
Memo TV3_V1 - CCMA
228
-
Upload
khangminh22 -
Category
Documents
-
view
0 -
download
0
Transcript of Memo TV3_V1 - CCMA


© 2006 Fundació La Marató de TV3
Direcció: Miquel Vilar
Coordinació i redacció:
Begonya Garcia i Batllori
Correcció i traduccions al castellà:
Francesc X. Navarro
Traduccions a l’anglès: Peter Harvey
Disseny gràfic: Zink comunicació
Impressió: Gramagraf
Tiratge: 1.100
Dipòsit legal:
L’elaboració d’aquesta Memòria ha estat
possible gràcies al suport d’Abertis.
Fundació La Marató de TV3
Ganduxer, 117
08022 Barcelona
Tel. 93 444 48 30
www.fundaciomaratotv3.cat
a/e: [email protected]

Presentacions 5
1. Fundació La Marató de TV3
Introducció 9
Òrgans de govern: Patronat i Comissió Assessora Científica 10
2. Tancament de comptes 13
3. La Marató de 2004
Introducció 15
Sensibilització i difusió 18
Comunicació 32
Organització i logística 33
Patrocini 39
4. Recerca biomèdica
Introducció 41
Convocatòria d’ajuts 42
Avaluació dels projectes de recerca 44
Adjudicació dels ajuts 49
Seguiment dels projectes 52
Finalització dels projectes. Simposium 53
La Marató del 1998. Resum de les memòries finals 57
Índex

PRESENTACIONS | 5
Celebrada La Marató s’obre un llarg pro c é s
de treball per a la Fundació. Es comença
amb el tancament dels donatius i amb la
convocatòria pública d’ajuts als projectes de
re c e rca que hauran de tractar sobre la
malaltia protagonista d’aquella edició. Un cop
avaluats tots aquests projectes, se seleccio-
nen els millors i es doten econòmicament. La
Fundació fa un seguiment curós de cada
p rojecte i examina, anualment, les memòries
elaborades pels investigadors, que tindran un
termini de tres anys per presentar els
resultats de la feina. Finalment, per tal que la
ciutadania conegui per a què han servit els
seus diners, la Fundació, amb la participació
dels investigadors, presenta, en un acte
públic, els avenços aconseguits. És llavors,
realment, quan es tanca el circuit que obre ,
cada any, el programa La Marató de TV3.
La Memòria que us presento recull tot aquest
p rocés. Durant l’exercici del 2004 va tenir lloc
la tretzena edició de La Marató i val a dir que
aquesta edició, dedicada al càncer, va tenir
un èxit de participació espectacular. Ha estat,
e n t re totes les celebrades, La Marató rècord
quant a recaptació. El tema triat va motivar
que científics de molt diversos àmbits hi
p resentessin 260 projectes, que van ser
avaluats per 169 avaluadors.
En el decurs d’aquest any, a més, la
Fundació ha celebrat l’acte de lliurament
d’ajuts als investigadors que van concórrer a
la convocatòria de La Marató 2003, dedicada
a les malalties respiratòries cròniques i,
finalment, el V Simposium, per retre comptes
a la societat dels avenços aconseguits en la
recerca sobre la diabetis, malaltia que va
centrar el programa de 1998.
Com veieu, la Memòria de tot allò que es
fa durant un any no és únicament una
enumeració de treballs començats i acabats
en els dotze mesos corresponents. Es tracta
d’una cadena molt més llarga i rigorosa que
exigeix constància en un seguiment lent i a la
força pacient en la visualització de resultats.
Comprèn les accions dutes a terme des del
juliol de 2004 fins al juny de 2005, per un
equip de gent que aprofito per felicitar. La
més important, sens dubte, és el programa
que dóna cos i passió a aquest trajecte de
gran recorregut. La Marató de TV3 s’ha
convertit en un d’aquells fenòmens que
configuren l’espai imaginari d’un col·lectiu
implicat, optimista i orgullós d’assumir els
reptes socials d’una manera diferent. La
Corporació Catalana de Ràdio i Televisió
aporta el mecanisme i configura l’engranatge
de servei públic pel qual va ser creada. Però
sense la participació ciutadana, que és al
capdavall el veritable motor de la cursa i de
qualsevol recerca biomèdica posterior, ni les
activitats referides en aquesta Memòria, ni
tots els avenços aconseguits fins al dia
d’avui haurien estat possibles. Us presento,
doncs, una petita mostra d’allò que tots
plegats hem aconseguit.
Trajecte de llarg
recorregut
Joan Majó, director general de la Corporació Catalana de Ràdio i Televisió
i president de la Fundació La Marató de TV3

FUNDACIÓ LA MARATÓ DE TV3
Aquesta Memòria comprèn el període juliol
de 2004-juny de 2005. Ha estat un període
ple d’esdeveniments molt positius en l’evolu-
ció del projecte de la Fundació La Marató
de TV3.
Primerament, en un acte públic que va tenir
lloc al Rectorat de la Universitat de
Barcelona, es van adjudicar les ajudes als
projectes de La Marató del 2003 dedicada a
les malalties respiratòries cròniques.
En segon lloc, el 18 de desembre de 2004
es va celebrar la tretzena edició de La
Marató de TV3, dedicada al càncer. El
programa va ser tot un èxit d’audiència i de
recaptació de recursos. S’hi van recollir 9
milions d’euros.
Després de la convocatòria pública de
projectes de recerca, un panel de 169
experts internacionals va avaluar els 260
projectes que s’hi havien presentat.
A més d’aquests actes fonamentals en la
vida anual de la Fundació La Marató de TV3,
també s’han produït una sèrie de canvis
significatius en l’assessorament científic
del Patronat. D’una banda, la Comissió
Assessora Científica del Patronat s’ha
ampliat, tot donant cabuda a científics
rellevants de tots els àmbits territorials de
Catalunya; més específicament: s’hi han
incorporat experts procedents de Girona,
Lleida, Tarragona i de la rodalia de
Barcelona. D’una altra banda, s’han introduït
millores en la metodologia d’avaluació del
tema per a la jornada anual de La Marató de
TV3. Finalment, el Patronat va rebre la
proposta de la Comissió Assessora Científica
d’implementar un nou procediment per a la
selecció del tema –“la crida pública”–, amb
la intenció de concedir un paper més notori a
la participació ciutadana.
En definitiva, el projecte de la Fundació La
Marató de TV3 s’actualitza d’una manera
continuada, tot dotant-se d’un rigor i una
participació més grans, atributs necessaris i
que suposen una resposta adequada al
nivell de solidaritat i generositat proporciona-
des pels ciutadans de Catalunya.
La Fundació,projecte de
renovació constant
D r. José J. Navas, vicepresident segon de la Fundació La Marató de TV3
i coordinador de la Comissió Assessora Científica

Quan el Patronat de la Fundació va acceptar
la proposta de la Comissió Assessora
Científica de dedicar La Marató del 2004 al
càncer, ja suposava que el tema era molt
adequat per tenir una bona resposta de la
ciutadania. L’antecedent del 1996, en què La
Marató va tractar, també, del càncer, ho feia
preveure, ja que els resultats van ser, alesho-
res, importants.
Es tracta d’una malaltia cruel i incurable,
s o b re la qual plana una gran sensibilitat
social. Per tant, tot feia pensar en la bondat
de la decisió, però difícilment podíem imaginar
els resultats d’aquesta tretzena edició.
És conegut que els resultats de La Marató de
TV3 estan en consonància amb les t e l e t h o n s
franceses, angleses, italianes o suïsses, no
només quant a mobilització social, sinó
també en audiència, i que recapta tants
recursos com la que més si es ponderen els
factors de població i hores de pro g r a m a .
És per això que hom va pensar que si es
mantenien els paràmetres habituals de La
Marató, ens podíem considerar satisfets.
Per això, quan vam tancar l’edició 13 de
La Marató amb una xifra de 8.712.000
e u ros, no ens en sabíem avenir. Per primera
vegada superàvem la xifra mítica de 1.000
milions de pessetes!
Però, és cert que hi havia moltes re f e r è n c i e s
anteriors que podien dur a pensar en un
resultat com aquest, començant pel nou
equip del programa, que va saber connectar
perfectament amb l’univers del món Marató.
En la campanya de sensibilització prèvia
s’havien fet 2.880 conferències a escoles i
c e n t res cívics per 220 professionals de la
medicina; els ajuntaments adherits van penjar
més de 900 pancartes, es van celebrar 502
activitats populars i 114.728 donants,
particulars i empreses van fer la seva aporta-
ció... Mai La Marató havia tingut una re s p o s t a
tan decidida com aquesta, sense oblidar- n o s
dels 592.000 euros aconseguits de diverses
e m p reses en concepte de patrocini que
cobrien, amb escreix, les despeses generals
de la Fundació.
Uns resultats tan positius ens obliguen a tenir
molt present el suport decidit de Bassat, La
Caixa, PricewaterhouseCoopers, Telefónica i
Tradia, empreses col·laboradores que any
rere any estan amb La Marató. Com també
de Barcelona de Serveis Municipals –gestora
del recinte del Palau Sant Jordi de
Barcelona–, Fira de Girona, Universitat de
Lleida i Port de Tarragona, que cedeixen els
seus espais.
I el reconeixement al Departament de
Benestar i Família, soci imprescindible en
l’organització del voluntariat i de la campanya
de sensibilització, i a l’Agència d’Avaluació
de Tecnologia i Recerca Mèdiques del
Departament de Salut, cooperadora
necessària en l’avaluació dels 260 projectes
de recerca biomèdica sobre el càncer
presentats i de les memòries científiques
anuals corresponents a convocatòries
anteriors.
PRESENTACIONS | 7
El tretze ens vadonar sort
Miquel Vi l a r, director de la Fundació La Marató de TV3


La Fundació La Marató de TV3 va ser creada
el 1996 per la Corporació Catalana de Ràdio
i Televisió amb dues finalitats principals:
· Fomentar i promoure la recerca biomèdica
d’excel·lència.
· Sensibilitzar sobre les malalties de què es
tracta en La Marató de TV3.
El foment i la promoció de la recerca
biomèdica d’excel·lència, entesa com la que
fan els investigadors en centres de recerca
acreditats, es desenvolupen, fonamental-
ment, gràcies als ingressos obtinguts amb el
programa La Marató de TV3.
La sensibilització i difusió de les malalties es
du a terme per mitjà de conferències
informatives a centres educatius i cívics,
campanyes i actes de participació ciutadana
en general.
El Departament d’Economia i Finances de la
Generalitat i PricewaterhouseCoopers
s’encarreguen de l’auditoria anual de la
Fundació.
FUNDACIÓ LA MARATÓ DE TV3 | 9
Fundació La Marató de TV3

El Patronat
És l’òrgan de govern i d’administració de la
Fundació i té la potestat decisòria de la
Fundació.
El Patronat, durant aquest exercici, va
aprovar, a proposta de la Comissió
Assessora Científica, la malaltia objecte de
La Marató del 2005: l’Alzheimer i altres
malalties del cervell, les bases de la
convocatòria d’ajuts a la recerca biomèdica
sobre el càncer i l’adjudicació de recursos
obtinguts en La Marató 2003 als projectes
de recerca sobre les malalties respiratòries
cròniques.
PRESIDENT:
Sr. Joan Majó
Director general de la Corporació Catalana
de Ràdio i Televisió
VOCALS:
Sra. Marta Aymerich
Directora del Consell Interdepartamental
de Recerca i Innovació Tecnològica (CIRIT)
Sr. Francesc Escribano
Director de Televisió de Catalunya.
Vicepresident primer
Sr. Marc Mateu
Director del programa La Marató de TV3
Sr. Jordi Menéndez
Vocal del Consell d'Administració de la
Corporació Catalana de Ràdio i Televisió
Sra. Montserrat Minobis
Directora de Catalunya Ràdio
Sr. José J. Navas Palacios
Departament de Salut. Vicepresident segon
Sr. Miquel Reniu
Vocal del Consell d’Administració de la
Corporació Catalana de Ràdio i Te l e v i s i ó
Sr. Antoni Segarra
Secretari general del Departament de
Benestar i Família
Sr. Esteve Mañá
Secretari, no patró
DIRECCIÓ EXECUTIVA
Sr. Miquel Vilar
Òrgans de govern
FUNDACIÓ LA MARATÓ DE TV3

FUNDACIÓ LA MARATÓ DE TV3 | 11
La Comissió Assessora Científica
Òrgan consultiu del Patronat. Els seus
membres –representants de les institucions
cientificomèdiques més importants del país–
es reuneixen periòdicament per convocatòria
del seu coordinador.
Des d’aquesta Comissió es proposen les
línies temàtiques per al programa La Marató
de TV3, els metges assessors del programa,
les accions per promoure la recerca
d’excel·lència i les bases per a la convocatò-
ria pública d’ajuts a la recerca.
Durant aquest exercici, la Comissió
Assessora Científica va proposar al Patronat
que l’Alzheimer i altres malalties del cervell
centressin La Marató del 2005, les bases de
la convocatòria per a la presentació de
projectes sobre el càncer i l’adjudicació dels
recursos als projectes finançats amb els fons
de La Marató del 2003, dedicada a les
malalties respiratòries cròniques.
Dr. José J. Navas Palacios
Departament de Salut (coordinador)
Dr. Josep M. Antó
Director de l’Institut Municipal d’Investigació
Mèdica
Dra. Marta Aymerich
Directora del Consell Interdepartamental
de Recerca i Innovació Tecnològica (CIRIT)
Dr. Lluís Blanch Torra
Director científic del Parc Taulí. Sabadell
Dr. Josep M. Borràs
Director científic de l’Institut Català d’Oncologia
Dr. Juan Emilio Feliu Albiñana
Director general de l’Institut de Recerca de
l’Hospital Universitari de la Vall d’Hebron
Dr. Alberto de Leiva Hidalgo
Director de l’Institut de Recerca de l’Hospital
de la Santa Creu i Sant Pau
Sr. Marc Mateu
Director del programa La Marató de TV3
Dr. Xavier Matias Guiu
Director científic de l’Hospital Arnau de Vi l a n o v a
Dr. Romà Pallarés
Director científic de la Fundació August Pi i
Sunyer
Dr. Ricardo Pujol
Director científic de l’Hospital Universitari
Germans Trias i Pujol
Dr. Wilfredo Ricart Engel
Cap de la Unitat d’Endocrí de l’Hospital Josep
Trueta de Girona
Dr. Joan Rodés
President de la Fundació Privada Clínic
per a la Recerca Biomèdica
Dra. Rosa Solà Alberich
Vicerectora d’Investigació i Institucions
Sanitàries de la Universitat Rovira i Virgili de
Ta r r a g o n a
Sr. Miquel Vilar
Director executiu de la Fundació La Marató de TV3
Dr. Ernest Viñolas
Comitè científic de la Fundació J. Gol i Gurina


TANCAMENT DE COMPTES | 13
La Fundació destina els fons recollits durant
el programa La Marató de TV3 a les dues
finalitats per a les quals va ser creada: el
foment de la re c e rca biomèdica d’excel·lència
i la posada en marxa de campanyes de
sensibilització i de difusió sobre les malalties
de què tracta el pro g r a m a .
En l’exercici 2004-2005, La Fundació va
obtenir un total de 9.022.034 euros,
provinents de donacions, patrocinis i altres
i d’interessos bancaris.
Els comptes anuals de l’exercici que
presentem són auditats per
PricewaterhouseCoopers Auditors i
pel Departament d’Economia i Finances.
Els diners procedents de donatius
s’esmercen, en la seva totalitat, en les
finalitats fundacionals de suport a la
recerca biomèdica i a la sensibilització
sobre la malaltia de què tracta La
Marató.
Les despeses d’administració de la
Fundació són cobertes amb els interes-
sos bancaris que generen els donatius
i, si la xifra obtinguda és insuficient, es
completa amb els recursos obtinguts
de patrocini.
Tancament
de comptes
Donatius 8.130.292,93
Patrocini i altres 584.751,84
Interessos bancaris i altres 306.989,09
TOTAL INGRESSOS 9.022.033,86
Ingressos
Aplicacions
Finalitats fundacionals (A i B): 8.598.169,39 95,30 %
A) Ajuts a projectes de recerca en càncer 7.730.478,60 85,68 %
B) Activitats de sensibilització i difusió 867.690,79 9,62 %
Despeses d'administració 423.864,47 4,70 %
TOTAL APLICAT 9.022.033,86 100,00 %


LA MARATÓ 2004 | 15
El Patronat de la Fundació va acordar que La
Marató de TV3 del 2004 tractés sobre el
càncer, tot acceptant la proposta de la
Comissió Assessora Científica.
La tretzena edició de La Marató va aconse-
guir una recaptació rècord de 8.715.045
euros, mitjançant els donatius, patrocinis i
altres fonts d’ingressos.
Amb una durada de prop de 14 hores, La
Marató de TV3 del 2004, celebrada el 19 de
desembre, va tenir una audiència televisiva
mitjana del 6,5%, és a dir, una mitjana de
410.000 espectadors, amb una quota mitjana
del 23,1%.
En aquesta edició, el nou equip de programa
va incorporar alguns canvis. Entre els més
significatius: un presentador únic, Ramon
Pellicer, que va conduir tot el programa; la
renovació musical, a càrrec de Marc Parrot,
o l’organització d’una activitat pròpia del
programa a l’Estació de França de
Barcelona, encapçalada per una escultura
gegant en forma de lletra M, que es va anar
omplint de les fotografies que tot al llarg de
la jornada van anar duent-hi els ciutadans.
La Marató 2004:
càncer
Donacions telefòniques* 91.285 5.728.252,71 �
Servicaixa 5.734 273.479,82 �
Internet, transferències
i ingressos directes 17.709 2.128.560,40 �
Patrocinis i altres – 584.751,84 �
TOTAL 8.715.044,77 �
Canals de donatius Nre. de donacions Euros
* Femarec destrueix els impresos
que emplenen els voluntaris el dia
de La Marató amb les dades
personals dels donants, un cop
s’han comptabilitzat els imports i
s’han resolt les incidències que hi
hagin pogut sorgir.
Aquesta acció es fa en compli-
ment del que estableix la Llei
orgànica 15/1999, de 13 de
desembre, de protecció de dades.


LA MARATÓ 2004 | 17
Recaptació de les tretze edicions de La Marató de TV3
Leucèmia 1992
Síndrome de Down 1993
Càncer 1994
Cor 1995
Cervell 1996
Genèriques hereditàries 1997
Diabetis 1998
Transplantaments 1999
Malalties mentals 2000
Sida 2001
Malalties Inflamatòries cròniques 2002
Malalties respiratòries cròniques 2003
Càncer 2004
1.230.128
2.352.669
3.035.331
2.040.443
4.107.795
4.172.090
3.945.421
4.685.110
4.511.808
4.653.496
4.518.315
4.279.265
8.715.045
Hores de programa 14 14
Audiència mitjana d’espectadors 410.000 298.000
Audiència mitjana (%) 6,5 4,9
Quota mitjana de pantalla (%) 23,1 18,9
Hores de programa especial a Catalunya Ràdio 4 4
Dades generals de La Marató de TV3 2004 2003

FUNDACIÓ LA MARATÓ DE TV3
Sensibilització i difusióLa Fundació participa, conjuntament amb el
Departament de Màrqueting de TV3 i de la
D i recció del programa La Marató de TV3, en
l’elecció de la imatge, els eslògans i el material
de difusió de la campanya de sensibilització,
difusió i publicitat de La Marató.
P u b l i c i t a t
Bassat Ogilvy&Mather Comunicación va ser
l’agència de publicitat responsable de la
creativitat de la campanya de l’edició 2004.
Campanya de sensibilització i difusió
La campanya es va iniciar el dia 2 de
novembre de 2004 i va anar adreçada a
centres educatius i cívics del territori català.
Es van impartir un total de 2.858 conferèn-
cies, repartides entre escoles i associacions,
donades per 220 professionals de la medici-
na, que prèviament havien estat formats per
la Fundació en una sessió informativa a
càrrec d’un expert en càncer, i en la qual
també se’ls va lliurar una documentació
elaborada especialment i un vídeo divulgatiu
que els va servir de suport en les xerrades.
Se’n van editar un total de 3.000 còpies en
VHS i 150 en DVD.
El Departament de Benestar i Família va
col·laborar en la coordinació d’aquesta
campanya.
Nombre de conferències escolars
Genèriques hereditàries 1997
Diabetis 1998
Transplantaments 1999
Malalties mentals 2000
Sida 2001
Malalties Inflamatòries cròniques 2002
Malalties respiratòries cròniques 2003
Càncer 2004
1.063
1.357
1.595
1.883
2.497
2.226
2.650
2.858

LA MARATÓ 2004 | 19
Un any més, TVC i la Fundació, juntament
amb el Departament de Benestar i Família
van convocar els centres educatius al
concurs Pinta La Marató per al foment del
coneixement dels joves sobre el càncer i de
l’esperit solidari que representa participar en
La Marató. En aquesta edició s’hi van
presentar 52 murals.
El grup de 1r CFGS d’Integració Social de
l’IES Ronda de Lleida va ser el guanyador
d’aquesta edició. Les aules de 1r B d’ESO del
Col·legi Santa Te resa de Jesús de Tarragona i
de 3r A i B d’ESO del Col·legi Calassanç
Escolapis Sant Martí de Barcelona, en van
quedar finalistes.El jurat estava format pels
pintors Elisabet Sabala i Joan Pere Vi l a d e c a n s ,
per Jaume Forés, del Departament de
Benestar i Família, i pels directors del pro g r a-
ma La Marató de TV3 i de la Fundació, Marc
Mateu i Miquel Vi l a r, re s p e c t i v a m e n t .
El premi va consistir en un cap de setmana a
l’alberg de la Generalitat l’Escala-Empúries per
a tots els alumnes participants en el mural,
sufragat pel Departament de Benestar i Família.
Adhesió dels ajuntaments
La promoció social de La Marató compta
amb el suport dels ajuntaments catalans que
dies abans del programa hi participen mitjan-
çant les pancartes que col·loquen als seus
carrers, tot motivant els ciutadans a
col·laborar.
A Albanyà, Alcoletge, Artés, Avinyonet del Penedès
B Barcelona, Bellaguarda, Bellmunt del Priorat, Blanes, Bràfim
C Calaf, Campdevànol, Canejan, Capellades, Castellbell i el Vilar, Castellet i
la Gornal, Castelló de Farfanya, Castelló d'Empúries, Cervera, Collbató,
Constantí, Corbera de Llobregat, Cornellà de Terri
E el Catllar, el Perelló, el Prat de Llobregat, el Soleràs
F Figueres, Fogars de la Selva, Fogars de Montclús
H Hostal Nou de Bianya, Hostal Nou de Bianya, Hostalric
L l’Albiol, la Pobla de Segur, L'Arboç, les Planes d'Hostoles, l'Espluga de
Francolí
M Masies de Voltregà, Masllorenç, Masquefa, Molins de Rei, Muntanyola
O Orpí
P Palol de Revardit, Pinell del Solsonès, Preixana, Premià de Mar
Q Querol
R Regencós, Riells i Viabrea, Riudellots de la Selva, Roquetes
S Sabadell, Salt, Sant Boi de Llobregat, Sant Celoni, Sant Cugat Sesgarrigues,
Sant Esteve Sesrovires, Sant Jaume dels Domenys, Sant Joan de Mediona,
Sant Joan Despí, Sant Martí Sarroca, Sant Mori, Sant Pere de Vilamajor,
Sant Sadurní d'Anoia, Sant Salvador de Guardiola, Sant Vicenç de Montalt,
Santpedor, Sarroca de Bellera
T Torregrossa, Tremp
V Vall Moll, Vallirana, Valls, Vandellós, Viladasens, Viladecans, Vilalba Sasserra,
Vilallonga del Camp
X Xerta
Municipis adherits per primera vegada a la campanya de les pancartes
Mural guanyador de la sisena edició del concurs Pinta La Marató.

FUNDACIÓ LA MARATÓ DE TV3
Almacelles IES Canigó 4t ESO C
Badalona ASPACE - Associació de la Paràlisi Centre d'Integració SocialCerebral
Col·legi Maristes Champagnat 3r ESO A
Col·legi Maristes Champagnat 3r ESO B
Col·legi Maristes Champagnat 3r ESO C
Barcelona Col·legi Calassanç -
Escolapis de Sant Mar tí 3r A i 3r B ESO
Escola Sant Medir 1r ESO
IES Maria Lluïsa Cura 1r Batxilllerat
IES Maria Lluïsa Cura 2n Batxillerat
Cadaqués SES Cap de Creus 1r ESO
Cassà de la Selva La Salle Cassà 4t ESO A
La Salle Cassà 4t ESO B
El Masnou Escola Bergantí 1r ESO
Girona Col·legi Bell-lloc del Pla 1r C ESO
Col·legi Bell-lloc del Pla 2n B ESO
Col·legi Bell-lloc del Pla 3r A ESO
Col·legi Bell-lloc del Pla 3r C ESO
Col·legi Bell-lloc del Pla 4t C ESO
La Selva del Camp SES Joan Puig i Ferreter 3r ESO
La Seu d'Urgell Càritas Diocesana d'Urgell Unitat d'Escolarització Compartida -ESO
Lleida IES Ronda CFGS 1r Animació Sociocultural
IES Ronda CFGS 1r Integració Social
IES Ronda CFGS 2n Animació Sociocultural
IES Ronda CFGS 2n Educació Infantil grup A
IES Ronda CFGS 2n Educació Infantil grup B
Mataró Escola Sol Ixent 4t ESO
Móra d'Ebre Col·legi Santa Teresa 2n ESO
Col·legi Santa Teresa 2n ESO
Navàs Escola Diocesana de Navàs 3r ESO A
Olot Escola Pia d'Olot 2n ESO A
Escola Pia d'Olot 2n ESO B
Escola Pia d'Olot 3r ESO B
Reus Col·legi La Presentació de Reus 2n ESO A
IES Salvador Vilaseca 2n ESO A
IES Salvador Vilaseca 2n ESO B
IES Salvador Vilaseca 3r ESO B
IES Salvador Vilaseca 3r ESO C
Roda de Ter IES Miquel Martí i Pol 3r ESO A1
IES Miquel Martí i Pol 3r ESO A2
IES Miquel Martí i Pol 3r ESO B1
IES Miquel Martí i Pol 3r ESO B2
IES Miquel Martí i Pol 3r ESO C1
IES Miquel Martí i Pol 3r ESO C2
IES Miquel Martí i Pol 3r ESO D1
IES Miquel Martí i Pol 3r ESO D2
Sabadell Centre de Secundària 1r CFGM d'Equips i Instal·lacions
Jaume Viladoms electròniques
Centre de Secundària CFGM Cures Auxiliars Infermeria
Jaume Viladoms
Centre de Secundària CFGM Gestió Administrativa
Jaume Viladoms
IES Castellarnau de Sabadell 2n Cures Auxiliars d'Infermeria
Tarragona Col·legi Santa Teresa de Jesús 1r ESO A
Col·legi Santa Teresa de Jesús 1r ESO B
Vilafranca del Penedès
Col·legi Sant Ramon de Penyafort 4t A ESO
Col·legi Sant Ramon de Penyafort 4t B ESO
Població Centre Curs
Centres educatius participants en el VI concurs Pinta La Marató

LA MARATÓ 2004 | 21
Els ajuntaments que es van adherir a La
Marató en l’edició 2004 van ser 527, dels
quals 110 ho van fer per primer cop. Les
pancartes penjades van ser 966.
A més d’aquesta crida, per aprovació del Ple
també hi va haver consistoris que van fer una
aportació econòmica directa. En aquesta
ocasió van fer-ho un total de 13 municipis:
l'Ametlla del Vallès, Banyeres del Penedès,
Benissanet, Casserres, Fornells de la Selva,
la Garriga, el Pinell de Brai, la Pobla de
Mafumet, Premià de Dalt, Salou, Sant
Llorenç d'Hortons, Sant Martí Centelles i
Santa Maria de Martorelles.
Adhesió dels teatres
La participació social per a la lluita contra les
malalties que centren La Marató també
preocupa les companyies de teatre i les
sales d’exhibició, que la nit abans del progra-
ma fan la lectura d’un manifest de suport.
En l’edició 2004, les sales on es va llegir
aquest manifest van ser: Brossa Espai
Escènic, Teatre Nacional de Catalunya,
Versus Teatre, SAT! Sant Andreu Teatre,
Teatre Tívoli, Club Capitol (Sala I i II), Teatre
Novedades, Teatre Borràs, Romea, Condal,
Villarroel Teatre, Teatre Auditori de
Granollers i Teatre Zorrilla de Badalona.
Activitats populars
El mateix dia de La Marató, però també en
les dates prèvies i posteriors a l’emissió del
programa, centenars d’associacions i entitats
de Catalunya organitzen diverses activitats
populars de suport, que apleguen multitud
de ciutadans. La Fundació els dóna suport
amb material de divulgació.
En l’edició de La Marató del 2004, es van
celebrar 502 actes festius, organitzats per
1.716 associacions i entitats del territori, que
van recaptar 589.269,51 euros.
Alt Urgell
Alt Empordà
BaixEmpordà
Gironès
Selva
VallèsOccidental
Maresme
Osona
Garrotxa Pla del’Estany
Ripollès
Cerdanya
VallèsOriental
Bages
Barcelonès
Baix Llobregat
Anoia
Alt Penedès
Garraf
BaixPenedès
Alt Camp
Conca deBarberà
TarragonèsBaixCamp
Baix Ebre
Montsià
Terra AltaRiberad’Ebre
Priorat
Segrià
Plad’Urgell Urgell
Segarra
Noguera
Solsonès
Pallars Sobirà
Pallars Jussà
Garrigues
Val d’Aran
Alta Ribagorça
Andorra
Berguedà
11
8
8
5
6
7
23
27
2 7
47
17
222
24
11
11
8
9
17
5
811
8
2
3 1
8
8
6
2 9
18
7
2
7
1
1
7
1
13
Activitats populars per comarq u e s
1
4
4

FUNDACIÓ LA MARATÓ DE TV3
Activitats populars celebrades per a La Marató de 2004
Alt Camp Alcover Ajuntament
Bràfim La Cleca
Cabra del Camp Associació de Dones de Cabra del Camp
Milà, el Ajuntament
Nulles Col·lectiu de Dones de Nulles
Cooperativa Agrícola Sant Isidre
Ajuntament
Pla de Santa Maria, el Comissió Pro Marató de TV3 del Pla de Santa Maria
Riba, la La Riba amb La Marató,
Paraularem
Casal Ribetà
Ajuntament
Rodonyà Col·lectiu de Dones El Taller
Valls Coral Espinavessa
Fundació Cultural Privada
Mare de Déu de Lledó
Vilabella Col·lectiu de Dones de Santa Àgata
Alt Empordà Agullana Grup de Voluntaris per a La Marató de TV3
Cadaqués Ciutadans de Cadaqués
Castelló d'Empúries Ajuntament i diverses entitats i associacions
Skydive Empuriabrava
Figueres Associació de Judo de l'Alt Empordà
Comerç Figueres-Comerç Ciutadà
Fundació Espor tiva Figueres
Fortià Grup de Teatre Pa amb Teatre
Navata Pessebre Vivent de Navata
Torremirona Club de Golf
Roses Fundació Roses contra el Càncer
Ajuntament
Viladamat Grup de Quina de Viladamat
Associació Juvenil Sant Feliu
AMPA CEIP Viladamat
Club de Futbol
Alt Penedès Avinyonet del Penedès Municipi d'Avinyonet del Penedès
Gelida Casal d'Avis Caixa Penedès
Coral Valldardina
Patronat Municipal d’Esports
Granada, la Associació Gent Gran
Societat Recreativa i Cultural Casal de la Granada
Olesa de Bonesvalls Grup de Dones les Bones Valls
Ordal Esplai Sol Naixent
Grup Excursionista Cresta Bocs
Grup Esplai Parroquial GEP-Vilafranca del Penedès
Santa Fe del Penedès Ajuntament de Santa Fe del Penedès
Grup Agàgio
Santa Margarida i els Monjos Penya Madridista els Monjos
Sant Cugat Sesgarrigues Societat Coral el Raïm
Grup de Joves Peluts i Pelats
Sant Martí Sarroca Agrupació Teatral i Dansaire Castell Sarroca
Junta Local AECC
Sant Pere de Riudebitlles Casal d'Avis Caixa Penedès de Sant Pere de Riudebitlles
Sant Quintí de Mediona Ajuntament
CF Mediona
CF Quintinenc
Sant Sadurní d'Anoia Centre de Fitness Equip 4
Ajuntament
Torrelavit Veïns de Torrelavit
Casal de la Gent Gran
Casal de Cultura
Coral Torrelavit
Vilafranca del Penedès Centre Comerç Penedes
Casal d'Avis del Tívoli de
Caixa Penedès
Castellers de Vilafranca
Grup d'Esplai Parroquial
de Vilafranca - GEM
Comarca Població Entitat

LA MARATÓ 2004 | 23
Unió Vinícola del Penedès (UVIPE)
Secció Sindical Intersindical CSC.
Hospital Comarcal Alt Penedès
Vilobí del Penedès AMPA Josep Madrenys
Ludoteca El Sol
Alt Urgell Bassella Museu de la Moto
Oliana Comissió de Festes d'Oliana
Seu d'Urgell, la Ciutadans de la Seu d'Urgell
Centre de Recursos Pedagògics de l’Alt Urgell
Alta Ribagorça Pont de Suert, el Coral Veus del Pont
Coral Infantil Edelweiss
Coral de la Gent Gran
Anoia Capellades Ajuntament
Esplai Aliret
Igualada Unió de Botiguers d'Igualada -
Comerç Ciutadà
Llacuna, la Pessebre Vivent de la Llacuna
Masquefa L’Alzinar-Societat Recreativa i Cultural
Òdena Ajuntament
Pobla de Claramunt, la Ajuntament
Barrufet Roig
ACCC
Grup de Teatre Jardí
Santa Margarida de Montbui Ajuntament
AECCC
Comissió de Festes
Grup Aixafarrocs
Vallbona d'Anoia Agrupament Escolta
Josep Masclans
Vilanova d'Espoia Centre Recreatiu i Cultural
de Vilanova d'Espoia
Bages Artés Artés Solidari amb La Marató
Avinyó AMPA de l'Escola Barnola d'Avinyó
Coordinadora d'Entitats Culturals
i Espor tives d'Avinyó
Balsareny Ajuntament
AECC
Comissió de la Festa dels Traginers
Castellbell i el Vilar Associació de Jubilats i Pensionistes de Castellbell
i el Vilar-Gent d'Edat
Fals Associació Cultural i Recreativa de Fals
Manresa Associació de Veïns Vic-Remei
IES Guillem Catà, branca Sanitària
Manresa, Mobilitza’t amb
La Marató
Monistrol de Calders Ajuntament
Cosidores de Patchwork de
Monistrol de Calders
Navarcles Unió de Botiguers de Navarcles
Ajuntament
Navàs Col·legi Sant Josep
Sant Fruitós de Bages Comissió Ciutadana per
La Marató de TV3
Sant Joan de Vilatorrada Casal de la Gent Gran
Santpedor Escola Llissach
Músics de la Catalunya Central i diverses entitats
i comerços de la població
Baix Camp Aleixar, l’ Associació de Dones ADA
Colla de Joves de l'Aleixar
Almoster Associació Almoster amb
La Marató
Ajuntament
Argentera, l’ Ajuntament
Borges del Camp, les Associació Cultural de
Dones Borgenques
Comarca Població Entitat

FUNDACIÓ LA MARATÓ DE TV3
Cambrils Ajuntament i diverses entitats i associacions locals
APA CEIP Joan Ardévol
Duesaigües Associació de Dones Duesaigües
Associació de Dones
Hospitalet de l’Infant, l’ Ajuntament i diverses entitats i associacions locals
Maspujols Associació de Dones la Dàli
Miami Platja Fòrum Miami
Club de Dansa Montserrat Sotero-Núria Esquius
Casal d’Avis de l’Hospitalet de l’Infant
Casal d’Avis de Miami Platja
Montbrió del Camp Ajuntament
Mont-roig del Camp Unió de Dones de Mont-roig del Camp
Col·lectiu de Poetes de Mont-roig i Miami
AMPA CEIP Mare de Déu de la Roca
Reus AMPA Col·legi Sant Josep
Associació de Veïns del Barr i de Sant Joan
Banda Simfònica del Conservatori
de la Diputació Tarragona
Escoles de música i Conservatori
de Grau Mitjà de Tortosa, Reus i Tarragona
Escola de Ball de Saló Pacha-Tubé
Escola Puigcerver (alumnes del
Taller de Teatre i del Consell de Delegats)
Pàmies Collector
Riudecanyes Grup de Dones de Riudecanyes
Riudecols Associació de Dones Puigcerver
Riudoms Taller d'Ocupació Baix Camp
L’Estudi. Centre de Dansa
Escola de Balls de Saló i Gimnàs
Ajuntament
Selva del Camp, la Associació l'Alzina i diverses entitats i associacions locals
Vandellòs Associació de Jubilats
Vilanova d’Escornalbou Associació de Dones de Vilanova d'Escornalbou
i l'Arbolet
Vilaplana del Camp Associació de Dones de Vilaplana
Vinyols i els Arcs Associació Cultural el Replà
Baix Ebre Ampolla, l’ Ajuntament
Benifallet Associació de Dones Meravelles
Jesús-Tortosa E.M.D. de Jesús
Perelló, el Agrupació Musical l'Emburgada,
Aula de Formació Musical
Perellonenca, Coral Santa Llúcia,
Coral de l’Associació de Jubilats del Perelló
Perelló, el AMPA CEIP Jaume II
Roquetes Associació de Dones de Roquetes
Tivenys Poble de Tivenys
Tortosa Escola Universitàr ia d'Infermeria
Verge de la Cinta
Baix Empordà Palafrugell Coral Mestre Sires de Palafrugell
Platja d'Aro Colla els Descarrilats de Platja d'Aro
Platja d'Aro Associació de la Gent Gran de la Vall d'Aro i S'Agaró
Sant Antoni de Calonge Ateneu Empordanès
Sant Feliu de Guíxols Casal Tueda APITE
Sant Feliu de Guíxols Club Aquàtic Xaloc
Piscina Municipal la Corxera
Colla Gegantera Guixolenca
Verges Moviment Cristià de la Gent Gran Pi Decreixent
Baix Llobregat Abrera Associació Contra el Càncer d'Abrera
i diverses entitats i associacions locals
Begues Spai d'Art
Collbató Entitats de Collbató
Cornellà de Llobregat Escola Santa Eulàlia
Esparreguera Associació Lluita Contra el Càncer d'Esparreguera
Gavà Casal d'Avis de Gavà
Martorell Unió de Botiguers Centre
Comercial la Vila
Comarca Població Entitat

LA MARATÓ 2004 | 25
Molins de Rei EITwo i diverses entitats i associacions
Olesa de Montserrat Associació Ariadna i diverses entitats i associacions locals
Prat de Llobregat, el Casal d'Avis Caixa Penedès el Prat
Santa Coloma de Cervelló Teatre Santa Coloma
Sant Boi de Llobregat Fundació Llor-Col·legi i Escola Espor tiva
Associació Juvenil 3,2,1 Acció
AMPA del CEI Rafael Casanova
Esplai la Barretina
Sant Climent de Llobregat Ajuntament i diverses entitats i associacions locals
Sant Esteve Sesrovires Coral Infantil Sant Esteve
Sant Feliu de Llobregat Agrupació Cultural Folklòrica Sant Feliu de Llobregat
Associació Balls d'Envelat Santfeliuenc
i diverses entitats i associacions locals
Sant Feliu de Llobregat Colla de Gegants de Sant Feliu
Sant Joan Despí Esbar t Dansaire Sant Joan Despí
i diverses entitats i associacions locals
Espai Físic. Escola de Dansa
Sant Just Desvern Comissió de festes del Barri Nord
Viladecans Agrupació Cultural Mossèn Cinto Verdaguer
Baix Penedès Albinyana Municipi d'Albinyana
Arboç, l’ Ajuntament
Bellvei La Muga, Associació de Dones
Calafell Confraria Pescadors de Calafell
Llorenç del Penedès CEIP les Cometes
Societat Cultural i Recreativa la Cumprativa
Coral Harmonia
Santa Oliva Ajuntament i diverses entitats i associacions locals
Sant Jaume dels Domenys Grup de Teatre de la Societat
Cultural i Recreativa de Sant Jaume dels Domenys
Sant Jaume dels Domenys Associació de Dones la Linyola
Vendrell, el Esplai Zig-Zag
Estudi de Dansa Tot Ritme
de Mar ta Sebares
Gimnàs Forma’t
Jove Baix Penedès i diverses entitats i associacions locals
Barcelonès Badalona Teatre Zorr illa
Conser vatori de Música
Badacentre
Barcelona Associació Catalana First Class 8
Centre Municipal de Vel Port Olímpic
Associació de Veïns Font de la Guatlla-Magòria
Associació Juvenil Mà Oberta
Cor Ober t Immaculada
Centre d'Esplai Josepets
Centre Moral i Cultural
del Poble Nou
Club d'Agility Ciutat Comtal
Associació Catalana de
Pitch & Putt
Colla Sant Medir Monumental
Cor de Cambra Impromptu i diverses entitats
i associacions del barr i de Gràcia
DiR-Clubs de Fitness
Escola Bonanova
Escola de Dansa Yisbell i
diverses entitats i associacions locals
Escola Peter Pan
Escola Universitària d'Infermeria
i Fisioteràpia Blanquerna
Esplai Sant Joan de Mata
Germandat de Trabucaires,
Geganters i Grallers de Sant Andreu
La Satànica de Sant Andreu
Grup Perelada
Marathon Catalunya i diverses
entitats i associacions locals
Comarca Població Entitat

FUNDACIÓ LA MARATÓ DE TV3
Mercat de Sarrià
Punt i Apart
Sants-Creu Coberta Centre
Comercial
UB Barça
Mountain Wilderness de
Catalunya i diverses entitats i associacions locals
Ràdio Club Quixots
Internacionals
ADOMA>45
AFANOC
Associació d’Excursionistes de Muntanya
Escola Sant Gregori
Marfil Santa Coloma
Mercat de l’Abaceria de Gràcia
Natació Sincronitzada Maristes
Teatre Llantiol
Per a l’altre Cor Cremat de Barcelona.
Ajut al Quart Món
Universitat Oberta de Catalunya
USAP de Barcelona
l’Independiente de Santander
Grup de Teatre de Psicologia de la UB
Vocalia de Discapacitats
Hospitalet de Llobregat, l’ SOMNIS. Ass. Catalana de Teatre de l'Hospitalet
Associació de Veïns Ildefons Cerdà
Casal d'Avis Caixa Penedès de l'Hospitalet de Llobregat
Casal d’Avis Sant Ramon
Caixa Penedès
Sant Adrià de Besòs Asociación de Mujeres Bienvenida
Associació Juvenil Barnavitas
Santa Coloma de Gramenet Escola d'Adults de Singuerlin
Berguedà Avià El poble d'Àvià
Unió Esportiva Avià
Berga Associació d'Excursionistes de Muntanya
Associació de Veïns els Quatre
Barris i diverses entitats i associacions locals
Cercs Ajuntament de la Vila de Cercs
i diverses entitats i associacions locals
Puig-reig Associació contra el Càncer de Puig-reig
Vilada Associació Juvenil Picant Cel
Associació de Dones de Fàtima
Ajuntament
Cerdanya Molina, la Couloir Extreme Sports
i diverses entitats i associacions locals
Conca de Barberà Barberà de la Conca Ajuntament i entitats de
Barberà de la Conca
Blancafort Comissió de Festes Blancafort
Espluga de Francolí, l’ Associació de Veïns
Urbanització Carreras
Casal de l'Espluga de Francolí
Col·legi del Carme de l'Espluga de Francolí
Montblanc Grup de Dones Afectades
de Càncer de Mama de Montblanc
Pira Anguera Associació Cultural
Solivella Grup Cultural de la Dona
Col·legi Públic Sagrat Cor
Garraf Sitges Casal d'Avis Sitges Caixa Penedès
Coral del Casal d’Avis de Cubelles
Vilanova i la Geltrú Associació de Gent Gran de la Mar
Garrigues Albi, l’ Associació de Dones l'Alosa
Associació de Gent Gran de
Sant Cosme i Sant Damià
Ajuntament
Arbeca Ajuntament i Associació de
Comarca Població Entitat

LA MARATÓ 2004 | 27
Dones l'Olivera i diverses
entitats i associacions locals
Borges Blanques, les Comissió La Marató de TV3
Grup Solidari de Castelldans
Cervià de les Garrigues Associació de Dones
Granadella, la Associació de Dones el Roure
Juneda Ajuntament i diverses entitats i associacions locals
Torms, els Societat Recreativa
Gran Casal dels Torms
Garrotxa Besalú Associació de Botiguers i Comerciants de Besalú
Olot Gimnàs Verd
Sant Feliu de Pallerols Ajuntament i entitats de Sant Feliu de Pallerols
Sant Jaume de Llierca Poligaris-Club d'Arquers de Sant Jaume de Llierca
Santa Pau Associació de Jubilats
Santa Pau Centre Cívic
Gironès Fornells de la Selva AMPA del Col·legi Públic
Forn d'Anells
Girona Gremi Artesà de Carnissers i Xarcuters
de les Comarques Gironines
Societat de Pesca Esportiva la Canya de Girona
02 Centre Wellness-Parc del Migdia
Llagostera Col·lectiu Pro Marató TV3
Club d’Handbol Llagostera
Llambilles Centre Cultural Sant Cristòfol
Sant Jordi Desvalls Agrupació de Trobades
Sardanistes Infantils
Maresme Alella Comissió de Festes d'Alella
Arenys de Mar Ajuntament i diverses entitats i associacions locals
Residència Geriàtrica d'Arenys de Mar i diverses
entitats i associacions locals
Arenys de Munt Ràdio Arenys de Munt
Cabrera de Mar Associació de Pensionistes i Gent Gran de Cabrera de Mar
Calella CAP Calella
Veïns del carrer Indústria
Coral Polifònica Granollers
Juventut Artística de Calella
Canet Unió de Botiguers i Comerciants
Dosrius Joventuts Socialistes del Maresme
Poliespor tiu
Malgrat de Mar Dinar 60 anys
Masnou, el Amics del Ferrocarril del Masnou (AFEM)
Parc Caramar
Gent del Masnou
Mataró Mataró Centre Comerç Ciutadà
Els Pastorets de La Marató
Montgat Mostra d’Entitats de Montgat
Ajuntament
Premià de Mar Ajuntament i diverses entitats i associacions locals
Centre l'Amistat
Kampikipugui
Senseoficinibenefici
Segle XXI
Santa Susanna Ajuntament i diverses entitats i associacions locals
Sant Cebrià de Vallalta Ajuntament i diverses entitats i associacions locals
Sant Pol de Mar Centre Cultural i d'Esbarjo
Sant Pol de Mar
Teià Ajuntament i diverses entitats i associacions locals
Tiana Taller de Cuina de Tiana
Parròquia
Taller Ocupacional Verge de l’Alegria
Ajuntament
Tordera Ajuntament i diverses entitats i associacions locals
Agrupació l’Amistat de
l’Alt Maresme
Vilassar de Dalt Associació Cultural Nat el 1951
Vilassar de Mar Casal de Curació Centre SocioSanitari de Vilassar de
Comarca Població Entitat

FUNDACIÓ LA MARATÓ DE TV3
Mar i Escola de Dansa Madó
Montsià Alcanar Associació Ajuda al Disminuït
Canareva
Amposta Ajuntament
Lliga contra el Càncer
de les comarques de Tarragona i Terres de l’Ebre
Freginals Associació de Dones de Freginals
Godall Mestresses de Casa de Godall
Sant Carles de la Ràpita Ràpita Endavant
Ulldecona Patronat de la Passió d'Ulldecona
Noguera Àger Associació de Dones de la
Vall d'Àger
AMPA Col·legi Andreu Ferran
Albesa Associació de Dones
Punt de Trobada
Artesa de Segre Gimnàs Altis
Ajuntament
Balaguer Associació Residència Geriàtr ica
Sant Domènec de Balaguer i
Associació de Dones d’Almatà
Balaguer Discoteca Sala Cats
Bellcaire d'Urgell Associació de Dones Isabelina de Bellcaire d'Urgell
Cubells Associació de Dones Mareselva
Menàrguens Associació de Dones la Clau de Menàrguens
Montgai Associació de Dones la Colomina de Montgai
Os de Balaguer Associació Juvenil Sampitot i entitats locals
Penelles Associació de Dones Il·lusió
Ponts Associació de Jubilats i Pensionistes de Ponts i Comarca
Dones Mig Segre (DOMISE)
Sentiu de Sió, la El poble de la Sentiu de Sió
Térmens Associació de Dones
Sant Joan de Térmens
Torrelameu Associació de Dones les Àguedes
Ajuntament
Vilanova de Meià Associació de Dones la Coma de Meià
Osona Manlleu Escola Carme Vedruna
Ràdio Manlleu
Masies Voltregà, les Ajuntament i diverses entitats
i associacions locals
Olost Casal d’Avis, Joves Músics i Noves Veus
Oristà Amics de la Marató del Municipi d'Oristà
Tona Escola Vedruna-Tona Fundació Paula Delpuig
Vic Centre Cívic Comerç Ciutadà de Vic
Hospitalitat Mare de Déu de Lourdes de Vic
Pallars Jussà Llimiana Ajuntament
Pobla de Segur, la Associació Cultura Comuna de Par ticulars de la Pobla de
Segur i diverses entitats i associacions locals
Associació Grup Escènic Lo Tamarro
Voluntaris Pobla de Segur
Salàs de Pallars Casal dels Avis de Salàs del Pallars
Tremp Gent de Tremp
Vall Fosca, la Veïns de la Vall Fosca-Amics de La Marató
Pallars Sobirà Sort Slot Club Sort i comerços locals
Pla de l’Estany Banyoles Ajuntament
Creu Roja Pla de l’Estany
Cornellà del Terri Ajuntament
Casal de Jubilats de Cornellà
de Terri i diverses entitats i associacions locals
Pla d’Urgell Bellvís Associació de Dones Pou de l'Hort
Golmés Associació de Dones Albada del Pla d'Urgell
Linyola Associació de Dones de Linyola (ADOLI)
Miralcamp Comissió de Festes de Miralcamp
Mollerussa Pub Cherookee
Associació de Dones Albada del Pla d’Urgell
Palau d'Anglesola, el Associació de Dones del Palau d'Anglesola
Comarca Població Entitat

LA MARATÓ 2004 | 29
Palau d'Anglesola, el Llar d'Infants Marte
Torregrossa Associació de Dones Busca
Temps i diverses entitats i associacions locals
Ajuntament i diverses entitats i associacions locals
Vilanova de Bellpuig Associació de Dones Anjua
i diverses entitats i associacions locals
Priorat Marçà Associació de Dones la Miloquera
Vilella Baixa, la Ajuntament i diverses entitats i associacions locals
Ribera d’Ebre Benissanet AMPA CEIP Antoni Nat
Flix Associació de Dones de Flix
Móra d'Ebre Ajuntament
Móra la Nova Associació Col·lectiu de Dones de Móra la Nova
KAU-KAUS i Aula de Música
Riba-roja d'Ebre Associació de Dones Riba-roja
Societat la Llum del Dia
Tivissa Col·lectiu Solidari Xino-Xano
Torre de l’Espanyol, la Associació de Dones Bassa d'Oli
Ripollès Planoles Associació de Dones de Planoles
Ribes de Freser Associació de Dones de la
Vall de Ribes i tot el poble
Associació de Jubilats i Pensionistes de Ribes de Freser
Teatre d'Emergència
Ripoll AMPA Escola Vedruna
Sant Joan de les Abadesses Associació de Jubilats
i Pensionistes de Sant Joan de les Abadesses
Segarra Cervera Creu Roja Joventut-Cervera i diverses entitats
i associacions locals
Guissona Escola de Dansa Montse Esteve
Ajuntament
Hostafrancs Associació de Dones d’Hostafrancs
Llor, el Associació de Veïns del Llor
Massoteres Ajuntament
Pallargues, les AMPA de l'Escola Pública de les Pallargues
Segrià Aitona Penya els Tabolls
Associació Mestresses de Casa
Associació de Sant Gaietà
Ajuntament
Grup d'Art Aitonenc
Alamús, els Associació de Puntaires Sant Martí
Associació de Dones dels Alamús
Albatàrrec Associació de Dones Àgates d'Albatàrrec
Associació de Puntaires
Club de Futbol
Ajuntament
Alcarràs Gotta Perfumeries
Penya Barcelonista
Alfarràs Poble d'Alfarràs
Alfés Associació de Dones d'Alfés
Almatret Associació de Dones l'Alba
Penya Barcelonista
Almenar Associació de Country d'Almenar
Artesa de Lleida Ajuntament i diverses entitats i associacions locals
Benavent de Segrià Ajuntament
Corbins Ajuntament i entitats i comerços de la vila
Granja d'Escarp, la Associació Juvenil Penya
Borrachitos
Lleida Associació de Veïns del Barri de Magraners
Circuit Urgellenc
Coral Sícor is
Grup Ioga Ananda
Centre d’Estudis Toni Prim
IES Ronda
Club Esportiu la Seu Vella
Joieria-Rellotgeria Horti’s
Danone
Comarca Població Entitat

FUNDACIÓ LA MARATÓ DE TV3
Residència Mixta la Seu
Massalcoreig Associació de Mestresses de Casa Llar d'Esbarjo
Montoliu de Lleida Associació Cultural i Recreativa de Montoliu de Lleida
Sarroca de Lleida Associació de Dones de
Sarroca de Lleida i diverses
entitats i associacions locals
Soses Ajuntament-Comissió de Festes
Sudanell Associació Pro Marató TV3 de Sudanell
Sunyer Associació de Dones Santa Llúcia
Torrefarrera Associació Cultural de Dones l'Amistat de Torrefarrera
Selva Arbúcies JAM
Blanes Ajuntament
Associació Penya Barcelonista Blanes
Escola de Dansa Daina i diverses entitats
i escoles de la població
Breda Esplai Plori Mai
Caldes de Malavella Club de Futbol Sala Femení de Caldes
Cellera de Ter, la Colònies per a la Gent
Gran de la Cellera
Lloret de Mar Esplai Bimbirimboines i diverses entitats i associacions locals
Massanes Associació Amics de Massanes
Osor Casal de Jubilats i Simpatitzants de la Vall d'Osor
Riells i Viabrea Riells i Viabrea amb
La Marató de TV3
Riudarenes AMPA del Col·legi Josep
Boada de Riudarenes-Secció
Gimnàstica Rítmica
Càmping Riudarenes
Esplai Parroquial Sant Martí de Riudarenes
i diverses entitats de la població
Riudellots de la Selva Esplai Parroquial
Ajuntament i altres entitats i associacions locals
Santa Coloma de Farners Club Hoquei Farners
Ajuntament i diverses entitats
i associacions esportives locals
Sant Hilar i Sacalm Associació de Pares de Família de Sant Hilari Sacalm
Associació Via Crucis Vivent de Sant Hilari Sacalm
Atletes de Sant Hilari
CasalxJoveshs
Isards per La Marató
Sils Ajuntament
Tossa de Mar Unió Atlètica Tossa i diverses associacions i entitats locals
Vidreres CEIP Salvador Espriu
Amics de La Marató de Vidreres
Solsonès Sant Llorenç de Morunys Associació Cultural l'Estaca
Solsona Consell Comarcal del Solsonès
Tarragonès Catllar, el Agrupació de Dones el Catllar
Constantí Ajuntament i altres entitats i associacions locals
Morell, el Esplai Parroquial del Morell
Nou de Gaià, la Ajuntament de la Nou de Gaià
Pobla de Mafumet, la Ajuntament
Comissió de Festes la Pobla de Mafumet
Tarragona Associació de Pares d'Alumnes del Col·legi Sagrat Cor
Comissió Infantil de la Federació Catalana de Judo
Escola Universitària d'Infermeria Rovira i Virgili
Institut Mediterrani del Disseny i la Moda
i diverses entitats i associacions locals
Associació Sant Magí Santa Tecla
Terra Alta Arnes Associació de Dones d'Arnes (ADDA)
Bot Associació de Dones de Bot
Les Veus de Bot
Fatarella, la Escola Municipal de Música de la Fatarella
Pobla de Massaluca, la Associació de Dones la Trobada
Vilalba dels Arcs Associació de Dones de Vilalba dels Arcs
Urgell Agramunt Associació de Dones l'Esbarjo
Comarca Població Entitat

LA MARATÓ 2004 | 31
Anglesola Mupassopipa
Bellpuig AV Sant Jaume de l'Avinguda de Catalunya
i diverses entitats i associacions locals
Castellserà Associació de Dones l'Esclat
Sant Martí de Maldà Associació de Dones Flor de Romaní
Gabinet de Crisi
Tàrrega Associació de Dones Arrel
Club d’Automibilisme Escuderia Tàrrega
Tornabous Associació de Dones Esperança de Tornabous
Vallès Occidental Castellar del Vallès Club d'Hoquei Castellar
Montcada i Reixac Associació de la Gent Gran
Ripollet Esplai la Gresca
Escola Anselm Clavé
Sabadell Agrupació de Comerciants Via de Massagué
Associació de Puntaires Ribot i Serra
Associació de Veïns Creu Alta i diverses entitats
i associacions locals
Carmelites Vedrunes de Sabadell
Escola Oficial d'Idiomes de Sabadell
Grup d’Amics de La Marató
IES-SEP Castellarnau
Joventut de la Faràndula
Sabadell Comerç Centre
Sabadell Cultura i diverses entitats i associacions locals
Santa Perpètua de Mogoda Ajuntament
Escola Bressol Municipal Can Folguera
Sant Cugat del Vallès Comissió de Festes de la Floresta - Sant Cugat
Marató AGA
Team Par tners
Ajuntament
Sant Llorenç Savall Sant Llorenç Solidari
Terrassa Casal de Gent Gran la Maurina
Cingle-Centre d'Ensenyaments Secundaris
Comerç Terrassa Centre - Comerç Ciutadà
Nens i Nenes del Programa Jugar i Llegir
Associació de Dones Bonaire
Oncolliga
Ekipo Parásito
Vacarisses Cor de Vacarisses
Vallès Oriental Aiguafreda DONAI-Associació Cultural
Dones d'Aiguafreda
Jugadors Veterans
Alumnes de l’Escola la Muntanya
Ajuntament
Ametlla del Vallès, l’ Agrupació Cultural de Dones l'Ametlla del Vallès
Caldes de Montbui Acció Cívica Calderina
Tot Música, Centres d'Ensenyament Artístic
IES Manolo Hugué
AV Can Rossell i els mitjans de comunicació locals
Canovelles Comissió Sant Antoni Abat
Cardedeu Comerç Ciutadà-Unió de Botiguers de Cardedeu
Garriga, la AMPA CEIP Puiggraciós
CEIP Tagamanent
Granollers Gran Centre Granollers
Martorelles Club de Futbol Martorelles
Santa Maria de UBIC
Palautordera
Sant Antoni de Vilamajor Colla Gegantera de Sant Antoni de Vilamajor
i diverses entitats i associacions locals
Vallromanes Ingreen-Grup Imbesòs
Vilanova del Vallès Comissió Marató Vilanova
Associació de Dones la Lluna
Andorra Sant Julià de Lòria Universitat d'Andorra
Comarca Població Entitat

FUNDACIÓ LA MARATÓ DE TV3
La Fundació canalitza la comunicació de la
campanya de sensibilització i del conjunt de
les accions que es duen a terme entorn de
La Marató a través dels mitjans de comuni-
cació i de les mateixes eines de divulgació
de què disposa.
Difusió
El dia 28 d’octubre de 2004, la Fundació va
convocar la premsa a l’Auditori de TVC, per
informar del contingut de la campanya de
sensibilització de La Marató, tot presentant
l’edició especial del DVD dirigit als centres
educatius i cívics.
La repercussió total que va tenir en els
mitjans de premsa escrita generalista i
comarcal la difusió de La Marató va ser de
485 impactes, mentre que un total de 40
emissores de ràdio i de televisions locals
van emetre reportatges i entrevistes
especials sobre La Marató.
Divulgació
Durant l’exercici, la Fundació va editar dos
n ú m e ros de la revista Món Marató – l ’ e d i c i ó
n ú m e ro12 (novembre de 2004), amb un tiratge
de 51.000 exemplars, i la número 13 (juny de
2005), amb 75.500 exemplars–, de distribució
gratuïta, publicació semestral i finançades amb
fons provinents de patro c i n a d o r s .
Una altra publicació editada durant aquest
període va ser la Memòria d’activitats.
Exercici juliol 2003-juny 2004, amb el resum
de totes les activitats dutes a terme i els
resultats econòmics obtinguts per la
Fundació durant l’exercici comptable.
El lloc web www.fundaciomaratotv3.org és
el mitjà amb el qual la Fundació informa
puntualment de totes i cadascuna de les
notícies que es generen al seu entorn, com
ara l’actualització de resultats obtinguts amb
els donatius, dades tècniques de la darrera
Marató, la convocatòria d’ajuts i els projectes
guanyadors o la celebració del Simposium,
entre d’altres.
Comunicació
Presentació de la campanya de sensibilització i
difusió de La Marató 2004 als mitjans de comuni-
cació, celebrada a l’Auditori de TVC.
Accions de difusió
Publicitat (pàgines d’anuncis): 45
Falques radiofòniques: 32
Encartaments (Avui,
La Vanguardia, El Periódico): 690.000
Banderoles (Badalona,
Banyoles, Barcelona,
Figueres, Girona, l’Hospitalet
de Llobregat, Lleida, Manresa,
Olot, Reus, Sabadell, Sant Joan
Despí, la Seu d’Urgell, Tarragona,
Tàrrega, Terrassa, Tortosa): 2.900

LA MARATÓ 2004 | 33
La planificació de les activitats prèvies al
programa es du a terme a partir del mes
de setembre des de la Comissió de
Coordinació, que organitza, també, la logísti-
ca de suport a La Marató.
Aquesta Comissió està formada, a més dels
representants de la Fundació i de TV3, pels
del Departament de Benestar i Família, “la
Caixa”, PriceWaterhouseCoopers, Telefónica
i Tradia, empreses col·laboradores habituals
amb La Marató de TV3.
Direcció General d’Accions Comunitàries i
Cíviques del Departament de Benestar i
Família: coordina les tasques de voluntariat i
col·labora en l’organització de la campanya
de sensibilització i difusió.
Bassat Ogilvy & Mather Comunicación:
responsable de la creativitat de la campanya
de promoció social i de difusió de La Marató.
Caixa d’Estalvis i Pensions de Barcelona:
organitza el processament de les dades
referents a les donacions i el cobrament
posterior i prepara la documentació de
tancament de La Marató i d’auditoria.
PricewaterhouseCoopers: estableix els
controls i les verificacions de les donacions,
amb l’auditoria posterior de la Fundació, tot
donant la màxima fiabilitat i transparència.
Telefónica: instal·la les línies telefòniques i
vetlla per les connexions necessàries per
rebre donacions. Aquestes línies estan distri-
buïdes entre les seus de Barcelona, Lleida,
Girona, Tarragona i els estudis de TV3.
Tradia: transporta el senyal d’àudio i vídeo
de les activitats exteriors mitjançant les
unitats mòbils desplaçades, amb enllaços
terrestres i via satèl·lit.
Seus territorials, línies telefòniques
i voluntaris
Els donatius a La Marató de TV3 es poden
fer per mitjà d’un ingrés directe o transferèn-
cia bancària, per Internet, per Servicaixa i
mitjançant una trucada al telèfon 905 111
213 el mateix dia del programa.
La Fundació posa en funcionament 5 seus
telefòniques repartides territorialment
–Barcelona: Palau Sant Jordi i Hall de TV3;
Palau Firal de Girona; Universitat de Lleida i
Cobert 1 del Port de Tarragona– per atendre
les trucades dels donatius econòmics que
fan els ciutadans.
Al capdavant de cadascuna de les seus hi ha
un responsable que s’hi incorpora durant un
període de dos mesos (novembre i desembre )
per coordinar les tasques necessàries per al
bon funcionament de La Marató.
Les seus disposen d’un determinat nombre
de línies que, en l’edició 2004, van ser
ateses per 2.337 voluntaris, procedents de
279 associacions i entitats de voluntariat.
Organització
i logística
Generalitat de CatalunyaDepartament de Benestar i Família

També hi participen com a voluntaris treballa-
dors de TVC, de Catalunya Ràdio i de les
altres empreses de la CCRTV, a més de
representants de la societat civil, empresarial
i del món polític.
A més, prop de 200 empreses i institucions
d’arreu de Catalunya donen, desinteressada-
ment, suport logístic en l’organització de les
seus des d’on es fa possible el programa.
Empreses que van donar suport
a La Marató 2004
FUNDACIÓ LA MARATÓ DE TV3
A totes les seus:
Aigua Sant Aniol; Bimbo; Bombons Blanxart;
Cadí Formatges i Mantega; Café Saula;
Casademont Indústria Càrnia; Caves Nadal;
Cepac, SL; ChupaChups, SA; Cobega, SA;
Danone, SA; Eckes Granini Iberica; Establiments
Viena; Freixenet; Frit Ravich, SL; Gelats
M e n o rquina; Jordi Pons, SL; Kenwood Iberica;
La Bolsera; Llet de Catalunya, SL; Massegur;
M e rc a b a rna, SA; Nestlé España, SA; Tr a n s p o r t s
A z k a r, SA; Veritas; Xocolates Las Comas.
Palau Sant Jordi
Apsa, Auxiliar Portuaria, SL; Aramark;
Associació Gatronòmica Bon Profit; Autoservicio
Milla; AVIS, Lloguer de cotxes; Badia Roca
Pastisser; Barcelona Promoció; Ben Fumat;
Casa Gay; Dasler; DHL Express; Dynamobel,
SA; Escola Bonanova; Flors Navarro; Focus, SA;
Foix de Sarrià; Fomento de Construcciones y
Contratas; Gremi de Flequers de Barcelona; IES
Maria Lluïsa Cura; J. Barzano, SA; MEYM,
Gestion Integral de Mantenimiento; Nostrum;
Novo Plastics la Primera; Pastisseria Escribà;
Radio Taxi 033; Semon; Servigrup; Te c n o
Antena Sistemas; To r res Alquilers de
Calefacción; Transports Metropolitans de
B a rcelona; UGT de Catalunya; Wall Vi d e o ,
Audiovisuals per a events.
TV3
Blachere; Ferfor; Mercabarna, SA; Serunión;
Telepizza.
Palau Firal de Girona
Avícola Maria, SA; Bonal, SA; Catalana de
Serveis; Ecosan Sanitaris Portàtils; Escola
d'Hostaleria de Girona; Flors Freesia; Fruites
Danés; Furgoempordà, SL - Llicència Hertz;
Fusté Burgas; Galeria 35; Girona Fruits S. Coop
C. Ltda.; Gremi de Flequers de les Comarques
Gironines; Hotel Carlemany; Hotel Costabella;
Lloguers i Muntatges Pous, SL; Massegur;
Miquel Alimentació Grup; Pastisseria Castelló;
Rètols Gispert; Securitas Seguridad España;
Servielèctric Girona, SL; Servis Complet, SA;
Tardós, SA; Videoson; Vivers Moner.
Universitat de Lleida
Valls Comercial Fripan; Signum Rètols;
Restaurant Tastapans; Restaurant Fonda del
Nastasi; Perfumeria La Primavera; Pastisseria
Tugues; Pastisseria Pons; Hotel NH Pirineos;
Fotografia Gómez Vidal; Escola d'Hostaleria;
ELECSON; COMERTEL SA.; Casa Piqué; Cafès
Batalla 2000; Associació Aspros; Ajuntament de
Lleida; Actel.
Cobert núm. 1 del Port de Tarragona
Automàtic Tarraco, SA; Cafeteria Leman;
Christian's Service; Civit Floristeria; Complex
Educatiu de Tarragona; Fira de Reus; Fruites
Civit, SL; Granja Gaià, SA; Gremi de Forners de
Tarragona; Guimerà, SA; Hotel Astari; Hotel
Urbis; Jordi Pons, SL; La Botiga del Cafè;
Marinada Tarragona, SL; Material per a
l'Hostaleria; Pastisseria Conde; Pastisseria Font;
Pastisseria Geneve; Pastisseria Palau;
Pastisseria Rabasó; Pastisseria Rovira;
Pastisseria Sanromà; Pastisseria Trill; Pastisseria
Viver; Pié Vídeo Sonido; Ralequip; Restaurant El
B a rquet; Restaurant Merlot; Servi-Repro .
Líniestelefòniques 542 50 50 75 717
Voluntaris 1.777 160 180 220 2.337
Procedència.Nombre detrucades(impulsos) 306.511 51.544 28.388 41.015 8.517 435.975
Barcelona* Girona Lleida Tarragona Altres Total
* S’hi sumen el Palau Sant Jordi i el Hall de TVC.
Distribució de les línies telefòniques i dels voluntaris

LA MARATÓ 2004 | 35
Els Alamús Ass. Dones Els Alamús
Alella Ass. de Dones Montserrat Roig
Algerri Ateneu Penya Tarros
Badalona Voluntaris Badalona
Badia del Vallès IES Ciutat Badia
A.C. Vermell i Negre
Balaguer Ass. Cultural d'Art 4
Banyoles IES Pla de l'Estany
Barcelona ACAIR
ACE Sansur
ADER Ass. Malalts de Ronyó
AFANOC - Ass. Nen amb Càncer
Agrupament Scout Gandhi
Àmbit Maria Corral
Àmbit Prevenció
AMPA Sant Joan de Ribera
APILC
APSOCECAT
ASEM
Ass. Amics Fires del Poble Sec
Ass. Benestar i Desenvolupament de Barcelona
Ass. Cívica d’Ajuda Mútua
Ass. Coordinadora ACA
Ass. de Dones Palas Atenea
Ass. Dones Artesanes
Ass. Gent Gran Bac de Roda
Ass. Nou Horitzó
Ass. Pere Relats per a Ancians
Ass. Taller Ocupacional Ariadna
La Caixa Barcelona
CAM Obres Socials Catalunya
Càritas Diocesana de Barcelona
Casal de Sords de Barcelona
CEIR – ARCO
Celíacs de Catalunya de Barcelona
Centre d'Esplai Santa Maria de Gràcia
CE Sant Isidor
Club Escolapis de Sarrià
Colla Els Patufets Sant Medir
Colla Sant Medir Els Pilans
Col·lectiu Congrés 81
Col·lectiu per la Igualtat en la Diversitat
Col·legi Sagrada Família de Barcelona
Col·legi Sagrat Cor de Sarrià
Comissió de Festes de Barcelona
Comitè Olímpic de Catalunya
Conex Barcelona
Consell Evangèlic de Catalunya
Coordinadora d'Usuaris de la Sanitat
Coordinadora Nacional de Jubilats i Pensionistes de Barcelona
La Dona, Diàleg i Catalunya
Ensems
Esade - Dept. d'Activitats Culturals
Escola Sant Ignasi - Sarrià
Escola Arco
Esplai El Drop
Esplai Natzaret i Betània
Fundació Ajuda i Esperança
Fundació Ajudant a Ajudar
Federació Catalana de Persones amb Retard Mental
Federació d'Ass. de Gitanes de Catalunya
Federació d'Ass. de Gitanes de Catalunya
Federació de Colles de Sant Medir
Fundació Esclerosi Múltiple de Barcelona
Població Entitat
Col·lectius de voluntariat amb La Marató

FUNDACIÓ LA MARATÓ DE TV3
Fundació Esmen
Fundació Ficat
Fundació Josep Carreras
Fundació Malalts Mentals de Catalunya
Fundació Pere Tarrés
Gramic
Grup Àgata
Grup de Dones del Casal de les Corts
Grup de Teatre Disbauxa
L’Hora de Déu
Hospital de Dia d'Esclerosi Múltiple de Barcelona
Hospitalitat Mare de Déu de Lourdes de Barcelona
Junta local AECC
Justícia i Pau
Llar Mare de Déu de Gràcia
Nexe Fundació
Once - Centre de Recursos Educatius
Patronat de Sant Galderic
Petit Ballet de Barcelona
Residència Jaume Batlle
Societat Coral l'Ideal d'en Clavé
SECOT
Servei Solidari
SISAL
Tutela, La
UE Sant Andreu
Unió Esportiva d'Horta
Bescanó Malalts de Crhon i Colitits Ulcerosa de Bescanó
La Bisbal d'Empordà Casa Cultural Andalucía de la Bisbal
Fundació Oncolliga de la Bisbal d'Empordà
Ràdio Bisbal
Les Borges Blanques Grup de Teatre Nissaga
Calaf Club Bàsquet Calaf
Caldes de Montbui Acció Cívica Calderina
Calella Obra de Maria de Calella
Cambrils Amics de la Gent Gran
Dones de Vilafor tuny
Canet de Mar Centre Sant Pere Apòstol
Canovelles Club d'Escacs Canovelles
Cardedeu CDU Multiserveis Gent Gran
Castellar del Vallès Fundació Obra Social Benèfica de Castellar del Vallès
Castellvell del Camp Unió de Dones de Castellvell
Cerdanyola del Vallès Ass. de Veïns Can Carau
Ass. Gent Gran de Cerdanyola del Vallès
Llar del Jubilat i del Pensionista de Cerdanyola del Vallès
Ràdio Club Vallès
Cervià de les Garrigues ADF Les Garrigues
Cornellà de Llobregat AFA Baix Llobregat
Ass. Cultural Amigas
L'Espluga de Francolí ADF l'Espluga de Francolí
Casal de l'Espluga de Francolí
Esplugues de Llobregat Espluga Viva
Voluntaris de Sant Joan de Déu
Falset OBF El Priorat
Figueres ACAS
Ass. de Voluntariat Figueres
Gelida Grup d'Esplai Mainada
Girona Amics de la UNESCO de Girona
Ass. de Veïns de Sant Narcís
ASSTEM
La Caixa Girona
Confrar ia Mare de Déu dels Dolors de Girona
Creu Roja de Girona
Població Entitat

LA MARATÓ 2004 | 37
Esclerosi Múltiple Girona
FADESIA
MIFAS
Voluntaris Penitenciaris de Girona
Granollers Col·lectiu d'Esplai als Barris
Parròquia Sant Esteve
L'Hospitalet de Llobregat AFEMHOS
Agrupació de Veïns B. Ciutat Comtal
ARC Vell Can Bori
Ass. de Disminuïts Físics de l’Hospitalet
Ass. Trasplantats Cardíacs de l'Hospitalet
Gent de Pau de l'Hospitalet de Llobregat
JIS Joves per la Igualtat i la Solidaritat
Vocalia Drogodependències
Voluntaris de Bellvitge
Igualada Ateneu Igualadí
ATLAS
Esplai Àuria
Grup de Lleure La Clau
Grup Sardanista Montserratí
Jorba AMPA Col·legi Montclar
Juneda Agrupació Sardanista de Juneda
La Llagosta Fórum de Debate Llagostense
Lleida AREMI
Ass. Aspros
Ass. de Dones Flor de Lis
Ass. Familiars Malalts Alzheimer de Lleida
Ass. Festa Moros i Cristians
A.V. Copa d'Or i Sot de Fontanet
A.V. Pda. Butséhit
La Caixa Lleida
Càritas Diocesana de Lleida
Coral Shalom
Esclat
FAPAC
Federació d'Ass. de Veïns de Lleida
Federació de Cases Regionals de Lleida
Jubilats i Pensionistes Arques de Rufea
Lliga contra el càncer de les comarques de Lleida
Org. de Veïns Pardinyes Overpard
Shalom Taller
Maçanet de la Selva Casal de Jubilats i Pensionistes de Maçanet de la Selva
Manlleu Comunitat de Propietaris de Manlleu
Grup Sardanista Ocellets del Ter
Manresa Ass. de Veïns Sagrada Família de Manresa
Martorell Ass. Arc de Sant Mar tí
Martorelles ADISMAR
Grup de Voluntaris l'Abraçada
Mataró Colles Sardanistes Repuntejant
Gimm - Grup Invàlids Mataró i Maresme
Voluntaris per al Futur de Mataró
Molins de Rei Centre d'Esplai Mib
Mollerussa Ass. de Dones Albada Pla d'Urgell
Consell de Dones Pla d'Urgell
Mollet del Vallès Ass. Chokorling
Montcada i Reixac Esplai SESA
Montserrat Servei Escolta de Montserrat
Móra d'Ebre Ass. de Dones Móra d'Ebre
Navarcles IBADA
Olot Fundació Oncolliga Olot
IES la Garrotxa
Ordal Esplai Sol Naixent
Població Entitat

FUNDACIÓ LA MARATÓ DE TV3
La Palma de Cervelló Moviment d'Apostolat Infantil La Palma
Parets del Vallès Solidaris Parets
El Pla de Santa Maria Catesplai Planenc
Pobla de Segur Ass. Cultural Amics del Poble d'Aransís
Polinyà Germandat de Polinyà
Prades Ass. Dones L'Abellera
El Prat de Llobregat Ass. Amics de la Unesco del Prat
Ass. de Veïns Sant Cosme
Reus Ass. de Veïns 25 de Setembre
Càritas Interparroquial de Reus
Esclerosi Múltiple Tarragona
Ripoll Grup Xarxa de Ripoll
Riudoms Esplai del Casal Riudomenc
La Roca del Vallès Esplai Xiruca
Rubí Club Minusvàlids Horitzó
Sabadell Ass. Moskitia
CAE La Llar del Vent
Casal Jubilat i Pensionistes de Campoamor
Esplai Can Colapi
Som-hi
Sallent Casal de Gent Gran de Sallent
Sant Adrià de Besòs Disfrisa
Sant Boi de Llobregat Fundació Marianao
Tots Som Santboians
Sant Cugat del Vallès Grup de Teatre Espiral
Grup de Voluntaris de l'HGC
Sant Cugat Sesgarrigues Ass. d’Assistència Social Sant Adjutor i
Sant Joan de Mediona Grup de Teatre l'Escorça
Sant Joan les Fonts Escola Mi Belles Ar ts
Sant Vicenç dels Horts Fundació Privada Iris
Societat Cultural la Vicenteta
Santa Coloma Cervelló Dones pels 4 Cantons
Santa Coloma de Gramenet Agrupació de Veïns Dte. VI del Fondo
ASED
Ass. Ajuda a l’Ancianitat de Santa Coloma
AV Can Mariner
Casa de Aragón de Santa Coloma
Centre Excursionista Puigcastellar
Grup de Dones de Can Mar iner
Santa Cristina d'Aro CEIP Pedralta
Santa Oliva Colla Gegantera de Santa Oliva
Santa Pau Ass. Cultural Lúdica la Trepa
Santa Perpètua de Mogoda Voluntaris Forestals
Tarragona A. E. Gelsto
Ass. de Veïns Campclar
Ass. Provincial Laringectomitzats de Tarragona
Ass. Residencial Palau
ASTAFANIAS
Ateneu Cultural de Dones de Tarragona
La Caixa Tarragona
Casal Cívic Camp Clar
CDC Torreforta
CERADAI
Colla Cossetània
Hospitalitat Mare de Déu de Lourdes de Tarragona
Parròquia de Sant Pau
Terrassa Ass. Coordinadora Ajuda Unida de Terrassa
Centre d'Esplai Sant Llorenç
Club de Lleure els Globus
Col·lectiu de Dones de Ca n’Anglada
Col·lectiu de Dones Viudes de Catalunya
Població Entitat

LA MARATÓ 2004 | 39
La Fundació du a terme una campanya de
captació de recursos econòmics de les
empreses per finançar la campanya de
sensibilització i de difusió de La Marató de
TV3 i per ajudar a sufragar les despeses del
seu funcionament.
Patrocini
A b e r t i s
A s t r a Z e n e c a
Comerç Ciutadà
D A M M
E s t e v e
F e c s a - E n d e s a
F I AT C
Gallina Blanca
Glaxo Smith Kline
L a c e r
M e rc a d o n a
Miquel Alimentació Grup
Mutua Manre s a n a
N o v a r t i s
O ro n a
P f i z e r
T i t a n
U v i p e
E m p reses que van patrocinar l’edició 2 0 0 4
Col·legi Joaquima de Vedruna AMPA
Coral Campus Terrassa
Lliga Reumatològica Terrassa
Voluntaris de Terrassa
Torà Càritas Parroquial de Torà
Torelló Càritas Interparroquial de Torelló
Torredembarra Fundació Pere Badia
Torrelles de Llobregat Ateneu Urgellenc
El Vendrell Assemblea Comarcal del Vendrell
Equipament Polivalent El Vendrell
Grup de Dones del Vendrell
Vic Càritas Diocesana de Vic
Vilabella Col·lectiu de Dones de Santa Àgata
Viladecans Centro de Formación de Adultos Edelia Fernández
Vilafranca del Penedès Caixa Penedès Voluntaris de Vilafranca
Camí Solidari
Castellers de Vilafranca
Ginesta
Vilajuïga Ass. de Dones de Vilajuïga
Vilanova i la Geltrú AMPA Escola Llebetx d’Adarró
Ass. de Voluntaris del Garraf
Societat Sardanista Dansaires Vilanovins
Vimbodí Grup de Grallers la Palanca
Xerta Ajuntament de Xerta
Població Entitat


L’aportació dels ciutadans a La Marató és
una important font de recursos per a la
recerca biomèdica d’àmbit català, per
l’impacte que té en les institucions de
recerca (formació i infraestructures) i per la
productivitat científica.
L’estudi de l’avaluació de l’impacte de la
recerca finançada per la Fundació La Marató
de TV3 demostra que hi ha un considerable
nombre de documents científics que superen
la mitjana internacional de citacions. Això
significa que els grups finançats per la
Fundació han fet treballs d’excel·lència, amb
visibilitat i interès internacional. A més,
aquest estudi també acredita que molts
d’aquests equips han aconseguit finança-
ment en convocatòries d’altres entitats a
partir dels treballs iniciats amb el suport de la
Fundació.
Aquesta Memòria conté les actuacions fetes
en l’àmbit de la recerca durant l’exercici juliol
de 2004-juny de 2005 i el resum de les
memòries finals presentades en el V
Simposium Fundació La Marató de TV3, en
què els investigadors que van finalitzar els
seus projectes sobre la diabetis i altres
malalties metabòliques (La Marató del 1998)
van presentar els seus resultats a la comuni-
tat científica i a la societat en general.
També hi ha totes les gestions –econòmi-
ques i científiques– que s’hi van dur a terme,
corresponents a diverses edicions de La
Marató i la convocatòria d’ajuts de La Marató
del 2004, dedicada al càncer, amb el resultat
de l’avaluació dels 260 projectes presentats,
segons la gestió de l’Agència d’Avaluació
de Tecnologia i Recerca Mèdiques, del
Departament de Salut.
En aquest període, la Fundació també va
aprovar l’adjudicació, a un total de 27 projec-
tes de recerca, dels fons obtinguts durant La
Marató del 2003, dedicada a les malalties
respiratòries cròniques.
RECERCA BIOMÈDICA | 41
Recerca biomèdica

FUNDACIÓ LA MARATÓ DE TV3
La convocatòria d’ajuts a la recerca sobre el
càncer va ser anunciada a la premsa i al web
de la Fundació, un cop aprovades les bases
pel Patronat a proposta de la Comissió
Assessora Científica.
S’hi van presentar un total de 260 projectes
de recerca, dels quals: 214 eren unitaris i 46
coordinats entre dos o més grups de
recerca de centres diferents. Per àrees de
recerca: 132 projectes corresponien a
recerca bàsica, 69 a recerca clínica, i 22 a
recerca epidemiològica i serveis sanitaris,
mentre que els altres 37, eren combinacions
d’aquestes àrees.
El nombre de projectes que van participar en
aquesta convocatòria representa una xifra
rècord de totes les celebrades per la
Fundació, la qual cosa demostra la importàn-
cia que va adquirint aquest concurs en
l’àmbit de la recerca biomèdica.
Convocatòria d’ajuts
Càncer. La Marató 2004

RECERCA BIOMÈDICA | 43
Centres Projectes Projectes Subprojectes* unitaris coordinats
Hospital Clínic i Provincial de Barcelona 33 6 2
Universitat de Barcelona 23 1 6
Hospital de la Santa Creu i Sant Pau 17 - 1
Institut Municipal d'Investigació Mèdica 17 5 1
Hospitals Vall d'Hebron 15 4 2
Institut Català d'Oncologia 14 3 3
Universitat Autònoma de Barcelona 12 2 2
Parc Científic de Barcelona 10 2 2
Ciutat Sanitària i Universitàr ia de Bellvitge 9 2 1
Hospital Universitari Germans Trias i Pujol 9 1 1
Consell Superior d'Investigacions Científiques 6 7 10
Comunitat Valenciana 5 - 6
Fundació Jordi Gol i Gurina 4 - 1
Centre de Regulació Genòmica 4 - -
Hospital Mútua de Terrassa 4 - -
Institut de Recerca Oncològica 4 5 -
Universitat de Girona 4 - 2
Consorci Hospitalari del Parc Taulí. Sabadell 3 - -
Hospital Josep Trueta de Girona 3 - -
Agència de Salut Pública de Barcelona 2 - -
Hospital General de l'Hospitalet 2 - -
Universitat Internacional de Catalunya 2 - -
Universitat Pompeu Fabra 2 1 3
Centre d'Investigació Cardiovascular de Barcelona 1 - -
Fundació Puigvert 1 - -
Hospital de Figueres 1 - -
Hospital de Mataró 1 - -
Hospital Universitari Joan XXIII. Tarragona 1 1 -
Hospital Universitari Sant Joan de Déu 1 2 -
Hospital Universitari Sant Joan. Reus 1 - -
Hospital Verge de la Cinta. Tortosa 1 - -
Illes Balears 1 1 -
Institut Català d'Investigació Química. Tarragona 1 - -
Hospital Universitari Arnau de Vilanova. Lleida - 1 1
Universitat Ramon Llull - 1 1
Universitat de Lleida - 1 -
Madrid - - 13
EUA - - 6
França - - 4
Aragó - - 2
Canàries - - 2
Galícia - - 2
País Basc - - 1
Universidad de Salamanca - - 2
Amsterdam University - - 1
CETIR Grup Mèdic - - 1
Cutaneous Cell Biology. Londres - - 1
Escola Politècnica Superior de Vic - - 1
Fundació irsiCaixa - - 1
Katholieke Universiteit Leuven. Bèlgica - - 1
Kurume University. Japó - - 1
Universidad de Granada - - 1
Universidad de Oviedo - - 1
Universitat Politècnica de Catalunya - - 1
University of Bergen - - 1
Total projectes unitaris 214
Total projectes coordinats 46
Total projectes presentats 260
* Aquests subprojectes formen part de projectes coordinats presentats per altres centres.
C e n t res dels equips de re c e rca que van presentar projectes a la convocatòria de La Marató 2004

FUNDACIÓ LA MARATÓ DE TV3
Avaluació dels
projectes de recerca
Càncer. La Marató 2004
El procés d’avaluació dels projectes de
recerca biomèdica s’inicia un cop s’han
registrat i revisat els projectes presentats dins
el termini de la convocatòria. L’Agència
d’Avaluació de Tecnologia i Recerca
Mèdiques (AATRM) és la responsable de dur
a terme la gestió d’aquest procés, seguint un
procediment que en garanteix el màxim de
transparència i objectivitat.
En aquest exercici es van avaluar els projec-
tes de recerca biomèdica sobre La Marató
del 2004, dedicada al càncer.
La selecció dels avaluadors es va fer, bàsi-
cament, per mitjà de tres fonts d’informació:
· Llista de professionals de reconegut prestigi
internacional en recerca sobre càncer, facili-
tat pels membres de la Comissió Assessora
Científica de la Fundació, pels membres del
Comitè Científic de l’AATRM i pels membres
del Consell Assessor del Pla Director
d’Oncologia de Catalunya.
· C e rca bibliogràfica amb la qual poder
valorar la trajectòria i producció científica
dels darrers anys dels avaluadors potencials.
· Revisió de la composició tant dels comitès
científics de les conferències internacionals
sobre el càncer celebrades darrerament
com dels comitès editorials de revistes
biomèdiques dedicades a l’oncologia.
Un cop revisada la llista resultant, es va
convidar 1.141 experts a participar-hi, dels
quals van re s p o n d re 407 i van acceptar 235.
Finalment, van ser 169 els especialistes que
van participar en aquest procés d’avaluació,
el 67% dels quals eren europeus.
El procés d’avaluació inclou dues fases per
a cada projecte: la valoració qualitativa, duta
a terme pels avaluadors, i la valoració quanti-
tativa, presentada per l’AATRM.
En una primera fase, dos avaluadors de
forma independent (peer-review) revisen un
projecte anonimitzat en funció de diversos
aspectes. Seguidament, els avaluadors
retornen el projecte a l’AATRM i reben la part
curricular de l’equip d’investigadors perquè
tornin a fer-ne una valoració amb un qüestio-
nari estructurat.
Si les valoracions dels dos avaluadors són
discordants, el projecte s’envia a un tercer,
que el revisa i valora sense conèixer els
resultats de les avaluacions prèvies. Els
resultats de l’avaluació qualitativa es basen
en la qualitat metodològica i científica i en la
rellevància del projecte, i se n’elaboren els
grups següents: finançables, finançables
amb reserves, dubtosos i no finançables.
La valoració quantitativa es fa per la mitjana
aritmètica de les dues (o tres) puntuacions
calculades amb les valoracions de cadascun
dels avaluadors, servint d’ajut a la classifica-
ció final dels projectes mitjançant les
tècniques estadístiques.
Finalment, es crea un comitè ad hoc, integrat
per 10 avaluadors europeus, els quals, en
una reunió a Barcelona, prioritzen els projec-
tes més ben classificats amb criteris explícits
i establerts a priori, per ser recomanats a la
Comissió Assessora Científica de la Fundació
perquè hi doni finançament. Finalment, és el
Patronat de la Fundació qui decideix els
projectes que rebran els fons recaptats amb
La Marató de TV3.

