
Poecilosclerida from the New Caledonia lagoon (Porifera: Demospongiae).
Upload
queenslandmuseumCategory
view
0download
0
Morphometric and Biochemical Differences Between Sympatric Populations of the Clathria "Spicata" Species Complex (Demospongiae: Poecilosclerida: Microcionidae) from Northern Australia
J.N.A. HOOPER, R.J. CAPON, C.P. KEENAN, and D.L. PARRY
Introduction
Morphological systematics of Recent Porifera are relatively difficult in compar-ison with other invertebrate phyla. On the one hand, morphological differences between supposedly distantly related higher taxa may appear to be minute or trivial, which is apparently a product of a long but conservative evolutionary history which was not produced by any radical morphological changes (Wiedenmayer 1977), but has produced an extremely diverse biochemical con-stitution (Bergquist 1978). Conversely, species may show a high degree of intraspecific morphological variability (e.g., Pansini 1982). In some cases that variability may be correlated directly with measurable differences in environ-mental factors (e.g., Palumbi 1984, 1986; Thompson and Murphy 1989), whereas in other cases species appear to be much more morphologically conservative across wide geographical ranges (e.g., Hooper 1987). Our knowledge of the effects of various environmental factors on intraspecific variability in sponges is still very limited (e.g., review in Simpson 1984), and it is not presently possible to construct models which may predict the behaviour of species (and their characters) under environmental changes (e.g., Simpson 1978; Palumbi 1984). These uncertainties have allowed authors to make quite different interpretations of the systematic importance of morphometric characters and levels of taxonomic divergence within the classification (cf. Hallmann 1912; Wilson 1921; Hooper 1991b). The present study examines a case in which only cryptic morphometric characters were able to consistently differentiate between two sympatric sibling species of Clathria, but species were otherwise clearly differentiated into two chemical populations. Methodology, descriptions of taxa and more complete results are presented elsewhere (Hooper et al. 1990).
Results
During the course of a larger study on the Microcionidae (Hooper 1991b), approximately 60 specimens of a soft, red-brown, branching or reticulate Clathria Schmidt were collected from various depths and localities along approximately 2300 km of the northwestern Australian coast (from Northwest Cape, Western Australia, to the Wessel Islands, Northern Territory). The "species" appeared to be relatively polymorphic in growth form, displaying up to four distinctive
J. Reitner and H. Keupp (Eds.) Fossil and Recent Sponges © Springer-Verlag Berlin Heidelberg 1991

272 J.N.A. Hooper et al.
morphotypes (Fig. 1). It also showed a few other apparently minor variations in morphological characters, such as the presence or absence of basal spination on megascleres and toxa thickness and curvature, whereas other features such as skeletal architecture, geometry of most spicules, spicule size, and live pigmen-tation were much more consistent, regardless of growth form type. Through a combination of those relatively consistent features, the "species" was certainly adequately defined and relatively easily differentiated from other Australasian Clathria. Intraspecific variability between specimens was remarkable, but cer-tainly no greater than known to occur in other sponge populations recorded in the literature (e.g., Pansini 1982). The "species" was characterized in particular by its skeletal architecture and localization of spicules within and around fibers. It was an obvious member of Hallmann's (1912) "spicata" species complex, which contained ten nominal species (Hooper 1991a), and the earliest available name for the taxon was Clathria lendenfeldi Ridley and Dendy (Hooper et al. 1990).
Subsequent biochemical analyses (general protein, carotenoid pigment and free amino acid profiles) and screening for antimicrobial activity showed that the "species" was in fact distinctly heterogeneous. Furthermore, two populations (chemotypes) were clearly differentiated, but those chemical populations did not appear to be correlated with any significant or obvious morphological key characters. Similarly, no correlations were found between those "chemical populations" and ecological, geographical or bathymetric distributions of sam-ples. The null hypothesis initially considered that the two sympatric populations were members of the same species (viz. C. lendenfeldi sensu lato), but this was tentatively rejected on the basis that electrophoretic "fingerprints" could con-sistently differentiate populations, and corroborated by other biochemical data. Much more rigorous systematics methods eventually provided some empirical evidence to support that conclusion, showing that there were in fact consistent (albeit relatively cryptic) morphometric differences between populations, and on that basis the two chemotypes were subsequently treated as sympatric sibling species. These were C., lendenfeldi Ridley and Dendy (1886), with six nominal species or subspecies as junior synonyms, and C. major Hentschel (1912) which was originally described as a subspecies of C. frondifera (Bowerbank) (Hooper 1991b). Morphometric and biochemical data which lead to those conclusions are summarized as follows.
Morphometrics
Shape (Fig.. 1). Four distinctive morphological forms were discovered for both species, showing an approximately similar range of gross morphological types: (1) branching (Fig. 1B,F), comprising 62% of specimens, with gross morphol-ogy of nominal species C. lendenfeldi, C. spicata Hallmann, and R. bispinosus Whitelegge; (2) clathrous, Echinoclathria-like (Fig. 1D,G), comprising 28% of specimens, and resembling the growth form of C. whiteleggei Dendy; (3) bushy, planar or multiplanar, lamellated fans (Fig. 1C,H); and (4) low growing ir-regularly bushy sponges (Fig. 1E), together comprising 10% of specimens.

