
Mercury in the eggs of aquatic birds from the Gulf of Gdansk and Wloclawek Dam (Poland)
12
1 23 Environmental Science and Pollution Research ISSN 0944-1344 Environ Sci Pollut Res DOI 10.1007/s11356-015-4154-y Mercury in the eggs of aquatic birds from the Gulf of Gdansk and Wloclawek Dam (Poland) Agnieszka Grajewska, Lucyna Falkowska, Emilia Szumiło-Pilarska, Julia Hajdrych, Marta Szubska, Tomasz Frączek, Włodzimierz Meissner, et al.
-
Upload
independent -
Category
Documents
-
view
2 -
download
0
Transcript of Mercury in the eggs of aquatic birds from the Gulf of Gdansk and Wloclawek Dam (Poland)

1 23
Environmental Science and PollutionResearch ISSN 0944-1344 Environ Sci Pollut ResDOI 10.1007/s11356-015-4154-y
Mercury in the eggs of aquatic birds fromthe Gulf of Gdansk and Wloclawek Dam(Poland)
Agnieszka Grajewska, LucynaFalkowska, Emilia Szumiło-Pilarska,Julia Hajdrych, Marta Szubska, TomaszFrączek, Włodzimierz Meissner, et al.

1 23
Your article is protected by copyright and
all rights are held exclusively by Springer-
Verlag Berlin Heidelberg. This e-offprint is
for personal use only and shall not be self-
archived in electronic repositories. If you wish
to self-archive your article, please use the
accepted manuscript version for posting on
your own website. You may further deposit
the accepted manuscript version in any
repository, provided it is only made publicly
available 12 months after official publication
or later and provided acknowledgement is
given to the original source of publication
and a link is inserted to the published article
on Springer's website. The link must be
accompanied by the following text: "The final
publication is available at link.springer.com”.

RESEARCH ARTICLE
Mercury in the eggs of aquatic birds from the Gulf of Gdanskand Wloclawek Dam (Poland)
Agnieszka Grajewska & Lucyna Falkowska & Emilia Szumiło-Pilarska &
Julia Hajdrych & Marta Szubska & Tomasz Frączek & Włodzimierz Meissner &
Szymon Bzoma & Magdalena Bełdowska & Andrzej Przystalski & Tomasz Brauze
Received: 15 November 2014 /Accepted: 19 January 2015# Springer-Verlag Berlin Heidelberg 2015
Abstract The aim of this paper was to assess the influence ofdiet on the concentrations of total mercury (HgTOT) in the eggsof aquatic birds. Trophic level was determined using stableisotopes (δ15N, δ13C). Analysis was carried out on eggs (laidin 2010–2012) belonging to two species of terns nesting at theRiver Vistula outlet on the Gulf of Gdansk and on herringgulls nesting both in Gdynia harbour and on the Vistula damin Wloclawek. The results show that seafood diet causes thehighest load of mercury, that which is transferred into ternseggs. The amounts of accumulated mercury obtained werefound to be different in the particular egg components withHgalbumen>Hgyolk>Hgmembrane>Hgshell. In the herring gulleggs, three stages of embryo development with varying levelsof mercury were determined. It was observed that mercuryreceived from the albumen and yolk was most effectivelyremoved when developing embryo into down.
Keywords Total mercury . Stable isotope analysis . Terns .
Herring gull . Bird eggs . Embryos . Coastal zone . SouthernBaltic Sea
Introduction
Mercury is an element that occurs naturally in the environ-ment. Human activity has resulted in the formation of addi-tional emission sources of this element, making it one of themost dangerous pollutants. Mercury emission from anthropo-genic sources, although on a global scale lower than naturalemission, may pose a much greater threat locally.Anthropogenic contamination of the environment with mer-cury is caused mainly by the burning of fossil fuels, predom-
Responsible editor: Philippe Garrigues
HighlightsHg concentration in avian eggs is determined by type and origin of food.Albumens contained the highest Hg concentrations due to Hg’s affinity to -SH groups.Herring gull embryos accumulate Hg in the liver and heart and sequester itin down.Herring gull chicks rapidly eliminate mercury after hatching.
A. Grajewska : L. Falkowska (*) : E. Szumiło-Pilarska :J. Hajdrych :M. Szubska :M. BełdowskaDepartment of Marine Chemistry and Environmental Protection,Faculty of Oceanography and Geography, University of Gdansk,Pilsudskiego 46, 81-387 Gdynia, Polande-mail: [email protected]
L. Falkowskae-mail: [email protected]
T. FrączekInstitute of Applied Radiation Chemistry, Faculty of Chemistry, LodzUniversity of Technology, Zeromskiego 116, 90-924 Lodz, Poland
W. MeissnerAvian Ecophysiology Unit, Department of Vertebrate Ecology andZoology, Faculty of Biology, University of Gdansk, Wita Stwosza59, 80-308 Gdansk, Poland
S. BzomaWaterbird Research Group KULING, Swierkowa 34/7,81-526 Gdynia, Poland
A. Przystalski : T. BrauzeDepartment of Vertebrate Zoology, Faculty of Biology andEnvironment Protection, Nicolaus Copernicus University, Lwowska1, 87-100 Torun, Poland
Environ Sci Pollut ResDOI 10.1007/s11356-015-4154-y
Author's personal copy

inantly hard coal and lignite (Wängberg et al. 2001). Mercuryis introduced into the water of the Baltic Sea from pointsources and via rivers (Saniewska et al. 2014). Most of themercury carried with river water becomes accumulated in thecoastal zone of the sea with only a small amount reaching theopen sea. Recent publications point to the warming of theclimate and the resultant increase in the frequency of extremeweather phenomena such as floods, as a factor that may con-siderably raise the amount of mercury transported into the seavia rivers (Saniewska et al. 2014).
Studies into the mercury present in the environment areundertaken due to its high toxicity. Its harmful influence de-pends on the route of introduction into the system and theform in which it is found, the dose and time of exposure.The ease of absorption results in the accumulation and mag-nification of mercury in the trophic chain. The studies of birdsin shore nesting areas have indicated threats resulting fromthis habitat and the food from which mercury becomes accu-mulated in their tissues and organs but also in eggs (Burgerand Gochfeld 2003; Braune 2007; Bond and Diamond 2009a,b). Long-lived predatory seabirds inhabiting the coastal zone,where human-generated contaminant levels may be high, ex-perience elevated Hg exposure and hence they are good indi-cators of the environmental pollution level (Burger 1993;Monteiro and Furness 1995).
Long-term exposure of a bird’s system to mercury mayhave a number of undesirable effects. Being a neurotoxin,mercury may influence behaviour and motor skills (Everset al. 2008; Kenov et al. 2010; Splading et al. 2000). It mayalso heighten the potential for infection by disease organisms(Scheuhammer et al. 2007). Of all the chemical forms of mer-cury, aquatic piscivorous birds are primarily exposed to themost toxic—methylmercury, and this is mainly through thediet (Thompson 1996; Wiener et al. 2003; Scheuhammeret al. 2007). Dietary ingested Hg is transported in blood andmay be accumulated in internal tissues (Hall et al. 1997). Birdshave nevertheless developed mechanisms which allow themto remove the accumulated mercury from their bodies. Themost effective mercury elimination takes place through incor-porating it into growing feathers during moulting (Burger1993; Becker et al. 1994; Burger and Gochfeld 1997; Bondand Diamond 2009a). It is estimated that between 70 and 93%of all mercury accumulated in the body becomes sequestratedin feathers (Braune and Gaskin 1987; Burger and Gochfeld1997; Bond and Diamond 2009a). Small amounts of mercuryare also excreted with guano, but its reintroduction to theenvironment is quicker (Yin et al. 2008). Females may addi-tionally eliminate accumulated mercury into eggs (Becker1992). The presence of mercury in bird eggs may result inlower hatchability, embryonic mortality and malformationsas well as lower chick survival (Burger and Gochfeld 1997;Scheuhammer et al. 2007; Heinz et al. 2009). MeHg predom-inates in avian eggs (Rumbold et al. 2001; Scheuhammer et al.