Avaluadors dels projectes de re c e rca presentats a la convocatòria de La Marató 2004
RECERCA BIOMÈDICA | 45
Adams, Jean School of Population and Health Sciences, Regne Unit
University of Newcastle upon Tyne
Adjei, Alex A. Mayo Clinic EUA
Agus, David Cedars-Sinai EUA
Ahlbom, Anders Karolinska Institute Suècia
Akdis, Cezmi A. Swiss Institute of Allergy and Suïssa
Asthma Research-Head of Immunology Dept
Almadori, Giovanni Università Cattolica del Sacro Cuore Itàlia
Andre, Fabrice Institut Gustave Roussy França
Andreakos, Evangelos Foundation for Biomedical Research of the Regne Unit
Academy of Athens, Greece and Imperial College
Antin, Joseph H. Dana-Farber Cancer Institute, EUA
Harvard Medical School
Auvinen, Anssi Tampere School of Public Health, Finlàndia
University of Tampere
Awada, Ahmad Jules Bordet Institute Bèlgica
Bardelli, Alberto Institute For Cancer Research and Itàlia
Treatment/University of Torino School of Medicine
Barkardottir, Rosa Björk Landspitali - University Hospital of Iceland Islàndia
Bentzen, Søren M. University of Wisconsin Medical School, EUA
Department of Human Oncology
Berr ino, Franco Istituto Nazionale Tumori Itàlia
Biroccio, Annamar ia Regina Elena Cancer Institute Itàlia
Bjerrum, Lars University of Southern Denmark Dinamarca
Blobel, Carl P. Hospital for Special Surgery, EUA
Weill Cornell Medical College
Boccardo, Francesco University of Genoa and National Itàlia
Cancer Research Institute
Boddy, Alan University of Newcastle Regne Unit
Bolumar Montrull, Francisco Universidad Miguel Hernández Espanya
Borreani, Claudia National Cancer Institute Itàlia
Bottomley, Andrew European Organisation for Research Bèlgica
and Treatment of Cancer (EORTC)
Braakhuis, Boudewijn J. M. VU University Medical Center Països Baixos
Brown, Julia Clinical Trials Research Unit, University of Leeds Regne Unit
Canellos, George Peter Dana-Farber Cancer Institute EUA
Harvard Medical School
Caramori, Gaetano Centro di Ricerca su Asma e BPCO, Itàlia
University of Ferrara
Cassidy, James Centre for Oncology & Applied Pharmacology, Regne Unit
University of Glasgow
Chang, Ting-Chang Chang Gung Memorial Hospital Taiwan
Linkou Medical Center
Chao, Ann Centers for Disease Control and Prevention Zàmbia
Chen, Wendy Y. Dana Farber Cancer Institute /
Brigham and Women's Hospital EUA
Chen, Xiaoxin North Carolina Central University (NCCU) EUA
Clarke, Paul R. University of Dundee Regne Unit
Cor tes, Jorge E. M.D. Anderson Cancer Center EUA
Darby, Sarah C. Cancer Research UK and University of Oxford Regne Unit
DeAngelo, Daniel J. Dana-Farber Cancer Institute EUA
Debatin, Klaus-Michael University Children's Hospital Alemanya
Delattre, Olivier INSERM França
Delgado Rodríguez, Miguel Universidad de Jaén Espanya
Denkert, Carsten Charité University Hospital, CCM, Alemanya
Institute of Pathology
Di Nicola, Massimo Istituto Nazionale Tumori Itàlia
Edwards, Dylan R University of East Anglia Regne Unit
Avaluador Centre País

FUNDACIÓ LA MARATÓ DE TV3
Ehrenstein, Michael University College London Regne Unit
Eisbruch, Avraham University of Michigan EUA
Expósito Hernández, José Servicio Andaluz de Salud Espanya
Fernández, Carmen Immunology, WGI, Stockholm University Suècia
Fernández-Luna, José Luis Hospital Universitario Marqués de Valdecilla Espanya
Feunteun, Jean Institut Gustave Roussy França
FitzGerald, J Mark Centre for Clinical Epidemiology and Evaluation Canadà
Fulda, Simone University Children's Hospital Alemanya
Garcia-Aguilar, Julio Department of Surgery-University of California EUA
Geboes, Karel University Hospital KULeuven Bèlgica
Georgoulias, Vassilis School of Medicine, University of Crete Grècia
Gisselsson, David Department of Clinical Genetics, Lund University Suècia
Giulotto, Elena Università di Pavia, Dipar timento di Itàlia
Genetica e Microbiologia
Gridelli, Cesare Division of Medical Oncology, Itàlia
"S.G. Moscati Hospital"
Groneberg, David A. Charité School of Medicine, Alemanya
Free and Humboldt-University
Grützmann, Robert University Hospital Carl Gustav Carus, Alemanya
Dep. of Surgery
Habib, Fouad K. The University of Edinburgh Regne Unit
Halaban, Ruth Yale University School of Medicine EUA
Hilders, Carina Reinier de Graaf Hospital Països Baixos
Hoban, Paul Institute for Science and technology in Medicine, Regne Unit
Keele University medical School
Hung, Mien-Chie The University of Texas M. D. Anderson EUA
Cancer Center
Incrocci, Luca Erasmus MC-Daniel den Hoed Cancer Center Països Baixos
Jäättelä, Marja Danish Cancer Society Dinamarca
Jones, Dan MD Anderson Cancer Center EUA
Jones, Peter Lloyd University of Pennsylvania School of Medicine EUA
Karin, Michael University of California - San Diego EUA
Keith, W. Nicol University of Glasgow Regne Unit
Kesari, Santosh Dana Farber Cancer Institute/ EUA
Harvard Medical School
Kiriakidis, Serafim Imperial College, Faculty of Medicine Regne Unit
Kirkali, Ziya Dokuz Eylul University School of Medicine Turquia
Kohno, Hiroyuki Department of Oncologic Pathology, Japó
Kanazawa Medical University
La Vecchia,Carlo Istituto di Ricerche Farmacologiche Mario Negri Itàlia
Ladányi, Andrea National Institute of Oncology Hongria
Lalloo, Fiona Department of Clinical Genetics, St Mary's Hospital Regne Unit
Lam, Stephen British Columbia Cancer Agency Canadà
Lee, Sug Hyung Department of Pathology, College of medicine, Corea
The Catholic University of Korea
Lemoine, Nick Barts & The London School of Medicine, Regne Unit
Queen Mary University of London
Leonetti, Carlo Regina Elena Institute for Cancer Research Itàlia
Lin, Jennifer Hsiang-Ling Department of Medicine, Brigham and Women's EUA
Hospital, Harvard Medical School
Liu, Wai Institute for Cancer Research Regne Unit
Liu, Xuedong University of Colorado-Boulder EUA
Lock, Richard Children's Cancer Institute Australia for Austràlia
Medical Research, University of New South Wales
Loehr, Matthias Univ. of Heidelberg, Mannheim Medical Faculty Alemanya
Lønning, Per Eystein Section of Oncology, Institute of Medicine, Noruega
Medical Faculty University of Bergen
Avaluador Centre País

RECERCA BIOMÈDICA | 47
Lorenzo Bermejo, Justo German Cancer Research Centre Alemanya
Lu, Xin Ludwig Institute For Cancer Research Regne Unit
Lubinski, Jan Pomeranian Medical University Polònia
Lui, Weng-Onn Stanford University EUA
Maitland, Norman J YCR Cancer Research Unit, University of York Regne Unit
Manjer, Jonas Dept. of Community health (also dept. of Surgery), Suècia
Malmö University Hospital, Lund University
Mantovani, Alberto Istituto di Ricerche Farmacologiche Mario Negri Itàlia
Markman, Maurie M.D. Anderson Cancer Center Regne Unit
Marsiglia, Hugo Institut Gustave Roussy França
Martin, Seamus Dept. of Genetics, Trinity College Irlanda
Massagué, Joan Sloan-Kettering Institute EUA
Maur i, Claudia University College London Regne Unit
Mazzolini, Guillermo D. Austral University Argentina
McTiernan, Anne Fred Hutchinson Cancer Research Center EUA
Miralbell, Raymond Service de Radio-oncologie, Hôpitaux Suïssa
Universitaires de Genève
Mischel, Paul S. UCLA School of Medicine EUA
Mocellin, Simone University of Padova Itàlia
Mollenhauer, Jan German Cancer Research Institute Alemanya
Mooi, Wolter J. Vrije Universiteit Amsterdam Països Baixos
Moran, Elizabeth Fels Institute for Cancer Research, EUA
Temple University School of Medicine
Morgan, David A. L. Nottingham City Hospital Regne Unit
Müller-Tidow, Carsten Dept. of Medicine, Hematology and Oncology Alemanya
Nelson, Stanley F. University of California, Los Angeles EUA
Oh, William Harvard Medical School and EUA
Dana-Farber Cancer Institute
O'Morain, Colm Adelaide and Meath Hospital Irlanda
Osman, Iman New York University School of Medcine EUA
Park, Keunchil Division of Hematology/Oncology, Corea
Dept of Medicine, Samsung Medical Center,
Sungkyunkwan University School of Medicine
Parmiani, Giorgio Istituto Nazionale Tumor i Itàlia
Pelucchi, Claudio Istituto di Ricerche Farmacologiche Mario Negri Itàlia
Peterlin, Borut Division of Medical Genetics, Eslovènia
University Medical Center
Peters, Godefridus J. VU University Medical Center Països Baixos
Plaschke, Jens University of Technology Dresden, Alemanya
Department of Surgical Research
Pollán Santamaría, Marina Centro Nacional de Epidemiología Espanya
Polsky, Daniel University of Pennsylvania EUA
Ponzetto, Carola University of Torino Itàlia
Puigserver, Pere Johns Hopkins School of Medicine EUA
Pützer, Brigitte Dept. of Vectorology and Exp. Gene Therapy, Alemanya
University of Rostock
Rachmilewitz, Daniel Shaare Zedek Medical Center Israel
Repetto, Lazzaro Istituto Nazionale di Riposo e Cura Itàlia
per Anziani - INRCA
Rome, Leonard H. David Geffen School of Medicine at EUA
UCLA and Jonsson Cancer Center
Rouzier, Roman IGR França
Roz, Luca Istituto Nazionale Tumor i Itàlia
Sahai, Erik Cancer Research UK Regne Unit
Said, Jonathan UCLA David Geffen School of Medicine EUA
Sancho Garnier, Hélène Faculté de Médecine, Université França
Sarbia, Mario Department of Pathology, Technical University Alemanya
Avaluador Centre País

FUNDACIÓ LA MARATÓ DE TV3
Sauerwein, Wolfgang University Duisburg-Essen Alemanya
Scagliotti, Giorgio Vittorio University of Torino - Department of Itàlia
Clinical & Biological Sciences - S. Luigi Hospital -
Regione Gonzole 10 -
Schmitt, Manfred Clinical Research Unit, Department of Alemanya
Obstetrics and Gynecology (Frauenklinik),
Technische Universitaet Muenchen
Schmitt, Fernando Carlos Medical Faculty of Porto University and IPATIMUP Por tugal
Schouten, Leo J. Department of Epidemiology, NUTRIM,
Maastricht University Països Baixos
Semenzato, Gianpietro Padua University School of Medicine Itàlia
Sharma, Ricky A. Royal Marsden Hospital Regne Unit
Shu, Xiao Ou Vanderbilt University EUA
Shuai, Ke University of California EUA
Singer, Christian University of Vienna Medical Center and Àustria
Harvard University School of Public Health
Slade, Martin Imperial College of Science Technology Regne Unit
and Medicine
Sorensen, Henrik Toft Department of Clinical Epidemiology, Dinamarca
Aarhus University Hospital
Span, Paul N. Radboud University Nijmegen Medical Centre Països Baixos
Sparreboom, Alex National Cancer Institute EUA
Spurr, Nigel K. GlaxoSmithKline Regne Unit
Starkschall, George The University of Texas M. D. Anderson EUA
Cancer Center
Stoecklein, Nikolas H. University Hospital Düsseldorf Alemanya
Sun, Ren University of California EUA
Sverremark-Ekström, Eva Dept of Immunology, The Wenner-Gren Suècia
Institute, Stockholm University
Syngal, Sapna Dana-Farber Cancer Institute EUA
Teixeira, Manuel R. Department of Genetics, Por tugal
Portuguese Oncology Institute-Porto
Terracini, Benedetto University of Torino Itàlia
Tímár, József Tumor Progression, National Institute of Oncology Hongria
Van Dekken, Herman Erasmus Medical Center Països Baixos
Vanhoefer, Udo Department of Internal Medicine, Alemanya
Marienkrankenhaus
Vernon, Sally W. University of Texas at Houston School of EUA
Public Health
Weiderpass Vainio, Norwegian Cancer Registry, Oslo, Norway, Noruega
Elisabete and Karolinska Institutet, Stockholm, Sweden
West, Catharine The University of Manchester EUA
Willis, Monte S. Univ. of Nor th Carolina EUA
Wilson, Sue Department of Primary Care,
University of Birmingham Regne Unit
Wu, Lily University of California EUA
Xu, Xiaochun UT MD Anderson Cancer Center EUA
Young, Lawrence S Division of Cancer Studies,
University of Birmingham Regne Unit
Yuan, Jian-Min USC/Norris Cancer Center, Keck School , EUA
of Medicine University of Southern California
Zhang, Ruiwen University of Alabama at Birmingham EUA
Zheng, Wei Vanderbilt Univesity Cancer Center EUA
Zinzani, Pier Luigi Institute of Hematology Seragnoli Itàlia
Avaluador Centre País

RECERCA BIOMÈDICA | 49
El Patronat de la Fundació va aprovar l’adju-
dicació dels fons obtinguts durant La Marató
del 2003, dedicada a les malalties respiratò-
ries cròniques, a 27 projectes, d’un total de
105 projectes presentats a concurs.
L’acte acadèmic públic de lliurament d’ajuts
es va celebrar el 17 de novembre de 2004
a l’Aula Magna de la Universitat de
Barcelona (UB).
El rector de la UB, Joan Tugores, va presidir
l’acte, acompanyat per la consellera de
Salut, Marina Geli, i pel conseller del
Departament d’Universitats, Recerca i
Societat de la Informació, Carles Solà. L’acte
també va comptar amb la presència del
president de la Fundació, Joan Majó, a més
del coordinador de la Comissió Assessora
Científica de la Fundació, José J. Navas, del
vicerector d’Estudiants de la UB, Joan
Guàrdia, i del director executiu de la
Fundació, Miquel Vilar.
En el decurs d’aquesta jornada, el Dr. Jordi
Roca, cap de Secció del Servei de
Pneumologia de l’Hospital Clínic de
Barcelona i professor titular de la Facultat de
Medicina de la UB, va fer una anàlisi del futur
de la recerca sobre les malalties pulmonars
obstructives cròniques (MPOC) i el seu
impacte assistencial, tot destacant la
importància que tindran en aquest futur els
diners procedents dels fons aconseguits
amb La Marató.
Investigadors que rebran finançament
Dr. Carlos Agustí García-Navarro
Hospital Clínic i Provincial de Barcelona
MPOC: Resposta inflamatòria durant la
deshabituació tabàquica
52.200 €
Dr. Josep M. Antó Boqué
Institut Municipal d'Investigació Mèdica
Caracterització fenotípica i evolució de la
malaltia pulmonar obstructiva crònica
(MPOC): estudi PAC-COPD
102.000 €
Dr. Josep Maria Argilés Huguet
Facultat de Biologia. UB
Estudi dels mecanismes moleculars respon-
sables del desgast muscular dels pacients
amb MPOC: implicacions terapèutiques
72.300 €
Dr. Josep M. Argimón Pallàs
DAP Costa de Ponent. Fundació Jordi Gol i
Gurina
Estudi de les variants genètiques (SNP)
relacionades amb deixar de fumar
179.563 €
Dr. Ferran Barbé Illa
Hospital Son Dureta. Palma de Mallorca
Cèl·lules progenitores endotelials circulants
en pacients amb síndrome d'apnees
obstructives del son. Implicacions en risc
cardiovascular
16.740 €
Dr. Joan Albert Barberà Mir
Hospital Clínic i Provincial de Barcelona
Estudi de la contribució de les cèl lules
progenitores vasculars en els mecanismes de
Adjudicació i acte
de lliurament d’ajuts
Malalties respiratòries cròniques. La Marató 2003

lesió i reparació vascular a la circulació
pulmonar en la malaltia pulmonar obstructiva
crònica (MPOC)
140.000 €
Dr. Jaume Canet Capeta
Hospital Universitari Germans Trias i Pujol
Malaltia respiratòria crònica i tabaquisme en
els pacients quirúrgics a Catalunya: perfil
epidemiològic, factors predictius de l'evolució
postoperatòria i qualitat de vida als tres
mesos
100.000 €
Dr. Pere Casan Clara
Hospital de la Santa Creu i Sant Pau
Dr. Pere Joan Riu Costa
Centre de Recerca en Enginyeria Biomèdica.
UPC
Monitorització no invasiva del patró respirato-
ri en pacients amb malaltia pulmonar
obstructiva crònica (MPOC)
101.750 €
Dra. Josefina Casas Brugulat
Institut d'Investigacions Químiques i
Ambientals de Barcelona
Dr. Antonio Delgado Cirilo
Facultat de Farmàcia. UB
Dr. Antonio Gómez Muñoz
Facultad de Ciencia y Tecnología.
Universidad del País Vasco
Patogènesi de la malatia pulmonar obstructiva
crònica: paper dels esfingolípids bioactius en
la regulació de l'apoptosi de les cèl·lules
alveolars i avaluació de quimioteques lipídi-
ques per a una potencial utilització terapèutica
234.342 €
Dr. Joan Artur Caylà Buqueras
Institut de Medicina i Salut. Agència de Salut
Pública de Barcelona
Tuberculosi en immigrants: una malaltia
importada?
145.651 €
Dr. José Antonio Domínguez Benítez
Hospital Universitari Germans Trias i Pujol
Inflamació local i sistèmica en pacients amb
malaltia pulmonar obstructiva crònica:
determinació de proteïnes de fase aguda i
citocines en l'exacerbació infecciosa
150.000 €
Dr. Esteve Fernández Muñoz
Institut Català d'Oncologia
Dr. Manel Nebot Adell
Institut de Medicina i Salut. Agència de Salut
Pública de Barcelona
Dr. José A. Pascual Esteban
Institut Municipal d'Investigació Mèdica
Símptomes respiratoris, asma i tabaquisme
passiu en escolars: efectivitat d'una interven-
ció individual, familiar i escolar
283.790 €
Dr. Jordi Gratacós Masmitjà
Hospital de Sabadell
Prevalença d'osteoporosi en homes amb
malaltia pulmonar obstructiva crònica.
Valoració dels factors de risc per osteoporosi
i utilitat de la densitometria per predir el risc
de fractures osteoporòtiques
109.062 €
Dr. Emmanouil Kogevinas
Institut Municipal d'Investigació Mèdica
Gens, factors ambientals i asma: un estudi
internacional
119.088 €
Dra. Concepción Larraz Gimeno
Centre de Salut Escola Graduada. Institut
Balear de Salut
Eficàcia de la rehabilitació respiratòria en
pacients amb MPOC moderada en atenció
primària i el manteniment dels beneficis als
dos anys
44.856 €
Dra. Marina Luquin Fernández
Facultat de Ciències. UAB
Dr. Carles Alonso Tarrés
Hospital General de l'Hospitalet
Noves eines per al diagnòstic i tractament de
la tuberculosi. Resposta immunològica
integrada respecte d’antígens lipídics i glicoli-
pídics de Mycobacterium tuberculosis
150.000 €
Dr. Frederic Manresa Presas
Hospital Universitari de Bellvitge
Colonització de les vies aèries dels pacients
amb MPOC greu per Aspergillus sp. Estudi
de prevalença i anàlisi dels factors associats
95.231 €
Dra. Concepción Montón Soler
Hospital de Sabadell
Aguditzacions infeccioses de la MPOC:
factors de risc per aïllar Pseudomonas
aeruginosa i influència dels gens de virulència
en la mortalitat
150.000 €
FUNDACIÓ LA MARATÓ DE TV3

RECERCA BIOMÈDICA | 51
Dr. Josep Morera Prat
Hospital Universitari Germans Trias i Pujol
Estudi de la colonització bacteriana bronquial
en la MPOC. Efecte del tractament antibiòtic
en l'eradicació i prevenció de les exacerba-
cions
137.640 €
Dr. Juan Jesús Pérez González
Escola Tècnica Superior d'Enginyeria
Industrial. UPC
Dr. Josep Castells Boliart
Institut Universitari de Ciència i Tecnologia.
Mollet
Identificació, síntesi i cribratge biològic de
nous caps de sèrie mitjançant una aproxima-
ció de disseny racional de fàrmacs per a
l'antagonisme de bradiquina com a estratè-
gia terapèutica antiasmàtica
180.300 €
Dr. Vicente Plaza Moral
Hospital de la Santa Creu i Sant Pau
Mecanismes inflamatoris i causes de l’asma
de risc vital d'instauració fulminant
46.350 €
Dra. Laura Pujols Tarrés
Hospital Clínic i Provincial de Barcelona
Dr. Cristóbal Mezquita Pla
Facultat de Medicina. UB
Efecte dels inhibidors reversibles del protea-
soma sobre els mecanismes implicats en la
inflamació i el remodelat en l'asma
100.000 €
Dr. Jordi Rello Condomines
Hospital Universitari Joan XXIII
Pneumònia associada a ventilació mecànica
en malalts amb MPOC. Factors de risc i
impacte evolutiu
150.000 €
Dr. Josep Roca Torrent
Hospital Clínic i Provincial de Barcelona
Atenció integrada i biopatologia de la MPOC
avançada
149.088 €
Dr. Robert Rodriguez Roisin
Hospital Clínic i Provincial de Barcelona
Dr. Antonio Román Broto
Hospital Universitari Vall d'Hebron
Correlacions anatomofuncionals de l'inter-
canvi gasós en malalts trasplantats de pulmó
per MPOC avançada
109.500 €
Dra. Beatriz Rosón Hernández
Hospital de Mataró
Estudi clínic de la importància de la infecció
vírica en l’exacerbació aguda de la malaltia
pulmonar obstructiva crònica amb èmfasi a
reduir amb seguretat l'ús d'antibiòtics
100.860 €
Dr. Antoni Torres Martí
Hospital Clínic i Provincial de Barcelona
Dra. Rosario Menéndez Villanueva
Hospital Universitari La Fe. València
Factors de risc i pronòstic de la pneumònia
adquirida a la comunitat en el pacient amb
malaltia pulmonar obstructiva crònica.
Impacte de la resposta inflamatòria
100.000 €

FUNDACIÓ LA MARATÓ DE TV3
Per concretar les condicions del finançament
dels projectes adjudicats i fer-ne efectius els
pagaments, la Fundació va signar un conveni
amb cadascun dels investigadors principals
premiats.
Els treballs de recerca tenen, com a màxim,
una durada de tres anys i el pagament es fa
anualment en les quantitats acordades en el
pla de finançament.
Els pagaments es fan una vegada emesa
l’avaluació positiva de la memòria econòmica
i de la científica corresponent.
El control econòmic i de despesa dels
projectes el du a terme la Fundació, i
l’avaluació de la memòria científica de l’anua-
litat la du a terme l’AATRM.
1996 1 14.424,29
1997 1
18.244,47
1998 1 2 25 67
58.107,07
1999 4 8 14
196.885,64
2000 2 14
308.719,73
2001 6 17
644.196,67
2002 15
478.248,74
2003 37 1.447.233,23
Total Memòries gestionades 213
Total finançat 3.166.059,84
Marató Convenis Memòries 1a. Memòries 2a. Mem. Finals Mem. Resum Diners(pagaments) anualitat anualitat Simposi destinats �
C o n t rol econòmic i científic de projectes de re c e rca. Exercici juliol 2004 - juny 2005
S e g u i m e n t
d e l s p r o j e c t e s

RECERCA BIOMÈDICA | 53
La Fundació organitza el Simposium amb la
finalitat de donar a conèixer de manera
pública i directa el resultat dels projectes de
recerca que són finançats amb els diners
recollits amb La Marató de TV3, a càrrec
dels investigadors principals que n’expliquen
els avenços obtinguts.
Dins el període d’activitats d’aquesta
Memòria, es va celebrar el 21 d’octubre de
2004, a la sala d’actes del Departament de
Salut a Barcelona, el V Simposium, sobre la
diabetis i altres malalties metabòliques, que
van centrar La Marató del 1998, organitzat
conjuntament amb l’Agència d’Avaluació de
Tecnologia i Recerca Mèdiques, del
Departament de Salut.
En aquest acte de debat científic sobre la
diabetis i altres malalties metabòliques, es
van presentar estudis sobre els mecanismes
causals de la malaltia, tot determinant els
aspectes patogènics i d’afectació cel·lular i
desenvolupant models animals i marcadors
precoços de les diverses patologies, entre
altres avenços exposats.
Les memòries finals d’aquests projectes es
van editar en un llibre en format CD-ROM, en
tres idiomes, destinat als especialistes i als
principals centres de recerca biomèdica.
A continuació es detallen els resums de les
memòries finals dels projectes finançats per
la Fundació sobre la diabetis i altres malalties
metabòliques, com també les dades que
resumeixen els indicadors bibliomètrics de la
producció científica d’aquests projectes.
Finalització delsprojectes. Simposium
Diabetis. La Marató 1998
Inauguració del V Simposium

FUNDACIÓ LA MARATÓ DE TV3
Identificació i anàlisi funcional de gens implicats en la fisiopatologia
de la diabetis mellitus de tipus 2 11 11 (100%)
Anàlisi molecular i funcional de mutacions al DNA mitocondrial
implicades en sordesa progressiva i diabetis 12 10 (83%)
La proteïna desacobladora mitocondrial UCP3,
nova diana terapèutica de l'obesitat i diabetis tipus 2 8 7 (88%)
Homocisteïna i malaltia: estudis genètics, bioquímics i clínics
de l'homocistinúria i del risc cardiovascular 9 7 (78%)
Dissecció d'un circuit transcripcional que controla la funció
de les cèl·lules productores d'insulina 6 6 (100%)
Estudi del sistema antioxidant i de l'estrès oxidatiu en pacients
fenilcetonúrics 6 6 (100%)
Estudi de la maquinària molecular secretora d'insulina
i de la translocació del transportador de glucosa GLUT4.
Canvis funcionals i d'expressió relacionats amb la diabetis 6 5 (83%)
OPTIDIAB: model d'investigació per a la gestió i economia de la salut,
i d'optimització de la qualitat de l'assistència integrada a la diabetis mellitus 5 5 (100%)
L'alteració del lloc d'unió del malonil-CoA a la carnitina palmitoil transferasa
com a agent causal de la diabetis mellitus no dependent d'insulina (NIDDM) 6 4 (67%)
La carnitina palmitoiltransferasa I (CPTI) com a diana
farmacològica a la diabetis tipus 2 4 4 (100%)
Estudi de l'efecte terapèutic de les neurotrofines en les neuropaties
diabètiques 3 3 (100%)
Diabetis mellitus, sobrecàrrega cel·lular de ferro i prevalença de
mutacions del gen HFE en la població espanyola 3 3 (100%)
Estudi dels factors determinants de l'amiloidogènesi a la cèl·lula beta
pancreàtica i la seva relació patogènica amb la diabetis tipus 2 4 3 (75%)
Paper modulador de la resposta biològica a la reacció de Maillard en
l'evolució de la nefropatia en un model experimental de diabetis 3 2 (67%)
Efecte de la hiperglicèmia en el fracàs primari del trasplantament d'illots
pancreàtics: contribució a la mort i la disfunció de les cèl·lules beta
trasplantades 3 2 (67%)
Adaptacions fisiològiques a la modificació del quocient oferta/demanda
d'insulina 3 2 (67%)
Anàlisi mutacional del domini intracel·lular "x1c" del gen del receptor 2
del factor de necrosi tumoral alfa en pacients amb diabetis mellitus tipus 2 3 2 (67%)
Expressió de VEGF i angiopoetina-2 en un model de retinopatia diabètica 2 2 (100%)
Antidiabètics orals. Nous procediments de síntesi d'azasucres inhibidors
de glucosidases 1 1 (100%)
Valoració de l'estrès oxidatiu i la fibrogènesi en la patogènia
de la nefropatia diabètica. Influència del control glicèmic i del
consum de tabac 1 1 (100%)
IFNbeta i predisposició genètica a la diabetis tipus 1: anàlisi comparativa
en el ratolí transgènic pINS-IFNbeta no obès diabètic (NOD) i
no obès resistent (NOR) 2 1 (50%)
Neogènesi dels illots pancreàtics: identificació de l'expressió diferencial
dels gens induïts per tungstat 2 1 (50%)
Caracterització fisicoquímica i funcional de la subfracció modificada,
electronegativa de les lipoproteïnes de baixa densitat (LDL[-])en
la diabetis mellitus. Influència del perfil glicèmic 1 1 (100%)
Virus de l'hepatitis C com a factor de risc per desenvolupar una diabetis
mellitus: estudi dels mecanismes etiopatogènics 1 1 (100%)
Aproximacions de teràpia gènica per a la diabetis mellitus centrades
en la manipulació genètica del pàncrees i teixits extrapancreàtics 1 1 (100%)
Anàlisi del paper dels activadors del plasminogen (PAs) a
la nefropatia diabètica. Establiment del model d'inducció experimental
de diabetis a ratolins Null pels PAs. 1 1 (100%)
TOTAL 109 92 (84%)
Nombre de treballs Documents Coberturadel projecte atribuïbles % dels treballs que
La Marató de TV3 s'han trobat documentsatribuibles
Convocatòria 1998: Diabetis i altres malalties metabòliques: literatura generada

RECERCA BIOMÈDICA | 55
43 3,91 18,2% 1,39 1,81 0,75 3,56 100,0%
143 14,30 20,0% 1,80 3,07 2,44 7,77 100,0%
10 1,43 57,1% 0,20 0,19 0,00 7,10 20,0%
4 0,57 57,1%
46 7,67 50,0% 2,31 1,34 1,31 8,624 0,0%
6 1,00 50,0% 0,29 0,38 0,58 1,62 0,0%
5 1,00 80,0% 0,00 0,00 4,04 0,0%
8 1,60 60,0% 0,00 0,00 0,57 50,0%
13 3,25 0,0% 2,16 3,53 0,60 6,18 33,3%
13 3,25 0,0% 0,69 1,11 0,34 4,88 66,7%
31 10,33 0,0% 4,53 8,55 1,74 8,18 100,0%
6 2,00 33,3% 0,28 0,36 0,0%
2 0,67 33,3% 0,43 0,43 0,69 1,57 0,0%
3 1,50 0,0% 2,03 3,48 100,0%
3 1,50 0,0% 2,45 3,74 0,0%
0 0,00 100,0% 0,00 0,00 0,00 5,49 0,0%
16 8,00 0,0%
6 3,00 0,0% 0,33 0,52 0,51 3,06 0,0%
1 1,0 0,0% 0,53 0,49 0,0%
1 1,00 0,0%
0 0,00 100,0%
2 2,00 0,0%
1 1,00 0,0% 0,33 0,46 0,0%
0 0,00 100,0%
1 1,00 0,0% 1,31 4,38 0,0%
1 1,00 0,0%
365 3,97 33,7% 1,13 1,61 1,17 5,58 39,1%
Citacions rebudes Citacions / Percentatge Share Share Citation Impact Mitjana del Percentatge dedocuments Documents de No Citació Internacional Espanyol Factor Rate Factor d'Impacte col·laboració atribuïbles internacional

FUNDACIÓ LA MARATÓ DE TV3
Dr. Jordi Alberch ViéFacultat de Medicina. Fundació Clínic
per a la Recerca Biomèdica
Dr. Jaume Arroyo BrosDAP Sabadell. Fundació Jordi Gol i Gurina
Dra. Begoña Campos Bonilla Facultat de Medicina. Fundació Clínic
per a la Recerca BiomèdicaDra. Anna Sanmartí Sala
Hospital Universitari Germans Trias i Pujol
Dra. Susana Balcells ComasFacultat de Biologia. UB
Dra. M. Antònia Vilaseca BuscáHospital Universitari Sant Joan de Déu
Dr. Xavier Pintó Sala Ciutat Sanitària i Universitària de Bellvitge
Dr. Ramon Bartrons Bach
Facultat d'Odontologia. CSUB
Dr. Joan M. V. Blasi CabúsFacultat de Medicina. CSUB
Dr. Joan Bosch Cartes
Facultat de Farmàcia. UB
Dra. Fàtima Bosch TubertFacultat de Veterinària. UAB
Dr. Jaume Campistol Plana
Hospital Universitari Sant Joan de Déu
Dr. Enric Esmatjes MompóHospital Clínic i Provincial de Barcelona
Dr. Xavier Estivill Pallejà
Centre de Genètica Mèdica i Molecular. IRO
Dr. Jorge Ferrer MarradesHospital Clínic i Provincial de Barcelona
Dra. Cristina Fillat Fonts
Centre de Genètica Mèdica i Molecular. IRO
Dr. Jordi Garcia FernàndezDra. Gemma Marfany Nadal
Facultat de Biologia. UB
Dr. Fausto García HegardtFacultat de Farmàcia. UB
Dr. Joan Gené Badia
DAP Baix Llobregat Litoral. Fundació Jordi Gol i Gurina
Dr. Néstor Gómez Trias
Facultat de Veterinària. UAB
Dr. Ramon Gomis de BarbaràHospital Clínic i Provincial de Barcelona
Dr. Joan J. Guinovart Cirera
Facultat de Química. UB
Dra. Cristina Hernández PascualHospital Universitari Vall d'Hebron
Dr. Albert J. Jovell Fernández
Fundació Biblioteca Josep Laporte. UAB
Dr. Alberto de Leiva HidalgoHospital de la Santa Creu i Sant Pau
Dr. Pedro F. Marrero González
Facultat de Farmàcia. UB
Dr. Cristóbal Mezquita PlaFacultat de Medicina. UB
Dr. Eduard Montanya Mias
Ciutat Sanitària i Universitària de Bellvitge
Dra. Anna Novials SardàFundació Sardà Farriol
Dr. Ramon Gomis de Barbarà en subrogació dela Dra. Elena Fernández Usac
Hospital Clínic i Provincial de BarcelonaDra. Anne Clark
Oxford University
Dr. Rafael Oliva VirgiliHospital Clínic i Provincial de Barcelona
Dr. Jordi Ordóñez Llanos
Hospital de la Santa Creu i Sant Pau
Dra. Rosanna Paciucci BarzantiHospital Universitari Vall d'Hebron
Dr. Joan Prat Corominas
Facultat de Medicina. UdL
Dr. Francisco X. Real ArribasInstitut Municipal d'Investigació Mèdica
Dr. Manuel Reina del Pozo
Facultat de Biologia. UB
Dr. Cristóbal Richart JuradoHospital Universitari Joan XXIII. Tarragona
Dr. Emili Ros Rahola
Hospital Clínic i Provincial de Barcelona
Dr. Bernat Sòria EscomsInstitut de Bioenginyeria. UMH. Alacant
Dr. Francesc Villarroya Gombau
Facultat de Biologia. UB
Dra. Marta Vives PiHospital Universitari Germans Trias i Pujol
Dr. Antonio Zorzano Olarte
Facultat de Biologia. UB
Investigadors principals

RECERCA BIOMÈDICA | 57
Títol del projecte
Estudi de l’efecte terapèutic de les neurotrofines en les neuropaties diabètiques
Investigador principalJordi Alberch Vié
InstitucióFundació Clínic per a la Recerca Biomèdica
Membres de l’equipAna López, Raquel Martín, Susana Pezzi, Núria Gavaldà, Jesús Torres, Pep Canals, Juan M. García, Esther Pérez, Miquel Bosch i J.R. Pineda.
99.660 €
MEMÒRIA FINAL
1. Resum del projecte originalLes neurotrofines són molècules amb capacitat deprotegir d’algunes neuropaties perifèriques experimen-tals i actualment ja han començat a utilitzar-se enalguns assaigs clínics. Aquest estudi ha estat dissen-yat per analitzar els efectes de la implantació quirúrgi-ca de fibroblastos transfectats que produeixenneurotrofines i també l’efecte d’alguns inductors de lasíntesi endògena de neurotrofines en la prevenció i/oreversió de les neuropaties diabètiques experimentals,com a pas previ per a la seva possible utilització enhumans. L’estudi es farà en ratolins amb neuropatiadiabètica induïda per estreptozotozina (neuropatiasensitiva de fibres primes).El tipus d’anàlisis i l’estratègia per al tractament ambimplants de fibroblastos es determinarà per compara-ció de les lesions observades en els animals experi-mentals amb els patrons d’alteracions perifèriques enratolins transgènics, en els quals els gens que codifi-quen per les diverses neurotrofines o els seusreceptors han estat inactivats. Es faran estudis tant detipus electrofisiològic com de l’estructura anatòmicafina dels nervis perifèrics, dels ganglis raquidis i de la
part anterior de la medul·la en tots els animals. Elsresultats que s’obtinguin poden ser de gran interèsamb vista a nous tractaments de les neuropatiesperifèriques en pacients amb diabetis basats en l’apli-cació de les neurotrofines.
2. ResultatsEls resultats obtinguts en aquest treball proporcionennoves evidències sobre les interaccions tròfiques i lasupervivència de les neurones sensorials que podenaportar estratègies interessants per al tractament o laprevenció de la neuropatia perifèrica en pacientsdiabètics. En resum, tot i que ja era conegut quemoltes neurones sensorials nociceptives afectades enla neuropatia diabètica responien al factor neurotròficNGF, que ja s’havia arribat a utilitzar en assaigs clínicsper al tractament de les esmentades neuropaties,nosaltres hem determinat que una altra fracció deneurones nociceptives respon al BDNF i que, per tant,també podria ser utilitzat en aquests tipus de tracta-ments.Segons dades recollides als Estats Units, al Canadà ien 16 països europeus, més del 25% dels pacientsdiabètics desenvolupen algun tipus de neuropatiaperifèrica (American Diabetes Society i Eurodiab IDDM

Complications Study; vegeu, per exemple, Tesfaye icols., 1996, Diabetologia, 39:1377-1384). La formamés comuna de neuropatia diabètica és la neuropatiasensoriomotora simètrica distal, que afecta lesneurones amb axons més llargs del sistema motor,sensorial i autonòmic. Un dels grups de neuronessensorials més afectats són les nociceptives,neurones de dimensió petita situades als ganglis del’arrel dorsal, i que contenen com a molècules activesCGRP/substància P o expressen diferencialment elmarcador de membrana IB4. La pèrdua de sensaciódolorosa en pacients diabètics els predisposa alesions, especialment als peus, que s’infecten ambfacilitat i cicatritzen malament, la qual cosa condueixen molts casos a la necessitat d’amputació.La família de les neurotrofines inclou quatre polipèp-tids altament relacionats, tant des del punt de vistaestructural com funcional: el factor de creixementnerviós (NGF), el factor neurotròfic derivat del cervell(BDNF), la neurotrofina 3 (NT-3) i la neurotrofina 4/5(NT-4/5). Aquestes molècules promouen la supervi-vència de poblacions de neurones tant del sistemanerviós central com perifèric. Interaccionen ambreceptors de membrana del tipus tirosina quinasa,denominats trkA, trkB i trkC, i la fosforilació d’aquestsreceptors amb la unió de la neurotrofina corresponentdesencadena una cascada de senyals intracel·lularsque condueixen, en darrera instància, a la supervivèn-cia de la neurona. L’especificitat neurotrofina-receptorés la següent: trkA és el receptor per NGF, tant BDNFcom NT-4/5 activen el receptor trkB, i NT-3 és ellligand de trkC.Els estudis realitzats en mutants nuls per les neurotro-fines i els seus receptors han demostrat que durant eldesenvolupament embrionari les neurones sensorialsnociceptives són completament dependents d’NGF(Kirstein i Fariñas, 2002, Cell Mol. Life Sci., 59: 1787-802). En el cas de la neuropatia diabètica experimen-tal, s’ha observat que l’NGF exogen és capaç derevertir les alteracions funcionals i estructurals de lesfibres nervioses produïdes per una diabetis induïda enmodels experimentals (Anand i cols., 1996, NatureMedicine, 26:703-707; McMahon i Priestley, 1995,Current Opinion in Neurobiology, 5:616-624; Riaz iTomlinson, 1996, Progress in Neurobiology, 49:125-143). A més, un assaig clínic fase III va demostrarl’absència de factors nocius del factor i la millora defuncions neurològiques, incloent-hi la percepció defred i calor mesurats com a sensació dolorosa (edició121 de la Reunió Anual American NeurologicalAssociation, Miami 1996). No obstant això, l’NGFpresentava alguns efectes perniciosos, com era laproducció d’hiperalgèsia en la pell a la zona on es feiala injecció.D’altra banda, en adults, una part de les neuronessensorials de dimensió petita segueixen expressanttrkA, però no altres. Això fa pensar que altres factorsneurotròfics diferents de l’NGF siguin els encarregatsde mantenir aquestes neurones. Durant el desenvolu-pament embrionari i en el moment del naixement, el70-80% de les neurones sensorials dels DRG expres-sen el receptor trkA i exigeixen la presència d’NGF pera la seva supervivència (Smeyne et al., 1994, Nature,368: 246-248; White et al., 1996, J. Neurosci., 16:4662-4672). Aquest grup està format per neurones demida petita o intermèdia i amb axons envoltats per
una fina capa de mielina (fibres tipus Ad) totalmentamielínics (fibres tipus C). La majoria expressen unneuropèptid associat a la nocicepció anomenat CGRP(calcitonin gene-related peptide). Tanmateix, 15 diesdesprés del naixement, només el 40% de lesneurones sensorials expressen trkA o CGRP. La restade la població original pateix un canvi fenotípic enaquest període del desenvolupament postnatal, totgenerant una nova població de neurones de dimensiópetita que constitueixen el 25-35% del gangli, quedeixen de dependre d’NGF, expressen c-ret i esmarquen amb IB4, una lecitina de la plantaBandeiraea simplicifolia (vegeu, per exemple, Silos-Santiago i cols., 1995, J. Neurosci., 15: 5929—5942).Vam decidir esbrinar si altres neurotrofines prenien elrelleu en el manteniment d’aquesta segona població.
1. En absència de BDNF es produeix una pèrduad’axons amielínics postnatalmentEls recomptes de fibres amielíniques en les arrelsdorsals i nervis raquidis mostren una pèrdua (49%) defibres amielíniques. Aquestes observacions demostrenque en animals mutants nuls per BDNF es perdenneurones sensorials amb fibres amielíniques de tipus C.
2. En absència de BDNF es produeix una pèrduade terminacions lliures, característiques de lainnervació de neurones sensorials petites nocicep-tives, en la pell
BDNF+/+ BDNF-/-
Quan s’analitza la innervació cutània, tant de la pellgruixuda (en la figura anterior) com de la pell pilosaamb inmunomarcatges per terminacions sensorialslliures (de fibres amielíniques o mielíniques de petitcalibre corresponents a neurones sensorials nocicep-tives petites), s’observa una disminució significativaen la intensitat del marcatge, que es correspon ambla pèrdua de fibres amielíniques observada en elsnervis raquidis.
3. En absència de BDNF es produeix una pèrduade neurones sensorials petites en els ganglis, tantpositives per CGRP com per IB4:Dins del conjunt de neurones sensorials petites, es
FUNDACIÓ LA MARATÓ DE TV3

poden distingir després del naixement:1) una població de neurones que expressen trkA iCGRP que tenen fibres C i són nociceptives.2) una població de neurones que no contenen trkAperò sí c-ret i que són positives en tincions amb lalectina IB4. Per tal de determinar amb més precisiól’efecte de l’absència de BDNF sobre neurones delsDRG amb terminacions lliures en la pell, analitzem elnombre de neurones positives per CGRP o per IB4 entalls a través del gangli L5 en animals BDNF+/+ iBDNF-/- de P15. En els mutants s’observa unareducció en el percentatge de neurones positives tantper CGRP com per IB4, tot indicant que ambduespoblacions contenen una fracció de neuronesdependents de BDNF per a la seva supervivència.
4. Algunes neurones sensorials petites passen dedependre d’NGF a dependre de BDNF durant lesdues primeres setmanes de vida
Supervivència de neurones dels ganglis sensorials del’arrel dorsal obtingudes de ratolins normals adiferents edats postnatals (P, dia postnatal) i mantin-gudes durant 48 hores en presència de diferentsfactors neurotròfics a 10 ng/ml o en absència total desuport neurotròfic extern (0 NT). Es representa elnombre de neurones (mitjana ± SEM) que sobreviuendesprés de 2 DIV partint d’un nombre igual deneurones sembrades en tots els pouets. S’observacom totes les neurones sensorials es fan progressiva-ment més independents de l’administració exògenade neurotrofines. La majoria de les neurones responena NGF a totes les edats. La resposta tant a BDNFcom a GDNF, que inicialment és molt minoritària,augmenta amb l’edat postnatal fins a fer-se tanimportant com la d’NGF.Les neurones que responen a BDNF també responena NGF, però es produeix un canvi de dependèncianeurotròfica en aquestes neurones d’NGF a BDNF.
Les neurones que responen a BDNF o GDNF sónpoblacions inicialment diferents i acaben sent unamica solapants.
RECERCA BIOMÈDICA | 59

Supervivència de les neurones CGRP-positives delsganglis sensorials de l’arrel dorsal obtingudes deratolins normals a diferents edats postnatals (P, diapostnatal) i mantingudes durant 48 hores en presènciade diferents factors neurotròfics a 10 ng/ml o enabsència total de suport neurotròfic extern (0 NT). Esrepresenta el nombre de neurones (mitjana ± SEM)que sobreviuen després de 2 DIV partint d’un nombreigual de neurones sembrades en tots els pouets.S’observa com les neurones sensorials que contenenCGRP es fan progressivament més independents del’administració exògena de neurotrofines. La majoriade les neurones responen a NGF a totes les edats. Laresposta tant a BDNF com a GDNF, que inicialment ésmolt minoritària, augmenta amb l’edat postnatal fins afer-se tan important com la d’NGF.
Inicialment, el BDNF no manté gaires neuronesCGRP+, però a més sembla que inhibeix l’expressióde CGRP.Supervivència de neurones IB4+ dels ganglissensorials de l’arrel dorsal obtingudes de ratolinsnormals a diferents edats postnatals (P, dia postnatal) imantingudes durant 48 hores en presència dediferents factors neurotròfics a 10 ng/ml o enabsència total de suport neurotròfic extern (0 NT). Esrepresenta el nombre de neurones IB4+ (mitjana ±SEM) que sobreviuen després de 2 DIV partint d’unnombre igual de neurones sembrades en tots elspous. S’observa com les neurones sensorials es fanprogressivament més independents de l’administracióexògena de neurotrofines. La majoria de les neuronesresponen a NGF a totes les edats. La resposta tant aBDNF com a GDNF que inicialment és menor queamb NGF, augmenta amb l’edat postnatal fins a fer-setan important com la d’NGF.
S’observa com es produeix un canvi en la dependèn-cia neurotròfica d’una part de les neurones IB4+d’NGF a BDNF, de tal manera que a partir de P5 sóndues poblacions separades de neurones IB4+, lesque responen a NGF i les que responen a BDNF.
Les neurones IB4+ que responen a BDNF o GDNFsón poblacions inicialment diferents i acaben sent unamica solapants.Quan les neurones solapants arriben a la independèn-cia neurotròfica d’aquests factors, cap de lescondicions tròfiques utilitzades modifica l’expressió deCGRP o IB4.
5. El BDNF in vivo s’expressa intensament en lesneurones dels ganglis i és el factor neurotròfic mésexpressat, tant al gangli com a la pell
Mesures per PCR quantitativa a temps real dels nivellsd’NGF, GDNF i BDNF als ganglis de l’arrel dorsal(DRG) i a la pell (skin).
6. Estudi de les variacions de receptors perifèricsen models de neuropaties diabètiquesS’han utilitzat models diferents de diabetis, com són:a) la inducció de la diabetis per estreptozotozina(STZ): es van tractar ratolins de la soca C57BL6 d’unmes d’edat amb STZ (225 mg/kg) intraperitoneal, dosiúnica o múltiple; b) animals diabètics genèticament:s’han utilitzat ratolins de la soca C57BL/Ks/Rj db/db(Janvier España S.L) amb una mutació espontàniamonogènica recessiva (procedència: CSAL Orleáns,1981) localitzada al cromosoma 4. En els animalshomozigots, la diabetis es desenvolupa en duesfases. Els primers símptomes clínics apareixen a unaedat de 4-5 setmanes: obesitat, polifàgia, poliuropoli-dipsia. Paral·lelament apareixen alteracions metabòli-
FUNDACIÓ LA MARATÓ DE TV3

ques: hiperinsulinèmia que comença a l’edat de 14dies i, després, hiperglucèmia. En animals d’edatavançada s'observa una degeneració de les cèl·lulesb -pancreàtiques, augment de la insulinèmia i de lahiperglucèmia no controlable per l’administraciód’insulina (Coleman, 1978; Berglund, 1978).L’anàlisi immunohistoquímic de mecanoreceptors (C.Pacini i C.Meissner) en animals amb neuropatia tòxicai metabòlica indica que els corpuscles de Meissnerestan presents en tots els grups d’animals analitzats(control, STZ, db/db i cisplatí) en els coixinets plantarsimmunomarcats amb la proteïna S100. Aparentmentno existeixen canvis estructurals. Amb referència alcorpuscle de Pacini, no s’observen diferències claresen els diferents grups d’animals. Tots ells estan bendesenvolupats i la immunoreactivitat és similar en totslos casos. L’única diferència podria estar en el marcat-ge per S100 dels nuclis de les cèl·lules de Schwann,que podria indicar un estat de divisió cel·lular. Si esconfirmés, seria interessant el fet que no hi haguésimmunoreacció per p75 en els corpuscles de Pacinidels animals tractats amb cisplatí, ja que coincidiriaamb dades bibliogràfiques que mostren que en casosde neuropatia es produeix una disminució en l’expres-sió d’aquest receptor i, per tant, una menor capacitatde regeneració nerviosa.
7. Estudi dels ganglis raquidis d’animals ambneuropatia metabòlica i tòxicaEs va fer una anàlisi immunohistoquímica dels ganglisraquidis per analitzar els canvis en el citoesquelet,proteïnes lligadores de calci, receptors de neurotrofi-nes i altres factors tròfics. Els resultats obtinguts mésinteressants són els següents:
1. Citoesquelet neuronal:Només els animals controls presenten IR per a -actina, localitzada en els axons neuronals.Només els animals amb neuropatia tòxica expressenb -tubulina en els cossos de les seves neuronesmitjanes i grans.El neurofilament de 200 kDa no fosforilat (NE-14)s’expressa en els axons de tots els tipus animals,però no en els cossos de les neurones grans delsanimals controls i amb neuropatia tòxica ni en lesmitjanes o petites dels tractats amb STZ. Els animalsamb diabetis congènita no presenten IR per aquestneurofilament o en els seus cossos neuronals.La IR per PGP 9.5 és igual en tots els casos exceptela seva localització al nucli de les neurones delsanimals tractats amb cisplatí.L’NF de 150 kDa i el de 200 kDa (policlonal) fosforilatss’expressen en els cossos neuronals dels animalstractats amb CisPt.L’NF de 200 kDa fosforilat RT 97 s’expressa en elsaxons dels 4 tipus animals, com també en els seuscossos, excepte aquells animals tractats amb STZ, enquè només s’observa localització axonal.
2. Proteïnes lligadores de calci:Les subunitats S100a i b semblen no expressar-se enels animals controls i sí en la resta, tot i que d’unamanera no molt clara.Amb referència a la proteïna completa S100, la sevaIR és molt similar en els 4 grups d’animals, tot i queno sembla expressar-se en cèl·lules glials en els
animals tractats amb STZ i, en tots els casos, lalocalització de la proteïna és nuclear.
3. Receptors de neurotrofines:La diferència més notable sembla la manca demarcatge glial amb p75 en els animals controls, opotser podria ser que es donés una sobreexpressiódel receptor esmentat en els animals amb patologies.De totes formes, s’haurien de fer més proves. El trkCno s’expressa en cap grup animal i el trkA només enels animals tractats amb cisplatí.
3. Rellevància i possibles implicacions clíniquesdels resultats finals obtingutsEls resultats obtinguts en el projecte han servit percaracteritzar les necessitats tròfiques de les neuronessensorials afectades en la neuropatia diabètica. Ja eraconegut que aquestes neurones nociceptivesresponen a l’NGF, i es van iniciar alguns assaigsclínics. Els nostres treballs defineixen que hi ha uncanvi en les respostes tròfiques d’aquestes neuronesd’NGF a BDNF. Així, proposem que el BDNF pot serun bon candidat per utilitzar-lo com a agent neuropro-tector de les neurones sensorials afectades en laneuropatia diabètica.
4. PublicacionsFariñas, I., K.R. Jones, L. Tessarollo, A.J. Vigers, E. Huang, M.
Kirstein, D.C. de Caprona, V. Coppola, C. Backus, L.F. Reichardt,
B. Fritzsch. Spatial shaping of cochlear innervation by temporally
regulated neurotrophin expression. J. Neuroscience, 21 (16),
6170-6180, 2001.
Leu, M., E. Bellmunt, M. Schwander, I. Fariñas, H.R. Brenner,
U. Muller. Erbb2 regulates neuromuscular synapse formation and
is essential for muscle spindle development. Development, 130,
2291-2301, 2003.
Ferrón, S., H. Mira, S. Franco, M. Cano-Jaimez, E. Bellmunt,
C. Ramírez, I. Fariñas, M.A. Blasco. Telomere shortening and
chromosomal instability abrogates proliferation of adult but not
embryonic neural stem cells. Development (en premsa)
Gónzalez-Martínez, T., D.F. Monjil, G. Germanà, F. De Carlos,
J. Cobo, M.E. del Valle, J.A. Vega. Immunohistochemical localiza-
tion of myelin basic protein in the human peripheral nervous
system, with special reference to the sensory corpuscles. (enviat
per a revisió)
Gónzalez-Martínez, T., S. Catania, B. Díaz-Esnal, M. Barbosa,
D.F. Monjil, G. Germanà, J.A. Vega. Human cutaneous Meissner
and Pacianian corpuscles display myelin basic protein immunore-
activity: fact or artefact? (enviat per a revisió)
Galeano, R.M., A. Germanà, T. González-Martínez, F. De
Carlos, H. Hidaka, M.E. del Valle, I. Silos-Santiago, J.A. Vega.
Characterization of dorsal root gangliaon sensory neurons expres-
sing neurocalcin in mice. Brain Res (en premsa)
De Carlos, F., A. Germanà, R.M. Galeano, D.F. Monjil, M.E. del
Valle, I. Fariñas, J. Cobo, J.A. Vega. Effects of neurotrophin-3
gene mutation in the expression of neurocalcin. (enviat per a
revisió)
RECERCA BIOMÈDICA | 61

MEMÒRIA FINAL
1. Resum del projecte original Objectius principals1) Demostrar que la polimedicació amb metformina,glibenclamida i insulina per al tractament de lahiperglucèmia de la diabetis mellitus tipus 2 (DM2) ésuna alternativa per aconseguir un bon controlglucèmic i per retardar el començament i l’evolucióde les complicacions.2) Demostrar que el tractament combinat ambmetformina i glibenclamida no té influències negativesen les complicacions macrovasculars de la DM2 i enel pronòstic de vida, en comparació amb altrespautes.3) Avaluar la relació cost-efectivitat de la polimedica-ció en comparació amb el tractament amb diversesdosis d’insulina.
Objectius secundaris1) Avaluar les possibles influències d’alguns delsfactors de risc cardiovascular no clàssics (homocisteï-na, fibrinogen, PAI-1, ferritina) en l’evolució de lescomplicacions cardiovasculars de la DM2.2) Avaluar el grau inicial de secreció i de sensibilitat ala insulina, com a factor de predicció de la resposta a
les diverses pautes terapèutiques, en un subgrup depacients.
Hipòtesi1) La polimedicació amb fàrmacs de mecanismed’acció diferent pot ser una alternativa vàlida per altractament de la DM2 amb menys efectes adversos(hipoglucèmies, augment de pes), en comparacióamb el tractament amb diverses dosis d’insulina.2) El tractament combinat amb metformina i gliben-clamida no augmenta la morbi-mortalitat de la DM23) La polimedicació, en comparació amb el tracta-ment amb diverses dosis d’insulina, té una millorrelació cost-efectivitat.
MètodeAssaig clinicoterapèutic en fase IV, prospectiu,multicèntric, aleatoritzat i controlat, atorgat a l’atenciódiabetològica (especialitzada o no), que incloudiabètics tipus 2 amb excés de pes, de diagnòsticrecent, sense complicacions i sense tractamentfarmacològic, en els quals s’estudiarà la secreció isensibilitat a la insulina inicials com a factor depredicció de la resposta terapèutica, com tambéd’altres variables d’efectivitat terapèutica i econòmi-ques.
FUNDACIÓ LA MARATÓ DE TV3
Títol del projecte
Estudi pilot de diabetis i obesitat a Catalunya
Investigadors principalsJaume Arroyo Bros · Begoña Campos Bonilla · Anna Sanmartí Sala
InstitucióDAP Sabadell. Fundació Jordi Gol i Gurina · Facultat de Medicina,
Fundació Clínic per a la Recerca Biomèdica · Hospital Universitari Germans Trias i Pujol
Membres de l’equipOlga Lóriz i Marisa Granada
141.642 €

Resultats esperatsNo increment de la morbi-mortalitat, menys efectesadversos i millor relació cost-efectivitat.
Impacte potencial esperatEls resultats d’aquest estudi permetran adoptaractituds terapèutiques definitives per al tractament dela diabetis mellitus tipus 2.
Modificacions de la metodologia i/o pla de treballoriginalA les anàlisis clíniques centralitzades realitzades allaboratori d’hormones de l’Hospital UniversitariGermans Trias i Pujol, que es demanen al principi i alfinal del estudi, s’ha cregut adient afegir-hi unaextracció transversal més (a l’any de la inclusió) ambla finalitat d’ampliar la informació sobre la evolució delprocés i fer una avaluació transversal de les dadesdisponibles.A més, s’ha considerat necessari estudiar una novacitoquina del teixit adipós (adiponectina) que semblaimplicada en el desenvolupament de la diabetis tipus2 i l’evolució de la resistència a la insulina i el risccardiovascular, com també una altra hormona tambérelacionada amb la diabetis i l’obesitat: la leptina. Desd’aquest punt de vista, té un interès especial avaluarel possible efecte de metformina en l’evolució de lesconcentracions d’adiponectina (ADPN) en elspacients diabètics amb excés de pes del nostreàmbit. També s’ha considerat necessari prescindir delsegon objectiu secundari ("avaluar el grau de secreciói de sensibilitat a la insulina, com a factor de predic-ció de la resposta a les diverses pautes terapèuti-ques") proposat inicialment, tenint en consideracióque s’ha iniciat un estudi multicèntric (estudi DESIRE,promogut per la indústria farmacèutica) que té elmateix objectiu però amb una n de 3.000 pacients.Hem considerat d’interès clínic substituir-lo per unavaloració de la resistència a la insulina (HOMA-IR) i deles característiques antropomètriques, biològiques icomorbilitat associada dels pacients, com a factorsde predicció de la resposta a les distintes pautes detractament (factors de predicció poc estudiats en laliteratura especialitzada).D’altra banda, les dades obtingudes dels diabèticsinclosos al nostre estudi (casos), hem cregut adientcomparar-les amb pacients no diabètics (controls)amb edat i índex de massa corporal (IMC) semblants,i així poder disposar també de valors normalsd’ADPN en la nostra població.
2. ResultatsFins ara, els resultats elaborats en aquest estudi pilotmostren la situació dels pacients a l’inici i la sevaevolució durant el primer any. No ha estat possible fercomparacions entre les branques de l’assaig perquèels estrictes criteris de bon control glucèmic han estatassolits per la majoria dels pacients i només uns pocscasos han presentat mal control metabòlic i hanpassat, segons protocol, a l’etapa d’assignació detractament per aleatorització. Aquest fet tampoc nopermet comparacions econòmiques sòlides entre elsdiversos tractaments. De manera majoritària, elspacients diabètics reclutats mantenen, segons lesrecomanacions actuals, un òptim control metabòlicamb dieta + exercici + metformina.
Mostra inicialLa mostra consta de 119 pacients que han estatreclutats en centres d’atenció primària (CAP) i 3hospitals. En aquest informe presentem les dadesreferents a 104 casos, que corresponen a un total de34 homes i 70 dones. Pel que fa a l’anàlisi inicial,destacarem els punts següents:
Descripció de paràmetres antropomètrics i delaboratoriEn el grup de pacients diabètics, el temps d’evoluciódes del diagnòstic de la malaltia va ser de 13± 6mesos. Un 20% són hipertensos amb tractamentfarmacològic i un 26% necessiten medicació hipolipe-miant.En les taules I i II es mostren els valors descriptius dela mostra inicial comparant homes i dones. Enaquestes taules destaca com a rellevant que en lanostra mostra, comparant per sexes, homes i donestenen un control metabòlic de la diabetis moltsemblant (sense diferències estadísticament significa-tives). Les dones tenen més IMC, concentracionsd’insulina més altes, més insulinoresistència (HOMA-IR), més colesterol i més elevats els marcadorsd’inflamació relacionats amb el risc cardiovascular:PCR i PAI-1. Pel que fa a l’ADPN i la leptina, el nivellde les dones es superior al dels homes.Contràriament, els homes tenen més índex cintura-maluc i més àcid úric.També en la taules I i II s’exposa la comparació entrecasos i controls no diabètics (n=56). De la compara-ció d’aquestes dues poblacions (que no presentendiferències estadísticament significatives d’IMC i edat,i a més cap de les dues tenia antecedents d’esdeve-niments cardiovasculars) es pot destacar que, encomparació amb els controls, els homes diabèticstenen de manera significativa (i com era de esperar)glucèmia en dejú i Hb glucada (Hb A1c) més altes,com també més insulinoresistència. Les donesdiabètiques, a més, tenen els triglicèrids més alts.Però en canvi, tant els homes com les dones ambDM2 tenen l’ADPN més baixa (figura 1).
Figura 1. Valors mitjans d’ln (adiponectina) segonssexe i presència de diabetis
RECERCA BIOMÈDICA | 63

Els quocients ADPN/Insulina i ADPN/HOMA sóncomparables entre homes i dones diabètiques itambé entre ambdós sexes dels no diabètics. Però sicomparem entre diabètics i no diabètics, ambdósquocients estan més elevats en els pacients nodiabètics. No hi ha altres diferències significatives,clíniques o biològiques, entre pacients i controls.
Taula 1.Rang mitjà i interquartil (percentils 25-75) de lesprincipals característiques clíniques de pacients ambdiabetis mellitus i grup control. S’utilizà la prova U deMann-Whitney per valorar les diferències entre homesi dones i entre pacients diabètics i controls.
PAS: pressió arterial sistólica.PAD: pressió arterial diastòlica.* Diferències entre homes i dones diabètics.** Diferències entre homes i dones controls.# Diferències entre homes diabètics i homes controls.$ Diferències entre dones diabètiques i homes controls.
Taula 2.
Rang mitjà i interquartil (percentils 25-75) de les principals
característiques analítiques de pacients amb diabetis mellitus i
grup control. S’utilitzà la prova U de Mann-Whitney per valorar les
diferències entre homes i mujeres i entre pacients diabèticos i
controls.
CRP: proteína reactiva C.* Diferències entre homes i dones diabètics.** Diferències entre homes i dones controls.# Diferències entre homes diabètics i homes controls.$ Diferències entre dones diabéticas i homescontrols.ADPN i resistència a la insulina
No es troben diferències significatives entre sexessegons les diferents ràtios d’ADPN per cada BMI,insulinèmia i HOMA-IR en els diabètics o en el grupcontrol.Si valorem de manera global (diabètics i controls) lesrelacions entre ADPN i els factors de risc cardiovas-cular, podem veure que en els homes, ADPN esrelaciona de manera negativa amb glucèmia en dejú,Hb A1c i àcid úric, però de manera positiva ambl’edat. En el cas de les dones, ADPN es relaciona demanera negativa amb índex cintura-maluc, BMI,glucèmia i insulina en dejú, Hb A1c, HOMA, triglicè-rids, àcid úric i PCR, però de manera positiva ambcolesterol HDL. No es troben relacions significativesen comparació amb edat, pressió arterial, colesteroltotal, leptina i homocisteïna.Les variables que es relacionen de manera significati-va amb ADPN s’utilitzen en un model de regressiómúltiple per determinar els principals factors quepoden influenciar les concentracions d’ADPN encada grup, segons sexes. En els homes, aquestmodel mostra com un 50,2% de la variabilitat en lesconcentracions d’ADPN es pot explicar en relacióamb els factors de predicció següents: presència dediabetis (36,21%), àcid úric (6,33) i edat (7,65%). Enel grup de dones, aquest model explica un 48,7% dela variabilitat d’ADPN, amb la insulina com a principali més potent variable de predicció, que pot explicarmés de la meitat de la variabilitat. La presència dediabetis participa també com a factor de predicció iconfirma la relació ajustada entre concentracionsd’ADPN i diabetis en ambdós sexes.
Descripció del risc cardiovascularRelacionat amb un dels objectius secundaris del’estudi, s’ha analitzat la presència o absènciad’FRCV clàssics i no clàssics en el grup de pacientsdiabètics. Una primera aproximació va consistir acalcular la prevalença de cadascun dels factors en elgrup, però també el nombre de factors presents encada individu. Mitjançant la fórmula clàssicad’Anderson de l’Estudi Framingham, els homes de lanostra mostra tenen un risc coronari total als 10 anys(0.15± 0.76) i les dones (0.12± 0.06), (entre el 15% yel 12%). Si apliquem la fórmula de l’Estudi Regicor(més adaptada a la població mediterrània), el risccoronari és més baix (0.06± 0.03 vs. 0.059± 0.03):6% del risc als 10 anys, però sense diferènciesestadísticament significatives. Així doncs, segons laprimera fórmula veiem que estem davant d’un riscmoderat i amb la segona fórmula (adaptada a lanostra població) parlaríem d’un risc lleu.Vam fer un càlcul del risc hipotètic: amb una interven-ció dràstica sobre tots els factors de risc modificablesprevistos en la fórmula d’Anderson, podríem disminuirel risc en una mitjana del 7.5% (IC95%: 5.1%-10%)per als homes i del 6% (IC95%: 4.8-7.2%) per a lesdones. D’això es desprèn que intensificant tracta-ments potser podríem arribar a disminuir-lo pràctica-ment a la meitat.
Mostra als 12 mesosEl nombre de pacients del qual disposem dades en lavisita de seguiment dels 12 mesos és de 63. Hanestat exclosos 33 pacients pels motius següents: 14per voluntat del pacient o no acudir als controls, 3 per
FUNDACIÓ LA MARATÓ DE TV3

canvi de domicili del pacient, 7 per intolerància a lametformina, 3 per presentar malaltia hepàtica, 3 permicroalbuminúria, 1 per voluntat del metge i 1 perpresentar mesotelioma en estadiatge avançat.La intervenció terapèutica amb metformina (1.700mg/dia = 2 comprimits/dia) ha aconseguit bon controlmetabòlic de la diabetis en 53 dels 63 pacients. Elsaltres 10 pacients han seguit les aleatoritzacionssegons protocol: 9 casos estan en la primera aleatorit-zació (2 pacients amb 3 comprimits de metformina i 7amb 2 comprimits de metformina i 1 de glibenclamida)i en un altre ha calgut fer la segona aleatorització,seguint 3 comprimits de metformina i insulina nocturnaper intentar aconseguir el control metabòlic establert.En aquest sentit, un 91,4% estan correctamentcontrolats (4,4 % de millora). Des de l’inici, hemmillorat el control metabòlic de la diabetis (Hbglucada) una mitjana de 0,47 en els homes i de 0,49en les dones.Respecte del control de la tensió arterial, hem trobatben controlats el 38,5% dels nostres pacients, un12% de millora respecte de l’inici.Pel que fa al control del perfil dels lípids, un 25%assolien l’objectiu establert, però un 13% perden elbon control.Pel que fa a la intervenció realitzada amb el tracta-ment amb 2 comprimits de metformina diaris, caldestacar que tal com veiem en les taules III i IV, tanten els homes com en les dones hauríem disminuïtamb significació estadística l’Hb A1c, la insulinèmia,el pèptid C i la insulinoresistència, com també laleptina i homocisteïna. També millora en el grup deles dones la PCR, malgrat que no va ser així en elshomes. També hi va haver una modesta però signifi-cativa millora pel que fa a l’IMC. En el que no hemtrobat diferències significatives ha estat quant al’ADPN (taules III i IV).
Taula III.
Comparació de les variables inicials i després d’un any d’interven-
ció amb metformina segons la prova t d’Student en el grup de les
dones.
Taula IV.
Comparació de les variables inicials i després d’un any d’interven-
ció amb metformina segons la prova t d’Student en el grup dels
homes.
3. Rellevància i possibles implicacions clíniquesdels resultats finals obtingutsDes que es va plantejar aquest estudi, el panoramaterapèutic de la DM tipus 2 ha canviat. D’una banda,s’han comercialitzat nous fàrmacs amb mecanismesd’acció diferents als disponibles fins fa ben poc i,d’una altra, hi ha un ús més habitual de la teràpiacombinada amb fàrmacs orals (que tenen mecanis-mes d’acció diferents) més insulina, com la proposa-da en la nostra hipòtesi de treball. També pel que faals teixits diana de la resistència a la insulina (fetge,múscul i teixit adipós) el seu protagonisme relatiu estàfent un canvi i, en aquest sentit, l’anomenada lipoto-xicitat dels àcids grassos lliures generada per ladisminució de la sensibilitat a la insulina en el teixitadipós guanya rellevància, tant pel que fa a lapatogènia de la resistència a la insulina com en ladisminució de la secreció d’aquesta per les cèl·lulesbeta del pàncrees. A més, la identificació de noveshormones sintetitzades pels adipòcits, com l’adipo-nectina, amb un paper encara no prou definit, peròles investigacions més recents de la qual semblenindicar que pot tenir efectes antiresistència a la insuli-na (antiinflamatoris, antiaterogènics, sensibilitzadors),situen el teixit adipós com una de les principalsdianes terapèutiques en les estratègies actuals deltractament de la DM tipus 2 amb excés de pes.Fetes aquestes consideracions, si valorem elsresultats obtinguts fins ara, podem destacar queaquest grup de pacients diabètics de diagnòsticrecent presenten ja quan es detecta la seva alteraciómetabòlica, un risc cardiovascular lleu-moderat.Els resultats del estudi UKPDS van demostrar, l’any1998, que la morbilitat cardiovascular en els pacientsdiabètics obesos tractats amb metformina disminuïade manera significativa. La teràpia amb metforminainstaurada des d’un principi, simultàniament amb ladieta i l’exercici físic, proporciona un control glucèmicmolt bo en la majoria dels pacients del nostre estudi(tot i els estrictes criteris de control demanats en elprotocol), amb una tolerància acceptable i sense capesdeveniment vascular durant aquest primer any deseguiment. Des d’aquest punt de vista, en elspacients diabètics tipus 2 amb excés de pes, lateràpia amb metformina ja en el moment del diagnòs-tic sembla una bona opció de tractament, si no hi hacap contraindicació.D’altra banda, hem pogut comprovar que les concen-tracions d’ADPN d’aquesta mostra de pacientsdiabètics són significativament més baixes que lesdels casos controls sense diabetis, però ambcaracterístiques antropomètriques similars. A més,després d’un any de tractament amb metformina, lesconcentracions d’ADPN no augmenten. Aquest és unfet demostrat per primera vegada en pacientsdiabètics del nostre àmbit (només hi ha una solapublicació sobre això en població nord-americana).Segons la literatura científica més recent, aquestanova citoquina del teixit adipós pot tenir un efecteprotector respecte del desenvolupament i l’evolució
RECERCA BIOMÈDICA | 65

de la DM tipus 2, com també una acció antiresistèn-cia a la insulina i protectora del risc cardiovascular. Enconseqüència, aquells fàrmacs que augmentin lesconcentracions d’ADPN poden ser de primeraelecció terapèutica en pacients amb DM tipus 2 iresistència a la insulina. Aquest és el cas de lesnoves glitazones, que actuen com a sensibilitzants ala insulina (de manera preferent en els teixits adipós imuscular), incrementen les concentracions d’ADPN imilloren la utilització de la glucosa i la lipotoxicitat. Sitenim en consideració que la teràpia amb metforminano millora les concentracions d’ADPN en aquestspacients diabètics amb excés de pes i resistència a lainsulina, però que els beneficis de la seva utilitzacióen pacients amb aquestes característiques ésindubtable, des del nostre punt de vista en aquesttipus de diabètics el tractament amb teràpiacombinada metformina + glitazona pot ser la primeraopció terapèutica, ja des del moment del diagnòstic.No obstant això, aquesta afirmació necessita sercorroborada per estudis a llarg termini.
4. PublicacionsPublicació en la revista Clinical Chemistry and Laboratory
Medicine (suplement especial juny de 2003): Association of
adiponectin with insulin resistance and cardiovascular risk factors
in Type 2 diabetes patients.
Referència bilbliogràfica: s252 Clin Chem Lab Med 2003; 41
Special Supplement, pp s1-S608, june 2003. En relació al
Congrés International IFCC-FESCC-Euromedlab’03 al juny del
2003 a Barcelona.
Publicació en la revista Diabetes and Metabolism (edició especial
de l’agost de 2003): Association of adiponectin with insulin
resistance and cardiovascular risk factors in Type 2 diabetes
patients.
Referència bibliografica: Diabetes Metab 2003,29,4S7-4S428. En
relació amb el "18th International Diabetes Federation Congress"
celebrat a París a l’agost de 2003.
Publicació en la revista Española de Obesidad, vol I, Nº 2-
Novembre de 2003: ¿Qué riesgo cardiovascular presentan los
pacientes con exceso de peso y diabetes mellitus tipo 2 incipiente?
En relació amb el VI Congrés Nacional de la Sociedad Española
de Obesidad (SEEDO) celebrat el novembre de 2003.
Publicació en la revista Atención Primaria de la Sociedad
Española de Medicina Familiar y Comunitaria (SEMFYC), vol 32
Suplement I:93-125: La Adiponectina protege del riesgo cardio-
vascular de los pacientes con diabetes mellitus tipo 2?
En relació amb el XXIII Congrés Nacional Semfyc celebrat a
Barcelona el desembre de 2003.
FUNDACIÓ LA MARATÓ DE TV3

RECERCA BIOMÈDICA | 67
Títol del projecte
Homocisteïna i malaltia: estudis genètics, bioquímics i clínics de l'homocistinúria i del risc cardiovascular
Investigador principalSusana Balcells Comas, Mª Antònia Vilaseca Buscà, Xavier Pintó Sala
InstitucióFacultat de Biologia. UB, Hospital Universitari Sant Joan de Déu, Ciutat Sanitària i Universitària de Bellvitge
Membres de l’equipEmili Corbella, Daniel Grinberg, Rafael Artuch, Joan Moreno, Eva López-Quesada, Carla Asteggiano, Mònica Cozar i Roser Urreizti.
71.549 €
MEMÒRIA FINAL
1. Resum del projecte originalObjectius originals del projecte global (inclou elstres subprojectes)La hiperhomocisteïnèmia greu és expressió d’unamalaltia metabòlica, l’homocistinúria, causada princi-palment per mutacions en el gen CBS (que codifical’enzim Cistationina-ß-Sintetasa). La hiperhomocisteï-nèmia moderada és molt més freqüent en la poblaciógeneral i està associada a un risc augmentat demalaltia cardiovascular. La causa d’aquesta hiperho-mocisteïnèmia no està totalment aclarida. Diversosgrups han demostrat que una variant en el gen de la5,10-metilenetetrahidrofolat reductasa (MTHFR) explicanomés un percentatge dels casos d’hiperhomocisteï-nèmia i que hi d’haver altres factors en l’etiologiad’aquest trastorn metabòlic. Nosaltres vam estudiaruna població espanyola d’individus amb infartsprematurs i els seus fills. Els nostres resultats tambéapunten a l’existència d’altres factors i, a més a més,indiquen que almenys un d’aquests factors és genètic.
HipòtesisLes hipòtesis que fonamenten el nostre treball són les
següents: variants del gen CBS, probablementdiferents de les que causen homocistinúria, poden serresponsables d’aquesta hiperhomocisteïnèmiamoderada tan freqüent. Alternativament, i possible-ment en una menor proporció de casos, poden estarimplicades altres variants dels gens MTHFR i MS.
D’altra banda, els fills de pacients cardiovasculars quepresenten elevades concentracions plasmàtiquesd’homocisteïna poden ser individus de risc de patolo-gia coronària, risc que es podria controlar mitjançanttractament amb folat.
Els objectius concrets del projecte són els següents
Estudiar les mutacions responsables del gen CBS enels malalts homocistinúrics espanyols.
Identificar variants relativament comunes del gen CBS,associades amb la hiperhomocisteïnèmia i amb el risccardiovascular.
En funció dels resultats obtinguts amb el gen CBS,s’analitzarien els gens MTHFR i MS.

FUNDACIÓ LA MARATÓ DE TV3
D’altra banda, es pretén:Avaluar la possible disfunció endotelial en individusamb hiperhomocisteïnèmia, el pare dels quals hagipatit una malaltia coronària prematura.
Igualment, s’estudiarà la resposta a diferents tracta-ments amb folat en funció del genotip d’aquests fills,tant per al gen MTHFR com per a les possiblesvariants que es trobin en el gen CBS.
2. ResultatsI- Caracterització de mutacions del gen CBS enpacients homocistinúricsa) Identificació de les mutacions.S’ha genotipat el gen CBS en 27 pacients homocisti-núrics de la Península ibèrica i s’han identificat lesmutacions en tots els casos. A continuació es presen-ta la llista de pacients amb les dues mutacions decadascun:
Pacient 01 (#02): T191M / T191MPacient 02 (#03): R125W / T191MPacient 03 (#05): R336C / 1566delGPacient 04 (#07): S349N / S349NPacient 05 (#08): R379Q / 1566delGPacient 06 (#09): T191M / T191MPacient 07 (#10): T191M / C275YPacient 08 (#11): L338P / L338PPacient 09 (#12): T191M / T191MPacient 10 (#13): T191M / T191MPacient 11 (#15): IVS1+1G>A / S349NPacient 12 (#16): T191M / T191MPacient 13 (#17): T191M / T191MPacient 14 (#18): R336C / R336CPacient 15 (#19): L456P / L456PPacient 16 (#20): T191M / T191MPacient 17 (#21): T191M / T191MPacient 18 (#23): G148R / G148RPacient 19 (#29): T191M / T191MPacient 20 (#31): T191M / T191MPacient 21 (#22): A226T / A226TPacient 22 (#25): P145L / P145LPacient 23 (#26): R336H / R336HPacient 24 (#28): T191M / M173VPacient 25 (#30): T191M / T353MPacient 26 (#33): T191M / T191MPacient 27 (#34): T191M / T191M
S’ha posat de manifest que la mutació T191M és,amb gran diferència, la més prevalent a la nostra àreageogràfica. Aquest resultat és nou, atès que a la restade països on s’han estudiat les mutacions de CBS enpacients homocistinúrics, l’espectre mutacionalobtingut ha estat totalment diferent. Concretament,s’hi havien descrit dues mutacions prevalents, G307Si I278T, que curiosament no hem trobat en cap delspacients ibèrics.
La gràfica següent mostra la freqüència relativa de lesmutacions trobades a la nostra àrea geogràfica. Lamutació T191M (canvi d’una treonina per una metioni-na a la posició 191 de la cadena aminoacídica)representa el 50% dels al·lels mutats.
b) Estudis funcionals de les mutacions: expressió de 6de les mutacions del gen CBS trobades en els pacientshomocistinúrics analitzats (resultats no publicats)Per comprovar que els diversos al·lels mutats sónrealment responsables de la patologia, hem fetestudis funcionals d’aquests i els hem comparat ambl’al·lel no mutat. Hem introduït cadascuna de les 6mutacions dins bacteris, en què s’ha expressat laproteïna mutada. A continuació, s’ha realitzat l’assaigenzimàtic de CBS i s’ha mesurat l’activitat de cadamutant. L’histograma següent il·lustra els resultatsobtinguts.
L’activitat enzimàtica de diverses proteïnes CBS s’hamesurat en presència o absència del seu reguladoral·lostèric AdoMet. pHCS3 és el clon que expressaCBS silvestre. Els clons portadors de mutacionsdiferents s’han obtingut per mutagènesi dirigida. Lessis mutacions estudiades són: T191M, 1566delG,L338P, L456P, S349N i R379Q.
Si bé les dues primeres ja estaven descrites a la litera-tura, no es disposava de cap dada sobre la sevaactivitat enzimàtica. Les quatre restants sónmutacions que hem trobat en els pacients espanyols ique ningú no havia reportat prèviament. Els valorss’expressen com a tant per cent del valor de referèn-cia del clon silvestre sense AdoMet.S’observa que totes les mutacions afecten severa-ment l’activitat enzimàtica, que en cap cas no arriba al15% del valor de referència.
II- Polimorfismes en gens del metabolisme del’homocisteïna i la seva relació amb malaltiescomunes de l’adultEn comptes de buscar noves variants polimòrfiques,es va optar per analitzar-ne deu ja descrites en la

literatura. A més a més, en comptes d’analitzar enprimer lloc polimorfismes de CBS i després elsd’altres gens, es van analitzar deu polimorfismes decinc gens del metabolisme de l’homocisteïna simultà-niament. Finalment, a més a més de valorar l’efected’aquests polimorfismes sobre la malaltia cardiovas-cular primerenca, també es va fer l’estudi d’aquestsrespecte la preeclàmpsia.
a) Polimorfismes en gens del metabolisme de l’homo-cisteïna i la seva relació amb la patologia cardiovascularS’han genotipat 10 polimorfismes en 5 gens relacio-nats amb el metabolisme de l’homocisteïna en 120pacients homes amb malaltia cardiovascular i 121controls. S’ha dut a terme una anàlisi estadística pervalorar si algun d’alguna d’aquestes variants genèti-ques polimòrfiques ajuda a explicar la malaltia cardio-vascular. El resultat obtingut indica que solament dosd’aquestes (ambdues del gen CBS) estarien relaciona-des amb la patologia cardiovascular, però ambefectes poc significatius:844 ins 68: l’al·lel amb la inserció estaria subrepresen-tat en els pacients.c.1080C>T: el genotip TT estaria subrepresentat enels pacients.
b) Polimorfismes en gens del metabolisme de l’homo-cisteïna i la seva relació amb la preeclàmpsiaCap dels 10 polimorfismes analitzats en 5 gensrelacionats amb el metabolisme de l’homocisteïna estroba associat a la preeclàmpsia en la nostra mostrade dones gestants.
III- Homocisteïna i preeclàmpsiaLes pacients amb preeclàmpsia mostren concentra-cions d’homocisteïna plasmàtica significativamentsuperiors que les gestants controls. S’observa unacorrelació negativa entre les concentracions d’homo-cisteïna plasmàtica i les concentracions de folat en elgrup control. Aquesta correlació no existeix en el grupde gestants amb preeclàmpsia.
Durant la preeclàmpsia, a més de l’elevació de lesconcentracions d’homocisteïna plasmàtica, s’observaun augment generalitzat de les concentracions de laresta d’aminoàcids, tot apareixent concentracionssimilars a les de fora de l’embaràs. Quan analitzemvalors relatius, fent desaparèixer el factor hemoconcen-tració propi de la malaltia, aquestes diferències amb lesgestants controls desapareixen. Malgrat això, nopodem oblidar que les concentracions absolutesd’homocisteïna són superiors en les preeclàmpsies, ique la hiperhomocisteïnèmia pot ser responsable d’unadisfunció endotelial mediada per l’estrès oxidatiu queprovoqui un dany important en la microvascularitzacióplacentària i a tot el sistema vascular matern.Hem trobat diferències significatives en la incidènciade preeclàmpsia en relació amb la intolerància alshidrats de carboni, però no s’ha pogut establir unaassociació entre aquesta intolerància i la concentracióplasmàtica d’homocisteïna.
La determinació d’homocisteïna plasmàtica juntamentamb l’estudi Doppler de les artèries uterines en elsegon trimestre de l’embaràs, n’augmenta la sensibili-tat en la predicció de complicacions obstètriques,
però l’especificitat, el valor predictiu positiu, el valorpredictiu negatiu, i la raó de probabilitat no varien.
IV- Estudi de l’efecte de la dieta rica en folat sobrela hiperhomocisteïnèmia i la funció endotelial enpacients cardiovascularsS’ha observat que tant la dieta rica en folat com elsuplement d’àcid fòlic han rebaixat l’homocisteïnaplasmàtica total de manera estadísticament significati-va (en un 8.6% i un 8%, respectivament; p<0.001).Les diferències entre la teràpia dietètica i la desuplement no han estat significatives.
També s’ha observat que ambdues formes de teràpiafeien elevar significativament la concentració plasmàticade folat (p< 0.001). L’elevació era més pronunciada enel cas dels suplements (p del factor interacció = 0.005).
El diàmetre de l’artèria braquial durant la hiperèmia vaaugmentar un 6,1% amb la dieta i un 6,5% amb elsuplement d’àcid fòlic. Ambdós increments sónestadísticament significatius respecte de la manca detractament (p=0.003) i no són estadísticamentdiferents entre si. En canvi, no s’han observat diferèn-cies en el diàmetre de l’artèria braquial basal abans odesprés de la teràpia.
3. Rellevància i possibles implicacions clíniquesdels resultats finals obtingutsI- Caracterització de mutacions del gen CBS enpacients homocistinúricsL’estudi mutacional dels pacients amb deficiència deCBS no havia estat realitzat a Espanya ni a la zona nordde Portugal. La rellevància del resultat epidemiològic ésque l’espectre mutacional obtingut difereix radicalmentdel d’altres zones europees. Cal, doncs, fer estudisgenètics dels pacients del nostre país, encara que ja sen’hagin fet d’altres a Europa o els EUA.
Aquest estudi permet un consell genètic adient i unpossible diagnòstic prenatal, si es dóna el cas. Lacorrelació fenotip-genotip que se n’ha derivat aportadades d’interès per al seguiment dels pacients, ja quejuntament amb l’estudi enzimàtic, informa sobre laresposta a la vitamina B6, coenzim d’aquesta reaccióenzimàtica i base primera del tractament dels pacients.
D’altra banda, aquest estudi ens ha permès fer elprotocol de diagnòstic i seguiment de l’homocistinúria,presentat al V Congrés d’Errors Congènits delMetabolisme, Madrid novembre del 2003, d’interèsper als clínics (pediatres, neuròlegs, oftalmòlegs, etc.que poden diagnosticar nous casos de la malaltia). Enaquest protocol, els Drs. Balcells i Grinberg (Dept.Genètica, UB) han realitzat els apartats de diagnòsticgenètic i la Dra. MA Vilaseca el de diagnòstic bioquí-mic i diferencial.En conjunt, aquest estudi ha permès renovar l’interèsde pediatres i neuròlegs en aquesta patologia rara.Això facilitarà que els nous casos es diagnostiquin i estractin d’una manera més primerenca, la qual cosa hade revertir en un millor pronòstic dels pacients. Si béel defecte metabòlic d’aquests pacients teòricamentes pot corregir, a la pràctica cal restringir severamentl’aportació proteica de la dieta i si aquesta restriccióes planteja quan el pacient ha fet una dieta completa
RECERCA BIOMÈDICA | 69

durant anys, el resultat és l’incompliment. Arribar adiagnòstics molt primerencs pot ajudar molt en aquestaspecte. Però per ara, com que la patologia presentapoques manifestacions en edats primerenques, sovintha passat desapercebuda d’entrada als ulls delsclínics. Esperem que aquest estudi i les publicacionsque en derivin ajudin a millorar aquest aspecte.
II- a) Estudi de polimorfismes de gens relacionatsamb el metabolisme de l’homocisteïna en pacientscardiovascularsMalgrat que els pacients cardiovasculars presentennivells d’homocisteïna total en plasma superiors alnormal, i que la hiperhomocisteïnèmia té un clarcomponent genètic, els polimorfismes dels cinc gensestudiats no sembla que aportin efectes significatius.Això cal dir-ho amb la prudència corresponent, ja quela potència estadística assolida no ha estat gaire gran.Caldrà veure, en el futur, els resultats de les metaanàli-sis que de ben segur es faran.
Hi ha dues hipòtesis no excloents. La primera és queper establir els nivells d’homocisteïna intervé unnombre molt gran de gens (que inclouen els que hemestudiat), cadascun dels quals aporta un petit efecteadditiu. La segona es que hi hagi un gen encara noidentificat que per si sol determini els nivells d’homo-cisteïna en un grau elevat.
Per ara, doncs, cap dels 10 polimorfismes estudiatsno té valor diagnòstic ni pronòstic de la patologiacardiovascular, i no tindria sentit clínic analitzaraquests genotips en nous pacients cardiovasculars.
II- b) Estudi de polimorfismes de gens relacionatsamb el metabolisme de l’homocisteïna en pacientsamb preeclàmpsiaDe manera semblant al que s’esdevé amb elspacients cardiovasculars, els polimorfismes dels cincgens estudiats no semblen aportar efectes significa-tius al risc de patir preeclàmpsia.
Hem de concloure que el factor genètic no pot serconsiderat com el factor determinant de la hiperhomo-cisteïnèmia observada en el grup de pacients ambpreeclàmpsia, almenys quant als polimorfismes quehem estudiat.
III- Homocisteïna i preeclàmpsiaMalgrat l’existència d’hiperhomocisteïnèmia en lesgestants amb preeclàmpsia, no hem pogut establir laseva relació causal amb la malaltia. La hiperhomocisteï-nèmia pot ser responsable d’un dany important en lamicrovascularització placentària i a tot el sistema vascularmatern, tot provocant disfunció endotelial. Tanmateix, lahiperhomocisteïnèmia també pot ser un marcador mésde la descompensació metabòlica de la malaltia.IV- Estudi de l’efecte de la dieta rica en folat sobrela hiperhomocisteïnèmia i la funció endotelial enpacients cardiovascularsLes nostres dades demostren que una dieta rica enfolats és igualment efectiva que l'àcid fòlic sintèticadministrat en forma de càpsules, tant per disminuirles concentracions plasmàtiques d'homocisteïna, comper millorar la funció de l'endoteli arterial. Aquestsresultats tenen transcendència en la mesura que la
hiperhomocisteïnèmia és un factor de risc indepen-dent de malaltia cardiovascular molt freqüent i lesevidències sobre el benefici del seu tractament estanaugmentant els últims anys. El fet que una dieta ricaen vegetals i altres aliments rics en folats tingui unefecte favorable és una dada més per insistir enl'adequació dels hàbits alimentaris de la població.
4. PublicacionsCouce M.L., S. Balcells, J. Dalmau, D.Grinberg, M.Rodés,
M.A. Vilaseca (2003). Pauta de diagnóstico y seguimiento de la
homocistinuria. Anales de Pediatría, supl (en premsa).
López-Quesada E, Also-Rayo E, Vilaseca MA. (2003).
Hiperhomocisteinemia durante el embarazo como factor de riesgo
de preeclampsia. Med Clin (Barc); 121:350-5.
Lopez-Quesada E, Vilaseca MA, Artuch R, Gómez E, Lailla JM
(2003). Homocysteine and other plasma amino acids in
preeclampsia and in pregnancies without complications. Clin
Biochem 36:185-92.
López-Quesada E, Vilaseca MA, Lailla JM (2003). Plasma total
homocysteine in uncomplicated pregnancy and in preeclampsia.
Eur J Obstet Gynecol Reprod Biol, 108:45-9.
López-Quesada E, Vilaseca MA, Vela A, Lailla JM. (2003)
Perinatal outcome prediction by maternal homocysteine and
uterine artery doppler velocimetry. Eur J Obstet Gynecol Reprod
Biol (acceptat, en premsa).
Eva López-Quesada, M. Antònia Vilaseca, Elena Gómez,
Josep M Lailla. (2004) Are plasma total homocysteine and other
amino acids associated with glucose intolerance in uncomplicated
pregnancies and preeclampsia? Eur J Obstet Gynecol Reprod
Biol (acceptat, en premsa)
Urreizti R., S. Balcells, M. Rodés, L. Vilarinho, A. Baldellou,
M.L. Couce, C. Muñoz, J. Campistol, X. Pintó, M. A. Vilaseca, D.
Grinberg (2003). Spectrum of CBS mutations in 16 homocystinu-
ric patients from the Iberian Peninsula: high prevalence of T191M
and absence of I278T or G307S. Human Mutation, 22:103.
Vilaseca MA, Vilarinho L, Zavadakova P, Vela E, Cleto E,
Pineda M, Coimbra E, Suormala T, Fowler B, Kozich V. (2003).
CblE type of homocystinuria: mild clinical phenotype in two
patients homozygous for a novel mutation in the MTRR gene.
J Inherit Metab Dis 26:361-9.
Also-Rallo E, López-Quesada E, Urreizti R, Vilaseca MA, Lailla
JM, Balcells S, Grinberg D. Polimorphisms of genes involved in
homocysteine metabolism in preeclampsia and in uncomplicated
pregnancies. Eur J Obstet Gynecol Reprod Biol (sotmès)
Pintó X, Vilaseca MA, Balcells S, Vila R, Artuch R, Corbella E,
Meco JF, Pujol R, Grinberg D. Treatment of
Hyperhomocisteinemia with a Folate-Rich Diet or Folic Acid from
Supplements. Clinical Nutrition (sotmès)
M.A. Vilaseca, M.L. Cuartero, M. Martinez de Salinas, N.
Lambruschini, X. Pintó, R. Urreizti, S. Balcells, D. Grinberg.
Successful pregnancies in pyridoxine-nonresponsive homocystinuria.
J Inherit Metab Dis (sotmès).
Publicacions addicionals relacionades amb el tema
López-Quesada EL, Vilaseca MA, González S. Homocisteína y
gestación. Med Clin (Barc) 2000; 115: 352-6.
Carlos Mainou Cid, Natalia García Giralt, Mª Antònia Vilaseca
Buscà, Imma Ferrer Codina, José Félix Meco López, Maite Palá,
Xavier Pintó Sala, Daniel Grinberg Vaisman, Susana Balcells Comas.
Hiperhomocisteinemia y polimorfismo 677C -> T de la metilen-
tetrahidrofolato reductasa como factor de riesgo cardiovascular
en la infancia. An Esp Pediatr 2002; 56: 402-8.
F Blanco, R Deulofeu, Mª A Vilaseca, P Chacón, E Dulin.
Determinació de homocisteína en plasma: metabolismo, metodo-
logía, interpretación de resultados y papel en la evaluación del
riesgo vascular. Quim Clin 2002;21,243-250.
FUNDACIÓ LA MARATÓ DE TV3

