
Neural Networks Underlying Epileptic Humming
12
Neural Networks Underlying Epileptic Humming *Fabrice Bartolomei, †Fabrice Wendling, *Jean-Pierre Vignal, *Patrick Chauvel, and *Catherine Lie ´geois-Chauvel *Service de Neurophysiologie Clinique et Laboratoire de Neurophysiologie et Neuropsychologie, INSERM EMI 9926, Universite ´ de la Me ´diterrane ´e, Marseille; and †Laboratoire Traitement du Signal et de L’Image, INSERM EPI 9934, Universite ´ de Rennes, Campus de Beaulieu, Rennes, France Summary: Purpose: Humming is a rare automatism occurring in partial seizures that has received little attention. Its study could shed light on the neural networks underlying melodic expression. In this study, we examined the anatomoelectroclini- cal correlates of humming during epileptic seizures Methods: Three patients undergoing presurgical stereoelec- troencephalography (SEEG) for medically intractable temporal lobe epilepsy were studied. Coherence analysis of SEEG ac- tivity was carried out to study the functional coupling of dif- ferent regions of the brain, whereas time–frequency (TF) analysis was conducted to assess epileptic discharge patterns. Changes in coherence were studied to identify the neural struc- tures/systems implicated in humming. Results: Humming began after the onset of seizures gener- ated in medial limbic regions of the temporal lobe. At seizure onset, coherence analysis showed an increase in amygdala– hippocampus coupling. Humming began after the onset of a rhythmic discharge over lateral regions of the superior temporal gyrus (STG). A highly significant increase in coherence was observed between prefrontal regions and the STG. TF analysis of the STG discharge showed a reproducible pattern with a single fundamental frequency and associated harmonics. This frequency was ∼6 Hz for two patients and 15 Hz for one pa- tient. Conclusions: These findings suggest that the occurrence of humming during epileptic seizures of the temporal lobe is as- sociated with activity in a neural network involving the STG and the inferior frontal gyrus. Key Words: Coherence— Superior temporal gyrus—Frontal lobe. Identifying the cerebral mechanisms underlying the clinical manifestations of epileptic seizures can provide insight into the physiologic processes underlying non- pathologic behavior in the normal brain. The study of abnormal perceptual processes, such as illusions or hal- lucinations, for example, has contributed to a better un- derstanding of how different regions in the normal brain process and integrate sensory information. Alterations in memory processing, such as “de ´ja ` vu” or the “dreamy state,” have similarly provided data on memory functions in the normal brain and have provided evidence in favor of a coactivation of the hippocampus and associative cortex in normal declarative memory (1,2). Although ab- normal motor or gestural behavior (i.e., forced acting automatisms) may involve some of the same processes as those underlying nonpathologic motor behavior, their pathophysiology remains largely unexplored. Generally accompanied by disturbances of conscious- ness, complex motor phenomena occurring with epileptic seizures have been observed and described but rarely studied in correlation with electrophysiologic data (3). With respect to disordered speech, language, and musical processing occurring with seizures, the receptive aspects of such processing have been more extensively studied than the expressive aspects. Complex auditory halluci- nations, such as musical hallucinations, are generally as- sociated with ictal discharges in the superior temporal gyrus (STG) and have generated a great deal of discus- sion on the perception of music and on the formation of auditory memories in the normal brain (4,5). Conversely, disordered verbal expression (i.e., vocalizations, verbal automatisms, or jargon) and the neural substrates that underlie it have received little attention in the literature [except where their study has been used to determine the hemispheric lateralization of the epileptogenic zone (6,7)]. Similarly, little is known of the mechanisms un- derlying humming or singing during epileptic seizures. With respect to these latter mechanisms, some evi- dence suggests that seizures associated with melodic singing originate in the temporal lobe (8,9). Singing was observed during postdischarges induced by the stimula- tion of medial temporal lobe structures (9). Various case Accepted April 11, 2002. Address correspondence and reprint requests to Dr. F. Bartolomei at Service de Neurophysiologie Clinique, CHU Timone-264 Rue st Pierre, 13005-Marseille, France. Epilepsia, 43(9):1001–1012, 2002 Blackwell Publishing, Inc. © International League Against Epilepsy 1001
-
Upload
independent -
Category
Documents
-
view
0 -
download
0
Transcript of Neural Networks Underlying Epileptic Humming

Neural Networks Underlying Epileptic Humming
*Fabrice Bartolomei, †Fabrice Wendling, *Jean-Pierre Vignal, *Patrick Chauvel, and*Catherine Liegeois-Chauvel
*Service de Neurophysiologie Clinique et Laboratoire de Neurophysiologie et Neuropsychologie, INSERM EMI 9926, Universitede la Mediterranee, Marseille; and †Laboratoire Traitement du Signal et de L’Image, INSERM EPI 9934, Universite de Rennes,
Campus de Beaulieu, Rennes, France
Summary: Purpose: Humming is a rare automatism occurringin partial seizures that has received little attention. Its studycould shed light on the neural networks underlying melodicexpression. In this study, we examined the anatomoelectroclini-cal correlates of humming during epileptic seizures
Methods: Three patients undergoing presurgical stereoelec-troencephalography (SEEG) for medically intractable temporallobe epilepsy were studied. Coherence analysis of SEEG ac-tivity was carried out to study the functional coupling of dif-ferent regions of the brain, whereas time–frequency (TF)analysis was conducted to assess epileptic discharge patterns.Changes in coherence were studied to identify the neural struc-tures/systems implicated in humming.
Results: Humming began after the onset of seizures gener-ated in medial limbic regions of the temporal lobe. At seizure
onset, coherence analysis showed an increase in amygdala–hippocampus coupling. Humming began after the onset of arhythmic discharge over lateral regions of the superior temporalgyrus (STG). A highly significant increase in coherence wasobserved between prefrontal regions and the STG. TF analysisof the STG discharge showed a reproducible pattern with asingle fundamental frequency and associated harmonics. Thisfrequency was ∼6 Hz for two patients and 15 Hz for one pa-tient.
Conclusions: These findings suggest that the occurrence ofhumming during epileptic seizures of the temporal lobe is as-sociated with activity in a neural network involving the STGand the inferior frontal gyrus. Key Words: Coherence—Superior temporal gyrus—Frontal lobe.
Identifying the cerebral mechanisms underlying theclinical manifestations of epileptic seizures can provideinsight into the physiologic processes underlying non-pathologic behavior in the normal brain. The study ofabnormal perceptual processes, such as illusions or hal-lucinations, for example, has contributed to a better un-derstanding of how different regions in the normal brainprocess and integrate sensory information. Alterations inmemory processing, such as “deja vu” or the “dreamystate,” have similarly provided data on memory functionsin the normal brain and have provided evidence in favorof a coactivation of the hippocampus and associativecortex in normal declarative memory (1,2). Although ab-normal motor or gestural behavior (i.e., forced actingautomatisms) may involve some of the same processes asthose underlying nonpathologic motor behavior, theirpathophysiology remains largely unexplored.
