt thống kê.
Kết quả đáp ứng nhanh của mẫu cấy mảnh lá có kích thước nhỏ (3 x 10 mm),
tỷ lệ mẫu tạo rễ cao hơn so với mẫu cấy mảnh lá có thể do mẫu cấy có kích thước
nhỏ, bề mặt tiếp xúc với môi trường lớn; nên tiếp nhận các chất từ môi trường nhanh
hơn. Tuy nhiên, chiều dài rễ ngắn hơn so với rễ từ mảnh lá ở cùng điều kiện nuôi cấy,
có thể do hàm lượng chất nội sinh trong mẫu ít hơn.
Nói chung, vật liệu mô lá, như một hệ thống nuôi cấy chứa protein sắc tố nhạy
sáng phytochrome, thường được sử dụng trong nghiên cứu tạo rễ bất định vì: (1)
auxin ngoại sinh có thể tác động dễ dàng đến loại tế bào có nhiều tiềm năng (tế bào
liên kết với mạch dẫn) dẫn đến phân chia, biệt hóa tạo một cơ quan nhất định trong
đó có rễ [174][175]; (2) lá còn chứa mô tế bào khuyết cũng có thể phản biệt hóa và
tái biệt hóa tạo sơ khởi rễ dưới tác động của auxin ngoại sinh trong quá trình nuôi cấy
như ở trường hợp tái sinh rễ Sainpaulia ionantha [103]; (3) lá cũng là cơ quan có khả
năng sinh tổng hợp auxin nội sinh - có thể có liên quan nhất định đến sự tạo sơ khởi
rễ [176]. Theo Yu và cộng sự (2017), rễ bất định tái sinh từ mảnh lá Arabidopsis
thaliana là kết quả của một số hoạt động sinh lý trong mô tế bào như: (1) Auxin nội
sinh được vận chuyển đến các tế bào có khả năng tái sinh (procambium và tế bào nhu
mô mạch), kích chuyển tái lập trình di truyền các tế bào này thành các tế bào ‘nguồn’;
(2) Các tế bào ‘nguồn’ phát triển thành sơ khởi rễ - nơi tích tụ auxin cao; (3) Các tế
bào sơ khởi rễ tiếp tục phân chia tạo mô phân sinh đầu rễ - nơi auxin giảm tích tụ; (4)
Đầu rễ đạt mức trưởng thành, tiếp tục phát triển ra khỏi mảnh lá [176].

101
Bảng 3.16. Ảnh hưởng của NAA và môi trường khoáng đến sự tạo rễ bất định trực
tiếp từ mảnh lá (3 x 10 mm), ở môi trường ½MS, 30 NSC.
NAA
(mg/L)
Môi trường
khoáng
Tỷ lệ mẫu tạo
rễ (%)
Số rễ/mẫu Chiều dài rễ
(mm)
0 MS 00,0o* 00,0s 0,00p
0,5 MS 63,33n 8,46q 8,23m
1 MS 66,67m 9,52mn 8,61k
2 MS 78,89j 12,39h 9,28i
3 MS 87,78e 15,78d 10,79g
4 MS 82,22f 13,46f 7,29l
5 MS 72,22h 10,69l 6,45o
0 ½MS 00,0o 00,0s 0,00p
0,5 ½MS 82,22h 9,67m 10,50h
1 ½MS 86,67fg 11,74i 11,48e
2 ½MS 93,33c 15,52e 12,32d
3 ½MS 100,00a 19,67a 16,29a
4 ½MS 96,67b 17,33b 9,41i
5 ½MS 88,89e 13,06g 8,48kl
0 B5 00,0o 00,0s 0,00p
0,5 B5 71,11l 6,89r 11,46ef
1 B5 81,11hi 8,22q 12,43d
2 B5 88,89e 9,33n 13,51c
3 B5 95,56b 11,50j 14,65b
4 B5 92,22cd 10,50l 10,41h
5 B5 85,56g 8,80o 9,31i
0 SH 00,0o 00,0s 0,00p
0,5 SH 65,56m 8,41qr 8,81j
1 SH 71,11l 9,52mn 9,28i
2 SH 80,00ij 12,46h 11,28f
3 SH 91,11d 16,59c 12,29d
4 SH 85,56g 13,36f 8,36l
5 SH 75,56k 11,04k 7,76n
*Các chữ cái khác nhau trong một cột thể hiện sự khác biệt có ý nghĩa ở mức
p ≤ 0, 05 trong phép thử Duncan. Số liệu (%) được chuyển đổi sang arcsin√𝑥 khi
phân tích thống kê.

102
Hình 3.36. Tạo rễ trực tiếp từ mảnh lá (3
x 10 mm) ở môi trường MS, có NAA, 30
NSC.
A. 0,5 mg/L B. 1 mg/L C. 2 mg/L
D. 3 mg/L E. 4 mg/L, F. 5 mg/L.
Hình 3.37. Tạo rễ trực tiếp từ mảnh lá (3
x 10 mm) ở môi trường ½MS, có NAA,
30 NSC.
A. 0,5 mg/L B. 1 mg/L C. 2 mg/L
D. 3 mg/L, E. 4 mg/L, F. 5 mg/L.
Hình 3.38. Tạo rễ trực tiếp từ mảnh lá (3
x 10 mm) ở môi trường B5, có NAA, 30
NSC.
A. 0,5 mg/L B. 1 mg/L C. 2 mg/L
D. 3 mg/L E. 4 mg/L, F. 5 mg/L.
Hình 3.39. Tạo rễ trực tiếp từ mảnh lá (3
x 10 mm) ở môi trường SH, có NAA, 30
NSC.
A. 0,5 mg/L B. 1 mg/L C. 2 mg/L
D. 3 mg/L E. 4 mg/L, F. 5 mg/L.
Hệ thống LMTB gồm những mẫu cấy có kích thước nhỏ (1 - vài mm), hàm
chứa ít chất điều hòa sinh trưởng nội sinh, nên sự phát sinh hình thái phụ thuộc chủ
yếu vào chất ĐHST bổ sung vào môi trường. Hệ thống này có ưu điểm là thời gian
phát sinh hình thái tương đối ngắn do đáp ứng nhanh với nuôi cấy, tỷ lệ phát sinh
hình thái cao, đa dạng về hình thái phát sinh tùy môi trường nuôi cấy. Do đó, hệ thống
này đã được ứng dụng hiệu quả trong thực tiễn nuôi cấy in vitro nhằm nghiên cứu cơ

103
bản về phát sinh hình thái, nhân giống cây trồng [178].
Kết quả của thí nghiệm tạo rễ bất định trực tiếp từ mô lá NGBCC cho thấy,
môi trường ½MS có 3 mg/L NAA được xem là rất thích hợp cho tạo rễ bất định trực
tiếp từ mô lá của NGBCC; ngược lại, IBA hoàn toàn không gây đáp ứng tạo rễ bất
định. Ling và cộng sự (2013) cho rằng trường hợp NAA có tác động tốt đối với tạo
rễ là do NAA được oxy hóa hiệu quả thành IAA để tế bào sử dụng [179]. Tác động
âm tính đối với tạo rễ có thể do IBA không được chuyển đổi thành IAA bởi quá trình
β-oxy hóa ở các thể peroxisome trong tế bào [180].
Đã ghi nhận NAA, ở dạng riêng lẻ hoặc kết hợp với auxin khác, là tác nhân
kích thích tạo rễ tốt từ mành lá nhiều loài thực vật được nuôi cấy in vitro như
Eurycoma longifolia khi dùng 3 mg/L NAA [181] hoặc 3 mg/L NAA (kết hợp với 1
mg/L IBA) là thích hợp nhất cho tạo rễ từ mảnh lá Gynura procumbens [182]. IBA
cũng là chất điều hòa sinh trưởng rất thích hợp trong sử dụng tạo rễ đối với nhiều cây
dược liệu [98]. Tuy nhiên, ở một số đối tượng thực vật, IBA (kể cả IAA) hoàn toàn
không thích hợp, ví dụ như ở trường hợp tạo rễ bất định từ mảnh lá Andrographis
paniculata trên môi trường MS chỉ tạo được rễ bất định khi sử dụng NAA (1 mg/L)
[183]; cũng nhận thấy chồi Diospyros crassiflora không tạo rễ bất định trên môi
trường ½MS có IBA (2,5 - 19,6 µM). Nói chung, loại và nồng độ auxin thích hợp cho
cảm ứng tạo rễ và kéo dài rễ bất định phụ thuộc chủ yếu vào loài thực vật [184].
Như đã trình bày mô lá tạo rễ bất định tốt nhất trên môi trường khoáng ½MS
ở tất cả các chỉ tiêu theo dõi so với các môi trường MS, B5 và SH. Theo Khalafalla
và cộng sự (2009), đáp ứng tạo rễ từ mô lá phụ thuộc vào thành phần khoáng cơ bản
của môi trường và hàm lượng đạm nitrate. Tương tự như một số loài thực vật khác,
theo chúng tôi, có thể do nhu cầu về khoáng đa lượng đối với NGBCC ở mức tương
đối thấp, cụ thể tổng khoáng đa lượng ở môi trường ½MS là thấp nhất (2.116 mg/L)
so với tổng khoáng đa lượng ở các môi trường MS, B5 và SH cao hơn nhiều, lần lượt
là 4.232 mg/L, 2.999 mg/L và 3.146 mg/L; và trong đó có thể nhu cầu về đạm nitrate
cũng ở mức thấp hơn – 1.775 mg/L ở ½MS so với các môi trường MS, B5 và SH -
cao hơn nhiều là 3.550 mg, 2.500 mg và 2.500 mg, theo thứ tự. Cũng theo Khalafalla
và cộng sự (2009), trong mối tương quan với đạm nitrate, đạm ammonium (NH4)
cũng đóng vai trò quan trọng trong tạo rễ và cho rằng số rễ Vernonia amygdalina tái
sinh từ mảnh lá trên môi trường B5 ít hơn so với môi trường MS có thể do thành phần

104
đạm ammonium (NH4)2SO4 trong môi trường B5 ít hơn (134 mg/L) ở môi trường MS
(NH4NO3, 1.650 mg/L) [185]. Ở NGBCC, số rễ tái sinh ít trên môi trường B5 cũng
có thể do yếu tố ammonium tương tự như trường hợp của các tác giả nêu trên. Trong
môi trường SH có đạm NH4PO4 phôi Panax quinquefolius nẩy mầm tạo rễ trụ tốt trên
môi trường có khoáng thấp - ½SH [36]; tương tự, chồi Panax ginseng (có nguồn gốc
phôi vô tính) phát triển rễ trụ tốt trên môi trường 1/3SH [41]. Rễ Panax ginseng tạo
sinh khối tốt trong môi trường SH chỉ có ½NH4PO4 [186]. Ở NGBCC, đáp ứng tái
sinh rễ không tốt có thể do nồng độ NH4PO4 ở môi trường SH không phù hợp hoặc
loại đạm ammonium này khác với NH4NO3, loại thích hợp hơn cho loài thực vật này
ở môi trường MS, ½MS hoặc/và khác với (NH4)2SO4 ở môi trường B5.
Như vậy, kết quả của nghiên cứu này cho thấy NAA có vai trò quan trọng
trong việc tạo rễ bất định trực tiếp từ mô lá, tỷ lệ % mẫu tạo rễ, số rễ/mẫu và chiều
dài rễ khác nhau chịu ảnh hưởng bởi các mức nồng độ của NAA và hàm lượng –
thành phần của môi trường khoáng.
3.1.1.2. Ảnh hưởng của nồng độ đường đến sự tạo rễ bất định trực tiếp từ mô lá
Ảnh hưởng của nồng độ đường đến sự tạo rễ bất định trực tiếp từ mảnh lá
(10 x 10 mm)
Kế thừa kết quả thí nghiệm trên, môi trường ½MS có bổ sung 3 mg/L NAA
được sử dụng để nghiên cứu xác định nồng độ đường thích hợp cho tạo rễ bất định.
Bảng 3.17. Ảnh hưởng của nồng độ đường đến sự tạo rễ bất định trực tiếp từ mảnh
lá (10 x 10 mm), ở môi trường ½MS, 30 NSC.
Nồng độ
sucrose (g/L)
Tỷ lệ mẫu
tạo rễ (%)
Số rễ/mẫu
Chiều dài rễ
(mm)
20 92,22b* 58,35b 14,63b
30 100,00a 70,50a 16,70a
40 87,78c 46,44c 12,26c
50 85,56d 36,24d 10,47d
*Các chữ cái khác nhau trong một cột thể hiện sự khác biệt có ý nghĩa ở mức
p ≤ 0, 05 trong phép kiểm định LSD. Số liệu (%) được chuyển đổi sang (x+0,5)1/2 khi
xử lý thống kê.
Ở 30 ngày sau nuôi cấy, kết quả thí nghiệm cho thấy các nghiệm thức có nồng
độ sucrose khác nhau cho kết quả khác biệt về mặt thống kê ở tất cả các chỉ tiêu theo

105
dõi, trong đó nồng độ sucrose 30 g/L là tối ưu cho tạo rễ ở mảnh lá có tỷ lệ mẫu tạo
rễ 100%, số rễ/mẫu 70,50; chiều dài rễ 16,70 mm (Bảng 3.17), ở nồng độ sucrose cao
50 g/L các chỉ tiêu theo dõi đều giảm (tỷ lệ mẫu tạo rễ 85,56%, số rễ/mẫu 36,24; chiều
dài rễ 10,47 mm) rễ hơi vàng, ít lông hút hơn, rễ ngắn hơn (Hình 3.40).
Mô tế bào thực vật nuôi cấy in vitro ít hoặc không có khả năng tự dưỡng nên
cần bổ sung nguồn carbon. Đường là nguồn carbon được sử dụng phổ biến, trong
nuôi cấy mô giúp hỗ trợ quá trình phân bào và phát sinh hình thái; ngoài ra, nồng độ
đường thích hợp còn làm gia tăng hàm lượng hợp chất thứ cấp trong rễ [98].
Hình 3.40. Ảnh hưởng của nồng độ đường đến sự tạo rễ từ mảnh lá (10 x 10 mm), ở
môi trường ½MS, 30 NSC.
A,B,C,D. Đĩa cấy mảnh lá tạo rễ ở môi trường ½MS có sucrose 20, 30, 40, 50 g/L.
Ảnh hưởng của nồng độ đường đến sự tạo rễ bất định trực tiếp từ mảnh lá
(3 x 10 mm)
Kết quả thí nghiệm cho thấy, sau nuôi cấy 30 ngày các mẫu cấy ở môi trường có
30 g/L sucrose tạo rễ tốt nhất với tỷ lệ mẫu tạo rễ 100%; số rễ/mẫu 22,65; chiều dài
rễ 15,40 mm; thấp nhất khi môi trường có bổ sung sucrose với nồng độ cao 50 g/L,
các chỉ tiêu theo dõi đều giảm 82,59%; số rễ/mẫu 9,37; chiều dài rễ 9,54 mm. Ở các
nghiệm thức có nồng độ sucrose 20 g/L, 40 g/L chỉ tiêu tỷ lệ mẫu tạo rễ, chiều dài rễ
có khác biệt thống kê nhưng số rễ/mẫu không khác biệt về thống kê. Từ kết quả này
cho thấy, các mức nồng độ sucrose khác nhau có ảnh hưởng khác biệt đến tạo rễ bất
định. Tỷ lệ % mẫu tạo rễ ở mẫu cấy mảnh lá có kích thước nhỏ (3 x 10 mm) cao hơn
mẫu cấy mảnh lá (10 x 10 mm) do đáp ứng của mẫu cấy với môi trường nuôi cấy tốt
hơn. Tuy nhiên, ở nghiệm thức nồng độ sucrose cao 50 g/L, ở mẫu cấy mảnh lá kích
thước nhỏ (3 x 10 mm) lại có tỷ lệ mẫu tạo rễ ít hơn so với mẫu cấy mảnh lá do ở
nồng độ đường cao làm áp suất thẩm thấu tăng, mô cấy thiếu nước ảnh hưởng đến
sinh lý của mô cấy.