RECERCA BIOMÈDICA | 71
MEMÒRIA FINAL
1. Resum del projecte originalEls últims anys el desenvolupament de modelsanimals transgènics ha demostrat que l’alteració del’expressió de gens específics pot corregir la resistèn-cia a la insulina (1,2) . El disseny d’aquests modelsexperimentals es fonamentava en la disminució de lasecreció de glucosa hepàtica o l’augment de la utilit-zació de glucosa pel teixit muscular, adipós o hepàtic(3,4). Tenint presents els resultats obtinguts fins almoment, aquest projecte proposava fer teràpia gènicaen models animals de resistència a insulina (modelsde ratolí db/db) o animals diabètics (induïda ambestreptozotocina), utilitzant un abordatge experimentalde transfecció gènica desenvolupat per un delsmembres de l’equip investigador (5-7). Aquest mètodepermet transferir gens amb un vector de poli(L-lisina)lligat a galactosa mitjançant el receptor d’asialoglico-proteïna. El projecte pretén posar a punt aquestametodologia per al tractament de la diabetis per mitjàde dos models experimentals que permetin disminuirla hiperglucèmia: augmentant el consum hepàtic deglucosa mitjançant la sobreexpressió del gen deglucoquinasa (1-4) i bloquejar la síntesi hepàtica denovo de glucosa amb la inhibició de l’expressió de la
PEPCK (1,10,11). A més, s’intentaria millorar latècnica assajant lligands alternatius que dirigissin elDNA al fetge dels animals, com els complexos serpin-enzim, que s’uneixen al receptor (8,9). Altres millorespotencials que es volien provar incloïen l’ús depromotors alternatius, tot intentant augmentar ladurada de l’expressió del transgèn al fetge. La majorinnovació que oferia aquesta tècnica era la condensa-ció de DNA utilitzant poli(L-lisina) unida covalentment aun lligand, per exemple galactosa. El complex DNA-poli(L-lisina)-lligand, injectat al sistema venós d’unanimal, és captat pel fetge de forma eficient sensenecessitat d’altres procediments invasius ni l’ús devectors vírics, que poden provocar resposta immuneen administrar-los repetidament.
Els objectius concrets que es plantejaren a l’inicid’aquest projecte van consistir a posar a punt lametodologia necessària per al tractament de ladiabetis amb dues estratègies experimentals quepermetessin fer disminuir la hiperglicèmia:
1. Augment del consum hepàtic de glucosamitjançant la sobreexpressió del gen de laglucoquinasa (1-4).Es pretenia crear una construcció quimèrica que
Títol del projecte
Teràpia gènica en animals diabètics
Investigadors principalsRamon Bartrons Bach
InstitucióFacultat d’Odontologia. CSUB
Membres de l’equipJosé Carlos Perales, Alícia Garcia, Jordi Bermúdez, Anna Vidal i Maria Molas.
98.936 €

contingués el promotor i els elements enhancer delcitomegalovirus (CMV) i el gen estructural de laglucoquinasa de rata i compactar-ho amb poli(L-lisina)galactosilada. Posteriorment, s’hauria d’introduiraquest complex en fetge de ratolins amb resistència ala insulina o diabetis tipus I mitjançant injecció venosadel complex DNA-lligand. Es determinarien la concen-tració de glucosa i cossos cetònics en plasma, comtambé l’expressió del gen en el fetge. Es farien estudisfuncionals mitjançant l’administració de glucosa, realit-zació de corbes de glucèmia i anàlisi de la insulinaplasmàtica i estudi del canvi metabòlic produït en elteixit hepàtic.
2. Inhibició de la síntesi hepàtica de novo deglucosa mitjançant la inhibició de l’activitat de lafosfoenolpiruvat carboxiquinasa (PEPCK) (10-11)Es pretenia dissenyar una quimera RNA/DNA quecontingués un únic desemparellament en un delscentres del promotor de la PEPCK de rata (AF2).Aquest centre en el promotor de la PEPCK és elresponsable de la regulació de la velocitat de latranscripció del gen, essent regulat per insulina imembres de les famílies de factors de transcripcióHNF/3 i C/EBP. Les quimeres RNA/DNA unides acomplexos amb poli(L-lisina) galactosidasa s’introdui-rien al fetge d’animals de laboratori mitjançant lainjecció venosa del complex. Aquest model s’utilitzaen ratolins db/db i en animals tractats amb estrepto-zotocina per tal d’avaluar les alteracions del seufenotip produïdes per la introducció del gen mutat dela PEPCK. Es determina la concentració de la glucosai cossos cetònics, a més a més de l’expressió delgen. Es realitzen estudis funcionals mitjançant l’admi-nistració de glucosa, determinació de corbes deglucèmia i anàlisi de la insulina plasmàtica. S’estudiael canvi metabòlic al teixit hepàtic.
3. Millorar la tècnica vectorialAssajant lligands alternatius que dirigeixin el DNA alfetge dels animals, com els complexos serpin-enzimque s’uneixen al seu receptor específic. Altres millorespotencials que s’assagen inclouen l’ús de promotorsalternatius, tot intentant augmentar la durada del’expressió del transgèn en el fetge.
2. Resultats1. Sobreexpressió del gen de la glucoquinasa in vivoA. Obtenció de plasmidis d’expressió i anàlisi del’expressió gènica amb diferents promotorsEn primer lloc s’obtingueren dos plasmidis d’expressióde glucoquinasa diferents: PEPCK-rGK, en què el gende la glucoquinasa està sota el control del promotorespecífic de fetge (PEPCK), i PCAGGS/GK, que estàcontrolat mitjançant el promotor de la ?-globina il’enhancer del citomegalovirus (CMV). Es va fer lacaracterització biofísica d’aquests plasmidis d’expres-sió compactats mitjançant poli-L-lisina i polietileniminagalactosilats (5). Es determinà la grandària de lespartícules obtingudes mitjançant espectroscòpia decorrelació fotònica (dispersió de la llum dinàmica). Lespartícules obtingudes amb la construcció PEPCK-rGKeren d’uns 40 nm de diàmetre, i amb la construccióPCAGGS/GK, de 30 nm. A continuació, s’analitzà lacapacitat d’expressió d’aquestes construccionsmitjançant la transfecció en cèl·lules d’hepatoma
humà (Huh-7 i FTO-2b). En el cas de la construccióPEPCK-rGK s’activà el promotor de la PEPCK ambcAMP (11). Els resultats obtinguts mostraren unaugment de l’activitat glucoquinasa del 150% encèl·lules Huh-7 transfectades amb el gen de laglucoquinasa controlat amb el promotor de la PEPCK.En canvi, la construcció PCAGGS/GK va permetreobtenir millors nivells d’expressió en cèl·lules Huh-7 i,a més, el promotor d’aquesta construcció no estàsotmès a silenciació epigenètica (figura 1). Lescèl·lules FTO-2b van resultar refractàries a la transfec-ció estàndard. A partir d’aquests experiments, vamseleccionar la construcció PCAGGS/GK com a einade treball in vivo.
B. Sobreexpressió del gen de la glucoquinasa in vivoen animals no diabèticsEn segon lloc, es van iniciar els experiments detransferència gènica al fetge en ratolins ICR normals.Es va desenvolupar el sistema de transfecció ambl’ajuda d’un vector control que expressa luciferasa(pLucSV). Aquests experiments inicials ens van mostrarque a 24 hores posttransfecció amb un plasmidicircular superenrotllat s'obté expressió del transgèn enfetge (figura 2). Així, subsegüentment, es va analitzar lasobreexpressió de glucoquinasa a curt termini. Vint-i-quatre hores postinjecció es van sacrificar els ratolins ies va analitzar la sobreexpressió de GK en els seusfetges, tpt mesurant-ne l'activitat GK, i quantificant eltransgèn mitjançant RT-PCR quantitativa. En aquestsassaigs no vam trobar nivells significativamentsuperiors d'activitat GK en els animals transfectatsamb la construcció pCAGGS/GK, atès que els animalsno eren diabètics (figura 3). No obstant això, en aplicarla tècnica d’RT-PCR, sí que vam detectar el transgènen els fetges (figura 4A), i en aplicar la tècnica de formaquantitativa vam constatar nivells significatius deltransgèn en els fetges analitzats (figura 4B).
C. Sobreexpressió del gen de la glucoquinasa in vivoen animals diabèticsEn tercer lloc, també vam estudiar l’activitat GK enfetges de ratolins ICR diabètics, obtinguts mitjançantinjecció amb STZ i transfectats amb una dosi única depCAGGS/GK a dia zero. L'anàlisi fenotípica delsanimals transfectats es va limitar inicialment a laglucèmia, mitjançant punció de vena de la cua aprimera hora del matí (glucèmia en alimentació), perpoder avaluar el curs temporal d'expressió en elnostre model experimental. Com es pot observar en lafigura 5, els animals transfectats amb 30 ?g depCAGGS/GK van respondre inicialment al tractamentamb una disminució significativa de la glucèmia,malgrat que en cap cas es van obtenir concentracionsequiparables als animals no diabètics. Es van utilitzarcom a controls, animals injectats amb un plasmidiirrellevant (pCMV-luciferasa). La durada de l'efecteterapèutic observat es limitava a les 3 primeressetmanes. Aquest efecte demostrà que la sobreex-pressió mitjançant vectors no vírics de glucoquinasaen fetge pot reduir significativament la hiperglicèmiadiabètica insulinodependent, però no reverteix elfenotip diabètic.
Com a estratègia complementària vam desenvoluparun vector de sobreexpressió d’una isoforma de
FUNDACIÓ LA MARATÓ DE TV3

glucoquinasa mutada en l’aminoàcid 456 (valina aalanina) de manera que la seva Km per a la glucosadisminueix (14).
D. Obtenció i anàlisi de la glucoquinasa mutadaA456VInicialment vam generar el cDNA de glucoquinasaA456V a partir del cDNA de glucoquinasa silvestre derata clonat en el vector d’expressió pCAGGS.Mitjançant la tècnica de PCR amb el Quikchange Site-Directed Mutagenesis Kit es va aconseguir el canvinucleotídic d’una timina per una citosina a la posició1370 del cDNA de glucoquinasa. Es va comprovarque aquest canvi nucleotídic havia tingut lloc amb laseqüenciació dels productes de la PCR.
A continuació, per comprovar que la construcciópCAGGS/GKA456V es transcriu correctament i s’obtéuna proteïna funcional, vam transfectar la construccióen cèl·lules Huh-7 mitjançant PEI (polietilenimina)lineal. Com a controls vam transfectar una construccióde pCAGGS amb glucoquinasa wild type i unaconstrucció de GFP (green fluorescent protein).Passades 48 hores de la transfecció vam determinarl’activitat glucoquinasa dels extractes cel·lulars ambun assaig enzimàtic amb diferents concentracions deglucosa. Tal com ens mostra la figura 6, l’activitat del’enzim mutat és més elevada que la del silvestre entotes les concentracions de glucosa, ja que, com aconseqüència del canvi aminoacídic, la glucoquinasamutada té una Km més baixa.
A partir d’aquí vam comprovar les nostres construc-cions in vivo amb la finalitat d’avaluar-ne les repercus-sions metabòliques. Mitjançant el mètode de transfe-rència gènica hidrodinàmic, es va transfectar laglucoquinasa A456V en ratolins ICR no diabètics a fide detectar si la glucoquinasa A456V causa hipoglucè-mia, com a resultat de la seva baixa Km, en compara-ció amb animals transfectats amb salí. Se n’analitzà laglucèmia i la insulinèmia, tant en dejuni com en alimen-tació. La figura 7A mostra com els ratolins transfectatsamb GKA456V presentaven glucèmies significativa-ment més baixes que els controls, tant en dejuni comen alimentació. En conseqüència, la insulinèmia tambéera menor (figura 7B).
Vista l'efectivitat en un animal no diabètic, vam injectarles construccions PCAGGS/GK i PCAGGS/GKA456Ven animals amb diabetis induïda mitjançant unainjecció amb estreptozotocina (STZ). Aquestesconstruccions van ésser linealitzades abans de lainjecció amb la finalitat d'obtenir una expressió a llargtermini, tal com està descrit a la bibliografia (13). Ambla injecció de 30 m g de cada construcció linealitzada,no es van observar efectes hipoglucemiants enanimals alimentats. Després d'un dejuni de 8 hores, laPCAGGS/GK no feia minvar la glucèmia, mentre que laPCAGGS/GKA456V aconseguia disminuir la glucèmiadurant els 2-3 dies postinjecció. Si s'incrementava ladosi a 60 m g de construcció PCAGGS/GKA456Vlinealitzada, l'efecte hipoglucemiant es prolongava finsals 10-11 dies postinjecció (figura 8).
Amb les dades de la caracterització metodològica imetabòlica obtingudes amb els anteriors experiments
hem redactat un manuscrit Hydrodynamic transfer ofthe glucokinase A456V gene in diabetic mice results inimproved glicemia control as compared to wild typeglucokinase, que serà sotmès a publicació.
2. Inhibició de l’activitat fosfoenolpiruvat carboxi-quinasa (PEPCK)Per a aconseguir els objectius plantejats es va dissen-yar una estratègia, utilitzant siRNA produïts ensistemes d'expressió endogen (pSHAG —obtingut dellaboratori del Dr. Hannon, CSHL-), per a la silenciacióhepàtica de la PEPCK (15). Utilitzant aquesta novaestratègia, els objectius proposats s'han pogut dur aterme i es resumeixen en els apartats següents.
L'estratègia d’inhibició específica de la PEPCK s'hacompletat amb el disseny d'experiments antisentit.Les siRNA (small-interference RNA) són molèculespetites de doble cadena d’RNA que indueixen ladegradació de l’RNA diana de manera específica. Pera aquests experiments hem dissenyat amb èxitplasmidis d'expressió d’RNA polimerasa III que utilit-zen el promotor de l'U6 humana per expressar unacadena d’RNA de grandària determinada dissenyadesper al seu autoplegament en una doble cadenamitjançant un hairpin o colze de plegament. Així, elplasmidi es compon de tres parts, dues fixes, elpromotor i el finalitzador que assegura la transcripciódel gen clonat, i una específica de silenciació d’RNAque es clona entre el promotor i el finalitzador ambseqüències específiques per a cada gen que es vulguisilenciar. Per a la silenciació de la PEPCK hem escollitdues seqüències de l’mRNA de la PEPCK per a lesquals s'han dissenyat dues seqüències de siRNA dedoble cadena que s'han clonat per a la seva expres-sió darrere del promotor de l'U6 humà en el plasmidid'expressió pSHAG.
Silenciació del gen de la PEPCK mitjançant siRNA invitroEls dos plasmidis d'expressió de siRNA s'hancotransfectat amb un plasmidi d'expressió de laPEPCK de rata en cèl·lules d’hepatoma Huh-7. S'hanutilitzat dosis creixents dels plasmidis de silenciaciócotransfectats amb una dosi fixa de plasmidi pC/EI-PCK i els nivells de PEPCK obtinguts en les cèl·lulesdesprés de 48 hores de transfecció s'han quantificatmitjançant Western blot. Com a control s'han utilitzatdues estratègies: la primera ha estat la cotransfecciódels plasmidis de silenciació amb el plasmidid'expressió per la GFP o bé per la luciferasa dePhotinus pyralis, i la segona, la cotransfecció d'unplasmidi de silenciació específic de la luciferasa dePhotinus pyralis. Cada transfecció ha estat normalitza-da per càrrega molar de DNA, utilitzant plasmidis buitspBluescript per igualar la quantitat de DNA transfectaten cada cas. L'anàlisi dels controls ha demostrat quela cotransfecció dels plasmidis de silenciació en capcas va produir una reducció dels nivells de PEPCKobtinguts (dades no mostrades). En canvi, la cotrans-fecció del plasmidi de silenciació PEPCKi664 vademostrar un reducció progressiva dels nivells dePEPCK obtinguts per Western blot, essent màxima lasilenciació amb relacions de cotransfecció d’1:6 delplasmidi d'expressió pCEI/PEPCK i del plasmidi desilenciació PEPCKi664 (figura 9). Un altre dels plasmi-
RECERCA BIOMÈDICA | 73

dis dissenyats per a la silenciació específica de laPEPCK, el PEPCKi482, no va ser capaç en cap casde demostrar silenciació específica de la PEPCKsobreexpressada. Els resultats mostrats a la figura 9són quantitatius i representen la mitjana obtinguda del’activitat PEPCK i densitometria de Western blot,obtinguts en dos experiments independents. ElWestern blot mostrat a la figura 9 és representatiudels experiments efectuats.
Silenciació mitjançant siRNA del gen de la PEPCK invivo.Per a la utilització de vectors d’expressió de siRNAper a la silenciació de gens endògens, aquestasilenciació ha de ser demostrada en sistemescel·lulars amb el background genètic adequat. ElsiRNA PEPCKi664 està teòricament dirigit al gen deratolí i rata, que són idèntics per a la seqüència diana,mentre que el gen humà no podria ser silenciat, atèsque conté una seqüència no idèntica. Per comprovarl'activitat silenciadora de la molècula PEPCKi664, hemutilitzat un model de ratolí no diabètic i un altre ambdiabetis, induïda mitjançant injecció amb STZ (en elqual la PEPCK endògena està altament sobreexpres-sada), per estudiar el fenotip que provocaria lasilenciació de la PEPCK en un context de diabetis.
S’ha utilitzat un mètode de transferència gènica novírica basat en la injecció de grans volums de lasolució de DNA (10% v/p) en la vena caudal (12,13).Aquest mètode de transferència gènica ha estatrecentment descrit per a la transfecció de gens enfetge amb una eficiència de fins al 40%. Les nostrestransfeccions utilitzant aquest mètode en els ratolinses van mostrar dosidepenents, tot obtenint unamàxima eficiència quan injectàvem 100 m g deplasmidi (dades no mostrades).
Per testar la silenciació in vivo es va cotransfectar unvector d’expressió de luciferasa (pGL3) amb el vectorcontrol de silenciació de la luciferasa de Photinuspyralis (pSHAG Ff). La cotransfecció a una ràtio 1:6(pGL3:pSHAG Ff) va produir una silenciació del 98%de l’activitat luciferasa en fetge, tot confirmant lafuncionalitat dels vectors de silenciació en fetge pelmètode de transferència hidrodinàmica.
En aquests experiments s'han injectat ratolinsdiabètics amb 100 mg de plasmidi d'expressióPEPCKi664 en la vena de la cua (n=12). Com acontrols, s'ha injectat 100 m g del plasmidi d'expres-sió pSHAG que expressa una seqüència controlcontra el gen de la luciferasa (n=12). Després de 72 h,els animals es van posar en dejuni durant 8 h iposteriorment es van obtenir mostres de sèrum, fetgei ronyó. Els resultats obtinguts es resumeixen en lafigura 10. En primer lloc, s'ha pogut observar unadisminució significativa (al voltant del 40%) de laglucèmia en els animals injectats amb la molèculaPEPCKi664 (figura 10A). Quan hem analitzat els nivellsde PEPCK a fetge i a ronyó, hem observat que alfetge hi ha una reducció d’un 20% (p< 0.05) en l’acti-vitat enzimàtica PEPCK, dada que es confirma perWestern blot (figura 10B). En canvi, en el ronyó no esveu una reducció significativa de PEPCK (dades nomostrades).
Metabòlicament no hem observat cap alteració delsnivells de metabòlits del metabolisme glucídic nilipídic. Els dipòsits de glicogen hepàtic en els animalstractats es mantenen inalterats: no hi ha variació enels nivells d’àcids grassos ni triglicèrids plasmàtics. Elsresultats obtinguts es mostren en la taula 1.
Aquests experiments es completaran amb un estudi allarg termini, mitjançant introducció del plasmidi desilenciació en fetge, utilitzant vectors vehiculats perreceptor. Així mateix, per obtenir un major percentatgeen el silenciació de la PEPCK i estudiar l’impactemetabòlic que se’n derivaria, es preveu aplicar unatècnica de transferència dels vectors de silenciaciómés efectiva, com podria ser l’ús de vectors virals, enconcret adenovirus, amb els quals es pot assolir unatransfecció gairebé total del fetge.
Una part dels resultats obtinguts en els estudisvectorials realitzats per al desenvolupament d’aquestapartat del projecte han estat publicats en CurrentGene Therapy (Molas, M., Gómez-Valadés, A.G., Vidal-Alabró, A., Miguel-Turu, M., Bermúdez, J., Bartrons,R., Perales J.C. "Receptor-Mediated Gene TransferVectors: Progress Towards Genetic Pharmaceuticals".Current Gene Therapy. 2003. 3 (5): 468-485.)
3. Caracterització de vectors de transferènciagènicaA. Disseny i caracterització de complexos de DNAmonocatenari per a la transferència gènica
La grandària final d'un vector de transferència gènicacompactat amb polications en complexos unimolecu-lars és directament proporcional a la massa d'àcidsnucleics que conté. Per aquesta raó hem aconseguitdissenyar i produir vectors de DNA de cadena simple,significativament més petits que els seus homòlegs dedoble cadena. Així mateix, hem identificat diferènciesquant al comportament biofísic durant la compactaciódel DNA en vectors de cadena simple comparat ambels de doble cadena. A més, s'han obtingut aquestespartícules compactades amb major abundància ihomogeneïtat quan el DNA compactat és monocate-nari. En absència de sals, la manera d'unió delspolications al DNA monocatenari difereix claramentrespecte de la unió al DNA bicatenari. En primer lloc,els complexos de DNA monocatenari compactat noes dissocien usant un gradient de NaCl de 0 a 1,5 M,mentre que els complexos de DNA de doble cadenaes dissocien fàcilment a altes concentracions de NaCl.En segon lloc, els complexos de DNA de cadenasimple s'agreguen significativament més lentamentquan són diluïts en solucions que contenen concen-tracions fisiològiques de sals i proteïnes en compara-ció amb els complexos de DNA de doble cadena.Finalment, vam demostrar que la transferència, eltransport nuclear i l’expressió del DNA monocatenaricompactat té lloc en cèl·lules en divisió, com tambéen cèl·lules postmitòtiques. La caracterització biofísicade vectors de DNA monocatenari compactat hapermès distingir certes característiques d'aquestsvectors que podrien millorar la disponibilitat del DNAen el nucli (vectors més petits i menys agregatius).Així, l'estudi funcional minuciós amb vectors de DNAmonocatenari, dirigits al receptor d’asialoglicoproteïna
FUNDACIÓ LA MARATÓ DE TV3

del fetge, permetrà investigar la viabilitat d'aquestsvectors per a teràpia gènica in vivo en el futur. Ambels resultats anteriors s'ha pogut redactar un articlecientífic publicat a Biochim. Biophys. Acta (Molas et al."Single-stranded DNA condensed with poly-L-lysineresults in nanometric particles that are significantlysmaller, more stable in physiological ionic strengthfluids and afford higher efficiency of gene delivery thantheir double-stranded counterparts". BBA. 2002. 1572(1): 37-44.)
B. Caracterització de copolímers de poli-lisina, triptò-fan i serina com a vectors de DNA compactat noagregatiusEl treball iniciat per l'equip s'ha basat en el disseny devectors estables en fluids fisiològics. Poli-L-lisina, poli-L-lisina-L-triptòfan i poli-L-lisina-L-serina són polímerscapaços de formar partícules compactades amb DNAplasmídic de grandària mínima en absència de sals.No obstant això, quan els complexos formats sóndiluïts en mitjans fisiològics, només els copolímers delisina i triptòfan i serina són capaços de romandre enforma de suspensió discreta sense agregació, en clarcontrast amb l'agregació observada en els complexosformats amb polímers de lisina i DNA. Encara així, elscopolímers de polilisina i lisina-serina no són capaçosde mediar transferència gènica per mitjà de receptorsespecífics. Solament les partícules obtingudes amb elcopolímer de lisina i triptòfan són capaces de transferirDNA mitjançant endocitosi mediada per receptor encèl·lules transfectades. Possiblement, la introduccióde dominis hidrofòbics (triptòfan) bloqueja l'agregaciócausada per l’atracció per forces de Van der Waals,característica de l'agregació de dispersions poliiòni-ques. La importància del treball descrit s'inscriu en eldesenvolupament de vectors amb característiquesfarmacològiques idònies per al seu ús en medicinahumana com a fàrmacs de DNA. S’ha presentat untreball (en procés de revisió) per ser publicat en larevista Journal of Gene Medicine (Gómez-Valadés, A.G., Molas, M., Vidal-Alabrò, A., Miguel-Turu, M.,Bermúdez, J., Bartrons, R., & Perales, J. C.Copolymers of poly-L-Lysine with serine and trypto-phan form stable DNA vectors: implications forreceptor-mediated gene transfer).
3. Rellevància i possibles implicacions clíniquesdels resultats finals obtingutsEl projecte ha permès posar a punt la metodologianecessària per poder realitzar teràpia gènica endiversos models animals de diabetis. Un dels greusproblemes dels diversos tipus de diabetis és la dificul-tat de controlar la hiperglucèmia, que comporta lamajoria de complicacions. La possibilitat de poderutilitzar noves metodologies terapèutiques, amb latransfecció de gens en el teixit hepàtic que facilitin elcontrol de la glucèmia, representa un pas moltimportant en el tractament d'aquesta malaltia. L'ús devectors no vírics, de baixa immunogenicitat i toxicitatnul·la, transportant gens reguladors del metabolismede carbohidrats i capaços de ser captats específica-ment pel teixit hepàtic, ha de permetre assolir larecuperació de la glucèmia en els diversos models dediabetis. Els resultats obtinguts en la realitzaciód'aquest projecte serveixen per posar a punt novesmetodologies que poden ser de gran utilitat per al
tractament i el control d'una de les malalties mésfreqüents al nostre entorn.
L’aplicació de la tecnologia de l’RNA d’interferènciaper a la restauració de la glucèmia en animalsdiabètics ofereix també noves possibilitats terapèuti-ques. La inhibició de l'activitat PEPCK ha estatpossible utilitzant siRNA. Aquest mètode ofereix unsresultats molt interessants que poden obrir noveslínies, referents al paper de la gluconeogènesi en lainstauració i el manteniment de la resistència a lainsulina en el fetge.
Figura 1. Activitat glucoquinasa. Transfecció amb vectors
d’expressió optimitzats.
Activitat glucoquinasa de cèl·lules d’hepatoma humàHuh-7 que han estat transfectades amb diversosvectors d’expressió. L’activitat GK està expressadacom el percentatge de sobreexpressió respecte de lescèl·lules no transfectades. PEPCK es refereix acèl·lules que han estat transfectades amb un vectord’expressió que conté el promotor del gen de laPEPCK. PEPCK cAMP són cèl·lules transfectadesamb el vector PEPCK-rGK estimulat amb cAMP.CAGGS correspon a cèl·lules transfectades amb laconstrucció PCAGGS/GK. CAGGS PEI són cèl·lulestransfectades mitjançant PEI amb la construccióPCAGGS/GK.
Figura 2. Activitat luciferasa in vivo. Transfecció de fetge de
ratolí ICR
Activitat luciferasa d'extractes proteics de fetge deratolins ICR transfectats amb un vector d'expressió deluciferasa de Photinus. Control; animals injectatssimplement amb solució salina 24 h abans de l'anàlisi(n=3). Transfectats; animals transfectats amb 30 µg
RECERCA BIOMÈDICA | 75

d'un vector d'expressió de luciferasa de Photinuspyralis (n=3). Els resultats s'han normalitzat per proteï-na total en l'assaig i s'han representat amb referènciaa l'activitat d'una recta patró de quantitats conegudesde luciferasa recombinant a l'assaig.
Figura 3. Activitat glucoquinasa en fetges de ratolí ICR transfec-
tats in vivo.
Activitat glucoquinasa en extractes proteics de fetgede ratolins ICR transfectats amb un vector d'expressióde glucoquinasa de rata (pCAGGS/GK). Lux/ (n=3);animals controls injectats amb 30 µg d'un vectord'expressió de luciferasa de Photinus pyralis 24 habans de l'anàlisi. GK (n=3); animals transfectats amb30 µg d'un vector d'expressió de glucoquinasa(pCAGGS/GK). Els resultats s'han normalitzat perproteïna total en l'assaig. Els resultats s'expressencom mitja +/- desviació estàndard de l'activitatglucoquinasa.
Figura 4. RT-PCR quantitativa específica del transgèn de
glucoquinasa de rata en RNA total de fetge de ratolí ICR transfec-
tat amb pCAGGS/GK.
A) RT-PCR amb primers específics del transgèntransfectat analitzat a partir d’RNA total de fetged’animals controls transfectats amb un plasmidid’expressió de luciferasa de Photinus (lux) o el vectorbuit pCAGGS (pCAGGS) i el control positiu de reaccióamb el plasmidi d’expressió in vitro (GK). El canalpCAGGS/GK correspon a un animal transfectat enfetge amb el plasmidi d’expressió pCAGGS/GK.B) RT-PCR quantitativa que mostra el nivell d’expressió
(mRNA) de glucoquinasa expressada pel transgèntransfectat en tres ratolins ICR amb un vector d’expres-sió de glucoquinasa de rata (pCAGGS/GK) comparatamb el nivell basal d’animals controls injectats amb 30µg d’un vector d’expressió de luciferasa de Photinus 24h abans de l’anàlisi. Els resultats s’han normalitzatmitjançant RT-PCR quantitativa d’actina de ratolí i s’hanajustat a una recta patró per GK i actina. Es mostrenels resultats per a cada animal.
Figura 5. Efecte sobre la glucèmia de la sobreexpressió de
glucoquinasa en ratolins ICR diabètics alimentats (induïda mitjan-
çant injecció d’STZ).
Ratolins ICR van ser transfectats amb un vector novíric d'expressió de glucoquinasa de rata(pCAGGS/GK) i un vector que sobreexpressa la lucife-rasa de Photinus pyralis. La glucèmia es va obtenir enels animals 2 h postrealimentació després d'un dejuniperllongat. La grandària mostral per al grup control(pLUX/) és de 12 animals, mentre que per al gruppGK és de 8 animals. Es presenten els resultats com amitjana +/- error estàndard (SEM). La diferència deglucèmia és significativa estadísticament (P £ 0,001)per al dia 14 i (P £ 0,007) per al dia 21 posttransfecció.
Figura 6. Comparació d’activitats de la glucoquinasa silvestre i la
glucoquinasa A456V amb diferents concentracions de glucosa.
S’ha realitzat l’assaig enzimàtic amb els extractesproteics de cèl·lules transfectades amb PCAGGS/GK iPCAGGS/GKA456V, amb diferents concentracions deglucosa. S’ha corregit l’activitat en funció de la quanti-tat de proteïna.
FUNDACIÓ LA MARATÓ DE TV3

Figura 7. Comparació de glucèmies i insulinèmies en dejuni i
alimentació.
A) Representació de la glucèmia en ratolins ICRnormals a les 24 hores postinjecció. El grup control haestat injectat amb solució salina i el grup GKA456Vamb la construcció PCAGGS/GKA456V. S’han realit-zat els mesuraments en situació d’alimentació idesprés d’un dejuni de 8 hores.B) Representació de la insulinèmia en els 2 grupsesmentats, en situació d’alimentació i després d’undejuni de 8 hores.
Figura 8. Efecte sobre la glucèmia de la sobreexpressió de
glucoquinasa silvestre i la mutada en ratolins ICR diabètics (per
injecció d’STZ) amb dejuni.
Ratolins ICR van ser transfectats amb pCAGGS/GK iPCAGGS/GkA456V. La glucèmia es va mesurar en elsanimals després de 8 h de dejuni. Es presenten elsresultats com a mitjana +/- error estàndard (SEM). Ladiferència de glucèmia entre el grup de gk 30 µg (Å£) igkA456V 30 µg (ܶ) és significativa estadísticament (*P £ 0,05; ** P £ 0.01) per al dia 2 i 3 posttransfecció.La diferència de glucèmia entre gk A456V 30 µg igkA456V 60 µg (Å°) és significativa estadísticamentfins al dia 11 posttransfecció.
Figura 9. Silenciació específica de la PEPCK mitjançant siRNA
in vitro.
La cotransfecció del plasmidi de silenciacióPEPCKi664 i del plasmidi d’expressió de la PEPCK derata demostra una reducció progressiva dels nivells dePEPCK obtinguts per Western blot a mesura ques’incrementa la dosi del plasmidi de silenciació, essentmàxima la silenciació amb ràtios de cotransfecció 1:1del plasmidi d’expressió pCEI/PEPCK i del plasmidi desilenciació PEPCKi664. Els resultats quantitatiusrepresenten l’activitat específica PEPcK (13) en extrac-tes de cèl·lules transfectades. El Western blot ésrepresentatiu de diferents experiments. Es potobservar com la banda específica, corresponent aPEPCK sobreexpresada, disminueix en intensitat amesura que s’incrementa la concentració de plasmidide silenciació en la transfecció.
Figura 10. Efectes metabòlics de la inhibició mitjançant siRNA de
l'expressió de la PEPCK endògena en animals diabètics.
Model diabètic de ratolí injectat amb STZ en el qual laPEPCK endògena està altament sobreexpressada. Enaquests experiments s'han injectat ratolins diabèticsamb 30 µg de plasmidi d'expressió PEPCKi664 en lavena de la cua (n=4) (pSHAG-664). Com a controls'ha injectat 30 µg del plasmidi d'expressió pSHAGque expressa una seqüència control contra el gen dela luciferasa (n=4) (pSHAG-Ff). Després de 72 h, elsanimals es van posar en dejuni durant 8 h i posterior-ment es van obtenir mostres de sèrum per a la sevaanàlisi. A) Glucèmia en sang (mg/dl). B) Triglicèrids ensèrum (mg/dl). C) ß-HBA en sèrum (µg/dl).
RECERCA BIOMÈDICA | 77

Taula 1. Concentració de metabòlits dels animals transfectats.
4. PublicacionsMolas, M.; Bartrons, R.; Perales, J. C. (2002).
Single-stranded DNA condensed with poly-L-lysine results in
nanometric particles that are significantly smaller, more stable in
physiological ionic strength fluids and afford higher efficiency of
gene delivery than their double-stranded counterparts. Biochim.
Biophys. Acta, 1572 (1): 37-44.
Molas, M., Gómez-Valadés, A.G., Vidal-Alabró, A., Miguel-Turu,
M., Bermudez, J., Bartrons, R., and Perales, J.C. (2003).
Receptor-mediated gene transfer vectors: Progress Towards
Genetic Pharmaceuticals. Current Gene Therapy, 3: 468-485.
Gómez-Valadés, A. G., Molas, M., Vidal-Alabrò, A., Miguel-
Turu, M., Bermúdez, J., Bartrons, R., & Perales, J. C. Copolymers
of poly-L-Lysine with serine and tryptophan form stable DNA
vectors: implications for receptor-mediated gene transfer. Journal
of Gene Medicine (manuscrit en fase de revisió).
Vidal-Alabró, A.; Gómez-Valadés, A.; Molas, M.; Duran, J.;
Bermúdez, J.; Bartrons, R.; Perales, J. C.
Hydrodynamic transfer of the glucokinase A456V gene in diabetic
mice results in improved glicemia control as compared to wild
type glucokinase. (manuscrit en preparació).
Gómez-Valadés, A.; Molas, M.; Vidal-Alabró, A.; Bermúdez, J.;
Bartrons, R.; Perales, J. C.
Overcoming fasting hyperglicemia in diabetes by means of shRNA
inhibition of the cytosolic phosphoenolpyruvate carboxikinase.
(manuscrit en preparació).
FUNDACIÓ LA MARATÓ DE TV3

RECERCA BIOMÈDICA | 79
MEMÒRIA FINAL
1. Resum del projecte originalObjectius generals i específicsEls objectius s'han desglossat en els dos gransapartats de què consta el projecte. D'una banda, elsaspectes relacionats amb la secreció d'insulina. D'unaaltra, els relacionats amb la translocació del GLUT4.
Secreció d’insulinaCaracteritzar la maquinària molecular secretora d’insu-lina en cèl·lules ß-pancreàtiques i BRIN-BD11, tant deles proteïnes SNARE com, encara més important, deles possibles proteïnes reguladores.
Utilitzar les cadenes lleugeres de neurotoxines clostri-dials per obtenir models cel·lulars en què la secrecióestà inhibida i permetre, d’una banda, estudiarseparadament la secreció purament dependent decalci i, d’una altra banda, buscar mecanismes ialternatives d’estímul i secreció d’insulina.
Estudiar el patró de fosforilació de proteïnes encèl·lules secretores d’insulina després del seu estímulamb glucosa, KCl i ionòfor de calci i el cicle associat ala secreció d’insulina d’algunes proteïnes (sobretot
munc-18 i rab3a). Aquest estudi es farà extensiu amodels animals experimentals de diabetis.
Estudiar l’expressió de les proteïnes implicades en lasecreció d’insulina en la cèl·lula ß-pancreàtica idiverses línies cel·lulars d’insulinomes, secretoresd’insulina, utilitzant substàncies estimulants o inhibido-res de la secreció i amb situacions experimentalsdiferents (hipòxia, presència de citoquines,..) Intentaridentificar els factors de transcripció implicats en elscanvis d’expressió proteica.
Translocació sensible a insulina del transportador deglucosa, GLUT4Caracteritzar la maquinària molecular responsable dela translocació del GLUT4 en fibres muscularsesquelètiques, adipòcits i, especialment, en fibresmusculars cardíaques. S’identificaran i caracteritzaranfuncionalment tant les proteïnes SNARE (tenint encompte les isoformes diferents de cadascuna), comles possibles proteïnes reguladores, algunes de lesquals s’han esmentat en l’apartat anterior de secreciód’insulina. Aquest apartat es complementaria ambestudis d’interacció entre les diverses isoformes ques’anessin identificant, per estudiar la seva afinitat en launió i formació de complexos.
Títol del projecte
Estudi de la maquinària molecular secretora d'insulina i de la translocació del transportador de glucosa glut4. Canvis funcionals i d'expressió relacionats amb la diabetis
Investigadors principalsJoan M.V. Blasi Cabús
InstitucióFacultat de Medicina. CSUB
Membres de l’equipMireia Martín, Adriana Raptis, Benjamín Torrejón, Inmaculada Gómez de Aranda, M. Eugenia Grandes, LaiaBahima, Laura Teixidó, Marc Elias, Àlex Soler, Artur Escalada, Carles Solsona, Jordi Marsal i Neus Latorrre.
72.392 €

Utilitzar les cadenes lleugeres de neurotoxines clostri-dials per obtenir models cel·lulars en què la transloca-ció estigui bloquejada parcialment o totalment i poderdissecar, pas a pas, el mecanisme de translocaciódependent d’insulina (en tots els casos) i tambéd’hipòxia en els cardiomiòcits.
Estudiar els cicles de fosforilació i defosforilació de lesproteïnes implicades en la translocació del GLUT4. Demanera especial s’estudiarien aquelles proteïnes ques’han proposat com a reguladores (entre les quals,munc-18).
Estudiar l’expressió de proteïnes responsables de latranslocació del transportador GLUT4 en condicionsdiferents d’estimulació per insulina i en els diversosmodels cel·lulars (i també en condicions d’hipòxia enel cas dels cardiomiòcits). Aquest estudi es faran tantin vitro, utilitzant cultius primaris i línies cel·lulars(adipòcits 3T3-L1), com in vivo, en models experimen-tals de diabetis (per exemple, mitjançant la injecciód’estreptozotocina).
Intentar identificar els senyals intracel·lulars quemodulen l’expressió d’aquestes proteïnes i els factorsde transcripció involucrats.
1.2 Breu síntesi del disseny, procediments imètodesModels per utilitzar: línies cel·lulars secretores d'insuli-na; adipòcits 3T3-L1; cultius cel·lulars primaris;fraccions de teixits extrets de rata per estudis bioquí-mics i morfològics.
Tècniques per utilitzar: cultius cel·lulars, producció deproteïnes recombinants, generació d'antisèrums,electroforesi i electrotransferències, PCR, immunohis-toquímica, mesurament de la secreció d'insulina i detranslocació del GLUT4.
Mesurament de resultats: mesuraments de tipusqualitatiu, basats en la qualitat del material d'estudi;posada a punt d’algunes tècniques, identificació delscomponents cel·lulars esmentats en els objectius,mesurament d'insulina i translocació de GLUT4.
1.3 Síntesi del pla de treball originalEl pla de treball ha consistit en la caracteritzacióbioquímica i morfològica de models d'estudi, enl'adquisició d'eines moleculars de treball i en lapreparació de material biològic (bàsicament líniescel·lulars i cultius primaris), i s’ha prosseguit ambestudis més funcionals, entre els quals la realització detransfeccions i la utilització de neurotoxines clostridialsper estudiar tant el procés de la secreció d'insulinacom el de la translocació del transportador de glucosaGLUT4. Es van fer els estudis per a la caracteritzacióalgunes de les isoformes de les proteïnes implicadesen la secreció d'insulina (SNARE) i la seva interaccióamb altres proteïnes amb possible funció reguladora.
Es van produir clons estables de línies cel·lularssecretores d’insulina unida a la proteïna fluorescentverda (GFP) com a model biològic per estudiar lasecreció regulada d’insulina.
Posteriorment es van estudiar els canvis en l'expressióde proteïnes implicades en la translocació deltransportador de glucosa GLUT4 durant la diferencia-ció d’una línia cel·lular preadipocitària i d’aquelles queintervenen en la maquinària secretora de la insulina,tant en cultius cel·lulars com en animals d'experimen-tació, en condicions d’altes concentracions deglucosa.
2. ResultatsDiferenciació de la línia cel·lular 3T3-L1Les cèl·lules de la línia 3T3-L1 són fibroblastos quetenen la propietat de diferenciar-se en adipòcits endeterminades condicions. Aquestes cèl·lules han estatutilitzades per estudiar diversos aspectes de l'acció dela insulina en cèl·lules diana, ja que en estat diferenciatsón sensibles a la insulina.
La forma d'aquestes cèl·lules varia sensiblementdesprés de la seva diferenciació, passant de tenir unaforma que podem definir com fibroblàstica, allargada iamb expansions cel·lulars, a una forma arrodonida, demida gran i amb nombroses gotes lipídiques al seucitoplasma.
Hem pogut demostrar que aquest canvi morfològic vaacompanyat de canvis en l'expressió de proteïnesimplicades en el tràfic de membranes.
Així, mentre que en les cèl·lules no diferenciadesnomés es pot detectar la presència d’SNAP-23 isintaxina 4, però no en canvi sintaxina 1, SNAP-25,VAMP-1 o VAMP-2, una vegada diferenciades, lescèl·lules expressen sintaxina 1 i VAMP2, a mésd’incrementar la quantitat de sintaxina 4 i cellubrevina,i s’observaren canvis en la distribució d’SNAP-23 isintaxina 4.
No expressen, però, ni SNAP-25 ni VAMP-1. Ambl'expressió d'aquestes proteïnes, les cèl·lules 3T3-L1tenen la competència per translocar GLUT4, des d’uncompartiment vesicular a la membrana plasmàtica, jaque formen els requeriments mínims de dos t-SNARE(sintaxina i SNAP-23) i una v-SNARE (VAMP-2).
Aquests canvis són paral·lels a l'aparició i expressiódel transportador de glucosa GLUT4.
Expressió de proteïnes SNARE implicades en lasecreció d’insulina en illots pancreàtics
Una part del projecte tenia com a objectiu caracterit-zar les proteïnes implicades en la secreció d’insulinaen illots pancreàtics. Treballs precedents s'han limitata identificar aquelles proteïnes que anteriormenthavien estat involucrades en secreció regulada ensistema neuroendocrí. En aquest apartat hem ampliatel nombre d’aquestes proteïnes en una aproximacióbioquímica i morfològica.
L’anàlisi per Western blot va mostrar la presència desintaxina 1, SNAP-25, VAMP-2 i munc-18 en quanti-tats considerables, SNAP-23 i sintaxina 4 en menorquantitat i l'absència de VAMP1. Paral·lelament, esvan processar mostres de pàncrees tant en criostatcom en parafina per localitzar i identificar les proteïnes
FUNDACIÓ LA MARATÓ DE TV3

en les cèl·lules dels illots pancreàtics i pàncrees exocrí.
Aquests resultats no han d'estranyar, ja que s'haviadescrit la sensibilitat de les cèl·lules beta a lescadenes lleugeres de les neurotoxines clostridials queactuen selectivament sobre la sintaxina 1, SNAP-25 iVAMP-2.
Caracterització de proteïnes SNARE en la líniacel·lular INS-1
De manera semblant als illots pancreàtics i les cèl·lules3T3-L1, hem caracteritzat bioquímicament i morfològi-cament les cèl·lules secretores d’insulina INS-1, ambresultats similars.
Obtenció de clons estables de línies cel·lulars queexpressen proinsulina-GFP
Es va construir un vector d'expressió en eucariotesper a la proteïna quimèrica GFP-preproinsulina. Laproteïna que s'expressa està formada per la GFP(Green Fluorescent Protein), que és una proteïnafluorescent innòcua i la proinsulina. Amb aquestaconstrucció es preveu fer la monitorització en viu de lainsulina en el seu trànsit dins la cèl·lula i mesurar laseva secreció per mètodes fluoromètrics.
Com a línies cel·lulars per obtenir clons estables s'hanutilitzat: INS-1 (secretora d'insulina), 29.3T (fibroblastde ronyó humà), SH-SY5Y (neuroblastoma humàsecretor de noradrenalina), 3T3-L1 (línia preadipocità-ria). Els diversos tipus cel·lulars es van triar per podermesurar la secreció d’insulina en cèl·lules secretoresque per si mateixes la secreten (INS-1), com també enlínies secretores que no ho fan, per poder compararels dos sistemes. Les línies fibroblàstiques i preadipo-citàries es van utilitzar per obtenir clons amb secrecióconstitutiva. El cas de la línia 3T3-L1 és especial, jaque després de la seva diferenciació a adipòcit, té lapropietat de translocar el transportador de glucosaGLUT4 de manera dependent d’insulina.
Es van transfectar les cèl·lules per lipofecció i es vanseleccionar els clons mitjançant l'antibiòtic G-418, jaque les cèl·lules que mantenen el vector en sónresistents.
En l’obtenció de clons transfectats estables, calassenyalar la dificultat a obtenir la viabilitat o almenysel manteniment a llarg termini dels clons de les INS-1.No obstant això, s'ha obtingut una quantitat mínimaque s'intenta mantenir. De la resta de línies cel·lulars,s'han pogut obtenir clons estables.
Caracterització morfològica d’isoformes de sintaxinaen ronyó de rates diabètiques i normals
En aquest apartat es pretenien estudiar canvis enl’expressió d’isoformes de sintaxina provocats per ladiabetis en teixits no directament sensibles a insulina,com el ronyó. Es van poder determinar les isoformes ila seva localització, però no es van observar diferen-cies significatives ni en la quantitat ni en la distribucióde les isoformes de sintaxina.Descripció de les afinitats en la unió d’isoformes de
sintaxina (involucrades en la secreció regulada) i altresproteïnes (munc-18a)
Una de les hipòtesis de treball es basa en el fet quel'existència d’isoformes de proteïnes SNARE pot sercausada per requeriments funcionals específics decada tipus cel·lular, bé sigui secretor o no. Així, en elsistema nerviós existeixen dues isoformes per a cadaSNARE implicada en la neurosecreció (sintaxina 1A i1B, sinaptobrevina 1 i 2, SNAP-25a i b). En canvi, enla cèl·lula b -pancreàtica, tant sols hi ha una solaisoforma de cadascuna (sintaxina 1A i sinaptobrevina2). Això pot fer pensar que la combinació d'aquestesisoformes és la més efectiva, o l’única que pot funcio-nar en secreció de productes peptídics.
Per això es van dissenyar experiments per estudiar lespossibles diferències d'afinitat entre isoformes deproteïnes SNARE i proteïnes associades.
Estudis previs assenyalen que la sinaptobrevina 2 ésla isoforma que s'uneix amb més afinitat a la sintaxina1A, mentre que la sintaxina 1B no presenta diferèn-cies significatives en la seva unió a les sinaptobrevines1 i 2. (SNAP-25 tampoc no presenta diferencies enl'afinitat). En el present treball es van intentar estudiarles afinitats entre les isoformes de sintaxina 1 i munc-18, una proteïna citosòlica que és essencial per a lasecreció i que interactua amb la sintaxina.
Els resultats mostren una major afinitat entre sintaxina1A i munc-18 in vitro (entre proteïnes recombinants).Els resultats també mostren que la localització demunc-18 en membrana plasmàtica es produeix per laseva interacció amb la sintaxina 1.
Mitjançant el mètode del triple híbrid, es va podercomprovar que munc-18 bloquejava la interacció desintaxina 1 amb la resta de proteïnes SNARE, totsuggerint una funció reguladora de munc-18 en laformació de complexos SNARE.
Descripció preliminar de canvis en l’expressió deproteïnes implicades en la secreció d’insulina en illotstrasplantats i en condicions hiperglicèmiques inormoglicèmiques
En col·laboració amb el laboratori del Dr. EduardMontanya, hem intentat caracteritzar el patród'expressió, mitjançant tècniques immunohistoquími-ques i de Western blot, de proteïnes implicades enl’exocitosi d’insulina en tres condicions experimentals:
Illots de rata que s'havien aïllat i trasplantat sota lacàpsula renal d'altres rates prèviament tractades ambestreptozotocina.
Pàncreas de rates pancreatomitzades.
Cultius d’illots pancreàtics amb concentracions altesde glucosa en el medi.
En tots els casos es van diferenciar dues situacions,normoglicèmia i hiperglicèmia. Els resultats mostrenque hi ha un paral·lelisme entre la quantitat d’insulina il'expressió de proteïnes implicades en la secreció que
RECERCA BIOMÈDICA | 81

es troben als grànuls secretors (sobretot de les v-SNARES). Mentre que en el cas de rates i condicionsnormoglicèmiques els nivells es poden considerarnormals, en rates i condicions hiperglicèmiques elsnivells d'insulina i també d’altres proteïnes presents alsgrànuls, com la sinaptobrevina i la cellubrevina, sónmenors o inexistents. Això indicaria que, juntamentamb la incapacitat o disminució en la producciód’insulina, les cèl·lules b d'illots que es mantenenhiperglicèmiques, també tenen disminuïda l'expressióde proteïnes normalment presents als grànulssecretors, tot suggerint que ni tan sols es deuenformar aquests.
Caracterització de les proteïnes implicades en lasecreció d'insulina en la línia cel·lular INS-1
La línia cel·lular secretora d’insulina INS-1 ha estatmolt utilitzada per a estudis de secreció de l’hormona.Aquesta línia cel·lular sintetitza la insulina, l’emmagat-zema en grànuls de secreció i l'allibera per exocitosicom a resposta a nivells elevats de glucosa.
Hem observat que aquesta secreció es veu significati-vament incrementada en activar conjuntament la viade la proteïna cinasa C amb TPA.
En aquesta part del treball s'han intentat identificar percentrifugació diferencial, immunotransferència itècniques morfològiques, les proteïnes SNARE i altresproteïnes relacionades amb la secreció d’insulina en lalínia INS-1. S’han analitzat en paral·lel cèl·lules INS-1 i29.3T transfectades amb preproinsulina-GFP.
3. Rellevància i possibles implicacions clíniquesdels resultats finals obtingutsEls coneixements generats en aquest projecte són detipus bàsic, incrementen el coneixement sobre:
La presència de proteïnes implicades en la transloca-ció del transportador de glucosa GLUT4 i en lasecreció d’insulina en diferents models cel·lulars.
El comportament de les cèl·lules diana de la insulinadurant el procés de la diferenciació i maduració.
Els canvis en l’expressió de proteïnes en les cèl·lulesß-pancreàtiques en condicions d’hiperglicèmia.
La interacció de proteïnes implicades en els proces-sos de translocació del transportador de glucosaGLUT4 (en adipòcits) i de secreció d’insulina (cèl·lulaß-pancreàtica).
Els resultats amb possibles implicacions clíniques ique criden més l’atenció són els relacionats amb:
Els canvis en l’expressió de proteïnes provocats perelevades dosis de glucosa.
Els canvis que acompanyen la diferenciació de teixitsdiana de la insulina, principalment l’augment enl’expressió de certes proteïnes i la seva redistribuciódintre de les cèl·lules.
Un dels nostres propòsits en el futur és determinar la
importància d’aquests canvis, és a dir, com podenafectar no solament la funció cel·lular (exposició detransportador de glucosa a la superfície cel·lular peracció de la insulina i la mateixa secreció d’insulina),sinó també la seva supervivència.
4. PublicacionsB. Torrejón, I. Gómez de Aranda and J. Blas SNARE expres-
sion and distribution during 3T3-L1 adipocyte differentiation.
FEBS Letters 512: 275-281 (2002)
F. Pérez-Brangulí, A. Muhaisen and J. Blasi
Títol: Munc 18a binding to syntaxin 1A and 1B isoforms defines
its localization at the plasma membrane and blocks SNARE
assembly in a three-hybrid system assay. Molecular and Cellular
Neuroscience 20: 169-180 (2002)
E.Martí, J. Blasi and I. Ferrer. Early induction of secretoneurin
expression following kainic acid administration at convulsant
doses in the rat and gerbil hippocampus. Hippocampus 12 (2):
174-185 (2002)
J. Aleu, J. Blasi, C. Solsona and J. Marsal. Calcium-dependent
acetylcholine release from Xenopus oocytes: Simultaneous
currents and acetylcholine release recordings.European Journal of
Neuroscience 16 (8): 1442-8 (2002)
M. A. Carmona, A. Martínez, A. Soler, J. Blasi, E. Soriano and
F. Aguado. Ca2+-Evoked Synaptic Transmission and
Neurotransmitter Receptor Levels are Impaired in the Forebrain of
trkB (-l-) Mice. Molecular and Cellular Neuroscience 22(2): 210-
226 (2003)
D. Reigada, I. Diez-Perez, P. Gorostiza, A. Verdaguer, I.
Gómez de Aranda, O. Pineda, J. Vilarrasa, Marsal, J., J. Blasi,
J. Aleu, C. Solsona. Control of Transmitter release by an
internal gel-matrix in synaptic vesicles. Proc. Natl. Acad. Sci
(USA) 100(6): 3485-3490 (2003)
En preparació:SNARE components in the insulin secretory cell line INS-1
Characterization of cells stably transfected with pre-pro-insulin-
GFP
Expression of SNARE proteins under different hyperglycemic
conditions
FUNDACIÓ LA MARATÓ DE TV3

RECERCA BIOMÈDICA | 83
MEMÒRIA FINAL
1. Resum del projecte originalL’objectiu últim del treball consisteix en la síntesienantioselectiva d’azasucres d’estructura polihidroxipi-peridínica (azaanàlegs de piranoses) amb activitatinhibidora de les glucosidases, que siguin d’utilitat enel tractament de la diabetis.
Per aconseguir aquest objectiu es pretén desenvolu-par una nova metodologia de síntesi, versàtil i eficaç,que permeti la preparació d’azasucres pertanyents adiverses sèries diastereomèriques, en les duespossibles configuracions absolutes (natural i nonatural), com també la introducció d’àtoms de fluor enles diverses posicions de l’anell.
Basant-nos en l’experiència adquirida en el nostrelaboratori durant els darrers anys en la síntesi enantio-selectiva de piperidines diversament substituïdes apartir de lactames bicícliques derivades de l’R- i del’S-fenilglicinol i en els precedents actualmentestablerts sobre relacions estructura-activitat d’azasu-cres com a inhibidors de glicosidases, el treball esdirigirà a estudiar els aspectes següents:
1. Desenvolupament de vies de síntesi alternatives,que superin les existents per determinats azasucresamb demostrada activitat com a inhibidors de glucosi-dases i d’aplicació en el tractament de la diabetis.
2. Síntesi de nous azasucres.
3. Extensió dels anteriors estudis a la síntesi d’azasu-cres en les dues possibles sèries enantiomèriques,amb l’objecte d’accedir tant a azasucres relacionatsamb els sucres de configuració natural com azasucresde configuració no natural. Això serà possible simple-ment utilitzant ja sigui R- o S-fenilglicinol (ambdósfàcilment assequibles) com a productes de partida.Aquesta possibilitat representa un clar avantatgerespecte d’altres mètodes previs descrits per a lasíntesi d’azasucres, en els quals com a productes departida s’utilitzen carbohidrats d’origen natural i, pertant, pertanyents a la sèrie D. Aquests mètodes nopermeten l’accés a azasucres de configuració nonatural.
Els compostos sintetitzats seran sotmesos a avaluacióbiològica a fi de determinar la seva activitat com ainhibidors de glucosidases i la seva potencial utilitat enel tractament de la diabetis. Les dades aportades per
Títol del projecte
Antidiabètics orals. Nous procediments de síntesi d'azasucres inhibidors de glucosidases
Investigadors principalsJoan Bosch Cartes
InstitucióFacultat de Farmàcia. UB
Membres de l’equipOriol Bassas, Oscar Lozano, Juan Aguilar, Marta Huguet, Antonia Gómez, María Pérez, Joan Bosch, MercedesAmat, Núria Llor, Josefina Badía i Laura Baldomà.
62.161 €

aquests assaigs, in vitro i in vivo ens permetranaprofundir en el coneixement de les relacions estruc-tura-activitat dels azasucres i, en conseqüència, en elconeixement del mecanisme de la inhibició enzimàticade les glucosidases, la qual cosa possibilitarà eldisseny de nous compostos amb, a priori, unpotencial superior com a antidiabètics.
2. ResultatsA continuació s’esquematitzen les seqüències sintèti-ques dutes a terme:
1) Preparació de cis-3,4-dihidroxipiperidines enantiopures
Reactius i condicions: (a) OsO4, NMO; (b)(CH3)2C(OCH3)2, p-TsOH, després BH3-THF; (c) H2 /Pd-C, MeOH-HCl.
2) Mètode per a la preparació enantioselectiva de 3-hidroxipiperidines enantiopures mitjançant la hidroxila-ció oxidativa d’una dihidropiridona derivada del fenilgli-cinol.
Reactius i condicions: (a) m-CPBA, CH2Cl2; (b) BH3,THF; (c) Pd(OH)2, Boc2O, AcOEt.
3) Procediment per introduir el substituent metoximetilpresent en la posició 2 en molts azasucres.
Reactius i condicions: (a) TMSCN, TiCl4; (b) 2-metil-cianocuprat de liti, BF3.Et2O; (c) O3, després NaBH4.
4) Mètode per a l’obtenció de 3,4,5-trihidroxipiperidi-nes enantiopures mitjançant hidroxilació oxidativad’una 2-piridona derivada de l’(R)-fenilglicinol.
Reactius i condicions: (a) m-CPBA; (b) OsO4, NMO;(c) (CH3)2C(OCH3)2, p-TsOH, després BH3, THF; (d)H2/Pd-C, MeOH després, MeOH-HCl; (e) MeOH-HCl.
5) Preparació d’azasucres en l’altra forma enantiomèri-ca a partir d’una 2-piridona derivada de l’(S)-fenilglicinol.
Reactius i condicions: (a) m-CPBA; (b) OsO4, NMO;(c) (CH3)2C(OCH3)2, p-TsOH, després BH3, THF; (d)H2/Pd-C, MeOH, després MeOH-HCl; (e) MeOH-HCl.
6) Introducció d’un àtom de fluor. Preparació d’hidro-xifluoropiperidines (fluoroazasucres) enantiopures.
Reactius i condicions: (a) H2/Pd-C, MeOH, desprésMeOH-HCl; (b) Boc2O, Et3N; (c) DAST; (d) H2/Pd-C,MeOH, després MeOH-HCl.
7) Procediment alternatiu per a la introducció del gruphidroximetil present en nombrosos azasucres:preparació de 3-hidroxi-(2-hidroximetil)piperidinesenantiopures.
Reactius i condicions: (a) NH(CH3)(OCH3).HCl, piridi-na; (b) BuLi, 1,3-ditià; (c) LDA, Br(CH2)2COOEt; (d)KOH, MeOH; (e) (R)-fenilglicinol; (f) NBS, acetona-H2O; (g) NaBH4, CeCl3, MeOH; (h) BH3, THF; (i) H2,Pd(OH)2, (t-BuOCO)2O.
8) Preparació de l’azasucre desoxi-D-gulonojirimicina.
FUNDACIÓ LA MARATÓ DE TV3
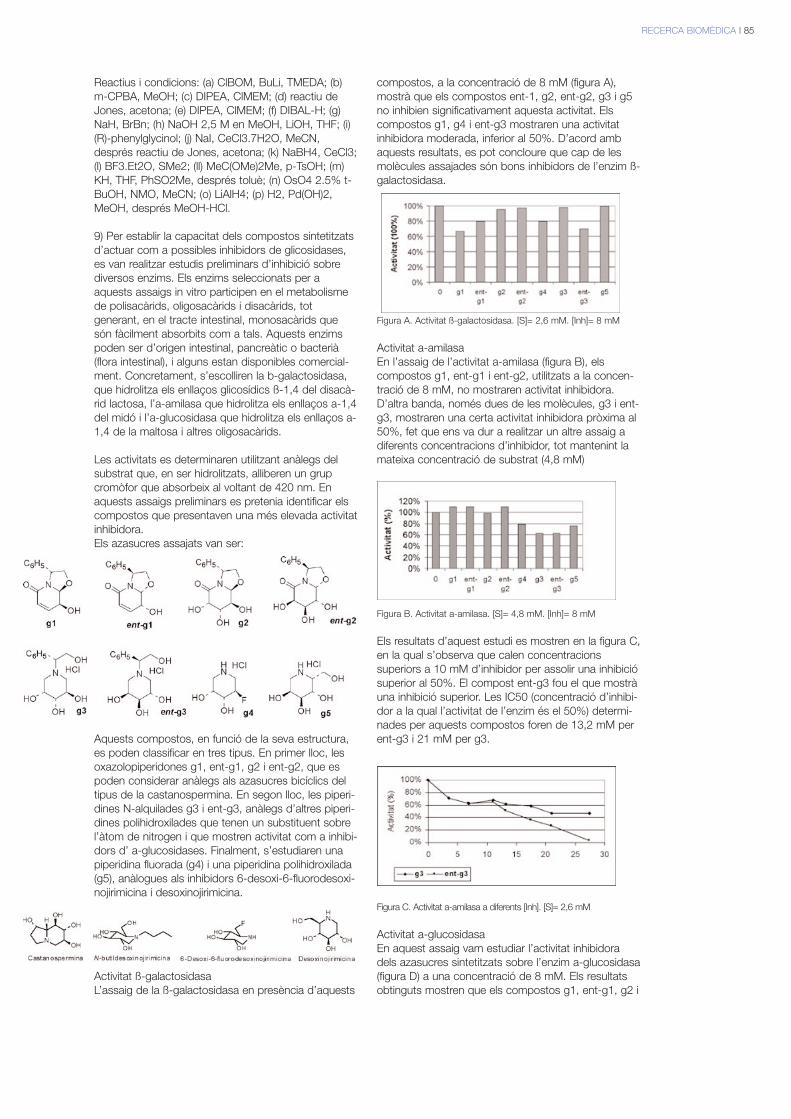
Reactius i condicions: (a) ClBOM, BuLi, TMEDA; (b)m-CPBA, MeOH; (c) DIPEA, ClMEM; (d) reactiu deJones, acetona; (e) DIPEA, ClMEM; (f) DIBAL-H; (g)NaH, BrBn; (h) NaOH 2,5 M en MeOH, LiOH, THF; (i)(R)-phenylglycinol; (j) NaI, CeCl3.7H2O, MeCN,després reactiu de Jones, acetona; (k) NaBH4, CeCl3;(l) BF3.Et2O, SMe2; (ll) MeC(OMe)2Me, p-TsOH; (m)KH, THF, PhSO2Me, després toluè; (n) OsO4 2.5% t-BuOH, NMO, MeCN; (o) LiAlH4; (p) H2, Pd(OH)2,MeOH, després MeOH-HCl.
9) Per establir la capacitat dels compostos sintetitzatsd’actuar com a possibles inhibidors de glicosidases,es van realitzar estudis preliminars d’inhibició sobrediversos enzims. Els enzims seleccionats per aaquests assaigs in vitro participen en el metabolismede polisacàrids, oligosacàrids i disacàrids, totgenerant, en el tracte intestinal, monosacàrids quesón fàcilment absorbits com a tals. Aquests enzimspoden ser d’origen intestinal, pancreàtic o bacterià(flora intestinal), i alguns estan disponibles comercial-ment. Concretament, s’escolliren la b-galactosidasa,que hidrolitza els enllaços glicosídics ß-1,4 del disacà-rid lactosa, l’a-amilasa que hidrolitza els enllaços a-1,4del midó i l’a-glucosidasa que hidrolitza els enllaços a-1,4 de la maltosa i altres oligosacàrids.
Les activitats es determinaren utilitzant anàlegs delsubstrat que, en ser hidrolitzats, alliberen un grupcromòfor que absorbeix al voltant de 420 nm. Enaquests assaigs preliminars es pretenia identificar elscompostos que presentaven una més elevada activitatinhibidora.Els azasucres assajats van ser:
Aquests compostos, en funció de la seva estructura,es poden classificar en tres tipus. En primer lloc, lesoxazolopiperidones g1, ent-g1, g2 i ent-g2, que espoden considerar anàlegs als azasucres bicíclics deltipus de la castanospermina. En segon lloc, les piperi-dines N-alquilades g3 i ent-g3, anàlegs d’altres piperi-dines polihidroxilades que tenen un substituent sobrel’àtom de nitrogen i que mostren activitat com a inhibi-dors d’ a-glucosidases. Finalment, s’estudiaren unapiperidina fluorada (g4) i una piperidina polihidroxilada(g5), anàlogues als inhibidors 6-desoxi-6-fluorodesoxi-nojirimicina i desoxinojirimicina.
Activitat ß-galactosidasaL’assaig de la ß-galactosidasa en presència d’aquests
compostos, a la concentració de 8 mM (figura A),mostrà que els compostos ent-1, g2, ent-g2, g3 i g5no inhibien significativament aquesta activitat. Elscompostos g1, g4 i ent-g3 mostraren una activitatinhibidora moderada, inferior al 50%. D’acord ambaquests resultats, es pot concloure que cap de lesmolècules assajades són bons inhibidors de l’enzim ß-galactosidasa.
Figura A. Activitat ß-galactosidasa. [S]= 2,6 mM. [Inh]= 8 mM
Activitat a-amilasaEn l’assaig de l’activitat a-amilasa (figura B), elscompostos g1, ent-g1 i ent-g2, utilitzats a la concen-tració de 8 mM, no mostraren activitat inhibidora.D’altra banda, només dues de les molècules, g3 i ent-g3, mostraren una certa activitat inhibidora pròxima al50%, fet que ens va dur a realitzar un altre assaig adiferents concentracions d’inhibidor, tot mantenint lamateixa concentració de substrat (4,8 mM)
Figura B. Activitat a-amilasa. [S]= 4,8 mM. [Inh]= 8 mM
Els resultats d’aquest estudi es mostren en la figura C,en la qual s’observa que calen concentracionssuperiors a 10 mM d’inhibidor per assolir una inhibiciósuperior al 50%. El compost ent-g3 fou el que mostràuna inhibició superior. Les IC50 (concentració d’inhibi-dor a la qual l’activitat de l’enzim és el 50%) determi-nades per aquests compostos foren de 13,2 mM perent-g3 i 21 mM per g3.
Figura C. Activitat a-amilasa a diferents [Inh]. [S]= 2,6 mM
Activitat a-glucosidasaEn aquest assaig vam estudiar l’activitat inhibidoradels azasucres sintetitzats sobre l’enzim a-glucosidasa(figura D) a una concentració de 8 mM. Els resultatsobtinguts mostren que els compostos g1, ent-g1, g2 i
RECERCA BIOMÈDICA | 85

ent-g3 no presenten activitat com a inhibidorsd’aquest enzim, mentre que g3, ent-g2, g4 i g5inhibien significativament. Dels quatre, els que presen-taven activitat inhibidora més rellevant eren g4 i g5.Per això es van fer nous assaigs a diferents concen-tracions d’inhibidor, per determinar la IC50.
Figura D. Activitat a-glucosidasa. [S]= 1,4 mM. [Inh]= 8 mM
La capacitat inhibidora dels compostos g5 i g4 va seranalitzada a concentracions inferiors a 8 mM. Tal coms’observa en la figura E, la piperidina fluorada g4presenta una IC50 de 2,5 mM i inhibeix completamentl’enzim a la concentració de 4 mM. Pel que fa a lapiperidina g5 (figura F), la IC50 és 250 mM i aconse-gueix la inhibició completa de l’enzim a la concentra-ció de 2 mM.
Figura E. Activitat a-glucosidasa. [S]= 1,4 mM
Figura F. Activitat a -glucosidasa. [S]= 1,4 mM
Com que g5 és un diastereòmer de la desoxinojirimici-na (DNJ), conegut inhibidor d’ a -glucosidases, es vafer un estudi comparatiu de la capacitat inhibidora d’ a-glucosidasa de S. cerevisiae d’aquests dos isòmers.L’estudi es va fer utilitzant la mateixa concentració desubstrat i diferents concentracions de l’inhibidorcomercial (figura G). Els resultats obtinguts mostrenque la IC50 de la DNJ, en aquest model, és 1,25 mMi, per tant, superior a la del nostre isòmer g5, que és de 0,25 mM.
Figura G. Activitat a-glucosidasa. [S]= 1,4 mM
Estudis preliminars de cinètica enzimàtica amb elcompost g5 actuant sobre l’activitat a -glucosidasa nohan permès demostrar que el mecanisme d’inhibiciósigui del tipus competitiu.
Les representacions de dobles inversos (Lineweaverand Burk) elaborades per altres autors per a la desoxi-nojirimicina indicava, per contra, que la inhibició era detipus competitiu i que molt probablement corresponiaa un anàleg de l’estat de transició del mecanismecatalític. En el nostre cas, aquesta afirmació no potésser sostinguda amb les dades obtingudes. Es potconcloure finalment que el compost g5 (desoxi-D-gulonojirimicina) sintetitzat en aquest treball escomporta com un millor inhibidor de l’activitat ?-glucosidasa que el compost natural abans esmentat(desoxinojirimicina) i que des d’aquesta perspectivapot ser un compost amb un valor afegit per a futursestudis d’antidiabètics orals de síntesi.
3. Rellevància i possibles implicacions clíniquesdels resultats finals obtingutsLa síntesi de noves molècules amb aplicacionsterapèutiques constitueix una àrea fonamental en elmón de la biomedicina actual. En l’àmbit dels antidia-bètics orals, hi ha molts productes naturals i derivatsque han donat informació d’interès per al coneixementdels requeriments estructurals per aconseguir determi-nades activitats farmacològiques. A partir d’aquestconeixement, la síntesi orgànica permet aproximar-sea noves estructures amb un interès i valors afegits.
En aquest context, el present projecte ha aplicat unanova metodologia de síntesi enantioselectiva perobtenir compostos d’interès com a antidiabètics oralsbasats en la inhibició de glucosidases. Aquestmecanisme impedeix l’alliberació de molècules deglucosa d’oligo o polisacàrids de la dieta, totdisminuint així l’absorció de molècules de glucosa. Larellevància rau fonamentalment en l’aplicaciód’aquests processos de síntesi per a l’obtenció demolècules de possible aplicació al camp dels antidia-bètics orals. Hem aconseguit un conjunt de compos-tos que han pogut ser assajats amb diversos enzimsamb activitat glucosidasa. Entre els quals cal destacardos possibles candidats per a futurs estudis d’interèsen la millora de molècules amb les indicades propie-tats farmacològiques, com són la piperidina fluoradag4 i la desoxi-D-gulonojirimicina g5, aquest últim ambla particularitat d’ésser millor inhibidor d’ a-glucosida-ses que la desoxinojirimicina.
FUNDACIÓ LA MARATÓ DE TV3

4. PublicacionsMercedes Amat, Núria Llor, Marta Huguet, Elies Molins,
Enrique Espinosa, Joan Bosch. "Unprecedented Oxidation of a
Phenylglycinol-Derived 2-Pyridone: Enantioselective Synthesis of
Polyhydroxypiperidines". Org. Lett. 2001, 3, 3257-3260.
Mercedes Amat, Carmen Escolano, Núria Llor, Marta Huguet,
Maria Pérez, Joan Bosch. "Stereoselective a-amidoalkylation
reactions of phenylglycinol-derived bicyclic lactams". Tetrahedron:
Asymmetry 2003, 14, 1679-1683.
Mercedes Amat, Marta Huguet, Núria Llor, Oriol Bassas,
Antonia Mª Gómez, Joan Bosch, Josefa Badia, Laura Baldoma y
Juan Aguilar. "Enantioselective synthesis of 1-deoxy-D-gulonoji-
rimycin from a phenylglycinol-derived lactam". Tetrahedron Lett.
2004 (enviat)
RECERCA BIOMÈDICA | 87

FUNDACIÓ LA MARATÓ DE TV3
Títol del projecte
Aproximacions de teràpia gènica per a la diabetis mellitus centradesen la manipulació genètica del pàncrees i teixits extrapancreàtics
Investigador principalFàtima Bosch i Tubert
InstitucióFacultat de Veterinària. UAB
131.271 €
MEMÒRIA FINAL
1. Resum del projecte originalObjectius i dissenyL’objectiu general d’aquest projecte és el desenvolu-pament de noves aproximacions de teràpia gènica pera la diabetis mellitus centrades en la manipulaciógenètica del pàncrees i teixits extrapancreàtics.Aquest objectiu pretén:
1) Estudiar la capacitat de disminuir la hiperglucèmiadiabètica mitjançant la producció regulada d’insulinamadura pel múscul esquelètic.
2) Investigar la capacitat de disminuir la hiperglucèmiai la cetoacidosi diabètiques mitjançant l’increment dela captació de glucosa i la disminució de l’alliberamentd’àcids grassos lliures pel teixit adipós.
3) Establir una nova aproximació terapèutica per a ladiabetes tipus 1 centrada en la regeneració delpàncrees endocrí.
Aquest objectiu general, el dividim en els objectiusespecífics següents:
1. Estudi in vivo de la capacitat de reduir la hiperglu-cèmia diabètica mitjançant la transferència del gende la insulina a múscul esquelètic utilitzant vectorsvirals i no virals.
1.1 Transferència de diversos gens quimèrics, quepermetran l’expressió regulada d’insulina, a músculesquelètic de ratolins diabètics mitjançant la utilitzacióde virus adenoassociats o lentivirus.
1.2 Transferència de diversos gens quimèrics, quepermetin l’expressió regulada d’insulina, mitjançant lainjecció directa a múscul esquelètic de ratolinsdiabètics de DNA en solució o en formulacionssòlides.
2. Estudi in vivo de la capacitat del teixit adipós dereduir la hiperglucèmia i la cetoacidosi diabèticamitjançant la utilització d’animals transgènics models.
2.1 Obtenció i anàlisi d’animals transgènics que expres-sin glucoquinasa (GK) en el teixit adipós. Aquestsanimals s’obtindran per microinjecció del gen quimèricaP2/GK a embrions d’una sola cèl·lula de ratolí.

2.2 Obtenció i anàlisi d’animals transgènics queexpressin el transportador de glucosa GLUT2 en elteixit adipós. Aquests animals s’obtindran per microin-jecció del gen quimèric aP2/GLUT2 a embrions d’unasola cèl·lula de ratolí.
2.3 Obtenció i anàlisi d’animals doble transgènics queexpressin GK i GLUT2 en el teixit adipós. Un copcaracteritzats, els animals que expressin GLUT2s’encreuaran amb els animals transgènics que expres-sin GK.
Tots aquests animals transgènics es tractaran ambestreptozotocina per estudiar la capacitat de contra-restar les alteracions causades per la manca d’insulina.
3. Estudi in vivo de la capacitat de regeneració delpàncrees endocrí en animals diabètics.
3.1 Estudi del mecanisme/s que donen lloc a unincrement de la replicació i neogènesi de cèl.lules ? enpàncrees d’animals transgènics que sobreexpressenIGF-I.
3.2 Estudi de la capacitat de vectors virals (virusadenoassociats [AAV] o lentivirus) per transferir el gende l’IGF-I a pàncrees endocrí in vivo.
Aquest estudi es durà a terme tant en ratolins com engossos diabètics.
3.3 Estudi del paper de l’IGF-II en la regeneració delpàncrees endocrí utilitzant animals transgènics quesobreexpressin IGF-II en cèl·lules ß.
2. ResultatsDurant el desenvolupament del projecte s’han dut aterme les activitats que s’especifiquen a continuació:
1. Estudi in vivo de la capacitat de reduir lahiperglucèmia diabètica mitjançant la manipulaciógenètica del múscul esquelètic.
1.1 Estudi in vivo de la capacitat de reduir la hiperglucè-mia diabètica mitjançant la transferència del gen de lainsulina a múscul esquelètic utilitzant vectors no virals.
En aquest estudi, en primer lloc vam examinar lacapacitat de contrarestar la hiperglucèmia diabèticamitjançant la producció constitutiva d’insulina pelmúscul esquelètic. Per això, vam transferir el genquimèric CMV/Insm a múscul esquelètic (tibialis igastrocnemius) de ratolí diabètic (tractat amb 5 dosisconsecutives de 40 mg/kg pes d’estreptozotocina[STZ]) mitjançant la injecció de DNA plasmídic ensolució i electrotransferència in situ. S’havia descritque DNA plasmídic injectat directament en solució amúscul esquelètic, prèviament tractat amb hialuroni-dasa, era captat per les miofibres, i que els gens queeren al vector plasmídic es podien expressar durantmés de dos mesos després de la transferència.Aquesta tècnica ens va permetre transferir al voltantdel 80% de les fibres musculars en els ratolinsdiabètics. Els estudis duts a terme en aquests animalsens indiquen que la insulina produïda pel múscul portaa una normalització de la glicèmia en dejuni, però no
contraresta la hiperglucèmia en estat d’alimentació.No obstant això, aquests animals són molt sensibles abaixes dosis d’insulina, de manera que presenten unanormalització de la glicèmia en condicions d’alimenta-ció després de l’administració de petites dosis del’hormona.
1.2 Estudi in vivo de la capacitat de reduir la hiperglu-cèmia diabètica mitjançant la transferència del gen dela glucoquinasa a múscul esquelètic utilitzant vectorsno virals.
També durant aquest període hem transferit el gen dela glucoquinasa (GK) a múscul esquelètic d’animalsdiabètics in vivo mitjançant electrotransferència deDNA plasmídic (pGK) que conté el gen quimèricCMV/GK. Aquests animals són hiperglucèmics enestat d’alimentació però presenten normoglucèmia endejuni. Amb tot, en condicions d’alimentació, entractar-los amb baixes dosis d’insulina soluble, aquestsanimals que expressen GK recuperen ràpidament lanormoglucèmia, mentre que animals controls diabèticscontinuen altament hiperglucèmics (Gene Ther.).
1.3 Trasplant de cèl·lules musculars que expressen elgen de la GK i anàlisi de la capacitat de contrarestarla hiperglucèmia diabètica.
En aquesta part del projecte, també vam obtenircèl·lules de mioblast que després de diferenciar amiotub expressaven GK (cèl·lules C2C12GK).Aquestes cèl·lules presenten un increment en lacaptació de glucosa dependent de la concentració delsucre del medi de cultiu. En trasplantar-les a ratolinssingènics C3H, aquestes cèl·lules no es dividien, jaque es diferenciaven a miotubs. A més, aquestsanimals no desenvolupaven en cap moment hipoglu-cèmia. En tractar amb STZ els ratolins trasplantatsamb miotubs que expressen GK, es va observar quees contrarestava la hiperglucèmia, polidípsia i polifàgia,mentre que els ratolins trasplantats amb cèl·lulescontrols desenvolupaven diabetis. Similarment, quanes van trasplantar miotubs controls a animalsdiabètics, aquests continuen hiperglucèmics. Encanvi, el trasplantament de miotubs que expressenGK en animals diabètics normalitza la glucèmia.Aquests resultats suggereixen que l’ús de cèl·lulesmusculars manipulades genèticament perquè expres-sin GK pot proporcionar una nova aproximació deteràpia gènica ex vivo per reduir la hiperglucèmiadiabètica que estaria regulada per glucosa (Hum.Gene Ther. [2002] 13:21225-2133).
1.4 Estudi in vivo de la capacitat de normalitzar lahiperglucèmia diabètica mitjançant la transferènciaconjunta del gen de la insulina i el gen de la GK amúscul esquelètic utilitzant vectors no virals i virals.
Com que la transferència del gen de la insulina o bé elde la GK només portava a la reducció parcial de laglicèmia i calia una teràpia combinada de teràpiagènica (transferència del gen insulina o el de la GK)conjuntament amb l’administració de dosis baixesd’insulina exògena, vam decidir investigar si amb unatransferència conjunta de dos gens (insulina i GK) espodia realment contrarestar la malaltia.
RECERCA BIOMÈDICA | 89

Per això, per tal de provar el concepte, es vanencreuar durant el primer any animals transgènics queexpressaven a múscul esquelètic el gen de la insulina(Tg MLC/Insm) amb animals que expressaven enaquest teixit l’enzim GK (Tg MLC/GK). Així, es vanobtenir animals doble transgènics que expressavenalhora insulina i GK a múscul esquelètic. En adminis-trar STZ a aquests animals, hem observat que sóncapaços de revertir la diabetis tipus 1, ja que esnormalitza la glucèmia, el consum de menjar i d’aigua,i els tests de tolerància a la glucosa i a la insulina, enpart com a resultat de la millora del metabolismemuscular i hepàtic. A més, aquests animals dobletransgènics diabètics no van desenvolupar en capmoment episodis d’hipoglucèmia.
Això ens indicava que la producció d’uns nivellsbasals d’insulina són suficients perquè la glucosapugui entrar al múscul esquelètic, ja que indueix lapresència del transportador GLUT4 a la membranaplasmàtica. Un cop dins, la GK fosforila la glucosaeficientment a causa de la seva elevada Km. Per tant,la GK és en realitat la que modula la captació de laglucosa arran de les seves característiques cinètiques,fet que permet que es metabolitzi més o menysglucosa dependent de la concentració del sucrecirculant. També ens evidencien que la producció denivells basals d’insulina de manera constant éssuficient per contrarestar eficientment el procésdiabètic i no cal la producció regulada de nivells altsd’hormona. En aquest model animal es demostra perprimera vegada com la cooperació entre dos gens(insulina i GK) permet obtenir una teràpia altamenteficient per a la diabetis.
Per tant, a partir d’aquests resultats, a continuacióvam iniciar una nova aproximació de teràpia gènicaper a la diabetis tipus 1 basada en la transferència delgen de la insulina i el de la GK a múscul esquelèticmitjançant la tècnica d’electrotransferència de DNAplasmídic a múscul esquelètic. En electrotransferiralhora el gen de la insulina i el gen de la GK a músculesquelètic, hem observat que es normalitza no nomésla glucèmia en animals diabètics en dejuni sinó tambéen animals alimentats. A més, no es va observar encap moment el desenvolupament d’hipoglucèmia.Aquests animals també presentaven una normalitzaciódel metabolisme de la glucosa, tant muscular comhepàtic, i de l’aigua i el menjar ingerits. Tots aquestsresultats indiquen que la cooperació de la insulinaproduïda pel múscul conjuntament amb la majorcaptació de glucosa per aquest teixit portava a larecuperació completa del procés diabètic. Per tant,que aquesta aproximació de teràpia gènica podria sermolt útil per a un tractament altament eficient per a ladiabetis, ja que permetria un millor control de l’home-òstasi de la glucosa en els pacients.
Per obtenir una teràpia a llarg termini, també vamtransferir a múscul esquelètic el gen de la GK i lainsulina mitjançant vectors virals. Per això, vam obtenirvectors adenoassociats de serotip 1, AAV1, quecontenien insulina (AAV1-Ins) i GK (AAV1-GK), ja ques’havia descrit que eren millors vectors per infectarmúscul esquelètic, que permetien l’expressió delsgens d’interès més enllà d’un any després de la
transducció i que en ratolins no s’havia observatresposta immune contra les cèl·lules infectades. Entransferir la GK i la insulina mitjançant vectors AAV1,vam observar una restauració completa de lanormoglicèmia, tant en condicions d’alimentació comde dejuni més enllà de sis mesos després de ladestrucció de les cèl·lules ß. En canvi, els ratolinscontrols diabètics no tractats no sobrevivien.L’expressió de GK i insulina permetia també lanormalització de l’estat general de l’animal, ja quepresentava una normalització del metabolismemuscular i hepàtic i de la ingesta de menjar i aigua, noperdia massa muscular i mantenia la massa del teixitadipós.
Per tant, en aquest estudi hem demostrat mitjançanttres aproximacions diferents en ratolins diabètics queuna teràpia gènica conjunta que permeti expressaralhora el gen de la insulina i de la GK porta no nomésa contrarestar la hiperglucèmia diabètica, sinó tambéa normalitzar totes les alteracions metabòliquesd’aquesta malatia. Així doncs, a partir d’aquestsresultats ens podem plantejar d’endegar el tractamentd’animals més grans, com ara gossos, com a pasprevi a l’aplicació d’una teràpia conjunta amb insulina iGK en pacients humans.
2. Estudi in vivo de la capacitat del teixit adipós dereduir la hiperglucèmia i la cetoacidosi diabèticamitjançant la utilització d'animals transgènicsmodels.La diabetis mellitus està caracteritzada per unahiperglucèmia crònica que porta a complicacionsmicrovasculars, macrovasculars i neurològiques. Unincrement en la utilització de glucosa pels teixits,mitjançant la sobreexpressió d’enzims reguladorsclaus en el transport o en la fosforilació de la glucosa,podria reduir la hiperglucèmia diabètica. L’objectiud’aquesta part del nostre projecte va ser determinar siun increment en la fosforilació de la glucosa en el teixitadipós blanc (WAT) portava a un increment en laconcentració de glicerol 3-P i en la formació de trigli-cèrids. Això molt probablement permetria reduir lahiperglucèmia diabètica, però també podria portar adesenvolupar obesitat i resistència a la insulina. Ambaquesta finalitat vam obtenir ratolins transgènics quesobreexpressaven l'enzim glucoquinasa en el teixitadipós sota el control del promotor del gen aP2(adipocyte lipid-binding protein gene) (ratolins transgè-nics aP2/GK). Els ratolins transgènics presentaven unsnivells d’mRNA de GK en WAT i en el teixit adipósmarró interescapular (BAT) aproximadament tresvegades superior als nivells d’mRNA de GK de fetgede ratolins controls. La presència d’activitat GK feiaque la captació basal de glucosa pel teixit adipósestigués incrementada dues vegades en els ratolinstransgènics. No obstant això, els ratolins transgènicsno mostraven cap alteració en el pes corporal ni en eldipòsit de greix epididimal respecte dels ratolinscontrol. A més, els nivells sèrics de glucosa, àcidsgreixosos lliures (FFA) i triglicèrids no estaven afectats.Els ratolins transgènics aP2/GK mostraven unincrement en la tolerància a la glucosa i una majorsensibilitat corporal a la insulina. Aquests resultatssuggereixen que l’expressió de GK en teixit adipósporta a un increment en l’oxidació de la glucosa i en la
FUNDACIÓ LA MARATÓ DE TV3

despesa energètica. Per tant, la transferència de GKal teixit adipós mitjançant vectors virals pot ser útil,d’una banda, per fer disminuir la hiperglucèmiadiabètica, i d’una altra banda, per contrarestarresistència a la insulina associada a la diabetis tipus 2.
3. Estudi in vivo de la capacitat de regeneració delpàncrees endocrí en animals diabètics.3.1 Estudis en animals transgènics que sobreexpres-sen IGF-I en cèl·lules ß.
En primer lloc, hem demostrat en animals transgènicsque sobreexpressen el gen de l’IGF-I en cèl·lules ß,que la producció local d’IGF-I en el pàncrees permetrevertir el procés diabètic, ja que en aquests animalses produeix un increment en la replicació i en laneogènesi de noves cèl·lules ß (J. Clin. Invest. (2002)109(9), 1153-1163).
No obstant això, com a continuació d’aquest estudivolíem determinar també si l’IGF-I expressat encèl·lules ß era capaç de contrarestar la respostaautoimmune i així demostrar que podia realmentrevertir la diabetis tipus 1. Per aquest motiu ens caliaen primer lloc disposar d’un model de diabetis autoim-mune i després estudiar si l’IGF-I el podia "curar".
Així s’ha caracteritzat la resposta de ratolins transgè-nics que expressen IFN-ß en cèl·lules ß debackground genètic CD1 a dosis molt baixes d’STZ.Hem observat que aquests ratolins són molt sensiblesa molt baixes dosis d’STZ (15-20 mg/Kg pes),presenten una gran infiltració limfocitària dels illots queporta a la destrucció de les cèl·lules ß i a diabetisoberta. Aquestes dosis no causen diabetis a ratolinscontrol. Per tant, aquests ratolins transgènics consti-tueixen un bon model de diabetis autoimmune queserà molt útil per assajar noves teràpies per a aquestamalaltia. D’altra banda, a fi d’estudiar si la producciód’IGF-I per les cèl·lules ß pot contrarestar la diabetisautoimmune, també hem obtingut animals dobletransgènics mitjançant l’encreuament dels ratolins queexpressen IFN-ß en cèl·lules ß amb ratolins transgè-nics que sobreexpressen IGF-I en aquestes cèl·lules.Aquests animals mostren que l’IGF-I és capaç decontrarestar l’efecte de l’IFNß i no desenvolupendiabetis després del tractament amb STZ. L’IGF-Iprotegeix les cèl·lules ß de la mort per apoptosi itambé indueix la replicació de les cèl·lules ß, i porta ala normalització de la massa de cèl·lules ß. A més amés, aquests estudis suggereixen que l’expressiólocal d’IGF-I pot protegir la cèl·lula ß contra la respos-ta inflamatòria i prevenir la diabetis autoimmune.
3.2 Estudis en animals transgènics que sobrexpressenIGF-II en cèl·lules ß.
Els factors de creixement similars a la insulina de tipusI i II (IGF-I i IGF-II) són polipèptids amb un gran podermitogènic i efectes antiapoptòtics. Estudis in vitrodemostren que l’IGF-II indueix la proliferació i ladiferenciació de les cèl·lules ß i està involucrat en laregulació del creixement i la diferenciació dels illots.Per això, hem estudiat si l’IGF-II expressat en cèl·lulesß era capaç de contrarestar el procés diabètic. Noobstant això, contràriament als ratolins transgènics
que sobreexpressen IGF-I, els ratolins transgènics queexpressen IGF-II en les cèl·lules ß mostraven hiperglu-cèmia i diabetis oberta després del tractament ambSTZ. Aquests ratolins eren hipoinsulinèmics i desenvo-lupaven polidípsia i polifàgia. Dos mesos després deltractament amb STZ les anàlisis morfomètriques delspàncrees dels ratolins transgènics mostraven unamarcada reducció de la massa de cèl·lula ß. Al contra-ri que els ratolins transgènics IGF-I, els ratolinstransgènics IGF-II no recuperaven els paràmetresalterats i no sobrevivien més de 4 mesos després deltractament.
Aquests resultats indiquen que l’expressió local d’IGF-II en les cèl·lules ß dels ratolins transgènics no contra-resta la diabetis tipus 1. Això ens suggereix que IGF-I iIGF-II poden actuar per mitjà de diversos mecanismesen les cèl·lules ß-pancreàtiques in vivo.
3.3 Transferència de gens d’interès a pàncrees in vivo.
La preservació i/o regeneració de cèl·lules ß a partir deprecursors podria contrarestar la diabetis tipus 1. Noobstant això, la dificultat de manipular genèticament elpàncrees in vivo suposa una gran limitació. També,durant aquest període, hem analitzat la possibilitat detransferir gens d’interès a illots pancreàtics in vivo.
En ratolins, la injecció directa al pàncrees o la injeccióretrògrada pel conducte pancreaticobiliar dels vectorsadenovirals indueix inflamació i toxicitat severa. Unbypass del fetge mitjançant un clamp de la circulacióportal ha demostrat ser efectiu per augmentarl’expressió dels adenovirus en teixits extrahepàtics.Així, hem estudiat la capacitat de transduir les cèl·lulesß-pancreàtiques de ratolí mitjançant l’administraciósistèmica d’adenovirus. Per això, es va bloquejar lavena porta i l’artèria hepàtica i es van injectar adenovi-rus que expressaven ß-galactosidasa (ß-gal) sota elcontrol del promotor CMV per la vena jugular. Elclamp circulatori es va mantenir durant 30 minutsdesprés de la injecció dels adenovirus. Els ratolins vanser sacrificats a diferents temps (7, 14 i 30 dies) peranalitzar l’expressió del transgèn amb una reacció dex-gal o bé, amb immunohistoquímica antiinsulina ianti-ß-gal. Set dies després de la injecció delsadenovirus, el pàncrees exocrí presentava poquescèl·lules (<5%) amb activitat ß-galactosidasa, mentreque la majoria (aproximadament 80-90%) dels illotspancreàtics mostraven cèl·lules positives per ß-gal.Mitjançant una doble immunohistoquímica antiinsuli-na/anti-ß-gal es va confirmar que les cèl·lules ß sóninfectades eficientment pels vectors adenovirals iexpressen els gens exògens d’interès. Aquestaaproximació pot ser útil per assajar noves aproxima-cions de teràpia gènica per a la diabetis mellituscentrades en la transferència de gens que indueixin lareplicació de les cèl·lules ß-pancreàtiques in vivo(Hum. Gene Ther., enviada versiò revisada).
També hem aconseguit transferir específicament elgen de la ß-galactosidasa, mitjançant vectors adenoví-rics, en pàncrees de gos sotmès a un clamp local dela circulació pancreàtica. Després de trencar elsvasos, els vectors virals s’injectaven per la vena o béper l’artèria pancreaticoduodenal i el clamp es
RECERCA BIOMÈDICA | 91

mantenia 10 min. Així, hem observat que cèl·lulespancreàtiques acinars, ductals i endocrines erentransduïdes en el pàncrees d’aquests gossos.Aquesta aproximació permetrà expressar gens d’inte-rès per regenerar el pàncrees endocrí en gossosdiabètics. Això ha estat possible mercès a lacol·laboració que vam establir amb l’Hospital ClínicVeterinari i amb el Departament de Cirurgia de lanostra Facultat.
4. Desenvolupament d’un model animal de compli-cacions oculars de la diabetis.El factor de creixement IGF-I s’ha associat amb lapatogènesi de la retinopatia diabètica, si bé encara noés del tot conegut. Al nostre laboratori vam observarque ratolins transgènics normoglicèmics/normoinsuli-nèmics que sobrexpressen IGF-I en la retina desenvo-lupaven la majoria de les alteracions que presenten elspacients diabètics amb patologia ocular. Un efecteparacrí de l’IGF-I en la retina iniciaria les alteracionsvasculars que progressaven de retinopatia no prolife-rativa a proliferativa i a despreniment de retina. Els ullsde ratolins transgènics de dos mesos d’edat mostra-ven la pèrdua de perícits i l’engruiximent de lamembrana basal dels capil·lars retinians. Animals desis mesos d’edat i més vells presentaven dilatació deles vènules, anomalies microvasculars intraretinals ineovascularització de la retina i de la cavitat del vitri.La neovascularització estava d’acord amb la induccióper IGF-I de l’expressió de VEGF (factor de creixe-ment de l’endoteli dels vasos) en les cèl·lules glials dela retina. A més, vam observar que l’IGF-I s’acumula-va en l’humor aquós, la qual cosa causava rubeosisde l’iris i adhesions entre la còrnia i l’iris que impedienel drenatge de l’humor aquós i portaven al desenvolu-pament de glaucoma. Igualment, tots aquests animalsdesenvolupaven cataractes. Tots aquests resultatssuggereixen que l’IGF-I té un paper important en eldesenvolupament de les complicacions oculars de ladiabetis. Per tant, aquests ratolins transgènics podenser utilitzats per estudiar els mecanismes que porten ala malaltia ocular diabètica i constitueixen un modelmolt adequat per assajar noves teràpies (J. Clin.Invest. (2004) 113:1149-1157).
3. Rellevància i possibles implicacions clíniquesdels resultats finals obtingutsEl nombre de pacients diabètics s’està incrementantmoltíssim a les societats amb estils de vida occidenta-litzats, i al nostre país un 10% de la població pateix lamalaltia. Si bé conèixer les causes desencadenants ésfonamental per intentar-ne la prevenció (aquestamalaltia és el resultat d’una interacció complexa entrefactors genètics i factors ambientals), desenvoluparnoves aproximacions terapèutiques més efectives pera la diabetis és clau a fi de poder reduir les greuscomplicacions secundàries de la malaltia (retinopatia,fallada renal, neuropaties, malalties cardiovasculars,etc.) i millorar la qualitat de vida dels pacients. A més,en tractar-se d’una malaltia crònica, els recursos quees destinen a la cura d’aquests malalts són moltelevats i representen una partida important delspressupostos sanitaris dels països. Els resultatsd’aquest projecte representen un pas més cap a unateràpia més efectiva per a la diabetis mellitus, tant detipus 1 com de tipus 2.
En el desenvolupament del projecte hem aconseguitels objectius que ens havíem plantejat. Així, d’unabanda, hem assajat una nova aproximació de teràpiagènica centrada en la transferència conjunta del gende la insulina i la glucoquinasa que s’ha demostratcom un sistema altament eficaç per regular l’homeòs-tasi de la glucosa en tres aproximacions diferents. Ésa dir, tant en animals doble transgènics que expresseninsulina i glucoquinasa, com en animals diabèticselectrotransferits amb aquests gens o bé que se’ls haadministrat vectors adenoassociats (AAV1GK + AAV1insulina) s’ha observat la normalització de la glicèmia idels altres paràmetres metabòlics alterats.L’aproximació de teràpia gènica no viral centrada enl’electrotransferència del gen de la insulina i el de laglucoquinasa a múscul esquelètic permet normalitzartots els paràmetres metabòlics alterats en models dediabetis. Aquesta aproximació és molt eficaç,fàcilment administrable i, a més, es pot regular la "doside gen" segons les necessitats de l’individu. Noobstant això, l’aproximació de teràpia gènica viral ambvectors AAV ens permet una correcció a llarg terminide les alteracions diabètiques. Com que la diabetis ésuna malaltia crònica, aquesta aproximació permetriadisminuir el nombre d’administracions dels vectors total llarg de la vida del pacient. Actualment el nostregrup ha iniciat un nou projecte, com a continuaciód’aquest projecte finançat per La Marató de TV3, quese centra en l’assaig d’aquestes aproximacions, quehan funcionat en ratolins, en animals més grans, comara gossos, per poder plantejar l’aplicació en pacientdiabètics en un futur proper.
D’altra banda, en aquest estudi també hem demostraten animals transgènics diabètics que l’IGF-I expressaten cèl·lules ß pot contrarestar la infiltració limfocitàriadels illots i la mort cel·lular per apoptosi, i pot induir lareplicació i la formació de noves cèl·lules ß, tot portanta la regeneració del pàncrees endocrí. Això indica quela transferència del gen de l’IGF-I al pàncrees in vivodurant la diabetis podria resultar en una teràpia curati-va per a aquesta malaltia. En aquest sentit, tambéhem aconseguit posar a punt la tecnologia pertransferir gens d’interès mitjançant vectors adenovíricsa pàncrees de ratolí i de gos in vivo. Actualmentestem administrant vectors virals que contenen el gende l’IGF-I a animals diabètics per comprovar siaquesta aproximació permet contrarestar la mort deles cèl·lules ß i regenerar el pàncrees durant ladiabetis.
4. PublicacionsM.George, E. Ayuso, A. Casellas, C. Costa, J.C. Devedjian and
F. Bosch. ß-Cell expression of insulin-like growth factor-I leads to
recovery from type 1 diabetes. J. Clin. Invest. (2002)109(9), 1153-
1163. Clau: A
P. Otaegui, M. Ontiveros, T. Ferre, E. Riu, R. Jimenez & F.
Bosch. Glucose-regulated glucose uptake by transplanted muscle
cells expressing glucokinase counteracts diabetic hyperglycemia.
Hum. Gene Ther. (2002) 13, 2125-2133. Clau: A
J. Ruberte, E. Ayuso, M. Navarro, A. Carretero, V. Nacher, V.
Haurigot, M. George, C. Llombart, A. Casellas, C. Costa. A.
Bosch & F. Bosch. Increased ocular levels of insulin-like growth
factor-I lead to diabetic eye disease. J. Clin. Invest. (2004) 113 (8),
149-1167. Clau: A
FUNDACIÓ LA MARATÓ DE TV3

A. Mas, J.Montane, M.Chillon, E. Riu, P. Otaegui & F. Bosch.
Increased insulin sensitivity by glucokinase expression in skeletal
muscle of diabetic mice. Gene Therapy (remesa versió revisada).
Clau: A
E. Ayuso, M.Chillon, J. Agudo, V. Haurigot, A. Bosch, A.
Carretero, P. Otaegui & F. Bosch. In vivo gene transfer to pancrea-
tic ß-cells by systemic delivery of adenoviral vectors. Hum. Gene
Ther. (remesa versió revisada). Clau: A
A. Mas, J.Montane, M. Chillón, X. Martínez, S. Muñoz, AM
Douar, O. Danos, E. Riu, P. Otaegui, & F. Bosch. Insulin and
glucokinase expression in skeletal muscle reverts type 1 diabetes.
Clau: A
A. Mas, J.Montane, P. Otaegui, E. Riu, M.Chillon & F. Bosch
Basal expression of insulin in skeletal muscle together with low
doses of soluble exogenous insulin maintains normoglycemia in
diabetic animals. Clau: A
A. Casellas, A. Salavert, E. Ayuso, M. Moya, M. Garcia, J.
Agudo & F. Bosch. Insulin-like growth factor-I expression in ß-cells
counteracts autoimmune diabetes. Clau: A
RECERCA BIOMÈDICA | 93

FUNDACIÓ LA MARATÓ DE TV3
Títol del projecte
Estudi del sistema antioxidant i de l'estrès oxidatiu en pacients fenilcetonúrics
Investigador principalJaume Campistol Plana
InstitucióHospital Universitari Sant Joan de Déu
74.586 €
MEMÒRIA FINAL
1. Resum del projecte originalObjectiu principalAvaluació del sistema antioxidant i de l’estrès oxidatiuen pacients fenilcetonúrics, estudiant els enzims isubstrats antioxidants i els productes resultants deldany oxidatiu en biomolècules, i comparant elsresultats obtinguts en els pacients amb hiperfenilalani-nèmia benigna en tractament i en controls sans.
Objectius específicsEstudi del possible augment de formació de radicalslliures en pacients fenilcetonúrics per mitjà de ladeterminació de les concentracions de marcadorsd’estrès oxidatiu (malondialdehid, m-tirosina i o-tirosi-na), comparant-los amb un grup de pacients ambhiperfenilalaninèmia benigna sense tractament i ambun grup control.
Valoració del sistema antioxidant per mitjà de l’anàliside les activitats dels enzims antioxidants (superòxiddismutasa, catalasa, glutatió peroxidasa, glutatióreductasa) i de les concentracions sèriques delssubstrats antioxidants (seleni, glutatió, tocoferol,
retinol, urat, hipoxantina, xantina, ascorbat, ubiquino-na i ubiquinol).
Relació dels resultats obtinguts en els dos puntsanteriors amb el control nutricional dels pacients.
Avaluació neuropsicològica dels pacients per mitjàd’examen clínic i proves complementàries i la sevarelació amb els resultats obtinguts en els dos primersobjectius.
MètodesEstudi de recerca no experimental transversal analíticen 60 pacients PKU, 40 HPA comparats amb 60controls sans de la mateixa edat i sexe. Avaluacióneuropsicològica i neurofisiològica completa.Avaluació nutricional. Control bioquímic periòdic:fenilalanina, aminoàcids. Estudi de l’estrès oxidatiu:enzims antioxidants (superòxid dismutasa, catalasa,glutatió peroxidasa, glutatió reductasa), substratsantioxidants (seleni, glutatió, tocoferol, retinol,ascorbat, urat, xantina, hipoxantina, ubiquinona iubiquinol), marcadors d’estrès oxidatiu (malondialde-hid, m-tirosina i o-tirosina).