2001; Bond and Diamond 2009a; Lavoie et al. 2010). Forincome breeders such as terns or gulls, the level of mercuryin their eggs reflects dietary uptake from the surrounding areain a short period before the eggs are laid (Monteiro andFurness 1995; Cotin et al. 2011). The embryo seems to bethe life stage at which seabirds are most sensitive to methyl-mercury (Heinz et al. 2006), but yet very little is known abouthow MeHg distributes in the avian embryo and recentlyhatched chicks (Rutkiewicz and Basu 2013). Heinz et al.(2009) suggest that sensitivity of avian embryos to methyl-mercury might be related to the taxonomic status of the bird.
The following factors can reflect differences in mercuryconcentrations between species: feeding habits, migratoryhabits, body size, life span, moult strategy and taxonomicinfluences on physiology (Walsh 1990). Moult, age and sex,meanwhile, can be described as intra-specific factors(Monteiro and Furness 1995). In eggs, significant declineswere also observed in connection with laying sequence(Becker 1992; Akearok et al. 2010).
Stable carbon (δ13C) and nitrogen (δ15N) isotope analysisreflects the consumer’s diet at the time of tissue synthesis andis commonly used to investigate trophic relationships and tounderstand foraging ecology of birds (Hobson and Clark1992a, b; Cotin et al. 2011; Burgess et al. 2013). δ13C canindicate a geographic foraging area by determining relativecontributions of marine and terrestial foods to the diet. In themarine environment, inshore and more productive oceanicareas are enriched with 13C, whereas offshore areas demon-strate considerably lower levels (Peterson and Fry 1987). δ15Nreflects trophic position as its values increase by 2–5 % oneach trophic level due to the preferential excretion of 14N withnitrogenous waste (Steele and Daniel 1978; Minagawa andWada 1984; Kelly 2000).
Alimentary differences resulting from the dissimilar natureof freshwater and brackish water basins will have an effect onthe mercury load delivered to the birds’ systems with food andby the same token will influence the mercury load transferredto the next generation by a female. For this reason, researchwas undertaken in order to determine the influence of diet onmercury concentrations in the eggs of aquatic birds from theregion of the Gulf of Gdansk and the dam on the River Vistulain Wloclawek. Another equally important aim was to explorethe depuration ability of the embryo and to determine themercury load that a newly hatched herring gull chick is bur-dened with at the very start of its life.
Material and methods
Sample collection and sampling area
The study was carried out in the years 2010–2012 using theeggs of three aquatic bird species: the sandwich tern (Sterna
Environ Sci Pollut Res
Author's personal copy

sandvicensis, Latham, 1787), the common tern (Sternahirundo, Linneaus, 1758) and the herring gull (Larusargentatus, Pontoppidan, 1763). All samples were collectedunder appropriate collecting permits.
Tern eggs were collected in the Mewia Lacha BirdSanctuary (54° 21′ N, 18° 56′ E) during the breeding seasonin the summer of 2010 and 2012. The reserve, established in1991 and measuring 150 ha in total, is located in the estuary ofthe River Vistula and includes sandy beaches, dunes, vastshallows with sand bars which appear and disappear periodi-cally, freshwater lakes, meadows and a forest (BirdLifeInternational 2013). The purpose of the reserve is to protectthe breeding, resting and feeding areas of rare tern species andbirds from the Charadriiformes and Anseriformes orders andalso to protect the natural scenery of the River Vistula moutharea (M.P. Nr 38, item 273). Indeed, the reserve is the onlybreeding area of the sandwich tern in Poland, the birds creat-ing shallow hole nests on offshore sand bars. The eggs used intests were collected because the breedings failed. In 2010, terneggs were collected after a sand bar was flooded, resulting inthe eggs being washed out of nests. The flood was one of themost severe ever recorded in Poland, and the peak of thefreshet on the River Vistula was the highest in 160 years(IMGW 2011). Twelve eggs belonging to the common ternand 69 to the sandwich tern were collected at that time. In2012, 43 sandwich tern eggs were collected after the colonywas destroyed by high storm waves. The size of the breedingcolony at that time was estimated at 276 sandwich tern pairsand 100 common tern pairs (Meissner et al. 2014). Owing tothe flood, there is no data concerning the size of the breedingcolony in 2010.
Thirty herring gull eggs (Larus argentatus, sensu lato) werecollected in 2011 at the breeding colony located on the RiverVistula dam inWloclawek (52° 39′N, 19° 08′E). Among themwere unfertilised and unhatched eggs, probably abandoned bythe parents. The breeding colony consisted of 138 pairs butwas a mixed colony of the herring gull, the Caspian gull(Larus cachinnans, Pallas, 1811) and hybrid specimens.Extra care was taken to make sure that the eggs selected forthe study belonged to the herring gull; however, owing to theclose proximity of the two species, the eggs need to be consid-ered to belong to the herring gull sensu lato. The damwas builtat Rkm 675 in 1969 as one of the six features regulating theflow of the lower Vistula. However, the project was not fin-ished and the remaining five elements were never constructed.The result of the dam, however, was the formation of the larg-est water reservoir in Poland—the Wloclawek Reservoir.
The material was complemented by samples collected in2010 in and around Gdynia harbour (54° 31′ N 18° 31′ E),where a herring gull breeding colony comprising over 10 nestswas found on the roof of a building. One unhatched egg wasfound there and two chicks which had died having fallen offthe roof.
Sample preparation
The eggs were collected into polyethylene Ziploc bags. In thelaboratory, they were washed and weighed. The material wasthen kept in a freezer at a temperature of −20 °C. Wherepossible, after shelling, the frozen eggs were separated intoalbumen and yolk. If that was not possible (e.g. owing todecomposition), the sample contained a whole egg. In a fewcases of sandwich tern eggs, it was only possible to partlyseparate a given structure after the eggs were opened (the restof the eggs, where albumen and yolk were mixed, wereanalysed separately), thus explaining why the numbers of al-bumen and yolk samples differ in Table 1.
Prior to analysis, all samples were lyophilised and weighedagain. The shells of tern eggs from the Mewia Lacha reservewere also used for tests. Shells with membranes were washedin deionised water. The membranes removed from the shellsof 10 sandwich tern eggs were washed in deionised waterthree times and then put in a drier at a temperature not exceed-ing 30 °C. With the other shells, parts of the shell were takenfor analysis from different places, without separating themembrane. All the shells (with and without membranes) werewashed in an ultrasonic bath containing 50% acetone solutionand then placed in a drier at a temperature not exceeding30 °C. The number of analysed samples is presented inTable 1.