106
Bảng 3.18. Ảnh hưởng của nồng độ đường đến sự tạo rễ bất định trực tiếp từ mảnh
lá (3 x 10 mm), ở môi trường ½MS, 30 NSC.
Nồng độ
sucrose (g/L)
Tỷ lệ mẫu
tạo rễ (%)
Số rễ/mẫu
Chiều dài rễ
(mm)
20 95,19b* 11,59b 13,61b
30 100,00a 22,65a 15,40a
40 91,11c 11,54b 11,53c
50 82,59d 9,37c 9,54d
*Các chữ cái khác nhau trong một cột thể hiện sự khác biệt ý nghĩa với p ≤ 0,05
trong phép thử LSD. Số liệu (%) được chuyển đổi sang (x+0,5)1/2 khi xử lý thống kê.
Hình 3.41. Ảnh hưởng của nồng độ đường đến sự tạo rễ trực tiếp từ mảnh lá (3 x 10
mm), ở môi trường ½MS, 30 NSC.
A,B,C,D. Đĩa cấy mảnh lá (3 x 10 mm) tạo rễ ở môi trường ½MS có sucrose 20, 30,
40, 50 g/L.
Như vậy, môi trường ½MS có bổ sung 3 mg/L NAA với 30 g/L sucrose là
thích hợp nhất cho tạo rễ bất định từ mô lá NGBCC. Kết quả này cũng phù hợp với
một số kết quả nghiên cứu sử dụng đường sucrose 30 g/L để tạo rễ bất định Panax
vietnamensis [187], Panax stipuleanatus [46], sucrose 30 g/L là tối ưu cho rễ Panax
ginseng phát triển [188].
3.1.1.3. Ảnh hưởng của điều kiện chiếu sáng đến sự tạo rễ bất định trực tiếp từ mô
lá
Ảnh hưởng của điều kiện chiếu sáng đến sự tạo rễ bất định trực tiếp từ
mảnh lá (10 x 10 mm)
Trong nghiên cứu tạo rễ bất định, ánh sáng được cho là tác nhân ức chế sự
hình thành rễ, nhất là giai đoạn hình thành sơ khởi rễ, điều kiện tối cần cho giai đoạn
đầu của quá trình ra rễ, chu kỳ tối thích hợp để rễ phát triển 3 - 10 ngày tùy thuộc vào
loài thực vật khác nhau [189]. Nhiều nghiên cứu tạo rễ bất định đạt kết quả tốt trong

107
điều kiện tối. Tuy nhiên ở đối tượng NGBCC, nhận thấy ánh sáng lại có tác động tích
cực đến sự thành rễ bất định từ mô lá. Số liệu (Bảng 3.19) cho thấy, các chỉ tiêu theo
dõi có sự khác biệt rõ rệt giữa các nghiệm thức. Ở điều kiện chiếu sáng, mẫu tạo rễ
với tỷ lệ rất cao, chiếm 95,56% (2.000 lux), 100% (4.000 lux); rễ hình thành với số
lượng nhiều nhất 68,80 rễ/mẫu (4.000 lux), thấp nhất mẫu cấy ở điều kiện tối. Các
mẫu cấy mảnh lá nuôi cấy ở cường độ chiếu sáng khác nhau, thì rễ hình thành có
chiều dài khác nhau, mẫu cấy mảnh lá ở điều kiện tối rễ hình thành dài nhất (19,77
mm), mẫu cấy mảnh lá ở điều kiện chiếu sáng 4.000 lux tạo rễ có chiều dài cũng khá
tốt (16,53 mm). Như vậy, sự khác biệt về các chỉ tiêu theo dõi ở các nghiệm thức cho
thấy, điều kiện sáng, tối có ảnh hưởng đến sự phát sinh rễ từ mảnh lá (Hình 3.42)
Bảng 3.19. Ảnh hưởng của điều kiện chiếu sáng đến sự tạo rễ bất định trực tiếp từ
mảnh lá (10 x 10 mm), ở môi trường ½MS, 30 NSC
Điều kiện chiếu
sáng (lux)
Tỷ lệ mẫu
tạo rễ (%)
Số
rễ/mẫu
Chiều dài
rễ (mm)
Đặc điểm rễ
4.000 100a* 68,80a 16,53b Rễ dài, nhiều lông hút, màu
trắng đục
2.000 95,56b 30,03b 15,37c Rễ dài, nhiều lông hút, màu
trắng
Tối 93,33c 27,20c 19,77a Rễ rất dài, ít lông hút, một số
rễ màu hơi vàng
*Các chữ cái khác nhau trong một cột thể hiện sự khác biệt ý nghĩa với p ≤ 0,05
trong phép thử LSD. Số liệu (%) được chuyển đổi sang (x+0,5)1/2 khi xử lý thống kê.
Hình 3.42. Ảnh hưởng của điều kiện chiếu sáng đến sự tạo rễ bất định từ mảnh lá
(10 x 10 mm), ở môi trường ½MS, 30 NSC.
A,B,C. Đĩa cấy mảnh lá tạo rễ ở cường độ chiếu sáng 4.000 lux, 2.000 lux, tối; D.
Mảnh lá tạo rễ ở môi trường ½MS có 3 mg/L NAA, 4000 lux. Thanh ngang 10 mm.

108
Ảnh hưởng của điều kiện chiếu sáng đến sự tạo rễ bất định trực tiếp từ
mảnh lá (3 x 10 mm)
Đối với mẫu cấy mảnh lá (3 x 10 mm), nhận thấy hiệu quả tạo rễ cao nhất ở
nghiệm thức có cường độ chiếu sáng 4.000 lux (100% mẫu tạo rễ; 21,96 rễ/mẫu; chiều
dài rễ 15,53 mm) so với mẫu cấy ở cường độ chiếu sáng 2.000 lux và điều kiện tối
(Bảng 3.20, Hình 3.41). Ở điều kiện chiếu sáng khác nhau, kết quả tạo rễ khác nhau
về tỷ lệ % mẫu tạo rễ, số rễ/mẫu, chiều dài rễ (mm).
Bảng 3.20. Ảnh hưởng của điều kiện chiếu sáng đến sự tạo rễ bất định trực tiếp từ
mảnh lá (3 x 10 mm), ở môi trường ½MS, 30 NSC.
Điều kiện chiếu
sáng (lux)
Tỷ lệ mẫu
tạo rễ (%)
Số
rễ/mẫu
Chiều dài
rễ (mm)
Đặc điểm rễ
4.000 100a* 21,96a 15,53a Rễ dài, nhiều lông hút, màu
trắng đục
2.000 94,44b 16,19b 13,43b Rễ ngắn, nhiều lông hút,
màu trắng
Tối 91,11c 14,11c 14,37c Rễ ngắn, nhiều lông hút,
một số rễ màu hơi vàng
*Các chữ cái khác nhau trong một cột thể hiện sự khác biệt ý nghĩa với p ≤ 0,05
trong phép thử LSD. Số liệu (%) được chuyển đổi sang (x+0,5)1/2 khi xử lý thống kê.
Hình 3.43. Ảnh hưởng của điều kiện chiếu sáng đến sự tạo rễ bất định từ mảnh lá
(3 x 10 mm), ở môi trường ½MS, 30 NSC.
A,B,C. Đĩa cấy mảnh lá tạo rễ ở cường độ chiếu sáng 4.000 lux, 2.000 lux, tối; D.
Mảnh lá tạo rễ ở môi trường ½MS có 3 mg/L NAA, 4000 lux. Thanh ngang 10 mm.
Như vậy, kết quả thí nghiệm cho thấy sự cảm ứng tạo rễ bất định từ mô lá,
hiệu quả cao nhất ở điều kiện chiếu sáng 4000 lux, các chỉ tiêu theo dõi đều có giá trị
cao nhất. Trong tự nhiên, sự tạo rễ bất định thuận lợi hơn khi ở điều kiện tối, nhưng
trong nuôi cấy mô với điều kiện môi trường tối ưu (khoáng, chất ĐHST,..), nhiều

109
nghiên cứu cho thấy sự phát sinh rễ bất định thuận lợi hơn ở điều kiện sáng (phụ
thuộc kiểu gen thực vật); ngoài ra, nhiều công bố cho thấy khi nuôi ở điều kiện sáng
mạnh, ở rễ nhiều loài thực vật hình thành cấu trúc tương tự như lục lạp (chloroplast-
like structure) có thể liên quan đến sự hình thành và tăng trưởng của rễ.
Đến nay đã ghi nhận nhiều nghiên cứu tạo rễ bất định với kết quả tốt với các
điều kiện chiếu sáng khác nhau như tối - ví dụ trường hợp nuôi cấy mảnh lá Cichorium
intybus [184], Centella asiatica [190], 40 µmol m-2 s-1 (3.000 lux) đối với Artemisia
amygdalina [191], và 4.000 - 5.000 lux ở nuôi cấy tạo rễ từ mô lá Vernonia
amygdalina [185]. Tương tự một số trường hợp nuôi cấy tạo rễ nêu trên, kết quả
nghiên cứu này cho thấy mảnh lá NGBCC tái sinh rễ bất định tốt nhất ở nghiệm thức
có cường độ chiếu sáng 4.000 lux.
3.1.1.4. Minh họa sự tái sinh rễ trực tiếp và khảo sát hình thái giải phẫu rễ tái
sinh trực tiếp
Như đã trình bày, trong nghiên cứu này rễ bất định hình thành từ mô lá được
định nghĩa theo cách tái sinh trực tiếp ngay ở lần nuôi cấy đầu tiên, khác với tái sinh
gián tiếp qua trung gian giai đoạn mô sẹo.
Hình 3.44. Minh họa cận cảnh sự tái sinh trực tiếp rễ bất định từ mảnh lá nuôi cấy
trên môi trường ½MS, 12 – 20 NSC.
A,B,C. Cận cảnh sự hình thành giai đoạn đầu của rễ đơn, cụm 2 rễ và cụm nhiều rễ ở
12 NSC; D,E,F. Sự phát triển tiếp theo của rễ đơn, cụm 2 rễ và cụm nhiều rễ, ở 20
ngày sau cấy.
Khá nhiều kết quả nghiên cứu về tái sinh trực tiếp rễ bất định từ mô lá đã được
ghi nhận như tạo rễ Ophiorrhiza prostrata dùng môi trường ½MS với 10,74 µM NAA

110
và 2,32 µM kinetin [, tạo rễ Centella asiatica trên môi trường MS có 1 mg/L IBA
[190], tạo rễ Rauwolfia serpentina trên môi trường MS có 1 mg/L PABA (para-amino
benzoic acid) và 4 mg/L NAA [192]. Rễ Gynura procumbens tái sinh trực tiếp trên
môi trường MS có bổ sung 3 mg/L IBA; ngược lại, ở môi trường MS có 3 mg/L NAA
rễ tái sinh gián tiếp thông qua mô sẹo [182]. Ở NGBCC đây là kết quả nghiên cứu
đầu tiên về tái sinh rễ trực tiếp từ vật liệu mảnh lá, được minh họa bằng các hình rễ
tái sinh sau 12 đến 20 ngày nuôi cấy qua chụp cận cảnh (Hình 3.44).
Các sơ khởi rễ (giai đoạn ngày thứ 10 sau cấy) cũng đã được khảo sát hình thái
giải phẫu với kết quả được trình bày dưới đây (Hình 3.45).
Hình 3.45. Hình thái giải phẫu sơ khởi rễ bất định hình thành trực tiếp từ mảnh lá.
A,B. Sơ khởi rễ hình thành ở vị trí gân phụ của lá và hình thái giải phẫu tương ứng;
C,D. Sơ khởi rễ hình thành không ở vị trí gân phụ của lá và hình thái giải phẫu tương
ứng (hình chụp dưới kính hiển vi soi nổi với độ phóng đại 20X) (GP: Gân phụ của lá,
BBD: Biểu bì mặt dưới của phiến lá, CR: Chóp rễ, M: vị trí bó mạch (cắt dọc), NMK:
Nhu mô khuyết phiến lá, NMG: Nhu mô giậu phiến lá; vòng tròn chỉ vị trí mạch dẫn
phụ của lá; thanh ngang 1 mm).
Sau 10 ngày nuôi cấy trên môi trường ½MS, dưới tác động của NAA (3 mg/L
NAA) nhận thấy mép cắt và mô ở vị trí sát mép cắt của mẫu phiến lá phù ra, sơ khởi
rễ hình thành trên bề mặt mẫu phiến lá gần mép cắt. Ở nghiên cứu này, có hai trường
hợp khác nhau về vị trí tạo sơ khởi rễ đã được ghi nhận, một là, sơ khởi rễ hình thành
ở ngay vị trí gân phụ của mẫu cấy với các cụm tế bào đang phân chia mạnh xuất phát
từ các tế bào ở vị trí gần kề mạch (Hình 3.45A,B), hai là, sơ khởi rễ hình thành ở vị
trí không có gân phụ - cũng ghi nhận có các cụm tế bào đang phân chia mạnh nhưng
xuất phát từ các tế bào ở vị trí lục mô khuyết (Hình 3.45C,D). Ở trường hợp 1 sơ khởi
rễ hình thành từ các tế bào liên kết với mô mạch, dưới tác động của NAA, các tế bào

111
này phân chia hình thành lớp tế bào tương tự tế bào tiền tượng tầng (procambial-like
cells) đóng vai trò như tế bào gốc có nhiều tiềm năng (pluripotent stem cells) trong
phân chia, phân hóa tạo sơ khởi rễ - tương tự kết quả nuôi cấy tạo rễ từ mảnh lá
Medicago truncatula dưới tác động của auxin NAA [104], nuôi cấy tạo rễ từ mảnh lá
Orthosiphon stamineus dưới ảnh hưởng của IAA /IBA/ NAA [193] và tạo rễ từ mảnh
lá Labisia pumila dưới tác động của IBA/NAA [179]. Trước đó, Samaj và cộng sự
(1999) cũng đã kết luận các lớp tế bào thuộc bó mạch (bundle sheet cells) chịu trách
nhiệm phân chia, phân hóa tạo rễ trực tiếp ở Helianthus occidentalis [194]. Ở trường
hợp 2, theo chúng tôi, sơ khởi rễ hình thành từ mô tế bào khuyết của mô thịt lá (spongy
parenchyma/mesophyll cells) tương tự trường hợp nuôi cấy tái sinh chồi/rễ từ mảnh
lá cây African violet (Sainpaulia ionantha) dưới ảnh hưởng của BA/ IBA [103]. Tái
sinh rễ bất định ở hai trường hợp trên, theo chúng tôi, được giải thích là do các tế bào
ở vị trí gần mạch và tế bào nhu mô khuyết, dưới tác động của sự di chuyển phân cực
auxin đã được kích hoạt thay đổi lập trình di truyền dẫn đến tái sinh rễ [15].
3.3.2. Tạo rễ bất định từ chồi
3.3.2.1. Tạo rễ bất định từ chồi có nguồn gốc từ đốt thân cây vườn ươm
Bảng 3.21. Ảnh hưởng của NAA, IBA đến sự tạo rễ bất định từ chồi có nguồn gốc từ
đốt thân cây vườn ươm, ở môi trường ½MS, 60 NSC.
Nghiệm
thức
Tỷ lệ chồi
tạo rễ (%)
Số
rễ/chồi
Chiều dài
rễ (mm)
Đặc điểm hình thái,
màu sắc rễ
ĐC 100 4,33c* 69,37c Rễ mảnh, có rễ hơi nâu, ít lông
hút, rễ nhánh dài
NAA 100 13,53a 80,00a Rễ có đường kính to, trắng,
nhiều lông hút, rễ nhánh ngắn
IBA 100 11,67b 74,56b Rễ có đường kính trung bình,
ít lông hút, rễ nhánh ngắn
*Các chữ cái khác nhau trong một cột thể hiện sự khác biệt ý nghĩa với p ≤
0,05 trong phép thử LSD.
Các đoạn thân sau khử trùng được nuôi cấy trên môi trường MS có 0,5 mg/L
BA và 2 mg/L kinetin. Kết quả cho thấy, dưới tác động kích thích của BA và kinetin,
sau nuôi cấy từ 20 đến 45 ngày, chồi nách hình thành và phát triển, đến ngày thứ 60
sau cấy đạt kích thước (7 – 10 mm), xuất hiện 4 – 5 lá, có hình thái đặc trưng. Chồi

112
nách được tách ra cấy chuyển sang môi trường tạo rễ bất định, nhận thấy phần gốc
mẫu cấy tiếp xúc với môi trường phù to do sùi mô sẹo, theo chúng tôi, đây là hiện
tượng thường gặp trong nuôi cấy mô cây thân gỗ.
Các chồi in vitro được nuôi cấy tạo rễ bất định dùng môi trường ½MS có hoặc
không bổ sung 0,2 mg/L IBA/NAA. Kết quả cho thấy trên môi trường không có
NAA/IBA, rễ hình thành ít, rễ dài, có rễ màu hơi nâu, có ít lông hút, rễ nhánh dài
(Hình 3.46A,D). Ở môi trường có IBA, chồi cũng tạo nhiều rễ, có ít lông hút, rễ nhánh
ngắn (Hình 3.46B,E). Trên môi trường có NAA, chồi tạo rễ nhiều, rễ có đường kính
to, dài, màu trắng, có nhiều lông hút, rễ nhánh cũng ngắn (Hình 3.46C,F). Kết quả
(Bảng 3.19) cho thấy 100% chồi nuôi cấy ở tất cả các nghiệm thức đều tạo rễ bất
định. Số rễ của chồi ở môi trường có NAA, IBA lần lượt là 13,53 và 11,67 so với ĐC
là 4,3; chiều dài rễ tương ứng là 80 mm và 74,56 mm, theo thứ tự (so với ĐC 69,37
mm).
Hình 3.46. Tạo rễ bất định từ chồi có nguồn gốc từ đốt thân, 60 NSC.
A,B,C. Cây 60 NSC ở môi trường ½MS (không có chất ĐHST); có 0,2 mg/L IBA;
có 0,2 mg/L NAA. D,E,F. Rễ hình thành dưới đáy bình tương ứng của A,B,C. G, H.
Cây hình thành ở môi trường ½MS có 0,2 mg/L IBA; 0,2 mg/L NAA. I. Cây từ phôi
vô tính (thanh ngang 20 mm).
Đến nay, đã ghi nhận được một số công bố kết quả nghiên cứu dùng chồi
(nguồn gốc từ thực địa) để tạo rễ bất định cây dược liệu, ví dụ, ngoài vật liệu mảnh
lá, đoạn thân, Martin và cộng sự (2008) còn sử dụng chồi Ophiorrhiza prostrata (30