Generally accompanied by disturbances of conscious-ness, complex motor phenomena occurring with epileptic
seizures have been observed and described but rarelystudied in correlation with electrophysiologic data (3).With respect to disordered speech, language, and musicalprocessing occurring with seizures, the receptive aspectsof such processing have been more extensively studiedthan the expressive aspects. Complex auditory halluci-nations, such as musical hallucinations, are generally as-sociated with ictal discharges in the superior temporalgyrus (STG) and have generated a great deal of discus-sion on the perception of music and on the formation ofauditory memories in the normal brain (4,5). Conversely,disordered verbal expression (i.e., vocalizations, verbalautomatisms, or jargon) and the neural substrates thatunderlie it have received little attention in the literature[except where their study has been used to determine thehemispheric lateralization of the epileptogenic zone(6,7)]. Similarly, little is known of the mechanisms un-derlying humming or singing during epileptic seizures.
With respect to these latter mechanisms, some evi-dence suggests that seizures associated with melodicsinging originate in the temporal lobe (8,9). Singing wasobserved during postdischarges induced by the stimula-tion of medial temporal lobe structures (9). Various case
Accepted April 11, 2002.Address correspondence and reprint requests to Dr. F. Bartolomei at
Service de Neurophysiologie Clinique, CHU Timone-264 Rue stPierre, 13005-Marseille, France.
Epilepsia, 43(9):1001–1012, 2002Blackwell Publishing, Inc.© International League Against Epilepsy
1001
reports have described “automatic” musical expressionduring seizures. Meiekord and Shorvon (10) described apatient whose seizures were accompanied with melodicexpression, which was variable from one seizure to thenext and which occurred after a discharge recorded fromright temporal scalp electrodes. The case of a womanwith an extensive left-hemispheric frontal tumor withparoxysmal episodes of musical hallucinations often as-sociated with the singing of familiar songs was reported(11). Although ictal EEG data were not acquired in thisstudy (therefore rendering anatomoelectroclinical corre-lations impossible), this observation suggests that frontalstructures may be implicated in singing during epilepticseizures. In a case involving whistling, the authors ofanother report (12) suggested a role of the temporal lobein musical expression, but again anatomic and electro-physiologic correlations were not obtained. The only au-thor to provide such correlations was Wieser (13), whoobserved ictal discharges in Heschl’s gyrus in a patientwho sang Portuguese songs during a temporal lobe statusepilepticus.
In the present study, we examined the anatomoelec-troclinical correlates of humming automatisms duringepileptic seizures in three patients undergoing presurgi-cal stereoelectroencephalographic (SEEG) evaluation formedically intractable temporal lobe epilepsy (TLE). Inaddition to visual analysis of SEEG recordings, coher-ence analysis of SEEG activity was conducted to studythe functional coupling of different regions in the brainduring different parts of the seizures. This method waspreviously found to be well suited to the study of cog-nitive processes in human and nonhuman subjects (14–17). Coherence describes the phase-locked componentshared by two signals and can detect large-scale corticalinteractions. In the present study, we examined the elec-trophysiologic changes in SEEG signals and the varia-tions of coherence values between different regionsimplicated in epileptic seizures to address the followingquestions: (a) What are the cerebral mechanisms of hum-ming during temporal lobe seizures? and (b) What in-sight do these mechanisms provide into the physiologicprocesses underlying humming and singing, and what istheir relation with the cerebral mechanisms underlyingmusic perception?
METHODS
SubjectsSubjects consisted of three patients (three women,
aged 31–44 years) undergoing preoperative evaluationfor the treatment of medically intractable epilepsy.Evaluation was based on clinical history, neurologic,neuropsychological, and magnetic resonance imaging(MRI) examination, and video-EEG recordings of sei-zures. These patients exhibited humming during sei-
zures. Two patients (cases 1 and 2) were diagnosed withcryptogenic TLE, associated with hippocampal sclerosisin case 1. The remaining patient (case 3) was diagnosedwith symptomatic TLE associated with a lesion of theleft middle and inferior temporal gyri.
Stereoelectroencephalography studyAfter completion of noninvasive presurgical proce-
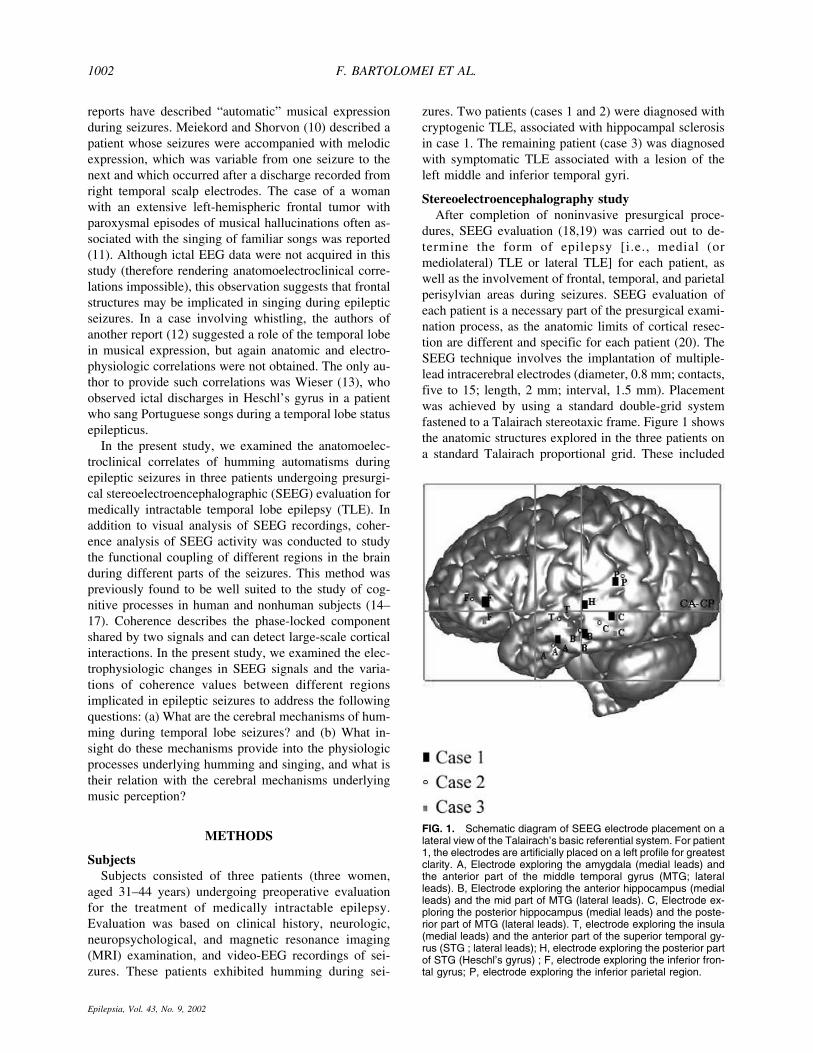
dures, SEEG evaluation (18,19) was carried out to de-termine the form of epilepsy [i.e., medial (ormediolateral) TLE or lateral TLE] for each patient, aswell as the involvement of frontal, temporal, and parietalperisylvian areas during seizures. SEEG evaluation ofeach patient is a necessary part of the presurgical exami-nation process, as the anatomic limits of cortical resec-tion are different and specific for each patient (20). TheSEEG technique involves the implantation of multiple-lead intracerebral electrodes (diameter, 0.8 mm; contacts,five to 15; length, 2 mm; interval, 1.5 mm). Placementwas achieved by using a standard double-grid systemfastened to a Talairach stereotaxic frame. Figure 1 showsthe anatomic structures explored in the three patients ona standard Talairach proportional grid. These included
