
Mudskippers (Literature Review)
71
Oceanogr. Mar. Biol. Annu. Rev., 1993, 31, 507-577 A. D. Ansell, R. N. Gibson and Margaret Barnes, Editors UCL Press MUDSKIPPERS DAVID A. CLAYTON Biology Department, Sultan Qaboos University, PO Box 36 Al Khod Postal Code 123, Muscat, Sultanate of Oman ABSTRACT The oxudercine gobies commonly known as mudskippers provide a rich source for comparative studies in adaptation to the littoral habitat. In spite of the paucity of information on reproduction, culturing techniques have improved in recent years and the developmental stages of the Chinese and Japanese species are now known. Morphological studies of mature fish havecentred on skeletal characteristics as an adjunct to taxonomy and on vision, respiration and excretion. Mudskippers are euryhaline and ammoniotelic with sophisticated biochemical pathways for ammonia detoxification operating to different extentsin the variousgenera. Free aminoacids play a central role in both excretion and osmoregulation. The relative importance of the respiratory surfaces of these facultative air-breathing fishes varies in air and water and also between genera. The surfaces include the gills and modified buccal, pharyngeal and opercular epithelia as well as limited, well vascularised areas of the skin. The respiratory rate and energy consumption are reduced during hypoxia, but the full extent of metabolic changes in aestivating or hibernating fishes has yet to be fullyexplored. Mudskippers usuallyinhabit tidal mudflats and mangroves, but can be found on sandy and rocky shores. Of the common genera, Boleophthalmus and Scartelaos are considered to be more aquatic than Periophthalmus and Periophthalmodon, but complex patterns of zonation indicate that more detailed ecological data are required on this topic as well as on the biotic com ponents and parasites. From simple tidal migrations to burrow construction and complex mud-walled polygonal mosaics, mudskippers exhibit a wide range of territorial behaviour which is matched by their diverse courtship and agonistic displays. INTRODUCTION The literature on the biology of intertidal fishes is extensive and has been reviewed by Gibson (1969, 1982, 1986). Despite the fact that observations on African, Australian and South American species are for the most part sadly lacking (Gibson, 1982), the best known examples of resident intertidal fishes are probably the predominantly Indo-Pacific mudskippers. In general, mudskippers can be defined as those fishes usually found moving about on the mudflats of creeks, estuaries and coastal waters when they are exposed at low tide (Qureshi & Bano, 1971). From the frontispiece of Higson's (1889) "A Naturalist in North Celebes" to Polunin's (1972) "Who says fish can't climb trees?" and the similar pictorial essay on "A fish out of water" (Fukuda, 1985), these perversely photogenic gobies, with their rich behavioural repertoire have caught the popular imagination (Sowerby, 1923; van Dijk, 1959; Whitley, 1960; MacNae, 1968a,c; Kaden, 1978) and warranted mention in museum (Inger, 1952; Whitley, 1968—a reprint
Transcript of Mudskippers (Literature Review)

Oceanogr. Mar. Biol. Annu. Rev., 1993, 31, 507-577A. D. Ansell, R. N. Gibson and Margaret Barnes, EditorsUCL Press
MUDSKIPPERS
DAVID A. CLAYTON
Biology Department, Sultan Qaboos University, PO Box 36 Al KhodPostal Code 123,Muscat, Sultanate of Oman
ABSTRACT The oxudercine gobies commonly known as mudskippers provide arich source for comparative studies in adaptation to the littoral habitat. In spite of thepaucity of information on reproduction, culturing techniques have improved in recentyears and the developmental stages of the Chinese and Japanese species are now known.Morphological studies of mature fish havecentred on skeletal characteristics as an adjunctto taxonomy and on vision, respiration and excretion. Mudskippers are euryhaline andammoniotelic with sophisticated biochemical pathways for ammonia detoxificationoperating to different extents in the various genera. Free amino acids play a central rolein bothexcretion and osmoregulation. The relative importance of the respiratory surfacesof these facultative air-breathing fishes varies in air and water and also between genera.The surfaces include the gills and modified buccal, pharyngeal and opercular epitheliaas well as limited, well vascularised areas of the skin. The respiratory rate and energyconsumption are reduced during hypoxia, but the full extent of metabolic changes inaestivating or hibernating fishes has yet to be fully explored. Mudskippers usuallyinhabittidal mudflats and mangroves, but can be found on sandy and rocky shores. Of thecommon genera, Boleophthalmus and Scartelaos are considered to be more aquatic thanPeriophthalmus and Periophthalmodon, but complex patterns of zonation indicate thatmore detailed ecological data are required on this topic as well as on the biotic components and parasites. From simple tidal migrations to burrow construction and complexmud-walled polygonal mosaics, mudskippers exhibit a wide range of territorial behaviourwhich is matched by their diverse courtship and agonistic displays.
INTRODUCTION
The literature on the biology of intertidal fishes is extensive and has beenreviewed by Gibson (1969, 1982, 1986). Despite the fact that observations onAfrican, Australian and South American species are for the most part sadlylacking (Gibson, 1982), the best known examples of resident intertidal fishes areprobably the predominantly Indo-Pacific mudskippers. In general, mudskipperscan be defined as those fishes usually found moving about on the mudflats ofcreeks, estuaries and coastal waters when they are exposed at low tide (Qureshi& Bano, 1971).
From the frontispiece of Higson's (1889) "A Naturalist in North Celebes" toPolunin's (1972) "Who says fish can't climb trees?" and the similar pictorialessay on "A fish out of water" (Fukuda, 1985), these perversely photogenicgobies, with their rich behavioural repertoire have caught the popular imagination(Sowerby, 1923; van Dijk, 1959; Whitley, 1960; MacNae, 1968a,c; Kaden,1978) and warranted mention in museum (Inger, 1952; Whitley, 1968—a reprint

508 DAVID A. CLAYTON
of the 1960 article) and zoo (Jes, 1972) bulletins. In some areas mudskippersare of economic importance, because they are eaten in China (Schreitmuller,1914), Taiwan (Liao et al, 1973), India (Das, 1934; Hora, 1935a; Siddiqi,1974; Qureshi & Bano, 1971) and provide an alternative fishery during themonsoon in Bombay (Mutsaddi & Bal, 1973). They are extensively cultured inTaiwan (Chen, 1976) and to some Malaysians the raw flesh of these fishes hasaphrodisiac properties (Johnstone, 1903). Among Europeans, however, themudskippers have remainedmerely interesting, if unusual, aquariumfish(Reichelt,1910; Schreitmuller, 1911; Rodewald, 1913; Simpson, 1955; Klausewitz,1968; Kluge, 1971; Von Anthouard & Mignot, 1973; Kaden, 1978; Hunziker,1985; Murdy, 1986; Schneider, 1990a,b). It is doubtful if there is any othergroup of fishes in which so much general interest is based on so little scientificknowledge.
There are four main reasons for this discrepancy. First, much of the popularwriting is based on only a few aspects of the fishes' biology. Parenthetically,some of this information was incorrect as in the case of the fishes' respiratoryadaptations, the misunderstanding of which began with Higson (1889) and onlyended with Sponder & Lauder (1981). Secondly, the scientific information is noteasily available, because it is written in a number of languages and spread overa wide variety of often inaccessible sources. As a consequence, relevant literature is often overlooked. For example, Yadav et al (1990) omitted thepertinent study of respiratory organs by Singh & Munshi (1969) and similarly,Lele & Kulkarni (1938, 1939) are not mentioned in the osteological study of Lee(1990). Thirdly, the very facet of their behaviour that attracted the originalinterest, their extreme manoeuvrability in an environment difficult of access forman, meant that they are hard to catch. Individuals caught by blowpipe (Higson,1889), weighted hooks or a 0 •22 air rifle (Clayton & Vaughan, 1986) are of littleuse for further study. Fish caught by traps or nets (Burhanuddin & Martosewojo,1979) or lassos (Khoo, 1966) are obviously preferable, but these are very inefficient techniques that often create unwanted habitat disruption. Consequently,in accounts of the ecology of tropical shores on which mudskippers are found,good qualitative data for other macrofaunal taxa are presented but only thepresence or absence of mudskippers is reported (Berry, 1972; Day, 1974;Sasekumar, 1974; Frith etai, 1976; Dorjes, 1978; Branch & Grindley, 1979;Nateewathana & Tantichodok, 1984). Even where the apparent redistribution ofmudskippers is discussed (Lipschitz et al, 1975) no quantitative data arepresented. Finally, the taxonomy of the group of fishes known as mudskippershas been in a state of confusion. Whilst this is partially a reflection of theproblematic systematics of gobies in general (Miller, 1973) it is also exacerbatedby problems of specific identification, as illustrated by Brillet's (1969a, 1975)study of the behaviour of Periophthalmus sobrinus and Al-Kadhomiy & Hughes'(1988) study of the comparative morphology of gill structure of Boleophthalmusboddarti. Wherever possible in this review correct specific designations basedon Murdy (1989) will be provided.
Previous reviews (Gibson, 1969, 1982, 1986) including mudskippers concentrated mainly on recent literature, but this synthesis attempts to include most ofthe literature irrespective of its age. This is partly because the literature is morelimited, but also because some of the earlier work has been misinterpreted orignored, despite continued relevance for present studies.

MUDSKIPPERS 509
TAXONOMY
Two brief examples of the taxonomic confusion will suffice to illustrate theproblems. Periophthalmus vulgaris was distinguished by Eggert (1935) and hasat least eight synonyms, the most common being P. koelreuteri (Pallas) andP. argentilineatus (Cuvier& Valenciennes). Eggert separated P. vulgaris fromthese on the form of the first dorsal fin which, although variable, did not havethe convex, fan-like or irregular shape of these species, and also by differencesin colour, pelvic fin and tooth form. Whitley (1931) separated P. vulgaris fromP. koelreuteri, but wrongly usedit for Euchoristopus kalolo (= Periophthalmuskalolo Lesson), which is unquestionably synonymous with P. koelreuteri(Milward, 1974). Whitley (1953, 1960, 1968) and MacNae (1968a,b) repeatedthe error. Furthermore, whilst Eggert (1935) recognised four geographicallyseparatedsubspecies of P. vulgaris, the diagnosticcharacteristicsprovided wereinadequate (Herre, 1941). The considerable intraspecific variation in fin spinesand rays (Milward, 1974) clearly contributed to this confusion as it has in theidentity of other mudskippers.
Boleophthalmus dussumieri (Valenciennes) and B. dentatus (Valenciennes)were originally described (Cuvier & Valenciennes, 1837) as differing in dentition and dorsal fin structure, the latter having stronger canines and united firstand second dorsal fins. These two species were later distinguished from a thirdspecies, B. chamiri Holly (Holly, 1929) on the basis of the possession of agreater number of teeth in the upper jaw. Tooth number is an age specificvariable, however, and the size and fusion of the first and second dorsal fins asex specific one (Berg, 1949). The correct designation, based on page priorityin Cuvier & Valenciennes (1837) is B. dussumieri (Berg, 1949). Nevertheless,and despite Khalafs (1961) caution, both names persist in more current literature (Al Nasiri & Hoda, 1975; Hoda, 1980; 1985). Furthermore, the relationship between this species and B. boddarti (commonly misspelt as boddaerti)remained problematical because the original descriptions were inadequate, and,as in other cases, the type specimens were no longer extant.
Before nomenclatural confusion of the sort illustrated above could be resolved,intraspecific variation in diagnostic characters needed to be assessed usingextended size range collections from all locations inhabited by each species. Thespecific determinations for most, if not all mudskippers, was based entirely onexternal features many of which have been shown to be of limited value and themore recent osteological criteria used in gobioid classification (Miller, 1973)needed to be incorporated in any such attempt.
Interrelationships between mudskipper taxa have recently been reassessedusing such criteria (Birdsong et al, 1988; Murdy, 1989) and have resulted ina complete revision of their taxonomy. The mudskipper genera are now allplaced in the Family Oxudercinae. The Tribe Oxudercini includes the generaParapocryptes (2 species), Apocryptodon (2) and Oxuderces (1) and the TribePeriophthalmini the genera Apocryptes (1), Pseudapocryptes (2), Zappa (1),Scartelaos (4), Boleophthalmus (5), Periophthalmodon (3) and Periophthalmus(12) (Murdy, 1989). This revision of the specific synonyms has reduced thenominal 103 species to a respectable 34, and will benefit future researchersmerely wishing to identify their experimental subjects. The revision also furtheremphasises the necessity for voucher specimens. The usual data on collectionshould be supplied and the latitude and longitude should be included in the

510 DAVID A. CLAYTON
locality information. In any publication authors should provide the museum'scatalogue number for the specimen(s). Current programmes of research will beable to rectify their subjects' identity, but continued care will be needed incomparisons made with other work. While some reassignments can be madeeasily others will be more difficult. Forexample Periophthalmus barbarus is theonly periophthalmid identified in west Africa and Periophthalmus modestus istheonly one inJapan. Conversely, Periophthalmus vulgaris and Periophthalmuskoelreuteri are no longer valid species names, anderroneously havebeen appliedto a number of extant species.
MORPHOLOGY
CHROMOSOMES
As an adjunct to the more usual morphological classificatory system for theanalysis of gobioid fishes, some karyotypic chromosomal analyses of mudskippers have been undertaken, some of which are tabulated in Nishikawa etal(1974). For Apocryptodon madurensis and Pseudapocryptes borneensis [ =Periophthalmus lanceolatusl] Verma (1968) records 48 as the diploid numberof chromosomes. There is agreement that the diploid number is 46 in P.cantonensis [ = P. modestus] (Nogusa, 1957, 1960; Arai & Kobayasi, 1973;Nishikawa etal, 1974). In Boleophthalmuspectinirostris (Nogusa, 1957, 1960;Nishikawa et al, 1974), B. dussumieri (Krishnaja & Rege, 1980; Verma,1968), andB. boddarti (Subrahmanyam, 1969; Verma, 1968; Manna & Prasad,1974) the number is also 46, but the detaileddescriptions of the Indian materialdiffer considerably. There is further confusion in that Manna& Prasad's (1974)identification of B. glaucus mustbe incorrectbecausethis species [ = Scartelaoscantoris] (Murdy, 1989) is only known from the Andaman Islands. The mostlikely appellation is Boleophthalmus boddarti. Subrahmanyam (1969) reportedthat the chromosomes of B. boddarti were all metacentric with a large hetero-morphic pair which could be the sex chromosomes. Verma (1968) failed to findany similar elements and Manna & Prasad (1974) recorded onlysix metacentricpairs, the remainder being submetacentric, acrocentric or telocentric. Onlyacrocentric chromosomes were found in B. dussumieri (Krishnaja & Rege,1980) and in B. pectinirostris and Periophthalmus modestus there were bothacrocentric and telocentric chromosomes (Nishikawa et al, 1974). Nevertheless,the chromosome structure of all three species would, as suggested by Krishnaja& Rege (1980) for Boleophthalmus dussumieri, provide good material for thedetection of chromosomal aberrations in mutagenic studies.
NERVOUS AND SENSORY SYSTEMS
Brain
The gross structure of the mudskippers' nervous system has largely beenignored. The dorsal structures of the brain are illustrated for Periophthalmuschrysospilos (Harms, 1935), P. barbarus [ = Periophthalmodon schlosseri](Lim, 1967), Periophthalmus koelreuteri, Pseudapocrytes lanceolatus (Datta& Das, 1980), Boleophthalmus boddarti (Lim, 1967; Datta & Das, 1980),Boleophthalmus sp. and Apocryptes sp. (Mookerjeeetal, 1950).The illustrations

MUDSKIPPERS 511
of Datta & Das (1980) are diagrammatic and while those of Lim (1967) areanatomically correct, they reveal little more than that the brain of the carnivorous Periophthalmus is larger than that of the herbivorous Boleophthalmus.As part of their study on rhythmic changes in neuroendocrine function of B.dentatus, Patel & Desai (1976) show micrographical sections of the hypothalamic hypophyseal region of the brain.
Mechanoreceptors
The mechanoreceptors of the lateral line system of mudskippers are limited toepidermal organs (neuromasts) as there are no lateral line canals (Afzelius,1956). Working with small Periophthalmus barbarus [as P. koelreuteri], hedetailed the location and nervous innervation of the neuromasts of the head.While most are embedded deeply in the epithelium, the two rear organs aresituated on elevations posterior to the eyes (Afzelius, 1956). He thought theywere innervatedby the ramus ophthalmicus superficialis, but is is now acceptedthat lateral line organs are innervated by the acoustic (VIII) nerve (Parker &Haswell, 1962) and it is likely that the nerve Afzelius observed was a sectionof the anterior lateral line nerve that joined the trigeminal (V). The greatestconcentration of neuromasts are along the lower jaw, and as part of a study oftheir function in these amphibious fishes, it would be interesting to compare thisdistribution with that in Boleophthalmus or Scartelaos (see p. 514). The structure of these organs as revealed by SEM would also prove instructive.
Without being able to determine the location of the sound receptor, Diessel-horst (1938) showed that Periophthalmus koelreuteri [?] detected air-bornesounds in the range 258-651 Hz. The sounds used were a conditioned stimulusfor food and elicited responses after 45 training presentations. However,becauseof the variable response, which in some cases was indicatedonly by eyemovement. Diesselhorst did not perform any discrimination tests. As a corollaryto this, the rapid escape response of many fishes is mediated by the Mauthnerreflex in which vibrational stimulation of the acoustic nerve initiates a strongmuscular response via the Mauthner cells (Diamond, 1971). The response ispresumed to assist in the avoidance of aerial predators, particularly diving orplunging ones. While the role of visual stimuli is acknowledged (Diamond,1971), it has not been investigated and the study of the mudskippers' startleresponse is likely to be doubly rewarding in the absence of vibrational cues.
Eyes
The dorsally protruding eyes of mudskippers are unique in a number of waysand while some of these were described by Graham (1971), it was withoutreference to the original literature, which in any case is rather confusing (Munk,1970). The generic designations, Boleophthalmus and Periophthalmus, reflectthe great mobility of the eyes of mudskippers, the musculature of which isdescribed for P. koelreuteri from Ceylon [ = P. kalolo or argentilineatus] byKarsten (1923), Periophthalmodon schlosseri and Boleophthalmus or Scartelaos[as Boleophthalmus sp. Val.] by Oliva & Skorepa (1970a,b respectively). Theeye muscles of mudskippers are similar to those of other fishes (Karsten, 1923),but because the length is greater than the diameter of the eyeball, they are longerthan those of deep sea fishes. Mudskippers also have clear anterior and posteriorpoints of muscle attachment (myodomes) which are more specialised in the

512 DAVID A. CLAYTON
carnivorous Periophthalmodon than the herbivorous Boleophthalmus (Oliva &Skorepa, 1970a, b). The eyes move independently of each other and also can beretracted into the head where they are covered by a ventrally placed lid-like skinfold (Karsten, 1923; Munk, 1970). This dermal cup (Graham, 1971) serves asa water reservoir for moistening the eye surface and is a characteristic used todistinguish Scartelaos, Boleophthalmus, Periophthalmus and Periophthalmodonfrom the other taxa within the Oxudercinae (Murdy, 1989). The cornea isdivided into distal and proximal lamellae which together correspond to thecornea of other vertebrates (Karsten, 1923). The outer lamella is presumed toprovide additional protection from desiccation and mechanical damage (Graham,1971).
The internal anatomy of the eye has been discussed for Periophthalmodonschlosseri (Volz, 1905a, b,c; Baumeister, 1913), Periophthalmusargentilineatus(Karsten, 1923; Harms, 1914, 1929; Munk, 1970), P. kalolo [as P. koelreuteri] (Hess, 1912, 1913; Munk, 1970), P. chrysospilos (Yew & Wu,1979), P. modestus [as P. cantonensis] (Yamamoto, 1931), Boleophthalmusboddarti (Volz, 1905a; Baumeister, 1913; Munk, 1970) and B. pectinirostris(Franz, 1910).
Despite the absence of the falciform process and the retractor lentis muscle(campanila Halleri) (Volz, 1905a,c) and contrary to Baumeister (1913), thearrangement of lens muscles is essentially the same as in other fishes and enableaccommodation to take place (Karsten, 1923). What form accommodationactually takes is debatable, for whereas Hess (1912, 1913) found that the unaccommodated eye of Periophthalmus argentilineatus in air focuses light frominfinity on (emmetropic) or slightly behind (hypermetropic) the retina, Yamamoto(1931) found the eye of P. modestus to be hypermetropic; slightly in air andstrongly so in water. In either case, the accommodatory range is probablyinsufficient to restore emmetric vision in water and the Periophthalmus eye mustbe considered to be adapted for aerial vision. It is likely that this generalisationalso applies to the other amphibious genera (Munk, 1970). The strongly curvedcornea and, in comparison with other fish, the flattened lens of Periophthalmusand Boleophthalmus also contribute to aerial adaptation (Graham, 1971). Thesegenera only differ in the shape of the pupillary aperture (horizontal slit and heart-shaped respectively) and the curvature of the retinal surface relative to theanatomical axis of the eye (Munk, 1970).
Periophthalmus has a 'ramp' retina which is tilted away from the lens suchthat the distance from the centre of the lens to the retina increases continuouslyin the vertical direction (Karsten, 1923; Munk, 1970). Besides accommodation,this is another method that allows objects at different distances from the eye tobe in focus. With a 'ramp' retina objects on the ground could be in focus on thedorsal part of the retina at the same time that a distant object located higher inthe visual field was in focus on the ventral retina. The density of visual cells inthe dorsal part of the retina is greater than that of the ventral part and in generalthe Periophthalmus retina is better developed than that of Boleophthalmus(Munk, 1970). Without specifying the mechanism, Munk (1970) stated that theeyes are light adapted and while Graham (1971) stated that they contain highconcentrations of pigment which gives protection from light and glare, Karsten(1923) found a strongly reflective argentea fibrosa, an adaptation for collectinglight, in Periophthalmus. The retina of Boleophthalmus pectinirostris has noretinomotor response (Zhang & He, 1989). There is no change from a dark-

MUDSKIPPERS 513
adapted state when it is exposed to a variety of different ambient light intensitiesand Zhang & He (1989) suggest that the pupillary response is entirely responsible for adaptation to different light conditions in this species. Contrary to thestatements of Harms (1929) and Graham (1971), rods and cones are found inboth the dorsal and ventral area of the retina. There are differences in theirdistribution, however, because Yew & Wu (1979) noted a decrease in both rodsand single cones and an increase in double cones from the anterior to the centreof the retina of P. chrysospilos.
Increased length of the outer segments of the visual cells also means thatobjects at various distances would be equally in or out of focus (Munz, 1971).Munk (1970) demonstrated that both Boleophthalmus {boddarti) and Periophthalmus (argentilineatus and kalolo) have horizontal band-shaped areae whichmay meet this criterion. The areae extend right across the retina slightly dorsalto the centreof the fundus where the retina and the visual cell layers are at theirthickest. In comparison with the rest of the retina, the photoreceptors (rods,cones and double cones) of the areae are longer and thinner and their densityis greatly increased. Possibly to avoid reducing the sensitivity of the areae, theoptic nerve fibres do not all converge on the optic disc as in other fishes, butare spread outovera largerpart of the retinal surface. The high density of visualcells in the areae increases the eyes' resolving power and lowers the thresholdof movement perception, especially in thehorizontal plane at right angles to theanatomical axisof the eye. Bothfeatures willassist in prey {Periophthalmus) andpredator detection {Boleophthalmus, Periophthalmus', Munk, 1970).
SKELETON
Detailed skeletal characteristics of mudskippers have been investigated by Lele& Kulkarni (1938, 1939), Birdsong etal (1988), Murdy (1989) andLee(1990).Eggert (1929a) and Harris (1961) concentrated on theappendicular skeleton andmuscular adaptations for locomotion on land. Pectoral fin musculature, forexample, in the form of the adbuctor superficialis muscle being divided intotwo sections, is a terrestrial adaptation. It is found in Periophthalmus, Periophthalmodon and Boloephthalmus, but not Scartelaos (Eggert, 1929b; Harris,1961; Murdy, 1989). Additionally, fin and girdle modifications of Periophthalmodon schlosseri [as Pseudapocryptes schlosseri] and Periophthalmus kalolo[as P. koelreuteri] are discussed by Mehta et al (1986, 1987), but like theaccounts of Venkateswarlu (1969) and Chatterjee & Siddiqi (1957) add littleto thebiology of theseanimals. Thedetails of the axial skeleton haveimportancefor taxonomy, the spinous dorsal fin pterygiophore formula being of particularimportance (Birdsong et al, 1988). From figures of the axial skeleton of P.barbarus (Lele & Kulkarni, 1939) and P. cantonensis (Lee, 1990) the formulae(Murdy, 1989) indicate that these two species shouldproperly be designated asP. kalolo andP. modestus respectively (Murdy, 1989). In comparison withothergobies, the pre-orbital shortening of mudskipper skulls is evident (Lele &Kulkarni, 1938; Murdy, 1989; Lee, 1990), as is the reduction in bones (Lele& Kulkarni, 1938; Afzelius, 1956). Morphologically, little attention has focusedon the pharyngeal plates. The marked differences in dentition of these platesrelate to different feeding strategies (Milward, 1974) but they may also havetaxonomic value.

514 DAVID A. CLAYTON
SKIN
The skin as a respiratory surface is discussed in the section on respiration(p. 541), but Bai & Kalyani (1960a,b,c) briefly give some detail of the skin ofB. boddarti. In their work on scale morphogenesis and regeneration Bai &Kalyani (1960a, b,c) report only that the concentration of Vitamin C (ascorbicacid) in the brain (14.6 mg-lOOg"1) of B. boddarti was considerably higherthan that of kidney (8.73), liver (2.09), skin (2.19), muscle (0.51) or blood(0.29). No explanation of the function of Vitamin C was presented although theysubsequently reported on imino acid concentrations (Bai & Kalyani, 1961) forwhich ascorbic acid is a necessary substrate in the conversion of proline tohydroxyproline. These compounds stabilise collagen and hydroxyproline is achemical marker for its presence. The concentration of the imino acids werehigher (16.45 g-lOOg"1) in scales than in skin ((13.91 g-100"1). High brainlevels of ascorbic acid are typical of marine fish, but only comparisons betweenthe different genera will show whether the distribution of Vitamin C in theremaining tissues, especially the skin, is of significance to the mudskippers'amphibious mode oflife. The glucose, fructose, galactose and xylose content ofthemuscles of B. pectinirostris (Yamazoe, 1970) andthe fatty acids of thebodyand liver of B. boddarti (Misra et al, 1983) have also been measured.
ALIMENTARY TRACT
Alimentary tract morphology has been histologically described for B. boddarti,Periophthalmodon schlosseri [as Periopthalmus barbarus] (Lim, 1967), Periophthalmus vulgaris [= P. argentilineatus] (Lim, 1971; Milward, 1974),Periophthalmodon schlosseri (Lim, 1971), P. freycineti [as P. schlosseri] andScartelaos histophorus (Milward, 1974). Miyazaki & Nakamura (1980) describethe muscles in the region of the tongue of Boleophthalmus pectinirostris andMehta etal (1990) compare the gross morphology of the head and position ofthe mouth of a number of oxudercine species and attempt to relate this to theirmethod of foraging.
The mouth is situated terminally on the lower part of the blunt snout and hasfleshy lips which are presumably well supplied with sensory cells, especially inthe herbivorous species. The jaw dentition is specialised such that carnivoreshave conical pointed teeth which may be of unequal size due to replacement(Milward, 1974). More spatulate, horizontally oriented teeth, as in Boleophthalmus, are more typical of herbivores. Boleophthalmus, Scartelaos andOxuderces possess a pairof large symphysial teeth (Lim, 1967; Milward, 1974;Murdy, 1989) which are probably used in agonistic situations (Lim, 1967).Pharyngeal plates also reflect diet because canine-like teeth, as illustrated forPeriophthalmus barbarus [as P. koelreuteri] (Sponder & Lauder, 1981) arefound in carnivores and smaller, more spatulate ones in Boleophthalmus (Lim,1967; Clayton, unpubl. data). Periophthalmodon andPeriophthalmus haveshorttuberculated gill rakers (Lim, 1971), while those of Boleophthalmus (Lim,1967) and Scartelaos (Milward, 1974) are much longer and more flexible. Therelative gut length has beendiscussed in relation to diet (p. 553). Only Milward(1974) notes any internal divisions of the alimentary tract, noting the presenceof a pyloric sphincter inperiophthalmid species. Along itsentirelength, the tractof all mudskippers studied shows much folding. It is greatest in the stomach and

MUDSKIPPERS 515
rectum, but Lim (1967) showed the stomach of Periophthalmon schlosseriwithout any folding and P. argentilineatus has none in its rectum (Lim, 1971).The only report dealing with enzymeactivity is that of Dhage & Mohamed(1977)in which the amylase activity of P. koelreuteri [IP. kalolo] decreased anteroposterior^ along the tract.
KIDNEY
Periophthalmus is one of the few teleost genera to retain in maturity functionalnephrons in the pronephros or head kidney, the gross and fine structure of whichhas been investigated by Safer et al (1982) and Safer & El-Sayed (1986). InP. waltoni [as P. koelreuteri] the paired renal corpuscles in the body of the headkidney and the single nephron of the anterior lobes are structurally indistinguishable from other kidney tubules. Haemopoietic tissue forms a supportivematrix throughout, with that of the anterior lobes being predominantly erythro-poeitic. Ultrastructurally, the nephronic tubuleconsists of two proximal segments,a distal segment and a collecting duct. In the proximal segments ciliated cellsassist in filtrate movement. Histochemical evidence that macromolecule andactive transport mechanisms are present (El-Sayed & Safer, 1985) is supportedby the anatomical evidence. Wandering cells that may be phagocytic are foundhere and in the proximal tubules of the body kidney (El-Sayed & Safer, 1986).The distal tubule is usually absent in marine teleosts and variably present inthose which are euryhaline. That of P. waltoni has fewer mitochondria and lesselaborate folding than euryhaline fishes adapted to sea water. While there waslittle enzyme activity in the distal tubule (El-Sayed & Safer, 1985), prior tosacrifice the fish were not adapted to freshwater and the distal tubule wasotherwise structurally designed for secretory activity. Furthermore, the distalsegment also contained many microbodies, the function of which is not yetknown (Safer & El-Sayed, 1985).
Also in the head kidney is the homologue of the adrenal gland. In Periophthalmus viridis [?] the suprarenal or chromaffin (medullary) cells are interspersed with the interrenal (cortical) cells that are located mainly around theposterior cardinal vein. Only adrenaline could be detected (Banerji, 1973).
REPRODUCTION, DEVELOPMENT AND GROWTH
REPRODUCTION
The most detailed study of the reproductive cycle in mudskippers is that ofBoleophthalmus dussumieri from Bombay (19°N) (Mutsaddi & Bai, 1970). Theperiod of active maturation (gonadal stages IV—VI) lasts from February to Mayin males and March to June in females and the fish spawn once a year over theperiod July to September. Further north, in Korangi Creek (25°N) however, B.dussumieri (Hoda, 1986a) and B. dentatus [ = B. dussumieri] (Hoda & Akhtar,1985) spawn twice a year, first in April to May and then in July to September.Working at an intermediate latitude (Jodia coast, 22°N), Soni & George (1986)comment that B. dentatus spawns once a year in January to February. Latitudinal differences are clearly inadequate to reconcile the disparate observationsand further work is necessary. The fish in the population sampled from Korangi(Hoda, 1986a) were smaller than those from Bombay (Mutsaddi & Bai, 1970)and reached (50%) maturity at about 70 mm. Fish from Bombay attained

