
Morphological Space Stability in Rivers under Different Disturbance Regimes
12
BioOne sees sustainable scholarly publishing as an inherently collaborative enterprise connecting authors, nonprofit publishers, academic institutions, research libraries, and research funders in the common goal of maximizing access to critical research. Morphological Space Stability in Rivers under Different Disturbance Regimes Author(s): Cecília G. Leal, Nara T. Junqueira, Carlos Bernardo M. Alves, and Paulo S. Pompeu Source: Copeia, 2014(1):149-159. 2014. Published By: The American Society of Ichthyologists and Herpetologists DOI: http://dx.doi.org/10.1643/CE-12-096 URL: http://www.bioone.org/doi/full/10.1643/CE-12-096 BioOne (www.bioone.org ) is a nonprofit, online aggregation of core research in the biological, ecological, and environmental sciences. BioOne provides a sustainable online platform for over 170 journals and books published by nonprofit societies, associations, museums, institutions, and presses. Your use of this PDF, the BioOne Web site, and all posted and associated content indicates your acceptance of BioOne’s Terms of Use, available at www.bioone.org/page/terms_of_use . Usage of BioOne content is strictly limited to personal, educational, and non-commercial use. Commercial inquiries or rights and permissions requests should be directed to the individual publisher as copyright holder.
-
Upload
independent -
Category
Documents
-
view
1 -
download
0
Transcript of Morphological Space Stability in Rivers under Different Disturbance Regimes

BioOne sees sustainable scholarly publishing as an inherently collaborative enterprise connecting authors, nonprofit publishers, academic institutions, research libraries, andresearch funders in the common goal of maximizing access to critical research.
Morphological Space Stability in Rivers under Different Disturbance RegimesAuthor(s): Cecília G. Leal, Nara T. Junqueira, Carlos Bernardo M. Alves, and Paulo S. PompeuSource: Copeia, 2014(1):149-159. 2014.Published By: The American Society of Ichthyologists and HerpetologistsDOI: http://dx.doi.org/10.1643/CE-12-096URL: http://www.bioone.org/doi/full/10.1643/CE-12-096
BioOne (www.bioone.org) is a nonprofit, online aggregation of core research in the biological, ecological, and environmentalsciences. BioOne provides a sustainable online platform for over 170 journals and books published by nonprofit societies,associations, museums, institutions, and presses.
Your use of this PDF, the BioOne Web site, and all posted and associated content indicates your acceptance of BioOne’s Terms ofUse, available at www.bioone.org/page/terms_of_use.
Usage of BioOne content is strictly limited to personal, educational, and non-commercial use. Commercial inquiries or rights andpermissions requests should be directed to the individual publisher as copyright holder.

Morphological Space Stability in Rivers under Different
Disturbance Regimes
Cecılia G. Leal1,2, Nara T. Junqueira2, Carlos Bernardo M. Alves3, andPaulo S. Pompeu2
Morphological characteristics of fish communities were compared in four rivers with different levels of anthropogenicdisturbance: Cipo, Curimataı, Pardo Grande, and Velhas. Discriminant function analysis indicated no morphologicaldifferences among the communities, suggesting a general morphological pattern for different river communities,regardless of their disturbance level. However, this pattern was shaped by different taxa in each river. Because of thepoorer condition of the Velhas River (substrate type, riparian vegetation, and anthropogenic disturbance), it seemsthat species morphological range is influencing the common morphological pattern. Even with notable differences inbiological condition, the Velhas River still accommodates the same range of morphological types, but not the samespecies pool found in well-preserved rivers within the basin. For this reason, the maintenance of tributaries, with theirparticular fish species and different physical habitat characteristics, is essential for the conservation of theichthyofauna on a basin scale. Moreover, the analysis of morphology and composition together allowed a more criticalapproach of disturbance effects and suggested that these community aspects respond differently.
SEVERAL aspects of a species niche can be inferred basedon morphological features that affect ecological per-formance (Wikramanayake, 1990; Winemiller, 1991;
Wainwright and Reilly, 1994). Neotropical fishes are highlydiversified morphologically, with different body shapes andmouth structures enabling them to exploit specific habitats(Lowe-McConnell, 1975). Species have specific requirementsregarding the habitat they occupy (e.g., feeding and repro-duction sites, physical and chemical water conditions), andspecies–environment relationships should strongly influencespecies permanence in a particular site. Consequently, thefinal composition of a community depends on both habitatrequirements and species–environment relationships (La-mouroux et al., 1999; Jackson et al., 2001; Lorion andKennedy, 2009).
The relationship between stream habitat complexity andfish species richness is well documented in the literature,and the combination of different substrates, water velocities(such as pools, runs, riffles), and depths are importantcomponents of habitat heterogeneity and consequentlyspecies habitat use (Gorman and Karr, 1978; Lamourouxet al., 1999; Langeani et al., 2005; Ferreira, 2007; Leal et al.,2011; Junqueira et al., 2012). More structurally complexhabitats, referred to as a mosaic of habitat patches (e.g.,shallow gravel bed rapids, mid-depth sandy shores, deep andslow-flowing muddy pool), provide proper areas for forag-ing, reproduction, and refuge from predators, and are thusmore attractive for fishes (Neves and Monteiro, 2003; Puseyand Arthington, 2003; Willis et al., 2005). Such structuralcomplexity of the aquatic environment interacts with bioticfeatures to further contribute to the community diversityand composition (Jackson et al., 2001).
Organic matter input from the riparian vegetation isparticularly important to instream physical habitat (Jacksonet al., 2001). The presence of wood debris can modify theflow locally and facilitate litter accumulation, resulting inareas with lower energetic demands and more options for
hiding and escaping predators, which are more attractive tofish. By increasing habitat heterogeneity, wood debris alsofacilitate species coexistence via niche partitioning. Finally,wood serves as a substrate for aquatic invertebrates andperiphyton, important food sources for the ichthyofauna(Angermeier and Karr, 1984; Hugueny, 1990; Jackson et al.,2001; Brooks et al., 2004; Ligeiro et al., 2009; Casatti et al.,2012).
Deforestation, road construction, dam installation, anddumping of pollutants usually modify stream physicalcharacteristics, resulting in habitat simplification andhomogenization alongside biodiversity loss. Such anthro-pogenic activities affect river morphology and vegetationcover, which both influence flow dynamics, the pattern bywhich rain reaches the channel and the amount ofparticulate matter that is carried to the river bed (Jacksonet al., 2001). Such disturbances lead to changes in substrate,current and depth, resulting in stream habitat complexityloss. In Brazil, most aquatic environments have already beenaltered by human activities and the consequences for thefish fauna are reflected in taxonomic compositional changesas well as functional ones (Casatti et al., 2012).
Based on the assumption that fish morphological charac-teristics have known functions that influence ecologicalperformance (e.g., swimming speed, maneuverability, andfeeding), ecological diversity in species assemblages can becoarsely inferred from the size of their morphological space(Winemiller, 1991; Douglas and Matthews, 1992). In thiscontext, the aim of this study was to test the hypothesis thatfish communities of rivers with different levels of anthropo-genic disturbance have different patterns of morphologicalspace occupation. Furthermore, we asked whether a moredegraded stream differs in its morphological hypervolume sizeand species dispersion in the morphological space from otherwell-preserved streams. We expected fish communities frommore impacted watercourses to differ in terms of its morpho-logical pattern, occupy less morphological space, and be less
1 Programa de Pos-graduacao em Ecologia Aplicada, Departamento de Biologia, Universidade Federal de Lavras–UFLA, CEP 37200-000, Lavras,Minas Gerais, Brazil; E-mail: [email protected]. Send reprint requests to this address.