2. ResultatsHem pogut constatar (vegeu publicacions adjuntes) undescens de les concentracions d’ubiquinona-10, quecausaven un augment de l’estrès oxidatiu en pacientsPKU amb tractament dietètic. A més a més, hemdemostrat la possible implicació d’altres componentsdel sistema antioxidant en relació amb el controlmetabòlic i nutricional dels pacients, com els enzimsantioxidants catalasa i glutatió peroxidasa. Tots aquestresultats estan resumits en la darrera publicació(pendent encara d’acceptació definitiva), en la quals’avaluen tots els resultats bioquímics i nutricionalsdurant els tres anys de durada de l’estudi.
Respecte dels estudis neuropsicològics, hem observatuna alteració de les funcions neurocognitives delspacients PKU en relació amb els hiperfenilalaninèmicsi especialment en el grup control. Tot i que el pacientPKU té un nivell intel·lectual gairebé normal, aquest és10 punts inferior al del grup control. Hem constatat, amés a més, puntuacions més baixes en funcionsvisuoespacials, motricitat fina, atenció i dificultats perinhibir la resposta. Al mateix temps, els pacients PKUtenen una major lentitud en el processament de lainformació que el grup control i els hiperfenilalaninè-mics. Estudis tan complets i comparant el tres grupsgairebé no s’havien publicat en la literatura internacio-nal.
3. Rellevància i possibles implicacions clíniquesdels resultats finals obtingutsEls nostres resultats refermen la necessitat de perllon-gar el tractament dietètic durant tota la vida enpacients fenilcetonúrics. Les concentracions elevadesde fenilalanina poden causar, a part d’alteracionsneurològiques i neuropsicològiques ben establertes,descompensacions dels sistemes de defensa antioxi-dant que, de manera mantinguda, podrien causaralteracions en teixits molt sensibles, com el sistemanerviós central. D’altra banda, el tractament dietèticcrònic pot ser responsable de deficiències en algunsantioxidants essencials, per la qual cosa la recomana-ció principal que es derivaria d’aquest estudi seria lanecessitat d’un control metabòlic i nutricionalbo/regular en pacients PKU per detectar i corregiraquestes deficiències. Un altre punt d’interès a partirdels nostres resultats és el fet d’aprofundir la valoracióneurocognitiva dels pacients amb hiperfenilalaninèmia,que tot i tenir un nivell intel·lectual normal, més alt queels PKU, tenen problemes neurocognitius menors,però molt superiors a la població general. L’estudid’aquest grup poblacional i les possibles implicacionsde la fenilalanina, dels neurotransmissors o delssistemes antioxidants queda pendent de noves investi-gacions en un futur molt proper i és un repte per a totsels professionals que treballem en aquest camp.
Per últim, les repercussions clíniques de la deficiènciad’antioxidants en la fenilcetonúria són més difícilsd’establir. Per una banda, en altres estudis s’hademostrat que un dany oxidatiu crònic estaria relacio-nat amb una major predisposició a desenvoluparcertes malalties cròniques i processos degeneratius. IPer altra banda, el sistema nerviós és un dels teixitsmés sensibles a l’estrès oxidatiu, per la qual cosa elsefectes adversos dels radicals lliures se sumarien als
propis de la malaltia (dany tòxic directe per la fenilala-nina i altres efectes relacionats). Un dels nostrespropòsits, a partir de les troballes d’aquest projecte,és relacionar algunes anomalies neurològiquespresents en pacients PKU, tot i mostrar un boncontrol metabòlic i dietètic, amb les alteracions jadocumentades en relació amb l’augment de l’estrèsoxidatiu.
4. PublicacionsArtuch R, Colomé C, Sierra C, Brandi N, Lambruschini N,
Campistol J, Ugarte D, Vilaseca MA. A longitudinal study of
antioxidant status in phenylketonuric patients. Clin Biochem. 2004
Mar;37(3):198-203.
Gassio R, Campistol J, Vilaseca MA, Lambruschini N, Cambra
FJ, Fuste E. Do adult patients with phenylketonuria improve their
quality of life after introduction/resumption of a phenylalanine-
restricted diet?. Acta Paediatr. 2003 Dec;92(12):1474-8.
Colomé C, Artuch R, Sierra C, Brandi N, Lambruschini N,
Campistol J, Vilaseca. MA. Plasma thiols and their determinants in
phenylketonuria. Eur J Clin Nutr. 2003 Aug;57(8):964-8.
Colomé C, Artuch R, Vilaseca MA, Sierra C, Brandi N,
Lambruschini N, Cambra. FJ, Campistol J. Lipophilic antioxidants
in patients with phenylketonuria. Am J Clin Nutr. 2003
Jan;77(1):185-8.
Pérez-Dueñas B, Cambra FJ, Vilaseca MA, Lambruschini N,
Campistol J, Camacho JA. New approach to osteopenia in
phenylketonuric patients. Acta Paediatr. 2002;91(8):899-904.
Colomé C, Artuch R, Vilaseca MA, Sierra C, Brandi N, Cambra
FJ, Lambruschini N, Campistol J. Ubiquinone-10 content in
lymphocytes of phenylketonuric patients. Clin Biochem. 2002
Feb;35(1):81-4.
Colomé C, Artuch R, Lambruschini N, Cambra FJ, Campistol
J, Vilaseca M. Is there a relationship between plasma phenylalani-
ne and cholesterol in phenylketonuric patients under dietary
treatment?. Clin Biochem. 2001 Jul;34(5):373-6.
Artuch R, Colomé C, Vilaseca MA, Sierra C, Cambra FJ,
Lambruschini N, Campistol J. Plasma phenylalanine is associated
with decreased serum ubiquinone-10 concentrations in phenylke-
tonuria.J Inherit Metab Dis. 2001 Jun;24(3):359-66.
Publicacions en procés de revisió
Rosa Gassió, Anna Sans, Rafael Artuch, Maria Antonia
Vilaseca, Jaume Campistol. Cognitive functions in classic phenyl-
ketonuria and mild hyperphenylalaninemia: Experience in a
pediatric population in Catalonia.
Gassio R, Fusté E, López-Sala A, Artuch R, Vilaseca MA,
Campistol J. School performance in PKU patients with early and
continuous treatment.
RECERCA BIOMÈDICA | 95

FUNDACIÓ LA MARATÓ DE TV3
MEMÒRIA FINAL
1. Resum del projecte originalLa nefropatia diabètica (ND) és una greu complicaciómicroangiopàtica de la diabetis mellitus. Aquestamalaltia té una etiopatogènia multifactorial, que incloua més de factors de risc clàssics com la hiperglicèmiao la hipertensió, d’altres com el tabaquisme, al qualcada vegada es dóna més importància.
HipòtesiLa nostra hipòtesi és que en l’etiopatogènia de la NDpoden estar implicats l’estrès oxidatiu i la fibrogènesi, ique aquests mecanismes de lesió poden estarinfluenciats pel control metabòlic i pel consum detabac. El nostre estudi el vam plantejar amb un modelin vitro de cultius cel·lulars de rata sotmesos adiverses condicions experimentals, tot valorant laproducció d’estrès oxidatiu i la fibrogènesi, per mitjàde la secreció d’isoprostans i de TGFß 1, respectiva-ment, en els sobrenedants de les cèl·lules.
El model experimental d’estudi va consistir en cultiusde cèl·lules mesangials de rata en cultiu primari quees van tractar amb extractes de fum de tabac. A més,
diverses concentracions de glucosa es van assajarcom a possible factor inductor de la secreció de TGFß1 en aquestes cèl·lules. El peròxid d’hidrogen tambés’hi va afegir com a control positiu.
Els objectius definits en aquest treball van serEstudi de la producció de TGFß 1 en un model in vitrode cèl·lules mesangials de rata, sotmeses a condicionsexperimentals diferents, és a dir, diverses concentra-cions d’extractes de fum de tabac, i diverses concen-tracions de glucosa en el medi de cultiu.
Estudi de la peroxidació lipídica en el mateix model decèl·lules sotmeses a les mateixes condicions experi-mentals i mesurada com a excreció de 8-epi-PGF2aen el medi de cultiu.
El pla de treball original era de dos anys de duradaDurant el primer any es van posar a punt lestècniques d´aïllament i cultiu de cèl·lules mesangialsde rata i l’extracció del fum de tabac. Es van fer uns20 aïllaments de 4 rates cadascun, que ens vanpermetre realitzar tots els experiments amb cèl·lulesentre els passis 3 i 6.
Títol del projecte
Valoració de l'estrès oxidatiu i la fibrogènesi en la patogènia de la nefropatia diabètica. Influència del control glicèmic
i del consum de tabac
Investigador principalEnric Esmatjes Mompó
InstitucióHospital Clínic i Provincial de Barcelona
Membres de l’equipJ. Clària, J.M. Campistol, P. Iñigo, A. Cases, S. Lario, C. Mur i J. Abian.
68.302 €

Durant el segon any es van fer els experiments i lesdeterminacions de TGFß 1 i de 8-epi-PGF2a en elssobrenedants dels cultius cel·lulars.
Durant els sis mesos addicionals de projecte, s’han fetnous experiments amb un inhibidor de la PKC i ambun antagonista de l’angiotensina II i s’ha valorat de lamateixa manera la producció de TGFß 1 i de 8-epi-PGF2a en els sobrenedants dels cultius cel·lulars, perveure com pot revertir el procés i quines vies patogè-niques hi estan implicades.
2. ResultatsEn una primera etapa, es va posar a punt la tècnicad’aïllament de cèl·lules mesangials, l’obtenció delsextractes de fum de tabac (EFT) i la determinació deTGFß 1 en sobrenedants de cèl·lules tractades. Esvan detectar nivells de TGFß 1 dins els límits dedetecció de la tècnica, i es va veure que no hi haviadiferències entre l’efecte dels extractes frescos i elscongelats (figura núm. 1). Es va fer l’estandaritzaciódels extractes per absorbància a 390 nm, el resultatsde la qual s'expressen en la figura núm. 2. Això ensva permetre veure que els diversos extractes no erenqualitativament diferents.
Vam fer uns 20 aïllaments a partir de 4 rates Sprague-Dawley cadascun, la qual cosa ens va permetre fertots els experiments entre el passi 3 i el passi 6.
Es van fer 7 extraccions de fum de tabac. Vammantenir els extractes concentrats i congelats a —80ºC, fins al moment de l’aplicació al medi de cultiu.
Un cop resolts els problemes de l’obtenció delscultius i de l’obtenció dels extractes, ens vam centraren la determinació de TGFß 1 i de 8-epiPGF2a . Per ala determinació de TGFß 1 es va utilitzar un kitd’ELISA de R& D, amb un límit de detecció de 7pg/ml. Per a la determinació de la 8-epiPGF2a , en unprincipi es va utilitzar un kit d’ELISA de Cayman, peròarran de la mala reproductibilitat del mètode vam optarper posar a punt una tècnica més complexa, quecombina un primer pas de cromatografia d’afinitat, i totseguit vam fer HPLC i espectrometria de masses, talcom havia estat descrit per altres autors abans.
En cada experiment es va determinar el nombre finalde cèl·lules després de cada tractament, i es va veureque no era constant. Podia haver-hi mort cel·lular acausa del tractament. Per aquest motiu, es vanexpressar tots els resultats referits a nombre decèl·lules finals (figura 2 i taula 1).
El mètode que vam utilitzar va ser un mètode colori-mètric senzill, d’exclusió de Trypan Blue, que correla-ciona el nombre de cèl·lules amb l’absorbància a 570nm (taula núm.1).
Respecte del nombre de cèl·lules, es va observar unaugment en els tractaments basal (glucosa 5 mM) iamb glucosa alta (glucosa 25 mM), una reducció en elcontrol positiu H2O2 150m M i no va haver-hi capvariació en els tractaments amb les diverses concen-tracions de tabac.
Els nivells de 8-epi-PGF2a i de TGFß 1 en condicionsbasals van ser 19 ± 17 pg / 106 cèl·lules i 11 ± 6,7ng/ 106 cèl·lules, respectivament. En afegir els extrac-tes de fum de tabac al medi de cultiu, es va observarun augment en els nivells de 8-epi-PGF2a que va sermàxim amb una concentració d’EFT de 0.1 m g/mL(27 ± 20pg / 106 cèl·lules), la qual cosa representa un156% d’augment respecte del nivell basal.
La incubació amb EFT va induir un augment en elsnivells de TGFß 1 en el medi de cultiu, que va sersignificatiu amb una concentració d’EFT d’1 m g/mL(82% d’augment, P = a .0116) i 10 m g/mL(67%d’augment, P = 0.054).
També va ser significatiu l’augment en els nivells deTGFß 1 en els dos controls positius, 25 mM deglucosa (P = 0.028) i H2O2 150 m M (P = 0.0006).
Així, després d’analitzar tots els resultats, s’haobtingut la figura núm. 3 i la figura núm. 4.
De l’última part del projecte, hem obtingut els resultatsde la figura 6, en la qual veiem que tant el Losartan, unantagonista dels receptors tipus 1 de l’angiotensina II,com un inhibidor específic de la PKC, disminueixen laproducció de TGFß 1 en aquestes condicions d’estrèsoxidatiu causat pel tabac. Això demostra la implicacióde dues vies patogèniques conegudes en aquestmodel de nefropatia diabètica in vitro (figura núm. 5).
Taula 1. Nombre de cèl·lules amb els diversos tractaments.
EFT: Extractes de fum de tabac.
Figura 1. Efecte dels EFT fresc i congelat sobre l’expressió de
TGFb 1 en cultius de cèl·lules mesangials de rata.
Figura 2. Absorbància dels diversos extractes de fum de tabac
(EFT) respecte de la seva concentració.
RECERCA BIOMÈDICA | 97

Figura 3. Alliberament de 8-epi PGF2a al medi de cultiu després
del tractament durant 24 hores amb extractes de fum de tabac
de diferents concentracions.
Figura 4. Alliberament de TGFb 1 al medi de cultiu després del
tractament durant 24 hores amb extractes de fum de tabac de
diferents concentracions.
Figura 5. Efecte dels EFT després de la incubació amb un inhibi-
dor de la PKC i amb un antagonista de l’angiotensinaII, Losartan.
3. Rellevància i possibles implicacions clíniquesdels resultats finals obtingutsEn aquest estudi es confirma l'efecte tòxic que té eltabac sobre les cèl·lules mesangials, fet que dónasuport a la hipòtesi que el tabac és un factor clar de riscper al desenvolupament de la nefropatia diabètica. Lesdades obtingudes suggereixen clarament que un delspossibles mecanismes patogènics del tabac és l'incre-ment en la síntensi de TGF-, molt probablement induïdaper l’estrès oxidatiu que genera el tabac. En conjunt,aquests experiments han de servir per reforçar laconducta dels clínics a l'hora de recomanar als malaltsamb diabetis l'abandonament del tabaquisme, unapossible via patogènica per a la nefropatia diabètica.
4. PublicacionsC. Mur, J. Clària, S. Rodela, S. Lario, J.M. Campistol, E. Titos ,
P. Iñigo, A. Cases, J. Abián, E. Esmatjes. Cigarette smoke
concentrate increases 8-epiPGF2aand TGFß 1 secretion in rat
mesangial cells. Life Science 2004 (en premsa).
FUNDACIÓ LA MARATÓ DE TV3

RECERCA BIOMÈDICA | 99
MEMÒRIA FINAL
1. Resum del projecte originalMutacions al DNA mitocondrial són responsables desordesa i/o diabetis. La sordesa és un defecteneurosensorial molt freqüent, ja que afecta el 10% dela població. Es valora que un 80% dels casos desordesa són de causa genètica, i entre aquests s’handescrit diverses mutacions al DNA mitocondrialassociades a sordesa. La mutació mitocondrial méscomuna responsable de sordesa neurosensorialprogressiva no sindròmica és la A1555G al gen del12S rRNA. Aquesta mutació també es relaciona ambuna major susceptibilitat en l’administració d’antibiò-tics aminoglicòsids, l’ototoxicitat dels quals és àmplia-ment coneguda. Altres mutacions mitocondrialss’associen a més a més de sordesa, a diabetis, comés el cas de la mutació A3243G en el gen deltRNALeu(UUR), la qual també s’ha associat al síndro-me de MELAS.
La patofisiologia i l’expressió fenotípica de lesmutacions mitocondrials són poc conegudes. No hiha una explicació clara de per què una mutació, tot iestar present en tots els teixits, causa alteracions teixitespecífiques ni per què es troba associada a trets
clínics, bioquímics i moleculars tan variables. Lesalteracions bioquímiques, funcionals i cel·lularscausades per aquestes mutacions poden estudiar-sedes d’un enfocament multidisciplinari, en modelscel·lulars i en models murins. A més a més, eldesenvolupament de models cel·lulars i murinspermetrà l’assaig de mesures farmacològiques deprevenció i teràpia d’aquestes patologies.
Objectius1) Tipificar les alteracions clíniques i bioquímiquespresents en pacients afectats d’hipoacúsia progressi-va i/o diabetis a causa de les mutacions A1555G iA3243G en el DNA mitocondrial.
2) Determinar les possibles alteracions clíniques ibioquímiques en individus asimptomàtics portadors dela mutació A1555G.
3) Determinar la presència d’altres mutacions en elDNA mitocondrial en pacients amb sordesa i diabetis,negatius per a les mutacions esmentades.
4) Determinar els haplotips del DNA mitocondrialassociats a la mutació A1555G.
Títol del projecte
Anàlisi molecular i funcional de mutacions al DNA mitocondrial implicades en sordesa progressiva i diabetis
Investigadors principalsXavier Estivill Pallejà
InstitucióCentre de Genètica Mèdica i Molecular. IRO
91.214 €

5) Desenvolupar i caracteritzar el metabolisme respira-tori de models cel·lulars portadors de la mutacióA1555G.
6) Desenvolupar un model murí de la mutaciómitocondrial A1555G per estudiar la fisiopatologiad’aquesta mutació en la sordesa.
7) Identificació dels factors genètics nuclears modifica-dors dels efectes de la mutació mitocondrial.
Disseny1) Tipificar les alteracions clíniques i bioquímiquespresents en pacients afectats d’hipoacúsia progressi-va i/o diabetis a causa de les mutacions A1555G iA3243G en el DNA mitocondrial.
Es tracta de detectar portadors de les mutacionsmitocondrials A3243G i A1555G en pacients afectatsd’hipoacúsia neurosensorial amb afectació predomi-nant de freqüències agudes i en pacients diabètics, oamb antecedents familiars materns de diabetis i/ohipoacúsia neurosensorial, com també en pacientsafectats d’hipoacúsia neurosensorial i MELAS. Lesmutacions A1555G i A3243G s’estudiaran mitjançantamplificació per PCR i anàlisi amb enzims de restricció.
Es realitzarà un estudi clínic i audiomètric complet,consistent en audiometria completa, timpanograma,reflexos acústics, productes de distorsió d’otoemis-sions acústiques i PEATC en els casos índexs iaudiometria tonal, productes de distorsió d’otoemis-sions acústiques i audiometria d’alta freqüència en elsfamiliars de primer grau. Així es vol determinar el patróaudiomètric dels pacients i buscar una manifestacióque pugui ser especifíca d’aquest trastorn, i també esvol verificar la localització de l’alteració a la còclea.
2) Determinar les possibles alteracions clíniques ibioquímiques en individus asimptomàtics portadors dela mutació A1555G.
Per a la sordesa causada per la mutació A1555G,s’han identificat 30 famílies, de les quals s’ha estès lagenealogia i identificat més de 250 individus amb lamutació però sense dèficit auditiu aparent (lapenetrància de la mutació és aproximadament del50%). Una mostra d’aquests individus sense afectacióauditiva aparent i dels pacients amb alteracions auditi-ves s’avaluen des del punt de vista clínic.
3) Determinar la presència d’altres mutacions en elDNA mitocondrial en pacients amb sordesa i diabetis,negatius per a les mutacions mencionades.
S’estudiarà el DNA mitocondrial de les mostrescorresponents a pacients amb sordesa i diabetis quesiguin negatius per a les mutacions A1555G iA3243G. Els estudis se centraran en la detecció dedelecions, mutacions específiques en gens potencial-ment implicats en la patologia i una anàlisi global delDNA mitocondrial utilitzant microxips.
4) Determinar els haplotips del DNA mitocondrialassociats a la mutació A1555G.
La caracterització haplotípica i d’haplogrups permetdefinir més de 250 haplogrups en la població europeaen els quals poder situar qualsevol mostra de DNAmitocondrial. Aquesta anàlisi es realitza mitjançantrestricció enzimàtica de diversos llocs polimòrfics iseqüenciació.
5) Desenvolupar i caracteritzar el metabolisme respira-tori de models cel·lulars portadors de la mutacióA1555G.
Es determinarà la concentració de proteïna mitocon-drial i es realitzaran proves de funcionalitat mitocon-drial a partir de mostres de sang de pacients amb lamutació i sense la mutació. Els estudis que es realit-zaran seran l’estudi de la capacitat oxidativa mitocon-drial i l’activitat enzimàtica individual de cadascun delscomplexos de la cadena respiratòria mitocondrial.
6) Desenvolupar un model murí de la mutaciómitocondrial A1555G per estudiar la fisiopatologiad’aquesta mutació en la sordesa.
La microinjecció de cèl·lules embrionàries (ES) deratolí, les quals poden ser modificades en l’estadi deblastocist, permet generar ratolins amb una mutacióespecífica en les cèl·lules germinals que es transmet ala descendència. De manera anàloga, les cèl·lules ESpoden utilitzar-se per generar ratolins amb mutacionsal DNA mitocondrial.
Per generar línies de cèl·lules ES amb la mutacióA1555G, s’obtindran cèl·lules híbrides (cíbrids) perfusió cel·lular, tot utilitzant com a donadors demitocondris cèl·lules de ratolí amb la mutacióA1555G. Es microinjectaran embrions de ratolíC57BL/6 de 3,5 dies (blàstula) amb cèl·lules ES ambla mutació A1555G. L’efecte de la mutació en elsratolins s’avaluarà mitjançant la realització d’estudisneurosensorials, electrofisiològics i bioquímics enteixits diferents.
7) Identificació dels factors genètics nuclears modifica-dors dels efectes de la mutació mitocondrial.
L’expressió clínica de la mutació A1555G es pot veureinfluenciada per múltiples factors, com són l’haplotipmitocondrial, factors ambientals, el genoma nuclear iefectes específics de teixit. Les dades epidemiològi-ques, bioquímiques i genètiques disponibles indiquenque els gens nuclears serien els principals determi-nants de les diferències fenotípiques observades.
Per identificar els loci cromosòmics de possibles gensnuclears modificadors implicats en sordesa a causade la mutació mitocondrial A1555G, es farà una anàlisiglobal del genoma en famílies espanyoles.S’estudiaran 8 famílies espanyoles, les quals presen-ten diversos afectats de sordesa neurosensorial,juntament amb altres famílies d’origen italià i arabois-raelià. En 3 famílies, diversos individus han estatexposats a aminoglicòsids, mentre que en les altres 5no hi ha antecedents d’exposició a aquests antibiò-tics. Es farà una anàlisi global del genoma amb els331 marcadors de l’ABI PRISMTM Linkage MappingSet (Applied Biosystems, Foster City, CA), els quals
FUNDACIÓ LA MARATÓ DE TV3

abasten tot el genoma. Els marcadors s’amplificaranmitjançant PCR i s’analitzaran en un seqüenciadorautomàtic. La genotipació es farà amb el programaGENOTYPERTM (Applied Biosystems).
Es van fer diversos tipus d’aproximacions estadístiques:
Anàlisis de lligament de dos punts.El Lod score entre el fenotip i els marcadors genèticses calcularan amb el programa MLINK de LINKAGE(Terwillinger and Ott, 1994) per a fraccions derecombinació iguals a 0.01, 0.05, 0.10, 0.15, 0.20,0.25, 0.30, 0.40.
Anàlisi de lligament de múltiples punts.L’anàlisi de lligament multipoint es farà amb el progra-ma GENEHUNTER.
Anàlisi de lligament amb parelles de germans afectats.Es faran amb el programa MAPMAKER/SIBS. Enaquest estudi, s’hi inclouran 12 parelles de germansafectats corresponents a famílies espanyoles, 7 parellesde famílies italianes i 65 del pedigrí araboisraelià.
Estudis paramètrics de lligament
L’anàlisi es farà utilitzant patrons d’herència recessius idominants amb penetrància reduïda. En el patród’herència autosòmic dominant la freqüència de l’al·lelmalalt es va considerar entre 0,1 i 0,4 amb unapenetrància de 0,8. Per al model recessiu, la freqüèn-cia de l’al·lel malalt es considerarà de 0,5 i lapenetrància de 0,8.
Estudis no paramètrics de lligament
Per identificar regions cromosòmiques potencialmentassociades a sordesa sense assumir cap modeld’herència, es farà un estudi de lligament genètic noparamètric.
Objectius específics assolits1) Els pacients i individus amb la mutació A1555Gavaluats no han mostrat cap alteració clínica o bioquí-mica addicional a la hipoacúsia. L’afectació auditiva ésde tipus endococlear i bilateral, s’acompanyad’acúfens en més de la meitat dels casos i l’afectacióés més severa per a freqüències agudes. La mutacióA1555G produeix alteracions en la fisiologia coclear atots els seus portadors, malgrat que amb diversosgraus d’afectació, fet que determina l’existència demanifestacions clíniques en funció de la seva severitat.
2) S’han fet estudis de funcionalitat mitocondrial enlimfòcits de pacients portadors de la mutacióA1555G, però no s’ha observat cap alteració respectedels controls.
3) No s’ha detectat cap cas portador de la mutacióA3243G.
4) S’han identificat dues mutacions noves al DNAmitocondrial associades a sordesa d’herència maternai se n’ha determinat l’haplotip associat.
5) S’han identificat diverses localitzacions nuclears per
a possibles gens modificadors de la mutació A1555G.
6) Una de les regions modificadores més rellevantpodria estar localitzada al cromosoma 8, tal com esva identificar en els estudis de lligament (LOD score4.1). L’estudi de l’estructura genòmica de 8p23 harevelat la possible implicació d’aquesta, en la manifes-tació fenotípica de la mutació. Aquesta regió estàflanquejada per duplicons que impliquen l’existènciad’una inversió de 4.7 Mb present en el 25% d’indivi-dus controls. Això seria un factor de confusió enestudis de lligament. També s’han localitzat clústersde defensines, polimòrfics en número de còpies, lesquals també podrien tenir un paper en la manifestaciófenotípica de la mutació.
7) S’ha iniciat l’estudi i la identificació d’altres regionscom a possibles localitzacions de gens modificadors.
8) S’ha obtingut un ajut del NIH en col·laboració ambel Dr. Nathan Fischel-Godsian de Los Angeles, ambqui es treballarà estretament per continuar l’estudi dela regió del cromosoma 8 i identificar altres regionsmodificadores. El projecte ha centrat la majoria delsesforços en la detecció i caracterització de gensmodificadors, a causa de l’elevat nombre de famíliesidentificades amb afectats i no afectats.
9) La part del model murí no s’ha pogut desenvoluparadequadament a causa de les dificultats intrínsequesen la mutagènesi del DNA mitocondrial. No obstantaixò, s’ha establert una col·laboració amb el grup delDr. Antonio Enríquez de la Facultat de Veterinària deSaragossa per desenvolupar aquesta part del projec-te. En aquests moments s’està treballant en lageneració de cèl·lules ES portadores de la mutaciómitocondrial. Cal esperar que aquest objectiu es puguidesenvolupar en un futur amb ajuts d’altres organismes.
2. Resultats1) Tipificar les alteracions clíniques i bioquímiquespresents en pacients afectats d’hipoacúsia progressi-va i/o diabetis a causa de les mutacions A1555G iA3243G en el DNA mitocondrial
2) Determinar les possibles alteracions clíniques ibioquímiques en individus asimptomàtics portadors dela mutació A1555G
Mutació A3243G:no s’ha detectat cap cas amb aquesta mutació.
Mutació A1555G:de 12 pacients amb hipoacúsia sense antecedentsfamiliars que van participar en l’estudi clínic, cap vaser portador de la mutació A1555G, mentre queaquesta es va trobar en 7 de 35 casos d’hipoacúsiaamb antecedents familiars. L’herència per via maternaera clara en totes les genealogies positives.
Del total de malalts afectats d’hipoacúsia neurosenso-rial postlocutiva amb antecedents familiars inclosos enl’estudi, el 21% van resultar portadors de la mutaciómitocondrial A1555G, que representa un percentatgemolt alt tenint en compte les nombroses alteracionsgenètiques conegudes que poden condicionar l’apari-
RECERCA BIOMÈDICA | 101

ció d’hipoacúsia. Això situa la mutació mitocondrialA1555G com un factor etiològic freqüent en el nostremedi de la hipoacúsia neurosensorial postlocutiva. Enels casos d’hipoacúsia neurosensorial en els quals nohi havia antecedents familiars, no es va trobar lamutació en cap cas.
L’important paper dels aminoglicòsids en la precipita-ció de l’aparició fenotípica de la hipoacúsia era un fetja bastant conegut i que es confirma en el nostreestudi. Es tracta d’una entitat que afecta per igualhomes i dones, però en el nostre estudi, en el casdels portadors asimptomàtics hi ha un major nombrede dones. Això té una justificació senzilla, perquè enser les dones les que transmeten la mutació, presen-ten més interès per ser estudiades.
La major part dels pacients estudiats són adults joves,però destaca la mitjana d’edat d’inici de la hipoacúsiaque es troba en edat pediàtrica (9 anys). La mitjanad’edat dels portadors asimptomàtics de la mutaciósupera amb escreix la mitjana d’edat d’inici de lahipoacúsia en els portadors simptomàtics, per la qualcosa hi ha d’haver un factor protector o un factorcoadjuvant en el desenvolupament de la hipoacúsia,no només l’exposició a aminoglicòsids. Es parla enpublicacions recents de gens nuclears que modifiquenl’expressió fenotípica.
La manifestació clínica més freqüent en els portadorssimptomàtics és òbviament la hipoacúsia en diversosgraus d’afectació, seguit de l’existència d’acúfens.Hem comprovat que, amb baixa incidència, podenexistir alteracions vestibulars.
També hem observat que els portadors asimptomàticspresenten alguna simptomatologia otològica, la mésfreqüent de les quals són els acúfens que es correla-cionen amb alguna petita alteració en l’audiometria,com és una caiguda mínima en aguts, que no tétraducció patològica, però sí que indica una certaalteració subclínica.
Els estudis audiomètrics practicats als portadorssimptomàtics revelen una hipoacúsia neurosensorialmoderada-severa, amb una afectació sempre majorde les freqüències agudes. Tots presenten una bonafunció de l’oïda mitjana. El reflex estapedial estàpresent en major proporció en les freqüències greus(500 Hz) i està absent a 4.000 Hz; això es justifica peruna major profunditat de la hipoacúsia en freqüènciesagudes.
Els PEATC presenten un allargament de l’ona I ambpreservació de les interlatències, cosa que ens central’origen del trastorn en un problema coclear senseafectació del nervi estatoacústic (VIII pc). Això esreferma pels resultats obtinguts amb els productes dedistorsió acústica, que són un reflex fidel del funciona-ment de les cèl·lules ciliades externes que, com s’hareflectit en els resultats, són absents en gairebé totsels portadors simptomàtics excepte en una freqüènciaen un individu.
Els estudis audiomètrics practicats als portadorsasimptomàtics revelen una fisiologia correcta de l’oïda
mitjana; dos malalts presenten una petita caiguda enaguts que es correspon amb una absència de reflexestapedial a 4.000 Hz. Els resultats obtinguts amb elsproductes de distorsió acústica revelen una disminu-ció de l’amplitud d’aquests si els comparem amb elsresultats que presenten els controls. Això denota undèficit en la fisiologia coclear, però que no és prousever com per ocasionar patologia.
3) Determinar la presència d’altres mutacions en elDNA mitocondrial en pacients amb sordesa i diabetis,negatius per a les mutacions esmentades
S’ha seqüenciat el gen del 12S rRNA per a un totald’11 mostres d’11 famílies amb patró d’herènciamaterna. Com a resultat, s’han trobat canvis jadescrits com a polimorfismes. S’ha identificat un canvino descrit (T1700C al gen del 16S rRNA), però que elfet de trobar-lo en moltes mostres fa pensar que estracta d’un polimorfisme. S’ha detectat un canvi nodescrit (T1243C al gen del 12S rRNA) en una solamostra en heteroplàsmia. S’ha detectat una mutacióen homoplàsmia en una família cubana (T1291C en elgen del 12S rRNA).
4) Determinar els haplotips del DNA mitocondrialassociats a la mutació A1555G
S’ha determinat l’haplotip de la mutació mitocondrialT1291C en col·laboració amb el grup de l’AntonioTorroni. Aquest haplotip sembla ser d’origen Africà-Cubà i està present en poblacions control no estudia-des per a sordesa.
5) Desenvolupar i caracteritzar el metabolisme respira-tori de models cel·lulars portadors de la mutacióA1555G
La taula següent resumeix els resultats de l’estudi delmetabolisme respiratori dels limfòcits de 4 pacientsamb la mutació A1555G:
FUNDACIÓ LA MARATÓ DE TV3

6) Desenvolupar un model murí de la mutaciómitocondrial A1555G per estudiar la fisiopatologiad’aquesta mutació en la sordesa
S’ha establert una col·laboració amb el grup del Dr.Antonio Enríquez de la Facultat de Veterinària deSaragossa per desenvolupar aquesta part del projec-te. En aquests moments s’està treballant en lageneració de cèl·lules ES portadores de la mutaciómitocondrial A1555G.
7) Identificació dels factors genètics nuclears modifica-dors dels efectes de la mutació mitocondrial.
Estudis paramètrics de lligament:L’anàlisi de lligament de dos punts amb el modelparamètric no va donar cap Lod score significatiu,però 10 regions van mostrar valors entre 1 i 1,8. UnLod score positiu dóna evidències a favor de lligamenten la zona en qüestió, però només quan el Lod scoreés superior a 3 es considera que hi ha lligament ambuna possibilitat d’error del 5%. L’anàlisi de lligamentamb GENEHUNTER amb els mateixos models no vadonar valors diferents als observats amb l’anàlisi dedos punts. Per tant, amb aquestes consideracions noes van obtenir localitzacions significatives en cap regiócromosòmica.
Resultats de l’anàlisi paramètrica de lligament
Estudis no paramètrics de lligament:Com a resultat es van identificar 10 regions cromosò-miques per a les quals hi ha una compartició d’al·lelssignificativa, amb valors de lligament multipoint entre0,5 i 1,7.
El marcador D8S277, el qual tenia el segon millorvalor de Lod score en les famílies espanyoles, se situaa uns 4 cM d’un altre marcador, el D8S262, el qualtenia un Lod score d’1,6 en un pedigrí araboisraeliàmolt extens. Per avaluar el pes que podria tenir
aquesta regió en concret, es va avaluar també en unafamília italiana. Com a resultat, els valors multipoint noparamètrics pel marcador D8S277, es van incremen-tar fins a 2,1. Es van analitzar també altres marcadorsamb Lod scores superiors a 0,5 en les famíliesespanyoles i l’araboisraelianes, tot obtenint resultatsnegatius en tots els casos.
Identificació i caracterització d’un possible locusmodificador a la regió 8p23:El locus del cromosoma 8 ha estat identificat com apossible localització d’un gen modificador, però caliaun mapatge més acurat de la regió candidat. S’hanidentificat nous marcadors polimòrfics utilitzant lainformació disponible a les bases de dades i s’hangenotipat les famílies portadores de la mutaciómitocondrial A1555G. L’anàlisi dels resultats de lagenotipació no ha aportat més informació al coneixe-ment de la zona ni del locus modificador. La figurasegüent esquematitza els marcadors utilitzats en elsdiferents anàlisis: en vermell apareixen elsmicrosatèl·lits utilitzats en la primera anàlisi i en verdels marcadors utilitzats per delimitar més la regió.
D’altra banda, també s’han identificat els genspresents a la regió, a fi d’estudiar la seva possiblecontribució al fenotip associat a la mutació mitocon-drial. Les famílies espanyoles que s’han recollit allaboratori s’han enviat al Dr. Fischel-Godsian de LosAngeles, perquè dugués a terme l’anàlisi dels gens dela regió. Al seu laboratori s’han intentat identificarvariants o mutacions presents en pacients i absentsen controls, en la major part dels gens de la regió, totobtenint només resultats negatius.
L’estudi de la seqüència d’aquesta regió va posar demanifest l’existència d’una estructura genòmicacomplexa, en la qual existeixen regions altamentrepetitives, les quals predisposen a reordenamentsgenòmics també freqüents. Aquesta estructuragenòmica pot estar implicada en la manifestaciófenotípica de la mutació A1555G i, per això, s’harealitzat una anàlisi in silico de la regió 8p23. En
RECERCA BIOMÈDICA | 103

aquesta regió, s’ha descrit una inversió de 5 Mbpresent en una proporció important en individus de lapoblació general (aproximadament 25%). A més amés, s’han descrit duplicacions i delecions en 8p23associades a diverses patologies. La caracteritzaciógenòmica d’aquesta regió ha permès identificar, abanda i banda de la inversió, parelles de duplicacionssegmentàries amb una organització complexa.Aquestes duplicacions segmentàries estan organitza-des en 16 subgrups, cada un dels quals conté entre 2i 5 duplicons de dimensió variable (5-200 kb) i unaidentitat de seqüència del 93-99%. Això resulta enuna imatge especular definida pels duplicons, queafavoreix l’aparició de diversos reordenamentsgenòmics. Grups de gens que codifiquen per defensi-nes es troben localitzats dins d’aquests duplicons.
La figura esquematitza les característiques principalsde la regió 8p23.
La regió on es localitzen els marcadors amb un Lodscore més alt associat al fenotip de sordesa es trobaal límit de la inversió. Aquest podria ser un factor deconfusió en els estudis de lligament a causa del’elevada proporció d’individus en la població generalque en són portadors. D’altra banda, la inversió per sisola també podria ser un dels factors modificadors dela mutació mitocondrial.
Un altre aspecte important que cal tenir en compte ésel nombre variable de còpies dels gens de defensines.S’ha intentat valorar el nombre de còpies en individuscontrol i portadors de la mutació per mitjà de l’anàliside microsatèl·lits duplicats, però no s’ha arribat a capconclusió. De totes maneres, s’ha pogut comprovarque el nombre de còpies d’aquests gens és variableen la població general, amb la qual cosa no es potdescartar un efecte dosidependent de les defensines,en el fenotip associat a la mutació A1555G. Ésimportant caracteritzar millor la zona per determinarquin és el paper d’aquesta regió com a factor modifi-cador de la mutació A1555G, per la qual cosa elsnostres estudis continuaran en aquesta direcció.
Identificació de localitzacions addicionals de gensmodificadors:És molt probable l’existència de més d’un factormodificador i s’han intentar identificar altres localitza-cions possibles. La identificació de noves famíliesportadores de la mutació és imprescindible per ferestudis de lligament a tot el genoma utilitzant nousmarcadors polimòrfics.
S’estan estudiant les altres zones que en l’anàlisiglobal del genoma tenien un Lod score superior a 1,5.La màxima atenció se centra en les regions següents:1q32, 3p11, 3p24, 6q25, 7p25, 8q11, 9q13, 15q14,15q22 i 17q23. Per determinar la contribució decadascuna d’aquestes zones com a possibles localit-zacions de factors modificadors, és convenient un
mapatge més fi de les diverses regions, per la qual cosaes genotiparan més marcadors al voltant de cadascunade les regions d’interès prèviament descrites.
3. Rellevància i possibles aplicacions clíniquesdels resultats finals obtingutsLa sordesa és un defecte neurosensorial moltfreqüent, ja que afecta el 10% de la població. Unaproporció important dels casos de sordesa tenencausa genètica, ja sigui monogènica o poligènica. Enl’actualitat no existeixen tractaments adequats per a lasordesa, ni es disposa de cap forma de prevenció.
La mutació mitocondrial A1555G constitueix un delsprincipals factors genètics de susceptibilitat per a lasordesa neurosensorial no sindròmica. Aquestamutació també es relaciona amb una major suscepti-bilitat a sordesa en l’administració d’antibiòticsaminoglicòsids i, per aquest fet, representa un delsmillors exemples de la interacció entre factorsgenètics i ambientals en el desenvolupament de lapatologia. La detecció precoç d’aquesta mutaciópermet una prevenció adequada i pel fet de tractar-sed’una forma de sordesa progressiva, la identificació dela mutació permet l’aplicació de teràpies pal·liatives i lareeducació del pacient molt abans que la pèrdua lisuposi dificultats importants en la seva vida quotidiana.
La informació epidemiològica resultant de l’estudi hapermès l’obtenció de dades fiables sobre la incidènciad’aquesta mutació en la població catalana. Alhora, lacaracterització de les alteracions clíniques i bioquími-ques presents en individus portadors de la mutacióA1555G, tant afectats com no, ha permès una localit-zació més concreta i una major comprensió delsproblemes del sistema auditiu que apareixen com aconseqüència d’aquest canvi al DNA mitocondrial.
D’altra banda, l’estudi dels mecanismes molecularssubjacents ens aporta informació que facilita lacomprensió de la interacció entre gens i entorn. Hi haevidències suficients de l’existència d’un o més gensmodificadors del fenotip associat a la mutacióA1555G. Això fa que ens trobem davant d’un modeld’estudi que permet entendre una mica millor lesinteraccions nucli-mitocondri, les quals determinenl’expressió fenotípica de les mutacions mitocondrialsen general.
4. PublicacionsDonaudy F, Snoeckx R, Pfister M, Zenner HP, Blin N, Di Stazio
M, Ferrara A, Lanzara C, Ficarella R, Declau F, Pusch CM,
Nurnberg P, Melchionda S, Zelante L, Ballana E, Estivill X, Van
Camp G, Gasparini P, Savoia A. Nonmuscle Myosin Heavy-Chain
Gene MYH14 Is Expressed in Cochlea and Mutated in Patients
Affected by Autosomal Dominant Hearing Impairment (DFNA4).
Am J Hum Genet. 2004 Apr;74(4):770-6. Epub 2004 Mar 10.
E Thönnissen, R Rabionet, ML Arbonés, X Estivill, K Willecke, T
Ott. Human connexin26 (GJB2) deafness mutations affect the
function of gap junction channels at different levels of protein
expression. Human Genetics 111:190-197 (2002)
N López-Bigas, ML Arbonés, X Estivill, L Simonneau
Expression profiles of the connexin genes, Gjb1 and Gjb3, in the
developing mouse cochlea. Mechanisms of Development 2:111-
115 (2002)
FUNDACIÓ LA MARATÓ DE TV3

R Rabionet, N Lopez-Bigas, ML Arbones, X Estivill. Connexin
mutations in hearing loss, dermatological and neurological
disorders. Trends in Molecular Medicine 8:205-212 (2002)
Y Bykhovskaya, H Yang, K Taylor, …, X Estivill, …., N Fischel-
Ghodsian. Modifier locus for mitocondrial DNA disease. Linkage
and linkage disequilibrium mapping of a nuclear modifier gene for
maternally inherited deafness. Genetics in Medicine 3:177-180
(2001)
N López-Bigas, S Melchionda, R de Cid, A Grifa, L Zelante, N
Govea, ML Arbones, P Gasparini, X Estivill. Identification of five
new mutations of PDS/SLC26A4 in Mediteranean Families with
hearing impairment. Human Mutation 18:548 (2001)
N López-Bigas, R Rabionet, E Martínez, O Bravo, J Girons, A
Borragán, M Pellicer, ML Arbonés, X Estivill. Mutations in the
mitochondrial tRNA Ser(UCN) and in the GJB2 (conexin 26) gene
are not modifiers of the age at onset and severity of hearing losss
in Spanish patients with the 12S rRNA A1555G mutation.
American Journal of Human Genetics 66:1465-1466 (2000)
Y Bykhovskaya, X Estivill, K Taylor, T Hang, M Hamon, ...., N
Fischel-Ghodsian. Candidate locus for a nuclear modifier gene for
maternally inherited deafness. American Journal of Human
Genetics 66:1905-1910 (2000)
A Torroni, F Cruciani, ..., N López-Bigas, R Rabionet, N Govea,
..., X Estivill, R Scozzari. The A1555G mutation in the 12S rRNA
gene of human mtDNA: Recurrent origins and founder events in
families affected by sensorineural deafness. American Journal of
Human Genetics 65:1349-1358 (1999)
RECERCA BIOMÈDICA | 105

FUNDACIÓ LA MARATÓ DE TV3
Títol del projecte
Dissecció d'un circuit transcripcional que controla la funció de les cèl·lules productores d'insulina
Investigador principalJorge Ferrer Marrades
InstitucióHospital Clínic i Provincial de Barcelona
100.017 €
MEMÒRIA FINAL
1. Resum del projecte originalAquest estudi es va plantejar poc després quediversos grups identifiquessin que mutacions en unconjunt de gens que codifiquen factors de transcripcióprodueixen una forma de diabetis anomenadamaturity onset diabetes of the young (MODY) i que escaracteritza per un defecte de la funció de cèl·lulesbeta del pàncrees. Aquests gens implicats en MODYsón HNF1alpha, HNF4alpha, Pdx1/IPF1, HNF1beta.La hipòtesi del projecte va ser que si els gens MODYsón tan importants per al funcionament de les cèl·lulesbeta, és prioritari comprendre de manera detallada laseva funció en aquestes cèl·lules. D’altra banda, atèsque tots aquests factors de transcripció donen lloc aun fenotip genètic similar, era versemblant que totsells participessin en una xarxa reguladora comuna.L’interès d’afrontar aquest problema és múltiple. Peruna part, es tracta de comprendre les bases molecu-lars d’una forma específica de diabetis, la qual cosapot tenir implicacions per al seu tractament. I per altrapart, com que aquests gens són tan importants per ala funció i diferenciació de les cèl·lules beta, la dissec-ció dels seus mecanismes d’acció és rellevant en la
producció artificial de noves cèl·lules beta per ateràpies destinades a diverses formes de diabetis.
El projecte es va centrar en l’aprofundiment sobre lafunció d’un d’aquests factors, HNF1alpha, el qual estàimplicat en la forma més freqüent de diabetis heredità-ria (MODY3). Els objectius concrets que ens vamproposar, per tant, va ser:
1) Identificar quins gens són controlats per HNF1alphaen les cèl·lules beta, en particular si altres gens regula-dors de la transcripció implicats en MODY són tambédianes d’HNF1alpha, ja que això donaria suport a lahipòtesi d’una xarxa reguladora comuna.2) Investigar els mecanismes d’activació transcripcio-nal emprats per HNF1a .3) Desenvolupar models genètics que permetin l’expres-sió selectiva d’HNF1a i les seves dianes en cèl·lulesbeta, amb la finalitat de determinar si la diabetis que esprodueix en ratolins que tenen inactivat el gen HNF1alpha és causada per una funció d’aquest factor restrin-gida a les cèl·lules beta ("autònom cel·lular").Vam proposar utilitzar ratolins amb inactivació del genHNF1a per analitzar els patrons d’expressió gènicadependents d’HNF1alpha, utilitzant immunoblots i

hibridacions de matrius de cDNA d’alta densitat.També es va proposar investigar com HNF1alpharegula la transcripció dels seus gens diana en lescèl·lules beta. L’estratègia escollida en aquell momentva ser estudiar la interacció amb coactivadorscandidats en el context d’un promotor de cèl·lula betadiana utilitzant models in vitro, i la recerca de novesinteraccions proteïna-proteïna mitjançant un sistemadoble-híbrid. Finalment vam proposar utilitzar unmodel transgènic per expressar HNF1alpha de formaselectiva en les cèl·lules beta de ratolins que tenenuna inactivació del gen HNF1alpha endogen.
2. ResultatsAquest projecte ha pogut assolir gairebé tots elsobjectius fixats, i ha conduït a troballes significativesen el camp de la regulació gènica de les cèl·lulesproductores d’insulina del pàncrees:
1) El descobriment d’un circuit transcripcional especí-fic de cèl·lules pancreàtiques que és controlat perHNF1a. Aquest circuit relaciona diferents factors detranscripció indispensables per a la diferenciaciópancreàtica. Un aspecte interessant d’aquesta xarxareguladora és que alguns dels gens que la formen hanestat prèviament implicats en la diabetis humanaautosòmica dominant (MODY).2) El descobriment del promotor alternatiu del genHNF4a que dirigeix la seva transcripció en les cèl·lulespancreàtiques. La importància d’aquesta troballa rauen el fet que s’ha demostrat prèviament que lesmutacions del gen HNF4a produeixen diabetis enhumans (MODY1) per un defecte de la funció de lescèl·lules pancreàtiques. Per tant, identifica la partd’aquest gen que controla la transcripció concreta-ment en les cèl·lules rellevants a la seva funció relacio-nada amb la diabetis. A més, vam demostrar queHNF1alpha (el gen que produeix MODY3) controla latranscripció del gen HNF4alpha a través d'aquestaregió gènica promotora.3) La identificació i caracterització d’una mutació en elpromotor nou d’HNF4 alpha en una família amb MODY1que destrueix la capacitat de regulació per HNF1alpha.Això ha permès demostrar no solament la importànciad’aquest promotor en humans, sinó que ha confirmat enhumans les nostres dades en ratolins, que HNF1alphaés un regulador del gen HNF4alpha, i addicionalmentmostra que aquest fet regulador és un componentindispensable de la funció d’HNF1alpha al pàncrees.4) La postulació d’un model, basat en les troballes dela xarxa gènica descoberta en aquest projecte, queexplica per què la pèrdua d’un sol al·lel del genHNF1alpha o HNF4alpha produeix una malaltiaespecíficament en les cèl·lules productores d’insulina.5) Des del punt de vista de la comprensió delsmecanismes d’activació de la transcripció perHNF1alpha, es van identificar canvis en la cromatinaproduïts per aquest factor en els seus gens diana,específicament en les cèl·lules productores d’insulina.Això va comportar el desenvolupament per primer copen el camp de la regulació gènica del pàncrees d’unsistema d’anàlisi in vivo aplicable al model animal.Aquesta estratègia va substituir altres metodologiesmés artificials basades en models in vitro proposadesen el nostre projecte inicial.L’estudi del ratolí transgènic per expressar HNF1alpha
selectivament en les cèl·lules beta es va complicar pelfet que aquests animals van presentar un fenotip deletalitat inesperat. Es va generar, per tant, un nousistema per expressar HNF1alpha selectivament enles cèl·lules beta de forma controlada en el temps.Aquests estudis encara estan en marxa. També estanencara en marxa estudis per estudiar noves interac-cions d’HNF1alpha en la cèl·lula beta.Finalment, val la pena esmentar que els resultatsderivats d’aquest projecte ens han permès dur aterme un estudi subsegüent que ha identificat unapoblació de cèl·lules mare tissulars del pàncrees i unanova xarxa transcripcional que opera en aquesta fasedel desenvolupament pancreàtic.
3. Rellevància i possibles implicacions clíniquesdels resultats finals obtingutsEn termes generals, aquests estudis han aportat nousconeixements fonamentals per comprendre elsmecanismes genètics detallats que controlen eldesenvolupament del pàncrees. El descobrimentd’una xarxa gènica que és imprescindible per a ladiferenciació pancreàtica, i que al mateix tempsrelaciona reguladors implicats en la diabetis humana,constitueix una diana òbvia per desenvolupar estratè-gies terapèutiques de diversos tipus. En primer lloc,per incrementar la funció de les cèl·lules beta enmalalts amb diabetis tipus 2 o MODY.En segon lloc, per completar el procés de diferencia-ció de les cèl·lules beta a partir de cèl·lules mareembrionàries o d’altres orígens. El promotor noud’HNF4alpha també constitueix més específicamentuna diana terapèutica per motius semblants. Tal coms’ha comentat anteriorment, els resultats derivatsd’aquest projecte han permès dur a terme un estudisubsegüent al nostre laboratori, el qual ha permèsidentificar una població de cèl·lules mare tissulars delpàncrees. Aquest tipus de troballa també té unaimportància capital en les estratègies terapèutiquesdestinades a generar cèl·lules beta artificials.
4. PublicacionsBoj SF, Parrizas M, Maestro MA, Ferrer J. (2001). A transcrip-
tion factor regulatory circuit in differentiated pancreatic cells. Proc
Natl Acad Sci USA 98(25): 14481-6.
Parrizas M, Maestro MA, Boj SF, Paniagua A, Casamitjana R,
Gomis R, Rivera F, Ferrer J. (2001) Hepatic nuclear factor 1-alpha
directs nucleosomal hyperacetylation to its tissue-specific
transcriptional targets. Mol Cell Biol 21(9): 3234-43.
Hansen SK, Parrizas M*, Jensen ML, Pruhova S, Ek J, Boj SF,
Johansen A, Maestro MA, Rivera F, Eiberg H, Andel M, Lebl J,
Pedersen O, Ferrer J*, Hansen T. ( 2002). Genetic evidence that
HNF-1alpha-dependent transcriptional control of HNF-4alpha is
essential for human pancreatic beta cell function.
J Clin Invest. 110(6): 827-33 (*coautor primer i primer autor de
correspondència, respectivament). Ferrer J. (2002). A genetic
switch in pancreatic beta-cells: implications for differentiation and
haploinsufficiency. Diabetes 51(8): 2355-62.
Servitja JM, Ferrer J. (2004). Transcriptional networks contro-
lling pancreatic development and beta cell function.. Diabetologia
(en prensa)
Maestro MA, Boj SF, Luco RF, Pierreux CE, Cabedo J, Servitja
JM, German MS, Rousseau GG, Lemaigre FP, Ferrer J. (2003).
Hnf6 and Tcf2 (MODY5) are linked in a gene network operating in
a precursor cell domain of the embryonic pancreas. Hum Mol
Genet 12(24): 3307-14
RECERCA BIOMÈDICA | 107

FUNDACIÓ LA MARATÓ DE TV3
MEMÒRIA FINAL
1. Resum del projecte originalLa síndrome de Wiskott-Aldrich (WAS) és una malaltiamonogènica lligada al cromosoma X que cursa ambèccema, immunodeficiència i trombocitopènia deplaquetes petites. El gen responsable de la malaltia ésel gen WASP i s’han descrit mutacions tot al llarg delgen en pacients WAS. El nostre grup, en el marcd’aquest projecte, ha contribuït significativament a laidentificació i caracterització de mutacions noves enaquests pacients. En població espanyola hem estudiat26 famílies i s’han identificat 17 mutacions diferents.Normalment, les dones són portadores de la malaltia,però són asimptomàtiques. Tanmateix, s’han descritalguns casos de dones portadores amb clínica. Elnostre grup ha demostrat que en alguns casos dedones amb trets de WAS, el que s’esdevé és unadesviació en el patró d’inactivació del cromosoma Xque afavoreix l’expressió de l’al·lel mutat.
L’única teràpia actual efectiva per a aquests pacientsés el trasplantament de moll de l’os, però ensituacions d’absència de donants histocompatiblesexisteix un risc de mortalitat entre el 20-30%, la qualcosa fa que siguin necessàries teràpies alternatives.
La transferència gènica de WASP en aquests pacientspodria ser una alternativa, atès que el gen es transferi-ria a les cèl·lules del mateix pacient i s’evitarien elsproblemes relacionats amb la malaltia de l’empeltcontra l’hoste. És important, però, en primer llocesbrinar la capacitat correctora de la transferènciagènica de WASP en les principals cèl·lules alterades.En aquest projecte hem avaluat la capacitat de latransferència del gen WASP per revertir les alteracionsfuncionals presents en cèl·lules B i T de pacients ambla síndrome de Wiskott-Aldrich. En cèl·lules B hempogut demostrar que la reintroducció de WASP, pertransducció retroviral en cèl·lules WAS deficientsrecupera les alteracions associades amb el citosqueletd’actina. Es recupera la capacitat de formar filopodisdesprés d’un estímul per bradiquinina, com també laformació de magnopodis i tecnopodis. Tanmateix,també hem observat que l’expressió de WASP indueixcanvis de caire transcripcional en altres gens i n’estemestudiant les conseqüències.
2. ResultatsI Estudi mutacional1. Deteccció de les mutacions en famílies espanyoles.
Mitjançant anàlisi per SSCP i seqüenciació directa,
Títol del projecte
Transferència del gen WASP, mediada per vectors retrovírics, a limfoblastos de pacients amb síndrome de Wiskott-Aldrich:
avaluació dels canvis metabòlics i funcionals
Investigadors principalsCristina Fillat Fonts
InstitucióCentre de Genètica Mèdica i Molecular. IRO
Membres de l’equipJosep Aran, Víctor Volpini i Núria Andreu.
29.903 €

s'han identificat 17 mutacions diferents en 26 famíliesestudiades.
Aquests resultats es recullen en Fillat et al., Am. J.Med. Genet (2001); Olivares et al., J. Pediat. Hemat.Onco (2002); Andreu et al., J. Hum. Genet. (2003);Andreu et al. (en preparació).
2. Estudi del patró d’inactivació del cromosoma X enuna dona portadora de WAS amb simptomatologiaclínica.
Els resultats obtinguts es recullen en Andreu et al.,Blood Cells Mol. Dis. (2003).
II Estudis de transferència gènica1. Caracterització dels retrovirus recombinantspLHWSN. Anàlisi de l'expressió
S'han caracteritzat els retrovirus pLHWSN. Els estudisd'expressió en cèl·lules Cos-7 han permès evidenciarla sobreexpressió de WASP mitjançant el retroviruspLHWSN tant per Western blot com per inmunofluo-rescència. L'anàlisi d'expressió també s'ha realitzat enels limfoblastos de pacients R86H i E133K transduïtsamb el vector pLHWSN. Els estudis de colocalitzaciódemostren com WASP i F-actina colocalitzen en lacèl·lula. Els estudis amb faloidina, un compost ques'uneix als filaments d'actina i permet visualitzar-los, jaque presenta coloració vermella, indicarien que lescèl·lules mutades R86H i E133K presenten un marcat-ge tènue i difús, essent més evident en les cèl·lulesE133K. En canvi, en les cèl·lules en què WASP sesobreexpressa, el marcatge és molt més intens.
2. Caracterització dels retrovirus recombinantspLHWSN. Estudis funcionals
Es va determinar la capacitat de les cèl·lules WASPmutades i les WASP corregides de formar filopodisper l’estímul bradiquinina. Només s’observà la capaci-tat formadora de filopodis en les cèl·lules que es vancorregir amb el retrovirus que contenia el gen WASP ino en les transduïdes amb el retrovirus control.
S’ha descrit que les cèl·lules hematopoètiques sóncapaces de formar un nou tipus de podis de funcióencara avui desconeguda (magnopodis, tecnopodis).
Vam estudiar aquest tipus d’estructures en el nostremodel i curiosament les cèl·lules WASP deficients sónincapaces de formar-los, mentre que en reintroduir elgen WASP es recupera aquesta capacitat, tot indicantla capacitat WASP de restablir el fenotip alterat.
3. Conseqüències transcripcionals de la sobreexpres-sió de WASP en limfòcits B
Vam voler estudiar si la reintroducció de WASP en unacèl·lula WASP negativa podia alterar el patró transcrip-cional d’alguns gens. Mitjançant la tecnologia de RAP-PCR es van identificar canvis en l’expressió dediversos gens. Per tècniques de Northern blot es vaconfirmar la reducció en els nivells de CTCF, i enaquest sentit hem iniciat estudis funcionals quepensem completar durant aquest any
4. Estudis de transferència génica en limfòcits T
S’han iniciat els estudis de transducció en cèl·lules Tper comprovar si les correccions fenotípiques o lesalteracions identificades en cèl·lules B es donaventambé en cèl·lules T. Inicialment vam iniciar el treballen cèl·lules T transformades amb el virus de l’HerpesSaimiri. Es van usar cèl·lules de dos pacients ambdues mutacions diferents en WASP, la IVS8+1 i laR86H , com també cèl·lules procedents d’unaportadora de WAS. Utilitzant un retrovirus controlportador del gen de resistència a neomicina vamaconseguir transduir les tres línies cel·lulars anterior-ment esmentades. Tanmateix, usant el retrovirus quecontenia el gen WASP només vam poder transduir lescèl·lules T procedents de la portadora i en cap casdels pacients. Aquests resultats suggeriren que algunainterferència es produiria entre el gen WASP i algunsdels gens que es podrien expressar com a resultat dela transformació amb el virus Herpes Saimiri. Això ensva obligar a reconsiderar els estudis en cèl·lules T ivam optar per iniciar-los en cèl·lules T primàriesobtingudes de pacients i sense cap modificacióposterior més que la del mateix cultiu. A fi d’incorpo-rar aquesta metodologia al laboratori, Núria Andreu vafer una visita al laboratori del Dr. Juan Bueren alCIEMAT a Madrid i amb la seva col·laboració hemposat a punt al laboratori la transducció de cèl·lules Tprimàries en individus control i som a punt d’iniciar latransducció de cèl·lules T de pacients WAS en quèpoder valorar la formació de podis i la regulació delgen CTCF.
3. Rellevància i possibles implicacions clíniquesdels resultats finals obtingutsUn dels aspectes que es deriven de la recerca ambaquest projecte i que té unes implicacions clíniquesdirectes és l’estudi molecular del gen WASP. El nostreés l’únic grup a l’Estat espanyol que fa aquest tipusd’anàlisi i això representa un ajut a les famíliesimportant, ja que confirma el diagnòstic clínic i a mésa més s’ofereix la possibilitat de realitzar, quan calgui,un diagnòstic prenatal. Malauradament, aquesta és
RECERCA BIOMÈDICA | 109

una tasca que, iniciada des de la recerca, després hade ser transferida a les unitats de diagnòstic delshospitals, i hem iniciat converses amb algunshospitals perquè es pugui dur a terme.
Un altre aspecte que té una possible implicacióclínica, però per a més endavant, és el treball realitzaten teràpia gènica i que encara està en procés.Aquesta és sens dubte una malaltia candidata a sersusceptible de tractament mitjançant teràpia gènica.Pertany al grup de malalties en què l’única teràpiaefectiva és el trasplantament de moll de l’os i enaquells casos en què no es disposa d’un donantcompatible el risc de mortalitat en el trasplantamentés encara molt alt, entre un 20-30%. Fent un balançrisc/benefici sobre l’aplicació de la teràpia gènica enaquests malalts, en un article recent Cavazzana-Calvoet al., Nature 427 779-781 (2004) clarament indicavaque, malgrat que igual que altres tractaments, lateràpia gènica tenia alguns efectes secundaris, no peraixò s’ha de deixar d’aplicar, ja que en algunspacients el benefici havia estat molt clar. El nostretreball creiem que contribueix a demostrar que amb lateràpia gènica podem recuperar alteracions cel·lularsimportants en les cèl·lules WAS i revertir així les princi-pals disfuncions.
4. PublicacionsC. Fillat, T. Español, M. Oset, M. Ferrando, X. Estivill, V. Volpini.
Identification of WASP mutations in 14 Spanish families with
Wiskott-Aldrich syndrome. Am. J. Med. Genetics (2001) 100:116-
121.
J.L. Olivares, F.J. Ramos, T. Olivé, C. Fillat, M. Bueno.
Autoinmune thyroiditis after bone marrow transplantation in a boy
with Wiskott-Aldrich syndrome. J. Pediat.. Hemat. Oncol (2002)
24:772-776.
N. Andreu, N. Pujol-Moix, L- Martínez-Lostao, M. Oset, E.
Muñiz-Díaz, X. Estivill, V. Volpini, C. Fillat. Wiskott-Aldrich syndro-
me in a female with skewed X-chromosome inactivation.
Blood Cells Mol. Dis. (2003) 31:332-337.
N. Andreu, C. Carreras, F. Prieto, X. Estivill, V. Volpini, C. Fillat.
Identification and characterization of a novel splice site mutation in
a patient with Wiskott-Aldrich syndrome. J. Hum. Genet. (2003)
48:590-593.
N. Andreu, N. Matamoros, A. Escudero, G. Fontan, X. Estivill,
V. Volpini, C. Fillat. Three novel mutations in Wiskott-Aldrich
syndrome. (manuscrit en preparació).
N. Andreu, J.M. Aran, T. Español, X. Estivill, C. Fillat.
Phenotypic correction of Wiskott-Aldrich syndrome defects in
lymphoblastoid B and T cells after retroviral transduction.
(manuscrit en preparació).
FUNDACIÓ LA MARATÓ DE TV3

RECERCA BIOMÈDICA | 111
MEMÒRIA FINAL
1. Resum del projecte originalL’objectiu principal d’aquest projecte era el descobri-ment i l’anàlisi de nous gens relacionats amb el genIPF-1 (Pdx1), que poguessin estar involucrats en ladiabetis mellitus tipus 2. El gen Pdx1 està involucraten la formació del pàncrees, regula el gen de la insuli-na, i mutacions seves són responsables d’un subtipusde diabetis (MODY4). Com que el gen Pdx1 formapart d’un dels complexos gènics ParaHox recentmentdescoberts, i que el genoma humà és resultat deduplicacions gèniques a gran escala en l’origen delsvertebrats, el projecte pretenia investigar si la resta degens ParaHox (Gsh1, Gsh2, Cdx1, Cdx2/3 i Cdx4),tots ells codificant per factors de transcripció, podriendesenvolupar algun paper regulador en el pàncreesendocrí, i si es podia detectar alguna relació funcionalentre els gens ParaHox i la regulació del gen de lainsulina, que es coneix regulat per Pdx1. Els gensamb homeobox (especialment els de tipus Hox-likecom els ParaHox) presenten una regulació complexaen la qual molts cops intervenen gens homòlegs o delmateix clúster. D'altra banda, malgrat que Pdx1 éscrucial per al desenvolupament del pàncrees, en
ratolins knockout per a aquest gen, tot i presentaragènesi del pàncrees s'han trobat cèl·lules disperses al'intestí que expressen insulina. L'anàlisi de la possibleunió dels ParaHox i de la regulació de l'expressió delgen de la insulina es trobà interessant per comprendreles bases moleculars de la diabetis mellitus tipus 2,com també per identificar nous gens candidats aresponsables de desordres diabètics, i per les sevespossibles aplicacions terapèutiques, fins i tot com afutures teràpies gèniques substitutives.
En concret, els objectius del projecte eren:1. Aïllament dels gens ParaHox de ratolí.2. Anàlisi del patró d'expressió de tots els gensParahox, incloent-hi Pdx1, en pàncrees de ratolí adulti en cultius cel·lulars de cèl·lules a i b-pancreàtiquesen condicions fisiològiques diferents.3. Anàlisi funcional del reconeixement i la interacciód’aquests factors de transcripció ParaHox amb elpromotor del gen de la insulina.4. Avaluació dels gens ParaHox com a possiblescandidats responsables de l’aparició de diabetis ianàlisi mutacional en pacients diabètics tipus 2.
Succintament, els gens ParaHox de ratolí s’havien
Títol del projecte
Cerca de gens relacionats amb l'IPF-1 (insulin promoter factor-1):implicacions en la regulació del gen de la insulina i en la diabetis de tipus 2
Investigador principalJordi Garcia-Fernàndez i Gemma Marfany Nadal
InstitucióFacultat de Biologia. UB
108.733 €

FUNDACIÓ LA MARATÓ DE TV3
d’aïllar emprant la informació dels bancs de dadespúblics i ser clonats en els vectors adequats. Per al’anàlisi de l’expressió es proposava emprar la tècnicad’hibridació in situ en talls histològics, combinada ambimmunodetecció per identificar les poblacionscel·lulars específiques que expressessin els gensParaHox en el pàncrees endocrí, si fos el cas, comtambé la utilització d’RT-PCR. El tercer objectiu esbasava principalment en l’estudi de cotransfeccionsen cèl·lules en cultiu derivades de cèl·lules alfa o beta-pancreàtiques, amb vectors que expressessin consti-tutivament les diverses proteïnes ParaHox, i devectors "reporters", on la regió promotora proximal delgen de la insulina dirigeix l’expressió del gen marcadorluciferasa.
2. ResultatsResumL’objectiu principal d’aquest projecte era el descobri-ment i l’anàlisi de nous gens relacionats amb el genIPF-1 (Pdx1), que poguessin estar involucrats en ladiabetis mellitus tipus 2. Hem estudiat l’expressió detots els gens dels complexos ParaHox, dels qualsforma part Pdx1, i hem estudiat la capacitat transacti-vadora de les proteïnes ParaHox sobre el promotordel gen de la insulina, en un model de cèl·lules encultiu derivades de pàncrees.
Els resultats del projecte es resumeixen succinta-ment en:
1) Els gens ParaHox Gsh1, Gsh2, Cdx1, Cdx2/3 iCdx4 s’expressen en el pàncrees endocrí adult.Majoritàriament ho fan en cèl·lules alfa pancreàtiques,productores de glucagó.2) Els gens ParaHox Pdx1, Gsh1, Gsh2, Cdx1,Cdx2/3 i Cdx4 s’expressen en cultius de líniescel·lulars endocrines, tant derivades de cèl·lules alfacom beta-pancreàtiques.3) Les proteïnes ParaHox Gsh1, Gsh2, Cdx1, Cdx2/3i Cdx4, a més de Pdx1, són factors de transcripciócapaços de regular el gen de la insulina en models delínies cel·lulars endocrines, tant derivades de cèl·lulesalfa com beta-pancreàtiques.
Consideració generalLa diabetis de tipus 2 és una malaltia molt prevalenten les poblacions occidentals, i es destinen esforçosimmensos a identificar gens candidats que potencial-ment estiguin involucrats en la malaltia. Aquest projec-te ha incidit en l’estudi de tots els factors de transcrip-ció membres dels complexos gènics ParaHox, delsquals forma part Pdx1, un activador transcripcionaldel gen de la insulina i gen responsable d’un subtipusde diabetis. L’organització en complexos gènics delsgens ParaHox, i l’anàlisi genòmica comparada queindica que tots deriven d’un gen ancestral comú, ensva fer hipotetitzar que podien tenir funcions relaciona-des. Així, hem estudiat l’expressió gènica i la capacitattransactivadora sobre la insulina de tots els gensParaHox. Sorprenentment, tots s’expressen ensubtipus particulars del pàncrees endocrí adult, i enlínies cel·lulars derivades de cèl·lules pancreàtiques.D’altra banda, hem mostrat que les proteïnes ParaHoxpoden regular el gen de la insulina.
Els nostres resultats són totalment inèdits, i no hireferències bibliogràfiques sobre la possible funciódels gens ParaHox Gsh1, Gsh2, Cdx1, i Cdx4 en laformació i el manteniment del pàncrees endocrí, nisobre la seva possible funció reguladora del gen de lainsulina. En conjunt, creiem que els nostres resultatsobren vies inèdites i prometedores de recerca, perquèsuggereixen que els gens ParaHox podrien estarimplicats en processos diabètics.
Exposició detallada dels resultats1) Expressió dels gens ParaHox: aspectes generals.
Els experiments d’RT-PCR dels gens ParaHox sobrepanells de cDNA de diversos estadis embrionaris i deteixits de ratolí adult (figura 1) mostren que Cdx2/3s’expressa a alts nivells durant els diversos estadis dedesenvolupament testats (7, 11, 15 i 17 dies post-coitum), mentre que els altres ParaHox estan mésrestringits en el temps. Cdx4 s’expressa nomésdurant els primers estadis del desenvolupament.L’expressió de Pdx1, Gsh1 i Gsh2 apareix més tarden el desenvolupament, i és detectable a partir dels11 dpc. Cdx-1, finalment, sembla relacionat ambprocessos de diferenciació, ja que s’expressa nomésa partir del dia 17. En teixits adults, l’expressió de totsels gens ParaHox és variada i complexa (figura 1).
2) Expressió dels gens ParaHox en línies cel·lularsderivades de pàncrees.
Els experiments d’RT-PCR sobre RNA obtingut de leslínies cel·lulars pancreàtiques beta-beta TC6 i alpha-TC1.9 han mostrat que, a baixes concentracions deglucosa (0-5 mM), les cèl·lules beta-TC6 expressen aalts nivells Pdx1, Cdx2/3 i Gsh1 i a baix nivell Cdx4.Els nivells d’expressió no canvien quan les cèl·lules esmantenen a altes concentracions (25 mM) de glucosa.En condicions de glucosa baixa, les cèl·lules alfa TC1expressen només Pdx1, mentre que, en augmentar laconcentració de glucosa, Cdx1, Gsh1 i Gsh2 tambéactiven la seva expressió.
3) Expressió dels gens ParaHox en pàncrees adult.
Mitjançant hibridacions in situ sobre talls histològicsde pàncrees de ratolí adult, hem esbrinat que tots elsgens ParaHox, incloent-hi el ja descrit Pdx1, s’expres-sen en el pàncrees endocrí adult amb relativesdiferències en les subpoblacions positives per leshibridacions (figura 2).

RECERCA BIOMÈDICA | 113
Per identificar les subpoblacions cel·lulars en cadacas, hem fet coimmunolocalitzacions sobre leshibridacions in situ, mitjançant anticossos antiglucagó(específic per a cèl·lules alfa) i antiinsulina (específicper a cèl·lules beta). En aquestes triples deteccionss’observa que Pdx1 és específic de cèl·lules beta iCdx2/3 de cèl·lules alfa, tal com està descrit en labibliografia. Les nostres dades, a més, revelen que elsaltres gens ParaHox s’expressen en cèl·lules alfa.
Particularment remarcable és el cas de Gsh-1 i Gsh-2,gens que només havien estat relacionats amb eldesenvolupament del sistema nerviós, i que lesnostres dades mostren que s’expressen en cèl·lulesalfa, ja que colocalitzen amb cèl·lules productores deglucagó (figura 3).
Figura 3. Hibridación in situ (señal negra) de Gsh1 (izquierda) o
Gsh2 (derecha) en cortes histológicos de páncreas, y coinmuno-
detección con anticuerpo antiglucagón (verde) o antiinsulina (rojo)
4) Capacitat transactivadora de les proteïnes Parahoxsobre el gen de la insulina.
Mitjançant triples transfeccions en els cultius cel·lularsbeta-TC6 i alfa-TC1 amb vectors controls, vectorsconstitutivament productors de les diverses proteïnesParaHox, i vectors reporters amb el promotor del gende la insulina, hem investigat les capacitats transacti-vadores de les diverses proteïnes ParaHox ensistemes en què el gen de la insulina es troba normal-ment inactiu (cèl·lules alfa) i on es troba actiu encondicions d’alta concentració de glucosa (cèl·lulesbeta a 25 mM de glucosa). En cèl·lules alfa, la majoriade proteïnes ParaHox, excepte Cdx1, activen latranscripció dirigida pel promotor del gen de la insuli-na. Els nivells d’activació són dependents de laconcentració de glucosa en el medi, i en la majoriadels casos són dependents de la concentració decada factor de transcripció ParaHox particular. Lesdades globals es resumeixen en la taula I.
Taula I. Activació del gen de la insulina per les proteïnes ParaHox
en cèl·lules alfa pancreàtiques TC1
En cèlules beta-pancreàtiques, Pdx1, tal com estàdescrit, té una capacitat transactivadora molt elevadasobre el promotor del gen de la insulina (taula II). Laresta de ParaHox, però, també són capaços d’activar(+) o reprimir (-) la construcció reportera a nivellsrellevants.
Taula II. Activació del gen de la insulina per les proteïnes ParaHox
en cèl·lules beta-pancreàtiquesTC6
En conjunt, els experiments de transfeccions indiquenque les proteïnes ParaHox, amb homeodominis d’unióal DNA emparentats, són capaces, en experiments desobreexpressió, d’unir-se al promotor del gen de lainsulina, tot activant o inhibint l’expressió, la qual cosasuggereix que en condicions fisiològiques tambédeuen estar involucrats en la regulació del gen de lainsulina.
3. Rellevància i possibles implicacions clíniquesdels resultats finals obtingutsAquest projecte d’investigació és eminentment deciència bàsica, i pretenia valorar la hipòtesi que gensrelacionats amb Pdx1, que ha estat associat ambpatologies diabètiques, poguessin estar involucrats enla formació, el manteniment o el funcionament de lescèl·lules endocrines pancreàtiques, i en la regulaciófina de la transcripció del gen de la insulina. En cas deser certa, la hipòtesi, aquests gens serien directamentdiana d’estudis més fins sobre la seva funció, icandidats a responsables de diabetis tipus 2. Elsnostres resultats han estat molt positius, en el sentitque es tracta, segons el nostre coneixement, de laprimera recerca que relaciona alguns dels gensParaHox amb el pàncrees endocrí i amb la regulaciódel gen de la insulina.
Aquest treball, nova línia d’investigació del grup inicia-da expressament amb aquest ajut, hauria de tenir unacontinuació natural en l’aprofundiment de la funciód’aquests gens en pàncrees, i en l’estudi de lesassociacions amb patologies diabètiques. En elsupòsit que a mitjà-llarg termini aquests treballs oferis-sin la identificació d’alguns ParaHox com a factorsresponsables o de risc de desenvolupament dediabetis, la rellevància biomèdica i les implicacionsclíniques d’aquest treball, en termes de deteccióprecoç de risc, o d’identificació de noves dianesterapèutiques, serien més que considerables.
4. PublicacionsRosanas A, Marfany, G & Garcia-Fernàndez, J.
All six ParaHox-cluster genes are expressed in subsets of pancre-
atic endocrine cells. (sotmès a publicació).
Rosanas A, & Garcia-Fernàndez, J. & Marfany, G
Transactivation of the insulin gene promoter by ParaHox transcrip-
tion factors. (en preparació).

FUNDACIÓ LA MARATÓ DE TV3
MEMÒRIA FINAL
1. Resum del projecte originalPresentació de la hipòtesi i descripció delsobjectius concrets del projecte i dels beneficiscientífics i tècnics.El projecte intenta conèixer el paper de la carnitinapalmitoil transferasa I (CPT I) i del malonil-CoA en lasecreció d’insulina en la cèl·lula ß del pàncrees.Existeixen dades en la literatura científica que intentendemostrar que el malonil-CoA és la molècula clau enla transducció del senyal dins la cèl·lula ß del pàncre-es, de tal manera que la seva concentració decideix sila cèl·lula ß ha de secretar o no insulina. Els experi-ments que s’han dissenyat en aquest projecte volendemostrar que l’actuació del malonil-CoA sobre laCPT I és decisiva. Per això es localitzaran els aminoà-cids responsables de la interacció del malonil-CoAamb la CPT I, es mutagenitzaran adequadament i estransfectaran cèl·lules d’insulinoma per veure de quinaforma l’alteració en l’oxidació dels àcids grassos ésdeterminant en l’alliberament d’insulina. A més a més,s’intentarà l’obtenció d’animals transgènics en elsquals el transgèn sigui la CPT I control, o la CPT Iamb diverses mutacions en les què l’actuació del
malonil-CoA estigui impedida. D’aquesta forma, nosolament es farà palesa aquesta hipòtesi, sinó quetambé es disposarà d’un conjunt d’animals diabèticsque serviran per realitzar experiments que mostrin lainfluència de diverses dietes en l’alliberament d’insuli-na. Els objectius són:
1) Determinar per anàlisi computacional, els llocs de laCPT I tant en fetge com en múscul que interaccionenamb el malonil-CoA.
2) Determinar experimentalment si els llocs seleccio-nats que presumiblement interactuen amb el malonil-CoA, efectivament ho fan. Així doncs, mutagenitzaremaquests llocs canviant l’aminoàcid responsabled’aquesta unió per un aminoàcid no carregat (alaninao glutamina). Després sobreexpressarem els cDNAmutats en llevat (Saccharomyces cerevisiae o Pichiapastoris) i determinarem si la CPT I mutada mantél’activitat enzimàtica i si ha desaparegut la inhibició permalonil-CoA, comparant-ho amb l’expressió del cDNAde la CPT control. Per a això farem cinètiques d’inhibi-ció a concentracions creixents de malonil-CoA icreixents també del substrat, decanoil-CoA, per veureel seu efecte en els diversos mutants.
Títol del projecte
L’alteració del lloc d'unió del malonil-CoA a la carnitina palmitoiltransferasa com a agent causal de la diabetis mellitus
no depenent d'insulina (NIDDM)
Investigador principalFausto García Hegardt
InstitucióFacultat de Farmàcia. UB
Membres de l’equipDolors Serra, Laura Herrero, M. Irene Vázquez, Assia Bentebibel,
Guillermina Asins, Antonio García, David Sebastián i Guillem Plasencia.
76.735 €

3) Transfectarem cèl·lules d’insulinoma amb els cDNAmutats i determinarem la producció i l’alliberamentd’insulina al medi en presència de secretagogs, comara la glucosa, àcids grassos, arginina i leucina soleso en combinació, tot determinant que la concentracióde malonil-CoA format en cada cas, no afecta l’allibe-rament d’insulina. La hipòtesi es basa en el fet que sesecretarà menor proporció d’insulina en aquellescèl·lules transfectades en haver alterat l’oxidaciónatural dels àcids grassos.
4) Produirem animals transgènics portadors deltransgèn corresponent, que en cada cas pot ser elcDNA de la CPT I amb l’aminoàcid mutat núm. 1,amb l’aminoàcid mutat núm. 2, o amb els dosaminoàcids mutats a la vegada, i comparant-hosempre amb animals transgènics control. Atès queexisteixen dos gens CPT I, el d’expressió en el músculi el d’expressió en el fetge, i que el de múscul té unaafinitat vers el malonil-CoA molt superior al de fetge(IC50= x100), produirem també animals transgènicsamb mutacions en els mateixos aminoàcids del gende múscul. Si bé el gen de la CPT I que s’expressa enpàncrees és el de fetge, atesa la major repercussió delmalonil-CoA en el gen de múscul, pot ser de moltinterès veure l’efecte d’una desaparició dels àcidsgrassos de la cèl·lula ß del pàncrees amb una cinèticamajor que la que presenta la CPT I de fetge. Ladiabetis produïda pot ser més intensa. Es patentaranels ratolins transgènics obtinguts, que serviran: 1) perconfirmar el mecanisme transductor del senyal per lasecreció d’insulina en la cèl·lula ßdel pàncrees i 2) coma mitjà ideal per assajar nous fàrmacs antidiabètics.
5) S’assajaran els animals diabètics transgènicsobtinguts, mitjançant dietes riques en diversosefectors, com glucosa, manosa, galactosa, leucina,arginina i palmitat, per veure la influència que tenenaquests secretagogs en l’alliberament d’insulina i en elmillorament de la diabetis induïda per les mutacionsen la CPT I.
6) S’analitzarà com les diverses mutacions en el gende la CPT I influencien en l’obertura o el tancamentdel canal de potassi en la cèl·lula b del pàncrees, uncop induïda la secreció d’insulina per diversossecretagogs. Es definirà l’esdeveniment dels succes-sos dins la cèl·lula b segons que es pugui establiraquesta dependència o no.
7) S’assajaran fàrmacs antidiabètics en aquestsratolins. S’elegiran segons si el seu lloc d’acció és elde la unió del malonil-CoA a la CPT I o bé si actuen alcentre catalític. Entre els compostos que actuen alnivell del lloc d’unió del malonil-CoA, s’assajaranl’Ofenicine, el 4-hidroxifenilglicòlic i el Ro-0187; totsaquests no produiran cap benefici en els transgènicsamb mutacions selectives. Altres agents antidiabèticsque s’assajaran seran els compostos (+) de palmitoil- idecanoil- carnitines, atès que ja s’ha provat que sóneficaços fins i tot en rates diabètiques aloxanitzadescom a inhibidors competitius dels substrats. Altrescompostos seran els derivats de carnitina, coml’aminocarnitina, N-acetil aminocarnitina, i l’anàleg del’aminocarnitina cíclica SDZ 269-456. S’espera queactuant fora del lloc d’unió del malonil-CoA, els
ratolins transgènics siguin ben útils per provar labondat del projecte.
2. ResultatsEstabliment d’un model previ estructural tridimen-sional (200 aminoàcids)Un objectiu que en el curs de la nostra recercaconsideràvem de gran utilitat va ser l’elaboració demodels estructurals de la CPT I i de la COT (el seuhomòleg que catalitza un àcid gras més curt). Aixòens facilitaria l’obtenció de mutants dirigits per a lacomprovació experimental dels aminoàcids importantsper a la funció catalítica i la sensibilitat al malonil-CoA.
Per a l’obtenció d’aquests models estructurals vamusar dos programes de threading, el THREADER2 i elFUGUE, a fi de trobar motlles estructurals nosolament per a CPT I, sinó per a CPT II, COT, CAT icolinaferases d’humans i rosegadors. Vam escollirnomés 200 aminoàcids al voltant de l’aminoàcidconsiderat clau en l’activitat catalítica, la histidina 473,(coordenades de CPT I), els aminoàcids compresosentre 368 i 567. Vam obtenir un clar candidat, que vaser l’enoil-CoA hidratasa.
Vàrem fer un alineament estructural d’un conjunt decarnitina aciltransferases amb l’enoil-CoA hidratasa ivam trobar que tot i no ser homòlegs per raó del’escassa identitat dels aminoàcids, sí que ho són perl’anomenada homologia remota, que té més encompte l’organització de les estructures secundàriesque no les seqüències d’aminoàcids. En la figura esmostra l’alineament trobat.
D’acord amb aquest alineament, es van crear modelstridimensionals de CPT I i COT usant l’enoil-CoA coma motlle. L’avaluació d’aquestes estructures va serfeta usant com a procediments el WhatCheck, elProCheck i el Prosa II. Són sistemes matemàtics quefan una avaluació de la realitat i concordança entre elsmodels.
El resultats van ser adequats, sense anormalitatsrellevants. Una representació de la concordança delsmodels estructurals 3D es mostra en la figurasegüent. S’hi veu clarament que tant la CPT I com laCOT o l’enoil-CoA hidratasa tenen localitzacions al’espai anàlogues, tot posant de manifest que el
RECERCA BIOMÈDICA | 115

model que apareix per a la CPT I és molt correcte.Aquest model, a més a més, permet la localització delsubstrat palmitoil-CoA o decanoil-CoA, ja que elmodel cristal·litzat d’enoil-CoA hidratasa va serobtingut amb una molècula d’octanoil-CoA que ésinhibidor de l’acció d’aquest enzim i que dóna lacasualitat que també és substrat de la carnitinaoctanoil transferasa. Les estructures espacials de COTi CPT I es donen en la figura i semblen quasi idènti-ques, la qual cosa indica que totes dues deuen serprobablement correctes.
2dub (cadena E) 2dubE / CPT I / COT
D’altra banda, la representació permet localitzar el lloccatalític i els aminoàcids que queden a prop del’enllaç CO-S-CoA , tal com es mostra en la figura dela ribbon plot representation.
CPT I COT
La histidina de color blau està molt a prop de l’àtomde sofre del coenzim A. La figura mostra igualment unaltre aminoàcid que tot i estar a 92 aminoàcids dedistància d’aquesta histidina, està molt pròxim en elmodel estructural. Precisament es va mutar aquestaalanina i el resultat va ser la pèrdua d’activitat catalíticade tots dos enzims, cosa que consolida el model.
Mutagènesi de les histidines H287 i H493 i del’A238Utilitzant com a motlle el cDNA de la CPT I obtinguten un cribratge d’una genoteca de cDNA, hem mutatels nucleòtids que codifiquen per les histidines 287 i493. Hem utilitzat els oligonucleòtids mutats
anomenats CPTIH277A i CPTIH483A, amb lesseqüències 5’AGT ATG GCG TGG ATG GTG TTGCCA GC3’ i 5'GTG GGC CAT TTG TGG GAG 3’,respectivament, seguint els protocols de Datta et al.(3). Els fragments de DNA mutats, un cop comprova-da l’absència de mutacions no desitjades, van serinserits al plasmidi corresponent (RIP-ß-globina).
Anàlisi dels aminoàcids responsables de la inhibi-ció de la CPT I per malonil-CoALes anàlisis fetes abans havien mostrat que probable-ment els aminoàcids d’interacció amb el malonil-CoAeren histidines. Per comparació de les histidines dediverses carnitina aciltransferases CPT I, amb d’altres,com la COT, es va deduir que els aminoàcids corres-ponents als H131 i H340 de la COT (1,2) eren elscandidats més probables d’inhibició per unió almalonil-CoA. Això va fer que utilizéssim aquestsaminoàcids models com a responsables de la inhibi-ció. L’anàlisi per docking de la CPT I va mostrar que lamolècula malonil-CoA s’insereix en una posiciópropera al canal catalític del palmitoil-CoA. La figuramostra la representació que resulta de la ubicació detotes dues molècules, palmitoil-CoA i malonil-CoA, iper comparació no és possible posar el malonil-CoAni en la CPT II ni en la CAT.
Figura 1. Representació de la superfície accessible al solvent dels
models per a COT (A i B), L-CPT I (B i C), CPT II (E) i CAT (F). En
A es representa el model per a la COT al qual s’ha afegit la
molècula de malonil-CoA per càlculs de docking. En B es
representa la mateixa molècula de COT, però en aquest cas
també es mostra la molècula de substrat decanoil-CoA
juntament a la de malonil-CoA. En aquest cas, la COT s’ha rotat
per mostrar millor la posició de tots dos, substrat i inhibidor. En
C i D es representa el mateix, però en aquest cas la molècula
proteica és la CPT I. S’han representat alguns aminoàcids
importants en la catàlisi i en la inhibició. En E es representa la
CPT II i en F la CAT. Es pot comprovar que el malonil-CoA no es
pot allotjar al locus, fet representat per una el·lipse verda.
FUNDACIÓ LA MARATÓ DE TV3

Establiment d’un model molt més acurat basat enl’homologia amb el cristall de la carnitina acetil-transferasa d’humà i de ratolíTal com s’ha mostrat anteriorment, els modelsproposats van ser construïts usant com a motllel’enzim enoil-CoA hidratasa, que de manera empíricas’havia trobat que podia tenir un plegament semblantal de la CPT I i COT. Però sobtadament el panoramava canviar, amb la publicació del cristall de la carnitinaacetiltransferasa (CrAT) d’humà i de ratolí per dosgrups diferents. Usant metodologies bioinformàtiques,i usant com a motlle la CrAT humana, vàrem obtenir elnou model de CPT I per aplicació d’homologia directa.Totes les carnitina aciltransferases es pleguen d’unamanera semblant i nosaltres vàrem fer ús d’aquestapropietat per obtenir el nou model que va ser publicatal Biochemical Journal el 08.01.2004 en la versióelectrònica. Aquest model va ésser validat per lamutagènesi d’aminoàcids molt propers al centrecatalític i també per les mutacions que produïenmalalties hereditàries o bé estaven molt a prop delcentre catalític o donaven inestabilitat al conjunt.
Una aportació d’interès al tema va ser que vamlocalitzar el pou hidrofòbic on s’introdueix la cadenalateral hidrocarbonada dels àcids grassos. Vamobservar que al voltant dels aminoàcids propers alcentre catalític hi havia tres glicines mentre que enaltres carnitina aciltransferases que catalitzaven àcidsgrassos de cadena curta hi havia tres aminoàcidsvoluminosos valina, metionina i fenilalanina. La localit-zació de les tres glicines produïa una boca molt ampladel pou hidrofòbic, tot permetent l’entrada de lacadena hidrocarbonada dels acil-CoA fins al fonsd’una manera molt operativa.
Expressió i cinètiques de la CPT I enSaccharomyces cerevisiaeEl plasmidi pYESCPTwt va ésser expressat en S.cerevisiae. La preparació de CPT I obtinguda conteniala mateixa activitat que les mitocòndries de fetge.Fetes les determinacions de l’activitat de l’enzim enrelació amb els seus substrats, carnitina i palmitoil-CoA, es va concloure que els cDNA mutats de CPT Itenien una activitat semblant respecte de l’enzimcontrol. Així doncs, les mutacions introduïdes al cDNA
de la CPT I no mostraven canvis importants en l’activi-tat de l’enzim, i van ser bons sistemes per a ladeterminació de la inhibició per malonil-CoA.
Les corbes que mostraven la dependència d’activitatper augment de concentracions de malonil-CoA, vanser molt valuoses. Els valors IC50 d’inhibició van ser9, 15, 95 i més de 1.500 ?M de malonil-CoA, fet queva mostrar la repercussió de les mutacions fetes al’enzim. Més dades referents a la influència de lesmutacions en l’activitat catalítica i en la sensibilitat amalonil-CoA es donen en els articles que s’indiquen.
Localització d’altres aminoàcids (no histidines) quepoden determinar la sensibillitat al malonil-CoALes anàlisis bioinformàtiques van ser dirigides en unaaltra direcció. Independentment de si les histidineseren aminoàcids importants com a elements desensibilitat al malonil-CoA, vam estudiar els determi-nants que podien ser definitius en la pèrdua o elguany de sensibilitat al malonil-CoA. La hipòtesi eraque una sèrie de carnitina aciltransferases erensensibles al malonil-CoA (L-CPT I, M-CPT I i COT) id’altres no ho eren (CAT, CPT II i colina aciltransfera-sa). Amb aquestes dades l’anàlisi bioinformàtica ensva dir que hi havia 5 aminoàcids que estaven en elprimer grup d’enzims (Thr314, Asn464, Ala478,Met593 i Cys608), i que en canvi eren substituïts peraltres aminoàcids que eren idèntics al segon grupd’enzims (Ser223, Asp363, Gly377, Ser490 i Ala505,coordenades de CPT II).
Vam preparar fins a vuit enzims mutants. El primer,l’anomenat pentamutant, contenia les cinc mutacionsen la mateixa proteïna, elT314S/N464D/A478G/M593S/C608A; a més a mésvam preparar els cinc mutants individuals T314S,N464D, A478G, M593S i C608A. Per últim vampreparar dos mutants més tots produïts en la metioni-na 593, M593A i M593E. Van ser expressats enSaccharomyces cerevisiae i assajats per activitat i persensibilitat al malonil-CoA. Tots els mutants tenienactivitat com la proteïna salvatge, però els resultatsd’inhibició per malonil-CoA eren diferents: mentre queels dos primers mutants no modificaren la sevasensibilitat al malonil-CoA, el tercer i cinquè produïenpetites alteracions de sensibilitat.
Pel contrari, els mutants que duien la metionina 593mutada (el pentamutant i els mutants individualsM593S, M593A i M593E), mostraven gairebé unainsensibilitat total al malonil-CoA. Aquests resultatseren molt bons, ja que durant els darrers anys,diversos grups arreu del món estaven buscant aquestresultat. Més important encara era que aquest mutantpermetia fer experiments d’apoptosi i de secreciód’insulina estimulada por glucosa amb cèl·lules betatransfectades amb aquests mutants i així mostrarienque la teoria de la transducció de senyal modificadaper la incidència dels àcids grassos restaria enfortida.Els resultats van ser publicats en el Journal ofBiological Chemistry el 2003.
Secrecció d’insulina per la cèl·lula ß del pàncreesEs van construir adenovirus recombinants en els qualsel cDNA inserit va ser el cDNA de la L-CPT I. A
RECERCA BIOMÈDICA | 117

continuació, cèl·lules en cultiu d’insulinoma humà vanser infectades amb aquests adenovirus i es vadeterminar l’activitat CPT I i la secreció d’insulina encondicions diferents. Es van realitzar controlsd’expressió mitjançant anàlisi d’immunoblots. Tambées van fer determinacions de l’oxidació de glucosa ipalmitat mesurant l’alliberament de CO2 marcatradioactivament a partir d’aquests substrats radioactius.
Els resultats mostren que l’oxidació de substrats ésdependent de l’expressió de la CPT I. Si les cèl·lulessón incubades amb glucosa, l’oxidació del palmitatminva. Igualment, quan les cèl·lules s’incuben ambetomoxir, l’oxidació de palmitat disminueix. Aquestsexperiments mostren que el sistema funciona tal coms’esperava. La determinació de la secreció d’insulinamostra a més a més que la sobreexpressió del cDNAde la CPT I redueix la secreció d’insulina. A 15 mMglucosa, en absència d’etomoxir, es produeix unadisminució del 40% en la secreció de l’hormona.
Obtenció dels plasmidis RIP-ß-globina-CPT I wt iRIP-ß-globina-CPT I doble mutant per a la produc-ció de ratolins transgènicsEl plasmidi RIP-ß-globina conté 567 nucleòtids (basescorresponents al promotor I de la insulina [RIP I] i alseu extrem més 5’ on es troben totes les seqüènciesque permeten l’expressió regulada i teixit-específicad’aquest gen). També conté part del gen de la ß-globina, des de la diana BamHI present en el segonexó de la ß-globina, seguit del segon intró i el tercerexó a més a més dels senyals de poliadenilació iseqüències a l’extrem 3’ no transcrites que es perllon-guen fins a 3000 pb (vegeu figura 2).
Fig. 2 Representació esquemàtica del gen RIP I-CPT I ß-globina.
El fragment XhoI/BamHI (-570 bp a +3 bp) del RIP I va ser lligat al
fragment BamHI/NotI del gen de la ß-globina. El fragment EcoRV
(+55 bp a +2700 bp) del cDNA de la L-CPT I fou introduït en el
lloc EcoRV del tercer exó del gen de la ß-globina. El fragment
XhoI/NotI de 5,4 kilobases que contenia el gen quimèric complet
va ser utilitzat per obtenir els ratolins transgènics. El triangle ( D )
representa el senyal de poliadenilació del gen de la ß-globina.
Aquest plasmidi conté una diana única EcoRV, queinterromp el tercer exó de la ?-globina i en la qual espot inserir qualsevol cDNA, de forma que la sevaexpressió en animals transgènics produirà transcritsestables, poliadenilats, sota el control del RIP I.Aquest plasmidi ha estat anteriorment utilitzat pelnostre grup de treball per a altres construccions ambmolt bons rendiments.
Els DNA amb les seqüències codificants de CPT I nts55 a 2700, tant la forma wild-type (wt) com la deldoble mutant (H277A/H483A) un cop fets roms ambl’enzim T4 DNA polimerasa, van ser inserits en ladiana EcoRV feta roma prèviament amb el mateixenzim. Es van comprovar les mutacions i la correcta
orientació. Els plasmidis de CPT I van ser digerits ambdos enzims, NotI i XhoI, la qual cosa va permetreobtenir un fragment de DNA, aprox. 5400 nts, que uncop purificat va ser utilitzat pel grup d’investigació quedirigeix la Dra F. Bosch a la Facultat de Veterinària dela Universitat Autònoma de Barcelona.
Producció d’animals transgènics portadors de la L-CPTI mutada i controlPer tècniques clàssiques d’obtenció d’animalstransgènics, es van produir dos tipus d’animalssegons el transgèn introduït: transgèn wt i el dobletransgèn mutat. En tots dos casos l’expressió del genva ser feta sota el control de RIP-I que dirigeixl’expressió específicament al pàncrees. El gen de la L-CPT I va ser introduït abans que el gen de la ß-globinaper assegurar l’expressió del transgèn. La produccióde ratolins transgènics va ser feta al Servei de RatolinsTransgènics de la Facultat de Veterinària de la UAB.
Per estudiar la sobreexpressió de la L-CPT I wt van serobtinguts sis animals founders i van ser encreuats ambanimals control. No es va observar expressió en capde les sis línies ni per experiments de RT-PCR ni perNorthern. Paral·lelament, es va injectar el transgèn deldoble mutant, però no es va poder obtenir cap founderdesprés d’un gran nombre d’animals analitzats.
Figura 3. Anàlisi per PCR de la F1 d’una de les línies trasngèni-
ques de la construcció CPT I no mutada. La presència del
transgèn RIP I-LCPT I en el DNA genòmic de les mostres de la
cua de ratolí va ser analitzada per PCR. L’amplificació de 1000 pb
amb els oligos RIPI.for i L-CPT I180.rev va mostrar la presència
del transgèn. L’amplificació amb els oligos COT428.for i XbaI.rev
produí una banda del gen de la COT de 400 pb que va ser utilit-
zada com un control intern de la PCR i de la qualitat del DNA
genòmic obtingut. Els carrils 1 i 2 representen animals control i els
carrils 4, 5 i 6, animals dels quals es vol conèixer si són transgè-
nics. El carril 3 és el control negatiu. El marcador de massa
molecular dels DNA es representa amb una M. Les mostres van
ser separades en un gel d’agarosa del 4,1%.
Detecció dels animals que han inserit el transgènEls founders, animals que han inserit el transgèn,s’han localitzat per Southern blot utilitzant una sondaobtinguda a partir del fragment microinjectat i tambéper PCR utilitzant dos oligonucleòtids que reconeixenel promotor de la insulina: RIP I.for (5’ GCA CTA GGCAAG TGT TTG G3’) y L-CPT I180.rev (5’TTC GTGGCT CAG GCG GAG GTC3’). S’ha seguit el protocolclàssic d’obtenció de DNA genòmic a partir d’un trosseccionat de la cua dels ratolins. En la figura 3observem un PCR representatiu.
FUNDACIÓ LA MARATÓ DE TV3

Analisi de l’expressió del transgèn en les diverseslíniesS’han obtingut 6 línies d’animals transgènics quecontenien el cDNA no mutat. No s’ha detectat cap deldoble mutant, havent-se microinjectat fins i tot més de100 oòcits. Els fundadors de les sis línies s’hanencreuat amb animals control per obtenir la F1, idesprés les generacions següents. Es va poder obtenirdescendència de només 5 de les línies de ratolins.S’han analitzat per PCR els animals de F1 qu
Fig. 4. Anàlisi per RT-PCR de l’expressió del transgèn en les 6
línies d’animals transgènics
Es va obtenir l’RNA total del pàncrees dels animals transgènics,
F1, de les 5 línies que van tenir descendència (carrils 2-7) i de
diversos teixits de la línia núm. 8 que no va tenir descendència
(carrils 8-21). En el PCR es van utilitzar dues parelles d’oligonucle-
òtids: una parella com a control intern que amplificava el gen de la
HMG-CoA sintasa i que produïa una banda de 300 pb, i un parell
d’oligolucleòtids específics per a l’expressió del transgèn, que
produïa una banda de 900 pb. En el carril 1 es mostra el control
negatiu. Dels carrils 8 al 21 es mostren els resultats del founder
núm. 8, de teixits diferents (fetge, testicle, cervell, múscul, melsa,
ronyó, pulmó, cor, teixit adipós blanc i intestí prim). El carril 22
mostra el resultat obtingut en analitzar el DNA genòmic d’un
animal control, per la qual cosa no apareix la banda (control
negatiu de PCR). En el carril 23 s’analitza el DNA genòmic d’un
animal transgènic (control positiu de PCR). En el carril 24 s’analit-
za el plasmidi microinjectat (control positiu de PCR).
van resultar transgènics, tot observant-se que enmolts casos eren mosaics, ja que el tant per cent detransgènics en la seva descendència era molt baix. Detots ells es va obtenir l’RNA del pàncrees i es vananalitzar per Northern blot i per RT-PCR, tècnica méssensible que ens permetia detectar nivells molt baixosd’expressió. En cap cas els animals van expressar eltransgèn, per la qual cosa no ens van servir per alsnostres objectius.En la figura 4 es mostra un RT-PCR representatiu. Elmascle, anomenat núm. 8, no ha tingut descendèn-cia, per això no hem pogut analitzar l’expressió deltransgèn en la seva descendència i ha calgut sacrifi-car-lo per observar si s’expressava el transgèn.
Test de tolerància a la glucosaEs va administrar 1 mg de glucosa per gram de pesde l’animal en dejú de 24 hores mitjançant unainjecció intraperitoneal. A diferents temps, es vanmesurar els nivells de glucosa en sang mitjançant elsistema Glucotrend de Boehringer Mannheim (ref.1623427).
Com s’observa en la figura 5, els controls segueixen
una pauta normal de tolerància a la glucosa, totrecuperant-se a les dues hores de l’administració.Tots els animals transgènics, excepte el núm. 8,també segueixen aquest comportament. L’animal 8mostra una tolerància a la glucosa, típica d’animalsdiabètics. A més a més, aquest animal tenia un pesun 45% superior a la mitjana dels animals de lamateixa edat i sexe. No obstant això, quan aquestanimal va ser analitzat (vegeu figura 3), no expressavatampoc el transgèn, ni en pàncrees, que és onesperàvem l’expressió ni tampoc en gònades, fet quehagués explicat la seva infertilitat. Per tant, vamconcloure que la inserció del transgèn en un llocinadequat va provocar l’esterilitat, el sobrepès i lacorba anòmala de tolerància a la glucosa, però queen cap cas allò va ser causat per l’expressió ectòpicade la CPT I.
Fig. 5. Test de tolerància a la glucosa de les 6 línies d’animals
transgènics i de 4 controls.
Es va administrar 1 mg de glucosa per gram de pes als diversos
animals i es van analitzar els nivells de glucosa en sang als temps
0, 30, 60, 90 i 120 minuts. Els animals transgènics són els
numerats com 8, 10.12, 13.2, 18.14 i 27.2. Els animals controls
s’han numerat com femella control 1 i 2, i mascle control 1 i 2.
L’animal 36 correspon a un possible transgènic doble mutant que
després no es va confirmar per Southern blot.
Estudis amb cèl·lules en cultiuPer a l’estudi de l’expressió de l’antisense de la L-CPTI en les cèl·lules INS-1E calia comprovar que elmodel cel·lular elegit era adequat.
Figura 6 Oxidació de glucosa en INS-1E.
Es va mesurar la producció de 14CO2 (6). Escomprovà que en augmentar la concentració deglucosa en el medi, l’oxidació de glucosa per les INS-1E també augmenta.
Oxidació de la glucosaL’oxidació de la glucosa es va mesurar com laproducció de 14CO2 després d’incubar les cèl·lulesINS-1E durant 1 h amb [U-14C]-glucosa en absènciad’àcids grassos (4). La figura mostra que les cèl·lulesINS-1E utilitzades tenen una oxidació de glucosa
RECERCA BIOMÈDICA | 119

FUNDACIÓ LA MARATÓ DE TV3
normal que augmenta en incrementar-se la concentra-ció de glucosa en el medi.
Oxidació de palmitatL’oxidació de palmitat es va mesurar com la produc-ció de 14CO2 després d’incubar les cèl·lules INS-1Edurant 1 h amb [1-14C]-àcid palmític (5), en presènciao no de 200 mM d’etomoxir, que és un inhibidorirreversible de CPTI. Això mostra que l’oxidació depalmitat disminueix en augmentar la concentració deglucosa i també en presència d’etomoxir que inhibeixla CPTI i, per tant, la ß-oxidació dels àcids grassos.
Fig. 7 Oxidació de palmitat en INS-1E.
Es va mesurar la producció de 14CO2 (5). Es comprova que en
augmentar la concentració de glucosa o en presència d’etomoxir,
inhibidor de la CPTI, l’oxidació de palmitat minva.
Construcció dels adenovirusL’adenovirus recombinant AdCA-L-CPTIas (antisense)va ser construït com es descriu en Miyake, 1996 (6).El AdCA-LacZ, utilitzat com a control, va ser cedit perIshihara (4).
El cDNA de L-CPTI subclonat en pBlueScript va serdigerit, fet rom i inserit en el lloc únic SwaI d’E1 delgenoma de l’adenovirus clonat en un còsmid. Aquestacasset amb L-CPTIas va ser cotransfectat en cèl·lulesHEK293 juntament amb el complex proteic de DNA-terminal de l’adenovirus digerit amb EcoT22I.L’adenovirus recombinant va ser generat per recombi-nació homòloga en les cèl·lules esmentades (vegeu lafigura). Els virus AdCA-L-CPTIas i AdCA-LacZ van seramplificats i purificats per ultracentrifugació amb CsCl.La titulació dels adenovirus es va dur a terme d’acordamb el protocol de Takara Biomedicals (ref. 6150).
3. Rellevància i possibles implicacions clíniquesdels resultats finals obtingutsEl concepte de glucolipotoxicitat està progressivamentemergent i vol descriure l’efecte d’àcids grassos iglucosa en la secreció d’insulina per la cèl·lula betadel pàncrees i en l’efecte d’aquests metabòlits en lamort de la cèl·lula beta del pàncrees. La nostra aproxi-mació ha consistit a confirmar aquesta teoria que témolt a veure amb el caràcter que relaciona l’obesitatde moltes persones i la diabetis tipus II (no dependentd’insulina).
Hem fet troballes de molt d’interès. En primer lloc,que la sobreexpressió de l’enzim CPT I, que és elregulador de l’oxidació d’àcids grassos en la mitocòn-dria, produeix una disminució de la secreció d’insulina.Per una altra banda, la sobreexpressió produeix unimportant augment de l’oxidació dels àcids grassos dela cèl·lula beta, fet posat també de manifest perquè lapresencia d’etomoxir (un inhibidor de la CPT I) produeix
una disminució de l’oxidació dels àcids grassos i unaugment de la secreció d’insulina en comparació ambel control de L-CPT I sobreexpressada.
Paral·lelament, l’obtenció de models de la CPT I, CPTII i CAT ens ha permès saber com els grups acil-CoAcom a substrats i el malonil-CoA com a inhibidorafecten l’estructura de la molècula.
Aquests fets també permeten conèixer millor lapossibilitat d’elaborar inhibidors farmacològics quepoden interactuar amb la molècula de CPT I perinhibir-la tant en la cèl·lula beta com al cor. Unaestratègia farmacològica s’està posant en marxa enmolts grups americans de cardiologia, per tal d’inhibirla CPT I amb inhibidors, com ara el tetradecil-glicil-CoA o etomoxir dins del cor per inhibir l’oxidaciód’àcids grassos i obligar el cor a oxidar glucosa, ambla consegüent baixada de concentracions i de millorcapacitat contràctil del mateix cor.
4. PublicacionsMiliar, A., Serra, D., Casaroli, R., Vilaró, S., Asins, G. i Hegardt,
F.G. Developmental changes in Carnitine octanoyltransferase gene
expression in intestine and liver of suckling rats Archives of
Biochemistry and Biophysics (2001) 385, 283-289
Nicot, C., Hegardt, F.G., Woldegiorgis, G., Haro, D i Marrero,
P.F. Pig liver carnitine palmitoyltransferase I, with low Km for
carnitine and high sensitivity to malonyl-CoA inhibition is a natural
chimera of rat liver and muscle enzymes Biochemistry (2001) 40,
2260-2266
Hegardt, F.G., Bach, M., Asins, G., Caudevilla, C. Morillas, M.,
Codony, C. i Serra, D. Post-transcriptional regulation of rat carniti-
ne octanoyltransferase Biochemical Society Transactions (2001)
29 316-320
Morillas, M., Gómez-Puertas, P., Roca, R., Serra, D., Asins, G.,
Valencia, A. i Hegardt, F.G. Structural model of the catalytic core
of carnitine palmitoyltransferase I and carnitine octanoyltransferase
(COT) Journal of Biological Chemistry (2001) 276, 45001-45008
Rubí, B., Antinozzi, P. Herrero, L., Ishihara, H., Asins, G., Serra,
D., Wollheim, C. W. i Hegardt, F.G. Adenovirus-mediated overex-
pression of liver carnitine palmitoyl transferase i in ins-1 cells.
effects on cell metabolism and insulin secretion Biochemical
Journal (2002) 364, 219-226
Morillas, M., Gómez-Puertas, P., Rubí, B., Clotet, J., Ariño, J.,
Valencia, A., Hegardt, F.G., Serra, D. y Asins, G. Structural model
of a malonyl-CoA binding site of carnitine octanoyltransferase and
carnitine palmitoyltransferase I. Mutational analysis of a malonyl-
CoA affinity domain Journal of Biological Chemistry (2002) 277,
11473-11480
Morillas, M., Gómez-Puertas, P., Bentebibel, A., Sellés, E.,
Casals, N., Valencia, A., Hegardt, F.G., Asins, G. y Serra, D.
Identification of conserved amino acid residues in rat liver carnitine
palmitoyltransferase I critical for malonyl-CoA inhibition Journal of
Biological Chemistry (2003) 278, 9058-9063
Morillas, M., López-Viñas, E., Valencia, A., Serra, D., Gómez-
Puertas, P., Hegardt, F.G. y Asins, G. Structural model of carnitine
palmitoyl transferase I based on the carnitine acetyltransferase
crystal Biochemical Journal (2004) Publicado el 8 de enero de
2004 en versión electrónica 10.1042/BJ20031373