113
– 50 mm) nuôi cấy trên môi trường đặc MS có 10,74 µM NAA và 2,32 µM kinetin
để tạo rễ, sau đó nuôi nhân thu sinh khối rễ nhằm nghiên cứu hợp chất thứ cấp
camptothecin [131]. Yusuf và cộng sự (2018) cũng đã sử dụng chồi nụ (bud)
Boesenbergia rotunda qua nuôi cấy trên môi trường đặc MS có 2 mg/L NAA để tạo
rễ - như là nguồn vật liệu dùng nghiên cứu pinostrobin [195]. Rễ bất định loài sâm
Java (Javanese ginseng) Talinum paniculatum được cảm ứng hình thành, thu thập từ
chồi nuôi cấy trên môi trường đặc MS có 10 µM IBA, 30 g/L đường nhằm mục đích
nghiên cứu sự tích lũy saponin [196].
3.3.2.2. Tạo rễ bất định từ chồi đốt thân cây in vitro
Sau cắt bỏ các lá, thân cây được cắt thành các đoạn thân mang chồi nách, do
thân cây in vitro có đường kính nhỏ, các chồi nách lại khá gần nhau nên khó cắt chính
xác các đoạn thân chỉ mang 1 chồi nách, do vậy ở ngày thứ 30 và 45 sau cấy nhận
thấy một số mẫu cấy có 2 chồi phát triển. Sau nuôi cấy 60 ngày, các chồi được tách
ra cấy sang môi trường tạo rễ bất định, nhận thấy thân chồi có đường kính nhỏ hơn
thân chồi từ cây vườn ươm, phần thân trong môi trường nuôi cấy phù ra, mô sẹo hình
thành nhưng ít hơn so với mẫu cấy cây vườn ươm, có thể do cây in vitro ở trạng thái
“trẻ” hơn vì đã được nuôi cấy trong thời gian khá dài ở điều kiện in vitro.
Bảng 3.22. Ảnh hưởng của NAA và IBA đến sự tạo rễ bất định từ chồi đốt thân cây
in vitro, ở môi trường ½MS, 60 NSC.
Nghiệm
thức
Tỷ lệ chồi
tạo rễ (%)
Số
rễ/chồi
Chiều
dài rễ
(mm)
Đặc điểm hình thái,
màu sắc rễ
ĐC 100 3,84c* 52,20c Rễ mảnh, dài, trắng, ít lông hút,
rễ nhánh dài
NAA 100 12,65a 64,45a Rễ có đường kính to, dài, trắng,
nhiều lông hút, rễ nhánh ngắn
IBA 100 10,47b 62,20b Rễ có đường kính trung bình, dài,
trắng, ít lông hút, rễ nhánh ngắn
*Các chữ cái khác nhau trong một cột thể hiện sự khác biệt ý nghĩa với p
≤ 0,05 trong phép thử LSD.
Tương tự trường hợp tạo rễ từ chồi có nguồn gốc cây vườn ươm, kết quả tạo
rễ cho thấy trên môi trường không có NAA/IBA (ĐC), rễ hình thành ít, rễ dài, ít lông

114
hút, rễ nhánh dài. Trên môi trường có IBA, chồi tạo nhiều rễ, rễ dài, ít lông hút, rễ
nhánh ngắn (Hình 3.47B); trên môi trường có NAA, chồi tạo nhiều rễ, rễ có đường
kính to, rễ dài, nhiều lông hút, rễ cũng ít phân nhánh (Hình 3.47C). Bảng 3.20 cho
thấy 100% chồi nuôi cấy ở tất cả các nghiệm thức đều tạo rễ bất định. Số rễ của chồi
ở môi trường có NAA, IBA lần lượt là 12,65 và 10,47 so với ĐC là 3,84; chiều dài rễ
tương ứng là 64,45 mm và 62,20 mm, theo thứ tự (so với ĐC 52,20 mm). Nhận thấy
số rễ, chiều dài rễ của chồi ở trường hợp này ít hơn, ngắn hơn so với rễ của chồi từ
cây vườn ươm có thể do đường kính chồi từ cây in vitro nhỏ hơn – ảnh hưởng đến
khả năng tạo rễ bất định.
Hình 3.47. Tạo rễ bất định từ chồi đốt thân cây in vitro, ở môi trường ½MS, 60 NSC
A. Thân cây in vitro cắt bỏ lá (vạch vàng vị trí cắt) B. Vật liệu tạo chồi nách C,D,E.
Chồi nách phát triển ở 30 NSC, 40 NSC, 60 NSC (thanh ngang trắng 3 – 4 mm) F,G.
Cây hình thành trên môi trường ½MS có 0,2 mg/L IBA; 0,2 mg/L NAA ở 60 NSC
(thanh ngang 20 mm).
Cây/chồi in vitro là vật liệu tiền đề rất thuận lợi trong thực hiện thí nghiệm vì
đây là nguồn mẫu sẵn ở trạng thái vô trùng, có thể thu các loại vật liệu khác nhau như
chồi ngọn/chồi nách, lá, thân, rễ dùng trực tiếp trong nhiều nghiên cứu khác nhau về
nhân giống, sự tích lũy hợp chất thứ cấp. Các chồi, có nguồn gốc chồi nách cây in
vitro Ophiorrhiza mungos qua nuôi cấy trên môi trường MS có 0,25 mg/L BA và 025
mg/L kinetin, tạo rễ bất định (trên môi trường ½MS có 100 mg/L than hoạt tính)
thành cây hoàn chỉnh được sử dụng trong nghiên cứu hợp chất thứ cấp kháng ung thư
camptothecin [197].

115
3.3.2.3. Tạo rễ bất định từ chồi có nguồn gốc phôi vô tính
Cây con từ phôi vô tính (Hình 3.48A), sau khi cắt bỏ phần rễ trụ, phần thân
mang các lá được cấy vào môi trường ½MS có chứa 0,1 mg/L NAA/IBA để tạo rễ
bất định. Sau 40 ngày nuôi cấy, trên môi trường không có chất điều hòa sinh trưởng
(NAA/IBA), chồi tạo rễ khá tốt, rễ ít lông hút, có số rễ/chồi là 3,13 với chiều dài rễ
trung bình là 25,67 mm (Hình 3.48B); môi trường nuôi cấy có 0,1 mg/L IBA, rễ dài,
ít lông hút, có rễ nhánh; môi trường có 0,1 mg/L NAA rễ hình thành nhiều, cũng có
hình thành rễ nhánh, nhưng rễ có nhiều lông hút đây là sự khác biệt khác biệt so với
rễ hình thành ở môi trường có IBA, (Hình 3.48C,D). Như vậy, ở môi trường tạo rễ có
bổ sung NAA/IBA hoặc không có chất ĐHST có ảnh hưởng khác biệt lên sự tạo rễ,
NAA hiệu quả hơn IBA.
Bảng 3.23. Ảnh hưởng của NAA và IBA đến sự tạo rễ bất định từ chồi có nguồn gốc
phôi vô tính, môi trường ½MS, 30 NSC.
Nghiệm
thức
Tỷ lệ chồi
tạo rễ (%)
Số
rễ/chồi
Chiều dài
rễ (mm)
Đặc điểm hình thái,
màu sắc rễ
ĐC 100 3,13c* 25,67b Rễ dài, trắng, ít lông hút, không có
rễ nhánh
IBA 100 4,37a 37,67a Rễ dài, trắng, ít lông hút, có rễ
nhánh
NAA 100 6,33b 39,78b Rễ dài, trắng, nhiều lông hút, gốc
rễ to, có rễ nhánh
*Các chữ cái khác nhau trong một cột thể hiện sự khác biệt ý nghĩa với p ≤
0,05 trong phép thử LSD.
Phôi nói chung, phôi vô tính hoặc phôi hạt nói riêng và dạng phát triển tiếp
theo của chúng (cây con) cũng là nguồn vật liệu được các nhà nghiên cứu sử dụng
trong một số trường hợp, ví dụ như trường hợp tạo phôi vô tính từ nuôi cấy mảnh lá
mầm phôi hạt sâm Mỹ Panax quinquefolius trên môi trường MS có 50 g/L đường,
tạo rễ trụ thành cây hoàn chỉnh trên môi trường 1/3SH, có bổ sung 0,25 mg/L NAA,
1% sucrose với mục đích nhân giống [36]. Baskaran và Jayabalan (2009) đã dùng
đoạn mô trụ dưới lá mầm (10 mm) cây từ phôi hạt Psoralea coryfolia (10 ngày tuổi)
nuôi cấy trên môi trường MS có bổ sung 4 µM IAA, NAA để tạo rễ bất định và sau

116
đó rễ bất định được nuôi nhân thu sinh khối trong môi trường ½MS có 3 µM IBA
dùng nghiên cứu hàm lượng psoralen [198].
Hình 3.48. Tạo rễ bất định từ chồi có nguồn gốc phôi vô tính, ở môi trường ½MS,
30 NSC.
A. Vật liệu tạo rễ bất định. Rễ bất định hình thành ở môi trường không có chất điều
hòa sinh trưởng (B); có 0,1 mg/L IBA (C); 0,1 mg/L NAA (D), (thanh ngang 10mm).
Như vậy, ở NGBCC, chồi cũng là vật liệu khá thuận lợi trong tạo rễ bất định
và NAA là tác nhân rất thích hợp trong kích thích tạo rễ từ chồi. Kết quả tác động của
NAA trong nghiên cứu này phù hợp với kết quả đáp ứng tích cực đối với tạo rễ của
mô lá trong điều kiện nuôi cấy có NAA như đã trình bày ở trên. Sự tác động của chất
điều hòa sinh trưởng thực vật lên mô lá, mô thân có sự khác biệt rất rõ nét, với mô lá
hoàn toàn không có biểu hiện cảm ứng với IBA ở các nồng độ khác nhau 0,5; 1; 2; 3;
4; 5 mg/L trên các môi trường nuôi cấy cơ bản là MS; ½MS; B5; SH. Nhưng với các
thí nghiệm sử dụng vật liệu chồi, IBA có tác động gây cảm ứng tạo rễ bất định trên
môi trường có ½MS. Điều đó chứng tỏ rằng, sự tác động của mỗi loại chất điều hòa
sinh trưởng là khác nhau và trên từng loại mô là khác nhau. Đồng thời, vật liệu mô lá
cho kết quả tạo rễ hiệu quả hơn so với vật liệu từ chồi, do vậy, chỉ rễ bất định từ mô
lá mới được sử dụng trong nhân rễ.
Akhami và cộng sự (2013) [101] đã ghi nhận hiện tượng IAA tăng lên ở vị trí
gốc đoạn thân mang chồi Petunia hybrida (trồng trong khay nhựa) ở thời điểm 2 h và
24 h sau cắt đoạn và kết luận sự phát sinh rễ bất định phụ thuộc vào đỉnh IAA ở thời
điểm 24 h và sự di chuyển phân cực của auxin - cảm ứng sự phân chia đầu tiên của
tế bào meristem dẫn đến tạo sơ khởi rễ, IAA giảm xuống sau thời điểm 24 h tạo thuận
lợi cho quá trình phát triển tiếp theo của rễ. Guan và cộng sự (2019) [101] cho rằng
rễ bất định ở đoạn thân mang chồi 2 dòng cà chua (dòng Alisa Craig và dòng chuyển
gen DR5pro:YFP trans, nuôi trong dung dịch dinh dưỡng Hoagland) bắt nguồn từ tế

117
bào vùng trụ bì (pericycle) ở gốc đoạn thân – kết quả tác động của mức auxin tăng
lên trong vòng 1 h sau cắt đoạn. Có sự phân bố auxin gia tăng ở pha khởi tạo rễ, tập
trung ở mô phân sinh đang phát triển. Tương tự, Guan và cộng sự (2020) [13] cho
rằng sự hình thành rễ bất định ở đoạn thân táo lùn Malus domestica bắt đầu bằng sự
phân chia và kéo dài của các tế bào tượng tầng, ở vị trí gần kề hai bó mạch; quá trình
này xảy ra ở điều kiện có mức tương đối cao của IAA trong mô.
Ở tạo rễ bất định từ chồi đốt thân cây vườn ươm/cây in vitro và từ chồi có
nguồn gốc phôi vô tính NGBCC, theo chúng tôi, cũng có cơ chế sinh lý tạo rễ tương
tự như ở kết quả nghiên cứu của các tác giả nêu trên với hai hệ thống vận chuyển
auxin riêng biệt nhưng có liên kết với nhau tương tự như: Hệ thống 1: Vận chuyển
auxin nhanh, không định hướng trong mô li-be cùng với sản phẩm quang hợp, và hệ
thống 2: Vận chuyển phân cực của auxin từ tế bào này sang tế bào khác dẫn đến sự
phân phối auxin không cân xứng, chậm, cần năng lượng, và có định hướng. Hệ thống
vận chuyển auxin trong mô li-be cung cấp auxin từ các vị trí tổng hợp (chủ yếu là ở
lá non) cho các cơ quan nhận; còn sự vận chuyển phân cực phân phối auxin một cách
chính xác cho tế bào, điều này cực kỳ quan trọng đối với sự hình thành ‘cực đại auxin
cục bộ’ (local auxin maxima), xảy ra chủ yếu trong các mô đang phát triển mạnh dẫn
đến kết quả tạo rễ bất định.
3.4. Nhân rễ bất định trong môi trường lỏng
3.4.1. Ảnh hưởng của NAA, IBA đến sự phân nhánh rễ
Ở thí nghiệm này, các rễ sơ cấp đơn (20 ngày tuổi, có nguồn gốc từ nuôi cấy
mảnh lá) sử dụng làm vật liệu nuôi cấy (Hình 3.49)
Hình 3.49. Vật liệu rễ bất định sơ cấp ở 20 NSC
A. Rễ đơn; B. Cụm 2 rễ; C. Cụm 3 rễ; D. Cụm 4 rễ (vạch ngang và dọc chỉ vị trí cắt
thu rễ đơn, thanh ngang 1 cm).

118
Bảng 3.24. Ảnh hưởng của NAA, IBA đến sự phân nhánh rễ, ở môi trường ½MS, 21
NSC.
Nồng độ
(mg/L)
Chiều dài
RSC
(mm)
Số RN
cấp 1
Chiều dài
RN cấp 1
(mm)
Số
RN
cấp 2
Đặc điểm hình thái, màu sắc rễ
NAA 1 18,20 d* 10,36b 3,40c 0,20c RN cấp 1 to, ngắn; đầu tròn, có
lông hút, rễ màu trắng xám.
2 19,72c 12,44a 3,90b 7,20b RN cấp 1 to, ngắn; đầu tròn, có
lông hút, rễ màu trắng xám. Có sự
hình thành RN cấp 2.
3 20,11c 12,95a 4,20a 7,33a RN cấp 1 to, ngắn; đầu tròn, có
lông hút; gốc, thân rễ sơ cấp và
đầu RN bị phù, rễ màu trắng xám.
Có sự hình thành RN cấp 2.
IBA 1 15,33e 2,62e 2,00e 0,10e RN cấp 1 nhỏ, dài; đầu RN thon,
không có lông hút, rễ màu hơi
vàng.
2 21,11b 4,43d 3,00d 3,33d RN cấp 1 nhỏ, dài, không có lông
hút, rễ màu hơi vàng. Có sự hình
thành RN cấp 2
3 22,43a 5,20c 3,00d 3,60c RN cấp 1 nhỏ, dài, không có lông
hút; gốc và thân rễ sơ cấp bị phù,
rễ hơi vàng. Hình thành RN cấp 2
*Các chữ cái khác nhau trong một cột thể hiện sự khác biệt có ý nghĩa ở
mức p ≤ 0,05 trong phép thử Duncan.
Kết quả (Bảng 3.22) cho thấy, nói chung rễ trong môi trường không bổ sung
NAA/IBA (đối chứng) không có khả năng phân nhánh (do vậy không đưa số liệu và
bảng), rễ nhánh trong môi trường có bổ sung NAA hình thành tốt hơn so với rễ trong
môi trường có IBA ở cả ba chỉ tiêu là số rễ nhánh cấp 1, cấp 2 và chiều dài rễ nhánh
cấp 1 ở 20 NSC. Lông hút rễ bị thoái hóa trong môi trường có IBA. Ở môi trường có
3 mg/L NAA, số rễ nhánh cấp 1 là cao nhất - 12,95 so với số rễ trong hai môi trường
có 1 mg/L, 2 mg/L NAA lần lượt là l0,36; 12,44. Đối với hai chỉ tiêu số RN cấp 1 và
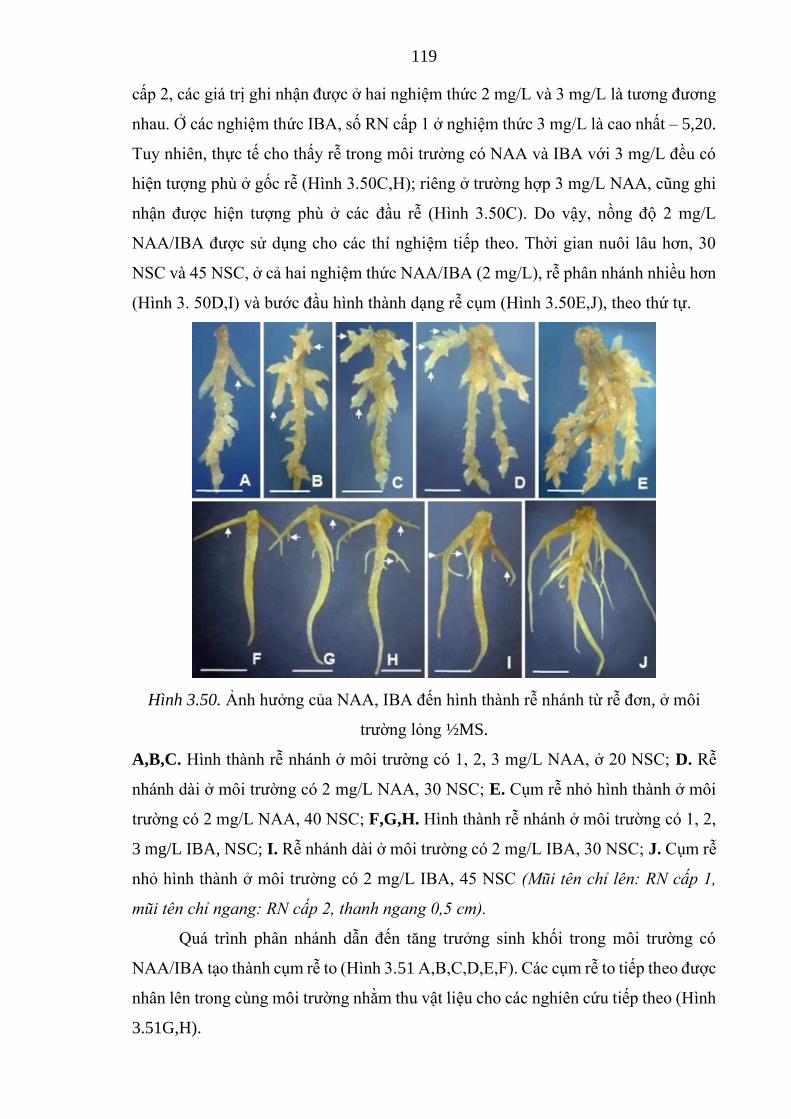
119
cấp 2, các giá trị ghi nhận được ở hai nghiệm thức 2 mg/L và 3 mg/L là tương đương
nhau. Ở các nghiệm thức IBA, số RN cấp 1 ở nghiệm thức 3 mg/L là cao nhất – 5,20.
Tuy nhiên, thực tế cho thấy rễ trong môi trường có NAA và IBA với 3 mg/L đều có
hiện tượng phù ở gốc rễ (Hình 3.50C,H); riêng ở trường hợp 3 mg/L NAA, cũng ghi
nhận được hiện tượng phù ở các đầu rễ (Hình 3.50C). Do vậy, nồng độ 2 mg/L
NAA/IBA được sử dụng cho các thí nghiệm tiếp theo. Thời gian nuôi lâu hơn, 30
NSC và 45 NSC, ở cả hai nghiệm thức NAA/IBA (2 mg/L), rễ phân nhánh nhiều hơn
(Hình 3. 50D,I) và bước đầu hình thành dạng rễ cụm (Hình 3.50E,J), theo thứ tự.
Hình 3.50. Ảnh hưởng của NAA, IBA đến hình thành rễ nhánh từ rễ đơn, ở môi
trường lỏng ½MS.
A,B,C. Hình thành rễ nhánh ở môi trường có 1, 2, 3 mg/L NAA, ở 20 NSC; D. Rễ
nhánh dài ở môi trường có 2 mg/L NAA, 30 NSC; E. Cụm rễ nhỏ hình thành ở môi
trường có 2 mg/L NAA, 40 NSC; F,G,H. Hình thành rễ nhánh ở môi trường có 1, 2,
3 mg/L IBA, NSC; I. Rễ nhánh dài ở môi trường có 2 mg/L IBA, 30 NSC; J. Cụm rễ
nhỏ hình thành ở môi trường có 2 mg/L IBA, 45 NSC (Mũi tên chỉ lên: RN cấp 1,
mũi tên chỉ ngang: RN cấp 2, thanh ngang 0,5 cm).
Quá trình phân nhánh dẫn đến tăng trưởng sinh khối trong môi trường có
NAA/IBA tạo thành cụm rễ to (Hình 3.51 A,B,C,D,E,F). Các cụm rễ to tiếp theo được
nhân lên trong cùng môi trường nhằm thu vật liệu cho các nghiên cứu tiếp theo (Hình
3.51G,H).

