
Microbial diversity of biological filters in recirculating aquaculture systems
8
Available online at www.sciencedirect.com Microbial diversity of biological filters in recirculating aquaculture systems Harold J Schreier 1,2 , Natella Mirzoyan 1 and Keiko Saito 1 Development of environmentally sustainable farming of marine and freshwater species using recirculating aquaculture systems (RASs) requires a complete understanding of the biological component involved in wastewater treatment. This component integrates biofilters composed of microbial communities whose structure, dynamics, and activities are responsible for system success. Engineering highly efficient, environmentally sound, disease-free, and economically viable systems necessitates a thorough knowledge of microbial processes involved in all facets of RAS biofilters and has only recently been the focus of comprehensive studies. These studies have included the application of molecular tools to characterize community diversity and have identified key processes useful for improving system performance. In this paper we summarize the current understanding of the microbial diversity and physiology of RAS biofilters and discuss directions for future studies. Addresses 1 Center of Marine Biotechnology, University of Maryland Biotechnology Institute, 701 E. Pratt St., Baltimore, MD 21202, USA 2 Department of Biological Sciences, University of Maryland Baltimore County, 1000 Hilltop Circle, Baltimore, MD 21250, USA Corresponding author: Schreier, Harold J ([email protected]) Current Opinion in Biotechnology 2010, 21:318–325 This review comes from a themed issue on Evironmental biotechnology Edited by Sharon Borglin and John van der Meer Available online 6th April 2010 0958-1669/$ – see front matter # 2010 Elsevier Ltd. All rights reserved. DOI 10.1016/j.copbio.2010.03.011 Introduction Over the past thirty years aquaculture has evolved from small-scale commerce to farming at a large-scale capacity, accounting for about 6% of fish available for human consumption in the 1970s to 47% in 2006 [1 ]. As popu- lation and per capita fish consumption increase, this growth is expected to continue. The aquaculture industry has been challenged to develop economically viable systems that not only produce species at high density but also must contend with limitations because of location, water availability and environmental impacts, among others [2–4]. By its nature recirculating aquacul- ture system (RAS) technology promises to be effective in overcoming these limitations and can provide sustainable farming of freshwater and marine fish. RASs effectively manage, collect and treat wastes that accumulate during fish growth and, under optimal conditions, do not require water replacement except to account for losses because of evaporation [4,5 ,6]. Developing efficient, productive, biologically secure, and disease-free RASs requires a thorough understanding of all life support processes from mechanical (oxygen, temperature, ozonation, UV, pH, and salinity) to the biological filtration systems. While mechanical processes can be monitored and controlled, biological filtration systems rely on the interaction of microbial communities with each other and their environment as a consequence of nutrient input (fish waste output) and, as such, are not easily controlled. Until approximately ten years ago, little was known about the microbial composition of these filters, which are often established as highly organized biofilm structures, and development of new technologies based on resident activities could not be done. Early studies focused on culturing and enrichment techniques, which are biased towards the <1% of microbial species able to grow under standard lab conditions [7]. Recent studies using molecular tools not only have allowed for evaluating microbial diversity but also have provided some insight into their activities that should lead to an understanding of community interactions. These approaches are certain to provide novel RAS biofilter arrangements as well as insight into new processes and tools to enhance and monitor these systems. In this paper we discuss RAS biofilter microbial diversity and processes and highlight areas for future consideration. Because of page limitations we focus on bacterial and archaeal com- munities and will not address eukaryotic microorganisms (rotifers, algae, fungi, etc.) that have complex roles in filtration as well as feed sources, for example, as com- ponents of biofloc or greenwater technologies. RAS biofilters Biofilter assemblies that reduce or eliminate harmful metabolic waste products have been tailored for specific fish species although generic systems have been devel- oped [4,5 ,6,8]. In general, biofilters are compartments that include a fixed medium for microbial attachment and growth, that is, via biofilms, or allow microbial growth to be suspended [9,10]. Regardless of application, for example, marine, freshwater, small aquaria, or large pro- duction systems, biofilters will integrate aerobic and anaerobic microbial processes for elimination of waste products — nitrogen in the form of ammonia excreted by Current Opinion in Biotechnology 2010, 21:318–325 www.sciencedirect.com
Transcript of Microbial diversity of biological filters in recirculating aquaculture systems

Available online at www.sciencedirect.com
Microbial diversity of biological filters in recirculatingaquaculture systemsHarold J Schreier1,2, Natella Mirzoyan1 and Keiko Saito1
Development of environmentally sustainable farming of marine
and freshwater species using recirculating aquaculture
systems (RASs) requires a complete understanding of the
biological component involved in wastewater treatment. This
component integrates biofilters composed of microbial
communities whose structure, dynamics, and activities are
responsible for system success. Engineering highly efficient,
environmentally sound, disease-free, and economically viable
systems necessitates a thorough knowledge of microbial
processes involved in all facets of RAS biofilters and has only
recently been the focus of comprehensive studies. These
studies have included the application of molecular tools to
characterize community diversity and have identified key
processes useful for improving system performance. In this
paper we summarize the current understanding of themicrobial
diversity and physiology of RAS biofilters and discuss
directions for future studies.
Addresses1Center of Marine Biotechnology, University of Maryland BiotechnologyInstitute, 701 E. Pratt St., Baltimore, MD 21202, USA2Department of Biological Sciences, University of Maryland BaltimoreCounty, 1000 Hilltop Circle, Baltimore, MD 21250, USA
Corresponding author: Schreier, Harold J ([email protected])
Current Opinion in Biotechnology 2010, 21:318–325
This review comes from a themed issue onEvironmental biotechnologyEdited by Sharon Borglin and John van der Meer
Available online 6th April 2010
0958-1669/$ – see front matter# 2010 Elsevier Ltd. All rights reserved.
