
Modification of chromium (VI) phytotoxicity by exogenous gibberellic acid application in Pisum...
15
1 23 Acta Physiologiae Plantarum ISSN 0137-5881 Volume 33 Number 4 Acta Physiol Plant (2011) 33:1385-1397 DOI 10.1007/s11738-010-0672- x Modification of chromium (VI) phytotoxicity by exogenous gibberellic acid application in Pisum sativum (L.) seedlings Savita Gangwar, Vijay Pratap Singh, Prabhat Kumar Srivastava & Jagat Narayan Maurya
-
Upload
independent -
Category
Documents
-
view
1 -
download
0
Transcript of Modification of chromium (VI) phytotoxicity by exogenous gibberellic acid application in Pisum...

1 23
Acta Physiologiae Plantarum ISSN 0137-5881Volume 33Number 4 Acta Physiol Plant (2011)33:1385-1397DOI 10.1007/s11738-010-0672-x
Modification of chromium (VI)phytotoxicity by exogenous gibberellicacid application in Pisum sativum (L.)seedlings
Savita Gangwar, Vijay Pratap Singh,Prabhat Kumar Srivastava & JagatNarayan Maurya

1 23
Your article is protected by copyright and
all rights are held exclusively by Franciszek
Górski Institute of Plant Physiology, Polish
Academy of Sciences, Kraków. This e-offprint
is for personal use only and shall not be self-
archived in electronic repositories. If you
wish to self-archive your work, please use the
accepted author’s version for posting to your
own website or your institution’s repository.
You may further deposit the accepted author’s
version on a funder’s repository at a funder’s
request, provided it is not made publicly
available until 12 months after publication.

ORIGINAL PAPER
Modification of chromium (VI) phytotoxicity by exogenousgibberellic acid application in Pisum sativum (L.) seedlings
Savita Gangwar • Vijay Pratap Singh •
Prabhat Kumar Srivastava • Jagat Narayan Maurya
Received: 11 August 2010 / Accepted: 7 December 2010 / Published online: 24 December 2010
� Franciszek Gorski Institute of Plant Physiology, Polish Academy of Sciences, Krakow 2010
Abstract Effects of exogenous gibberellic acid (GA; 10
and 100 lM) application on growth, protein and nitrogen
contents, ammonium (NH4?) content, enzymes of nitrogen
assimilation and antioxidant system in pea seedlings were
investigated under chromium (VI) phytotoxicity (Cr VI;
50, 100 and 250 lM). Exposure of pea seedlings to Cr and
100 lM GA resulted in decreased seed germination, fresh
and dry weight and length of root and shoot, and protein
and nitrogen contents compared to control. Compared to
control, Cr and 100 lM GA led to the significant alteration
in nitrogen assimilation in pea. These treatments decreased
root and shoot nitrate reductase (NR), glutamine synthetase
(GS) and glutamine 2-oxoglutarate aminotransferase
(GOGAT) activities (except 50 lM Cr alone for GOGAT)
while glutamate dehydrogenase (GDH) activity and NH4?
content increased. Compared to control, the root and shoot
activities of superoxide dismutase (SOD) and ascorbate
peroxidase (APX) increased (except APX activity at
250 lM Cr ? 100 lM GA) while catalase (CAT), gluta-
thione reductase (GR) and dehydroascorbate reductase
(DHAR) activities were decreased (except GR at 100 lM
GA alone) following exposure of Cr and 100 lM GA.
Total ascorbate and total glutathione in root and shoot
decreased by the treatments of Cr and 100 lM GA while
their levels were increased by the application of 10 lM GA
compared to Cr treatments alone. It has been reported that
application of 10 lM GA together with Cr alleviated
inhibited levels of growth, nitrogen assimilation and anti-
oxidant system compared to Cr treatments alone. This
study showed that application of 10 lM GA counteracts
some of the adverse effects of Cr phytotoxicity with the
increased levels of antioxidants and sustained activities of
enzymes of nitrogen assimilation; however, 100 lM GA
showed apparently reverse effect under Cr phytotoxicity.
Keywords Antioxidants � Chromium (VI) phytotoxicity �Gibberellic acid � Nitrogen assimilation �Pisum sativum (L.)
Abbreviations
APX Ascorbate peroxidase
CAT Catalase
DHAR Dehydroascorbate reductase
GDH Glutamate dehydrogenase
GOGAT Glutamine 2-oxoglutarate aminotransferase
GR Glutathione reductase
GS Glutamine synthetase
NR Nitrate reductase
SOD Superoxide dismutase
Introduction
Chromium (Cr) contamination of soil and water is one of
the most serious environmental threats leading to losses in
crop yield and hazardous effects on health when it enters
the food chain (Vernay et al. 2007). Chromium is
Communicated by G. Klobus.
S. Gangwar (&) � J. N. Maurya
Department of Plant Science, MJP Rohilkhand University,
Bareilly 243006, India
e-mail: [email protected]
V. P. Singh � P. K. Srivastava
Ranjan Plant Physiology and Biochemistry Laboratory,
Department of Botany, University of Allahabad,
Allahabad 211002, India
123
Acta Physiol Plant (2011) 33:1385–1397
DOI 10.1007/s11738-010-0672-x
Author's personal copy

continuously being released in the environment from lea-
ther tanning, steel plating and dyeing industries and thus
has lead to a continuous increase in available Cr in water
and soil (Shanker et al. 2004). Chromium possesses
valence numbers ranging from 0 to 6, of which the trivalent
chromium (Cr, III) and hexavalent chromium (Cr, VI) are
more common in Cr-polluted substrates (Kotas and Stasi-
cka 2000). Hexavalent Cr is highly toxic because of its
ability to cross plasma membrane with powerful oxidizing
capability (Shanker et al. 2004). Chromium (VI) phyto-
toxicity inhibits seed germination, growth and photosyn-
thesis, decreases uptake of nutrient elements, alters water
balance, and affects nitrogen and sulfur metabolism
(Shanker et al. 2004; Vernay et al. 2007; Kumar and Joshi
2008; Schiavon et al. 2008). It has been reported that toxic
property of Cr(VI) originates from the formation of reac-
tive oxygen species (ROS), i.e. superoxide radical (O2�-),
hydrogen peroxide (H2O2) and hydroxyl radical (�OH), and
these ROS present in higher concentrations produce cyto-
toxic effects due to their ability to oxidize lipids, proteins
and nucleic acids (Shanker et al. 2004; Panda 2007; Pandey
et al. 2009b). In order to mitigate deleterious effects of
ROS, plants possess complex defense mechanisms. Such
defense mechanisms involve both enzymatic and non-
enzymatic antioxidants (Panda 2007). The simultaneous
action of various antioxidant enzymes viz. superoxide
dismutase (SOD, EC 1.15.1.1), catalase (CAT, EC
1.11.1.6), glutathione reductase (GR, EC 1.6.4.2), dehy-
droascorbate reductase (DHAR, EC 1.8.5.1), ascorbate
peroxidase (APX, EC 1.11.1.11), etc. is essential for reg-
ulation of ROS levels within cell (Shanker et al. 2004;
Panda 2007). Non-enzymatic antioxidants such as ascor-
bate and glutathione also play important role in preventing
oxidative stress (Noctor and Foyer 1998).
To fulfill the food demand of increasing population,
alternative strategies are needed to alleviate the adverse
effects of metal phytotoxicity on crops. Chemical appli-
cation and agronomical crop management practices have
been used to alleviate the metal phytotoxicity with a little
bit of success. Exogenous application of plant hormones
may be an alternative strategy to induce the capability
within plants to face successfully the detrimental situation
of metal phytotoxicity. Plant hormones are a group of
chemical messengers that regulate plant growth and
development. Gibberellic acid (GA), one of the key plant
hormones influences seed germination, stem elongation,
leaf expansion and reproductive development (Hooley
1994; Matsuoka 2003). Studies have shown that exogenous
application of GA provides protection to plants against
abiotic stresses and increases crop yield (Tuna et al. 2008;
Wen et al. 2010). However, Fuchs and Lieberman (1968),
Mori and Schroeder (2004) and Celik et al. (2007) have
shown that if GA is applied in excess, it results in increased
ethylene production, ROS generation and alteration in
defense mechanisms, respectively, which, in turn, cause
tissue damage and retard growth. Though there have been
many studies which show effects of exogenous application
of GA on plants and animals, however, a lot of contra-
dictory results can be found in the literature. Very few
studies have been carried out to investigate plants respon-
ses to exogenous application of GA during their early stage
of growth under stress conditions. Therefore, we have
undertook this study to investigate the effects of exogenous
GA application on growth, nitrogen assimilation and anti-
oxidant system in Pisum sativum L. seedlings during their
early stage of growth under Cr(VI) phytotoxicity. This
paper is mainly focused on the mechanisms by which GA
influences pea seedling responses under Cr(VI)
phytotoxicity.