120
Hình 3.51. Rễ tăng trưởng sinh khối liên tục qua quá trình phân nhánh của rễ đơn.
A,B. Bình rễ sơ cấp nuôi trong môi trường có 2 mg/L NAA, 2 mg/L IBA, 20 NSC
(thanh ngang 2 cm); C,D. Rễ đơn phân nhánh tạo cụm, sau cấy chuyền, 30 NSC,
(thanh ngang 1 cm); E,F. Cụm rễ to, sau cấy chuyển, 60 NSC (thanh ngang 2 cm);
G,H. Rễ được nhân sinh khối, sau cấy chuyền, 60 NSC (thanh ngang 2 cm).
Trong nuôi cấy in vitro, khả năng nhân rễ bất định phụ thuộc vào việc bổ sung
auxin (loại và nồng độ) vào môi trường nuôi cấy [98] trong đó các auxin như IBA và
NAA được sử dụng phổ biến ở dạng riêng rẽ hoặc phối hợp với nhau; ngoài ra, nhân
rễ còn phụ thuộc vào yếu tố quan trọng khác là họ/ loài thực vật nghiên cứu [198].
Tác động lắc cơ học tạo điều kiện cho mô hấp thu chất ĐHST, chất dinh dưỡng,
nước,… đồng đều (không tạo gradient phân bố), trao đổi khí đều hơn, khác với trường
hợp mô nuôi cấy trên môi trường đặc [142]. Đáp ứng phân nhánh của rễ trong nuôi
cấy mang tính quyết định đối với tăng trưởng sinh khối [184]. Hiện tượng phù ở gốc
rễ hoặc/và đầu rễ trong trường hợp dùng NAA, IBA với nồng độ 3 mg/L ở nghiên
cứu này, theo chúng tôi, sẽ dẫn đến giảm năng suất sinh khối rễ tương tự như kết quả
ghi nhận được ở nuôi lỏng rễ Cichorium intybus [184]. NAA được ghi nhận là thích
hợp trong kích thích phân nhánh rễ đối với khá nhiều loài cây dược liệu như
Andrographis paniculata [199], Eurycoma longifolia [181]. NAA 1 mg/L, 2 mg/L là
các nồng độ tối ưu cho nhân rễ Fagonia indica [200], rễ Boesenbergia rotunda [195],
theo thứ tự. IBA cũng rất thích hợp cho nuôi cấy rễ bất định; nhận thấy 2 mg/L IBA
là nồng độ tối ưu cho phát triển sinh khối rễ khá nhiều loài cây dược liệu như Vernonia
amygdalina [185]. Ở NGBCC, kết quả nghiên cứu ở thí nghiệm này phù hợp với kết
quả công bố của một số tác giả đã sử dụng 2 mg/L NAA/IBA như đã nêu trên để nhân
rễ thông qua hiện tượng tạo rễ nhánh.

121
3.4.2. Ảnh hưởng của nồng độ đường đến sự tăng trưởng sinh khối rễ
Kết quả (Bảng 3.25) cho thấy trong môi trường có NAA, khối lượng tươi rễ
rễ đạt giá trị cao 2,10 g; 2,16 g lần lượt ở hai nghiệm thức có 30 g/L; 40 g/L sucrose;
hệ số nhân tương ứng là 7,00 và 7,20; tuy nhiên các giá trị sinh khối và hệ số nhân
giữa hai nghiệm thức này khác biệt không có ý nghĩa về mặt thống kê. Khối lượng
rễ, hệ số nhân thấp nhất lần lượt là 1,62 g; 5,40 ở nghiệm thức 20 g/L sucrose. Tương
tự ở môi trường có IBA, khối lượng tươi đạt 1,80 g; 1,86 g lần lượt ở hai nghiệm thức
30 g/L, 40 g/L sucrose; hệ số nhân tương ứng là 6,00; 6,21 – cao hơn so với nghiệm
thức 30 g/L sucrose (1,45 g; 4,85). Do vậy, 30 g/L sucrose được sử dụng cho các
nghiên cứu tiếp theo.
Bảng 3.25. Ảnh hưởng của nồng độ đường đến sự tăng trưởng sinh khối rễ, ở môi
trường ½MS, 30 NSC.
Chất
ĐHST
Nồng độ
sucrose
(g/L)
KLT rễ
thu nhận
(g)
Hệ số
nhân sinh
khối (lần)
Đặc điểm hình thái, màu sắc rễ,
cụm rễ
NAA 20 1,62c* 5,40c Rễ nhỏ, có lông hút; cụm rễ màu
trắng xám, mềm
30 2,10a 7,00a Rễ to, có lông hút; cụm rễ màu
trắng xám, mềm
40 2,16a 7,20a Rễ to, có lông hút; cụm rễ màu
trắng xám, mềm
IBA 20 1,45d 4,83d Rễ nhỏ, không có lông hút; cụm
rễ màu hơi vàng đục, cứng
30 1,80b 6,00b Rễ to, không có lông hút; cụm rễ
màu hơi vàng đục, cứng
40 1,86b 6,20b Rễ to, không có lông hút; cụm rễ
màu hơi vàng đục, cứng
*Các chữ cái khác nhau trong một cột thể hiện sự khác biệt có ý nghĩa ở
mức p ≤ 0,05 trong phép thử Duncan.
Có sự khác biệt về hình thái, màu sắc rễ/cụm rễ nuôi trong môi trường có NAA,
IBA. Nói chung, ở cả hai trường hợp môi trường có NAA, IBA với nồng độ sucrose
cao (30 - 40 g/L), sợi rễ to so với rễ ở môi trường có 20 g/L sucrose. Như đã nêu ở

122
trên, rễ ở môi trường bổ sung NAA có nhiều lông hút ở phần đầu rễ; ngược lại, lông
hút rễ khó quan sát ở môi trường có IBA; cụm rễ ở môi trường bổ sung NAA có màu
trắng xám, cụm rễ ở môi trường chứa IBA có màu hơi vàng đục (Hình 3.52).
Hình 3.52. Ảnh hưởng của nồng độ đường đến tăng trưởng sinh khối rễ ở môi
trường ½MS, 30 NSC.
A,B,C. Cụm rễ nuôi ở môi trường ½MS bổ sung 2 mg/L NAA, có sucrose 20, 30, 40
g/L; D,E,F. Cụm rễ nuôi ở môi trường bổ sung 2 mg/L IBA, có sucrose 20, 30, 40
g/L. Thanh ngang 2 cm.
Ngoài môi trường khoáng và chất ĐHST, sự tăng trưởng của rễ bất định còn
chịu ảnh hưởng của đường - nguồn carbon quan trọng trong trao đổi chất nhằm tăng
sinh khối thông qua sự hình thành rễ nhánh và nồng độ sử dụng thích hợp tùy thuộc
vào loài thực vật. Theo nhiều tài liệu, nồng độ đường sử dụng dao động từ 20 - 50
g/L, trường hợp cao hơn thường tạo áp suất thẩm thấu bất lợi cho tăng trưởng rễ [100].
Nồng độ đường 20 g/L là tối ưu đối với nuôi rễ Gynura procumbens [182], 30 g/L
thích hợp cho nuôi rễ khá nhiều loài thực vật như Hypericum perporatum [100]. Nồng
độ đường sucrose khá cao (50 g/L) lại thích hợp cho nuôi lỏng rễ bất định Eurycoma
longifolia [181], rễ Boesenbergia rotunda [195]. Rễ NGBCC tăng trưởng tốt trong
môi trường có 30 g/L sucrose - phù hợp với kết quả công bố của một số tác giả trên.
Ngoài ra, nồng độ đường khác nhau còn ảnh hưởng đến sinh tổng hợp hợp chất thứ
cấp đối với cây dược liệu [98], do vậy kết quả này cũng tạo tiền đề cho nghiên cứu
tiếp theo về hợp chất thứ cấp ở rễ NGBCC.

123
3.4.3. Ảnh hưởng của khối lượng rễ nuôi cấy đến tăng trưởng sinh khối của rễ
Kết quả (Bảng 3.26, Hình 3.53) cho thấy trong môi trường có 2 mg/L NAA, ở
nghiệm thức cấy 2% (w/v) rễ đạt hệ số nhân sinh khối cao nhất (9,96) so với các
nghiệm thức cấy 0,5%, 1% và 3% (hệ số nhân tương ứng là 6,74; 8,13 và 9,15). Ở
nghiệm thức 3% tuy thu được khối lượng tươi rễ cao nhất (19,22 g) nhưng hệ số nhân
thấp hơn so với nghiệm thức cấy 2%. Tương tự, trong môi trường có 2 mg/L IBA,
kết quả cũng cho thấy hệ số nhân sinh khối rễ đạt cao nhất ở trường hợp cấy 2% (khối
lượng tươi rễ và hệ số nhân sinh khối lần lượt là 12,85 g và 9,18) so với các trường
hợp còn lại.
Bảng 3.26. Ảnh hưởng của khối lượng rễ nuôi cấy đến tăng trưởng sinh khối rễ, ở
môi trường ½MS, 45 NSC.
Chất
ĐHST
KLR nuôi
cấy ban đầu
(g)
KLT rễ
thu nhận
(g)
Hệ số
nhân sinh
khối (lần)
Đặc điểm hình thái, màu sắc rễ,
cụm rễ
NAA 0,35 (0,5%) 2,36g* 6,74g Rễ dài, có lông hút; cụm rễ màu
trắng xám, mềm
0,70 (1%) 5,69e 8,13e Rễ dài, có lông hút; cụm rễ màu
trắng xám, mềm
1,40 (2%) 13,95c 9,96a Rễ dài, có lông hút; cụm rễ màu
trắng xám, mềm
2,10 (3%) 19,22a 9,15c Rễ dài, có lông hút, cụm rễ màu
trắng xám, mềm
IBA 0,35 (0,5%) 2,22g 6,34g Rễ ngắn, không có lông hút;
cụm rễ màu hơi vàng đục, cứng
0,70 (1%) 5,16f 7,37f Rễ ngắn, không có lông hút;
cụm rễ màu hơi vàng đục, cứng
1,40 (2%) 12,85d 9,18b Rễ ngắn, không có lông hút;
cụm rễ màu hơi vàng đục, cứng
2,10 (3%) 18,03b 8,58d Rễ ngắn, không có lông hút;
cụm rễ màu hơi vàng đục, cứng
*Các chữ cái khác nhau trong một cột thể hiện sự khác biệt có ý nghĩa ở
mức p ≤ 0,05 trong phép thử Duncan.

124
Hình 3.53. Ảnh hưởng của khối lượng rễ nuôi cấy đến tăng trưởng sinh khối rễ, ở
môi trường ½MS, 45 NSC.
A,B,C,D. Khối lượng rễ nuôi cấy ban đầu 0,5%, 1%, 2%, 3% trong môi trường có 2
mg/L NAA; E,F,G,H. Khối lượng rễ nuôi cấy ban đầu 0,5%, 1%, 2%, 3%, trong môi
trường có 2 mg/L IBA. Thanh ngang 2 cm.
Khối lượng rễ ban đầu dùng nuôi cấy (inoculum density/size) là yếu tố quan
trọng ảnh hưởng đến chỉ số tăng trưởng (growth index) của vật liệu nuôi cấy, chỉ số
tăng trưởng giảm ở trường hợp dùng khối lượng rễ cao không phù hợp [201]. Ngoài
ra, khối lượng rễ cấy còn ảnh hưởng đến sự tích lũy hợp chất thứ cấp trong điều kiện
nuôi cấy nhất định [202]. Đối với nuôi lỏng lắc, khối lượng rễ sử dụng thường dao
động từ 0,1% đến 5% (w/v) tùy thuộc vào loài thực vật nghiên cứu. Cụ thể khối lượng
rễ 0,1% là thích hợp để nuôi rễ Mondia whitei [198], 1% cho nuôi rễ cây dược liệu
Echinacea angustifolia [202]; 1,6% đối với rễ sâm Panax ginseng [203], khối lượng
rễ 2% là tốt nhất đối với nuôi rễ Panax ginseng, Gynura procumbens, theo thứ tự
[182]. Nhưng ở trường hợp Curcuma amada, rễ đạt tỷ lệ tăng trưởng sinh khối cao
nhất khi được nuôi cấy với khối lượng rễ nuôi cấy cao là 5% [93]. Kết quả khối lượng
rễ 2% là thích hợp đối với nhân rễ NGBCC và cũng phù hợp với kết luận về thông số
nuôi cấy này của một số tác giả đã nêu ở trên.
3.4.4. Khảo sát diễn biến tăng trưởng sinh khối rễ theo thời gian
Kết quả (Biểu đồ 3.1, Hình 3.54) cho thấy rễ nuôi trong môi trường NAA tăng
sinh khối chậm ở giai đoạn 7 NSC (sinh khối đạt 2,62 g, hệ số nhân 1,87), tăng trưởng
rất nhanh ở ba giai đoạn 21, 28 và 35 NSC, khối lượng tươi rễ đạt lần lượt 7,90 g;
13,92 g và 17,42 g; hệ số nhân tương ứng là 5,64; 9,94 và 12,44. Ở các thời điểm 42
và 49 NSC, rễ đạt giá trị khối lượng tối đa lần lượt là 18,26 g và 18,47 g, tuy nhiên
khối lượng tươi rễ ở thời điểm 49 NSC khác biệt không có ý nghĩa về mặt thống kê