2 Laboratorio de Ecologia de Peixes, Departamento de Biologia, Universidade Federal de Lavras–UFLA, CEP 37200-000, Lavras, Minas Gerais,Brazil; E-mail: (NTJ) [email protected]; and (PSP) [email protected].
3 Nuvelhas–Projeto Manuelzao, Universidade Federal de Minas Gerais–UFMG, Avenida Antonio Carlos, 6627, Pampulha, CEP 31270-901, BeloHorizonte, Minas Gerais, Brazil; E-mail: [email protected].
Submitted: 3 August 2012. Accepted: 14 September 2013. Associate Editor: J. F. Schaefer.F 2014 by the American Society of Ichthyologists and Herpetologists DOI: 10.1643/CE-12-096
Copeia 2014, No. 1, 149–159

dispersed in the morphological space due to homogeneoushabitat conditions leading to more similar morphologies.
MATERIALS AND METHODS
Study area.—The Velhas River, one of the major tributariesof the Sao Francisco River, is located in the central region ofMinas Gerais state, crossing the transition zone between theAtlantic Forest and Cerrado (savanna) biomes. Currently,the ichthyofauna of the Velhas River basin includes at least115 species (Alves and Pompeu, 2005); this number willlikely increase pending descriptions of new species that havebeen documented in the region.
The drainage basin of the Velhas River extends 761 kmand covers an area of 27.867 km2 (CETEC, 1983; PLANVASF,1986). In the first 100 km from its source, domestic andindustrial waste is dumped into the river from themetropolitan region of Belo Horizonte (MRBH hereafter),which over the years has contributed to the pollution andsiltation of the river. Several cities that occur along theVelhas River and use its water contribute to its degradation.
The fieldwork in this study was conducted in four rivers of theVelhas River basin (Fig. 1). Certain stretches of each river werechosen based on their similar sizes and discharge rates, thusaiding comparison (Table 1). Although many cities are locatednearby, three of the rivers are well preserved and can beconsidered references within the basin (Pompeu et al., 2005).On the other hand, the main Velhas River, upstream of theMRBH, receives domestic sewage from the Itabirito municipal-ity and mining waste from the region. The site is heavily silted,and riparian vegetation is sparse or absent. The differencebetween studied stretches regarding habitat condition wasclearly observed in the field (Fig. 2) and was confirmed throughthe application of the Rapid Assessment Protocol of HabitatDiversity. The protocol was proposed by Hannaford et al. (1997)and modified by Callisto et al. (2002) to better fit to tropicallotic streams. The protocol enables the quantification of aspectssuch as water, substrate, riparian vegetation, erosion, siltation,and channel flow in relation to anthropogenic impacts. Theevaluation results in a final score which reflects the level ofdisturbance (0–40 represents severely degraded streams, 41–60substantially altered, and 61–100 least impacted ones; Table 1).Based on this protocol Cipo, Curimataı, and Pardo Grandescored 78–88 and were classified as least impacted rivers, andVelhas scored 32 confirming its severely degraded condition.
Data sampling.—Data were collected during the dry seasonin September 2007. Each river was sampled over two days,resulting in a total of eight consecutive days in the field. In a500 m stretch in each river, the following habitat categorieswere sampled: rapids, beaches, submerged roots, and leafpacks. Each individual habitat was considered a samplingsite, resulting in at least 20 samples in each river, with theexception of the Cipo River, which only contained 12sample sites. During the day, fish were captured using bagseines (6 m long, 2.6 m high, 5 mm mesh) and semi-circularhand nets (80 cm in diameter, 1 mm mesh). Additionally wesampled the main channel of each river using gillnets (twosets of eight nets measuring 10 m long and 3 to 12 cmbetween opposite knots) that were placed at dusk andremoved the next morning. The individuals collected werefixed in 10% formalin. In the laboratory, they wereidentified to species, and then fixed in 70% ethanol. Smallindividuals (,5 cm) of the genus Hypostomus were difficultto identify and thus were recorded as Hypostomus spp. group.
Habitats were characterized in terms of depth, watervelocity and substrate type, and this information was usedto represent the rivers’ environmental characteristics (Ta-ble 1). Depth and average water velocity were measured inthe center of the habitat using a flow meter, with thevelocity measured at 60% of the depth. A sample of substratewas collected for laboratory granulometric analysis, and thedominant substrate was defined as the substrate at the 50th
percentile of the size gradient based on granulometry. Thefollowing substrate classes were considered: silt+clay(,0.063 mm), sand (0.063–2 mm), gravel (.2–16 mm),pebble (.16–300 mm), and boulder (.300 mm; Suguio,1973). The presence of leaf packs and submerged roots wasnoted.
The morphometric analyses were performed using tenindividuals (whenever possible) of each species from eachriver. For Phalloceros uai and Poecilia reticulata, only non-pregnant females were measured to avoid differences due tosexual dimorphism. The Hypostomus spp. were not consid-ered in these analyses because, as previously mentioned, itwas not always possible to identify to species level.Individuals smaller than 15 cm were measured with digitalcalipers (0.01 mm precision) and larger specimens with aruler (0.1 mm precision). Sixteen linear and five areameasurements were obtained from each specimen (Fig. 3).To calculate the area, the fins and the bodies of the fish were
Fig. 1. Rio das Velhas basin map, southeastern Brazil, showing thesampling points in the Curimataı, Pardo Grande, Cipo, and Velhas rivers.Geographic coordinate system datum: WGS, 1984; base: Geominas,1997; adapted: Manuelzao Project, 1999–2009; execution: Daniel N.Rodrigues under the supervision of Lussandra M. da Silva, NUVELHAS–Transdisciplinary and Interinstitutional Center for Restoration of Rio dasVelhas Basin.
150 Copeia 2014, No. 1
superimposed on graph paper to outline the shape of thefish. Size corrections consisted of regressing each length orarea against a linear measure or area of body. All measure-ments were used in the calculation of morphologicalattributes related to fish habitat use and locomotion aspects(Table 2). Pelvic, dorsal, and caudal fin attributes for taxalacking those fins (Gymnotiformes) were recorded as 0.
Data analysis.—To search for similarity in family composi-tion, a pairwise comparison of shared families among riverswas carried out through Jaccard’s similarity index. Speciesdistribution in morphological space was assessed via acorrelation-based principal component analysis (PCA),which aids simultaneous evaluation of several morpholog-ical attributes. The highest morphological attribute load-ings (values greater than 0.70) were retained for axesinterpretation. Axes with eigenvalues higher than unitywere retained for interpretation (Gatz, 1979a; Watson andBalon, 1984). Next, a step-by-step backward discriminantfunction analysis (DFA) was applied to detect differencesbetween each river community. In this analysis family wasincluded as a categorical variable to account for a possiblephylogenetic effect in the pattern found. The stepwiseprocedure was chosen as it can predict group membership,as well as identify which variables contribute most to groupseparation. Moreover DFA is particularly suitable for highlycorrelated variables, such as morphological attributes(Burns and Burns, 2008). For both PCA and DFA analysiswe considered the mean value of each morphologicalattributes for each species, and used Statistica 10.0 software(StatSoft, 2011).