In 10 herring gull eggs (nine from the Wloclawek dam andone from Gdynia harbour), developing embryos were found.Internal organs (liver and heart) and downwere collected fromboth the embryos and the 2-week-old chicks. Owing to thefragility of the developing embryos, the internal organs werecollected after the sample had been lyophilised, while thechicks were dissected. Their organs were then lyophilised.Depending on their size, embryos from the Wloclawek damwere divided into three groups (three stages of embryonicdevelopment: early, middle, late). The first group comprisedonly one, the smallest embryo. Out of the remaining eight,four were assigned to group 3, having pipped their eggshells.That meant the chicks were fully developed and ready tohatch. The down collected from embryos and chicks waswashed in an acetone solution in an ultrasonic bath and thendried at a temperature not exceeding 30 °C.
Stable isotope analysis
In selected lyophilised albumens (10 of the sandwich tern, 5 ofthe common tern and 5 of the herring gull), an analysis ofstable isotopes δ15N and δ13C was carried out using theSercon 20-22 Continuous Flow Isotope Ratio MassSpectrometer (CF-IRMS) coupled with Sercon SL elementalanalyser for simultaneous carbon-nitrogen-sulfur (NCS) anal-ysis. The albumens selected for analysis were chosen so as torepresent the whole range of total mercury concentration in
Environ Sci Pollut Res
Author's personal copy
the samples. Weighted into tin capsules were 1.0±0.2 mg ofsamples, and 0.5–0.75 mg of vanadium pentoxide was addedas a catalyst. An in-house standard thiobarbituric acid (δ15N=0.11±0.9 (Air), δ13C=28.35±0.06 (PDB)) was used as areference.
The results were expressed as differences in isotopic ratiosas parts per thousand (‰) according to the equation
δX ¼ RSample
RStandard
� �−1
� �˙1000
where X represents 15N or 13C and is the corresponding15N/14N or 13C/12C ratio.
Total mercury analysis
In the analysed samples, the concentration of total mercurywas assayed using an AMA-254 atomic absorption spectrom-eter. Weighed amounts of the biological material were placedin fired nickel boats. Each sample was assayed in two or threerepeats. An empty fired nickel boat was used as a blank sam-ple. A sample, having been automatically inserted into thefurnace, was dried (120 °C/300 s) and mineralised (550 °C/180 s). After sample inflammation, mercury was captured intothe amalgamator then released from it as a result of heating upand gradually transferred onto the detector. The absorbancemeasurement occurred at a wavelength of 253.6 nm. The du-ration of each measurement cycle was 60 s.
The method precision was measured using a certified stan-dard BCR414 prepared on the basis of plankton and BCR463prepared on the basis of tuna. The precision was 5 %.Accuracy expressed as mercury recovery was established at96.7 %, while the limit of quantification (LOQ) amounted to0.075 ng Hg g−1 d.w.
Statistical analysis
The statistical processing of the obtained results was per-formed using Microsoft Excel and statistical programmes. Inorder to assess the normality of the distribution of HgTOT
results in the studied samples, the Shapiro-Wilk test was used.When comparing the results for isotopes, the one-wayANOVA was used, followed by the Tukey’s test for variousn. In the rest of the cases, in order to test the significance ofdifferences, non-parametric tests were used: the Mann-Whitney U test, the Kruskall-Wallis test and the Wilcoxontest. All statistical analyses were performed at a confidencelevel of 95 %. When dividing embryos into groups (early,middle, late), only the size of the embryo was taken intoaccount.
Results
Stable isotope
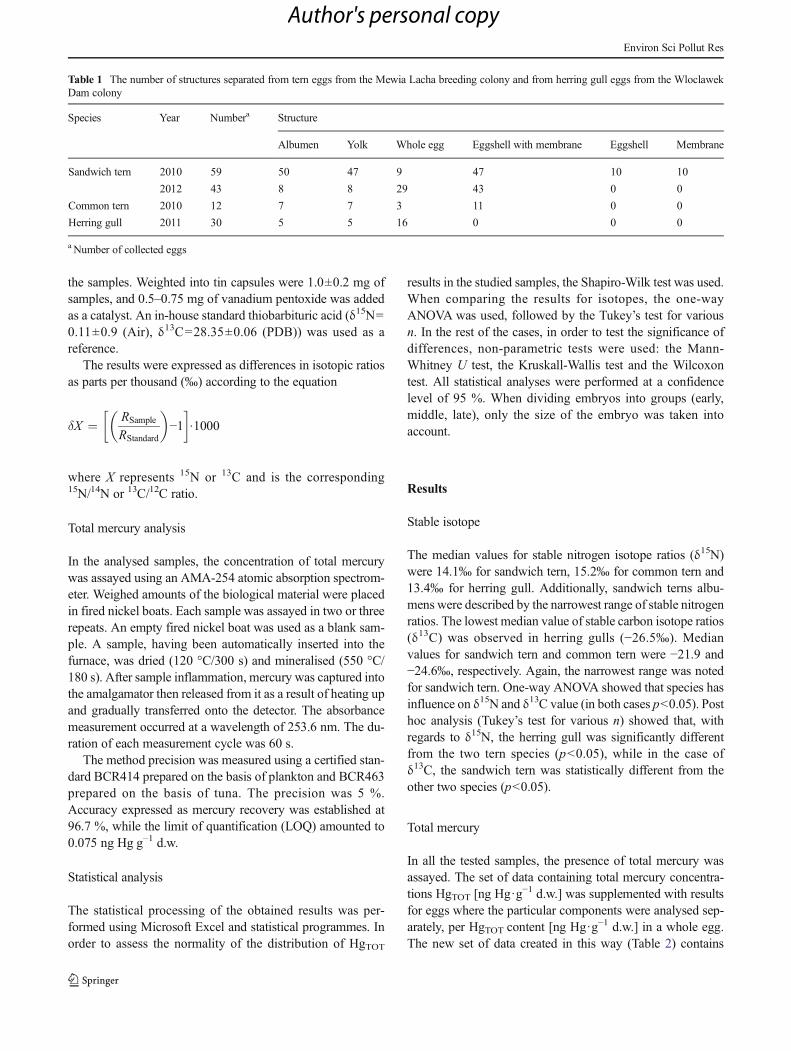
The median values for stable nitrogen isotope ratios (δ15N)were 14.1‰ for sandwich tern, 15.2‰ for common tern and13.4‰ for herring gull. Additionally, sandwich terns albu-mens were described by the narrowest range of stable nitrogenratios. The lowest median value of stable carbon isotope ratios(δ13C) was observed in herring gulls (−26.5‰). Medianvalues for sandwich tern and common tern were −21.9 and−24.6‰, respectively. Again, the narrowest range was notedfor sandwich tern. One-way ANOVA showed that species hasinfluence on δ15N and δ13C value (in both cases p<0.05). Posthoc analysis (Tukey’s test for various n) showed that, withregards to δ15N, the herring gull was significantly differentfrom the two tern species (p<0.05), while in the case ofδ13C, the sandwich tern was statistically different from theother two species (p<0.05).