FIG. 1. Schematic diagram of SEEG electrode placement on alateral view of the Talairach’s basic referential system. For patient1, the electrodes are artificially placed on a left profile for greatestclarity. A, Electrode exploring the amygdala (medial leads) andthe anterior part of the middle temporal gyrus (MTG; lateralleads). B, Electrode exploring the anterior hippocampus (medialleads) and the mid part of MTG (lateral leads). C, Electrode ex-ploring the posterior hippocampus (medial leads) and the poste-rior part of MTG (lateral leads). T, electrode exploring the insula(medial leads) and the anterior part of the superior temporal gy-rus (STG ; lateral leads); H, electrode exploring the posterior partof STG (Heschl’s gyrus) ; F, electrode exploring the inferior fron-tal gyrus; P, electrode exploring the inferior parietal region.
F. BARTOLOMEI ET AL.1002
Epilepsia, Vol. 43, No. 9, 2002

medial temporal structures (amygdala and hippocam-pus), the STG, the middle temporal gyrus (MTG), aninferior parietal region, and prefrontal regions of the in-ferior frontal gyrus (IFG).
The electrophysiological study of hummingduring seizures
The SEEG correlates of humming were studied in thethree patients. SEEG signals were visually analyzed andquantitatively processed according to two methods: (a)time–frequency (TF) distributions were computed to in-vestigate the evolution of signal spectral features as afunction of time, and (b) coherence was measured tocharacterize functional couplings between different brainstructures during epileptic seizures and humming.
Time–frequency representation of SEEG signalsTF representations are now well established, and an
intensive literature at both theoretic and applied levels isavailable (21). In the present study, the smoothed pseudoWigner-Ville distribution (SPWVD) was chosen to char-acterize the TF content of depth-EEG signals recorded inepilepsy patients. Previous studies have shown that thismethod, which provides separable smoothing in time andfrequency, allows the temporospectral content of SEEGsignals to be visualized with a good resolution in bothtime and frequency (22).
Coherence analysis of SEEG signalsThe relation between two areas in the brain may be
investigated through the study of the statistical relationbetween signals generated by these two areas. Coherenceanalysis belongs to the set of linear methods commonlyused to estimate this statistical coupling (23). Thismethod quantifies the interdependence (linear correla-tion) between two signals as a function of time and fre-quency. This information is not only capital in thecharacterization of the functional coupling betweenstructures that give rise to the two considered signals but
also is very difficult to acquire through visual inspectionof the signals.
In brief, coherence may be seen as the generalizationof a linear correlation coefficient computed on two timeseries. It quantitatively reflects the extent to which twosignals are related one to the other at a given frequency.Coherence values range from 0 to 1: values close to 1indicate coincident, or “coherent,” activities, whereasvalues close to 0 indicate independent activities, at theconsidered frequency. From coherence, a quantity easierto interpret (referred to as the average coherence) may beproduced by averaging coherence values within a givenfrequency range. We recently introduced a new bias-reduced estimator of the average coherence, obtained bysubtracting a bias computed on long-duration simulatedtime series from coherence values computed on signalsto be analyzed [see (24,25) for a description of thismethod].
This estimator is used in the present study. Averagecoherence values were computed within a user-selectedfrequency band every 0.64 s and over a 12.8-s windowsliding on SEEG signals sampled at 200 Hz.
Statistical analysis of coherence valuesCoherence values were compared before and during a
given period of interest to study variations in the func-tional coupling of different cerebral structures.
Two seizures accompanied by humming were studiedfor each patient. Coherence values from different periodsof interest were compared to identify the neural struc-tures involved at the onset of seizure activity as well asduring humming (Fig. 2). To identify epileptogenicstructures, values obtained 20 s before seizure onset(PreI) were compared with values obtained after seizureonset (I). For the identification of the neural structuresimplicated in humming, values obtained 20 s beforehumming (PreH) were compared with values obtainedafter the onset of humming (H). At each of the four
FIG. 2. Illustration of the four periodsstudied by coherence analysis of signal:Curve showing changes in coherence val-ues during seizure in patient 1. Valueswere calculated between bipolar signalsfrom electrodes in the right superior tem-poral gyrus (STG) and electrodes in thefrontal lobe (F). The numbers in parenthe-sis indicate the various periods of study:(1) the preictal period (preI), (2) the initialictal period (I), (3) the prehumming period(PreH), and (4) the humming period (H).The duration of each period was 20 s.
NEURAL NETWORKS AND HUMMING 1003
Epilepsia, Vol. 43, No. 9, 2002

periods, interactions between regions of the brain con-taining intracerebrally implanted electrodes were studied.
The cerebral regions implicated in humming wereidentified as those regions for which coherence valueswere higher at H than at PreH. Bipolar derivations cor-responding to the greatest variations in coherence wereretained for statistical analysis.
A methodologic problem associated with studying co-herence comes from the fact that coherence values arecorrelated in time (i.e., a sliding window is used). Co-herence data cannot be used directly in statistical teststhat evaluate differences in the distributions of two in-dependent samples. In the present study, this problemwas resolved by decorrelating the coherence series bymeans of an inverse autoregressive (AR) filtering opera-tion. In brief, this operation is based on the assumptionthat a coherence series can be assimilated to a filterednoise with a normal distribution and that, as such, pass-ing it through the inverse of its estimated AR filter re-sults in normal noise distribution with zero mean andvariance. After AR filtering, statistical tests of conform-ity may be executed on the two series of values.