125
so với khối lượng tươi rễ thu nhận ở 42 NSC (do đã ở thời kỳ ổn định); ngoài ra, nhận
thấy ở giai đoạn 49 NSC, một số rễ có hiện tượng hóa nâu (Hình 3.55) có thể do hàm
lượng dinh dưỡng môi trường đã giảm nhiều. Tương tự, rễ nuôi trong môi trường IBA
cũng có xu hướng tăng sinh khối rất nhanh ở 28 NSC và đạt mức tối đa ở 42 NSC
(Hình 3.54G,H) (khối lượng tươi rễ lần lượt là 12,83 g và 16,91 g; hệ số nhân tương
ứng là 9,16 và 12,08), đến giai đoạn 49 NSC nhận thấy khối lượng rễ tươi thu nhận
được không có sự khác biệt về mặt thống kê so với thời điểm 42 NSC. Rễ nuôi trong
môi trường có NAA phát triển nhanh hơn về khối lượng ở từng thời điểm và từng
nồng độ so với rễ trong môi trường có IBA (Hình 3.54G,H).
Biểu đồ 3.1. Diễn biến tăng trưởng sinh khối rễ bất định theo thời gian, 7 – 49 NSC.
Nhằm thu sinh khối ở mức tối ưu dùng cho các mục đích khác nhau, đối với
bất kỳ hệ thống nuôi cấy (lỏng lắc, bioreactor) và vật liệu nuôi cấy nào (tế bào, phôi,
rễ,..), khảo sát động học tăng trưởng theo thời gian của vật liệu nuôi cấy là yêu cầu
quan trọng. Trong điều kiện nuôi lỏng lắc, nhiều tài liệu cho thấy giai đoạn rễ đạt giá
trị khối lượng tối đa ở một thời điểm nuôi cấy nhất định tùy thuộc vào loài thực vật,
cụ thể Ahmad và cộng sự (2020) ghi nhận 3 pha tăng trưởng của rễ bao gồm pha mở
đầu (7 – 9 ngày), pha tăng trưởng rất nhanh (12 - 27 ngày) và đạt mức ổn định từ 27
2.62k
5.13i
7.90g
13.92e
17.42b
18.26a 18.47a
2.18l
4.52j
7.15h
12.83f
16.12d
16.91c 17.07bc
1.87k
3.66i
5.64g
9.94e
12.44b13.04a 13.19a
1.56l
3.23j
5.11h
9.16f
11.50d12.08c 12.19bc
0.00
4.00
8.00
12.00
16.00
20.00
0.00
2.00
4.00
6.00
8.00
10.00
12.00
14.00
16.00
18.00
20.00
7 14 21 28 35 42 49
Hệ
số n
hân
sin
h k
hố
i (l
ần)
Khố
i lư
ợng r
ễ th
u n
hận
(g)
Thời gian (ngày)
Khối lượng rễ thu nhận (NAA) Khối lượng rễ thu nhận (IBA)
Hệ số nhân sinh khối (NAA) Hệ số nhân sinh khối (IBA)
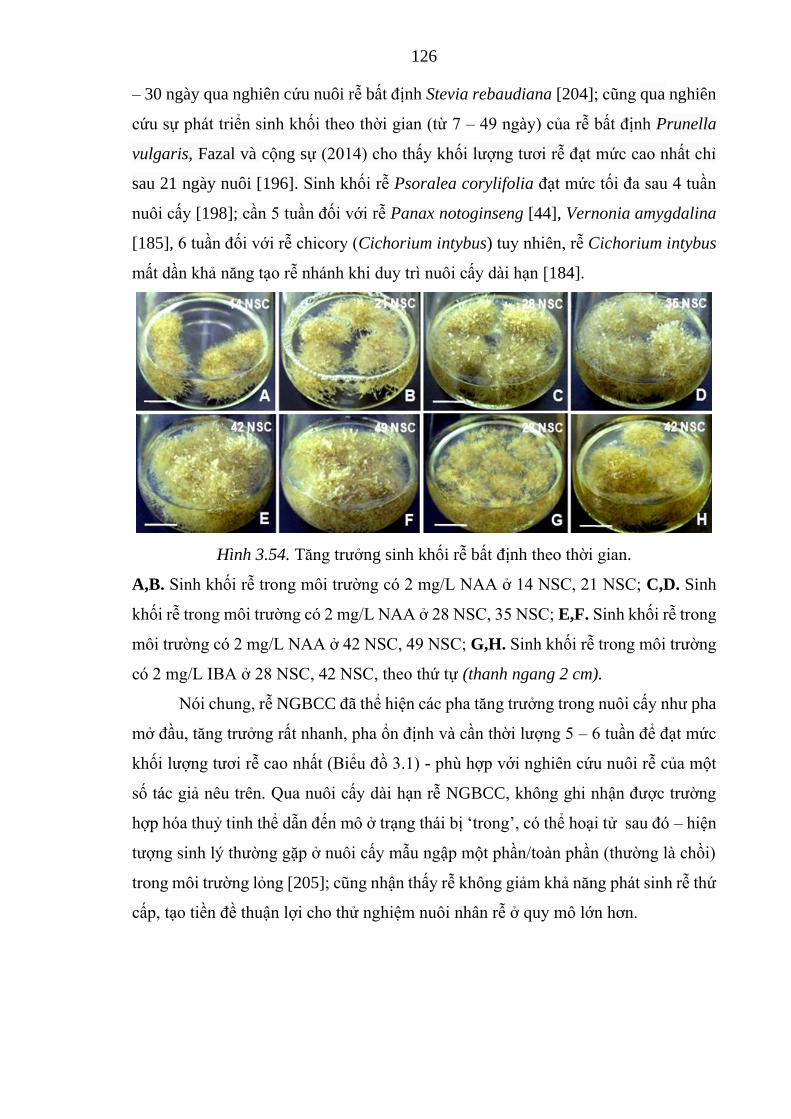
126
– 30 ngày qua nghiên cứu nuôi rễ bất định Stevia rebaudiana [204]; cũng qua nghiên
cứu sự phát triển sinh khối theo thời gian (từ 7 – 49 ngày) của rễ bất định Prunella
vulgaris, Fazal và cộng sự (2014) cho thấy khối lượng tươi rễ đạt mức cao nhất chỉ
sau 21 ngày nuôi [196]. Sinh khối rễ Psoralea corylifolia đạt mức tối đa sau 4 tuần
nuôi cấy [198]; cần 5 tuần đối với rễ Panax notoginseng [44], Vernonia amygdalina
[185], 6 tuần đối với rễ chicory (Cichorium intybus) tuy nhiên, rễ Cichorium intybus
mất dần khả năng tạo rễ nhánh khi duy trì nuôi cấy dài hạn [184].
Hình 3.54. Tăng trưởng sinh khối rễ bất định theo thời gian.
A,B. Sinh khối rễ trong môi trường có 2 mg/L NAA ở 14 NSC, 21 NSC; C,D. Sinh
khối rễ trong môi trường có 2 mg/L NAA ở 28 NSC, 35 NSC; E,F. Sinh khối rễ trong
môi trường có 2 mg/L NAA ở 42 NSC, 49 NSC; G,H. Sinh khối rễ trong môi trường
có 2 mg/L IBA ở 28 NSC, 42 NSC, theo thứ tự (thanh ngang 2 cm).
Nói chung, rễ NGBCC đã thể hiện các pha tăng trưởng trong nuôi cấy như pha
mở đầu, tăng trưởng rất nhanh, pha ổn định và cần thời lượng 5 – 6 tuần để đạt mức
khối lượng tươi rễ cao nhất (Biểu đồ 3.1) - phù hợp với nghiên cứu nuôi rễ của một
số tác giả nêu trên. Qua nuôi cấy dài hạn rễ NGBCC, không ghi nhận được trường
hợp hóa thuỷ tinh thể dẫn đến mô ở trạng thái bị ‘trong’, có thể hoại tử sau đó – hiện
tượng sinh lý thường gặp ở nuôi cấy mẫu ngập một phần/toàn phần (thường là chồi)
trong môi trường lỏng [205]; cũng nhận thấy rễ không giảm khả năng phát sinh rễ thứ
cấp, tạo tiền đề thuận lợi cho thử nghiệm nuôi nhân rễ ở quy mô lớn hơn.

127
KẾT LUẬN VÀ KIẾN NGHỊ
KẾT LUẬN
Trên cơ sở ứng dụng phương pháp nuôi cấy mô tế bào thực vật, lần đầu
tiên trên đối tượng NGBCC, đã xác định được:
Sự phát sinh phôi vô tính trực tiếp từ mô lá hiệu quả nhất ở môi trường đặc SH
có bổ sung 5 mg/L NAA, 0,25 mg/L BA, 50 g/L sucrose, 10% nước dừa; điều kiện
chiếu sáng 4.000 lux. Phát sinh phôi gián tiếp qua mô sẹo mảnh lá ở môi trường đặc
SH có 2 mg/L NAA, 0,25 mg/L BA, 30 g/L sucrose.
Môi trường nuôi cấy lỏng SH có bổ sung 2 mg/L NAA, 0,25 mg/L BA, 50 g/L
sucrose, 10% nước dừa ở điều kiện chiếu sáng 4.000 lux là tối ưu cho sự phát sinh
phôi thứ cấp, với khối lượng phôi nuôi cấy 2% và kích thước phôi nuôi cấy ~ 10 mm
là hiệu quả nhất. Phôi thứ cấp hình thành trong môi trường lỏng lắc hiệu quả hơn so
với môi trường đặc.
Sự tăng trưởng của cây con từ phôi vô tính hiệu quả nhất ở môi trường ½MS
(20 g/L sucrose), cây con sinh trưởng bình thường ở điều kiện ex vitro.
Rễ bất định phát sinh trực tiếp từ mô lá hiệu quả cao nhất trên môi trường đặc
½MS có bổ sung 3 mg/L NAA, 30 g/L sucrose, điều kiện chiếu sáng 4.000 lux.
Thu nhận sinh khối rễ bất định thông qua sự tạo rễ thứ cấp bằng phương pháp
nuôi lỏng lắc trong môi trường ½MS có bổ sung 2 mg/L NAA, 30 g/L sucrose; khối
lượng rễ nuôi cấy ban đầu thích hợp là 2% đạt hiệu quả cao nhất.
Rễ bất định phát sinh từ chồi đốt thân (cây vườn ươm/in vitro) nuôi cấy trên
môi trường thích hợp ½MS có 0,2 mg/L NAA có 20 g/L sucrose. Chồi có nguồn gốc
phôi vô tính tạo rễ hiệu quả qua nuôi cấy trên môi trường đặc ½MS có 0,1 mg/L NAA
(20 g/L sucrose).
Khảo sát hình thái giải phẫu phôi vô tính (ở các giai đoạn phát triển khác nhau)
và rễ bất định bằng phương pháp nhuộm kép đã được thực hiện có kết quả.
Các kết quả nghiên cứu trên góp phần phát triển hướng nghiên cứu về cơ sở
sinh lý của tính toàn thế của tế bào thực vật nói chung, ở đối tượng Schefflera
octophylla nói riêng.

128
KIẾN NGHỊ
Nghiên cứu nuôi nhân phôi vô tính, rễ bất định ở quy mô lớn bằng hệ thống
bioreactor.
Sử dụng sản phẩm sinh khối phôi vô tính (từ kết quả nhân phôi theo cơ chế tạo
phôi thứ cấp), rễ bất định trong nghiên cứu nhân giống, thu nhận các hợp chất thứ cấp
phục vụ lĩnh vực y dược/mỹ phẩm và trong một số nghiên cứu quan trọng khác như
bảo quản nguồn gen, biến nạp gen,...

129
DANH MỤC CÔNG TRÌNH
1. Huỳnh Thị Lũy, Nguyễn Hữu Hổ, Bùi Văn Lệ, Tạo phôi vô tính trực tiếp
từ nuôi cấy in vitro mô lá cây Ngũ gia bì chân chim Schefflera octophylla (Lour.)
Harms, Kỷ yếu Hội nghị Công nghệ Sinh học toàn quốc, TP. HCM, 2019, 405-410.
2. Huỳnh Thị Lũy, Nguyễn Hữu Hổ, Bùi Văn Lệ, Tạo rễ bất định trực tiếp từ
mô lá cây Ngũ gia bì chân chim Schefflera octophylla (Lour.) Harms nuôi cấy in
vitro, Tạp chí Công nghệ Sinh học, 2021, 19(3), 495-507.
3. Huỳnh Thị Lũy, Nguyễn Hữu Hổ, Bùi Văn Lệ, Nhân rễ bất định cây Ngũ
gia bì chân chim Schefflera octophylla (Lour.) Harms bằng phương pháp nuôi lỏng
lắc, Tạp chí Nông nghiệp và Phát triển Nông thôn, 2021, 6(3), 12-20.

130
TÀI LIỆU THAM KHẢO
1. Lã Đình Mỡi, Châu Văn Minh, Trần Văn Sung, Phạm Quốc Long, Phan Văn
Kiệm, Trần Huy Thái, Trần Minh Hợi, Ninh Khắc Bản, Lê Mai Hương, Họ
Nhân sâm (Araliaceae Juss.) - Nguồn hoạt chất sinh học đa dạng và đầy triển
vọng ở Việt Nam, Tuyển tập Hội nghị khoa học toàn quốc lần thứ 5 về Sinh thái
và Tài nguyên Sinh vật, Viện Hàn Lâm Khoa học và Công nghệ Việt Nam,
2013, 1152-1158.
2. J. Kitajima, M. Shindo, Y. Tanaka, Two new triterpenoid sulfates from the
leaves of Schefflera octophylla, Chemical Pharmaceutical Bulletin, 1990, 38(3),
714-716.
3. Y.L. Li, D.X. Shi, M.L. Wang, Tissue culture and rapid propagation of
Schefflera octophylla, Plant Physiology Communications, 2004, 40(2), 193.
4. Y.L. Li, P.P. But, V.E.C. Ooi, Antiviral activity and mode of action of
caffeoylquinic acids from Schefflera heptaphylla (L.) Frodin, Antiviral
Research, 2005, 68(1), 1-9.
5. Y.L. Li, R.W. Jiang, L.M.S. Ooi, P.P.H. But , V.E.C. Ooi, Antiviral
triterpenoids from the medicinal plant Schefflera heptaphylla. Phytother
Research, 2007, 21(5), 466-470.
6. Y.L. Li, C.M. Yeung, L.C.M. Chiu, Y.Z. Cen, V.E.C. Ooi, Chemical
composition and antiproliferative activity of essential oil from the leaves of a
medicinal herb, Schefflera heptaphylla, Phytother Research, 2009, 23(1), 140-
142.
7. Y.F. Chen, S.H. Tao, F.L. Zeng, L.W. Xie, Z.B. Shen, Antinociceptive and anti-
inflammatory activities of Schefflera octophylla extracts, Journal
Ethnopharmacology, 2015, 171, 42-50.
8. X. Liu, Y. Niu, J. Liu, R. Xu, M. Shi, W. Kang, Efficient extraction of anti-
inflammatory active ingredients from Schefflera octophylla leaves using ionic
liquid-based ultrasonic-assisted extraction coupled with HPLC, Molecules,
2019a, 24(16), 2942.
9. X. Liu, J. Dong, Q. Liang, H.M.D. Wang, X.H. Liu, Coagulant effects and
mechanism of Schefflera heptaphylla (L.) Frodin, Molecules, 2019b, 24(24),
4547.

131
10. Đỗ Huy Bích, Đặng Quang Chung, Bùi Xuân Chương, Nguyễn Thượng Dong,
Đỗ Trung Đàm, Phạm Văn Hiền, Vũ Ngọc Lộ, Phạm Duy Mai, Phạm Kim Mẫn,
Đoàn Thị Thu, Nguyễn Tập, Trần Toàn, Cây thuốc và động vật làm thuốc ở Việt
Nam, Nhà xuất bản Khoa học và Kỹ thuật, Viện Dược liệu, 2006, 2, 411-414.
11. Đặng Thị Thanh Tâm, Nghiên cứu tạo rễ tơ của cây Tam thất, Ngũ gia bì chân
chim và thử nghiệm quá trình sản xuất sinh khối bằng bioreactor, Báo cáo tổng
hợp đề tài KC.04.TN10/11-15 thuộc Chương trình KH&CN trọng điểm cấp nhà
nước (KC.04/11-15), Trường Đại học Nông nghiệp Hà Nội, 2012.
12. W. Gaoyin, W. Xiaoli, W. Xiao, L. Xian, W. Yi, Changes in biochemistry and
histochemical characteristics during somatic embryogenesis in Ormosia henryi
Prain, BioRxiv, 2020; https://doi.org/10.1101/2020.09.21.307009.
13. L. Guan, Y. Li, K. Huang, Z.M. Cheng, Auxin regulalation and MdPIN
expression during adventitious root initiation in apple cuttings, Hoticulture
Reseach, 2020, 7: 143.
14. S.K.Verma, A.K.Das, S.Gantait, S. Gurel, E. Gurel, Influence of auxxin and its
polar transport inhibitor on the development of somatic embryos in Digitalis
trojana, 2018, 8 (2), 99.
15. Y.H. Su, L.P. Tang, X.Y. Zhao, X.S. Zhang, Plant cell totipotency: Insights into
cellular reprogramming, Plant Biology Journal of Integrative, 2021, 63, 228–
240.
16. N. Vidal, C. Sanchez, Use of bioreactor systems in the propagation of forest
trees, Engineering in Life Sciences, 2019, 19 (12), 896-915.
17. B. Qiang, J. Miao, N. Phillips, K. Wei, Y. Gao, Recent advances in the tissue
culture of American ginseng (Panax quinquefolius), Chemistry and
Biodiversity, 2020, 17(10), e2000366.
18. F. Engelmann, In vitro conservation of tropical plant germplasm - a review,
Euphytica, 1991, 57, 227-243.
19. Y. Guan, S.G. Li, X.F. Fan, Z.H. Su, Application of somatic embryogenesis in
woody plants, Front Plant Science, 2016, 7, 938.
20. V. Pikulthong, T. Teerakathiti, A. Thamchaipenet, S. Peyachoknagul,
Development of somatic embryos for genetic transformation in Curcuma longa