DOI 10.1016/j.copbio.2010.03.011
IntroductionOver the past thirty years aquaculture has evolved fromsmall-scale commerce to farming at a large-scale capacity,accounting for about 6% of fish available for humanconsumption in the 1970s to 47% in 2006 [1!]. As popu-lation and per capita fish consumption increase, thisgrowth is expected to continue. The aquaculture industryhas been challenged to develop economically viablesystems that not only produce species at high densitybut also must contend with limitations because oflocation, water availability and environmental impacts,among others [2–4]. By its nature recirculating aquacul-ture system (RAS) technology promises to be effective inovercoming these limitations and can provide sustainable
farming of freshwater and marine fish. RASs effectivelymanage, collect and treat wastes that accumulate duringfish growth and, under optimal conditions, do not requirewater replacement except to account for losses because ofevaporation [4,5!!,6].
Developing efficient, productive, biologically secure, anddisease-free RASs requires a thorough understanding ofall life support processes from mechanical (oxygen,temperature, ozonation, UV, pH, and salinity) to thebiological filtration systems. While mechanical processescan be monitored and controlled, biological filtrationsystems rely on the interaction of microbial communitieswith each other and their environment as a consequenceof nutrient input (fish waste output) and, as such, are noteasily controlled. Until approximately ten years ago, littlewas known about the microbial composition of thesefilters, which are often established as highly organizedbiofilm structures, and development of new technologiesbased on resident activities could not be done. Earlystudies focused on culturing and enrichment techniques,which are biased towards the <1% of microbial speciesable to grow under standard lab conditions [7]. Recentstudies using molecular tools not only have allowed forevaluating microbial diversity but also have providedsome insight into their activities that should lead to anunderstanding of community interactions. Theseapproaches are certain to provide novel RAS biofilterarrangements as well as insight into new processes andtools to enhance and monitor these systems. In this paperwe discuss RAS biofilter microbial diversity and processesand highlight areas for future consideration. Because ofpage limitations we focus on bacterial and archaeal com-munities and will not address eukaryotic microorganisms(rotifers, algae, fungi, etc.) that have complex roles infiltration as well as feed sources, for example, as com-ponents of biofloc or greenwater technologies.
RAS biofiltersBiofilter assemblies that reduce or eliminate harmfulmetabolic waste products have been tailored for specificfish species although generic systems have been devel-oped [4,5!!,6,8]. In general, biofilters are compartmentsthat include a fixed medium for microbial attachment andgrowth, that is, via biofilms, or allow microbial growth tobe suspended [9,10]. Regardless of application, forexample, marine, freshwater, small aquaria, or large pro-duction systems, biofilters will integrate aerobic andanaerobic microbial processes for elimination of wasteproducts — nitrogen in the form of ammonia excreted by
Current Opinion in Biotechnology 2010, 21:318–325 www.sciencedirect.com

fish, and carbon and nitrogen accumulated from uneatenfeed and fecal matter [9–13]. The major reactions associ-ated with RAS biofilters are summarized in Table 1.
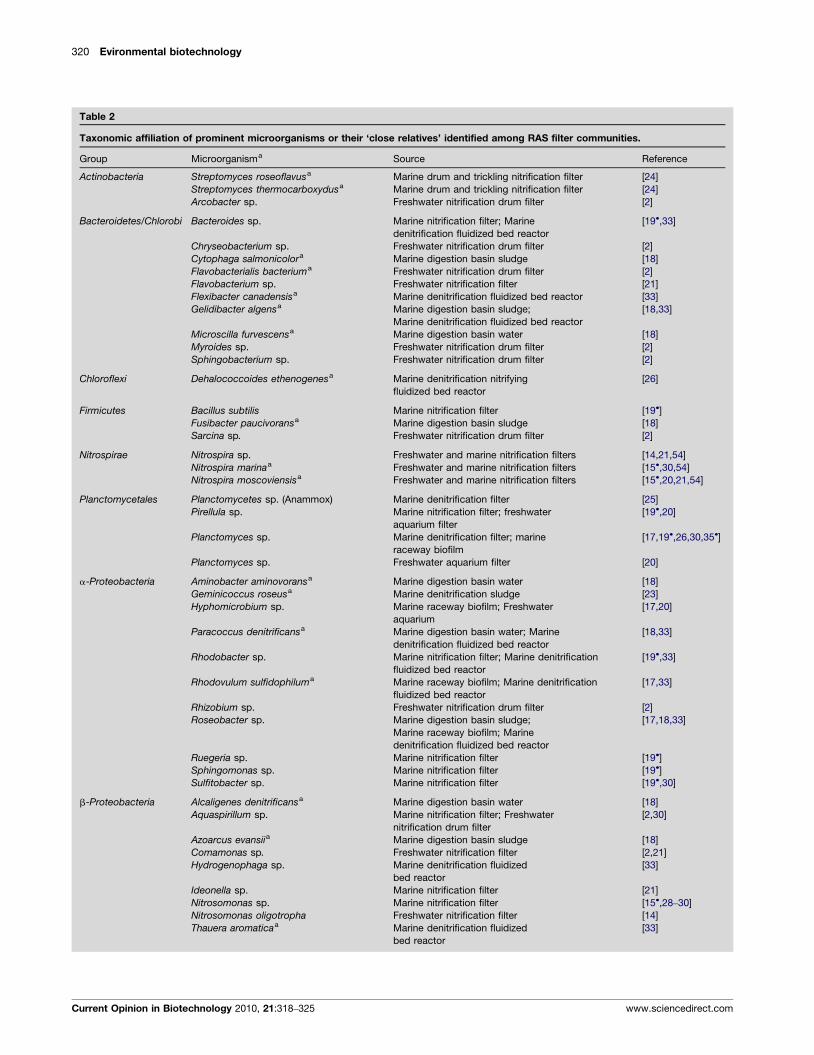
Microbial community diversityCurrent understanding of RAS biofilter microbial diver-sity in both freshwater and marine systems is based onstudies using 16S rRNA and functional gene-specificprobes or 16S rRNA gene libraries rather than culture-based techniques. Biofilter bacteria include members ofActinobacteria, Bacterioidetes/Chlorobi, Firmicutes, Nitros-pirae, Planctomycetales, and Proteobacteria (Table 2). Chlor-oflexi and Synergistetes have also been found in a few cases(Table 2). Recent studies have also suggested involve-ment of Archaea [14,15!,16].
