
Relationality in Alice Munro, Joan Riviere, Jane Campion and ...

edited by
Ofer Bar-Yosef&
François R. Valla
InternatIonal Monographs In prehIstory
Archaeological Series 19
Natufian Foragers in the Levant
Terminal Pleistocene Social Changes in Western Asia

Library of Congress Cataloging-in-Publication Data
Natufian foragers in the Levant : terminal Pleistocene social changes in Western Asia / edited by Ofer Bar-Yosef & François Valla. pages cm. -- (Archaeological series / International Monographs in Prehistory ; 19) Papers from a symposium held in 2009. Includes bibliographical references. ISBN 978-1-879621-45-9 (paperback : acid-free paper) -- ISBN 978-1-879621-46-6 (hard cover : acid-free paper) 1. Natufian culture--Middle East--Congresses. 2. Hunting and gathering societies--Middle East--Congresses. 3. Pleistocene-Holocene boundary--Congresses. 4. Social archaeology--Middle East--Congresses. 5. Social change--Middle East--History--To 1500--Congresses. 6. Excavations (Archaeology)--Middle East--Congresses. 7. Middle East--Antiquities--Congresses. I. Bar-Yosef, Ofer. II. Valla, François Raymond. GN774.3.N38N28 2013 306.3›640956--dc23 2013035516
© 2013 by International Monographs in PrehistoryAll rights reserved
Printed in the United States of AmericaAll rights reserved
Paperback:ISBN 978-1-879621-45-9Hard Cover:ISBN 978-1-879621-46-6
This book is printed on acid-free paper. ∞
International Monographs in PrehistoryAnn Arbor, MichiganU.S.A.
Printed with the support of the American School of Prehistoric Research (Peabody Museum, Harvard University)

Table of Contents
List of Contributors ................................................................................................................... vii
Preface – The Natufian Culture in the Levant: Twenty Years LaterOfer Bar-Yosef and François R. Valla ...............................................................................xv
Acknowledgements ....................................................................................................................xix
Northern Levant
Natufian Lifeways in the Eastern Foothills of the Anti-Lebanon MountainsNicholas J. Conard, Knut Bretzke, Katleen Deckers, Andrew W. Kandel, Mohamed Masri, Hannes Napierala, Simone Riehl and Mareike Stahlschmidt ..............................1
The Natufian of Moghr el-Ahwal in the Qadisha Valley, Northern LebanonAndrew Garrard and Corine Yazbeck ..............................................................................17
The Natufian of Southwestern Syria Sites in the Damascus ProvinceKurt Felix Hillgruber ........................................................................................................28
The Natufian Occupations of Qarassa 3 (Sweida, Southern Syria)Xavier Terradas, Juan José Ibáñez, Franck Braemer, Lionel Gourichon and Luis C. Teira ...................................................................................................................................45
The Early Natufian Site of Jeftelik (Homs Gap, Syria)Amelia del Carmen Rodríguez Rodríguez, Maya Haïdar-Boustani, Jesús E.González Urquijo, Juan José Ibáñez, Michel Al-Maqdissi, Xavier Terradasand Lydia Zapata ..............................................................................................................61
Fish in the Desert? The Younger Dryas and its Influence on the Paleoenvironment at Baaz Rockshelter, Syria
Hannes Napierala .............................................................................................................73
Preliminary Results from Analyses of Charred Plant Remains from a Burnt Natufian Building at Dederiyeh Cave in Northwest Syria
Ken-ichi Tanno, George Willcox, Sultan Muhesen, Yoshihiro Nishiaki, YousefKanjo and Takeru Akazawa..............................................................................................83
Southern Levant
El-Wad
Spatial Organization of Natufian el-Wad through Time: Combining the Results of Past and Present Excavations
Mina Weinstein-Evron, Daniel Kaufman and Reuven Yeshurun ...................................88

iv
The Last Natufian Inhabitants of el-Wad TerraceNoga Bachrach, Israel Hershkovitz, Daniel Kaufman and MinaWeinstein-Evron..............................................................................................................107
Domestic Refuse Maintenance in the Natufian: Faunal Evidence from el-Wad Terrace, Mount Carmel
Reuven Yeshurun, Guy Bar-Oz, Daniel Kaufman and Mina Weinstein-Evron ...........118
Natufian Green Stone Pendants from el-Wad: Characteristics and Cultural ImplicationsDaniella E. Bar-Yosef Mayer, Naomi Porat and Mina Weinstein-Evron ......................139
Eynan
The Final Natufian Structure 215-228 at Mallaha (Eynan), Israel: an Attempt at Spatial Analysis
François R. Valla, Hamoudi Khalaily, Nicolas Samuelian, Anne Bridault, Rivka Rabinovich, Tal Simmons, Gaëlle Le Dosseur and Shoshana Ashkenazi ....................146
A Study of two Natufian Residential Complexes: Structures 200 and 203 at Eynan (Ain Mallaha), Israel
Nicolas Samuelian ..........................................................................................................172
Graves in Context: Field Anthropology and the Investigation of Interstratified Floors and Burials
Fanny Bocquentin, Teresa Cabellos and Nicolas Samuelian ........................................185
Obsidian in Natufian Context: the Case of Eynan (Ain Mallaha), IsraelHamoudi Khalaily and François R. Valla ......................................................................193
Flint Knapping and its Objectives in the Early Natufian. The Example of Eynan- Ain Mallaha (Israel)
Boris Valentin, François R. Valla and Hugues Plisson with the collaboration of Fanny Bocquentin ...........................................................................................................203
Searching for the Functions of Fire Structures in Eynan (Mallaha) and their Formation Processes: a Geochemical Approach
Ramiro J. March ..............................................................................................................227
Avifauna of the Final Natufian of EynanTal Simmons ....................................................................................................................284
Bone Ornamental Elements and Decorated Objects of the Natufian from MallahaGaëlle Le Dosseur and Claudine Maréchal ...................................................................293
Reconstruction of the Habitats in the Ecosystem of the Final Natufian Site of Ain Mallaha (Eynan)
Shoshana Ashkenazi .......................................................................................................312