516 DAVID A. CLAYTON
maturity at 96-110mm. Similarly, the fecundity of Korangi specimens waslower (970-4113) than that of the females from Bombay (1028-7197). Fecundity of 15-50 g B. pectinirostris was much higher at 10 000-23 000 (Zhang etal., 1989) and in China (Fujian coast, 24°N) this species spawns only once ayear (Xie & Zhang, 1990). Direct comparison between the Indian and Chinesedata is difficult because different measures (length, weight) of size were used. Asweight relationships are unreliable (see p. 517), both measures shouldbe givenin future. In the Ganges estuary at Alampur in west Bengal (22°25'N: 86°40'E),Acentrogobius, Boleophthalmus Periophthalmodon and Periophthalmus spawnbetween May or June and September or October. This is the monsoon periodwhen pH and salinity are reduced and turbidity, temperature and the standingcrop of plankton increases (Sircar & Har, 1975).
Harms' (1935) supposition of viviparity in Boleophthalmus and Periophthalmusfrom Batavia (Jakarta) needs verification, for in other species eggs are laid ina burrow. The adhesion of eggs to the walls and roof of the egg chamber isachieved by filamentous attachment threads in P. chrysospilos (Harms, 1929),P. kalolo (Magnus, 1972), P. cantonensis [ = P. modestus] (Kimura, 1958),Boleophthalmus boddarti (Jones, 1937) and B. pectinirostris (Chen & Ting,1984; Liao et al, 1973; Zhang et al, 1987, 1989; Zhang & Zhang, 1988;Hong et al, 1988). According to Brillet (1976) the eggs of Periophthalmussobrinus [ = P. argentilineatus] are without threads, but as the eggs were onlycollected from burrows, it is possible that they were not noticed. Similarly,Asano (1936) omitted to mention their presence despite presenting a figureof pre-hatch eggs on the wall of the egg chamber of Periophthalmus sp. [ =P. modestus], a species which clearly possess them (Kimura, 1958).
DEVELOPMENT
Periophthalmids appear to require a longer period to hatch than do boleoph-thalmids. Hatching times for P. modestus are reported as 104 h (Kimura, 1958)and 170 h (Kobayashi et al, 1971) after fertilisation. The hatching period forPeriophthalmus sobrinus [ = P. argentilineatus. ] as calculated from data inBrillet (1976) is 115 h. Liao et al (1973) found that the eggs of Boleophthalmuspectinirostris hatched between 66 h and 86 h after fertilisation, data that aresupported by Chen & Ting's (1984) value of 66 h and by the value of 88 h givenby Zhang et al (1989). Shorter hatching times were due to higher rearingtemperatures, which may also have contributed to larval mortality. The researchof the last three groups of authors and of Chen (1976, 1982) and Qiu (1989) hasbeen directed towards the artificial propagation of larval fish for commercialfarming. Previously, Pearse (1932) using animals ready to spawn, had littlesuccess in fertilisation of eggs and none in hatching eggs. Kimura (1958) wasunsuccessful in inducing ovulation in Periophthalmus modestus by the use offrog cerebral hypophysis extract. Greater successwas achievedin Boleophthalmuspectinirostris where injections of carp pituitary and synchorin (Liao et al,1973), human chorionic gonadotrophin (HCG; Chen & Ting, 1984) and HCGand luteinising hormone releasing hormone and pimozide (Hong & Wang, 1989)and combinations thereof (Zhang et al, 1989) produced varying degrees ofsuccess. In all cases milt was provided from surgically removed testes.
Only Zhang et al. (1987, 1989) systematically controlled egg rearing conditions and reported that the optimum temperature and salinity for hatching were

MUDSKIPPERS 517
28°C and 15-25%o respectively. Outside these values hatch rates droppeddramatically. The failure of eggs to hatch at higher salinities is undoubtedly acontributing factor in the disparity between the potential and the realised nicheof these species. In the Red Sea Periophthalmus is absent from the mangrovesof the Sinai, an apparently suitable habitat (Fishelson, 1971; Por et al, 1977),but where salinity can reach 40-70%o (Por, 1974).
Larval stages successfully hatched from eggs are illustrated by Kimura(1958), Kobayashi etal (1971), Zhangetal. (1987), Hongetal (1988), Zhang& Zhang (1988), Zhang et al (1989) and Yhang & Zhang (1990).
Both Kobayashi et al (1972) and Zhang et al (1987, 1989) were successfulin rearing larval fish beyond the fifth day and present feeding regimes for thedeveloping larvae. In both cases, the diet was similar, but diverged as theperiophthalmid was offered and began to take meat between 40-50 days afterhatching when the 14-15mm larvae became substratum bound and amphibious(Kobayashi et al, 1972). In Boleophthalmus pectinirostris the equivalent ageand total length for the young to become benthic was 42 days and 20 mm(Zhang et al, 1989). Field observations support these data on the size at whichmudskippers become amphibious. The smallest amphibious Periophthalmusargentilineatus found in the mangrove are 11-13 mm (Brillet, 1976). Seasonalcollection of 15-30 mm Boleophthalmus pectinirostris fry from the mudflatswas the only way to obtain stocks for commercial rearing until successful eggculture was achieved (Chen, 1976). The fry were placed in specially preparedponds of 0.1-1.0 hectares at densities of between 30-50 000 fry-hectare"1.Beforethe fish reach a size suitable for sale the ponds requiredcareful pre- andpost-stocking management over 1-2 years (Chen, 1976). As fish growth isgreatest above 28°C (Chen, 1982) tropical pond culturing could reduce thismanagement period (Macintosh, 1982).
GROWTH
The only data on longevity of mudskippers is for Boleophthalmus dussumierifrom the west coastof the Indian continent where, from a study of otolith ringsof B. dentatus [ = B. dussumieri], Soni & George (1986) showed that 130 mmtotal length fish were 2 years old. Both Hoda (1986b, 1987) and Soni & George(1986)present length/weightrelationships expressed in the form of the equationW= aLb, but the analyses are not comparable because Soni & George usedtotal length (TL) and Hoda used standard length (SL). Furthermore, the equationpresented in Soni & George (1986) is clearly incorrect. The fish in the sampledpopulation ranged from 20-133 mm TL and 0.1-10.5 g, yet the equation givesthe weight of an 80 mm fish as 256 g. Hoda also gave equations for therelationship between SL and TL but these also seem unreliable because the slopeand intercept values in the equations for the combined male and female data setsare at variance with those of the separate (male and female) data sets. This ismost obvious in the equation for B. dentatus where similar intercept and slopevalues for male (6.21, 0.734 respectively) and female (6.184, 0.730) changeconsiderably in the combined data set (6.957, 0.743). Additionally, separateanalyses for B. dussumieriand B. dentatus were presented (Hoda, 1986b, 1987)and the latter name is a junior synonym of the former (Murdy, 1989). Moreinformation on length/weight relationships would be of value to other areasof investigation, for size related changes in structure or function complicate

518 DAVID A. CLAYTON
comparative studies (see sections on Osmoregulation, p. 524, and Respiration,p. 532). The data on the range of standard length of the specimens Murdy (1989)examined in his taxonomic revision of the Oxudercinae provide some guidelinesfor the limits of comparisons between species (see also p. 540).
EXCRETION
As amphibious fishes, the mudskippers' excretory mechanisms are particularlyinteresting, but as they are only just being revealed, the full potential of thisaspect of the fishes' physiology in relation to their degree of amphibiousness hasyet to be fully explored. Basic information on the rates of excretion undercomparable conditions is required to complement the available and sometimescontradictory data (Table I). Allowing for differences in experimental conditions, there is still considerable variation in the rates of excretion of ammoniaand urea. Although retaining similar percent levels of urea excretion, the sameauthorities working on the same species show several-fold changes in the absolutelevels of ammonia and urea excretion {B. pectinirostris and Periophthalmusmodestus: Morii et al., 1978, 1979 in Table I). The ammonia: urea ratio {% ureaTable I) shows no correlation with the fishes presumed water dependence{Scartelaos > Boleophthalmus > Periophthalmus). Unfortunately, measurementswere made at a variety of salinities which has a marked influence on this ratioas shown in P. modestus by Iwata et al (1981) (Table I). Additionally, morestudies need to be done on Scartelaos and Pseudapocryptes, especially as thelatter has a response to the drying out of its habitat that is similar to lungfishes(Hora, 1935a, b). Furthermore, because there are several closely related generawithin a single subfamily, better comparative material can be obtained forPseudapocryptes than for any lungfish.
The basic issue in excretion has centred on differences in nitrogen metabolismbetween the mudskippers' aquatic or surface phases of activity. In marine teleoststhere are a number of waste nitrogen compounds and in mudskippers over 96%is in the form of ammonia and urea (Morii et al., 1978). More than three-quarters of waste nitrogen is excreted by the gills, a pathway presumably notavailable to a mudskipper out of water (Gregory, 1977). Setting the experimentalparadigm, Gordon et al. (1969) compared the 24 h rate of ammonia and ureaexcretion between starved Periophthalmus sobrinus [ = argentilineatus orkalolo] which had been confined in sea water for 24 h with that of similar fishwhich had spent the first 12 h out of water before being returned to sea water.They found no differences in these 24 h rates. At this point the fish exposed toair had only been back in sea water for 12 h, but when the excretion rate of bothcompounds was calculated at the end of 24 h in sea water, it was found to bedouble that of the 24 h rate for fish maintained continuously in sea water. Thissuggested to Gordon et al (1969) that nitrogenous waste, accumulated in thebody during aerial exposure, was released on return to water. Further calculation demonstrated that while the rate of ammonia production only doubledwhen the fish was out of water, there was a 3.5 fold increase in the rate ofurea production (Gordon et al., 1969; Gordon, 1970). This shift of the ammonia: urea ratio in the direction of urea was also shown for P. modestus [asP. cantonensis] and Boleophthalmus pectinirostris (Morii et al, 1978).Gregory (1977) showed that in sea water, Periophthalmus novaeguineaensis [asP. expeditionium], P. gracilis and Scartelaos histophorus all had comparable

Table I
Ammonia and urea excretion in mudskippers. For ease of comparison all data are converted to /ig-g-24 h"1. aData from 24 hgraphical presentation.
Species Ammonia-N Urea-N % Urea Fish Temperature Fed (F) or % Reference(Mg-g-24h_l) (Mg-g-24h_l) size
(g)(°C) starved
(S)sea
water
Scartelaos histophorus 156 37 19 1-5 Room s 100 Gregory, 1977
Boleophthalmus 34 4 10 33-46 20+2 s 25 Morii et al., 1978pectinirostris 104 11 9 36-45 28 + 2 s 25 Morii et al., 1979a 2
aoGO
Periophthalmus gracilis 184 31 14 1.0 Room s 100 Gregory, 1977
P. novaeguineaensis 259 129 33 1-5 Room s 100 Gregory, 1977
P. sobrinus 165 242 59 0.5-15.0 26.5 + 3.5 s 100 Gordon et al., 1965Gordon et al., 1969
m
on
259 40 13 3.0-15.0 26.5±3.5 s 40 Gordon et al., 1965
P. modestus 2117 4973 55 0.5-5.0 21+2 s 100 Gordon et al., 19782150 8467 66 0.5-5.0 21+2 F 100 Gordon et al., 1978106 11 9 4.0-6.3 20±2 s 25 Morii et al., 1978440 60 2 3.8-6.5 29±2 s 25 Morii et al., 1981a222 55 20 1.0-3.0 20±1 s 20 Iwata et al., 1981167 40 19 1.0-3.0 20±1 s 50 Iwata et al., 1981101 62 38 1.0-3.0 20±1 s 80 Iwata et al., 198123 28 55 1.0-3.0 20+1 s Air Iwata et al., 1981

520 DAVID A. CLAYTON
levels of ammonia excretion to that of Periophthalmus sobrinus, but considerably lower levels of urea excretion (Gordon et al., 1965). Urea excretion,expressed as a percentage of total excretion in these three species, is thereforeconsiderably lower than that of P. sobrinus (Gregory, 1977; see also Table I).No air exposure trials were done, but Gregory (1977) found that the fish weredeficient in enzymes necessary for the production of urea by the ornithine-ureacycle. He suggested that, alternatively, the urea could have been produced bypurine catabolism because the enzymes necessary for thebreakdown of uricacidare present in the periophthalmids. Neither this suggestion (seealsoChew & Ip,1987) nor the supposition by Gordon et al (1968) that P. sobrinus exhibits ashift towards ureotelism while out of water have found support.
When nitrogen excretion is monitored over 4-5 days in both P. modestusandBoleophthalmus pectinirostris the general responseis that urea levels remainfairly constant while ammonia initially declines and thereafter increases (Moriiet al., 1978). This pattern does not depend on whether the fish are in or outof water although the absolute levels of excretion in water are several timesgreater. A compensatory increase in the rate of nitrogenous excretion when fishreturn to water (Gordon et al, 1968) has been partially confirmed in moredetailed studies. Ignoring the high excretory levels reported for Periophthalmusmodestus in comparison with other authors (Table I), Gordon et al. (1978) keptfish out of water for 16.5 h and found that the rate of urea-N excretion showeda three-fold increase above control (in sea water) rates for the first 4 h afterreturn to sea water and a two-fold increase in ammonia-N excretion for the first5 h. Working witha 6-h sampling programme Morii etal. (1979) monitored theexcretion on return to water of P. modestus and Boleophthalmus pectinirostrisafter 12, 24, 36, or 48 h out of water. For the shortest period out of water (12h), for example there was an approximate 1.5 fold increase in ammonia-Nexcretion above control rates in both Periophthalmus modestus and Boleophthalmuspectinirostris during the first 6 h back in water. Contrary to Gordon et al.(1978), no change in urea rates were found. As the period out of water increasedthere was an increase in the rates of excretion of both products during this initialperiod (Morii et al., 1979). This trend was most pronounced for ammonia inB. pectinirostris, but in the absenceof statistical analysis the significance of anyof these trends is not known.
Morii et al. (1978) also attempted to show the partitioning of urea andammonia excretion between the gills, skin and cloaca. When Periophthalmusmodestuswas out of water, there was no great difference between concentrationsexcreted through the skin and cloaca. In anuric (cloaca closed) P. modestus andBoleophthalmus pectinirostris in water the nitrogen concentration excreted bythe skin (body behind head) was numerically higher in urea than ammonia. Thereverse was found in the gills (actually head and gills). Verification of these dataand conclusions are required, if only because the data show a 10-fold increaseof urea production by anuric fish when compared with that of normal fish inwater.
In the absence of any change in the rate of urea excretion on return to water,Morii et al. (1978) suggest that the ammonia accumulated during the period outof water was not converted to urea. The data on blood concentration are consistent with this interpretation, although again statistical confirmation would bebeneficial. In Periophthalmus modestus urea concentration was lower in fish outof water than in fish in water, and decreased with longer periods out of water.

MUDSKIPPERS 521
For Boleophthalmus pectinirostris the data show an opposite trend, but in bothspecies, fish out of water exhibit a lower ratio of urea-N/ammonia-N than thosein water (Morii et al, 1979). These results were replicated by Morii (1979) andextended for 5 days instead of two and show a broadly similar distribution andchange with time of the levels of urea-N and ammonia-N in the liver, muscle,skin and gill tissues. After 3 days out of water, ammonia levels increaseddramatically in muscle, skin, and especially gill tissue, of B. pectinirostris. It islikely that these data represent pathological changes because fish in this groupdied (Morii, 1979). Similarly, Periophthalmus modestus that died in sea waterare also included in the data sets. Certainly Boleophthalmus pectinirostris islikely to have a much lower tolerance to excretory product level than Periophthalmus modestus as both urea and ammonia concentrations in the latter areseveral times greater than in the former species (Morii, 1979; Morii et al,(1979). The absence of any accumulation of urea in any of the tissues of eitherfish can, however, be taken as confirmation that ammonia is not converted tourea during the period out of water (Morii, 1979). Certainly in P. modestus keptout of water the distribution of urea was more uniform among the tissues(muscle 34 jag-g-day-1; liver 39 /xg-g-day-1; 5-day data, Morii, 1979; muscle65 /xg-g-day"1; liver 29 /xg-g-day-1; 7-day data, Iwata et al., 1981) thanammonia where the concentration was greatest in muscle (142, 186 ^g-g-day-1)and lowest in liver (52, 59 /xg-g-day-1, Morii, 1979; Iwata et al., 1981respectively). As the liver accounts for less than 2% and the muscle for 48%of the body weight in P. modestus (Iwata, 1988), muscle is likely to be the majorstorage site of ammonia during the period out of water (Iwata et al, 1981).
In the absence of ureotelism, the detoxification of nitrogenous waste appearsto be achieved through the metabolism of non-essential amino acids which aremore usually thought to be associated with osmoregulation (Iwata et al, 1981).In comparison with fish kept in 20% sea water, and paralleling the change inammonia concentration, there was a significant doubling of total free aminoacids (FAA) in the muscle of P. modestus out of water. Markedly increasedlevels of the non-essential amino acids alanine, aspartate and glutamate andespecially taurine were found, such that the FAA comprised about 70% of thetotal increment of nitrogen (above that of fish in water); ammonia and urea madeup only 21% and 4% respectively (Iwata et al, 1981).
The enzyme glutamate dehydrogenase (GDH) also plays a central role inremoving ammonia from the tissues of fish subjected to conditions of watershortage including being out of water (Iwata & Kakuta, 1983). This enzymecatalyses reductive animation of a-keto-glutarate and Iwata et al. (1981) andIwata & Kakuta (1983) measured its activity in the reductive amination (orglutarate forming) direction in the skin, gills, muscle and liver of P. modestus.In fish kept out of water the activity of muscle GDH was four times higher thanin 20% sea water. Liver GDH responded similarly, but was about five timeshigher inactivity than that of muscle (Iwata etal., 1981; Iwata & Kakuta, 1983).The kinetic constant Km (substrate concentration at which the reaction rate ishalf maximum) of GDH for NH4+ in muscle is lower than that of liver, skinand gill GDH, permitting a rapid response of the enzyme to tissue ammoniaconcentration.
GDH activity in the liver was also greater than in the muscle (and gills) ofBoleophthalmus boddarti and Periophthalmodon schlosseri (Chew & Ip, 1987).Enzyme activity in the reductive amination direction was 16 {Boleophthalmus

522 DAVID A. CLAYTON
boddarti) and 20 {Periophthalmodon schlosseri) times greater than in the oxidative deamination (or ammonia forming) direction indicating that liver GDHis not involved in ammoniagenesis, but is likely to be central to the regulationof amino acid pool in the whole body (Chew & Ip, 1987). The metabolicpathways for the production of ammonia from other FAA is also discussed byChew & Ip (1987). Chew & Ip (1990) confirm that liver GDH activity is higherthan that of muscle in Boleophthalmus boddarti and also showed that GDHactivity was greater, but of a similar pattern in Periophthalmus chrysospilos. ForBoleophthalmus boddarti, however, the enzyme activity in the aminationdirection was 32 times greater than in the deamination direction: double thatreported previously (Chew & Ip, 1987). It is difficult to relate the doubling ofthe amination/deamination ratio to salinity since Chew & Ip (1990) found nodifference within liver or muscle GDH activity between fish exposed for 3 daysto either 10% or 80% sea water. In the earlier experiment, the fish were maintained in 50% sea water for 1 day prior to enzyme analysis (Chew & Ip, 1987).This is possibly why the earlier paper was not mentioned in the later one.
Aspartate transaminase (or glutamate-oxaloacetate transaminase, GOT) and,to a much lesser extent, alanine transaminase (or glutamate pyruvate transaminase, GPT) activity was also much greater in the liver than in the gills ormuscle of B. boddarti and Periophthalmodon schlosseri (Chew & Ip, 1987).While the activity of GOT was also higher than that of GPT in Periophthalmusmodestus, their activity was higher in the muscle rather than the liver (Iwata,1988). The relative activity of these enzymes suggest that aspartate (= FAAaspartic acid) rather than alanine is the major substrate for ammoniagenesis inthe muscle and liver mitochondria of Boleophthalmus boddarti (Chew & Ip,1987) and Periophthalmus modestus (Iwata, 1988) and the liver of Periophthalmodonschlosseri (Chew & Ip, 1987). In support of this suggestion, the aspartatecontent was significantly less than that of alanine in the muscle of Boleophthalmusboddartiand Periophthalmodonschlosseri (Siau & Ip, 1987). Under thecatalytic action of mitochondrial GOT, aspartate and a-ketoglutarate formoxaloacetate and glutamate, the latter being deaminated by GDH to yield a-ketoglutarate and ammonia. In muscle of P. schlosseri the FAA glutamine is themost effective substrate for ammoniagenesis, but the low glutaminase activity(which could catalyse ammonia and glutamate formation) and other datasuggested to Chew & Ip (1987) that an alternative pathway for ammoniagenesisinvolving an initial transamination to a-ketoglutaramate which is hydrolysed toa-ketoglutarate. A more parsimonious explanation of the low glutaminaseactivity found by Chew & Ip (1987) may be that, contrary to expectations, themuscles were actually subjected to ammonia loading or stress.
By comparing Periophthalmus modestus out of water with those subjected toan increased ammonia load by being kept in NH4C1 solution, Iwata (1988)further confirmed the role of FAA in ammonia detoxification. The increases in
the qualitatively similar amino acids (glutamate, alanine, glutamine and glycine)under both conditions strongly suggest that their production was triggered byammonia rather than dehydration stress although the impairment of gas exchangemay be the reason for the greater increase in alanine in the ammonia-stressedfish (Iwata, 1988). In contrast to the situation in Boleophthalmus boddarti andPeriophthalmodon schlosseri (Chew & Ip, 1987), glutamine synthetase in themuscles of Periophthalmus modestus was actively involved in the removal ofammonia by catalysing its use in the synthesis of glutamine from glutarate (Iwata,

MUDSKIPPERS 523
1988). Chew & Ip (1987) maintained their fish in 50% sea water but, as Iwataet al (1981) demonstrated, salinity changes have consequences for levels ofammonia and urea excretion. This raises the general observation that fish rearingconditions are extremely importantin determining the adaptiveresponseof enzymesystems to physiological change. It is not clear why most studies (Table I) haveused fish adapted to 20% sea water unless it is to maximise ammonia and ureaproduction between the adapted fish and those kept out of water. Following theoriginal experimental paradigm, biochemical investigation of fish returning towater after being emersed would be most instructive. Whatever further details arisefrom such studies, including the role of taurine (Iwata etal, 1981; Iwata, 1988),it is clear that mudskippers have a greater tolerance than other water-breathingfish including gobies. This ammonia tolerancecan be accountedfor by their abilityto synthesise non-essential amino acids (Iwata, 1984, 1988). P. modestus waskept in very different conditions (Iwata, 1988) to the other species (Chew & Ip,1987) and it is possible that glutamate synthetase also becomes active in speciesof Boleophthalmus, Scartelaos and Periophthalmodon under conditions ofextreme ammonia stress. Further comparisons that could be made include theassay of brain enzyme activity. In Periophthalmus modestus there is a powerfulGDH-glutamine synthetase ammonia detoxification system (Iwata, 1988). Itwould be instructive to know if this is a general mechanism or specific to thosespecies which have a high tissue tolerance for ammonia.
SALINITY TOLERANCE AND OSMOREGULATION
SALINITY TOLERANCE
Mudskippers can generally be regarded as euryhaline and percent mortalityis the simplest and crudest measure of mudskippers' response to changes insalinity. Typically, groups of fish have been placed in water of salinity rangingfrom 0—100% sea water for a number of days during which deaths weremonitored (Table II). Leaving aside ethical issues, the execution of even thissimple procedure leaves much to be desired and experimental procedures needto be standardised and carefully described before good comparative data can beobtained. The timing of analytical procedures used to investigate any physiological mechanisms of adaptation to changing environmental conditions isimportant, if only in determining the sequence of such mechanisms. Vagueexperimental protocols (Bhan & Mansuri, 1978a; 5, 10 or 15 days acclimation?), experimental design (Gordon et al, 1965; adaptation period variedbetween 2 and several days), or independent non-comparable measures ofresponse (Mansuri & Bhan, 1978; Chew & Ip, 1990; Table II) at best simplyconfuse the analysis. At worst, and given the reported mortality of fish indifferent salinities as described below, it may mean that pathological changes arebeing observed. Mansuri & Bhan (1978) report high mortality of Periophthalmusdipes in 50 and 70% sea water, but little in 0% and 100% sea water. Converselyat 0% and 100% salinities, P. chrysospilos and Boleophthalmus boddartiexhibited their highest mortality but survived well (only 8% mortality) in 50%and 80% sea water (Chew & Ip, 1990). While Mansuri etal (1982) showed thatB. dussumieri [as B. dentatus] had a better survival rate at all salinities thanPeriophthalmus dipes, mortality in both species was greatest in 50% and 70%
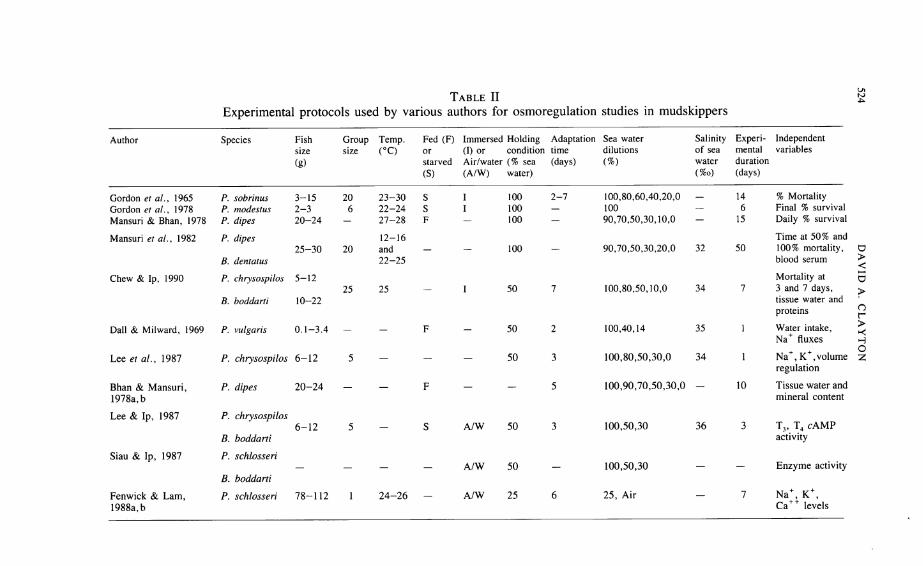
Table II
Experimental protocols used by various authors for osmoregulation studies in mudskippers
Author Species Fish Group Temp. Fed (F) Immersed Holding Adaptationsize size (°C) or (I) or condition time(g) starved Air/water (% sea (days)
(S) (A/W) water)
Sea water
dilutions
(%)
Salinityof sea
water
(%o)
Experimental
duration
(days)
Independentvariables
100,80,60,40,20,0100
90,70,50,30,10,0
—
14
6
15
% MortalityFinal % survival
Daily % survival
90,70,50,30,20,0 32 50
Time at 50% and
100% mortality,blood serum >
100,80,50,10,0 34 7
Mortality at3 and 7 days,tissue water and
proteins
S>
nr
100,40,14 35 1 Water intake,Na+ fluxes
>
HO
100,80,50,30,0 34 1 Na+, K+,volumeregulation
100,90,70,50,30,0—
10 Tissue water and
mineral content
100,50,30 36 3 T3, T4 cAMPactivity
100,50,30- -
Enzyme activity
25, Air-
7 Na+, K+,Ca++ levels
Gordon et al., 1965Gordon et al., 1978Mansuri & Bhan, 1978
Mansuri et al., 1982
Chew & Ip, 1990
Dall & Milward, 1969
Lee et al., 1987
Bhan & Mansuri,1978a, b
Lee & Ip, 1987
Siau & Ip, 1987
Fenwick & Lam,1988a, b
P. sobrinus 3—15
P. modestus 2—3
P. dipes 20-24
P. dipes25-30
B. dentatus
P. chrysospilos 5—12
B. boddarti 10-22
P. vulgaris 0.1—3.4
P. chrysospilos 6—12
P. dipes 20-24
P. chrysospilos
B. boddarti
P. schlosseri
B. boddarti
P. schlosseri
6-12
78-112
20
6
20
25
23-30 S
22-24 S
27-28 F
12-16
and —
22-25
25 -
100
100
100
100
50
2-7
—
F—
50 2
- - -50 3
-
F- -
5
-S A/W 50 3
- -
A/W 50
24-26 A/W 25 6