We used Euclidean distances in morphological spaceamong species to test the hypothesis that more impactedwatersheds occupy less space in the ecomorphologicalhypervolume. For each river community, a normalizedEuclidean distance matrix was constructed between the
pairs of species from the morphological attributes accordingto the following equation:
Djk~Xn
i~1
xij{xik
� �2" #1=2
where Djk is the Euclidean distance between species j and k, n isthe number of attributes, and xij and xik are the values ofattribute i for the pair of species j and k. In the case of normalizeddistance, the data were adjusted for the Gaussian distributionwith a mean of zero and a standard deviation of one:
x’ik~ xik{XX�
k
� �=DPk,
where xik is the attribute mean k for the species I, Xk is the meanfor all xik, and DPk is the standard deviation for the attribute k.
In each mirrored Euclidean distance matrix, the distance ofeach species to the nearest neighbor (NND) was determined,which corresponds to the lowest value of each column. Themean of these NND values is the species dispersion index inmorphological space (Ricklefs et al., 1981). The mean of eachcolumn represents the centroid of each species, and the mean ofall species’ centroids represents the community centroid (CC).The difference between these values is the species’ distance tothe community centroid (CD), and the mean of the CD valuesmodule is an estimate of the relative size of the morphologicalhypervolume occupied by the community (Winemiller, 1991).The differences between the CD and the NND means betweenrivers were tested through ANOVA followed by the post hocTukey test using Statistica 10.0 software (StatSoft, 2011).
RESULTS
Fish captured included 4,051 individuals from 65 species, 13families, and five orders (Table 3). The greatest species richnesswas found in the Curimataı River (43), followed by the Pardo
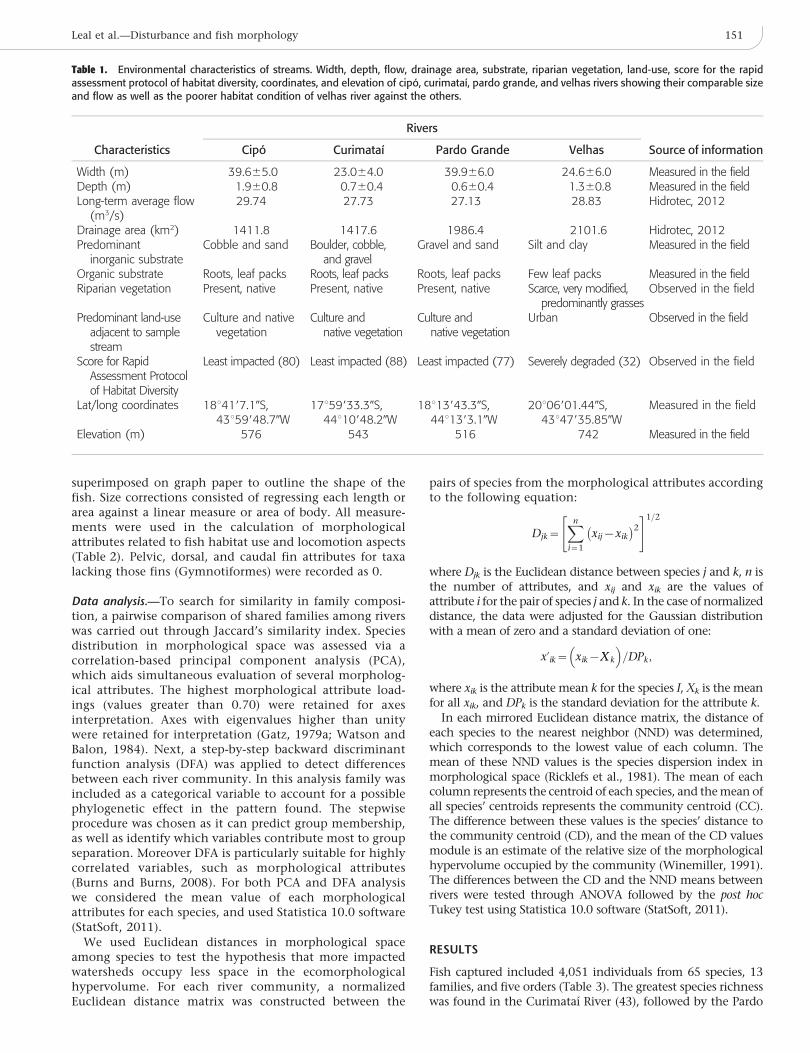
Table 1. Environmental characteristics of streams. Width, depth, flow, drainage area, substrate, riparian vegetation, land-use, score for the rapidassessment protocol of habitat diversity, coordinates, and elevation of cipo, curimataı, pardo grande, and velhas rivers showing their comparable sizeand flow as well as the poorer habitat condition of velhas river against the others.
Characteristics
Rivers
Source of informationCipo Curimataı Pardo Grande Velhas
Width (m) 39.665.0 23.064.0 39.966.0 24.666.0 Measured in the fieldDepth (m) 1.960.8 0.760.4 0.660.4 1.360.8 Measured in the fieldLong-term average flow
(m3/s)29.74 27.73 27.13 28.83 Hidrotec, 2012
Drainage area (km2) 1411.8 1417.6 1986.4 2101.6 Hidrotec, 2012Predominant
inorganic substrateCobble and sand Boulder, cobble,
and gravelGravel and sand Silt and clay Measured in the field
Organic substrate Roots, leaf packs Roots, leaf packs Roots, leaf packs Few leaf packs Measured in the fieldRiparian vegetation Present, native Present, native Present, native Scarce, very modified,
predominantly grassesObserved in the field
Predominant land-useadjacent to samplestream
Culture and nativevegetation
Culture andnative vegetation
Culture andnative vegetation
Urban Observed in the field
Score for RapidAssessment Protocolof Habitat Diversity
Least impacted (80) Least impacted (88) Least impacted (77) Severely degraded (32) Observed in the field
Lat/long coordinates 18u4197.10S,43u59948.70W
17u59933.30S,44u10948.20W
18u13943.30S,44u1393.10W
20u06901.440S,43u47935.850W
Measured in the field
Elevation (m) 576 543 516 742 Measured in the field
Leal et al.—Disturbance and fish morphology 151

Grande (32), Cipo (29), and Velhas (26). In the Cipo, Curimataı,and Pardo Grande rivers, most species were Characiformes(58.6%, 52.4%, and 74.2%, respectively) followed by Silur-iformes (34.5%, 42.9%, and 22.6%, respectively). No cyprino-dontiforms were recorded. An inverse pattern was found in the
Velhas River, with more siluriform species (46.2%) thancharaciforms (34.6%), and two cyprinodontiform species. Alsoin the Velhas River, none of the following migratory specieswere collected: Brycon nattereri, Brycon orthotaenia, Leporinusreinhardti, Leporinus taeniatus, Pimelodus maculatus, Prochilodus
Fig. 2. Sampling points at Cipo (A), Curimataı (B), Pardo Grande (C), and Velhas rivers (D, E). The first three pictures show the well-preservedriparian vegetation of Cipo, Curimataı, and Pardo Grande. Velhas River site shows a grass-dominated riparian vegetation and a remarkable presenceof anthropogenic disturbances such as houses (D, E), walking trails (D), fences (D), trash (E), as well as a silted area in the middle of the channelprobably due to the past presence of a bridge (E).