132
L. and Curcuma mangga Valeton & Zijp. Agriculture and Natural Resources,
2016, 50, 276-285.
21. F. Afreen, S.M.A. Zobayed, T. Kozai, Photoautotrophic culture of Coffea
arabusta somatic embryos: Development of a bioreactor for large-scale plantlet
conversion from cotyledonary embryos. Annals of Botany, 2002, 90, 21-29.
22. H. Etienne, D. Breton, J.C. Breitler, B. Bertrand, E. Déchamp, R. Awada, P.
Marraccini, S. Léran, E. Alpizar, C. Campa, P. Courte, F. Georget, J.P. Ducos,
Coffee somatic embryogenesis: How did research, experience gained and
innovations promote the commercial propagation of elite clones from the two
cultivated species? Frontiers in Plant Science, 2018, 9, 01630.
23. Nguyễn Trung Thành và Paek Kee Yoeup, Nhân nhanh rễ bất định Nhân sâm
Panax ginseng C. A. Meyer: Ảnh hưởng của một số nhân tố lý hóa lên sự tăng
trưởng sinh khối và sản phẩm trao đổi chất ginsenosides, Tạp chí Khoa học
ĐHQGHN, Khoa học Tự nhiên và Công nghệ, 2008, 24, 318-323.
24. Dương Tấn Nhựt, Trần Hiếu, Nguyễn Thị Nhật Linh, Hoàng Thanh Tùng,
Nguyễn Bá Nam, Nguyễn Phúc Huy, Vũ Quốc Luận, Vũ Thị Hiền. Tối ưu hóa
quá trình nhân nhanh và tích lũy saponin của rễ bất định sâm Ngọc Linh (Panax
vietnamensis Ha et Grushv.) trong các hệ thống nuôi cấy, Tạp chí Công nghệ
Sinh học, 2015, 13(3), 853-864.
25. Y. Wang, S. Jia, Z. Wang, Y. Chen, S. Mo, N.N. Sze, Planning considerations
of green corridors for the improvement of biodiversity resilience in suburban
areas, Journal of Infrastructure Preservation and Resilience, 2021, 2:6.
26. H.G.A. Engler and K.A.E. Prantl, Die Natürlichen Pflanzenfamilien, 1894,
3(8): 38, Angelmann Publisher.
27. Tự điển Bách khoa Dược học, Nhà xuất bản Từ điển Bách khoa Hà Nội, 1999,
433-434.
28. Trần Văn Sung, G. Adam, A sulphated triterpenoid saponin from Schefflera
octophylla. Phytochemistry, 1991, 30 (8), 2717-2720.
29. Trần Văn Sung, J. Peter, Katalinic, G. Adam, A bisdesmosidic triterpenoid
saponin from Schefflera octophylla, Phytochemistry, 1991a, 30 (11), 3717-
3720.
30. Trần Văn Sung, Steglich, G. Adam, Triterpenoid glycosides from Schefflera

133
octophylla, Phytochemistry, 1991b, 30 (7), 2349-2356.
31. Trần Văn Sung, C. Lavaud, A. Porzel, W. Steglich, G. Adam, Triterpenoids and
their glycosides from the bark of Schefflera octophylla, Phytochemistry, 1992,
31(1), 227-231.
32. C. Wu, Y.H. Duan, M.M. Li, W. Tang, X. Wu, G.C. Wang, Triterpenoid
saponins from the stem barks of Schefflera heptaphylla, Planta Medica, 2013,
79(14), 1348-1355.
33. C. Wu, Y.H. Duan, W. Tang, M.M. Li, X. Wu, G.C. Wang, W.C. Ye, G.X.
Zhou, New ursane-type triterpenoid saponins from the stem bark of Schefflera
heptaphylla, Fitoterapia, 2014, 92, 127-132.
34. S. Arya, I.D. Arya, T. Eriksson, Rapid multiplication of adventitious somatic
embryos of Panax ginseng, Plant Cell Tissue and Organ Culture, 1993, 34, 157-
162.
35. Y.E. Choi, J.H. Jeong, C.K. Shin, Hormone-independent embryogenic callus
production from ginseng cotyledons using high concentrations of NH4NO3 and
progress towards bioreactor production. Plant Cell Tissue and Organ Culture,
2003, 72(3), 229-235.
36. S. Zhou and D.C. Brown, High efficiency plant production of North American
ginseng via somatic embryogenesis from cotyledon explants, Plant Cell Reports,
2006, 25(3), 166-173.
37. Y.L. Kim, K. Kim, O.R. Lee, D.C. Yang, High frequency of plant regeneration
through cyclic secondary somatic embryogenesis in Panax ginseng. Journal
Ginseng Research, 2012, 36(4), 442-448.
38. Vũ Thị Hiền, Vũ Quốc Luận, Nguyễn Phúc Huy, Nguyễn Bá Nam, Bùi Văn
Thế Vinh, Thái Xuân Du, Dương Tấn Nhựt, Phát sinh phôi trực tiếp từ lá, cuống
lá và thân rễ cây sâm Ngọc Linh (Panax vietnamensis Ha et Grushv.), Tạp chí
Sinh học, 2014, 36(1), 277-282.
39. Vũ Thị Hiền, Vũ Quốc Luận, Nguyễn Phúc Huy, Nguyễn Bá Nam, Nguyễn Thị
Kim Loan, Nguyễn Thanh Sang, Vũ Thị Thủy, Nguyễn Hồng Hoàng, Thái Xuân
Du, Dương Tấn Nhựt, Sử dụng kỹ thuật nuôi cấy lớp mỏng tế bào trong nghiên
cứu quá trìmh phát sinh hình thái của cây sâm Ngọc Linh (Panax vietnamensis
Ha et Grushv.) in vitro, Tạp chí Khoa học và Phát triển, 2015, 13(4), 657-664.

134
40. Mai Trường, Trần Thị Ngọc Hà, Phan Tường Lộc, Lê Tấn Đức, Trần Trọng
Tuấn, Đỗ Đăng Giáp, Bùi Đình Thạch, Nguyễn Thị Thanh, Phạm Đức Trí,
Nguyễn Đức Minh Hùng, Nguyễn Văn Kết, Trần Công Luận, Nguyễn Hữu Hổ,
Nghiên cứu tạo và nhân phôi soma sâm Ngọc Linh (Panax vietnamensis Ha et
Grushv.) trong môi trường lỏng, Tạp chí Khoa học và Phát triển, 2014, 12(7),
1085-1095.
41. J.Y. Zhang, H.J. Sun, I.J. Song, T.W. Bae, H.G. Kang, S.M. Ko, Y.I. Kwon,
I.W. Kim, J. Lee, S.Y. Park, P.O. Lim, Y.H. Kim, H.Y. Lee, Plant regeneration
of Korean wild ginseng (Panax ginseng Meyer) mutant lines induced by γ-
irradiation (60Co) of adventitious roots. Journal Ginseng Research. 2014, 38(3),
220-225.
42. Đỗ Mạnh Cường, Hoàng Thanh Tùng, Hoàng Đắc Khải, Vũ Quốc Luận,Vũ Thị
Hiền, Trương Thị Bích Phượng, Dương Tấn Nhựt, Increasing the somatic
embryogenesis frequency of Panax vietnamensis Ha et Grushv. by disinfection
of leaf explant using nano silver and the addition of nano silver in culture
medium, Vietnam Journal of Biotechnology, 2020, 18, 517-527.
43. Nguyễn Thị Ngọc Hương, Trần Hùng, Trương Thị Đẹp, Biến đổi hình thái trong
phát sinh phôi soma thông qua nuôi cấy mô sẹo Tam thất hoang (Panax
stipuleanatus H.T.Tsai et K.M.Feng), Tạp chí Công nghệ Sinh học, 2018, 16(2),
279-284.
44. X. Gao, C. Zhu, W. Jia, W. Gao, M. Qiu, Y. Zhang, P. Xiao, Induction and
characterization of adventitious roots directly from the explants of Panax
notoginseng, Biotechnology Letters, 2005, 27(22), 1771-1775.
45. Mai Trường, Trần Thị Ngọc Hà, Phan Tường Lộc, Lê Tấn Đức, Trần Trọng
Tuấn, Đỗ Đăng Giáp, Bùi Đình Thạch, Nguyễn Thị Thanh, Phạm Đức Trí,
Nguyễn Đức Minh Hùng, Nguyễn Văn Kết, Trần Công Luận, Nguyễn Hữu Hổ,
Một số kết quả nghiên cứu tạo và nuôi rễ bất định sâm Ngọc Linh (Panax
vietnamensis Ha et Grushv.), Tạp chí Công nghệ Sinh học, 2014, 12(2), 355-
364.
46. Nguyễn Thị Ngọc Hương, Trần Hùng, Trương Thị Đẹp, Tìm hiểu các biến đổi
hình thái trong sự phát sinh rễ Tam thất hoang (Panax stipuleanatus H.T.Tsai
et K.M.Feng) nuôi cấy in vitro và bước đầu định tính oleanolic acid trong rễ

135
tạo thành, Tạp chí Công nghệ Sinh học, 2016, 14(1), 49-54.
47. Nguyễn Thị Nhật Linh, Nghiên cứu nuôi cấy rễ thứ cấp sâm Ngọc Linh (Panax
vietnamensis Ha et Grushv.) và khảo sát ảnh hưởng của một số elicitor lên sự
tích lũy saponin, Luận án TS chuyên ngành Sinh lý học Thực vật (Mã số:
62.42.01.12), Trường ĐH Khoa học Huế - ĐH Huế, 2017.
48. Z. Gui, Z. Guo, S. Ke, R.M. Skirvin, Somatic embryogenesis and plant
regeneration in Acanthopanax senticosus, Plant Cell Reports, 1991, 9(9), 514-
516.
49. Y.E. Choi, J.W. Kim, W.Y. Soh, Somatic embryogenesis and plant
regeneration from suspension cultures of Acanthopanax koreanum Nakai, Plant
Cell Reports, 1997, 17(2), 84-88.
50. S.Y. Park, J.K. Ahn, W.Y. Lee, H.N. Murthy, K.Y. Paek, Mass production
of Eleutherococcus koreanum plantlets via somatic embryogenesis from root
cultures and accumulation of eleutherosides in regenerants, Plant Science,
2005, 168(5), 1221-1225.
51. A.M. Shohael, H.N. Murthy, E.J. Hahn, K.Y. Paek, Methyl jasmonate induced
overproduction of eleutherosides in somatic embryos of Eleutherococcus
senticosus cultured in bioreactors. Electronic Journal of Biotechnology, 2007,
10(4), 633-637.
52. J.M. Seo, C.G. Shin, Y.E. Choi, Mass production of adventitious roots of
Eleutherococcus sessiliflorus through the bioreactor culture, Journal of Plant
Biotechnology, 2003, 5(3), 187-191.
53. E.J. Lee and K. Y. Paek, Effect of nitrogen source on biomass and bioactive
compound production in submerged cultures of Eleutherococcus koreanum
Nakai adventitious roots, Biotechnology Progress, 2012, 28(2), 508-514.
54. A. Sliwinska, O. Olszowska, M. Furmanowa, A. Nosov, Rapid multiplication
of Polyscias filicifolia by secondary somatic embryogenesis, In vitro Cellular
and Developmental Biology, Plant, 2008, 44(2), 69-77.
55. Trịnh Việt Nga, Nguyễn Đức Minh Hùng, Bùi Minh Trí, Ngô Thị Anh Khôi,
Triệu Thị Bích, Phạm Thị Minh Tâm, Nguyễn Hữu Hổ, Nuôi nhân in vitro phôi
vô tính cây Đinh lăng lá nhỏ (Polyscias fruticosa (L.) Harms) trên hệ thống nuôi
cấy lỏng lắc, Tạp chí Nông nghiệp và Phát triển Nông thôn, 2019, 5, 25-34.

136
56. T.P. Murali, P.K. Rajeevan, K. Sheeja, V.V. Ranjan, In vitro propagation and
improvement of foliage plants, Final Report, Department of Pomology and
Floriculture, College of Horticulture, Kerala Agricultural University,
Velhmikkara, India, 2000, 52 pages.
57. A.S. Misra, Micropropagation studies with some ornamental plants, PhD
Thesis in Botany, Department of Botany, Faculty of Science, Maharaja
Sayajirao University of Baroda, Vadodara, India, 2001, 390002.
58. Thuc Dinh Ngoc and Thanh Le Nguyen, Chemical composition and
antimicrobial activity of essential oils from the leaves and stems of Schefflera
arboricola (Hayata) Merr. collected in Vietnam. Journal of Essential Oil
Bearing Plants, 2019, 22(5), 1401-1406.
59. O.R. Baghbidi and A. Jowkar, Micropropagation of dwarf Schefflera
[Schefflera arboricola (Hayata) Merr.] via direct shoot regeneration. Advances
in Horticultural Science, 2018, 32(2), 205-212.
60. B. Potduang, C. Chongsiriroeg, Y. Benmart, R. Giwanon, W. Supatanakul, S.
Tanpanich, Biological activities of Schefflera leucantha, African Journal of
Traditional, Complementary and Alternative Medicines, 2007, 4 (2), 157-164.
61. Y. Chirakiattikun and S. Prakaisrithongkham, Micropropagation of Schefflera
leucantha, Thai Science and Technology Journal, 2004, 12(2), 34-40.
62. T.B.C. Da Silva, C.O. D’Sousa Costa, A.F.C. Galvao, L.M. Bomfim, A.B.C.
Da C. Rodrigues, M.C.S. Mota, A.A. Dantas, T.R. Dos Santos, M.B.P. Soares,
D.P. Bezerra, Cytotoxic potential of selected medicinal plants in northeast
Brazil, BMC Complementary and Alternative Medicine, 2016, 16(1), 199.
63. E.T.H Franco, L.B. Gavioli, A.G. Ferreira, In vitro regeneration of
Didymopanax morototoni, Brazilan Journal Biology, 2006, 66(2A), 455-462.
64. G. Lloy and B. McCown, Commercially feasible micropropagation of moutain
leurel, Kalmia latifolia, by use of shoot tip culture. Combined Proceedings -
International Plant Propagator Society. Soc, 1980, 30(5), 421-427.
65. H.A. Méndez-Hernández, M. Ledezma-Rodríguez, R.N. Avilez-Montalvo, Y.L.
Juárez-Gómez, A. Skeete, J. Avilez-Montalvo, C. De-la-Peña, V.M. Loyola-
Vargas, Signaling overview of plant somatic embryogenesis. Frontiers in Plant
Science, 2019, 10,77.

137
66. A. Smertenko and P.V. Bozhkov, Somatic embryogenesis: life and death
processes during apical-basal patterning, Jounal of Experimental Botany,
2014, 65 (5), 1343-1360.
67. A. Iantcheva, A. Barbulova, M. Vlahova, A. Atanassov, Primary asymmetric
division and embryo formation in a single cell suspension of embryogenic
Medicago falcata, Biotechnology and Biotechnological Equipment, 2004,
18(3), 27-33.
68. S. Von Arnold, I. Sabala, P. Bozhkov, J. Dyachok, L. Filonova, Developmental
pathways of somatic embryogenesis, Plant Cell Tissue and Organ Culture, 2002,
69, 233-249.
69. S. Kadokura, K. Sugimoto, P. Tarr, T. Suzuki, S. Matsunaga, Characterization
of somatic embryogenesis initiated from the Arabidopsis shoot
apex, Developmental Biology, 2018, 442 (1), 13-27.
70. M. Zhang, A. Wang, M. Qin, X. Qin, S. Yang, S. Su, Y. Sun, L. Zhang, Direct
and indirect somatic embryogenesis induction in Camellia oleifera Abel,
Frontiers in Plant Science, 2021, 12, 644389.
71. L. Kharwanlang, MC. Das, S. Kumaria, P. Tandon, High frequency somatic
embryos induction from the rhizome explant of Panax pseudoginseng Wall.
using thin cell layer section. International Journal of Applied Biology and
Pharmaceutical Technology, 2016a, 7(3), 32-40.
72. O.T. Kim, T.S. Kim, D.S. In, K.H. Bang, Y.C. Kim, Y.E. Choi, S.W. Cha, N.S.
Seong, Optimization of direct somatic embryogenesis from mature zygotic
embryo78s of Panax ginseng C.A. Meyer, Journal of Plant Biology, 2006, 49(5),
348-352.
73. Y.J. Kim, K. Kim, O.R. Lee, D.C. Yang, High frequency of plant regeneration
through cyclic secondary somatic embryogenesis in Panax ginseng, Journal of
Ginseng Research, 2012, 36(4), 442-448.
74. D.T. Nhut, B.V.T. Vinh, T.T. Hien, N.P. Huy, N.B. Nam, H.X. Chien, Effects
of spermidine, proline and carbohydrate sources on somatic embryogenesis
from main root transverse thin cell layers of Vietnamese ginseng (Panax
vietnamensis Ha et. Grushv.), African Journal of Biotechnology, 2012, 11(5),
1084-1091.