Morphometric and Biochemical Differences
273
Fig. 1. Morphological variability in Clathria lendenfeldi (A-E) and C. major (F-I). (A NTM Z1339; B NTM Z0875; C NTM Z2701; D NTM Z1011; ENTM Z1870; FNTM Z1409; GNTM Z2091; HNTM Z2096; I NTM Z1755)
Colour. Pigmentation of both species was relatively consistent and stable, with the same range of hues: vivid red (Munsell 2.5R 4-5/10), bright red (2.5R 5/ 12-5R 4/10), bright red-orange (5R 7/8), mauve-red (2.5R 4/8-5R 5/6) or pale red- brown (5 R 6/6) in shallow-water specimens, whereas typical pigmentation was absent in deeper water specimens, varying from light brown (2.5YR 6/6), gray-brown (2.5YR 7/4) or light gray-brown (7.5YR 6-7/4-6). Several deeper-

274 J.N.A. Hooper et al.
water specimens were gray-brown with a turquoise (oxidized copper) ectosomal tinge (2.5B 7/4). Preserved material was beige, brown, gray-brown or muddy gray. Pigment was alcohol soluble. The presence of reddish colouration in shal-low-water populations of both species (intertidal-20 m) may be indicative of the functional photoprotective role of the carotenoid pigments, whereas those pig-ments would not be expected to be as predominant in specimens from deeper coastal waters (39-108 m).
Skeletal Structure (Fig. 2). The development of the ectosomal skeleton (consisting of brushes of small, basally spined auxiliary subtylostyles) partially obscured by a paratangential subectosomal skeleton (brushes of larger, basally smooth or spined auxiliary subtylostyles) appeared to vary significantly in both species, and spicimens of both species spanned the nominal genera Thalysias (or Rhaphidophlus) and Clathria. These data and similar evidence derived from population studies on other northern Australian microcionids (Hooper 1991b) provided sufficient reason to abandon the taxonomic differentiation between the genera Clathria and Thalysias.
a) 13% of specimens lacked an ectosomal skeleton entirely (true Clathria condition);
b) where present, the major proportion of dermal brushes were of subectosomal origin, and in many cases those structures masked the presence of smaller ectosomal subtylostyles, characteristic of the Thalysias condition;
c) other specimens exhibited a gradual progression from Clathria to Thalysias states, with a poorly developed paratangential ectosome (64%), or with occasional erect brushes of both ectosomal and subectosomal spicules, mainly in thicker regions (18%), to obvious well-developed Thalysias structures (5% of those specimens);
d) 6% of specimens had significant quantities of inorganic detritus on the ectosome and scattered throughout the mesohyl, but never incorporated into fibers (cf. the Clathriopsamma condition).
In the subectosomal region:
a) 87% of specimens had characteristically poorly developed subectosomal regions, identical in both species;
b) 5% showed marked structural differences between ectosomal and subecto-somal regions (these differences were most obvious in thicker regions of branches (e.g., surface projections);
c) 8% had a well-developed subectosomal region, characteristic of the Thalysias condition.
a) 26% of specimens had an undifferentiated and open extra-axial re-ticulation of fibers and spicule tracts;
b) 14% had markedly plumose and dendritic extra-axial primary tracts, connected by pauci- or aspicular (reticulate) fibers;
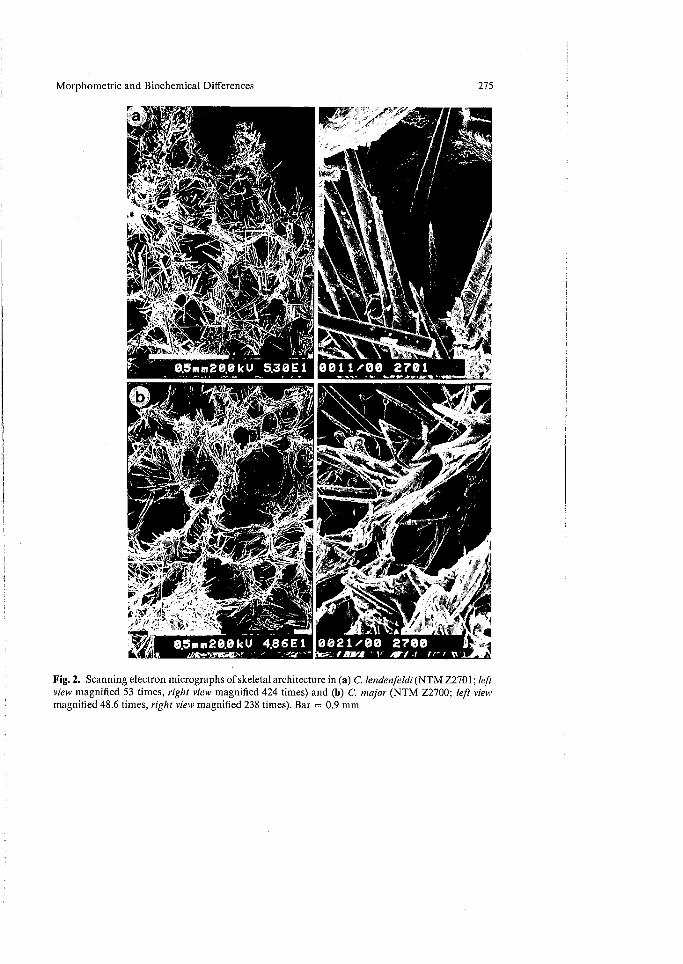
Fig. 2. Scanning electron micrographs of skeletal architecture in (a) C. lendenfeldi (NTM Z2701; left view magnified 53 times, right view magnified 424 times) and (b) C. major (NTM Z2700; left view magnified 48.6 times, right view magnified 238 times). Bar = 0.9 mm
Morphometric and Biochemical Differences 275
0011/00 2701
08ne20.131“) 4,86E1 uss"
0021/00 2700 1,1101M ' Al re° I V

276 J.N.A. Hooper et al.
c) 60% exhibited an extra-axial structure somewhere .between those two extremes, with reticulate fiber and plumo-reticulate spicule skeletons.