152 Copeia 2014, No. 1

costatus, and Salminus franciscanus. Curimatids and prochilo-dontids were collected only in the tributaries, and the non-native Poecilia reticulata occurred only in the Velhas River.
A pairwise comparison of shared families among the riversby Jaccard’s index was generally high; 0.80 for Curimataı vs.Pardo Grande, 0.71 for Curimataı vs. Cipo, and 0.56 forPardo Grande vs. Cipo. The lowest values were found for theVelhas River comparisons: 0.40 vs. Cipo River, 0.44 vs.Curimataı River, and 0.50 vs. the Pardo Grande River.
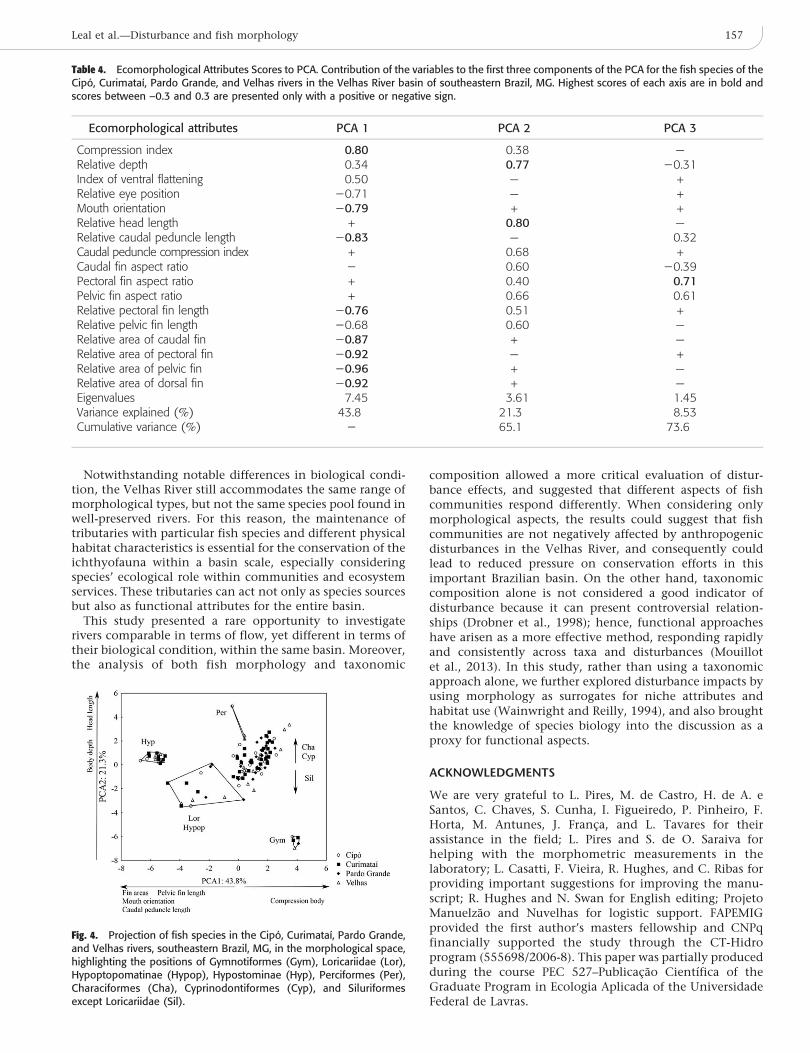
The first three axes of the PCA were retained forinterpretation, as their eigenvalues were higher than unity(Table 4). The first axis explained 43.8% of data variationand was negatively influenced by the pelvic fin relative area,dorsal fin relative area, caudal fin relative area, caudalpeduncle relative length, mouth orientation, relative pecto-ral fin length, and relative eye position and was positivelyinfluenced by the compression index. The highest scores forthis axis included species from the order Gymnotiformes,which have no pelvic, dorsal, or caudal fins (Fig. 4). On theother hand, the lowest scores belonged to species ofHypostominae, which have a ventrally oriented mouthand a dorsoventrally flattened body, resulting in low valuesof the compression index. The second axis, accounting for21.7% of the variance, had a higher positive contribution tothe relative head length and relative height. Thus, thespecies with higher scores on this axis were characiforms,cyprinodontiforms, and perciforms, all of which have adeeper body when compared to other orders. Finally, thethird axis only represented 8.4% of data variation and wasnegatively influenced by pectoral fin aspect ratio.
The species distribution in the first two axes of morpho-logical space shows that the four rivers communities occupya similar area, with a large overlap among them (Fig. 5). Thediscriminant function analysis reflected this pattern andshowed no statistical difference among the communities,resulting in no variables retained in the model. Morpholog-ical space for Pardo Grande River in the first two PCA axesseemed smaller than the other rivers because there was no
record of species belonging to the subfamily Hypostominae.However these graphs are bidimensional, while the mor-phological hypervolume is multidimensional and did notdiffer between rivers. Furthermore, despite the differences infamily composition between Velhas and the other rivers(Jaccard’s index presented above), family was not retained inthe DFA model.
Considering the Euclidian distance measurements be-tween pairs of species, there were no statistical differencesbetween Cipo (CD 5 1.08), Pardo Grande (CD 5 1.01),Curimataı (CD 5 0.84), and Velhas (CD 5 0.68) (F3,1 5 1.80;P 5 0.15). The species dispersion (NND mean) in themorphological space was only significantly different be-tween Pardo Grande (2.88) and Curimataı (2.00) river (F3,1 5
2.76; P 5 0.04). Cipo and Velhas rivers accounted for 2.64and 2.10, respectively.
DISCUSSION
Although the studied rivers are part of the same basin, theupper Velhas site differs in fish family composition from theother rivers. Ibanez et al. (2009) mentioned that similarity incommunity composition reflects common evolutionaryprocesses because phylogenetically close species are proba-bly ecologically similar. Part of the variation in taxonomiccomposition might reflect ecological aspects specific to eachriver, including physical characteristics such as velocity,substrate, and depth, which affect the habitats occupied byspecies (Gorman and Karr, 1978). Different communities areexpected in each river because local environmental gradi-ents, natural or human driven, and intra and inter-specificinteraction determine which species will become part of thecommunity (Jackson et al., 2001).
The most remarkable differences in family compositionfor the studied stretches could be due to poorer ecologicalconditions in Velhas River compared to the tributaries.Velhas River has a history of degradation and becameheavily polluted in the 20th century. It has been recognizedas the most polluted river of Minas Gerais state, as its basinhas the largest metropolitan region (MRBH in its upperportion), the highest gross domestic product, and the largesthuman population in the Sao Francisco basin (one of Brazil’slargest rivers; Alves and Pompeu, 2005). Such severeecological degradation has resulted in the local extinctionof several fish species in Velhas River, when comparingrecent records to historic data from 160 years ago (Alves andPompeu, 2005). In the present study, migratory species werenot captured at this site, unlike the other rivers. This resultcan be attributed to the downstream pollution released fromthe MRBH, which might act as a barrier to colonization inthis portion of the basin (Pompeu et al., 2005). Moreover,detritivorous species, such as Curimatidae and Prochilodon-tidae, were also not recorded at the Velhas River site. Theriver sediment is probably poor for these species; the inputof allochthonous plant matter is low due to scarce riparianvegetation, composed mainly of grasses and shrubs. Fur-thermore, detritivorous species may be affected by the sandybottom of the river and its constant flow, which preventsalgal growth. Casatti et al. (2006) suggest that someCurimatidae are negatively affected by siltation becausethey require special soft substrate for foraging. Similarly, theabsence of Hypoptopomatinae, species strongly related tosubmerged roots (Casatti and Castro, 1998), probably resultsfrom the depletion of riparian vegetation, consistent withthe fact that submerged roots were not found in this river.