MUDSKIPPERS 525
sea water. Additionally both species had the greater survival times in winter(12-16°C) than in the summer (22-25°C). This apparent confirmation of theearlier data for Periophthalmus (Mansuri & Bhan, 1978) must be offset againstthe large discrepancy between the two data sets. Mansuri & Bhan (1978) foundthat some fish survived for 15 days in both salinities (50% sea water, 90%mortality; 70% sea water, 70% mortality) whereas in later experiments(Mansuri et al, 1982) all fish died within 2 (summer) or 4 (winter) days.
If, as the data on summer and winter survival suggests, temperature is animportant variable, then the discrepancy is even more surprising because the fishsurvived longer at high temperatures (27-28°C; Mansuri & Bhan, 1978).The specific identity of P. dipes is also unclear. P. dipes is a synonym of P.argentilineatus, but at 12—16 cm and 20-24 g, the specimens seem to be toolarge to be this species.
The difference between the results obtained by Chew & Ip (1990) and Mansuri& Bhan (1978) could be related to the experimenters' choice of holding conditions for the fish prior to the salinity change. P. chrysospilos and Boleophthalmus boddarti were able to move between air and 50% sea water (= 17%osalinity; Chew & Ip, 1990), while Periophthalmus dipes was maintained in100% sea water of unknown salinity (Mansuri & Bhan, 1978). As P. dipes cantolerate salinity changes in the range of 24%o—47%o salinity (Bhan & Mansuri,1978a), the acclimation salinity is of some importance. Where P. sobrinus[ = kalolo or argentilineatus] all died within 3 days of being transferred from100% sea water (= 34%o salinity), if they were first acclimated in 20% seawater for 6 days, there was no mortality (Gordon et al, 1965). Especially ingroups, mortality among totally submerged mudskippers may be due toasphyxiation rather than salinity stress (cf. Hora, 1935b).
OSMOREGULATION
The body water content of mudskippers is maintained at constant levels over awide range of salinities. Only P. chrysospilos in air and deionised water showedany significant change (decrease and increase respectively) in water content after3 days of exposure to a variety of salinities (Lee et al., 1987), supporting dataon P. sobrinus where, however, the significant increase of muscle water contentin fresh water over the 100% sea water condition was reported only frommoribund fish (Gordon et al, 1965). Nevertheless, P. sobrinus [ = kalolo orargentilineatus] showed a 10-15% weight increase following transfer to allsalinities, which Gordon et al, (1965) attributed to the immersion of the fish.For fish directly transferred to fresh water, the increase was 20% and the fishdied within 3 days, but if previously adapted to 20% sea water, there was nochange in weight (Gordon et al, 1965). In air in direct sunlight, P. sobrinuslost 12% body weight before dying after 50 min, whereas in shade and highhumidity the body weight loss could be doubled and the fish survived for 24 h(Gordon et al, 1969).
In P. dipes [?= P. argentilineatus] the percent water content of muscle, gill,liver, heart and kidney remained stable at all salinities including fresh water,whereas in comparison with the levels in 100% se? water, sodium, potassium,and to a lesser extent, calcium, but not phosphorus, were reduced in these tissues(Bhan & Mansuri, 1978a). The total protein and fat content of red and whitemuscle, kidney and gills was reduced, however, especially at lower (< 30%)

526 DAVID A. CLAYTON
salinities (Mansuri & Bhan, 1978). In most salinities white muscle used glycogenrather than fat as the main energy source (Bhan & Mansuri, 1978b) and succinicdehydrogenase activity was lower than in other tissues (Bhan & Mansuri,1978c). Gills of fish adapted to 10% sea water and fresh water exhibited thehighest level of succinic dehydrogenase activity (Bhan & Mansuri, 1978c).
The regulation of plasma osmolarity (and NaCl concentration) was nearlyperfect in P. sobrinus kept for up to 6 days in 20—100% sea water (Gordon etal, 1965). Only dying fish that had been directly transferred to fresh watershowed a significant dilution of their blood. On the basis that when previouslyadapted to 20% sea water, a single fish in fresh water maintained osmolarity inthe normal range, Gordon et al (1965) concluded that the species was probablya perfect osmoregulator. No other mudskipper approaches this level of performance. After 3 days in 100% sea water plasma osmolarity of P. chrysospilosreached 440 mmol-kg"1 a significant increase above the 324 mmol-kg"1control level offish free to enter 50% sea water (Lee etal, 1987). No significantchanges were found for fish kept in 30, 50 or 80% sea water (Lee et al., 1987),but after 7 days in 80% sea water the 315 mosmolal value reported by Chew& Ip (1990) for P. chrysospilos kept under similar control conditions represented a significant increase above the 299 mosmohal control level. Furthermore,the plasma osmolarity of 260 mosmolal of fish in 10% sea water indicated asignificant dilution of the blood. Chew and Ip (1990) did not test their fish in100% sea water but additionally showed that plasma osmolarity of Boleophthalmusboddarti (219 mosmolal) was significantly less than that of Periophthalmuschrysospilos and also decreased (to 252 mosmolal) in 10% sea water after 7days. However, as the decrease in 10% sea water was greater in P. chrysospilos,and Boleophthalmus boddarti was able to maintain its plasma osmolarity (275mosmolal) in 80% sea water, it is likely that extracellular aniso-osmoticregulatory mechanisms are more efficient in B. boddarti (Chew & Ip, 1990),supporting the suggestion by Mansuri et al. (1982) that Boleophthalmus was abetter regulator than Periophthalmus.
In comparison with the condition in 100% sea water, after a day Boleophthalmus boddarti showed increased plasma levels of sodium at all salinitiesexcept for fresh water where there was a decrease (Mansuri et al., 1982). Forpotassium and calcium there was no similar simple relationship between salinityand plasma concentration. Plasma potassium increased greatly in 50% and 70%sea water (the salinities in which most fish died) and calcium only decreased infresh water (Mansuri et al, 1982). Whether the increased plasma sodium isaccounted for by the reduced intracellular (tissue) levels (Bhan & Mansuri,1978a) is a moot point, but there was little correlation between intra- andextracellular levels of potassium and calcium (Bhan & Mansuri, 1978a; Mansuriet al., 1982). If only because of the absence of the units of measurement, the dataof Mansuri et al., (1982) must be questionable. Truly comparative data with asimilar control condition are not available, but in Periophthalmus chrysospilosafter 1 day plasma sodium levels were similar (80—82 mM) at salinities above(100%, 80% sea water) and below (30% sea water) the freely accessible 50%sea water control condition, where the plasma concentration was significantlyhigher at 92mM (Lee et al., 1987). After 3 days for sodium and 5 days forpotassium, however, a more stable state was reached in which plasma concentrations were higher than those of the control condition, such that after 5 dayssodium values were between 134-151 mM and potassium between 14-16.8 mM

MUDSKIPPERS 527
(values calculated from Figure in Lee et al., 1987). These data need reconcilingwith the osmolarity data presented in the same paper. In comparison with controlfish, a 3-day restriction to 50% sea water produced no change in osmolarity (Leeet al, 1987; Chew & Ip, 1990) while 3 days in 100% sea water significantlyincreased it (Lee et al, 1987). Plasma levels of sodium and potassium did notreflect this lack of change and were increased to similar levels above control inboth conditions. The removal (in 50% sea water) or addition (in 100% sea water)of other osmotically active substances must be implicated. The presence of NPS(Ninhydrin Positive Substances) including FAA in tissues is clearly related tothe regulation of intracellular fluid content, but they may also be involved inplasma osmotic regulation, although Lee et al, (1987) found no significantchanges in NPS levels of the liver and muscles of the fish after 3 days. Fish inhigher salinities for 7 days, however, showed higher levels of NPS in bothtissues. There was a similar direct relationship between NPS and salinity after7 days in P. modestus (Iwata et al, 1981). Chew & Ip (1990) confirm this 7-dayfinding and additionally showed that the decrease in plasma osmolarity of P.chrysospilos was accompanied by a decrease in the level of NPS in musclecompared with control fish. Furthermore, GDH activity in the aminatingdirection was significantly greater in fish maintained in 80% sea water than infish in 10% sea water for 7 days.
The differences between short term (1 day) and long term (7 days) plasmalevels of sodium and potassium were related to changing membrane permeabilitysuch that both sodium influx (k) and efflux (k) rate constants increased withincreasing salinity; the latter linearly (Lee et al, 1987). The efflux rate constantof P. argentilineatus [as P. vulgaris] also showed a linear relationship tosalinity (Dall & Milward, 1969), but the slope was much shallower, possiblybecause the fish were ligated, excluding urinary excretion. There was an effluxof 15—20% from the body and fins, the remainder was from the branchialregion. While individual rates were variable , drinking of P. argentilineatus wasinversely related to salinity such that the rate in 100% sea water (35 %o) wastripled in 14% sea water (5%o; Dall & Milward, 1969). In comparison with acardinal fish, the mudskipper had similar efflux rates but a reversed drinkingregime and Dall & Milward (1969) concluded that the wide salinity tolerancecoupled with behavioural adaptations were adequate to account for salt regulation in Periophthalmus. For example, adaptation to low salinities could beachieved by remaining out of water for long periods so that hydration could beoffset by evaporation (Dall & Milward, 1969).
Studies of osmoregulation and excretion frequently used animals that were notfed prior to sacrifice while in captivity, and care is required in interpretingresults from starved fish. The depletion of muscle glycogen reported in P. dipes[? P. argentilineatus] as a result of exposure to different levels of salinity (Bhan& Mansuri, 1978c) was also reported for Boleophthalmus boddarti as a responseto starvation (Lim & Ip, 1989). In Bhan & Mansuri's (1978c) experiments withPeriophthalmus dipes the fish were fed while in captivity (Table II) but this isthe exception. If only because of the variety and inter-relationships between thebiochemical pathways concerned, the effects of fasting need to be assessedindependently of those of osmoregulation and excretion.
In P. chrysospilos the short term (1 day), regulation of plasma sodium andpotassium, as evidenced by the increased Na+ and K+ efflux rates with increasing salinity, was matched by increased Na+K+ATPase activity in the

528 DAVID A. CLAYTON
gills. This active mechanism that requires energy to regulate ionic control wasfurther reflected in the increased oxygen consumption rate at higher salinities(Lee etal, 1987).
Plasma osmolarity increases in mudskippers out of water, presumably becauseof dehydration, but varied experimental protocols again leave much to bedesired. Gordon et al. (1978) subjected P. modestus to rapid (20% body weightchange, h"1) and slow (2%h_1) dehydration in air at 25-28°C. After the fishhad lost 20% of their body weight in the latter condition there was an increaseof 40% and 60% in plasma osmotic and sodium concentrations respectively. Thefish dehydrated so rapidly that they became moribund and the experiments lastedonly 23 h, but the 24 h air exposure of P. chrysospilos did not result in anyweight loss and only a 16% increase in plasma osmolarity (Lee et al, 1987).Despite the difference between species, the (unspecified) conditions were apparently far less severe in the latter experiment. By constantly moistening filterpaper, Fenwick & Lam (1988a) were able to maintain Periophthalmodonschlosseri in air at 25 ± 1°C for 7 days and body weight and plasma sodium andpotassium concentration remained at control levels. Only calcium levels increased significantly and measurement of influx and efflux rates showed a netuptake of calcium by P. schlosseri in water, whereas in air the efflux rate waseffectively zero (Fenwick & Lam, 1988b). This suggests that the gills are theprimary site of calcium efflux and that P. schlosseri is able to performconsiderable cutaneous calcium transport. Being able to prevent the increase incalcium plasma concentration of fish maintained in air, it appears that calcitoninonly has a regulatory effect under conditions of hypercalcemia and is otherwisenot involved in osmoregulation (Fenwick & Lam, 1988a).
HORMONAL CONTROL OF OSMOREGULATION
Using histological evidence of the synthesis and utilisation of a variety of neuroendocrine glands in Boleophthalmus dentatus, Patel & Desai (1976) correlatedgland activity with environmental conditions at capture. They made monthlycollections of fish at low tide from July to October; half were sacrificed immediately and the rest after being kept on the shore until they had been immersedfor an hour by the flood tide. Examination of the hypothalamic neurohypophyseal complex, the caudal neurosecretory system, the corpuscles of Stannius,inter-renal cells and thyroid gland showed that their activities were correlatedwith the water temperature and salinity at sacrifice. Water pH was also measuredand found to vary in the same way as salinity, but was otherwise ignored.
The changes in neuroendocrine activity were dependent on the relative salinityand temperature of the tides and are best interpreted in terms of the ebb and floodtides of October against those of the remaining months. In comparison withflood tides, the ebb tides of October had a marginally higher salinity (28 %o,+0.6%o) and a lower temperature (26°C, -1.7°C) and all neuroendocrineglands, except the thyroid, were in a phase of synthesis. In the October floodtides this situation was reversed, such that products were being utilised whilethe thyroid, having been secretory, entered its phase of synthesis. For the Julyto September period the relationships between the activity of the neuroendocrineglands and environmental conditions were maintained, except that it was theflood tides that exhibited marginally higher salinity (median 36.9%o, range,25.6-37.2%o: + 1.6%o, 0.1-2.1%o) and lower temperature (27°C, 27-28°C:

MUDSKIPPERS 529
-2°C, 0.7-2°C). Patel & Desai (1976) relate the neuroendocrine response onlyto salinity, but in view of the wide salinity and temperature ranges that these fishcan tolerate, the level and speed of the response for such small environmentalchanges needs to be verified. Nevertheless, some evidence for thyroidal secretionat higher salinities and prolactin secretion from the eta cells of the rostral parsdistalis (Patel & Desai, 1976) at lower salinities, is provided by Lee & Ip (1987)who showed that plasma thyroxine (T4), 3,5,3'-triido-L-thyroxine (T3), prolactin and cyclic adenosine 3',5'-monophosphate (cAMP) are implicated incontrol of osmoregulation in Periophthalmus chrysospilos and Boleophthalmusboddarti. Furthermore, in comparison with fish in 15% and 100% sea water, thenuclear and cell sizes of prolactin—but not growth-hormone-secreting, cells ofthe pituitary of P. chrysospilos were considerably enlarged (Ogasawara et al.,1991).
Prolactin acts to reduce salt loss and in P. chrysospilos plasma concentrationincreased significantly in 30% and 0% sea water and in fish out of water.Whatever the mechanism of salt reduction induced by prolactin, it is difficult toaccount for the significantly reduced plasma concentration when the fish wassubmerged in 50% sea water. This was the same concentration as the controlcondition, and was not matched by an equivalent reduction in the other 'higher'salinity (100% sea water). In Boleophthalmus boddarti plasma prolactinconcentration only increased above control values in 100% sea water, but themechanism of prolactin action in hyperosmotic regulation is unclear. Relativeto the control condition, the level of T4 in Periophthalmus chrysospilos washigher in air and, irrespective of salinity, was reduced while the fish weresubmerged. T4 helps the fish to cope with terrestrial stress and is otherwise notinvolved in osmoregulation and the intermediate position of the T4 concentration in control fish is to be expected. In Boleophthalmus boddarti, T4 wasonly elevated in fish out of water and how terrestrial adaptation is mediated byT4 in either fish is unknown. T3 appeared to be unaffected by changing salinityin Periophthalmus chrysospilos while in Boleophthalmus boddarti it was higherin 100% sea water and reduced in 30% sea water and in air suggesting that itis of importance in osmoregulation in the latter but not the former species (Lee& Ip, 1987). Consistent with the function of plasma cAMP in stimulatingchloride secretion are the findings, in fully submerged fish, that levels increasewith increasing salinity in Periophthalmus chrysospilos (30, 50 100% sea water)and in Boleophthalmus boddarti in 100% sea water. The absence of anydifference between control cAMP levels where the fish is free to move into and
out of 50% sea water and those of B. boddarti submerged in 30% and 50% seawater is explained by the suggestion that these conditions approximate to thatof the natural habitat where the fish spends most of its time swimming along thewater's edge half submerged (Lee & Ip, 1987). This explanation was also offeredfor the absence of any change of plasma T4 concentrations in B. boddarti at allsea water concentrations, including 100%.
RESPIRATION AND RESPIRATORY SURFACES
HISTORICAL ASPECTS
The investigation of the mudskippers' respiratory capabilities was initiated byHigson (1889). Based on the observation that mudskippers always kept their tails

530 DAVID A. CLAYTON
in water he believed that these fish used caudal respiration. Crude experimentalsupport for this supposition was provided by Haddon (1889). He noted thatblood circulation to the tail in Periophthalmus was strong and showed that a fishwhich had its tail painted with gold size died while control fish did not (Haddon,1889). These finding were dismissed (Rauther, 1910; Harms, 1929; Hora, 1933;Das, 1935), but did draw attention to the role of cutaneous respiration in thesefishes.
Vascularisation of the skin epithelium necessary for cutaneous respiration wasdemonstrated (Rauther, 1910; Harms, 1929; Schottle, 1932) but, contrary toGraham's (1976) assertion, no mudskipper has any specialisation of the caudalregion for respiratory purposes. The general question of the partitioning ofrespiratory activity between the gills and accessory respiratory organs,however, continued to interest researchers. Das's (1933, 1934) work is anexample of the methodology adopted in early investigations of air breathingfishes. Like the airbreathers, Pseudapocryptes lanceolatus comes to the surfaceto swallow air and Das attempted to drown them by keeping them submergedand denying them access to the surface (Das, 1934). Whilst these mudskipperssurvived considerably longer than other species such as Clarias batrachus andAnabas testudineus, they still died. This was probably due to deoxygenation ofthe water (Hora, 1935b) although in some conditions, osmotic stress may beresponsible (Ogasawara etal., 1991). Nevertheless, the idea persisted that thesefish were obligate airbreathers and Willem & Boelaert (1937) used Rauther's(1910) finding of a vascularised diverticulum in the buccopharyngeal cavity ofPeriophthalmus to suggest that mudskippers utilise both the gills for aquatic andthe buccopharyngeal cavity for aerial respiration. Both modes of respirationwere considered to operate simultaneously irrespective of whether the fisheswere in or out of water. Thus, respiration in immersed P. papilio [ = P. barbarus]involved the presence of a supply of air from the surface. Willem & Boelaert(1937) argued that the lamellar arrangement of Arch I and its clear separationfrom the second arch supported this suggestion. Furthermore, eye retraction,which was often accompanied by slight bulging of the opercular region, wasthought to be a mechanism whereby the the two media, water and air, could bemixed. Eye retraction depressed the ceiling of the buccal cavity compressing thebuccopharyngeal air and moving the interface between the air and the watermeniscus.
The suggested presence of air in immersed fish has not been pursued, but thealternative suggestion, that water remained in the buccopharyngeal space duringterrestrial excursions (Stebbins & Kalk, 1961), was seriously considered. Theynoted that P. sobrinus gulped air on land and, following Willem & Boelaert(1937), discussed ways in which the air and water could be mixed to providean adequate oxygen supply to the gills. While Gordon et al. (1968) disputed thatP. sobrinus held water during terrestrial excursions, MacNae (1968a, b)believed that mudskippers did not need any accessory respiratory organs beyondtheir gills and dismissed the earlier (Rauther, 1910; Schottle, 1932) histologicalevidence. Later P. modestus [as P. cantonensis] was reported as maintainingbuccal water on land (Gordon et al., 1978), but the cineradiographic study ofSponder & Lauder (1981) showed conclusively that it was not necessary duringterrestrial excursions in P. barbarus [as P. koelreuteri]. What the exact role,if any, retained water plays in respiration or feeding (Sponder & Lauder, 1981)remains a moot point but in common with other marine fishes, mudskippers are
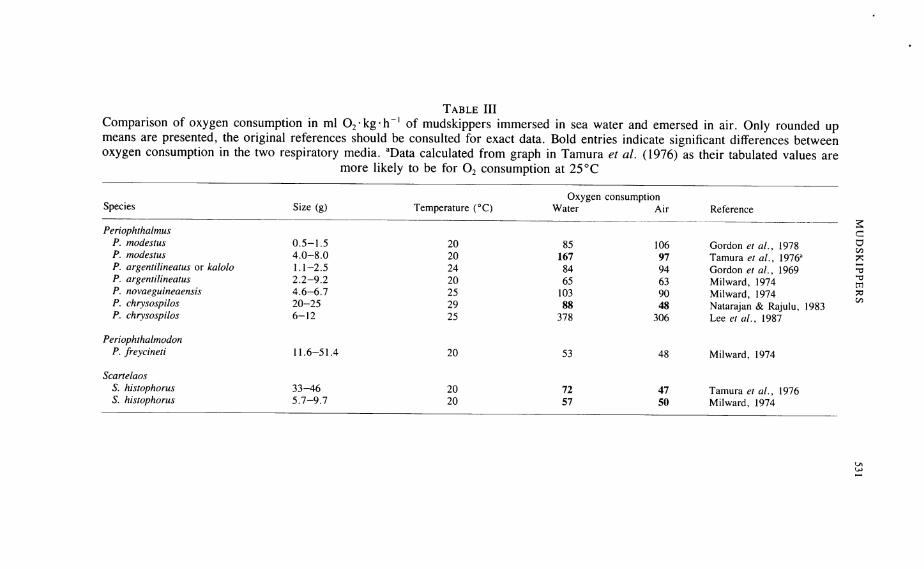
Table III
Comparison of oxygen consumption in ml 02-kg'h_1 of mudskippers immersed in sea water and emersed in air. Only rounded upmeans are presented, the original references should be consulted for exact data. Bold entries indicate significant differences betweenoxygen consumption in the two respiratory media. aData calculated from graph in Tamura et al. (1976) as their tabulated values are
more likely to be for 02 consumption at 25°C
Oxygen consumptionSpecies Size (g) Temperature (°C) Water Air Reference
Periophthalmus2C
P. modestus 0.5-1.5 20 85 106 Gordon et al., 1978 a
P. modestus 4.0-8.0 20 167 97 Tamura et al., 1976aC/2
5P. argentilineatus or kalolo 1.1-2.5 24 84 94 Gordon et al., 1969 *TD
P. argentilineatus 2.2-9.2 20 65 63 Milward, 1974P. novaeguineaensis 4.6-6.7 25 103 90 Milward, 1974 *3
P. chrysospilos 20-25 29 88 48 Natarajan & Rajulu, 198300
P. chrysospilos 6-12 25 378 306 Lee et al., 1987
PeriophthalmodonP. freycineti 11.6-51.4 20 53 48 Milward, 1974
Scartelaos
S. histophorus 33-46 20 72 47 Tamura et al., 1976S. histophorus 5.7-9.7 20 57 50 Milward, 1974

532 DAVID A. CLAYTON
now considered as facultative air breathers, respiring aerially with gills, modifiedbuccal or pharyngeal epithelium and a vascularised skin (Graham, 1976, see alsop. 536).
Despite their obvious suitability, however, the contribution of studies of mudskippers to an understanding of respiratory adaptation of marine air-breathingfishes has been minimal, accounting for less than 15% of species mentioned inreviews of the topic (Graham, 1976; Bridges, 1988). The respiratory rates ofmudskippers confined to either the aquatic or aerial respiratory medium (TableIII) shows that they also conform to the generalisation that respiration rates ofmarine species in water are equal to or greater than the aerial rate (Bridges,1988). This relationship also holds for Periophthalmus, Boleophthalmus andScartelaos freely able to select aerial or aquatic respiration (Tamura et al., 1976;Niva et al., 1979, Natarajan & Rajulu, 1983).
RESPIRATION IN AIR AND WATER
Oxygen consumption in water and in air increases with increasing temperature.For Periophthalmus modestus [as P. cantonensis] (10—30°C; Gordon et al.,1978; 10-15°C, Tamura et al., 1976), P. argentilineatus [as P. vulgaris] andPeriophthalmodon freycineti [as Periophthalmodon schlosseri] (20—35°C;Milward, 1974) and Scartelaos histophorus [as Boleophthalmus chinensis](10—15°C; Tamura et al., 1976) there are no significant differences between theaerial and aquatic rates. By contrast, however, the aerial rate is significantly lessthan the aquatic rate for Periophthalmus modestus and Scartelaos histophorusin the temperature range 20—35°C (Tamura et al., 1976) and for S. histophorusin the range 15—35°C where the disparity increased with increasing temperature(Milward, 1974). Gordon etal. (1978) could offer no reasonable explanation forthe differences between their result and that of Tamura et al. (1976): the stateof thermal acclimation including a seasonal one and the possibility of populationdifferences seem unlikely. In Periophthalmuschrysospilos however, smaller fishhave a greater dependence on cutaneous rather than branchial respiration (Lowet al., 1990) and the small (0.5-1.5g) P. modestus used by Gordon et al. (1978)could have been aerially more efficient than the larger (4—8 g) fish used byTamura et al. (1976). The physiological findings for Australian Scartelaoshistophorusmatch ecological reality in that the fish is usually confined to surfacepools at low tide (Milward, 1974). A similar explanation could be offered forJapanese S. histophorus, except that when the relative role of the gill and skinare considered, it appears that the gill of Scartelaos has a greater ability foroxygen uptake than that of the more terrestrial Periophthalmus (Tamura et al.,1976).
Except for P. chrysospilos, which showed no change (Lee et al, 1987), whenmudskippers could choose between respiring in air or water, the total oxygenconsumption increased over that recorded in either medium separately. For P.modestus and Scartelaos histophorus both the aquatic and aerial rates werereduced when compared with the single medium values (Tamura et al., 1976).In Periophthalmus koelreuteri, Natarajan & Rajulu (1983) found no differencein the values while for Boleophthalmus boddarti there was an increase in theaquatic rate when the animal was free to choose. The aerial rate in air alonewas not calculated (Niva et al., 1979). Without some method of comparinglevels under the different conditions, the relative contribution of activity to the

MUDSKIPPERS 533
differences observed in oxygen consumption is difficult to determine. As fishmove between the respiratory media, oxygen consumption may well be expectedto be higher and whereas fish become quiescent under water it is not clear ifbasal rates were measured in all cases. That oxygen consumption can beextremely variable is demonstrated by the routine metabolic rates in sea waterof Periophthalmus sobrinus (Gordon et al, 1978). During the 5-h period ofmeasurement, the fish would remain quiescent, but at irregular intervals wouldswim for variable periods. The routine metabolic rate varied between 160—430ml 02*kg'h_1 and for small fish the maximum rate was five times larger thanthe resting metabolic rate. In air the resting oxygen consumption rate of P.barbarus could be more than tripled during activity (Hillman & Withers, 1987).
The bimodal oxygen uptake reported under free choice conditions (Tamura etal., 1976; Niva et al., 1979; Natarajan & Rajulu, 1983) is simply the percentvalues of total oxygen obtained from the two media. It is invariably greater fromwater than air and is not to be confused with either cutaneous and branchial
respiration or the bimodal respiration of air-breathing freshwater fish wheremost oxygen is taken aerially and most carbon dioxide released aquatically(Graham, 1976). Indeed, no work on mudskippers has attempted to partition gasexchange in this way and only Tamura et al. (1976), by preventing gill ventilation, have strictly compared the relative contribution of cutaneous and branchialrespiration. In reporting on respiratory quotients, only Schottle (1932) providesany information on carbon dioxide release. In air the mean oxygen consumptionof P. argentilineatus [as P. vulgaris for a single small specimen and P. dipusfor two larger ones] was 151.4 ml 02.kg-l.rT1 and the mean carbon dioxideoutput 109.5 ml COrkg~1.h~1 with a corresponding RQ of 0.78. For a singlePeriophthalmodon schlosseri the values were 123.5, 90.23 and 0.72 respectively.
When the body behind the gills and the head are separated by a thin rubberseptum, oxygen consumption anterior and posterior to the septum can bemeasured independently. Thus in water Periophthalmus argentilineatus andScartelaos histophorus obtain about 20—25% and 10—17% respectively fromrespiratory surfaces behind the head (Milward, 1974). In Periophthalmussobrinus [ = argentilineatus or kalolo] the equivalent value was slightly lessthan half, but by assuming that the head represented about a quarter of the totalbody surface, Teal & Carey (1967) suggest that 60% of oxygen is exchangedthrough the skin. Unfortunately, their tabulated results cannot be meaningfullyinterpreted and do not substantiate their conclusions. Total cutaneous respirationin P. modestus and Scartelaos histophorus accounted for 48% ± 12% and36% ± 18% in water and 76% ± 10% and 43% ± 11% respectively in air(Tamura et al., 1976). In air, gill oxygen uptake was much reduced being only31% of the 'standard' for uptake in water in Periophthalmus and 58% of thatin Scartelaos; aerial skin respiration remained much higher at 96% and 78%respectively. Given that oxygen uptake in both species in air was reduced toabout 60% of that in water, it appears that the reduction in Periophthalmus waslargely due to a reduction in gill uptake. For Scartelaos, although the gillreduction was the greater, oxygen uptake was also reduced via the skin.
Partitioning studies are necessarily restrictive of fish movement, but theexperimental conditions in the free choice situation also introduce limitationsthat have implications for the partitioning of oxygen uptake between water andair. By only providing a small 5-cm diameter semi-circular hole in the centreof the float separating the aquatic and aerial parts of their respirometer, Niva