The choanosomal architecture of both species was identical and equally as variable (Fig. 2):
a) 46% of specimens had an openly reticulate fiber and spicule skeleton, without axial condensation or obvious axial and extra-axial differentiation (95% of that material had more-or-less regular reticulate architecture);
b) 13% had very slightly axially condensed fibers with plumose spicule tracts; c) 23% had an obvious axial condensation; d) 18% had multiple axial cores in the choanosome, which produced several
series of radiating plumose or plumo-reticulate extra-axial tracts.
' The presence of heavily echinated fibers by both acanthostyles and principal megascleres, particularly at fiber nodes, was a consistent feature of both species, and characteristic for Hallmann's (1912) "spicata" group of Clathria in general. The inclusion of spherical pigment granules dispersed throughout the mesohyl was common to many specimens, but that feature was not always present: 61% of specimens had abundant pigmented bodies, mostly confined to the periphery, whereas in 8% those bodies were scattered evenly throughout the mesohyl, in 13% they were rare, and in 18% of specimens they were absent entirely.
Megasclere Geometry (Fig. 3).
1. The bases of choanosomal principal megascleres in both species were typically spined, although fewer examples of entirely smooth spicules were also found in most (85%) specimens. The proportion of entirely smooth choanosomal megascleres ranged from 0-4% of spicules (44% of specimens), 5-10% (25%), 11-20% (27%), up to 40% of megascleres sampled (6% of specimens). In only one specimen smooth principal subtylostyles were predominant. Rare ex-amples of strongylote principal megasclere modifications were observed in few specimens (8%). Similarly, there was a tendency for acanthostyles and choanosomal subtylostyles to intergrade, with intermediate forms relatively common (Fig. 3). Both categories of spicules were found echina-ting fibers. The proportion of choanosomal principal megascleres with partly spined shafts ranged from 0-4% of spicules (48% of specimens), 5-10% (22%), 11-20% (25%), up to 32% of megascleres sampled (5% of specimens).
2. The geometry and distribution of larger subectosomal subtylostyles was relatively consistent in lendenfeldi. The majority of those samples had microspined subtylote bases, whereas fewer had smooth bases. The proportion of subectosomal subtylostyles with smooth bases ranged from 0-4% of spicules (46% of specimens), 5-10% (25%), 11-20% (26%), up to 40% of megascleres sampled (3% of specimens). In contrast, in specimens of major, up to 80% of larger auxiliary megascleres had spinose and hastate apices, the remainder being the usual fusiform and smooth forms.

Morphometric and Biochemical Differences 277
Fig. 3. Megascleres microscleres and skeletal structures of (a) C. lendenfeldi (NTM Z2095) and (b) C. major (NTM Z0858)
3. The bases of ectosomal auxiliary subtylostyles were invariably spined. They were morphologically similar to subectosomal spicules, but differed in size and distribution in the skeleton. Specimens of major had a variable proportion (30-80%) of apically spined and hastate spicules, and that feature was one of very few consistent characters differentiating the two species. In one specimen of major ectosomal megascleres were greatly reduced, and the dermal skeleton was composed mainly of larger auxiliary subtylostyles, and in another, both classes of auxiliary spicule were practically absent, replaced almost entirely by acanthostyles and choanosomal styles protruding through the surface.
4. The form and even spination of acanthostyles did not vary between specimens, regardless of seasonal or geographical distribution of samples. Acanthostyles of major were also significantly larger than those of lendenfeldi (F = 5.38, P< 0.0005), which appeared to be due to the greater elongation of the aspinose neck region on spicules.
Microsclere Geometry (Fig. 3).
1. The geometry of isochelae was entirely homogeneous within and between species. Isochelae were always present in samples but they often varied in abundance, ranging from abundant (3%), common (86%), uncommon (8%), to rare (3% of specimens). In addition, 18% of specimens had a few contort

278 J.N.A. Hooper et al.
isochelae (between 4-16% of spicules sampled), but those forms were not considered to be characteristic for the species. Isochelae of major were slightly smaller than those of lendenfeldi (F = 1.56, P< 0.0025), although their geometry was identical.
2. Both species had toxa microscleres which varied from long thin straight morphs to short thin recurved morphs (Fig. 3aG-F; 3bE-F), although the thickness and extent of curvature differed between species. The separation of toxas into two distinct morphological categories in either species was artificial, since many intermediate forms were present. Toxas varied greatly in length and abundance between samples of both species: in 36% of specimens they were rare, 23% uncommon, 26% moderately common, and in only 15% of specimens they were noticeably abundant. Toxas of major had a distinctly different geometry (Fig. 3bE) and thickness (F = 2.66, P< 0.0005) from those of lendenfeldi, and that feature appeared to be a consistent cryptic morphological feature differentiating the two taxa.