RECERCA BIOMÈDICA | 121
MEMÒRIA FINAL
1. Resum del projecte originalObjectiusObjectiu general:Comparar l’absentisme laboral entre els diabètics ambun bon control de la malaltia i la població general ambhistòria clínica oberta als centres d’atenció primària.Objectius específics:
Descriure els episodis d’incapacitat temporal (IT) delstreballadors diabètics i no diabètics segons edat,gènere, diagnòstic, ocupació, condicions de treball iduració.
Analitzar el paper que tenen en l’absentisme laboraldels treballadors diabètics, l’evolució i el control de lamalaltia, l’aparició de complicacions i l’educació.
DissenyEs farà un estudi de cohorts històriques.
Població i àmbit de l’estudiLa població d’estudi són uns 409.137 treballadorsocupats entre 16 i 65 anys amb història clínica obertaals centres d’atenció primària de la Regió Sanitària
Costa de Ponent. El nombre d’individus inclosos serà1.247 persones diabètiques i 1.247 no diabètiques,tot assumint un error del 5% i una potència del 80%.
Pla de treballElaboració del protocol de treball.
Adaptació del model de qüestionari i ensinistramentde les persones encarregades de l’administració delsqüestionaris.
Realització de les sessions d’informació als equipsd’atenció primària de les diverses àrees bàsiques desalut.
Realització de les enquestes en les àrees bàsiques desalut, recollida d’informació i processament informàticde les dades.Anàlisi dels resultats. Elaboració dels resultatsobtinguts i redacció del document final de l’estudi.
2. Resultats1. Resultats generalsEn l’estudi es van incloure 1.113 pacients diabètics i1.320 individus de la població general que reuniencriteris d’inclusió. De les variables sociodemogràfiques
Títol del projecte
Diabetis i treball. Anàlisi de la influència del control de la malaltia i de les condicions de treball en l'absentisme laboral de les persones diabètiques
Investigadors principalsJoan Gené Badia
InstitucióDAP Baix Llobregat Litoral. Fundació Jordi Gol i Gurina
Membres de l’equipÀngels Company, Javier Funes, Oriol Fàbrega i Jordi Castejón.
28.825 €

de les poblacions estudiades, destaca un predominidels homes, tant en el grup de la població diabètica(76%) com en la població general (82%) (p = 0,001).La mitjana d’edat ha estat de 50,9 anys (DE 9,7 anys)i de 49,7 anys (DE 9,5 anys), respectivament (IC 95%50,4 — 51,5; 49,23 — 50,25; p = 0,02).
L’anàlisi de les característiques sociolaborals mostraque existien diferències estadísticament significativesen la distribució de l’ocupació en la població diabèticai en la població general, (p = 0,001), amb un predomi-ni dels treballadors de la indústria en ambdós grups(34,8%; 37,0%). També destacava en la poblaciódiabètica una més gran proporció de treballadors noqualificats (17,5%; 12,7%), i una menor proporciód’empleats administratius (7,8%; 11,2%). Els sectorseconòmics majoritaris en ambdós grups van ser elcomerç, venda i reparació de vehicles de motor(14,6%; 12,6%), la construcció (10,1%; 11,7%) i eltransport i comunicació (10,1%; 9,9%).
Es va observar que existien diferències estadística-ment significatives en comparar la grandària de lesempreses. Així, els treballadors diabètics treballavenen empreses de menys de 9 treballadors (32,5%;28,2%), mentre que els treballadors de la poblaciógeneral estaven ocupats en empreses de més de 500treballadors (19,9%; 26,5%) (p = 0,003). De la restade variables sociolaborals estudiades, destacava queel 76,2% dels pacients diabètics i el 74,3% de lapoblació general tenien una antiguitat en la sevaocupació igual o superior als 10 anys, sent la mitjanade 20,8 anys (DE 13 anys) (IC 95% 20,0- 21,6) i de20 anys (DE 12,4 anys) (IC 95% 19,3 — 20,7),respectivament; la mitjana d’antiguitat a l’empresa erade 16,9 anys (DE12 anys) (IC 95% 16,2 — 17,6) i de16,8 anys (DE11,9 anys) (IC 95% 16,2 — 17,4); i queel 37,3% dels treballadors diabètics i el 31,6% de lapoblació general treballaven més de 40 hores a lasetmana, sent la mitjana d’hores treballades a lasetmana de 43,7 (DE 11,7) (IC 95% 43,0 — 44,4) i de42,4 (DE 9,1) (IC 95% 42,0 — 43,0), respectivament,tot existint diferències estadísticament significativesentre ambdós grups (p = 0,003). També es vaobservar que ambdues poblacions tenien majoritària-ment un horari de treball fix diürn (81,9%; 81,0%). Encomparar la distribució de les poblacions en relacióamb el tipus de contracte de treball, es va observarque en la població diabètica un 12,8% (8% en lapoblació general) tenien contracte temporal i que el21,4% eren treballadors autònoms (18,0% en lapoblació general) (p>0,00005).
De l’anàlisi dels factors de risc percebuts pels treballa-dors relacionats amb les condicions ambientals,ergonòmiques i organitzatives, destaca que el 29,2%dels treballadors diabètics i el 30,0% de la poblaciógeneral referia exposició a soroll, el 26,3% i el 27,9%,respectivament, exposició a fums, pols, o fibres al’aire, i el 25,8% i el 25,3% utilitzaven substànciesperilloses a la feina. Es van observar diferènciesestadísticament significatives entre ambdós grups enrelació amb l’exposició a baixes temperatures (5,4%;3,3%; p = 0,03), una il·luminació inadequada (13,1%;9,7%; p = 0,04), i la possibilitat d’obrir les finestres(37,0%; 50,9%; p = 0,001). Quan es va analitzar el
grup de treballadors exposats a substàncies perillo-ses, es va observar l’existència de diferències estadís-ticament significatives entre el grup de treballadorsdiabètics i la població general en l’exposició a metalls(10,0%; 4,7%; p = 0,02), dissolvents (30,0%; 47,9%;p = 0,00001), i altres substàncies químiques (37,0%;50,9%; p = 0,001).
En relació amb les condicions ergonòmiques, s’obser-và que el 74,9% dels treballadors diabètics i el 71,7%de la població general realitzava moviments repetitius,el 69% i el 70,3% treballaven dempeus la major partde la jornada laboral, i que el 38,5% i el 40,3%adoptava postures incòmodes. De les condicionsorganitzatives, destaca que el 53,7% dels treballadorsdiabètics i el 55,4% de la població general treballavaamb persones alienes a l’empresa.
Pel que fa a l’exposició a factors de risc psicosocial,podem observar que el 50,3% dels pacients diabèticsi el 52,5% de la població general presentaven unademanda de treball alta, el 45,3% i el 47,2% uncontrol baix, i un 44,3% i un 58%, respectivament, unsuport social baix. En relació amb el suport social, esvan trobar diferències estadísticament significatives (p= 0,00001) entre ambdós grups. El risc psicosocial,és a dir, aquell amb alta demanda, baix control i baixsuport social, va estar present en l’11,3% delstreballadors diabètics i en el 14,2% de la poblaciógeneral (p = 0,004).
L’anàlisi de les variables relacionades amb la diabetisva mostrar que l’evolució de la malaltia de la poblacióestudiada va ser de 6,7anys de mitjana (DE 6,5) (IC95% 5,31 — 7,1) i que són els equips metge/ssainfermer/a d’atenció primària els que fan el control dela malaltia (68,40%). En relació amb l’educació sanità-ria, els casos estudiats van manifestar haver rebutinformació referida a la malaltia (80, 2%), sobre comrealitzar el control (91%), i com prevenir les complica-cions (79,9%). En relació al tipus de tractament queutilitzaven per al control de la diabetis, es va observarque el 38,1% seguien una pauta terapèutica ambantidiabètics orals i dieta, sent el 23,2% els que feientractament amb insulina i dieta. Per a aquells que estractaven amb insulina, la mitjana d’anys d’insulinitza-ció va ser de 7,34 (DE 8,27 anys) (IC 95% 6,35 —8,32). El 73,85% es practicaven anàlisi de glucèmiacapil·lar.
L’anàlisi de les variables referides al grau de controlmetabòlic va mostrar que el 31,7% dels treballadorsdiabètics tenien un bon control metabòlic, mentre queen el 35,2% no es van disposar de dades quepermetessin avaluar el seu grau de control metabòlic.
En relació amb la presència de complicacions de lamalaltia, es va observar que existia informació clínicaen menys del 40% dels casos estudiats. Es vadetectar retinopatia (7,3%), macroangiopatia (6,4%),alteracions renals (alteracions analítiques 9,0%,nefropatia 0,63%), alteracions vasculars perifèriques(4,4%) i neuropatia (2%).
De l’anàlisi de les variables relacionades amb elmaneig de la diabetis en el medi laboral, va destacar
FUNDACIÓ LA MARATÓ DE TV3

que el 79,5% dels treballadors diabètics podienadaptar les seves ingestes a l’horari laboral, sent eltipus de feina (46,5%) la principal causa per nomantenir un horari d’àpats fix. També es va observarque el 18,8% dels treballadors diabètics estudiatstreballaven en empreses que tenien menjadorscol·lectius, els quals eren utilitzats pel 46% d’aqueststreballadors. El 89,65% podia triar un menú ques’adaptava a la seva diabetis.
El 54,5% dels treballadors diabètics insulinodepen-dents (n = 288) s’administraven la insulina durantl’horari de treball, dels quals el 87,9% disposava detemps suficient per al control i l’administració de lainsulina. No tenir pauses (52,63%) va ser el motiuprincipal per no tenir temps suficient per al control il’administració de la insulina. El lloc més freqüent peral control i l’administració de la insulina va ser: elslavabos (34,4%). En el 73,24% dels casos el llocreunia les condicions higièniques necessàries i el91,72% disposava de suficient intimitat.
En relació amb les variables que fan referència al graude coneixement de la malaltia entre els companys, esva observar que en el 72,5% dels casos els seuscompanys coneixien que eren diabètics, sent el princi-pal motiu per no comunicar-ho el fet de preservar laintimitat (25,8%). En el 60,5% de los casos els seuscaps coneixien la, malaltia, sent també el fet depreservar la seva intimitat (13,8%) la causa principalper no comunicar-ho.
Les dades referides a la freqüentació sanitària mostrenque la mitjana de visites al metge de capçalera de lapoblació diabètica (4,24 visites, DE 4,26; IC 95% 4 —4,5) respecte de la població general (2,52 DE 2,51; IC95% 2,33 — 2,7) van ser estadísticament significati-ves (p <0,0005). També es van comportar de manerasimilar les dades referides a la consulta d’infermeria(3,1 visites, DE 3,3; IC 95% 2,8 — 3,3; 0,6 DE 1,54;IC 95% 0,5 — 0,7 p <0,0005).
Els treballadors diabètics van presentar una taxad’incidència d’incapacitat temporal del 27,3%(n=304), sent del 17,4% (n=230) per a la poblaciógeneral.
Es van observar diferències estadísticament significati-ves entre el grup de treballadors diabètics i el de lapoblació general en relació amb el percentatge detreballadors que van presentar almenys un episodid’incapacitat temporal durant el període d’estudi (p<0,001). La mitjana d’episodis d’incapacitat temporalva ser de 0,3 i de 0,2, i la durada dels episodis de16,7 dies i de 9,5 dies, respectivament. Per compararel nombre d’episodis d’incapacitat temporal entre elstreballadors diabètics i la població general es va utilit-zar la prova U Mann-Whitney, que va donar diferèn-cies estadísticament significatives (p< 0,001). Tambées va utilitzar aquesta prova estadística per compararentre ambdues poblacions el nombre de dies d’inca-pacitat temporal, i es van trobar diferències estadísti-cament significatives (p= 0,001). La distribució delsepisodis d’incapacitat temporal segons les categoriesdiagnòstiques que els van ocasionar mostra que esvan observar diferències estadísticament significatives
en el grup de malalties endocrines nutricionals,metabòliques i trastorns de la immunitat (6,2%, IC95% 3,8 — 6,8; 0,36%, IC 95% 0 — 1,1; p <0,0001), els trastorns mentals (3,6%, IC 95% 1,8 —5,5; 7,5%, IC 95% 4,3 — 10,7; p < 0,05), i en lesmalalties de l’aparell digestiu (3,9%, IC 95% 1,9 —5,8; 9,0%, IC 95% 5,6 — 12,5; p < 0,01). També esva trobar significació estadística en el grup del qual noconstava el diagnòstic (11,6%, IC 95% 8,4 — 14,8;5,6%, IC 95% 2,9 — 8,4; p < 0,01).
2. Anàlisi de la població diabètica i no diabèticaamb un episodi o més d’incapacitat temporalL’anàlisi de les dades referides al grup de treballadorsdiabètics i de la població general que van presentaralmenys 1 episodi d’incapacitat temporal va mostrarun predomini d’homes tant en el grup de la poblaciódiabètica (77%) com en el de la població general(79,6%), amb una mitjana d’edat de 51,5 anys (DE9,6 anys) (IC 95% 50,4 — 52,5) i de 50,8 anys (DE9,2 anys) (IC 95% 49,8 — 52,2), respectivament.
Les característiques sociolaborals mostraven que enrelació amb l’ocupació existia un predomini detreballadors de la indústria (36,5% població diabètica,42,2% de la població general) seguit dels treballadorsno qualificats (20,1%; 37,1%, respectivament). Elssectors econòmics que ocupaven majoritàriamentambdós grups van ser el comerç, venda i reparacióde vehicles de motor (12,2%; 11,4%), i el transport i lacomunicació (11,6%; 10,0%). El 25% dels treballadorsdiabètics i el 30% de la població general treballavenen empreses de més de 500 treballadors, i el 24% i el22,2%, respectivament, en empreses petites d’entre 1i 9 treballadors. De la resta de variables sociolaboralsestudiades, destaca que el 76,6% dels pacientsdiabètics i el 75,2% de la població general tenien unaantiguitat en la seva ocupació superior o igual als 10anys, sent la mitjana de 20,3 anys (DE 13 anys) (IC95% 18,8- 21,8), i de 20,8 anys (DE 12,7 anys) (IC95% 19,1 — 22,4), respectivament; l’antiguitat mitjanaa l’empresa va ser de 17,3 anys (DE 11,9 anys) (IC95% 15,9- 18,6) i de 17,2 anys (DE 11,6 anys) (IC95% 15,7 — 18,7), respectivament; i que el 28,6%dels treballadors diabètics i el 25,7% de la poblaciógeneral treballaven més de 40 hores a la setmana,sent la mitjana d’hores treballades a la setmana de43,2 (DE 10) (IC 95% 42,1 — 44,4) i de 42,1 (DE 7,3)(IC 95% 41,2 — 43,1), respectivament. També es vaobservar que ambdues poblacions tenien majoritària-ment un horari de treball fix diürn (77%; 79,1%), i queel 73,3% dels treballadors diabètics i el 78,3% de lapoblació general tenien un contracte de treball indefi-nit, mentre que 13,5% i el 6,9%, respectivament,tenien un contracte de treball temporal.
L’anàlisi de les condicions de treball a partir delsfactors de risc percebut mostrava, en l’apartat decondicions estructurals i ambientals, que el 33,9%dels treballadors diabètics i el 36,5% de la poblaciógeneral referia exposició a soroll, el 27,3% i el 34,3%,respectivament, exposició a fums, pols, o fibres al’aire, i el 27% i el 30,4% utilitzaven substànciesperilloses a la feina. Quan es va analitzar el grup detreballadors exposats a substàncies perilloses, es vaobservar que el 26,3% dels treballadors diabètics i el
RECERCA BIOMÈDICA | 123

41,4% de la població general referien exposició adissolvents. En relació amb l’exposició a altressubstàncies químiques, van haver-hi diferènciesestadísticament significatives entre ambdós grups(32,9%; 54,3%; p = 0,03).
De l’apartat de condicions ergonòmiques, va destacarque el 78,9% dels treballadors diabètics i el 77,7% dela població general realitzava moviments repetitius, el68,8% i el 74,8% treballaven dempeus la major partde la jornada laboral, i que el 45,4% i el 50,4%adoptava postures incòmodes. Les condicionsorganitzaves van mostrar que el 47% dels treballadorsdiabètics i el 49,1% de la població general treballavaamb persones alienes a l’empresa.
Les dades referides a l’exposició a factors de risc detipus psicosocial mostra que el 54,6% dels pacientsdiabètics i el 58,7% de la població general presenta-ven una demanda de treball alta, el 49,3% i el 46,3%un control baix, i el 47,1% i el 59%, respectivament,un suport social baix, sent aquestes diferènciesestadísticament significatives (p = 0,007). El càlcul delrisc psicosocial va mostrar que el 13,8% dels pacientsdiabètics i el 19,1% de la població general presenta-ven una situació de risc, és a dir, alta demanda, baixcontrol i baix suport social.
3. Anàlisi de la influència de l’evolució i control dela malaltia, l’aparició de complicacions i l’educaciósanitària sobre l’absentisme laboral dels treballa-dors diabèticsPer analitzar el paper, en l’absentisme laboral (presèn-cia o absència d’incapacitat temporal durant elperíode d’estudi) dels treballadors diabètics, quehavien tingut l’evolució i el control de la malaltia,l’aparició de complicacions i l’educació sanitària, es vafer una anàlisi bivariable que va mostrar diferènciesestadísticament significatives en relació amb el controlmetabòlic (p = 0,005), la presència de macroangiopa-tia (p< 0,0005), i la presència de patologia nefrològica(p= 0,02) entre el grup que no va presentar incapacitattemporal i el que va presentar incapacitat temporal.
Posteriorment es va efectuar una anàlisi multivariant del’absentisme laboral segons un model de regressiólogística amb el mètode d’introducció de variablesEnter que va incloure les variables amb significacióestadística en l’anàlisi bivariable, el nombre de visites il’edat i el sexe. Segons aquest model, es va observarassociació estadísticament significativa entre el nombrede visites i la presència d’incapacitat temporal (OR:1,14 per cada visita més a l’any; p<0,001), l’existènciade macroangiopatia i presència d’incapacitat temporal(OR: 3,65; p <0,001), i entre manca de dades referidesa l’existència d’alteracions nefrològiques i la presènciad’incapacitat temporal (OR: 0,70; p = 0,04; així doncs,que no constessin dades sobre nefropaties s’associa-va a no presentar incapacitat temporal).
4. Anàlisi de l’absentisme laboral dels treballadorsdiabètics amb un bon control de la malaltia i lapoblació general amb història clínica oberta alscentres d’atenció primària.
Es va efectuar una anàlisi comparativa de l’absentis-
me laboral entre els treballadors diabètics amb un boncontrol de la malaltia i la població general, ambhistòria clínica oberta als centres d’atenció primària.
Els treballadors diabètics amb un bon control de lamalaltia (n =353) van presentar una taxa d’incidènciad’incapacitat temporal del 26% (n= 92), sent del17,4% (n=230) per a la població general. Es vanobservar diferències estadísticament significatives (p<001) entre el grup de treballadors diabètics amb unbon control de la malaltia i el de la població general enrelació amb el percentatge de treballadors que vanpresentar almenys un episodi d’incapacitat temporaldurant el període d’estudi. La mitjana d’episodisd’incapacitat temporal va ser de 0,3 i de 0,2, i la delnombre de dies de durada dels episodis de 13,9 i 9,5,respectivament. Per comparar el nombre d’episodisd’incapacitat temporal entre els treballadors diabèticsamb un bon control de la malaltia i la població generales va utilitzar la prova de U Mann-Whitney, i lesdiferències van ser estadísticament significatives (p<0,001).També es va utilitzar aquesta prova estadísticaper comparar entre ambdues poblacions el nombrede dies d’incapacitat temporal, i es van trobar diferèn-cies estadísticament significatives (p= 0,001).Posteriorment es va efectuar una anàlisi multivariantde l’absentisme laboral segons un model de regressiólogística amb el mètode d’introducció de variablesEnter que va incloure les variables amb significacióestadística en l’anàlisi bivariable, la presència dediabetis, el nombre de visites i l’edat i el sexe. En laregressió logística es va observar associació estadísti-cament significativa entre la presència de diabetis ipresència d’incapacitat temporal (OR: 1,60; p=0,007), el nombre de visites i presència d incapacitattemporal (OR: 1,20 per cada visita més a l’any;p<0,001), i entre ser un treballador autònom i presèn-cia d’incapacitat temporal (OR: 0,50; p= 0,001; totpresentant els treballadors autònoms menys episodisd’incapacitat temporal).
La distribució dels episodis d’incapacitat temporalsegons les categories diagnòstiques que els vanocasionar van mostrar diferències estadísticamentsignificatives entre ambdós grups (p= 0,0018). Elsprincipals motius diagnòstics van ser les malaltiesinfeccioses i parasitàries (18,3%), les malalties del’aparell locomotor i del teixit conjuntiu (13,9%) i lesmalalties de l’aparell respiratori (9,6%) per al grup detreballadors diabètics amb un bon control, i lesmalalties de l’aparell locomotor i del teixit conjuntiu(22,2%), les malalties de l’aparell respiratori (14,7%) iles malalties infeccioses i parasitàries (12%) per a lapoblació general.
FUNDACIÓ LA MARATÓ DE TV3

RECERCA BIOMÈDICA | 125

FUNDACIÓ LA MARATÓ DE TV3

RECERCA BIOMÈDICA | 127
3. Rellevància i possibles implicacions clíniquesdels resultats finals obtingutsLa població diabètica ocupada atesa als centresd’atenció primària es caracteritza per:
Ser majoritàriament homes de mitjana d’edat elevada.Aquestes dades estarien motivades per una majortaxa d’ocupació dels homes i una major prevalençade la diabetis en els grups de major edat.
El perfil sociolaboral del treballador diabètic és moltsimilar al de la població general i es caracteritza així:treballador de la indústria que desenvolupa la sevaocupació predominantment en empreses petites delsector del comerç, venda i reparació de vehicles demotor, amb experiència laboral, que treballa en horarifix diürn i més de 40 hores setmanals i que majorità-riament té un contracte de treball fix. Tot i això, entreels treballadors diabètics existeixen més treballadors noqualificats, treballen en menor proporció en empresesgrans i tenen una major precarietat en la feina.
Les condicions de treball observades a partir delsriscos de treball percebudes pels treballadorsdiabètics mostren una distribució similar a la de lapoblació general, i es caracteritzen per un nombreimportant de treballadors exposats a riscos de tipusfísics (sorolls, vibracions...), a fums, pols o fibres al’aire, o a la utilització de substàncies perilloses, i peruna baixa proporció de treballadors diabètics ensituació de risc per risc de tipus psicosocial, malgratque presenten una elevada demanda psíquica i físicade treball. També destaca el major suport social rebut.
El perfil de la malaltia mostra que es tracta dediabètics amb aproximadament 7 anys d’evolució,que han rebut informació sanitària referida a lamalaltia, sobre com fer el control i sobre les complica-cions, que realitzen autocontrols de glucèmia capil·lar,i que majoritàriament efectuen tractament ambantidiabètics orals i dieta, tot i que els tractats ambinsulina representen aproximadament una quarta partdels treballadors diabètics ocupats.
FUNDACIÓ LA MARATÓ DE TV3

El control de la malaltia el realitzen bàsicament elsmetges i infermeres d’atenció primària. En aproxima-dament el 65% dels casos es disposa d’informaciósobre el control metabòlic, sent bo en aproximada-ment la meitat d’aquests casos. Existeix unsubregistre de les complicacions de la malaltia enl’HCAP.
La freqüentació sanitària dels treballadors diabètics almetge de capçalera i a la infermera va ser superiorque la de la població no diabètica.
Una gran part dels treballadors diabètics tenen unabona integració laboral. Així, en relació amb el maneigi control de la diabetis al medi laboral, s’ha observatque la majoria de treballadors diabètics estudiatspodien adaptar les ingestes a l’horari de treball, i sentel tipus de feina el motiu principal per no poder dur-lesa terme.
També s’ha observat que aproximadament la meitatdels treballadors que feien tractament amb insulina, sel’administraven durant l’horari de treball, tot disposanten la majoria dels casos de temps suficient per fer elcontrol i l’administració. La raó principal per no poderfer-la va ser no disposar de pauses per a l’aplicació. Ellloc més habitual per a l’administració és els lavabos.
En una proporció important de treballadors diabètics,els companys de feina i els caps coneixen la presèn-cia de la malaltia, tot i que en el cas dels capsaquesta proporció és notablement inferior. Les princi-pals causes per no comunicar-ho són preservar laintimitat i evitar la discriminació laboral.
L’absentisme laboral dels treballadors diabètics atesosals centres d’atenció primària, es caracteritza per:
Les característiques demogràfiques, les sociolaborals iles condicions de treball percebudes pels treballadorsdiabètics amb almenys un episodi d’incapacitattemporal són similars a les dels conjunt treballadorsdiabètics atesos a l’atenció primària de salut.
Més 2/3 dels treballadors diabètics no van presentarcap episodi d’incapacitat temporal durant el períoded’estudi.
La taxa d’incidència, el nombre d’episodis i la duradadels episodis d’incapacitat temporal és notablementsuperior en els treballadors diabètics respecte de lapoblació general.
La presència de macroangiopatia diabètica i elnombre de visites als professionals sanitaris s’hanmostrat com factors associats a absentisme laboralper IT. També destaca que el fet de no disposar dedades clíniques sobre nefropatia s’associa a absènciad’incapacitat temporal.
Els principals motius de salut que han ocasionat elsepisodis d’incapacitat temporal han estat els mateixosper als treballadors diabètics i per a la poblaciógeneral i no han estat patologies relacionades amb lescomplicacions de la diabetis.
La incapacitat temporal en el grup de treballadorsdiabètics amb un bon control metabòlic es caracterit-za per:
A. Presentar també una major taxa d’incidència d’IT, iun major nombre i durada dels episodis d’IT que lapoblació general. Tot i això, la durada dels episodisd’IT és menor que la que presenta la població diabèti-ca ocupada atesa a l’atenció primària de salut.
B. S’ha observat associació entre el grau de controlmetabòlic i la presència d’incapacitat temporal,malgrat que es perd la significació estadística enincloure en el model la presència de la malaltia i elnombre de visites als professionals sanitaris o el tipusde contracte.
C. Un altre element per destacar és que els treballa-dors diabètics amb un mal control presenten unamajor taxa d’incidència d’incapacitat temporal que laque presenten els treballadors amb un bon controlmetabòlic, i que aquesta és gairebé el doble que ladels treballadors de la població general.D. La presència de la malaltia i el nombre de visites alsprofessionals sanitaris s’han mostrat com factorsassociats a absentisme laboral per IT en aquest grup.D’altra banda, el fet de ser un treballador autònom escomporta com un factor protector en relació amb lapresència d’IT.
E. Els principals motius diagnòstics que ocasionen elsepisodis d’IT dels treballadors diabètics amb un boncontrol metabòlic són els mateixos que els de lapoblació diabètica ocupada i que els dels treballadorsocupats de la població general.
RECERCA BIOMÈDICA | 129

FUNDACIÓ LA MARATÓ DE TV3
MEMÒRIA FINAL
1. Resum del projecte originalLa resistència a la insulina és un component fonamen-tal de la patogenesi de la diabetis tipus II i també estàassociada a altres disfuncions fisiològiques, com aral’obesitat o la hipertensió. En cèl·lules adiposes, laresistència a la insulina provoca un increment en lalipòlisi i, com a conseqüència, un augment dels àcidsgrassos circulants, la qual cosa pot derivar en proble-mes vasculars.
El TNFa (Tumor Necrosis Factor a) és una citoquinaque provoca una àmplia varietat de respostes biològi-ques en diversos teixits. A part de les seves propietatsoncolítiques, s’ha observat que les elevades concen-tracions de TNFa associades a diferents estats patolò-gics, estan relacionades amb la resistència a la insuli-na. És per això que s’ha proposat el TNFa com unmediador clau en la resistència a la insulina relaciona-da amb la obesitat. Un altre dels factors que s’haproposat com a mediador de la resistència a la insuli-na és l’estrès oxidatiu.
Els mecanismes moleculars pels quals es produeix la
intercomunicació entre les vies de senyalització de lainsulina i el TNFa no són completament coneguts; noobstant això, es coneixen una sèrie de quinases quesón activades tant pel TNFa com per l’estrès oxidatiu.Aquest grup de quinases pertanyen a la família de lesMAP quinases i són anomenades genèricamentquinases activades per estrès (stress activated proteinkinases). Per tant, un dels objectius d’aquest projecteconsisteix a determinar quines són les quinasesactivades tant pel TNFa com per l’estrès oxidatiu encèl·lules 3T3-L1 diferenciades en adipòcits. Per dur aterme aquest estudi s’utilitzarà la tècnica de Westernblot utilitzant anticossos contra les formes fosforila-des/actives d’aquests enzims.
Els efectes provocats per la insulina estan mediatsprincipalment per dues vies, la de la MAP quinasa(ERK1/ERK2) relacionada amb els efectes metabòlics ila via d’activació de la PKB a través de la PI3 quinasa,relacionada amb el transport de glucosa per mitjà delstransportadors Glut4. Es per això que un altre delsobjectius del treball consisteix a determinar com elTNFa i l’estrès oxidatiu modulen l’activació d’aquestesdues vies. Així mateix, s’estudiarà si alguna de lesquinases d’estrès activades per aquests factors està
Títol del projecte
Estudi de l'activació de quinases d'estrès pel TNF-alfa i estrès oxidatiuen adipòcits: intercomunicació amb el senyal generat per la insulina
Investigador principalNéstor Gómez Trias
InstitucióFacultat de Veterinària. UAB
Membres de l’equipMercè Unzeta i Lourdes Garcia.
61.423 €

implicada en la modulació de les vies estimulades perla insulina. Per això, s’utilitzaran inhibidors selectiusd’aquests enzims.
D’altra banda, existeixen una sèrie d’enzims produc-tors d’aigua oxigenada dels quals encara esdesconeix el paper fisiològic. Un d’aquests enzims,anomenat SSAO (semicarbazide sensitive amineoxidase) metabolitza amines biògenes i és moltabundant en teixit adipós. En aquest context, hi ha lapossibilitat que en determinats estats patològics enels quals augmenti la concentració dels substratsendògens d’aquest enzim, l’increment en la producciód’aigua oxigenada generi resistència a la insulina. Pertant, també s’estudiarà si la presència de substratsd’aquest enzim provoca l’activació de les quinasesd’estrès en cultius diferenciats de cèl·lules 3T3-L1.
2. ResultatsEs va determinar com el TNFa i l’H2O2 activaven elsdos tipus de MAP quinases estimulades per estrès,anomenades p38 i JNK. En el cas de la p38, s’activa-va als 5-10 min i retornava als nivells basals als 20min, mentre que amb H2O2 l’activació va ser mésgradual, tot observant-se ja als 5 min i arribant a unmàxim als 60-90 min. Pel que fa a la JNK, el TNFal’activava als 5 min, tot arribant a un màxim als 10min, que es mantenia constant almenys fins a 1 hora,mentre que l’activació per H2O2 es començava aobservar als 30 min i arribava al màxim a l’hora. Capd’aquestes quinases va ser activada per insulina. Aixímateix, la incubació de les cèl·lules amb diversesconcentracions de substrats de l’enzim SSAO,tampoc no indueix l’activació de les quinases d’estrès.
Atès que es volien determinar els efectes del TNFa il’H2O2 sobre les MAP quinases (ERK1/2) i la PKBestimulades per insulina, també es va determinarquina era la cinètica d’activació d’aquests enzimsquan les cèl·lules eren estimulades per cadascund’aquests estímuls.
En el cas d’ERK1/2, el TNFa l’estimulava per arribar aun màxim als 10 min i retornava a nivells basals als 20min, mentre que l’H2O2 l’activava als 5 min, arribavaa un màxim als 20 min i retornava lentament als nivellsbasals, tot observant-se encara un lleu activació a les2 hores. L’activació d’aquest enzim per la insulina vaser més ràpida, en arribar al màxim als 5 min, idavallant posteriorment fins a nivells pràcticamentbasals als 20 min, si bé una petita fracció encara esmantenia activa al cap d’una hora.
La PKB no era estimulada pel TNFa, mentre quel’H2O2 l’activava als 5 min, arribava a un màxim als10 min i desapareixia després de 30 min d’estimula-ció. Pel que fa a la insulina, activa la PKB, arriba a unmàxim als 5 min, que es manté constant aproximada-ment durant una hora, després de la qual hi ha unalenta davallada, de tal manera que a les 2 horesencara s’observava un cert grau de fosforilaciód’aquest enzim.
Per veure si el TNFa bloquejava l’activació de la MAPquinasa per la insulina, les cèl·lules es van incubaramb aquest compost durant diversos períodes de
temps (30-5 hores) i a continuació es van tornar aestimular amb insulina durant 5 min. Es van escolliraquests temps perquè a partir de 30 min l’activaciód’ERK1/2 pel TNFa ja ha retornat als nivells basals, ien el cas de la insulina, ja que és el punt on es trobael màxim d’activació. En aquests experiments no esva observar cap efecte del TNFa sobre l’activitatestimulada per insulina. Experiments similars es vandur a terme amb H2O2, si bé en aquest cas es vaobservar un bloqueig complet en l’activació de la MAPquinasa estimulada per la insulina. Aquest bloqueigs’observa fins a les dues hores i a partir d’aquí esrecupera la resposta a la insulina. Així mateix, tambées va determinar quin era l’efecte que provocavasobre ERK1/2 un pretractament de les cèl·lules ambinsulina. Com en el cas de l’H2O2, es va observar quela insulina també produïa un bloqueig en l’activacióposterior d’ERK1/2 per la mateixa hormona, si bé enaquest cas a les 5 hores les cèl·lules encara erenrefractàries a l’estimulació per insulina.
Assajos de la mateixa naturalesa es van dur a termeen el cas de la PKB. En aquest cas es va observarque la preincubació de les cèl·lules amb TNFa o insuli-na no tenia cap efecte en la posterior estimulaciód’aquest enzim amb insulina. La manca d’efecte delTNFa sobre l’activació de la PKB per insulina es vacorroborar amb els estudis realitzats mesuranttransport de glucosa, ja que està descrit que aquestdepèn de l’activació de la PKB. En aquest cass’observa una manca d’efecte del TNFa sobre eltransport de glucosa estimulat per la insulina. Pel quefa a l’H2O2, es va observar que produïa un lleubloqueig en l’activació de la PKB per insulina duranttots els temps assajats.
A continuació es van fer una sèrie d’estudis perdeterminar quina via de senyalització era la responsa-ble del bloqueig que provocaven tant la insulina coml’H2O2 en l’activació d’ERK1/2 per insulina. Per a aixòes van incubar les cèl·lules durant 30 min amb inhibi-dors específics d’algunes d’aquestes vies, prèviamentals tractaments descrits anteriorment.
El pretractament amb l’inhibidor específic de la p38,SB203580, no va produir cap efecte sobre el bloqueigen l’activació d’ERK1/2 per insulina originat perl’H2O2. Això descartava aquesta quinasa com amediadora d’aquest bloqueig. En el cas de la insulina,això ja quedava descartat pel fet que aquestahormona no activa aquesta via.
El mateix resultat es va obtenir utilitzant l’inhibidor dela JNK, SP600125, amb la qual cosa es descartava laparticipació d’aquest enzim en aquest fenomen.
Per veure si aquests efectes eren provocats per mitjàde la PKB, es va utilitzar la wortmanina, un inhibidorde la PI3 quinasa que bloqueja l’activació de la PKB,com també de la p70S6quinasa. Els resultatsobtinguts indiquen que aquesta tampoc no seria la viaresponsable del bloqueig observat.
Com que no existeixen inhibidors coneguts d’ERK1/2i que els inhibidors de la via actuen bloquejant l’activa-ció de MEK1/2, l’enzim responsable de l’activació
RECERCA BIOMÈDICA | 131

d’ERK1/2, es va optar per utilitzar un inhibidor de larsk (p90 ribosomal S6 kinase). Aquesta és una de lesquinases activades per ERK. Aquest inhibidoranomenat GF109203X també actua com a inhibidorde la PKC. La manca d’efecte d’aquest inhibidorpermet descartar aquestes dues quinases com lesimplicades en el bloqueig.
Aquests resultats no exclouen la possibilitat queaquests efectes siguin provocats per la mateixa MAPquinasa. Com que els inhibidors de MEK no es podienutilitzar perquè bloquegen l’activació de les ERK, esva optar per una estratègia alternativa. Aquesta vaconsistir a mesurar l’activació de ras, un componentde la via de les ERK situat per sobre de MEK.
Aquesta proteïna s’activa a través de la seva unió aGTP i és aquesta la forma que interacciona i promoul’activació de raf. Així, per mesurar l’activació de ras,es van fer assajos en què la forma activa de ras (ras-GTP) es precipitava i la quantitat de ras actiu esdeterminava mitjançant Western blot. Com que rasinteracciona amb raf únicament quan està unit a GTP,per precipitar aquesta forma selectivament es va utilit-zar una proteïna de fusió en què el domini d’unió a raspresent en raf està unit a GST. Tal com es podiaesperar, l’estimulació de les cèl·lules amb insulinadurant 2 min va provocar un augment en la concen-tració de ras-GTP. La preincubació d’aquestescèl·lules amb H2O2 o insulina bloquejava aquestaactivació, però la incubació prèvia amb el compost PD98059 va revertir el bloqueig provocat per la insulina.Aquest compost actua bloquejant l’activació de MEK id’aquestes dades es pot concloure que la resistènciaa la insulina provocada per la mateixa insulina ol’H2O2 estan mediades per vies diferents. Així, mentreque la resistència provocada per la mateixa insulinaestà mediada per la MAP quinasa, en el cas del’H2O2 el mecanisme és diferent.
En la majoria dels treballs en què s’ha estudiat laresistència a la insulina, aquest fenomen s’haassociat a la fosforilació en Ser/Thr de l’IRS1. Enaquests treballs s’ha suggerit que la fosforilació del’IRS1 en Ser/Thr provoca una disminució de lafosforilació en Tyr i, com a conseqüència, unbloqueig en la resposta posterior a la insulina. S’hanidentificat diverses Ser que són fosforilades enresposta a estímuls que poden provocar resistènciaa la insulina. Una d’aquestes, la Ser307, ha estatproposada com la implicada en la resistència a lainsulina mediada pel TNFa i la insulina. És per aixòque es va estudiar la cinètica de fosforilaciód’aquesta Ser en resposta al TNFa , H2O2 i insulina.En els tres casos es va donar una cinètica similar, enla qual, la Ser307 es fosforilava després de 10 mind’estimulació, i encara romania fosforilada als 60min. No obstant, en cap dels casos es pot correla-cionar la fosforilació d’aquesta Ser amb els efectessobre la MAP quinasa i la PKB. Per altra banda, elgrau de fosforilació observat era molt baix quan escomparava amb el que es dóna en la línia cel·lularHEK293. Això indicava que la resistència a la insuli-na atribuïda a la fosforilació de l’IRS1 no es correla-cionava amb els efectes sobre la PKB i la MAPquinasa. Per veure si aquest fenomen era específic
de la línia cel·lular 3T3-L1, es van repetir els experi-ments en les cèl·lules HEK293.
Malauradament, en aquestes cèl·lules la insulina noactiva la MAP quinasa i, per tant, només es vanestudiar els efectes sobre la PKB. En aquest cas,únicament l’H2O2 bloquejava la posterior activació dela PKB i únicament a temps llargs (2-5 hores). Aixòcontrasta amb el fet que la fosforilació de l’IRS en laSer307 és transitòria en el cas de l’H2O2, on desapa-reix completament a les dues hores, mentre que en elcas del TNFa i la insulina encara s’observa un alt graud’IRS1 fosforilat a les 5 hores d’incubació. A més, laSer307 es fosforilava amb més intensitat quan lescèl·lules s’estimulaven amb TNFa o insulina. Per tant,en aquestes cèl·lules tampoc s’observa una correlacióentre la fosforilació d’IRS i el bloqueig de la PKB.Aquests resultats indiquen que la resistència a lainsulina provocada pels diversos estímuls, almenysdurant els temps assajats, no és causada per lafosforilació de la Ser307 de l’IRS.
3. Rellevància i possibles implicacions clíniquesdels resultats finals obtingutsLa resistència a la insulina ha estat concebuda comun dels factors que contribueix a la diabetis de tipus2. El mecanisme molecular d’aquest fenomen ha estatestudiat per diversos grups i s’ha proposat, d’unamanera generalitzada, que està associat a una fosfori-lació d’IRS1 en Ser/Thr. Aquesta proteïna actua comun adaptador en la via de transducció de senyals dela insulina, on és fosforilada en tirosines pel receptorde la insulina, tot generant llocs d’ancoratge per aaltres proteïnes. Així, s’ha proposat que la fosforilacióen serines faria disminuir la fosforilació en tirosines, totbloquejant el senyal. S’han identificat diversos factorsque estimulen la fosforilació en serines, entre els qualss’inclouen el TNFa, determinats tipus d’estrès, comtambé la mateixa insulina. S’han identificat diversesSer fosforilades en IRS1 i s’han proposat diversesquinases com les responsables d’aquesta fosforilació.Entre les Ser fosforilades destaca la Ser307 i ha estatdemostrat que és fosforilada per dues quinasesestimulades pel TNFa, la JNK i la IKK1. Pel que fa a laJNK, s’ha demostrat que és una quinasa estimuladaper diversos tipus d’estrès, la qual cosa apunta lapossibilitat que sigui la implicada en la resistència a lainsulina induïda per estrès oxidatiu. Però malgrat lagran quantitat d’estudis realitzats sobre la fosforilaciód’IRS, pràcticament no hi ha estudis dedicats aestudiar les quinases situades per sota d’IRS1 quesón activades per la insulina. En el nostre, s’hanestudiat dues d’aquestes vies, la MAP quinasa i laPKB. Aquesta última és estimulada per la PI3 quinasai esta relacionada amb el transport de glucosa através de Glut4. En el nostre treball es demostra queen les condicions en les quals IRS1 es fosforila enserines i que, per tant, disminueix la fosforilació entirosina, el TNFa no produeix cap efecte en capd’aquestes vies. Així mateix, es demostra que l’efectemés rellevant provocat per la insulina i l’H2O2 és elbloqueig de la MAP quinasa, i només s’observa unlleu bloqueig en l’activació de la PKB per l’H2O2.L’efecte de la insulina sobre la MAP quinasa estàmediat per la mateixa MAP quinasa i se situa persobre de ras, mentre que en el cas de l’H2O2 aquest
FUNDACIÓ LA MARATÓ DE TV3

bloqueig és originat per un mecanisme diferent, si béno es pot atribuir a cap de les dues quinases activa-des per estrès (p38 i JNK). En aquestes condicions,tots aquests factors estimulen la fosforilació de laSer307, la qual cosa sembla descartar que aquestafosforilació estigui relacionada amb l’efecte sobre laMAP quinasa. Tot això sembla indicar que la fosforila-ció de la Ser307 d’IRS no estaria implicat en elsefectes a curt termini en la resistència a la insulinaprovocada pels factors estudiats i que possiblementtindria un paper a temps més llargs. Per tant, enfuturs estudis sobre la resistència a la insulina, caldràtenir en compte aquest fenomen i demostrar que elscanvis en la fosforilació d’IRS1 estan associats acanvis en l’activació de les quinases situades per sotad’aquesta proteïna.
RECERCA BIOMÈDICA | 133

FUNDACIÓ LA MARATÓ DE TV3
MEMÒRIA FINAL
1. Resum del projecte originalIntroduccióLa diabetis mellitus és una malaltia crònica degenerati-va que s’associa amb l’obesitat. Es caracteritza perun grau major o menor de resistència a la insulina iper una pèrdua total o parcial de la funció insulinose-cretora. El metabolisme dels hidrats de carboni té unpaper clau en la producció d’insulina. Aquestahormona és produïda per les cèl·lules beta del pàncre-es, que s’agrupen amb altres cèl·lules endocrines,talment com si es tractés d’un cabdell. La poblaciócel·lular que forma aquests cabdells cel·lulars, vista almicroscopi òptic s’assembla a un illot, per la qualcosa es descriuen com illots pancreàtics.
La població de cèl·lules beta replica i es diferencia enl’estat embrionari i en els primers dies postnaixement.No obstant això, aquesta població no romandràestable de per vida, sinó que, ans al contrari, es potmultiplicar a partir de les cèl·lules beta ja formades(replicació), o bé augmentar a partir de precursorscel·lulars poc diferenciats situats als ductes pancreà-tics, procés anomenat neogènesi.
La regulació fisiològica d’aquests processos encarano és del tot coneguda. Sabem molt poc dels factorsque participen en aquests procés ni tampoc el tipusde cèl·lules progenitores. S’han establert dues hipòte-sis, la primera accepta que les cèl·lules beta esformen a partir de cèl·lules ductals o acinars pancreà-tiques. La segona preveu la diferenciació de cèl·lulespluripotencials localitzades al voltant de l’epiteli ductal.
En estudis previs vam demostrar que l’administracióde tungstat sòdic a diferents models animals diabèticsera capaç de normalitzar les concentracions deglucosa en la sang per dos mecanismes. Un d’ellsfacilita el metabolisme de la glucosa al teixit hepàtic il’altre millora la funció de l’illot pancreàtic, gràcies a larecuperació de la massa cel·lular beta.
En aquest projecte hem investigat el procés de plasti-citat cel·lular induïda per l’administració de sals detungstat als animals diabètics i hem identificatl’expressió diferencial dels gens que la regulen.Tanmateix, en el curs de la nostra recerca vamobservar un tercer efecte de l’administració de les salsde tungstat, la pèrdua de pes, per la qual cosa vaminvestigar aquest efecte com un possible camí
Títol del projecte
Neogènesi dels illots pancreàtics: identificació de la expressió diferencial dels gens induïts per tungstat
Investigador principalRamon Gomis de Barbarà
InstitucióHospital Clínic i Provincial de Barcelona
Membres de l’equipEva Aguilera, Jordi Altirriba, Albert Barberà, Sílvia Barceló, Anna Bosch, Aida Caballero, Judith Cabedo, Ignasi Canals, Silvia Casas,
Marc Claret, Ignacio Conget, Helena Corominola, Imma Coronado, M. J. Coves, Isabel Crespo, Alicia de Diego, Enric Esmatjes, SilviaFernández, Reina Fernández, Jordi Ferrer, Lilian Flores, Ainhoa García, Rosa Gasa, Marta Julià, Maria Lucas, M.A. Maestro, Melina
Mara, Anna Martín, Nuria Marzo, Eugenia Mato, Concepció Mora, Silvia Moreno,Rosa Morigino, Belén Nadal, José L. Navarro, NuriaPalau, Marcelina Parrizas, Sandra Piquer, Manel Puig, Joan M. Servitja, Gemma Sesmilo, Judith Viaplana i Josep Vidal.
109.685 €

terapèutic per al tractament de la diabetis mellitustipus 2.
2. ResultatsNormalització de la glucèmia i recuperació de lainsulinèmia en rates diabètiquesTal com es mostra a la figura 1, les rates diabètiquestractades amb tungstat sòdic normalitzen la glucèmiaa partir del quart dia de tractament. Aquest efecteantidiabètic del tungstat sòdic es manté els 28 dies dedurada del tractament. Aquesta normalització de laglicèmia s’acompanya per una recuperació de lesxifres d’insulinèmia. En el model de les rates nSTZ,aquesta recuperació és del 100%, i en altres modelspot oscil·lar entre el 20 i el 50%.
Aquesta situació metabòlica va acompanyada d’unapèrdua de pes. No obstant això, els efectes sobre elsparàmetres metabòlics i hormonals són independentsde qualsevol pèrdua de pes.
Anàlisi morfològica i funcional del teixit pancreàticEn els diversos models animals diabètics en els qualshem estudiat l’efecte antidiabètic de l’administració detungstat sòdic, hem observat una recuperació de lamassa beta cel·lular. Aquesta recuperació s’ha produïtper un procés de neogènesi, tal com pot observar-seen la fotografia de la figura 2, i sense observar-hi capaltra modificació en la població cel·lular endocrina oexocrina. Aquesta massa beta cel·lular de novo ésfuncionant. S’acompanya per un augment en l’expres-sió del factor de transcripció PDX-1, que resulta críticen el procés de diferenciació cel·lular.
En alguns models animals, el procés de neogènesis’acompanya d’un procés d’apoptosi, d’una intensitatmenor però que contribueix a mantenir la plasticitatcel·lular de l’illot en el ventall de la normalitat.
Quan després de 28 dies de tractament vam suprimirel tractament, observàvem que la recuperació de lamassa beta cel·lular és estable i manté l’animaldiabètic en normoglicèmia durant llarg temps sensenecessitat de cap tractament; és a dir, l’animal s’hacurat (vegeu figura 3).
Mecanisme d’acció del tungstat sobre el procés deneogènesiEn els diversos estudis en què es va investigar l’efectedel tungstat sobre el procés de neogènesi de l’illot, noes va observar un augment en la quantitat de proteïnaPDX-1. No obstant això, és ben conegut que l’activitatde PDX-1 com a factor de transcripció que regulal’expressió de determinats processos de plasticitatcel·lular està regulat pel grau de fosforilació. D’acordamb aquest coneixement previ, vam investigar si lessals de tungstat sòdic tenien la capacitat d’incremen-tar el grau de fosforilació de PDX-1. Tal com es potobservar en la figura 4, el tractament d’animalsdiabètics amb tungstat aconsegueix fosforilar PDX-1,procés que és mediat per un augment de la fosforila-ció de P-38 i que té com conseqüència la transloca-ció de PDX-1 al nucli (figura 5) i l’augment en l’expres-sió de l’m-RNA de la insulina als illots (figura 4).
Efectes antiobesitat del tractament amb tungstatUn dels objectius secundaris del projecte era investi-gar l’efecte del tungstat sobre el pes. Els resultats vanmostrar que els animals tenien una pèrdua de pesque no estava relacionada amb cap efecte tòxic, perla qual cosa es va dissenyar un projecte que tinguéscom a objectiu principal l’efecte antiobesitat de lessals de tungstè. Els resultats es mostren a la figura 6.Com s’hi pot observar, els animals obesos responienel tractament i normalitzaven el seu pes. Aquestefecte no afectava el creixement, la pèrdua de greixosper via intestinal, sinó que era causat per un augmenten la termogènesi, tal com es va comprovar pelconsum d’oxigen. Tanmateix, en els estudis molecu-lars es va veure un augment en l’activitat de les proteï-nes desacobladores (figura 7), que vam vincular a unaugment en la taxa de fosforilació del factor detranscripció PPAR-gamma (figura 8).
Figura 1. En la figura es mostren els valors de glicèmia dels
animals diabètics (quadrats buits), diabètics tractats (quadrats
plens) i animals no diabètics, tant si estan tractats (rodones
plenes) com no tractats (rodones buides). * p? 0.05 i ** p? 0.001
Figura 2. Expressió d’insulina i PDX-1 en algunes seccions de
teixit pancreàtic. En les finestres a i b, el tint verd és un marcador
d’insulina (x400). Es pot veure com es forma un illot nou, a partir
de cèl·lules del ductus pancreàtic. En les finestres c, d, e, f, el tint
verd és PDX-1 i el vermell insulina; c) és un illot normal; d) un illot
diabètic; e) un illot diabètic d’un animal tractat. Es pot observar un
augment de PDX-1 extraillot; f) és un illot ja normalitzat després
de 28 dies de tractament (x630).
RECERCA BIOMÈDICA | 135

Figura 3. Glicèmia de les rates tractades (quadrats negres)
després d’haver suprimit el tractament. En quadrats buits
representem la glicèmia dels animals diabètics no tractats. Pot
observar-se que als 15 dies d’haver suprimit el tractament, els
animals segueixen amb les glucèmies normals. *p<0,001
Figura 4. Western blots (2 D) obtinguts a partir d’illots pancreàtics
d’animals tractats i no tractats amb tungstat (4 dies de tracta-
ment). a) el tungstat disminueix el PI del PDX-1, la qual cosa és
un indicador de fosforilació; b) activació de P-38 pel tungstat. DT
(animals diabètics tractats), DU (animals diabètics no tractats); c)
efecte del tungstat sobre l’mRNA d’insulina.
Figura 5. En la figura es visualitza com el tractament amb tungstat
transloca PDX-1 als nuclis.
Figura 6. Efecte de les sals de tungstat sobre el pes d’animals als
qual es va induir obesitat per una dieta rica amb nutrients hiperca-
lòrics (dieta de cafeteria) ***p<0,001.
Figura 7. Consum d’oxigen (a) i expressió gènica de les proteïnes
desacobladores en teixit adipós marró interescapular (b), teixit
adipós blanc epididimari (c) i múscul (d). *p<0,05.
FUNDACIÓ LA MARATÓ DE TV3

Figura 8. Efecte del tungstat sobre l’expressió i fosforilació de
PPAR-gamma i MAPK.
3. Rellevància o aplicabilitat clínica dels resultatsfinals obtingutsEls resultats obtinguts tenen un doble interès. D’unabanda, es confirma l’efecte antidiabètic de les sals detungstat. D’una altra, es coneix molt millor el mecanis-me d’actuació de les sals de tungstat com a antidia-bètic. Així sabem que part de l’efecte antidiabètic éscausat per un augment de la massa de cèl·lules betaproductores d’insulina, per mitjà d’un increment de laneogènesi, procés que consisteix en la transformacióde cèl·lules ductals en cèl·lules de naturalesa endocri-na secretores d’insulina.
El manteniment i/o la recuperació de les cèl·lules betaés una estratègia clau per avançar en el tractament dela diabetis. En la diabetis mellitus tipus 1 hi ha unapèrdua gairebé total de cèl·lules beta productoresd’insulina i en la diabetis mellitus tipus 2, aquestareducció encara que sigui parcial augmenta tot al llargde la malaltia. Ambdós defectes es poden resoldreamb un tractament capaç de regenerar el teixit. D’acíla rellevància de l’estudi, especialment si consideremque en aquests moments estem en curs d’obtenirresultats en què mostrem amb l’ús d’arrays quins sónels determinants moleculars que regulen el procés deneogènesis i estem capacitats perquè en un futurproper siguem capaços d’identificar noves dianesterapèutiques i possibles nous productes amb finali-tats terapèutiques.
També hi ha un altre impacte addicional. En eltranscurs de l’acció que hem desenvolupat en aquestprojecte, hem estat capaços de descobrir un efecteantiobesitat de les sals de tungstat. Aquest efecte hadonat opció a patentar l’invent, cosa que hem fet permitjà de l’Agència de Patents de la Universitat deBarcelona. A més, aquesta patent ha estat adquirida ala Universitat per una multinacional farmacèutica i enaquest moment s’està acabant la fase clínica 1 enhumans per seguir el camí de la seva possible aplica-ció terapèutica. Hem d’assenyalar que això suposaque hem superat les fases de toxicitat prèvia. Aquestaés una de les poques patents generades al nostrepaís que està ja superant la fase 1 en humans.
Per tot això, creiem que aquesta recerca pot tenir enun futur no llunyà un clar benefici per a la poblacióque pateix diabetis mellitus i obesitat.
4. PublicacionsJ Fernández-Álvarez, A Barberà, B Nadal, S Barceló-Batllori, S
Piquer, M Claret, JJ Guinovart and R Gomis. Stable and functional
regeration of pancreatic beta-cell population in nSTZ-rats treated
with tungstate. Diabetologia (2004) 47:470-477
A Barberà, H Del Zotto, J Fernández-Álvarez, B Nadal, JJ
Guinovart, JJ Gagliardino and R Gomis. Induction of beta cell
neogenesis by tungstate treatment en STZ- diabetic rats. (sotmès
per a publicació)
M. Claret, H Corominola, I Canals, J Saura, S Barcelo-Batllori,
JJ Guinovart and R Gomis. Tungstate decrease body weight and
adiposity through increased thermogenesis and lipid oxidation in
obese rats. (sotmès per a publicació)
Enguany hi ha quatre manuscrits en preparació.
RECERCA BIOMÈDICA | 137

FUNDACIÓ LA MARATÓ DE TV3
MEMÒRIA FINAL
1. Resum del projecte originalObjectius1. Determinar el paper de la glucosa 6-fosfat (G6P) idels enzims que en controlen la síntesi i la degradacióen el control de la translocació de la glucogen sintasa(GS) hepàtica.
1.1 Estudi de la translocació de la GS en cèl·lules nohepàtiques.1.2 Efecte de la G6P i de la GK sobre la translocacióde la sintasa.1.3 Avaluació de la sobreexpressió de la GK en latranslocació de la GS endògena en hepatòcits de rata.1.4 Sobreexpressió de la GK pancreàtica.1.5 Estudi de la sobreexpressió de la G6Pasa en latranslocació de la GS.
2. Estudiar el mecanisme de regulació de la glucoqui-nasa (GK) per la seva proteïna reguladora en fetge(GKRP).2.1 Impacte metabòlic de la sobreexpressió de laGKRP en hepatòcits de rata.
2.2 Determinar la localització intracel·lular de la GKRP.
2.3 Estudiar les alteracions de la GKRP en hepatòcitsprocedents de rates genèticament diabètiques (ZDF).
3. Determinar el paper de la G6P i dels enzims que encontrolen la síntesi i la degradació en la secreciód’insulina estimulada per glucosa.3.1 Efecte de la sobreexpressió de la GK pancreàtica ihepàtica en cultius de línies cel·lulars derivades decèl·lula ß.3.2 Efecte de la sobreexpressió de la GKRP en cultiusde línies cel·lulars derivades de cèl·lula ß.3.3 Efecte de la sobreexpressió de la G6Pasa encultius de línies cel·lulars derivades de la cèl·lula ß.3.4 Estudi de la glucotoxicitat en la cèl·lula ß.
MetodologiaPreparació d’adenovirus recombinants per a lasobreexpressió d’enzims.
Ús de la GFP en la construcció de proteïnes de fusióper localitzar-les intracel·lularment.
Tècniques de biologia molecular.
Títol del projecte
La glucosa 6-fosfat com a metabòlit clau en els mecanismes deregulació de la glicèmia
Investigador principalJoan J. Guinovart i Cirera
InstitucióFacultat de Química. UB
73.357 €

Immunohistoquímica i immunocitoquímica.Western i Northern blot.Cultiu d’hepatòcits i de línies cel·lulars.
2. Resultats1.1La línia cel·lular FTO2B és una línia cel·lular d’hepato-ma de rata que té com a característica que és incapaçd’acumular quantitats detectables de glucogen quans’incuba en presència de glucosa. A més, la localitza-ció de la glucogen sintasa hepàtica (LGS) no varia coma resposta al tractament amb glucosa, amb la qualcosa es troba distribuïda pel citoplasma de la mateixamanera que els cultius primaris d’hepatòcits incubatsen absència de glucosa (figura 1).
Figura 1. Distribució de la LGS en la línia cel·lular FTO2B. S’ha
emprat un anticòs específic contra GS de fetge.
A. Cèl·lules FTO2B sense glucosa.B. Cèl·lules FTO2B incubades amb glucosa 30 mMC. Cèl·lules FTO2B infectades amb adenorvirus quecodifiquen per a GK (AdCMV-GK) incubades enabsència de glucosa.D. Cèl·lules FTO2B infectades amb adenovirus quecodifiquen per a GK (AdCMV-GK) incubades ambglucosa 30 mM.E. Cèl·lules FTO2B infectades amb adenovirus quecodifiquen per a HKI (AdCMV-GK) incubades enabsència de glucosa.F. Cèl·lules FTO2B infectades amb adenovirus quecodifiquen per a HKI (AdCMV-GK) incubades ambglucosa 30 mM.
La sobreexpressió de GK en FTO2B i el seu tracta-ment amb glucosa origina la síntesi de glucogen (figura2), tot produint-se translocació de la GS a la perifèriacel·lular (figura 1). Aquest efecte és específic de GK,atès que l’hexoquinasa I (HKI), que catalitza la mateixareacció, no és capaç d’induir el canvi de localitzaciósubcel·lular de LGS (figura 1). En conseqüència,s’aconsegueix recuperar el fenotip que s’observa enhepatòcits en cultiu només en presència de GK.
Figura 2. Acumulació de glucogen en cèl·lules FTO2B que
sobreexpressen GK i HKI en presència de 10 mM glucosa.
La distribució subcel·lular de glucogen queda palesaper mitjà d’un anticòs específic contra aquest polisa-càrid.
A. Cèl·lules FTO2B tractades amb glucosa 10 mMB. Cèl·lules FTO2B infectades amb adenorvirus quecodifiquen per a GK (AdCMV-GK) tractades ambglucosa 10 mM.C. Cèl·lules FTO2B infectades amb adenovirus quecodifiquen per a HKI (AdCMV-GK) tractades ambglucosa 10 mM.
1.2S’ha fet la sobreexpressió en cèl·lules FTO2B de HKI,GK, LGS i la isoforma muscular de la glucogen sintasa(MGS) per mitjà de l’ús d’adenovirus recombinants. Amés, s’han dut a terme combinacions dels dosenzims fosforiladors de glucosa (GK i HKI) ambcadascuna de les isoformes de GS.
En un primer estudi es va comprovar que la sobreex-pressió de la HKI i/o LGS, els dos enzims expressatsconstitutivament per aquestes cèl·lules, no produeixun increment significatiu en els nivells de LGS activa nien el contingut total de glucogen. En canvi, lasobreexpressió de la GK originava una activació de laLGS tant endògena com exògena, de manera doside-pendent de glucosa. Aquests processos originavenuna acumulació de glucogen. Com a contrast, la MGSsobreexpressada s’activa eficientment per la G6Pproduïda tant per la HKI com per la GK. A partird’aquests resultats, es va aprofundir en l’estudi de latranslocació de la GS muscular en absència i presèn-cia de glucosa (figura 3).
Els resultats indicaven la presència de dos reservorisintracel·lulars de G6P a la cèl·lula: l’un, que eraaccessible per a les dues isoformes de la GS i que ésgenerat per la GK, i un altre conjunt, en què la G6P ésproduïda per la HKI del qual la LGS és exclosa.
Quant al fenomen de translocació de LGS, es vaobservar que la sobreexpressió de la GK originava unincrement dels nivells de G6P i una acumulació deLGS en la perifèria cel·lular. En conseqüència, es
RECERCA BIOMÈDICA | 139

produïa l’inici de la síntesi de glucogen en la perifèriacel·lular. Amb l’objectiu de determinar si la respostaera un efecte exclusiu de la GK, es van posar a puntexperiments en què se sobreexpressava la HKI, ambla qual cosa s’obtenien nivells de G6P semblants alsobtinguts mitjançant la sobreexpressió de GK. Laincubació de les cèl·lules que sobreexpressen HKIamb glucosa no originava un canvi en la localitzaciósubcel·lular de la LGS ni tampoc no es produïal’acumulació de glucogen. Aquests resultatsdemostren que només un increment dels nivells deG6P generada a partir de GK és capaç d’originar elscanvis de localització subcel·lular de la LGS queacompanyen l’inici de la síntesi de glucogen (figura 4).
Figura 3. Distribució d’LGS i gucogen en cèl·lules FTO2B infecta-
des amb el virus que codifica per a MGS (AdCMV-MGS).
A. Cèl·lules FTO2B sense glucosa. Detecció ambl’anticòs contra MGS.B. Cèl·lules FTO2B tractades amb glucosa 30 nM.Detecció amb l’anticòs contra MGS.C. Cèl·lules FTO2B infectades amb adenovirus quecodifiquen per a MGS en absència de glucosa.Detecció amb l’anticòs contra glucogen.D. Cèl·lules FTO2B infectades amb adenovirus quecodifiquen per a MGS tractades amb glucosa 30 mM.Detecció amb l’anticòs contra glucogen.
Figura 4. Distribució de la LGS (en vermell) i de la GK (en verd) en
cèl·lules FTO2B infectades amb l’adenovirus que codifica per a la
GK i incubades amb glucosa 10 mM.
1.3LGS endògena d’hepatòcits de rata control es localit-za a prop de la membrana cel·lular quan s’indueix lasíntesi de glucogen com a resposta a la incubacióamb glucosa. En el cas de cèl·lules que sobreexpres-sen GK, aquesta translocació s’accentua però ésdifícil de quantificar. En canvi, la sobreexpressió de HKI no provoca un increment en la translocació de laLGS tal com s’esdevenia amb GK. Aquestes dadesremarquen la importància de l’enzim que catalitza lasíntesi de G6P en la localització subcel·lular de LGS ien la deposició de glucogen.
1.4Els nostres experiments indiquen que la isoformapancreàtica de GK, que només es diferencia del’hepàtica en els seus primers 15 aminoàcids, estimu-lava d’una manera semblant en la isoforma hepàtica latranslocació de la GS en presència de glucosa encondicions de deposició de glucogen.
1.5Els nostres resultats mostren que dosis creixents dedihidroxiacetona (DHA) eren capaces d’induir unaacumulació de G6P i de glucogen en hepatòcitscontrols, la qual cosa indica que aquest substratgluconeogènic és eficient per a l’activació de la LGS.Aquest resultat contrasta amb la situació en la qual laG6P és generada a partir de HKI, en la qual nos’indueix la deposició de glucogen ni l’activació de laLGS.
La sobreexpressió de nivells creixents de G6Pasa(mitjançant infecció amb adenovirus recombinants), enhepatòcits en presència de glucosa o substratsgluconeogènics (lactat o DHA), induïa una disminuciódels nivells de G6P, la qual cosa s’associava a unareducció en la deposició de glucogen. Aquestareducció era superior quan DHA era el substratemprat (fins a un 30% i un 40% respecte dels valorscontrols amb lactat o DHA, respectivament). Enconclusió, la sobreexpressió de la G6Pasa va sercapaç de fer minvar la síntesi de glucogen en totes lescondicions assajades.
En contrast, l’increment dels nivells de G6P mitjançantla sobreexpressió de la GK en hepatòcits incubatsamb DHA no va tenir cap efecte significatiu sobre ladeposició de glucogen, i no es van incrementar elsnivells de G6P ni l’activitat de la LGS.
S’ha aprofundit l’estudi de la dependència que existeixentre la síntesi de glucogen i la concentracióintracel·lular de G6P. Els nostres resultats indiquenque independentment dels substrats emprats, hi hauna correlació entre el contingut de glucogen i elsnivells de G6P en hepatòcits. El pendent de la rectadepèn del substrat emprat, en demostrar les nostresdades que la glucosa és el substrat més eficient quanta l’acumulació de glucogen. Una explicació possibled’aquestes diferències podria venir del fet que ladegradació del glucogen sigui diferent en les duescondicions.
FUNDACIÓ LA MARATÓ DE TV3

Està demostrat que la glucosa és un inhibidor potentde la glucogen fosforilasa hepàtica (GP). D’aquestamanera, el cicle fútil que s’estableix entre la LGS i laGP durant el procés de síntesi de glucogen estàreduït. Això permet que la LGS activa sintetitzi moltmés glucogen en presència de glucosa. Per respon-dre aquesta pregunta hem utilitzat un anticòs quereconeix específicament la forma fosforilada en laserina 14 de la GP, que indica un estat actiu d’aquestenzim. Els nostres resultats demostren que els nivellsde GP activa eren més elevats en hepatòcits en cultiuincubats amb 10 mM DHA que amb 5 o 25 mMglucosa (figura 5)
Figura 5. Anàlisi per immunoblot de la fosforilació de GP en serina
14 (forma activada) i de la GP total en hepatòcits incubats amb
glucosa o DHA.
Posteriorment es va fer l’estudi de la translocació dela LGS en aquestes condicions i es va analitzar l’efec-te de la modulació dels nivells de G6P en expressar-se diverses proteïnes implicades en el metabolismedel glucogen (figura 6). La LGS translocava a la perifè-ria cel·lular en presència de glucosa i DHA, d’acordamb un paper directe de l’esmentada translocació enla deposició de glucogen. Amb tot, no s’observavaaquesta translocació en hepatòcits que sobreexpres-saven G6Pasa i que, per tant, presentaven nivellsbaixos de G6P i una deposició reduïda del polisacàrid.
Figura 6. Distribució de la LGS en hepatòcits en cultiuincubats sense glucosa (control), glucosa 10 mM,glucosa 25 mM i DHA 10 mM.
2.1Hem construït un adenovirus recombinant quepermet l’expressió de la proteïna reguladora de laglucoquinasa (GKRP), el qual s’ha utilitzat peravaluar l’impacte metabòlic de la sobreexpressiód’aquesta proteïna en hepatòcits primaris de rata encultiu.
La sobreexpressió de la GKRP va produir unadisminució de la GK llibre, la qual cosa indicava quela GKRP s’unia a la GK en l’hepatòcit intacte. Amés, es va observar una disminució en la fosforila-ció de glucosa, en la síntesi de glucogen i en laglicòlisi. Per tant, es va aconseguir modificar la utilit-
zació de glucosa per l’hepatòcit mitjançant el canvide la relació molar GK/GKRP.
Els nostres primers estudis van mostrar que elcoeficient de control de la GKRP en la síntesi deglucogen era negatiu i alt, per sobre de la unitat envalor absolut (-1.2), mentre que el seu coeficient decontrol en la glucòlisi era inferior. Probablement, aixòés d’aquesta manera perquè ambdues vies tenen unadependència diferent per la G6P, ja que la LGSnecessita la G6P produïda per la GK per activar-se.
En conseqüència, aquests primers experimentsdemostraven que la utilització de glucosa per l’hepa-tòcit depèn de la relació molar GK/GKRP, ja que lasobreexpressió d’aquesta darrera en hepatòcits derata origina una disminució en l’afinitat de l’hepatòcitper la glucosa. Tot seguit vam començar a fer experi-ments in vivo amb l’objectiu d’escatir el paper fisiolò-gic de la relació entre la GK i la seva proteïna regula-dora. S’han fet experiments amb rates en dejunidurant 3 dies que permetien observar disminucionsimportants en la relació GK/GKRP. Després d’aquestdejuni s’aïllaven hepatòcits d’aquests animalsparal·lelament amb d’altres de rates que havien estatalimentades ad libitum. En els hepatòcits d’animals endejuni, la relació GK/GKRP era un 60% inferior a ladels aïllats d’animals alimentats. El descens en larelació molar GK/GKRP induïda pel dejuni era paral·lel auna disminució en la fosforilació de glucosa (figura 7).
Figura 7. Relació entre els valors de síntesi de glucogen i la
fosforilació de glucosa en hepatòcits.
Animals en dejuni
Animals alimentats
D’una manera sorprenent, no es va observar unareducció en la síntesi de glucogen en aquestasituació. D’altra banda, els nivells de PEPCK erensignificativament més alts en els animals en dejuni,mentre que no s’observaven diferències en l’activitatG6Pasa entre els grups assajats (figura 8). Enconseqüència, es pot concloure que durant elsperíodes de dejuni, l’enzim PEPCK, en trobar-seaugmentat, origina un increment en la taxa gluconeo-genètica que produirà un increment en la síntesi deglucogen, la qual cosa compensaria la disminució dela síntesi d’aquest polisacàrid per mitjà de la fosforila-ció de glucosa.
Aquests resultats demostren que en la situació in vivo,el fetge podria adaptar el seu consum de glucosa a
RECERCA BIOMÈDICA | 141

les condicions d’alimentació de l’organisme per mitjàde la modulació de la relació molar GK/GKRP, sensecomprometre-hi les reserves de glucogen hepàtic. Pertant, un possible paper fisiològic de la GKRP seriamodular el consum de glucosa en sang pel fetgemitjançant la inhibició/activació de la GK en funció dela relació molar GK/GKRP.
Figura 8. Nivells de PEPCK en hepatòcits aïllats d’animals en
dejuni perllongat i alimentats.
Animals en dejuni
Animals alimentats
2.2Es van fer diverses construccions que van permetrel’expressió heteròloga en hepatòcits primaris de laGKRP fusionada a GFP (proteïna verda fluorescent).La localització intracel·lular de la fusió GKRP-GFP vaser sempre nuclear en les diverses condicions experi-mentals realitzades (figura 9A). Les immunofluorescèn-cies d’hepatòcits no transfectats també van revelaruna concentració nuclear de la GKRP hepàticaendògena, tant amb baixa com alta concentració deglucosa (figura 9B).
Figura 9. Efecte de la fusió de la GFP en la localització subcel·lular
de la GKRP i GK. A) Localització intracel·lular de les proteïnes de
fusió GFP-GK i GKRP-GFP amb baixa (1a columa) i alta (2a
columna) concentració de glucosa. B) Localització intracel·lular de
la GK i GKRP endògenes amb baixa (1a columna) i alta (2a
columna) concentracions de glucosa. Barra. 10 µM.
Hi ha evidències experimentals que indiquen que enpresència d’altes concentracions de glucosa es dónauna translocació parcial de la GKRP vers el citoplas-ma (Mukhtar et al., FEBS Lett. 462:453). Els experi-ments d’immunofluorescència mostrats en la figura 10no permeten la quantificació de la translocació de laGKRP, per la qual cosa es va dur a terme la separacióde les fraccions nuclear i citoplasmàtica d’hepatòcitsincubats a dues concentracions de glucosa, 5 mM i25 mM. Posteriorment es va analitzar l’abundància deGKRP per mitjà de Western blot. La figura 10 mostra
una disminució en la intensitat de la banda correspo-nent a la fracció nuclear en els hepatòcits incubatsamb glucosa 25 mM en comparació amb els incubatsamb glucosa 5 mM. En canvi, no es va observar unincrement significatiu en la fracció citoplasmàtica coma resposta a l’augment de la concentració de glucosa.
Figura 10. Anàlisi per Western blot de la translocació de la GKRP.
Fracció nuclear (1) i citoplasmàtica (2) d’hepatòcits incubats amb
glucosa 5 mM. Fracció nuclear (3) i citoplasmàtica (4) d’hepàtocits
incubats amb glucosa 25 mM.
2.3Es van fer estudis comparatius de localitzaciósubcel·lular de GKRP en hepatòcits aïllats de ratesdiabètiques ZDF en diverses situacions metabòliques.Els nostres resultats indiquen que no hi ha diferènciessignificatives entre els animals sans i diabètics en elventall de situacions experimentals assajades.
3. Experiments no programats3.1Hem sobreexpressat la fructosa 1,6 bisfosfatasa enhepatòcits i hem observat que la seva localitzaciósubcel·lular respon a les condicions metabòliques.L’enzim transloca entre el nucli i el citoplasma. S’haestudiat la relació entre la seva distribució subcel·lular ila de la glucogen sintasa (treball en preparació).
3.2Mitjançant l’ús de la nova metodologia que hemdesenvolupat, hem demostrat que hi ha una xarxacomplexa d’intermediaris del metabolisme de laglucosa. Hem trobat que la magnitud del canvi en l’úsde G6P és una propietat funcional de la xarxametabòlica que es reflecteix en l’acumulació deproductes arran dels canvis compensatoris que esduen a terme en la reacció reversa dels cicles fútils.Aquests efectes no són predictibles fàcilment i elsnostres resultats indiquen que per comprendre lesvies metabòliques cal una aproximació global.
En aquest treball s’analitza simultàniament la distribu-ció de fluxos metabòlics des de G6P vers glucosa,lactat, síntesi de glucogen i metabolisme de lespentoses en hepatòcits de rata incubats amb glucosai diversos precursors gluconeogènics (glutamina,fructosa o lactat).
Quan es comparen els canvis en la distribució defluxos per mitjà de les diverses vies i per mitjà de lesreaccions directa i inversa dels cicles fútilsGK/G6Pasa i glicòlisi/gluconeogènesi en condicionesd’incubació amb glucosa sola o en combinació ambun precursor gluconeogènic, s’observen diferènciesimportants en la distribució de fluxos segons quinsigui el precursor gluconeogènic afegit. Els resultatsmostren que els cicles fútils no tan sols canvien ladirecció del flux de substrat mitjançant el cicle, sinóque també minimitzen l’impacte dels grans canvis en
FUNDACIÓ LA MARATÓ DE TV3

la disponibilitat de nutrients sobre les concentracionsde metabòlits intermediaris.
El fet que la incubació d’hepatòcits en presència defructosa, lactat o glutamina resulti en diverses distribu-cions, demostra que hi ha un fenotip metabòliccaracterístic diferent per a cadascun dels substratsgluconeogènics analitzats.
3. Rellevància i possibles implicacions clíniquesdels resultats finals obtingutsEls resultats presentats en aquest projecte hanpermès contribuir a l’aprofundiment de l’estudi delsmecanismes de regulació de l’acumulació i la utilitza-ció de la glucosa, com també de les seves alteracionsen la patologia diabètica.
A més, aquest estudi ha subratllat la importància de laglucoquinasa i la glucogen sintasa com a possiblesdianes per al desenvolupament de nous fàrmacs iteràpies contra la diabetis.
4. PublicacionsDe la Iglesia N, Mukhtar M, Seoane J, Guinovart JJ & Agius L
The role of the regulatory protein of glucokinase in the glucose
sensory mechanism of the hepatocyte. J. Biol. Chem. 275:
10597-10603 (2000)
Gomis RR, Cid E, García-Rocha M, Ferrer JC & Guinovart JJ
Liver glycogen synthase but not the muscle isoform differentiates
between glucose 6-phosphate produced by glucokinase or
hexokinase. J. Biol. Chem. 277:23246-52. (2002)
Gomis RR, Favre C, García-Rocha M, Fernández-Novell JM,
Ferrer JC, Guinovart JJ. Glucose 6-phosphate produced by
gluconeogenesis and by glucokinase is equally effective in activa-
ting hepatic glycogen synthase. J Biol Chem. 278:9740-6. (2003)
Ferrer JC, Favre C, Gomis RR, Fernández-Novell JM, García-
Rocha M, de la Iglesia N, Cid E, Guinovart JJ. Control of glycogen
deposition. FEBS Lett. 546:127-32. (2003)
Marin S, Lee WN P, Bassilian S, Lim S, Boros LG, Centellles
JJ, Fernández-Novell JM, Guinovart JJ, Cascante M. Dynamic
profiling of the glucose metabolic network in fasted rat hepatocy-
tes using [1,2-13C2]-glucose. Biochem. J. (en premsa 2004)
Yáñez AJ, García-Rocha M, Bertinat R, Concha II, Guinovart
JJ, Slebe JC. Subcellular localization of liver FBPase is modulated
by glucose, dihydroxyacetone and insulin. (en preparació).
RECERCA BIOMÈDICA | 143

FUNDACIÓ LA MARATÓ DE TV3
MEMÒRIA FINAL
1. Resum del projecte originalObjectiusObjectiu 11.1 Confirmar que els pacients infectats pel VHCtenen una prevalença més elevada de diabetis ointolerància a la glucosa que els pacients amb altreshepatopaties considerant les variables que podeninfluenciar el desenvolupament de diabetis (edat, sexe,antecedents familiars de diabetis, índex de massacorporal i grau histològic de l’hepatopatia). A més dela prevalença de diabetis s’avaluarà la prevalença deglucèmia anòmala en dejú (nous criteris de l’AmericanDiabetes Association) i d’intolerància a la glucosa.
1.2 Estudiar els possibles mecanismes patogènicspels quals el VHC pot contribuir a desenvolupar unadiabetis mitjançant la determinació d’autoanticossosrespecte de la cèl·lula beta, l'estudi de la secreciód’insulina i l'avaluació de la insulinoresistència.
Objectiu 2Determinar el genotip del VHC i estudiar si existeixassociació entre genotip i la resta de paràmetres
analitzats: prevalença de diabetis, autoanticossosrespecte de la cèl·lula beta, insulinosecreció, insulino-resistència i dipòsits de ferro.
Objectiu 33.1 Avaluar si existeix un augment de citoquinesproinflamatòries relacionades amb la insulinoresistèn-cia en els pacients infectats pel VHC.
3.2 Determinar si hi ha relació entre els dipòsits deferro i la secreció d’insulina i/o insulinoresistència.
3.3 Estudiar si la presència d’heterozigosi pel gen del’hemocromatosi és un factor de risc per desenvolupardiabetis en produir un major increment dels dipòsitsde ferro en els pacients infectats pel VHC.
3.4 Analitzar si la reducció dels dipòsits de ferromitjançant sagnia millora la secreció d’insulina i/o lainsulinoresistència en un subgrup del pacientsinfectats pel VHC. Aquest punt es desenvoluparà encas que es demostri que l’augment dels dipòsits deferro està implicat en el desenvolupament de diabetisen els pacients infectats pel VHC.
Títol del projecte
Virus de l'hepatitis C com a factor de risc per desenvolupar unadiabetis mellitus: estudi dels mecanismes etiopatogènics
Investigador principalCristina Hernández Pascual
InstitucióHospital Universitari Vall d'Hebron
41.036 €

DissenyEl disseny de l'estudi és del tipus cas-control.
2. ResultatsObjectiu 1Prevalença de les alteracions hidrocarbonades en elpacients infectats pel VHC.Hem demostrat que la prevalença d’alteracionshidrocarbonades (diabetis i glucèmia anòmala en dejú)són superiors en els pacients amb infecció pel VHC(n=498) que en els pacients amb altres malaltieshepàtiques anti-VHC negatius (n=144).
En considerar el grau de lesió hepàtica, vam observarque els pacients amb hepatitis crònica pel VHCpresentaven més alteracions hidrocarbonades que elspacients amb hepatitis crònica d’altres etiologies anti-VHC negatius (32% vs. 12%; p=0.0003). En analitzarels pacients amb transaminases normals (fasesinicials), la prevalença de diabetis persistia significati-vament elevada en els pacients infectats pel VHC(24% vs. 5%; p=0.003). En canvi, en els pacients ambcirrosi, malgrat que la prevalença d’alteracionshidrocarbonades era més elevada en els pacientsamb infecció pel VHC, la diferència no fou estadística-ment significativa (40% vs. 36%; p=n.s.).
En l’estudi de regressió logística, la infecció per VHC esrelacionà de manera independent amb la presènciad’alteracions hidrocarbonades tant en el total delspacients (OR: 2.26, IC95%: 1.47-3.70) com en elscasos d’hepatitis crònica (OR: 4.26, IC95%: 2.03-8.93).
En un subgrup de pacients amb hepatitis crònica nodiabètics pels criteris de glucèmia basal (50 anti-VHCpositius i 50 amb altres hepatopaties) es va fer unTTOG (test de tolerància oral a la glucosa). En elspacients anti-VHC es van diagnosticar 9 casos dediabetis (18%) i 15 casos d’intolerància a la glucosa(30%), percentatges molt superiors als detectats en elgrup anti-VHC negatiu (diabetis: 4%, intolerància a laglucosa: 18%; p= 0.02 i p=0.04, respectivament).
En conjunt, aquests resultats demostren que elspacients amb infecció pel VHC són un grup d’alt riscper desenvolupar una diabetis i que els mecanismespels quals la infecció causa la diabetis ja operen enfases inicials.
Aquests resultats es publicaren el mes de maig de2004 a la revista Diabetes Care (Lecube A, HernándezC, Genescà J, Esteban JI, Jardí R, Simó R. Highprevalence of glucose abnormalities in patients withhepatitis C virus infection. A multivariate analysisconsidering the liver injury).
Estudi dels mecanismes etiopatogènicsEstudi de l’autoimmunitat respecte de la cèl·lula betapancreàticaCom és conegut, el VHC s’associa amb diversesmalalties autoimmunitàries (crioglobulinèmia, glomeru-lonefritis, porfíria cutània tardana, síndrome deSjögren, hepatitis autoimmune), i a més, mostrahomologia amb la decarboxilasa de l’àcid glutàmic(GAD), que és un antigen dels illots betapancreàtics.Per això, vam avaluar si la infecció pel VHC podria
desencadenar una resposta autoinmunitària respectedels illots pancreàtics que contribuís a produir unadiabetis. En resum, l’estudi d’autoanticossos respectede la cèl.lula beta es realitzà mitjançant un cribratgeinicial de tots els individus amb la determinació d’anti-cossos anti-GAD (GADA) i anti-IA2. En les mostres enquè resultaren positius també es determinaren els isletcell antibodies (ICA). Els GADA i els IA-2 es determi-naren per RBA (radio binding assay) en fase fluida i elsICA per immunofluorescència indirecta sobre pàncreeshumà del grup O. La prevalença d’autoanticossosobtinguda fou: 0% en el grup de pacients diabèticsinfectats pel VHC, 1.4% en el grup de pacients nodiabètics infectats pel VHC, i 0.4% en un grup controlconstituït per 273 subjectes sans equiparats per edat isexe. Per tant, la infecció pel VHC no s’associa ambun increment dels marcadors autoimmunitaris respec-te de la cèl·lula betapancreàtica, o amb altresparaules, els mecanismes autoimmunitaris no tenenun paper rellevant en l’etiopatogènia de la diabetismellitus associada amb el VHC (Piquer S, HernándezC, Enriquez J, Ross A, Esteban I, Genescà J,Bonifacio E, Puig-Domingo M, Simó R. "Islet Cell andthyroid antibody prevalence in patients with hepatitis Cvirus infection: effect of treatment with interferon". JLab Clin Med 2001;137: 38-42).
Estudi de la secreció i de la resistència a la insulinaHem comparat la secreció d’insulina (pèptid-C basal iresposta al test del menjar estàndard i del glucagó i.v)i la resistència a la insulina (insulinèmia basal, HOMA)entre un grup de pacients anti-VHC positiu (n=40) i unaltre anti-VHC negatiu (n=20) amb hepatitis crònica.S’han estudiat pacients no diabètics per avaluar quinsson els efectes deleteris inicials del VHC sobre elmetabolisme de la glucosa. Es tracta d’un estudi cas-control i els pacients s’han equiparat per les variablesque poden influir en el desenvolupament de diabetis(edat, sexe, índex de massa corporal, índex cintura-maluc i antecedents familiars de diabetis). En resum,hem demostrat que els pacients infectats pel VHCpresenten un augment de la resistència a la insulina(HOMA grup anti-VHC positiu: 4.47±2.23 vs. HOMAgrup anti-VHC negatiu: 2.58±1.74; p=0.009) peròsense detectar diferències en la reserva pancreàticadesprés dels estímuls administrats (menjar estàndard iglucagó i.v).
S’està elaborant un article per comunicar aquestsresultats.
Objectiu 2Anàlisi dels genotips del VHC i de la seva relacióamb la prevalença d’alteracions del metabolismehidrocarbonat:La distribució de genotips detectada en la nostrapoblació fou la següent: 207 pacients amb genotip 1(83.8%), 12 pacients amb genotip 2 (4.8%), 18pacients amb genotip 3 (7.3%), i 10 pacients ambgenotip 4 (4%). No vam detectar diferències en laprevalença d’alteracions hidrocarbonades entre elsgenotipus (genotip 1: 39.1 %, genotip 2: 33%,genotip 3: 16%, genotip 4: 33%; p = n.s.). A més, enanalitzar separadament els pacients amb hepatitiscrònica equiparats per edat, la prevalença fou moltsimilar entre els genotip (genotip 1: 20%, genotip 2:
RECERCA BIOMÈDICA | 145

14.3%, genotip 3: 20% , genotip 4: 22.2%; p=n.s.).Aquests resultats estan pendents de publicació a larevista Diabetes Care (Lecube A, Hernández C,Genescà J, Esteban JI, Jardí R, Simó R. Highprevalence of glucose abnormalities in patients withhepatitis C virus infection. A multivariate analysisconsidering the liver injury).
Objectiu 3Estudi de citoquines proinflamatòries:En un estudi cas-control (40 pacients anti-VHCpositius i 20 anti-VHC negatius no diabètics), s’hanestudiat les citoquines proinflamatòries següents:TNF-alfa, els receptors solubles del TNF-a (sTNFR1 isTNFR2) i la IL-6. Hem observat un augment d’aques-tes citoquines (pg/ml) en els pacients amb hepatitiscrònica pel VHC en comparació amb subjectes ambaltres malalties hepàtiques (TNF-a : 6.47±3.44 vs.2.71±3.06 p=0.007; sTNFR1: 174±34 vs. 149±27,p=0.04; sTNFR2: 300±78 vs. 241±81, p=0.04; IL-6:3.78±1.45 vs. 2.07±1.02, p=0.002). A més, laconcentració dels receptors del TNF-alfa es correla-cionà de manera directa i estadísticament significativaamb la resistència a la insulina (HOMA). Per tant, elsnostres resultats suggereixen que la causa principalper la qual el VHC produiria més diabetis seria perl’augment de la resistència a la insulina secundària auna producció incrementada d’aquestes citoquines.Estem elaborant un article per comunicar els resultatsd’aquest estudi cas-control.
Estudi del metabolisme del ferroTant la diabetis com la infecció pel VHC s'hanassociat a augments de la ferritina sèrica. Per tant,vam avaluar quina era la contribució de la diabetis enl'augment de ferritina detectat en els pacients infectatspel VHC. Vam reclutar de manera prospectiva un totalde 619 subjectes, que es van dividir en quatre grupsen funció de la presència o no de diabetis i d'infecciópel VHC: grup A (anti-VHC positiu i amb diabetis,n=46), grup B (anti-VHC negatiu i amb diabetis,n=236), grup C (anti-VHC positiu i sense diabetis,n=173) i grup D (anti-VHC negatiu i sense diabetis;grup control, n=164). A més de la ferritina, es vandeterminar altres paràmetres sèrics del metabolismedel ferro i es va descartar l'hemocromatosi per estudigenètic. En resum, vam observar que les concentra-cions de ferritina en el grup A eren significativamentsuperiors a les detectades en els altres grups (A>B,p<0.01; A>C, p<0.001; A>D, p<0.001). Els pacientsdiabètics (grup B) mostraven concentracions de ferriti-na superiors a les del grup D (p=0.001), però elspacients anti-VHC positiu sense diabetis (grup C)presentaven concentracions de ferritina similars a ladels controls (grup D). A més, en l'estudi multivariablela presència de diabetis fou un predictor de lesconcentracions de ferritina, mentre que la infecció pelVHC no es relacionà amb les concentracions de ferriti-na. Per tant, concloem d'aquest estudi que és lapresència de diabetis la que explicaria l'augment deferritina comunicat en els pacients infectats pel VHC.Tal com s'ha esmentat, aquests resultats estant enavaluació per a una publicació a la revista JAMA.
3. Rellevància i possibles implicacions clíniquesdels resultats finals obtingutsEn aquest estudi hem demostrat que el risc de patiruna diabetis en els pacients amb hepatitis crònica pelVHC és 4 vegades superior al de la població general i,per tant, és un grup en el qual el despistatge dediabetis s’ha de recomanar de forma periòdica.
En l’actualitat, la infecció pel VHC és la causa principalde malaltia hepàtica en la majoria dels països i s’esti-ma que un 3% de la població mundial està infectada.A més, arran de les campanyes sanitàries, cadavegada es diagnostiquen més casos d’infecció enpacients asimptòmatics, en fase de baix grau defibrosi hepàtica. En estudis recents, s’ha posat demanifest que la progressió vers una hepatopatiaavançada és menor a la considerada prèviament. Defet, es considera que la durada mitjana entre lainfecció i el desenvolupament d’una cirrosi és de 46anys per als homes infectats a l’edat de 25 anys.Entre les dones infectades a la mateixa edat, menysdel 30% progressaran a cirrosi després de 50 anysd’infecció. Per tant, en els pacients infectats pel VHCs’han d’implementar les mesures terapèutiques perassolir els mateixos objectius de bon control glucèmici de la resta de factors de risc cardiovascular que a lapoblació general.
Hem observat que la diabetis en els pacients infectatspel VHC es produeix per un augment de la resistènciaa la insulina secundari a una major producció decitoquines proinflamatòries (especialment TNF-alfa).En canvi, no hem observat defectes en la secreciód’insulina, almenys en les fases inicials de la infecció.Aquests resultats ens suggereixen que els antidiabè-tics d’elecció en aquests pacients seran els quemillorin la resistència a la insulina i, sobretot, aquellsfàrmacs que redueixin la producció de TNF-alfa.
Hem detectat que l’augment de ferritina observat enels pacients amb infecció pel VHC es produeix aexpenses dels pacients diabètics (més del 30% delspacients infectats són diabètics). Mitjançant ladeterminació dels receptors solubles de la transferrina,que són un bon marcador dels dipòsits de ferro i noes comporten com a reactants de fase aguda, hemdemostrat que l’elevació de ferritina a la diabetis tipus2 no reflecteix un augment dels dipòsits de ferro, sinóel procés inflamatori subjacent a la diabetis. Des delpunt de vista de la pràctica clínica, aquesta troballaens indica que la ferritina no és un paràmetre útil en ladiagnosi de l’anèmia ferropènica en els pacientsdiabètics tipus 2.
4. PublicacionsPiquer S, Hernández C, Enríquez J, Ross A, Esteban I,
Genescà J, Bonifacio E, Puig-Domingo M, Simó R. Islet Cell and
thyroid antibody prevalence in patients with hepatitis C virus
infection: effect of treatment with interferon. J Lab Clin Med
2001;137: 38-42.
C.Hernández, A.Lecube, J.Genescà, J.Mesa. R.Simó. Iron
stores as the link between diabetes and hepatitis C virus infection:
case-control study. 60th Scientific Sessions of the American
Diabetes Association (ADA). San Antonio, 9-13 de junio de 2000.
Diabetes 2000; 49 (suppl 1): A343.
Lecube A, Hernández C, Genescà J, Esteban JI, Jardí R,
FUNDACIÓ LA MARATÓ DE TV3

Carreras A, Mesa A, Simó R. Insulin resistance is associated to
iron stores in non-diabetic patients with hepatitis C infection.
62nd Scientific Sessions of the American Diabetes Association
(ADA). San Francisco, junio 2002. Diabetes 2002;51 (suppl 2):
A585
Lecube A, Hernández C, Genescà J, Carreras A, Mesa A,
Simó R. Resistencia a la insulina y depósitos de hierro en los
pacientes infectados por el virus de la hepatitis C. 44 Congreso
de la Sociedad Española de Endocrinología y Nutrición. Alicante,
mayo 2002. Endocrinología y Nutrición 2002;49(supl. 1):24
Lecube A, Hernández C, Genescà J, Esteban JI, Jardí R, Simó R.
Prevalencia de alteraciones hidrocarbonadas en los pacientes
infectados por el virus de la hepatitis C. XVI Congreso de la
Sociedad Española de Diabetes. Cádiz, marzo 2002. Avances en
Diabetología 2002;18 (Supl.1): 37.
Lecube A, Hernández C, Genescà J, Esteban JI, Jardí R, Simó R.
High prevalence of glucose abnormalities in patients with hepatitis
C virus infection. A multivariate analysis considering the liver injury.
Diabetes Care (en premsa)
Hernández C, Lecube A, Carrera A, Simó R. Soluble transferrin
receptors and ferritin in type 2 diabetic patients. Diabetic Medicine
(en premsa)
Lecube A, Hernández C, Genescà J, Esteban, JI, Jardí R,
García L, Simó R. Diabetes is the main factor accounting for the
high ferritin levels detected in patients with hepatitis C virus
infection. (enviat per a publicació).
A.Lecube, C.Hernández, L.García, J.Genescà, J.Mesa, R.Simó.
Diabetes is the main factor related to the high ferritin levels
detected in patients with Hepatitis C virus infection. 63rd Scientific
Sessions of the American Diabetes Association. New Orleans,
juny 2003. Diabetes 2003; 52 (suppl 1): A541
Lecube A, Hernández C, Genescà J, Mesa J, Simó R. High
prevalence of glucose abnormalities in patients with hepatitis C
virus infection. A large cohort study considering the liver damage.
Congrés: 18th Congress of the International Diabetes Federation.
24-29 d’agost. París. Diabetologia 2003; 46 (suppl 2): A148.
RECERCA BIOMÈDICA | 147