v
Southern Levant - other sites
Wadi Hammeh 27: an open-air ‘base-camp’ on the Fringe of the Natufian ‘homeland’Phillip C. Edwards, Fanny Bocquentin, Sue Colledge, Yvonne Edwards, Gaëlle Le Dosseur, Louise Martin, Zvonkica Stanin and John Webb ...........................................319
Art Items from Wadi Hammeh 27Janine Major ...................................................................................................................349
The Final Epipaleolithic / PPNA site of Huzuq Musa (Jordan Valley)Dani Nadel and Danny Rosenberg .................................................................................382
Natufian Settlement in the Wadi al-Qusayr, West-Central JordanMichael Neeley ................................................................................................................397
The Steppic Early Natufian: Investigations in the Wadi al-Hasa, JordanDeborah I. Olszewski ......................................................................................................412
The Natufian of the Azraq Basin: An AppraisalTobias Richter and Lisa A. Maher ..................................................................................429
Chert Procurement Patterns And Exploitation Territory: Case Study From Late Natufian Hayonim Terrace (Western Galilee, Israel)
Christophe Delage ...........................................................................................................449
A Faunal Perspective on the Relationship between the Natufian Occupations of Hayonim Cave and Hayonim Terrace
Natalie D. Munro ............................................................................................................463
The Natufian at Raqefet CaveGyörgy Lengyel, Dani Nadel and Fanny Bocquentin ....................................................478
Hof Shahaf: A New Natufian Site on the Shore of Lake KinneretOfer Marder, Reuven Yeshurun, Howard Smithline, Oren Ackermann, Daniella E. Bar-Yosef Mayer, Anna Belfer-Cohen, Leore Grosman, Israel Hershkovitz, Noa Klein and Lior Weissbrod ...............................................................................................505
The Life History of Macrolithic Tools at Hilazon Tachtit CaveLaure Dubreuil and Leore Grosman ..............................................................................527
General Reviews, Climate and Interpretations
Breaking the Mould: Phases and Facies in the Natufian of the Mediterranean ZoneAnna Belfer-Cohen and A. Nigel Goring-Morris ...........................................................544
Ruminations on the Role of Periphery and Center in the NatufianA. Nigel Goring-Morris and Anna Belfer-Cohen ...........................................................562

vi
The Natufian and the Younger DryasDonald O. Henry .............................................................................................................584
Scaphopod Shells in the Natufian CultureAldona Kurzawska, Daniella E. Bar-Yosef Mayer and Henk K. Mienis ......................611
The Natufian Chronological Scheme – New Insights and their ImplicationsLeore Grosman ................................................................................................................622
Natufian Foragers and the ‘Monocot Revolution’: A Phytolith PerspectiveArlene M. Rosen ..............................................................................................................638
Lithic Technology in the Late Natufian – Technological Differences between ‘Core-area’ and ‘Periphery’
Hila Ashkenazy ...............................................................................................................649
Variability of Lunates and Changes in Projectile Weapons Technology during the NatufianAlla Yaroshevich, Daniel Kaufman, Dmitri Nuzhnyy, Ofer Bar-Yosef and Mina Weinstein-Evron..............................................................................................................671
Specialized Hunting of Gazelle in the Natufian: Cultural Cause or Climatic Effect?Guy Bar-Oz, Reuven Yeshurun and Mina Weinstein-Evron .........................................685
Commensalism: was it Truly a Natufian Phenomenon? Recent Cntributions from Ethnoarchaeology and Ecology
Lior Weissbrod, Daniel Kaufman, Dani Nadel, Reuven Yeshurun and Mina Weinstein-Evron..............................................................................................................699

463
Introduction
Hayonim Cave and Hayonim Terrace are well-known Natufian occupations in the Lower Galilee of the southern Levant (Fig. 1; Bar-Yosef 1991; Belfer-Cohen 1988; Valla et al. 1989, 1991). The sites are situated very close together—the terrace is only 20 meters from the cave’s mouth. Nevertheless, because of a gap in stratigraphy, the relationship between the two sites and their contemporaneity remains unknown. Hayonim Cave and Terrace were excavated independently by two research teams and thus, are most often treated separately in published reports. Nevertheless, the proximity of Hayonim Cave and Terrace suggests that they should be considered as two parts of the same site. Here I use faunal data to test two hypotheses regarding the relationship between Hayonim Cave and Terrace. These include (1) the two areas are part of the same site and potentially served different functions (i.e., the cave was used more for special activities and the terrace was used more as a multi-purpose habitation area (Bar-Yosef and Belfer-Cohen 2012; Boyd 1992); and (2) the two areas represent separate, non-contemporaenous occupations. Hayonim Cave is a karstic cave that initially contained four chambers, although two have since collapsed (Bar-Yosef et al. 2005). The Natufian occu-pation is situated in the easternmost chamber atop a long prehistoric sequence of human occupation that extends from the Acheuleo-Yabrudian to the Late Natufian phase (Bar-Yosef et al. 2005). Exca-vations of the Natufian layer were directed by Ofer Bar-Yosef in the late 1960s and 1970s. Bar-Yosef and a multi-disciplinary international team re-visited the Natufian layer in the 1990s as part of a larger project focused on the Middle Paleolithic occupation in the cave. The Natufian occupation at Hayonim Cave includes both Early and Late Natufian phases. The Early Natufian component is home to significant building activity that include the manufacture of multiple structures, built hearths,
A Faunal Perspective on the Relationship between the Natufian
Occupations of Hayonim Cave and Hayonim Terrace
Natalie D. Munro
slab-lined pits and graves. The construction of built features largely ceased during the Late Natufian, although human burial activities continued in this phase (Bar-Yosef 1991). The recovery of several caches containing basalt pestles, quartzite ham-mer stones, dentalia beads, gazelle horn cores, and partially worked bovid ribs along the northeastern wall of the cave suggest that the cave continued to serve as a locus for other activities during the
Fig. 1. Map showing the locations of Hayonim Cave and Hayonim Terrace in the Lower Galilee of Israel.