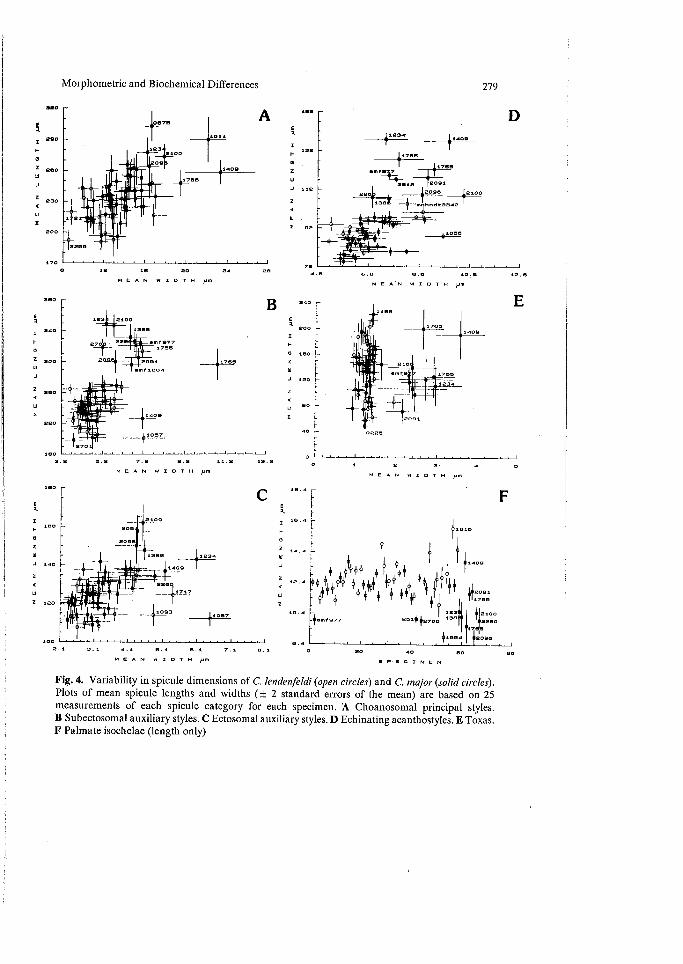
Variability in Spicule Dimensions (Fig. 4). In general, lendenfeldi had a high intraspecific variability in spicule size, for all spicule categories (Fig. 4A-F), but that variability was as consistent within groups as it was between groups (subdivided on the basis of locality, seasonal, bathymetric, and growth form classes). In fact, only the width of choanosomal styles showed any statistically significant differences between seasonal distributions of samples (analyzed for samples from the Darwin region only, P< 0.05) (Hooper et al. 1990). In other words, the species was equally as variable throughout Western Australian, Northern Territory, Queenslands, and Indonesian populations. In contrast, most megascleres in most specimens of lendenfeldi were generally (statistically) smaller than those in major. That trend was clearly indicated in visual plots of mean spicule lengths and widths ( ± 2 standard errors) (Fig. 4), but it was not an absolute difference and the two species showed some overlap. Statistical differences between the species were significant for acanthostyles only (Hooper et al. 1990), but those low significance levels are probably due to high levels of within-group variability.
Reproduction
Only three specimens of lendenfeldi and one of major were recorded with reproductive products, over 4 years observations. Viviparous larvae were oval to elongate, measuring 210-313 x 190-212 pm, and 290-330 X 380-397 p.m, respectively, and fecund individuals were collected from both the Darwin and Cobourg Peninsula regions of the Northern Territory (April-May, September- October). Larvae contained well-developed juvenile styles, isochelae and toxas located predominantly near the posterior pole and forming radial tracts, in addition to abundant spongin, and scattered pigment granules, identical with those observed in the adult. Patterns of ciliation on larvae were not observed (preserved material).
bsoee
72 10.6 12.6
MEA'N WIDTH pm
PA
MEAN WIDTH pm MEAN 1.1/071.1
4. I S. I 7. I 2.1 2.1
1,1234
11057
Morphometric and Biochemical Differences
.2 A
279
D 11234' 11402
1756
il7as
io
m,
1E 20." IT lwaoo
12 12 20 24 we
MEAN W /DTH
f1216
0 s Z
•
la .4
i
11402 0 43 1„44 11T,
r 4;1, 4.1 2091 w
•
30.4 [9i "
if
m 261#12700
20 40 60 SO MEAN WIDTH pm SPECIMEN
Fig. 4. Variability in spicule dimensions of C. lendenfeldi (open circles) and C. major (solid circles). Plots of mean spicule lengths and widths (± 2 standard errors of the mean) are based on 25 measurements of each spicule category for each specimen. A Choanosomal principal styles. B Subectosomal auxiliary styles. C Ectosomal auxiliary styles. D Echinating acanthostyles. E Toxas. F Palmate isochelae (length only)
'244. r 7 2 11/22(54

280 J.N.A. Hooper et al.
Ecology and Biogeography
Clathria lendenfeldi (sensu stricto) has been recorded previously from the Pacific coast of Australia and the Gulf of Manaar, India. Other species synonymized with lendenfeldi (Hooper et al. 1989) are also known to have a widespread distribution throughout (a) the Indian Ocean (C. spicata-W estern Australia, Cargados Ca-rajos, and Gulf of Aden; C. whiteleggei-Saya de Malha, Natal coast, Gulf of Aden and Arabian coast); (b) Arafura and Timor Seas (C. coppingeri var. aculeata-Aru I., Indonesia), and (c) Pacific coast of Australia (R. bispinosus -NSW coast). The present study found the species to be relatively abundant throughout the northwest coast of Australia. C. major has been previously recorded only from Aru I., Indonesia (Hentschel 1912), but in the present study it was found to have a relatively widespread distribution throughout northwest Australian waters, comparable to (and sympatric with) C. lendenfeldi.
Shallow-water specimens of both species (0-19 m depth) were found on the sides, tops, and in crevices of rock and dead coral heads, invariably exposed to currents (personal observations). Deeper-water specimens (39-108 m depth) were exposed to currents, attached to flat rock or coral substrate, with at least the basal portion of stems covered by silt or shell-grit (unpublished CSIRO in situ trawl photographs, P Young pers. commun ) Polychaete infestations were common in both species, and 41% (lendenfeldi) and 36% (major) of sections examined had at least one polychaete worm in the mesohyl [ Typosyllisspongicola (Scyllidae)]. One specimen of lendenfeldi contained numerous isopods on the external surface and burrowing into the sponge, and another was infested with ophiuroids, whereas one specimen of major was covered entirely with epiphytic algae.