138
75. A.H. Naing, C.K. Kim, B.J. Yun, J.Y. Jin, K.B. Lim, Primary and secondary
somatic embryogenesis in Chrysanthemum cv. Euro. Plant Cell Tiss Organ
Culture, 2013, 112, 361-368.
76. R. Zegzouti, M.F. Arnould, J.M. Favre, Histological investigation of the
multiplication step in secondary somatic embryogenesis of Quercus robur L.,
Annals of Forest Science, 2001, 58(6), 681-690.
77. H.S.M. Khierallah and N.H. Hussein, The role of coconut water and casein
hydrolysate in somatic embryogenesis of date palm and genetic stability
detection using RAPD markers. Research in Biotechnology, 2013, 4(3), 20-28.
78. Tô Thị Nhã Trầm, Trương Phi Yến, Tôn Trang Ánh, Hoàng Thanh Tùng, Hà
Thị Mỹ Ngân, Dương Tấn Nhựt, Phát sinh phôi soma cây đinh lăng lá xẻ nhỏ
(Polyscias fruticosa L. Harms) thông qua nuôi cấy mẫu lá ex vitro, Tạp chí
Công nghệ Sinh học, 2020, 18(3), 497-506.
79. G.I. Nic-Can and V.M. Loyola-Vargas, The role of the auxins during somatic
embryogenesis, Somatic Embryogenesis: Fundamental Aspects and
Applications, 2016, 171-181.
80. M.A. Abohatem, Y. Bakil, M. Baaziz, Plant regeneration from Somatic
Embryogenic Suspension Cultures of Date Palm, Methods in Molecular
Biology, 2017, 1637, 203-214.
81. T. Keshvari, A. Najaphy, D. Kahrizi, A. Zebarjadi, Callus induction and
somatic embryogenesis in Stevia rebaudiana Bertoni as a medicinal
plant, Cellular and Molecular Biology, 2018, 64(2), 46-49.
82. O. Novák and K. Ljung K, Zooming in on plant hormone analysis: tissueand
cell-specific approaches, Annual review of Plant Biology, 2017, 68, 323-348.
83. X. Yang and X. Zhang, Regulation of somatic embryogenesis in higher plants.
Critical Reviews Plant Science, 2010, 29, 36-57.
84. R.E. Márquez-López, C.A. Pérez-Hernández, Á. Kú-González, R.M. Galaz-
Ávalos, V.M. Loyola-Vargas, Localization and transport of indole-3-acetic
acid during somatic embryogenesis in Coffea canephora. Protoplasma, 2018,
255, 695-708.
85. B. Jones, S.A. Gunneras, S.V. Petersson, P. Tarkowski, N. Graham, S. May, K.
Dolezal, G. Sandberg, K.Ljung, Cytokinin regulation of auxin synthesis in

139
Arabidopsis involves a homeostatic feedback loop regulated via auxin and
cytokinin signal transduction, Plant Cell, 2010, 22(9), 2956-2969.
86. V. M. Loyola-Vargas and N. Ochoa-Alejo, Somatic Embryogenesis. An
Overeview, Somatic Embryogenesis: Fundamental Aspects and Applications,
2016, 1-8.
87. D. Leljak-Levanic, S. Mihaljevic, and N. Bauer, Somatic and zygotic embryos
share common developmental features at the onset of plant embryogenesis. Acta
Physiologiae Plantarum, 2015, 37, 1-14.
88. L. Michalczuk, T.J. Cooke, J.D. Cohen, Auxin levels at different stages of carrot
somatic embryogenesis, Phytochemistry, 1992, 31, 1097-1103.
89. D.M. Ribnicky, N. IliC, J.D. Cohen, T.J. Cooke, The effects of exogenous
auxins on endogenous indole-3-acetic acid metabolism - The implications for
carrot somatic embryogenesis, Plant Physiol, 1996, 112, 549-558.
90. B. Ayil-Gutiérrez, R.M. Galaz-Ávalos, E. Peña-Cabrera, V.M. Loyola-Vargas,
Dynamics of the concentration of IAA and some of its conjugates during the
induction of somatic embryogenesis in Coffea canephora, Plant Signaling &
Behavior, 2013, 8, e26998.
91. H.H. Chung, J.T. Chen, W.C. Chang, Plant regeneration through direct
somatic embryogenesis from leaf explants of Dendrobium. Biologia Plantarum,
2007, 51, 346-350.
92. W.P. Gow, J.T. Chen, W.C. Chang, Effects of genotype, light regime explants
possion and orientation on direct somatic embryogenesis from leaf explants of
Phalaenopsis orchids, Acta Physiologiae Plantarum, 2009, 31(2), 363-369.
93. C.S. Raju, A. Aslam, K. Kathiravan, P. Palani, A. Shajahan, Direct somatic
embryogenesis and plant regeneration from leaf sheath explants of mango
ginger (Curcuma amada Roxb.), In Vitro Cellular & Developmental Biology –
Plant, 2014, 50, 752-759.
94. Y. Wang, F. Chen, Y. Wang, X. Li, H. Liang, Plant regeneration from immature
embryos of Tapiscia sinensis Oliv., an endemic and endangered species in
China, Hortscience, 2014, 49(12), 1558-1562.
95. Bùi Văn Thế Vinh, Vũ Thị Thủy, Thái Thương Hiền, Đỗ Khắc Thịnh, Dương
Tấn Nhựt, Nghiên cứu hình thái giải phẫu và cấu trúc phôi trong quá trình phát

140
sinh phôi vô tính sâm Ngọc Linh (Panax vietnamensis Ha et Grushv.), Tạp chí
Khoa học và Phát triển, 2014, 7, 1140-1148.
96. H.J. Shen, J.T. Chen, H.H. Chung, W.C. Chang, Plant regeneration via direct
somatic embryogenesis from leaf explants of Tolumnia Louise Elmore ‘Elsa’.
Botanical Studies, 2018, 59, 4.
97. H. Liang, Y. Xiong, B. Guo, H. Yan, S. Jian, H. Ren, X. Zhang, Y. Li, S. Zeng,
K. Wu, F. Zheng, J.A. Teixeira da Silva, Y. Xiong, G. Ma, Shoot organogenesis
and somatic embryogenesis from leaf and root explants of Scaevola sericea.
Scientific Reports, 2020, 10, 11343.
98. E. Rahmat and Y. Kang, Adventitious root culture for secondary metabolite
production in medicinal plants, A Review, Journal of Plant Biotechnology,
2019, 46(3), 143-157.
99. J. Yu, W. Liu, J. Liu, P. Qin, L. Xu, Auxin control of root organogenesis from
callus in tissue culture, Frontiers in Plant Science, 2017, 8, 1835.
100. X.H. Cui, H.N. Murthy, C.H. Wu, K.Y. Paek, Sucrose-induced osmotic stress
affects biomass, metabolite, and antioxidant levels in root suspension cultures
of Hypericum perforatum L., Plant Cell Tissue and Organ Culture, 2010, 103,
7-14.
101. A.H. Ahkami, M. Melzer, M.R. Ghaffari, S. Pollmann, M.G. Javid, F.
Shahinnia, M.R. Hajirezaei, U. Druege, Distribution of indole-3-acetic acid in
Petunia hybrida shoot tip cuttings and relationship between auxin transport,
carbohydrate metabolism and adventitious root formation, Planta, 2013, 238,
499–517.
102. L. Guan, R. Tayengwa, Z. Cheng, W.A. Peer, A.S. Murphy, M. Zhao, Auxin
regulates adventitious root formation in tomato cuttings, Plant Biology, 2019,
19, 435.
103. K.H. Lo, Factor affecting shoot organogenesis in leaf disc culture of African
violet, Scientia Hoticulturae, 1997, 72(1), 49-57.
104. R.J. Rose, X.D. Wang, K.E. Nolan and B.G. Rolfe, Root meristems in Medicago
truncatula tissue culture arise from vascular-derived procambial-like cells in a
process regulated by ethylene, Journal of Experimental Botany, 2006, 57(10),
2227-2235.

141
105. T. Murashige and F. Skoog, A revised medium for rapid growth and bioassay
with tobacco tissue culture, Physiol Plant, 1962, 15, 473-497.
106. O.L. Gamborg, R.A. Miller, K. Ojima, Nutrient requirement of suspensions
cultures of soybean root cells, Experimental cell research, 1962, 50(1), 151-158.
107. R.U. Schenk and A.C. Hildebrandt, Medium and techniques for induction and
growth of monocotyledonous and dicotyledonous plant cell culture, Canadian
Journal Botany, 1972, 50, 199-204.
108. S. Merkle, W. Parrott, B. Flinn, Morphogenic Aspects of Somatic
Embryogenesis, In vitro Embryogenesis in Plants (Current Plant Science and
Biotechnology in Agriculture), 1995, 20, 155-203.
109. R. Fernández-Da Silva, L. Hermoso-Gallardo, A. Menéndez-Yuffá, Primary
and secondary somatic enbryogenesis in leaf sections and cell suspensions of
Coffea arabica cv. Catimor, Interciencia, 2005, 30(11), 694-698.
110. H.M.S. Aboshama, Direct somatic embryogenesis of pepper (Capsicum
annuum L.), World Journal of Agricultural Sciences, 2011, 7 (6), 755-762.
111. E. Minyaka, N. Niemenak, Fotso, A. Sangare, D.N. Omokolo, Effect of MgSO4
and K2SO4 on somatic embryo diffirentiation in Theobroma cacao L., Plant Cell
Tissue and Organ Culture, 2008, 94(2), 149-160.
112. M.S.D. Ibrahima, R.S. Hartatib, R. Rubiyoa, A. Purwitoc, S. Sudarsono, Direct
and indirect somatic embryogenesis on Arabica coffee (Coffea arabica),
Indonesian Journal of Agricutural Science, 2013, 14(2), 79-86.
113. Y. Aoshima Y, Efficient embryogenesis in the callus of tea (Camellia sinensis)
enhanced by the osmotic stress or antibiotics treatment, Plant Biotechnol, 2005,
22, 277-280.
114. C. Anjaneyulu, B. Shyamkuma, C. Giri, Somatic embryogenesis from callus
cultures of Terminalia chebula Retz.: An important medicinal tree, Trees –
Structure and function, 2004, 18(5), 547-552.
115. Y.J. Kim, M.K. Kim, J.S. Shim, R.K. Pulla, D.C. Yang, Somatic embryogenesis
of two new Panax ginseng cultivars, Yun-Poong and Chun-Poong, Russian
Journal of Plant Physiology, 2010, 57(2), 283-289.
116. A. Prades, M. Dornier, N. Diop, J.P. Pain, Coconut water uses, composition and
properties: A review, Fruits, 2012, 67, 87–107.

142
117. Mai Trường, Trần Thị Ngọc Hà, Phan Tường Lộc, Lê Tấn Đức, Trần Trọng
Tuấn, Đỗ Đăng Giáp, Bùi Đình Thạch, Phạm Đức Trí, Nguyễn Đức Minh Hùng,
Nguyễn Thị Thanh, Nguyễn Văn Kết, Trần Công Luận, Nguyễn Hữu Hổ,
Nghiên cứu nuôi cấy mô sẹo có khả năng sinh phôi và mô phôi sâm Ngọc Linh
(Panax vietnamensis Ha et Grushv., Tạp chí Sinh học, 2013, 35(3se), 145-157.
118. J.Y. Kim, P.B. Adhikari, C.H. Ahn, D.H. Kim, Y.C. Kim, J.K. Han, S. Kondeti,
Y.E. Choi, High frequency somatic embryogenesis and plant regeneration of
interspecific ginseng hybrid between Panax ginseng and Panax quinquefolius,
Journal Ginseng Research, 2019, 43,38-48.
119. M. Ali, A. Mujib, D. Tonk, N. Zafar, Plant regeneration through somatic
embryogenesis and genome size analysis of Coriandrum sativum L,
Protoplasma, 2017, 254(1), 343-352.
120. W.M. Irene, H.L. Alumiro, K.K. Asava, C.O. Agwanda, S.E. Anami, Effects of
genotype and plant growth regulators on callus induction in leaf cultures of
Coffea arabica L. F1 hybrid, Journal Plant Biochemistry Physiology, 2019, 7,
236.
121. P. Giridhar, V. Kumar, E.P. Indu, G.A. Ravishankar, A. Chandrasekar,
Thidiazuron induced somatic embryogenesis in Coffea arabica L. and Coffea
canephora P ex Fr, Acta Botanica Croatica, 2004, 63 (1), 25–33.
122. N. Burbulis, A. Blinstrubiene, V. Jonytiene, In vitro regeneration from leaf
explants of Petunia hybrida L. Propag Ornam Plants, 2015, 15, 47–52.
123. A. Blinstrubiene, N. Burbulis, V. Jonytiene, R. Masiene R, Evaluation of factors
affecting direct organogenesis in a somatic tissue culture of Sinningia speciosa
(Lodd.) Hiern, Agronomy, 2020, 10(11), 1783.
124. Y. Zhang, T.A. Bozorov, D.X. Li, P. Zhou, X.J. Wen, Y. Ding, D.Y. Zhang, An
efficient in vitro regeneration system from different wild apple (Malus sieversii)
explants, Plant Methods, 2020, 16,56.
125. S.B. Mostafiz, A. Wagiran, Efficient callus induction and regeneration in
selected indica rice, Agronomy, 2018, 8, 77.
126. X. Deng, Y. Xiong, J. Li, D. Yang, J. Liu, H. Sun, H. Song, Y. Wang, J. Ma, Y.
Liu, M. Yang, The establishment of an efficient callus induction system for lotus
(Nelumbo nucifera), Plants, 2020, 9(11),1436.

143
127. S.L. Zhong, S.G. Zhong, Morphological and ultrastructural characteristics of
the embryogenic callus of American ginseng, Chin J Bot, 1992, 4, 92-98.
128. M.D. Gaj, Factors influencing somatic embryogenesis induction and plant
regeneration with particular reference to Arabidopsis thaliana (L.) Heynh,
Plant Growth Regulation, 2004, 43: 27–47.
129. S. Hussein, R. Ibrahim, A.L.P. Kiong, Somatic embryogenesis: an alternative
method for in vitro micropropagation, Iranian Journal of Biotechnology, 2006,
4(3), 156-161.
130. Y. Sun, X. Zhang, S. Jin, S. Liang, Y. Nie, Somatic embryogenesis and plant
regeneration in wild cotton (Gossypium klotzschianum), Plant Cell Tissue
Organ Culture, 2003, 75, 247-253.
131. K.P. Martin, Plant regeneration through direct somatic embryogenesis on seed
coat explants of cashew (Anacardium occidentale L.), Science Horticulturae,
2003, 98(3), 299-304.
132. M. Umehara and H. Kamada H, Development of the embryo proper and the
suspensor during plant embryogenesis, Plant Biotechnology, 2005, 22, 253-260
133. O.J. Parra-Penalosa and G.O. Cancino-Escalante, Evaluation of induction of
somatic embryogenesis from cotyledonary leaves of banana passion fruit
(Passiflora mollissima) L.H Bailey, Respuestas, 2019, 24(3), 31-38.
134. SA. Kahrawat and S. Chand, Continuous somatic embyogenesis and plant
regeneration from hypocotyl segments of Psoralea corylifolia Linn., an
endangered and medicinally important Fabaceae plant, Current Science
Assocation, 2001, 81, 1328-1331.
135. D. Sharada, P.S. Krishna, N.R. Swamy, Plant Regeneration via somatic
embryogenesis in Solanum nigrum L. (Black nightshade) (Solanaceae),
Biotechnology Journal International, 2019, 23(1), 1-9.
136. E. Corredoira, A. Ballester, A.M. Vieitez, Proliferation, maturation and
germination of Castanea sativa Mill. somatic embryos originated from leaf
explants, Annals of Botany, 2003, 92(1), 129–136.
137. T. Hazubska-Przybyl, E. Ratajczak, A. Obarska, E. Pers-Kamczyc, Different
roles of auxins in somatic embryogenesis efficiency in two Picea species,
International Journal of Molecular Sciences, 2020, 21(9), 3394.