For inverse filtering, an AR model was estimated fromthe coherence series that were to be compared. Modelorder was determined from an Akaike criterion (26).Model order 6 was often found to be appropriate. Re-sulting innovations were used in a classic F test (27) forsignificantly different variances. The F test tests the hy-pothesis that two samples have different variances (i.e.,that the null hypothesis is false). In this test, the con-structed statistical F is the ratio of one variance to an-other, such that ratios significantly less or greater than 1will indicate significant differences. Small p values cor-respond to significantly different variances of the com-pared series and, by extension, to significantly different
series of coherence values. For the present study, thelevel of significance was defined as p < 0.05.
RESULTS
Clinical dataClinical findings are summarized in Tables 1 and 2. In
all cases, the onset of humming occurred between 3 and20 s (mean, 13 s) after seizure onset. In cases 1 and 3, thehummed melody was an unfamiliar tune that was similaracross seizures. In case 2, humming episodes were char-acterized by a rhythmic repetition of the syllable “la” anda less distinct melody. The duration of humming epi-sodes was greater in case 2 (35 s) than in case 1 (20 s)and case 3 (16 s). The clinical profile of each patient’shumming episodes was the same across seizures.
SEEG dataThe SEEG data clearly indicate a temporal lobe origin
of epileptic seizures for all patients (Table 1). The epi-leptogenic zone was in the left hemisphere in two casesand in the right hemisphere in one case. Seizures weregenerated in medial temporal structures (amygdala andhippocampus) in case 1, and in anterior neocortical struc-tures (anterior and middle regions of the MTG) and me-dial structures in cases 2 and 3. Seizures were classifiedas medial TLE (MTLE) or medial–lateral TLE (MLTLE)seizures (25).
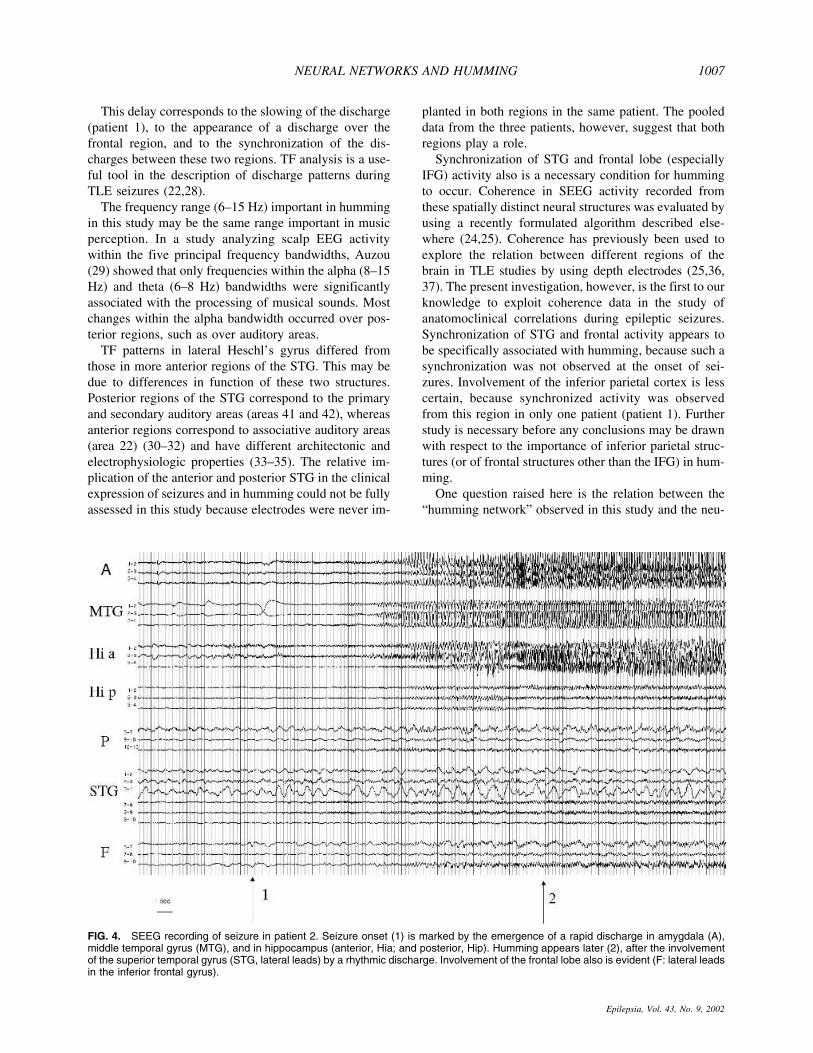
The occurrence of humming did not coincide with theinitial phase of the ictal discharge. The most notablefinding in all patients was that humming began immedi-ately after the appearance of a rhythmic discharge overthe lateralmost regions of the STG (i.e., over associativeauditory areas) as well as the emergence of slow-wavesdischarge over the frontal lobe electrodes (Figs. 3 and 4).This rhythmic discharge appeared during the propagation
TABLE 1. Clinical findings
Patients Sex HandnessAge(yr) MRI/pathology
Type ofTLE
Clinical symptoms during seizures (chronology)
1 2 3
1 F RH 31 HA R-MTLE Epigastric sensation,nausea, anxiety
Loss of contact Smile, humming,rubefaction
2 F RH 44 N L-LMTLE Groaning, swallowing Loss of contact Humming3 F RH 32 Gangliogloma anterior
part MTGL-LMTLE Smile, sniffing Loss of contact
(partial), hummingMydriase, rubefaction,
aphasia
SEEG data and propagation of seizures
PatientsNumber of
recorded seizuresNumber of
seizures with HSA
Chronology of anatomic structures involvement
A Ant H Post H MTG STG F P
1 7 5 1 (S, LVD) 1 (S, LVD) 2 (S) 3 (SW) 4 (SW) 5 (Sl W) 5 (Sl W)2 3 3 1 (LVD) 1 (LVD) 1 (LVD) 2 (SW) 4 (SW) 5 (Sl W) NE3 2 2 1 (S) 2 (S) 3 (S) 1 (S) 4 (S) 5 (Sl W) 5 (Sl W)
F, female; M, male; RH, right-handed; HA, hippocampal atrophy; N, normal; MTG, middle temporal gyrus; R-MTLE, right mesial temporal lobeepilepsy; L-MLTLE, left mesial temporal lobe epilepsy; L-LMTLE, left lateral–mesial temporal lobe epilepsy.