Significant differences in microbial communities havebeen found in RAS biofilters, culture water, and commu-nities representing unique and complex environments[17,18,19!]. Because every fish species introduces itsown unique microbial flora, filter diversity varies from
RAS to RAS [20]. Differences in communities also reflectutilization of UV disinfection and ozonation treatments,which play significant roles in perturbing these commu-nities [17] but appear to have little effect in deep layers offilter biofilms [16,19!].
As RAS culture water is mixotrophic, biofilms develop viathe segregation of individual community members intodistinct layers, which change according to nutritionalconditions, regardless of biofilter compartment. Fluor-escence in situ hybridization (FISH) using species-specific 16S rRNA gene probes has provided details ofthe development and composition of RAS biofilm layers.Weitz et al. [17] monitored biofilm formation in a marineRAS and found g-Proteobacteria to be the most abundantgroup followed by a-Proteobacteria and b-Proteobacteria; b-Proteobacteria, Bacteroides, and Planctomycetales wereamong the first to settle. Similar results were obtainedby Itoi et al. [21], who examined 16S rRNA gene clonelibraries made from outer layers of freshwater aquariumfilters, with Nitrospira sp. restricted to outside layers.
Microbial diversity of aquaculture biofilters Schreier, Mirzoyan and Saito 319
Table 1
Primary activities associated with RAS biofiltration units and participating microorganisms.
Process Reaction Microorganisma
Freshwater Marine
NitrificationAmmonium oxidation NH4
+ + 1.5O2 ! NO2" + 2H+ + H2O Nitrosomonas oligotropha [14] Nitrosomonas sp. [15!,29]
Nitrosomonas cryotolerans [15!,30]Nitrosomonas europaea [23,28]Nitrosomonas cinnybus/nitrosa [15!]Nitrosococcus mobilis [15!]
Nitrite oxidation NO2" + H2O! NO3
" + 2H+ + 2e" Nitrospira spp. [14,21,54]Nitrospira marinaa [54] Nitrospira marinaa [15!,30]Nitrospira moscoviensisa
[20,21,54]Nitrospira moscoviensisa [15!]
DenitrificationAutotrophic(sulfide-dependent)
S2" + 1.6NO3" + 1.6H+ !
SO42" + 0.8N2(g) + 0.8H2O
Thiomicrosporia denitrificans [33]Thiothrix disciformisa [33]Rhodobacter litoralisa [26]Hydrogenophaga sp. [26,33]
Heterotrophic 5CH3COO" + 8NO3" + 3H+ !
10HCO3" + 4N2(g) + 4H2O
Pseudomonas fluorescens [22]Pseudomonas sp. Pseudomonas stutzeri [19!]Comamonas sp. [21] Pseudomonas sp. [30,32]
Paracoccus denitrificans [18]
Dissimilatory nitratereduction to ammonia(DNRA)
NO3" + 2H+ + 4H2 ! NH4
+ + 3H2O Various Proteobacteria and Firmicutes
Anaerobic ammoniumoxidation (Anammox)
NH4+ + NO2
" ! N2(g) + 2H2O Planctomycetes spp. [35!]Brocadia sp.a [25]
Sulfate reduction SO42" + CH3COO" + 3H+ !
HS" + 2HCO3" + 3H+
Desulfovibrio sp.,Dethiosulfovibrio sp.,Fusibacter sp., Bacteroides sp. [18]
Sulfide oxidation HS" + 2O2 ! SO42" + H+ Thiomicrospira sp. [26]
Methanogenesis 4H2 + H+ + HCO3" ! CH4(g) + 3H2O Methanogenic Archaea [Mirzoyan and
Gross, unpublished]
a Microorganisms identified solely on the basis of partial 16S rRNA gene or functional gene sequences.
www.sciencedirect.com Current Opinion in Biotechnology 2010, 21:318–325
320 Evironmental biotechnology
Table 2
Taxonomic affiliation of prominent microorganisms or their ‘close relatives’ identified among RAS filter communities.
Group Microorganisma Source Reference
Actinobacteria Streptomyces roseoflavusa Marine drum and trickling nitrification filter [24]Streptomyces thermocarboxydusa Marine drum and trickling nitrification filter [24]Arcobacter sp. Freshwater nitrification drum filter [2]
Bacteroidetes/Chlorobi Bacteroides sp. Marine nitrification filter; Marinedenitrification fluidized bed reactor
[19!,33]
Chryseobacterium sp. Freshwater nitrification drum filter [2]Cytophaga salmonicolora Marine digestion basin sludge [18]Flavobacterialis bacteriuma Freshwater nitrification drum filter [2]Flavobacterium sp. Freshwater nitrification filter [21]Flexibacter canadensisa Marine denitrification fluidized bed reactor [33]Gelidibacter algensa Marine digestion basin sludge;
Marine denitrification fluidized bed reactor[18,33]
Microscilla furvescensa Marine digestion basin water [18]Myroides sp. Freshwater nitrification drum filter [2]Sphingobacterium sp. Freshwater nitrification drum filter [2]
Chloroflexi Dehalococcoides ethenogenesa Marine denitrification nitrifyingfluidized bed reactor
[26]
Firmicutes Bacillus subtilis Marine nitrification filter [19!]Fusibacter paucivoransa Marine digestion basin sludge [18]Sarcina sp. Freshwater nitrification drum filter [2]
Nitrospirae Nitrospira sp. Freshwater and marine nitrification filters [14,21,54]Nitrospira marinaa Freshwater and marine nitrification filters [15!,30,54]Nitrospira moscoviensisa Freshwater and marine nitrification filters [15!,20,21,54]
Planctomycetales Planctomycetes sp. (Anammox) Marine denitrification filter [25]Pirellula sp. Marine nitrification filter; freshwater
aquarium filter[19!,20]
Planctomyces sp. Marine denitrification filter; marineraceway biofilm
[17,19!,26,30,35!]