144
138. D.E. Sakr and R.M.S. Sayed, Morpho- histological observations on somatic
embryogenesis in mature embryo derived callus of Oryza sativa L. cv. Sakha,
101, Journal of Scientific Reseach in Science, 2018, 35(1), 126-141.
139. J.L.D. Cipriano, A.C.F. Cruz, K.C. Mancini, E.R. Schmildt, J.C. Lopes, W.C.
Otoni, R.S. Alexandre, Somatic embryogenesis in Carica papaya as affected by
auxins and explants, and morphoanatomical-related aspects. Anais da
Academia Brasileia de Ciencias, 2018, 90(1), 385-400.
140. E. Grzebelus, M. Szklarczyk, R. Baranski, An improved protocol for plant
regeneration from leaf and hypocotyl-derived protoplasts of carrot. Plant Cell
Tissue Organ Culture, 2012, 109, 101-109.
141. J. Friml, A. Vieten, M. Sauer, D. Weijers, H. Schwartz, T. Hamann, R. Offringa,
G. Jürgens, Efflux-dependent auxin gradients establish the apical basal axis of
Arabidopsis. Nature, 2003, 426, 147-153.
142. G.D. Ascough and C.W. Fennell, The regulation of plant growth and
development in liquid culture, South African Journal of Botany, 2004, 70(2),
181–190.
143. J.A. Teixeira da Silva, J. Dobranszki, Plant thin cell layers: update and
perspectives, Folia Hort, 2015, 27(2), 183-190.
144. V.T. Hien, N.P. Huy, B.V.T. Vinh, H.X. Chien, H.T. Tung, N.B. Nam, V.Q.
Luan, D.T. Nhut. Somatic embryogenesis from leaf transverse thin cell layer
derived-callus of Vietnamese ginseng (Panax vietnamensis Ha et Grushv.), Tạp
chí Công nghệ Sinh học, 2016, 14(1), 63-73.
145. N. Sabooni and A. Shekafandeh, Somatic embryogenesis and plant
regeneration of blackberry using the thin cell layer technique, Plant Cell Tissue
Organ Culture, 2017, 130: 313–321.
146. J.E. Scherwinski-Pereira, R.S. da Guedes, Jr P.C.P Fermino, T.L. Silva, F.H.S.
Costa, Somatic embryogenesis and plant regeneration in oil palm using the thin
cell layer technique, In Vitro Cellular and Development Biology Plant, 2010,
46, 378-385.
147. M. Pandey, U. Dhar, S.S. Samant, M.V. Shirgurkar, S.R. Thengane, Recurrent
somatic embryogenesis and plant regeneration in Angelica glauca Edgew., a

145
critically endangered medicinal plant of the Western Himalaya, Journal of
Horticultural Science & Biotechnology, 2011, 86(5), 493–498.
148. I. Sivanesan, M.S. Son, S. Jana, B.R. Jeong, Secondary somatic enbryogenesis
in Crocus vernus (L.) Hill, Propagation of Ornamental Plants, 2012, 12(3), 163-
170.
149. G. Wang, C. Xu, S. Yan, B. Xu, An efficient somatic embryo liquid culture
system for potential use in large-scale and synchronic production of Anthurium
andraeanum seedlings, Fronties in Plant Science, 2019, 10,29.
150. M. Malik, Comparison of different liquid/solid culture systems in the production
of somatic embryos from Narcissus L. ovary explants, Plant Cell Tissue and
Organ Culture, 2008, 94, 337–345.
151. A.L. De Rezende Maciel, F.A. Rodrigues, M. Moacir Pasqual, C.H.S. De
Carvalho, Large-scale, high-efficiency production of coffee somatic embryos,
Crop Breeding and Applied Biotechnology, 2016, 16, 102-107.
152. T.S. Mariani, Sjafrullatif, N.G. Wardjo, H. Miyake, Initiation and proliferation
of globular somatic embryos of oil palm (Elaeis guineensis, Jacq.) observed by
inverted and transmission electron microscopy. International Journal of Basic
& Applied Sciences, IJBAS-IJENS 2014, 14(1), 1-7.
153. C. Guillou, A. Fillodeau, E. Brulard, D. Breton, S. De Faria Maraschin, D.
Verdier, M. Simon, J.D. Ducos, Indirect somatic embryogenesis of Theobroma
cacao L. in liquid medium and improvement of embryo-to-plantlet conversion
rate, In Vitro Cellular & Developmental Biology, Plant, 2018, 54, 377-391.
154. B.B. Misra and S. Dey, Culture of East Indian sandalwood tree somatic
embryos in air-lift bioreactors for production of santalols, phenolics and
arabinogalactan proteins, AoB Plants, 2013, 5, plt 025.
155. L. García-Águila, R. Gómez-Kosky, Y. Alvarado Capó, Z. Sarría Hernández,
N. Albany, J. Vílchez, M. Reyes Vega, B. Pérez Pérez, A. Rodríguez
Concepción, Effect of inoculation density of somatic embryos for obtaining
plantain FHIA-21 (AAAB) cultivars, Cultivos Tropicales, 2016, 37(1), 57-64.
156. R.R. Nair and S.D. Gupta, High-frequency plant regeneration through cyclic
secondary somatic embryogenesis in black pepper (Piper nigrum L.), Plant Cell
Reports, 2006, 24(12), 699-707.

146
157. S. Te-chato and A. Hilae, High-frequency plant regeneration through
secondary somatic embryogenesis in oil palm (Elaeis guineensis Jacq. var.
Tenera). Journal of Agricultural Technology, 2007, 3(2), 345-357.
158. S. Hussein, R. Ibrahim, A.L.P. Kiong, N.M. Fadzillah, S.K. Daud,
Micropropagation of Eurycoma longifolia Jack via formation of somatic
embryogenesis, Asian Journal of Plant Sciences, 2005, 4(5),472-485.
159. T. Cosic, B. Vinterhalter, D. Vinterhalter, N. Mitic, A. Cingel, J. Savic, B.
Bohanec, S. Ninkovic, In vitro plant regeneration from immature zygotic
embryos and repetitive somatic embryogenesis in kohlrabi (Brassica oleracea
var. gongylodes), In Vitro Cellular & Developmental Biology – Plant, 2013, 49(3),
294-303.
160. M.S.D. Ibrahima, R.S. Hartatib, Rubiyoa, A. Purwitoc, Sudarsono. Direct and
indirect somatic embryogenesis on Arabica coffee (Coffea arabica), Indonesian
Journal of Agricultural Science, 2013, 14(2), 79-86.
161. E. Corredoira, A. Ballester, M. Ibarra, A.M. Vieitez, Induction of somatic
embryogenesis in explants of shoot cultures established from adult Eucalyptus
globulus and E. saligna × E. maidenii trees. Tree Physiology, 2015, 35, 678-
690.
162. G. Daigny, H. Paul, R.S. Sangwan, B.S. Sangwan-Norreel, Factors influencing
secondary somatic embryogenesis in Malus x domestica Borkh. (cv 'Gloster
69'), Plant Cell Reports, 1996, 16(3-4), 153-157.
163. M. Al Shamari, H.Z. Rihan, M.P. Fuller, An effective protocol for the
production of primary and secondary somatic embryos in cauliflower (Brassica
oleraceae var. Botrytis), Agri Res & Tech, 2018, 14(1), 38-46.
164. J.W.H. Yong, L. Ge, Y.F. Ng, S.N. Tan, The chemical composition and
biological properties of coconut (Cocos nucifera L.) water, Molecules, 2009,
14, 5144-5164.
165. S.M. Khalil, K.T. Cheah, E.A. Perez, D.A. Gaskill, J.S. Hu, Regeneration of
banana (Musa spp. AAB cv. Dwarf Brazilian) via secondary somatic
embryogenesis, Plant Cell Reports, 2002, 20,1128-1134.
166. H.K. Moon, S.Y. Park, Y.W. Kim, S.H. Kim, Somatic embryogenesis and
plantlet production using rejuvenated tissues from serial grafting of a mature

147
Kalopanax septemlobus tree, In Vitro Cellular & Developmental Biology.-
Plant, 2008, 44(2), 119-127.
167. I.M. Al-Khayri, Somatic Embryogenesis of Date Palm (Phoenix dactylifera L.)
Improved by Coconut Water. Biotechnology, 2010, 9(4), 477-484.
168. Y. Chirakiattikun and S. Prakaisrithongkham, Micropropagation of Schefflera
leucantha. Thai Science and Technology Journal, 2004, 12(2), 34-40.
169. J.L. Yang, E.S. Seong, M.J. Kim, B.K. Ghimire, W.H. Kang, C.Y. Yu, C.H,
Direct somatic embryogenesis from pericycle cells of broccoli (Brassica
oleracea L. var. Italica) root explants. Plant Cell Tissue and Organ Culture,
2010, 100(1), 49-58.
170. K.D. Xu, W. Wang, D.S. Yu, X.L. Li, J.M. Chen, B.J. Feng, Y. Zhao, M.J.
Cheng, X.X. Liu, C.W. Li, NAA at a high concentration promotes efficient plant
regeneration via direct somatic embryogenesis and SE-mediated
transformation system in Ranunculus sceleratus, Scientific Reports, 2019, 9,
18321.
171. M. Sukhada, K.Y. Prathibha, H.D. Soumya, R.M. Ranganath, The origin and
development of secondary somatic embryos in banana cv. Nanjangud Rasbale
(Silk GP. Rasthali, AAB). Acta Horticulture, 2010, 865, 349-351.
172. E.U. Kurczynska, M.D. Gaj, A. Ujczak, E. Mazur, Histological analysis of
direct somatic embryogenesis in Arabidopsis thaliana (L.) Heynh. Planta, 2007,
226(3), 619-628.
173. A. Onay, Histology of somatic embryogenesis in cultured leaf explants of
pistachio (Pistacia vera L.), Turkisk Journal Botany, 2000, 24, 91-95.
174. N. Imin, M. Nizamidin, D. Daniher, K.E. Nolan, R.J. Rose and B.G. Rolfe.
Proteomic analysis of somatic embryogenesis in Medicago truncatula, Plant
Physiology, 2005, 137, 1250-1260.
175. C. Thomas, D. Meyer, C. Himber C and A. Steinmetz, Spatial expression of a
sunflower SERK gene during induction of somatic embryogenesis and shoot
organogenesis. Plant Physiology and Biochemistry, 2004, 42, 35-42.
176. S.V. Eduardo, In vitro root induction and plumule explants of Helianthus
annuus, Environmental and Experimental Botany, 1998, 39, 271-277.
177. J. Yu, W. Liu, J. Liu, P. Qin, L. Xu, Auxin control of root organogenesis

148
from callus in tissue culture, Frontiers in Plant Science, 2017, 8, 1835.
178. Dương Tấn Nhựt, Một số phương pháp, hệ thống mới trong nghiên cứu công
nghệ sinh học thực vật, NXB Nông nghiệp TP. HCM, 2010, 218 tr.
179. A.P.K. Ling, K.P. Tan, S. Hussein, Comparative effects of plant growth
regulators on leaf and stem explants of Labisia pumila var, Alata. Journal of
Zhejiang University-SCIENCE B (Biomedicine & Biotechnology), 2013, 14(7),
621-631.
180. K. Roberts, Auxin, In: Handbook of Plant Science, John Wiley & Sons Ltd.,
USA, 2007, 352-360.
181. S. Hussein, A.P.K. Ling, T.H. Ng, R. Ibrahim and K.Y. Paek, Adventitious roots
induction of recalcitrant tropical woody plant, Eurycoma longifolia. Romanian
Biotechnological Letters, 2012, 17(1), 7026-7025.
182. M.Z. Saiman, N.R. Mustafa, A.E. Schulte, R. Verpoorte, Y.H. Choi, Induction,
characterization, and NMR-based metabolic profiling of adventitious root
cultures from leaf explants of Gynura procumbens, Plant Cell Tissue Organ
Culture, 2012, 109, 465–475.
183. S.N. Sharmaa, Z. Jhaa and R.K. Sinha, Establishment of in vitro adventitious
root cultures and analysis of andrographolide in Andrographis paniculata,
Natural Product Communications, 2013, 8(8), 1045-1047.
184. S. Nandagopal and B.D.R. Kumari, Effectiveness of auxin induced in vitro root
culture in chicor, Journal of Central European Agriculture, 2007, 8(1), 73-80.
185. M.M. Khalafalla, H.M. Daffalla, H.A. El-Shemy and E. Abdellatef,
Establishment of in vitro fast-growing normal root culture of Vernonia
amygdalina - a potent African medicinal plant, African Journal of
Biotechnology, 2009, 8(21), 5952-5957.
186. K.W. Yu, W.Y. Gao, E.J. Hahn and K.Y. Paek, Effects of macro elements and
nitrogen source on adventitious root growth and ginsenoside production in
ginseng (Panax ginseng C.A. Meyer), Journal of Plant Biology, 2001, 44(4),
179-184.
187. Nguyễn Thị Liễu, Nguyễn Trung Thành, Nguyễn Văn Kết, Nghiên cứu khả
năng tạo rễ bất định của sâm Ngọc Linh (Panax vietnamensis Ha et Grushv.)
trong nuôi cấy in vitro, Tạp chí Khoa học ĐHQGHN, Khoa học Tự nhiên và

149
Công nghệ, 2011, 27, 30-36.
188. H.J. Kim, E.J. Chang, H.I. Oh, Saponin production in submerged adventitious
root culture of Panax ginseng as affected by culture conditions and elicitors.
Asia Pacific Journal Molecular Biology Biotechnology, 2005, (13), 87-91.
189. D.J. James, Adventitious root formation in vitro in apple rookstock (Malus
pumila), Phyosilogia Plantarum, 1983, 57, 213 -218.
190. A.P.K. Ling, M.F. Chin, S. Hussein, Adventitious roots production of Centella
asiatica in response to plant regulators and sucrose concentration, Medicinal
and Aromatic Plant Science and Biotechnology, 2009a, 3(1), 36-41.
191. F. Taj, M.A. Khan, H. Ali and R.S. Khan, Improved production of industrially
important essential oils through elicitation in the adventitious roots of Artemisia
amygdalina, Plants, 2019, 8(10), 430.
192. V.P. Pandey, E. Cherian and G. Patani, Effect of growth regulators and culture
conditions on direct root induction of Rauwolfia serpentina L. (Apocynaceae)
Benth by leaf explants, Tropical Journal of Pharmaceutical Research, 2010,
9(1), 27-34.
193. A.P.K. Ling, K.M. Kok, S. Hussein and S.L. Ong SL, Effects of plant growth
regulators on adventitious roots induction from different explants of
Orthosiphon stamineus, American-Eurasian Journal of Sustainable Agriculture,
2009b, 3(3), 493-501.
194. J. Samaj, M. Bobak, D. Kubosnikova, J. Kristin, E. Kolarik, M. Ovecka,
A. Blehova, Bundle sheet cells are responsible for direct root regeneration from
leaf explants of Helianthus occidentalis, Journal Plant Physiology, 1999, 154,
89-94.
195. N.A. Yusuf, N.S.M. Rahim, S.Z.A. Azhar, K.A. Ghani, S. Sommano, N. Khalid,
Adventitious root cultures of Boesenbergia rotunda as a source of pinostrobin.
International Journal on Advanced Science, Engineering and Information
Technology, 2018, 8(2), 377-383.
196. H. Fazal, B.H. Abbasi, N. Ahmad, Optimization of adventitious root culture for
production of biomass and secondary metabolites in Prunella vulgaris L,
Applied Biochemistry and Biotechnology, 2014, 174(6), 2085-2095.

150
197. P.S. Kaushik, M.K. Swamy, S. Balasubramanya, M. Anuradha, Rapid plant
regeneration, analysis of genetic fidelity and camptothecin content of
micropropagated plants of Ophiorrhiza mungos Linn. - a potent anticancer
plant, Journal of Crop Science and Biotechnology, 2015, 18, 1-8.
198. P. Baskaran and N. Jayabalan, Psoralen production in hairy roots and
adventitious roots cultures of Psoralea coryfolia, Biotechnology Letters, 2009,
31, 1073-1077.
199. N. Praveen, S.H. Manohar, P.M. Naik, A. Nayeem, J.H. Jeong, H.N. Murthy,
Production of andrographolide from adventitious root cultures of Andrographis
paniculata, Current Science, 2009, 96(5), 694-697.
200. T. Khan, B.H. Abbasi, M.A. Khan, M. Azeem, Production of biomass and
useful compounds through elicitation in adventitious root cultures of Fagonia
indica. Industrial Crops and Products, 2017, 108(1), 451-457.
201. P.K. Silja and K. Satheeshkumar, Establishment of adventitious root cultures
from leaf explants of Plumbago rosea and enhanced plumbagin production
through elicitation, Industrial Crops and Products, 2015, 76, 479-486.
202. H. Wu, Y.H. Dewir, E.J. Hahn, K.Y. Paek, Optimization of culturing conditions
for the production of biomass and phenolics from adventitious roots of
Echinacea angustifolia, Journal of Plant Biology, 2006, 49(3), 193-199.
203. J.Y. Zhang, T.W. Bae, K.H. Boo, H.J. Sun, I.J. Song, C.H. Pham, M. Ganesan,
D.H. Yang, H.G. Kang, S.M. Ko, K.Z. Riu, P.O. Lim, H.Y. Lee, Ginsenoside
Production and morphological characterization of wild ginseng (Panax ginseng
Meyer) mutant lines induced by g-irradiation (60Co) of adventitious roots,
Journal Ginseng Research, 2011, 35(3), 283-293.
204. A. Ahmad, H. Ali, H. Khan, A. Begam, S. Khan, S.S. Ali, N. Ahmad, H. Fazal,
M. Ali, C. Hano, N. Ahmad and B.H. Abbasi, Effect of gibberellic acid on
production of biomass, polyphenolics and steviol glycosides in adventitious root
cultures of Stevia rebaudiana (Bert.), Plants, 2020, 9, 420.
205. M. Ziv, Quality of micropropagated plants - vitrification. In Vitro Cellular
Biology, 1991, 27, 64–69.

0
PHỤ LỤC
Thành phần môi trường MS (Murashige và Skoog, 1962), SH (Schenk và
Hildebrandt, 1972) và B5 (Gamborg và cs., 1968)
Thành phần MS (mg/L) SH (mg/L) B5 (mg/L)
Khoáng đa lượng
KNO3 1.900 2.500 2.500
NH4NO3 1.650
NH4H2PO4 300
(NH4)2SO4 134
MgSO4.·7H2O 370 400 250
CaCl2·2H2O 440 200 150
KH2PO4 170
NaH2PO4·H2O 150
Khoáng vi lượng
MnSO4·H2O 10,0 10,0
MnSO4·4H2O 22,3
KI 0,83 1,0 0,75
H3BO3 6,2 5,0 3,0
ZnSO4·7H2O 8,6 1,0 2,0
CuSO4·5H2O 0,025 0,2 0,025
Na2MoO4·2H2O 0,25 0,1 0,25
CoCl2·6H2O 0,025 0,1 0,025
FeSO4·7H2O 27,8 15,0 27,8
Na2EDTA 37,3 20,0 37,3
Vitamin
Nicotinic acid 0,5 5,0 1,0
Pyridoxine-HCl 0,5 0,5 1,0
Thiamine-HCl 0,1 5,0 10,0
myo-Inositol 100 1.000 100
Glycine 2.0
Copyright © 2022 FDOKUMEN




