Total mercury
In all the tested samples, the presence of total mercury wasassayed. The set of data containing total mercury concentra-tions HgTOT [ng Hg·g−1 d.w.] was supplemented with resultsfor eggs where the particular components were analysed sep-arately, per HgTOT content [ng Hg·g−1 d.w.] in a whole egg.The new set of data created in this way (Table 2) contains
Table 1 The number of structures separated from tern eggs from the Mewia Lacha breeding colony and from herring gull eggs from the WloclawekDam colony
Species Year Numbera Structure
Albumen Yolk Whole egg Eggshell with membrane Eggshell Membrane
Sandwich tern 2010 59 50 47 9 47 10 10
2012 43 8 8 29 43 0 0
Common tern 2010 12 7 7 3 11 0 0
Herring gull 2011 30 5 5 16 0 0 0
aNumber of collected eggs
Environ Sci Pollut Res
Author's personal copy

results for 92 sandwich tern eggs, 10 common tern eggs and21 herring gull eggs. There were no statistically relevant dif-ferences (Mann-Whitney U test, p>0.05) between sandwichtern eggs collected in 2010 and 2012 in the Mewia LachaSanctuary. However, the Kruskall-Wallis analysis of varianceshowed statistically relevant differences between the studiedspecies (p<0.05). The multiple post hoc analyses showed thatHgTOT concentrations in the eggs of the sandwich tern and thecommon tern were significantly higher than HgTOTconcentra-tions in the eggs of the herring gull (p<0.05) but that therewere no differences between HgTOT concentrations in terneggs. The highest HgTOT concentration values were found insandwich tern eggs, for which the two highest values (3384.1and 3213.4 ng Hg·g−1 d.w.) were classed as extreme and thefollowing four (2750.5; 2525.0; 2332.1; 2311.0 ng Hg·g−1
d.w.) as outlying. In the eggs of the herring gull, the highesttwo values were also characterised as outlying (1196.1 and1136.3 ng Hg·g−1 d.w.).
In the eggs of all the studied species, statistically relevantdifferences were found between the mercury concentrations inthe albumen and in the yolk (Table 3). The Wilcoxon test(p<0.05) was performed only for eggs in which it was possi-ble to assay HgTOT in both the albumen and yolk (nST=53pairs, nCT=7 pairs, nHG=5 pairs). The concentrations in thealbumens were an order of magnitude higher than mercuryconcentrations in the yolks, and HgTOT concentrations in theyolks did not exceed the minimum HgTOT values in the albu-mens for any of the species. In the eggs of the sandwich tern,the albumens were from 2.3 to 91.2 times richer in mercury
(median value 25.3). The highest concentration ratiosHgalbumen/Hgyolk were determined in eggs where the mercurycontent in the yolk was the lowest, while low ratios werecharacteristic of eggs with high mercury content. A similartendency was observed for the other studied species. The al-bumens of the common tern and the herring gull eggs werecharacterised, respectively, by mercury concentrations thatwere 2.6–19.0 (median value 11.6) and 9.1–20.8 (median val-ue 16.6) higher compared with the yolks. In the case of sand-wich tern eggs, a statistically significant dependence was ob-served between the HgTOT concentration in the albumen andthat in the yolk (rSpearmann=0.35; p<0.05).
The results obtained for the external components of terneggs (Table 4) showed that mercury concentrations in theshells were two orders of magnitude lower than in the mem-branes. The Wilcoxon test (p<0.05) showed that the com-pared groups were significantly different, although the con-centration ranges assayed in the eggshells with membranewere wide for both species. The Mann-Whitney U testshowed no statistically relevant difference between sand-wich and common tern. No results were obtained where,within one egg, the mercury concentration in the eggshelland membrane was higher than the concentration in theyolk.
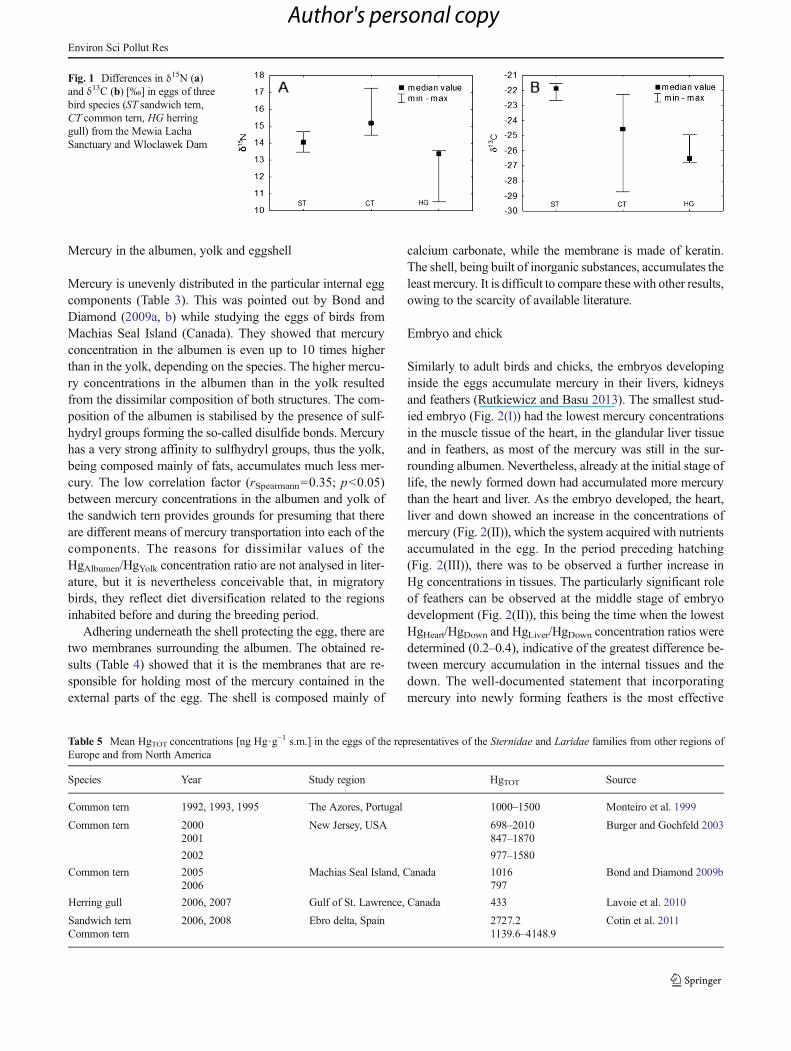
Apart from assaying mercury concentrations in the partic-ular egg components, analyses were conducted on the internalorgans and the down of embryos found in the eggs of theherring gull. The results for total mercury concentrations innine embryo livers from the Wloclawek Dam nesting colony(Fig. 2.) were contained within the range of 240.6–1677.0 ngHg·g−1 d.w. Similar values were obtained for the hearts:160.3–1643.2 ng Hg·g−1 d.w. Mercury concentrations in thedown had the widest range: 458.1–2860.8 ng Hg·g−1 d.w.