Materials and methods
Plant materials and culture conditions
Uniform sized pea seeds (Pisum sativum L. cv. Azad P-1)
were surface sterilized in 10% (v/v) sodium hypochlorite
solution for 5 min. Then they were washed thoroughly and
soaked for 2–4 h in distilled water. The imbibed seeds were
placed in Petri plates (radius 7.3 cm and surface area
167.42 cm2:150 mm RivieraTM). Bottom of each Petri
plate was lined with Whatman No-1 filter paper which was
moistened with either 20 ml of 0.5 strength Hoagland’s
solution only (control) or with 20 ml of each selected
concentration of Cr and GA alone as well as in combina-
tion prepared in 0.5 strength Hoagland’s solution (Arditti
and Dunn 1969). Petri plates were kept in a growth
chamber for seed germination for 3 days under dark con-
ditions (relative humidity of 50–60% at 25 ± 2�C). After
3 days, seedlings of each Petri plate were exposed with a
light irradiance of 150 lmol photons m-2 s-1 for next
8 days in same growth chamber (in which seeds were
germinated) under same humidity and temperature condi-
tions. The photoperiod was 12/12 h (day/night regime)
daily. After 11 days, root and shoot samples from control
and treated seedlings were harvested and different param-
eters were analyzed immediately. In the present study,
following combinations of Cr and GA were made: only 0.5
strength Hoagland’s solution (control), 50 lM Cr, 100 lM
Cr, 250 lM Cr, 10 lM GA, 100 lM GA, 50 lM Cr ?
10 lM GA, 100 lM Cr ? 10 lM GA, 250 lM
Cr ? 10 lM GA, 50 lM Cr ? 100 lM GA, 100 lM
Cr ? 100 lM GA and 250 lM Cr ? 100 lM GA.
Therefore, 12 Petri plates were arranged one for each
combination. Each Petri plate contained 25 healthy and
uniform sized seeds. These selected combinations were
1386 Acta Physiol Plant (2011) 33:1385–1397
123
Author's personal copy

given four times during 11 days growth of pea seedlings at
an interval of 3 days, i.e. first treatment was given during
seed sowing (on the 1st day), second on the 3rd day, third
on the 6th day and fourth on the 9th day in 0.5 strength
Hoagland’s solution. All the Petri plates were kept wet by
the addition of 0.5 strength Hoagland’s solution daily or as
per requirement to avoid limitation of nutrients.
Seed germination and growth parameters
Seed germination (%) was determined by counting the
number of germinated seeds per 25 seeds. Growth param-
eters (fresh and dry weight, and length of root and shoot)
were measured after harvesting control and treated pea
seedlings randomly.
Determination of total protein, total nitrogen and NH4?
contents
Root and shoot total protein was determined by following
the method of Lowry et al. (1951) using bovine serum
albumin as standard. Total nitrogen content of each sample
was quantified using Kjeldahl method (Lang 1958). For the
measurement of root and shoot NH4? content, Nessler’s
reagent method of Molins-Legua et al. (2006) was used.
Assays of nitrogen assimilating enzymes
Nitrate reductase activity was assayed according to the
method of Debouba et al. (2006). One unit of enzyme
activity is defined as 1 nmol NO2- formed/mg protein/h.
glutamine 2-oxoglutarate aminotransferase activity also
called glutamate synthase was measured by the method of
Singh and Srivastava (1986). One unit of enzyme activity is
defined as 1 nmol NADH oxidized/mg protein/min. For
measurement of glutamine synthetase activity the method
of Lillo (1984) was used. One unit of enzyme activity is
defined as 1 nmol c-glutamylhydroxamate formed/mg
protein/min. Glutamate dehydrogenase activity was
assayed by the method of Singh and Srivastava (1983). One
unit of enzyme activity is defined as 1 nmol NADH oxi-
dized/mg protein/min.
Assays of antioxidant enzymes
Fresh root and shoot samples (1 g) from control and treated
seedlings were homogenized in ice cold mortar and pestle
using 10 ml of 50 mM potassium phosphate buffer (pH
7.0) containing 1 mM EDTA and 1% (w/v) polyvinylpyr-
rolidone. In case of APX and DHAR activities, 1 mM
ascorbic acid and 2 mM 2-mercaptoethanol were added
into above buffer, respectively. After centrifugation
(20,000g for 10 min at 4�C), the supernatant was used for
determination of SOD, APX, GR, CAT and DHAR
activities.
Superoxide dismutase activity was assayed by the
nitroblue tetrazolium (NBT) reduction method of Gian-
nopolitis and Reis (1977). One unit of SOD activity is
defined as the amount of enzyme which is required to cause
50% inhibition in the reduction of NBT. Ascorbate per-
oxidase activity was assayed using the method of Nakano
and Asada (1981). One unit of enzyme activity is defined as
1 nmol ascorbate oxidized/mg protein/min. Glutathione
reductase activity was estimated by recording decrease in
absorbance at 340 nm due to the oxidation of NADPH
(Schaedle and Bassham 1977). One unit of enzyme activity
is defined as 1 nmol NADPH oxidized/mg protein/min.
Catalase activity was determined by the method of Aebi
(1984). One unit of enzyme activity is defined as 1 nmol
H2O2 decomposed/mg protein/min. Dehydroascorbate
reductase activity was determined by measuring the
increase in absorbance at 265 nm due to the reduction of
dehydroascorbate (DHA) into reduced ascorbate (AsA)
(Nakano and Asada 1981). One unit of enzyme activity is
defined as 1 nmol DHA reduced/mg protein/min.
Measurements of total ascorbate and total glutathione
levels
Total ascorbate in control and treated seedlings was
determined by the method of Gossett et al. (1994). Total
ascorbate was calculated using standard curve prepared
with L-ascorbic acid. Total glutathione was determined by
the enzyme-recycling method of Brehe and Burch (1976).
The amount of total glutathione was calculated using a
standard curve prepared with reduced glutathione (GSH).
Statistical analysis
All the data were subjected to analysis of variance and
expressed as mean ± SE of three independent experi-
ments. Statistical significance of the means was compared
by the Duncan’s multiple range test at P \ 0.05 signifi-
cance level using SPSS-10 software. In each experiment,
ten seedlings were taken randomly for the measurements of
fresh and dry weight and length of root and shoot while for
other parameters three seedlings were used.
Results
Seed germination, fresh and dry weight, and length of root
and shoot were used to assess the impact of exogenous GA
application on growth of pea seedling under Cr phytotox-
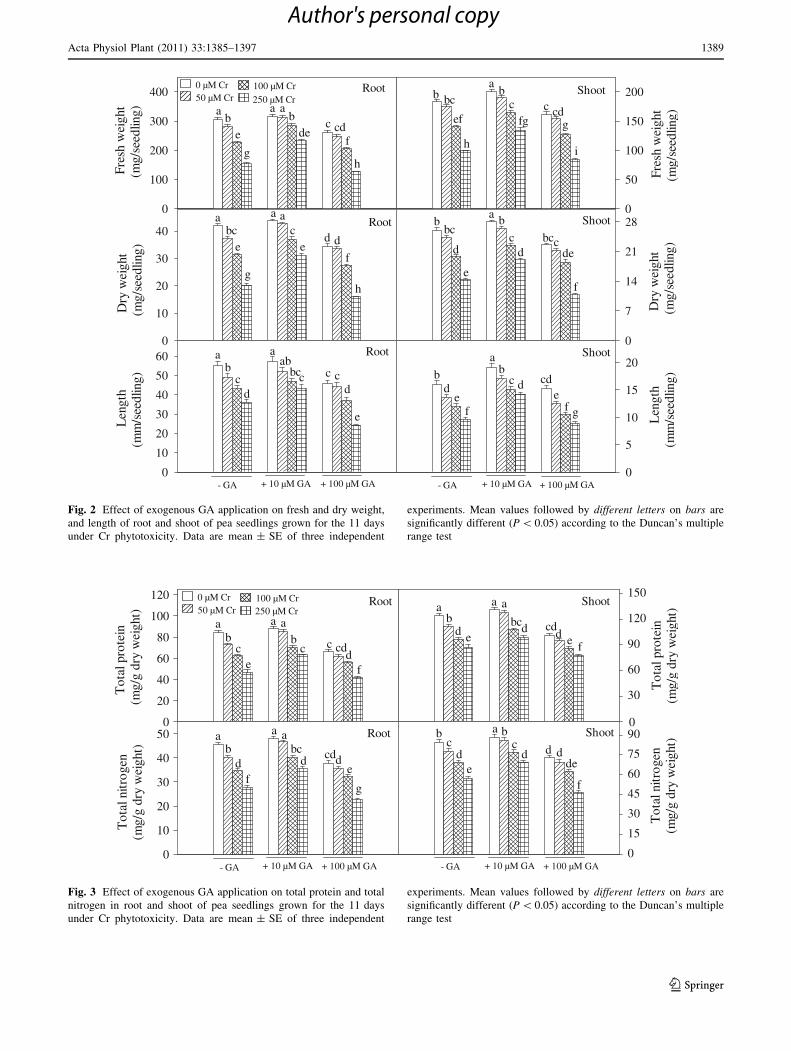
icity. As shown in Figs. 1 and 2, Cr (50, 100 and 250 lM)
and GA (100 lM) decreased seed germination, fresh and
Acta Physiol Plant (2011) 33:1385–1397 1387
123
Author's personal copy

dry weight, and length of root and shoot compared to
control. Moreover, it was noticed that combined applica-
tion of Cr and 100 lM GA further decreased seed germi-
nation and growth of pea seedlings. Maximum decrease in
seed germination and growth parameters was noticed under
the treatment of 250 lM Cr ? 100 lM GA (seed germi-
nation decreased by 44%, root and shoot fresh weight
decreased by 59 and 54%, root and shoot dry weight
decreased by 62 and 58%, and length of root and shoot
decreased by 56 and 45%, respectively, compared to con-
trol). On the contrary, it was reported that addition of
10 lM GA together with Cr increased the germination of
seeds (equal to the control), and alleviated inhibited levels
of growth in pea seedlings compared to Cr treatments alone
(Figs. 1, 2). Moreover, it was observed that 10 lM GA
alone did not significantly influence length of root; how-
ever, length of shoot was significantly increased by this
dose compared to control (Fig. 2).