Numbers indicate the chronology of involvement during seizure. (), aspect of the discharge; S spikes; LVD, fast low-voltage discharge; Sl W, slowwaves; SW, spike–waves; A, amygdala; Ant H, anterior part of the hippocampus; Post H, posterior part of the hippocampus; MTG, middle temporalgyrus; STG, superior temporal gyrus; F, inferior frontal gyrus; P, inferior parietal cortex; gray areas, periods of humming.
F. BARTOLOMEI ET AL.1004
Epilepsia, Vol. 43, No. 9, 2002

of ictal activity to other medial structures and to theMTG. In case 1, secondary involvement of medial re-gions of Heschl’s gyrus was observed. In this same pa-tient, humming did not occur with every seizure, andwhen humming was absent, discharges were entirely lim-ited to medial structures and did not propagate to tem-poral neocortical regions.
Coherence analysisBy the use of coherence, we attempted to correlate the
recorded signals between the different brain regions, be-fore and during humming. Table 3 shows the main re-sults from statistical analyses of coherence valuesbetween different cerebral regions after seizure onset(PreI vs. I) and after the onset of humming (PreH vs. H)The results from each patient were generally consistentacross seizures.
Seizure onset was marked by a significant increase incoherence between medial temporal structures (i.e., theamygdala and the hippocampus). For the two patientswith MLTLE (cases 2 and 3), coherence increased be-tween medial structures and anterior regions of the MTG,consistent with previous data (25). These structures makeup the “epileptogenic network” (Fig. 5a). Seizures werenot associated with significant variations in coherencebetween frontal and temporal regions, frontal and pari-etal regions, or medial temporal regions and the STG.
Humming, conversely, was associated with a signifi-cant increase in coherence between prefrontal regions(lateral leads in the IFG region) and the STG (lateralleads). In cases 1 and 2 (seizure 2), coherence also in-creased between the STG and parietal structures, as wellas (albeit to a lesser extent) between parietal and frontalregions.
A “humming network” constructed from coherencevalues is schematized in Fig. 5b. This network is distinctfrom the network implicated in epileptic zone (Fig. 5a).Patient 1 did not hum in every seizure. Study of theinteractions in this case show that the synchrony betweenSTG and the IFG did not occur (Fig. 6).
Time–frequency analysisSTG discharges observed in patients exhibiting hum-
ming automatisms were analyzed by using the TF algo-
rithm. In all cases, humming was associated with theappearance of a particular TF pattern in this region(Table 2; Fig. 7). A low or medium fundamental fre-quency (along with higher harmonic frequencies) wasobserved. This frequency was ∼6 Hz in cases 2 and 3 and15 Hz in case 1. Harmonic frequency was 40 Hz in case1, which was markedly different from that observed incases 2 and 3.
DISCUSSION
To our knowledge, this study is the first to explore theanatomic and electrophysiologic correlates of hummingduring epileptic seizures. Intracerebral recordings ob-tained in this study suggest that humming in TLE is nothemisphere dependent, because it was observed in pa-tients with left or right TLE. Interestingly, humming oc-curred with a significantly later onset than that of seizureactivity (occurring after ictal activity propagated out ofthe epileptogenic zone) and was associated with a dif-ferent type of discharge. In all cases, the epileptogeniczone was localized in limbic temporal and/or anteriortemporal neocortical structures [corresponding to the lo-calization of the epileptogenic zone in MTLEs andMLTLEs (25)], and seizure onset was characterized byfast ictal discharges. SEEG activity observed at the onsetof humming, conversely, was characterized by a lower-frequency rhythmic discharge in other temporal and ex-tratemporal structures.
An appreciation of the entire set of neural structuresthat may be implicated in humming was not possible inthis study, because electrodes were implanted in one re-gion of interest only. For the same reason, the extent towhich both cerebral hemispheres are implicated in hum-ming could not be assessed within the same patient.
However, our findings suggest that humming occursonly under a certain definable set of conditions. First,ictal activity in anteromedial limbic regions must be fol-lowed by discharge activity in the STG, and the fre-quency of this discharge activity must be within a certainrange (∼6 Hz in the anterior STG and 15 Hz in theposterior STG). The humming starts 0–4 s after the onsetof discharge in STG (Figs. 3 and 4; Table 2).
TABLE 2. SEEG correlations with humming and time–frequency characteristics of STG discharges
Patients/seizures
Lag time ofhumming occurrence
after seizure onset(seconds)
Lag time of MTGinvolvement after
seizure onset(seconds)
Lag time of STGinvolvement after
seizure onset(seconds)
Dominant frequencyof TF pattern over
STG electrodes(onset and end, Hz)
TF patternduration over
STG electrodes(seconds)
Hummingduration(seconds)
1/1 18 11 18 15–12 10 301/2 20 10 20 15–12 18 262/1 7 0 6 7.2–6.3 25 162/2 3 0 3 7.6–6.6 22 183/1 16 1 12 6.9–3.6 36 373/2 16 1 12 7.4–4 37 35
NEURAL NETWORKS AND HUMMING 1005
Epilepsia, Vol. 43, No. 9, 2002

FIG. 3. SEEG recording of seizure in patient 1. A: Seizure onset with first clinical symptoms (1). Seizure starts by a fast low-voltagedischarge in amygdala (A) and in hippocampus (anterior, Hia; and posterior, Hip). B: The ictal activity spreads to the lateral temporalneocortex (MTG, middle temporal gyrus). A rhythmic activity is then seen over the leads exploring the lateral part of the inferior frontalgyrus (F), the superior temporal gyrus (STG, lateral leads 8–9, 9–10) and the parietal region (P). Humming appears in 2 (arrow).
F. BARTOLOMEI ET AL.1006
Epilepsia, Vol. 43, No. 9, 2002
This delay corresponds to the slowing of the discharge(patient 1), to the appearance of a discharge over thefrontal region, and to the synchronization of the dis-charges between these two regions. TF analysis is a use-ful tool in the description of discharge patterns duringTLE seizures (22,28).
The frequency range (6–15 Hz) important in hummingin this study may be the same range important in musicperception. In a study analyzing scalp EEG activitywithin the five principal frequency bandwidths, Auzou(29) showed that only frequencies within the alpha (8–15Hz) and theta (6–8 Hz) bandwidths were significantlyassociated with the processing of musical sounds. Mostchanges within the alpha bandwidth occurred over pos-terior regions, such as over auditory areas.
TF patterns in lateral Heschl’s gyrus differed fromthose in more anterior regions of the STG. This may bedue to differences in function of these two structures.Posterior regions of the STG correspond to the primaryand secondary auditory areas (areas 41 and 42), whereasanterior regions correspond to associative auditory areas(area 22) (30–32) and have different architectonic andelectrophysiologic properties (33–35). The relative im-plication of the anterior and posterior STG in the clinicalexpression of seizures and in humming could not be fullyassessed in this study because electrodes were never im-
planted in both regions in the same patient. The pooleddata from the three patients, however, suggest that bothregions play a role.