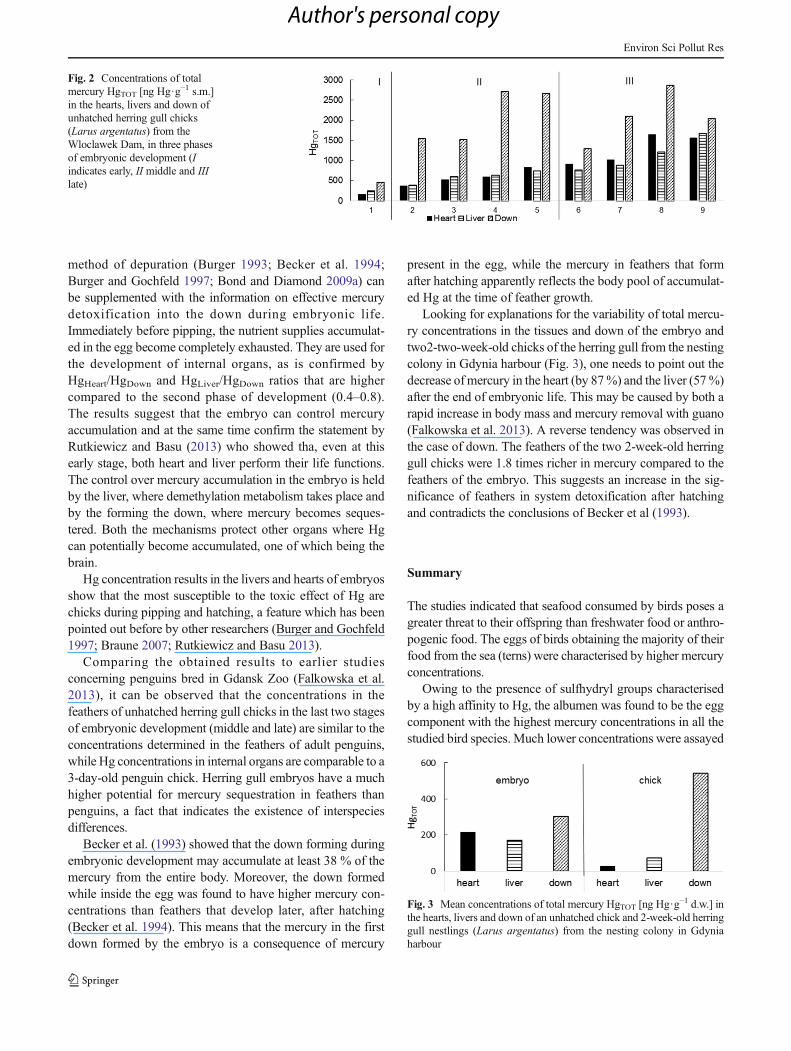
In the case of the herring gull embryo from the colony atGdynia harbour (Fig. 3.), total mercury concentrationamounted to 171.6 ng Hg·g−1 d.w. in the liver, 216.2 ngHg·g−1 d.w. in the heart and 303.4 ng Hg·g−1 d.w. in thedown. In the tissues and down of the two fortnight-old herringgull chicks from the same colony, respective mercury concen-trations amounted to 58.1 and 87.9 ng Hg·g−1 d.w. in the liver,21.5 and 32.8 ng Hg·g−1 d.w. in the heart and 409.0 and678.0 ng Hg·g−1 d.w. in the down.
Table 2 Characterisation of total mercury concentrations HgTOT [ng Hg·g−1 d.w.] in the eggs of selected aquatic bird species from the MewiaLacha Sanctuary (Gulf of Gdansk) and the Wloclawek Dam in 2010–2012
Species Samples x±SD MdQ1−Q3
Min–max range
Sandwich tern 92 1154.2±573.2 1021:3791:0−1364:5 244.9–3384.1
Common tern 10 1239.4±571.7 1054:6850:9−1942:1 499.3–2048.5
Herring gull 21 560.0±300.6 416:4365:6−660:2 140.7–1196.1
x arithmetic mean, SD standard deviation, Md– median value, Q1 lower
quartile, Q3 top quartile
Table 3 Characterisation of totalmercury concentrations HgTOT[ng Hg·g−1 d.w.] in the internalcomponents of the egg of selectedaquatic bird species from theMewia Lacha Sanctuary (Gulf ofGdansk) and theWloclawek Damin 2010–2012
Symbols as in Table 2
Species Structure Samples x±SD MdQ1−Q3
Min–max range
Sandwich tern Albumen 58 2940.6±1048.3 2798:22242:1−3553:6 1026.6–7288.6
Yolk 55 160.7±162.5 115:878:4−156:1 18.8–971.9
Common tern Albumen 7 3748.4±1476.2 3786:72523:6−4795:9 1916.7–5949.9
Yolk 7 459.4±473.9 246:9168:6−559:7 143.3–1479.6
Herring gull Albumen 5 1502.5±387.1 1549:71456:8−1652:9 897.1–1956.1
Yolk 5 96.2±12.4 99:798:7−101:3 74.7–106.3
Environ Sci Pollut Res
Author's personal copy

Discussion
The influence of aquatic birds’ diet on Hg concentrationin eggs
The surface water of the Southern Baltic is characterised bylow Hg concentration values (median value 1.2 ng·dm−3)(Bełdowski et al. 2009; Murawiec et al. 2007). Mercury con-centrations measured in river mouths, however, can be almostthree times higher (median value 3.3 ng·dm−3) (Saniewskaet al. 2010). The Wloclawek Dam region is a polluted area,and the Wloclawek Reservoir has a considerable capacity forpollutant accumulation, resulting from the fact that it is thelargest basin receiving industrial and communal pollutiontransported by the Vistula River from the highly industrialisedareas of southern and central Poland (Gierszewski 2008). Itwas calculated that the accumulated sediments in theWloclawek Reservoir contain 46 tonnes of mercury(Bojakowska 1999). In addition, highmercury concentrations,particularly in the muscles of predatory fish caught in thebasin, testify to the constant presence of large amounts ofmercury (Stężycka et al. 2005).
Terns of the Sterna genus are birds whose diet is derivedfrom a water environment. On the basis of pellets found in thetern breeding colony in the Mewia Lacha Sanctuary, it can beconcluded that the birds that nest there feed mainly on smallfish or juvenile fish (predominantly the lesser sand eel, spratand herring, but also garfish, smelt and fish of the Cyprynidaefamily) and invertebrae (Bzoma, unpublished data).
Natural sources of food for herring gulls are fish, insectsand other invertebrae. However, they are also often describedas opportunistic predators and scavengers and, unlike terns,they also like to eat anthropogenic food, found for example inlandfills (Meissner et al. 2007). In the egg incubation andchick feeding period though, most of its diet is constitutedby fish. Roach (Rutilus rutilus) is the food of choice for gullsfrom the Wloclawek Dam colony, the proportion of bream(Abramis brama), carp (Carpinus carpio) and tench (Tincatinca) representing just 5–10 % of their diet. However, while
feeding chicks, there are three times more insects of theCarabidae family included in the diet than usual(Przystalski, unpublished data).
According to stable nitrogen (δ15N) isotope analysis(Fig. 1a), it can be concluded that the herring gull occupiesthe lowest trophic position of the three studied species. Thisspecies is the most strongly linked to land food sources (low-est median δ13C). The common tern occupies the highest tro-phic level, although its food comes from both the land and thesea. The wide range of δ13C values for the common tern indi-cates that these birds collect their food across the largest area(Fig. 1b). The sandwich terns were the most strongly connect-ed to the marine environment, as confirmed by the dataconcerning the distribution of the three studied species and,therefore had a more limited feeding area. While the herringgull and common tern are widespread water birds which canalso be observed in inland areas, the sandwich tern isdistributed exclusively along ocean and sea coasts. This fitsin with the assertion of Burger and Gochfeld (1997) that birdswhich fed on larger or more marine fish have higher levels ofmercury in their eggs and allows for the conclusion that a dietof anthropogenic food and natural food from freshwater envi-ronment was chiefly responsible for the lower mercury con-centrations found in the eggs of these gulls.