534 DAVID A. CLAYTON
et al. (1978) will have increased aquatic activity and limited aerial respirationto gulping air. Furthermore, cutaneous respiration under such conditions wouldbe reduced. Similarly, in the respirometer used by Tamura et al. (1976), theopportunity for aerial cutaneous respiration would be less than that available tofish in more natural conditions.
If only because it is the only known example of diving brachycardia in fishes,the reduction in heart rate of Periophthalmodon freycineti [as P. australis](Garey, 1962) should be re-examined. The presumed confirmation (Gordon etal, 1969; Graham, 1976) by Bandurski et al. (1968) was no more than thenormal brachycardiac response of fish to hypoxia. Indeed, it was an incidentalobservation made during a preliminary experiment where the heart rate ofP. freycineti [as P. australis] in an aerated bag was noted to change from95-110 beats-min-1 to 65 beats-min-1 after substitution of a nitrogen gasphase (Bandurski et al, 1968). Working with Periophthalmus sobrinus [ = P.argentilineatus or kalolo], Gordon et al.. (1969) were unable to demonstrateany similar brachycardia and the fish actually showed an initial, but slightincrease in heart beat frequency when transferred between the two media,irrespective of the direction of transfer. The only finding in common with Garey(1962) was that the heart rate decreased after 3-4min in either water or air. Asa result of electrode insertion, however, the fish were dying during recording(Gordon etal., 1969) and even this similarity is suspect. The whole problem willundoubtedly benefit from the improved methodologies now available.
RESPONSES TO HYPOXIA
Garey (1962) suggested that diving brachycardia in mudskippers was anadaptation to hypoxia in their burrows and reported that the mud in one burrowwas almost free of oxygen. In the water in burrows of P. modestus the dissolvedoxygen concentration was certainly low, ranging between 0.2 and 0.7 ml02.1-1and the inertness of P. modestus in closed jars of sea water when the oxygenlevel fell to between 0.7 and 1.0 ml 02.1~' lent further support to the idea thatmudskippers had a limited tolerance to hypoxia (Gordon et al., 1978). In asimilar experimental situation, Niva et al. (1979) found that Boleophthalmusboddarti died when the dissolved oxygen in the water fell to 2.3 to 1.8 ml02.1_I. However, mudskippers spend long periods in their burrow at high tideand at low tide in inclement weather (Clayton & Vaughan, 1988; Lim & Ip,1989) and burrows contain an adequate supply of oxygen for eggs to hatch(Kobayashi et al., 1971). Replication of the 'closed jar' experiment withPeriophthalmus chrysospilos also produced inert fish when the oxygen level inthe sea water fell after 2 h to 0.75 ml 02.1_1 (Chew et al., 1990). Furtherexperiments showed that this response was not, as Gordon et al. (1978) concluded, due to a limited tolerance for hypoxia. Fish were exposed to varyingdegrees of environmental hypoxia by substituting part of the oxygen by anequivalent amount of nitrogen. The more usual response of fish to hypoxia isto increase the respiratory rate (Bridges, 1988), but in P. chrysospilos the rateat 1.0 ml 02.1~' of dissolved oxygen was approximately a quarter of that offish under normoxia and the respiratory rate decreased sharply within 5 min ofexposure with the result that at 0.8 ml 02.1_I, P. chrysospilos was able tosurvive for at least 6 h (Chew et al., 1990). Rather, it is possible that the 'closedjar' fish succumbed to the increased carbon dioxide level and concomitant pH

MUDSKIPPERS 535
increase in the sea water (Chew et al, 1990). The possibility that fish built upan oxygendebt during hypoxic exposure was expressed by Teal & Carey (1967)who reported a doubling of respiration in the period following return tonormoxic conditions. As the anaerobic end product of carbohydrate metabolism,evidence of high lactate (lactic acid) levels would provide support for thissuggestion, but as Teal & Carey (1967) did not measure it, it was left toBandurski et al. (1968) to demonstrate that in Periophthalmodon freycinetianoxic conditions more than doubled brain and tripled muscle resting values oflactate. During hypoxia, blood lactate levels in Periophthalmus chrysospilosshowed a 6-fold increase (control value 1.23 ^mol-ml"1), but the respiratoryrate was only slightly increased during recovery in normoxic conditions. Thislow level of oxygen debt repayment suggested that there must have been adepressionof glycolytic activityand energy consumption during hypoxia (Chewet al., 1990). This was confirmed by Ip et al. (1991a) who compared a varietyof compounds including ATP, glycogen and lactate in P. chrysospilos exposedto normoxic (environmental) hypoxia and severe exercise (functional hypoxia).Experimental hypoxia produced no increase in lactate above the normoxiccontrol condition and only when exercised did anaerobic metabolism occur, withATP and glycogen decreasing and lactate increasing in muscle tissue. Thismudskipper's response to hypoxia thus involves slowing of the respiratory rateand a reduced energy consumption. It does not involve glycolysis and previousreports of lactate accumulation (Bandurski etal, 1968; Gordon etal., 1969) arelikely to be due to functional hypoxia. After 6 h hypoxic exposure, however,lactate did accumulate in the gills of P. chrysospilos (Ip et al., 1990) andPeriophthalmodon schlosseri (Ip & Low, 1990). Lactate was not found toaccumulate in branchial tissue of Boleophthalmus boddarti, suggesting that thisfish has biochemical adaptations for dealing with hypoxia that are different fromthose of Periophthalmus and Periophthalmodon (Ip et al., 1990). While lactateaccumulation indicates anaerobiosis, neither Periophthalmus chrysospilos norBoleophthalmus boddarti are anaerobic animals because there was no evidenceof the metabolic products of glycolysis in such animals: no branchial succinateor proprionate (Ip et al., 1990) or ethanol (Chew et al., 1990) was found.As suggested earlier for studies of nitrogenous excretion, similar studies onPseudapocryptes wouldbe informative. If the observation that Periophthalmodonschlosseri [as Pseudapocryptes schlosseri] is also capable of aestivation(becoming torpid for periods during the hottest part of the year; Hora, 1933) canbe confirmed, a strong comparative study could be undertaken, especially ifthe torpor induced during the cold of winter (hibernation response) of Periophthalmus (Kobayashi et al., 1971) and Scartelaos [as Boleophthalmus](Tamura et al, 1976) are included (see also p. 545).
OXYGEN TRANSPORT
Virtually no work on oxygen transport in the blood has been undertaken inmudskippers. Venkateswarlu (1966) reports that the total amount of iron in theblood of Periophthalmus schlosseri [ = Periophthalmodon schlosseri] andBoleophthalmus boddarti are approximately the same (27—45 mg-100 ml"1and 25-42 mg-100 ml"1 blood respectively) and greater than that of the fullyaquatic goby Glossogobius giuris. Without any indication of the number ofindividuals or their weight the data are of minimal value as are those on oxygen

536 DAVID A. CLAYTON
consumption at an unspecified temperature. For two specimens of Periophthalmus sp., Pradhan (1961) reported that the amount of haemoglobin (16.5 and15 g-100 ml"1 blood for 21.2 and 61.5 g fish respectively) was greater thanthat of a variety of freshwater fishesof equivalent size. Vivekanandan& Pandian(1979) report on the leukocyte (15.2 x 103-mm"3), erythrocyte (3.032 X106-mm-3) count and haemoglobin concentration (14.8 ml-100 ml"1 blood) inBoleophthalmus boddarti. The haemoglobin and red blood corpuscle (RBC)counts were significantly correlated. The highest values were found in males,the lowest in juveniles and intermediate levels in females. The same measureswere repeated over 48 h for males with free access to water and air (control)and for those restricted to either the aquatic or the aerial medium. The waterand air values of the percentage of haemoglobin and RBC counts were reducedin comparison with the control. It should be noted, however, that the fishexposed to air died within 51 h. All of these studies would benefit from morecontrolled repetition. Similar values for the blood parameters of B. boddartiwere reported by Manickam & Natarajan (1985). Variables measured includederythrocyte count, haemoglobin concentration, haemocrit, mean cell volume,mean cell haemoglobin, mean haemoglobin concentration, oxygen capacity andstandard bicarbonates, all of which were higher in B. boddarti than inPeriophthalmodon schlosseri [as Pseudapocryptes schlosseri] (Manickam &Natarajan, 1985). The authors also report, but without statistical analysis, that(for both species) there were clear-cut sex differences with male valuesexceeding those of females. Additional data on oxygen dissociation curves andits associated Bohr effect would be instructive.
RESPIRATORY SURFACES
The main gas exchange surfaces in mudskippers include the gills, buccopharyngeal and opercular cavity membranes and the skin (Graham, 1976) butthe fins (Milward, 1974) and the nasal sac (Rauther, 1910) have also beenproposed as suitable structures.
Gills
Gill structure has been investigated by light (Rauther, 1910; Schottle, 1932;Singh & Munshi, 1969; Milward, 1974; Hughes & Munshi, 1979; Al-Kadhomiy& Hughes, 1988) and electron microscopy (Welsch & Storch, 1976; Hughes &Munshi, 1979; Low et al., 1988; Yadav et al., 1990). The general (Schottle,1932; Das, 1934) and specific (Niva et al., 1981; Al-Kadhomiy & Hughes, 1988)gill blood circulation and musculature (Schottle, 1932; Willem & Boelaert,1937; Singh & Munshi, 1969) of some mudskippers has also been described.
Morphometric measurements of the gills of a number of species have beentaken and are summarised in Table IV. The gill areas of a number of mudskippers were calculated by Schottle (1932) and re-presented by Graham (1976),but are omitted here as only the length and not the weight of the fish wererecorded (see also p. 540). As with earlier tabulated summaries (Hughes & Al-Kadhomiy, 1986; Low et al., 1990), to aid comparison between species, themeasurements presented are those calculated for specimens of a weight dictatedby Tamura & Moriyama (1976). Theirs was the first major study of mudskippergill morphometries, but does not include regression slopes that describe gill
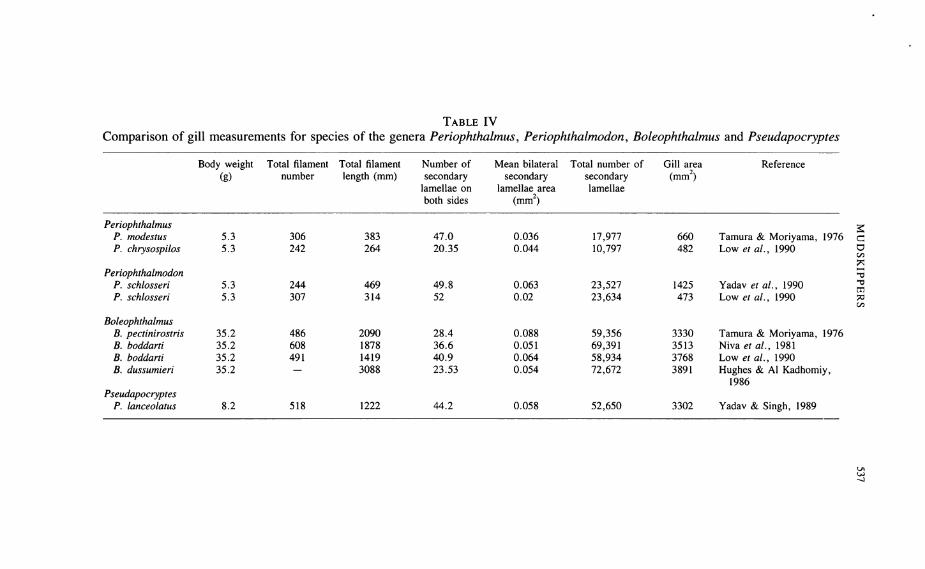
Table IV
Comparison of gill measurements for species of the genera Periophthalmus, Periophthalmodon, Boleophthalmus and Pseudapocryptes
Body weight Total filament Total filament Number of Mean bilateral Total number of Gill area Reference
(g) number length (mm) secondarylamellae on
both sides
secondarylamellae area
(mm2)
secondarylamellae
(mm2)
Periophthalmus 2P. modestus 5.3 306 383 47.0 0.036 17,977 660 Tamura & Moriyama, 1976 cP. chrysospilos 5.3 242 264 20.35 0.044 10,797 482 Low et al., 1990 O
Periophthalmodon ^3P. schlosseri 5.3 244 469 49.8 0.063 23,527 1425 Yadav etal., 1990 gP. schlosseri 5.3 307 314 52 0.02 23,634 473 Low et al., 1990 *
BoleophthalmusB. pectinirostris 35.2 486 2090 28.4 0.088 59,356 3330 Tamura & Moriyama, 1976B. boddarti 35.2 608 1878 36.6 0.051 69,391 3513 Niva et al., 1981B. boddarti 35.2 491 1419 40.9 0.064 58,934 3768 Low et al., 1990B. dussumieri 35.2
—3088 23.53 0.054 72,672 3891 Hughes & Al Kadhomiy,
1986
PseudapocryptesP. lanceolatus 8.2 518 1222 44.2 0.058 52,650 3302 Yadav & Singh, 1989

538 DAVID A. CLAYTON
measurements in relation to fish weight. Consequently Hughes & Al-Kadhomiy(1986) and Low et al. (1990) used their regression equations to produce valuesfor the various gill parameters so they could directly compare their results withthose of maximum sized Scartelaos histophorus [as Boleophthalmus chinensis]and Periophthalmus modestus [as P. cantonensis] (53 & 8.8 g respectively) ascalculated by Tamura & Moriyama (1976). However, Tamura & Moriyama'sdata were presumed dimensions, calculated from regressions in body weight forAnabas testudineus and Saccobranchusfossilis. Instead of using such adulterateddata, it is more realistic to use the averaged data set values of the gill parameterspresented by Tamura & Moriyama (1976).
Table IV therefore replicates the comparisons made by Hughes & Al-Kadhomiy(1986) and Low et al. (1990) at the more appropriate (35.2 g & 5.3 g) fish size.The total filament number reported for Tamura & Moriyama (1976) by Lowet al. (1990) and in Table IV remains the same because Low et al. (1990)erroneously presented the averaged values (of 35.2 & 5.3 g) and not those ofthe maximum sized (53 & 8.8 g) fish. Data for small (1—6 cm, 0.75—2.3 g)Periophthalmodon schlosseri from the Andman (Andaman) islands (Yadav etal., 1990) is included in the Table, but it is likely that this is an incorrectdesignation. P. schlosseri can exceed 20 cm and 100 g and given the smallsize of the specimens, it is more likely that the fish was Periophthalmodonseptemradiatus or Periophthalmus minutus, both of which are from the correctgeographical area (Murdy, 1989). Less circumstantial, however, is the evidenceof the structure of the gills. In this case, branched filaments seem to be a featureof the gills of Periophthalmodon, being found in P. freycineti (Milward, 1974)and P. schlosseri (Low et al, 1990), but not in the specimens investigated byYadav et al. (1990).
When presented for a hypothetical fish of a given weight, the calculated valuesfor total gill area within boleophthalmids and within periophthalmids are similar(Table IV). The exception is the high value obtained for P. schlosseri whichresults from the increased filament length and bilateral secondary lamellae areareported for this species (Yadav et al. (1990).
Gill morphometric data enabled Hughes & Al-Kadhomiy (1986) to distinguishbetween genera such that when values for their fish of appropriate weight werecompared with that of Tamura & Moriyama (1976), their data on Boleophthalmusdussumieri [as B. boddarti] matched that of Scartelaos histophorus andBoleophthalmus boddarti better than that of Periophthalmus modestus. Whilethey were correct to conclude that their fish was a boleophthalmid and not aperiophthalmid, the comparison of gill parameters between species based onvalues obtained from regression equations should be treated with considerablecaution. By way of illustration, Table V provides values of gill areas calculatedfrom regression equations. The Table clearly shows that gill area increases withincreasing weight and that generally the gill areas of the boleophthalmids aregreater than those of the other genera. This is certainly true among fish of 20g and greater where, for example, the gill area of a 30-g P. chrysospilos is only77% that of an equivalent sized Boleophthalmus dussumieri. B. dussumieri alsohas a gill area of a similar magnitude to other 30-g boleophthalmids. At themean weight of Periophthalmus chrysospilos (5.3 g), the comparison would leadto the conclusion that Boleophthalmus dussumieri has a gill area closer to thatof Periophthalmus chrysospilos and Periophthalmodon schlosseri than the otherboleophthalmids. At this size the gill area of Periophthalmus chrysospilos is

Table V
Gill areas (A) for fishes of differentweights (W) calculatedfrom regression equations. Fish weightsare given at arbitrary 10-g intervalsexcept that 5.3, 35.2 and 53 g are the averaged and presumed maximum weight individuals from Tamura & Moriyama (1976). aYadav& Singh (1989) measured the gills of 35 fish (1-21 g), but then reduced the data to seven averaged body weights from which the
regression equations were calculated.
Species Regression Gill area (mm2) calculated Fish used in
equation for fish of weight (g;} calculation Reference5.3 10 20 30 35.2 53 N Weight
(g)
Length(mm)
Boleophthalmus dussumieri A = 92.654W10496 533 1039 2150 3290 3891 5979 14 3.6-35.4 40-240 Hughes & Al Khadomiy 1986B. boddarti A = 281.28W°-709 918 1439 2353 3136 3513 4695 9 1-12 55-110 Niva et al., 1981B. boddarti A = 679W04812 1515 2056 2870 3487 3768 4588 10 2-35 _ Low et al., 1990Pseudapocryptes lanceolatus A=607.8W0827 2376 4081 7240 10124 11555 116209 7 1-21 75-165 Yadav & Singh, 1989aPeriophthalmus chrysospilos A = 97.6W°'9577 482 885 1720 2536 2955 4373 9 2-13 — Low et al., 1990Periophthalmodon schlosseri A=100W09312 473 853 1627 2374 2755 4033 7 3-111
—Low et al., 1990
aa00
2
oo

540 DAVID A. CLAYTON
90% that of Boleophthalmus dussumieri. Nevertheless, the results appear tosupport the generalisation that gills are reduced in more terrestrial species(Graham, 1976). Similarly, in comparing 10-g fish, Low etal. (1990) concludethat the natural preference of B. boddarti for an aquatic environment can beexplainedby their having more than double the gill area of either Periophthalmuschrysospilos or Periophthalmodon schlosseri. Furthermore, Pseudapocrypteslanceolatus can be considered the most aquatic as the gill area is greater thanthat of all the other genera (Table V). This conclusion is partially at variancewith that of Yadav & Singh (1989). They calculated gill area for fish of a lengthequivalent to specimens of Periophthalmus vulgaris [ = P. argentilineatus] andBoleophthalmus viridis [ = Scartelaos histophorus] for which Schottle (1932)provided data. On this basis Pseudapocryptes was more aquatic than Periophthalmus having more than 1.5 times the gill area, but equivalent to Scartelaoswith which it had a comparable gill area. These are inappropriate comparisonsbecause species have different length/weight relationships and will have widelydifferent weights at a similar length. As a corollary to this observation, the datain Table V for Periophthalmus chrysospilos and Pseudapocryptes lanceolatusabove 20 g have no biological significance because neither of these species growto such sizes.
Similar comparisons could have been presented for other gill variables andwould have also shown the size dependent nature of the conclusions drawn. Themeasurement of gills is a time-consuming operation and so small data sets areto be expected. However, while regression equations are precise descriptions ofthe relationship between two variables, they can be influenced considerably byone data point, especially in small data sets (Clayton, 1990). As a minimum,data sets need to include the complete size range of the species with measurements of equal numbers of fish at each size. Niva et al. (1981) only have onefish heavier than 10 g while Low et al. (1990) have only three and Hughes &Al-Kadhomiy (1986) only two that are lighter than 10 g. The measurements ofany one of these fish will have a greater influence on the slope of the regressionline than that of the other fish.
The cause for caution in the interpretation of data derived from regressionequations does not imply that size dependence is only a statistical artefact. Thegrowth related changes in the size or form of the respiratory surfaces are of clearphysiological relevance. Greater spacing between secondary lamellae is anadaptation to aerial exposure, reducing the collapse of the gill in air (Graham,1976) and is a characteristic of all mudskippers so far investigated. This is theonly parameter for which there is a consistently negative slope value for theregression equation for all species (Niva et al., 1981; Hughes & Al-Kadhomiy,1986; Low et al., 1990; Yadav et al., 1990) indicating that the frequency ofsecondary lamellae decreases with increasing body weight. Low et al. (1990)found the decrease was greater for Periophthalmus chrysospilos (slope —0.237)than for Periophthalmodon schlosseri (—0.052) which was similar to the moreaquatic Boleophthalmus boddarti (-0.031). Other features of the gill ofPeriophthalmodon schlosseri, however, exhibited greater terrestrial adaptation.The total filament length and gill area was similar to that of Periophthalmuschrysospilos and reduced in comparison with that of Boleophthalmus boddarti(Low et al., 1990). The branched filaments reduce the risk of collapse and thetissue fusion between secondary lamellae may reduce the risk of dehydration(Schottle, 1932; Low et al., 1988). Other authors report slope values of a similar

MUDSKIPPERS 541
magnitude for B. boddarti (-0.083, Niva et al., 1981) and Periophthalmodonschlosseri (-0.048, Yadav et al., 1990), but for Boleophthalmus dussumierithe slope (-0.229, Hughes & Al-Kadhomiy, 1986) is similar to that ofPeriophthalmus chrysospilos. In other respects Boleophthalmus gills are betteradapted to aquatic respiration because the filaments are longer and more of thesecondary lamellae are aligned parallel to the respiratory water current (Low etal., 1988). Conversely, the branched filaments of Periophthalmodon (Milward,1974; Low etal., 1988) and the interlamellar fusion (Schottle, 1932; Low etal.,1988; Yadav et al., 1990) will impede water flow and reduce their gill efficiencyin water.
Gill area is reduced in species that are more dependent on aerial respiration(Graham, 1976) and is reflected in the low values of the gilhskin area ratiofor Periophthalmus (0.27-0.46 for P. argentilineatus [as vulgaris and dipus],P. kalolo [as koelreuteri] and P. chrysospilos (Schottle, 1932; Low et al.,1990) and for Periophthalmodon schlosseri (0.22-0.5; Low et al., 1990) whencompared with that of Scartelaos histophorus (0.72; [as Boleophthalmusviridis], Schottle, 1932) and B. boddarti (0.67-0.77; Low et al., 1990).Contrary to the generalisation, however, are the lower values (0.48-0.52)reported for B. boddarti (Niva et al., 1981). Additionally, the ratio forPeriophthalmus chrysospilos and Periophthalmodon schlosseri increased withincreasing fish size (Low et al, 1990), a finding that argues for the greateramphibiousness of smaller individuals except that the largest ratio is stillrelatively low.
Skin
The skin of mudskippers undoubtedly has a respiratory function (Rauther, 1910;Schottle, 1932), but the gill:skin area ratio is a crude method of measuring itscontribution to total respiration. The capacity for gaseous exchange is variablylocated and dependent on adequate skin vascularisation.
Beginning with the species with most skin vascularisation, Periophthalmusargentilineatus, the blood circulation is greatest on the skin of the upper andfront sides of the head and gill covers, reduced in the tail and body and isvirtually absent from the ventral surfaces (Schottle, 1932). In the region of scalepockets, blood vessels in finger-like projections of the dermis (corium) rise tothe epidermal layer where they branch umbrella-like into several intra-epithelialcapillaries running parallel to the surface. Intra-epithelial capillaries also occurin P. modestus [as P. cantonensis] (Maekawa et al., 1968). In Periophthalmodonschlosseri where the epidermis is thicker, the dermal papillae are histologicallymore complicated, but also have intra-epithelial branches and capillaries so closeto the surface that the epidermis can be seen microscopically to bulge out.
Scartelaos histophorus [as Boleophthalmus viridis] has the simplest form ofskin respiration that consists of blood vessels in flat dermal papillae that lie overthe scales. Whereas the epidermis on the head and trunk has several layers ofcells, it is reduced to one layer of very flat turgor cells above the scales, whichare confined to the caudal region. More highly differentiated papillae, withpresumably better respiratory properties, are found on the outer surface of theoperculum, on the dorsal surface of the head and in a narrow zone along theupper trunk to the beginning of the second dorsal fin. The opercular buttonpapillae consist of a single capillary loop towards the surface while elsewhere

542 DAVID A. CLAYTON
the skin papillae are larger and contain a network of capillaries.In B. boddarti there are large diameter button-shaped papillae on the dorsal
surface of the head and to the side of the start of the first dorsal fin. Additionally,the fish has greatly differentiated true respiratory papillae each associated witha single scale. They are distributed over the upper side of the head up to a linejoining the ventral point of attachment of the branchiostegal membrane to thecorner of the mouth and on the side of the trunk in a wedge-shaped zone pointingcaudo-dorsally up to the middleof the seconddorsal fin. These papillaeare alsofound in the skin of the basal section of the ventral fins. Where the blood vesselsbreak through the dermis into the epidermis, the efferent and afferent vesselsalways run close together and parallel to the scale before bending upwards intothe papilla (Schottle, 1932). The possibility that the positioning of these vesselsmay act as a counter-current heat exchanger is worth exploring. The centrally-located arteriole branches into a large capillary bed which connects to peripheralring-like venules. As both vessels are pigmented this arrangement is easilyvisible and has also been found in B. dussumieri [as B. dentatus] by Salih &Al Jaffery (1980) and Al Kadhomiy & Hughes (1988) [as B. boddarti].
Whereas there is a standard methodology for the measurement of gill and skinarea, the other accessory respiratory surfaces are less amenable to quantificationand, therefore, seem to have been somewhat neglected in the discussion of thephysiology of respiration. Only Niva et al. (1981) measured the opercularchamber area, finding that its rate of growth was less than that of either the gillsor the skin.
Accessory organs
There are many anatomical descriptions of the accessory organs (Rauther, 1910;Harms, 1929; Schottle, 1932; Das 1933, 1934; Willem & Boelaert, 1937,Marlier, 1938; Milward, 1974; Singh & Munshi, 1969; Klika & Tesik, 1980;Singh et al., 1989; Singh et al., 1990; Yadav et al., 1990). In all species ofmudskipper the buccal and pharyngeal cavities and the opercular space outsidethe gill arches are large and capable of considerable extension for retentionof air. Additionally, the extent of the opercular chamber is increased in one{Boleophthalmus, Pseudapocryptes and Scartelaos) or two directions {Periophthalmus and Periophthalmodon). Ventro-anteriorly, a diverticulum extendsforwards beneath the branchial arches and the floor of the buccal cavity. Thisventral extension (Schottle, 1932) is found in all species and has subsequentlybeen called the infra- (Singh & Munshi, 1969) or inferior branchial recess(Yadav et al, 1990). Dorsally a flat pharyngeal side chamber has a wide connection to the opercular cavity through the dorsally restricted opening of the firstgill slit. The opening is restricted because the gill arch is fused to the pharynxwall by a ventro-medial membrane which is vascularised on the opercular side(Schottle, 1932). The dorsal extension of the opercular chamber is also calledthe pharyngeal diverticulum (Klika & Tesik, 1980) or the suprabranchial chamber(Singh & Munshi, 1969; Yadav et al., 1990). Neither the dorsal extension northe restricted opening of the gill arch is present in Boleophthalmus or Scartelaos.
Information relating to the extent of vascularisation of these accessory organscan be derived from two sources: the descriptions of the histology of the accessory organs (Rauther, 1910; Harms, 1929; Schottle, 1932, Singh & Munshi,1969; Milward, 1974; Klika & Tesik, 1980) and that of the blood vessels

MUDSKIPPERS 543
supplying them. The latter have been described in Periophthalmus argentilineatus(Schottle, 1932; Singh & Munshi, 1969), Periophthalmodon schlosseri, Scartelaoshistophorus (Schottle, 1932), Boleophthalmus boddarti (Schottle, 1932; Niva etal, 1981) and Pseudapocryptes lanceolatus (Das, 1934; Singh etal., 1990). Forall species, the pharyngeal and opercular chamber, including the branchiostegalapparatus which greatly contributes to the inflation capability of the opercularchamber, and the ventral extension are well vascularised as are the dorsalextensions in those species that possess them. Contrary to the generalisationmade earlier that an increase in area of more heavily vascularised epithelium isaccompanied by a reduction of gill area in increasingly amphibious species(Schottle, 1932), the buccal cavity is vascularised to a variable degree. WhileScartelaos histophorus and Boleophthalmus boddarti have fewer intra-epithelialblood vessels than the well vascularised buccal epithelia of Periophthalmuschrysospilos and Periophthalmodon schlosseri, Schottle (1932) found almostnone in Periophthalmus argentilineatus and P. modestus [as P. cantonensis].
Singh et al. (1989) found that the development of the vascularised respiratoryepithelium of the buccal, opercular and suprabranchial chambers was broadlysimilar in Periophthalmodon schlosseri, Boleophthalmus boddarti and Pseudapocryptes lanceolatus. The gill material of the embryonic fourth gill arch givesrise to a separate gill mass on either side of the pharynx which later begins toform the respiratory epithelium of all three chambers. In the suprabranchialchamber, gill lamellae from the dorsal ends of the arches also contribute towardsthe respiratory epithelium (Singh et al., 1989). In P. lanceolatus and Periophthalmodon schlosseri the gill tissue transferred from the arches, and particularlythe first gill arch, forms a series of longitudinal antero-posteriorly orientedridges on the inner surface of the operculum (Singh et al., 1990). The ridgesare more pronounced in Pseudapocryptes lanceolatus. As the ridges were foundto be rich in carbonic anhydrase, Singh et al (1990) consider them to be anorgan for the release of carbon dioxide that is independent of oxygen uptake.
From the large number of published light and electron micrographs of therespiratory surface (see pp. 536, 542), it is clear that the air/blood barrier is onlya few /xm thick, and where it has been measured, variability in width is apparent.The range of skin diffusion distances must be presumed to vary from that in therespiratory papillae to that in the non-vascularised sections of the epithelia. Thepresence of specialised cells including mucus, chloride and acidophil cells(Hughes & Munshi, 1979; Welsch & Storch, 1976) and mitochondria-rich cells(Hughes & Al Kadhomiy, 1986) all increase the diffusion distance. The diffusiondistances in the gills of P. argentilineatus are given as 0.9 pan (Welsch & Storch,1976) and between 1—3.2 /xm (Singh & Munshi, 1969) while for Boleophthalmusboddarti the mean and range are 1.43 /xm and 0.1—1.56 /xm respectively(Hughes & Munshi, 1979; Niva et al., 1981). For B. dussumieri the diffusiondistance of the gill membranes ranged between 4-5 /xm and 10—12 /xmdepending on the site of measurement (Hughes & Al Kadhomiy, 1986), but evenat its thickest, the distances were generally less than that for skin of the trunk(mean 121 /xm range 4-230 /xm) or snout (50, 5-144 /xm ) or the outer (725,700-1200 /xm) or inner operculum (9, 4-230 /xm ) (Al Kadhomiy & Hughes,1988), the last values being the only ones available for any accessory respiratorysurface.
There are profuse networks of capillaries in the dorsal, pelvic, caudal andparticularly the pectoral fins of Australian mudskippers (Milward, 1974).