Synchronization of STG and frontal lobe (especiallyIFG) activity also is a necessary condition for hummingto occur. Coherence in SEEG activity recorded fromthese spatially distinct neural structures was evaluated byusing a recently formulated algorithm described else-where (24,25). Coherence has previously been used toexplore the relation between different regions of thebrain in TLE studies by using depth electrodes (25,36,37). The present investigation, however, is the first to ourknowledge to exploit coherence data in the study ofanatomoclinical correlations during epileptic seizures.Synchronization of STG and frontal activity appears tobe specifically associated with humming, because such asynchronization was not observed at the onset of sei-zures. Involvement of the inferior parietal cortex is lesscertain, because synchronized activity was observedfrom this region in only one patient (patient 1). Furtherstudy is necessary before any conclusions may be drawnwith respect to the importance of inferior parietal struc-tures (or of frontal structures other than the IFG) in hum-ming.
One question raised here is the relation between the“humming network” observed in this study and the neu-
FIG. 4. SEEG recording of seizure in patient 2. Seizure onset (1) is marked by the emergence of a rapid discharge in amygdala (A),middle temporal gyrus (MTG), and in hippocampus (anterior, Hia; and posterior, Hip). Humming appears later (2), after the involvementof the superior temporal gyrus (STG, lateral leads) by a rhythmic discharge. Involvement of the frontal lobe also is evident (F: lateral leadsin the inferior frontal gyrus).
NEURAL NETWORKS AND HUMMING 1007
Epilepsia, Vol. 43, No. 9, 2002

ral systems underlying the processing of music in gen-eral. Most studies on the cerebral underpinnings of musicperception suggest that both hemispheres play an impor-tant role, but that the two hemispheres process differentaspects of music. The right hemisphere plays a crucialrole in the processing of contour/interval information
(necessary in the discrimination of melody), whereas theleft hemisphere is specific for intervals processing(38,39). Networks involving posterior temporal and pre-frontal regions also have been described in music-perception studies. A coherence study of scalp EEG data(16) reported coherent frontal and temporoparietal activ-
FIG. 5. Schematic diagram of the epilep-togenic network and “humming network” inthe three patients with humming duringtemporal lobe epilepsy (TLE) seizures(cases 1–3) according to coherence varia-tions. Only significant and consistent coher-ence changes are represented (arrows). a:the epileptogenic zone included the amyg-dala and hippocampus (and the middletemporal gyrus in case 2 and 3). STG, su-perior temporal gyrus; F, inferior frontal gy-rus; P, inferior parietal cortex; A, amygdala;Hi, hippocampus. b: “humming” network.The role of STG as well as its relation withthe prefrontal cortex is shown. Interactionswith parietal cortex have been depicted be-cause they were noted in the two patientswith electrodes in this region. STG, superiortemporal gyrus; F, inferior frontal gyrus; P,inferior parietal cortex; A, amygdala; Hi, hip-pocampus.
TABLE 3. Statistical analysis of coherence variations in patients with humming
Coherencebetween
Patient 1 Patient 2 Patient 3
Pre I/I Pre HSA/HSA Pre I/I Pre HSA/HSA Pre I/I Pre HSA/HSA
STG-F NS 0.00074 NS 0.00006 NS 0.025NS 0.0006 NS 0.00002 NS 0.001
STG-P NS 0.0027 NS NS — —NS 0.0085 NS 0.004 — —
STG-HI NS NS NS NS NS NSNS NS NS 0.010 NS NS
STG-A NS NS NS NS NS NSNS NS NS NS NS NS
A-HI 0.0022 NS 0.0079 NS 0.035 NS0.0002 NS 0.0050 NS 0.0001 NS
STG-MTG NS NS NS NS NS 0.023NS NS NS NS NS 0.030
MTG-HI NS NS 0.032 0.008 0.018 NSNS NS 0.0001 0.0001 0.0045 NS
F-HI NS NS NS NS NS NSNS NS NS NS NS NS
P-HI NS NS NS NS — —NS NS NS NS — —
F-MTG NS NS NS 0.0009 NS NSNS NS NS 0.004 NS NS
A-F NS NS NS NS NS NSNS NS NS NS NS NS
A-MTG NS NS 0.0015 NS 0.0001 0.022NS NS <0.0001 NS 0.0001 NS
MTG-P NS NS NS NS — —NS NS NS NS — —
P-F NS 0.0030 NS 0.046 — —NS 0.019 NS 0.02 — —
The interactions between brain regions are represented in the left column. The PreI/I and PreH/H periods are studied in each patient (see definitionin the text). The results from two seizures in each patient are reported. A strong interaction between STG and F is invariably found for the two seizuresin each patient.
STG, superior temporal gyrus; F, inferior frontal gyrus; P, inferior parietal cortex; HI, anterior hippocampus; A, amygdala; MTG, middle temporalgyrus (mid part); NS, nonsignificant interaction (p > 0.05).
F. BARTOLOMEI ET AL.1008
Epilepsia, Vol. 43, No. 9, 2002

FIG. 6. Average coherence estimated on signals recorded from the superior temporal gyrus (STG) and the inferior frontal gyrus (IFG)in four seizures (patient 1). Estimation uses the same parameters in the four cases (window duration, 12 s ; sliding step, 0.64 s; frequencyband for averaging, 0–25 Hz). In each recording, seizure onset is marked by the white arrow. In the two first seizures (top), humming isobserved and occurs at time t = 115 s (seizure 1) and t = 120 s (seizure 2). A significant coherence increase is measured in both cases.In the two other seizures (down), no humming sign was observed. Correspondingly, no significant modification of coherence wasmeasured on signals from the same structures.
NEURAL NETWORKS AND HUMMING 1009
Epilepsia, Vol. 43, No. 9, 2002

ity in subjects listening to Mozart. In another study,variations occurring within the same frequency band-width were observed in scalp EEG activity recordedfrom temporal and frontal electrodes (29).