Fig. 3. Schematic representation of the linear morphometric measure-ments: standard length (SL), maximum body height (MBH), bodymidline height (BMH), maximum body width (MBW), caudal pedunclelength (CPL), caudal peduncle height (CPH), caudal peduncle width(CPW), head length (HL), head height (HH), eye height (EH), caudal finwidth (CW), pectoral fin length (PtL), pectoral fin height (PtH), pelvic finlength (PvL), pelvic fin height (PvH) and the calculated areas: body area(BA), dorsal fin area (DA), caudal fin area (CA), pectoral fin area (PtA),and pelvic fin area (PvA) for Velhas River basin fish species.
Leal et al.—Disturbance and fish morphology 153

Besides taxonomic composition differences, we expectedthe fish community from Velhas River to differ in terms ofits morphological pattern, occupying less space in themorphological space. We also expected the community tobe less dispersed in the morphological space due tohomogeneous habitat conditions leading to more similarmorphologies. Willis et al. (2005) found greater morpho-logical variation and spacing of species within the morphol-ogy space in more structurally complex habitats. However,the discriminant function analysis based on morphologicalattributes indicated no morphological difference among thefour fish communities.
These results highlight the existence of a commonmorphological pattern among different fish communitiesfrom similar size rivers within the basin, independent oftheir disturbance level and taxonomic composition. Thiscould indicate that the river sites are not as different aspreviously assumed or that species have a wider morpho-logical range or plasticity to adapt to various environmentalfluctuations. Based on Velhas River’s poorer condition(substrate type, riparian vegetation, and anthropogenicdisturbance), it seems that species’ morphological range orplasticity is influencing the common morphological pat-tern. From 26 fish species registered in Velhas River, 19
Table 2. Morphological attributes and their ecological interpretations.
Morphological attributes Calculations Ecological interpretations
Compression index CI 5 MBH/MBW High CI values indicate laterally compressed fishes inhabitingenvironments with slow-flowing waters (Watson and Balon, 1984).
Relative depth RD 5 MBH/SL RD is inversely related to water velocity and directly related to thefish’s ability to perform vertical movements (Gatz, 1979a).
Index of ventralflattening
IVF 5 BMH Low IVF values indicate fishes living in environments with fast-flowing waters; these characteristics enable the fish to stay inposition with no swimming (Hora, 1930).
Relative eye position REP 5 EH/HH REP is assumed to be related to vertical habitat preference (Gatz,1979b) and high values suggest benthic fishes with dorsallylocated eyes (Watson and Balon, 1984).
Mouth orientation Angle formed between the tangentialplane to both lips and the perpendicularplane to the longitudinal axis of the bodywhen the mouth is open (Gatz, 1979b).
MO is characterized as follows: superior 5 between 10u and 80u;terminal 5 90u; inferior 5 between 100u and 170u and ventral 5
180u (Freire and Agostinho, 2001). Degree values were convertedto a decimal scale where 1u 5 60 (Casatti and Castro, 2006).
Relative head length RHL 5 HL/SL Directly related to prey size, high values of RHL suggests speciesthat are able to feed on relatively large prey (Gatz, 1979b).
Relative caudalpeduncle length
RCPL 5 CPL/SL Relatively long peduncles indicate fishes with good swimming abilitiesinhabiting turbulent waters (Gatz, 1979b; Watson and Balon,1984).
Caudal pedunclecompression index
CPCI 5 CPH/CPL The CPCI is inversely related to the range of swimming motion;high values indicate compressed peduncles, and this isassumed to be typical of less active swimmers (Gatz, 1979b).
Caudal fin aspect ratio CFAR 5 CW2/CA The CFAR is directly proportional to the amount of swimming by afish (Gatz, 1979b).
Pectoral fin aspect ratio PtFAR 5 PtL/PtW High values suggest long fins typical of fishes with remarkableswimming ability (Watson and Balon, 1984).
Pelvic fin aspect ratio PvFAR 5 PvL/PvW Low values are observed in fishes that use their pelvic fins for brakingand swimming forward, while high values are observed in fishes thatuse their pelvic fins to swim backward and to maintain their positionin the water column (Gatz, 1979b).
Relative pectoralfin length
RPtFL 5 PtL/SL High values suggest fishes that are able to perform many slowmaneuvers and inhabit slow-flowing waters (Gatz, 1979b).
Relative pelvicfin length
RPvFL 5 PvL/SL The RPvFL relates to the choice of habitat and is longer in rocky habitat-dwelling species and shorter in nektonic species (Gatz, 1979b).
Relative caudalfin area
RCFA 5 CA/BA High values indicate fins capable of quick propelling movements,typical of the swimming style of benthic fish (Watson and Balon,1984).
Relative pectoralfin area
RPtFA 5 PtA/BA High values indicate slow swimmers using fins to performmaneuvers, although high values may also indicate fishesinhabiting fast-flowing waters that use these fins as waterdeflection surfaces to help maintain their body close to thesubstrate (Watson and Balon, 1984).
Relative pelvic fin area RPvFA 5 PvA/BA Benthic fishes have relatively large RPvFA (Gatz, 1979b).Relative dorsal
fin areaRDFA 5 DA/BA The dorsal fin provides fishes with stability, and dorsal fins with small
areas are assumed to be more efficient in fast-flowing waters(Gosline, 1971).