544 DAVID A. CLAYTON
Following Harms (1929) and Schottle (1932), Milward (1974) was unable tofind any histological support for any respiratory function of the nasal cavity assuggested by Rauther (1910) after he noted the sac-like enlargement of theolfactory chamber in mudskippers. Datta & Das (1980) revealed no respiratoryfunction in their detailed anatomical study of the olfactory apparatus ofPeriophthalmus koelreuteri [?= P. novemradiatus], Boleophthalmus boddartiand Pseudapocryptes lanceolatus.
TEMPERATURE REGULATION
The range of temperature to which intertidal amphibious fishes are exposed areamong the greatest met by any fish. Within wide tolerance limits, mudskippersare extremely eurythermal using body colour changes (Stebbins & Kalk, 1961),evaporative cooling and behavioural thermoregulation (Tytler & Vaughan,1983). Generally, body temperatures match those of the mud or surface waterfrom which the fish were caught (Stebbins & Kalk, 1961; Burhanuddin &Martosewojo, 1979; Tytler & Vaughan, 1983) although Gordon et al (1968)found no correlation between substratum and body temperature. Upper tolerancelimits have been measured in a variety of ways. In full sunlight at shade temperature of 43-44°C (Gordon etal, 1969), Periophthalmus sobrinus died within 50min when their body temperatures were 33-35°C (Gordon et al., 1968, 1969).In water P. sobrinus stopped breathing at a mean temperature of 41.9°C (n = 5,range 40.4—42.3), but subsequently recovered when placed in water at 31°C(Stebbins & Kalk, 1961). They did not note the time of exposure, but in waterat 38-39°C, P. waltoni survived for 9 min before becoming comatose (Al-Naqi,1977). Survival time decreased with increasing temperature until at 43 °C it wasonly 48 s. All fish subsequently recovered at lower temperatures (Al-Naqi,1977). Lower limits are equally vague, P. sobrinus was immobilised at 10°Cand recovered at 20°C (Stebbins & Kalk, 1961) and opercular movements, butnot respiration, of P. modestus and Scartelaos histophorus ceased at 5°C andbegan again at 10°C (Tamura et al., 1976). At 4°C Periophthalmus waltonibecame comatose after 77 s and at -1.5°C coma ensured after 26 s. Fish were
resilient in that after 95 min exposure at 4°C and 0°C, recovery at roomtemperature took 97 and 215 min respectively; 24 min at —1.5°C required 58min recovery time (Al-Naqi, 1977).
The burrow is an important temperature refuge especially during periods ofhigh surface temperature, when differences can be as high as 11°C (Tytler &Vaughan, 1983). Fish avoid high surface water temperatures (37°C, Stebbins& Kalk, 1961) and under these conditions it would be expected that the durationof terrestrial excursions would decrease and refuging increase, but no convincing data are available. At the other extreme when surface temperaturesare below that of the burrow, Periophthalmus waltoni and Boleophthalmusdussumieri in Kuwait (29 °N) only emerge when the temperature rises to about10-11 °C, equalling that of the burrow (Tytler & Vaughan, 1983). Other mechanisms of behavioural thermoregulation include basking in the shallower,warmer edges of standing pools in cold weather and following the advancing tideor climbing onto shaded structures that project from the tidal flat in the splashzone when the weather is hot (Tytler & Vaughan, 1983). Rolling onto the sideis also assumed to be involved in thermoregulation (Stebbins & Kalk; 1961,

MUDSKIPPERS 545
Khoo, 1966; Clayton & Vaughan, 1988) and Boleophthalmus engage in pectoralfin flapping in which the well spread fin is first moved anteriorly and applied tothe opercular region and then placed posteriorly against the flank (Clayton &Vaughan, 1988). This action may also serve to keep the skin or fin rays moist,but Stebbins & Kalk (1961) thought that the similar action in Periophthalmushelped to mix the air and water retained in the opercular chambers. The skinand fins could be moistened for both evaporative cooling and cutaneousrespiratory purposes, and the former possibility is supported by the observationthat B. boddarti roll onto the side at which a jet of air is directed (Ip et al.,1991b).
So far only relatively short term responses to environmental temperatureshave been discussed, but mudskippers also exhibit long term responses toseasonal fluctuations in temperature. These responses have been little studied,but are more likely to be physiological than behavioural. In tropical regions(23 °N) Pseudapocryptes and Periophthalmodon probably aestivate, retreatinginto their burrows for extended periods during the hottest part of the year(Hora, 1933, 1935a).
Further north in Japan (33 °N), deep burrow temperature in the cold seasondropped to 5—5.6°C and Periophthalmus modestus remained in their burrowsfrom November to March, only emerging when the air temperature reached18°C (Kobayashi et al., 1971). For Scartelaos histophorus air emergencetemperature was 22°C and was not reached until May (Tamura et al., 1976).Understanding how these species cope with the low burrow temperature for suchlong periods would be interesting.
ECOLOGY
HABITAT AND DISTRIBUTION
The distribution of mudskippers is commonly assumed to be based on thepresence of a suitable muddy substratum (MacNae, 1968a; Quereshi & Bano,1971) but they are also found on other substrata. At Laboean (= Labuhan) andPopole Island (=Popau Popole, 6°24'S:105°48'E) along the west coast ofJava, Periophthalmus kalolo [as P. harmsi and P. koelreuteri] were found on(sea?) grass surfaces and on sandy-rocky shores (Harms, 1929; Eggert, 1935)and at Perbaoegan (= Perbaungan, 2°03'N:99°58'E) P. chrysospilos "onlyoccurs on the sandy beach despite the presence of a large muddy area not faraway" (Eggert, 1935). P. argentilineatus [as P. kalolo] inhabits rock strewnshores in the New Hebrides and Polynesia (Vanuato region, 16°S:168°E)(McCulloch & Ogilby, 1919). Similarly, Gordon etal. (1968) found P. sobrinus[ = P. argentilineatus or kalolo] on cobblestone beaches and on the rockycoastline of Nosy Be (13°24'S:48°17'E) off the west coast of Madagascar.Mangroves and intertidal mud and sand flats were equally populated both here(Gordon et al., 1968) and at Inhaca Island (26°01'S:32°58'E) off the east coastof Africa (Stebbins and Kalk, 1961). P. modestus [as P. cantonensis] was foundon sand, gravel and mud substrata in the Gum river in Korea (36°N:126°43'E)(Ryu & Lee, 1979). Despite such observations and the certainty of Eggert'sstatement, MacNae is probably correct in suggesting that such records were ofwandering immature individuals. This was true of mudskippers he collected

546 DAVID A. CLAYTON
from a rocky promontory adjacent to mangrove areas (MacNae, 1968b).Distribution according to size was reported by Ryu & Lee (1979) and additionaldata on the size and maturity of mudskippers taken from non-muddy areas wouldhelp resolve the issue. MacNae's explanation may also account for the presenceof Boleophthalmus sp. [ = boddarti or dussumieri] on the rocky littoral of PortOkha (22°28'N: 69°05'E) where they were found on the Okha and Balapur reefsbut not the ones at Dwarka, Adatra and Hanuman. The mudskippers were in themidlittoral zone of both reefs, but only at Balapur was the environment muddy(Gopalakhrishnan, 1970).
Distribution is also assumed to be based on a species' ability to live out ofwater (Hora, 1935a; MacNae, 1968a, b; Gunderman & Popper, 1984). Speciesof Apocrytes and Scartelaos are considered to live in very soft mud around meansea level while Boleophthalmus spp. are found higher on the shore. The distribution of Periophthalmus and Periophthalmodon spp. are at the level of thehighest tides and beyond (Hora, 1935a; MacNae, 1968a; Berry, 1972). Pseudapocryptes is considered as a semi-aquatic form and in the Ganges Delta is confined to spring tide pools. Of all the oxudercine genera, however, Pseudapocryptesis best adapted to withstand drought, becoming torpid at the bottom of its 2 mdeep burrow when the pools dry up (Hora, 1935a).
Thus on the open mudflats of Kuwait (29°20'N:47°50'E) Periophthalmuswaltoni [as P. koelreuteri] is found in a zone extending from above mean highhigh water (MHHW) to below mean low high water (MLHW). This zoneoverlaps that of Boleophthalmus dussumieri [as B. boddarti] which extendsfrom above MLHW to mean sea level (MSL). Scartelaos tenius [as S. viridis]is found only in areas above mean high low water (MHLW) where standingwater remains on the surface at low tide. At the upper limits of its distributionS. tenius overlaps into the area occupied by Boleophthalmus dussumieri (Clayton,1986). Apocrytes madurensis is also commonly found in this zone (Clayton &Abo Seedo, 1986). As in all reports of mudskipper distributions, however, thiszonation is based on qualitative data. A more quantitative assessment of thedistribution of Periophthalmus waltoni and Boleophthalmus dussumieri on thesame mudflats, used the number of burrows as an indicator of fish density (AlTaher, 1990). The problems of density estimates based on burrow counts arewell known (cf. crab density: Warren, 1990) and equally apply to the study ofmudskippers. In this study, however, each species' burrow was easily identifiedand it was known that a foraging P. waltoni commonly occupied a home rangeof 4-5m2 in which the fish used several burrows (Clayton & Snowdon, unpubl.data). Al Taher (1990) was able to confirm the relative positions of the specieson the shore and additionally demonstrated a seasonal variation in distribution.Between May and September the mean number of Periophthalmus waltoniburrows along two transects was 4.15-25m-2 but increased to 8.55-25m~2between November and March: the increase occurring at MHHW and mean hightide level (MHTL). As fewer burrows were found at MLHW in this latterperiod, it is probable that there was a seasonal migration of fish up the shoreduring the cooler months when longer periods of emersion would be lessstressful to the fish.
On other mudflats, this simple overlapping zonation is confounded in anumber of ways. The most obvious of these is the absence of a generic representative from the series. Periophthalmodon is only found to the east of thenortheastern coast of India (Murdy, 1989) although P. septemradiatus [as P.

MUDSKIPPERS 547
tridecemradiatus] is also reported from Korangi Creek (24°48'N:67°08'E) onthe coast of Pakistan (Qureshi & Bano, 1971; Hoda & Akthar, 1983; Hoda,1985) as well as from Dubla Island (= Dubla shoal, 21°44'N:89°32'E) inBangladesh (Qureshi & Bano, 1971) Nevertheless, the genus is absent fromKuwait mudflats. The Arabian Gulf also marks the most westward extent of
the distribution of Boleophthalmus and Scartelaos and only Periophthalmus{argentilineatus, kalolo, barbarus) are found to the west.
Whether the absence of one or more genera from a particular locality meansthat the remaining genera extend their niche is one area for future study. Inthe mixed species {Avicennia, Bruguira, Rhizophora) mangrove forests of theMorrumbene (23°45'S:35°20'E) (Day, 1974) and Mngazane (31°42'S:29°25'E) (Branch & Grindley, 1979) only Periophthalmus argentilineatus [asP. sobrinus] is found. In terms of indicating the zonation of this mudskipperwithin the estuaries, the data are inconclusive. The only indication thatP. argentilineatus may occur extensively, both within mangroves and out on theopen mudflats comes from its absence from a list of 11 species (1 annelid, 3molluscs, 7 crustaceans) which were only found among mangroves (Day, 1974).The extent of the niche of P. argentilineatus [as P. koelreuteri] in the Avicenniamangroves of Melita Bay (15°16'N:39°49'E) in the Red Sea is similarlyunclear (Fishelson, 1971).
In mangrove forests at Ku Yao Yai (8°N:98°30'E) in southern ThailandPeriophthalmus koelreuteri [ = ?P. kalolo] is confined to the mangrove andScartelaos histophorus [as Periophthalmus viridis] the open mudflat (Nateewathana & Tantichodok, 1984). Similarly in those of An Nam Bor on PhuketIsland (7°51'N:98°25/E) where P. argentilineatus [as P. vulgaris] is foundwith Scartelaos histophorus [as S. viridis], the latter species occupies the openmudflat and the former is found in the Rhizophora and Sonnertia mangroveforest (Frith et al., 1976). At Koh Surin Nua on Surin Island (9°25'N:98°50'E), Periophthalmus argentilineatus [as P. vulgaris] is also found in theBruguira and Rhizophora mangrove forests, but Periophthalmus koelreuteri[ = IP. kalolo] is the species present on the mudflat. (Frith, 1977).
On a more localised scale, the ecological correlates of the presence or absenceof a species would be instructive. Why for example is Boleophthalmus presentin the mangrove forests of Ao Nam Bor but absent from Kih Surin Nua? It isunlikely that competitive exclusion by Scartelaos koelreuteri at the latterlocation is responsible for its absence from the mudflat and other explanationsneed to be investigated. Similarly, as Periophthalmodon is absent from theselocations leaving a vacant niche in the landward edge of the mangrove, why isPeriophthalmus argentilineatus only found in Ao Nam Bor (Frith et al., 1976),but not at Koh Surin Nao (Frith, 1977)? Confusion over classification ofequivalent mangrove zones is unlikely as both studies were carried out byFrith. Nursall (1981) attributed the disappearance of P. novaeguineaensis [asP. expeditionium] and increased occurrence of Periophthalmodon freycineti[as P. schlosseri] in mangroves near Townsville (19°13'S:146°48'E) between1972and 1976to topographical changes. The former species preferred to inhabitsteep banks of the tidal channel. As siltation occurred, these became shallow,sluggish, low-sided creeks that were favoured by the latter species.
The mangroves are another confounding variable in the simple picture ofmudskipper zonation because they provide a considerably increased range ofhabitats. Nevertheless, the generalised picture of zonation of mudskippers seems

548 DAVID A. CLAYTON
to hold within Malaysian mangroves (Berry, 1972). Here, Scartelaos andBoleophthalmus are found in the mangrove pioneer and foreshore zones andPeriophthalmus and Periophthalmodon occupy the mangrove forest and itsterrestrial edge. However, while Periophthalmodon is the only fish to be foundin the driest zone, which is subjected to tides on only 2—3 days a year, it is alsofound in the pioneer and foreshore mudflat zones where the residents experiencedaily tidal immersion. A similar generic association is recorded from PortSwettenham [Pelabohan Kelang, 2°57'N:101°24'E) where Periophthalmodonalso occurs with Boleophthalmus in the streams and channels that create greatertopographical variation and tidal penetration of the mangrove (Sasekumar,1974).
ASSOCIATIONS
In Japan, the estuarine Apocryptodonpunctatus [as A. bleekeri] lives in alpheidshrimp burrows (Dotsu, 1961). In Malaysian mangroves the association ofScartelaos viridis and the crab Macrophthalmus latreilli and Boleophthalmusboddarti with Metaplax crenulatus, Uca coarctata and U. dussumieri was firstreported by MacNae (1968a) and subsequently by Berry (1972) and Chapman(1976). The complex faunal zonation of mangrove and mudflat inhabitants andthe limited geographical distribution of individual species mean that mudskipperfaunal associations are difficult to construct with any degree of accuracy andthe absence of Macrophthalmus latreilli and Uca coarctata from faunal liststhat include mudskippers (Sasekumar, 1974; Frith et al., 1976; Frith, 1977;Nateewathana & Tantichodok, 1984) advise caution in attempting such generalisations. Additionally, while Metaplax crenulatus is found in association withBoleophthalmus it also occurs with Periophthalmodon (Berry, 1972; Sasekumar,1974) and Scartelaos (Frith et al., 1976). Nevertheless, in the same way thata specific mangrove fauna is a point for discussion, there may be some validassociations between mudskippers and other faunal elements. Furthermore,since fish distributions are given only in terms of their presence or absence froma particular transect or station, it is not possible to judge fully the level ofassociation. At present the best that can be attempted is to point out some genericrather than specific associations. The latter can be explored during additionaldetailed survey work. While associations probably involve all macrofaunalgroups, including polychaetes and molluscs, existing observations are restrictedto the burrowing surface-active crustaceans.
The general association of mudskippers and decapod crustaceans, particularlygrapsids and ocypodids has long been recognised, but the xanthids Erycarcinusnatalensis and E. orientalis, the synalpheid Alpheus sp. (usually A. eurythrosineor A. crassimanus), the pagurids Clibanarius padavensis and Diogenes avarusand the callianasids Thalassina anomola and Upogebia sp. (rarely identified)also need to be included (Table VI). Scartelaos has the fewest associations,but this simply may reflect limited sampling at lower tidal levels. Nevertheless,an investigation of the joint distribution of mudskippers and the crustaceangenera {Clibanarius, Alpheus, Uca, Macrophthalmus, Metaplax) that occurwith them could providegreater insight into habitat selection and utilisation bythe fishes because, in the case of Macrophthalmus and Uca, much moreinformation is available on the ecological correlates of their distribution (cf.Jones, 1984).
MUDSKIPPERS 549
Table VI
Associations between mudskipper genera and burrowing crustaceans. Speciesnames are only used if they are given by all authors. Data drawn from Gopala-krishnan, 1970; Berry, 1972; Day, 1974; Sasekumar, 1974; Frith etal., 1976;Frith, 1977; Dorjes, 1978; Branch & Grindley, 1979; Nandi & Choudhury,
1983; Nateewathana & Tantichodok, 1984.
Periophthalmodon Periophthalmus Boleophthalmus Scartelaos
Callianassidae
Thalassina anomala X X X
Upogebia sp. X X X
PaguridaeClibanarius padavensis X X X X
Diogenes avarus X X X X
SynalphidaeAlpheus spp. X X X X
Xanthidae
Eurycarcinus sp. X X X
OcypodidaeUca rosea X X X
U. lactea annulipes X X
U. vocans X X
U. mani X X X
U. dussumieri X X X
U. triangularis X X X
Tylodiplax tetratylophora X X X
Ilyoplax spp. X X X
Macrophthalmus spp. X X X X
GrapsidaeChiromanthes spp. X
Metaplax spp. X X X X
Sesarma spp. X X X
PREDATORS
One common explanation for the amphibious mode of life adopted by mudskippers has been that it has enabled them to exploit new food resources (Inger,1952) and to escape marine competitors or predators (Pearse, 1929, 1933;Stebbins & Kalk, 1961). At high tide, however, mudskippers are availableto such predators. Boleophthalmus tenuis [ = Scartelaos tenuis] are taken byHydrophis cyanocinctus, blue banded sea snakes (Volsloe, 1939) and the catfishArius sagar (Sasekumar et al, 1984). Stonefish, Leptosyanceja melanostigmaand possibly catsharks, Chiloscyllium griseus are predators of Boleophthalmusboddarti [ = B. dussumieri] (Clayton & Vaughan, 1988). In captivity Periophthalmodon schlosseri eat Scartelaos viridis (Khoo, 1966), and although this maybe a laboratory artefact, the possibility that mudskippers eat other fish is supported by field observations of the predation of Periophthalmus barbarus by thesame species of Periophthalmodon (Lim, 1971). Furthermore, fish remains inthe stomachs of Periophthalmodon schlosseri [ = P. freycineti] were tentativelyidentified as of periophthalmid origin (Milward, 1974).

Table VII
Heron, bittern and egret predators of mudskippers in the Sundarban region of the Ganges delta (based on Mukherjee, 197la, b)
Predator species No. birds % diet Total
examined as fish species N
fish eaten
Periophthalmus Boleophthalmus Other fishesRank N Rank N Rank N Family
Smaller egret 220 95 16 1292 1 225Egretta intermedia intermedia
Eastern large egret 70 80 24 1166 1 109Egretta alba modesta
Little egret 138 66 25 1073 2 95Egretta garzetta garzetta
Purple heron 70 57 32 963 3 81Ardea purpurea manilensis
Indian grey heron 76 34 24 1303 2 203Ardea cinerea rectirostris
Little green heron 26 29 10 108 1 55Butorides striatus chloriceps
Chestnut bittern 8 19 7 80 1 40
Ixobrychus cinnamomeus
Night heron 78 10 7 70 1 32Nycticorax n. nycticorax
67
8 41
86
76
147
2
3
202
183
CyprinodontidaeBagridae
2
2
1
3
106
106
212
86
MugilidaeCyprinodontidae
BagridaeAmbassidae
a><
5>
1
2
300
198
BagridaeAnguillidae
nr>
1
3
210
176
BagridaeCyprinodontidae
HO
2
•3
19
11
CyprinidaeBagridae
•2 10 Channidae
•2 10 Mastocembelidae
18 Bagridae

MUDSKIPPERS 551
In an extensive survey of the stomach contents of the water birds of theSundarban region of the Ganges delta (Mukherjee 1971a, b), mudskippers werethe commonest fishes taken by herons, bitterns and egrets (Table VII). Of the61 prey species of fishes only the bagrid Mystus guilo, the anguillid Anguillabengalensis and the cyprinodonts Oryzias melastigmus and Aplocheilus panchaxwere as numerically significant as Periophthalmus in the diet of these birds.Only Periophthalmus, however, was consumed by all the birds and indeed thethree least piscivorous species (little green heron, chestnut bittern and nightheron) can almost be considered as mudskipper specialists because mudskippersconstituted such a large percentage of the piscivorous portion of their diet. Thesmaller egret additionally takes Scartelaos histophorus [as Boleophthalmusviridis] (Mukherjee, 1971b). On Singapore Island Halcyon chloris humii, thewhite-collared kingfisher is largely a bird of the mangrove belt and associatedmudflats and takes crabs and Periophthalmus. As an occasional visitor to theintertidal mudflats Alcedo atthis bengalensis, the Indian common kingfisher,occasionally takes small Periophthalmus schlosseri [? = Periophthalmodonschlosseri] (Gibson-Hill, 1948). Mudskippers are also taken by gulls and terns(Clayton & Vaughan, 1986).
Especially in mangrove areas, there are a number of reptiles that seem to takecrabs and other invertebrates in their prey (Berry, 1972) and which are likelypredators of mudskippers. Morton & Morton (1983) suggest that Bennett'swater snake, the viper Enhydris bennetti, from Hong Kong may include mudskippers in its diet and in Madagascar the skink Cryptoblepharus boutoni takessmall Periophthalmus koelreuteri [ = P. kalolo or argentilineatus], grapsidcrabs and insects in its daily foraging forays into the intertidal zone (Fricke,1970). Boleophthalmus boddarti is second only to the swimming crab as apreferred food of the Ganges estuarine crocodile Crocodylus porosus, at leastin captivity (Banerjee et al, 1989). In Malaysia, snakes (Eggert, 1935) monitorlizards and fishing hawks take mudskippers (Johnstone, 1903).
On the Solomon Islands, Slooff & Marks (1965) caught female mosquitoesAdes (Geoskusea) longiforceps that were gorged on the blood of Periophthalmussp. [ = P.kalolo or argentilineatus].
PARASITES
Parasitologically, mudskippers are somewhat neglected, but in most investigationsreported to date, new species of parasite have been described. At the generallevel, larval ascarid nematodes infect the guts of Periophthalmodon schlosseri(Khoo, 1966) and cestode, agamofilarial and acanthocephalid cysts have beenfound in Periophthalmus koelreuteri (Pearse, 1933), Periophthalmodon schlosseriand Boleophthalmus boddarti (Khoo, 1966). Unusually all these three specieswere found to be infested with Gnathia sp., a parasitic copepod of the gills(Pearse, 1933) but heavy infestation of parasitic copepods in the gills andbuccal cavity of Pseudapocryptes lanceolatus were also present (Das, 1934). InPeriophthalmus cantonensis [ = P. modestus] stomach spirurioids and duodenallarval cestodes were also noted by Pearse (1932). In Boleophthalmus boddartiand B. viridis [ = Scartelaos histophorus] no external infections on the skin orgills could be found by Choudhury & Nandi (1973). However, the former, butnot the latter, was found to contain protozoan parasites. In the gut and gallbladder two species of myxosporidian sporozoans were identified and a third

552 DAVID A. CLAYTON
species was recorded for the first time from an estuarine as opposed to a freshwater fish (Choudhury & Nandi, 1973). Pearse (1933) also noted the presenceof intestinal flagellates in Boleophthalmus boddarti and Morii & Kasama (1989)have reported on the bacterial flora in the digestive tracts of B. pectinirostris andPeriophthalmus cantonensis [ = P. modestus]. Only some of the bacteria foundin the mud substratum on which the fish live were found in the digestive tracts,Vibrio or coryneforms being the commonest. It was suggested that osmoregulatory sea water is the main factor controlling the intestinal flora of the fish(Morii & Kasama, 1989). From P. barbarus in west Africa a new species ofacanthocephalan parasite (Troncy & Vassiliades, 1974 [as P. papilio] and ahemiurid trematode (Fischthal & Thomas, 1971 [as P. koelreuteri]) weredescribed. In the mesentries of Periophthalmodon schlosseri from Singapore,Khan & Ip (1988) also found and described a new acanthocephalan species.
POLLUTION STUDIES
Benthic organisms are useful in monitoring heavy metal pollution as they oftenconcentrate potential pollutants in their tissues. In two marine areas surroundedby heavy industrialisation, the Ariake Sea in Japan and Bombay Harbour inIndia, the tissues of Boleophthalmus pectinirostris (Uchida et al., 1971) and B.boddarti (Patel et al., 1985) respectively, have been assayed for heavy metallevels. The Indian study was more comprehensive, but of the elements commonto both studies, iron accumulation was 10-15 times greater than that of zinc.Cadmium appeared not to be accumulated to any great extent by these fishes.In the Indian study mercury was not sampled, but the oversight was correctedby Patel & Chandry (1988) and Mahajan & Srinivasan (1988), the latter authorsfinding greater accumulation of mercury (2.6 /xg-g"1 dry weight) in B. boddartiin the 4 months following the monsoon than in the 4 months before it (2.36fxg-g~x). This was also true of sediment values and was accounted for by aninput of freshwater containing industrial effluent into the sea. The effects ofmercury and fluoride poisoning on mudskippers have been investigated throughexperimental exposure of the fishes to the pollutants. In B. dussumieri [asB. dentatus] from the west coast of India, mercury had an inhibitory effect onintestinal (Lakshmi et al., 1991) and gill ATPases (Lakshmi et al., 1990).Mercury pollution adjacent to industrial areas was chronic because its concentration was more than twice that required to kill the fish (Lakshmi et al., 1991).Also from the Gujarat coast of western India, B. dussumieriexposed to sublethaldoses of sodium fluoride exhibited cell membrane rupture, degeneration andvacuolarisation of the liver (Shaikh & Hiradhar, 1987) and inhibition of liverand muscle acid and alkaline phosphatase activity (Shaikh & Hiradhar, 1988).
B. boddarti from Bombay Harbour (Bangera & Patel, 1984) and Periophthalmus schlosseri [ = Periophthalmodon schlosseri] {Paid et al., 1975) havebeen used to monitor levels of radionuclides. Tissue concentrations of theelements recorded in all these studies on pollution monitoring were greater thanthat found in the water column, but were often considerably less than that foundin the sediments.
FEEDING
The behaviour of Periophthalmus and Periophthalmodon when catching decapodcrustaceans consists of a swift tail-powered lunge (Khoo, 1966; Lim, 1971),

MUDSKIPPERS 553
after which the fish may or may not return to the water (Stebbins & Kalk, 1961;Gordon et al., 1969; Sponder & Lauder, 1981), the latter issue being of interestto respiratory physiologists (see p. 530). Appetitive behaviour is variable. InPeriophthalmus waltoni waiting, stalking and fast approaches are all used, but,successful or not, they avoid returning to the attack site for a long time, possiblybecause prey in the area do not re-surface immediately the fish has moved away(Clayton & Snowden, unpubl.).
Boleophthalmids graze on the surface sediment with distinctive side to sidemovements of the anterior body and skim off the algae and diatoms that are theirpredominant food items (Khoo, 1966; Lim, 1967; Clayton & Vaughan, 1988).Feeding occupies two-thirds of the time for which B. dussumieri is active,choosing areas of its territory where the diatom densities are high (Clayton &Wright, 1989). Scartelaids also feed by scraping the top layer of the sediment(Khoo, 1966) but, additionally, may arch slightly into the air before biting intothe substratum (pers. obs.). Whether these lunges are directed at prey movements is not known.
Generally, mudskippers of the genera Periophthalmus and Periophthalmodonare considered as carnivores while those of Boleophthalmus are herbivores andScartelaos are omnivores. There is no information on the other genera althoughthe structure of their dentition, including that of their pharyngeal plates (seeSponder & Lauder (1981) for SEM of plate of P. barbarus), should clarify theissue. While Apocrytodon madurensis is very small (40-50 mm), its pharyngealplates (and the manner of feeding) are very similar to that of Boleophthalmusdussumieri (Clayton, unpubl. data). Relative gut lengths for the different generaare consistent with the generalisation (Pearse, 1932; Table VIII). Differences inthe way in which the ratios were calculated prevent cross-study comparisons,but within studies (Table VIII) the carnivorous species have the shortest,omnivores intermediate and herbivores the longest intestinal length. Two otherspecies are herbivores: Apocryptes bato with a ratio of 2-0 (Hora, 1935a) andPseudapocrytes lanceolatus [as Apocryptes lanceolatus] for which recordedvalues are 3-43 (Pearse, 1932) and 2-7 (Hora, 1935a).
Table VIII
Relative gut lengths for genera of mudskippers
Periophthalmus Periophthalmodon Boleophthalmus Scartelaos Reference
0.5-0.6 1.09 2.55 1.266 Khoo, 1966
0.76 — — — Pearse, 1932
0.64 0.43 1.45 — Pearse, 1933_ 0.6 — — Hora, 1935a
0.39 0.8 — — Lim, 1967
1.0 — — 2.0 Lim, 1971
0.28-0.45 0.42—
0.6 Milward, 1974
The detailed study of the diet, however, reveals that all the fish are omnivorous to some extent and that there are dietary differences between juvenilesand adults. Mutsaddi & Bai (1969a) examined the stomach contents of over 1000specimens of Boleophthalmus boddarti collected from the Bombay coast atmonthly intervals over a year. The fish ingest algae, diatoms, polychaetes,nematodes, crustaceans and teleost eggs. Monthly percent occurrence values

554 DAVID A. CLAYTON
showed that crustaceans formed the dominant food for juvenile fish (total n for1 year = 330, 16—95 mm SL, no gonadal development), while for adults(n = 680, 96-185 mm) plant material, particularly diatoms, were dominant.There was also considerable seasonal variation in their diet. Crustacean larvae
were common in January and February but were superseded by copepods inMarch and April. Nematodes were present in the diet mainly in July to Octoberand diatoms were only a significant component from July to December. Therewas similar variation in the adults' diet; teleost eggs only appearing in July andnematodes predominantly in September to December. These variations in dietprobably reflect the seasonal availability of the prey items rather than any selectivefeeding process. This was demonstrated for diatoms where there was a goodcorrelation between the percent occurrence in the stomach and their availabilityin the mud (Mutsaddi & Bai, 1969a). Prince Jayaseelan & Krishnamurthy (1980)record that Boleophthalmus sp. [ = dussumieri] from the Pichavaran mangroves(11°29'N:20°49'E) is a herbivore, but later (Krishnamurthy et al., 1984)mentioned that it ingested nematodes and missed a good opportunity to verifyif the seasonal variations of nematode species abundance was reflected in themudskippers' diet.
Based on the percentage of empty stomachs, juveniles apparently have a lowerfeeding intensity than adults (Mutsaddi & Bai, 1969a). This result contrasts withthat of Sarker et al. (1980) on 159 Pseudapocryptes dentatus [ = Boleophthalmusdussumieri] from the Shatt Al Arab in the northern Arabian Gulf. They foundthat food consumption, as measured by weight of stomach contents per grambody weight, was least in the largest fish (160—209 mm total length). Thecollection was made on a single day (26 November 1976) and no data onseasonal variation are available. This may partially explain why Sarker et al(1980) failed to find any animal material in the fishes' diet. As strict herbivoresonly benthic diatoms were taken including, in decreasing order of importancein the diet, the bacillariophytes Pleurosigma, Navicula, Nitzschia, and Synedra,the chlorophyte Closteriopsis, and the cyanophyte Oscillatoria (Sarker et al,1980). Using the same collection offish, Pankow & Hug (1979) provided taxo-nomic information on the ingested diatoms and identified 114 species, of whichNitzschia (23 spp.) and Navicula (15 spp.) were the dominant genera. Diatomsof the genera Nitzschia, Navicula and Oscillatoria were also predominant in thediet of Boleophthalmus boddarti in Malaysia (Lim, 1967). In addition algae,fungal material and some nematodes were eaten. In contrast to the findings ofMutsaddi & Bai (1969a), Khoo (1966) found that 80% of the diet of 32-41 mmjuveniles consisted of diatoms; in 20-40 mm SL juveniles Lim (1967) found thediet to be strictly bacillariophylic. The similarities and differences between thediets of Boleophthalmus boddarti and Scartelaos viridis [ = S. histophorus] arewell illustrated by Khoo (1966). In both species the diatoms Oscillatoria sp.,Pleurosigma spp., Nitzschia sp. and the cyanobacta Lyngbya sp. are importantcomponents of the diet. Both also take harpacticoid copepods, chromadoroidmolluscs and nematodes but these form a significant part of the diet only inScartelaos (Khoo, 1966). The diet of Scartelaos histophorus from Australia(Milward, 1974) is also clearly that of an omnivore. Diatoms and nematodeswere the major food items in all sizes offish (19-112 mm SL) while ostracodsand harpacticoid copepods may be less important in the larger fish (66-112mm). Algal filaments, dipteran larvae, kinorhynchs and crustacean nauplii werealso included in the diet (Milward, 1974).