It is known that mercury measured in eggs reflects thebirds’ diet during egg formation (Monteiro and Furness1995; Cotin et al. 2011). Moreover, during laboratory exper-iments, Heinz et al. (2009) suggested that all of the examinedCharadriformes (three tern species and two gull species) pre-sented the same sensitivity in embryos to methylmercury, sug-gesting certain similarities in Hgmetabolism in closely relatedspecies such as gulls and terns. Hence, despite differencesresulting from the different physiologies of the studied spe-cies, it may be assumed that environmental conditions and thefood consumed by females will have the greatest influence onthe differentiation of mercury concentrations in the studiedeggs.
In the eggs of terns, Monteiro et al. (1999), Burger andGochfeld (2003) and Bond and Diamond (2009b) assayedtotal mercury concentrations at a level comparable to the con-centrations found in the eggs of terns from the nesting sites inthe Mewia Lacha Sanctuary on the Gulf of Gdansk. Mercuryconcentrations in the eggs of gulls from the St. Lawrence Bay(Lavoie et al. 2010) also displayed similar values to thosefrom the Wloclawek Dam colony (Table 5). It is worth men-tioning here the measurement results obtained by Cotin et al.(2011), who found high mercury concentration values in theEbro delta in Spain. The authors explain that a chemical plantis located 90 km from the river outlet, and it is considered to bethe main source of pollution in the region. Moreover, theyattribute high concentrations in sandwich tern eggs to its su-perior ability to acquire food from the sea in comparison toother species.
Table 4 Characterisation of total mercury concentrations HgTOT [ng Hg·g−1 s.m.] in the external egg components of terns from the nesting colonyin the Mewia Lacha Sanctuary
Species Structure Samples x±SD MdQ1−Q3
Min–maxrange
Sandwichtern
Eggshell 10 0.6±0.5 0:50:2−0:7 0.1–1.6
Membrane 10 21.0±28.6 11:36:1−20:9 3.1–98.8
Eggshell withmembrane
93 50.1±37.0 43:223:4−64:0 4.5–216.6
Commontern
Eggshell withmembrane
10 53.6±26.1 54:332:9−66:1 12.9–110.3
Symbols as in Table 2
Environ Sci Pollut Res
Author's personal copy
Mercury in the albumen, yolk and eggshell
Mercury is unevenly distributed in the particular internal eggcomponents (Table 3). This was pointed out by Bond andDiamond (2009a, b) while studying the eggs of birds fromMachias Seal Island (Canada). They showed that mercuryconcentration in the albumen is even up to 10 times higherthan in the yolk, depending on the species. The higher mercu-ry concentrations in the albumen than in the yolk resultedfrom the dissimilar composition of both structures. The com-position of the albumen is stabilised by the presence of sulf-hydryl groups forming the so-called disulfide bonds. Mercuryhas a very strong affinity to sulfhydryl groups, thus the yolk,being composed mainly of fats, accumulates much less mer-cury. The low correlation factor (rSpearmann=0.35; p<0.05)between mercury concentrations in the albumen and yolk ofthe sandwich tern provides grounds for presuming that thereare different means of mercury transportation into each of thecomponents. The reasons for dissimilar values of theHgAlbumen/HgYolk concentration ratio are not analysed in liter-ature, but it is nevertheless conceivable that, in migratorybirds, they reflect diet diversification related to the regionsinhabited before and during the breeding period.
Adhering underneath the shell protecting the egg, there aretwo membranes surrounding the albumen. The obtained re-sults (Table 4) showed that it is the membranes that are re-sponsible for holding most of the mercury contained in theexternal parts of the egg. The shell is composed mainly of
calcium carbonate, while the membrane is made of keratin.The shell, being built of inorganic substances, accumulates theleast mercury. It is difficult to compare these with other results,owing to the scarcity of available literature.
Embryo and chick
Similarly to adult birds and chicks, the embryos developinginside the eggs accumulate mercury in their livers, kidneysand feathers (Rutkiewicz and Basu 2013). The smallest stud-ied embryo (Fig. 2(I)) had the lowest mercury concentrationsin the muscle tissue of the heart, in the glandular liver tissueand in feathers, as most of the mercury was still in the sur-rounding albumen. Nevertheless, already at the initial stage oflife, the newly formed down had accumulated more mercurythan the heart and liver. As the embryo developed, the heart,liver and down showed an increase in the concentrations ofmercury (Fig. 2(II)), which the system acquired with nutrientsaccumulated in the egg. In the period preceding hatching(Fig. 2(III)), there was to be observed a further increase inHg concentrations in tissues. The particularly significant roleof feathers can be observed at the middle stage of embryodevelopment (Fig. 2(II)), this being the time when the lowestHgHeart/HgDown and HgLiver/HgDown concentration ratios weredetermined (0.2–0.4), indicative of the greatest difference be-tween mercury accumulation in the internal tissues and thedown. The well-documented statement that incorporatingmercury into newly forming feathers is the most effective
Fig. 1 Differences in δ15N (a)and δ13C (b) [‰] in eggs of threebird species (ST sandwich tern,CT common tern, HG herringgull) from the Mewia LachaSanctuary and Wloclawek Dam
Table 5 Mean HgTOT concentrations [ng Hg·g−1 s.m.] in the eggs of the representatives of the Sternidae and Laridae families from other regions ofEurope and from North America
Species Year Study region HgTOT Source
Common tern 1992, 1993, 1995 The Azores, Portugal 1000–1500 Monteiro et al. 1999
Common tern 2000 New Jersey, USA 698–2010 Burger and Gochfeld 20032001 847–1870
2002 977–1580
Common tern 2005 Machias Seal Island, Canada 1016 Bond and Diamond 2009b2006 797
Herring gull 2006, 2007 Gulf of St. Lawrence, Canada 433 Lavoie et al. 2010
Sandwich tern 2006, 2008 Ebro delta, Spain 2727.2 Cotin et al. 2011Common tern 1139.6–4148.9
Environ Sci Pollut Res
Author's personal copy
method of depuration (Burger 1993; Becker et al. 1994;Burger and Gochfeld 1997; Bond and Diamond 2009a) canbe supplemented with the information on effective mercurydetoxification into the down during embryonic life.Immediately before pipping, the nutrient supplies accumulat-ed in the egg become completely exhausted. They are used forthe development of internal organs, as is confirmed byHgHeart/HgDown and HgLiver/HgDown ratios that are highercompared to the second phase of development (0.4–0.8).The results suggest that the embryo can control mercuryaccumulation and at the same time confirm the statement byRutkiewicz and Basu (2013) who showed tha, even at thisearly stage, both heart and liver perform their life functions.The control over mercury accumulation in the embryo is heldby the liver, where demethylation metabolism takes place andby the forming the down, where mercury becomes seques-tered. Both the mechanisms protect other organs where Hgcan potentially become accumulated, one of which being thebrain.
Hg concentration results in the livers and hearts of embryosshow that the most susceptible to the toxic effect of Hg arechicks during pipping and hatching, a feature which has beenpointed out before by other researchers (Burger and Gochfeld1997; Braune 2007; Rutkiewicz and Basu 2013).