154 Copeia 2014, No. 1
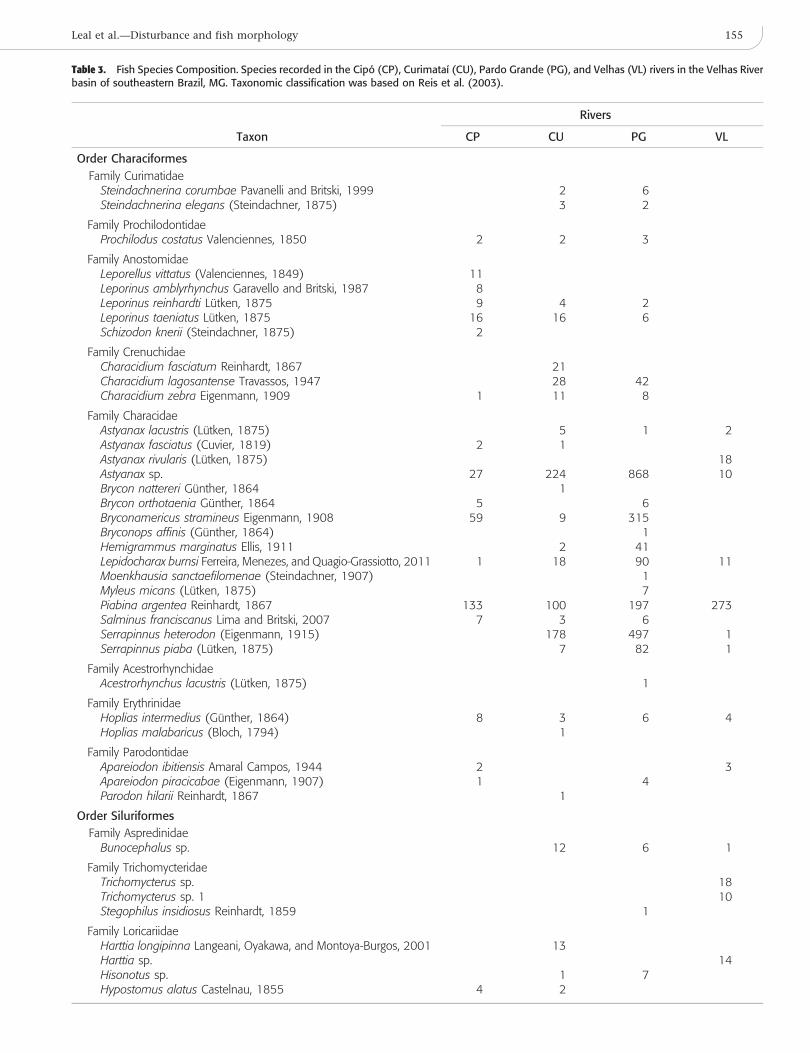
Table 3. Fish Species Composition. Species recorded in the Cipo (CP), Curimataı (CU), Pardo Grande (PG), and Velhas (VL) rivers in the Velhas Riverbasin of southeastern Brazil, MG. Taxonomic classification was based on Reis et al. (2003).
Taxon
Rivers
CP CU PG VL
Order Characiformes
Family CurimatidaeSteindachnerina corumbae Pavanelli and Britski, 1999 2 6Steindachnerina elegans (Steindachner, 1875) 3 2
Family ProchilodontidaeProchilodus costatus Valenciennes, 1850 2 2 3
Family AnostomidaeLeporellus vittatus (Valenciennes, 1849) 11Leporinus amblyrhynchus Garavello and Britski, 1987 8Leporinus reinhardti Lutken, 1875 9 4 2Leporinus taeniatus Lutken, 1875 16 16 6Schizodon knerii (Steindachner, 1875) 2
Family CrenuchidaeCharacidium fasciatum Reinhardt, 1867 21Characidium lagosantense Travassos, 1947 28 42Characidium zebra Eigenmann, 1909 1 11 8
Family CharacidaeAstyanax lacustris (Lutken, 1875) 5 1 2Astyanax fasciatus (Cuvier, 1819) 2 1Astyanax rivularis (Lutken, 1875) 18Astyanax sp. 27 224 868 10Brycon nattereri Gunther, 1864 1Brycon orthotaenia Gunther, 1864 5 6Bryconamericus stramineus Eigenmann, 1908 59 9 315Bryconops affinis (Gunther, 1864) 1Hemigrammus marginatus Ellis, 1911 2 41Lepidocharax burnsi Ferreira, Menezes, and Quagio-Grassiotto, 2011 1 18 90 11Moenkhausia sanctaefilomenae (Steindachner, 1907) 1Myleus micans (Lutken, 1875) 7Piabina argentea Reinhardt, 1867 133 100 197 273Salminus franciscanus Lima and Britski, 2007 7 3 6Serrapinnus heterodon (Eigenmann, 1915) 178 497 1Serrapinnus piaba (Lutken, 1875) 7 82 1
Family AcestrorhynchidaeAcestrorhynchus lacustris (Lutken, 1875) 1
Family ErythrinidaeHoplias intermedius (Gunther, 1864) 8 3 6 4Hoplias malabaricus (Bloch, 1794) 1
Family ParodontidaeApareiodon ibitiensis Amaral Campos, 1944 2 3Apareiodon piracicabae (Eigenmann, 1907) 1 4Parodon hilarii Reinhardt, 1867 1
Order Siluriformes
Family AspredinidaeBunocephalus sp. 12 6 1
Family TrichomycteridaeTrichomycterus sp. 18Trichomycterus sp. 1 10Stegophilus insidiosus Reinhardt, 1859 1
Family LoricariidaeHarttia longipinna Langeani, Oyakawa, and Montoya-Burgos, 2001 13Harttia sp. 14Hisonotus sp. 1 7Hypostomus alatus Castelnau, 1855 4 2
Leal et al.—Disturbance and fish morphology 155

(73%) were common to at least one of the tributaries. Williset al. (2005) found that fish assemblage morphologicalcentroids (interpreted as average morphological character-istics) were not correlated with habitat complexity, suggest-ing that fishes do not systematically shift toward differentmeans of using space, locomotion, and resources in relationto habitat complexity.
The other seven species were exclusive to Velhas River,despite belonging to the same morphological type as othertributary species, suggesting that they may have differentenvironmental requirements. For example, the morpholog-ically similar Poeciliidae (found only in Velhas River) andAnostomidae (exclusive to the tributaries) have differenttolerances to pollution and physical degradation. Therefore,it is likely that species composition differs as a result ofreplacement, whereby more sensitive and specialist speciesare replaced by more tolerant species along the gradient ofdegradation (Casatti et al., 2009, 2012). As proposed byJackson et al. (2001), the distribution of a species within its
potential range is a combination of historical and biogeo-graphical factors (e.g., colonization, speciation) and contem-poraneous aspects (e.g., biotic interactions, environmentalgradients) on a smaller scale. The combination of thesefactors would limit the group of species observed in eachcommunity. Thus, in degraded streams, it is not onlymorphological aspects related to habitat use that affect thepermanence of fish species, but also their full range ofbiological attributes (e.g., physiology, migratory condition,species interactions).
The Velhas River basin morphological pattern is shaped bydifferent groups of species. In the Cipo River, the deepestand slowest river, the Anostomidae were the richest,whereas in the Pardo Grande, composed mainly of sandybeaches, several species of small Characidae were registered.In the Curimataı River, characterized by rocky substrate andrapids, nine species of Hypostominae were found. Finally, inthe Velhas River, the fastest of the four rivers (Santos et al.,2009), the Siluriformes predominated.
Taxon
Rivers
CP CU PG VL
Hypostomus cf. auroguttatus 24Hypostomus francisci (Lutken, 1874) 22 83 51Hypostomus garmani (Regan, 1904) 4 1Hypostomus lima (Lutken, 1874) 16Hypostomus margaritifer (Regan, 1908) 6 17Hypostomus macrops (Eigenmann and Eigenmann, 1888) 7 36Hypostomus aff. paulinus 11Hypostomus sp. 3Group Hypostomus spp. 1 4 3 36Otocinclus xakriaba Schaefer, 1997 3 12Rineloricaria sp. 1 1 12
Family PseudopimelodidaeCephalosilurus fowleri Haseman, 1911 1 1
Family HeptapteridaeCetopsorhamdia iheringi Schubart and Gomes, 1959 3 2 1Imparfinis minutus (Lutken, 1874) 2 8 5Rhamdia quelen (Quoy and Gaimard, 1824) 1
Family PimelodidaeBagropsis reinhardti Lutken, 1874 4Pimelodus maculatus Lacepede, 1803 2 1 3Pimelodus pohli Ribeiro and Lucena, 2006 9 2
Order Gymnotiformes
Family SternopygidaeEigenmannia virescens (Valenciennes, 1836) 1 3 2 2Sternopygus macrurus (Bloch and Schneider, 1801) 1
Order Cyprinodontiformes
Family PoeciliidaePhalloceros uai Lucinda, 2008 73Poecilia reticulata Peters, 1859 24
Order Perciformes
Family CichlidaeAustraloheros facetus (Jenyns, 1842) 1 5Geophagus brasiliensis (Quoy and Gaimard, 1824) 2
Total richness 29 43 32 26
Table 3. Continued.