MUDSKIPPERS 555
There are differences in diet between juveniles and adults. In a study of thecombined age and locality differences in the feeding of P. barbarus [nowPeriophthalmodon schlosseri] (Lim, 1967), those individuals inhabiting abrackish water stream habitat took Formicidae, Coleoptera, dipteran nymphsand molluscs, the last being the only food item in common with those individualsthat lived downstream in an estuarine habitat. Crabs of the genera Sesarma andUca, polychaetes and fish fry completed the diet of these estuarine fish. In thestream habitat, ants were eaten by the juveniles and beetles by the adults; in theestuarine habitat polychaetes were only consumed by juvenile fish (Lim, 1967).
The catholic diet of species of Periophthalmus can partially be attributed totheir extreme opportunistic approach to prey items. Crabs (61 %), insects (29%),of which over a quarter were ants, small gastropods (9%), nemertine worms,Orthoptera and arachnids were taken by Periophthalmus vulgaris [ = P.argentilineatus] (Lim, 1971). Also from the same general area, Macintosh(1979) identified the crab prey of P. vulgaris [ = P. argentilineatus] as smallindividuals of Uca rosea, while Frith & Brunenmeister (1980) report that thesame species took small U. forcipata. In terms of percentage by volume ofstomach contents, shrimp (25%), spiders (12%), copepods (10%), snails (9%) andcrabs (8%), were the commonest items taken by Periophthalmusargentilineatus[as P. cantonensis] from India (Pearse, 1932). As in other periophthalmidsinsect larvae, ostracods and amphipods completed the diet. Foraging P. waltoniin Kuwait take the small ocypodid crabs Tylodiplax indica, Cleistostoma dotil-leforme and Ilyoplax stevensi (Al Taher, 1990; as P. koelreuteri) and Uca sp.,Alpheus sp., nematodes and polychaetes (El Zaidy et al., 1975; as P. chrysospilos) and along the Gujurat coast of north western India, Siddiqi (1974) reportsthat the gut content of P. waltoni was mostly small-shelled molluscs. OnMadagascar, P. sobrinus [ = P. argentilineatus or kalolo] takes polychaetes{Polydora, Dendronereis), crustaceans {Upogebia sp., Uca spp. including U.chlorophthalmus; Dotilla fenestrata), tanaids, and insects including ephydridflies and dipteran larvae (Stebbins & Kalk, 1961; MacNae & Kalk, 1962).
Secondary sources (e.g. Lim, 1971) quote Khoo (1966) as recording Periophthalmus chrysospilos from Singapore as a herbivore because only diatoms,filamentous algae and other plant material were found in the stomachs of sixindividuals. Khoo (1966) qualifies these findings, however, by pointing out thatthe relative gut length of this species is that of a carnivore and that the other twoindividuals examined, which were from a different location, contained crustaceans {Ilyoplax sp.), harpacticoid copepods and polychaetes. It was concludedthat the fish has the ability temporarily to utilise plant food sources when unableto take animal prey. The reason for the inability to take animal prey, however,remains obscure.
In a study of Queensland's mudskippers (Milward, 1974), the diet of threesympatric species of Periophthalmus were compared. Arthropods dominated thediet and included harpacticoid copepods, brachyurans, dipteran larvae andadults. Tanaids, ostracods, collembolans and ants were also eaten and the commonest non-arthropod items were polychaetes and gastropods. Nematodes,foraminiferans and algal filaments were present in insignificant amounts.Copepods and dipteran larvae were mostly taken by the small fish and adultinsects and crabs by the large ones. This size difference in diet applied bothwithin and between species such that P. expeditionium [ = P. novaeguineaensis](22-95 mm SL) was intermediate in size between P. gracilis (18—64 mm) and

556 DAVID A. CLAYTON
P. vulgaris [ = P. argentilineatus] (14—104 mm) and resulted in some resourcepartitioning (Milward, 1974).
Species of Periophthalmodon are crab specialists. Only Lim (1971) mentionsthe presence of ants and small bivalve molluscs as part of the diet of P. schlosserithe major portion being composed of crabs of the genera Metaplax, Ilyoplax,Sesarma and Uca. Similarly, the diet of Periophthalmodon freycineti [asP. schlosseri] consisted predominantly of crabs of the genera Macrophthalmusand Uca with a few ants and fish. The fish were probably smaller periopthalmids(Milward, 1974; see also p. 549). In terms of volumetric composition ofidentifiable food items, crabs also dominated the diet of Periophthalmodonschlosseri from Klang (3°24'N:101°23'E) (Sasekumar etal., 1984). However,the sipunculid Phascolosoma arcuatum was also common and mangrove plantdetritus and gastropods were present in small quantities (Sasekumar et al,1984). Khoo (1966) only found sesarmid crabs in the diet of Periophthalmodonschlosseri while Macintosh (1979) found only Uca dussumieri and U. rosea. Ina series of prey presentations, only 15—25 mm carapace width fiddler crabs wereeaten by Periophthalmodon; crabs larger than 30 mm carapace width wereattacked and sometimes injured but not ingested. It was suggested this size-dependent predation was one reason for the excess of large males in the fiddlercrab population (Macintosh, 1979).
BEHAVIOUR
GENERAL
There are many short descriptive accounts of general behaviour (Petit, 1921;Champeau, 1951; van Dijk, 1959, 1960, 1978; Klauzewitz, 1967; MacNae,1968c; Vestergaard, 1972; Burhanuddin & Martosewojo, 1979), locomotion(van Dijk, 1960; De & Nandi, 1984), courtship and nesting (Petit, 1922, 1928;Freitas, 1961a, b; Brillet, 1969a; Magnus, 1972) and aggression and territoriality(Mutsaddi & Bai, 1969b; Brillet, 1969b,c; Nursall, 1974; Clayton & Vaughan,1982). Similarities in the general aspects of the biology and life cycles ofsmall substratum-bound fish (Gibson, 1969, 1982) can be extended to includebehaviour. In Madagascar the behaviour of Periophthalmus sobrinus [ = P.argentilineatus] and Lophalticus kirkii, an amphibious blenny that inhabitsrocky shores, have many behavioural characteristics in common (Brillet, 1986).There are also important differences, but these might have been less marked hadmudskippers from a similar rocky habitat been studied (cf. Gordon et al, 1968;.see p. 545).
As well as providing a detailed functional analysis of skeletal and musculatureinvolvement in fin adaptations for terrestrial locomotion, Harris (1961) alsodescribes the behaviour. In water P. barbarus [as P. koelreuteri] employspaddling, fast swimming and skimming whereas on land, they use crutching,skipping and climbing. Apart from climbing, Boleophthalmus dussumieri [as B.boddarti] exhibits similar locomotory behaviour (Clayton & Vaughan, 1988).
RHYTHMIC BEHAVIOUR
While Gordon et al. (1968) found that Periophthalmus sobrinus [ = argentilineatusor kalolo] was active at all hours, Stebbins & Kalk, (1961) and Magnus (1972)

MUDSKIPPERS 557
only reported diurnal activity. The study of activity rhythms tends to support thelatter observations. Al Naqi (1977) found that P. waltoni [as P. chrysospilos]was strongly diurnal. Under a 12L:12D regime in the laboratory it showed twoactivity peaks in the light period, the larger starting at the beginning of theperiod, the smaller occurring towards the end of it. Under continuous light ordark a similar periodicity of activity was maintained, but at a much lower levelin the dark (Eissa et al., 1978). Behavioural rhythms have also been investigatedin Boleophthalmus pectinirostris [as B. chinensis] (Ishibashi, 1972; Ishibashi& Nishikawa, 1973) and Periophthalmus modestus [as P. cantonensis](Nishikawa & Ishibashi, 1973,1975a,b). Using a photocell to record entry andexit from an artificial burrow Ishibashi (1972) demonstrated that, under constantconditions, Boleophthalmus pectinirostris was rhythmically active. Its peaks ofactivity coincided with the time of low tide on the shore from which it wascollected and overall activity was low at times of neap tides and high at springtides. This 15-day activity cycle was confirmed under conditions of continuouslight or darkness by Ishibashi & Nishikawa (1973). They briefly report that thefish have both a circadian and a tidal component to their rhythm, but a moredetailed verification is required. The equally brief report on endogenous rhythmsin Periophthalmus (Nishikawa & Ishibashi, 1975a) explains some of the falsestarts in experimental procedure (19-h feeding entrainment and operant conditioning) leading to their later work on entrainment of activity by feeding(Nishikawa & Ishibashi, 1975b). Under conditions of continuous darkness, thefree running activity apparently showed some synchronisation with times of lowtide, but the data were only averaged on a 24-h and not a tidal periodicity. Clearsynchronisation of activity was achieved with 12-h (for 88 days) and 24-h (for22 days) feeding regimes, the anticipation of which was evident from pre-feeding bursts of activity, particularly under the shorter regime (Nishikawa &Ishibashi, 1975b).
BURROWING AND TERRITORIALITY
It has long been known that mudskippers build burrows (Petit, 1922, 1928;Harms, 1935; Champeau, 1951), but the common observation offish migratingacross the intertidal zone following changing tidal levels (Hora, 1935a; Stebbins& Kalk, 1961; Khoo, 1966; Gordon etal., 1968; MacNae, 1968b) made the ideaof territoriality in terms of a permanent burrow occupancy difficult to accept(Gibson, 1969). Thus, in his short study of mudskippers in Australia, Nursall(1981) who cited MacNae (1968a), was of the opinion that none is territorial. The mudskippers concerned were Periophthalmus argentilineatus [asP. vulgaris] and P. gracilis, Periophthalmus which migrate with the tide, andP. minutus [as Periophthalmus sp.] and Periophthalmodon freycineti [asP. schlosseri], species that were associated more clearly with identifiableburrows. These observations contrast with those showing that Periophthalmusargentilineatus and P. gracilis in the same study area, construct and inhabitburrows (Milward, 1974). Milward was not quoted by Nursall (1981). Kobayashiet al (1971) imply that P. modestus [as P. cantonensis] also migrates with thetide, resting on the edge of the high tide line. However, they spend the coldseason in burrows and subsequently use them for spawning.
Of the two forms of territories of P. sobrinus [ = P. argentilineatus ] inMadagascar, one involves fish moving between high shore burrows on the bank

558 DAVID A. CLAYTON
and low shore feeding territories in the tidal channel bed (Brillet, 1975). Moreconventional, permanent residency territories occur in areas away from bankedchannels. Territory size depends on fish density but each usually contains asingle burrow system from which the territory is defended (Brillet, 1975),although in P. waltoni there are several burrow systems in a single territory(Clayton & Snowden, unpubl.). Single burrow systems were also found interritories of Boleophthalmus (Mutsaddi & Bai, 1969b; Clayton & Vaughan,1986, 1988). As well as resident burrow occupants that are territorial, populations also contain non-territorial errant fish that may range more widely over themudflat. In B. dussumieri, territorial individuals live in well-defined, contiguous, polygonal mud-walled territories (Clayton & Vaughan 1982, 1988). Ifresidents are removed, smaller errant individuals occupy vacant territorieswithin one or two tidal cycles (Clayton & Vaughan, 1986). This division alsoapplies to Periophthalmus argentilineatus where errant fish are usually smallerand less aggressive than the territorial ones (Brillet, 1975). In captivity thedifference results in the development of a dominance system comprisingdominant, territorial burrow occupants and subordinate non-territorial ones(Brillet, 1969c, 1975). Thus fish that follow the tide are likely to be errantindividuals, including juveniles and adults unable to establish dominance.
The core, but not the geometric centre, of the territory is the burrow. Burrowstructure and construction behaviour has been described for P. modestus (Kobashiet al., 1971; Matoba & Dotsu, 1977), P. sobrinus [ = P. argentilineatus orkalolo] (Freitas, 1961b; Brillet, 1969a, 1976), Boleophthalmus dussumieri(Clayton & Vaughan, 1986) and Periophthalmodon schlosseri (Harms, 1929;Verwey, 1930). The generalised structure consists of a lower, near verticaltunnel connected to the surface by two, less steep entrances. The variability ofburrow architecture has been illustrated schematically (Brillet, 1969a, 1976;Matoba & Dotsu, 1977), photographically in cut away section (Kobashi et al,1971) and by casts (Freitas, 1961b; Clayton & Vaughan, 1986); the last authorsclearly showing that tunnels of smaller infauna anastomose with those of thefish. The specific architecture is so profoundly affected by local topography andtidal conditions (Brillet, 1969a, 1976) that generalisations of species-specificconstructions (MacNae, 1968b) are to be treated with caution. While surfaceembellishments variably consist of turrets and saucer-like depressions with orwithout an elevated rim, in some species there are none at all (Milward, 1974).Species identification should not be attempted on burrow structure alone. Thesituation is exacerbated in situations where burrows may be appropriated byother species. Additionally, fish escape down the nearest burrow at the approachof an observer and may not be the true occupant: this may also account for thereports of paired fish (Freitas, 1961b; MacNae, 1968b). Structural modificationsrelated to breeding are unlikely to be directly related to surface structures (cf.MacNae, 1968b), but consist of diverticulae from the base of the descendingtunnel in which the eggs are laid (Kobashi et al., 1971; Brillet, 1976). Burrowdepth can exceed 1.5 m (Clayton & Vaughan, 1986) and the general assumptionis that they descend to the water table, but Brillet (1976) noted that eggs requireperiods out of water for successful hatching. However, while the breedingburrows of Periophthalmusmodestus contain an egg chamber (Kobayashi et al.,1971), Matoba & Dotsu (1977) also showed that, irrespective of the diameterof the original, the opening of the breeding burrow is restricted by muddeposition to such an extent that females may have to squeeze through an

MUDSKIPPERS 559
opening smaller than themselves. Construction and maintenance is performed byremoving mouthfuls of mud from the burrow and depositing them on the surface(Petit, 1928; Brillet, 1969a; Clayton & Vaughan, 1988) where, in periophthalmids,they may be used in turret construction or burrow modification (Brillet, 1969a;Matoba & Dotsu, 1977). Boleophthalmus dussumieri is unique in that the mudgobbets are used to construct mud walls at some distance from their burrow.Neighbouring fish may contribute to building the same wall (Clayton &Vaughan, 1986). Wall construction behaviour is density dependent and servesto reduce aggression in neighbouring territorial fish (Clayton, 1987). Burrowsfunction as a refuge, observation post and nest (Brillet, 1975). Additionally,because mudskippers are poor swimmers, the burrow will be an importantrefuge from piscivorous predators at high tide (Milward, 1974).
HABITAT SELECTION
Habitat selection has not been studied in juvenile fishes, despite its relevance toresolving the issue of occupancy of non-muddy shores. The salinity preferencesof adult Periophthalmus sobrinus (Gordon et al., 1968) and P. waltoni (Al-Naqi,1977) were assessed by choice trials in groups of fish pre-adapted to differentlevels of salinity. Both studies showed that seawater-adapted fish preferred theterrestrial habitat to any dilution of sea water and that 100% sea water wasgenerally the least preferred salinity. In a group choice experiment without pre-adaption, P. modestus [as P. cantonensis] also preferred land over water of anysalinity (Gordon et al, 1985). Fish also preferred water at 30 °C over water at10, 20 or 35°C and darkness to light. In terms of behavioural selection ofmicrohabitat, these results are difficult to interpret, but in any case are confounded by social groupings and inappropriate analyses. Dominance relations incaptive groups would interfere with fish distribution and the preference for theterrestrial habitat could have simply resulted from the fact that it occupiedconsiderably more of the choice chamber than did the water.
SOCIAL BEHAVIOUR
Social behaviour in mudskippers is similar to that of other substratum boundfishes, involving erection of fins in a variety of combinations and leaping intothe air (Gibson, 1982). Courtship has been described in Periophthalmus (Khoo,1966; Brillet 1969b, 1970, 1980a, 1984; Magnus, 1972) and Boleophthalmus(Khoo, 1966; Lim, 1967; Clayton & Vaughan, 1988). As in many speciesagonistic elements occur during courtship, but these have been separatelydescribed for Periophthalmus (Brillet, 1969c,d, 1970, 1975, 1980b, 1981,1983) and Boleophthalmus (Lim, 1967; Clayton, 1987; Clayton & Vaughan,1988). The territorial status of interacting fish determines the nature of the socialinteraction. Errant or subordinate fish exhibit a marked lack of aggression ininteractions with each other and do not resist or display to an attack by adominant fish. In established territories, dominant fish avoid each other or usea ritualised approach, but directly attack subordinate ones (Brillet, 1969c,d).The direct attack is similar to the approach to a prey and is without fin erectionwhereas the ritualised approach is slower and less direct with fin displaysoccurring at closer approach. In prolonged encounters the fish circle each otherin an 'O' formation (Brillet, 1969c). In a more quantitative analysis of agonisticbehaviour, 16 non-exclusive elements were reported (Brillet, 1980a, 1981). Fish

560 DAVID A. CLAYTON
that initiated agonistic encounters always chased their opponents and invariablyused tail-assisted bounds in doing so. Dorsal fin erection, posturing by raisingthe body on pelvic and pectoral fins and physical contact including head-buttingand biting were also common. The last two elements were difficult to distinguish, because they were extremely fast, erupting from essentially static displaypostures. Tail undulation and butting or biting were much reduced in juvenilefish (43—55 mm TL) that initiated encounters when compared with adults(90—100 mm TL). The recipient of such attacks rarely reciprocated, onlyoccasionally erecting its dorsal fins (Brillet, 1980b). The influence of sex andsize was assessed by recording victories and defeats among small captive groups(Brillet, 1981). Larger individuals, irrespective of sex, dominated smallerindividuals but during encounters between fish of the same size males dominatedfemales. Males initiated and won a greater percentage of encounters thanfemales. Dorsal fin erection constituted the greatest percentage of all displaysin small males and least in large males with that of females being intermediate.In contests initiated by large males, dorsal fin erection of the opponent was lowirrespective of whether it was female or a large or small male. When respondingin a contest initiated by a female or a small male, large males gave a strongerresponse than females (Brillet, 1981).
Size was also an important factor in resolution of intraspecific agonisticencounters in four species of Periophthalmus (Nursall, 1974). The specificidentity of these fish was only known later (Nursall, 1981) and in Nursall (1974)P. argentilineatus, P. novaeguineaensis, P. gracilis and P. minutus were described as BB, RS, SBB and RF respectively. While the largest species P. argentilineatus dominated all the other species, P. novaeguineaensis, the second-ranked in size retreated before the smaller P. gracilis and P. minutus, which inturn ignored each other. Species-specific differences in agonistic behaviour areevident because, whereas P. argentilineatus rarely raised the first dorsal finalone (Brillet, 1980b), the periophthalmids observed by Nursall (1981) showedvarious combinations of first, second and caudal fin erection in their displays.Nevertheless, and in agreement with Brillet's (1981) observations, the moreaggressive species used such displays more frequently. Combinations of first andsecond dorsal and caudal fin displays were a feature of the agonistic displays ofBoleophthalmus dussumieri (Clayton & Vaughan, 1988). While chromatophorebody colour changes have been noted in courtship (Magnus, 1972; Matoba &Dotsu, 1977), only Nursall (1981) indicates that colour change is involved inagonistic display.
While there are clear species-specific differences in courtship (Brillet, 1984),the general organisation of mudskipper courtship is similar for B. dussumieri(Clayton & Vaughan, 1988), Periophthalmus kalolo [as P. koelreuteri] (Brillet,1984), P. argentilineatus (Brillet, 1969c) and P. modestus (Matoba & Dotsu,1977). The last authors provided diagrammatic representations of the male-female interactions during the complete sequence of the 'nuptial parade' asBrillet (1969b) so aptly describes it.
From the vicinity of their burrows, males advertise their presence by leapinginto the air with all fins fully spread and, at the approach of a female, give aseries of substratum bound displays. These displays, which involve orientationof the body in relation to the female, mouth gaping and buccopharyngealexpansion or contraction and fin display, may be extended, repeated or becomeaggressive depending on the responsiveness of the female. The female's role is

MUDSKIPPERS 561
largely passive requiring only that she remains stationary or follows the male.Furthermore, the fewer dorsal fin erections she gives, the more likely thecourtship is to progress. After successful display, which in periophthalmidsinvolves some tail waving, the male leads the female to the burrow. InBoleophthalmus tail waving has become a more pronounced static displaytermed quivering (Clayton & Vaughan, 1988). The male usually enters the nestfirst and spawning can take up to 5 h.
Many agonistic elements occur during courtship, sometimes leading to itstermination especially if interrupted by the male chasing an intruder (Brillet,1970). Dorsal fin erection is an agonistic act that must be absent from the laterstages of successful courtship such as tail beating or quivering (Brillet, 1969b,1980a; Clayton & Vaughan, 1988). The dorsal fin displays always involve bothdorsal fins in P. argentilineatus, whereas in the sympatric species P. kalolo, theymay be raised independently as well as together (Brillet, 1984). The absence offirst dorsal fin erection when male P. kalolo are close to females is seen as a
mechanism that leads to reproductive isolation of the two species. The abilityto erect the fins independently, on the other hand, is seen as a mechanism thatincreases the chances of successful courtship. In P. argentilineatus there is noscope for signalling different levels of aggression (Brillet, 1984).
CONCLUSIONS
That mudskippers can provide a rich source of comparative studies in adaptationto the littoral habitat should now be apparent. While some suggestions for futurestudy have been mentioned in the relevant sections, there are a number of issuesthat bear repetition. The problems of specific identification should be diminishedby the recent taxonomic revision of the Oxudercinae. If researchers also providevoucher specimens the confusion that previously hindered progress should alsobe reduced. Now that the artificial propagation of larval fish is possible, thepotential for commercial rearing of mudskippers has increased. Successfulaquaculture, however, requires that all aspects of reproductive biology areunderstood and more data on natural reproduction and larval development areneeded. The study of larval stages and the factors that determine benthicsettlement would also contribute towards understanding adult distribution andhabitat selection, two areas where there is currently little information. Indirectlythese studies may benefit the investigation of the physiological mechanismsof respiration, excretion and osmoregulation. It is likely that some of the conflicting results in these physiological studies are related to variations inresponsiveness and/or the mechanisms of adaptation that are dependent onfactors of age or size. More control of these variables is certainly required, asis attention to antecedent experimental conditions. That these are important isborne out by the tolerance to a wide range of salinities exhibited by fish that havefree access to water and air when compared with those confined to one medium.This also illustrates that the mudskippers' behavioural control mechanisms arewell developed, a feature more typically associated with thermoregulation thanwith other physiological systems. The full potential of comparative study inthese closely related gobies has yet to be explored. Few data on Pseudapocryptesand Apocryptes are available, yet the former have considerable capabilities toresist drought. In northern latitudes the cold winters induce mudskippers to

562 DAVID A. CLAYTON
hibernate, while in subtropical and tropical latitudes the summer heat causesthem to aestivate. The partitioning of gaseous exchange between the variouscomponents of the respiratory system, and the characteristics of blood transportmechanisms need to be elucidated. Mudskippers provide wide opportunities forecological and behavioural studies. Unfortunately, their pelagic larvae andparticularly the difficulty of access to breeding burrows, prevent any realisticmeasures of reproductive success from being made and exclude any seriousinvestigation of evolutionary issues. Nevertheless, mudskippers are eminentlysuitable for studies of the proximate causes of territorial, courtship and agonisticbehaviour. Studies of resource partitioning and predator-prey relationships aretwo areas for further study.
ACKNOWLEDGEMENTS
This review was initially supported by Grant SZ032 from the University ofKuwait, continued at the Plymouth Marine Laboratory and completed at SultanQaboos University.
REFERENCES
Note: References or parts of references enclosed in brackets [ ] are translations of theoriginal text.
Afzelius, B. A., 1956. Seitenorgane und Schleimkanalknocken bei Periophthalmuskoelreuteri und Gobius minutus. Zeitschrift fur Anatomie und Entwicklungs-geschichte, 119, 470-484.
Al-Kadhomiy, N. K. & Hughes, G. M. 1988. Histological study of different regions ofthe skin and gills in the mudskipper Boleophthalmus boddarti with respect to theirrespiratory function. Journal of the Marine Biological Association of the UnitedKingdom, 68, 413-422.
Al-Naqi, Z. A., 1977. Ecological and physiological studies on the mud-skipper fish,Periophthalmus chrysospilos, inhabiting the muddy shores of Kuwait. MSc thesis,Kuwait University, Kuwait, 133pp.
Al-Nasiri, S. K. & Hoda, S. M. S., 1975. Survey of the fish fauna of Shatt-Al-Arab(from Abu Al-Khasib to Karmat Ali). Bulletin of the Basrah Natural HistoryMuseum, 2, 36—46.
Al Taher, E. Y., 1990. The impact of the Doha west power station effluent on the uppereulittoral benthos of western Sulaibikhat Bay, Kuwait. MSc Thesis, KuwaitUniversity, Kuwait, 208pp.
Arai, R. & Kobayasi, H., 1973. A chromosome study on thirteen species of Japanesegobiid fishes. Japanese Journal of Ichthyology, 20, 1-16.
Asano, N., 1936. The egg laying habits of the South Sea mudskipper, Periophthalmussp. Ecological Review (Sendai), 2, 137—1139.
Bai, P. A. & Kalyani, M., 1960a. On the composition of the scales and skin ofBoleophthalmus boddarti Pall, and Mugil dussumieri (Val.). Journal of AnimalMorphology and Physiology, 7, 71-77.
Bai, P. A. & Kalyani, M., 1960b. Studies on ascorbic acid in fish tissues I. Journal ofAnimal Morphology and Physiology, 7, 162—165.
Bai, P. A. & Kalyani, ML, 1960c. Studies on the ascorbic acid content of fish tissues.II. Journal of the Zoological Society of India, 12, 216—219.

MUDSKIPPERS 563
Bai, P. A. & Kalyani, M., 1961. The relative content of imino acids in the skin andscales of some teleosts. Journal of Animal Morphology and Physiology, 8,126-129.
Bandurski, R. S., Bradstreet, E. & Scholander, P. F., 1968. Metabolic changes inthe mud-skipper during asphyxia or exercise. Comparative Biochemistry andPhysiology, 24, 271-274.
Banerjee, R., Nandi, N. C. & Raut, S. K., 1989. Food and feeding habits of the estuarinecrocodile Crocodylus porosus Schneider in captivity. Acta Biologica CracoviensiaSeries Zoologica, 30, 95—98.
Banerji, T. K., 1973. A comparative histological investigation on the interrenal gland ofsome Indian teleosts. Anatomische Anzeiger, 133, 20—32.
Bangera, V. S. & Patel, B., 1984. Natural radionuclides in sediment and in arcid clam(Anadara granosa L.) and gobiid mudskipper {Boleophthalmus boddaerti Cuv. &Val.). Indian Journal of Marine Science, 13, 5—9.
Baumeister, L., 1913. Uber die Augen der Schlammspringer {Periophthalmus undBoleophthalmus). Zoologische Jahrbucher, Abteilungfur Anatomieund Ontogenieder Tiere, 35, 341-354.
Berg, L. S., 1949. [Freshwater fishes of Iran and neighbouring countries]. Studiesof the Zoological Institute of the Academy of Science of the USSR Moscow, 8,753-758. (In Russian).
Berry, A. J., 1972. The natural history of west Malaysian mangrove faunas. MalayaNature Journal, 25, 135-162.
Bhan, S. & Mansuri, A. P., 1978a. Adaptations to osmotic stress in the marineeuryhaline teleost Periophthalmus dipes: Tissue water & mineral content. IndianJournal of Marine Science, 7, 134—136.
Bhan, S. & Mansuri, A. P., 1978b. Adaptations to osmotic stress in the marineeuryhaline teleost Periophthalmus dipes Bleeker: II. Changes in glycogen and fatlevels of tissues. Geobios, 5, (1), 3—8.
Bhan, S. & Mansuri, A. P., 1978c. Adaptations to osmotic stress in the marine euryhaline teleost Periophthalmus dipes: III Changes in tissue succinic dehydrogenaseenzyme levels. Journal of Animal Morphology and Physiology, 25, 132-138.
Birdsong, R. S., Murdy, E. O. & Pezold, F. L., 1988. A study of the vertebral columnand median fin osteology in gobioid fishes with comments on gobioid relationships.Bulletin of Marine Science, 42, 174—215.
Branch, G. M. & Grindley, J. R., 1979. Ecology of southern African estuaries. Part XIMngazana: a mangrove estuary in Transkei. South African Journal ofZoology, 14,3, 150-170.
Bridges, C. R., 1988. Respiratory adaptations in intertidal fish. American Zoologist, 28,79-96.
Brillet, C, 1969a. Etude du comportement constructeur des poissons amphibiesPeriophtalmidae. Terre et Vie, 23, 496—520.
Brillet, C, 1969b. Premiere description de la parade nuptiale des poissons amphibiesPeriopthalmidae. Comptes Rendus Hebdomadaires des Seancesde VAcademie desSciences, Serie D, 269, 1889-1892.
Brillet, C, 1969c. Observations sur le comportement agressif des poissons amphibiesperiophtalmes {Periophtalmus koelreuteri Pallas). Comptes Rendus Hebdomadairesdes Seances UAcademie des Sciences, Serie D, 269, 1114—1117.
Brillet, C, 1969d. Comportement territorial et hierarchie sociale chez les poissonsamphibies du genre Periophtalmus (Teleosteen, Gboioidae). Comptes RendusHebdomadaires des Seances de VAcademie des Sciences, Serie D, 269, 1322-1325.
Brillet, C, 1970. Relations entre comportement sexual, territoire et agressivitie chez lesperiophtalmes. Comptes Rendus Hebdomadaires des Seances de VAcademie desSciences, Serie D, 269, 1507-1510.