FUNDACIÓ LA MARATÓ DE TV3
MEMÒRIA FINAL
1. Resum del projecte originalS’ha ideat un instrument específic per a l’avaluació dela qualitat de la informació sobre diabetis mellitus (DM)que contenen els llocs web d’Internet identificats ambuna estratègia de cerca específica. Els resultats del’anàlisi s’han comparat amb el contingut de duesguies de pràctica clínica sobre DM seleccionades perla metodologia AGREE. S’ha fet una anàlisi transversaldescriptiva i una altra de seguiment longitudinal en lainformació obtinguda pels principals cercadorsd’Internet triats, fins a un total de 8, per la qual cosaes va adoptar la perspectiva del pacient o usuari delsserveis sanitaris en el procés de cerca.
2. ResultatsS’han recuperat 74 llocs web avaluables no repetits, ien 64 casos (86,2%) la revisió de temes ha estat eltipus d’informació recuperada amb més freqüència.En 53 llocs web (71,6%) no es va identificar explícita-ment l’audiència a la qual anava adreçada la informa-ció. Quant a la procedència del lloc web recuperat, 19documents (25,7%) corresponien a les organitzacionssanitàries, i 17 (23%) a les societats mèdiques especia-
litzades, i de 16 (22,5%) no s’ha pogut identificar.
Pel que fa a aspectes de l’estructura de la informaciórecuperada, no s’esmenta: el nom de l’autor o autorsde la informació en 41 llocs web (55,4%), ni lescredencials dels autors en 51 (68,9%), ni la presènciade patrocinadors en 66 (89,2%). No consta la data decreació del lloc web en 70 llocs web recuperats(94,6%) ni la d’actualització en 59 (70,3%).
En 49 llocs web (66,2%) no hi ha detallada cap citacióbibliogràfica que completi o aporti més informació alsusuaris de la informació recuperada. En 68 (91,9%) nos’identifica en els llocs web cap codi de conducta ovalidació. En relació amb el rigor científic, no s’explici-ta en 71 (95,9%) l’existència d’un procés de revisióeditorial i no s’identifica en 44 (59,5%) alguna paraularelacionada amb la classificació de l’evidència científi-ca segons que sigui assaig clínic o aleatoritzat o béalgun altre tipus d’evidència. En 46 (62,2%) nos’ofereix la possibilitat d’ampliar més informació ambels autors per mitjà d’una adreça de correu electrònic,telèfon o enllaç amb un altre lloc web.
Pel que fa a aspectes del contingut de la informació
Títol del projecte
Investigació sobre la qualitat de la informació sobre diabetis mellitusdisponible per internet. Construcció i validació d'un instrument
d'avaluació de la qualitat
Investigador principalAlbert J. Jovell Fernández
InstitucióFundació Biblioteca Josep Laporte. UAB
28.474 €

recuperada, en 49 llocs web recuperats i avaluats(66,2%) la informació feia referència al concepte ietiologia de la DM, en 25 (33,8%) a diagnòstic, en30 (40,5%) a tractament, en 9 (12,2%) a prevenció ien 22 (29,7%) a complicacions. Seguint les recoma-nacions de les guies de pràctica clínica publicades iemprades com a estàndard de comparació enaquest treball, la informació es va considerarcompleta en 26 llocs web de concepte i etiologia(35,1%), en 14 de diagnòstic (18,9%), en 18 detractament (23,3%), en 5 (6,8%) de prevenció i en 5dels de complicacions (6,8%).
S’ha trobat una associació positiva significativa, r=0,410(p=0,000) entre la posició segons el criteri del cercador ila posició segons el criteri de qualitat elaborat. Laposició d’una pàgina en un cercador i la seva qualitaten relació amb la resta de pàgines obtingudes amb elmateix cercador estan associades positivament —commillor posició, més qualitat de la informació. Així, lespàgines que es troben en millors posicions ocupen lesprimeres posicions segons el criteri de qualitat, és a dir,tenen puntuacions més altes.
La mitjana de posicions que variarien les pàgines sis’ordenessin per cada cercador en funció de laqualitat de la informació és igual per a tots elscercadors. Sí s’han trobat diferències entre tipusd’informació en relació amb la discrepància entre elcriteri actual i el criteri de qualitat proposat. Si latemàtica de la informació és el diagnòstic (p=0,007) icomplicacions (p=0,023), hi ha evidència que elnombre de posicions seria més alt, és a dir queaquestes pàgines es veurien afavorides en adoptar elcriteri de qualitat proposat.
El càlcul de la variable "diferència de rangs" es potinterpretar com el nombre de posicions que variaria lapàgina en una cerca per Internet si les pàginesobtingudes en el cercador s’ordenessin en funció dela qualitat de la informació en comptes del criteri delcercador. Si es calcula l’interval de confiança per a lavariable "diferència", es pot afirmar amb una confiançadel 95% que el nombre de posicions que, com amitjana, variaria una pàgina si es considerés el criteride qualitat relativa, no seria superior a 1,53 (cap amillor o pitjor); així, és molt probable que la pàginaarribés a canviar dues posicions. L’interval de confian-ça del 95% (I95%:-1,53,1,53) està centrat en el 0, perla qual cosa no hi ha evidència, amb la informació del’estudi, que el criteri de cada cercador discrepi delcriteri de reordenar les pàgines obtingudes segons laseva puntuació en el qüestionari.
Sobre la qualitat de les pàgines web recuperades, nohi ha evidència per afirmar que és diferent segons elcercador emprat per recuperar la informació (p=0,51).En relació amb l’audiència de la pàgina web, no s’hidetecten diferències significatives, amb una probabilitatno significativa però improbable (p=0,082), ni tampocsegons la procedència de la pàgina web (p=0,168). Laqualitat de la informació és diferent si el tipus d’infor-mació de la pàgina és de diagnòstic (p=0,001), tracta-ment (p=0,000) o complicacions (p=0,001). La qualitatmillora si la pàgina conté informació sobre diagnòstic,tractament i complicacions.
Dels 11 mesos en què pot canviar l’evolució d’unapàgina, com a mitjana les pàgines estan 7 mesossense variar de posició, en 2 milloren la posició i en 2l’empitjoren. Aquesta troballa és congruent amb el fetque, com a mitjana, les pàgines mantenen durant 4mesos la tendència que havien tingut entre el primer iel segon mes d’observació.
Les pàgines canvien una mitjana de 5,15 vegades detendència (de guanyar posicions a perdre-les omantenir-les, de mantenir-les a guanyar-les o perdre-les...), amb la qual cosa cada tendència té una duradamitjana de 2,69 mesos.
3. Rellevància i possibles implicacions clíniquesdels resultats finals obtingutsEn l’anàlisi que s’ha fet, la posició d’una pàgina webde DM en la jerarquia del cercador està relacionadaamb el seu nivell de qualitat, obtingut per mitjà delprotocol específic d’avaluació. Aquest fet es mantéconstant per a tots els cercadors analitzats i durant totel temps de durada de l’estudi.
Les pàgines o llocs web que tracten dels temes dediagnòstic, complicacions o tractament de la DMestan infravalorades en els cercadors, per la qual cosala seva posició en la jerarquia fóra superior si estigues-sin ordenades per criteris de qualitat basats enl’evidència científica.
Cap cercador entre els analitzats destaca sobre elsaltres en la selecció del nivell de qualitat de les diversespàgines web, de les que hi ha en les primeres 30posicions, que ha estat l’objecte de l’anàlisi.
El nivell de qualitat d’un lloc o pàgina web augmentasi l’abast temàtic és més gran, tot destacant en unabast parcial les que inclouen diagnòstic, tractament icomplicacions.
Les posicions jeràrquiques dels llocs web en lesprimeres 30 posicions no varien durant els mesos, iaquest fet és constant per a tots els cercadorsseleccionats. Durant els 11 mesos de seguiment, enuna mitjana de 7 mesos les pàgines web no variende posició.
La qualitat de la informació sobre diabetis per Internetés possiblement d’una qualitat superior que la trobadaper a altres patologies descrites en la bibliografia.
4. PublicacionsJovell E, Jovell AJ. Calidad de la información en Internet. Siete
Días Médicos num 600
RECERCA BIOMÈDICA | 149

FUNDACIÓ LA MARATÓ DE TV3
MEMÒRIA FINAL
1. Resum del projecte originalLa hipòtesi general del Projecte OPTIDIAB ésdemostrar que en el nostre àmbit sanitari l’ús delsrecursos avançats de les tecnologies de la informació(TI) i de la telemedicina (TM) contribueixen d’unamanera impactant i eficient en els costos en l’optimit-zació del tractament de la diabetes mellitus (DM).L’equip investigador del Projecte OPTIDIAB estàcoordinat pel GBT (Grup de Bioenginyeria iTelemedicina, Universitat Politècnica de Madrid), ambuna experiència llarga i acreditada en recerca tecnolò-gica de l’àrea de les telecomunicacions, com a equiptecnològic principal, i amb el Grup Investigador en TI iTM del Servei d’Endocrinologia i Nutrició de l’Hospitalde Sant Pau com a grup de recerca coordinador delprojecte.
Tot seguit es resumeixen les activitats dutes a termefins ara en el si del Projecte:A.1. Desenvolupament del cercle de qualitat(DiabCare)S’ha estructurat el cercle de qualitat (CQ) correspo-nent (metges, infermeres especialistes en diabetis, un
analista de sistemes informàtics i una administrativa).El programa de gestió clínica elaborat ad hoc(DiabCare Data for Windows) ha avaluat en total 141indicadors de qualitat assistencial i els ha transferit alprimer servidor per fer factible l’anonimització de lesdades i fer-ne l’anàlisi estadística (paquet SPSS).L’avaluació obtinguda ha permès reconèixer deficièn-cies en l’emplenament del registre i la identificació depacients considerats d’alt risc. L’avaluació vacomprendre també els indicadors determinants del’estatus de la Declaració de Saint Vincent. Com acentre coordinador a Espanya del Consorci DiabCareQuality Network (DQN) de la Comissió Europea, s’hadonat servei de promoció, entrenament i ajuda logísti-ca a un total de 27 centres i institucions sanitàries (24hospitals i 3 centres d’atenció primària), tant aCatalunya com a la resta de l’Estat.
A.2. Programa especial de diabetes i embaràsEl programa Optidiab té un interès especial en laproblemàtica determinada per la gestació complicadaper diabetis, o diabetis gestacional (DG), per la qualcosa participa en l’equip internacional responsable del’elaboració d’una base de dades i un programainformàtic de registre europeu i d’avaluació de qualitat
Títol del projecte
OPTIDIAB: model de recerca per a la gestió i l'economia de la salut,i d'optimització de la qualitat de l'assistència integrada
a la diabetis mellitus
Investigador principalAlberto de Leiva Hidalgo
InstitucióHospital de la Santa Creu i Sant Pau
96.343 €

assistencial en el problema de la gestació complicadaper diabetis (HIB/BIS, DM and Pregnacy Clinic). Elprograma registra i analitza paràmetres rellevants de lahistòria obstètrica prèvia, consell preconcepcional,benestar maternal i fetal, evolució del part, i ha facilitattant la reclassificació de la diabetis gestacionalpostpartum com l’estudi del curs evolutiu de latolerància a la glucosa i altres paràmetres clínics imetabòlics d’aquestes dones en el seu seguimentpostpartum. Es va fer una prova pilot que va incorpo-rar l’avaluació dels paràmetres registrats en la HIB enl’elaboració d’algoritmes d’ajuda per a l’optimitzaciódel tractament metabòlic de la DG. En un primerexperiment, el sistema informatitzat suggereix laproposta algorítmica corresponent, basada en lamonitorització glucèmica i els antecedents terapèuticsprevis. Davant d’aquesta situació, l’expert defineix si laproposta és difícil d’interpretar, si la informaciódisponible és suficient per prendre la decisió, o si s’had’ajornar durant un període curt de temps, i obtenirmentrestant més informació complementària. Aquestaprova va permetre detectar la necessitat d’introduirajustaments terapèutics en el 92% de les pacients, totacceptant el grup d’experts consultats el 80% delsalgoritmes proposats per la xarxa neuronal de predic-ció (intel·ligència artificial).
A.3. Programa de qualitat assistencial: sistemesDiabCard i QualidiabEl Projecte DiaCard ha dut a terme l’especificació d’unsistema d’informació clínica en una targeta intel·ligent(microxip), que incorpora tant el registre clínic electrò-nic com la monitorització i la conducció de l’assistèn-cia del pacient; un aspecte fonamental d’aquestaaplicació tecnològica va ser facilitar la comunicació il’intercanvi d’informació entre els diversos professio-nals sanitaris, amb caràcter interactiu, com tambéentre el pacient i els diversos nivells de l’assistènciaclínica. Els anys 2000 i 2001 es va dissenyar unavariant de la targeta intel·ligent original, en llengüesfrancesa, castellana i catalana ("Sistema Qualidiab"),que a hores d’ara està acceptat com a eina bàsica alspaïsos de l’Amèrica Central i del Sud, d’acord amb elsprincipis de la Declaració de les Amèriques sobrel’assistència en la malaltia diabètica. El programa delsistema, desenvolupat a la nostra institució encol·laboració amb la Fundació Diabem, va ampliard’una manera considerable les possibilitats prèviamentdesenvolupades en el Programa DiabCare, totgenerant-se un total de 19 unitats o mòduls per a lavariant d’aplicació a la Unitat Hospitalària de Diabetis, i9 mòduls per a l’execució als centres d’atencióprimària. La dificultat a disposar de garanties demanteniment del sistema (fabricació de les targetesseleccionades, lectors electrònics d’aquestes, encrip-tament de la informació, compatibilitat, etc.), elcompromís econòmic determinant, i el desenvolupa-ment ràpid d’altres tecnologies més avançades(mòdem, Internet, entre d’altres) van determinar lainterrupció del desenvolupament del sistemaDiabCard en favor del desenvolupament del sistemesDiabTel, T-IDDM i M2DM.
A.4. Programa de telemedicina: sistema DiabTelDissenyat, originalment, des del punt de vista tecnolò-gic per GBT, l’arquitectura del sistema disposa de dos
components principals: a) l’estació de treball o UnitatMèdica (UM), i b) l’estació receptora o Unitat delPacient (UP), tot comunicant-se ambdues d’unamanera interactiva per mitjà de la xarxa telefònicapública (mòdem). La UM es va instal·lar en un PC isuporta les funcions de telemonitorització, teleassis-tència, i també l’educació o informació sanitària delpacient i el públic en general; pot incorporar instru-ments d’ajuda en la presa de decisions, la històriaclínica electrònica i l’elaboració adequada d’informesclínics. La UP es va instal·lar en un PC Palmtop, HP.Es tracta d’un ordinador de butxaca, fàcil d’usar, quefa possible l’enregistrament, la monitorització metabò-lica, dietètica i del règim de vida, les consultes telemà-tiques, l’enviament de missatges, la presa dedecisions respecte de les modificacions/ajustamentsde la terapèutica.
A.5. Programa de telemedicina: sistemes T-IDDM iM2DMEl sistema T-IDDM utilitza la cooperació de la UnitatMèdica (UM) i la Unitat Pacient (UP) connectades en lanostra experiència per Internet. La UM ajuda el clínicresponsable de l’assistència a definir el règim d’insuli-na del pacient, d’acord amb l’avaluació periòdica dela informació clínica facilitada pel pacient. La UP ajudael pacient a la monitorització diària de la glucèmia(recepció directa de la determinació per mitjà delglucòmetre; ús de l’algoritme escaient d’ajustamentde la dosi d’insulina diària, segons l’esmentadaautomonitorització; activitat física practicada i alimen-tació ingerida). Les dues unitats operen habitualmentd’una manera asincrònica. La UM integra la coopera-ció de diversos servidors: 1) base de dades (Oracle);2) abstracció temporal (Common Lisp); 3) anàlisi dedades (Common Lisp); 4) ajuda en la decisió(Common Lisp); servidor web (Common Lisp). Elsistema d’ajuda en la decisió incorpora la metodologiade xarxa neuronal probabilística d’acord amb el càlculde la resposta glucèmica diària del pacient i elconcepte d’abstracció temporal. La UP utilitzaWindows amb un port en sèrie secundari RS232 percomunicar-se amb el reflectòmetre de glucosa, iconté: a) l’agenda diària de valors glucèmics (possibili-tat d’expressió gràfica), dieta, protocol insulínic,glucosúria/cetonúria, dades personals del pacient(filiació, dades clíniques i antropomètriques), b) lanormativa de consulta terapèutica; c) missatgeselectrònics; d) comunicacions; e) el sistema adminis-trador de l’aplicació. La base de dades és Paradox5.0. La verificació tècnica del sistema es va dur aterme per mitjà del procés de simulació de pacients,tot resultant satisfactòria. El Projecte M2DM, un altreconsorci de TM de la CE investiga la potencialitat delsserveis de multiaccés en l’assistència de la DM, ambla finalitat de proveir un servei de televigilància,integrant-hi amb bona relació cost-eficiència equipsde comunicació diversos (web, telèfon mòbil, telèfonestàndard, Internet, GPRS, etc.). Les diverses modali-tats de comunicació s’integren en un servidor organit-zador de multiaccés. El sistema permet eines d’ajudaper a la decisió (models matemàtics), anàlisi automàti-ca de les dades, generació automàtica d’alarmespreestablertes. L’avaluació clínica només va inclourepacients tractats amb insulina, de manera convencio-nal o amb infusió continuada, visitats ja fos d’una
RECERCA BIOMÈDICA | 151

manera estàndard o bé experimental (randomització)als 0, 3, 6, 9, 12 mesos. Els paràmetres d’avaluacióvan ser: indicadors clínics, aspectes organitzatius ieconòmics, qüestionaris de qualitat de vida i satisfac-ció del pacient, usabilitat i avaluació tècnica.
A.6. Sistema de documentació i informació endiabetis de l’Hospital de la Santa Creu i Sant PauUna aplicació d’interès i benefici immediat en l’assis-tència dels pacients diabètics, resultant del ProjecteOPTIDIAB, ha estat l’elaboració, en col·laboració ambel Servei d’Informació i Gestió del Departamentd’Informàtica i Comunicació de l’Hospital d’un sistemaglobal d’informació i documentació electrònica, al quales pot accedir, amb l’autorització corresponent, permitjà del portal de la pàgina web, i que té l’índexsegüent de continguts.
El sistema conté:a) El conjunt d’informes clínics elaborats pels profes-sionals sanitaris per respondre a les necessitats de lesdiverses àrees assistencials, com a garantia dequalitat i bases de dades (n=12).
b) El conjunt de documents de caire informatiu per alsmateixos malalts diabètics i les seves famílies (n=13).
c) Els protocols electrònics d’assistència en la diabetisadreçats a l’atenció primària (n=7).
2. ResultatsB. 1. Programes de qualitat assistencialB. 1.1. Programa DiabCareDurant el primer any del projecte es van incloure enl’estudi 870 pacients, atesos entre l’1 de gener i el 31de desembre. La mostra va representar el 79% deltotal de pacients registrats amb codificació diagnòsticapertinent, que van atorgar el consentiment correspo-nent després de la informació donada per l’equip derecerca. Tot seguit s’esmenten les troballes principals:
1. La població de pacients amb DM-2 va presentaruna prevalença superior de ceguesa, cardiopatiaisquèmica, malaltia cerebrovascular, insuficiència renalcrònica avançada, i amputació major d’extremitatsinferiors respecte dels pacients amb DM-1.
2. EL 88% dels pacients practicava l’automonitoritza-ció domiciliària de la glucèmia capil·lar, amb unafreqüència superior en els pacients amb DM-1.
3. El 96% dels pacients diabètics va ser objected’avaluació ocular (AV, fons ocular, etc.), per l’endocri-nòleg iy/o l’oftalmòleg.
4. El 35% del total de pacients presentava retinopatiadiabètica, corresponent en el 85% d’aquests a laforma no proliferativa. Entre els pacients amb DM-2 varesultar més freqüent la presència de cataractes,maculopatia diabètica, i malaltia ocular avançada.
5. Es va fer exploració física dels peus en el 95% delspacients, el 38% dels quals mostrava alguna/esanormalitats, amb predomini de l’alteració de lapalestèsia, la resposta sensitiva al monofilament, il’absència de polsos pedis.
6. El 20% dels pacients va comparèixer durant l’anyamb una (o més) urgència/ies metabòliques, sobretothipoglucèmcia en pacients amb DM-1.
7. El 90% dels pacients diabètics tipus 1 rebienteràpia insulínica intensificada. El 3,4% d’aquestasubpoblació utilitzava bombes d’infusió contínuad’insulina (BICI).
8. La majoria dels pacients amb DM-2 rebia tracta-ment farmacològic antihiperglucemiant. Només el13% restava amb tractament exclusivament higienico-dietètic. Dels que rebien tractament insulínic, el 78%administraven dues o més dosis diàries; el 2,6%d’aquests estaven inclosos en el programa BICI.
9. El 9,4% dels subjectes amb DM-1 i el 45% delssubjectes amb DM-2 s’administraven tractamentfarmacològic antihipertensiu de més o menys comple-xitat. Rebien agents antihipertensius el 79% delscasos d’hipertensió associada a DM-1 i el 57% delsd’hipertensió associada a DM-2.
10. El 23% dels individus amb DM-2 eren tractatsamb agents antihiperlipemiants. El perfil de lípidsplasmàtics es va considerar acceptable en el 64%dels pacients amb DM-1 i el 21% dels pacients ambDM-2.
11. Només el 12% dels pacients amb DM-1 versus el13% dels pacients amb DM-2 mostrava un controlmetabòlic òptim (HbA1c<6,5%).
Una informació més àmplia i la discussió d’aquestsresultats aquí enumerats, es pot consultar en eldocument "Gestió clínica informatitzada de les presta-cions sanitàries al pacient amb diabetis mellitus.Projecte Optidiab. Fase I: desenvolupament continuatde la qualitat assistencial (Programa DiabCare). Tesina,programa de màster en Economia de la Salut i GestióSanitària, Universitat Pompeu Fabra, que va merèixerla màxima qualificació (desembre, 2000).
B.1.2. Programa DiabCard/QualidiabEls resultats obtinguts per l’experiència de l’equipinvestigador en aquest programa indiquen que elPrograma DiabCard ofereix un sistema de registreclínic, avaluació de qualitat assistencial, i promoció dela comunicació interprofessional i pacient-sistemasanitari, interessant d’explotar. Les proves fetes ambprototips diversos van palesar deficiències tecnològi-ques (bàsicament de maquinari) determinants deperllongació de la durada mitjana de la visita mèdica(18 min per al grup control versus 27 minuts per algrup DiabCard), i d’acceptabilitat limitada pels clínicsque hi van col·laborar. A tot això, cal afegir-hi aspecteslogístics que van superar l’esfera del protocol derecerca clínica proposat (necessitat de tenir lacol·laboració d’una empresa industrial especialitzadaen la producció de targetes amb microxip, crear unmercat per a aquest producte, i aspectes de seguretatper resoldre, com ara l’encriptament selectiu de lainformació i protecció de dades individuals, a més deles limitacions pressupostàries relacionades amb ladotació per a OPTIDIAB). Per tot plegat, l’equip investi-gador va decidir no continuar les experiències en
FUNDACIÓ LA MARATÓ DE TV3

aquest terreny, malgrat les perspectives positives d’unprograma amb una acollida excel·lent i amb satisfacciódels pacients. Després de l’experiència de Barcelona,altres grups europeus han continuat avaluant DiabCard(a Perugia, Viena, Corbeil, Kassel, Tessalònica), conjun-tament amb un altre projecte de la CE (Cardiolink).D’altra banda, actualment el programa es manté actiuen diversos països de l’Amèrica Llatina (variantQualidiab, generada inicialment per la col·laboracióentre DiabCard-Espanya i DiabCard-França).
B.1.3 Programa especial de diabetis i embaràsL’experiència del Sistema OPTIDIAB (full d’informacióbàsic per a diabetis i embaràs; protocols de gestió iajuda en la decisió clínica en diabetis i embaràs, entrealtres components) ha resultat especialment fructíferaen el desenvolupament d’iniciatives diverses, que handeterminat avenços importants, tant en la gestió del’atenció clínica a l’embarassada amb diabetis, comen la creació d’una eina de gran vàlua per a unaavaluació més precisa i completa de la qualitatassistencial oferta a les gestants amb diabetis,escaientment objectivada per mitjà de l’anàlisi delsparàmetres de benestar maternofetal. L’equip derecerca ha descrit amb detall les característiques deles aplicacions especials del Programa Optidiab a lagestió clínica i qualitat assistencial en la gestaciócomplicada amb diabetis, en un capítol de revisió d’unllibre editat, al qual remetem per ampliar els contingutsdescrits en aquesta secció (A de Leiva et al. "Qualityof Care for the woman with diabetes and pregnancy".En: M Hod, L Jovanovic, GC Di Renzo, A de Leiva, OLanger [ed.] Textbook of Diabetes andPregnancy,Martin Dunitz. London, 2003).
S’indiquen tot seguit els resultats més rellevantsobtinguts. La informació ampliada es pot consultar enel conjunt d’articles originals publicats com aconseqüència de les recerques relacionades ambaquest projecte, ressenyats en aquest document.
B.1.3.1. Sistema d’ajuda en la decisió en el platerapèutic de la diabetis gestacionalEn col·laboració amb el grup investigador GBT es vaavaluar el sistema d’ajuda a la gestió clínica, basat enl’operació de la xarxa probabilística causal centradaen l’efectivitat de l’ajustament de la dosi terapèuticad’insulina i de les característiques de la dieta.Aproximadament el 80% de les propostes generadespel sistema d’"intel·ligència artificial" van ser accepta-des pels experts consultats. L’experiència adquiridaen aquest estudi ha servit de base per als procedi-ments de predicció automatitzada inclosos en progra-mes terapèutics posteriors. (Per a informació addicio-nal, consulteu l’article original derivat d’aquestarecerca, M E Hernando et al. "Evaluation of DiabNet, adecision support system for therapy planning ingestational diabetes." Computer Methods andPrograms in Biomedicine, 2000;62:235-248).
B.1.3.2. Efectivitat del personal d’infermeria en eltractament de la diabetis gestacionalAmb l’ajuda dels registres clínics informatitzats i elsalgoritmes de suport a la gestió terapèutica metabòli-ca de la diabetis gestacional, s’ha fet una avaluacióretrospectiva que va comparar l’índex de tractament
amb insulina i l’evolució perinatal en dones ambdiabetis gestacional tractades per l’endocrinòleg, ambels mateixos paràmetres formulats pel personald’infermeria especialitzat en diabetis. En l’estudi no esvan observar diferències objectives entre la pràcticad’un o altre professional ni en l’índex de tractament, nien l’evolució perinatal. Aquest resultat contribueix apotenciar el paper actiu del personal especialitzatd’infermeria en el tractament metabòlic de la donaamb diabetis gestacional. (Per a més informació,consulteu l’article derivat d’aquesta recerca, A.García-Patterson et al. "Nurse-based management inpatients with gestational diabetes". Diabetes Care,2003;26:998-1001).
B.1.3.3. Aplicació a l’estudi de reproductibilitat deldiagnòstic de diabetis mellitusEls registres clínics HIB i història clínica electrònica dediabetis i gestació han esdevingut la base d’un estudiadreçat a la reproductibilitat del diagnòstic de DM,segons el criteri de l’OMS (1999) en un seguimentmitjà de 6,2 anys, per a un grup de 696 dones amb eldiagnòstic previ de DG. Una vegada classificada lapacient com a DM després d’una sobrecàrrega oralde glucosa, se li va fer un segon test 3 mesosdesprés (interval mitjà). 44 dones van presentar untest diagnòstic DM en el primer test (12 diagnostica-des per la glucèmia de dejuni, 10 per la hiperglucèmiapostsobrecàrrega, 10 per la combinació de lesdeterminacions basal i postcàrrega i 3 per hiperglucè-mia simptomàtica). El segon test va confirmar eldiagnòstic de DM en el 60% dels casos i en el 56,3%dels casos diagnosticats en absència de simptomato-logia. La reproductibilitat diagnòstica va ser del 33%en el cas d’hiperglucèmia de dejuni, 40% en el casd’anormalitat postcàrrega i del 100% quan es vademostrar anormalitat tant en la determinació basalcom postcàrrega. Quan, en canvi, no es va confirmarel diagnòstic de DM, es va poder constatar l’existèn-cia d’intolerància a la glucosa (de dejuni a postcàrre-ga) en el 75% de les dones diagnosticades amb DMen el primer test, i en el 83,3% dels casos diagnosti-cats per hiperglucèmia postcàrrega. En termesglobals, entre les dones diagnosticades en el primertest, es va documentar tolerància anormal a laglucosa (intolerància a la glucosa de dejuni/postpran-dial o DM) en el segon test en el 83,3% dels inicial-ment diagnosticats per hiperglucèmia de dejuniexclusiva, en el 90% dels diagnosticats postcàrrega, ien el 100% dels diagnosticats per ambdues anormali-tats, basal i postcàrrega. En termes generals, endones asimptomàtiques, el diagnòstic es va poderconfirmar en el 90,6% dels casos. (Per a més informa-ció, es pot consultar l’article original derivat d’aquestarecerca, M. Albareda et al, "Reproducibility of diabetesmellitus diagnosis (WHO 1999 criteria) in women".Acta Diabetol 2004;41:14-17)B.1.3.4 Aplicació a l’estudi evolutiu de la tolerància ala glucosa en la població amb diabetis gestacionalLa població de 696 dones amb antecedent de DG jaesmentada va ser seguida en el temps, tot estudiantla progressió a DM/tolerància anormal a la glucosa i laidentificació de factors predictius. La població d’inte-rès i la població de 70 dones controls van ser avalua-des postpartum i en intervals de 5 anys. El riscacumulatiu de DM va ser del 13,8% i del 42,4% el risc
RECERCA BIOMÈDICA | 153

de tolerància anormal a la glucosa, comparats amb el0% i el 2,8% en el grup control. Els factors de predic-ció investigats (antecedent d’hiperglucèmia esporàdi-ca/intolerància a la glucosa, edat de gestació en eldiagnòstic, IMC previ a l’embaràs) van mostrar uncomportament no lineal. El factor determinant de lamajor fracció atribuïble de risc en el grup de poblaciógeneral va ser l’IMC preembaràs. (Per a informaciómés àmplia sobre aquest apartat, consulteu l’articleoriginal derivat d’aquesta recerca, M Albareda et al.,"Diabetes and Abnormal Glucose Tolerance in womenwith previous gestational diabetes". Diabetes Care2003;26:1199-1205).
B.1.3.5 Aplicació a l’anàlisi del benestar maternofetal:predicció de malformacions congènites en la diabetisgestacionalAmb l’ajuda dels registres clínics HIB i història clínicaelectrònica de la Clínica de Diabetis i Embaràs, s’hainvestigat la freqüència de malformacions congènites(MC) en una cohort de 2.060 nounats de mares ambdiabetis gestacional. Es va analitzar la relació ambpredictors potencials, tot emprant anàlisis de regressiólogística. La freqüència d’MC va ser: 6% per a malfor-macions menors i 3,8% per a malformacions majors(cardíaques, 1,4%; renal/urinàries, 0,8%; esquelèti-ques, 0,7%; hipospàdies, 0m8%; sistema nervióscentral, 0,2%; paladar/llavi leporí, 0,2%; tractedigestiu, 0,1%; d’altres, 0,3%). Els predictors princi-pals van resultar: IMC preembaràs (el més important),l’edat de gestació en el diagnòstic, i el valor glucèmicals 60 minuts en el TTOG. (Per a una informació mésàmplia sobre aquest apartat, consulteu l’article originalderivat d’aquesta recerca, A García-Patterson et al.,"In human gestational diabetes mellitus congenitalmalformations are related to pre-pregnancy bodymass index and to severity of diabetes". Diabetologia2004;47:509-514).
B.2. Programes de telemedicinaB.2.1. Sistema DiabTelHem avaluat el sistema DiabTel (protocols telemàtics,gestió clínica, control metabòlic, qualitat de vida) enun protocol encreuat de 6 mesos de durada amb ungrup de pacients amb DM-1. Entre altres paràmetres,s’hi van determinar: nombre de comunicacionsregistrades, interval d’aquestes, tramesa de missatgesper l’equip assistencial i pels pacients, transmissió devalors glucèmics, ajustaments diaris de les dosisd’insulina, pràctiques d’exercici físic, modificacionsdietètiques. L’avaluació va demostrar que els metgesvan fer un nombre més gran de canvis terapèuticsdurant la fase experimental (DiabTel) que no durant lafase control; es va copsar una tendència a la reduccióde l’HbA1c (al límit de la significació estadística,malgrat la curta durada de l’estudi) i sense augmentde la freqüència de presentació d’hipoglucèmies.
La recerca va poder demostrar l’operació correctadel sistema en la pràctica clínica diària i els beneficisderivats d’una comunicació millor i més freqüententre metge i pacient. A parer dels investigadors, elsistema ofereix possibilitats interessants per explorarmés àmpliament entre els pacients inclosos enprogrames de teràpia intensiva, particularmentembaràs i tractament amb BICI.
(Per a una informació més detallada, consulteu l’articleoriginal derivat d’aquesta recerca, EJ Gómez et al.,"Telemedicine as a tool for intensive management ofdiabetes: the DIABTel experience". Computer Methodsand Programs in Biomedicine 69 (2002):163-177).
B.2.2. Sistema T-IDDMLa validació del sistema ha seguit dues fases. En lafase de verificació, s’ha comprovat el funcionamenttècnic de cada component del prototipus, tant en lespropietats relacionades amb l’ús com amb les especi-ficacions funcionals; va merèixer un interès especial lainterconnexió entre UP i UM, amb l’ajuda d’un procésde simulació, tot comprovant la resposta del programad’intel·ligència artificial davant de diversos escenaris.
La fase de validació clínica es va estendre durant uninterval de 12 mesos, tot mostrant una operaciócorrecta, i el benefici d’una comunicació més eficiententre el pacient i l’equip sanitari, objectivada perl’anàlisi de la transferència, emmagatzematge iprocessament de dades. L’efecte beneficiós sobre elcontrol metabòlic (9% de reducció de l’HbA1cmitjana, de 7,77 a 7,1%) observat no va arribar asuperar el criteri de significació estadística, que sí esva complir per a la reducció de la dosi insulínica diària.
(Per a una informació més detallada, consulteu l’articleoriginal derivat d’aquesta recerca, R Bellazzi et al., "Atelemedicine support for diabetes management : theT-IDDM project". Computer Methods and Programs inBiomedicine 2002;69:147-161)
B.2.3. Sistema M2DML’avaluació s’ha fet mitjançant una mostra de 22pacients durant 2 anys. Els pacients han enviat untotal de 36.801 dades glucèmiques; com aconseqüència, s’han fet 29 canvis de tractament quehan afectat 12 pacients. Com a mitjana, els pacientshan emprat la interfície de la pàgina web durant uns45 minuts per setmana, i el metge ha invertit uns 3minuts setmanals per pacient consultat. Els pacientshan emprat el sistema amb moderació, tot adaptant elnombre i la durada dels accessos a les seves necessi-tats i a la situació metabòlica concreta. L’acceptabilitatdel sistema ha estat positiva per als usuaris en un80% i la manifestació ha estat negativa només en un5% dels casos. La puntuació obtinguda a favor del’ús futur de TM ha estat del 4,7(0,7)/5,0, valoracióque va abastar aspectes organitzatius, econòmics,clínics, qualitat de vida i accessibilitat. El metge vallegir els missatges rebuts en menys de 24 hores en el82% dels casos i en menys de l’interval de 5 dies enel 100% dels casos. Amb tot, el veritable impacteorganitzatiu del sistema no es podrà determinar finsque el sistema no formi part de la rutina clínica.El sistema ha exercit un impacte positiu en el controlmetabòlic, demostrat per la reducció mitjana del’HbA1c d’un punt i mantinguda durant tot l’any queva durar l’experiència clínica. Aquesta afirmació es vaveure reforçada per la correlació constatada entre lavariació de l’hemoglobina glicada i el nombre demissatges (R=-0,645, p=0,04) enviats pels pacients.La millora del control metabòlic no es va associar ambaugment del nombre d’hiperglucèmies registrades.
FUNDACIÓ LA MARATÓ DE TV3

(Per ampliar la informació, consulteu l’article originalderivat d’aquesta recerca, R Bellazzi et al., "Design,methods, and evaluation directions of a Multi-AccessService for the management of diabetes mellituspatients". Diabetes, Technology and Therapeutics2003;5:621-629. També: Angel García Olaya,Arquitectura Multiacceso de Telemedicina para elcuidado compartido para pacientes diabéticos [tesidoctoral]. Universidad Politécnica de Madrid, 2004)
3. Rellevància i possibles implicacions clíniquesdels resultats finals obtingutsEl Projecte OPTIDIAB, desenvolupat en aquestarecerca, esdevé un model original de gestió clínica,dissenyat per a l’optimització de l’assistència delspacients amb diabetis mellitus. La rellevància i aplica-bilitat clínica que té són evidents, i la posada enpràctica en qualsevol institució sanitària ha de contri-buir en gran mesura a millorar el nivell de la qualitatassistencial i el grau de satisfacció del pacient.
La iniciativa responsable del disseny, l’organització i laposada en pràctica del Programa Optidiab compleixamb els postulats de la Declaració de Saint Vincent(OMS, FID, octubre de 1989), pedra angular en l’esta-bliment de programes de desenvolupament de laqualitat assistencial en l’atenció diabetològica, essentles seves fites principals millorar la salut i incrementarla qualitat i la quantitat de vida dels pacients amb DM.Per a l’acompliment d’aquesta missió important, calintensificar l’activitat investigadora adreçada a lanormalització metabòlica i a la prevenció de complica-cions de la malaltia. El Projecte OPTIDIAB empra elsinstruments potents de les noves tecnologies de lainformació per adreçar-los al servei de l’optimitzacióterapèutica de la diabetis mellitus i la reduccióconsegüent de la seva alta morbi-mortalitat.
Aquest projecte de recerca ha assolit, entre altresobjectius, l’organització i l’operació del programa dequalitat assistencial de la diabetis mellitus al serveidels més de 3.000 pacients diabètics atesos regular-ment al Centre de Diabetis de l’Hospital de la SantaCreu i Sant Pau. Igualment, ha ajudat, en la mesurapossible, a la introducció del programa en un nombrealt de centres hospitalaris de l’Estat, i a diversoscentres d’atenció primària, per mitjà de sessionsinformatives, cursos i seminaris específics, i d’unadisponibilitat continuada d’assessorament clínic i desuport informàtic.
El Programa de Qualitat Assistencial (basat en elProjecte Europeu DiabCare Quality Network) abastatot el ventall de procediments bàsics de l’assistènciadiabetològica, resulta fàcil d’usar, i permet la introduc-ció completa i vàlida de les dades en forma estandar-ditzada, seleccionades a partir d’una llista determina-da. Es recomana que les dades siguin recollides ambuna periodicitat mínima anual, i que el mateix equipresponsable de l’assistència faci una anàlisi local delsresultats. A partir d’aquesta anàlisi s’identifiquen elsfactors del procés assistencial susceptibles de correc-ció per modificar els resultats no desitjats. Un procedi-ment informàtic assequible, senzill, anonimitza lesdades i les agrega i, eventualment, també en fa l’anàli-si comparada amb la xarxa de centres col·laboradors.
La repetició del procediment i la retroalimentaciópermet la continuïtat del progrés de la qualitatassistencial.
L’ús del Programa Optidiab ha representat fins arauna eina molt beneficiosa per a la gestió clínica alnostre equip i per als nostres pacients. Ha contribuït apromoure i esperonar el col·lectiu d’infermeria expertaen diabetis, fins al punt de ser capaç d’objectivar queel seu impacte professional en el tractament metabòlicde la dona amb diabetis gestacional és tan excel·lentcom la del mateix endocrinòleg, i, de retruc, en larepercussió associada al benestar maternofetal.
La implantació del Projecte OPTIDIAB ha aconseguitintegrar els procediments innovadors de la telemedici-na amb els de l’atenció tradicional de la diabetis. Haincorporat la telemonitorització, la teleconsulta i lateleassistència amb la gestió del coneixement i l’auto-matització d’algoritmes terapèutics, per bé quemodulats per la supervisió professional.
OPTIDIAB ha fet factible l’operació satisfactòria d’unaarquitectura de multiaccés de telemedicina capaçd’aplegar tecnologies d’informatització ubiqua, CTI(telefonia i ordinadors) i Internet, la qual cosa facilitaque els actors implicats puguin accedir a una històriaelectrònica comuna i eines de comunicació icol·laboració determinants en l’assistència comparti-da. Aquest sistema de TM forneix als pacients méspossibilitats d’accés als recursos sanitaris; especial-ment, al consell i la supervisió del diabetòleg.
La reducció de prop d’un punt de l’hemoglobinaglicada esdevé un benefici de repercussions extraordi-nàries en el curs evolutiu de les complicacions, iaquest benefici no sembla que representi una càrregalaboral excessiva per als professionals.
En definitiva, OPTIDIAB s’ha mostrat com un sistemaeficaç tant per millorar el control metabòlic de ladiabetis com la qualitat de l’assistència clínica oferta aaquest col·lectiu tan important de malalts crònics.L’extensió d’aquest benefici sanitari a la generalitat depacients diabètics ha de representar un objectiu percomplir els anys vinents.
4. PublicacionsBellazzi, R. Larissa, C. Montani, S. Riva, A. Stefanelli, M.
D’Annunzio, G. Lorini, R. Gomez, E.J. Hernando, E. Brugués, E.
Cermeño, J. Corcoy, R. de Leiva, A. Cobelli, C. Nucci, G. del
Prato, S. Maran, A. Kilkki, E. Tuominen, J. A telemedicine support
for diabetes management: the T-IDDM Project. Computer
Methods and Programs in Biomedicina 69 (2002) 147-161.
Gómez, E. J, Hernando, M.E., Garcia, A., del Pozo, F., Corcoy,
R., Brugués, E., Cermeño J., y de Leiva A. Telemedicine as a tool
for intensive management of diabetes: the DIABTel experience.
Comp,. Meth. and Prog. in Biomedicine, 69(2002) 163-177.
Riccardo Bellazzi, Marco Arcelloni, Giuliana Bensa, Hannes
Blankenfeld, Eulàlia Brugués, Ewart Carson, Claudio Cobelli,
Derek Cramp, Giuseppe d’Annunzio, Pasquale De Cata, Alberto
de Leiva, Tibor Deutsch, Pietro Fratino, Carmine Gazzaruso,
Angel Garcìa, Tamás Gergely, Enrique Gómez, Fiona Harvey,
Pietro Ferrari, Elena Hernando, Maged Kamel Boulos, Cristiana
Larizza, Hans Ludekke, Alberto Maran, Gianluca Nucci, Cristina
Pennati, Stefano Ramat, Abdul Roudsari, Mercedes Rigla, Mario
RECERCA BIOMÈDICA | 155

Stefanelli. Design, methods and evaluation directions of a multi-
access service for the management of diabetes mellitus patients.
Diabetes, Technology and Therapeutics, Vol 5, Number 4, 2003
p621-629.
Garcia-Patterson, A. Martin, E. Ubeda, J. María, M.A.
Adelantado, J.M. Ginovart, G. de Leiva, A. Corcoy, R. Nurse-
based Management in Patients With Gestational Diabetes.
Diabetes Care, Vol 26, Num 4, April 2003. p 998-1001.
Albareda, M. de Leiva A. Ciorcoy, R. Reproductibility of
diabetes mellitus diagnosis (WHO 1999 criteria) in women. Acta
Diabetol (2004) 41:14-17.
Albareda, M. Caballero, A. Badell, G. Piquer, S. Ortiz, A. de
Leiva, A. Corcoy, R. Diabetes and Abnormal Glucose Tolerante in
Women With Previous Gestational Diabetes. Diabetes Care, vol
26, Num 4, Abril 2003, p1199-1205.
García-Patterson, A, Erdozain, L. Ginovart, G. Adelantado, J.M.
Cubero, J.M. Gallo, G. de Leiva, A. Corcoy, R. In human gestatio-
nal diabetes mellitus congenital malformations are related to pre-
pregnancy body mass index and to severity of diabetes.
Diabetologia (2004) 47:509-514.
Capítol de llibre de Leiva, A. Corcoy, R. Brugués, E. Quality of
care for the woman with diabetes in pregnancy. En: M Hoh, L
Jovanovic, GC DiRenzo, A deLeiva, O Langer (editors), Textbook
of Diabetes and Pregnancy 2003, MD Martin Dunitz (Taylor & Francis
Group plc, London and New York); Chapter 44, p 539-553.
Resums publicatsBrugués E, Cermeño J, y Mercedes Rigla en representación de
los Consorcios EuroDiabCare Q-Net y DiabCare España.
Actividades en España de los Consorcios Europeos de
Investigación DiabCare Q-Net y DiabCard (Resultados del período
1998-2000). Av. Diabetol 2000; 16 (Supl. 1): 58-A.
Corcoy R, Cermeño J, Brugués E, Hernando E, Gómez E, de
Leiva A y Consorcio T-IDDM. Prueba piloto de la utilización del sistema
T-IDDM en la práctica clínica. Av. Diabetol 2000; 16 (Supl. 1): 58-A.
J. Cermeño, R. Corcoy, E. Brugués, E. Hernando, E. Gómez,
A. de Leiva. Utilidad Clínica del Sistema DIABtel en el seguimiento
ambulatorio de pacientes tipo 1. Av. Diabetol 2000; 16 (Supl. 1): 59-A.
Cermeño J, Corcoy R, Brugués E, de Leiva A. Calidad de la
información usada en el sistema Diabcare. Endocrinología 2000;
47 (Supl. 1): 32-A.
Brugués E, Corcoy R, Cermeño J, Rigla M, Hernando E,
Gómez E, de Leiva A. Telematic management of Type 1 diabetes:
The T-IDDM Project. Diabetologia 2000; 43 (Suppl. 1): 906-A.
Bellazzi R, Brugués E, Cobelli C, de Leiva A, del Prato S,
Gómez E, Hernando E, Stefanelli M, Tuominen J. T-IDDM.
Reports on the results of the ELI funded Project.
Diabetes, Nutrition and Metabolism 2000; 13: 244-A.
Bellazi R, Brugués E, de Leiva A, del Prato S, Gómez E,
Hernando E, Roudsari A, Stefanelli M and the M2 DM Project
Consortium. Multi-access services for DM management: The M2
DM Project. Diabetes, Nutrition and Metabolism 2000; 13. 249-A.
Gómez E, Hernando E, Corcoy R, Cermeño J, Brugués E, de
Leiva A, del Pozo F. Evaluation of the Diabtel Telemedicine System.
Diabetes, Nutrition and Metabolism 2000; 13: 250-A.
J. Cermeño, R. Corcoy, E. Brugués, E. Hernando, E. Gómez,
A. de Leiva. Feasibility of Diabtel Use in the follow up of Type 1
Diabetic Patients. Diabetes Research and Clinical Practice 2000;
50: S210-A (P811).
Brugués E, Corcoy R, Cermeño J, Rigla M, de Leiva A.
Telematic management of Type 1 DM: The T-IDDM Project.
Diabetes Research and Clinical Practice 2000; 50: S57-A (P 286).
Cermeño J, Corcoy R, Brugués E, de Leiva A.
Quality of the Information used by the Diabcare System.
Diabetes Research and Clinical Practice 2000; 50: S57-A (P 288).
E. Brugués, J. Cermeño, R. Corcoy, E. Hernando, E. Gómez-
Aguilera, A. García, and A. de Leiva. Telemedicine may assist
diabetic subjects to improve metabolic outcome and promote
patient’s empowerment. Diabetologia 2001; 44 (Suppl. 1): 224:A-59.
JM. Cubero, M. Hernandez, E. Brugués, J. Cermeño, R.
Corcoy and A. de Leiva. Metabolic outcome achieved by intensive
management in european diabetes centers are far from derirable
targets. Diabetologia 2001;44 (Suppl. 1):59:A17.
G. Gallo, J. Cermeño, E. Brugués, M. Rigla, R.Corcoy and A.
de Leiva. The burden of type 2 diabetes compared to the burden of
type 1 diabetes in patients undergoing intensive and specialized care.
Diabetologia 2001; 44 (Suppl. 1): 56:A-16.
E. Brugués, J. Cermeño, R. Corcoy, E. Hernando, E. Gómez-
Aguilera, A. García, y A. de Leiva. La telemedicina pot facilitar la
millora dels resultats metabòlics i la autonomia dels pacients amb
diabetis mellitus. VI Congrés i I Curs PreCongrés de l’Associació
Catalana de Diabetes 2001; SP22:115.
JM. Cubero, M. Hernández, E. Brugués, J. Cermeño, R.
Corcoy, y A. de Leiva. El control metabòlic aconseguit amb tracta-
ment intensiu als centres europeus de diabetis és lluny dels
objectius desitjables. VI Congrés i I Curs PreCongrés de
l’Associació Catalana de Diabetes 2001; SP9:107.
Rigla, M. Brugues, E. Gómez, E. Hernando, E. García, A.
Perdices, J. Torralba, V. de Leiva, A. Supervised Autonomy with
Diabetes Self-Management by using Telemedicine Allows Better
Glycemic Control in Patients with Diabetes. Endocrine Practice:
vol 9 (Suppl 1), Marh/April 2003: 27:A70.
Rigla, M. Brugues, E. Gómez, E. Hernando, E. García, A.
Perdices, J. Torralba, V. de Leiva, A. The use of multi-access
service for the diabetes management allows better glycaemic
control n insulin treated diabetic patients. Diabetología, Vol 46.
Supp 2. August 2003: A426:1246.
Blankkenfeld, H. Arcelloni, M. Bellazzi, R. Brugues, E. Cobelli, C.
d’annunzio, G. de Leiva, A. Fratino, P. Harvey, F. Hernando, H. Ferrari,
P. Maran, A. Pennati, C. Schumm-Draeger, P.M. Stefanelli, M. Clinical
evaluation of a multi-access service for the management of
diabetic patients with insulin theraphy (M2DM Project).
Diabetología, Vol 46. Supp 2. August 2003: A427:1247.
FUNDACIÓ LA MARATÓ DE TV3

RECERCA BIOMÈDICA | 157
MEMÒRIA FINAL
1. Resum del projecte originalLa diabetis tipus 2 està assolint trets d’epidèmia enles societats occidentals. Les evidències experimen-tals obtingudes els darrers anys permeten afirmar queels àcids grassos lliures (FFA), actuant probablementcom a molècules senyalitzadores, tenen un impacteprofund en el funcionament de la cèl·lula b. A mésd’aquest paper fisiològic, avui dia sabem que l’exposi-ció crònica de la cèl·lula b a nivells alts d’FFA potproduir una hiperinsulinèmia basal, i també unaproducció deficient d’insulina com a resposta a laglucosa, fenotip que caracteritza la diabetis tipus 2.
Actualment també hi ha evidències que una acumula-ció Acil-CoAs (reflectida per un augment de triglicèridsen la cèl·lula muscular) pot ser un factor important enla gènesi de la resistència a la insulina, possiblementinterferint en la captura i el metabolisme de la glucosainduïts per insulina. L’escenari actual en estudi és queen etapes primerenques, tant de l’obesitat com de ladiabetis tipus 2, l’acumulació de greixos en múscul,en cèl·lula b o en els dipòsits de greixos, pot ser unmecanisme responsable de la resistència a la insulina ide la hiperinsulinèmia compensatòria. En aquestes
condicions, particularment quan el teixit adipósesdevé refractari a l’efecte antilipolític de la insulina, laproducció excessiva de VLDL en el fetge facilita eldesenvolupament de la dislipidèmia i augmenta elgrau de resistència a insulina. En individus predispo-sats genèticament, aquestes condicions podenpredisposar a lipotoxicitat en la cèl·lula b, amb elsubsegüent desenvolupament obert de la diabetis.
La nostra hipòtesi és que el control dels nivells d’FFAen etapes primerenques de la diabetis tipus 2permetria reduir la hiperinsulinèmia i la resistència ainsulina. L’objectiu global d’aquest projecte és estudiarels mecanismes de regulació transcripcional ial·lostèric dels gens clau en el control de l’oxidaciómitocondrial dels FFA, com són els isotipus L i M de laCPT I (carnitina palmitoiltransferasa I), en la recerca dedianes terapèutiques que millorin els tractamentsantidiabètics disponibles actualment.
Recentment s’han aïllat els promotors dels gensimplicats en la regulació de la b-oxidació dels àcidsgrassos en diversos teixits, L-CPTI i M-CPTI, alhoraque s’han desenvolupat sistemes d’expressió quepermeten l’anàlisi de les característiques cinètiques deCPTI recombinants. Tenint en compte el paper dels
Títol del projecte
La carnitina palmitoiltransferasa I (CPTI) com a diana farmacològica a la diabetIs tipus 2
Investigadors principalsPedro F. Marrero González
InstitucióFacultat de Farmàcia. UB
69.786 €

FFA en els diversos estadis de la malaltia, és plausiblede pensar que l’estudi dels mecanismes molecularsque controlen la degradació dels àcids grassos puguifacilitar l’elaboració d’estratègies farmacològiquesdestinades a pal·liar el desenvolupament de la malaltia
Els objectius concrets són els següents1. Determinar l’efecte de les tiazolidinediones (TZD) enla b-oxidació dels àcids grassos i en l’expressió delgen M-CPTI en teixits de rates normals i diabètiques.Estudiar la influència de PPARg sobre els efectesproduïts per PPARa en el promotor de l’M-CPTI humà.
2. Estudiar els mecanismes moleculars que dirigeixenl’expressió del gen L-CPTI al fetge i a la cèl·lula b.
3. Determinar, per mitjà de l’anàlisi de proteïnesrecombinants, la relació que hi ha entre l’estructuraprimària de les CPTI i els seus paràmetres cinètics.Especialment la sensibilitat dels enzims al malonil-CoA.
El pla de treball per assolir els objectius és elsegüent1. Determinar l’efecte de les tiazolidinediones (TZD):
1.1 En l’expressió del gen M-CPTI en cèl·lules i teixitsde rates normals i diabètiques.
1.2 En la b-oxidació dels àcids grassos en cèl·lules.
2. Estudiar la influència de PPARg sobre els efectesproduïts per PPARa en el promotor del gen M-CPTIhumà.
3. Estudiar els mecanismes moleculars que controlenl’expressió del gen L-CPTI en fetge i cèl·lula b.
4. Determinar la relació entre l’estructura primària deles CPTI i els seus paràmetres cinètics. Especialmentla sensibilitat dels enzims al malonil-CoA.
4.1 Aïllar el cDNA de l’M-CPTI de porc.
4.2 Expressar L-CPTI i M-CPTI de porc i humà enPichia pastoris.
4.3 Estudis de l’estructura primària de la CPTI.
2. ResultatsObjectiu 1Determinar l’efecte de les tiazolidinediones (TZD) en lab-oxidació dels àcids grassos i en l’expressió del genM-CPTI en teixits de rates normals i diabètiques.Estudiar la influència de PPARg sobre els efectesproduïts per PPARa en el promotor del M-CPTI humà.
En aquest objectiu hem demostrat que:
Els receptors nuclears PPARa i PPARg regulen demanera diferencial l’expressió musculoespecífica delgen M-CPTI humà.
PPARa produeix un efecte sinèrgic amb els factorsmiogènics: Mef2 i MyoD. D’aquesta manera, l’expres-sió d’aquest gen serà màxima en múscul, que utilitzacom a substrat metabòlic els àcids grassos. Amb tot,
PPARg, que també activa l’expressió del gen, no téun efecte sinèrgic amb els factors miogènics; amb laqual cosa la seva activació, dependent de lligands,pot bloquejar l’activitat transcripcional del gen enmúscul mediada per PPARa i, per tant, la b-oxidaciómitocondrial. Aquest fenomen molecular, prèviamentno descrit, ha estat comprovat bioquímicament encultius primaris de cèl·lules musculars humanes i enteixits de ratolí (dades no publicades).
Objectiu 2Estudiar els mecanismes moleculars que dirigeixenl’expressió del gen L-CPTI en el fetge i la cèl·lula b
En aquest objectiu hem demostrat que:
El gen L-CPT és diana del receptor d’hormonatiroïdal.
L’isotip de rata del gen L-CPTI és un gen dianad’hormona tiroïdal. En cèl·lules HepG2, la transcripciódel gen està reprimida en presència del receptor TRa,i s’activa en presència de l’hormona T3. El receptorTR s’uneix, en forma d’homodímers o heterodímersamb RXR, a una seqüència present en la posició —2935 del promotor de rata i que està conservada enel promotor del gen humà. L’efecte de la inhibició perTR pot ser revertit per l’ús d’inhibidors d’histonesdesacetilases (dades no mostrades, i vegeu publica-cions: Barrero et al., 2000).
El gen CPTII és diana de PPAR
El gen CPTII té, en el seu promotor pròxim a l’inici detranscripció, un element de resposta a PPAR. Aquestelement (PPRE) és característic, perquè el segon delshalf sites del DR-1 està molt poc conservat (vegeupublicacions: Barrero et al., 2003). Aquest resultat i elsobtinguts abans pel nostre grup (Mascaró et al., 1998;Rodríguez et al., 1994) i per altres autors (Gulik et al.,1994; Schoonjans et al., 1995) permeten establir unmodel de regulació de l’oxidació dels àcids grassosmediat per PPAR, tal com s’indica en la figura 1. Enaquest model, destaca que el mecanisme pel qual elsLCFA (àcids grassos de cadena llarga) activen el genL-CPTI en cèl·lules Min-6 sembla independentd’agonistes de PPAR, malgrat que hem pogutcaracteritzar elements d’unió (PPRE) per a aquestreceptor nuclear (vegeu més endavant).
FUNDACIÓ LA MARATÓ DE TV3

Figura 1. Activació dels gens responsables de l’oxidació dels
àcids grassos de cadena llarga i cetogènesi per PPARa.
L’acil-CoA sintetasa ((Schoonjans et al., 1995), CPTIbo M-CPTI (Mascaró et al., 1998), CPTII (Barrero et al.,2003), Medium chain acilCoA deshidrogenasa (Guliket al., 1994), i HMG-CoA sintasa mitocondrial(Rodríguez et al., 1994) són gens diana de PPAR. L-CPTI (CPTIa) és un gen diana d’àcids grassos permitjà d’un mecanisme encara no determinat.
En cèl·lules Min-6, el gen L-CPTI de rata és dianad’àcids grassos però no d’agonistes de PPARa.El tractament de cèl·lules Min-6 amb àcids grassos decadena llarga, com el linoleic o l’oleic, indueixenl’expressió del gen endogen L-CPTI. Aquesta respos-ta es bloqueja en presència d’actinomicina D. Enaquestes cèl·lules s’expressa PPARa Malgrat això,diversos agonistes de PPARa no indueixen l’expressiódel gen endogen (dades no mostrades). Aquestresultat suggereix que el gen L-CPTI és diana d’àcidsgrassos per mitjà d’un mecanisme transcripcionalindependent de PPARa. Per estudiar aquest mecanis-me vam aïllar el promotor de l'L-CPTI de rata (vegeupublicacions Barrero et al., 2000) i vam caracteritzar-loper mitjà d’assajos de gen reporter amb el control dediverses variants del promotor objecte d’estudi.Tanmateix, cap de les construccions de gen reporterassajades respon al tractament amb àcid linoleic(dades no mostrades). Aquest conjunt de resultatsens permeten plantejar que: 1) o bé no hem clonatprou regió promotora, tot deixant fora de l’anàlisi elselements responsables del control per àcids grassos;2) o bé la regulació transcripcional del gen és unefecte complex que necessita el DNA cromatinitzat(vegeu més endavant).
El gen L-CPTI humà és diana d’inhibidors d’histonadesacetilases.L’àcid gras de cadena curta n-butirat i el TSA (uninhibidor específic d’histona desacetilases) indueixenl’expressió del gen en termes d’mRNA en cèl·lulesCaCo-2 (dades no mostrades). L’estudi del promotorpròxim del gen humà i de rata mostra que ambdóssón sensibles al TSA. Aquest efecte es constata tanten cèl·lules CaCo-2 com en cèl·lules HepG2. L’anàliside la seqüència promotora mostra l’existència de llocsd’unió per als factors de transcripció de la família deproteïnes Sp1. Per experiments de retardament de lamobilitat electroforètica i de mutagènesi dirigida, hemcomprovat que la unió de proteïnes de la família Sp1és necessària per a l’activitat transcripcional del gen,alhora que és la responsable de mediar en la respostaals inhibidors d’histona desacetilases. La resposta aaquests inhibidors es correlaciona amb l’afinitat ambquè les proteïnes de la família Sp1 s’uneixen aaquests elements (dades no mostrades).
Estudi in situ del promotor L-CPTI humà.Per mitjà de l’ús de la tècnica d’immunoprecipitacióde cromatina (ChIP), hem estudiat les proteïnes ques’uneixen in vivo al promotor humà del gen endogend’L-CPTI. Aquests experiments (figura 2) mostren queanticossos contra TRb o PPARa són capaçosd’immunoprecipitar la cromatina present a 4.000 pbde l’origen de transcripció (zona A), mentre queanticossos elicitats contra Sp1 o NFY són capaços
d’immunoprecipitar el promotor pròxim del gen (zonaB). Aquests experiments de ChIP concorden amb lapresència dels elements que actuen en cis a lesregions corresponents del promotor i que han estatcaracteritzats en aquest estudi. Cal destacar quel’element de resposta a PPAR, el PPRE, ha estatcaracteritzat per tècniques in vitro, però aquestelement no és funcional en els assajos de transfecciótransitòria. En la figura 2 també s’observa que elsanticossos contra Sp1 i NFY són capaços d’immuno-precipitar, a més, el promotor llunyà del gen (zona B).Aquests resultats suggereixen la formació d’unaestructura d’ordre superior, un loop, en la cromatinadel promotor i podrien explicar en part la manca deresposta a PPARa en experiments clàssics del genreporter
Figura 2. ChIP del promotor LCPTI humà.
En la part superior s’indica d’una manera esquemàticael promotor del gen humà; els elements cis caracterit-zats en aquest treball (TRE, PPRR, Sp1 i NFY) s’indi-quen englobats en caixes. NFY ha estat caracteritzatin silico. Les fletxes indiquen la posició dels primersemprats per amplificar el DNA després de la immuno-precipitació de la cromatina, amb la qual cosa s’ampli-fiquen fragments denominats Zona A o Zona B. En lapart inferior es mostra el resultat de l’amplificació perPCR d’ambdues zones. Cada carril correspon a l’anti-còs específic emprat o anticossos control (-).
Objectiu 3Determinar, per mitjà de l’anàlisi de proteïnes recombi-nants, la relació que hi ha entre l’estructura primàriade les CPTI i els seus paràmetres cinètics.Especialment la sensibilitat dels enzims al malonil-CoA.
En aquest objectiu hem demostrat que:L-CPTI de porc és una quimera natural de lesformes L- i M-L-CPTI de porc recombinant mostra una Km per a lacarnitina característica dels isotipus de fetge (L-), peròuna inhibició per al malonil-CoA (IC50) característicadels isotipus musculars (M-). O sigui que és unaquimera natural i es converteix en una eina útil, talcom havíem proposat, per a la producció de quimeresentre formes hepàtiques que siguin catalíticamentactives i, per tant, que permetin determinar zones dela proteïna responsables de la sensibilitat al malonil-CoA (vegeu figura 3 i publicacions: Nicot et al., 2001).
M-CPTI de porc presenta característiques cinètiquesde la forma L-M-CPTI de porc recombinant mostra una sensibilitat al
RECERCA BIOMÈDICA | 159

malonil-CoA (IC50) i una Km per a la carnitina,característica dels isotips de fetge (L-) (vegeu figura 3).O sigui que és una eina útil per a la producció dequimeres entre formes musculars que amb un alt graud’identitat siguin catalíticament actives i, per tant,permetin determinar zones de la proteïna responsa-bles de la sensibilitat al malonil-CoA i de la Km per acarnitina.
Figura 3. Paràmetres cinètics de diversos isotipus de CPTI de
diverses espècies.
Les proteïnes recombinants van ser expressades enPichia pastoris, i se’n van determinar els paràmetrescinètics. La Km per al palmitoil-CoA va ser semblantper a tots els enzims assajats. (A) sensibilitat almalonil-CoA determinada com IC50. (B) Km per alsubstrat carnitina.
La interacció de l’extrem amino i carboxil d’L-CPTIparticipa en la seva sensibilitat al malonil-CoAAquest és un resultat clar del nostre estudi. L-CPTI deporc (L-P) té una IC50 de 141 nM, mentre que unaquimera d’L-CPTI que conté l’extrem amino de porc iel carboxil de rata (L-P/R) té una IC50 de 2 mM. Ladeleció dels primers 18 aminoàcids en ambduesproteïnes genera enzims poc sensibles al malonil-CoA,amb una IC50 semblant, al voltant de 150 mM. Comque els 18 aminoàcids de l’extrem amino d’L-P i L-P/Rsón idèntics, i el grau de sensibilitat al malonil-CoA vedeterminat pel domini carboxílic, aquests resultatsimpliquen una interacció efectiva entre ambdósdominis que determina l’afinitat del malonil-CoA delenzim (vegeu publicacions: Nicot et al., 2002).
3. Rellevància i possibles implicacions clíniquesdels resultats finals obtingutsAquest era una projecte de ciència bàsica, per la qualcosa no n’esperàvem el desenvolupament de molècu-les per al tractament de la diabetis tipus 2. Amb tot,creiem que hem fet dues aportacions rellevants:
1. Hem introduït el concepte d’especificitat depromotor per poder explicar la paradoxa de l’activacióper receptors nuclears del tipus PPAR. Aquestaparadoxa consisteix en el fet que tant els gens quecontrolen la lipogènesi, com els que controlen l’oxida-ció dels àcids grassos, contenen elements de respos-
ta, els PPRE, als quals es poden unir, tant PPARgcom PPARa. Aquesta paradoxa es pot explicar enpart per dos fenòmens: el patró d’expressió teixitespecífic dels diversos PPAR, i l’existència de lligandsespecífics en determinats teixits per als diversosPPAR. Tanmateix, en circumstàncies patològiques,com la diabetis tipus 2, s’ha reportat l’expressió dePPARg en múscul on és abundant PPARa. Nosaltrespensem que els resultats obtinguts amb aquestprojecte afegeixen una tercera explicació a aquestaparadoxa, la ja esmentada especificitat del promotor.Així, el promotor de l’M-CPTI humà seria un paradig-ma de com els diversos receptors poden exercirefectes contraposats d’acord amb les sinergies queun dels isotipus (PPARa) és capaç d’establir ambfactors miogènics i que l’altre isotipus (PPARg) seriacapaç de bloquejar. Aquest model seria capaç, a més,d’explicar els efectes de les tiazolidinediones, com ainhibidors de l’oxidació d’àcids grassos de maneraespecífica en múscul.
2. Hem demostrat que la sensibilitat del malonil-CoAd’L-CPTI és dependent de la interacció del seuextrem amino i carboxil. Aquesta és una observacióimportant per al futur desenvolupament d’estratègiesque permetin determinar amb exactitud la unió del’inhibidor al·lostèric.
4. PublicacionsMaría José Barrero, Nuria Camarero, Pedro F. Marrero and
Diego Haro.Control of human carnitine palmitoyltransferase II gene
transcription by peroxisome proliferator-activated receptor through
a partially conserved peroxisome proliferator-responsive element.
Biochem J. A (369, 721-729, 2003)
Carine Nicot, Joana Relat, Gebre Woldegiorgis, Diego Haro
and Pedro F. Marrero. Pig liver carnitine palmitoyltransferase.
Chimera studies show that both the N- and C-terminal regions of
the enzyme are important for the unusual high malonyl-CoA
sensitivity. J Biol Chem. A (277, 10044-10049, 2002)
Carine Nicot, Fausto G. Hegardt, Gebre Woldegiorgis, Diego
Haro and Pedro F. Marrero. Pig Liver Carnitine Palmitoyltransferase
I, with Low Km For Carnitine And High Sensitivity to Malonyl-CoA
Inhibition, is a Natural Chimera of Rat Liver and Muscle Enzymes.
Biochemistry. A (40, 2260-2266, 2001)
María J. Barrero, Pedro F. Marrero and Diego Haro.
Regulation of the Rat Liver Carnitine Palmitoyltransferase I gene
transcription by Thyroid Hormone Biochem. Biophys. Res. Comm.
A (279, 81-88, 2000)
FUNDACIÓ LA MARATÓ DE TV3

RECERCA BIOMÈDICA | 161
MEMÒRIA FINAL
1. Resum del projecte originalL’objectiu principal del projecte consistia a investigarels canvis d’expressió del VEGF i Angiopoetina-2 enl’angiogènesi ocular induïda per la hiperglucèmia i lahipòxia. En la retinopatia diabètica s’han descrit canvisvasculars consistents en l’apoptosi de cèl·lulesendotelials i periendotelials, per una banda, i en laproliferació i l’augment de la permeabilitat de l’endote-li, per una altra. S’ha observat, a més, que la hiperglu-cèmia i la hipòxia estan directament implicades en lapatogènia d’aquests fenòmens i que el VEGF és unmediador responsable de l’angiogènesi i de l’augmentde permeabilitat vascular.
Recentment s’ha vist la implicació de les angiopoeti-nes en els fenòmens d’apoptosi/supervivència de lescèl·lules de la paret vascular. Partint d’aquest fet i deles observacions del nostre laboratori sobre la funciódel VEGF i l’Angiopoetina-2 en els fenòmens d’angio-gènesi i regressió vascular, com també de la caracte-rització que hem realitzat d’una nova formad’Angiopoetina-2, l’Ang-2B, ens proposem investigarl’expressió de VEGF/angiopoetines durant l’angiogè-nesi ocular de l’embrió de pollastre en condicions
d’hiperglucèmia i d’hipòxia i establir la correlació ambels canvis vasculars observats. Arran de les limitacionsdel tractament actual de la retinopatia diabètica il’impacte social de la malaltia (la diabetis afecta un 5%de la població i causa ceguesa en un 5% delsdiabètics), considerem que les nostres investigacionspoden contribuir a oferir una alternativa de tractamentfarmacològic basada en el manteniment d’una relacióadequada VEGF/angiopoetines.
2. Resultats1. Caracterització de les diverses isoformes del’Angiopoetina-2La tasca inicial del projecte consistia a completar lacaracterització de les diverses isoformesd’Angiopoetina-2. La primera isoforma havia estatcaracteritzada per primera vegada al nostre laboratori.Per caracteritzar totes les possibles isoformes hemseqüenciat el gen complet de l’Ang-2 de pollastre.Aquest gen té una extensió de 46 kb i consisteix en 9exons separats per 8 introns. L’empalmament(splicing) alternatiu del gen dóna lloc a tres transcritsdiferents que hem anomenat Ang-2A, Ang-2B i Ang-2C. Hem demostrat que les mateixes isoformes delpollastre s’expressen també en humans. Les isofor-mes codifiquen proteïnes amb un domini fibrinogen
Títol del projecte
Expressió de VEGF i Angiopoetina-2 en un model de retinopatia diabètica
Investigadors principalsCristóbal Mezquita Pla
InstitucióFacultat de Medicina. UB
Membres de l’equipJoaquim Bosch, Agustín García, Joaquim Mateo, Jovita Mezquita, Montserrat Pau, Pilar Carmona i Betlem Mezquita.
115.827 €

like comú a l’extrem C-terminal de la proteïna ques’uneix al receptor Tie-2 i dominis N-terminalsdiferents. Les isoformes A i B s’expressen en teixitsangiogènics, mentre que la seva expressió no esdetecta en teixits adults vascularment quiescents.
2. Secreció de les diverses isoformesd’Angiopoetina-2La manca de pèptid senyal a la isoforma Ang-2Bplanteja el dubte de si aquesta isoforma pot sersegregada per la cèl·lula. Per resoldre aquesta qüestióhem construït plasmidis que contenien les seqüènciesde les diverses isoformes de l’Angiopoetina-2 fusiona-des a l’epítop Flag. Amb aquests plasmidis hemtransfectat cèl·lules CHO. Quaranta-vuit hores desprésde la transfecció s’han recollit, d’una banda, els lisatscel·lulars i, d’altra banda, el medi en el qual s’havienincubat les cèl·lules. S’han detectat per Western blotles diverses isoformes de l’Angiopoetina-2 utilitzant unanticòs antiFlag. S’ha observat que nomésl’Angiopoetina-2A i l’Angiopoetina-2C es troben en elsobrenedant de les cèl·lules transfectades. Per tant,aquestes dues isoformes són les úniques que sónsecretades per les cèl·lules.
3. Expressió del VEGF, l’Angiopoetina-2 i altresgens durant l’angiogènesi ocular fisiològica del’embrió de pollastreEn intentar caracteritzar l’expressió de VEGF iAngiopoetina-2 i altres gens durant l’angiogènesiocular fisiològica de l’embrió de pollastre, ens vamadonar de la dificultat d’interpretar els resultats acausa de l’excessiva heterogeneïtat de la mostrabiològica. Igualment, els experiments in vivo perestudiar els efectes de la hipòxia i la hiperglucèmiaresultaven inadequats en afectar-se simultàniamenttots els teixits oculars. En tractar de resoldre aquestsproblemes, ens vam adonar de les diferènciesessencials entre l’angiogènesi ocular dels mamífers i lade les aus. Mentre que la retina dels mamífers ésaltament vascular, la part interna de la retina de lesaus és avascular. El subministrament sanguini a laretina avascular de les aus es realitza mitjançant unòrgan vascular anomenat pecten oculi. La simplicitatd’aquest òrgan, constituït principalment per endoteli,cèl·lules periendotelials i macròfags, ens oferia unaoportunitat única d’investigar l’angiogènesi ocular enun nou model que no havia estat prèviament explorat.Per aquest motiu vam reorientar els objectiusproposats cap a la caracterització de l’expressiógènica de molècules angiogèniques i antiangiogèni-ques del pecten oculi.
A més a més de proposar la utilització d’aquest noumodel per investigar l’angiogènesi in vivo, hemcultivat el pecten in vitro en condicions quepermeten obtenir els RNA mitjancers intactesdesprés de 16 h d’incubació.
4. Expressió de molècules angiogèniques i antian-giogèniques durant el desenvolupament del pectenoculi in vivoLes isoformes de VEGF-A 166, 146 i 122 s’expressenen el pecten d’embrions E10, E12, E16 i E20. Laquantitat expressada no varia de manera important.La quantitat augmenta lleument fins al dia E16.
El receptor VEGF-R1 no es gairebé visible per RT-PCRen el pecten d’embrions E10 i augmenta progressiva-ment en embrions E12, E16 i E20.
El receptor VEGF-R2 té una expressió molt baixa en elpecten d’embrions E10 i E12 i augmenta en embrionsE16 i E20.
L’expressió de les isoformes de l’Angiopoetina-2augmenta des del dia E10 a E16 i persisteix elevadael dia E20. S’observa un predomini de la isoformaAng-2C.
L’expressió de l’Angiopoetina-1 augmenta des del diaE10 al E16 i és particularment elevada el dia E16.
L’expressió de la N-cadherina, una proteïna endotelialsobreexpressada en tumors en fase invasiva,augmenta des del dia E10 al E12 i decau ulteriorment.
L’expressió del factor PEDF, un factor inhibidor del’angiogènesi ocular segregat per l’epiteli pigmentari,s’expressa dèbilment els dies 10 i 12 i augmentanotablement els dies E16 i E20.
5. Expressió de molècules angiogèniques i antian-giogèniques en cultius de pecten oculiHem elegit el pecten oculi d’embrions E15 percultivar-lo in vitro perquè expressa en abundància lesmolècules següents angiogèniques i antiangiogèni-ques: VEGF-A (isoformes 166, 146 i 122), VEGF-R1(Flt-1), VEGF-R2 (KDR), Ang-1, Ang-2 (A i C) i factorinhibidor de l’angiogènesi PEDF. Els pecten envoltatsper humor vitri van ser incubats, en una atmosfera deCO2 5%, en medi EBM-2 (Clonetics) complementatper sèrum de pollastre al 10% i gentamicina (0.05mg/ml). Augmentant el temps d’incubació fins a 16hores s’observen canvis en l’expressió de les diversesmolècules angiogèniques i antiangiogèniques.L’expressió de VEGF i VEGF-R1 augmenta amb eltemps d’incubació. Els transcrits de les diversesisoformes del receptor VEGF-R1, el que codifica laforma completa del receptor de membrana i dostranscrits que codifiquen la forma soluble (el dominiextracel·lular), augmenten amb el temps d’incubació.A causa de l’existència de dos transcrits de la formasoluble no havia estat prèviament descrita, els hemcaracteritzat per seqüenciació.
L’expressió d’Angiopoetina-2 no varia amb el tempsd’incubació i l’expressió del factor inhibidor de l’angio-gènesi ocular PEDF disminueix. No coneixem elmecanisme dels canvis d’expressió de les molèculesangiogèniques i antiangiogèniques en funció deltemps d’incubació. Una possibilitat podria ser lahipòxia existent durant el temps d’incubació, queincrementaria l’expressió de VEGF i VEGF-R1 idisminuiria la del factor PEDF, tal com ha estat descriten altres teixits. No obstant això, l’Ang-2, que ésinduïda per la hipòxia i el VEGF, no augmenta quans’incrementa el temps d’incubació.
6. Efecte de la inhibició del proteasoma enl’expressió de molècules angiogèniques i antian-giogèniques en cultius de pecten oculi
FUNDACIÓ LA MARATÓ DE TV3

Els inhibidors reversibles del proteasoma mostren unaacció antiangiogènica i es troben en fase d’assaigclínic per al tractament del càncer i malalties inflama-tòries, entre d’altres. La possible utilitat terapèuticad’aquests inhibidors per induir la regressió vascular enla retinopatia diabètica va constituir la raó principal perintroduir un nou objectiu en el projecte. Aquestobjectiu consistia en la utilització del nou modeldescrit anteriorment per estudiar l’efecte d’un inhibidorreversible del proteasoma, el pèptid boronat MG262,sobre l’expressió de les molècules angiogèniques iantiangiogèniques anteriorment esmentades. Enpresència de l’inhibidor disminueix lleument l’expressióde VEGF i Ang-2, no es modifica l’expressió de VEGF-R2 i Ang-1 i disminueix dràsticament l’expressió delreceptor VEGF-R1, principalment del transcrit quecodifica la forma complerta del receptor tirosinaquina-sa. Respecte dels transcrits que codifiquen la formasoluble del receptor VEGF-R1, d’acció antiangiogèni-ca, s’observa que l’inhibidor no disminueix l’expressiódel transcrit més petit, amb el 3’UTR curt, i disminueixla del transcrit de la forma soluble amb el 3’UTR llarg.
7. Efecte de la inhibició del proteasoma enl’expressió de VEGF-R1 en cultius de cèl·lulesendotelials microvascularsLes cèl·lules microvasculars de dermis humanaexpressen el receptor VEGF-R1, tant el transcritcorresponent a la forma completa com els dostranscrits corresponents a la forma soluble. Enpresència de l’inhibidor del proteasoma MG262 esredueix l’expressió de les diverses formes.
8. Efecte de la inhibició del proteasoma enl’expressió de VEGF-R1 en cultius de macròfagsA diferència del receptor VEGF-R2 que té una expres-sió molt restringida, principalment a cèl·lules endote-lials, el receptor VEGF-R1 s’expressa també alsmacròfags. Atesa la importància de la participaciódels macròfags en els fenòmens de l’angiogènesi, ensvam proposar investigar l’expressió del receptorVEGF-R1 en aquestes cèl·lules en presència o enabsència de l’inhibidor del proteasoma MG262. Quanels macròfags no han estat activats, l’expressió delreceptor VEGF-R1 no és gairebé visible. Ben alcontrari, quan els macròfags són tractats amb lipopo-lisacàrid (LPS) s’indueix de forma potent l’expressióde les diverses isoformes del receptor VEGF-R1,particularment del transcrit més curt que codifica laforma soluble. Aquesta inducció és completamentbloquejada per l’inhibidor del proteasoma. Si el tracta-ment amb l’inhibidor del proteasoma es fa una vegadaha tingut lloc la inducció amb LPS, s’observa unamarcada reducció de la forma completa, una reducciódel transcrit llarg de la forma soluble i la relativaconservació del trànscrit curt de la forma soluble.
Els resultats obtinguts amb els macròfags són similarsals que s’obtenen amb el pecten i corroboren que lainhibició reversible del proteasoma fa disminuirl’expressió dels transcrits que codifiquen la formacompleta del receptor i mantenen la concentració deltranscrit que codifica la forma soluble del receptorVEGF-R1. Considerant el paper dels macròfags enl’angiogènesi i la funció clau atribuïda al receptorVEGF-R1 dels macròfags, l’observació per primera
vegada al nostre laboratori dels efectes dels inhibidorsreversibles del proteasoma, tot disminuint la formaangiogènica del VEGF-R1 i preservant la forma antian-giogènica del receptor soluble, pot tenir en el futurrellevància terapèutica.
3. Rellevància i possibles implicacions clíniquesdels resultats finals obtingutsEs caracteritza l’estructura del gen de l’Angiopoetina-2i de les diverses isoformes dels RNAm del genLa importància funcional de l’Angiopoetina-2 en elsfenòmens d’angiogènesi i regressió vascular justifical’interès de caracteritzar per primera vegada el gen del’Angiopoetina-2 i de descriure les isoformes expres-sades del gen que hem anomenat A, B i C. Solamentsón segregades les isoformes A i C. La B es minorità-ria i no és segregada.
Es proposa un nou model, el cultiu de l’òrganvascular pecten oculi, per investigar l’angiogènesi ila regressió vascularLa utilització del pecten oculi in vitro com a model perinvestigar l’angiogènesi i la regressió vascular ésl’aportació més interessant dels resultats del projecte,ja que permet, a diferència dels cultius de cèl·lulesendotelials i macròfags, estudiar l’expressió coordina-da del conjunt de molècules angiogèniques i antian-giogèniques que intervenen en la formació de l’òrganvascular en condicions experimentals diferents. Hemestudiat l’expressió de VEGF i els seus receptorsVEGF-R1 i VEGF-R2, de les angiopoetines 1 i 2, de laN-cadherina i dels inhibidors de l’angiogènesi PEDF itrombospondina-1 durant el desenvolupament delpecten i durant la incubació in vitro.
En cultius de pecten oculi, la inhibició del proteaso-ma fa disminuir l’expressió del receptor demembrana angiogènic VEGF-R1 i no disminueix laforma soluble antiangiogènica del mateix receptorLa possible utilitat terapèutica d’aquests inhibidors perinduir la regressió vascular en la retinopatia diabèticava constituir la raó principal per estudiar l’efecte d’uninhibidor reversible del proteasoma, el pèptid boronatMG262, sobre l’expressió de les molècules angiogèni-ques i antiangiogèniques anteriorment esmentades.L’observació més interessant, amb possibles implica-cions terapèutiques, ha estat la disminució de formadràstica de l’expressió del receptor VEGF-R1 d’accióangiogènica mentre no disminueix la forma solubleantiangiogènica del mateix receptor.
En cultius de cèl·lules endotelials microvasculars imonòcits/macròfags, amb***1* la inhibició delproteasoma disminueix l’expressió del receptor demembrana angiogènic VEGF-R1 i no disminueix laforma soluble antiangiogènica del mateix receptorEls resultats observats utilitzant el model del pectenoculi han estat corroborats pels efectes de l’inhibidorsobre cultius de cèl·lules endotelials i macròfags.L’inhibidor reversible del proteasoma MG262 potinhibir totalment l’activació dels macròfags pel lipopoli-sacàrid (LPS) o reduir l’expressió de la tirosinquinasaVEGF-R1 una vegada els macròfags han estatactivats. El transcrit de la forma soluble antiangiogèni-ca del receptor no disminueix de forma important coma conseqüència del tractament. La disminució de la
RECERCA BIOMÈDICA | 163

forma angiogènica del VEGF-R1 i la relativa preserva-ció de la forma soluble antiangiogènica pot tenir en elfutur rellevància terapèutica.La prioritat dels resultats esmentats, que poden tenirrepercussió terapèutica, sobre altres aspectes quefiguraven en el projecte van determinar un canvi en lesprioritats assolides. L’efecte de la hipòxia, hiperglucè-mia i els estudis in situ, són ara més senzills amb elnou model proposat.
4. PublicacionsMezquita J., Mezquita, P., Pau M., Mezquita, B., Francone, V.,
Vilagrasa, X. and Mezquita C. Genomic structure and alternative
splicing of chicken Angiopoietin-2. Biochem Biophys Res
Commun, 275: 643-651 (2000).
Mezquita J, Mezquita B, Pau, M and Mezquita, C. Down-
regulation of Flt-1 gene expression by the proteasome inhibitor
MG262. J. Cell. Biochem. 89: 1138-47 (2003).
Mezquita B, Mezquita J, Pau M and Mezquita, C. The reversible
proteasome inhibitor MG262 down-regulates the expression of VEGF
and its high affinity receotir Flt-1 in human monocytes/macrophages.
Keystone Symposia. Angiogenesis: Novel Basic Science Insights
and Human Therapy. Abstract book, 69 (2004).
FUNDACIÓ LA MARATÓ DE TV3

RECERCA BIOMÈDICA | 165
MEMÒRIA FINAL
1. Resum del projecte originalObjectiusL’objectiu del projecte és investigar els efectes delete-ris de la hiperglicèmia en el trasplantament d’illotspancreàtics, i la prevenció amb insulina. S’estudiaranespecialment uns dies crucials, que són els inicialsdesprés del trasplantament, en què es determinaranels efectes de la hiperglicèmia en la mort i funció delsillots trasplantats i s’investigaran potencials mecanis-mes causants del dany sobre les cèl·lules beta.
Els objectius específics del projecte són investigar si:
1) L’exposició a hiperglicèmia després del trasplanta-ment origina un augment de la mort (apoptosi i/onecrosi) de les cèl·lules beta trasplantades, compara-da amb la dels illots trasplantats en receptorsnormoglicèmics. La mort cel·lular s’estudiarà en elsprimers dies després del trasplantament i en els illotsexposats a hiperglicèmia crònica.
2) Es produeix una major pèrdua de massa beta elsprimers dies després del trasplantament en els illotsexposats a hiperlgicèmia en comparació amb els illots
trasplantats en receptors normoglicèmics.3) Els illots trasplantats mostren, els primers diesdesprés del trasplantament, una alteració en la funcióbeta (expressió del gen de la insulina, contingutd’insulina, secreció d’insulina), que pugui estaragreujada als illots exposats a hiperglicèmia encomparació amb els illots trasplantats en receptorsnormoglicèmics.
4) Hi ha una major expressió de citoquines en elsempelts d’illots trasplantats en receptors hiperglicè-mics, comparant-ho amb els illots trasplantats enreceptors normoglicèmics, que contribueixin a l’efectedeleteri de la hiperglicèmia en el pronòstic deltrasplantament.
DissenyTrasplantament isogènic d’una massa subòptimad’illots pancreàtics en la càpsula renal de receptorsdiabètics per estreptozotocina, tractats amb insulina(normoglicèmics) o no tractats (hiperglicèmics).
Es determinarà, en l’empelt: apoptosi beta (TUNEL),replicació beta (immunohistoquímica) i massa beta(morfometria), secreció i contingut d’insulina (RIA),contingut de DNA (fluorimetria), expressió del gen
Títol del projecte
Efecte de la hiperglicèmia en el fracàs primari del trasplantament d'illots pancreàtics: contribució a la mort i disfunció de les cèl·lules beta-trasplantades
Investigadors principalsEduard Montanya Mias
InstitucióCiutat Sanitària i Universitària de Bellvitge
80.947 €

d’insulina (Northern blot) i expressió gènica de citoqui-nes (RT-PCR).
2. Resultats1. El manteniment de la normoglicèmcia en elperíode immediat previ i posterior al trasplantamentd’illots permet millorar el pronòstic del trasplanta-ment en les dues espècies estudiades: ratolí (figura1a) i rata (figura1b)Els animals van ser trasplantats amb una massa betainsuficient per restablir la normoglicèmia en elsreceptors diabètics (100 illots en ratolins C57Bl/6 i500 illots en rata Lewis). Els receptors que no vanrebre tractament amb insulina van romandre hipergli-cèmics durant el seguiment (dies 3, 10 i 30 desprésdel trasplantament), mentre que els tractats ambinsulina, els dies previs i posteriors al trasplantamentvan assolir la normoglicèmia definitiva després de sersuspès el tractament amb insulina.
Figura 1a. Evolució metabòlica en ratolins diabètics trasplantats.
Els grups 1-3 no van rebre tractament amb insulina, els grups 4-6
van ser tractats des de 7 dies abans del trasplantament fins a 10
dies després.
Figura 1b. Evolució metabòlica en rates Lewis diabètiques
trasplantades. El grup normoglicèmic va ser tractat amb insulina
des de 7 dies abans del trasplantament i va aconseguir mantenir
la normoglicèmia després de ser suspès el tractament 14 dies
després del trasplantament. El grup hiperglicèmic va rebre el
mateix nombre d’illots, però no va ser tractat amb insulina, i es va
mantenir hiperglicèmic durant l’estudi.
La demostració de l’efecte beneficiós de la insulina endues espècies diferents (ratolí el primer any, rata el segon)suggereix que aquest efecte és generalitzable, i que espodria donar també en el trasplantament d’illots humans.
2. Els primers dies després del trasplantament télloc un augment en la mort de cèl·lules betatrasplantades. La mort de cèl·lules beta s’esdevéper necrosi i apoptosi. Els mecanismes de mortcel·lular (apoptosi i necrosi) són semblants en lesdues espècies estudiades (rata i ratolí). (figura 2)3. L’exposició crònica de l’empelt a hiperglicèmia
s’acompanya d’una elevada apoptosi de lescèl·lules beta que no es detecta en els empelts, quemantenen la normoglicèmia del receptor. Aquestaalta apoptosi va anar acompanyada d’una reduccióde la massa beta de l’empelt, que no va esdevenir-se en els animals normoglicèmics (figura 2).Els primers dies després del trasplantament es vaidentificar la presència d’extenses àrees de necrosi enels empelts (al voltant del 30% del teixit endocrí del’empelt al cap de 3 dies del trasplantament) tant enels receptors normoglicèmics (26,8%) com hiperglicè-mics (30,7%). No es va detectar necrosi en etapesposteriors del seguiment (dies 10 i 30).
Els primers dies després del trasplantament, elsempelts van mostrar un augment en l’apoptosi de lescèl·lules beta respecte dels illots del pàncrees controlscom també dels illots immediatament després del’aïllament. L’augment de l’apoptosi es va detectar tantentre els grups d’animals hiperglicèmics com en elsnormoglicèmics als 3 i 10 dies després del trasplanta-ment. Tanmateix, 30 dies després del trasplantamentes van observar diferències entre els grups: mentreque en els empelts dels animals hiperglicèmics l’apop-tosi de les cèl·lules beta es mantenia elevada, en elsempelts dels animals normoglicèmics l’apoptosi vaminvar a nivells basals.
Figura 2. Apoptosi de les cèl·lules beta trasplantades en ratolí
C57Bl/6. Els grups 1-3 no van rebre tractament amb insulina i van
romandre hiperglicèmics durant l’estudi. Els grups 4-6 van ser
tractats amb insulina des de 7 dies abans del trasplantament fins a
10 dies després i es van mantenir normoglicèmics durant l’estudi.
4. El manteniment de la normoglicèmia els primersdies després del trasplantament permet reduirl’apoptosi de les cèl·lules beta dels illots trasplan-tats en rates però no en ratolí, tot indicant ladiversa sensibilitat als efectes de la hiperglicèmiaen espècies diferents
Figura 3. Apoptosi els primers dies després del trasplantament
isogènic en rates Lewis.
Al cap de 3 dies del trasplantament es va detectar un
FUNDACIÓ LA MARATÓ DE TV3

augment significatiu de l’apoptosi en les cèl·lules betatrasplantades, en comparació amb les cèl·lules betadel pàncrees i amb l’apoptosi als illots immediatamentdesprés de l’aïllament. L’apoptosi va ser més gran alsillots trasplantats en animals hiperglicèmics) (n ) queno en els receptors diabètics tractats amb insulina (n ),que es mantenien normoglicèmics.
5. Els primers dies després del trasplantament,l’augment de la mort per necrosi i apoptosi estradueix en la pèrdua massiva de teixit trasplantat,que representa al voltant del 60% de la massainicial (figura 4)La massa de cèl·lules beta recuperada en els empeltsal cap de 3 dies del trasplantament va ser un 60%inferior a la inicialment trasplantada, tant en animalsexposats a hiperglicèmia com en els normoglicèmicspel tractament amb insulina. En el grup tractat ambinsulina, la massa beta no es va modificar 10 diesdesprés del trasplantament comparada amb la del dia3, però va augmentar significativament després desuspendre el tractament amb insulina per fer front a lanova demanda metabòlica per mantenir la normogli-cèmcia, en concordança amb la reducció de l’apoptosique es mostra en la figura 2. En canvi, al cap de 30dies del trasplantament, la massa beta dels empeltsexposats a hiperglicèmcia sostinguda s’havia reduïtsignificativament comparada amb la trasplantada inicial-ment, d’acord amb la persistència de l’alta apoptositrobada en el grup exposat a hiperglicèmia crònica.
Figura 4. Evolució de la massa beta trasplantada.
Els grups 1-3 no van rebre tractament amb insulina ivan romandre hiperglicèmics durant l’estudi. Els grups4-6 van ser tractats amb insulina des de 7 dies abansdel trasplantament fins a 10 dies després i es vanmantenir normoglicèmics durant l’estudi.
6. Immediatament després de l’aïllament ja esdetecta l’expressió del gen de la citoquina proinfla-matòria IL-1b als illots, i aquesta expressióaugmenta en els empelts els primers dies desprésdel trasplantament. L’expressió d’IL-1b es manifes-ta tant als illots trasplantats en receptors normogli-cèmics com en els hiperglicèmicsTot just després de l’aïllament es va detectar l’expressiódel gen d’IL-1b als illots, i aquesta expressió va augmen-tar els primers dies després del trasplantament, malgrattractar-se d’un empelt singènic en el qual es descartavala presència de rebuig o recurrència de l’autoimmunitat.Els resultats indiquen que l’inici del procés inflamatori jas’esdevé durant l’aïllament, i que aquest augmenta elsprimers dies després del trasplantament. Aquest procés
inflamatori pot contribuir a la mort de les cèl·lulestrasplantades descrites en l’apartat 2.
Figura 5. Expressió del gen d’IL-1b en empelts singènics.
Es van estudiar illots acabats d’aïllar i empelts delsdies 1, 3 i 7 després del trasplantament. Els resultatscom a valors relatius d’IL-1b , expressats en unitatsarbitràries (quocient IL-1b /ciclofilina).
7. L’expressió del gen de la forma induïble de lasintasa de l’òxid nítric (iNOS) s’identifica igualmentals illots immediatament després de l’aïllament, iaquesta expressió augmenta als empelts elsprimers dies després del trasplantament (figura 7).
8. Hi ha una correlació significativa entre l’expres-sió del gen d’IL-1b i el d’iNOS als illots i alsempelts, la qual cosa suggereix que IL-1b , permitjà de la inducció d’iNOS, pot tenir un paper enla mort dels illots identificada els primers diesdesprés del trasplantament.D’acord amb els resultats de l’expressió del gen d’IL-1b mostrats en l’apartat anterior, es va detectarl’expressió del gen iNOS als illots després de l’aïlla-ment, i un augment els primers dies després deltrasplantament. La correlació positiva entre l’expressiódel gen d’IL-1b i el d’iNOS (figura 7) suggereix quel’expressió del gen d’IL-1b està implicat en la del gend’iNOS, i l’activació d’aquesta via d’inducció de mortcel·lular als illots trasplantats.
Figura 6. Expressió d’iNOS en illots i empelts.
RECERCA BIOMÈDICA | 167

A: les línies 1-6 corresponen a mostres d’illots immedia-tament després de l’aïllament. B: la línia 7 correspon aun trasplantament isogènic d’illots de 3 dies, la línia 8 aun al·lotrasplantament de 5 dies, i C+ a illots de ratatractats durant 6 hores amb IL-1b (control positiu).
Figura 8. Representació gràfica de la relació lineal entre l’expressió
del gen d’IL-1b i el gen d’iNOS.
Els cercles indiquen les unitats arbitràries de l’expres-sió d’ambdós gens per a cada mostra individuald’empelts d’illots.
9. S’ha identificat la presència d’IL-1b en elsempelts els primers dies després del trasplanta-ment i s’ha establert que el principal origen cel·lulard’IL-1b son macròfags de l’empelt.Un cop identificada l’expressió del gen d’IL-1b , es vainvestigar la presència de la citoquina en els empelts iel seu origen cel·lular. Per això es va fer una dobletinció amb un anticòs anti-IL-1b que va confirmar lapresència de cèl·lules positives en tots els empelts iamb un anticòs anti-CD68 que identificà elsmacròfags com el principal, però no pas l’únic, tipuscel·lular productor de la citoquina
Figura 9. Detecció d’IL-1b als macrófags de l’empelt per mitjà de
doble tinció amb un anticòs anti-CD68 visualitzat amb diaminoben-
zidina al microscopi òptic (A) i un anticòs anti-IL-1b identificat i un
anticòs secundari marcat amb TRIC (B). Les imatges corresponen a
la mateixa secció d’un empelt singènic al cap de 3 dies del trasplan-
tament i les fletxes indiquen macròfags positius per a IL-1b .
En resum, en aquest projecte s’ha demostrat que elsprimers dies després del trasplantament d’illots esprodueix una pèrdua important del teixit trasplantat.Més de la meitat de la massa beta trasplantada es vaperdre arran d’un augment en la mort cel·lular beta
tant per processos d’apoptosi com de necrosi. Enaquests primers dies després del trasplantament vamidentificar l’expressió dels gens d’IL-1b i iNOS enl’empelt, tot indicant la presència d’un procés inflama-tori al lloc d’implantació dels illots que podria tenir unpaper en l’augment de l’apoptosi i necrosi detectada.L’augment en l’expressió dels gens d’IL-1b i iNOS esva produir tant en animals diabètics hiperglicèmicscom en animals diabètics normoglicèmics pel tracta-ment amb insulina. Els animals hiperglicèmics presen-taven una expressió menys elevada dels gens d’IL-1bi iNOS que els normoglicèmics, tot suggerint lapossible existència d’una limitació en l’augment del’expressió d’aquests gens en els empelts exposats ahiperglicèmia. L’augment en l’expressió dels gensd’IL-1b i iNOS amb una correlació significativa entreells suggereix que aquests gens participen en elsfenòmens inflamatoris i la mort cel·lular que té lloc elsprimers dies després del trasplantament. La identifica-ció de la presència de la citoquina els primers diesdesprés del trasplantament indica que l’expressió delgen d’IL-1b va donar lloc a l’expressió de la proteïnaen l’empelt, i confirma la presència d’un procésinflamatori no lligat a rebuig els primers dies desprésdel trasplantament i que té lloc fins i tot als empeltssingènics. El patró temporal d’expressió que hemidentificat suggereix que el procés inflamatori s’iniciaamb l’aïllament dels illots i s’exacerba amb el trasplan-tament, tot assolint molecularment el seu nivell màximel primer dia després del trasplantament i donant lloca la presència d’IL-1b en l’empelt. Considerant que lacitotoxicitat d’IL-1b sobre les cèl·lules beta està benestablerta, les nostres dades en conjunt suggereixenque IL-1b té un paper en la destrucció massiva decèl·lules beta que s’esdevé els primers dies desprésdel trasplantament. La caracterització d’aquestsfenòmens obre una via per al desenvolupamentd’estratègies que permetin millorar la supervivènciadels illots trasplantats.
3. Rellevància i possibles implicacions clíniquesdels resultats finals obtingutsLa detecció d’una molt important mort cel·lular elsprimers dies després del trasplantament ens hapermès identificar un període crític després deltrasplantament vers el qual dirigir estratègies terapèu-tiques que permetin millorar el pronòstic de l’empelt.La identificació que l’apoptosi participa en la mort elsprimers dies del trasplantament d’illots obre un campde recerca per conèixer els mecanismes de destruc-ció de les cèl·lules beta trasplantades en aquestperíode crític inicial. La identificació de l’elevadaexpressió del gen d’IL-1b i iNOS en els empelts, i ladetecció de la presència d’IL-1b en els empelts, és unpas en aquest sentit, atès que ens ofereix la possibili-tat d’elaborar estratègies adreçades al bloqueig de laseva acció amb la intenció de millorar la supervivènciadel teixit trasplantat i, en conseqüència, de l’empelt, ien les quals actualment el nostre grup ja estàtreballant. D’altra banda, la identificació d’una apopto-si beta més gran en illots exposats a hiperglicèmiacrònica pot tenir importància en la comprensió delsmecanismes patogenètics que menen al desenvolu-pament de la diabetis mellitus, i particularment de ladiabetis tipus 2.
FUNDACIÓ LA MARATÓ DE TV3