156 Copeia 2014, No. 1
Notwithstanding notable differences in biological condi-tion, the Velhas River still accommodates the same range ofmorphological types, but not the same species pool found inwell-preserved rivers. For this reason, the maintenance oftributaries with particular fish species and different physicalhabitat characteristics is essential for the conservation of theichthyofauna within a basin scale, especially consideringspecies’ ecological role within communities and ecosystemservices. These tributaries can act not only as species sourcesbut also as functional attributes for the entire basin.
This study presented a rare opportunity to investigaterivers comparable in terms of flow, yet different in terms oftheir biological condition, within the same basin. Moreover,the analysis of both fish morphology and taxonomic
composition allowed a more critical evaluation of distur-bance effects, and suggested that different aspects of fishcommunities respond differently. When considering onlymorphological aspects, the results could suggest that fishcommunities are not negatively affected by anthropogenicdisturbances in the Velhas River, and consequently couldlead to reduced pressure on conservation efforts in thisimportant Brazilian basin. On the other hand, taxonomiccomposition alone is not considered a good indicator ofdisturbance because it can present controversial relation-ships (Drobner et al., 1998); hence, functional approacheshave arisen as a more effective method, responding rapidlyand consistently across taxa and disturbances (Mouillotet al., 2013). In this study, rather than using a taxonomicapproach alone, we further explored disturbance impacts byusing morphology as surrogates for niche attributes andhabitat use (Wainwright and Reilly, 1994), and also broughtthe knowledge of species biology into the discussion as aproxy for functional aspects.
ACKNOWLEDGMENTS
We are very grateful to L. Pires, M. de Castro, H. de A. eSantos, C. Chaves, S. Cunha, I. Figueiredo, P. Pinheiro, F.Horta, M. Antunes, J. Franca, and L. Tavares for theirassistance in the field; L. Pires and S. de O. Saraiva forhelping with the morphometric measurements in thelaboratory; L. Casatti, F. Vieira, R. Hughes, and C. Ribas forproviding important suggestions for improving the manu-script; R. Hughes and N. Swan for English editing; ProjetoManuelzao and Nuvelhas for logistic support. FAPEMIGprovided the first author’s masters fellowship and CNPqfinancially supported the study through the CT-Hidroprogram (555698/2006-8). This paper was partially producedduring the course PEC 527–Publicacao Cientıfica of theGraduate Program in Ecologia Aplicada of the UniversidadeFederal de Lavras.
Table 4. Ecomorphological Attributes Scores to PCA. Contribution of the variables to the first three components of the PCA for the fish species of theCipo, Curimataı, Pardo Grande, and Velhas rivers in the Velhas River basin of southeastern Brazil, MG. Highest scores of each axis are in bold andscores between –0.3 and 0.3 are presented only with a positive or negative sign.
Ecomorphological attributes PCA 1 PCA 2 PCA 3
Compression index 0.80 0.38 —Relative depth 0.34 0.77 20.31Index of ventral flattening 0.50 — +Relative eye position 20.71 — +Mouth orientation 20.79 + +Relative head length + 0.80 —Relative caudal peduncle length 20.83 — 0.32Caudal peduncle compression index + 0.68 +Caudal fin aspect ratio 2 0.60 20.39Pectoral fin aspect ratio + 0.40 0.71Pelvic fin aspect ratio + 0.66 0.61Relative pectoral fin length 20.76 0.51 +Relative pelvic fin length 20.68 0.60 —Relative area of caudal fin 20.87 + —Relative area of pectoral fin 20.92 — +Relative area of pelvic fin 20.96 + —Relative area of dorsal fin 20.92 + —Eigenvalues 7.45 3.61 1.45Variance explained (%) 43.8 21.3 8.53Cumulative variance (%) 2 65.1 73.6
Fig. 4. Projection of fish species in the Cipo, Curimataı, Pardo Grande,and Velhas rivers, southeastern Brazil, MG, in the morphological space,highlighting the positions of Gymnotiformes (Gym), Loricariidae (Lor),Hypoptopomatinae (Hypop), Hypostominae (Hyp), Perciformes (Per),Characiformes (Cha), Cyprinodontiformes (Cyp), and Siluriformesexcept Loricariidae (Sil).
Leal et al.—Disturbance and fish morphology 157

LITERATURE CITED
Alves, C. B. M., and P. S. Pompeu. 2005. Historical changesin the rio das Velhas fish fauna–Brazil. American FisheriesSociety Symposium 45:587–602.
Angermeier, P. L., and J. R. Karr. 1984. Relationshipsbetween woody debris and fish habitat in a small warm-water stream. Transactions of the American FisheriesSociety 113:716–726.
Brooks, A. P., P. Gehrke, J. D. Jansen, and T. B. Abbe. 2004.Experimental reintroduction of woody debris on theWilliams River, NSW: geomorphic and ecological respons-es. River Research and Applications 20:513–536.
Burns, R., and R. Burns. 2008. Discriminant analysis,p. 589–608. In: Business Research Methods and StatisticsUsing SPSS. R. Burns and R. Burns (eds.). SAGE Publica-tions Ltd, London.
Callisto, M., W. Ferreira, P. Moreno, M. D. C. Goulart, andM. Petrucio. 2002. Aplicacao de um protocolo de avaliacaorapida da diversidade de habitats em atividades de ensino epesquisa (MG-RJ). Acta Limnologica Brasiliensia 14:91–98.
Casatti, L., and R. M. C. Castro. 1998. A fish community of theSao Francisco River headwater riffles, southeastern Brazil.Ichthyological Exploration of Freshwaters 9:229–242.
Casatti, L., and R. M. C. Castro. 2006. Testing theecomorphological hypothesis in a headwater riffles fishassemblage of the rio Sao Francisco, southeastern Brazil.Neotropical Ichthyology 4:203–214.
Casatti, L., C. P. Ferreira, and F. R. Carvalho. 2009. Grass-dominated stream sites exhibit low fish species diversityand dominance by guppies: an assessment of two tropicalpasture river basins. Hydrobiologia 632:273–283.
Casatti, L., F. Langeani, A. M. Silva, and R. M. C. Castro.2006. Stream fish, water and habitat quality in pasturedominated basin, southeastern Brazil. Brazilian Journal ofBiology 66:681–696.
Casatti, L., F. B. Teresa, T. Goncalves-Souza, E. Bessa, A. R.Manzotti, C. da S. Goncalves, and J. de O. Zeni. 2012.From forests to cattail: How does the riparian zone influencestream fish? Neotropical Ichthyology 10:205–214.
CETEC–Fundacao Centro Tecnologico de Minas Gerais.1983. Diagnostico ambiental do estado de Minas Gerais.Serie de Publicacoes Tecnicas/SPT-010:1, Belo Horizonte.