Planctomyces sp. Freshwater aquarium filter [20]
a-Proteobacteria Aminobacter aminovoransa Marine digestion basin water [18]Geminicoccus roseusa Marine denitrification sludge [23]Hyphomicrobium sp. Marine raceway biofilm; Freshwater
aquarium[17,20]
Paracoccus denitrificansa Marine digestion basin water; Marinedenitrification fluidized bed reactor
[18,33]
Rhodobacter sp. Marine nitrification filter; Marine denitrificationfluidized bed reactor
[19!,33]
Rhodovulum sulfidophiluma Marine raceway biofilm; Marine denitrificationfluidized bed reactor
[17,33]
Rhizobium sp. Freshwater nitrification drum filter [2]Roseobacter sp. Marine digestion basin sludge;
Marine raceway biofilm; Marinedenitrification fluidized bed reactor
[17,18,33]
Ruegeria sp. Marine nitrification filter [19!]Sphingomonas sp. Marine nitrification filter [19!]Sulfitobacter sp. Marine nitrification filter [19!,30]
b-Proteobacteria Alcaligenes denitrificansa Marine digestion basin water [18]Aquaspirillum sp. Marine nitrification filter; Freshwater
nitrification drum filter[2,30]
Azoarcus evansiia Marine digestion basin sludge [18]Comamonas sp. Freshwater nitrification filter [2,21]Hydrogenophaga sp. Marine denitrification fluidized
bed reactor[33]
Ideonella sp. Marine nitrification filter [21]Nitrosomonas sp. Marine nitrification filter [15!,28–30]Nitrosomonas oligotropha Freshwater nitrification filter [14]Thauera aromaticaa Marine denitrification fluidized
bed reactor[33]
Current Opinion in Biotechnology 2010, 21:318–325 www.sciencedirect.com

In a few cases, enrichment studies have supportedsequence-based identifications and clarified roles forone or more microorganisms including denitrifyingPseudomonas [22] and Geminicoccus spp. [23], secondarymetabolite-producing Streptomyces sp. [24], anaerobicammonium oxidizing (anammox) Planctomycetales [25],and oxic and anoxic sulfide oxidizers, Thiomicrospiraand Rhodobacteraceae spp., respectively [26]. Phylogeneticanalyses based on 16S rRNA genes sequences obtainedafter denaturing gradient gel electrophoresis (DGGE) ofPCR-amplified segments have been augmented bysequencing 16S rRNA clone libraries [19!,20]. Finally,comparing DGGE patterns of 16S rRNA genes obtainedfrom whole DNA preparations to reverse-transcribed 16SrRNA (RT-PCR) has provided some information aboutphylotypes active under specific environmental con-ditions [26].
The following is a discussion of key microorganisms infreshwater and marine RAS biofilters and their associatedprocesses as shown in Table 1.
NitrificationCommon to all RASs, the nitrification biofilter is criticalfor maintaining sub-lethal ammonia concentrations —between 0.1 and <3 mg/L (NH3-N) depending onparameters including species, pH, and salinity [27] —with the ionized form, NH4
+, being relatively harmless[27]. Ammonia oxidation is associated with cultivars or16S rRNA sequences ofNitrosomonas sp. andNitrosococcussp. (b-Proteobacteria and g-Proteobacteria, respectively) inall systems examined (Table 1), with the latter dominat-ing brackish or marine nitrification systems [14,15!,28–30]. In addition, one or more Archaea-related species
(Crenarchaeota) have been detected using archaeal ammo-nia monoxygenase gene (amoA) probes [14,15!,16], buttheir role in nitrification has yet to be determined.
The initial product of ammonia oxidation, nitrite, may betolerated at high levels by aquatic species, although manyare adversely affected by long-term exposure to low levels(<0.2 mg/L NO2
"-N) [27]. Nitrite and ammonia oxi-dation occur together in nitrification biofilters andb-proteobacterial Nitrospira sp. is the dominant nitriteoxidizer (Table 1). a-Proteobacterial Nitrobacter sp. hasnot been detected in RASs presumably because of its lowsubstrate affinity compared to Nitrospira sp. [16]. Invol-vement of novel nitrite-oxidizing species, which mayinclude Archaea, has been suggested because of theabsence of Nitrospira-specific targets in some enrichmentcultures [14,15!,16].
Heterotrophic and autotrophic denitrificationUnder anaerobic conditions (in dedicated compartmentsor deep inside biofilm layers), the combination of highnitrate/nitrite levels and organic carbon sources —derived from uneaten feed and fish waste or providedas a supplement [31] — promotes heterotrophic denitri-fication in freshwater and marine RASs [12] (Table 1).Several 16S rRNA gene phylotypes related to denitrifyingheterotrophs, including Pseudomonas sp., Aquaspirillumsp., and various a-Proteobacteria, were found in anaerobicnitrate-stimulated enrichments derived from marine RASnitrification communities, which likely contributed tonitrate-reducing activities [22,30]. Denitrifying Pseudomo-nas sp. appear to be the most abundant g-Proteobacteria inmarine RAS nitrification filters, with strains related toPseudomonas stutzeri and Pseudomonas fluorescens identified
Microbial diversity of aquaculture biofilters Schreier, Mirzoyan and Saito 321
Table 2 (Continued )
Group Microorganisma Source Reference
g-Proteobacteria Acinetobacter sp. Freshwater nitrification filters [2,21]Aeromonas sobria/hydrophilaa Freshwater denitrification drum filter [2]Dechloromarinus chlorophilusa Marine digestion basin sludge [18]Edwardsiella sp. Freshwater nitrification drum filter [2]Marinobacter sp. Marine nitrification filter; Marine digestion basin sludge [26,32]Microbulbifer sp. Marine raceway biofilm [17]Nitrosococcus mobilis Marine nitrification filter [15!]Pseudoalteromonas sp. Marine nitrification filter [19!]Pseudomonas sp. Marine nitrification filter; Marine raceway biofilm;
Freshwater nitrification drum filter; Marinetrickling filter
[2,17,19!,21,22,32]
Pseudomonas stutzeri Marine nitrification filter [19!]Thiococcus sp. Marine raceway biofilm [17]Thiothrix disciformisa Marine denitrification fluidized bed reactor [33]Vibrio sp. Marine nitrification filter [19!,32]
d-Proteobacteria Desulfovibrio caledoniensisa Marine digestion basin sludge [18]
e-Proteobacteria Thiomicrospira (Sulfurimonas) denitrificansa Marine denitrification fluidized bed reactor [33]
Synergistetes Dethiosulfovibrio sp. Marine digestion basin sludge [18]
Crenarchaeota Nitrosopumilus maritimusa Marine denitrification filter [14,15!,16]
a Microorganisms identified solely on the basis of partial 16S rRNA gene or functional gene sequences.