464
Natalie D. Munro
Late Natufian (Belfer-Cohen 1991; Bar-Yosef and Belfer-Cohen 2012). Hayonim Terrace is situated on the uppermost of four terraces directly outside the mouth of the cave (Valla et al. 1991). The site contains a variety of features, including two structures, one living floor, one possible storage pit, a garbage pit, and 6 graves including 9 human burials (Valla et al. 1991). Two large mortars, one still standing in situ were also recovered during excavation. The density of material remains, in particular lithics, is higher than in the cave (Bar-Yosef and Belfer-Cohen 2012). The burials of Homo 7, 8 and 10 are of particular interest due to their association with two canid interments that have been interpreted as early evidence for dog domestication (Tchernov and Valla 1997). Earlier excavations at Hayonim Terrace by Donald Henry in the 1970s recovered material dating to the Early Natufian and Geometric Kebaran periods (Henry et al. 1981; Henry and Leroi-Gourhan 1976). The faunal assemblage from this occupation was not analyzed by the author and thus this discussion focuses exclusively on the Late Natufian occupations of the cave and terrace. Here I test hypotheses about the relationship between Hayonim Cave and Terrace by comparing results from numerous taphonomic and economic analyses and by considering potential “social markers” deriving from the Late Natufian faunal assemblages. The assemblages are especially well suited for comparison because comprehensive taphonomic and economic analyses of both were undertaken by the author using the same collec-tion and recording procedures. Moreover, the size of the assemblages is sufficiently large (Hayonim Cave Late Natufian NISP 5,899; Hayonim Terrace Late Natufian NISP = 10,425) to examine not only general economic trends through analyses such as relative taxonomic abundance, but more specialized analyses such as mortality profiles skeletal-part representation, carcass preparation, and the spatial distribution of various taxa. The analyses are designed to identify common-alities and differences between the assemblages. Similarity in prey procurement and preparation techniques is expected if the two areas were occu-pied by the same human group, although functional differences in the use of cave and terrace could result in notable differences in the disposal or use of certain body-parts. In particular, butchery and food preparation techniques are two of the best indicators of group identity or affiliation as these practices represent conservative aspects of human behavior (Crabtree 1990; Hesse 1986). Members of
the same group tend to butcher, and cook animals in similar ways. Finally the representation of animal body-parts that may serve as social markers is also expected to be similar if the areas were occupied by the same group (see below).
Quality of Preservation
Before past human behaviors can be recon-structed from the Hayonim Cave (HAYC) and Terrace (HAYT) assemblages, the impact of ta-phonomic processes and the resulting quality of preservation of the assemblages must be assessed. This is of particular importance, since despite the proximity of the sites, the assemblages derive from very different depositional environments. It must be established whether differences in the assem-blages result from human behavior or taphonomic history. The cave provides a protective setting that is reflected in the very good preservation of bone including very fine tissues such as nasal and foetal bone. In contrast, the terrace assemblage derives from an open-air environment, and was undoubtedly subjected to more water percolation than the cave. This is indicated by the adherence of concretions to nearly all bone surfaces. The concretions are made of precipitated calcium carbonates that combined with sediment to form a thin crust on the bone surfaces. Following excavation, most of the bones from the terrace were soaked in a light acetic acid bath to remove the concretions—but this treatment was not always successful. To control for the pres-ence of concretions when calculating frequencies of surface damage at Hayonim Terrace, the percent of the surface obscured by concretions was recorded for each bone specimen. Several analyses were undertaken to compare the quality of bone preservation in the cave and on the terrace (Table 1). Most of these capitalize on differences in bone density, and assume that as preservation worsens, the bias toward more resis-tant, high-density bone portions will become more pronounced (Lyman 1984, 1993, 1994). The analyses include: (1) regression of gazelle bone survivorship against bone density (Lyman 1994); (2) relative representation of proximal (low-density) and distal (high-density) ends of gazelle humerus and tibia; (3) gazelle bone- (low density) versus tooth-based MNI (high density) of the cranium (Stiner 1994); and (4) %completeness of dense compact bones (astragalus and third phalanx) that do not contain in-bone nutrients and are not expected to be broken during butchery (Marean 1991). Breakage of these compact bones is most likely caused by postdepo-

465
A Faunal Perspective on…Hayonim Cave and Hayonim Terrace
sitional processes. It is expected that under ideal conditions of preservation there will be no relation-ship between bone survivorship and bone density; the representation of low- and high-density bones will be equal; and %completeness of non-nutritive elements will be 100%. Table 1 indicates a significant density-medi-ated bias in both the Hayonim Cave and Terrace gazelle assemblages. In both cases low-density bone portions are underrepresented in comparison to high-density portions. Four of the remaining five indices show that this bias is more pronounced on the terrace than in the cave: the ratio of low to high-density bone specimens is greater on the ter-race than in the cave; and the dense astragali and third phalanges are less complete. The fifth index, however, shows the opposite pattern. Lower-density cranial bones are better preserved in relation to teeth on the terrace than in the cave. The results for this index indicate small differences between tooth- and bone-based MNIs at both sites (HAYT =.964; HAYC=.818); and variability is insignificant and may be related to small sample sizes (Appendix 2). Agreement in the other four indices point to a more pronounced density-mediated bias in the Hayonim Terrace assemblage. This bias must be considered when comparing economic data from the two areas, particularly when skeletal-part representation favors higher density elements on the terrace than in the cave.
Economic Strategies
The economic strategies employed by the inhabitants of Hayonim Cave and Terrace are compared to determine the likelihood that these assemblages were created by the same human population. Here, the term economic is used to re-fer to those activities that are aimed at procuring or preparing food—in particular human hunting practices, carcass transport and processing, and food preparation. Numerous analyses were undertaken, but only the most informative are presented here.
These include: (a) relative taxonomic abundance indices (broad taxonomic groups, ungulates and small game); (b) gazelle body-part representation; (c) gazelle mortality profiles; and (d) small game preparation techniques including tortoise butchery and hare burning patterns. The array of animal taxa represented at Hayonim Cave and Hayonim Terrace are nearly identical, although Hayonim Cave has a much richer assemblage of avian species (Appendix 1). Only taxa demonstrably captured and utilized by humans are investigated here (see Munro 2001 for details). Like all Natufian assemblages in the Mediterranean zone, the Hayonim Cave and Terrace ungulates are dominated by the mountain gazelle (Gazella gazella), although several other species—fallow deer (Dama mesopotamica); red deer (Cervus elaphus); wild boar (Sus scrofa); wild goat (Capra aegagrus) and wild cattle (Bos primigenius) play secondary roles. Several small carnivore species are represented at both sites, most importantly, the red fox (Vulpes vulpes); the jungle cat (Felis chaus) and a few mustelid species (Martes foina; Vormela peregusna; Meles meles). The most common small game taxa include the Mediterranean spur-thighed tortoise (Testudo graeca); the cape hare (Lepus cap-ensis) and the chukar partridge (Alectoris chukar). A number of other bird taxa are also present—those that were most likely captured by humans include several species of waterfowl (i.e., Anas sp, Anser sp, Fulica atra) and Falconiformes (i.e., Buteo buteo; Aquila chrysaetos; Appendix 1). The taxa are first, divided into three broad groups determined by biological relatedness (un-gulates, carnivores) and similarities in physical and ecological characteristics, namely body size (small game). Significant differences in the repre-sentation (proportion of NISP) of the three groups are observable in the two assemblages (Fig. 2a). Although carnivores make up similar proportions of the assemblages (5.6-5.8%), there is a disparity in the representation of ungulates and small game. Un-gulates are better represented at Hayonim Terrace
Table 1. Measures of in situ attrition at Hayonim Cave (HAYC LN) and Hayonim Terrace (HAYT LN). All values refer to the gazelle assemblage only. See Appendix 2 for sample sizes (MNE)
IN SITU ATTRITION SUMMARY HAYC LN HAYT LNCorrelation between bone density and MNE 0.308 0.228Bone:Tooth-Based MNI 0.818 0.964% Complete Astragalus (MNE) 0.840 0.720% Complete Third Phalanx (MNE) 0.771 0.434Proximal Humerus: Distal Humerus (MNE) 0.139 0.072Proximal Tibia: Distal Tibia (MNE) 0.800 0.231