Results shown in Fig. 3 indicated that exposure of pea
seedlings to Cr and 100 lM GA decreased both total
protein and total nitrogen in root and shoot compared to
control. Exposure of pea seedlings to 250 lM
Cr ? 100 lM GA decreased protein content in root and
shoot by 51 and 38% and nitrogen content by 50 and 45%,
respectively, compared to control. On the contrary, sup-
plementation of 10 lM GA together with Cr increased both
total protein and total nitrogen compared to Cr treatments
alone (Fig. 3).
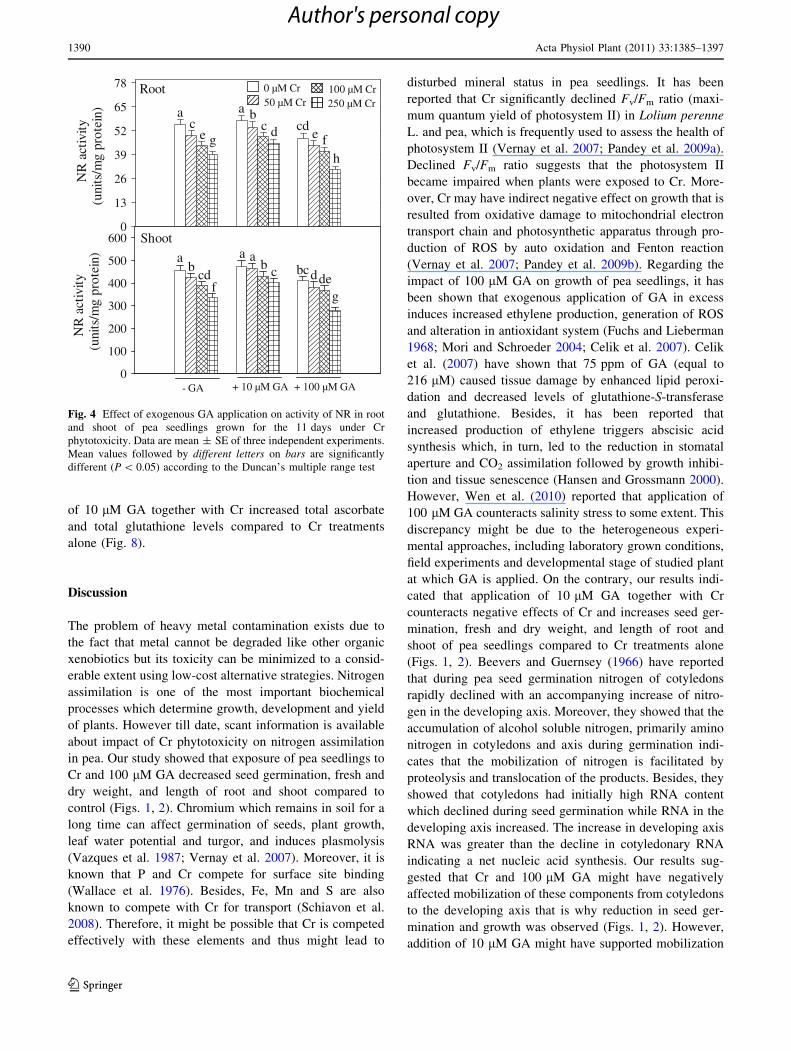
As shown in Fig. 4, treatments of pea seedlings with Cr
and 100 lM GA alone as well as in combination decreased
NR activity compared to control. However, application of
10 lM GA together with Cr increased NR activity com-
pared to Cr treatments alone (Fig. 4). Results pertaining to
the NH4? content are shown in Fig. 5. Exposure of pea
seedlings to single and combined treatments of Cr and
100 lM GA increased NH4? content in root and shoot
compared to control as it was increased by 79 and 74%,
respectively, under the treatment of 250 lM Cr ? 100 lM
GA. However, 10 lM GA alone did not significantly
influence NH4? content in root and shoot compared to
control. Besides, when 10 lM GA was applied together
with Cr, it appreciably lowered NH4? content in root and
shoot compared to Cr treatments alone (Fig. 5). Results
related to the enzymes of NH4? assimilation are depicted
in Fig. 5. Results showed that Cr and GA alone as well as
in combination increased GDH activity in root and shoot
compared to control. On the contrary, Cr and 100 lM GA
alone as well as in combination decreased GOGAT and GS
activities (except 50 lM Cr alone for GOGAT activity) in
root and shoot compared to control. Reduction in GOGAT
and GS activities was higher in roots than shoots. Exposure
of pea seedlings to 250 lM Cr ? 100 lM GA led to a
decrease in root and shoot GOGAT activity by 44 and 35%
and in GS activity by 41 and 37%, respectively, compared
to control. However, supplementation of 10 lM GA
together with Cr either increased GOGAT and GS activi-
ties over control values or restored them over Cr treatments
alone (Fig. 5).