www.sciencedirect.com Current Opinion in Biotechnology 2010, 21:318–325

in 16S rRNA clone libraries and enrichments [19!,22,32].Very few studies have examined denitrifying populationsin freshwater RASs, although Pseudomonas and Comamo-nas spp. are likely participants [21].
In addition to heterotrophic denitrifying microorganisms,marine RAS biofilters contain representatives of theBacteroidetes, Proteobacteria, Furmicutes and Synergistetescapable of sulfide-dependent autotrophic denitrification(Tables 1 and 2) [17,18,19!,26,33]. This activity is stimu-lated by the high sulfate in seawater and is part of a sulfur-based oxidation/reduction cycle established in anaerobicRAS compartments that drives both organic waste diges-tion and nitrate reduction. In this cycle, microorganismsoxidize organic waste to reduce sulfate to sulfide, which isused for sulfide-dependent denitrification to produce N2
gas (Table 1) [5!!,18,34!!]. Sulfate-reducing fermentativeDesulfovibrio, Dethiosulfovibrio, Fusibacter, and Bacterioi-detes, were identified in a marine RAS sludge digestioncompartment [18].
Elimination of toxic sulfide — and, consequently, sulfateregeneration — may occur aerobically by sulfide-oxidiz-ing species including Thiomicrospira sp. [26]. Underanoxic conditions, sulfide oxidation is likely facilitatedby chemolithotrophic nitrate-reducing sulfide-oxidizingbacteria, including e-proteobacterial Thiomicrospora (Sul-furimonas) denitrificans, b-proteobacterial Hydrogenophagasp., g-proteobacterial Thiothrix sp., and the metabolicallydiverse a-proteobacterial Rhodobactereacea having highsequence similarity to Paracoccus, Rhodovulum, Roseobac-ter, and Ruegeria spp. [26,33].
Dissimilatory nitrate reduction to ammonia(DNRA)RAS biofilter compartments having high C/N ratios andelevated sulfide are capable of supporting DNRA, whichwas detected by decreased nitrate and nitrite with aconcomitant increase in ammonia [12]. Nitrate-reducingrelatives of a-Proteobacteria (Paracoccus and Aminobacterspp.), b-Proteobacteria (Alcaligenes and Azoarcus spp.), andd-Proteobacteria (Desulfovibrio sp.) have been found underDNRA-favorable conditions suggesting involvement ofthese microorganisms in this activity [18].
Anaerobic ammonium oxidation (anammox)In addition to denitrifyers, 16S rRNA gene sequences and15N tracer studies have provided evidence for anammoxPlanctomycetales in marine RAS biofilters (Table 1)[5!!,35!,36]. Anammox bypasses a portion of the denitri-fication reaction by direct conversion of ammonia andnitrite to N2 and can lead to significant savings in oper-ational costs and reducing N2O emissions [37]. For RAS,the anammox pathway is important in removing ammo-nia, although it cannot replace denitrification because ofits sensitivity to organic acids and very slow doublingtimes (>11 days). Therefore, a combination of anammox
and denitrification (termed ‘denammox’ or ‘deamox’)[5!!,35!,36] likely contributes to nitrogen waste manage-ment in RAS anaerobic biofilters, similar to their role inthe environment and as applied in industrial wastewatertreatment plants [37–39].
MethanogenesisWhile aerobic and anaerobic microbial activities of RASbiofilters significantly decrease nitrogen and organic mat-ter, complete removal of dissolved and solid organic wastesis a major challenge of an RAS operation. Solid organicwaste — suspended matter originating from uneaten feedand fish feces — is often thickened and used for landapplications or composting. For freshwater systems, thisapproach has environmental consequences and faces regu-latory scrutiny. Marine systems pose an additional com-plication because of the presence of concentrated salts,which would prohibit its use in agriculture applications.Elimination of organic waste from marine RASs usingresident microbial communities is currently under study,with a focus on anaerobic digestion reactors that promoteactivity of methanogenic Archaea [5!!]. Although high saltconcentrations associated with marine RAS sludge mayinhibit methane-producing Archaea, methanogenicactivity has been detected for several marine RASs atlevels lower than those observed for domestic and indus-trial sludge digestion [40]. The archaeal community inbrackish aquaculture sludge digesters consist of membersof both Euryarchaeota and Crenarchaeota, with evidence formethanogenic representatives of the Methanosarcinaceaeobtained from 16S rRNA gene sequences (Mirzoyan andGross, manuscript in preparation).