466
Natalie D. Munro
(54.7%; HAYC = 29.4%) while small game are more common in the cave (64.8%; HAYT = 38.7%). The difference between the small game and ungulate fractions at the two sites is about the same (25%). A closer look at the relative abundance of gazelle and tortoise, the most common taxa in these two groups, shows that these two taxa are driving this pattern—gazelle are far more abundant on the terrace (63.3%; HAYC = 36.4%) while tortoise are more abundant in the cave (63.6%; HAYT =36.7%)(Fig. 2b).
The relative representation of small, medium and large ungulates are similar in the two assem-blages (Fig. 3a). In both cases, the ungulate fraction is dominated by the small ungulates (HAYC = 90.7%; HAYT = 92.0%), which are composed nearly exclusively of gazelle. Medium and large ungulates are rare and together compose less than 10% of the ungulate fraction in each assemblage. Although sample sizes of medium and large ungulates are small (HAYT = 348; HAYC = 88), there are dif-ferences in their representation at the two sites. In particular, wild cattle are proportionally more abundant on the terrace, while wild boar and red
deer are more abundant in the cave (Fig. 3b). The presence of two concentrations of cattle remains on the Terrace (in a probable trash pit; feature 7 –square N34; and in fire pit 8—squares K/L 35) is likely driving this pattern.
The tortoise makes up a similar proportion of the small game fraction at the two sites, despite differences in its overall abundance in each as-semblage (Fig. 4). Differences are visible in the relative proportion of hares and birds; birds are rare at Hayonim Terrace (7.3% of small game assemblage) compared to the cave (16.1% of small game assemblage). Given their fragile interiors and thin bone structure, the poor representation of birds at Hayonim Terrace is likely related to taphonomic processes at the site. Concretions can
Fig. 2a. Relative abundance of broad taxonomic groups (ungulates, carnivores and small game) at Hayonim Cave and Hayonim Terrace.
Fig. 2b. Relative abundance of gazelle and tortoises out of total Hayonim Cave and Hayonim Terrace assemblages. See Appendix 1 for NISP values.
Fig. 3a. Relative abundance of large, medium and small-bodied ungulates at Hayonim Cave and Hayonim Terrace. Includes both animals iden-tified to genus and body-size categories. See Appendix 1 for NISP values.
Fig. 3b. Relative abundance of ungulate species at Hayonim Cave and Hayonim Terrace. Includes only those specimens assigned to genus and species. See Appendix 1 for NISP values.

467
A Faunal Perspective on…Hayonim Cave and Hayonim Terrace
be particularly harmful to thin-walled bones—once they fill a bone’s hollow interior, the pressure of the concretions can cause long bone shafts to explode.
Gazelle mortality profiles were calculated based on bone fusion (Davis 1980; Table 2, Fig. 5) and tooth wear and eruption data (Munro et al. 2009; Fig. 6). The fusion data indicate that prior to 12 months of age gazelle survivorship was higher at Hayonim Terrace (78.1-98.7%) than in the cave (57.0-80.0%). This pattern evens out once gazelles reach 13 months of age—close to 75% of gazelles in both assemblages survived to this age. These results indicate that there are fewer very young gazelle (<12 months) in the terrace assemblage than in the cave, although the number of older juvenile gazelle (12-18 months) in the two assemblages is equal. This may reflect differential human behavior or a taphonomic bias that more severely impacted very young bone tissue on the terrace. As they grow, bones become increasingly more mineralized and increase in density. Young bone tissue is very
porous and thus more subject to postdepositional destruction than the older juvenile and adult bone tissue. The under-representation of young gazelle on the terrace may thus result from increased destruction of young bone tissue due to the more aggressive postdepositional processes indicated previously. Once the bones become more mineral-ized, the difference between the sites disappears.
Like the fusion results the gazelle tooth wear data indicate slightly fewer juveniles (0-18 mos) in the Hayonim Terrace (39%) than the Hayonim Cave assemblage (50%). Adults (>18 months) represent 50% of the Hayonim Cave and 61% of the Hayonim Terrace gazelles (Fig. 6). Despite this difference, the representation of juveniles is higher than would be expected in a natural population at both sites, suggesting similar acquisition strategies for the two assemblages. The gazelle body-part profiles from Hayonim Cave and Terrace are also similar (Fig. 7). The pattern at both sites is typical of density-mediated assemblages: the axial skeleton is under-repre-sented while other anatomical regions are more even. In both assemblages, the head is not as well represented as the appendicular skeleton. The head
Fig. 4. Relative abundance of common small game groups (tortoises, hares and partridges) at Hay-onim Cave and Hayonim Terrace. See Appendix 1 for NISP values.
Table 2. Gazelle bone fusion data for Hayonim Terrace and Hayonim Cave. Data used in survivorship curve (Fig. 5)
Age of Fusion HAYT LN HAYC LNEpiphysis in Months MNE %Fused MNE %FusedFirst Phalanx - Proximal 6.5 186 83.3 72 56.9Tibia - Distal 9 41 78.1 11 72.7Calcaneum - Distal 12 75 98.7 15 80.0Femur - Proximal 13 60 68.3 8 75.0Metapodial - Distal 14 10 59.8 64 59.4
Fig. 5. Gazelle survivorship curves for Hayonim Cave and Hayonim Terrace indicating the per-centage of gazelle that survived to a given age. Survivorship rates are based on the proportion of fused epiphyses of elements that fuse at the ages indicated on the X-axis. See data in Table 2.