4. PublicacionsBiarnés M , Montolio M, Nacher V, Raurell M. Soler J,
Montanya E.Hyperglycemia and beta cell death in syngeneically
transplanted islets. Diabetes 2002;51:66-72
Montanya E. El trasplante de islotes de páncreas como terapia
en la diabetes tipo 1. Endocrinologia 2001;48:223-225.
Berná G, León-Quinto T, Enseñat-Waser R, Montanya E, Martin
F, Soria B. Stem Cells and Diabetes. Biomed Pharmacother
2001;55:206-12.
Biarnés M, Montolio M, Téllez N, Soler J, Montanya E.
Apoptosis and IL-1ßeta mRNA expression in syngeneic rat islet
grafts. Diabetologia 2001;44(Suppl 1):A144.
Montolio M, Biarnés M, Téllez N, Soler J, Montanya E.
Caspase inhibition reduces initial beta cell apoptosis in syngenei-
cally transplanted islets. Diabetologia 2002;45(Supl 2):A138
Montolio M, Montanya E. Increased beta cell apoptosis in
transplanted islets is reduced with caspase inhibitors. Avances en
Diabetologia 2002;18:189.
Montolio M, , Téllez N, Biarnés M, Soler J, Montanya E.
Short term islet culture with the caspase inhibitor z-VAD.fmk
reduces beta cell apoptosis after transplantation and improves the
metabolic outcome of the graft. (enviat per a publicació)
Biarnés M, Montolio M, Téllez N, Escoriza J, Soler J, Montanya E.
Increased expression of IL-1ß and iNOS in early syngeneic islet
trasplantation. (enviat per a publicació).
RECERCA BIOMÈDICA | 169

FUNDACIÓ LA MARATÓ DE TV3
MEMÒRIA FINAL
1. Resum del projecte originalIntroduccióEls dipòsits anomenats d’amiloide que s’han detectatals illots pancreàtics de pacients afectats de diabetistipus 2, estan formats per l’agregació i precipitació del’amilina. Aquest és un pèptid de 37 aminoàcids quees fabrica a les cèl·lules b dels illots pancreàtics i s’alli-bera paral·lelament amb la insulina. S’ha demostratque l’amilina exerceix accions perifèriques antagonis-tes de la insulina i, per tant, el seu paper fisiològic potcontribuir a la regulació del metabolisme de laglucosa. El que no es coneix és la causa que provocala conversió de l’amilina en fibres insolubles que esdipositen al pàncrers i formen els dipòsits d’amiloide.La presència d’aquests dipòsits exerceix un papercrític en la progressiva alteració i destrucció de lescèl·lules b que es desenvolupa tot al llarg del tempsen la diabetis tipus 2. La nostra hipòtesi de treballestableix que una alteració en la funció de la cèl·lula bque comporti canvis en la producció, el processamenti la secreció de l’amilina, poden ser crítics per iniciar laseva transformació en forma de fibres insolubles, queprecipiten i donen lloc als dipòsits d’amiloide pancreà-tic, típics de la diabetis humana. La recerca de
defectes en el gen de l’amilina, com també l’estudi del’expressió, regulació i activitat de l’enzim iduronat-2-sulfatasa, implicat en la modulació del substrat on esdiposita l’amilina, poden ser rellevants per entendreels mecanismes moleculars de la formació d’amiloideals illots pancreàtics en la diabetis tipus 2.
Objectius específics1. Investigar la presència de possibles mutacions en elgen de l’amilina en pacients afectats de diabetis tipus2 i en població control sense diabetis.
2. Estudi de les característiques clíniques delspacients portadors de mutacions per establir correla-ció entre el fenotip i el genotip.
3. Estudio del pedigrí familiar de los sujetos portadoresde mutaciones para determinar la forma de transmisión.
4. Investigar l’activitat del promotor en un sistema invitro, mitjançant tècniques de mutagènesi i detransfecció en línies cel·lulars de ratolí.
5. Investigar la localització i la regulació de l’enzimIduronat-2-Sulfatasa (IDU) en la cèl·lula b pancreàticahumana i de ratolí.
Títol del projecte
Factors determinants de l'amiloïdogènesi en la cèl·lula b pancreàtica i la seva relació patogènica amb la diabetis tipus 2
Investigadors principalsAnna Novials Sardà · Ramon Gomis de Barbarà en subrogació de la
Dra. Elena Fernández Usac · Anne Clark
InstitucióFundació Sardà Farriol·Hospital Clínic i Provincial de Barcelona·Oxford University
280.408 €

Disseny, procediments i metodologia. PoblacióS’ha comptat amb la col·laboració de 677 persones,356 de les quals tenien diabetis tipus 2, 207 erencontrols sense diabetis i la resta (114) tenien altrestipus de diabetis. Tots ells, amb l’acceptació prèvia i lasignatura del consentiment informat, han ceditmostres de sang per a obtenció de l’ADN.Extracció de l’ADN genòmic segons tècniques Standard
Anàlisi de polimorfismesS’analitza l’ADN mitjançant la tècnica d’SSCP.
Seqüenciació d’ADNLes mostres amb patró de migració anormal sónsotmeses a seqüenciació automàtica.
Estudi de l’activitat del promotor del gen de l’amilinaMitjançant tècniques de transfecció a línies cel·lulars(assaig activitat del gen luciferasa) s’estudia l’efecte demutacions en la regió promotora per veure si s’alterala transcripció genètica.
Estudi de l’enzim IDS als illots pancreàtics humansi de ratolí1. Anàlisi de la localització específica cel·lular de l’IDSen teixit humà per tècniques de PCR i d’immunofluo-rescència.
2. Anàlisi dels senyals metabòlics que regulenl’expressió del gen de l’IDS per tècniques de PCR atemps real.
3. Determinació de l’efecte de l’IDS sobre la funciósecretora dels illots pancreàtics mitjançant tècniquesde transfecció amb lentivirus i microscòpia electrònica.
2. ResultatsDetecció de mutacions en el promotor del gen del’amilinaL’estudi genètic ha detectat la presència d’una mutacióen heterozigosi en la regió promotora del gen de l’amili-na, consistent en el canvi Gà A en posició —132 bp delpunt d’inici de transcripció del gen. Aquesta variant haestat detectada en 38 de 356 pacients amb diabetistipus 2 i en 3 de 207 controls no diabètics. La freqüèn-cia del genotip GA és significativament superior en lapoblació diabètica que en la població control (10.7% vs1.4%, P < 0.001, odds ratio: 8.3).
Característiques clíniques dels pacients portadorsde mutacióL’anàlisi de les característiques clíniques diferencialsdels 38 pacients afectats de diabetis tipus 2 portadorsde la mutació, respecte dels 318 pacients sensedetecció de la mutació, ha demostrat diferènciessignificatives en relació amb les xifres de tensióarterial. El 74% de diabètics portadors de la mutaciótenen HTA, a diferència del grup no portador, en quèes detecta HTA en un 57% de la població. Aquestadiferència és estadísticament significativa (p<0.05%).Això vol dir que els pacients portadors de la mutaciótenen tendència a ser més hipertensos.
Estudi familiarS’han estudiat fins a l’actualitat 31 familiars de primer
grau (fills i germans dels portadors). Hem detectat lamutació en 16 persones, la qual cosa representa un50% dels familiars. Aquesta troballa indica la transmis-sió hereditària de l’anomalia genètica. De les personesportadores de la mutació, una d’elles és diabètica i lesaltres dues presenten un trastorn d’intolerància a laglucosa (estadi de prediabetis).
La detecció de la mutació en familiars no diabèticsens indica, atès que són població jove, que caldràprendre mesures preventives i fer el seguiment de laseva evolució, per evitar l’aparició de la diabetis.
Estudi in vitro de l’activitat del promotorL’activitat transcripcional del promotor de l’amilina,tant de la seqüència normal com de la mutada, s’haestudiat utilitzant l’anomenat assaig de luciferasa,descrit en els apartats anteriors. El promotor portadorde la mutació presenta el doble d’activitat que elpromotor salvatge, és a dir que la seqüència normal(p<0.001). L’activitat d’ambdós promotors es regulaper senyals que provenen del metabolisme de laglucosa i que al mateix temps reclama la presència decalci en el medi, per a la seva funcionalitat.
El resultat d’aquests experiments són prou convin-cents per concloure que la mutació G/A provoca unaugment de l’activitat de la transcripció del gen del’amilina en un sistema in vitro. L’interès d’aquestatroballa serà poder-ho demostrar en un futur en unsistema in vivo, és a dir, en les persones portadoresde la mutació.
Localització i regulació del enzim IDS en la cèl·lulapancreàticaL’enzim iduronat-2-sulfatasa (IDS) és l’encarregat dedegradar els proteoglicans, substàncies localitzades ala membrana basal. La seva activitat principal éseliminar el grup sulfat del dermatan i de l’heparansulfat (compostos de la membrana basal). L’objectiuprincipal d’aquest treball és investigar l’expressió i laregulació del gen IDS als illots pancreàtics de ratolí.
El gen de l’IDS s’expressa específicament als illotspancreàtics, tant en cèl·lules beta com en cèl·lules alfai es localitza als lisosomes del citoplasma cel·lular. Elsnivells d’mRNA d’IDS dels illots cultivats amb diferentsconcentracions de glucosa, augmenta de formadependent de la dosi. La sobreexpressió del gen del’IDS no provoca cap canvi en el contingut d’insulinani en la resposta secretora dels illots respecte del’estimulació de glucosa. En canvi, quan es bloquejal’expressió d’IDS als illots, es produeix una disminuciósignificativa en el contingut d’insulina (35 % versus elcontrol, p< 0.005).
Podem concloure que l’enzim IDS (iduronat-2-sulfata-sa) es localitza i s’expressa específicament als illotspancreàtics. Tanmateix, es regula per glucosa i pot serun enzim essencial en el manteniment de la funció dela cèl·lula beta.
3. Rellevància i possibles implicacions clíniquesdels resultats finals obtingutsL’interès de trobar un marcador genètic que defineixiun risc de patir diabetis en una població determinada,
RECERCA BIOMÈDICA | 171

ve donat pel fet que es podran establir una sèrie demesures sanitàries preventives en aquest subgrup depoblació.
A mesura que puguem confirmar la importància de lamutació trobada en el gen de l’amilina com amarcador genètic, es podran desenvolupar estratègiesterapèutiques de futur, pel que fa a la recerca de nousfàrmacs que inhibeixin el procés anomenat d’amiloïdo-gènesi pancreàtica o de formació de precipitats de lamolècula d’amilina, responsables finals de la formaciódels dipòsits d’amiloide típics de la diabetis tipus 2.
Aquest raonament es justifica plenament, atès que sesap que al llarg dels anys d’evolució de la diabetis,l’acumulació d’aquests dipòsits produeix toxicitat idestrucció de les cèl·lules pancreàtiques, amb lapèrdua consegüent de la seva funció principal, que ésfabricar insulina.
Els resultats dels experiments in vitro que actualmentestà duent a terme el nostre grup de recerca, comtambé altres grups d’investigació, avalaran —comesperem— els plantejaments de la nostra hipòtesiclínica.
4. PublicacionsNovials A., Rojas, I., Franco C., Casamitjana R., Fernández
Usac E., Gomis R. A novel mutation in Islet Amyloid Polypeptide
gene promoter is associated with type 2 diabetes mellitus.
Diabetologia 2001, 44:1064-1065.
Rojas I., Gomis R., Casals E., Quinto Ll., Franco C., Novials A.
Polymorphism in intron 2 of islet amyloid polypeptide gene is
associated with lower low-density lipoprotein cholesterol in
mondiabetic subjects and in type 2 diabetic patients. Endocrine
2002, 19:185-9.
Novials A., Kistauri A., Chico A., Gomis R. Letter To: Poa NR,
Cooper GJS, Edgar PF: Amylin gene promoter mutations predis-
pose to Type 2 diabetes in New Zealand Maori. Diabetologia 46:
574.578. Diabetologia 2003, 1708-1709.
Novials A., Mato E., Lucas M., Franco C., Ribas M.,
Santisteban P., Gomis R. Mutation at the position —132 in the
islet Amyloid Polypeptide (IAPP) gene promoter enhances the
basal transcriptional activity through a new CRE-like binding site.
(en revisió per publicar a la revista Diabetologia).
Coronado-Pons I., Novials A., Clark A., Gomis R. Identification
of Iduronate-2-Sulfatase (IDS) in mouse pancreatic islets. (en
revisió per publicar a la revista American Journal Physiology).
FUNDACIÓ LA MARATÓ DE TV3

RECERCA BIOMÈDICA | 173
MEMÒRIA FINAL
1. Resum del projecte originalObjectius1. Determinació de la prevalença i tipus de diabetismellitus en malalts diagnosticats d’hemocromatosihereditària.1.1 Selecció de la mostra:Inclusió d’una mostra de 50 malalts consecutiusdiagnosticats d’hemocromatosi hereditària a l’HospitalClínic i Provincial.
1.2 Determinacions bioquímiques: glucosa, test detolerància de glucosa, insulinèmia, ferritina sèrica iíndex de saturació de transferrina (IST) a tots elsparticipants.
1.3 Anàlisi genètica (anàlisi del gen HFE en recerca deles mutacions C282Y i H63D) a tots els participants.Es disposa ja del genotip corresponent a 47 malaltsamb hemocromatosi hereditària.
1.4 Resposta dels malalts al tractament per flebotomiaquant a l’evolució o prevenció de la diabetis.2. Prevalença de disfuncions en el metabolisme del
ferro i de mutacions del gen HFE en 500 malaltsconsecutius amb diabetis tipus 2.
2.1 Selecció dels malalts: 500 malalts consecutiusamb diabetis tipus 2 que acudeixen espontàniament al’HCP. Es disposen ja de 200 malalts catalogats en lanostra Unitat de Diabetis.2.2 Determinacions bioquímiques: glucosa, test detolerància de glucosa, insulinèmia, ferritina sèrica i índexde saturació de transferrina (IST) a tots els malalts.
2.3 Inici o seguiment dels tractaments convencionalshabituals de la diabetis.
2.4 Anàlisi genètica (anàlisi del gen HFE en recerca deles mutacions C282Y i H63D) a tots els malalts.
2.5 Determinació de la prevalença de mutacions delgen HFE en malalts amb diabetis tipus 2.
2.6 Tractament per flebotomia dels malalts detectatsamb un excés de ferro (ferritina sèrica ? 500m g/ml i/oIST ? 55%) o bé homozigots per a mutacions del genHFE i seguiment prospectiu de la possible majoria oestabilització de seu estat diabètic.
Títol del projecte
Diabetis mellitus, sobrecàrrega cel·lular de ferro i prevalença de mutacions del gen HFE en la població espanyola
Investigador principalRafael Oliva Virgili
InstitucióHospital Clínic i Provincial de Barcelona
78.183 €

2.7 Avaluació de la possible relació cost-eficàcia de ladeterminació de mutacions del gen HFE i d’altera-cions dels paràmetres del metabolisme del ferro en eldiagnòstic etiològic d’alguns dels casos de diabetismellitus tipus 2.
3. Determinació de la relació cost-eficàcia del cribrat-ge genètic molecular (anàlisi del gen HFE) poblacio-nal i de familiars de primer grau de malalts ambhemocromatosi en la detecció de malalts homozigotsC282Y potencialment afectats de diabetis i altera-cions hepàtiques.3.1 Selecció de la mostra:
Inclusió d’una mostra de 5.000 controls poblacionals.
Inclusió de familiars de primer grau d’afectats.
3.2 Cribratge bioquímic: glucosa, ferritina sèrica i IST atots els malalts.
3.3 Anàlisi genètica (anàlisi del gen HFE en recerca deles mutacions C282Y i H63D) a tots els malalts.
3.4 Convocatòria a tots els malalts amb hiperglucè-mia, ferritina sèrica ? 500m g/ml i/o IST ? 55%, o/ihomozigots C282Y o/i heterozigots C282Y ambl’objectiu de fer: avaluació clínica completa, arbregenealògic, interrogació sobre antecedents familiars,hàbits tòxics i realització d’una nova anàlisi de sang(determinació de GOT, GPT, FA, bilirubina, albúmina,hemograma, virus C i B, temps de protrombina, testde tolerància de glucosa, insulinèmia).
3.5 Biòpsia hepàtica en dos supòsits:
a) Si persisteixen les alteracions del metabolisme delferro practicada tres mesos després i en absència dequalsevol altra causa d’hemocromatosi secundària.
b) En individus homozigots o heterozigots només si hiha manifestacions clíniques o bioquímiques de lamalaltia.
3.6 Avaluació de l’eficàcia (com anys de vidaguanyats) del cribratge genètic molecular (anàlisi delgen HFE) de poblacions i de familiars de primer graud’afectats en la prevenció de la diabetis mellitus id’altres alteracions en malalts portadors de mutacionsdel gen HFE.
3.7 Avaluació del cost del cribratge genètic molecular(anàlisi del gen HFE).
3.8 Determinació de la relació cost-eficàcia.
4. Identificació de factors reguladors de l’expressiódel gen HFE com a base del desenvolupament defàrmacs o teràpies alternatives.4.1 Identificació de proteïnes reguladores i elementsreguladors de l’expressió del gen HFE.
4.1.1 Identificació d’elements conservats en l’evolució:seqüenciació de la regió promotora del promotor delgen HFE en diverses espècies model.4.1.2 Seqüenciació de la regió promotora del gen HFE
en malalts d’HH sense mutacions C282Y.
4.1.3 Estudis de band-shift i de footprinting.
4.2 Generació d’un model de transcripció in vitro.
2. Resultats
Quant a l’estudi de pacients amb diabetis i a l’estudipoblacional, s’ha recollit i analitzat el gen HFE en latotalitat de les mostres (Sánchez et al., 2003).
En aquest treball, s’hi han inclòs 5.370 controlspoblacionals, a més dels 512 controls prèviamentestudiats en un treball pilot (Sánchez et al., 1998).S’ha analitzat la presència de possibles mutacionsC282Y i H63D en tots els participants i els nivells deferro, ferritina i transferrina. Això ha permès trobar 8 (5homes i 3 dones) donants que són homozigotsC282Y (0,15%) i 74 heterozigots compostosC282Y/H63D (1,38%), a part de determinar acurada-ment la freqüència poblacional dels al·lels mutants enla nostra població (3,16%±0,46% per a la mutacióC282Y i 20,80%±1,09% per a la mutació H63D;Sánchez et al., 2003). Hi havia 4 dels 8 homozigotsC282Y, tot homes, que presentaven nivells de ferritinai de saturació de transferrina elevats. En canvi, no s’hadetectat cap dona amb un increment en aquestsparàmetres. Només 1 dels 74 heterozigots compostosC282Y/H63D presentava nivells de ferritina i desaturació de transferrina elevats (penetració del’1,35% per a aquest genotip).
El nostre estudi també prediu, segons la llei de Hardy-Weinberg, que la freqüència del genotipC282Y/C282Y a Espanya és d’1 en 1.004 i que lafreqüència dels heterozigots compostos(C282Y/H63D) és d’1 en 72. Pel que fa a la rellevàn-cia, l’expressió bioquímica de la malaltia en la nostrapoblació és aproximadament del 80% dels homes (4de 5) detectats amb el genotip C282Y/C282Y. Pertant, podem concloure que a Espanya hi ha aproxima-dament unes 40.684 persones C282Y homozigot(40.847.371 [cens 2001] x 1 / 1004 = 40.684) i563.693 persones heterozigot compostes(C282Y/H63D) que no saben que tenen risc dedesenvolupar hemocromatosi. D’aquestes, aproxima-dament uns 23.882 homes i unes 8.136 donesexpressen o expressaran bioquímicament la malaltia sino s’hi du a terme cap mena de prevenció.
L’estudi de les regions promotores 3’ humana i deratolí ha donat lloc a diverses publicacions (Sánchezet al., 2000; 2001). En concret, es descriu la identifi-cació de proteïnes reguladores i d’elements regula-dors de l’expressió del gen HFE, i la seqüenciació deles regions promotores corresponents del gen humà,de ratolí i de la rata, i la regió promotora del gen HFEen malalts HH sense mutacions C282Y, tot aportantuna valuosa informació bàsica referent als mecanis-mes d’expressió del gen HFE. Addicionalment, quanta la caracterització dels extrems 3’ i 5’ del gen HFE,s’han seqüenciat diversos clons (EST) identificats a lesbases de dades amb la finalitat de caracteritzar moltmillor l’inici de transcripció del gen. A més, per RT-PCR i RACE s’ha aconseguit estendre la regió 3’ del
FUNDACIÓ LA MARATÓ DE TV3

gen i identificar noves variants d’splicing.
També s’ha fet un estudi de la relació cost-eficàcia,que va ser presentat al World Congress on IronMetabolism (Bioiron 2001), i previsiblement aquestesdades seran enviades per ser pubicades (Oliva et al.,2004). Un altre aspecte afrontat ha estat l’anàlisiclínica dels pacients amb diabetis tipus 2 i la sevarelació amb els genotips HFE (Oliva et al., 2004).L’aspecte més rellevant d’aquest objectiu ha estat ladetecció d’una edat d’inici més precoç de la diabetistipus 2 en els pacients amb els genotips heterozigotscompostos (C282Y/H63D), heterozigots C282Y, ohomozigots C282Y, i de la presència d’una majorincidència de retinopàtia o de nefropatia diabètica.
3. Rellevància i possibles implicacions clíniquesdels resultats finals obtingutsEl cribratge de 5.370 donants de sang per a lesmutacions C282Y i H63D del gen de l’hemocromatosiha permès determinar acuradament la freqüènciapoblacional dels al·lels mutants en la nostra població(3.16%±0.46% per a la mutació C282Y i20.80%±1.09% per a la mutació H63D; Sánchez etal., 2003). També és rellevant haver determinat que lesmutacions del gen HFE són responsables del 85,1%dels casos d’hemocromatosi al nostre país (Sánchezet al., 2000). Aquestes dades poden resultar molt útilsen la planificació de les necessitats sanitàries sobrehemocromatosi, atès que permeten predir que aEspanya hi ha aproximadament 40.684 personesC282Y homozigots i 563.693 persones heterozigotscompostes (C282Y/H63D) que no saben que tenenrisc de desenvolupar hemocromatosi.
Si a aquestes dades, hi afegim els resultats assolitssobre l’expressió bioquímica de la malaltia en la nostrapoblació (un 80% dels homes detectats amb elgenotip C282Y/C282Y i un 1,35% dels C282Y/H63D;Sánchez et al., 2003), també podem predir que uns23.882 homes i unes 8.136 dones expressen oexpressaran bioquímicament la malaltia al nostre paíssi no es fa res per prevenir-ho. Aquestes dades sónmolt rellevants, perquè un excés crònic de ferro al’organisme com el que es dóna en l’hemocromatosicondueix a cirrosi hepàtica i risc incrementat decàncer de fetge. També comporta un risc incrementatde diabetis mellitus i de cardiomiopatia, entre altresalteracions. Totes aquestes alteracions es podenprevenir si el risc de desenvolupar hemocromatosi esdetecta de manera precoç (per exemple mitjançant elcribratge genètic) i, en cas d’augment dels nivells deferro, s’inicia una prevenció. En aquest cas, la preven-ció consisteix a iniciar un tractament per mitjà dedonacions de sang periòdiques per reduir, isubsegüentment mantenir, els nivells de ferro dins dela normalitat.
L’estudi inicial de la relació cost (expressat en euros)-eficàcia (expressat en anys de vida guanyats) del’anàlisi genètica molecular indicava que l’estratègia decribrar tota la població de 35-40 anys i de salvar videsés dominant respecte de l’estratègia de no fer res,sempre que el cost de l’anàlisi genètica no superi els10 euros. No obstant això, aquestes dades van sercalculades inicialment basades en publicacions
internacionals i assumint una penetrància clínica deles mutacions que causen hemocromatosi del 50%(20% en dones i 80% en homes). Cal remarcar queestudis recents indiquen la penetrància de lesmutacions del gen HFE podria ser molt més baixa quela inicialment estimada. Per tant, és previsible que elcost del cribratge haurà de baixar molt més per assoliruna relació cost-eficàcia dominant. Des del punt devista tècnic, es podrien assolir costos almenys quant amagnituds inferiors per mitjà de tècniques d’espectro-metria de masses o d’altres tècniques de genotipat agran escala. Previsiblement, aquests resultats espublicaran en una revista internacional aquest any(Oliva et al., 2004). Aquests resultats haurien de servirper ajudar a planificar les necessitats del nostresistema sanitari.
També de rellevància pràctica immediata ha estat ladetecció d’una edat d’inici més precoç de la diabetistipus 2 en els pacients amb els genotips heterozigotscompostos (C282Y/H63D), heterozigots C282Y, ohomozigots C282Y, i de la presència d’una majorincidència de retinopatia o de nefropatia diabètica(Oliva et al., 2004. The HFE gene is associated to anearlier age of onset and to the presence of diabeticnephropathy in diabetes mellitus type 2. Sotmès).L’aplicació clínica més immediata d’aquesta troballaés que cal fer un seguiment més detallat dels pacientsque iniciïn una diabetis tipus 2 i que tinguin genotipsde risc (C282Y/H63D, heterozigots C282Y, o homozi-gots C282Y).
A part de tots aquests resultats potencialment aplica-bles a curt termini, amb el projecte també hem contri-buït substancialment a l’avenç dels aspectes derecerca bàsica. Així, hem completat la seqüència delgen HFE humà i hem descobert i descrit diversesnoves variants d’splicing (Sánchez et al., 2001).Aquests avenços poden tenir aplicacions a mitjà i llargtermini en estimular el desenvolupament de tracta-ments més eficaços per a les alteracions del metabo-lisme del ferro.
4. PublicacionsSánchez M, Bruguera M, Quintero E, Barrio Y, Mazzara R,
Rodés J and Oliva R (2000). Hereditary Hemochromatosis in
Spain. Genetic Testing, 4, 171-176.
Sánchez M, Bruguera M, Rodés J and Oliva R (2001)
Complete characterization of the 3' region of the human and
mouse hereditary hemochromatosis HFE gene and detection of
novel splicing forms. Blood Cells, Molecules, and Diseases 27,
35-43
Sánchez, M.; Villa, M.; Ingelmo, M.; Sanz, C.; Bruguera, M.;
Ascaso, C.; Oliva, R. (2003). Population screening for hemochro-
matosis: a study in 5370 Spanish blood donors. Journal of
Hepatology 38, 745-750.
Oliva R, Novials A, Sánchez M, Villa M, Ingelmo M, Recasens
M, Ascaso C, Bruguera M, and Gomis R (2004). The HFE gene is
associated to an earlier age of onset and to the presence of
diabetic nephropathy in diabetes mellitus type 2. (sotmès).
Oliva R, Pereira A, Rodés J, Bruguera M (2004)
Cost-effectiveness analysis for population screening for hemoch-
romatosis in Spain. (article en preparació).
RECERCA BIOMÈDICA | 175

FUNDACIÓ LA MARATÓ DE TV3
MEMÒRIA FINAL
1. Resum del projecte originalObjectius específics1. Per analitzar la primera hipòtesi, que existeixenmecanismes fisiopatològics que incrementen laproducció d'LDL(-) en la diabetis:
Aïllar LDL(-) en pacients diabètics (tipus 1 i 2) i d’indivi-dus controls normolipèmics.
Analitzar la proporció plasmàtica, estructura icomposició d'LDL(-) dels pacients diabètics (tipus 1 itipus 2). Comparar aquestes característiques ambl'LDL nativa.
Comparar els resultats obtinguts d’LDL(-) i LDL nativadels pacients diabètics (tipus 1 i tipus 2) amb els d'LDL(-) i LDL nativa d'individus controls normolipèmics.
2. Per analitzar la segona hipòtesi, que les LDL(-) delspacients diabètics són proaterogèniques:
Avaluar les propietats biològiques d'LDL(-) de pacientsdiabètics (tipus 1 i tipus 2) que estiguin directamentrelacionades amb el desenvolupament de la lesió
ateromatosa. Comparar-les amb les d'LDL nativa.
Comparar les propietats biològiques d'LDL(-) i LDLnativa de pacients diabètics (tipus 1 i tipus 2) amb lesd'LDL(-) i LDL nativa d'individus controls, normolipèmics.
3. Per analitzar la tercera hipòtesi, que els mecanis-mes de producció d'LDL(-) poden ser diferents en ladiabetis tipus 1 i tipus 2:
Comparar l'estructura, composició i propietats biològi-ques d'LDL(-) dels pacients diabètics tipus 1 amb lesdels tipus 2. Comparar, així mateix, les mateixescaracterístiques de l'LDL nativa entre els pacientsdiabètics tipus 1 i 2.
4. Per analitzar la quarta hipòtesi, que l'optimitzaciódel control glucèmic pot influenciar els mecanismes deproducció d'LDL(-) en la diabetis:
Fer tots els estudis que pertoquin a diabètics tipus 1 itipus 2 en situació de mal i bon control glucèmic.Comparar els resultats obtinguts en ambduessituacions.
Títol del projecte
Caracterització fisicoquímica i funcional de la subfracció modificada,electronegativa de les lipoproteïnes de baixa densitat (LDL(-)) en la
diabetis mellitus. Influència del perfil glicèmic
Investigadors principalsJordi Ordóñez Llanos
InstitucióHospital de la Santa Creu i Sant Pau
108.588 €

DissenyLa mostra de pacients serà de 30 individus ambdiabetis mellitus tipus 1 i 30 tipus 2. La selecció delspacients es durà a terme d’acord amb les característi-ques següents:
a) Pacients amb diabetis tipus 1.
Tots seran individus de nou diagnòstic, sense teràpiaprèvia amb insulina, amb nivells d'hemoglobinaglicada (HbA1c) superiors al 9% i absència decetoacidosi. Es descartaran els pacients que seguei-xin teràpia hipolipemiant.
b) Pacients amb diabetis tipus 2.
Tots seran individus sense teràpia prèvia amb insulina iamb nivells d'HbA1c superiors al 9%. Es descartaranels pacients amb complicacions vasculars associadesa la diabetis i els que segueixin teràpia hipolipemiant.
c) Grup control d'individus sans.
Seran individus normolipèmics, normoglicèmics i nofumadors, tot constituït el grup, personal voluntari del'Hospital. Estarà integrat per dos subgrups ambcaracterístiques similars d'edat, sexe i índex de massacorporal als diabètics tipus 1 i 2.
Tipus d'estudiLongitudinal. Tots els pacients seran estudiats ensituació de mal control glicèmic i després de l’optimit-zació (2-4 mesos). D'aquesta manera es valoraràl'efecte d'hiperglicèmia sobre les característiques del'LDL(-).
Tipus d'intervencióTots els pacients amb diabetis mellitus tipus 1 seraninclosos en un programa de teràpia intensiva ambmúltiples dosis d'insulina. L'objectiu és assolir nivellsd’HbA1c normals o pròxims a la normalitat (<6%), icom a mínim dos punts per sota del valor inicial. Elspacients amb diabetis tipus 2 també seran tractatsamb insulina en una o més dosis, segons les caracte-rístiques del pacient. Els objectius són similars als delsdiabètics tipus 1.
El plasma obtingut serà congelat a -80° C. Unavegada optimitzat el control glicèmic, s'obtindrannoves mostres de plasma que també seran congela-des a -80° C fins a la realització de les anàlisispertinents. S'avaluarà l'estructura i interacció ambreceptors cel·lulars i amb cèl·lules endotelials del'LDL(-) de diabètics tipus 1 i tipus 2, tot comparantaquestes característiques amb les d'LDL nativad'aquests mateixos pacients i amb les d'LDL(-) inativa d'individus control. Els diabètics s'analitzaranen mal i bon control glicèmic. Les tècniques delaboratori inclouen la composició en lípids, apoproteï-nes, antioxidants i paràmetres d'oxidació de lesfraccions lipoproteïques, estudis d'afinitat al receptord’LDL en cultius de fibroblastos i, finalment, efectesobre la producció de molècules proinflamatòries comles quimioquines IL-8 i la MCP-1 per cultius decèlul·les endotelials.
2. ResultatsAssociació entre LDL(-), perfil lipidic i paràmetresde control glicèmicEn el treball adjunt (Sánchez-Quesada et al., J ClinEndocrinol Metab 2001;86:3243-3249) hemdemostrat que l’LDL(-) està augmentada en els dostipus de diabetis, però probablement mitjançantmecanismes diferents. Els resultats especificats (taules1-4 i figura 1) es mostren també en la publicació. Lesdades antropomètriques, els paràmetres de controlglicèmic i el perfil lipídic dels pacients diabètics i delsindividus controls es mostren a les taules 1 i 3. Lacomposició en lípids i proteïnes de les LDL delsdiversos grups es mostra a les taules 2 i 4. La figura 1mostra la relació entre LDL(-) i LDL glicada en els dosgrups de pacients DM abans i després de l’optimitza-ció del control glicèmic.
En el cas de la diabetis tipus 1, el control glicèmic téuna important relació amb els nivells elevats d’LDL(-),com es dedueix de la seva disminució després de lanormalització de la glicèmia i de la relació directa ambla proporció d’LDL glicada (figura 1). A més, nosembla que hi hagi relació amb els processos demodificació oxidativa, com indica la falta de canvis enla resistència a l’oxidació de l’LDL (Sánchez-Quesadaet al., Diabetologia 1996;39:1469-1476).
Per la seva banda, l’LDL(-) en la diabetis tipus 2 no esmodifica posteriorment a l’optimització del controlmetabòlic (taula 2 i figura 1), al contrari del quesucceeix amb l’LDL glicada (figura 1), tot indicant quela glicèmia no és rellevant en la generació d’LDL(-). Enaquest cas, els processos oxidatius semblen tenirmés importància, ja que l’LDL total de diabètics tipus2 és més susceptible a oxidar-se que la d’individussans (taula 2).
Caracterització fisicoquímica i biològica de l’LDL(-)Característiques físiquesCarrega elèctrica: la càrrega elèctrica es va determinarrutinàriament en cada experiment mitjançant electrofo-resi en agarosa, amb la finalitat de comprovar que laseparació cromatogràfica de les subfraccions d’LDLva funcionar correctament. Les fraccions LDL(-) vanpresentar més mobilitat electroforètica que les LDL(+),malgrat que no es van observar diferències entre elsdiversos grups estudiats (figura 2).
Grandària: la grandària de les fraccions d’LDL va serdeterminada mitjançant electroforesi en gradientd’acrilamida no desnaturalizant (GGE). A diferència delque s’observava en individus controls, en què l’LDL(-)presenta una grandària lleument inferior a l’LDL(+)(figura 3 i Sánchez-Quesada et al., J Lipid Res2002;43:699-705), la banda principal d’LDL(-) té unagrandària similar a la de l’LDL(+) en els dos grups depacients DM, i aquesta no es veu modificada peltractament amb insulina. No obstant això, hi hadiverses observacions remarcables. Primerament,mentre que les fraccions LDL de DM tipus 1 presen-ten un diàmetre similar a l’observat en els individuscontrols (diàmetre > 26 nm, que correspon a unfenotip A, no aterogènic), les LDL de DM tipus 2presenten un diàmetre igual o inferior a 25,5 nm, quecorrespon a un fenotip B d’elevat risc aterogènic. En
RECERCA BIOMÈDICA | 177

segon lloc, les LDL(-) dels dos grups DM en bon i malcontrol glicèmic presenten bandes d’elevat diàmetre(> 30 nm) que són causades per un important graud’agregació. Aquests resultats es presenten a la taula5 i a la figura 3a i 3b. Aquestes observacionssuggereixen que l’LDL(-) podria tenir un importantpaper aterogènic, ja que l’agregació de l’LDL és undels elements determinants de la seva retenció iacumulació a paret arterial.
Susceptibilitat a l’agregació: els resultats obtinguts al’apartat anterior referits al major nivell d’agregació del’LDL(-) van ser confirmats monitoritzant la susceptibili-tat a l’agregació per agitació intensa a 680 nm, ja quetotes les LDL(-) es van agregar més fàcilment que noles seves respectives LDL(+) (figura 4).
Integritat física de l’apoB: es va comprovar rutinària-ment per mitjà d’electroforesi d’acrilamida en presèn-cia d’SDS (SDS-PAGE). No es van observar fragmen-tació de l’apoB ni presència de proteïnes contami-nants (figura 5).
Composició químicaLípids i apoproteïnes: les diferències entre LDL(+) iLDL(-) en els dos grups de diabètics són semblants ales trobades en altres grups d'individus, com normoli-pèmics o hipercolesterolèmics. Bàsicament, l’LDL(-) téun contingut augmentat de triglicèrid, apoE, apoC-III,àcid siàlic i àcids grassos no esterificats (NEFA) quanes compara amb l’LDL(+) (taula 6). D’altra banda, elsDM tipus 2 presenten en l’LDL(-) un menor continguten colesterol total que els DM tipus 1 (fet que tambées correspon amb la menor grandària de la partícula),com també menor contingut en NEFA que l’LDL(-) delgrup control (taula 6). La millora del control glicèmicno exerceix modificacions significatives en la composi-ció de les LDL.
Paràmetres oxidatiusAntioxidants: pel que fa a les diferències entre diabetistipus 1 i tipus 2 i el grup control, els DM tipus 2 tenenmenys contingut d’antioxidants en les dues fraccionsd’LDL que els DM tipus 1 i els controls. No hi hadiferències entre LDL(+) i LDL(-) en cap dels grups nies va observar efecte del control glicèmic (taula 7).
Aldehids: el malondialdehid (MDA) està lleugeramentaugmentat en els dos grups de DM respecte del grupcontrol, però sense assolir diferències significatives.No hi ha diferències entre LDL(+) i LDL(-) en cap delsgrups ni s’observà efecte del control glicèmic (taula 7).Altres aldehids característics de l’LDL oxidada, com el4-hidroxinonenal o el 4-hidroxihexenal, no forendetectables, fet que indica que en general el nivelld’oxidació de les LDL és molt baix.
Susceptibilitat a l’oxidació: en tots els grups les LDL(-)foren més resistents a oxidar-se que les LDL(+), comindica la fase de latència de la cinètica de formació dediens conjugats (lag phase) (taula 7). Respecte de lesdiferències entre grups, els DM tipus 1 es vancomportar de manera similar al grup control, mentreque les dues fraccions d’LDL dels DM tipus 2 erenmés susceptibles a l’oxidació que els controls.Aquests resultats coincideixen amb el menor contin-
gut en antioxidants de les LDL dels DM tipus 2. No esva observar cap efecte del control glicèmic.
Activitat PAF-AH: la determinació de l’activitat PAF-AHno estava contemplada en la memòria original delprojecte, però resultats recents obtinguts pel nostre grup(Benítez et al., Circulation 2003;108:92-96) indiquen queaquest enzim s’associa preferentment a l’LDL(-), fet pelqual es va quantificar aquesta activitat enzimàtica en lesmostres de pacients DM. L’activitat PAF-AH era entre 5 i6 vegades més alta en les fraccions LDL(-) respecte del’LDL(+) (taula 7). Els resultats d’activitat van ser corrobo-rats mitjançant Western blot (figura 6). No es van trobardiferències significatives entre els diversos grups.Respecte del control glicèmic, l’activitat PAF-AH vadisminuir significativament en els DM tipus 1 però no enels tipus 2 (taula 7).
Paràmetres de glicèmiaPercentatge d’LDL glicada: no es van observardiferències en la proporció d’LDL glicada entre lesfraccions LDL(+) i LDL(-) en els tres grups estudiats(taula 8). Ambdós grups de diabètics van presentarmés proporció d’LDL glicada que el grup control iaquesta proporció va disminuir després de l’optimitza-ció del control glicèmic en les fraccions LDL(+), sensecanvis en les fraccions LDL(-).
Fructosamina: la determinació de fructosamina (mètodede Roche Diagnostics) en les fraccions d’LDL va oferirresultats similars als obtinguts amb la determinació delpercentatge d’LDL glicada (taula 8), malgrat que, acausa de treballar a prop del límit de sensibilitat delmètode, originalment dissenyat per treballar amb sèrumo plasma total, les diferències significatives observadesno van coincidir plenament amb les obtingudes amb elmètode de la proporció de LDL glicada.
Determinació del contingut en monosacàrids: s’intentàadaptar un mètode comercial (BioRad, Monosacharidedetection kit) de determinació de monosacàrids basaten electroforesi i marcatge fluorescent. Tanmateix, elsdiversos intents per posar a punt el sistema van resultarinfructuosos i no aplicables a la quantificació demonosacàrids en lipoproteïnes. A causa de l’elevat preudel kit (800 euros aproximadament) i la manca degaranties d’obtenir uns resultats òptims, i desprésd’assajar amb dos kits diferents, vam desistir decontinuar explorant aquesta opció.
Interacció amb receptors cel·lulars d’LDLReceptor d’LDL (apoB/E): les LDL(-) de tots els grupsestudiats mostren una pèrdua d'afinitat pel receptord'LDL (rLDL) en cultius de fibroblastos, respecte deles fraccions LDL(+) (figura 7). En el cas dels DM tipus1, les dues fraccions presenten un afinitat extrema-ment baixa, similar a la mostrada per l’LDL(-) del grupcontrol. Després del tractament amb insulina, l’LDL(-)va augmentar lleument l’afinitat per l’rLDL, sensecanvis en la fracció LDL(+). Respecte dels DM tipus 2,l’LDL(-) va presentar una baixa afinitat per l’rLDL,similar o fins i tot menor que la de l’LDL(-) de controls,mentre que l’LDL(+) va presentar una afinitat intermè-dia entre l’LDL(+) i l’LDL(-) del grup control. No es vanobservar diferències després de l’optimització delcontrol glicèmic.
FUNDACIÓ LA MARATÓ DE TV3

Receptor Scavenger tipus A (SRA): l’acumulaciód’esters de colesterol mediada per l’SRA enmacròfags P388D1, no va mostrar diferències signifi-catives entre les dues fraccions d’LDL, ni entre elsdiversos grups d’individus (dades no mostrades).
Producció de molècules inflamatòries en cèl·lulesendotelialsCitotoxicitat: l’observació directa per mitjà de micros-còpia òptica, com també els resultats de l’LDH (viabili-tat superior al 90% en tots els casos), indiquen que enles nostres condicions de cultiu les dues fraccionsd’LDL no produeixen toxicitat cel·lular. Aquestaabsència de toxicitat és important, ja que el nostreinterès és demostrar que l’LDL(-) pot induir en lescèl·lules endotelials (HUVEC) la producció de molècu-les inflamatòries (com s’indica en els apartatssegüents) mitjançant mecanismes específics no lligatsa processos de necrosi i/o apoptosi.
Producció de quimioquines: les LDL(-) de tots elsgrups estudiats mostren capacitat per induir l'allibera-ment d’IL-8 i MCP-1 per cultius de cèl·lules endotelials(figura 8). No es va observar capacitat per induir laproducció de PAI-1 (dades no mostrades). Nos'observen diferències significatives entre els dosgrups de diabètics. Respecte del control glicèmic,l’única diferència que es pot observar (per bé quesense assolir significació estadística, P<0.06) és unadisminució en l’alliberament d’MCP-1 en l’LDL(-) deDM tipus 1; aquesta diferència podria estar relaciona-da amb la disminució de PAF-AH, ja que el nostregrup té indicis que suggereixen que la PAF-AH potestar implicada en la inducció de quimioquines permitjà de l’LDL(-).
Conclusions1. La proporció d’LDL(-) està augmentada en els dostipus de diabetis. Tanmateix, mentre que en la DMtipus 1 l’optimització del control glicèmic produeix unadisminució d’aquesta proporció, en la DM tipus 2 nos’observa cap efecte del tractament amb insulina.
2. Malgrat que el control glicèmic fa disminuir laproporció d’LDL(-) en la DM tipus 1, el mecanisme nosembla ser un efecte directe de la glicació no enzimà-tica. Aquest raonament es fonamenta en duesobservacions: a) l’LDL(-) de DM no presenta més nivellde glicació que l’LDL(+) i b) l’LDL dels DM tipus 2estaria sotmesa als mateixos processos de glicació deles LDL dels tipus 1, però, tanmateix la proporciód’LDL(-) no es veu modificada per la millora delcontrol glicèmic.
3. La caracterització fisicoquímica de l’LDL(-) en elsdos tipus de diabetis mostra que les diferènciesrespecte de l’LDL(+) són similars a les observades enl’LDL(-) d’individus controls, és a dir, contingutaugmentat de triglicèrid, NEFA, àcid siàlic, apoB,apoE, apoC-III, PAF-AH, més susceptibilitat a l’agre-gació i més resistència a l’oxidació. No es vanobservar evidències de modificació oxidativa entreLDL(+) i LDL(-).
4. La major susceptibilitat a l’oxidació de les duesfraccions d’LDL de DM tipus 2, com també el menor
contingut en antioxidants d’aquests individus coinci-deix amb els resultats generalment descrits a la biblio-grafia.
5. Diverses característiques esmentades en l’apartat 3s’han associat a un major risc cardiovascular, incloent-hi el contingut augmentat en triglicèrid, NEFA, apoC-III, apoE, PAF-AH i susceptibilitat a l’agregació.
6. La pobra afinitat pel receptor d’LDL de les duesfraccions d’LDL en els dos grups de DM suggereixenun mecanisme addicional que contribueix a l’augmentde risc cardiovascular a la diabetis.
7. Igualment, la major proporció d’LDL(-) en els dostipus de diabetis pot implicar un elevat potencialinflamatori per mitjà de la inducció de la producció dequimioquines relacionades amb els events inicials dereclutament de leucòcits durant el desenvolupamentde les lesions arterioscleròtiques.
3. Rellevància i possibles implicacions clíniquesdels resultats finals obtingutsEls resultats obtinguts suposen un aprofundiment enels mecanismes proaterogènics de l’LDL(-) enpacients diabètics. D’una banda, es confirma que unasèrie de característiques aterogèniques presents enl’LDL(-) d’individus normolipèmics i hipercolesterolè-mics (de Castellarnau et al., Arterioscler Thromb VascBiol 2000;20:2281-2287; Sánchez-Quesada et al., JLipid Res 2002;43:699-705; Sánchez-Quesada et al.,Atherosclerosis 2003;166:261-270) també estanpresents en l’LDL(-) dels pacients diabètics.
D’altra banda, la similitud entre les característiquesaterogèniques observades entre els diversos grupsd’individus suggereixen que una de les nostres hipòte-sis inicials d’un origen diferent per a la formaciód’LDL(-) pot ser errònia. En aquest sentit, l’observaciód’una activitat PAF-AH augmentada en l’LDL(-) de totsels grups (individus normolipèmics, hipercolesterolè-mics i diabètics tipus 1 i tipus 2) (Benítez et al.,Circulation 2003;108:92-96) apunta cap a una relaciódirecta entre l’existència de processos inflamatoriscrònics, subjacents a l’arteriosclerosi, i un augment dela proporció d’LDL(-). En aquest sentit, cal remarcarque la PAF-AH és un enzim que augmenta la sevaexpressió en resposta a processos inflamatoris i quela seva concentració plasmàtica s’ha associat a unincrement del risc cardiovascular (Packard et al., NEngl J Med 2000;343:1148-1155).
Respecte de les implicacions clíniques que es podendesprendre dels resultats obtinguts, òbviament l’apli-cació d’aquests coneixements al pacient diabèticencara està llunyana. Tanmateix, s’obren possiblesestratègies d’intervenció a mitjà termini, gràcies alpossible ús d’inhibidors de la PAF-AH com a fàrmacsantiaterogènics, tal com alguns grups ja estanexplorant (Leach et al., Farmaco 2001;56:45-50). Enaquest sentit, resultats preliminars obtinguts al nostrelaboratori indiquen que la inhibició de la PAF-AHelimina la capacitat de l’LDL(-) d’induir l’expressió dequimioquines proinflamatòries.
RECERCA BIOMÈDICA | 179

4. PublicacionsJosé Luis Sánchez-Quesada, Antonio Pérez, Assumpta Caixàs,
Mecedes Rigla, Amalia Payés, Sònia Benítez, Jordi Ordóñez-
Llanos. Effect of glycemic optimization on electronegative low
density lipoprotein in diabetes: relation to nonenzymatic glycosyla-
tion and oxidative modification. Journal of Clinical Endocrinology
and Metabolism 2001;86:3243-3249.
FUNDACIÓ LA MARATÓ DE TV3

RECERCA BIOMÈDICA | 181
MEMÒRIA FINAL
1. Resum del projecte originalNombroses evidències indiquen la participació delsistema d’activació del plasminogen, directament o béper disminució de la fribrinòlisi, en l’engruiximent deles membranes basals (BM) i l’augment de la matriuextracel·lular (ECM) trobats en aquestes patologies.L’augment dels esdeveniments microtrombòtics i unareducció de la fibrinòlisi pot, directament o bé indirec-ta, estimular la producció de factors de creixement ide l’ECM.
Objectiu principalAquest estudi es proposa de determinar quin és elpaper de l’activador del plasminogen del tipus tissular(t-PA), la urocinasa (u-PA), la plasmina i l’inhibidor delsactivadors del plasminogen tipus I (PAI-1) en aquestsprocessos patològics, per mitjà de l’ús d’animalsknockout (KO) per a alguns dels gens essencials en elsistema plasminogen/plasmina.
Iespecíficament:1) S’induirà la diabetis en ratolins KO per als gens t-PA, u-PA, dobles KO t-PA/u-PA, i en els KO per al gen
PAI-1, i també en ratolins controls wild type per mitjàd’injecció d’estreptozotocina. Les alteracions alsronyons dels diversos animals diabètics seran analit-zats després de diversos temps de malaltia.
2) S’analitzaran els efectes patològics de la induccióde diabetis.
1. Estudi de la fibrinòlisi mitjançant la determinaciódels nivells de t-Pa, u-PA i plasmina i la seva activitaten el plasma dels ratolins diabètics.
2. Determinació dels efectes morfològics i histològicsen ronyó mitjançant anàlisi histològica convencional id’expressió per immunohistoquímica de: col·lagen IV,fibronectina, t-PA, u-PA, PAI-1, fibrinogen/fibrina.
3. Anàlisi de l’expressió dels gens esmentats per RT-PCR semiquantitativa i/o Western bloting.
La nostra hipòtesi formula que en ratolins KO per al’activador del tipus tissular (t-PA), la inducció dediabetis i l’absència d’aquesta proteasa, crucial per ala degradació de proteïnes de la matriu extracel·lular,indueixen lesions en ronyons d’un grau més alt. En
Títol del projecte
Anàlisi del paper dels activadors del plasminogen (PAs) a la nefropatia diabètica. Establiment del model d'inducció experimental de diabetis en ratolins null per als PAs
Investigador principalRosanna Paciucci Barzanti
InstitucióHospital Universitari Vall d’Hebron
67.035 €

aquesta mateixa línia, els ratolins PAI-1 -/- podrienestar protegits contra aquestes lesions. Els resultatsobtinguts d’aquests experiments podrien plantejarfuturs tractaments amb PA recombinants i fer minvarla mortalitat de malalts diabètics amb aquestespatologies.
2. ResultatsEn aquesta memòria final es resumeix el treball dut aterme durant el projecte.
Inducció de diabetisS’ha induït la diabetis per mitjà d’una injecció únicaintraperitoneal d’estreptozotocina (STZ) en dosi de180 mg/kg de pes en ratolins KO i wild-type de lamateixa soca (C57BI/6). La inducció diabètica quehem obtingut ha variat entre un 20% i un 72%.Aquesta inducció s’ha confirmat al cap de 7 i 14 diesdesprés de la injecció d’STZ per anàlisi de la glicèmia.En la figura 1 es resumeix el nombre d’animals queformen part de l’estudi. Com es pot observar, hemobtingut entre 5 i 7 ratolins per soca de knockout, quehan suportat la malaltia durant temps diferents (aparei-xen ratolins amb períodes de malaltia de 30 dies i finsa 100 dies). L’obtenció d’un nombre limitat de ratolinsamb malaltia ha tingut com a causa diversos factors:per exemple, el fet que els animals malalts es vaginmorint i que sovint la decisió de sacrificar-los abansque mori (i es perdi) es contraposa a la de deixar-losobreviure durant més dies. Per això, una part delsratolins induïts i amb malaltia s’han perdut.
Anàlisi de la glicèmia en la soca de ratolins C57BI/6No es coneixen els valors de glicèmia en els animalsknockout per als gens t-PA, u-PA i PAI-1. Per això,hem analitzat la glicèmia en els diversos animalsknockout, en condicions normals o en dejuni amb unmediador portàtil de glucosa. En la figura 2 s’indiquenaquests valors (mg/dL) en ratolins de 2 mesos d’edat.S’ha emprat un mínim de 5 ratolins per soca deknockout en cada condició. Com s’observa en elpanell de l’esquerra, hi ha diferències clares entrel’estat en dejuni (starved) comparat amb l’estatalimentat (fed) en totes les soques estudiades. En elcas dels PAI-1-/-, aquestes diferències són mésacusades. En canvi, no s’han observat diferènciessignificatives comparant els diversos KO en un estatdeterminat, per exemple en "dejuni". En el panell de ladreta, s’ha analitzat l’estat de la glicèmia en diversosmoments del dia en animals alimentats. Com es potobservar, no hi ha diferències significatives quedepenguin del moment del dia en què es faci elmesurament de la glicèmia. Tampoc no s’han consta-tat diferències entre mascles i femelles durant el dia.Aquests resultats ens han permès determinar que elsvalors de glicèmia repetidament superiors a 200mg/dL en ratolins inoculats amb STZ alimentatspermeten identificar la inducció de diabetis.
Anàlisi immunohistoquímica de ronyons en parafinaHem estudiar l’expressió de fibrinogen/fibrina (Fbg),fibronectina (Fn). col·lagen I i col·lagen IV en ronyonsd’animals sans i amb malaltia emprant anticossosespecífics sobre talls de teixits parafinats. Els animalscontrols eren animals de la mateixa edat no tractatsamb STZ. Les figures 3-6 són imatges representatives
dels resultats obtinguts amb cada KO per a l’expres-sió de fibrinogen (Fbg) (figura 3 i 4) o de fibronectina(figura 5 i 6). Els animals controls C57BI/6 (WT) sansmostren una distribució esperada de la tinció, en laparet vascular i als espais intersticials sense acumula-cions de Fbg en glomèruls. En els animals diabètics(en la figura es mostra un diabètic amb 70 dies demalaltia), en alguns glomèruls la tinció per a Fbg ésmés intensa en comparació amb els animals sans. Enla mateixa figura, a la part inferior apareix la tinció pera Fbg en la soca u-PA-/-. En els animals sans espoden veure nivells de Fbg lleument més altscomparats amb els controls sans WT, tant en elsespais intersticials com en algun glomèrul. En elsanimals u-PA diabètics amb 54 dies de diabetis, tantla intensitat de la tinció com el nombre de glomèrulspositius augmenta, tot indicant que la manca d’u-PAprovoca un augment de la deposició de Fbg en ronyóquan apareix la diabetis.
En la figura 4 es mostren els resultats derivats de latinció per a Fbg en les soques t-Pa-/- (panell superior)i PAI-1-/- (panell inferior). En els animals t-PA-/- sans,tant la intensitat de la tinció com el nombre deglomèruls positius són més alts que en la soca controlWT. L’expressió de Fbg augmenta clarament en elsanimals diabètics, en els quals es concentra alsglomèruls i afecta la majoria. En canvi, en els animalsPAI-1-/-, l’expressió de Fbg és amb prou feinesdetectable, tant en els animals sans com en elsanimals diabètics.
En resum, en els animals sans knockout per a u-PA it-PA, però no els PAI-1-/-, s’observen nivells augmen-tats de Fbg. Aquestes diferències són més evidentsen els animals diabètics u-PA-/- i, sobretot, en els t-PA-/- diabètics.
Els resultats derivats de l’estudi de l’expressió de lafibronectina (Fn) en animals WT i u-PA-/- es mostrenen la figura 5. Com s’esperava, l’expressió d’Fn en elsratolins WT es detecta en la paret vascular i en lamembrana basal, a més d’una lleu tinció als espaisintersticials. No s’observen grans canvis en presènciade malaltia. En canvi, els ratolins u-PA-/- presentenuna intensitat de reacció més gran tant en l’estat sacom de diabetis. L’augment de tinció és més palès alsespais intersticials i menys en glomèruls.
En la figura 6 es mostren els resultats obtinguts per aFn en els animals t-PA-/- i PAI-1-/-. D’una manerasemblant al que s’observa per a la tinció de Fbg, enels animals t-PA-/- sans hi ha més acumulació d’Fntant en els espais intersticials com en glomèruls. Enanimals diabètics, aquestes acumulacions d’Fnaugmenten d’una manera més evident que en altressoques.
Els animals PAI-1-/- sans mostren una disminuciód’expressió d’Fn, i no es detecten canvis substancialsen els animals diabètics.
En resum, l’estudi de l’expressió de proteïnes de lamatriu extracel·lular com a fibrinogen i fibronectina enronyons de ratolins knockout, reflecteix l’acumulaciód’aquestes proteïnes sobretot en els ratolins t-PA-/-.
FUNDACIÓ LA MARATÓ DE TV3

La condició diabètica incrementa aquesta acumulació.Els ratolins PAI-1-/-, d’acord amb el que s’haviaformulat en la hipòtesi de treball inicial, presentennivells de Fbg i Fn inferiors respecte dels altresanimals.
Anàlisi dels nivells de cistatina C en la sangd’animals knockout, sans i diabèticsEstudis recents han demostrat que la cistatina C, unaproteïna bàsica de 13 kDa amb funció inhibidora deles cisteïnes proteases, es pot emprar com un noumarcador de la taxa d’infiltració glomerular (AmericanJ Kidney Disease, 36:205-207, 2000; Clin Chem48:699-707, 2002). Aquesta proteïna podria represen-tar un marcador endogen ideal per mesurar la GFR(glomerular filtration rate) perquè detecta reduccionssubclíniques en la GFR, ben al contrari d’altresmarcadors. Per avaluar si els ratolins KO sans idiabètics tenen lesions que ja afecten la funcionalitatrenal, hem determinat els nivells de cistatina C en lasang. Per això s’ha recollit plasma dels ratolinsindicats en la figura 1 i se n’han estudiat els nivells decistatina C seguint la tècnica d’immunofelometria. Enhumà, hem comprovat que els valors de cistatina C,que corresponen a nivells normals de creatinina, són2.2 mg/L i els valors en el cas d’una patologia són 3.1vegades més alts (6.95 mg/L):
CreatininaMg/dL 1.75 7.38
CystMg/L 2.22±0.05 6.95±0.11
No hi ha dades en la literatura que permetin conèixerel valor de la cistatina C en ratolins. Amb els nostresresultats hem esbrinat que els valors d’aquesta proteï-na en el plasma de ratolins C57BI/6 sans és aproxi-madament 46 µg/L (figura 7).
Els ratolins KO mostren valors una mica més alts (lesmitjanes varien de 60 a 75 µg/L, entre 1.3 i 1.7vegades més alts). En els ratolins t-PA Ko sans elsvalors són més alts (75 µg/L), com també en els PAI-1-/- diabètics (77 µg/L) respecte dels controls WT.
Aquests resultats, que s’han de comprovar, indicarienuna afectació, molt incipient, de la funcionalitat renalen els KO per a t-PA i PAI-1. Les alteracions histològi-ques observades microscòpicament per morfometria iper immunohistoquímica en els ratolins KO t-PA i PAI-1 sans i diabètics, també refermen una afectacióincipient de la funcionalitat del ronyó.
Análisis morfométrico de tejido de riñón enanimales KO con y sin diabetesEl análisis morfométrico indica que existen gradosvariables de aumento de la matriz mesangial yengrosamientos de membranas basales que afectande forma global o segmentaria a los glomérulos. Haybiopsias con lesiones prácticamente inaparentes,mientras que en otras se aprecia una afectación del100% de los glomérulos. La severidad de las lesioneses muy similar en las distintas biopsias de una misma
cepa. Las lesiones que se observan son totalmentecompatibles con nefropatía incipiente. En la figura 8 semuestran ejemplos de resultados obtenidos continciones con ácido tricrómico en los diferentes KO.Se aprecian en los ratones t-PA-/- lesiones incipientesen diabetes (aumento de la matriz mesangial yengrosamientos de membranas basales). Estaslesiones son visibles ya en los animales sanos de lamisma edad (5-6 meses). La figura 9 muestra unejemplo de tinción para los animales PAI-1-/-. Essorprendente constatar que mientras estos ratonesestán "protegidos" frente a la acumulación de proteí-nas como fibrina y fibronectina, como hemos observa-do anteriormente, no lo están para proteína tipocolágeno, evidenciado mediante la tinción con ácidotricrómico. En estos animales, el riñón, en condicionesbasales, es ya distinto al de los WT, porque presentaáreas de expansión mesangial de carácter segmenta-rio antes de inducir la diabetes experimental.
En las figuras 10-11 se resumen los datos del análisismorfométrico y se comparan las variables analizadas(diámetro glomérulo, superficie glomérulo, superficiemesangio, intersticio) por cada ratón KO y WT, con ysin enfermedad. Es evidente que existen alteracionesdel mesangio y del intersticio en los animales condiabetes vs. sus controles (animales sanos de lamisma edad) como muestra la figura 10, panelesinferiores. Para los KO de u-PA, no se observandiferencias significativas. Las lesiones del mesangio ymembrana basal mesangial y del intersticio son másintensas en los PAI-1 que WT y t-PA-/-. Se observanprocesos de fibrosis intersticial y del mesangio en losKO para PAI-1 y t-PA no diabéticos.
Resumiendo estos resultados, podríamos decir que:1) en la diabetes existen alteraciones significativas delmesangio e intersticio en la mayoría de los animalesKO analizados, y 2) los animales PAI-1 y t-PA presen-tan alteraciones antes de inducir la diabetes.
Estos resultados sugieren que los ratones KO paraPAI-1 y para t-PA pueden espontáneamente desarro-llar alteraciones en la morfología del glomérulo, conconsiguiente alteración de la funcionalidad renal. Enanimales de mayor edad, este fenómeno podríaacentuarse más. Para comprobar esta hipótesis,hemos analizado animales de 1 año de edad encomparación con animales jóvenes. Como se indicaen la figura 11, los animales de 1 año de edad KOpara PAI-1 y, sobre todo, los KO para t-PA presentanaumentos muy significativos del diámetro y superficiedel glomérulo, del mesangio y del intersticio.
Así, podemos concluir que los animales KO para t-PAy para PAI-1 desarrollan espontáneamente alteracio-nes en la morfología del riñón. Estas alteraciones soncompatibles con una disminución de la funcionalidadrenal, como también sugieren las observacionesanteriores (aumento de cistatina C y acúmulos defibrina y colágeno).
Actualmente estamos confirmando las diferenciasobservadas a nivel cuantitativo y por medio deinmunohistoquímica con anticuerpos específicos paracolágeno I, III y IV.
RECERCA BIOMÈDICA | 183

Estas últimas observaciones, inesperadas según lahipótesis inicial, no tienen una explicación directa.Además, no se han descrito en la literatura alteracio-nes de la función o alteraciones de la morfología delriñón en los animales KO PAI-1 o t-PA, por lo tantoestamos delante de un proceso no descrito previa-mente, de cuyo estudio esperamos obtener informa-ción válida para entender los mecanismos queintervienen en el establecimiento de la patología renalen los diabéticos.
3. Rellevància i possibles implicacions clíniquesdels resultats finals obtingutsAmb els resultats obtinguts, podem concloure que hiha diferències morfològiques en el ronyó dels animalsKO per als gens PAI-1 i t-PA. Les acumulacions deproteïnes de la matriu s’observen molt clarament enels ratolins més grans d’1 any d’edat sense malaltia,com també en els animals més joves amb diabetisinduïda. Aquestes alteracions suggereixen una afecta-ció incipient de la funció glomerular. L’augment delsnivells de cistatina C detectat en plasma dels KO pera t-PA i PAI-1 confirmaria aquestes observacions, totindicant que l’ablació dels gens t-PA i PAI-1 originafenòmens de reorganització de la matriu mesangialque pot dur a una afectació de la funció glomerular.
És important d’assenyalar que aquestes alteracions nos’havien descrit prèviament en aquests ratolins KO. Lafunció de t-PA per al manteniment de la morfologianormal glomerular sembla més rellevant que la d’u-PA.D’una manera sorprenent i contràriament a la hipòtesiformulada primerament, l’ablació de PAI-1 empitjora lapatologia del ronyó en la diabetis, tot provocantl’acumulació de matriu mesangial de composiciódiferent de la present en els KO de t-PA. En resum,aquests resultats suggereixen que la funció de t-Pa iPAI-1 són importants per a la funció glomerular normal.
Cal remarcar que el mesurament dels nivells decistatina C en plasma de ratolins amb patologiesrenals no s’ha descrit abans. Aquesta tècnica aprofitamolt poc volum de mostra i podria ser un marcadorútil també en aquells pacients en què un volum mésgran de mostra pot representar un problema.
FUNDACIÓ LA MARATÓ DE TV3

RECERCA BIOMÈDICA | 185
MEMÒRIA FINAL
1. Resum del projecte originalObjectiusL’objectiu global és examinar l’evolució de la reaccióde Maillard (glicació-glicoxidació) i la resposta biològi-ca contra aquesta durant el desenvolupament de lanefropatia diabètica, emprant diversos models experi-mentals.
Aquest objectiu general es desglossa en els objectiusespecífics següents:
1. Mesurar la concentració de marcadors de lareacció de Maillard i d’oxidació, com també dels seusprecursors, en teixits renals, plasma i orina en modelsde la nefropatia diabètica i de pacients amb insuficièn-cia renal derivada de diabetis, i també l’efecte dediverses modalitats terapèutiques en els paràmetresanteriors.
2. Identificar dianes proteiques de la reacció deMaillard i d’oxidació en models experimentals de lanefropatia diabètica.
3. Avaluar nous efectes biològics de la reacció deMaillard com a moduladors del risc de desenvolupa-ment de la nefropatia.
DissenyPer assolir els objectius del projecte, es va optar perl’ús d’un rang ampli de models. Així, les hipòtesis escontrastaren en cultius de línies cel·lulars endotelials(ECV304), entre d’altres, també en models animals dediabetis, com la induïda per estreptozotocina i la rataZucker (Lepr-/-; fa/fa) i finalment per l’anàlisi demostres (plasma, orina i dialitzat peritoneal) depacients amb nefropatia diabètica.
Per al mesurament de les concentracions demarcadors de la reacció de Mailllard i d’oxidació s’hanemprat tècniques cromatogràfiques d’alt rendiment —HPLC per a l’anàlisi dels precursors 3-desoxiglucoso-na (3-DG), glioxal (GO) i metilglioxal (MGO); GC/MSper a la quantificació dels marcadors d’oxidació,glicoxidació i lipoxidació. Per a la identificació dedianes proteiques de lesió oxidativa i glicoxidativa,s’han utilitzat electroforesi preparativa i mapatgepeptídic per MALDI, aplicant-hi també anàlisi perWestern blot amb diversos anticossos monoclonals i
Títol del projecte
Paper modulador de la resposta biològica a la reacció de Maillard en l'evolució de la nefropatia en un model experimental de diabetis
Investigador principalJoan Prat Corominas
InstitucióFacultat de Medicina. UdL
Membres de l’equipManel Portero, M. Cristina Ruiz, Sílvia Garcia, M. Josep Bellmunt i Reinald Pamplona.
52.840 €

policlonals anti-AGE generats com a resultat delprojecte. Finalment, per establir mecanismes demodulació de risc de desenvolupament de nefropatia,s’han introduït manipulacions farmacològiques(basades en antioxidants, IECA, ARA-II i quelantsd’agents dicarbonílics), genètiques (transgènesi deproteïnes desacoblants mitocondrials) i teràpies renalssubstitutives (diàlisi peritoneal).
2. ResultatsReacció de Maillard en models experimentals denefropatia diabètica: modulació experimental iterapèuticaLes evidències obtingudes per GC/MS van provar quela inducció de diabetis per estreptozotocina en rata(com a model de la diabetis tipus I) produïaincrements significatius en les concentracions defurosina proteica, indicador de fases inicials de lareacció, en citosols hepàtics i renals. Pel que fa almesurament de productes derivats de fases mésavançades com carboximetilisina (CML), carboxietilisi-na (CEL) i malondialdehid-lisina (MDAL) es demostràque l’efecte de la diabetis és heterogeni, en funció delteixit i fracció subcel·lular analitzats. Així, en mitocòn-dria hepàtica, però no en renal o cardíaca, tots elsmarcadors de glicoxidació (CML i CEL) i el de lipoxi-dació (MDAL) mostraven increments significatius ambla diabetis.
En aquest sentit, també es va avaluar l’efecte de ladiabetis en els nivells de marcadors d’oxidació directaproteica com els semialdehids glutàmic (GSA) iaminoadípic (AASA). Els resultats demostraren, perprimer cop, l’existència d’aquestes modificacions enfraccions mitocondrials de diversos teixits i que enmitocòndries de fetge, hi hagué una disminució en elsnivells d’oxidació per la diabetis, mentre que en les decor no. En clar contrast, les mitocòndries renals sí quedemostraren un increment en els nivells d’oxidació. Espot suggerir, d’aquests resultats, que la lesió oxidati-va, glicoxidativa i lipoxidativa proteiques depenen, enla mitocòndria, de factors específics de cada teixit. Enaquest sentit, es va determinar que la dotació enproteïna desacoblant 3 (UCP3), un dels mecanismesmediadors de la fuita mitocondrial de protons, potésser un dels moduladors d’aquests tipus de lesió,perquè la concentració dels seus marcadors augmen-tà de manera significativa en mitocondris de músculesquelètic de ratolins defectius genèticament enUCP3 respecte dels controls no modificats. Aquest fetes féu palès especialment en l’AASA i GSA. En canvi,la sobreexpressió d’UCP3 no va induir una menorproducció de lesió oxidativa.
En el context de la nefropatia de la diabetis tipus II, esvan obtenir mostres d’escorça renal d’un modelexperimental (rata de tipus Zucker, Lepr-/-; fa/fa).Aquests animals, amb un dèficit en el receptor deleptina, desenvolupen una patologia similar a ladiabetis de tipus II humana amb obesitat i proteïnúria,al marge de diversos trets anatomopatològics moltsemblants a la nefropatia diabètica humana. Enestudiar la concentració de marcadors derivats de lareacció de Maillard, d’oxidació i de lipoxidació respec-te de controls no obesos (ZuckerFa/fa), s’objectivà lapresència d’aquests productes, tot evidenciant que la
hiperglucèmia condueix a un increment significatiu enles concentracions de tots ells, especialment en GSA iMDAL. Aquests resultats són compatibles amb unincrement de l’estrès oxidatiu induït per la diabetis.Per tal com els àcids grassos poliinsaturats sónmolècules amb una alta susceptibilitat a l’oxidació, escomprovà que la diabetis indueix una disminució moltmarcada en l’índex de peroxidisabilitat i en el contin-gut en dobles enllaços membranaris, al marge decanvis molt marcats en àcids grassos concrets, comels derivats de la família dels n-6. Aquests canvispoden atribuir-se, principalment, als efectes del dèficitinsulínic sobre els enzims responsables de la síntesid’aquests àcids grassos (desaturases i elongases),malgrat que no es pot excloure un efecte de consumd’aquests per un increment de l’estrès oxidatiu.
Per aprofundir en el possible paper de la reacció deMaillard en el desenvolupament de la nefropatiadiabètica en humans, es mesuraren per HPLC lesconcentracions plasmàtiques de 3-desoxiglucosona(3-DG), glioxal (GO) i metilglioxal (MGO), compostosde baix pes molecular precursors de la formació deCML, CEL i altres productes de la reacció de Maillard.Així es va demostrar que la concentració plasmàticad’aquests compostos està fortament influenciada perla funció renal. Els pacients sotmesos a diàlisi (pertant, amb funció renal complementada terapèutica-ment), mostraren concentracions de 3-DG, GO i MGOsignificativament menors respecte de pacients ambinsuficiència renal no sotmesos a diàlisi. Entre altrestroballes d’aquesta població, destacaren els efectesde l’hàbit tabàquic, tot incrementant els nivells de GO iMGO; i del tipus de tractament substitutiu, atès queels pacients sotmesos a diàlisi peritoneal mostravenincrements significatius en els nivells de 3-DG i MGOrespecte dels pacients sotmesos a hemodiàlisi ambfuncions renals residuals equivalents.
En concordança amb aquestes dades, es palesà queels pacients sotmesos a diàlisi peritoneal mostrennivells de lesió oxidativa significativament superiors alspacients en hemodiàlisi. Per aprofundir aquestsaspectes, es va avaluar per espectrometria de massesla presència dels intermediaris 3-DG, GO i MGO i elscanvis en les seves concentracions durant eldesenvolupament de la diàlisi peritoneal. Els resultatsdemostraren que la seva concentració en aquestmedis terapèutics fou marcadament superior a la quehom pot trobar al plasma. Això és pel processamenttèrmic emprat per a l’esterilització d’aquests líquids i lapresència de sals, emprades per al tamponament delpH, durant aquest procés, ja que en comparar-losamb les solucions denominades de doble cambra onels estabilitzadors de pH s’esterilitzen separadament,aquests exhibien unes concentracions de marcadorsderivats de 3-DG, GO i MGO significativamentinferiors. D’altra banda, aquestes troballes suggerei-xen l’absorció d’aquests intermediaris amb potencialsimplicacions negatives. En quantificar la lesió derivadade la reacció de Maillard i d’oxidació, s’ha objectivatque les proteïnes presents en el dialitzat d’aquestspacients mostren un elevat grau de modificació. Enconjunt, aquests resultats demostren que la formacióde productes derivats de la reacció de Maillard en lamembrana peritoneal, relacionables amb la insuficièn-
FUNDACIÓ LA MARATÓ DE TV3

cia de la diàlisi que es desenvolupa amb el temps enaquests pacients, pot tenir com a causa l’alta concen-tració dels precursors anteriorment esmentats.
En relació amb els productes derivats d’aquestsintermediaris i la influència de la diabetis en la sevaconcentració plasmàtica, la de pirrolina (un marcadorderivat de 3-DG) mostrà una relació significativa ambdiversos paràmetres de funcionalisme renal, enpacients amb insuficiència renal. Així, es demostrarencorrelacions significatives amb la concentració decreatinina, l’aclariment de creatinina, la durada de lanefropatia i la concentració de triacilglicèrids (unindicador de la dislipèmia diabètica).
En aquesta població també s’obtingueren dades querelacionen la lesió oxidativa no enzimàtica de proteïnesplasmàtiques i la funció renal en pacients ambdiabetis, tot reforçant els resultats dels models experi-mentals animals. Es demostrà un increment de la lesióoxidativa en dependència del grau d’insuficiènciarenal, mesurada per la creatininèmia i/o l’aclariment decreatinina. Així, també s’objectivà que la concentracióde marcadors de lesió oxidativa proteica en plasma ésmés alta en els pacients sotmesos a diàlisi que en elspacients (amb insuficiència renal) que encara nonecessitaven aquesta terapèutica. En avaluar lainfluència de malalties vasculars (en relació amb elpossible paper fisiopatològic de l’estrès oxidatiu en lanefropatia), es va percebre un increment en la lesióoxidativa en els pacients amb aquesta tipologia.
Així mateix, s’objectivà que els pacients sotmesos atractament amb inhibidors de l’enzim conversord’angiotensina II tenen nivells significativament inferiorsde lesió oxidativa en proteïnes plasmàtiques mesuratsper ELISA. Fruit d’aquesta troballa s’analitzà, en elmodel de rata Zucker, la variació induïda per diversosfàrmacs renoprotectors en la concentració demarcadors de reaccions no enzimàtiques en escorçarenal. El tractament amb un inhibidor de l’enzimconversor d’angiotensina posà de manifest que elsseus efectes respecte de la nefropatia diabètica vanaparentment més enllà que el control de la tensióarterial. Així, es demostrà que indueix de formadosidepenent, disminucions i fins i tot reversionscompletes en la concentració de marcadors d’oxida-ció proteica com GSA i de marcadors de glicoxidaciócom la CML. Val a dir, però, que els nivells d’altresmarcadors com AASA, MDAL i CEL no s’afectarentant com els anteriors, tot suggerint l’especificitat deformació d’aquests productes, i que, en aquest cas,les reaccions derivades de l’augment de la concentra-ció de glucosa cel·lular (CEL) no es veuen afectadesper vies dependents d’angiotensina. El bloqueig delreceptor d’angiotensina també dugué a disminucionssignificatives en les concentracions de GSA, CML iAASA proteics. Cal destacar que les dades obtingu-des amb CEL i MDA-Lys suggereixen que les viesdependents del receptor d’angiotensina AT1 per sesón més determinants per a la seva formació que lacirculació d’angiotensina II, atès que la reversióobtinguda amb el bloqueig del receptor no es donàamb inhibició de l’enzim conversor de l’angiotensina.Alternativament, hom podria suposar una funcióprotectora vers els efectes deleteris dels alts nivells de
glucosa intracel·lular mediada pel subtipus AT2 delreceptor d’angiotensina, no inhibit pel fàrmac emprat.
D’altra banda, en emprar un tractament experimental,l’AVE7688,un inhibidor de vasopeptidases, que afectatant l’enzim conversor de l’angiotensina com l’endo-peptidasa neutral, i que per tant potencia els efectesrenoprotectors de la bradiquinina, s’evidencià lareversió cap a la normalitat de les vies d’oxidació(GSA), de glicoxidació (CEL i CML) i de lipoxidació(MDAL), en la mesura que els nivells d’AASA nos’afectaren significativament. Aquesta situaciódivergeix clarament dels efectes d’aquests fàrmacs enla composició lipídica, on només s’observaren efectesparcials en determinats àcids grassos (18:2 i 20:4),sense retorn a la situació normal.
Identificació de dianes proteiques de la reacció deMaillard i d’oxidació en models experimentals denefropatia diabèticaEn el model de diabetis experimental induïda perestreptozotocina s’assolí el mesurament i la identifica-ció parcial de dianes d’estrès oxidatiu en fraccionssubcel·lulars. Així, les anàlisis de distribució de la lesióper Western blot en mitocòndria, la font cel·lular mésimportant de radicals lliures, permeteren observar quementre la diabetis no indueix augments generalitzatsen oxidació en proteïnes concretes en fetge (comsuggerien les anàlisis quantitatives per GC/MS), sí quehi ha increments específics en ronyó. Una de lesproteïnes lesionades de forma específica en mitocòn-dries renals fou identificada com la isoforma mitocon-drial de l’aspartat-aminotransferasa. Experiments invitro amb la forma citosòlica i la mitocondrial del’enzim demostraren que l’oxidació catalitzada permetalls condueix a una pèrdua de la seva activitat, ique la isoforma mitocondrial es més resistent a lainactivació oxidativa. També, els resultats de l’anàliside l’activitat in vivo demostraren que la diabetisinduïda per estreptozotocina s’associa, de formaespecífica per òrgan, a canvis marcats en les activitatsaspartat aminotransferasa citosòlica i mitocondrial.Així, en citosols hepàtics i renals, hi hagué unincrement de l’activitat (molt evident en el fetge) ques’estén també en fraccions mitocondrials hepàtiques icardíaques. Tot i això, cal destacar que en mitocondrisrenals, on s’identificà la lesió oxidativa de l’aspartataminotransferasa mitocondrial, es demostrà unapèrdua significativa d’aquesta activitat amb la diabetis.A causa de l’important paper d’aquest enzim en lallançadora de malat, un dels mecanismes de manteni-ment energètic cel·lular, es pot suggerir una nova viapatogènica d’actuació de la hiperglucèmia per aldesenvolupament de la nefropatia.
D’altra banda, en examinar el perfil de lesió oxidativapresent en les mostres d’escorça renal del modelZucker, es confirmaren les dades obtingudes perGC/MS, i es palesà un increment generalitzat de lacarbonilació de proteïnes (modificació oxidativa), queés especialment evident en una regió compresa entreels 50 i 75 kD.
Per tal de comparar aquest tipus de modificacióproteica amb la derivada de la reacció de Maillard, esvan realitzar Western blot de mostres similars amb
RECERCA BIOMÈDICA | 187

anti-CEL i un anticòs específic de modificacionsderivades de peroxidació lipídica. Els resultatsrefermaven un increment menys marcat de glicoxida-ció que de peroxidació lipídica i d’oxidació proteica.En el cas de la glicoxidació, els guanys es donen enbandes al voltant de 50 i 60 kD, mentre que en el casde peroxidació lipídica els guanys es donen en proteï-nes de pes aparent per sota de 50 kD i en proteïnesde més de 150 kD. Aquests fets suggereixen que elsdiversos processos derivats d’oxidació poden tenirdianes diferents.
La preparació d’anticossos policlonals contra produc-tes AGE va permetre, en conjunció amb l’aplicació demetodologies de proteòmica, la identificació de proteï-nes modificades per productes derivats de la reaccióde Maillard en homogenats tissulars, tot donant-se enproteïnes de diverses fraccions cel·lulars, incloent-hicitosol (catalasa i anhidrasa carbònica), mitocondri(piruvat carboxilasa, carbamil fosfat sintasa I) i d’altres,com peroxisomes i reticle endoplàsmic. Cal remarcartambé que la majoria de proteïnes identificadesposseeixen una vida mitja estimada de 30 h, totsuggerint-se que el procés de modificació es du aterme per precursors altament reactius, com GO iMGO. Es demostrà també que la modificació no ésexclusiva de proteïnes amb funció estructural (com lacadena pesant de miosina), sinó que hi ha tambéenzims modificats (sintasa d’àcids grassos, glucogenfosforilasa i transcetolasa, entre d’altres), essentdestacable el fet que un percentatge estimabled’aquests pertanyen al metabolisme de carbohidrats.
Avaluació de nous efectes biològics de la reaccióde Maillard com a moduladors del risc de desenvo-lupament de la nefropatia.Per avaluar si els alts nivells de precursors presents enpacients diabètics amb o sense nefropatia du aefectes biològics implicables en la patogènesi de lanefropatia diabètica, s’incubaren diverses líniescel·lulars amb altes concentracions de glucosa i/o deprecursors de la reacció de Maillard. Els resultatsdemostraren que les altes concentracions de glucosa(en increments relatius equivalents als presents in vivoper a diabetis) indueixen una pèrdua de capacitat deresposta a l’EGF, amb pèrdua de capacitat autofosfo-riladora i tirosina cinasa del receptor, de reclutamentd’intermediaris de senyalització amb mòduls SH2,com la proteïna SHC, tot suggerint que la modificacióinduïda per la glucosa està unida covalentment alreceptor. Aquest fenomen està relacionat ambincrements d’estrès oxidatiu cel·lular i no està induïtper canvis en pressió osmòtica. No s’evidenciaren,però, diferències en l’estat de fosforilació d’EGFR enaortes de rates amb diabetis tipus I induïda perestreptozotocina, ni en l’associació d’SHC, tot i existirmenys fosforilació d’una banda amb un pes aparentde 40 kD en les aortes dels models diabètics, ievidenciar-se una forma d’EGFR en aortes diabèti-ques amb diferent motilitat electroforètica, totsuggerint una modificació induïda per la diabetis en laproteïna.
La relació entre la hiperglucèmia i les complicacionscròniques de la diabetis, incloent-hi la nefropatia, potincloure alteracions en la senyalització cel·lular, com
evidencien els experiments precedents. En aquestalínia, es va hipotetitzar que els AGE poden interferirdirectament o bé modificar els receptors de factors decreixement. Així, en una sèrie d’experiments amb GO iMGO, es demostrà que ambdós compostos interferei-xen amb la capacitat de senyalització d’EGF mediatper EGFR, de forma temps i dosi dependent. Aixímateix, aquests precursors d’AGE inhibien el recluta-ment d’intermediaris de senyalització, com la fosfoli-pasa C i la senyalització ulterior per MAPK. A més, elGO i el MGO disminuïen de forma directa, enimmunoprecipitats d’EGFR i en fraccions membranalsenriquides en EGFR, la capacitat de respostad’aquest receptor al seu lligand. Addicionalment, elsprecursors d’AGE alteraven l’activitat fosfatasa defosfotirosina. Aquests efectes de GO i MGO podienatribuir-se a la formació d’entrecreuaments en l’EGFR,mediada per la reacció de Maillard, atès que un inhibi-dor d’aquesta va prevenir aquests fenòmens de formaparcial. Experiments addicionals demostraren que GOi MGO alteraven també la capacitat de resposta delPDGFR. Aquest fenomen s’associà, en termes demodificació proteica avaluada per espectrometria demasses, a un increment del derivat específic de GOen proteïnes, la CML. En conjunt, aquestes troballesdemostren un nou mecanisme patogènic de lescomplicacions cròniques diabètiques, basat en lamodificació de receptors amb activitat tirosina cinasaper intermediaris aldehídics generats per la reacció deMaillard secundària a la hiperglicèmia.
3. Rellevància i possibles implicacions clíniquesdels resultats finals obtingutsEn tractar-se d’un estudi de tipus bàsic, un delsaspectes més rellevants d’aquest treball ha estatcontribuir al coneixement de les vies patogèniquesbàsiques que relacionen la hiperglucèmia amb eldesenvolupament de la nefropatia. En aquest sentit,s’han proposat nous mecanismes, tot involucrant lareacció de Maillard i l’d’oxidació proteica: la inactivacióoxidativa de l’aspartat aminotransferasa mitocondrialrenal, entre d’altres, i la modulació de les vies senyalit-zades per receptors tirosina cinasa. Totes dues vieshan de permetre, quan se’n completi la recerca, ladefinició de paràmetres clínics d’aparició precoç en lamalaltia diabètica (com concentració plasmàticaelevada de precursors de la reacció de Maillard) quepuguin ésser emprats com a marcadors de complica-ció o del risc de patir-la, tot ajudant a la identificaciód’aquelles persones més susceptibles al desenvolupa-ment de la malaltia. També tenen una utilitat terapèuti-ca potencial, en haver establert punts de possibleintervenció farmacològica, per retardar la progressióde la nefropatia un cop instaurada. La rellevància delsegon dels mecanismes, a més, ultrapassa l’etiopato-gènia de la nefropatia, atès que podria intervenir en eldesenvolupament de l’anomenat "peu diabètic",primera causa d’amputació no traumàtica en la nostrasocietat, a través de la interferència amb senyalscel·lulars de creixement epitelial i vascular. Finalment,en el camp de la terapèutica, els resultats ajudaran aldesenvolupament de fàrmacs en fase experimental(inhibidors de la peptidasa) o a l’ús de fàrmacs jaemprats (ARA-II, IECA), d’una forma més racional. Aixímateix, els resultats amb diàlisi peritoneal són aplica-bles per a l’establiment de pautes d’instauració i
FUNDACIÓ LA MARATÓ DE TV3

millores en la formulació de les solucions de diàlisiencaminades a minimitzar els seus efectes secunda-ris, i millorar així la qualitat de vida d’aquestespersones.
4. PublicacionsRuiz MC, Portero-Otin M, Pamplona R, Requena JR, Prat J,
Lafarga MA, Borras M, Bellmunt MJ. Chemical and immunological
characterization of oxidative nonenzymatic protein modifications in
dialysis fluids. Perit Dial Int. 2003 23(1): 23-32.
Portero-Otin M, Pamplona R, Bellmunt MJ, Ruiz MC, Prat J,
Salvayre R, Negre-Salvayre A. Advanced glycation end product
precursors impair epidermal growth factor receptor signaling.
Diabetes. 2002 51(5): 535-42.
Brand MD, Pamplona R, Portero-Otin M, Requena JR,
Roebuck SJ, Buckingham JA, Clapham JC, Cadenas S. Oxidative
damage and phospholipid fatty acyl composition in skeletal
muscle mitochondria from mice underexpressing or overexpres-
sing uncoupling protein 3. Biochem J. 2002;368(Pt 2):597-603.
Pamplona R, Portero-Otin M, Bellmunt MJ, Gredilla R, Barja G.
Aging Increases Nepsilon-(Carboxymethyl)lysine and caloric
restriction decreases Nepsilon-(malondialdeyde)lysine in rat heart
mitochondrial proteins. Free Radical Res, 2002, 36(1), 47-54.
Ruiz MC, Portero-Otin M, Pamplona R, García S, Ayala V,
Bellmunt MJ, Prat J. Increased oxidative damage in kidney
mitochondria from diabetic rats: potencial role of aspartate-
aminotransferase in nephropathy development.
(sotmès 2004)
RECERCA BIOMÈDICA | 189

FUNDACIÓ LA MARATÓ DE TV3
MEMÒRIA FINAL
1. Resum del projecte originalObjectiu principalAnalitzar el paper del factor de transcripció p48 en elsprocessos de diferenciació i proliferació del pàncreesendocrí.
Hipòtesi de treball
p48, que està implicat en l'activació de l'expressió degens específics de cèl·lules acinars en l'adult, té unpaper en la proliferació dels precursors del pàncreesendocrí i exocrí, com també en la proliferació de lescèl·lules epitelials d'ambdós components del pàncrees.
Mètodes1) s'analitzarà en detall l'expressió de p48 en precur-sors pancreàtics i s'establiran models in vitro dediferenciació endocrina;
2) s'identificaran proteïnes capaces d'interaccionaramb p48 (interacció in vitro i dobles híbrids);
3) es caracteritzaran les rutes i molècules implicadesen els efectes de p48 sobre la proliferació cel·lular;
4) es generarà un ratolí knockout condicional de p48.
Resultats esperatss'espera determinar la capacitat de p48 per regular laproliferació i diferenciació de les cèl·lules pancreàti-ques endocrines.
Impacte potencial esperat1) un millor coneixement dels mecanismes queregulen el nombre i la funció de les cèl·lules beta alpàncrees,
2) l'establiment de noves vies per a la modulació de laproliferació i la diferenciació de les cèl·lules beta o delsseus precursors.
2. Resultats1. Analitzar el paper funcional de p48 en la diferen-ciació exocrina i endocrina
1.1 Analitzar l'expressió de p48 durant el desenvolu-pament precoç del pàncrees
Els estudis que hem dut a terme al nostre laboratoriutilitzant els anticossos de què disposàvem contra laproteïna p48 completa han posat de manifest una
Títol del projecte
Paper del factor de transcripció p48 en el desenvolupament, la proliferació i la diferenciació del pàncrees endocrí
Investigadors principalsFrancisco X. Real Arribas
InstitucióInstitut Municipal d'Investigació Mèdica
130.305 €

distribució inesperada i sorprenent: en embrions e9.5-e10.5, p48 es detecta preferentment en el citoplasmade les cèl·lules que constitueixen el primordi pancreà-tic. Dades similars han estat obtingudes en altreslaboratoris utilitzant la nostra preparació d'anticossos,com també anticossos antipèptid obtinguts en el grupde R. MacDonald. Més endavant, durant la transiciósecundària que té lloc cap al dia e13.5, p48 esdetecta en el nucli de les cèl·lules exocrines, com enel pàncrees adult. El conjunt d'aquestes troballessuggereix que p48 exerceix una funció diferent enaquesta etapa molt precoç del desenvolupamentpancreàtic. Calen estudis addicionals per confirmaraquesta troballa.
D'altra banda, també hem posat a punt els assaigsper a la detecció d'altres factors de transcripció imolècules implicades en diferenciació pancreàticaexocrina (Pbx-1, Mist-1, amilasa) i endocrina (Pdx1,Pax6, neuroD, insulina, nestina, glucagó, polipèptidpancreàtic) en teixit parafinat.
També hem dissenyat pèptids sintètics corresponentsals dominis N- i C-terminals de p48 per a la produccióde nous anticossos que facilitin la detecció de laproteïna al pàncrees embrionari. Malauradament,aquests anticossos no són tan efectius com els quehem generat contra la proteïna p48 sencera. Perobviar aquest problema, més recentment hempreparat les construccions per produir dues proteïnesde fusió corresponents als dominis N-terminal i C-terminal de p48, respectivament, sense el dominibHLH. També hem produït anticossos contra unpèptid sintètic corresponent a l'extrem N-terminal dep64, el partner de p48 al pàncrees exocrí; aquestsanticossos s'estan analitzant actualment.
1.2. Analitzar l'expressió de p48 en les cèl·lulesductals del pàncrees en un model experimental denesidioblastosi associat a diabetis.
Aquests estudis han estat portats a terme encol·laboració amb el grup del Dr. R. Gomis, del'Hospital Clínic. La posada a punt de la detecció dep48 en teixit de parafina ha permès dur a termedobles marcatges analitzant l'expressió de p48 i insuli-na. Així, hem posat en evidència l'existència d'unapoblació poc abundant, però molt claramentdemostrable, de cèl·lules que expressen p48 enl’àmbit nuclear i insulina en el citoplasma en els illotsde rates diabètiques per tractament neonatal ambestreptozotocina després del tractament amb tungstè.Aquests resultats suggereixen que durant la neogène-si d'illots es pot detectar una població cel·lular que ésp48+ insulina+ mai identificada en el pàncrees adult.Aquests resultats són importants perquè (juntamentamb la recent demostració pel grup de Chris Wrightque les cèl·lules endocrines provenen de precursorsp48+) suggereixen que aquestes podrien constituir lescèl·lules progenitores endocrines de l’adult.
1.3. Caracteritzar models in vitro per a l'estudi de lafunció de p48 en la transició exocrina/endocrina
Hem generat transfectants estables de p48 encèl·lules AR42J i hem demostrat que la proteïna p48
se sobreexpressa en aquestes cèl·lules, havent-hi unaugment dels nivells d'amilasa en les cèl·lulestransfectades en comparació amb els controls. Encanvi, no hem demostrat l'activació de l'expressió degens endocrins com ara la insulina.
Els experiments que havíem proposat utilitzant lescèl·lules MSL-G2 han estat descartats, ja que el Dr. O.Madsen, al laboratori del qual s'han aïllat i caracterit-zat les poblacions derivades d'aquestes cèl·lules, ensha indicat que es tracta d'un model relativament pocreproduïble en altres laboratoris com a conseqüènciad’exigència de sèrum fetal de vedella molt específicd'aquestes cèl·lules. D'altra banda, ell mateix ha utilit-zat els nostres anticossos anti-p48, com tambéassaigs d’RT-PCR per demostrar que p48 nos'expressa de forma espontània en cap de les líniesanalitzades.
1.4. Estudiar el paper de p48 en la regulació del'expressió i la funció de molècules d'adhesió en elpàncrees.
En aquest subobjectiu, hem analitzat la capacitat dep48 per regular l’activitat del promotor de l’E-cadheri-na en un panell de cèl·lules pancreàtiques utilitzanttransfeccions transitòries i assaigs del promotor.Aquests estudis han demostrat que, efectivament,p48 i neuroD poden regular l’activitat del promotor del’E-cadherina en cèl·lules pancreàtiques amficrines i/oductals. Aquests estudis s'han dut a terme encol·laboració amb la Dra. Amparo Cano (IIB/CSIC,Madrid).
D'altra banda, hem generat transfectants estables dep48 en cèl·lules RWP-1 amb fenotip ductal i hemanalitzat els canvis en el patró global d'expressiógènica associats a l'expressió de p48. L'anàlisi demicroarrays ha demostrat una inducció d'un augmentdels nivells d’RNA de diverses proteïnes implicades enadhesió cel·lular i en la funció del citoesquelet, quehan d'ésser confirmats utilitzant Q-RT-PCR. Caldestacar la claudina-4 (component de les unionsestretes). Altres transcrits han mostrat una disminuciódels seus nivells en els transfectants; cal destacar lacadherina EGF LAG3 i altres proteïnes implicades enel citoesquelet. La rellevància fisiològica d'aquestscanvis està essent adreçada actualment utilizantexperiments de transfecció de p48 en altres líniescel·lulars i experiments de knock down utilizant siRNA.A continuació es mostra una taula amb els resultatsmés rellevants de l'anàlisi de microarrays.
Taula 1. Canvis d'expressió gènica més rellevants observats en
cèl·lules RWP-1 transfectades de forma estable amb el cDNA de
p48 (comparació amb els transfectants amb vector buit i anàlisi
als 15 dies de la transfecció).
RECERCA BIOMÈDICA | 191