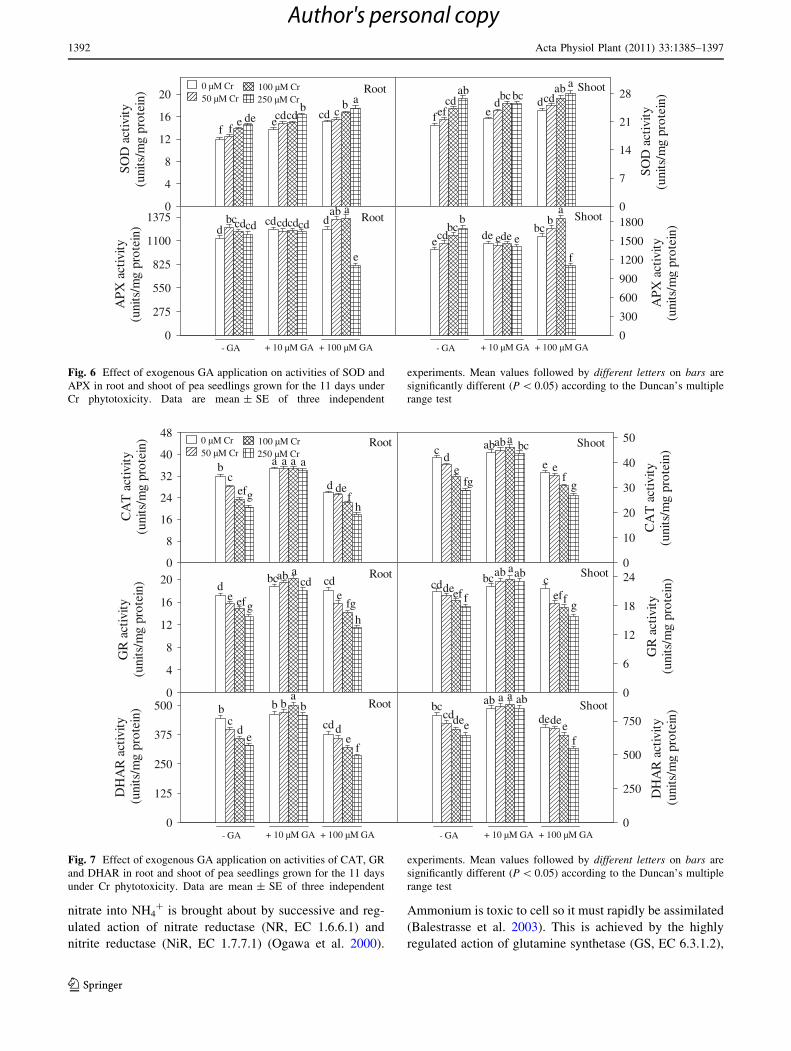
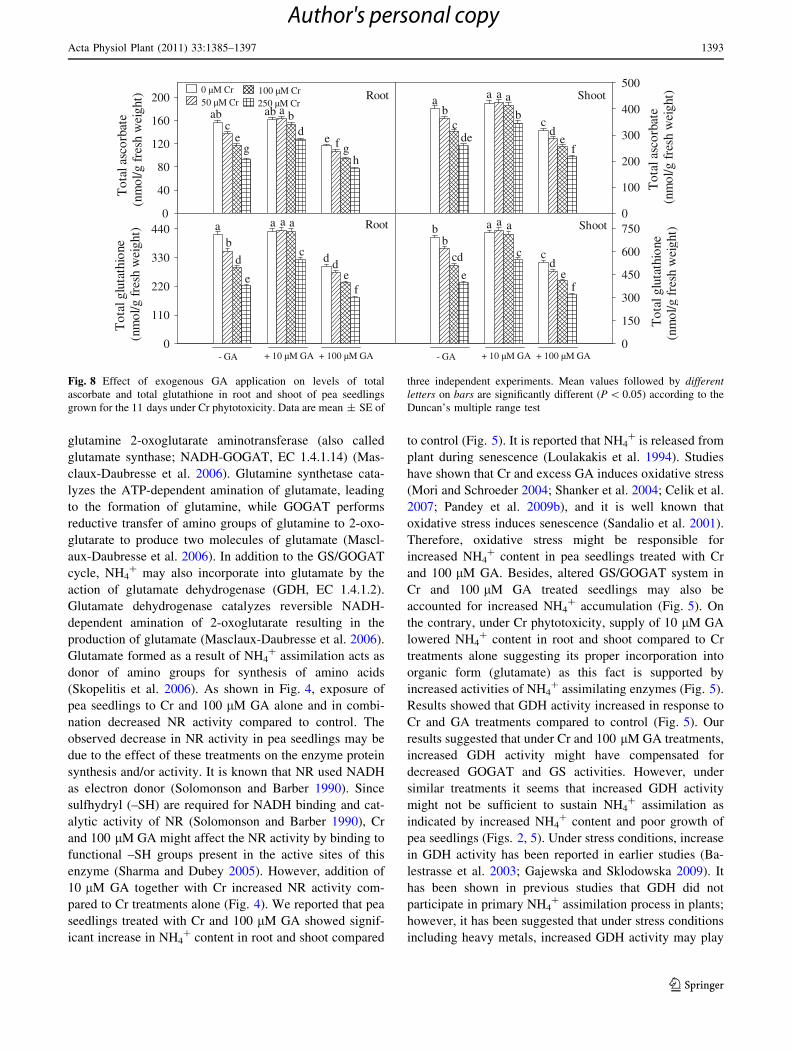
Results for antioxidant enzyme activities are shown in
Figs. 6 and 7. Exposure of pea seedlings to Cr and GA
alone as well as in combination increased SOD and APX
activities (except root and shoot APX activity at 250 lM
Cr ? 100 GA treatment; decreased by 28 and 19%,
respectively) in root and shoot compared to control
(Fig. 6). On the contrary, exposure of pea seedlings to Cr
and 100 lM GA led to the decrease in CAT, GR and
DHAR activities (except GR activity at 100 lM GA alone)
compared to control (Fig. 7). Treatment of pea seedlings
with 250 lM Cr ? 100 lM GA produced decrease in root
and shoot CAT activity by 44 and 36%, in GR activity by
33 and 25% and in DHAR activity by 36 and 31%,
respectively, compared to control. On the contrary, under
Cr phytotoxicity, addition of 10 lM GA increased CAT,
GR and DHAR activities in root and shoot even above their
respective control values (Fig. 7). Though CAT, GR and
DHAR activities did not show much variation in
Cr ? 10 lM GA treatments however, they were signifi-
cantly higher than Cr treatments alone (Fig. 7).
Ascorbate and glutathione were tested due to their
involvement in several useful metabolic processes and their
capability to reduce ROS. As shown in Fig. 8, treatments
of pea seedlings with Cr and 100 lM GA alone as well as
in combination decreased total ascorbate and total gluta-
thione levels in root and shoot compared to control.
However, glutathione was more affected than ascorbate. It
was noticed that exposure of pea seedlings to 250 lM
Cr ? 100 lM GA declined root and shoot ascorbate level
by 51 and 46% and glutathione level by 58 and 54%,
respectively, compared to control. On the contrary, supply
Seed
ger
min
atio
n
(%
)
0
30
60
90
120
150
+ 10 µM GA + 100 µM GA
a a a a aabc
f
c cdef
g
0 µM Cr50 µM Cr
100 µM Cr
250 µM Cr
- GA
Fig. 1 Effect of exogenous GA application on pea seed germination
under Cr phytotoxicity. Data are mean ± SE of three independent
experiments. Mean values followed by different letters on bars are
significantly different (P \ 0.05) according to the Duncan’s multiple
range test
1388 Acta Physiol Plant (2011) 33:1385–1397
123
Author's personal copy
Fre
sh w
eigh
t(m
g/se
edlin
g)
0
100
200
300
400
Fre
sh w
eigh
t(m
g/se
edlin
g)
0
50
100
150
200
Dry
wei
ght
(mg/
seed
ling)
0
10
20
30
40
Dry
wei
ght
(mg/
seed
ling)
0
7
14
21
28
Len
gth
(mm
/see
dlin
g)
0
10
20
30
40
50
60
Len
gth
(mm
/see
dlin
g)
0
5
10
15
20a a
abb bcc cc
d
cd
e
ab
cbd d
e
cde
f f g
Root
Root Shoot
Root Shoot
ab
e
g
a ab
dec cd
f
h
b bc
ef
h
ab
cfg
c cdg
i
abc
e
g
a ac
ed d
f
h
bbc
d
e
a b
cd
bccde
f
0 µM Cr50 µM Cr
100 µM Cr
250 µM Cr
+ 10 µM GA + 100 µM GA + 10 µM GA + 100 µM GA
Shoot
- GA - GA
Fig. 2 Effect of exogenous GA application on fresh and dry weight,
and length of root and shoot of pea seedlings grown for the 11 days
under Cr phytotoxicity. Data are mean ± SE of three independent
experiments. Mean values followed by different letters on bars are
significantly different (P \ 0.05) according to the Duncan’s multiple
range test
T
otal
pro
tein
(m
g/g
dry
wei
ght)
0
20
40
60
80
100
120
T
otal
pro
tein
(m
g/g
dry
wei
ght)
0
30
60
90
120
150
T
otal
nitr
ogen
(m
g/g
dry
wei
ght)
0
10
20
30
40
50
T
otal
nitr
ogen
(m
g/g
dry
wei
ght)
0
15
30
45
60
75
90
Root Shoot
Root Shoot
ab
ce
a ab
c c cdd
f
ab
d e
a a
bcd cdd e f
ab
df
a abc
d cdde
g
bc
de
bc
d d dde
f
0 µM Cr50 µM Cr
100 µM Cr250 µM Cr
+ 10 µM GA + 100 µM GA + 10 µM GA + 100 µM GA
a
- GA- GA
Fig. 3 Effect of exogenous GA application on total protein and total
nitrogen in root and shoot of pea seedlings grown for the 11 days
under Cr phytotoxicity. Data are mean ± SE of three independent
experiments. Mean values followed by different letters on bars are
significantly different (P \ 0.05) according to the Duncan’s multiple
range test
Acta Physiol Plant (2011) 33:1385–1397 1389
123
Author's personal copy
of 10 lM GA together with Cr increased total ascorbate
and total glutathione levels compared to Cr treatments
alone (Fig. 8).
Discussion
The problem of heavy metal contamination exists due to
the fact that metal cannot be degraded like other organic
xenobiotics but its toxicity can be minimized to a consid-
erable extent using low-cost alternative strategies. Nitrogen
assimilation is one of the most important biochemical
processes which determine growth, development and yield
of plants. However till date, scant information is available
about impact of Cr phytotoxicity on nitrogen assimilation
in pea. Our study showed that exposure of pea seedlings to
Cr and 100 lM GA decreased seed germination, fresh and
dry weight, and length of root and shoot compared to
control (Figs. 1, 2). Chromium which remains in soil for a
long time can affect germination of seeds, plant growth,
leaf water potential and turgor, and induces plasmolysis
(Vazques et al. 1987; Vernay et al. 2007). Moreover, it is
known that P and Cr compete for surface site binding
(Wallace et al. 1976). Besides, Fe, Mn and S are also
known to compete with Cr for transport (Schiavon et al.