Biochemical Systematics
Proteins (Fig. 5). From an electrophoretic study of 276 specimens, consisting of 13 species of Microcionidae, 15 Raspailiidae, and 23 other species of Axinellida, it was found that protein banding patterns of sponges were generally more difficult to detect in comparison with other marine taxa, but the silver-staining technique of Merril et al. (1982) as modified by Shaklee and Keenan (1986), proved to be particularly useful in detecting these low-concentration proteins. This technique also produced color differences in many bands, and these differences were found to be useful in determining whether or not proteins with the same relative mobilities (Rm) were in fact homologous. Banding patterns of all species examined were shown to be relatively consistent regardless of geo-graphic, seasonal, or other differences between samples (Hooper 1991b). Visual interpretation of "fingerprints" from ten specimens of C. lendenfeldi and seven of C. major showed that each species had quite different general protein banding patterns (Fig. 5). Intraspecific variability was relatively low, with mean levels of similarity between samples [ 1 standard error of the mean, (and coefficient of variation)] being 85.86% +1.31 (CoV = 4.58) and 85.90% ± 0.95 (CoV = 2.71) for

Morphometric and Biochemical Differences
281
r4
sic
H •
240 -
•220 -
200
180
160
140
120
100
80
60
40
20 -
a • .
b
. . • • •
• •
. •
• $
•
2 2 2 2 2 2 2 2 2 2 1 0 0 0 0 0 2 2 3 3 5 9 0 0 1 2 5 5 3 9 9 9 9 5 6 8 8 0 7 9 9 0 6 0 1 3 0 3 4 5 9 1 6 8 3 8 1 6 0 0 6 2
Clathria (Thalysias) lendenfeldi Clafbria (Thalysias) major
SPECIMEN
Fig. 5a,b. Intra- and interspecific comparisons in electrophoretically detectible protein profiles, showing the major relative mobility (Rm, millimeters) banding pattern of proteins in samples of C. lendenfeldi and C. major, collected from different localities and in different seasons
lendenfeldi and major, respectively. An interspecific comparison in protein banding patterns between lendenfeldi and major produced and average Gower metric similarity index of 80.4% between samples. This index included all relative mobility bands, regardless of consistency or possible origin from symbiotic organisms. A further comparison was made between all 13 microcionid species from northern Australia (Fig. 6a), in which similarity indices were averaged for each species, and in which only consistent relative mobility bands were con-sidered. From those data it was noted that for electrophoretic proteins at least, lendenfeldi and major were not necessarily the most closely related species, as would be otherwise expected from their postulated close morphological affinities.
Carotenoid Pigments (Table 1, Fig. 6b). The utilization of HPLC in the present study, over methods based on TLC (Lee and Gilchrist 1985) enabled much more

282
a
COVER METRIC SIMILARITY 60% 65% 70% 75% 80%
ELECTROPHORETIC PROTEINS Clathria frhalysiasl ratan:WU
(11.14) Clathria Malystaal coppinperi
(11s6) • Clathria (Thalywies) u.sp.372
Clathria (Clathriopsamme) tubtrosa (140)
Clathria (Vhalffiviss) lendenfoldi (11.10)
Clathria (Thalyviasl vulpina (Mel)
Clathria (Tbilisi.) major (W)
• Clathria (Thalywias) u.sp.606 (11s5)
Clathria llsociellal *countries (14)
• Clathria (rhalysias) u.sp.610 (11,3)
• Clathria (Thalywias) taxi: ore (11.12)
Clathria (Thalysiaa) ablating (11■25)
• Intho (Plocamilla) ridlayi ONO
60% 65% 70% 75% 80%
GOWER METRIC SIMILARITY 90% 92% 94% 96% 98% 100%
CAROTENOID PIGNIDITS Clathria (Thalywias) &biotin•
(11.14) Clathria (Thaliwissl vulpins
(11.6) Clathria (lhalysias) landenfaldi
(11.4) Clathria (Thalysias) usjor
Clathria (Isociellal acceatrica (1.7)
Clathria (Thalywies) copping's! (1.3)
• Clathria (Thalysies) nm.610 (11.3)
• Clathria (Thalyviss) toxilara (1S)
• Clathria (rhalysias) nm•606 (5.6)
Clathria (Clathriopsammal tuberose (1•71
Clathria (Thalysdasl reinvardti WU)
• Antho (nominal ridlityi (11s6)
90% 92% 94%. 96% 98% 100%
GOWER METRIC SIMILARITY 40% 50% 60% 70% 80% 90% 100% 1 I
FREE AMMO ACIDS
J.N.A. Hooper et al.