Pathogens and probioticsCultivars and 16S rRNA sequences have revealed thepresence of pathogenic relatives of Vibrio, Erwinia, Cox-iella, and Aeromonas spp., among others in RAS biofilters[2,17,19!,32]. While antibiotic therapy is often used totreat pathogenic microorganisms, enhanced resistanceand its consequences [41] have necessitated incorporatingUV and ozone treatments to safeguard against infection.RAS biofilters, however, allow for accumulation of patho-gens within protective biofilms [42,43], which could resultin contamination of fish and fish consumer upon detach-ment. Recent approaches for inhibiting pathogen coloni-zation have focused on using probiotic bacteria that maycompete for nutrients, produce growth inhibitors, or,quench cell-to-cell communication that allows for settlingwithin biofilms [44,45,46!!]. Probiotic bacteria includeBacillus, Pseudomonas [46!!], and Roseobacter spp. [47],and bacteria related to these have been identified inRAS biofilters (Table 2), although any role in influencingcolonization remains to be determined.
Conclusions and future perspectivesCharacterization of RAS biofilter communities has pro-vided the foundation for understanding microbial pro-
322 Evironmental biotechnology
Current Opinion in Biotechnology 2010, 21:318–325 www.sciencedirect.com

cesses involved in managing and maintaining efficientand stable wastewater treatment and water quality con-trol. Culture-independent methods, which have, for themost part, relied on 16S rRNA gene analyses, and enrich-ment studies have revealed the presence of phylogeneti-cally diverse microorganisms and identified importantparticipants in functionally distinct regions of RAS bio-filters. Identifications have also established the presenceof important physiologies, for example, anammox andmethanogenesis, which are being applied to RAS tech-nology to achieve efficient, environmentally friendly, andeconomically viable systems.
As RAS technology evolves, its success will lie, in part,not only in continuing to identify essential microorgan-isms associated with specific processes, but also in un-derstanding how they contribute within their consortiumto achieve efficient and stable activities. To that end,future studies that include application of 16S rRNA genemicroarrays [48,49] will provide a broader representationof biofilter communities in different compartments andwill yield information about spatial and temporalrelationships within and between compartments. Thiswill be necessary for assessing the presence (or absence)of critical contributors to a particular process or forevaluating the development of biofilter communitiesin start-up systems. In addition, genetic analysis of com-munities will lead to the development of molecular toolsfor evaluating biofilter conditions and predicting systemfailure. Thus, quantitative assessments of process-specific gene expression, for example, ammonia mono-xygenase (amoA), nitrite reductase (nirS and nirK), anddissimilatory sulfate reductase (dsrA), for nitrification,denitrification, and sulfate reduction, respectively, forthe entire population by microarray-based profiling[50,51] will allow for evaluation of activities throughoutthe system.Given the diversity ofmicroorganisms associ-ated with a particular process, it is clear that many willcontribute to the observed activities and an integrated‘systems biology’ approach should enable functional andphysiochemical relationships to be developed. Finally,metagenomic approaches [52,53] that provide detailedinformation about metabolic functionality will assist inthe design of efficient RASs and bioaugmentation strat-egies.
AcknowledgementsFunding was provided, in part, by grants from the U.S. Israel BinationalAgriculture Research and Development Fund (MB-8720-08) and the U.S.National Oceanic and Atmospheric Administration (NOAA), NationalMarine Aquaculture Initiative (NMAI) (NA08OAR4170821). This iscontribution number 10-200 from the Center of Marine Biotechnology.
References and recommended readingPapers of particular interest, published within the period of review,have been highlighted as:
! of special interest
!! of outstanding interest
1.!
Pulvenis JF: The State ofWorld Fisheries and Aquaculture (SOFIA).Rome: FAO Fisheries and Aquaculture Department; 2009:.http://www.fao.org/fishery/sofia/en (online).
A biennial publication of the United Nations Food and Agriculture Orga-nization (FAO), this advocacy document evaluates world fisheries andaquaculture, reviewing production, utilization, and trade trends, as well asanalysis of selected issues of economic and environmental importance.
2. Schneider O, Chabrillo-Popelka M, Smidt H, Haenen O, Sereti V,Eding EH, Verreth JAJ: HRT and nutrients affect bacterialcommunities grown on recirculation aquaculture systemeffluents. FEMS Microbiol Ecol 2007, 60:207-219.
3. Wik TEI, Linden BT, Wrammer PI: Integrated dynamicaquaculture and wastewater treatment modelling forrecirculating aquaculture systems. Aquaculture 2009, 287:361-370.
4. Zohar Y, Tal Y, Schreier HJ, Steven CR, Stubblefield J, Place AR:Commercially feasible urban recirculating aquaculture:addressing the marine sector. In Urban Aquaculture. Edited byCosta-Pierce B, Desbonnet A, Edwards P, Baker D. CABIPublishing; 2005:159-171.
5.!!
Tal Y, Schreier HJ, Sowers KR, Stubblefield JD, Place AR, Zohar Y:Environmentally sustainable land-based marine aquaculture.Aquaculture 2009, 286:28-35.
This paper reports the development of a fully contained, land-based, nearzero discharge marine RAS for growth of gilthead seabream in a formatthat integrates aerobic and anaerobic microbial processes includingdenitrification, anammox, and methanogenesis.
6. Gelfand I, Barak Y, Even-Chen Z, Cytryn E, van Rijn J, Krom MD,Neori A: A novel zero discharge intensive seawaterrecirculating system for the culture of marine fish. J WorldAquacult Soc 2003, 34:344-358.
7. Keller M, Zengler K: Tapping into microbial diversity. Nat RevMicrobiol 2004, 2:141-150.
8. Shnel N, Barak Y, Ezer T, Dafni Z, van Rijn J: Design andperformance of a zero-discharge tilapia recirculating system.Aquacult Eng 2002, 26:191-203.
9. Avnimelech Y: Bio-filters: the need for an new comprehensiveapproach. Aquacult Eng 2006, 34:172-178.
10. Gutierrez-Wing MT, Malone RF: Biological filters in aquaculture:trends and research directions for freshwater and marineapplications. Aquacult Eng 2006, 34:163-171.
11. Eding EH, Kamstra A, Verreth JAJ, Huisman EA, Klapwijk A:Designandoperationofnitrifying tricklingfilters in recirculatingaquaculture: a review. Aquacult Eng 2006, 34:234-260.