The prominent role of the STG in the perception ofmusic has similarly been described in numerous studies(35,38,40–42). Whereas the posterior STG in both hemi-spheres has been shown to play a role in music percep-tion, it is unclear whether the same (bilateral) structuresare implicated in humming and singing. One case ofmotor amusia consecutive to a right temporal lobe hem-orrhage involving the Heschl’s gyrus was reported in amusic teacher, who complained of having difficulty sing-ing and playing musical instruments after the cerebralattack (43). Another study (44) suggested that, whereasthe left hemisphere is primarily implicated in motor con-trol and articulation during singing, the right hemisphereplays a major role in nonverbal types of musical expres-sion such as humming. Data from several positron emis-sion tomography (PET) studies (41,45) also suggest arole of the frontal lobe in the perception of music. Ac-tivation of right-hemispheric frontal regions is observedonly after completion of a cognitive task, whereas pas-sive listening appears to activate the STG in both hemi-spheres (although to a greater degree in the righthemisphere). The same study (46) suggests that whereasperceptual processing is carried out in the primary audi-tory cortex, auditory working memory involves tempo-ro–frontal interactions. Left inferior frontal regions havebeen found to be activated during familiarity tasks (46)and song perception (47). Several PET studies have re-
ported left inferior frontal activations during tasks requir-ing a semantic response (48–50). In a recent study, Perryet al. (51) reported cerebral activity in frontal areas andin right Heschl’s gyrus during singing. In sum, abundantdata suggest that left and right inferior frontal regions, aswell as the STG, are important in humming.
The participation of the frontal lobe in the perceptionand expression of music is likely dependent on fibersconnecting the prefrontal cortex to superior temporal au-ditory areas. Such connective fibers have been describedin primates in numerous studies (52–55). In a recentanatomic study, authors reported the existence of fibersprojecting from anterior and posterior auditory regions torostral prefrontal and dorsolateral prefrontal/premotor re-gions, respectively (56). These connections seem to behierarchically organized, because few projections enterthe frontal lobe from the lateral and medial auditory belts(i.e., from primary and secondary auditory areas). Mostprojections originate from parabelt regions and anteriorregions of the temporal lobe (i.e., associative regions)(56,57). In humans, activation of the IFG with a prefron-tal stream from rostral/parabelt auditory regions to ros-tral/orbital prefrontal areas is a consistent finding infunctional MRI studies on semantic encoding and audi-tory attention (58,59).
In conclusion, our findings suggest that the occurrenceof humming during epileptic seizures of the temporallobe is associated with activity in the STG and the IFG,which are related within a neural network. This “hum-ming” network is separate from the network implicatedin the epileptogenic zone. These findings are consistent
FIG. 7. Time–frequency distribution of the SEEG signal recorded during the ictal discharge involving the superior temporal gyrus in twopatients (a and b) disclosing middle-range frequency during humming. In both cases, a rupture (onset indicated by the arrow) is observedin the evolution of the spectral content of the signal: a new narrow-band component (dominant frequency ∼15 Hz in a, dominant frequency∼6 Hz in b) appears in the signal and holds for the whole duration of humming.
F. BARTOLOMEI ET AL.1010
Epilepsia, Vol. 43, No. 9, 2002

with behavioral evidence implicating temporal and pre-frontal regions in music perception and anatomic evi-dence showing the existence of connective pathwaysbetween the prefrontal lobe and the STG.
Acknowledgment: We thank P. J. M. Scarabin for stereo-taxic placement of the electrodes and Dr. Jean Regis for hishelp in illustration (3D rendering of the brain MRI). We thankDr. C. Allaire for her help in the clinical evaluation of thepatients in the study and Kimberley Giraud for the revision ofthe English version of this article and useful comments. Wealso thank Helene Leroy for the musical score transcription ofhumming.
REFERENCES
1. Bancaud J, Brunet-Bourgin F, Chauvel P, et al. Anatomical originof deja vu and vivid “memories” in human temporal lobe epilepsy.Brain 1994;117:71–90.
2. Gloor P. Experiential phenomena of temporal lobe epilepsy: factsand hypotheses. Brain 1990;113:1673–94.
3. Chauvel P, Delgado-Escueta A. Frontal lobe seizures and epilep-sies. Adv Neurol 1992;57:750.
4. Wieser H. Ictal manifestations of temporal lobe epilepsy. Adv Neu-rol 1991;55:301–15.
5. Penfield W, Perrot P. The brains record of auditory and visualexperience: a final summary and discussion. Brain 1963;86:595–696.
6. Gabr M, Luders H, Dinner D, et al. Speech manifestations inlateralization of temporal lobe seizures. Ann Neurol 1989;25:82–7.
7. Serafetinides E, Falconer M. Speech disturbances in temporal lobeseizures: a study in 100 epileptic patients submitted to anteriortemporal lobectomy. Brain 1963;86:333–46.
8. Theodore W, Porter R, Penry J. Complex partial seizures: clinicalcharacteristics and differential diagnosis. Neurology 1983;33:1115–21.
9. Fish D, Gloor P, Quesney F, et al. Clinical response to electricalbrain stimulation of the temporal and frontal lobes in patients withepilepsy. Brain 1993;116:397–414.
10. Meierkord H, Shorvon S. Variation of a theme: singing as anepileptic automatism. J Neurol Neurosurg Psychiatry 1991;54:1114–6.
11. Vidailhet M, Serdaru M, Agid Y. Singing in the brain: a new formof complex partial seizure? J Neurol Neurosurg Psychiatry 1989;52:1306.
12. Tan E, Ciger A, Zileli T. Whistling epilepsy: a case report. ClinElectroencephal 1990;21:110–11.
13. Wieser H. Temporal lobe or psychomotor status epilepticus. Elec-troencephalogr Clin Neurophysiol 1980;48:558–72.
14. Engel A, Konig P, Kreiter A, et al. Interhemispheric synchroniza-tion of oscillatory neuronal responses in cat visual cortex. Science1991;252:1177–9.
15. Laurent G, Wehr M, Davidovitz H. Temporal representation ofodours in an olfactory network. J Neurosci 1996;15:3837–47.
16. Sarthien J, Petsche H, Rappelsberger P, et al. Synchronisationbetween prefrontal and posterior association cortex during humanworking memory. Proc Natl Acad Sci U S A 1998;95:7092–5.
17. Sarnthein J, vonStein A, Rappelsberger P, et al. Persistent patternsof brain activity: an EEG coherence study of the positive effect ofmusic on spatial-temporal reasoning. Neurol Res 1997;19:107–16.
18. Chauvel P, Vignal J, Biraben A, et al. Stereoelectroencephalogra-phy. In: Pawlik G, Stefan H, eds. Multimethodological assessmentof the epileptic forms. New York: Springer Verlag, 1996:270.
19. Bancaud J, Talairach J, Bonis A, et al. La stereoelectroencepha-lographie dans l’epilepsie: informations neurophysiopatho-logiques apportees par l’investigation fonctionnelle stereotaxique.Paris: Masson, 1965.
20. Talairach J, Bancaud J, Szickla G, et al. Approche nouvelle de lachirurgie de l’epilepsie: methodologie stereotaxique et resultatstherapeutiques. Neurochirurgie 1974;20(supp 1):1–240.