468
Natalie D. Munro
is better represented in the cave than the terrace, due to the presence of a cache of male horn cores (MNE >18) deposited during the Late Natufian occupation (Belfer-Cohen 1991; Munro 2001). Over-all, the body part pattern suggests that residents of both the cave and terrace transported complete gazelle carcasses, with the exception in many cases, of the heads (Munro 2001).
Patterned breakage of tortoise plastrons at Hayonim Cave and Terrace indicates that the inhab-itants repeatedly used the same technique to open tortoises and access their meat prior to consumption (Fig. 8). At both sites articulated anterior portions of the plastron have been recovered. The posterior end of these portions is characterized by a spiral break that crosscuts three plastron segments (the right and left hypoplastron and the entoplastron) and must have occurred when the bone was still fresh. A similar pattern has also been found at the nearby Late Natufian site at Hilazon Tachtit (Grosman and Munro 2007; Grosman et al. 2008) where the recovery of a number of complete tortoise shells enabled reconstruction of the butchering sequence. Repeated breaks across the segments that form the bridge between the carapace and the plastron indicate that the remaining plastron was pulled away from the carapace following the removal of the anterior plastron. This method of butchery would have exposed the tortoise’s flesh with the secondary benefit of leaving the carapace intact. This is of special relevance at Hayonim Cave where many tortoise carapaces were ground around the edges potentially for their use as vessels. The similarity in the sequence used to butcher tortoises at different sites, points to commonality in tortoise preparation techniques despite differential use of the tortoise shells at the cave and terrace (see below). Likewise, intensive patterns of burning on hare bones from both sites points to commonalities in food preparation techniques. In both assemblages, hare bones are burned in significantly higher pro-
Fig. 6. Proportion of adult and juvenile gazelles at Hayonim Cave (MNI=30) and Hayonim Terrace (MNI=36) based on the state of eruption and wear of the lower deciduous fourth premolar and the lower third molar. Note that because of small sample sizes, the Hayonim Cave Early and Late Natufian assemblages are combined for this analysis.
Fig. 7. Representation of gazelle anatomical units at Hayonim Cave and Hayonim Terrace. Anatomical units defined following Stiner (1994). See Appendix 2 for MNE values of individual gazelle elements.

469
A Faunal Perspective on…Hayonim Cave and Hayonim Terrace
portions than any other taxon (38.1% at Hayonim Terrace, mean=24.8%; 27.1% at Hayonim Cave, mean=18.7%). This non-random burning pattern suggests a shared cooking strategy specific to hares that resulted in high rates of burning on the bones. The economic analyses presented here demon-strate significant similarities between the cave and the terrace assemblages with the exception of: (a) lower proportions of tortoise in relation to gazelle on the terrace; (b) lower proportions of birds in relation to hares on the terrace; and (c) lower proportions of juvenile compared to adult gazelles on the terrace. As previously discussed, the latter two points are most likely related to taphonomic processes that resulted in more severe attrition on the terrace than in the cave. The question now remains, are the differences between the Hayonim Cave and Terrace assemblag-es small enough to justify their consideration as a single assemblage? The answer is certainly yes, given the results presented here, except perhaps, the relative abundance of gazelle and tortoise which differ significantly between the two areas. We must also consider if the degree of variability in the representation of gazelles and tortoises falls within the expected range of intrasite variability. To investigate this question, spatial variability in the distribution of gazelles and tortoises is examined at Hayonim Terrace.
Figs. 9 and 10 map the respective distributions of gazelle and tortoise relative abundance at Hay-onim Terrace. Each square on the map represents a 50x50 cm subsquare from the excavated site. Darker squares indicate higher proportions of gazelle or tortoise than lighter squares. Both maps indicate considerable variation in the distribution of each
taxa across the excavated area. In particular, the abundance of gazelle is much higher in the northern half of the excavation area than in the south. In contrast, tortoises are substantially more abundant in the southern half of the site than they are in the north. The magnitude of the difference in the rep-resentation of these two species springs into focus when the relative abundances for the northern and southern halves of the site are compared (Fig. 11) to those from Hayonim Cave (Fig. 2a and 2b). Gazelles are only 10% more common than tortoises in the northern half of the site (55.0%), while they are 50% more common than tortoises in the southern half of the excavated area (75.0%; Fig. 11). Recall that gazelles are ca. 25.0% less common than tortoises at Hayonim Cave. The difference in the representation of tortoises and gazelle between the northern and the southern halves of the occupation area are thus on an even greater order of magnitude as those between the cave and the terrace. Thus, differences in the proportions of gazelle and tortoises on the cave and terrace fit within the range of variation expected for a single Natufian site. Nevertheless, the proportion of tortoises at Hayonim Cave is consistently greater than at Hayonim Terrace. The difference in the proportions of tortoises may be related to a functional difference in the use of the cave, in particular, to the secondary use of tortoise shells as vessels (see below). In summary, there are extensive similarities between the faunal assemblages from Hayonim Cave and Terrace. The differences that do exist can be explained by taphonomic factors or normal intra-site variability. The similarities in the faunal assemblages suggest that the occupants of these sites were part of the same local population that shared long-term traditions. The evidence convincingly establishes a link between the people inhabiting the two areas at Hay-onim. Nevertheless, Natufian residents throughout the Mediterranean zone shared cultural tradition through population interaction and exchange of knowledge without necessarily occupying the same residential sites. It is possible that given the close affiliation of regional Natufian populations, the faunal assemblages from other Natufian sites in the Mediterranean zone may also fall within the range of variation exhibited by the assemblages at Hayonim Cave and Terrace. If the link between the two sites is to be ascertained, it may be helpful to also consider faunal remains that served as “social markers” (Belfer-Cohen and Goring-Morris herein). Social markers are material remains that reflect social identity (Reitz and Wing 1999) including
Fig. 8. Typical spiral fracture across the anterior tortoise plastrons from Hayonim Terrace (a) and Hayonim Cave (b).

470
Natalie D. Munro
affiliation with a specific group, and thus should be unique to particular assemblages either in their presence/absence or in their abundance. For example, there is substantial variability in the relative representation of decorative items such as ornaments and other material forms of artistic expression among Natufian sites (Belfer-Cohen 1991; Le Dosseur and Maréchal herein). These have been interpreted as markers of group identity in the Natufian context. If these ornaments do rep-resent group identity, we would expect the same social markers to show up in sites occupied by the same group unless these markers are used only in specific contexts (i.e. graves) that are restricted to special locations. One unique feature of the Hayonim Cave fau-na is the presence of numerous modified tortoise carapace segments in both the Early and Late Naufian layers. In the Late Natufian deposit 8.3% (n=1533) of the carapace segments exhibit some kind of anthropogenic modification. Modifications
include use wear in the form of polish and striations in the carapace interior; grinding on the distal edge on pleural segments; pierced nuchal bones; and cut and ground peripheral segments. The study of these carapace segments enabled the reconstruction of the modification process. First, the plastron was broken at its anterior end and then pulled back from the carapace. The peripheral carapace segments with the exception of the nuchal bone were then removed from the neural and pleural segments along their proximal suture. Next, the external edge of the remaining articulated pleural and neural segments was ground, and then the remaining vessel was used for an unknown function that produced polish and multi-directional striations in the shell’s interior. No residues are visible on the carapace bone segments. Interestingly although modified carapace seg-ments are nearly absent from the terrace assem-blage, nine (0.3% of all carapace segments) segments exhibit modifications similar to those seen in the cave. These segments originate from a variety of
Fig. 9. Map indicating spatial distribution of gazelle remains at Hayonim Terrace. Each square represents a 50x50 cm excavation unit. The colors depict variation in the percentage of gazelle specimens out of all taxa recovered from the subsquare—the color becomes increasingly darker as the percentage of tortoise increases. Figure drafted by Austin Hill; site underlay adapted from Valla et al. (1991).