2008). Therefore, it might be possible that Cr is competed
effectively with these elements and thus might lead to
disturbed mineral status in pea seedlings. It has been
reported that Cr significantly declined Fv/Fm ratio (maxi-
mum quantum yield of photosystem II) in Lolium perenne
L. and pea, which is frequently used to assess the health of
photosystem II (Vernay et al. 2007; Pandey et al. 2009a).
Declined Fv/Fm ratio suggests that the photosystem II
became impaired when plants were exposed to Cr. More-
over, Cr may have indirect negative effect on growth that is
resulted from oxidative damage to mitochondrial electron
transport chain and photosynthetic apparatus through pro-
duction of ROS by auto oxidation and Fenton reaction
(Vernay et al. 2007; Pandey et al. 2009b). Regarding the
impact of 100 lM GA on growth of pea seedlings, it has
been shown that exogenous application of GA in excess
induces increased ethylene production, generation of ROS
and alteration in antioxidant system (Fuchs and Lieberman
1968; Mori and Schroeder 2004; Celik et al. 2007). Celik
et al. (2007) have shown that 75 ppm of GA (equal to
216 lM) caused tissue damage by enhanced lipid peroxi-
dation and decreased levels of glutathione-S-transferase
and glutathione. Besides, it has been reported that
increased production of ethylene triggers abscisic acid
synthesis which, in turn, led to the reduction in stomatal
aperture and CO2 assimilation followed by growth inhibi-
tion and tissue senescence (Hansen and Grossmann 2000).
However, Wen et al. (2010) reported that application of
100 lM GA counteracts salinity stress to some extent. This
discrepancy might be due to the heterogeneous experi-
mental approaches, including laboratory grown conditions,
field experiments and developmental stage of studied plant
at which GA is applied. On the contrary, our results indi-
cated that application of 10 lM GA together with Cr
counteracts negative effects of Cr and increases seed ger-
mination, fresh and dry weight, and length of root and
shoot of pea seedlings compared to Cr treatments alone
(Figs. 1, 2). Beevers and Guernsey (1966) have reported
that during pea seed germination nitrogen of cotyledons
rapidly declined with an accompanying increase of nitro-
gen in the developing axis. Moreover, they showed that the
accumulation of alcohol soluble nitrogen, primarily amino
nitrogen in cotyledons and axis during germination indi-
cates that the mobilization of nitrogen is facilitated by
proteolysis and translocation of the products. Besides, they
showed that cotyledons had initially high RNA content
which declined during seed germination while RNA in the
developing axis increased. The increase in developing axis
RNA was greater than the decline in cotyledonary RNA
indicating a net nucleic acid synthesis. Our results sug-
gested that Cr and 100 lM GA might have negatively
affected mobilization of these components from cotyledons
to the developing axis that is why reduction in seed ger-
mination and growth was observed (Figs. 1, 2). However,
addition of 10 lM GA might have supported mobilization
NR
act
ivity
(u
nits
/mg
prot
ein)
0
13
26
39
52
65
78 Root
ac
e g
a bc d cd
e fh
0 µM Cr50 µM Cr
100 µM Cr250 µM Cr
N
R a
ctiv
ity
(uni
ts/m
g pr
otei
n)
0
100
200
300
400
500
600 Shoot
ab
cdf
a ab c bcdde
g
- GA + 10 µM GA + 100 µM GA
Fig. 4 Effect of exogenous GA application on activity of NR in root
and shoot of pea seedlings grown for the 11 days under Cr
phytotoxicity. Data are mean ± SE of three independent experiments.
Mean values followed by different letters on bars are significantly
different (P \ 0.05) according to the Duncan’s multiple range test
1390 Acta Physiol Plant (2011) 33:1385–1397
123
Author's personal copy

of these components from cotyledons to the developing
axis under Cr phytotoxicity as supported by the data of
seed germination, fresh and dry weight, and length of root
and shoot (Figs. 1, 2).
Protein and nitrogen contents may be considered as
important indicators to assess growth performance of plants
under stress conditions. Chromium and 100 lM GA alone
as well as in combination decreased both total protein and
total nitrogen contents in root and shoot compared to
control (Fig. 3). Their contents were more affected in roots
than shoots. Decrease in protein content could be a con-
sequence of increased protein degradation and/or a
decrease in protein synthesis. Balestrasse et al. (2003) have
shown that under cadmium stress, decrease in protein
content was related with increased protease activity in
soybean. Similar to protein content, decreased level of
nitrogen has also been reported in range of crops under
stress conditions (Shen et al. 1994; Tuna et al. 2008).
Declined level of nitrogen might be due to the decreased
nitrate absorption efficiency of roots as evidenced from
decreased NR activity (Figs. 3, 4). In the present study,
however, addition of 10 lM GA together with Cr increased
both total protein and total nitrogen contents compared to
Cr treatments alone (Fig. 3).
Nitrogen is an essential plant macronutrient. The avail-
ability of nitrogen has a major influence on crop yield.
Nitrate is the common source of nitrogen available to
plants. Once nitrate is being absorbed by plants it may be
reduced in roots, stored in the vacuoles, or transferred to
the shoots before being processed. The conversion of
N
H4+
con
tent
(µm
ol/g
fres
h w
eigh
t)
0
2
4
6
N
H4+
con
tent
(µm
ol/g
fres
h w
eigh
t)
0
1
2
3
4
G
DH
act
ivity
(u
nits
/mg
prot
ein)
0
25
50
75
100
G
DH
act
ivity
(u
nits
/mg
prot
ein)
0
14
28
42
56
70
G
OG
AT
act
ivity
(un
its/m
g pr
otei
n)
0
4
8
12
16
G
OG
AT
act
ivity
(un
its/m
g pr
otei
n)
0
3
6
9a a aa
bc bccde
c d
f
a ab
a a
cbc bc ce d
f
Root Shoot
Root Shoot
Root Shoot
g fd
b
gfgef ed d
bca
g ef
cb
g ef efdcd
b a
gef
cab
fdecd c
dec c
a
gefbc
a
fgefcd bc
efbcb
a
G
S a
ctiv
ity
(uni
ts/m
g pr
otei
n)
0
7
14
21
28
G
S ac
tivity
(u
nits
/mg
prot
ein)
0
30
60
90
120
150toohStooRab a bc c
de de d
f
deeg
a a abc b
cd d d ddee
f
0 µM Cr50 µM Cr
100 µM Cr250 µM Cr
- GA- GA + 10 µM GA + 100 µM GA + 10 µM GA + 100 µM GA
e
Fig. 5 Effect of exogenous GA application on NH4? content, and
activities of GDH, GOGAT and GS in root and shoot of pea seedlings
grown for the 11 days under Cr phytotoxicity. Data are mean ± SE of
three independent experiments. Mean values followed by differentletters on bars are significantly different (P \ 0.05) according to the
Duncan’s multiple range test
Acta Physiol Plant (2011) 33:1385–1397 1391
123
Author's personal copy
nitrate into NH4? is brought about by successive and reg-
ulated action of nitrate reductase (NR, EC 1.6.6.1) and
nitrite reductase (NiR, EC 1.7.7.1) (Ogawa et al. 2000).
Ammonium is toxic to cell so it must rapidly be assimilated
(Balestrasse et al. 2003). This is achieved by the highly
regulated action of glutamine synthetase (GS, EC 6.3.1.2),
S
OD
act
ivity
(u
nits
/mg
prot
ein)
0
4
8
12
16
20
S
OD
act
ivity
(u
nits
/mg
prot
ein)
0
7
14
21
28
A
PX a
ctiv
ity
(uni
ts/m
g pr
otei
n)
0
275
550
825
1100
1375
A
PX a
ctiv
ity
(uni
ts/m
g pr
otei
n)
0
300
600
900
1200
1500
1800
Root Shoot
Root Shoot
f fe de ecdcd
bcd c
b a
fefcd
ab
edbcbc dcd
ab a
dbccdcd cdcdcdcd d
ab a
eecd
bcb
de ede ebc
ba
f
0 µM Cr50 µM Cr
100 µM Cr250 µM Cr
+ 10 µM GA + 100 µM GA + 10 µM GA + 100 µM GA- GA- GA
Fig. 6 Effect of exogenous GA application on activities of SOD and
APX in root and shoot of pea seedlings grown for the 11 days under
Cr phytotoxicity. Data are mean ± SE of three independent
experiments. Mean values followed by different letters on bars are
significantly different (P \ 0.05) according to the Duncan’s multiple
range test
C
AT
act
ivity
(u
nits
/mg
prot
ein)
0
8
16
24
32
40
48
C
AT
act
ivity
(u
nits
/mg
prot
ein)
0
10
20
30
40
50
G
R a
ctiv
ity
(uni
ts/m
g pr
otei
n)
0
4
8
12
16
20
G
R a
ctiv
ity
(uni
ts/m
g pr
otei
n)
0
6
12
18
24
DH
AR
act
ivity
(u
nits
/mg
prot
ein)
0
125
250
375
500
DH
AR
act
ivity
(u
nits
/mg
prot
ein)
0
250
500
750b b b
c
a
d cd
b
ed
ef
ab a abc
cdab
de dedee ef
Root Shoot
Root Shoot
Root Shoot
bc
efg
a a a a
d def
h
cd
efg
ababa bc
e ef
g
de efg
bcab acd cd
efg
h
cddeef f
bcab aabc
eff g
0 µM Cr50 µM Cr
100 µM Cr250 µM Cr
+ 10 µM GA + 100 µM GA- GA + 10 µM GA + 100 µM GA- GA
Fig. 7 Effect of exogenous GA application on activities of CAT, GR
and DHAR in root and shoot of pea seedlings grown for the 11 days
under Cr phytotoxicity. Data are mean ± SE of three independent
experiments. Mean values followed by different letters on bars are
significantly different (P \ 0.05) according to the Duncan’s multiple
range test
1392 Acta Physiol Plant (2011) 33:1385–1397
123
Author's personal copy
glutamine 2-oxoglutarate aminotransferase (also called
glutamate synthase; NADH-GOGAT, EC 1.4.1.14) (Mas-
claux-Daubresse et al. 2006). Glutamine synthetase cata-
lyzes the ATP-dependent amination of glutamate, leading
to the formation of glutamine, while GOGAT performs
reductive transfer of amino groups of glutamine to 2-oxo-
glutarate to produce two molecules of glutamate (Mascl-
aux-Daubresse et al. 2006). In addition to the GS/GOGAT
cycle, NH4? may also incorporate into glutamate by the
action of glutamate dehydrogenase (GDH, EC 1.4.1.2).