C 40% 50% 60% 70% 80%
• Antho (Plocaailla)
Clathria (Thalysias) liondenteldi
Clathria (Dendrocial Manias
Clathria (rhalysias) abietina (IRS Z2642)
Clathria (Thalysias1 abletina OM DI 24)
Clathria (Thalysiul Matins (11711 Z2611)
Clathria (thalysisslaajor
Clathria (Thalysias) vulpine
Clathria (Isociells) eccentrics
L— Clathria (Clathriopsamma) tuberosa
90% 100%

Morphometric and Biochemical Differences 283
Table 1. Carotenoid pigment profiles, showing the distribution (retention times; Rt minutes) and relative abundance (%) of carotenoids in samples of C. lendenfeldi and C. major, collected from different localities and in different seasons. ( + = trace detected only)
Retention Time (Rt) (min) 1
C. lendenfeldi 2 3
Sample"
4 C. major
1 2
1.74-1.78 26.1 11.8 2.9 4.1 1.94-1.98 51.3 20.1 7.1 29.6 6.3 13.9 2.43-2.58 1.1 1.3 2.82-2.97 3.04-3.17 7.6 3.58-3.77 4.23-4.41 2.5 1.1 4.82-5.36 3.0 33.9 8.9 35.5 5.7 5.5 5.40 5.61-5.75 5.90-6.51 + 7.6 14.6 13.3 10.6 6.80-7.18 7.24-7.55 7.70-8.06 + 2.5 6.0 8.7 6.9 5.9 8.47-8.88 + 5.0 1.6 8.94 9.62 9.90-10.53 12.0 21.5 44.4 4.2 42.3 61.8
10.84-11.07 17.4 11.32-11.65 12.08-12.12 12.34-12.43 12.72-12.97 5.1 7.1 0.8 2.3 3.4 14.61-14.82 + 5.0 11.0 1.0 2.1 2.2
a Samples: 1. C. lendenfeldi, NTM Z2093, Darwin region, dry season. 2. C. lendenfeldi, NTM Z2259, Darwin region, wet season. 3. C. lendenfeldi, NTM Z2386, Darwin region, pre-dry season. 4. C. lendenfeldi, NTM Z2503, Cobourg Peninsula region, dry season.
I. C. major, NTM Z2512, Cobourg Peninsula region, dry season. 2. C. major, NTM Z2506, Cobourg Peninsula, dry season.
Fig. 6. Numerical similarity (%) in general protein electrophoresis (a), carotenoid pigment profiles (b) and free amino acids (c) of representative Australasian Clathria species, based on a cluster analysis using the Gower metric similarity index and UPGMA (group-average) sorting (* incrusting species; N number of specimens analyzed)

284 J.N.A. Hooper et al.
accurate detection of even minute differences in both the composition and concentration of carotenoids between samples. However, conclusions derived from both these studies were based on phenetic comparisons between samples and species, which must assume that retention time (RT) peaks with the same or similar numerical values represent homologous proteins, whereas more sophis-ticated analyses are capable of molecular recognition of carotenoids (e.g., Liaaen-Jensen et al. 1982). Consequently, reliable conclusions from these data can only be made for comparisons based on multiple samples of individual species. From an examination of 178 samples, consisting of 12 species of Microcionidae, 13 Raspailiidae, and 12 other species of Axinellida, it was found that levels of intraspecific variability between samples were generally much lower than those recorded for electrophoretically detectable proteins (Hooper 199 lb). Four specimens of C. lendenfeldi and two of C. major were analyzed (Table 1), and mean levels of similarity between samples were 95.53% +0.73 (CoV = 1.32) and 95.30% respectively. The average level of similarity between the two species was 92.7%, and on the basis of their carotenoid pigment characteristics at least, the two taxa appeared to be more closely related to each other than they were to any of the other 10 species of Microcionidae examined (Fig. 6b). Another important difference found between C. lendenfeldi and C. major was that the water-soluble fraction of sample extracts in major only showed antibiotic activity against B. subtilis. The biological activity observed was not pronounced, nor have analyses proceeded further to enable the identification of active molecules, but activity was consistent for both samples of the species and may be an accurate representation.
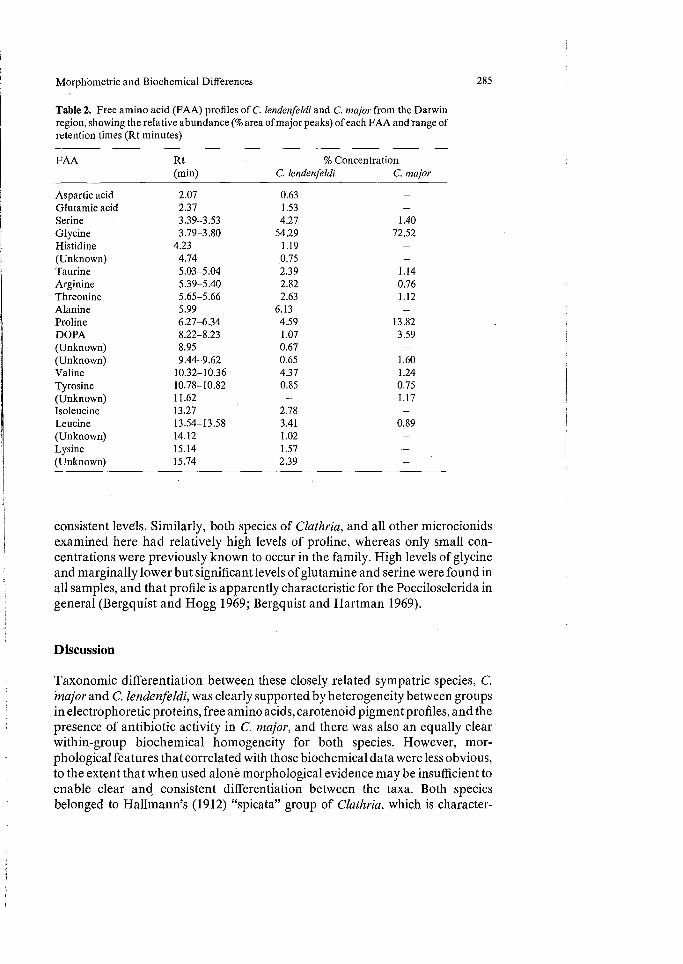
Free Amino Acids (Table 2, Fig. 6c). Free amino acid (FAA) analyses provided accurate molecular comparison between samples, and these data are also directly comparable with results of earlier studies (Bergquist and Hogg 1969; Bergquist and Hartman 1969). Thirty-nine specimens, consisting of nine species of Microcionidae, nine Raspailiidae, and 12 other species of Axinellida were examined. All species were clearly differentiated from each other by the dis-tribution and concentrations of FAA's, and these profiles did not appear to vary significantly between conspecific populations (checked for multiple samples of Clathria abietinus) (Fig. 6c). Substantial differences were noted to occur between FAA's in C. lendenfeldi and C. major (Table 2). C. lendenfeldi had a greater diversity of amino acids, although many of those occurred only in relatively low con-centrations (and which less sophisticated methods of detection would probably record as merely trace quantities), and FAA's such as serine, alanine and leucine were relatively more common. By comparison, C. major had fewer FAA's but significantly higher concentrations of proline, DOPA, and glycine, in particular (per given unit weight of sample); the glycine:proline ration was 11:1 in C. lendenfeldi and 5:1 in C. major. In general, the FAA profiles of the 2 species of Microcionidae examined here, the other 7 species examined (Fig. 6c), and the three species known from previous studies (Bergquist and Hogg 1969; Bergquist and Hartman 1969) were in relatively close agreement. A significant difference between those studies was the low concentrations of taurine in present samples, whereas earlier results showed that the amino acid occurred at relatively high and