471
A Faunal Perspective on…Hayonim Cave and Hayonim Terrace
contexts within the excavation area. Clearly, there was some movement of modified tortoise segments between the cave and terrace in prehistory. How-ever, it is difficult to ascertain if the low numbers
of modified segments reflect the restricted use of modified shells for specific activities undertaken only in the cave, or rather, a minimal connection (due to non-contemporaneous habitation) between the occupants of the terrace and the cave. The presence of modified carapace segments within all contexts in both the Early and Late Natufian layers at Ha-yonim Cave attests to the longevity of this practice within the cave, although the incidence of striated pieces is significantly higher in Early Natufian contexts (20.0% of carapace fragments; n=1027). Furthermore, no modified tortoise segments have been reported from other Epipaleolithic or Neolithic contexs in the southern Levant, although modified tortoise shells are known from elsewhere in the Middle East (Late Epipaleolithic at Dederiyeh Cave, Syria [Gourichon pers. com. 2010]; Early Neolithic tortoises from Ganj Dareh, Iran [Munro unpublished data]).
Fig. 10. Map indicating spatial distribution of tortoises at Hayonim Terrace. Each square represents a 50x50 cm excavation unit. The colors depict variation in the percentage of tortoise specimens out of all taxa recovered from the subsquare—the color becomes increasingly darker as the percentage of gazelle increases. Figure drafted by Austin Hill; site underlay from Valla et al. (1991).
Fig. 11. The relative abundance of gazelle and tor-toises at Hayonim Cave, the northern (Rows L-N) and southern (Rows I-K) areas of the Hayonim Terrace excavation.
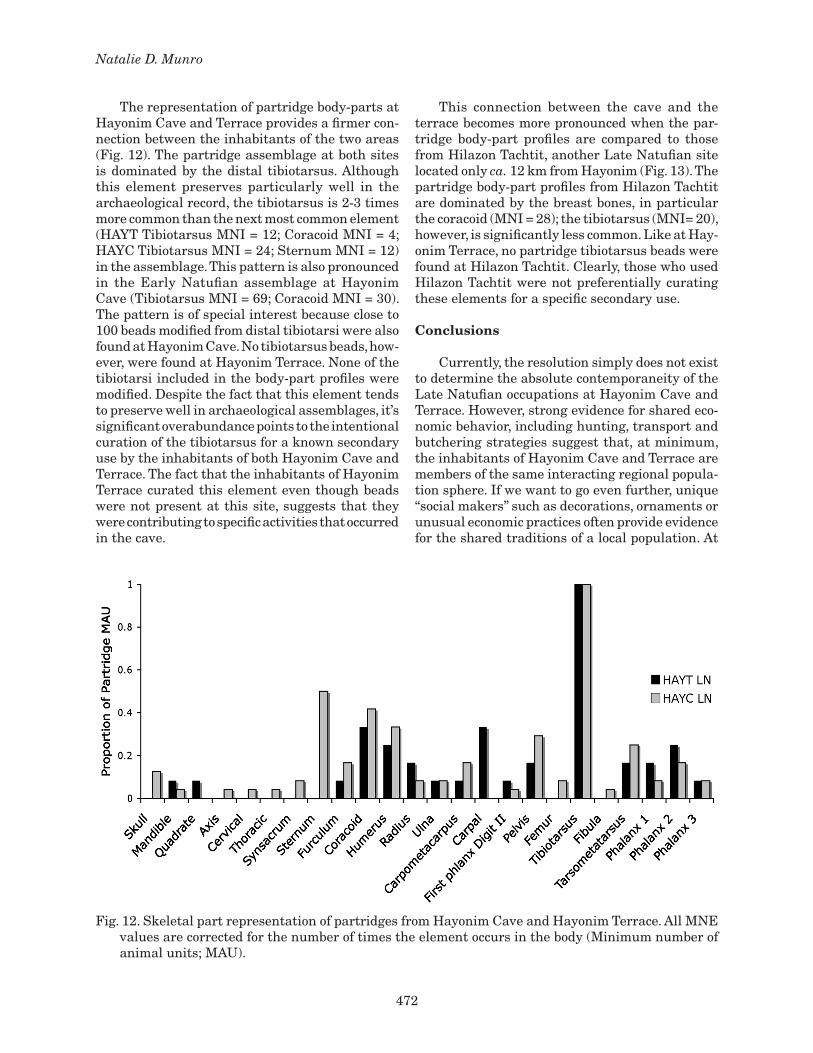
472
Natalie D. Munro
The representation of partridge body-parts at Hayonim Cave and Terrace provides a firmer con-nection between the inhabitants of the two areas (Fig. 12). The partridge assemblage at both sites is dominated by the distal tibiotarsus. Although this element preserves particularly well in the archaeological record, the tibiotarsus is 2-3 times more common than the next most common element (HAYT Tibiotarsus MNI = 12; Coracoid MNI = 4; HAYC Tibiotarsus MNI = 24; Sternum MNI = 12) in the assemblage. This pattern is also pronounced in the Early Natufian assemblage at Hayonim Cave (Tibiotarsus MNI = 69; Coracoid MNI = 30). The pattern is of special interest because close to 100 beads modified from distal tibiotarsi were also found at Hayonim Cave. No tibiotarsus beads, how-ever, were found at Hayonim Terrace. None of the tibiotarsi included in the body-part profiles were modified. Despite the fact that this element tends to preserve well in archaeological assemblages, it’s significant overabundance points to the intentional curation of the tibiotarsus for a known secondary use by the inhabitants of both Hayonim Cave and Terrace. The fact that the inhabitants of Hayonim Terrace curated this element even though beads were not present at this site, suggests that they were contributing to specific activities that occurred in the cave.
This connection between the cave and the terrace becomes more pronounced when the par-tridge body-part profiles are compared to those from Hilazon Tachtit, another Late Natufian site located only ca. 12 km from Hayonim (Fig. 13). The partridge body-part profiles from Hilazon Tachtit are dominated by the breast bones, in particular the coracoid (MNI = 28); the tibiotarsus (MNI= 20), however, is significantly less common. Like at Hay-onim Terrace, no partridge tibiotarsus beads were found at Hilazon Tachtit. Clearly, those who used Hilazon Tachtit were not preferentially curating these elements for a specific secondary use.
Conclusions
Currently, the resolution simply does not exist to determine the absolute contemporaneity of the Late Natufian occupations at Hayonim Cave and Terrace. However, strong evidence for shared eco-nomic behavior, including hunting, transport and butchering strategies suggest that, at minimum, the inhabitants of Hayonim Cave and Terrace are members of the same interacting regional popula-tion sphere. If we want to go even further, unique “social makers” such as decorations, ornaments or unusual economic practices often provide evidence for the shared traditions of a local population. At
Fig. 12. Skeletal part representation of partridges from Hayonim Cave and Hayonim Terrace. All MNE values are corrected for the number of times the element occurs in the body (Minimum number of animal units; MAU).