Glutamate dehydrogenase catalyzes reversible NADH-
dependent amination of 2-oxoglutarate resulting in the
production of glutamate (Masclaux-Daubresse et al. 2006).
Glutamate formed as a result of NH4? assimilation acts as
donor of amino groups for synthesis of amino acids
(Skopelitis et al. 2006). As shown in Fig. 4, exposure of
pea seedlings to Cr and 100 lM GA alone and in combi-
nation decreased NR activity compared to control. The
observed decrease in NR activity in pea seedlings may be
due to the effect of these treatments on the enzyme protein
synthesis and/or activity. It is known that NR used NADH
as electron donor (Solomonson and Barber 1990). Since
sulfhydryl (–SH) are required for NADH binding and cat-
alytic activity of NR (Solomonson and Barber 1990), Cr
and 100 lM GA might affect the NR activity by binding to
functional –SH groups present in the active sites of this
enzyme (Sharma and Dubey 2005). However, addition of
10 lM GA together with Cr increased NR activity com-
pared to Cr treatments alone (Fig. 4). We reported that pea
seedlings treated with Cr and 100 lM GA showed signif-
icant increase in NH4? content in root and shoot compared
to control (Fig. 5). It is reported that NH4? is released from
plant during senescence (Loulakakis et al. 1994). Studies
have shown that Cr and excess GA induces oxidative stress
(Mori and Schroeder 2004; Shanker et al. 2004; Celik et al.
2007; Pandey et al. 2009b), and it is well known that
oxidative stress induces senescence (Sandalio et al. 2001).
Therefore, oxidative stress might be responsible for
increased NH4? content in pea seedlings treated with Cr
and 100 lM GA. Besides, altered GS/GOGAT system in
Cr and 100 lM GA treated seedlings may also be
accounted for increased NH4? accumulation (Fig. 5). On
the contrary, under Cr phytotoxicity, supply of 10 lM GA
lowered NH4? content in root and shoot compared to Cr
treatments alone suggesting its proper incorporation into
organic form (glutamate) as this fact is supported by
increased activities of NH4? assimilating enzymes (Fig. 5).
Results showed that GDH activity increased in response to
Cr and GA treatments compared to control (Fig. 5). Our
results suggested that under Cr and 100 lM GA treatments,
increased GDH activity might have compensated for
decreased GOGAT and GS activities. However, under
similar treatments it seems that increased GDH activity
might not be sufficient to sustain NH4? assimilation as
indicated by increased NH4? content and poor growth of
pea seedlings (Figs. 2, 5). Under stress conditions, increase
in GDH activity has been reported in earlier studies (Ba-
lestrasse et al. 2003; Gajewska and Sklodowska 2009). It
has been shown in previous studies that GDH did not
participate in primary NH4? assimilation process in plants;
however, it has been suggested that under stress conditions
including heavy metals, increased GDH activity may play
T
otal
asc
orba
te
(nm
ol/g
fre
sh w
eigh
t)
0
40
80
120
160
200
T
otal
asc
orba
te
(nm
ol/g
fres
h w
eigh
t)
0
100
200
300
400
500
T
otal
glu
tath
ione
(n
mol
/g fr
esh
wei
ght)
0
110
220
330
440
T
otal
glu
tath
ione
(n
mol
/g fr
esh
wei
ght)
0
150
300
450
600
750
toohStooR
Root Shoot
abc
eg
ab a bd
e f gh
ab
cde
a a a
bc
de
f
ab
d
e
a a a
c d de
f
bb
cd
e
a a a
c cd
ef
0 µM Cr50 µM Cr
100 µM Cr250 µM Cr
+ 10 µM GA + 100 µM GA- GA + 10 µM GA + 100 µM GA- GA
Fig. 8 Effect of exogenous GA application on levels of total
ascorbate and total glutathione in root and shoot of pea seedlings
grown for the 11 days under Cr phytotoxicity. Data are mean ± SE of
three independent experiments. Mean values followed by differentletters on bars are significantly different (P \ 0.05) according to the
Duncan’s multiple range test
Acta Physiol Plant (2011) 33:1385–1397 1393
123
Author's personal copy

an important in relieving the pressure of accumulating
toxic amount of NH4? and also in the replenishment of
glutamate pool (Srivastava and Singh 1987; Syntichaki
et al. 1996; Skopelitis et al. 2006). Glutamine synthetase
and GOGAT both are considered prime enzymes for NH4?
assimilation. Decrease in their activities (except 50 lM Cr
alone for GOGAT) was observed following single and
combined exposure of Cr and 100 lM GA (Fig. 5). Under
similar treatments, decrease in GOGAT and GS activities
suggests impairment in NH4? assimilation as indicated by
decreased protein and nitrogen contents, and increased
NH4? content (Figs. 3, 5). Reduction in both GOGAT and
GS activities has been attributed to oxidative modifications
of these enzyme proteins (Balestrasse et al. 2006). As Cr
and excess GA both are known to stimulate generation of
ROS which might be related partially to the oxidative
destruction of these enzymes (Mori and Schroeder 2004;
Celik et al. 2007; Kumar and Joshi 2008; Pandey et al.
2009b). However, addition of 10 lM GA together with Cr
increased GOGAT and GS activities in root and shoot
compared to Cr treatments alone indicating proper incor-
poration of NH4? into glutamate (Fig. 5).
Under stress conditions, greater production of ROS
results into oxidative damage. In such conditions, the
combined action of antioxidants is critical in mitigating
ROS-mediated oxidative damage. Superoxide dismutase is
a ubiquitous metalloenzyme in aerobic organisms catalyses
disproportionation of more toxic O2�– leading to the for-
mation of comparatively less toxic H2O2 (Salin 1988).
Ascorbate peroxidase is an important enzyme of ascorbate–
glutathione cycle and decomposes H2O2 into H2O and O2
using ascorbate as electron donor (Asada 1999). It is
known that if cellular level of H2O2 is regulated, it acts as
systemic intracellular signal for induction of defense sys-
tem while its higher and un-regulated level leads to oxi-
dative stress (Hernandez et al. 2004). As shown in Fig. 6,
Cr and GA alone as well as in combination increased SOD
and APX activities in root and shoot (except APX activity
at 250 lM Cr ? 100 lM GA) compared to control.
However, in case of Cr ? 10 lM GA treatments, there was
no much variation in APX activity when compared to
control. Under Cr and 100 lM GA treatments, our results
suggested that increased SOD and APX activities might not
be sufficient to prevent oxidative stress as indicated by
reduced growth of pea seedling (Figs. 2, 6). However,
better growth of pea seedlings grown under Cr ? 10 lM
GA treatments suggested that SOD and APX activities
probably keep ROS under control mitigating oxidative
stress when compared to Cr treatments alone (Figs. 2, 6).