564 DAVID A. CLAYTON
Brillet, C, 1975. Relations entre territoire et comportement agressif chez Periophthalmussobrinus Eggert (Pisces, Periophthalmidae) au laboratoire et en milieu naturel.Zeitschrift fur Tierpsychologie, 39, 283-331.
Brillet, C, 1976. Structure du terrier, reproduction et comportement des jeunes chez lepoisson amphibie Periophthalmus sobrinus Eggert. Terreet la Vie, 30, 465-483.
Brillet, C, 1980a. Comportement sexuel du poisson amphibie Periophthalmus sobrinusEggert. Ses rapports avec le comportement agonistique. Review Ecologie {Terre etVie), 34, 427-468.
Brillet, C, 1980b. Comportement agonistique du poisson amphibie Periophthalmussobrinus Eggert. Analyse quantitative-1. Frequence des divers elements desrepertoire. Biology of Behavior, 5, 297-315.
Brillet, C, 1981. Comportement agonistique du poisson amphibie Periophthalmussobrinus Eggert. Analyse quantitative-II. Influence de la taile et du sexe surl'establissement des relations de dominance/subordination et sur l'utilisation durepertoire agonistique. Biology of Behavior, 6, 35-57.
Brillet, C, 1983. Agonistic behaviour in the amphibious fish Periophthalmus sobrinusEggert. Biology of Behavior, 8, 49-66.
Brillet, C, 1984. Etude comparativede la parade nuptialechez deux especes de poissonsamphibies sympatriques (Pisces, Periophthalmidae). Comptes Rendus Hebdomadairesdes Seances de VAcademie des Sciences, Serie D, 298, 347-350.
Brillet, C, 1986. Notes on the behaviour of the amphibious fish Lophalticus kirkiiGunther (Pisces Salariidae). Comparison with Periophthalmus sobrinus Eggert.Review Ecologie (Terre et Vie), 41, 361-375.
Burhanuddin & Martosewojo, S. 1979. [Observations on the natural history of ikangelodok Periophthalmus koelreuteri in Par Island]. In, Proceedingsof theSeminaron Mangrove Forest Ecosystems, Jakarta, edited by S. Soemodihardjo et al,Indonesia, pp. 86-92. (In Malay with English abstract).
Champeau, M. F., 1951. Des moeurs de Periophthalmus koelreuteri gobiide de l'OuestAfrica. Deuxieme Conference International des Africanists de VOuest, Bissau,1947, 3, 271-277.
Chapman, V. J., 1976. Mangrove Vegetation. J. Cramer, Berlin, 448 pp.Chatterjee, T. K. & Siddiqi S. Z., 1957. Periophthalmus weberi Eggert (Pisces:
Gobiidae), a rare gobioid fish from Indian waters. Matsya, 1, 82 only.Chen, T. P., 1976. Culture of the mudskipper. In, Aquaculture Practices in Taiwan.
Fishery News (Books) Ltd, Farnham, England, 62 pp.Chen, T. P., 1982. Mudskipper culture. Thai Fish Gazette, 35, 321-324.Chen, Y.-D., & Ting, Y.-Y., 1984. [The preliminary report on inducing breeding
experiments with mudskipper, Boleophthalmus pectinirostris (L.) by injections ofHCG.] Bulletin ofthe Taiwan Fisheries Research Institute, No. 37, 133-144. (InChinese with English abstract).
Chew, S. F. & Ip, Y. K., 1987. Ammoniagenesis in mudskippers Boleophthalmusboddarti and Periophthalmus schlosseri. Comparative Biochemistry and Physiology, 87B, 941-948.
Chew, S. F. & Ip, Y. K., 1990. Differences in the responses of two mudskippers,Boleophthalmus boddaerti and Periophthalmus chrysospilos to changesin salinity.Journal of Experimental Zoology, 256, 227—231.
Chew, S. F., Lim, A. L. L., Low, W. P., Lee, C. G. L., Chan, K. M. & Ip, Y. K.,1990. Can the mudskipper Periophthalmus chrysospilos tolerate acute environmental hypoxic exposure? Fish Physiology and Biochemistry, 8, 221-227.
Choudhury, A. & Nandi, N. C, 1973. Studies on myxosporidian parasites (Protozoa)froman estuarine gobiidfish of westBengal. Proceedings of the Zoological Societyof Calcutta, 26, 45-55.
Clayton, D. A., 1986. Ecology of mudflats with particular reference to those of thenorthernArabianGulf. In, Marine Environment andPollution: Proceedings of theConference on the Marine Environment and Pollution, edited by R. Halwagy

MUDSKIPPERS 565
et al, Kuwait University, Kuwait Foundation for Advancement of Science andEnvironmental Protection Council, Kuwait, pp. 83-96.
Clayton, D. A., 1987. Why mudskippers build walls. Behaviour, 102, 185-195.Clayton, D. A., 1990. Crustacean allometric growth: A case for caution. Crustaceana,
58, 270-290.Clayton, D. A. & Abo Seedo, F. 1986. Osteichthyes. In, D. A. Jones, A Field Guide
to the Sea Shores ofKuwait and the Arabian Gulf. University of Kuwait, BlandfordPress, Kuwait, pp. 167-175.
Clayton, D. A. & Vaughan, T. C, 1982. Pentagonal territories of the mudskipperBoleophthalmus boddarti (Pisces: Gobiidae). Copeia, 1982, 233-235.
Clayton, D. A. & Vaughan, T. C, 1986. Territorial acquisition in the mudskipperBoleophthalmus boddarti (Pisces: Gobiidae) on the mudflats of Kuwait. Journalof Zoology, London, 209, 501-519.
Clayton, D. A. & Vaughan, T. C, 1988. Ethogram of Boleophthalmusboddarti (Teleostei:Gobiidae), a mudskipper found on the mudflats of Kuwait. Journal ofthe University of Kuwait, (Science), 15, 115-138.
Clayton, D. A. & Wright, J. M., 1989. Mud-walled territories and feeding behaviourof Boleophthalmus boddarti (Pisces: Gobiidae) on the mudflats of Kuwait. Journalof Ethology, 7, 91-95.
Cuvier, G. L. G. C. D. & Valenciennes, A., 1837. Histoire Naturelle des Poissons,Vol 12, F. G. Levrault, Paris, 509 pp.
Dall, W. & Milward, E., 1969. Water intake, gut absorption and sodium fluxes in amphibious and aquatic fishes. Comparative Biochemistry and Physiology, 30, 247—260.
Das, B. K., 1933. On the bionomics, structure and physiology of respiration in anestuarine air-breathing fish, Pseudapocryptes lanceolatus (Bloch and Schneider),with special reference to a new mode of aerial respiration. Current Science, 1,389-393.
Das, B. K., 1934. The habits and structure of Pseudapocryptes lanceolatus, a fish in thefirst stages of structural adaption to aerial respiration. Proceedings of the RoyalSociety, Series B, 115, 422-430.
Datta, N. C. & Das. A., 1980. Anatomy of the olfactory apparatus of some Indiangobioids (Pisces: Perciformes). Zoologische Anzeiger, 205, 241-252.
Day, J. H., 1974. The ecology of Morrumbene Estuary Mocambique. Transactions ofthe Royal Society of South Africa, 41, 43-97.
De, J. K. & Nandi, N. C, 1984. A note on the locomotory behaviour of the mudskipperBoleophthalmus boddarti. Indian Journal of Fisheries, 31, 407—409.
Dhage, K. P. & Mohamed, O. M. M. J., 1977. Study of the amylase activity inmudskipper Periophthalmus koelreuteri (Pallas). Indian Journal of Fisheries, 24,129-134.
Diamond, J., 1971. The Mauthner cell. In, Fish Physiology, Vol. 5, edited by W. S. Hoar& D. J. Randall, Academic Press, London, pp. 265—346.
Diesselhorst, G., 1938. Horversuche an Fishen ohne Weberschen Apparat. ZeitschriftfurVergleichende Physiologie, 25, 748—783.
Dorjes, J., 1978. Sedimentologische und faunistiche Untersuchungen an Watten inTaiwan. II. Faunistiche und aktuopalaontologische Studien. SenckenbergianaMaritima, 10, 117-143.
Dotsu, Y., 1961. The bionomics and life history of the gobioid fish. Apocryptodon bleekeri(Day). Bulletin of the Faculty of Fisheries, Nagasaki University, 10, 127-131.
Eggert, B., 1929a. Bestimmungstabelle und Beschreibung des Arten der FamilyPeriophthalmus. Zeitschrift fur Wissenschaftliche Zoologie, 133, 398—409.
Eggert, B., 1929b. Die Gobiidenflosse und ihre Anpassung an das Landleben. Zeitschriftfur Wissenschaftliche Zoologie, 133, 411—440.
Eggert, B. 1935. Beitrag zur Systematik, Biologie und geographischen Verbreitung derPeriophthalminae. Zoologische Jahrbucher, Abteilung fur Systematik, Okologieund Geographie, 67, 29—114.

566 DAVID A. CLAYTON
Eissa, S. M. El-Zaidy, S. & Al-Naqi, Z., 1978. Circadian activity rhythm of mudskipper fish, Periophthalmus chrysospilos (Bleeker) in Kuwait. Bulletin of theFaculty of Science, Cairo University, 48, 213-225.
El-Sayed, N. K. & Safer, A. M., 1985. Enzyme activity in the kidney of the mudskipperPeriophthalmus koelreuteri (Pallas). Journalof the University ofKuwait, (Science),12, 245-256.
El-Sayed, N. K. & Safer, A. M., 1986. Wandering cells in the nephron of the mudskipper Periophthalmus koelreuteri (Pallas). Cell Biology International Reports,10, 755-762.
El-Zaidy, S., Eissa, S. M., & Al-Naqi, Z., 1975. Ecological studies on the mud-skipperfish Periophthalmus chrysospilos (Bleeker) in Kuwait. Bulletin of the Faculty ofScience, Cairo University, 48, 113—134.
Fenwick, J. C. & Lam, T. J., 1988a. Effects of calcitonin on plasma calcium andphosphate in the mudskipper, Periophthalmodon schlosseri (Teleostei), in waterand during exposure to air. General and Comparative Endocrinology, 70, 224-230.
Fenwick, J. C. & Lam, T. J., 1988b. Calcium fluxes in the teleost fish Tilapia(Oreochromis) in water and in both water and air in the marble goby {Oxyeleotris)and the mudskipper (Periophthalmodon). Physiological Zoology, 61, 119-125.
Fischthal, J. H. & Thomas, J. D., 1971. Some hemiurid trematodes of marine fishesfrom Ghana. Proceedings of the Helminthological Society of Washington, 38,181-190.
Fishelson, L., 1971. Ecology and distribution of the benthic fauna in the shallow watersof the Red Sea. Marine Biology, 10, 113-133.
Franz, V., 1910. Die japanischen Knockenfische der Sammlungen Harberer und Doflein.Abhandlungen Bayerische Akademie der Wissenschaften, Mathematisch-Natur-wissenschaftliche Klasse, Supplement, 4, 1—32.
Freitas, A. J., 1961a. Alqumas notas sobre a reproducao do Periophthalmus sobrinus.Boletim do Instituto de Investigacao Cientica de Mocambique, 2, 73—76.
Freitas, A. J., 1961b. Uma nota sobre a estructura dos ninhos do peixe Periophthalmussobrinus. Boletimdo Instituto de Investigacao Cienticade Mocambique, 2, 389—390.
Fricke, H. W., 1970. Die okologische Spezialisierung der Eidechse Crytoblepharusboutoni cognatus (Boettger) auf das Leben in der Gezeitenzone (Reptilia,Skinkidae). Oecologia (Berlin), 5, 380-391.
Frith, D. W., 1977. A preliminary list of macrofauna from a mangrove forest andadjacent biotopes at Surin Island, Western Peninsular Thailand. Phuket MarineBiological Centre, Research Bulletin, 17, 1-14.
Frith, D. W., Tantanasiriwong, R. & Bhatia, O., 1976. Zonation and abundance ofmacrofauna on a mangrove shore, Phuket Island. Phuket Marine BiologicalCentre, Research Bulletin, 10, 1-37.
Frith, D. W. & Brunenmeister, S., 1980. Ecological and population studies of fiddlercrabs (Ocypodidae, Genus Uca) on a mangrove shore at Phuket Island, Westernpeninsular Thailand. Crustaceana, 39, 157-183.
Fukuda, T., 1985. A fish out of water. Discover, 6, 56-61.Garey, W. F., 1962. Cardiac responses of fishes in asphyxic environments. Biological
Bulletin (Woods Hole, Massachusetts), 122, 362—368.Gibson, R. N., 1969. The biology and behaviour of littoral fish. Oceanography and
Marine Biology: an Annual Review, 7, 367—410.Gibson, R. N., 1982. Recent studies on the biology of intertidal fishes. Oceanography
and Marine Biology: an Annual Review, 20, 363-414.Gibson, R. N., 1986. Intertidal teleosts: life in a fluctuating environment. In, The
Behaviour of Teleost Fishes, edited by T. J. Pitcher, Croom Helm, London, pp.388-408.
Gibson-Hill, C. A., 1948. A note on the food habits of three kingfishers occurring onSingapore Island. Journal of the Bombay Natural History Society, 48, 146-152.

MUDSKIPPERS 567
Gopalakrishnan, P., 1970. Some observations on the shore ecology of the Okha coast.Journal of the Marine Biological Association of India, 12, 15-34.
Gordon, M. S., 1970. Patterns of nitrogen excretion in amphibious fishes. In, Urea andKidney, edited by K. Schmidt-Nielson, Excerpta Medica, Amsterdam, pp. 238-242.
Gordon, M. S., Boetius, J., Boetius, I., Evans, D. H., McCarthy, R. & Oglesby, L.,1965. Salinity adaptation in the mudskipper fish Periophthalmus sobrinus.Hvalradets Skrifter, 48, 85-93.
Gordon, M. S., Boetius, J., Evans, D. H. & Oglesby, L. C, 1968. Additional observations on the natural history of the mudskipper, Periophthalmus sobrinus. Copeia,1968, 853-857.
Gordon, M. S., Boetius, I., Evans, D. H., McCarthy, R. & Oglesby, L. C, 1969.Aspects of the physiology of terrestrial life in amphibious fishes. I. The mudskipper, Periophthalmus sobrinus. Journal ofExperimental Biology, 50, 141—149.
Gordon, M. S., Gabaldon, D. J. & Yip, A. Y.-W., 1985. Exploratory observations onmicrohabitat selection within the intertidal zone by the Chinese mudskipper fishPeriophthalmus cantonensis. Marine Biology, 85, 209—215.
Gordon, M. S., Ng, W. W.-S. & Yip, A. Y.-W., 1978. Aspects of the physiology ofterrestrial life in amphibious fishes. III. The Chinese mudskipper Periophthalmuscantonensis. Journal of Experimental Biology, 72, 57—75.
Graham, J. B., 1971. Aerial vision in amphibious fishes. Fauna (Rancho Mirage,California), 3, 14-23.
Graham, J. B., 1976. Respiratory adaptations of marine air-breathing fishes. In Respirationof Amphibious Vertebrates, edited by G. M. Hughes, Academic Press, London,pp. 165-187.
Gregory, R. B., 1977. Synthesis and total excretion of waste nitrogen by fish of thePeriophthalmus (mudskipper) and Scartelaos families. Comparative Biochemistryand Physiology, 57A, 33-36.
Gundermann, N. & Popper, D. M., 1984. Notes on the Indo-Pacific mangal fishes andon mangrove related fisheries. In, Hydrobiology of the Mangal, edited by F. D.Por & I. Dor, W. Junk, The Hague, pp. 201-206.
Haddon, A. C, 1889. Zoological notes from the Torres Straits: Caudal respiration inPeriophthalmus. Nature, 39, 285 only.
Harms, J. W., 1929. Die Realization von Genen und die consecutive Adaptation. 1:Phasen in der Differenzierung der Anlagenkomplexe und die Frage der Landtier-werdung. Zeitschrift fur wissenschaftliche Zoologie, 133, 211-397.
Harms, J. W., 1935. Die Realisation von Genen und die consecutive Adaptation. 4:Mitteilung Experimentell hervorgerufener Medienwechsel: Wasser zu Feuchtluftbwz. zu Trockenlaft bei Gobiiformes (Gobius, Boleophthalmus und Periophthalmus). Zeitschrift fur wissenschaftliche Zoologie, 146, 417-462.
Harms, W., 1914. Uber die Augen der am Grunde der Gewasser lebenden Fische.Zoologische Anzeiger, 44, 35-41.
Harris, V. A., 1961. On the locomotion of the mudskipper Periophthalmus koelreuteri(Pallas) (Gobiidae). Proceedings of the Zoological Society of London, 134,107-135.
Herre, A. W. C. T., 1941. A list of fishes from the Andaman Islands. Memoirs of theIndian Museum, 13, 331—403.
Hess, C, 1912. Untersuchungen zur vergleichenden Physiologie und Morphologie desCiliarringes. Zoologische Jahrbiicher Supplement, 15, 155—175.
Hess, C, 1913. Gesichtssin. In, Winterstein's Handbuch der Vergleichenden Physiologie, 4, 555-840.
Higson, S. J., 1889. A Naturalist in North Celebes. Murray, London.Hillman, S. S. & Withers P. C, 1987. Oxygen consumption during aerial activity in
aquatic and amphibious fish. Copeia, 1987, 232—234.Hoda, S. M. S., 1980. A contribution to the gobioid fishes of Pakistan. Proceedings of
the First Pakistan Congress of Zoology B. 469-482.

568 DAVID A. CLAYTON
Hoda, S. M. S., 1985. Fishes of the coast of Pakistan. Pakistan Agriculture, 7, 38—44.Hoda, S. M. S., 1986a. Maturation and fecundity of the mudskipper Boleophthalmus
dussumieri Cuv. & Val. from the Karachi Coast. Mahasagar—Bulletin of theNational Institute of Oceanography, 19, 73-78.
Hoda, S. M. S., 1986b. On the growth of Boleophthalmus dussumieri C.V. and B.dentatus C.V. (Family Gobiidae) from the Karachi coast of Pakistan. Biologia(Lahore), 32, (Suppl.), 55-62.
Hoda, S. M. S., 1987. Relative growths of body parts and length-weight relationshipsin Boleophthalmus dussumieri and B. dentatus of Karachi Coast. Indian Journalof Fisheries, 34, 120-127.
Hoda, S. M. S. & Akhtar, Y., 1983. Mudskippers from mangrove swamps of Karachicoast. In, Mangroves of Pakistan. Proceedings of the National Workshop onMangroves held at Karachi 8-10 August, 1983. Pakistan Agricultural ResearchCouncil, Islamabad, pp. 49-52.
Hoda, S. M. S. Akhtar, Y., 1985. Maturation and fecundity of mudskipper Boleophthalmusdentatus in the northern Arabian Sea. Indian Journal of Fisheries, 32, 64—74.
Holly, M., 1929. Beitrage zur Kenntnis der Fischfauna Persiens. Zoologische Anzeiger,85, 183-185.
Hong, W.-S., Dai, Q.-G., Zhang, Q.-Y., Zang, J. & Cai, Y.-Y., 1988. [Observationson the early development of the mudskipper Boleophthalmus pectinirostris(Linnaeus).] Tropic Oceanology/Redai Haiyang, 5, (2), 1—8. (In Chinese withEnglish abstract).
Hong, W.-S. & Wang, Y.-L., 1989. [Introduction of ovulation in female mudskipper(Boleophthalmus pectinirostris) by HCG analogue ofLHRH & pimozide. ] ChinaJournal of Oceanology & Limnology, 7, 283—287. (In Chinese with Englishabstract).
Hora, S. L., 1933. Respiration in fishes. Journal ofthe Bombay Natural History Society,36, 538-560.
Hora, S. L., 1935a. Ecology and bionomics of the gobioid fishes of the Gangetic delta.International Congress of Zoology, 12, 841—865.
Hora, S. L., 1935b. Physiology, bionomics and evolution of the air breathing fishes ofIndia. Transactions of the National Institute of Science, India, 1, 1-16.
Hughes, G. M. & Al-Kadhomiy, N. K., 1986. Gill morphometry of the mudskipperBoleophthalmus boddarti. Journal of the Marine Biological Association of theUnited Kingdom, 66, 671-682.
Hughes, G. M. & Munshi, J. D., 1979. Fine structure of the gills of some Indianair-breathing fishes. Journal of Morphology, 160, 169—194.
Hunziker, R. E., 1985. Gobies for freshwater and brackish aquaria. Tropical FishHobbyist, 34, 10-18.
Inger, R. F., 1952. Walking fishes of south eastern Asia travel on land. Chicago NaturalHistory Museum Bulletin, 23, 4—7.
Ip, Y. K., Chew, S. F. & Low, W. P., 1991a. Effects of hypoxia on the mudskipper,Periophthalmus chrysospilos (Bleeker, 1853). Journal ofFish Biology, 38, 621—623.
Ip, Y. K., Chew, S. F. & Ping, C. T., 1991b. Evaporation and the turning behaviourof the mudskipper Boleophthalmus boddaerti. Zoological Science (Tokyo), 8,621-623.
Ip, Y. K. & Low, W. P., 1990. Lactate production in the gills of the mudskipperPeriophthalmodon schlosseri exposed to hypoxia. Journal of ExperimentalZoology, 253, 99-101.
Ip, Y. K., Low, W. P., Lim, A. L. L. & Chew, S. F., 1990. Changes in lactate contentin the gills of the mudskippers Periophthalmus chrysospilos and Boleophthalmusboddaerti in response to environmental hypoxia. Journal of Fish Biology, 36,481-488.
Ishibashi, T., 1972. [Periodical behaviour of mudskippers.] Zoological Magazine(Tokyo), 81 409-410. (In Japanese).

MUDSKIPPERS 569
Ishibashi, T. & Nishikawa, M., 1973. [Behavioural rhythms of gobioid fish, Boleophthalmus chinensis (Osbeck) with special reference on the effect of light condition].Zoological Magazine (Tokyo), 82, 311 only. (In Japanese).
Iwata, K., 1984. A high ammonia tolerance in the mudskipper Periophthalmus cantonensis. Zoological Science, 1, 877 only.
Iwata, K., 1988. Nitrogen metabolism in the mudskipper Periophthalmus cantonensis:changes in free amino acids and related compounds in various tissues underconditions of ammonia loading, with special reference to its high ammoniatolerance. Comparative Biochemistry and Physiology, 91A, 499—508.
Iwata, K. & Kakuta, I., 1983. A comparison of catalytic properties of glutamatedehydrogenase from liver and muscle between amphibious Periophthalmus cantonensis and water-breathing gobiid fishes Tridentiger osbcurus obscurus. Bulletinof the Japanese Society of Scientific Fisheries, 49, 1903—1908.
Iwata, K., Kakuta, I., Ikeda, M., Kimoto, S. & Wada, N., 1981. Nitrogen metabolismin the mudskipper Periophthalmus cantonensis: a role of free amino acids indetoxification of ammonia produced during its terrestrial life. Comparative Biochemistry and Physiology, 86A, 589—596.
Jes, H., 1972. Eine Uferanlage fur Schlammspringer und Winkerkrabben im KolnerAquarium am Zoo. Zeitschriftefur Kolner Zoo, 15, 111—117.
Johnstone, J., 1903. Report on the marine fishes. Fasciculi Malayenses: Zoology, 12,293-303.
Jones, D. A., 1984. Crabs of the mangal ecosystem. In, Hydrobiology of the Mangal,edited by F. D. Por & I. Por, W. Junk, The Hague, pp. 207-209.
Jones, S., 1937. Observations on the breeding habits and development of certain brackishwater fishes of Adyar, Madras. Proceedings of the Indian Academy ofScience, 5,261-289.
Kaden, J., 1978. Schlammspringer sind ganz anders. Aquarien-Magazin, 12, 552—555.Karsten, H., 1923. Das Auge von Periophthalmus koelreuteri. Jenaische Zeitschriftfur
Naturwissenschaft, 59, 115-154.Khalaf, K. T., 1961. The Marine and Freshwater Fishes of Iraq. Ar Rabitta Press,
Baghdad, 143 pp.Khan, M. M. & Ip, Y. K., 1988. Pallisentis singaporensis (Acanthocephala: Quad-
rigyridea) from the mudskipper Periophthalmodon schlosseri in Singapore.Journal of Singapore National Academy of Sciences, 17, 24—27.
Khoo, K. G., 1966. Studies on the biology of periophthalmid fishes in Singapore. BScthesis, University of Singapore, 61 pp.
Kimura, M., 1958. Early development of the mudskipper Periophthalmus cantonensis(Osbeck). Bulletin of the Japanese Society of Scientific Fisheries, 23, 754-757.
Klausewitz, W., 1967. Uber einige Bewegungsweisen der Schlammspringer (Periophthalmus). Natur und Museum, 97, 6, 211—222.
Klausewitz, W., 1968. Some observations on East African mudskippers. Tropical FishHobbyist, 16, 52-65.
Klika, E. & Tesik, I., 1980. Die Ultrastruktur der Schleimhaut des akzessorischenLuftatmungsorganes des Schlammspringerfisches Periophthalmus barbarus. GiTFachzeitschrift fur das Laboratorium, 24, 189—196.
Kluge, K., 1971. Gologo. Aquarien Den Hag, 42, 108-109.Kobayashi, T., Dotsu, Y. & Takita, T., 1971. [Nest and nesting behaviour of the mud
skipper Periophthalmus cantonensis, in Ariake Sound. ] Bulletin of the FacultyofFisheries, Nagasaki University, 32, 27-39. (In Japanese with English abstract).
Kobayashi, T., Dotsu, Y. & Miura, N., 1972. [Egg development and rearing experiments of the larvae of the mudskipper Periophthalmuscantonensis. ] Contributionsfrom the Fisheries Experimental Station ofNakasagi University, No. 35, 49-62.(In Japanese with English abstract).
Krishnaja, A. P. & Rege, M. S., 1980. Some observations on the chromosomes ofcertain teleosts using a simple method. Indian Journal of ExperimentalBiology,18, 268-270.

570 DAVID A. CLAYTON
Krishnamurthy, K., Sultan Ali, M. A. & Prince Jeyaseelan, M. A., 1984. Structure anddynamics of the aquatic food web community with special reference to nematodesin mangrove ecosystems. In, Proceedings of the Asian Symposium on MangroveEnvironment-Research & Management, edited by E. Soepadmo et al, Universityof Malaya and UNESCO, Kuala Lumpur, pp. 429-452.
Lakshmi, R., Kundu, R. & Mansuri A. S. P., 1990. Toxicity of mercury to mudskipperBoleophthalmus dentatus C. & V. Part I. Changes in the activity of ATPases ingills. Acta Hydrochimica et Hydrobiologica, 18, 605—611.
Lakshmi, R., Kundu, R., Thomas, E. & Mansuri, A. S. P., 1991. Mercuric chlorideinduced inhibition of different ATPases in the intestine of Boleophthalmusdentatus. Ecotoxicology and Environment Safety, 21, 18—24.
Lee, C. G. L. & Ip, Y. K., 1987. Environmental effect on plasma thyroxine (T4),3,5,3'-triodo-L-thyronine (T3), prolactin and cyclic adenosine 3',5'-monophosphate(cAMP) content in the mudskippers Periophthalmuschrysospilos and Boleophthalmusboddarti. Comparative Biochemistry and Physiology, 87A, 1009—1014.
Lee, C. G. L., Low, W. P. & Ip, Y. K., 1987. Na+, K+ and volume regulation in themudskipper, Periophthalmus chrysospilos. Comparative Biochemistry andPhysiology, 87A, 439-448.
Lee, C.-L., 1990. [Osteological study of the mudhopper, Periophthalmus cantonensis(Perciformes, Gobiidae) from Korea]. Korean Journal ofZoology, 33, 402-410.(In Korean with English abstract).
Lele, S. H. & Kulkarni, R. D., 1938. The skeleton of Periophthalmus barbarus (Linn.).I. The skull. Journal of the University of Bombay, New Series, 6, 76—91.
Lele, S. H. & Kulkarni, R. D., 1939. The skeleton of Periophthalmus barbarus (Linn.).II. Branchial arches, vertebral column and appendicular skeleton. Journal of theUniversity of Bombay, New Series, 7, 123—134.
Liao, I.-C, Chao, N.-H., Tseng, L.-C. & Kuo, S.-C, 1973. [Studies on the artificialpropagation of Boleophthalmus chinensis (Osbeck)—I: Observations on embryonicdevelopment and early larvae.] J.CR.R. Fish Series, No. 15, 29-42.
Lim, A. L. L. & Ip, Y. K., 1989., Effect of fasting on glycogen metabolism and activitiesof glycolytic and gluconeogenic enzymes in the mudskipper Boleophthalmusboddaerti. Journal of Fish Biology, 34, 349—367.
Lim, G. S., 1967. Studies on the feeding biology of Periophthalmus barbarus (Linneus)and Boleophthalmus boddaerti (Pallas) Perciformes—Periophthalmidae. BScthesis, University of Malaya, 37 pp.
Lim, L. S., 1971. The feeding biology of Periophthalmus vulgaris (Eggert) and Periophthalmodon schlosseri (Pallas) Perciformes, Periophthalmidae. MSc thesis,University of Malaya, 33 pp.
Lipschitz, H. D., Berjak, P., Pammenter, N. W., & Davey, J. E., 1975. A note on theapparent redistribution of crabs in Saco mangrove swamp Inhaca Island,Mocambique. South African Journal of Science, 71, 55-57.
Low, W. P., Ip, Y. K. & Lane, D. J. W., 1990. A comparative study of the gillmorphometry in the mudskippers Periophthalmus chrysospilos, Boleophthalmusboddaerti and Periophthalmodon schlosseri. Zoological Science (Tokyo), 7,29-38.
Low, W. P., Lane, D. J. W. & Ip, Y. K., 1988. A comparative study of terrestrialadaptations of the gills in three mudskippers—Periophthalmus chrysospilos,Boleophthalmus boddaerti, and Periophthalmodon schlosseri. Biological Bulletin(Woods Hole, Massachusetts), 175, 434-438.,
Macintosh, D. J., 1979. Predation of fiddler crabs (Uca spp.) in estuarine mangroves.Biotrop Special Publication No. 10, Mangrove and Estuarine Vegetation in SouthEast Asia, edited by P. B. L. Srivastava, Serdang, Malaysia, pp. 101-110.
Macintosh, D. J., 1982. Fisheries and aquaculture significance of mangrove swamps withspecial reference to the Indo-West Pacific region. In, Recent Advances inAquaculture, edited by J. F. Muir & R. Roberts, Croom Helm, London, pp. 3—36.