Morphometric and Biochemical Differences 285
Table 2. Free amino acid (FAA) profiles of C. lendenfeldi and C. major from the Darwin region, showing the relative abundance (% area of major peaks) of each FAA and range of retention times (Rt minutes)
FAA Rt (min)
% Concentration C. lendenfeldi C. major
Aspartic acid 2.07 0.63 Glutamic acid 2.37 1.53 Serine 3.39-3.53 4.27 1.40 Glycine 3.79-3.80 54.29 72.52 Histidine 4.23 1.19 - (Unknown) 4.74 0.75 - Taurine 5.03-5.04 2.39 1.14 Arginine 5.39-5.40 2.82 0.76 Threonine 5.65-5.66 2.63 1.12 Alanine 5.99 6.13 - Proline 6.27-6.34 4.59 13.82 DOPA 8.22-8.23 1.07 3.59 (Unknown) 8.95 0.67 - (Unknown) 9.44-9.62 0.65 1.60 Valine 10.32-10.36 4.37 1.24 Tyrosine 10.78-10.82 0.85 0.75 (Unknown) 11.62 - 1.17 Isoleucine 13.27 2.78 - Leucine 13.54-13.58 3.41 0.89 (Unknown) 14.12 1.02 Lysine 15.14 1.57 (Unknown) 15.74 2.39
consistent levels. Similarly, both species of Clathria, and all other microcionids examined here had relatively high levels of proline, whereas only small con-centrations were previously known to occur in the family. High levels of glycine and marginally lower but significant levels of glutamine and serine were found in all samples, and that profile is apparently characteristic for the Poecilosclerida in general (Bergquist and Hogg 1969; Bergquist and Hartman 1969).
Discussion
Taxonomic differentiation between these closely related sympatric species, C. Major and C. lendenfeldi, was clearly supported by heterogeneity between groups in electrophoretic proteins, free amino acids, carotenoid pigment profiles, and the presence of antibiotic activity in C. major, and there was also an equally clear within-group biochemical homogeneity for both species. However, mor-phological features that correlated with those biochemical data were less obvious, to the extent that when used alone morphological evidence may be insufficient to enable clear and consistent differentiation between the taxa. Both species belonged to Hallmann's (1912) "spicata" group of Clathria, which is character-

286 J.N.A. Hooper et al.
ized in having mostly aspiculose (uncored) fibers heavily echinated by both acanthostyles and choanosomal principal subtylostyles, an ordered extra-fiber skeleton consisting of more or less plumose ascending spicule tracts composed of both choanosomal and subectosomal megascleres (producing the "spicate" arrangement from which the group derives its name), and the presence of choanosomal and subectosomal subtylostyles which project through (and mostly mask) the ectosomal skeleton.
Shape and surface features were found to be the least consistent of all characters examined, although it was possible to define variability to within finite ranges, and several nominal Clathria species which had been previously differentiated from other members of the "spicata" group on the basis of growth form (e.g., C. whiteleggei, C. spicata) were found to be clear synonyms of C. lendenfeldi (Hooper 1991a). Similarly, many features which have been used previously in the traditional identification of species (and even genera) of Microcionidae were not found to be particularly reliable in indicating new taxa. Those features were shared between these sibling species, in numerous other microcionid species complexes (Hooper 1991 b), and it was concluded that for this group only combinations of characters appear to be reliable indicators.
Of the 148 nominal Australasian Microcionidae species examined, including 16 species which occurred relatively commonly in the northwestern Australian region, C. lendenfeldi and C. major were clearly the most morphologically diverse sponge species of that group, so much so that some specimens were assigned to either taxon only with hesitation. Those anomalous examples differed from typical forms of both species only in details of questionable systematic importance (megasclere basal spination, isochelae size, degree of axial condensation of the skeleton). Specimens referred here to major differed from the remainder only by the geometry and thickness of toxas (cf. Fig. 3aF-G; 3bE-F), in the generally marginally, statistically) larger size of most megascleres (Fig. 4A-F), and in the presence of apically microspined auxiliary megascleres, similar to those found in a small group of other Clathria [viz. C. (Clathriopsamma) australiensis (Carter) from southern Australia, C. (Clathria) parthena (de Laubenfels) from California, and C. haplotoxa (Topsent) from north Africa]. In other details, C. major was equally as variable as C. lendenfeldi, and both species had sympatric geograph-ical, ecological, and bathymetric distributions, similar ranges of growth forms, skeletal architecture, and spiculation. As shown above, it is highly improbable that those morphological and biochemical differences merely represent phe-notypic variability between populations of a single species, whereas it is quite likely that they do represent fixed differences in the sponge genotype. De-spite the impracticalities associated with "field identifications" of that species complex, it is considered here that a combination of cryptic morphological characters, statistically different morphometric features, and biochemical differences between the taxa clearly differentiate them at the species level of classification.