473
A Faunal Perspective on…Hayonim Cave and Hayonim Terrace
Hayonim Cave and Terrace the presence of high proportions of partridge distal tibitotarsi suggest that the Hayonim area was inhabited by such a local population, but that there was differentiation in the activities performed in and out of the cave.
Acknowledgements:
Thanks first and foremost to Ofer Bar-Yosef and François Valla not only for organizing and inviting me to the very productive second Natufian conference, but for providing permission to study the Hayonim Cave and Hayonim Terrace assem-blages. Rivka Rabinovich generously provided assistance and work space for many years in the Zooarchaeology Lab in the Department of Evolution, Systematics and Ecology at the Hebrew University of Jerusalem. Rivka also oversaw the cleaning of the Hayonim Terrace material. Thanks to Austin (Chad) Hill for investing numerous hours into the preparation of the spatial distribution maps for Hayonim Terrace. Finally, thanks to Ofer Bar-Yosef, Anna Belfer-Cohen, Leore Grosman, Gideon Hart-man, Liora Horwitz, Laura Niven, Mary Stiner and François Valla for fruitful discussions about these assemblages. This research was supported by a National Science Foundation grant (SBS 0618937),
an NSF dissertation improvement grant (SBR-9815083) and a University of Connecticut research foundation large grant.
References Cited
Bar-Yosef, O. 1991 The archaeology of the Natufian layer at
Hayonim Cave. In The Natufian Culture in the Levant, edited by O. Bar-Yosef and F. R. Valla, pp. 81-92. International Monographs in Prehistory, Ann Arbor.
Bar-Yosef, O and A. Belfer-Cohen2012 The Natufian in Hayonim Cave and the
Natufian of the Terrace. In Les Fouilles de la Terrasse d’Hayonim (Israel) 1980-1981 et 1985-1989. F.R. Valla, ed. Pp. 471-519. Memoires et Travaux du Centre de Re-cherche Francais a Jerusalem 10. Paris: De Boccard.
Bar-Yosef, O., Belfer-Cohen, A., Goldberg, P., Kuhn, S. L., Meignen, L., Vandermeersch, B. and S. Weiner
2005 Archaeological Background to Hayonim Cave and Meged Rockshelter. In The Faunas of Hayonim Cave, Israel: A 200,000-year record of Paleolithic Diet,
Fig. 13. Skeletal part representation of partridges from Hilazon Tachtit. All MNE values are corrected for the number of times the element occurs in the body (Minimum number of animal units; MAU).

474
Natalie D. Munro
Demography, and Society, by M. C. Stiner, pp. 17-38. American School of Prehistoric Research Bulletin 48, Peabody Museum of Archaeology and Ethnology, Harvard University, Cambridge.
Belfer-Cohen, A. 1988 The Natufian Settlement at Hayonim
Cave: A Hunter-gatherer Band on the Threshold of Agriculture. Ph.D. disser-tation, The Hebrew University of Jeru-salem.
1991 The Natufian in the Levant. Annual Review of Anthropology 20:167-186.
Belfer-Cohen, A and A. N. Goring-Morrisherein Breaking the Mould: Phases and Facies in
the Natufian of the Mediterranean Zone. Edited by O. Bar-Yosef and F. R. Valla. International Monographs in Prehistory, Ann Arbor.
Boyd, B.1992 The transformation of knowledge: Late
Epipalaeolithic (Natufian) mortuary practices at Hayonim, Western Galilee. Archaeological Review from Cambridge 11/1:19-38.
Crabtree, P. 1990 Zooarchaeology and complex societies:
some uses of faunal analysis for the study of trade, social status, and ethnicity. Archaeological Method and Theory 2:155-205.
Davis, S. J. 1980 A note on the dental and skeletal ontogeny
of Gazella. Israel Journal of Zoology 29:129-134.
Grosman, L. and N. D. Munro2007 The sacred and the mundane: domestic
activities at a Late Natufian burial site in the Levant. Before Farming 4/4:1-14.
Grosman, L., Munro, N. D. and A. Belfer-Cohen2008 A 12,000 year old burial from the
southern Levant (Israel): a case for early shamanism. Proceedings of the National Academy of Sciences 105/46:17665-17669
Henry, D. O. and A. Leroi-Gourhan1976 The excavation of Hayonim Terrace:
an Interim Report. Journal of Field Archaeology 3:391-406.
Henry, D. O., Leroi-Gourhan, A. and S. Davis1981 The excavation of Hayonim Terrace: an
examination of Terminal Pleistocene climatic and adaptive changes. Journal of Archaeological Science 8:33-58.
Hesse, B. 1986 Animal use at Tel Miqne-Ekron in
the Bronze Age and Iron Age. Bulletin of the American School of Oriental Research 264:17-27.
Le Dosseur, G. and C. Maréchalherein Bone ornamental elements and decorated
objects of the Natufian from Mallaha. Edited by O. Bar-Yosef and F. R. Valla. International Monographs in Prehistory, Ann Arbor.
Lyman, R. L. 1984 Bone density and differential survivor-
ship of fossil classes. Journal of Anthro-pological Archaeology 3:259-299.
1993 Density-mediated attrition of bone as-semblages: new insights. In From Bones to Behavior: Ethnoarchaeological and Experimental Contributions to the Inter-pretation of Faunal Remains, edited by J. Hudson, pp. 324-341. Occasional paper No. 21. Center for Archaeological Inves-tigations, Southern Illinois University. Carbondale.
1994 Vertebrate Taphonomy. Cambridge University Press, Cambridge.
Marean, C. W. 1991 Measuring the postdepositional destruc-
tion of bone in archaeological assemblag-es. Journal of Archaeological Science 18:677-694.
Munro, N. D. 2001 A Prelude to Agriculture: Game Use
and Occupation Intensity During the Natufian Period in the Levant. Ph.D. dissertation, University of Arizona, Tucson.
2004 Zooarchaeological measures of hunting pressure and occupation intensity in the Natufian: implications for agricultural origins. Current Anthropology 45:S5-S33.
Munro, N. D., Bar-Oz, G. and A. Stutz2009 Aging Mountain Gazelle (Gazella gazel-
la): Refining Methods of Tooth Eruption and Wear and Bone Fusion. Journal of Archaeological Science 36:752-763.
Reitz, E. and E. Wing1999 Zooarchaeology. Cambridge University
Press, Cambridge.Stiner, M. C.
1994 Honor Among Thieves: A Zooarchaeo-logical Study of Neandertal Ecology. Princeton University Press, Princeton.