Comparing the obtained results to earlier studiesconcerning penguins bred in Gdansk Zoo (Falkowska et al.2013), it can be observed that the concentrations in thefeathers of unhatched herring gull chicks in the last two stagesof embryonic development (middle and late) are similar to theconcentrations determined in the feathers of adult penguins,while Hg concentrations in internal organs are comparable to a3-day-old penguin chick. Herring gull embryos have a muchhigher potential for mercury sequestration in feathers thanpenguins, a fact that indicates the existence of interspeciesdifferences.
Becker et al. (1993) showed that the down forming duringembryonic development may accumulate at least 38 % of themercury from the entire body. Moreover, the down formedwhile inside the egg was found to have higher mercury con-centrations than feathers that develop later, after hatching(Becker et al. 1994). This means that the mercury in the firstdown formed by the embryo is a consequence of mercury
present in the egg, while the mercury in feathers that formafter hatching apparently reflects the body pool of accumulat-ed Hg at the time of feather growth.
Looking for explanations for the variability of total mercu-ry concentrations in the tissues and down of the embryo andtwo2-two-week-old chicks of the herring gull from the nestingcolony in Gdynia harbour (Fig. 3), one needs to point out thedecrease ofmercury in the heart (by 87%) and the liver (57%)after the end of embryonic life. This may be caused by both arapid increase in body mass and mercury removal with guano(Falkowska et al. 2013). A reverse tendency was observed inthe case of down. The feathers of the two 2-week-old herringgull chicks were 1.8 times richer in mercury compared to thefeathers of the embryo. This suggests an increase in the sig-nificance of feathers in system detoxification after hatchingand contradicts the conclusions of Becker et al (1993).
Summary
The studies indicated that seafood consumed by birds poses agreater threat to their offspring than freshwater food or anthro-pogenic food. The eggs of birds obtaining the majority of theirfood from the sea (terns) were characterised by higher mercuryconcentrations.
Owing to the presence of sulfhydryl groups characterisedby a high affinity to Hg, the albumen was found to be the eggcomponent with the highest mercury concentrations in all thestudied bird species. Much lower concentrations were assayed
Fig. 2 Concentrations of totalmercury HgTOT [ng Hg·g−1 s.m.]in the hearts, livers and down ofunhatched herring gull chicks(Larus argentatus) from theWloclawek Dam, in three phasesof embryonic development (Iindicates early, II middle and IIIlate)
Fig. 3 Mean concentrations of total mercury HgTOT [ng Hg·g−1 d.w.] in
the hearts, livers and down of an unhatched chick and 2-week-old herringgull nestlings (Larus argentatus) from the nesting colony in Gdyniaharbour
Environ Sci Pollut Res
Author's personal copy

in yolks, while the shell, composed of inorganic substances,displayed the lowest Hg content.
The increase mercury concentration in tissues and internalorgans of the embryo as it develops is the result of the absorp-tion of supplies accumulated in the egg. Mercury was mosteffectively incorporated into the down forming while insidethe egg. However, the detoxification route into the first downprobably ends before the nutrient supplies in the egg becomefully exhausted and incorporated into the internal organs, asindicated by an increase in the HgHeart/HgDown andHgLiver/HgDown ratios in the final phase of embryonic life(0.4–0.8) in comparison with the preceding phase (0.2–0.4).Once hatched, a herring gull chick underwent rapid detoxifi-cation and, within 2 weeks of life, Hg concentrations haddropped in the internal organs: by 87 % in the heart and by57 % in the liver.
Acknowledgments The present paper was created with the financialsupport of the Polish Ministry of Science and Higher Education withinthe framework of the research project No. N N304 161637.
References
Akearok JA, Hebert CE, Braune BM, Mallory ML (2010) Inter- andintraclutch variation in egg mercury levels in marine bird speciesfrom the Canadian Arctic. Sci Total Environ 408:836–840
Becker PH (1992) Eggmercury levels decline with the laying sequence incharadriiformes. Bull Environ Contam Toxicol 48:762–767
Becker PH, Furness RW, Henning D (1993) Mercury dynamics in youngcommon tern (Sterna hirundo) chicks from a polluted environment.Ecotoxicology 2:33–40
Becker PH, Henning D, Furness RW (1994) Differences in mercury con-tamination and elimination during feather development in gull andtern broods. Arch Environ Contam Toxicol 27:162–167
Bełdowski J,MiotkM, Pempkowiak J (2009)Mercury fluxes through thesediment water interface and bioavailability of mercury in southernBaltic Sea sediments. Oceanologia 51:263–285
BirdLife International (2013) Important Bird Areas factsheet: Vistula riv-er mouth. Downloaded from http://www.birdlife.org/ on 04/01/2013
Bojakowska I, Gliwicz T, Sokołowska G, Strzelecki R, Wołkicz S (1999)Investigation of bottom sediments inWłocławek Reservoir, the prop-osition to solve the problem. Synthesis. Polski Instytut Geologiczny(manuscrupt in Polish)
Bond AL, Diamond AW (2009a) Total and methyl mercury contamina-tions in seabird feathers and eggs. Arch Environ ContamToxicol 56:286–291
Bond AL, Diamond AW (2009b) Mercury concentrations in seabird tis-sues from Machias Seal Island, New Brunswick, Canada. Sci TotalEnviron 407:4340–4347
Braune BM (2007) Temporal trends of organochlorines and mercury inseabirds eggs from the Canadian Arctic, 1975–2003. Environ Pollut148:599–613
Braune BM, Gaskin DE (1987) Mercury levels in Bonaparte’s gulls(Larus Philadelphia) during autumn molt in the Quoddy region,New Brunswick, Canada. Arch Environ Contam Toxicol 16:539–549
Burger J (1993) Metals in avian feathers: bioindicators of environmentalpollution. Rev Environ Contam Toxicol 5:203–311
Burger J, Gochfeld M (1997) Risk, mercury levels, and birds: relatingadverse laboratory effects to field biomonitoring. Environ Res 75:160–172
Burger J, GochfeldM (2003) Spatial and temporal patterns inmetal levelsin eggs of common terns (Sterna hirundo) in New Jersey. Sci TotalEnviron 311:91–100
Burgess NM, Bond AL, Hebert CE, Neugebauer E, Champoux L (2013)Mercury trends in herring gull (Larus argentatus) eggs fromAtlantic Canada, 1972–2008: temporal trend or dietary shift?Environ Pollut 172:216–222