Similarly, Shanker et al. (2004) and Panda (2007) have
reported decreased growth despite increased SOD and APX
activities. Catalase is another key antioxidative enzyme
which decomposes H2O2 (Corpas et al. 1999). Exposure of
pea seedlings to Cr and 100 lM GA produced reduction in
CAT activity in root and shoot compared to control
(Fig. 7). Decrease in CAT activity suggested possible delay
in H2O2 scavenging which, in turn, causes oxidative
damage as judged by reduced growth (Figs. 2, 7). Romero-
Puertas et al. (2002) have reported that under cadmium
stress, decrease in CAT activity was due to the oxidative
stress-mediated modification of this enzyme protein and
subsequently oxidized protein could be the target of spe-
cific peroxisomal proteases. Therefore, Cr- and 100 lM
GA-mediated decrease in CAT activity may be attributed
to its oxidative modification. However, addition of 10 lM
GA together with Cr increased CAT activity compared to
Cr treatments alone indicated its greater efficiency in
removing of H2O2 as reflected by better growth of pea
seedlings grown under Cr ? 10 lM GA treatments when
compared with Cr treatments alone (Figs. 2, 7). Glutathi-
one reductase and DHAR are important enzymes of
ascorbate–glutathione cycle required for regenerating
reduced glutathione (GSH) and reduced ascorbate (AsA),
respectively, and maintaining cellular redox balance
(Noctor and Foyer 1998). We observed that exposure of
pea seedlings to single and combined treatments of Cr and
100 lM GA decreased GR and DHAR activities (except
100 lM GA alone for GR) in root and shoot compared to
control (Fig. 7). Under such treatments, decrease in GR
and DHAR activities suggested dys-functioning of ascor-
bate–glutathione cycle as indicated by decreased levels of
ascorbate and glutathione (Figs. 7, 8). Decrease in GR and
DHAR activities can lead to shift in cellular redox balance
which, in turn, may cause disturbances in ROS scavenging
processes. Decrease in GR activity (a member of flavoen-
zyme family) may be due to the enzyme protein damage as
reported for Brassica sp. under cadmium stress (Nouairi
et al. 2009). It has been reported that DHAR was labile in
the absence of thiols and inactivated at higher H2O2 levels
(Hossain and Asada 1984); therefore, Cr and 100 lM GA
may also cause inactivation in DHAR activity through
oxidative stress (Celik et al. 2007; Pandey et al. 2009b). On
the contrary, application of 10 lM GA together with Cr
increased GR and DHAR activities compared to Cr treat-
ments alone indicating important role of these enzymes in
preventing oxidative stress through enhanced levels of
ascorbate and glutathione (Figs. 7, 8).
Ascorbate and glutathione are two major non-enzymatic
antioxidants involved in scavenging of ROS directly or
indirectly (Noctor and Foyer 1998). It is known that under
stress conditions plants can adjust ascorbate and glutathi-
one levels by modulating the regeneration and biosynthesis
of ascorbate and glutathione. We observed that exposure of
pea seedlings to single and combined treatments of Cr and
100 lM GA decreased total ascorbate and total glutathione
levels in root and shoot compared to control (Fig. 8).
1394 Acta Physiol Plant (2011) 33:1385–1397
123
Author's personal copy

Under similar treatments, decreased ascorbate and gluta-
thione levels would have led to weakening of the antioxi-
dant potential thereby culminating into oxidative damage
as judged by reduced growth of pea seedlings (Figs. 2, 8).
It is reported that ascorbate is used for the hydroxylation of
proline residues during extensive synthesis, and implicated
in root elongation, the cell cycle regulation and cell wall
formation while physiological significance of glutathione
in plants as well as in animals is due to its involvement in
sulfur metabolism and in controlling cellular heavy metal
concentrations (Scheller et al. 1987; Arrigoni 1994; Kerk
and Feldman 1995; Cordoba-Pedregosa et al. 1996; Lap-
partient and Touraine 1996). Recently, Chao et al. (2010)
and Shan and Liang (2010) showed that increased levels of
ascorbate and glutathione protect rice and Agropyron
cristatum from oxidative stress. Therefore, their decreased
levels may cause Cr and 100 lM GA toxicity symptoms in
pea. Decrease in cellular ascorbate and glutathione levels
may primarily be associated with decreased DHAR and GR
activities as reported in the present study when seedlings
were exposed to Cr and 100 lM GA (Figs. 7, 8). More-
over, decrease in ascorbate and glutathione levels may also
be due to their increased oxidation for scavenging of ROS
and/or prevention of their formation (Shanker et al. 2004;
Pandey et al. 2009b).On the contrary, addition of 10 lM
GA together with Cr increased total ascorbate and total
glutathione levels in root and shoot of pea compared to Cr
treatments alone (Fig. 8). Thus, application of 10 lM GA
under Cr phytotoxicity reflects its important role in
inducing expression of ascorbate and glutathione to protect
pea seedlings from oxidative stress. The increased levels of
ascorbate and glutathione in Cr ? 10 lM GA treated pea
seedlings might have led to better antioxidant system to
combat Cr phytotoxicity.
Conclusions
Our results showed that exogenous application of GA (10
and 100 lM) led to different changes in pea seedlings
under Cr phytotoxicity. Our study pointed out that Cr and
100 lM GA alone as well as in combination decreased
growth and alters nitrogen assimilation in pea seedlings
compared to control. Chromium- and 100 lM GA-medi-
ated adverse effects on growth and nitrogen assimilation
may be attributed to decreased levels of antioxidants. On
the contrary, it was reported that application of 10 lM GA
together with Cr was able to alleviate Cr phytotoxicity
appreciably. This 10 lM GA-mediated amelioration of Cr
phytotoxicity may be assigned to the better antioxidant
system and sustained activities of enzymes of nitrogen
assimilation. Furthermore, these data suggest that GA may
play different roles based on its exogenous concentrations
and plant species used under specific developmental and
environmental conditions.
Acknowledgments We wish to thank The Head, Department of
Plant Science, MJP Rohilkhand University, Bareilly, India for pro-
viding necessary laboratory facilities to carry out this work. We are
also thankful to UGC, India for financial support.
References
Aebi II (1984) Catalase in vitro. Methods Enzymol 105:121–126
Arditti J, Dunn A (1969) Environmental plant physiology—experi-
ments in cellular and plant physiology. Holt, Rinehart and
Winston Inc, New York
Arrigoni O (1994) Ascorbate system in plant development. J Bioenerg
Biomemb 26:407–419
Asada K (1999) The water-water cycle in chloroplasts: scavenging of
active oxygen and dissipation of excess photons. Annu Rev Plant
Physiol Plant Mol Biol 50:601–639
Balestrasse KB, Benavides MP, Gallego SM, Tomaro ML (2003)
Effect of cadmium stress on nitrogen metabolism in nodules and
roots of soybean plants. Funct Plant Biol 30:57–64
Balestrasse KB, Gallego SM, Tomaro ML (2006) Oxidation of the
enzymes involved in ammonium assimilation plays an important
role in the cadmium-induced toxicity in soybean plants. Plant
Soil 284:187–194
Beevers L, Guernsey FS (1966) Changes in some nitrogenous
components during the germination of pea seeds. Plant Physiol
41:1455–1458
Brehe JE, Burch HB (1976) Enzymatic assay for glutathione. Anal
Biochem 74:189–197
Celik I, Tuluce Y, Isik I (2007) Evaluation of toxicity of abscisic acid
and gibberellic acid in rats: 50 days drinking water study.
J Enzym Inhib Med Chem 22:219–226
Chao YY, Hong CY, Kao CH (2010) The decline in ascorbic acid
content is associated with cadmium toxicity of rice seedlings.
Plant Physiol Biochem 48:374–381
Cordoba-Pedregosa MC, Gonzalez-Reyes JA, Sanadillas MS, Navas
P, Cordoba F (1996) Role of apoplastic and cell-wall peroxidases
on the stimulation of root elongation by ascorbate. Plant Physiol
112:1119–1125
Corpas F, Palma JM, Sandalio LM, Lopez-Huertas E, Romero-
Puertas MC, Barroso JB (1999) Purification of catalase from pea
leaf peroxisomes: identification of five different isoforms. Free
Rad Res 31:235–241
Debouba M, Gouia H, Suzuki A, Ghorbel MH (2006) NaCl stress
effects on enzymes involved in nitrogen assimilation pathway in
tomato ‘‘Lycopersicon esculentum’’ seedlings. J Plant Physiol
163:1247–1258
Fuchs Y, Lieberman M (1968) Effects of kinetin, IAA and gibberellin
on ethylene production and their interactions in growth of
seedlings. Plant Physiol 43:2029–2036
Gajewska E, Sklodowska M (2009) Nickel-induced changes in
nitrogen metabolism in wheat shoots. J Plant Physiol
166:1034–1044
Giannopolitis CN, Reis SK (1977) Superoxide dismutase. I. occur-
rence in higher plants. Plant Physiol 59:309–314
Gossett DR, Millhollon EP, Cran LM (1994) Antioxidant response to
NaCl stress in salt-sensitive cultivars of cotton. Crop Sci
34:706–714
Hansen H, Grossmann K (2000) Auxin-induced ethylene triggers
abscisic acid biosynthesis and growth inhibition. Plant Physiol
124:1437–1448
Acta Physiol Plant (2011) 33:1385–1397 1395
123
Author's personal copy

Hernandez JA, Escobar C, Creissen G, Mullineaux PM (2004) Role of
hydrogen peroxide and the redox state of ascorbate in the
induction of antioxidant enzymes in pea leaves under excess
light stress. Funct Plant Biol 31:359–368
Hooley R (1994) Gibberellins: perception, transduction responses.