12. van Rijn J, Tal Y, Schreier HJ: Denitrification in recirculatingsystems: theory and applications. Aquacult Eng 2006,34:364-376.
13. Stickney RR: Recirculating water systems. In Encyclopediaof Aquaculture. Edited by Stickney RR. John Wiley and Sons;2000: 722-731.
14. Pedersen L-F, Pedersen PB, Nielsen JL, Nielsen PH: Peraceticacid degradation and effects on nitrification in recirculatingaquaculture systems. Aquaculture 2009, 296:246-254.
15.!
Foesel BU, Gieseke A, Schwermer C, Stief P, Koch L, Cytryn E, dela Torre JR, van Rijn J, Minz D, Drake HL et al.: NitrosomonasNm143-like ammonia oxidizers and Nitrospira marina-likenitrite oxidizers dominate the nitrifier community in a marineaquaculture biofilm. FEMS Microbiol Ecol 2008, 63:192-204.
The authors present a quantitative analysis of ammonia and nitrite-oxidizing bacteria in a marine RAS nitrification biofilter using 16S rRNAand amoA probes. The very low fraction of Crenarchaeaota amoA signalsleads them to speculate that the contribution of these Archaea toammonia oxidation in their RAS is minimal.
16. Urakawa H, Tajima Y, Numata Y, Tsuneda S: Low temperaturedecreases the phylogenetic diversity of ammonia-oxidizingarchaea and bacteria in aquarium biofiltration systems. ApplEnviron Microbiol 2008, 74:894-900.
17. Wietz M, Hall MR, Hoj L: Effects of seawater ozonation onbiofilm development in aquaculture tanks. Syst Appl Microbiol2009, 32:266-277.
Microbial diversity of aquaculture biofilters Schreier, Mirzoyan and Saito 323
www.sciencedirect.com Current Opinion in Biotechnology 2010, 21:318–325

18. Cytryn E, Gelfand I, Barak Y, van Rijn J, Minz D: Diversity ofmicrobial communities correlated to physiochemicalparameters in a digestion basin of a zero-dischargemariculture system. Environ Microbiol 2003, 5:55-63.
19.!
Michaud L, Lo Giudice A, Troussellier M, Smedile F, Bruni V,Blancheton JP: Phylogenetic characterization of theheterotrophic bacterial communities inhabiting a marinerecirculating aquaculture system. J Appl Microbiol 2009,107:1935-1946.
A culture-dependent and culture-independent analysis of communitiesfrom different compartments of a marine RAS biofilter that highlights thediscrepancy between the two approaches and points out the lack ofrichness in diversity when considered separately.
20. Sugita H, Nakamura H, Shimada T: Microbial communitiesassociated with filter material in recirculating aquaculturesystems of freshwater fish. Aquaculture 2005, 243:403-409.
21. Itoi S, Ebihara N, Washio S, Sugita H: Nitrite-oxidizing bacteria,Nitrospira, distribution in the outer layer of the biofilm fromfilter materials of a recirculating water system for the goldfishCarassius auratus. Aquaculture 2007, 264:297-308.
22. Borges MT, Sousa A, De Marco P, Matos A, Honigova P,Castro PM: Aerobic and anoxic growth and nitrate removalcapacity of a marine denitrifying bacterium isolated from arecirculation aquaculture system. Microb Ecol 2008, 55:107-118.
23. Foesel BU, Gossner AS, Drake HL, Schramm A: Geminicoccusroseus gen. nov., sp. nov., an aerobic phototrophicAlphaproteobacterium isolated from a marine aquaculturebiofilter. Syst Appl Microbiol 2007, 30:581-586.
24. Guttman L, van Rijn J: Identification of conditions underlyingproduction of geosmin and 2-methylisoborneol in arecirculating system. Aquaculture 2008, 279:85-91.
25. Tal Y, Watts JE, Schreier HJ: Anaerobic ammonium-oxidizing(anammox) bacteria and associated activity in fixed-filmbiofilters of a marine recirculating aquaculture system. ApplEnviron Microbiol 2006, 72:2896-2904.
26. Cytryn E, van Rijn J, Schramm A, Gieseke A, de Beer D, Minz D:Identification of bacteria potentially responsible for oxic andanoxic sulfide oxidation in biofilters of a recirculatingmariculture system. Appl Environ Microbiol 2005, 71:6134-6141.
27. Stickney RR:Nitrogen. In Encyclopedia of Aquaculture. Edited byStickney RR. John Wiley and Sons; 2000:586-590.
28. Hovanec TA, DeLong EF: Comparative analysis of nitrifyingbacteria associated with freshwater and marine aquaria. ApplEnviron Microbiol 1996, 62:2888-2896.
29. Paungfoo C, Prasertsan P, Burrell PC, Intrasungkha N, Blackall LL:Nitrifying bacterial communities in an aquaculture wastewatertreatment system using fluorescence in situ hybridization(FISH), 16S rRNA gene cloning, and phylogenetic analysis.Biotechnol Bioeng 2007, 97:985-990.
30. Tal Y, Watts JE, Schreier SB, Sowers KR, Schreier HJ:Characterization of the microbial community and nitrogentransformation processes associated with moving bedbioreactors in closed recirculated mariculture systems.Aquaculture 2003, 215:187-202.
31. Morrison MM, Tal Y, Schreier HJ: Stimulating denitrification in amarine recirculating aquaculture system biofilter usinggranular starch as a carbon source. Int J Recirculating Aquacult2008, 9:23-41.
32. Leonard N, Blancheton JP, Guiraud JP: Populations ofheterotrophic bacteria in an experimental recirculatingaquaculture system. Aquacult Eng 2000, 22:109-120.
33. Cytryn E, Minz D, Gelfand I, Neori A, Gieseke A, De Beer D, VanRijn J: Sulfide-oxidizing activity and bacterial communitystructure in a fluidized bed reactor from a zero-dischargemariculture system. Environ Sci Technol 2005, 39:1802-1810.