Cotin J, García-Tarrasón M, Sanpera C, Jover L, Ruiz X (2011) Sea,freshwater or saltpans? Foraging ecology of terns to assess mercuryinputs in a wetland landscape: The Ebro Delta. Estuarine. CoastalShelf Sci 92:188–194
Evers DC, Savoy LJ, DeSorbo CR, Yates DE, Hanson W, Taylor KM,Siegel LS, Cooley JH, Bank MS, Major A, Munney K, Mower BF,Vogel HS, Schoch N, Porkas M, Goodale MW, Fair J (2008)Adverse effects from environmental mercury loads on breedingcommon loons. Ecotoxicology 17:69–81
Falkowska L, Reindl AR, Szumiło E, Kwaśniak J, Staniszewska M,BełdowskaM, Lewandowska A, Krause I (2013)Mercury and chlo-rinated pesticides on the highest level of the foodweb as exemplifiedby herring from the southern Baltic and African penguins from zoo.Water Air Soil Pollut 224:1549–1563
Gierszewski P (2008) Koncentracja metali ciężkich w osadach zbiornikawłocławskiego jako wskaźnik hydrodynamicznych warunkówdepozycji. Landform Anal 9:79–82 (in Polish)
Hall ED, Bodaly RA, Fudge RJP, Rudd JWM, Rosenberg DM (1997)Food as the dominant pathway of methylmercury uptake by fish.Water Air Soil Pollut 100:13–24
Heinz GH, Hoffman DJ, Kondrad SL, Erwin CA (2006) Factors affectingthe toxicity of methylmercury injected into eggs. Arch EnvironContam Toxicol 50:264–279
Heinz GH, Hoffman DJ, Klimstra JD, Stebbins KR, Kondrad SL, ErwinCA (2009) Arch Environ Contam Toxicol 56:129–138
Hobson KA, Clark RG (1992a) Assessing avian diets using stable iso-topes I: turnover of 13C in tissues. Condor 94:181–188
Hobson KA, Clark RG (1992b) Assessing avian diets using stable iso-topes II: factors affecting diet–tissue fractionation. Condor 94:189–197
IMGW (2011) In: Maciejewski M, Ostojski MS, Walczykiewicz T (eds)Dorzecze Wisły: monografia powodzi – maj – czerwiec 2010.IMGW – PIB, Warszawa, in Polish
Kelly JF (2000) Stable isotopes of carbon and nitrogen in the study ofavian and mammalian trophic ecology. Can J Zool 78:1–27
Kenov KP, Hines RK,MeyerMW, Suarez SA, Gray BR (2010) Effects ofmethylmercury exposure on the behavior of captive–reared commonloon (Gavia immer) chicks. Ecotoxicology 19:933–944
Lavoie RA, Champoux L, Rail J, Lean DRS (2010) Organochlorines andbrominated flame retardants and mercury levels in six seabird spe-cies from the Gulf of St. Lawrence (Canada): relationships withfeeding ecology and molt. Environ Pollut 158:2189–2199
Meissner W, Staniszewska J, Bzoma S (2007) Abundance, species com-position and age structure of gulls Laridae in the Gulf of Gdańskarea during non-breeding season. Notatki Ornitologiczne 48:67–81(in Polish)
Meissner W, Bzoma S, Zięcik P, Wybraniec M (2014) Nesting ofSandwich Tern Sterna sandvicensis in Poland in 2006–2013. OrnisPolonica 55:96–104 (in Polish)
Minagawa M, Wada E (1984) Stepwise enrichment of 15N along foodchains: further evidence and the relation between δ15N and animalage. Geochim Cosmochim Acta 48:1135–1140
Monteiro LR, Furness RW (1995) Seabirds as monitors of mercury in themarine environment. Water Air Soil Pollut 80:851–870
Monteiro LR, Granadeiro JP, Furness RW, Oliveira P (1999)Contemporary patterns of mercury contamination in the
Environ Sci Pollut Res
Author's personal copy

Portuguese Atlantic inferred frommercury concentrations in seabirdtissues. Mar Environ Res 47:137–156
Murawiec D, Gajecka A, Bełdowska M, Falkowska L (2007)Investigation on mercury concentration levels in coastal and off-shore waters of the Gulf of Gdańsk. Oceanol Hydrobiol Stud 36:83–97
Peterson BJ, Fry B (1987) Stable isotopes in ecosystem studies. AnnuRev Ecol Syst 18:293–320
Rumbold DG, Niemczyk SL, Fink LE, Chandraesekhar T, Harkanson B,Laine KA (2001) Mercury in eggs and feathers of great egrets(Ardea albus) from the Florida Everglades. Arch Environ ContamToxicol 41:501–507
Rutkiewicz J, Basu N (2013) Methylmercury egg injections: part 1—tissue distribution of mercury in the avian embryo and hatchling.Ecotoxicol Environ Saf 93:68–76
Saniewska D, Bełdowska M, Bełdowski J, Saniewski M, Kwaśniak J,Falkowska L (2010) Distribution of mercury in different environ-mental compartments in the aquatic ecosystem of the coastal zone ofSouthern Baltic Sea. J Environ Sci 22:1144–1150
Saniewska D, Bełdowska M, Bełdowski J, Saniewski M, Falkowska L(2014) Mercury loads into the sea associated with extreme flood.Environ Pollut 191:93–100
Scheuhammer AM, Perrault JA, Bond DE (2001) Mercury, methylmer-cury, and selenium concentrations in eggs of common loons (Gaviaimer) from Canada. Environ Monit Assess 72:79–94
Scheuhammer AM, Meyer MW, Sandheinrich MB, Murray MW (2007)Effects of environmental methylmercury on the health of wild birds,mammals, and fish. Ambio 36:12–18
Splading MG, Frederick PC, McGill HC, Bouton SN, Richey LJ,Schumacker IM, Blackmore CGM, Harrison J (2000) Histologic,neurologic, and immunologic effects of methylmercury in captivegreat egrets. J Wildl Dis 36:423–435
Steele KW, Daniel RM (1978) Fractionation of nitrogen isotopes by an-imals: a further complication to the use of variations in the naturalabundance of 15N for tracer studies. J Agric Sci 90:7–9
Stężycka E, Bzdęga J, Pawlikowska K, Siwicki A (2005) Content ofmercury in fish from Wisła between 1998 and 2002. Probl HigEpidemiol 86:27–29 (in Polish)
Thompson DR (1996) Mercury in birds and terrestrial mammals. In:Bayer WN, Heinz GH, Redmon-Norwood AW (eds)Environmental contaminants in wildlife: interpreting tissue concen-trations. SETAC Special Publication Series. CRC Press, BocaRaton, pp 341–356
Walsh PM (1990) The use of seabirds as monitors of heavy metalsin the marine environment. In: Furness RW, Rainbow PS (eds)Heavy metals in the marine environment. CRC Press, NewYork
Wängberg I, Munthe J, Pirrone N, Inverfeldt A, Bahlman E, Costa P,Ebinghaus R, Feng X, Ferrara R, Gardfelt K, Kock H, LanzillottaE, Mamane Y, Mas F, Melamed E, Osnat Y, Prestbo E, Sommar J,Schmolke S, Dpain G, Spovieri F, Tuncel G (2001) Atmosphericmercury distribution in Northern Europe and in Mediterranean re-gion. Atmos Environ 35:3019–3025
Wiener JG, Krabbenhoft DP, Heinz GH, Scheuhammer AM (2003)Ecotoxicology of mercury. In: Hoffman DJ, Rattner BA, BurtonGA Jr, Cairns J Jr (eds) Handbook ecotoxicology, sd ed. CRCPress, Boca Raton, pp 409–463
Yin X, Xia L, Sun L, Luo H, Wang Y (2008) Animal excrement: apotential biomonitor of heavy metal contamination in the marineenvironment. Sci Total Environ 399:179–185
Zarządzenie Ministra Ochrony Środowiska, Zasobów Naturalnych iLeśnictwa z dnia 9 października 1991 r. w sprawie uznania zarezerwaty przyrody (M.P. Nr 38, item 273) (in Polish)
Environ Sci Pollut Res
Author's personal copy




