21. Cohen L. Time frequency analysis. Englewood Cliffs, NJ:Prentice-Hall, 1995.
22. Wendling F, Shamsollahi M, Badier J, et al. Time-frequencymatching of warped depth-EEG seizure observations. IEE TransBiomed Eng 1999;46:601–5.
23. Zaveri H, Williams W, Sackellares J, et al. Measuring the coher-ence of intracranial electroencephalograms. Clin Neurophysiol1999;110:1717–25.
24. Wendling F, Le Bouquin Jeannes R, Bellanger J, et al. Extractionde signatures epileptiques et mesure de coherence dans des obser-vations SEEG: etude preliminaire. Innovation Technol Biol Med1998;19:59–75.
25. Bartolomei F, Wendling F, Vignal J, et al. Temporal lobe epilep-sies: identification of subtypes using coherence analysis in stereo-electroencephalography. Clin Neurophysiol 1999;110:1741–54.
26. Shibata R. Selection of the order of an autoregressive model byAkaike’s information criterion. Biometrika 1976;63:117–26.
27. Brandt S. Statistical and computational methods in data analysis.Amsterdam, North Holland, 1983.
28. Franaszczuk P, Bergey G, Durka P, et al. Time-frequency analysisusing the matching pursuit algorithm applied to seizures originat-ing from the mesial temporal lobe. Electroencephalogr Clin Neu-rophysiol 1998;106:513–21.
29. Auzou P, Eustache F, Etevenon P, et al. Topographic EEG acti-vations during timbre and pitch discrimination tasks using musicalsounds. Neuropsychologia 1995;33:25–37.
30. Galaburda A, Sanides F. Cytoarchitectonic organization of the hu-man auditory cortex. J Comp Neurol 1980;190:597–610.
31. Braak H. Architectonics of the human telencephalic cortex. Berlin:Springer Verlag, 1978.
32. Brodmann K. Vergleichende Lokalisationslehre der Grosshirn-rinde in ihren Prinzipien dargestelld auf Grund des Zellenbaues.Leipzig: Barth, 1909.
33. Liegois-Chauvel C, Musolino A, Chauvel P. Localization of theprimary auditory area in man. Brain 1991;114:139–53.
34. Liegeois-Chauvel C, Musolino A, Badier J, et al. Evoked-potentials recorded from the auditory cortex in man: evaluation andtopography of the middle latency components. ElectroencephalogrClin Neurophysiol 1994;92:204–14.
35. Liegeois-Chauvel C, Peretz I, Babaı M, et al. Contribution of dif-ferent cortical areas in the temporal lobes to music processing.Brain 1998;121:1853–67.
36. Duckrow R, Spencer S. Regional coherence and the transfer ofictal activity during seizure onset in the medial temporal lobe.Electroencephalogr Clin Neurophysiol 1992;82:415–22.
37. Gotman J, Levtova V. Amygdala-hippocampus relationships intemporal lobe seizures: a phase coherence study. Epilepsy Res1996;25:51–7.
38. Peretz I. Processing of local and global musical information byunilateral brain-damaged patients. Brain 1990;113:1185–205.
39. Peretz I, Kolinsky R, Tramo M, et al. Functional dissociationsfollowing bilateral lesions of auditory cortex. Brain 1994;117:1283–301.
40. Samson S, Zatorre R. Contribution of the right temporal lobe tomusical timbre discrimination. Neuropsychologia 1994;32:231–40.
41. Zatorre R, Evans A, Meyer E. Neural mechanisms underlyingmelodic perception and memory for pitch. J Neurosci 1994;14:1908–19.
42. Zatorre R. Functional specialization of human auditory cortex inmusical processing [editorial]. Brain 1998;121:1817–8.
43. Takeda K, Bandou M, Nishimura Y. Motor amusia following aright temporal lobe hemorrhage: a case report. Rinsho Shinkeigaku1990;30:78–83.
44. Cadalbert A, Landis T, Regard M, et al. Singing with and withoutwords: hemispheric asymmetries in motor control. J Clin Exp Neu-ropsychol 1994;16:664–70.
45. Zatorre R, Perry D, Beckett C, et al. Functional anatomy of musicalprocessing in listeners with absolute pitch and relative pitch. ProcNatl Acad Sci U S A 1998;95:3172–7.
46. Platel H, Price C, Baron J, et al. The structural components ofmusic perception: a functional anatomical study. Brain 1997;120:229–43.
NEURAL NETWORKS AND HUMMING 1011
Epilepsia, Vol. 43, No. 9, 2002

47. Zatorre R, Halpern A, Perry D, et al. Hearing in the mind’s ear:a PET investigation of musical imagery and perception. J CognNeurosci 1996;8:29–46.
48. Petersen S, Fox P, Posner M, et al. Positron emission tomographicstudies of the cortical anatomy of processing of single words.Nature 1988;331:585–9.
49. Petersen S, Fox P, Posner M, et al. Positron emission tomographicstudies of the processing of single words. J Cogn Neurosci 1989;1:153–70.
50. Raichle M, Fiez J, Vadeen T, et al. Pratice-related changes inhuman brain functional anatomy during nonmotor learning. CerebCortex 1994;4:8–26.
51. Perry D, Zatorre R, Petrides M, et al. Localization of cerebralactivity during simple singing. Neuroreport 1999;10:3979–84.
52. Barbas H. Architecture and cortical connection of the prefrontalcortex in the rhesus monkey [Review]. Adv Neurol 1992;57:91–115.
53. Barbas H. Organisation of cortical afferent input to orbitofrontalareas in the rhesus monkey. Neuroscience 1993;56:841–64.
54. Pandya D, Hallett M, Kmukherjee S. Intra and interhemisphericconnections of the neocortical auditory system in the rhesus mon-key. Brain Res 1969;14:49–65.
55. Petrides M, Pandya D. Association fiber pathways to the frontalcortex from the superior temporal region in the rhesus monkey.J Comp Neurol 1988;273:52–66.
56. Hackett T, Stepniewska I, Kaas J. Prefrontal connections of theparabelt auditory cortex in macaque monkeys. Brain Res 1999;817:45–58.
57. Romanski L, Bates J, Goldman-Rakic PS. Auditory belt and para-belt projections to the prefrontal cortex in the rhesus monkey.J Comp Neurol 1999;403:141–57.
58. Gabrieli J, Poldrack R, Desmond J. The role of left prefrontalcortex in language and memory. Proc Natl Acad Sci U S A 1998;95:906–13.
59. Braver T, Cohen J, Nystrom L, et al. A parametric study of pre-frontal cortex involvment in human working memory. Neuroimage1997;5:49–62.
F. BARTOLOMEI ET AL.1012
Epilepsia, Vol. 43, No. 9, 2002




