Douglas, M. E., and W. J. Matthews. 1992. Does morphol-ogy predict ecology? Hypothesis testing within a freshwa-ter stream fish assemblage. Oikos 65:213–224.
Drobner, U., J. Bibby, B. Smith, and J. B. Wilson. 1998.The relation between community biomass and evenness:
Fig. 5. Projection of the first two principal components based on 17 fish morphological attributes for the Cipo, Curimataı, Pardo Grande, and Velhasrivers in the Velhas River basin of southeastern Brazil, MG. Species that comprise each river community are highlighted with filled circles, and theremaining species are indicated by empty squares.
158 Copeia 2014, No. 1

What does community theory predict, and can thesepredictions be tested? Oikos 82:295–302.
Ferreira, K. M. 2007. Biology and ecomorphology of streamfishes from the rio Mogi-Guacu basin, southeastern Brazil.Neotropical Ichthyology 5:311–326.
Freire, A. G., and A. A. Agostinho. 2001. Ecomorfologia deoito especies dominantes da ictiofauna do reservatorio deItaipu Parana/Brasil. Acta Limnologica Brasiliensis 13:1–9.
Gatz, A. J. 1979a. Community organization in fishes asindicated by morphological features. Ecology 60:711–718.
Gatz, A. J. 1979b. Ecological morphology of freshwaterstream fishes. Tulane Studies in Zoology and Botany21:91–124.
Gorman, O. T., and J. R. Karr. 1978. Habitat structure andstream fish communities. Ecology 59:507–515.
Gosline, W. A. 1971. Functional Morphology and Classificationof Teleostean Fishes. University of Hawaii, Honolulu.
Hannaford, M. J., M. T. Barbour, and V. H. Resh. 1997.Training reduces observer variability in visual-basedassessments of stream habitat. Journal of North AmericanBenthological Society 16:853–860.
Hidrotec. 2012. Atlas digital das aguas de Minas. Ruralmi-nas/Igam/UFV. Available at: http://www.atlasdasaguas.ufv.br. Accessed in 15 August 2013.
Hora, S. L. 1930. Ecology, bionomics and evolution of thetorrential fauna, with special reference to the organs ofattachment. Philosophical Transactions of the RoyalSociety of London 28:171–282.
Hugueny, B. 1990. Richesse des peuplements de poissonsdans le Niandan (haut Niger, Afrique) en fonction de lataille de la riviere et de la diversite de milieu. RevueHydrobiologie Tropicale 23:351–364.
Ibanez, C., J. Belliard, R. M. Hughes, P. Irz, A. Kamdem-Toham, N. Lamouroux, P. A. Tedesco, and T. Oberdorff.2009. Convergence of temperate and tropical stream fishassemblages. Ecography 32:1–13.
Jackson, D. A., P. R. Peres-Neto, and J. D. Olden. 2001. Whatcontrols who is where in freshwater fish communities—theroles of biotic, abiotic, and spatial factors. Canadian Journalof Fisheries and Aquatic Sciences 58:157–170.
Junqueira, N. T., C. G. Leal, C. B. M. Alves, and P. S.Pompeu. 2012. Morphological diversity of fish along therio das Velhas, Minas Gerais, Brazil. Neotropical Ichthy-ology 10:417–424.
Lamouroux, N., H. Capra, M. Pouilly, and Y. Souchon.1999. Fish habitat preferences in large streams of southernFrance. Freshwater Biology 42:673–687.
Langeani, F., L. Casatti, H. S. Gameiro, A. Belluco-do-Carmo, and D. C. R. Feres. 2005. Riffle and pool fishcommunities in a large stream of southeastern Brazil.Neotropical Ichthyology 3:305–311.
Leal, C. G., N. T. Junqueira, and P. S. Pompeu. 2011.Morphology and habitat use by fishes of the Rio dasVelhas basin in southeastern Brazil. Environmental Biol-ogy of Fishes 90:143–157.
Ligeiro, R., A. S. Melo, and M. Callisto. 2009. Spatial scaleand the diversity of macroinvertebrates in a Neotropicalcatchment. Freshwater Biology 55:424–435.
Lorion, C. M., and B. P. Kennedy. 2009. Riparian forestbuffers mitigate the effects of deforestation on fishassemblages in tropical headwater streams. EcologicalApplications 19:468–479.
Lowe-McConnell, R. H. 1975. Fish Communities in Trop-ical Freshwaters. Longman, New York.
Mouillot, D., N. A. J. Graham, S. Villeger, N. W. H. Mason,and D. R. Bellwood. 2013. A functional approach revealscommunity responses to disturbances. Trends in Ecologyand Evolution 28:167–177.
Neves, F. M., and L. R. Monteiro. 2003. Body shape and sizedivergence among populations of Poecilia vivipara incoastal lagoons of south-eastern Brazil. Journal of FishBiology 63:928–941.
PLANVASF (Plano Diretor para o Desenvolvimento doVale do Sao Francisco). 1986, Companhia de Desenvol-vimento do Vale do Sao Francisco, Brasılia.
Pompeu, P. S., C. B. M. Alves, and M. Callisto. 2005. Theeffects of urbanization on biodiversity and water qualityin the Rio das Velhas Basin, Brazil. American FisheriesSociety Symposium 47:11–22.
Pusey, B. J., and A. H. Arthington. 2003. Importance of theriparian zone to the conservation and management offreshwater fish: a review. Marine and Freshwater Research54:1–16.
Reis, R. E., S. O. Kullander, and C. J. Ferraris, Jr. 2003.Checklist of the Freshwater Fishes of South and CentralAmerica. Edipucrs, Porto Alegre, Brazil.
Ricklefs, R. E., D. Cochran, and E. R. Pianka. 1981. Amorphological analysis of the structure of communities oflizards in desert habitats. Ecology 62:1474–1483.
Santos, H. de A. e, P. S. Pompeu, S. F. Cunha, M. Leite, andC. Chaves. 2009. Hydrodynamic and physical assessmentfor the four reaches in Velhas River basin, Minas Gerais,p. 1–10. In: Proceedings of the 7th International Sympo-sium on Ecohydraulics. Conception.
StatSoft, Inc. 2011. STATISTICA (data analysis softwaresystem), version 10. www.statsoft.com
Suguio, K. 1973. Introducao a Sedimentologia. EdgardBlucher/EDUSP, Sao Paulo.
Wainwright, P. C., and S. M. Reilly. 1994. EcologicalMorphology: Integrative Organismal Biology. Universityof Chicago Press, Chicago.
Watson, D. J., and E. Balon. 1984. Ecomor-phological analysis of taxocenes in rainforest streamsof northern Borneo. Journal of Fish Biology 25:371–384.
Wikramanayake, E. D. 1990. Ecomorphology andbiogeography of a tropical stream fish assemblage:evolution of assemblage structure. Ecology 71:1756–1764.
Willis, S. C., K. O. Winemiller, and H. Lopez-Fernandez.2005. Habitat structural complexity and morphologicaldiversity of fish assemblages in a Neotropical floodplainriver. Oecologia 142:284–295.
Winemiller, K. O. 1991. Ecomorphological diversificationin lowland freshwater fish assemblages from five bioticregions. Ecological Monographs 61:343–365.
Leal et al.—Disturbance and fish morphology 159




