Plant Mol Biol 26:1529–1555
Hossain MA, Asada K (1984) Purification of dehydroascorbate
reductase from spinach and its characterization as thiol enzyme.
Plant Cell Physiol 25:85–92
Kerk NM, Feldman LJ (1995) A biochemical model for initiation and
maintenance of the quiescent center: implications for organiza-
tion of root meristems. Plant Dev 121:2825–2833
Kotas J, Stasicka Z (2000) Chromium occurrence in the environment
and methods of its speciation. Environ Pollut 107:263–283
Kumar S, Joshi UN (2008) Nitrogen metabolism as affected by
hexavalent chromium in sorghum (Sorghum bicolor L.). Environ
Exp Bot 64:135–144
Lang CA (1958) Simple microdetermination of Kjeldahl nitrogen in
biological materials. Anal Chem 30:1692–1694
Lappartient AG, Touraine B (1996) Demand-driven control of root
ATP sulfurylase activity and sulphate uptake in intact Canola.
Plant Physiol 111:147–157
Lillo C (1984) Diurnal variations of nitrite reductase, glutamine
synthetase, glutamate synthase, alanine amino transferase and
aspartate amino transferase in barley leaves. Physiol Plant
61:214–218
Loulakakis KA, Roubelakis-Angelakis KA, Kanellis AK (1994)
Regulation of glutamate dehydrogenase and glutamine synthe-
tase in avocado fruit during development and ripening. Plant
Physiol 106:217–222
Lowry OH, Rosebrough NJ, Farr AL, Randall RJ (1951) Protein
measurement with the folin phenol reagent. J Biol Chem
193:265–275
Masclaux-Daubresse C, Reisdorf-Cren M, Pageau K, Lelandias M,
Grandjean J, Valadier MH, Feraud M, Jouglet T, Suzuki A
(2006) Glutamine synthetase-glutamate synthase pathway and
glutamate dehydrogenase play distinct roles in the sink source
nitrogen cycle in tobacco. Plant Physiol 140:444–456
Matsuoka M (2003) Gibberellin signaling: how do plant cells respond
to GA signals? J Plant Growth Regul 22:123–125
Molins-Legua C, Meseguer-Lloret S, Moliner-Martinez Y, Campıns-
Falco P (2006) A guide for selecting the most appropriate
method for ammonium determination in water analysis. Trends
Anal Chem 25:282–290
Mori IC, Schroeder JI (2004) Reactive oxygen species activation of
plant Ca2? channels. a signaling mechanism in polar growth,
hormone transduction, stress signaling and hypothetically mech-
anotransduction. Plant Physiol 135:702–708
Nakano Y, Asada K (1981) Hydrogen peroxide is scavenged by
ascorbate specific peroxidase in spinach chloroplasts. Plant Cell
Physiol 22:867–880
Noctor G, Foyer CH (1998) Ascorbate and glutathione: keeping
active oxygen under control. Annu Rev Plant Physiol Plant Mol
Biol 49:249–279
Nouairi I, Ammar WB, Youssef NB, Miled DDB, Ghorbal MB,
Zarrouk M (2009) Antioxidant defense system in leaves of
Indian mustard (Brassica juncea) and rape (Brassica napus)
under cadmium stress. Acta Physiol Plant 31:237–247
Ogawa K, Soutome R, Hiroyama K, Hagio T, Ida S, Nakagawa H
(2000) Co-regulation of nitrate reductase and nitrite reductase in
cultured spinach cells. J Plant Physiol 157:299–306
Panda SK (2007) Chromium-mediated oxidative stress and ultra-
structural changes in root cells of developing rice seedlings.
J Plant Physiol 164:1419–1428
Pandey V, Dixit V, Shyam R (2009a) Chromium (VI) induced
changes in growth and root plasma membrane redox activities in
pea plants. Protoplasma 235:49–55
Pandey V, Dixit V, Shyam R (2009b) Chromium effect on ROS
generation and detoxification in pea (Pisum sativum) leaf
chloroplasts. Protoplasma 236:85–95
Romero-Puertas M, Palma JM, Gomez M, del Rıo LA (2002)
Cadmium causes the oxidative modification of proteins in pea
plants. Plant Cell Environ 25:677–686
Salin ML (1988) Toxic oxygen species and protective systems of the
chloroplast. Physiol Plant 72:681–689
Sandalio LM, Dalurzo HC, Gomez M, Romero-Puertas MC, del Rıo
LA (2001) Cadmium-induced changes in the growth and
oxidative metabolism of pea plants. J Exp Bot 52:2115–2126
Schaedle M, Bassham JA (1977) Chloroplast glutathione reductase.
Plant Physiol 59:1011–1012
Scheller HV, Huang B, Hatch E, Goldsbrough PB (1987) Phytochel-
atin synthesis and glutathione levels in response to heavy metals
in tomato cells. Plant Physiol 85:1031–1035
Schiavon M, Pilon-Smits EAH, Wirtz M, Hell R, Malagoli M (2008)
Interaction between chromium and sulfur metabolism in Bras-sica juncea. J Environ Qual 37:1536–1545
Shan C, Liang Z (2010) Jasmonic acid regulates ascorbate and
glutathione metabolism in Agropyron cristatum leaves under
water stress. Plant Sci 178:130–139
Shanker AK, Djanaguiraman M, Sudhagar R, Chandrashekar CN,
Pathmanabhan G (2004) Differential antioxidative response of
ascorbate glutathione pathway enzymes and metabolites to
chromium speciation stress in green gram (Vigna radiata (L.)
R. Wilczek. Cv CO 4) roots. Plant Sci 166:1035–1043
Sharma P, Dubey RS (2005) Modulation of nitrate reductase activity
in rice seedlings under aluminium toxicity and water stress:
role of osmolytes as enzyme protectant. J Plant Physiol
162:854–864
Shen Z, Shen Q, Liang Y, Liu Y (1994) Effect of nitrogen on the
growth and photosynthetic activity of salt-stressed barley. J Plant
Nutr 17:787–789
Singh RP, Srivastava HS (1983) Regulation of glutamate dehydro-
genase activity by amino acids in maize seedlings. Physiol Plant
57:549–554
Singh RP, Srivastava HS (1986) Increase in glutamate synthase
(NADH) activity in maize seedlings in response to nitrate and
ammonium nitrogen. Physiol Plant 66:413–416
Skopelitis DS, Paranychianakis NV, Paschalidis KA (2006) Abiotic
stress generates ROS that signal expression of anionic glutamate
dehydrogenases to form glutamate for proline synthesis in
tobacco and grapevine. Plant Cell 18:2767–2781
Solomonson LP, Barber MJ (1990) Assimilatory nitrate reductase;
functional properties and regulation. Annu Rev Plant Physiol
Plant Mol Biol 41:225–253
Srivastava HS, Singh RP (1987) Role and regulation of L-glutamate
dehydrogenase during defence response in maize. Phytochem-
istry 26:597–610
Syntichaki KM, Loulakakis KA, Roubelaki-Angelakis KA (1996) The
amino acid sequence similarity of plant glutamate dehydroge-
nase to the extremophilic archaeal enzyme conforms to its stress-
related function. Gene 168:87–92
Tuna AL, Kaya C, Dikilitas M, Higgs D (2008) The combined effects
of gibberellic acid and salinity on some antioxidant enzyme
activities, plant growth parameters and nutritional status in
maize plants. Environ Exp Bot 62:1–9
Vazques MD, Poschenrieder C, Barcelo J (1987) Chromium VI
induced structural and ultrastructural changes in bush bean
plants (Phaseolus vulgaris L.). Ann Bot (Lond) 59:427–438
1396 Acta Physiol Plant (2011) 33:1385–1397
123
Author's personal copy

Vernay P, Gauthier-Moussard C, Hitmi A (2007) Interaction of
bioaccumulation of heavy metal chromium with water relation,
mineral nutrition and photosynthesis in developed leaves of
Lolium perenne L. Chemosphere 68:1563–1575
Wallace A, Soufi SM, Cha JW, Romney EM (1976) Some effects of
chromium toxicity on bush bean plants grown in soil. Plant Soil
44:471–473
Wen F, Zhang Z, Bai T, Xu Q, Pan Y (2010) Proteomics reveals the
effects of gibberellic acid (GA3) on salt-stressed rice (Oryzasativa L.) shoots. Plant Sci 178:170–175
Acta Physiol Plant (2011) 33:1385–1397 1397
123
Author's personal copy




