MUDSKIPPERS 571
MacNae, W., 1968a. Mudskippers. African Wildlife, 22, 240-248.MacNae, W., 1968b. A general account of the fauna and flora of mangrove swamps and
forests in the Indo-West-Pacific region. Advances in Marine Biology, 6, 73-270.MacNae, W., 1968c. Mangroves and their fauna. Australian Natural History, 7, 17-21.MacNae, W. & Kalk, M., 1962. The ecology of the mangrove swamps at Inhaca Island,
Mocambique. Journal of Animal Ecology, 50, 19-34.Maekawa, K., Fukuda, Y., Okada, Z. & Imamura, K., 1968. [Intraepithelial blood
capillaries in the air breathing organs of fish.] Bulletin of the Tokyo MedicalCollege, 26, 793-799. (In Japanese with English abstract).
Magnus, D. B. E., 1972. On the ecology of the reproduction behaviour of Periopthalmuskalolo Lesson at the east African coast. Proceedings ofthe International Congressof Zoology, 17, Theme 6, 1-4.
Mahajan, B. A. & Srinivassan, M., 1988. Mercury pollution in the estuarine regionaround Bombay. Environmental and Technological Letters, 9, 331—336.
Manickam, P. & Natarajan, G. M., 1985. Observations on the blood parameters of twoair breathing mudskippers of South India. Journal ofCurrentBioscience, 2, 19-22.
Manna, G. K. & Prasad, R., 1974. Chromosome analysis in three species of fishesbelonging to family Gobiidae. Cytologia, 39, 609—628.
Mansuri, A. P. & Bhan, S., 1978. Adaptations to the osmotic stress in the marineeuryhaline teleost, Periophthalmus dipes IV. Changes in total protein levels ofvarious tissues. Journal of Animal Morphology and Physiology, 25, 139—146.
Mansuri, A. P., Bhan, S. & Mathai, T., 1982. Salinity tolerance and survival of mudskippers, Boleophthalmus dentatus and Periophthalmus dipes of Saurashtra coast.Journal of Animal Morphology and Physiology, 29, 142—148.
Marlier, G., 1938. Considerations sur les organes accessoires servant a la respirationaerienne chez les teleosteens. Annales de la Societe Royale Zoologique deBelgique, 69, 163-185.
Matoba, M. & Dotsu, Y., 1977. Pre-spawning behaviour of the mudskipper Periophthalmus cantonensis in Ariake Sound. Bulletin of the Faculty of Fisheries,Nagasaki University, No. 43, 23-33.
McCulloch, A. R. & Ogilby, J. D., 1919. Some Australian fishes of the family Gobiidae.Records of the Australian Museum, 12, 193—291.
Mehta, R., Mehta, H. S. & Devi, K., 1986. Evolutionary modifications in the girdlesand fins of the gobiid fish Periophthalmus schlosseri (Pallas). Journal of theAndaman Science Association, 2, 47—50.
Mehta, R., Mehta, H. S. & Devi, K. 1987., Adaptational modifications of pelvic finsin some gobiid fishes. Journal of the Andaman Science Association, 3, 152—154.
Mehta, R., Mehta, H. S. & Devi, K., 1990. Structure and shape of mouth in somegobioid fishes. Environment and Ecology, 8, 668—671.
Miller, P. J., 1973. The osteology and adaptive features of Rhyacichthys aspo (Teleostei:Gobioidei) and the classification of gobioid fishes. Journal of Zoology, London,111, 397-434.
Milward, N. E., 1974. Studies on the taxonomy, ecology and physiology of Queenslandmudskippers. PhD thesis, University of Queensland, 276 pp.
Misra, S., Dutta, A. K., Dhar, T., Ghosh, A., Choudhury, A. & Dutta, J., 1983. Fattyacids of the mudskipper Boleophthalmusboddaerti. Journal ofthe Science ofFoodand Agriculture, 34, 1413-1418.
Miyazaki, M. & Nakamura, T., 1980. [On the shape of muscles of the tongue ofBoleophthalmus pectinirostris.] Journal Kurume Medical Association, 43, 807-811. (In Japanese with English abstract).
Mookerjee, H. K., Ganguly, D. N. & Mookherji, P. S., 1950. Study on the structuresof the brains of some Indian fishes in relation to their feeding habits. Proceedingsof the Zoological Society, Bengal, 3, 119-152.
Morii, H., 1979. Changes with time of ammonia and urea concentrations in the bloodand tissue of mudskipper fish, Periophthalmus cantonensis and Boleophthalmus

572 DAVID A. CLAYTON
pectinirostris kept in water and on land. Comparative Biochemistry andPhysiology, 64A, 235-243.
Morii, H. & Kasama, K., 1989. [Bacterial flora in the digestive tracts of mudskipperfishes Boleophthalmus pectinirostris and Periophthalmus cantonensis. ] NipponSuisan Gakkaishi, 55, 733-741. (In Japanese with English abstract).
Morii, H., Nishikata, K. & Tamura, O., 1978. Nitrogen excretion of mudskipper fishPeriophthalmus cantonensis and Boleophthalmus pectinirostris in water and onland. Comparative Biochemistry and Physiology, 60A, 189-193.
Morii, H., Nishikata, K. & Tamura, O., 1981. Ammonia and urea excretion frommudskipper fishes Periophthalmus cantonensis and Boleophthalmus pectinirostristransferred from land to water. Comparative Biochemistry and Physiology, 63A,23-28.
Morton, B. & Morton J., 1983. The Sea Shore Ecology of Hong Kong. Hong KongUniversity Press, Hong Kong, 350 pp.
Mukherjee, A. K., 1971a. Food-habits of water-birds of the Sundarban, 24-ParganasDistrict, West Bengal, India—II. Herons and bitterns. Journal of the BombayNatural History Society, 68, 37-64.
Mukherjee, A. K., 1971b. Food-habits of water-birds of the Sundarban, 24-ParganasDistrict, West Bengal, India—III. Egrets. Journal of the Bombay Natural HistorySociety, 68, 691-716.
Munk, O., 1970. On the occurrence and significance of horizontal band-shaped retinalareae in teleosts. Videnskabelige Meddelelserfra Dansk Naturhistorisk Foreningi Khobenhavn, 133, 85-120.
Munz, F. W., 1971. Vision: Visual pigments. In, Fish Physiology, Vol. 5, edited byW. S. Hoar & D. J. Randall, Academic Press, London, pp. 1-32.
Murdy, E. O., 1986. Mudskippers of Malaysia. The lords of the mudflat. Freshwaterand Marine Aquarium, November 1986, 20-23.
Murdy, E. O., 1989. A taxonomic revision and cladistic analysis of the oxudercinegobies (Gobiidae: Oxudercinae). Records of the Australian Museum, Supplement11, 1-93.
Mutsaddi, K. B. & Bai, D. V., 1969a. Food and feeding of Boleophthalmus dussumierei(Cuv. & Val.). Journal of the University of Bombay, 38, 42—55.
Mutsaddi, K. B. & Bai, D. V., 1969b. Some observations on habits and habitat ofBoleophthalmus dussumierei (Cuv. and Val.). Journal of the University ofBombay, 39, 33-41.
Mutsaddi, K. B. & Bai, D. V., 1970. Maturation and spawning of Boleophthalmusdussumierei. Journal of the University of Bombay, 39, 58-76.
Mutsaddi, K. B. & Bai, D. V., 1973. Gobies from the intertidal region of Bombay.Indian Journal of Fisheries, 20, 476-486.
Nandi, S. & Choudhury, A., 1983. Quantitative studies on the benthic macrofauna ofSagar Island, intertidal zone, Sunderbans, India. Mahasager—Bulletin of theNational Institute of Oceanography, 16, 409—414.
Natarajan, G. M. & Rajulu, G. S., 1983. Bimodal oxygen uptake in the amphibiousmud-skipper Periophthalmus koelreuteri (Pallas) (Gobiidae: Teleostei). JournalofAnimal Morphology and Physiology, 30, 177—180.
Nateewathana, A. & Tantichodok, P. 1984. Species composition, density and biomassof macrofauna of a mangrove. In, Proceedings of the Asian Symposium on theMangrove Environment-Research & Management, edited by E. Soepadmo et al,University of Malaya and UNESCO, Kuala Lumpur, pp. 258-285.
Nishikawa, M. & Ishibashi. T., 1973. [Behavioural rhythms of gobioid fish, Boleophthalmus chinensis (Osbeck) with special reference to the effect of light condition. ]Zoological Magazine (Tokyo), 82, 311 only. (In Japanese).
Nishikawa, M., & Ishibashi, T., 1975a. [Endogenous rhythm and time sense in the mudskipper Periophthalmus cantonensis (Osbeck)]. Zoological Magazine (Tokyo),84, 354 only. (In Japanese).

MUDSKIPPERS 573
Nishikawa, M. & Ishibashi, T., 1975b. [Entrainment of the activity rhythm by the cycleof feeding in the mud-skipper Periophthalmus cantonensis]. Zoological Magazine(Tokyo), 84, 184-189. (In Japanese with English abstract).
Nishikawa, S., Amaoka, K. & Nakanishi K., 1974. A comparative study of chromosomesof twelve species of gobioid fishes in Japan. Japanese Journal ofIchthyology, 21,61-71.
Niva B., Ojha, J. & Munshi, J. S. D., 1979. Bimodal oxygen uptake in relation to bodyweight of the amphibious mud-skipper Boleophthalmus boddaerti (Pall.). IndianJournal of Experimental Biology, 17, 752-756.
Niva, B., Ojha, J. & Munshi, J. S. D., 1981. Morphometries of the respiratory organsof an estuarine goby. Boleophthalmusboddaerti. Japanese Journal ofIchthyology,27, 316-326.
Nogusa, S., 1957. Chromosome studies in Pisces VII. A comparative study of thechromosomes in six species of the Gobiidae. Japanese Journal of Ichthyology, 6,141-146.
Nogusa, S., 1960. A comparative study of the chromosomes in fishes with particularconsiderations on taxonomy and evolution. Memoires of Hyogo University,Agriculture, 3, 1—62.
Nursall, J. R., 1974. Character displacement and fish behaviour, especially in coral reefcommunities. American Zoologist, 14, 1099-1118.
Nursall, J. R., 1981. Behaviour and habitat affecting the distribution of five species ofsympatric mudskippers in Queensland. Bulletin ofMarine Science, 31, 730-735.
Ogasawara, T., Ip, Y. K., Hasegawa, S., Hagiwara, Y. & Hirano, T., 1991. Changesin prolactin cell activity in the mudskipper Periophthalmus chrysospilos inresponse to hypotonic environment. Zoological Science (Tokyo), 8, 89-95.
Oliva, O. & Skorepa, V., 1970a. The eye muscles in Periophthalmus Bloch et Schneider,1801 and Chauliodus Bloch et Schneider, 1801 (Osteichthyes). Vestnik Cesko-slovenske Spolecnosti Zoologicke Acta Societatis Zoologicae Bohemoslovacae, 34,50-57.
Oliva, O. & Skorepa, V., 1970b. The eye muscles in Argyropelecus olfersi (Cuvier,1829) and Boleophthalmus Val., 1837. Vestnik Ceskoslovenske Spolecnosti Zoologicke Acta Societatis Zoologicae Bohemoslovacae, 34, 234-239.
Pankow, H. & Hug, M. F., 1979. Diatoms in the stomach content of Pseudapocryptesdentatus a mudskipper from the Shatt-al-Arab estuary (Iraq). WissenschaftlicheZeitschschrift der Wilhem-Pieck-Universitat Rostock. Mathematisch-Naturwissen-schaftliche Reihe, 6, 547-554.
Parker, T. J. & Haswell, W. A., 1962. A Textbook ofZoology. Macmillan, London, 952 pp.Patel, B., Bangera, V. S., Patel, S. & Balani, M. C, 1985. Heavy metals in the Bombay
harbour area. Marine Pollution Bulletin, 16, 22—28.Patel, B. & Chandry, J. S., 1988. Mercury in the biotic and abiotic matrices along
Bombay coast. Indian Journal of Marine Science, 17, 55-58.Patel, B. & Desai, K., 1976. Tidal rhythms and cyclic changes in the neuroendocrine
complex of Boleophthalmus dentatus Cuv. & Val. Annals of Zoology (Agra), 1269-95.
Patel, B., Mulay, C. D. & Ganguly, A. K., 1975. Radioecology of Bombay harbour—atidal estuary. Estuarine and Coastal Marine Science, 3, 13—42.
Pearse, A. S., 1929. Observations on certain littoral and terrestrial animals at Tortugas,Florida, with special reference to migrations from marine to terrestrial habitats.Papers from the Tortugas Laboratory ofthe Carnegie Institute of Washington, No.391. 205-223.
Pearse, A. S., 1932. Observations on the ecology of certain fishes and crustaceans alongthe bank of the Malta River at Port Canning. Records of the Indian Museum,Calcutta, 34, 289-298.
Pearse, A. S., 1933. The gobies at Paknam. Journal of the Siamese Natural HistorySociety, Supplement, 9, 173—178.

574 DAVID A. CLAYTON
Petit, M. G., 1921. Observations sur certains poissons des cotes de Madagascar pres-entant une adaptation a la locomotion terrestre. Bulletin du Museum Nationald'Histoire Naturelie Zoologique, 28, 216—220.
Petit, M. G., 1922. Les Periophthalmes, poissons fouisseurs. Bulletin du MuseumNational d'Histoire Naturelie Zoologique, 27, 404-408.
Petit, M. G., 1928. Novelle observations sur la biologie des Periophthalmes. BulletinduMuseum National d'Histoire Naturelle Zoologique, 34, 197-199.
Polunin, I., 1972. Who says fish can't climb trees? NationalGeographic Magazine, 141,84-92.
Por, F. D., 1974. The hard-bottom mangroves of Sinai. Israel Journal ofZoology, 23,211 only.
Por, F. D., Dor, I, & Amir, A., 1977. The mangal of Sinai: Limits of an ecosystem.Helgoldnder wissenschaftliche Meersuntersuchungen, 30, 295—314.
Pradhan, S. V., 1961. A study of blood of a few Indian fishes. I. Haemoglobin. Proceedings of the Indian Academy of Sciences, Section B, 54, 251-256.
Prince Jeyaseelan, M. A. & Krishnamurthy, K., 1980. Role of mangrove forest ofPichavaram as fish nursery. Proceedings ofthe Indian National Science Academy,Part B, 46, 548-553.
Qui, L.-Y., 1989. [Observation on two species of Periophthalmidae along Jiangsu coast].Oceanologia et Limnologia Sinica,20, 577-580. (In Chinese with English abstract).
Qureshi, M. R. & Bano, A., 1971. Common gobioid fishes of sub-families Perioph-thalminae and Apocrypteinae. Pakistan Journal of Science, 23, 143-146.
Rauther, M., 1910. Die akzessorischen Atmungsorgane der Knochenfische. Ergebnisseder Fortschritte der Zoologie, 2, 517—585.
Reichelt, J., 1910. Boleophthalmus pectinirostris L. und Periophthalmus koelreuteri(Pall.) Bl. Blatterfur Aquarien- und Terrarienkunde, 21, 309-311.
Rodewald, C, 1913. Boleophthalmus pectinirostris L. (?). Blatterfur Aquarien- undTerrarienkunde, 24, 565—566.
Ryu, B. & Lee, J. 1979. [The life form of Periophthalmus cantonensis in the Gum riverin summer.] Bulletin of the Korean Fisheries Society, 12, 71—78. (In Korean).
Safer, A. M. A. & El-Sayed, N. K., 1986. The fine structure of the nephronic tubuleof the mudskipper Periophthalmus koelreuteri (Pallas). Journal of Morphology,187, 109-121.
Safer, A. M. A., Tytler, P. & El-Sayed, N., 1982. The structure of the head kidney inthe mudskipper, Periophthalmus koelreuteri (Pallas). JournalofMorphology, 174,121-131.
Salih, M. S. & Al-Jaffery, A., 1980. Morphological study of Boleophthalmus dentatusCuvier & Valenciennes, 1837 with reference to its feeding habits. I. Habitat,external features and integument. Bulletin of the Biological Research Center,Baghdad., 12, 11-44.
Sarker, A. L., Al-Daham, N. K. & Bhatti, M. N., 1980. Food habits of the mudskipper,Pseudapocryptes dentatus (Val.). Journal of Fish Biology, 17, 635—639.
Sasekumar, A., 1974. Distribution of macrofauna in a Malayan mangrove shore. Journalof Animal Ecology, 43, 52—69.
Sasekumar, A., Ong, T. L. & Thong, K. L., 1984. Predation of mangrove fauna bymarine fishes. In, Proceedings of the Asian Symposium on the MangroveEnvironment-Research & Management, edited by E. Soepadmo et al, Universityof Malaya and UNESCO, Kuala Lumpur, pp. 378-384.
Schneider, H., 1990a. Schlammspringer in der Mangrove. Aquarium (Minden), 249, 19—23.Schneider, H., 1990b. Das Naturerlebnis: Schlammspringer in der Mangroven. Aquaria
(St. Gallen), 35, 287-295.Schottle, E., 1932. Morphologie und Physiologie der Atmung bei wasser-, schlamm- und
landlebenen Gobiiformes. Zeitschriftfur Wissenschaftliche Zoologie, 140, 1-114.Schreitmuller, W., 1911. Neues uber Boleophthalmus pectinirostris Linne. Blatter fur
Aquarien- und Terrarienkunde, 22, 29—31.

MUDSKIPPERS 575
Schreitmuller, W., 1914. Boleophthalmus pectinirostris Linne und Boleophthalmusboddaerti Pallas. Blatter fur Aquarien- und Terrarienkunde, 25, 41-43.
Shaikh, Y. A. & Hiradhar, P. K., 1987. Histological observations on changes inducedin some tissues of the edible mudskipper Boleophthalmus dussumieri exposed tosublethal concentrations of sodium fluoride. Journal of Animal Morphology andPhysiology, 34, 69-75.
Shaikh, Y. A. & Hiradhar, P. K., 1988. Alterations in the acid and alkaline phosphatasequantitiesin fluoride-exposed estuarine goby Boleophthalmus dussumieri. Fluoride,21, 131-136.
Siau, H. & Ip, Y. K., 1987. Activities of enzymes associated with phosphoenolypyruvatemetabolism in the mudskippers, Boleophthalmus boddaerti and Periophthalmusschlosseri. Comparative Biochemistry and Physiology, 88B, 119—125.
Siddiqi, S. A., 1974. Natural distribution and ecology of dark blotched mudskipper,Periophthalmus waltoni Koumans (Pisces: Gobiidae). Journal of the MarineBiological Association of India, 16, 843-844.
Simpson, D. A., 1955. Fish out of water. Aquarium Journal, 26, 181-183.Singh, B. N. & Munshi, J. S. D., 1969. On the respiratory organs and mechanics of
breathing in Periophthalmus vulgaris (Eggert). Zoologische Anzeiger, 183, 92-110.Singh, B. R., Mishra, A. P., Prasad, M. S. & Singh, I., 1989. Development of neo-
morphic air-breathing organs in three genera of estuarine gobies. GegenbaursMorphologische Jahrbuch, 135, 529-540.
Singh, B. R., Yadav, A. N., Prasad, M. S., Mishra, A. P. & Singh, I., 1990. Neo-morphic organ for C02 elimination in air-breathing teleosts. European Archivesof Biology, 101, 257-268.
Sircar, A. K. & Har, S. P., 1975. Ecology of an estuarine breeding ground of somegobioid fishes in west Bengal, India. In, Special Symposium on Marine Sciences,edited by B. Morton & J. R. Lee. Pacific Science Association, GovernmentPrinter, Hong Kong, pp. 63-64.
Slooff, R. & Marks, E. N., 1965. Mosquitoes (Culicidae) biting a fish (Periophthalmidae).Journal of Medical Entomology, 2, 16 only.
Soni, V. C, & George, B., 1986. Age determination and length weight relationship inthe mudskipper Boleophthalmus dentatus. Indian Journal ofFisheries, 33, 321—234.
Sowerby, A. De C, 1923. The mud skipper. China Journal of Science and Arts, 1,343-345.
Sponder, D. L. & Lauder, G. V., 1981. Terrestrial feeding in the mudskipper Periophthalmus (Pisces: Teleostei) : A cineradiographic analysis. Journal of Zoology,London, 193, 517-530.
Stebbins, R. C. & Kalk, M., 1961. Observations on the natural history of the mudskipper Periophthalmus sobrinus. Copeia, 1961, 18—27.
Subrahmanyam, K., 1969. A karyotypic study of the estuarine fish Boleophthalmusboddaerti (Pallas) with calcium treatment. Current Science, 38, 437-439.
Tamura, S. O., Morii, H. & Yuzuriha, M., 1976. Respiration of the amphibious fishesPeriophthalmus cantonensis and Boleophthalmus chinensis in water and on land.Journal of Experimental Biology, 65, 97-107.
Tamura, O. & Moriyama, T., 1976. On the morphological feature of the gill of amphibious and air breathing fishes. Bulletin of the Faculty of Fisheries, NagasakiUniversity, 41, 1—8.
Teal, J. M. & Carey, F. G., 1967. Skin respiration and oxygendebt in the mudskipper,Periophthalmus sobrinus. Copeia, 1967, 677-679.
Troncy, P.-M. & Vassiliades, G., 1974. Acanthocephales parasites de poissonsd'Afrique. Bulletin de VInstitut Fondamental d'Afrique Noire, Serie A: SciencesNaturelles, 36, 902-910.
Tytler, P. & Vaughan, T., 1983. Thermal ecology of the mudskipper, Periophthalmuskoelreuteri (Pallas) and Boleophthalmus boddarti (Pallas) of KuwaitBay. Journalof Fish Biology, 23, 327-337.

576 DAVID A. CLAYTON
Uchida, Y., Enomoto, N. & Miyaguchi, M., 1971. [On the heavy metal contents inmarine products from Ariake Sea.] Agricultural Bulletin of Saga University, 32,45—49. (In Japanese with English abstract).
Van Dijk, D. E., 1959. A fish that walks, jumps and burrows. Nucleon, 1, 36—38.Van Dijk, D. E., 1960. Locomotion and attitudes of the mudskipper, Periophthalmus,
a semi-terrestrial fish. South African Science, 56, 158—162.Van Dijk, D. E., 1978. Transition from aquatic to semi-terrestrial life in vertebrates: the
gobioid mudskipperPeriophthalmus, as model. IsraelJournal ofMedical Science,14, 905-906.
Venkateswarlu, T., 1966. A note on the oxygen carrying capacity of the blood in threegobiid fishes. Naturwissenschaften, 12, 3 only.
Venkateswarlu, T., 1969. On Periophthalmus schlosseri Bl. Schn. from Vellar estuary,Portonovo, S. India. Science and Culture, 35, 492-493.
Verma, G. K., 1968. Studies on the structure and behaviour of chromosomes of certainfresh water and marine gobid fishes. National Academy of Science of India, 37,178 only.
Verwey, J., 1930. Einiges uber die Biologie Ost-Indischer Mangrovekrabben. Treubia,12, 167-261.
Vestergaard, K., 1972. The biology of some mangrove fish. In, Anonymous, The StudyTour to East Africa, 1970, University of Copenhagen, pp. 317-319.
Vivekanandan, E. & Pandian, T. J., 1979. Erythrocyte count and haemoglobin concentration of some tropical fishes. Journal ofMadurai Kamaraj University (Science),8, 71-75.
Volsoe, H., 1939. The sea snakes of the Iranian Gulf and the Gulf of Oman. Witha summary of the biology of the sea snakes. In, Danish Investigations in IranPart 1, edited by K. Jessen & R. Sparch, Einar Munksgaard, Copenhagen, pp.9-45.
Volz, W., 1905a. Uber das Auge von Periophthalmus und Boleophthalmus. Mitteilungender Naturforschenden Gesellschaft in Bern, 108—111.
Volz, W., 1905b. Zur Kenntnis des Auges von Periophthalmus und Boleophthalmus.Zoologische Jahrbiicher, Abteilung fur Anatomie und Ontogenie der Tiere, 22,331-346.
Volz, W., 1905c. Sur l'oeil de Periophthalmus et Boleophthalmus. Archives des SciencesPhysiques et Naturelles (Geneve), 4, 579—580.
Von Anthouard, M. & Mignot, A., 1973. Schlammspringer. Aquarien Terrarien, 25,330-333.
Warren, J. H., 1990. The use of open burrows to estimate abundances of intertidalestuarine crabs. Australian Journal of Ecology, 15, 277—280.
Welsch, U. & Storch, V., 1976. Elektronenmikroskopische Beobachtungen am Kiemen-epithel des amphibischen Teleosteers Periophthalmus vulgaris. Zeitschrift furMikroskopisch-Anatomische Forschung, 90, 447—457.
Whitley, G. P., 1931. New names for Australian fishes. Australian Zoologist, 6, 310—334.Whitley, G. P., 1953. Fishes collected by the Australian museum expedition, 1952.
Records of the Australian Museum, 23, 123—132.Whitley, G. P., 1960. The mud-skipper. The Education Gazette, 54, 142-143.Whitley, G. P., 1968. Mud-skippers. The Australian Museum Leaflet, 17, 1-6.Willem, V. & Boelaert, R., 1937. Les manoeuvres respiratoires de Periophthalmus.
Bulletin de L'Academie Royale des Sciences des Lettres et des Beaux-Arts deBelgique (Series 5), 23, 942-959.
Xie, X.-G. & Zhang, Q.-Y., 1990. [Studies on the gonadal development in femalemudskipper, Boleophthalmus pectinirostris]. Journal ofOceanography in TaiwanStraits, 9, 217-221. (In Chinese with English abstract).

MUDSKIPPERS 577
Yadav, A. N., Prasad, M. S. & Singh, B. R., 1990. Gross structure of the respiratoryorgans and dimensions of the gill in the mudskipper, Periophthalmodon schlosseri(Bleeker). Journal of Fish Biology, 37, 383-392.
Yadav, A. V. & Singh, B. R., 1989. Gross structure and dimensions of the gill in anair breathing estuarine goby, Pseudapocryptes lanceolatus. Japanese Journal ofIchthyology, 36, 252-259.
Yamamoto, S., 1931. Die statische Refraktion des Auge verchiedener Tiere. Journal ofOkayama Medical Society, 43, 1407-1425.
Yamazoe, Y., 1970. Nutritional studies on fish in Ariake Bay (IV). Free sugars composition of Mutugoro (Boleophthalmus pectinirostris) and Warasubo (Odontam-blyops rubicundis). Journal of the Japanese Society of Food and Nutrition, 23,21-23. (In Japanese with English abstract).
Yang, K.-G. & Zhang, Q.-Y., 1990. [Development stages of larval, juvenile andyoungmudskippers Boleophthalmus pectinirostris.] Journal of Xiamen University,Natural Science, 29, 76-80. (In Chinese with English abstract).
Yew, D. T. & Wu, M. F., 1979. Ratio of rods and different cones in fishes of shallowsea. Anatomische Anzeiger, 145, 155-160.
Zhang, H.-G. & He, D., 1989. [The comparison of the retino-motor response ofEpinephalus fario and Boleophthalmus pectinirostris under various light conditions.] Journalof Xiamen University, Natural Science, 28, 647-650. (In Chinesewith English abstract).
Zhang, Q.-Y., Hong, W.-S., Dai, Q.-N., Cai, Y.-Y. & Zhang, J., 1987. Studies on theartificial propagation and larval rearing of the mudskipper (Boleophthalmuspectinirostris L.).] Journal ofXiamen University, Natural Science, 26, 366-373.(In Chinese with English abstract).
Zhang, Q.-Y., Hong, J.-L., Dai, Q.-N., Zhang, J., Cai, Y.-Y., & Huang, J.-L., 1989.Studies on induced ovulation, embryonic development and larval rearing of themudskipper (Boleophthalmus pectinirostris). Aquaculture, 83, 375-385.
Zhang, Q.-Y. & Zhang, J., 1988. [On the feeding, growth and survival of larval mudskipper (Boloeophthalmus pectinirostris)]. Journal of Fisheries, China, 12,203-211. (In Chinese with English abstract).




