Morphometric and Biochemical Differences 287
Acknowledgements. We wish to thank Daniel Low Choy, Stephen Brand, Diane Barton, Peter Scammels, and Paul Lynden for technical assistance during various aspects of this study, and Russell Hanley (NT Museum) for identifying polychaete worms. Shirley Stone (BMNH), Klaus RUtzler (USNM), Dietrich Kuhlmann (ZMB), Manfred Grasshoff (SMF), Claude Levi (MNHN), Lester Cannon (QMU), and Loisette Marsh (WAM) kindly provided numerous type specimens and other material used in this study. Peter Young (CSIRO Fisheries) generously provided access to his in situ trawl photographs of sponges from the Northwest Shelf. The Northern Territory University Planning Authority, Northern Territory Museum, University of Queensland and Australian National Uni-versity provided funds and equipment for biochemical analyses. The Australian Heritage Com-mission and CSIRO Fisheries are thanked for providing funds and shiptime, enabling collections to be made from the Cobourg Peninsula and the Northwest Shelf, respectively.
References
Bergquist PR (1978) Sponges. Hutchinson, London, pp 268 Bergquist PR, Hartman WD (1969) Free amino acid patterns and the classification of the Demo-
spongiae. Mar Biol 3(3):247-268 Bergquist PR, Hogg JJ (1969) Free amino•acid pattern in Demospongiae: a biochemical approach to
sponge classification. Cah Biol Mar 10:205-220 Hallmann EF (1912) Report on the Sponges obtained by the FIS "Endeavour" on the Coasts of New
South Wales, South Australia, Queensland, and Tasmania, 1909-10. Part 1. Zool Res Fish Exp FIS "Endeavour" 1909-10, (2): 117-300
Hentschel E (1912) Kiesel- und Hornschwamme der Aru und Kei-Inseln. Abh Senckenb Naturfbrsch Ges (1912):295-448
Hooper JNA (1987) New Records of Acarnus Gray (Porifera: Demospongiae: Poecilosclerida) from Australia, with a synopsis of the genus. Mem Queensl Mus 25(1):71-105
Hooper JNA (1991a) Character stability, systematics and affinities between Microcionidae (Poecilosclerida) and Axinellida. Proc 3rd Int Conf Biol Sponges. Smithson Contrib Zool: (in press)
Hooper JNA (1991b) Revision of the Australasian Microcionidae (Porifera :Demospongiae). Invert Tax (in preparation)
Hooper JNA, Capon RJ, Keenan CP, Parry DL (1990) Biochemical and morphometric differences of two sympatric sibling species of Clathria (Porifera: Demospongiae: Microcionidae) from northern Australia. Invert Tax 4(1):123-148
Lee WL, Gilchrist BM (1985) Carotenoid patterns in twenty-nine species of sponges in the order Poecilosclerida (Porifera: Demospongiae): a possible tool for chemosystematics. Mar Biol 86:21-35
Liaaen-Jensen S, Renstrom B, Ramdahl T, Hallenstvet M, Bergquist P (1982) Carotenoids of Marine Sponges. Biochem Syst Ecol 10(2): 167-174
Merril CR, Goldman G, van Keuren ML (1982) Simplified silver protein detection and image enhancement methods in polyacrylamide gels. Electrophoresis 3:17-23
Palumbi SR (1984) Tactics of acclimation: morphological changes of sponges in an unpredictable environment. Science 225 (4669): 1478-1480
Palumbi SR (1986) How body plans limit acclimation: responses of a demosponge to wave force. Ecology 67(1):208-214
Pansini M (1982) Notes on some Mediterranean Axinella with description of two species. Boll Mus Ist Biol Univ Genova 50-51:79-98
Ridley SO, Dendy A (1886) Preliminary report on the monaxonida collected by the HMS "Challenger". Ann Mag Nat Hist (5)18:325-351, 470-493
Shaklee JB, Keenan CP (1986) A practical laboratory guide to the techniques and methodology of electrophoresis and its application to fish fillet identification. CSIRO Mar Lab Aust Rep (177):1-59
Simpson TL (1978) The biology of the marine sponge Microciona prolifera (Ellis and Solander). 3. Spicule secretion and the effect of temperature on spicule size. J Exp Mar Biol Ecol 35(1):31-42

288 J.N.A. Hooper et al.: Morphometric and Biochemical Differences
Simpson TL (1984) The cell biology of sponges. Sprihger, Berlin Heidelberg New York Tokyo, pp 662 Thompson JE, Murphy PT (1989) Environmentally induced variation in the diterpene composition
of a Great Barrier Reef sponge. Proc 3rd Int Conf Biol Sponges. Smithson Contrib Zool (in press) Wiedenmayer F (1977) The Nepheliospongiidae Clarke, 1900 (Demospongea , Upper Devonian to
Recent) an ultraconservative, chiefly shallow-marine sponge family. Eclogae Geol Hely 70(3):885-918
Wilson HV (1921) The genus Raspailia and the independent variability of diagnostic features. J Elisha Mitchell Sci Soc Univ N Carolina Chapel Hill 37:54-60
Copyright © 2022 FDOKUMEN




