475
A Faunal Perspective on…Hayonim Cave and Hayonim Terrace
Tchernov, E. and F. R. Valla1997 Two new dogs, and other Natufian dogs,
from the southern Levant. Journal of Archaeological Science 24:65-95.
Valla, F. R., Le Mort, F. and H. Plisson1991 Les fouilles en cours sur la Terrase
d’Hayonim. In The Natufian Culture in
the Levant, edited by O. Bar-Yosef and F. R. Valla, pp. 93-110. International Monographs in Prehistory, Ann Arbor.
Valla, F. R., Plisson, H. and R. Buxo I Capdevila1989 Notes préliminaires sur les fouilles
en cours sur la Terrasse d’Hayonim. Paléorient 15/1:245-257.
Appendix 1. Number of Identified Specimens (NISP) of economic taxa represented at Hayonim Cave and Hayonim Terrace. *Counts of skeletal elements from 2 canid burials not included in total. Indet. = indeterminate.
COMMON NAME LATIN NAME HAYT LN HAYC LNUngulates:Mountain gazelle Gazella gazella 5008 883Wild goat Capra aegagrus 35 7Aurochs (wild cattle) Bos primigenius 93 4Roe deer Capreolus capreolus 26 1Fallow Deer Dama mesopotamica 90 20Red deer Cervus elaphus 19 13Cervid Cervidae 13 28Wild pig Sus scrofa 85 43
Small Ungulate 123 574
Medium Ungulate 108 51
Large Ungulate 12 10
Huge Ungulate 5 0
Carnivores:Jungle cat Felis cf. chaus 52 51Leopard Panthera pardus 0 2Red fox Vulpes vulpes 373 153*Wolf/dog Canis sp. 31 3Mustelid Musetlidae 7 8Polecat Vormela peregusna 7 5Stone marten Martes foina 8 15Badger Meles meles 21 20Carnivore Indet. Carnivora indet. 8 59
Small Carnivore Indet. 63 0
Small Game:Cape hare Lepus capensis 740 417Hedgehog Erinaceus europaeus 13 6Diurnal Raptors Falconiformes indet. 55 40Golden eagle Aquila chrysaetos 9 0Eurasian sparrowhawk Accipiter nisus 0 14Buzzard Buteo buteo 34 38Kestrel Falco tinnunculus 0 1
Vulture indet. 1 3

476
Natalie D. Munro
COMMON NAME LATIN NAME HAYT LN HAYC LNPartridges and Quails Phasinidae 0 2Chukar Partridge Alectoris chukar 69 231Common Quail Coturnix coturnix 0 7Owls Strigidae 1 6Jay Garullus sp. 0 1Pigeons Columbiformes indet. 4 8
Waterfowl 0 7
Great bustard Otis tarda 0 5
Small Bird 13 12
Medium Bird 59 158
Large Bird 33 30
Huge Bird 8 9
Mediterranean Spur-Thighed Tortoise Testudo graeca 2987 2542Fish Pices indet. 15 8Breams Sparids 8 0
Large Mammal 0 15
Medium Mammal 77 226
Small Mammal 112 178
TOTAL 10425 5914
Appendix 1. (Continued…)
Appendix 2. NISP and MNE values for gazelle elements and long bone portions from Hayonim Cave and Hayonim Terrace
HAYT LN HAYC LN NISP MNE NISP MNEHorn and Head: Horn 79 24 62 20 Skull 92 21 62 9 Mandible 164 27 47 7 Teeth Upper 207 17 67 9 Teeth Lower 337 28 73 11
Axial Skeleton: Atlas 2 1 7 3 Axis 5 2 4 2 Cervical 38 11 31 4 Thoracic 44 16 26 9 Lumbar 55 17 36 11 Caudal 9 9 2 2 Sacrum 0 0 1 1 Sternum 2 1 8 1 Rib 75 16 147 54

477
A Faunal Perspective on…Hayonim Cave and Hayonim Terrace
HAYT LN HAYC LN NISP MNE NISP MNE
Forelimb: Scapula-glenoid fossa 71 57 14 11 Scapula-blade 27 4 17 4 Humerus-proximal 16 6 9 5 Humerus-distal 130 83 44 36 Humerus-shaft 57 27 46 27 Radius-proximal 54 36 38 22 Radius-distal 14 6 13 7 Radius-shaft 16 4 48 18 Ulna-proximal 73 62 30 24 Metacarpal-proximal 33 16 25 14 Carpal-cuneiform 31 30 5 5 Carpal-lunate 48 48 6 6 Carpal-magnum 38 38 7 6 Carpal-scaphoid 64 62 12 12 Carpal-unciform 41 40 7 7
Hindlimb: Pelvis-ilium 30 26 12 10 Pelvis-ischium 37 12 7 4 Pelvis-pubis 58 49 12 12 Pelvis-acetabulum 129 12 27 10 Femur-proximal 72 65 9 3 Femur-distal 43 26 16 8 Femur-shaft 21 4 18 6 Tibia-proximal 19 9 12 8 Tibia-distal 51 39 24 10 Tibia-shaft 28 7 43 7 Patella 51 50 13 13 Astragalus 96 82 29 24 Calcaneum 66 45 31 24 Navicular cuboid 41 18 19 17 Ext&Mid cuneiform 74 73 2 2 Lateral malleolus 38 37 3 3 Metatarsal-proximal 47 20 25 15
Toes: Phalanx 1 645 266 151 89 Phalanx 2 598 341 122 92 Phalanx 3 281 219 74 65 Seasamoid 527 525 19 19 Metapodial-distal 417 184 132 57 Metapodial -shaft 39 12 99 28
Appendix 2. (Continued…)
Copyright © 2022 FDOKUMEN




