34.!!
Sher Y, Schneider K, Schwermer C, van Rijn J: Sulfide-inducednitrate reduction in the sludge of an anaerobic digester of azero-discharge recirculating mariculture system. Water Res2008, 42:4386-4392.
This study analyzes the physiochemical characteristics of anaerobiccompartments of a zero-discharge marine RAS and establishes relation-ships with sulfate-reducing and nitrate-dependent sulfide-oxidizingpopulations identified in previous studies (Refs. [18,26,33]).
35.!
Lahav O, Tackoubov D, Zelikson R, Mozes N, Tal Y, Tarre S:Quantification of anammox activity in a denitrification reactorfor a recirculating aquaculture system. Aquaculture 2009,288:76-82.
This paper describes fluorescence in situ hybridization and anammoxactivity experiments with denitrification biofilms from a marine RAS. Theauthors hypothesize that the anammox-related bacteria identified in theirsystem biofilters were derived from the cultured fish species (giltheadseabream), as significant quantities of identical anammox-relatedsequences were detected in fish feces.
36. Kalyuzhnyi S, Gladchenko M, Mulder A, Versprille B: DEAMOX —new biological nitrogen removal process based on anaerobicammonia oxidation coupled to sulphide-driven conversion ofnitrate into nitrite. Water Res 2006, 40:3637-3645.
37. Paredes D, Kuschk P, Mbwette TSA, Stange F,Muller RA, Koser H:New aspects of microbial nitrogen transformations in thecontext of wastewater treatment — a review. Eng Life Sci 2007,7:13-25.
38. Strous M, Heijnen JJ, Kuenen JG, Jetten MSM: The sequencingbatch reactor as a powerful tool for the study of slowlygrowing anaerobic ammonium-oxidizing microorganisms.Appl Microbiol Biotechnol 1998, 50:589-596.
39. Schmidt I, Sliekers O, Schmid M, Bock E, Fuerst J, Kuenen JG,Jetten MS, Strous M: New concepts of microbial treatmentprocesses for the nitrogen removal in wastewater. FEMSMicrobiol Rev 2003, 27:481-492.
40. Mirzoyan N, Parnes S, Singer A, Tal Y, Sowers KR, Gross A:Quality of brackish aquaculture sludge and its suitability foranaerobic digestion and methane production in an upflowanaerobic sludge blanket (UASB) reactor. Aquaculture 2008,279:35-41.
41. Smith P: Antimicrobial resistance in aquaculture. Rev SciTechnol 2008, 27:243-264.
42. Sharrer MJ, Summerfelt ST, Bullock GL, Gleason LE, Taeuber J:Inactivation of bacteria using ultraviolet irradiation in arecirculating salmonid culture system. Aquacult Eng 2005,33:135-149.
43. King RK, Flick GJ, Pierson MD, Smith SA, Boardman GD,Coale CW: Identification of bacterial pathogens in biofilms ofrecirculating aquaculture systems. J Aquat Food Prod Technol2004, 13:125-133.
44. Defoirdt T, Boon N, Sorgeloos P, VerstraeteW, Bossier P:Quorumsensing and quorum quenching in Vibrio harveyi: lessonslearned from in vivo work. ISME J 2008, 2:19-26.
45. Defoirdt T, Boon N, Sorgeloos P, Verstraete W, Bossier P:Alternatives to antibiotics to control bacterial infections:luminescent vibriosis in aquaculture as an example. TrendsBiotechnol 2007, 25:472-479.
46.!!
Kesarcodi-Watson A, Kaspar H, LateganMJ, Gibson L:Probioticsin aquaculture: the need, principles andmechanisms of actionand screening processes. Aquaculture 2008, 274:1-14.
A comprehensive summary of the research and methodology of probio-tics in aquaculture with an emphasis on shellfish aquaculture.
47. Bruhn JB, Nielsen KF, Hjelm M, Hansen M, Bresciani J, Schulz S,Gram L: Ecology, inhibitory activity, and morphogenesis of amarine antagonistic bacterium belonging to the Roseobacterclade. Appl Environ Microbiol 2005, 71:7263-7270.
48. Green BD, Keller M: Capturing the uncultivated majority. CurrOpin Biotechnol 2006, 17:236-240.
49. DeSantis TZ, Brodie EL, Moberg JP, Zubieta IX, Piceno YM,Andersen GL: High-density universal 16S rRNA microarrayanalysis reveals broader diversity than typical clone librarywhen sampling the environment. Microb Ecol 2007, 53:371-383.
50. Urakawa H, Matsumoto J, Inaba K, Tsuneda S: DNA microarraymediated transcriptional profiling of Nitrosomonas europaea
324 Evironmental biotechnology
Current Opinion in Biotechnology 2010, 21:318–325 www.sciencedirect.com

in response to linear alkylbenzene sulfonates. FEMS MicrobiolLett 2008, 282:166-173.
51. Wu L, Thompson DK, Li G, Hurt RA, Tiedje JM, Zhou J:Development and evaluation of functional gene arrays fordetection of selected genes in the environment. Appl EnvironMicrobiol 2001, 67:5780-5790.
52. Tringe SG, von Mering C, Kobayashi A, Salamov AA, Chen K,Chang HW, Podar M, Short JM, Mathur EJ, Detter JC et al.:
Comparative metagenomics of microbial communities.Science 2005, 308:554-557.
53. Roland JS, Marco G: Genomics of biological wastewatertreatment. Microb Biotechnol 2008, 1:333-340.
54. Hovanec TA, Taylor LT, Blakis A, Delong EF: Nitrospira-likebacteria associated with nitrite oxidation in freshwateraquaria. Appl Environ Microbiol 1998, 64:258-264.
Microbial diversity of aquaculture biofilters Schreier, Mirzoyan and Saito 325
www.sciencedirect.com Current Opinion in Biotechnology 2010, 21:318–325




