2. Identificació de proteïnes que interaccionenamb p48En el decurs del darrer any del projecte hem analitzatla capacitat de p48 per interaccionar amb altresproteïnes que podrien contribuir al seu rol en ladiferenciació i proliferació pancreàtiques.Específicament, hem demostrat la interacció amb lesproteïnes p75/E2A i p64/HEB utilitzant anàlisi d’immu-noprecipitació i Western blotting, hem confirmat lainteracció descrita en l’assaig de doble híbrid ambRBPjK, tot fent-la extensible a cèl·lules pancreàtiques.Vam hipotetitzar que p48 podria interaccionar amb laproteïna pRb, atenent al seu paper en proliferaciócel·lular, malgrat que la seqüència primària no contéels motius LXCXL característics de proteïnes queinteraccionen amb Rb, com ara MyoD; en assaigsd’immunoprecipitació no hem pogut demostrar lainteracció in vivo d’ambdues proteïnes.
Els darrers mesos hem generat les construccions peral cribratge de doble híbrid d'interactors de p48 i hemidentificat cDNAs que estan essent seqüenciatsactualment. Esperem disposar d'aquesta informacióen el decurs dels propers 3 mesos.
3. Paper de p48 en la regulació de la proliferaciócel·lular3.1 Identificar les vies moleculars implicades en l'efec-te antiproliferatiu de p48
3.2 Identificar gens diana de p48 implicats en laregulació de la proliferació cel·lular
El conjunt de treballs realitzats en el decurs d’aquestprojecte ha permès demostrar l’efecte antiproliferatiude p48, que té lloc per mitjà d’una perturbació de laprogressió de la fase G1 a S del cicle cel·lular i queno exigeix de manera absoluta la funcionalitat d’Rb ip53, ja que els efectes antiproliferatius es podenpalesar en cèl·lules SaOs (que tenen Rb inactiu) i encèl·lules 3T3 p53-/-. D’altra banda, hem demostratque els clons transfectats amb p48 de forma establepresenten una sobreexpressió de p21CIP/KIP enl’RNA i proteïna, com també una sobreexpressió dep27 exclusivament a la proteïna. Aquestes dadesestan d’acord amb l’efecte de p48 d’augmentaraproximadament dues vegades l’activitat delpromotor de p21 sense que s’hagin evidenciatefectes sobre altres promotors d’inhibidors de Cdks,incloent-hi el de p27. Així, també s’ha demostrat queels clons transfectants de p48 presenten unadisminució dels nivells d’RNA de ciclina D2, però nod’altres ciclines, que podria contribuir al defecte deprogressió de G1 a S. Aquests canvis han estatconfirmats utilitzant RT-PCR quantitativa (SyBrgreen). Aquestes alteracions moleculars s’associenamb un augment, en els transfectants, de la formad’Rb hipofosforilada. S’han caracteritzat els dominisde la proteïna p48 implicats en aquests efectes, totdemostrant-se que és la regió C-terminal la respon-sable dels efectes antiproliferatius, sense que calguiel domini bHLH. Aquesta mateixa construcció té unefecte dominant negatiu quan s'utilitza en cotrans-fecció amb un plasmidi reporter amb el promotor del'elastasa. Per aquesta raó, és probable que elsefectes antiproliferatius de p48 no siguin directes
(gràcies a la seva capacitat de regulació transcripcio-nal), sinó indirectes.
La rellevància de l'efecte antiproliferatiu de p48 vindriarefermada per la troballa d'un augment del 50% de laproporció de cèl·lules Ki-67+ en les cèl·lules acinarsdels ratolins p48+/-. Aquestes troballes es fan extensi-bles als complexos ductals que típicament s'associenamb la pancreatitis crònica (tant en teixit humà comen models animals, com ara els ratolins MT-TGF alfa).
També hem dut a terme experiments per determinar sip48 és capaç d’induir apoptosi en cèl·lules pancreàti-ques (activació de caspasa-3, aparició d'un pic sub-G0 en citometria de flux), però no hem trobat capevidència a favor de la participació d’aquest mecanis-me en l’efecte inhibidor del creixement de p48.D'acord amb aquests resultats, no hem observatcanvis en els nivells d'apoptosi en el pàncrees deratolins p48+/- utilitzant la tècnica de TUNEL.
Els resultats de les anàlisis de canvis globals enl'expressió gènica s'han dut a terme com s'ha indicatanteriorment i els resultats més rellevants es mostrenen la taula 1.
4. Obtenció d'un ratolí knockout condicional delgen p48Aquesta part del projecte ha estat realitzada enestreta col·laboració amb el laboratori del Dr. PierreSavatier. Com ja s’havia esmentat en les memòriesanteriors, ens hem vist obligats a abandonar l’estratè-gia inicialment proposada per a la construcció delvector de recombinació homòloga perquè es produïade forma sistemàtica una deleció d’una part de laconstrucció durant la darrera etapa de lligament delsfragments que contenien les seqüències 5’ i 3’flanquejants del gen. Aquests experiments foren dutsa terme en els dos laboratoris implicats en el projecteutilitzant una àmplia varietat de mètodes per a lapurificació del DNA i per al lligament. Una discussióextensa amb el Dr. Savatier ens va portar a decidir dedesestimar aquesta estratègia, perquè vam intentareliminar el problema de recombinació durant el clonat-ge utilitzant múltiples mètodes en els dos laboratorissense èxit.
El projecte no ha estat completament desestimat.També en col·laboració amb el Dr. Savatier, estemintentant generar nous vectors que inclouen seqüèn-cies frt, en comptes de seqüències lox, per a larecombinació i escissió induïble del gen p48.L’estratègia que ara considerem òptima i que permetaprofitar tot el treball realitzat fins ara consisteix en l’úsd’un plasmidi diferent, el pFLN, que és més petit —facilitant el treball i potencialment disminuint laprobabilitat de deleció per recombinació— i que contéuna seqüència loxP i dues seqüències frt flanquejantel gen de resistència neo. Això permetria escindir elgen de resistència per transfecció de les cèl·lules ambFLP recombinasa sense afectar les seqüències loxP.En aquest vector es clonarien els mateixos fragmentsque hem preparat per a l’estratègia inicial: el 3’ flankdel gen de p48 i el 5’ flank-loxP/BspEI-p48.
Altres activitats no incloses en el projecte inicial
FUNDACIÓ LA MARATÓ DE TV3

Caracterització de les alteracions pancreàtiques en unmodel de diabetis associada a la inactivació dels gensE2F1 i E2F2En el decurs del darrer any del projecte hem dut aterme una col·laboració amb la Dra. Ana Zubiaga, dela Universitat del País Basc, per caracteritzar lesalteracions pancreàtiques en els ratolins KO per alsgens E2F1 i E2F2. Aquests ratolins presenten unadiabetes de l'adult que s'acompanya d'insuficiènciapancreàtica exocrina. Els ratolins moren als pocsmesos d’edat i la malaltia és més greu en els masclesque en les femelles. En aquest model es produeix unapèrdua ràpida de cèl·lules beta, com també unapèrdua, desestructuració i alteració displàstica delpàncrees exocrí. Aquestes alteracions s’acompanyend’un augment de la proliferació cel·lular i un augmentde l’apoptosi en les primeres setmanes després delnaixement que, finalment, comporten la pèrdua delteixit funcional. Aquestes evidències indiquen queE2F1 i E2F2 són necessaris per al manteniment del’homeòstasi funcional pancreàtica en l’adult. Aquestsratolins podrien constituir un bon model per utilitzarestratègies de transplantament de cèl·lules diferencia-des acinars i/o ductals.
Paper de la via de Notch en la diferenciaciópancreàticaEn un treball de col·laboració amb el grup del Dr. StevenLeach, hem contribuït a la demostració que l'activacióde la via de Notch està implicada en el mantenimentd'un pool de cèl·lules indiferenciades en el pàncrees ique aquesta ruta participa tant en la regulació de ladiferenciació endocrina com en la diferenciació exocrinautilitzant una combinació de models cel·lulars in vitro itransducció gènica en peix zebra.
Objectius específics assolits1. Demostració de l'expressió de p48 en el pàncreesembrionari abans de la diferenciació exocrina.
2. Identificació de l'existència d'una població decèl·lules que coexpressen p48 i insulina en models deneogènesi dels illots i de regeneració pancreàticapostadministració d'estreptozotozina i tungstè.
3. Identificació dels efectes de p48 sobre elspromotors de gens codificant proteïnes implicades enadhesió cel·lular i de canvis en l'expressió de proteïnesd'adhesió cel·lular en el model de cèl·lules RWP-1.
4. Identificació dels mecanismes moleculars pels qualsp48 indueix un efecte antiproliferatiu inhibidor delcreixement cel·lular.
5. Caracterització dels efectes de p48 sobre apoptosien cèl·lules pancreàtiques.
6. Identificació dels dominis de p48 implicats en elsseus efectes antiproliferatius.
7. Identificació de gens regulats (induïts i reprimits) perp48 en cèl·lules RWP-1 mitjançant microarrays.
8. Cribratge d'interactors de p48 utilitzant doblehíbrid.
9. Generació de nous anticossos contra p48 utilitzantpèptids sintètics i proteïnes de fusió-GST.
10. Identificació de polimorfismes genètics en p48 quepodrien estar associats a malaltia.
Modificacions de la metodologia i el pla de treballoriginal i justificació
Les modificacions que s’han incorporat en la metodo-logia i el pla de treball original han estat discutides enles memòries anteriors. La més important de totes esrefereix a l’estratègia per a la generació del ratolí KOde p48 induïble, objectiu que fins ara no ha estatpossible assolir. Les darreres discussions amb el Dr.Savatier, com també les darreres informacions sobreel paper de p48 com un gen important en l’establi-ment del destí pancreàtic durant el desenvolupament,ens fan refermar la idea que aquest és un objectiumolt important que hem de perseguir utilitzant novesestratègies experimentals. Per aquesta raó, mantin-drem els esforços amb la col·laboració amb el Dr.Savatier per assolir aquest objectiu. D’altra banda,hem mantingut converses amb el Dr. Ray MacDonald(Southwestern University, Dallas, EUA) per a l’obtenciód’un model induïble d’inactivació de p48 utilitzant unaestratègia similar a la que el seu grup ha descrit per algen Pdx1. En aquest cas, la regió codificant de Pdx1es va substituir pel cDNA del regulador Tet-off perknock in i l’expressió de Pdx1 es va aconseguir pertransgènesi sota el control del promotor de Pdx1induïble per doxiciclina. El Dr. MacDonald ha estatpersuadit de la importància de fer el mateix tipusd’experiment amb el gen p48 i aquest treball es realit-zarà en paral·lel en col·laboració amb nosaltres.
3. Rellevància i possibles implicacions clíniquesdels resultats finals obtinguts
Diverses evidències derivades del nostre treball id'altres investigadors posen de manifest la importàn-cia del gen p48 en patologia pancreàtica. D'unabanda, s'ha demostrat clarament que —tal comhaviem hipotetitzat— p48 s'expressa en els precur-sors pancreàtics i que totes les cèl·lules pancreàtiquesadultes es deriven d'aquests. D'altra banda, p48 ésnecessari per a la correcta especificació pancreàtica,d'acord amb els experiments del grup de ChrisWright. L'efecte antiproliferatiu de p48, demostrat perprimera vegada pel nostre grup en diversos tipuscel·lulars, posa de manifest que —almenys en elpàncrees adult— p48 podria contribuir d’una manerafonamental a l'homeòstasi tissular i que aquestsefectes tenen lloc fonamentalment per mecanismesque no exigeixen p53. Aquestes troballes podrien tenirimportància en la carcinogènesi pancreàtica. La nostrahipòtesi actual és que la pèrdua d'expressió de p48associada a l'aparició de complexos ductals eliminariaun mecanisme de regulació negativa del cicle cel·lularen el pàncrees exocrí, i faria les cèl·lules més suscep-tibles de progressar cap a l'adquisició d'un fenotipneoplàstic. Finalment, hem identificat diversospolimorfismes genètics en p48, la qual cosa permetràl'anàlisi del seu possible paper patogènic. Això,juntament amb la recent descripció que un locusresponsable de diabetis neonatal està a 10p12 —on
RECERCA BIOMÈDICA | 193

es localitza el gen p48— referma la idea que aquestgen pot constituir un factor de susceptibilitat heredità-ria a la diabetis.
4. PublicacionsSoria B, Skoudy A , Martin F. From stem cells to beta cells:
new strategies in cell therapy of diabetes mellitus. Diabetologia
2001; 44:407-415.
Skoudy A, Rovira M, Savatier P, Martín F, Leon-Quinto T, Soria
B, Real FX. Transforming growth factor beta, fibroblast growth
factor, and retinoid signalling pathways promote exocrine gene
expression in mouse embryonic stem cells. Biochem J 2004;
379:749-756.
Iglesias A, Murga M, Laresgoiti U, Skoudy A, Bernales I,
Fullaondo A, Moreno B, Lloreta J, Field SJ, Real FX, Zubiaga A.
Diabetes and exocrine pancreatic insufficiency in E2F1/E2F2
compound mutant mice reveals essential roles for these genes in
pancreatic growth and differentiation. J Clin Invest 2004 (en
premsa).
Esni F, Ghosh B, Biankin AV, Lin JW, Yu X, MacDonald RJ,
Civin CI, Real FX, Ball DW, Leach SD. Notch inhibits exocrine
differentiation in developing mouse and zebrafish pancreas.
Development 2004 (en premsa).
León-Quinto, Jones J, Skoudy A, Burcin M, Soria B.
Directed differentiation of mouse embryonic stem cells towards
insulin-producing cells. Diabetologia 2004 (en premsa).
Rodolosse A, Chalaux E, Skoudy A, Adell A, Hagège H, Real FX.
The pancreas-specific transcription factor p48 couples cell
differentiation to cell proliferation. Gastroenterology (en re-revisió).
Pujal J, Capellà G, Real FX. The Wnt pathway is active in a
subset of pancreatic cancers. (sotmès a publicació).
FUNDACIÓ LA MARATÓ DE TV3

RECERCA BIOMÈDICA | 195
MEMÒRIA FINAL
1. Resum del projecte originalLa hipòtesi experimental en què es basa el projecteés la següentLa dislipèmia diabètica és causada, com a mínimparcialment, per una funció anòmala de la lipoproteïnalipasa. Aquesta anomalia provoca un augment de lespartícules VLDL i LDL en la circulació, amb l’incrementconsegüent del risc aterogènic. La presència deformes al·lèliques de la lipoproteïna lipasa podria serun element determinant en la regulació del transport icaptació de lipoproteïnes, juntament amb el receptorde les lipoproteïnes de baixa densitat, en la paretarterial (transcitosi cel·lular), com també en la formacióde les cèl·lules escumoses (captació cel·lular) i enl’eliminació de romanents a nivell hepàtic (captacióhepàtica), és a dir, en aquells processos que estanimplicats en les primeres fases de l’arteriosclerosi.
Per verificar la hipòtesi anterior, ens plantegem elsobjectius bàsics següents1. Clarificar el paper de la lipoproteïna lipasa i delsreceptors de lipoproteïnes, especialment del receptorVLDL en la captació endotelial de lipoproteïnes,
determinant els mecanismes cel·lulars de la transcitosiendotelial. Determinar si els mecanismes implicats enla unió, internalització i transport de lipoproteïnesdurant el desenvolupament de l’arteriosclerosi sónsusceptibles de regulació.
2. Determinar els mecanismes cel·lulars implicats en launió i captació de lipoproteïnes per les cèl·lulesmusculars de l’íntima arterial.
3. Estudiar els efectes que provoca en el metabolismei en la captació de lipoproteïnes la presència deformes al·lèliques de la lipoproteïna lipasa que estroben amb una freqüència superior en la poblaciódiabètica i que s’associen amb una aterogenitat mésgran, fent èmfasi especial en els mecanismes detransport, secreció i recaptació de lipoproteïnes.
2. ResultatsLa descripció dels resultats aconseguits es fa per acadascun dels apartats del pla de treball.
A. Unió, endocitosi i/o transcitosi de lipoproteïnes encèl·lules endotelials i musculars llises. Paper de la LPL,i la seva aterogenicitat.
Títol del projecte
Lipoproteïna lipasa i receptor de les lipoproteïnes de baixa densitat i la seva relació amb la diabetis
Investigador principalManuel Reina del Pozo
InstitucióFacultat de Biologia. UB
77.459 €

La major quantitat de resultats en aquest apartatcorrespon al desenvolupament de nous sistemescel·lulars en cultiu corresponents tant a cèl·lulesendotelials com musculars llises, sempre d’origenhumà i que es mantenen al laboratori com a cultiusprimaris.
1. Transcitosi de la LPL i LDL en cèl·lules endote-lials. Estudi dels mecanismes cel·lularsResultats aconseguits:
Establiment de cultius de cèl·lules endotelials ambcèl·lules de llits vasculars diferents, els més importantsdels quals són els de les endotelials de cordó umbili-cal humà en cultiu primari, les cèl·lules de la líniaHMEC, endotelials de microvascultura de la pellhumana, i les cèl·lules de microvasculatura cerebralbovina.
Estandardització de l’obtenció de cultius primaris decèl·lules endotelials de cordó umbilical humà a partirde cordons. Aquest procés ens permet obtenir novescèl·lules, a passi molt baix, amb periodicitat setmanal.
Establiment de les condicions de creixement enmonocapa sobre dispositius Transwell i caracteritzaciódel model quant a la seva activitat de barrera(permeabilitat, flux, marcadors d’unions estretes, etc.).
Estudi de les cinètiques de pas de lipoproteïnesnormals (LDL nadiues) i modificades (oxidades,mínimament oxidades, electronegatives).
Estudis sobre la influència de la preincubació demonocapes cel·lulars de cèl·lules endotelials (especial-ment HUVEC i HMEC) amb lipoproteïnes diferents(LDL nadiues, oxidades, mínimament modificades) enles seves propietats de barrera, tant en les modifica-cions en el flux paracel·lular (mesurat per pas demarcadors com inulina i manitol) com en eltranscel·lular (mesurat per pas de peroxidasa). Elsefectes observats depenen de la situació d’estimula-ció de la monocapa cel·lular (amb histamina o trombi-na) i de la forma de lipoproteïna, tot incrementant-senotablement el pas en el cas de les formes oxidades.Les LDL estudiades (per sota de 200 m g apoB/mLsense preincubació o 100 m g/mL apoB/mL ambpreincubació de 24 h) incrementa la permeabilitat dela monocapa cel·lular comparada amb cèl·lules notractades, però no arriba als valors obtinguts amb untractament amb trombina. Tanmateix, altres fraccionsde lipoproteïnes (LDL(+), LDL(-), ox-LDL incremententant el transport paracel·lular com transcel·lular.
2. Paper del VLDL-R en l’endoteli. Està implicat enla transcitosi?Resultats aconseguits:
Clonatge de les dues formes del receptor humà deVLDL (VLDL-R+ i -).
Establiment de línies cel·lulars MDCK amb expressióestable de les dues isoformes del receptor de leslipoproteïnes de molt baixa densitat (VLDL-R).
Desenvolupament de nous anticossos contra el
receptor de les lipoproteïnes de molt baixa densitat,específicament contra dos pèptids sintètics correspo-nents a seqüències N i C-terminals.
Posada a punt del sistema ligand blotting per a ladeterminació de nivells de VLDL-R.
Mapatge d’expressió de les isoformes del receptor endiferents teixits humans i en diferents línies cel·lulars.
Determinació de la distribució intracel·lular delreceptor en línies cel·lulars MDCK amb expressióestable de les dues isoformes. Diferenciació endispositius de polarització. Inmunocitoquímica. Esconfirma una distribució específica en els diversosdominis extracel·lulars de les cèl·lules polaritzades.
Determinació de les propietats d’unió de lipoproteïnesnadiues (VLDL, LDL...) a superfícies cel·lulars delsclons MDCK amb expressió estable. Es detecta unamajor unió de VLDL a les cèl·lules amb expressió dela forma O-glucosilada. La unió de LDL és molt baixao inexistent, però ambdues formes de receptor podenunir i internalitzar VLDL. Aquestes lipoproteïnes sónconcentrades perinuclearment.
No s’ha detectat implicació del VLDL-R en la transci-tosi de lipoproteïnes en el model MDCK polaritzat enmonocapa sobre dispositius Transwells.
3. Mecanismes de retenció i captació de lipoproteï-nes per les cèl·lules musculars llises de l’íntimaarterial humanaResultats aconseguits:
Establiment de cultius de cèl·lules musculars lliseshumanes prominents de llits vasculars diferents: aorta,coronària, femoral, caròtida, vena uterina.
S’han realitzat estudis de cinètica de captació delipoproteïnes (LDL nadiues, oxidades, mínimamentmodificades).
S’ha estudiat la unió específica d’un factor de creixe-ment, el PDGF, en les seves diverses isoformes a lasuperfície cel·lular de cèl·lules musculars llises endiferents estadis de propagació, migració o extensió,tot relacionant-ho amb l’expressió de receptors debaixa afinitat però alta capacitat, com són els proteo-glicans d’heparan sulfat o condroïtinsulfat.
S’han clonat i caracteritzat dos proteoglicans de lafamília dels sindecans, concretament els sindecans 2 i3, tot analitzant-se la seva distribució cel·lular enteixits i en tipus cel·lulars i la seva influència en ladistribució del citoesquelet d’actina.
B. Estudi de la biologia cel·lular de les diversesformes al·lèliques de la LPL, la LH i apoB.Aquests estudis, clàssics al nostre laboratori, hancontinuat centrats en tres formes al·lèliques comunesde la lipoproteïna lipasa: Asp9, Asn291 y Stop 446.Durant la darrera anualitat no s’ha trobat cap novaforma mutant de la LPL, de la LH o de l’apoBdiferents de les ja conegudes.
FUNDACIÓ LA MARATÓ DE TV3

S’ha inclòs en aquest apartat l’estudi de dos gensque codifiquen per a proteoglicans del tipus heparansulfat, concretament de la família dels sindecans.Anteriorment, al laboratori s’havia clonat el sindecà 2humà. En aquest treball s’ha clonat i caracteritzat elsindecà 3 humà i es disposa de clons que contenenel sindecà 1 humà. El clonatge i caracterització delsindecà 3 ha estat publicat recentment (Berndt et al.[2001] J. Cellular Biochemistry 82: 246-259).
Resumint, el treball realitzat es concreta en:
1. Determinacions de mutacions en el gen LPL,possibles causants de la hiperlipèmia de tipus I
S’han determinat les mutacions causants d’hiperlipèmiade tipus I en un total de 12 famílies que presentenafectació d’hiperlipoproteïnèmia de tipus I. En aquestes,s’ha determinat la presència de la mutació Gly188à Gluen 11 i de Ile205à Ser en la darrera. S’han estudiat elsheterozigots en les diverses famílies.
Els resultats aconseguits són:Estudi del comportament cel·lular de diferents isofor-mes de la LPL amb alteracions del seu transportcel·lular. Han estat estudiades les isoformes de la LPLsegüents: G139E, G139Q, G142E, G142A, G142Q,A158T, S193R, N43A, G142E, S172C, K338St.
2. Estudi de les formes al·lèliques de la LPL i de laLH que presenten alterat el processamentintracel·lularHan estat estudiades les isoformes de la LPLsegüents: G139E, G139Q, G142E, G142A, G142Q,A158T, S193R, N43A, G142E, S172C, C418St,K428St, C438St, C418G, C438G, C418G/C438G,S447St. Les conclusions obtingudes són lessegüents:
i. Els residus 142 i 172 de la lipoproteïna lipasa sóncrítics per al manteniment de l’activitat catalítica del’enzim, com també per al seu transport intracel·lular.Canvis en el residu 172 provoquen la retencióintracel·lular al reticle endoplasmàtic i el seu direccio-nament cap a lisosomes. Un processament similars’ha observat en el cas de canvis G142A, G142E iG142Q.
ii. Canvis en el residu 139 (G139E, G139Q, G139 A)provoquen la impossibilitat d’adquisició de l’estructuradimèrica característica d’aquest enzim al reticleendoplasmàtic.
iii. Canvis en el residu 43 (N43A) són determinants pera l’activitat enzimàtica i la secreció normal de l’enzim.
iv. Una anàlisi del domini C-terminal amb mutants desubstitució i de truncament (C418St, K428St, C438St,C418G, C438G, C418G/C438G) mostra que malgratque la integritat del pont disulfur no és crítica per al’activitat de l’enzim, truncacions d’aquest dominiprodueixen l’agregació de la proteïna al reticleendoplasmàtic per exposició de regions hidrofòbiquesinternes de l’estructura de "b -barrel" del domini.
v. Tan sols hem analitzat dues mutacions que es
presenten com a polimorfismes en la població. És elcas dels canvis N291Si N447St. En el primer cas, laproteïna secretada amb normalitat té una activitatespecífica que és la meitat de la proteïna salvatge. Enel segon cas, la proteïna formada té una activitatenzimàtica superior a la salvatge.
3. Estudi de la biologia cel·lular de formes polimòr-fiques del pèptid senyal de la apoBEs van analitzar tres isoformes de l’apolipoproteïna B.Es va poder comprovar que les isoformes del pèptidsenyal de l’apolipoproteïna B presentaven petitesdiferències en la seva distribució intracel·lular mitjan-çant inmunocitoquímica. Posteriorment s’hademostrat que la isoforma ApoB-B24 (pèptid senyalde 24 aa) se secreta amb menys eficiència que laisoforma ApoB-B27. Aquestes diferències podrientenir com a causa un increment en el transportretrògrad.
4. Clonatge i caracterització del sindecà 3 humà,senyalització mediada per sindecansCom a receptors de lipoproteïnes de baixa afinitat ialta capacitat i de coreceptors, els proteoglicans desuperfície cel·lular tenen una gran importància. Allaboratori es va clonar el cDNA del sindecà de tipus 2,i en aquest projecte s’ha clonat i analitzat el sindecàde tipus 3. Les conclusions d’aquest estudi són lessegüents:
El sindecà de tipus 3 humà té una important homolo-gia de seqüència amb els descrits en altres espècies,malgrat que en el seu possible pèptid senyal (AA 7-23) s’observen fortes discrepàncies.
S’expressa fortament en cervell, melsa, còlon iglàndula adrenal.
La seva expressió temporal en sistemes cel·lularsindueix la formació de llargs filopodis and microspikesen un procés dependent de la presència de sucres enla cadena proteica.
Una anàlisi del paper de la cua citoplasmàtica mostraque la regió anomenada C1 és crítica per induir laformació de filopodis.
Una sobreexpressió estable de sindecà 3 truncat en laseva cua citoplasmàtica en sistemes cel·lulars redueixla capacitat proliferativa del cultiu, com també lesseves propietats d’adhesió i migració. Aquests efecteses relacionen amb l’augment de fibres d’estrès i decontactes focals, tot indicant possiblement unasenyalització mitjançant Rho GTPases i proteïnes de lafamília ERM.
En l’anàlisi de la funcionalitat i el paper intracel·lular delsindecà 2, s’han estudiat les seves interaccions ambel citoesquelet d’actina. Hem demostrat que elsindecà 2 interacciona directament amb la ezrina,proteïna de la família ERM, amb una KD de 0,71 m M,en una seqüència diferent a la prèviament descrita(DEGSYD).
RECERCA BIOMÈDICA | 197

3. Rellevància i possibles implicacions clíniquesdels resultats finals obtingutsAquest treball vol trobar evidències i explicacions de lahipertrigliceridèmia, que és l’alteració lipídica méscomuna en la diabetes mellitus tipus II o no insulino-depenent. Aquesta hipertrigliceridèmia ha estatproposada com una de les causes desencadenantsde la diabetes 2 (Mingrone et al., 1999, Diabetes 48 :1258; Yang et al., 2003, Hum. Mutat 21-453). És peraquest motiu que s’han estudiat en profunditat algunsdels mecanismes moleculars que regulen els nivells detriglicèrids circulants: activitat d’enzims hidrolítics comla lipoproteïna lipasa, activitat de receptors de baixa id’elevada afinitat com és el cas del VLDL-R i delsproteoglicans de tipus sindecà. La descripciód’aquests mecanismes i la caracterització dels detallsde funcionament moleculars ens permeten entendremillor els processos de determinació dels nivellscirculants de triglicèrids.
Quant a l’aplicació clínica dels resultats obtinguts, espot fer èmfasi en el paper de les variants de la lipopro-teïna lipasa. En diversos estudis ha estat descrit lamajor presència de formes al·lèliques de la lipoproteï-na lipasa en poblacions de pacients afectats dediabetis 2, especialment les isoformes d’A71T, V181I,N291S, G188E i I194T, G242L, F351L, S447 (Zhanget al. [1997] Clin Sci 93:335). L’explicació de lahipertrigliceridèmia en els heterozigots portadors decanvis en el gen de la lipoproteïna lipasa ésconseqüència o bé de l’absència d’activitat hidrolíticade triacilglicèrids (per exemple G188E) o de laformació de proteïnes que tenen capacitat de retencióde les lipoproteïnes a l’endoteli, però que hidrolitzenamb menor eficàcia (per exemple S291E) (Knudsen etal., 1997, Eur J. Clin Invest. 27 : 928). La presència devariants no actives de la LPL en heterozigosi en lapoblació podria suposar una resposta diferent ensituacions d’obesitat moderada i/o resistència a lainsulina que incrementés el risc de desenvolupamentde diabetis II i/o aterosclerosi.
4. PublicacionsChristine Berndt, Ricardo P. Casaroli-Marano, Senén Vilaró i
Manuel Reina (2001). Cloning and Characterization of Human
Syndecan 3. Journal of Cellular Biochemistry 82:246-259.
Contreras, H.R.; Fabre, M.; Franés, F.; Casaroli-Marano, R.P.;
Rocamora, N.; García-Herreros, A.; Reina, M. y Vilaró, S. (2001)
Syndecan expression in colorectal cancer derived HT29 M6
epithelial cells induces a migratory phenotype Biochem Biophys
Res Commun 286:742-751
Granés, F.; Berndt, Ch.; Roy, Ch.; Mangeat, P.; Reina, M. y
Vilaró, S. (2003) Identification of a novel Exrin binding site in
syndecan-2 cytoplasmic domain. FEBS Lett 547:212-216.
Sabater, D.; Benítez, S.; Magrane, J.; De Castellarnau, C.;
Vilaró, S.; Reina, M.; Casaroli-Marano, R.P. (2004) Two isoforms of
very low density lipoprotein receptor (VLDLr) mediate binding and
uptake of very low density lipoprotein (VLDL) in MDCK cells.
Atherosclerosis 5 (1):77 (abstract presentado en el 74tt Congress
of the European Atherosclerosis Society, 17-20 de abril de 2004,
Sevilla, España)
De Castellarnau, C.; Casalà, C.; Reina, M.; Casaroli-Marano,
R.P.; Benítez, S. y Vilaró, S. (2004) Effect of different lipoproteins
(LDLs) on endothelial barrier function. Atherosclerosis 5 (1):74
(abstract presentado en el 74tt Congress of the European
Atherosclerosis Society, 17-20 de abril de 2004, Sevilla, España)
FUNDACIÓ LA MARATÓ DE TV3

RECERCA BIOMÈDICA | 199
MEMÒRIA FINAL
1. Resum del projecte originalObjectiudeterminar si l’existència de variants genètiques de laregió crítica d’unió de les proteïnes TRAF al receptor ede l’FNT alfa (x1c) s’associen a una major susceptibili-tat genètica al desenvolupat de la DM2.
Dissenyanàlisi d’associació poblacional en un estudi transversal.
Àmbit de l’estudipoblació d’origen caucàsic mediterrani, atesa al’Hospital Joan XXIII de Tarragona.
Subjectes d’estudi330 pacients afectats de diabetis mellitus tipus 2 icontrols sans.
Instrumentacióamplificació de fragments d’ADN mitjançant PCR iseqüenciació directa del producte amplificat.Radioimmunoassaig. Enzimoimmunoassaig.
Resultats: regió "x1c" sense mutacions detectables enambdues poblacions. Anàlisi dels exons 10, 9, 8 i t(regió intracel·lular del receptor), sense mutacionsdetectables. Anàlisi de l’exó 6 (mutació 196T<C) i intró4 (CA repeat) associades a perfil lípid aterogènic.Mutació —174 G<C de la IL6 associada a majorprevalença de DM2.
Conclusionsla regió "x1c" no presenta mutacions. La mutació 196T<C de l’exó 6 s’associa a un perfil lípid aterogènic. Elgen de la IL6 pot participar en la susceptibilitat DM2en caucàsics.
2. ResultatsLa regió "x1c" del receptor del factor de necrositumoral alfa, no va mostrar mutacions detectables tanten la població control com en la població afecta dediabetis tipus 2. S’han analitzat els exons 10, 9, 8 i 7(regió intracel·lular del receptor) sense observarmutacions útils per a l’anàlisi de polimorfismesgenètics. Respecte de l’exó 6 (mutacions 196 T<C) iintró 4 (CA repeat), s’ha observat una associació entredeterminades variants polimòrfiques amb un perfillípidic proaterogènic.
Títol del projecte
Anàlisi mutacional del domini intracel·lular “x 1c” del gen del receptor 2 del factor de necrosi tumoral alfa en pacients amb diabetis mellitus tipus 2
Investigador principalCristóbal Richart Jurado
InstitucióHospital Universitari Joan XXIII. Tarragona
Membres de l’equipJoan Vendrell, Montserrat Broch, Cristina Gutiérrez, Immaculada Simón, Ana Megía, Lluís Gallart, Wifredo Ricarti José M. Fernández-Real.
73.647 €

Es va estudiar la participació de variants a la regió delpromotor del gen del factor de necrosi tumoral alfa(FNTalfa) en l’aparició de complicacions cardiovascu-lars entre la població amb diabetis tipus 2. Vamobservar que l’existència d’un al·lel en la posició —308 del promotor d’aquest gen s’associava a unamajor prevalença de cardiopatia isquèmica, preferent-ment entre les dones amb diabetis tipus 2.
Així mateix, vam avaluar el paper que podien tenir lesmutacions 196T>G exon6/ CA-repeat de l’intró 4 delgen del receptor 2 de l’FNT i els nivells plasmàtics deles fraccions solubles del receptor 1 i 2 de l’FNT en lapatogènesi de les complicacions microvasculars queapareixen en el context de la diabetis tipus 2. Vampoder observar que els pacients amb nefropatiadiabètica presentaven nivells més elevats en plasmad’ambdues fraccions solubles, però sense que lesmutacions estudiades predisposessin a un major riscgenètic.
Per últim, s’ha estudiat una mutació a la regió delpromotor del gen de la IL6 (mutació —174 G<C) en lanostra població de pacients amb diabetis tipus 2 is’ha trobat una associació entre els portadors delgenotip GG i la presència de diabetis. Aquestaassociació també es va veure en una població d’indisPima, naturals d’Arizona.
Conclusions: les nostres troballes donen suport al’existència d’una activació del sistema inflamatori enla patogènesi de la diabetis tipus 2 i de les complica-cions cròniques que se’n deriven, mediat per diversescitocines proinflamatòries, tant des del vessant sèriccom genètic.
3. Rellevància i possibles implicacions clíniquesdels resultats finals obtingutsEls resultats d’aquest projecte suposen un pas mésen el coneixement de les causes genètiques queintervenen en el procés de l’aparició de la diabetismellitus tipus 2. En un nivell pràctic no impliquen capconsideració, però sí que obren un nou camí per afuturs estudis d’associació entre els nous marcadorsdescrits, especialment la IL-6, en altres poblacions. Elcaràcter preliminar d’aquestes dades no justifica, demoment, un cribratge massiu de les mutacionstrobades entre la població de risc, per identificarpersones susceptibles, tenint en compte, a més amés, que es tracta d’una malaltia poligènica amb unaimportant interacció ambiental.
4. PublicacionsFernández-Real JM, Láinez B, Vendrell J, Rigla M, Castro A,
Peñarroja G, Broch M, Pérez A, Richart C, Engel P, Ricart W.
Shedding of TNF-alfpha receptors, blood pressure, and insulin
sensitivity in type 2 diabetes mellitus. Am J Physiol Endocrinol
Metab 2002; 282:E952-959
Vozarova B, Fernández-Real JM, Knowler WC, Gallart L,
Hanson RL, Gruber JD, Ricart W, Vendrell J, Richart C, Tataranni
PA, Wolford JK. The interleukin-6 (-174) G/C promoter polymor-
phism is associated with type-2 diabetes mellitus in Native
Americans and Caucasians. Hum Genet 2003;112:409-13
Vendrell J, Fernández-Real JM, Gutiérrez C, Zamora A, Simón
I, Bardají A, Ricart W, Richart C. A polymorphism in the promoter
of the tumor necrosis factor-alfpha gene (-308) is associated with
coronary heart disease in type 2 diabetic patients. Atherosclerosis
2003; 167:257-264
Vendrell J, Broch M, Fernández-Real JM, Gutiérrez C, Simón I,
Megia A, Gallart L, Ricart W, Richart C. Tumor necrosis factor
receptors (TNFRs) in type 2 diabetes. Analysis of soluble plasma
fractions and genetic variations of TNFR2 gene in a case-control
study. Diabetic Medicine 2004 (en premsa).
FUNDACIÓ LA MARATÓ DE TV3

RECERCA BIOMÈDICA | 201
MEMÒRIA FINAL
1. Resum del projecte originalObjectiu principalEstudiar els increments de la glucèmia (GLU) i triglicè-rids (TG) postprandials amb un reflectòmetre portàtilen persones amb xifres límit de TG basals entre 100 i200 mg/dl (TGL), amb o sense GLU alterada en dejú(GAD=110-125 mg/dl) o amb diabetis mellitus tipus 2(DM2) amb bon control glucèmic (HbA1c <8,0%), i encontrols sans amb GLU normal i TG <100 mg/dl.
Objectius secundaris1. Comprovar que és factible avaluar el perfil diürn deTG en sang capil·lar al domicili del pacient.
2. Definir quin moment durant l’estat postprandial ésel millor per detectar l’amplificació de TG.
3. Definir la normalitat del perfil diürn de GLU i TG enel grup control.
4. Relacionar el perfil diürn i el pic màxim de GLU i TGamb diversos marcadors de risc aterogènic i ateros-clerosi preclínica.
DissenyEstudi transversal de pacients ambulatoris i controls sans.
Àmbit de l’estudiConsultes externes de Clínica de Lípids en un hospitaluniversitari.
PacientsHomes (H) i dones (D) no fumadors, normocolestero-lèmics (cLDL<150 mg/dl) i amb pressió arterial normal(<140/90 mm Hg), distribuïts en 4 grups (n=18-20 pergrup): 1) control, 2) TGL, 3) TGL+GAA, 4) TGL+DM2.
MètodesExtracció de sang en dejú per a GLU, insulina,lipoproteïnes, àcids grassos lliures (AGL), vitamina E ales LDL, grau d’oxidació de les LDL (oxLDL) i prova detolerància oral a la glucosa amb determinació desensibilitat a la insulina (HOMA-S). Mesuramentsdomiciliaris amb reflectòmetre de GLU i TG a sangcapil·lar en dejú, a les 2, 4 i 6 h d’esmorzar i dinar, i al’hora d’anar al llit (8 determinacions en 16 h), undia/setmana x 2. Mesurament de la vasodilataciódependent (VDDE) i independent (VDIE) de l’endotelien l’artèria braquial i del gruix del complex íntima-
Títol del projecte
Estudi pilot de la lipèmia i glucèmia postprandials domiciliàries enpersones amb xifres de triglicèrids entre 100 i 200 mg/dl, amb i senseglucèmia alterada en dejú.
Investigador principalEmili Ros Rahola
InstitucióHospital Clínic i Provincial de Barcelona
Membres de l’equipJordi Gich, Daniel Zambón, Mercè Serra, Ramon Deulofeu, Berenice Cortés, Ingrid Spandau, Ana Pérez-Heras,Montserrat Portas, Maria Vela, Rosa Gilabert, María J. Ortega i Isabel Núñez.
23.621 €

mitjana (GIM) de la caròtida i de l’artèria femoralsuperficial per ecografia d’alta resolució en mode B.
ResultatsS’han estudiat 74 persones (40 H, 34 D, d’edats 27-76 anys): 20 controls, 18 TGL, 18 TGL+GAA i 18TGL+DM2. L’edat dels grups era similar, excepte en elgrup DM2, amb més edat. Els resultats demostrenque és factible efectuar mesuraments domiciliaris deTG en sang capil·lar. Les àrees sota la corba (ASC) de16 h de GLU i TG augmenten en la seqüènciaDM2>TGL+GAA>TGL>control (P<0,001). En tots elsgrups la màxima amplificació de GLU es registra a les24 h i la de TG a les 19 h (4-5 h després del dinar). Hiha una excel·lent correlació entre el pic de GLU il’ASC de GLU (r=0,90, P<0,001) i entre el pic de TG il’ASC de TG (r=0,935, P<0,001). Els límits alts delsperfils diürns de GLU i TG (P95 de l’ASC del grupcontrol) són fins a 1574 i 2438 mg/dl x 16 h, respecti-vament; els del pic de GLU i TG són fins a 115 i 232mg/dl. Els individus dels tres grups amb TGL tenen unperfil de TG patològic en >50% dels casos. Els AGL ila insulinèmia augmenten en l’ordreDM2>TGL+GAA>TGL>control, mentre que l’HOMA-Sdisminueix en l’ordre invers. No hi ha diferències entreels grups respecte d’homocisteïna, vitamina E a lesLDL i ox-LDL. La funció endotelial i el GIM de lescaròtides i femorals estan alterats als 3 grups de TGLen comparació amb els controls (P<0,01). El perfildiürn de TG es correlaciona directament (P<0,05) ambpes, perímetre de cintura, GLU basal, HbA1c, TGbasal, Apo B, oxLDL i GIM de caròtides i femorals, iinversament (P<0,01) amb el cHDL i HOMA-S. Perregressió lineal múltiple, la GLU i els TG basals són elsúnics predictors independents de l’ASC de TG (r2ajustada 0,518, P=0,004). D’altra banda, el perímetrede cintura (negatiu) i l’HOMA-S (positiu) són els únicspredictors independents de la VDDE (r2 ajustada0,281, P<0,001) i de la VDIE (r2 ajustada 0,131,P<0,01), respectivament.
ConclusionsEl perfil diürn domiciliari de TG en sang capil·larproporciona informació rellevant sobre el risc aterogè-nic en persones sense factors de risc convencionals iamb mínimes alteracions de GLU i TG en dejú, i enpacients amb DM2 ben controlada. Una sola determi-nació a les 4-5 h del dinar proporciona una informaciósimilar. El perfil diürn alterat de TG capil·lars ésfreqüent en les persones amb TGL i s’associa ambuna agrupació de factors de risc relacionats amb lasíndrome de resistència a la insulina i aterosclerosipreclínica. Per això, la presència de TGL amb o senseGAD implica ja un risc cardiovascular augmentat, totsuggerint que aquests trets precoços de la síndromemetabòlica mereixen atenció terapèutica. Malgrat unbon control glucèmic, la majoria de pacients ambDM2 i TGL en dejú tenen un perfil diürn de TG moltalterat, tot indicant la necessitat de tractament especí-fic. L’obesitat visceral, una alteració central en lasíndrome metabòlica, s’associa fortament amb disfun-ció endotelial.
2. ResultatsTaula 1. Característiques basals dels 4 grups de l’estudi.
Dades en mitjana±DE excepte Lp(a): mitjana (P25 —p75) i test de Kruskal-Wallis
Comparacions entre grups per test post-hoc deBonferroni:
a p<0,05 comparat amb el grup controlb p<0,05 comparat amb el grup TGLc p<0,05 comparat amb el grup TGL+GAAd p<0,05 comparat amb el grup TGL+DM2
Els valors de GLU i TG varien entre els grups perdefinició, perquè són les variables emprades perconstruir-los.
Taula 2. Paràmetres metabòlics i oxidabilitat de les LDL en els 4
grups
Dades com a mitjana±DE excepte àcids grassoslliures: mitjana (P25 — p75) i test de Kruskal-Wallis
TTOG, test de tolerància oral a la glucosa;* P<0,05 comparat amb els grups control i TGL (testpost-hoc de Bonferroni)
En fer un TTOG amb determinació de glucèmia a los120´, es van detectar 2/16 (16,5%) subjectes del grupTGL i 5/15 (33%) del grup TGL+GAA amb un TTOGpatològic, indicatiu de DM2.
Taula 3. Distribució dels genotips de l’Apo E
FUNDACIÓ LA MARATÓ DE TV3

La distribució de genotips de l’Apo E és similar entreels grups i no difereix de la descrita en la poblaciógeneral espanyola.
Taula 4. Perfil diürn de TG i GLU en sang capil·lar. Valors
màxims (pic) i àrees sota la corba de 16 hores (8:00 h a 24:00
h) en els 4 grups
Dades com a mitjana ± DE.
Comparacions mitjançant test post-hoc de Bonferroni:
a P <0,05 respecte del grup controlb P <0,05 respecte del grup TGLc P <0,05 respecte del grup TGL+GAA
En tots els grups el pic de GLU s’esdevé a les 24:00h, 4 h després del sopar, mentre que el pic de TGs’observa a les 19:00 h, 4-5 hores després del dinar.
D’una manera previsible, els pacients amb DM2presenten xifres de GLU i TG superiors a les delsaltres grups durant el dia. No obstant això, calremarcar que el perfil diürn de TG en sang capil·lar ésmolt patològic en el grup DM2 malgrat un bon controlglucèmic i unes xifres de TG en dejú relativamentnormals (100-200mg/dL en les anàlisis convencionalsde laboratori amb condicions correctes preextracció).Tanmateix, durant el perfil diürn observem que elsvalors basals (en dejú al domicili) són sovint superiorsals 200 mg/dL, i durant el dia resten molt por sobrede los 200 mg/dL.
Taula 5. Límit superior de la normalitat (mitjana + DE o percentil
95) de les àrees sota de les corbes i els pics postprandials de
GLU i TG d’acord amb els resultats del grup control
Taula 6. Proporció de subjectes en cada grup amb àrea sota de la
corba o pic de GLU i TG per sobre del percentil 95 dels valors del
grup control
La proporció de subjectes amb valors patològicsaugmenta seguint l’ordre TGL+DM2>TGL+GAA>TGL.Destaca que més del 50% de subjectes en tots elsgrups tenen hiperlipèmia postprandial. També calremarcar que menys del 50% dels pacients amb DM2tenen un ABC de GLU patològic, tot confirmant el seubon control metabòlic.
Taula 7. Correlacions bivariants (Pearson) entre els valors del perfil
diürn de TG i GLU en sang capil·lar i factors de risc cardiovascu-
lar. Valor del coeficient de regressió (r) i significació estadística
Només s’indiquen les correlacions estadísticament significatives (P
<0,05).
a P <0,05; b P <0,01; c P <0,001. En negreta,correlació amb més potència estadística per a cadauna de les quatre variables.
Com es podia suposar les correlacions més potentss’observen entre els perfils de GLU i TG i el controlglucèmic (HbA1c) i TG basals, respectivament.
Taula 8. Correlacions entre les ABC i els pics de GLU i TG durant
els perfils diürns en sang capil·lar.
Per tant, les determinacions aïllades de TG a les 4hores del dinar o de la GLU a les 24:00 hores fornei-xen un mesurament fiable del perfil diürn, és a dir, elperfil diürn de TG en sang capil·lar es pot substituiramb avantatges per una única determinació 4 horesdesprés del dinar, i el de GLU per una sola determina-ció a les 24 hores.
Taula 9. Correlacions bivariants (Pearson) entre els paràmetres de
reactivitat vascular i els factors de risc. Valor del coeficient de
regressió (r) i significació estadística
RECERCA BIOMÈDICA | 203
Només s’indiquen les correlacions estadísticamentsignificatives (P <0,05).
a P <0,05; b P <0,01; c P <0,001. En negreta,correlació amb més potència estadística per acadascuna de les dues variables.
Les correlacions més fortes es van observar entreVDDE i perímetre de cintura (inversa) i entre VDIE isensibilitat a la insulina (directa). En sengles anàlisis deregressió logística en passos successius amb lesvariables de reactivitat vascular com a variablesdependents, els dos paràmetres es mostren com apredictors independents, amb la qual cosa cap delsaltres entra en el model. En el cas de la VDDE, elcoeficient beta estandarditzat per al perímetre decintura és—0,481, amb una r2 corregida de 0,220(P<0,001). Per a la VDNTG, el coeficient betaestandarditzat és 0,414, amb una r2 corregida de0,152 (P=0,004). Figura 1. Vasodilatació dependent (VDDE) i independent (VNTG)
de l’endoteli en l’artèria braquial en els 4 grups. Els valors (mitjana
i EEM) són percentatges del calibre basal del vas després de la
isquèmia i l’aplicació sublingual de nitroglicerina, respectivament.
Les diferències entre grups són significatives (ANOVA).
* P <0,005 respecte del grup control.Figura 2. Gruix màxim mitjà del complex íntima-mitjana en la
caròtida i l’artèria femoral superficial en els 4 grups. Valorsen
mitjanes i EEM. Les diferències entre grups són
significatives(ANOVA).
* P <0,005 respecte del grup control.
Figura 3. Línia de regressió i interval de confiança del 95% de la
correlació entre la vasodilatació dependent de l’endoteli (VDDE) i
el perímetre de cintura (r=0,481, P <0,001).
3. Rellevància i possibles implicacions clíniquesdels resultats finals obtingutsEls resultats obtinguts permeten afirmar que unadeterminació de TG en sang en dejú i en condicionspreextracció estàndard mostrant valors entre 100 i200 mg/dL (xifres que actualment no es considerenper intervenir) ja es tradueix en un cert risc aterogènic,que està relacionat amb les elevacions dels TG sèricsdurant el dia. La determinació del perfil diürn de TG ensang capil·lar amb el reflectòmetre és una einaadequada per avaluar aquest important aspecte delmetabolisme lipídic. Atès que l’obtenció de 8 mostresde sang capil·lar durant el dia pot ser feixuga per aalguns pacients i que hi ha una correlació excel·lententre l’àrea sota la corba de TG i el pic màxim de TG,el perfil diürn es pot substituir beneficiosament peruna sola determinació 4 hores després del dinar. Entreles troballes, destaca que una bona part dels pacientsamb HTG límit amb o sense GAA tenen pics de TGsuperiors als 300 mg/dL, mentre que els diabètics,malgrat un bon control metabòlic i uns TG en dejúentre 100 i 200 mg/dL, fan pics màxims de 400mg/dL de mitjana.
Això és important perquè gairebé mai s’insisteix en elcontrol lipídic amb dieta o fàrmacs hipolipidemiants enpacients pertanyents als 3 grups estudiats d’HTGL,GAA i DM-2. Aquest estudi suggereix que la síndromemetabòlica, tot i expressant-se amb un fenotip lleu, jacomporta un alt risc aterogènic que està en gran partmediat per la hiperlipèmia postprandial. En aquestsentit, la utilització del reflectòmetre per determinar elperfil diürn de TG en sang capil·lar pot ser una eina dediagnosi molt útil per millorar la definició de risccardiovascular i tractar-lo de manera adient enpacients seleccionats amb un fenotip lleu de risc a lesanàlisis convencionals en dejú.
Per tant, els resultats tenen una important utilitatpràctica i aplicabilitat en l’àrea de la prevenció cardio-vascular.
4. PublicacionsRos E, Rodríguez-Villar C, Núñez I, Gilabert R, Conget I.
Endothelial dysfunction in healthy subjects without cardiovascular
risk factors but with variable adiposity is related to visceral fat
accumulation. 35th Annual Meeting of the European Society for
Clinical Investigation, Barcelona, 2-5 de mayo de 2001 (Eur J Clin
Invest 2001;31 [Supl. 1]:26).
Rodríguez-Villar C, Casals E, Núñez I, Gilabert R, Conget I, Ros E.
Disfunción endotelial en pacientes con mínimas alteraciones del
metabolismo lipídico e hidocarbonado. XIV Congreso Nacional de
la Sociedad Española de Arteriosclerosis, Barcelona, 2-5 de mayo
de 2001 (Clin Invest Arterioscler 2001;13 [Supl. 2]:41-42).
Rodríguez-Villar C, Conget I, Núñez I, Gilabert R, Mercadé I,
Casals E, Ros E. Perfiles diurnos de trigliceridemia y glucemia
capilar como marcadores de riesgo cardiovascular. XXVII
Congreso Nacional de Diabetes, Cádiz, 2-4 de marzo de 2002.
(Avances en Diabetología, vol 18, supl.1.)
Rodríguez-Villar C, Conget I, Núñez I, Gilabert R, Mercadé I,
Casals E, Ros E. Perfiles diurnos de trigliceridemia y glucemia
capilar en controles sanos, sujetos con hipertrigliceridemia leve y
diabéticos. Relación con aterosclerosis preclínica.
XV Congreso Nacional de la Sociedad Española de
Atherosclerosis, Salamanca, 1-4 de mayo de 2002. (Clin Invest
Arterioscler 2002;14 [Supl. 2]:20).
FUNDACIÓ LA MARATÓ DE TV3

Gilabert R, Núñez I, Rodríguez-Villar C, Ros E.
Endothelial dysfunction in subjects with early stages of the
metabolic syndrome. Association with visceral adiposity and
insulin resistance. 74th Congress of the European Atherosclerosis
Society, Sevilla, 17-20 de abril de 2004. (Atherosclerosis Suppl
2004;5:51).
Cofán M, Rodríguez-Villar C, Deulofeu R, Núñez I, Gilabert R,
Ros E. Daytime capillary triglyceride profiles in health, borderline
hypertriglyceridemia, and new-onset type 2 diabetes.
74th Congress of the European Atherosclerosis Society, Sevilla,
17-20 de abril de 2004. (Atherosclerosis Suppl 2004;5:68).
RECERCA BIOMÈDICA | 205

FUNDACIÓ LA MARATÓ DE TV3
MEMÒRIA FINAL
1. Resum del projecte original
Objectiu principalcaracterització experimental de les adaptacions fisiolò-giques a la modificació del quocient entre oferta idemanda d’insulina.
Objectius secundariscaracterització biofísica, fisiològica i molecular del’oferta i la demanda d’insulina en els diversos modelsexperimentals; recerca de senyals plasmàtics,caracterització del quocient oferta/demanda i delfactor de seguretat del sistema, modelatge i tracta-ment farmacològic dels desequilibris entre l’oferta i lademanda d’insulina.
Hipòtesi de treballen condicions fisiològiques existeix un equilibri entrel’oferta i la demanda d’insulina, mentre que, indepen-dentment de l’etiologia, els distints quadres diabèticstenen en comú un desequilibri entre l’oferta i lademanda d’insulina.
Mètodes i resultats: pancreatectomia parcial, determi-
nació de la massa cel·lular beta i no beta, tolerància ala glucosa, caracterització de l’oferta (acoblamentestímul-secreció) i caracterització de la demanda.
Impacte potencial esperatmillorar els procediments diagnòstics i terapèutics enla prediabetis.
2. Resultats1. Obtenció del model amb disminució de massacel·lular betaDesenvolupament del model: s’ha desenvolupat unmodel de ratolí pancreatectomitzat un 60% (60% Px).
Es van estudiar quatre grups d’animals:i) grup Px14: animals estudiats als 14 dies de la 60%Px (n= 30);
ii) grup Px60: animals estudiats als 60 dies de la 60%Px (n= 14);
iii) grup control 14: animals "controls operats" delmateix pes i edat que els Px14 (n= 12), i
iv) grup control 60: animals "controls operats" delmateix pes i edat que els Px14 (n= 8).
Títol del projecte
Adaptacions fisiològiques a la modificació del quocient oferta/demanda d’insulina
Investigadors principalsBernat Sòria Escoms
InstitucióInstitut de Bioenginyeria. UMH. Alacant
125.041 €

Caracterització de l’oferta: es va analitzar en els grupsPx14 i control 14 la resposta secretora d’insulina enresposta a concentracions creixents de glucosa. Amés, es va analitzar, en els mateixos grups, la respos-ta dinàmica d’insulina en presència de concentracionsestimuladores de glucosa.
Estudi de l'activitat dels canals K(ATP): es vanmesurar els corrents unitaris del canal de potassidependent d’ATP (canal KATP) en registres de trossosde membrana plasmàtica escindits en la configuracióinside-out. No existeixen diferències significatives,entre les cèl·lules ß de ratolins Px14 i control 14, pelque fa a la inhibició del corrent del canal KATP induïdaper l’ATP. A més, no es van observar diferènciessignificatives entre els corrents unitaris, dels canalsKATP, de les cèl·lules ß obtingudes dels ratolins Px14 icontrol 14. Finalment, tampoc no es van observardiferències significatives entre les resistències d'entra-des observades en les cèl·lules ß procedentsd'ambdós grups de ratolins. Tots aquestes dadessuggereixen que, entre els possibles mecanismesd'adaptació que apareixen en resposta a la disminucióde la massa de cèl·lules ß, no figuren els canvis en elnombre de canals de KATP o en la probabilitatd'obertura d’aquests.
Estudis de la metabolizació de la glucosa: atès queels canals no es veuen afectats, l'altre pas del procésd’acoblament estímul-secreció que pot ser suscepti-ble de tenir modificacions, és la metabolizació de laglucosa. Aquests estudis indiquen que en els illotsprocedents dels ratolins Px14, la metabolització de laglucosa comença a concentracions més baixes. Amés, en aquests mateixos illots hi ha una majormetabolització de la glucosa. Totes aquestes dadessuggereixen que el mecanisme d'adaptació probable-ment tingui lloc en la metabolització de la glucosa, ipreferentment de la glucoquinasa.
Caracterització de la demanda: per a estudiar lademanda, es van mesurar en els quatre grups elsnivells circulants d’insulina.
Captació de glucosa en resposta a insulina enadipòcits i múscul: en absència d’insulina, tant per alsadipòcits, com per a les fibres musculars, la captacióde glucosa no es veia afectada en cap dels grupsd'animals estudiats (Px14, Px60, control 14 i control60). En canvi, la captació induïda per 1.25 nM insulinaera significativament més alta en els adipòcits i fibresmusculars dels animals Px14 que en la resta delsgrups d'animals. A més, per a tots els grupsd'animals, la captació de glucosa en presència d’1.25nM insulina sempre va ser superior a l'obtinguda enabsència d’insulina. Aquestes dades suggereixen quel'adaptació central, per si sola, no és suficient per aexplicar la normoglucèmia que tenen els animals enels quals hi ha una disminució de la massa de cèl·lulesß, com també una menor secreció d’insulina i pèptidC. Els mecanismes mitjançant el quals els animals delgrup Px14 controlen la seva glucèmia també passaper una menor resistència a la insulina.
Expressió del nivell de transportadors d’insulina enresposta al múscul: l'augment de la captació de
glucosa observada en els animals Px14 pot ser acausa de:
i) la presència d'un major nombre de transportadorsde glucosa (GLUT4) en la membrana dels adipòcits ifibres musculars;
ii) una major activitat dels transportadors, i
iii) ambdues coses. Aquestes dades suggereixen quela major captació de glucosa observada en elsmúsculs i adipòcits dels animals Px14, no és causadaper una major expressió de transportadors de glucosa(GLUT4), sinó probablement a una activitat augmenta-da d’aquests.
2. Obtenció del model amb augment de la massacel·lular betaCaracterització del model: es van aïllar 150 illots deLangerhans i es van trasplantar en l’interior de lacàpsula renal. A les 4 setmanes del trasplantament elsillots trasplantats es van recuperar per estudiar-los.
Caracterització de l’oferta: es va estudiar el senyal decalci en illots recuperats després de 4 setmanes delseu trasplantament. En eixos illots es van analitzar lesrespostes a secretagogs nutrients com la glucosa, asecretagogs no nutrients com la tolbutamida, i aagonistes muscarínics com el carbacol.
3. Obtenció d’animals transgènics hexoquinasa II iglucogen sintetasaDurant el darrer any s'han desenvolupat una sèrie demodels transgènics que permeten l'anàlisi de la contri-bució dels diversos teixits a l'augment de la resistèn-cia a la insulina. Aquests resultats, publicats desprésde l'inici d’aquest projecte, han justificat el canvid'aquest objectiu. Amb aquesta hipòtesi de treball esvan plantejar diversos models d’animals transgènics.La idea principal consistia a modificar l’equilibriintracel·lular entre els processos de lipòlisi i els delipogènesi que ocorrien dintre de la cèl·lula ß. L’enzimclau que regula aquest procés és la carnitina palmitoiltransferasa 1 (CPT-1). Es van generar ratolins transgè-nics amb CPT-1. Curiosament, no van poder obtenir-se ratolins homozigòtics entre F1 i F3, solamentanimals hemizigòtics. A pesar d’això, es va comprovarla tolerància a la glucosa dels distints animals hemizi-gòtics mitjançant un test d’injecció intraperitoneal deglucosa.
4. Resultats amb apoptosiEstudis preliminars amb animals transgènics sobreex-pressant el gen de la carnitina palmitoil transferasa-1(CPT-1) en cèl·lula ?, semblaven suggerir que aquestenzim podria estar implicat en processos de mortcel·lular. La CPT-1 és l’enzim clau en la ?-oxidaciólipídica a nivell mitocondrial, tot permetent el seuaccés a aquest orgànul on són degradats per ?-oxidació. Les evidències acumulades fins al momentsemblaven suggerir que l’acumulació de lípids esterifi-cats en la cèl·lula ? (procés que ocorre en els casosde diabetis associada a obesitat) implicaven disfun-cions severes en aquest tipus cel·lular. Per a verificaraquesta hipòtesi, vam decidir estudiar el fenomen enuna línia cultivada de cèl·lula ? (INS-1). Les anàlisis de
RECERCA BIOMÈDICA | 207

cicle cel·lular mitjançant citofluorimetria de flux vanmostrar que les cèl·lules incubades en presènciad’àcids grassos presentaven un alt percentatge de lapoblació en un estadi subdiploide, molt similar al ques’observava quan les cèl·lules s’incubaven en presèn-cia d’estaurosporina (un agent que causa mort endiversos tipus cel·lulars). Les anàlisis de fragmentaciód’ADN mostren un patró en escala, fet que suggereixque la incubació en presència d’elevades concentra-cions d’àcids grassos indueix fenòmens d’apoptosi omort cel·lular programada.
5. Paper del calci en l’expressió gènica de lescèl·lules de l’illotEn la cèl·lula b -pancreàtica, diversos nutrients, com laglucosa o els àcids grassos, modulen l’expressió degens específics d’expressió ràpida, i majoritàriamentaquesta regulació es dependent de calci. Els ionsCa2+ ions són capaços de regular la transcripció degens diferents per la via d’un complex sistema desenyals restringits en l’espai i codificats temporalment.En aquest projecte, hem utilitzat un prototip demicroscopi confocal puntual per mesurar leselevacions de Ca2+ en el citosol i el nucleoplasma decèl·lules de l’illot de Langerhans estimulades durantbreus períodes amb nutrients diferents, el qual varevelar que un mateix estímul genera senyals quepresenten una amplitud major en el nucli respecte delcitosol. Utilitzant la tècnica de FISH, que permetl’estudi del control de la transcripció en cèl·lules indivi-duals, hem visualitzat l’expressió del gen c-myc enfunció d’aquests estímuls. Els resultats ens indiquenque l’expressió de c-myc en les cèl·lules de l’illot ésmolt sensible a l’amplitud dels senyals de calci,particularment en el nucli cel·lular. Aquests estudisindiquen que els nutrients poden modular l’expressióde gens específics mitjançant rangs d’amplituddefinits i associats al seu senyal de calci característic.
6. Caracterització d’illots humansAïllament d’illots humans: els illots humans que es vanobtenir per a aquest estudi procedien de donantshumans. Els donants eren d’ambdós sexes i amb unaedat compresa entre 10 i 55 anys. El temps d’isquè-mia freda va ser de 8 hores. Els illots es van aïllarseguint el protocol d’Edmonton.
Contingut i secreció d’insulina en resposta a glucosa:després de l’aïllament dels illots, es van cultivar a 37ºC i amb 5% CO2 durant 3 dies en medi Miami. Elcontingut de proteïnes i la dimensió dels illots aïllatspel protocol d’Edmonton és molt paregut al delsratolins. Respecte del contingut d’insulina, el seu valorés 3 vegades inferior al que tenen els illots de ratolí.Els illots humans mostren una secreció d’insulinabasal a concentracions no estimuladores de glucosa.Quan s’incrementa la concentració de glucosa perdamunt de 5 mM (1 g/l), els illots incrementen lasecreció d’insulina fins a concentracions màximes de22 mM (4 g/l). Després d’una setmana de cultiu, elsillots perdien la capacitat de secretar insulina enresposta a glucosa, tot mantenint la secreció basal.En aquest sentit, el rang de sensibilitat en resposta ala glucosa és igual a la dels illots de ratolí i al d’altresmolts mamífers, els quals regulen l’homeòstasi deglucosa entre nivells de 0.8 i 3 g/l. Els illots són
estables durant una setmana en cultiu, i per tant espot esperar a obtenir més donants en cas de no tenirsuficients illots amb un sòl aïllament per fer el trasplan-tament. Els valors totals de la secreció d’insulina sónsimilars als de ratolí.
Resposta del senyal intracel·lular de Ca2+ en presèn-cia de concentracions estimuladores de glucosa: enaquestos illots es va estudiar la resposta del senyalintracel·lular de Ca2+ en les poblacions cel·lulars pelseu patró de calci, i tal com ocorre amb els illots deratolí, les cèl·lules b incrementaren el seu senyal decalci en resposta a concentracions estimuladores deglucosa, mentre que les cèl·lules a van silenciar laseva resposta. Per un altre costat, les cèl·lules dmantenen el senyal de Ca2+ .
7. Obtenció del model d’Octodon degus hiperglu-cèmicObtenció de la soca hiperglucèmica d’Octodondegus: l’Octodon degus és un rosegador, d’origenxilè, que desenvolupa una malaltia que es pot identifi-car amb la diabetes tipus 2 dels humans.Fonamentalment, presenta hiperglucèmia espontània,cataractes i amiloidosi en els illots de Langerhans. Pera establir una colònia d’animals hiperglucèmics, s’hanestudiat 300 animals de la colònia existents a l’anima-lari durant un període de 9 mesos. Durant aquesttemps se’ls ha mesurat les xifres de glucèmia en dejúi s’han identificat 10 animals clarament hiperglucè-mics. Finalment, aquests animals (4 mascles i 6femelles) s’han encreuat entre ells i amb animalslleument hiperglucèmics (en els casos en els quals nohi havia parelles per a ells) amb el fi d’establir lacolònia d’Octodons hiperglucèmics.
Caracterització de l’oferta-demanda en Octodondegus controls: es va estudiar la resposta secretorad’insulina en 5 Octodons mascles normoglucèmics,de 4-5 mesos d’edat, en resposta a concentracionscreixents de glucosa, a altres secretagogs nutrientscom el ketoisocaproat. A més, es va analitzar laresposta dinàmica d’insulina en presència de concen-tracions estimuladores de glucosa.
8. Obtenció del model de malnutrició proteinoener-gèticaLa malnutrició proteinoenergètica no només afecta elspaïsos subdesenvolupats. Als països industrialitzats,ens vam trobar igualment poblacions afectades. Unamalnutrició comporta sovint alteracions de l’homeòs-tasi glucídica. Açò va ser clarament evidenciat enpacients amb Kwashiorkor que presentaven hipoglu-cèmia. Per altra banda, una malnutrició està associa-da en la majoria dels casos a una intolerància a laglucosa, com també a una resposta insulinosecretoradisminuïda, anomalies aquestes més patents en nensamb mancances proteiques. D’aquesta manera, moltsnens amb marasme presenten una tolerància a laglucosa lleument disminuïda o normal, amb unasecreció d’insulina en el mateix sentit, mentre quenens amb Kwashiorkor presenten una clara intoleràn-cia a la glucosa, com també una hipoinsulinèmia.Aquestes observacions han conduït a implicar lamalnutrició, fonamentalment proteica, en l’etiologia dela diabetis. Tenint en compte tot el que s’ha esmentat
FUNDACIÓ LA MARATÓ DE TV3

anteriorment i la dificultat d’estudiar la malnutrició enl’home in situ, cal utilitzar models animals. A més, elsmodels animals permeten estudiar l’impacte d’una odiverses mancances específiques, ja que proporcio-nen l’oportunitat de realitzar estudis nutricionalsrigorosament controlats. En el nostre estudi, hemelegit un model de malnutrició fonamentalment protei-ca. La restricció elegida en el nostre model ha estatde 2/3 en proteïnes (5%) i 1/3 en calories (65%), sentla rata l’espècie animal utilitzada. En aquest treball,estem interessats a investigar el control nerviós de lasecreció d'insulina de forma aïllada, sense la influènciad’altres factors que puguin intervenir en la resposta.Per a això, analitzàrem el control directe exercit pelsprincipals neurotransmissors del sistema nervióssimpàtic i parasimpàtic (noradrenalina i acetilclolina,respectivament) sobre la secreció d'insulina d’illotsaïllats de rates amb malnutrició. En primer lloc, hemestudiat la resposta insulínica d’illots aïllats de ratesamb malnutrició i controls respecte de les concentra-cions de glucosa esmentades prèviament. Elsresultats mostren una resposta insulinosecretora moltdisminuïda en els illots de rates amb malnutrició. A lavista d’aquests resultats, vam investigar si les altera-cions funcionals dels illots procedents de rates ambmalnutrició es podien explicar per una disminució delseu contingut en insulina. Aquest últim paràmetre es vatrobar netament disminuït en illots procedents de ratesamb malnutrició respecte del contingut d’insulina enillots procedents de rates controls. Atesa la importàn-cia d’aquesta diferència, vam representar la secreciórespecte del contingut específic de cada tipus d’illots.Tal com s’observa, fins i tot en utilitzar aquesta formad’expressió, les diferències entre els dos grups d’illotspersisteixen. Per altra banda, s’observa que la respos-ta secretora màxima d’illots de rates amb malnutricióés inferior a l’observada en illots de rates controls, sentla concentració de glucosa a la qual s’obté la meitatd’aquesta resposta màxima, superior en illots de ratesamb malnutrició respecte dels controls.
3. Rellevància o aplicabilitat clínica dels resultatsfinals obtinguts1. L’anàlisi de la fisiopatologia en situacions dedesequilibri de l’oferta i la demanda d’insulina:
a) Ens permet conèixer les adaptacions fisiològiquesdurant el procés d’instal·lació de la diabetis o prediabetis.
b) Introdueix un model per a estudiar la progressió dela malaltia.
c) Pot ser utilitzat per assajar noves estratègiesterapèutiques.
2. L’obtenció de cèl·lules productores d’insulina apartir de cèl·lules pluripotencials pot ser d’utilitat enl’elaboració de noves teràpies, per exemple la teràpiacel·lular de la diabetis mellitus.
3. S’ha iniciat la caracterització dels illots humans, pertal de saber si les observacions amb models animalses poden traslladar al pacient diabètic.S’ha iniciat lacaracterització dels illots humans, per tal de saber siles observacions amb models animals es podentraslladar al pacient diabètic.
4. PublicacionsAny 2000
Gil A, Segura J, Pertusa JAG, Soria B (2000) Monte Carlo
simulation of 3-D buffered Ca2+ diffusion in neuroendocrine cells.
Biophysical Journal 78: 13-33
Soria B, Roche E, Berná G, León-Quinto T, Reig JA, Martin F
(2000) Insulin-secreting cells derived from embryonic stem-cells
normalize glycaemia in streptozotocin-induced diabetic mice.
Diabetes 49: 157-162
Soria B, Andreu E, Berná G, Fuentes E, Gil A, León-Quinto T,
Martín F, Montanya E, Nadal A, Reig JA, Ripoll C, Roche E,
Sanchez-Andrés JV, Segura J (2000) Engineering Pancreatic
Islets. Pflügers Archiv- Eur. J. Physiol. 440: 1-18
Quesada I, Martín F, Soria B (2000) Polarized and sustained
submembranal Ca2+ microgradients in pancreatic islet cells are
modulated by nutrient secretagogues. Journal of Physiology.
525(1): 159-167
Roche E, Maestre I, Martín F, Fuentes E, Casero J, Reig JA,
Soria B (2000) Nutrient toxicity in pancreatic b -cell dysfunction. J.
Physiol. Biochem. 56(2): 119-128
Charollais A, Gjinovci A, Huarte J, Bauquis J, Nadal A, Martín F,
Andreu E, Sanchez-Andrés JV, Calabrese A, Bosco D, Soria B,
Wollheim CB, Herrera PL, Meda P (2000) Junctional communica-
tion of pancreatic-beta cells contributes to the control of insulin
secretion and glucose tolerance. Journal of Clinical Investigation
106: 235-243
Segura J, Gil A, Soria B (2000) Modelling study of exocytosis in
neuroendocrine cells: influence of the geometrical parameters.
Biophys. J. 79: 1771-1786
Nadal A, Soria B (2000) Imaging intracellular calcium in living
tissue by laser-acanning confocal microscope. En: Calcium: The
molecular basis of calcium action in biology and medicine. ISBN
0-7923-6421 (eds. R Pochet, R Donato, J Haiech, C Heizmann, V
Gerke) pp. 653-663. Kluwer academic publishers, Dordecht,
Holanda
Nadal A, Laribi O, Ropero AB, Maillet M, Fuentes E, Soria B
(2000) Non-genomic actions of estrogens and xenoestrogens by
binding at a plasma membrane receptor unrelated to estrogen
receptor alpha and estrogen receptor beta.Proceedings of the
National Academy of Sciences 97(21): 11603-11608
Any 2001McLennan AG, Barnes LD, Blackburn GM, Brenner Ch,
Guranowsky A, Miller AD, Rovira JM, Rotllán P, Soria, B, Tanner
JA, Sillero A (2001) Recent progess in the study of the intracellular
function of diadenosine-polyphosphates. Drug Development
Research 52: 249- 259
Soria B, Skoudy A, Martin F (2001) From stem cells to b -cells:
new strategies in cell therapy of diabetes mellitus. Diabetologia
44: 407- 415
Berná G, León-Quinto T, Enseñat-Wasser R, Montanya E,
Martín F, Soria B (2001) Stem cells and Diabetes. Biomed.
Pharmacother. 55: 206- 212
Soria B (2001) In-vitro differentiation of pancreatic ?-cells
Differentiation 68: 205-219
Nadal A, Ropero AB, Fuentes E, Soria B (2001) The oestrogen
plasma membrane receptor: nuclear or unclear Trends in
Pharmacological Sciences 22: 597-599
Soria B (2001) Trasplante de islotes pancreáticos y de celulas
diferenciadas a partir de células madre. Avances en Diabetología
17: 121-128
Berna G, León-Quinto T, Fuentes E, Andreu E, Nadal A, Roche
E, Martin F, Reig JA, Soria B Ingeniería Celular y Diabetes Mellitus
Rev Clin Esp 201: 548-556
Andreu E, Pomares R, Soria B, Sanchez Andrés JV (2001)
Homogeneity in the electrical activity pattern as a function of
RECERCA BIOMÈDICA | 209

intercellular coupling in cell networks. Lect. Notes Comp. Sci.
2084: 14-20.
Any 2002Ropero, AB, Soria B, Nadal A (2002) A membrane non classi-
cal estrogen receptor triggers rapid differential actions in the
endocrine pancreas Molecular Endocrinology 16 (3): 497-505
Quesada I, Rovira JM, Martín F, Roche E, Nadal A, Soria B
(2002) Nuclear KATP channels trigger nuclear Ca2+ transients and
gene expression. Proc Natl Acad Sc USA 99(14) : 9544- 9549
Quesada I, Fuentes E, Viso-León MC, Soria B, Ripoll C, Nadal
A (2002) Low doses of the endocrine disruptor bisphenol-A and
the native hormone 17b estradiol rapidly activate the transcription
factor CREB. FASEB J 16(12): 1671-1673
Enseñat-Waser R, Martín F, Barahona F, Vázquez J, Soria B,
Reig JA (2002) "Direct visualization by confocal fluorescent
microscopy of the permeation of myristoylated peptides through
the cell membrane" IUBMB Life 54: 33- 36
Reig JA, Enseñat-Waser R, Roche E, Soria B (2002) "Terapia
Celular en la Diabetes Mellitus" Endocrinol Nutr 49(8): 260-268
Sòria B, Juan V (2002) Cèl·lules mare, embrions i clonació: el
naixement d’un nou paradigma? Annals de Medicina 85: 71-72
Any 2003Maestre I, Jordan J, Calvo S, Reig JA, Cena V, Soria B, Prentki
M, Roche E. (2003) Mitochondrial Dysfunction Is Involved in
Apoptosis Induced by Serum Withdrawal and Fatty Acids in the
beta-Cell Line Ins-1. Endocrinology 144(1): 335-345.
Quesada I, Fuentes E, Andreu E, Meda P, Nadal A, Soria B
(2003) On-line analysis of gap junctions reveals more efficient
electrical than dye coupling between islet cells. American Journal
of Physiology 282: E980 - E987.
Martín F, Jones J, Vaca P, Berná G, Soria B (2003) Production
de cellules sécrétant l’insuline à partir de cellules souches.
Flammarion Médecine-Sciences. Journés de Diabetologie 2002.
Diabètologie 2003: 23 -30. Flammarion ISSN 0075-4439; ISBN 2-
257-10842-6.
Fernández-Llebrez P, Bermúdez-Silva F, León-Quinto T, Martín
F, Soria B, Nadal A, Pérez J (2003) Bovine subcomissural organ
displays spontaneous and synchronous intracellular calcium
oscillations. Brain Res 977(1): 90-96
Roche E, Burcin MM, Esser S, Rüdiger M, Soria B (2003) The
use of gating technology in bioengineering insulin-secreting cells
from embryonic stem cells. Cytotechnology 41: 145-151.
Roche E, Sepulcre MP, Ensenat-Waser R, Maestre I, Reig JA,
Soria B. (2003) Bio-engineering inslulin-secreting cells from
embryonic stem cells: a review of progress. Med Biol Eng
Comput. 41(4):384-91.
Vaca P, Berna G, Martin F, Soria B (2003) Nicotinamide induces
both proliferation and differentiation of embryonic stem cells into
insulin-producing cells. Transplant Proc. 35(5): 2021-3.
Any 2004
Quesada, I, Martin F, Roche E, and Soria, B (2004) Nutrients
induce different Ca2+ signals in cytosol and nucleus in pancreatic
B-cells. Diabetes 53: S86-S95.
Soria B, Quesada I, Ropero AB, Pertusa JAG, Martín F, Nadal
A (2004) Novel players in pancreatic islet signaling: from membra-
ne receptors to nuclear channels. Diabetes 53: S86-S91.
Anouchka Skoudy, Meritxell Rovira, Pierre Savatier, Franz
Martin, Trinidad León-Quinto, Bernat Soria and Francisco X. Real
(2004) TGF?, FGF and retinoid signalling pathways promote
pancreatic exocrine gene expression in mouse embryonic stem
cells. Biochem J 379: 749-756.
Nadal A, Ropero AB, Fuentes E, Soria B, Ripoll C (2004)
Estrogen and xenoestrogen actions on endocrine pancreas from
ion channel modulation to activation of nuclear function. Steroids
(en premsa).
León-Quinto T, Jones J, Vaca P, Berná G, Skoudy A, Real FX,
Martín F, Soria B (2004) In-vitro directed differentiation and
selection of mouse embryonic stem cells into islet precursors.
Diabetología 48 (en premsa).
FUNDACIÓ LA MARATÓ DE TV3

RECERCA BIOMÈDICA | 211
MEMÒRIA FINAL
1. Resum del projecte originalDades tant genètiques com fisiològiques indiquen queel gen UCP3 és un gen candidat d'obesitat i diabetistipus II i que la proteïna UCP3 està estretamentrelacionada amb la utilització d'àcids grassos pel teixitmuscular amb finalitat oxidativa i, en general, amb ladespesa energètica en aquest teixit. Defectes enl'expressió o activitat d'UCP3 estarien relacionats ambel desenvolupament de resistència a la insulina enl'obesitat i diabetis tipus II, amb la qual cosa UCP3constituiria una diana potencial d'acció terapèutica enaquestes patologies.
Amb la perspectiva de poder actuar sobre la funcióbiològica d'UCP3 en el múscul i així promoure l'ús desubstrats metabòlics amb finalitat oxidativa, es podenplantejar tres nivells diferents d'actuació: a) influenciarl'expressió del gen UCP3 endogen, b) influenciar laproteïna UCP3 ja existent, c) sobreexpressar ectòpica-ment el gen UCP3. L'avenç en qualsevol d'aquests tresàmbits exigeix l'obtenció de models cel·lulars i animalsexperimentals capaços de proporcionar informació útil iaplicable en última instància a humans.
Els objectius específics que es plantegen en aquestprojecte són:
1. Caracterització de l'acció de fibrats, tiazolidinedio-nes i altres molècules sobre l'expressió del gen UCP3en múscul esquelètic.
2. Obtenció d'un model de ratolins transgènics útil pera l'estudi de la regulació i acció de fàrmacs sobre elgen UCP3 en humans.
3. Obtenció de models cel·lulars per a l'anàlisi del'activitat de la proteïna UCP3. Identificació demolècules que afectin l'activitat i de dominis funcionalde la proteïna UCP3.
4. Obtenció de vectors adenovírics dirigint l'expressiód'UCP3, eines potencials de teràpia gènica delmetabolisme oxidatiu muscular.
2. ResultatsObjectiu 1Hem identificat els fibrats (activadors de PPARalfa)com les principals molècules activadores del genUCP3 en múscul del ratolí adult, mentre que els
Títol del projecte
La proteïna desacobladora mitocondrial UCP3, nova diana terapèuticade l'obesitat i la diabetis tipus 2
Investigador principalFrancesc Villarroya Gombau
InstitucióFacultat de Biologia. UB
101.831 €

agents antidiabètics tiazolidindiones (activadores dePPARgamma) no mostren acció. Això no és a causad’una insensibilitat del gen UCP3, sinó pel contexttissular que determina la presència o no de receptorPPARgamma, ja que en adipòcits marrons, onPPARgamma és present, l'expressió d'UCP3 éssensible a les tiazolidindiones tot i que en un menorgrau que als fibrats. D'altra banda, la sensibilitat delgen UCP3 als fibrats es veu condicionada per l'estatfisiològic de l'animal i és particularment gran quan elsnivells circulants d'àcids grassos lliures són baixos, talcom s'esdevé, per exemple, durant l'alletament. Elreceptor PPARalfa és el mitjancer principal delsefectes d’àcids grassos i fibrats sobre la regulació delgen UCP3. No obstant això, PPARdelta pot tenir unpaper compensador, com hem demostrat en elsestudis emprant ratolins knockout per a PPARalfa.
Objectiu 2S'han establert els mecanismes de regulació de latranscripció del gen UCP3 humà mitjançant aproxima-cions in vivo i in vitro. El promotor del gen humàUCP3 és controlat per dos factors principals:receptors PPAR, que li confereixen sensibilitat a àcidsgrassos i fibrats, i MyoD, que està implicat enl’expressió específica a múscul esquelètic. Ambdóselements tenen una potent interacció funcionalmediada per coactivadors amb activitat histonaacetilasa. Aquests factors interactuen amb la zona 5’no-codificant proximal del gen humà. No obstant això,els estudis amb transgènics indiquen que existeixenelements distals en 5’ o bé en introns que sónnecessaris per a l’expressió màxima del gen. El modelde ratolins transgènics contenint el gen complet UCP3és millor model que la construcció que només conté laregió 5’ promotora no codificant per tal de provarfàrmacs amb potencial efecte sobre el gen UCP3 humà.
Objectiu 3El model d'estudi d'expressió induïble d’UCP3 encèl·lules en cultiu (293 amb UCP3 sota el control delsistema Tet-on) és adient per a estudis funcionalsd’UCP3, ja que dóna lloc a modificacions regulades(per àcids grassos i GDP), no artefactuals, delpotencial de membrana mitocondrial i de la respiració,La presència d’UCP3 al mitocondri, juntament ambefectes esperables sobre la bioenergètica cel·lular(desacoblament moderat) dóna lloc a altres efectes enl’homeòstasi cel·lular, tot produint especialment unasensibilització respecte d’estímuls de l’apoptosi.
Objectiu 4La sobreexpressió d’UCP3 en cèl·lules muscularshumanes mitjançant el vector adenovíric desenvolupatdóna lloc a una disminució del potencial de membra-na mitocondrial sense deplecionar la cèl·lula d’ATP.Promou la utilització oxidativa per la cèl·lula deglucosa i d'àcids grassos, però en fer competirambdós substrats, afavoreix la utilització dels àcidsgrassos. Si es provoca una expressió ectòpicad’UCP3 al fetge de ratolí de manera experimental, nohi ha efectes metabòlics deleteris.
3. Rellevància i possibles implicacions clíniquesdels resultats finals obtingutsEls resultats obtinguts tenen com a rellevància princi-
pal, haver proporcionat:
a) Informació de tipus bàsic sobre quina funció faUCP3 en el múscul i com pot repercutir això en laseva consideració com a potencial diana terapèuticaper a la diabetis tipus II i/o obesitat. Augmentant laquantitat d’UCP3 al mitocondri es constaten efectesmolt prometedors en aquest sentit (es promou l’oxida-ció d’àcids grassos sense impedir l’oxidació deglucosa, el desacoblament obtingut no depleciona lacèl·lula d’ATP...), però se’n detecten d’altres (l’afavori-ment de l’apoptosi amb certs estímuls) que reclama-ran una anàlisi posterior, ja que no és clar si poden serbeneficiosos o deleteris.
b) La identificació de quines vies podrien emprar-seper influenciar l’expressió d’UCP3: l’activació dereceptors PPAR, principalment PPARalfa, i per tantassenyalant a mol·lècules derivades dels fibrats com afàrmacs candidats de qualsevol intent farmacològicd’induir UCP3. UCP3 apareix com una diana fins arano coneguda dels efectes beneficiosos dels fibrats, nonomés com a hipolipemiants sino també com apromotors de sensibilitat a la insulina.
c) La disponibilitat de nous models animals (transgè-nics que expressen el gen UCP3 humà), cel·lulars(cèl·lules 293 amb expressió d’UCP3 humana induïbleper tetracicilina) i biotecnològics (adenovirus quedirigeixen l’expressió de la proteïna UCP3 humana).Aquests models podran ser emprats per estudisposteriors a aquest projecte per aprofundir en elpaper d’UCP3 i la seva implicació en les disfuncionsmetabòliques com la diabetis tipus II o l’obesitat.
4. PublicacionsPedraza, N.; Solanes, G.; Carmona, M.C; Iglesias, R.; Viñas,
O.; Mampel, T.; Vázquez, M.; Giralt, M.; Villarroya, F. Impaired
expression of the uncoupling protein-3 gene in skeletal muscle
during lactation. Fibrates and troglitazone reverse lactation-
induced down-regulation of the uncoupling protein-3 gene
Diabetes 49, 1224-1230, 2000
Solanes, G.; Pedraza, N..; Iglesias, R.; Giralt, M.; Villarroya, F.
The human uncoupling protein-3 gene promoter requires MyoD
and is induced by retinoic acid in muscle cells. FASEB Journal 14,
2141-2143, 2000
Pedraza, N.; Solanes, G.; Iglesias, R.; Vázquez, M.; Giralt, M.,
Villarroya, F. Differential regulation of uncoupling protein-2 and -3
gene expression in brown adipose tissue during lactation in mice
Biochemical Journal, 355 105-111, 2001
García-Martínez C, Sibille B, Solanes, G.; Darimonet C, Mace
K, Villarroya F, Gomez-Foix, A.M. Overexpression of UCP3 in
cultured human muscle lowers mitochondrial membrane potential,
raises ATP/ADP ratio and favors fatty acids vs. glucose oxidation
FASEB Journal, 15, 2033-2035, 2001
Villarroya F, Brun S, Giralt M, Cámara Y, Solanes G, Iglesias R.
Gene expression of leptin and uncoupling proteins: molecular
end-points of fetal development Biochemical Society Transactions
29, 76-79, 2001
Villarroya, F. ¿La proteína desacoplante mitocondrial UCP3, un
nuevo actor en el metabolismo muscular de los ácidos grasos?
Clínica e Investigación en Arteriosclerosis 15, 114-123, 2003
Villarroya, F., Iglesias, R., Giralt.M. Retinoids and retinoid
receptors in the control of energy balance: novel pharmacological
strategies in obesity and diabetes Current Medicinal Chemistry
11, 1241-1253, 2004
FUNDACIÓ LA MARATÓ DE TV3

Solanes, G., Pedraza, N., Iglesias, R., Giralt, M., Villarroya,F.
Functional relationship between MyoD and PPAR-dependent
regulatory pathways in the control of human uncoupling protein-3
gene transcription. Molecular Endocrinology, 17,1944-58, 2003
Dejean, L, Camara Y, Sibille B, Solanes, G. Villarroya,F.
Uncoupling protein-3 sensitizes cells to the mitochondrial-
dependent stimulus of apoptosis. Journal of Cell Physiology. 2004
(en premsa).
RECERCA BIOMÈDICA | 213

FUNDACIÓ LA MARATÓ DE TV3
MEMÒRIA FINAL
1. Resum del projecte originalObjectiu principalL’objectiu principal és determinar el paper de l’interfe-ró beta (IFN-beta) en l’etiopatogènia de la diabetismitjançant la generació d’un ratolí transgènic consan-guini que expressi IFN-beta a les cèl·lules beta, enbackground diabetogènic (NOD) i resistent (NOR).
Objectius secundarisDeterminar:1) L’efecte de l’IFN-beta en la incidència i el desenvo-lupament de la diabetis en un model espontani dediabetis autoimmune (NOD) i en una soca resistent aaquesta malaltia (NOR).
2) L’efecte de l’expressió d’IFN-beta a la cèl.lula diana,tant en NOD com NOR a nivell fenotípic i de funció.
3) La influència del transgèn en l’expressió d’autoantí-gens tímics (tolerància central).
4) La caracterització de la transferència de la malaltiaentre els possibles animals diabètics i els resistents
generats (tolerància perifèrica).
Hipòtesi de treballL’interferó beta és una citocina inflamatòria antiviralproduïda per les cèl·lules epitelials que s’ha detectatals pàncrees de pacients amb diabetis tipus I. El ratolítransgènic CD1 que expressa IFN-beta selectivamenta les cèl·lules beta sota la regulació del promotor delgen de la insulina de rata (pIns-IFNbeta) és un modelper estudiar el paper d’aquesta citocina en l’etiopato-gènia de la diabetis tipus I, malaltia que desenvolupenun 25% d’aquests individus (amb presència d’insulitis ihiperexpressió d’MHC de classe I a les cèl·lulesinsulars). Es tracta d’un ratolí no consanguini generatper a interessos del Departament de Bioquímica de laFacultat de Veterinària (UAB) a partir d’un ratolíC57BL6/SJL que es va encreuar repetidament amb elratolí CD1 (outbred o no consanguini). La nostrahipòtesi es basa en el fet que, en un backgroundgenètic amb predisposició a la diabetis, l’expressiód’IFN-beta podria afectar la generació de precursors(tolerància central), la funció de les cèl·lules T, B imacròfags en perifèria i la generació d’un microam-bient inflamatori, tot incrementant la susceptibilitat al’atac autoimmune, i podria modular l’expressió de
Títol del projecte
IFN-beta i predisposició genètica a la diabetis tipus 1: anàlisi comparativa en el ratolí transgènic PINS-IFNbeta
no obès diabètic (NOD) i no obès resistent (NOR)
Investigador principalMarta Vives-Pi
InstitucióHospital Universitari Germans Trias i Pujol
Membres de l’equipAurora Alba, Raquel Planas, Jorge Carrillo, M. Carmen Puertas, Xavier Pastor, Rosa Ampudia,
Jaume Fernández-Llamazares, Ricard Pujol-Borrell i Joan Verdaguer.
70.198 €

l’autoantígen al pàncrees i al timus. La generaciód’aquest transgènic consanguini en un backgrounddiabetogènic (Non Obese Diabetic, NOD) i resistent(Non Obese Resistant, NOR) ens permetria estudiarl’expressió d’aquesta citocina d’efecte antiviral en elmodel de diabetis per excel·lència i en el seu homòlegresistent i així conjugar en un mateix model el paperdels factors ambientals i genètics.
Mètodes (disseny)Retroencreuaments durant 10 generacions defemelles NOD o NOR amb mascles CD1 pIns-IFNbeta. Seguiment molecular del transgèn per PCR apartir de DNA i emprant primers específics de l’IFN-beta humà. La correcta expressió del transgèn enanimals retroencreuats es determinarà per Northern oRT-PCR a partir d’RNA d’illots. Anàlisi demicrosatèl·lits dels retroencreuaments per determinarla presència d’al·lels NOD per PCR del DNA genòmic.Seguiment metabòlic amb control de pes i glicèmia.Seguiment immunològic per histologia convencional(H/E), immunohistologia (doble IFL indirecta) i anàlisien microscopi d’UV i tractament d’imatges (morfome-tria). Determinació de subpoblacions limfocitàriesmitjançant citometria de flux.
Estudis metabòlics in vitro dels illots de NOD i NORtransgènics pIns-IFNbeta examinant la resposta a laglucosa i determinació de secreció/contingut d’insuli-na (RIA). Determinació del grau d’expressió d’autoan-tígens insulars al timus mitjançant RT-PCR semiquanti-tativa. Transferència limfocitària a partir de limfòcitsesplènics aïllats per depleció amb anticossosmonoclonals i boles magnètiques.
Pla de treball1. El disseny del protocol inclou una primera etapad’uns 20 mesos per a la generació dels animalstransgènics en background NOD i NOR a l’estabularide l’Hospital Universitari Germans Trias i Pujol.
2. Establiment i caracterització immunològica delsmodels NOD pIns-IFNbeta i NOR pIns-IFNbeta. Enuna primera fase i a partir de N7 (excepte en l’estudidels autoantígens tímics que es farà en arribar a N10).La durada prevista és de 9 mesos.
3. Un cop aconseguits els individus homozigots (N10)s’avaluarà l’expressió d’autoantígens en timus. Ladurada prevista és de 6 mesos.
4. Un cop acabat el punt 1, iniciarem els experimentsde transferència de la malaltia. Aquesta part incloupreparació dels limfòcits del donant, transferènciapròpiament dita i seguiment del receptor. La duradaprevista és de 6 mesos.
Resultats esperatsAtès que disposem de resultats preliminars del ratolípIns-IFNbeta en background CD1, esperem quel’expressió d’aquesta citocina en el ratolí NOD acceleril’aparició de diabetis, mentre que a l’homòleg resistentNOR cal esperar que es desencadeni una diabetis perefecte de la inflamació local.
Impacte potencial esperat
Els resultats contribuirien a completar l’estudi delpaper de l’IFN-beta en la immunologia de la diabetis.Les dades podrien aplicar-se a la immunoteràpiaespecífica basada en l’administració de bloquejantsde citocines (anticossos monoclonals, receptorssolubles...) en el model animal i, a la llarga, en humans(immunoteràpia en una fase concreta de la prediabetis).
2. ResultatsGeneració del model amb canvi de backgroundgenètic mitjançant seguiment per microsatèl·lits
Per accelerar l’adquisició del background genètic, esvan identificar una sèrie de marcadors de tipusmicrosatèl·lit que diferencien —per dimensió del’amplímer generat per PCR de DNA— els ratolinsNOD i els NOR dels CD-1. Aquests marcadors esresumeixen a la taula 1. La identificació i utilitzaciód’aquests marcadors és la base d’un treball enpreparació (A. Alba, J. Carrillo, MC. Puertas, R.Planas, X. Pastor, R. Ampudia, C. Mora, R. Pujol-Borrell, J. Verdaguer, M. Vives-Pi. PCR-basedmicrosatellite analysis and genetic monitoring fordifferentiation of mice strains Non Obese Diabetic andNon Obese Resistant after backcrosses with transge-nic mice) i d’una presentació en un congrés nacional.
Taula 1. Marcadors microsatèl·lit que diferencien NOD i NOR de
CD-1
Incidència de diabetis
Un cop establertes les colònies en condicions SPF, laincidència de diabetis a la colònia original RIP-IFN-beta va ser del 22% en mascles i del 0% en femellesa les 23 setmanes d’edat (seguiment fins a 30setmanes). El primer pic de manifestació clínica dediabetis es va donar a les 8 setmanes d’edat (figura 1).
L’anàlisi de la generació resultant del desè retroen-creuament (BC10) amb NOR i amb NOD va mostraruna acceleració important en el moment d’aparició dela malaltia i un augment en la incidència. El 57% delsmascles i 22% de femelles RIP-IFN-beta NOR erendiabètics a les 7 setmanes d’edat, ott situant-se el
RECERCA BIOMÈDICA | 215
Fig. 1 Incidència de diabetis en RIP-IFN-beta en backgrounds
genètics diferents
primer pic d’aparició de la malaltia entre les 3 i les 4setmanes d’edat, coincidint amb el moment deldeslletament. Un 60% dels ratolins mascles RIP-IFN-beta NOD i un 25% de les femelles van manifestar ladiabetis a partir de les 3 setmanes d’edat. Lesmanifestacions clíniques van ser les mateixes que elmodel original: poliúria, polidípsia i pèrdua de pes, totconfirmant-se la diabetis amb determinacions deglicemia i glucosúria. Aquests resultats suggereixenque l’IFN-beta expressat al microambient de la cèl·luladiana productora d’insulina accelera l’autoimmunitaten un background genètic de susceptibilitat i trenca laprotecció del background resistent.
Insulitis scoreEl grau d’infiltració limfocitària (insulitis score) es vacalcular d’acord amb la intensitat de la insulitis (mínim0, màxim 4). Els pàncrees dels ratolins RIP-IFN-betaCD-1 de 4 a 18 setmanes d’edat van mostrar unafeble infiltració insular en un 26% dels illots, totcontrastant amb la marcada insulitis dels transgènicsen background NOD i NOR, en els quals més del85% dels illots estaven infiltrats, la majoria de manerasevera.
En la figura 2 es mostra el baix grau d’insulitis delmodel transgènic en background CD-1 comparat ambel model en background NOR (el model transgènicNOD té també el mateix grau que el transgènic NOR).
Figura 2. Grau d’infiltració insular (insulitis score) en ratolins
transgènics segons background genètic i desenvolupament de
diabetis.
Metabolisme de la insulinaPer eliminar la possibilitat que una reducció en lacapacitat productora d’insulina en els animals transgè-nics fos la causa de la malaltia, es va determinar laglicèmia i la insulinèmia, com també el contingutd’insulina pancreàtica en ratolins no diabètics (figura3). Pel que fa a la glicèmia, no es van detectar diferèn-cies significatives en els animals transgènics respectedels seus controls a les 4-20 setmanes d’edat,excepte en animals en inici de la diabetis. El nivelld’insulinèmia també va ser normal, igual que laconcentració d’insulina pancreàtica (contingut d’insuli-na en els illots). Els animals diabètics, evidentment,tenien nivells elevats de glicèmia i baixos d’insulinèmiai contingut d’insulina pancreàtica. Aquests resultatssuggereixen que el desenvolupament de la diabetis noestà causat per una toxicitat directa o errada metabò-lica de la cèl·lula beta com a conseqüència deltransgèn. Per confirmar aquesta dada es van dur aterme tests intraperitoneals de tolerància a la glucosaque van resultar normals, comparats amb els ratolinscontrols (no transgènics d’igual background genètic).Això demostra que la inserció del transgèn o eltransgèn en si, no té un efecte tòxic en els ratolins idóna suport al fet que som davant una diabetisautoimmunitària
Figura 3. Assajos metabòlics en els ratolins transgènics: a)
glicèmia, b) insulinèmia, c) contingut d’insulina pancreàtica.
Morfologia dels illotsUna de les troballes dels estudis de morfometria vaser que els illots dels animals RIP-IFN-beta enbackground original (CD-1) tenen una mida més gran
FUNDACIÓ LA MARATÓ DE TV3

que els seus controls a causa d’un major nombre decèl·lules endocrines. L’àrea mitjana dels illots delsanimals transgènics va ser de 17.499 µm2 respectedels 13.962 µm2 dels controls no transgènics, la qualcosa significa que els animals transgènics tenen elsillots un 25% més grans que els no transgènics;aquestes diferències són estadísticament significati-ves. La densitat cel·lular va ser la mateixa en els dosmodels (taula 2).
La composició de l’illot, quant a contingut de diversossubtipus de cèl·lules endocrines, va ser normal. Nos’han trobat diferències en la taxa de replicació(mitjançant la tècnica de BrdU), grau d’apoptosi en elsillots ni dimensió individual de les cèl·lules beta entreels dos grups. Ens proposem continuar els estudisper aclarir el motiu d’aquest augment de dimensió i laseva possible relació amb l’interferó beta.
Taula 2. Àrea i nombre de cèl·lules insulars (estudis de morfometria)
Immunopatologia: hiperexpressió d’MHC de classeI i fenotip de les cèl·lules limfomononuclearsinfiltrantsEn caracteritzar els models generats histopatològica-ment, es van descriure diferències importants en elgrau i el tipus d’infiltrat en funció del backgroundgenètic. Com a característica comuna, independent-ment del background genètic, tots els individustransgènics hiperexpressaven molècules d’MHC declasse I a les cèl·lules endocrines insulars i als endote-lis de l’illot. Tot seguit es detalla el grau i la composicióde l’infiltrat en funció del background genètic.
1. RIP-IFN-beta CD-1 (model original): en animalssans, el grau d’infiltració insular era molt baix, i lescèl·lules predominants eren limfòcits T, macròfags ialguna cèl·lula dendrítica (figura 4). En animalsdiabètics, es va observar un major grau d’insulitis, totpredominant les cèl·lules T CD4+ al voltant i dins l’illot.En animals diabètics, la composició o proporció delsdiversos subtipus cel·lulars en l’infiltrat era la mateixa,tot augmentant (lògicament) el percentatge d’illotsamb destrucció total de les cèl·lules beta (dada afavor del caràcter autoimmune de la malaltia).
2. RIP-IFN-beta NOD i NOR (models generats)
El patró de la insulitis observada en els dosbackgrounds genètics generats (NOD i NOR) va sermolt similar (cal recordar que aquests animalscompartien la majoria del genoma). Així doncs, elcanvi de background cap a un model diabetogènic oresistent va suposar un augment de l’infiltrat enanimals sans (prediabètics) i un canvi en la composi-ció d’aquest, tot augmentant el nombre de cèl·lules Bque formen clústers al voltant de l’illot. Aquest canvi,esperat en NOD arran del genotip diabetogènic, vasorprendre quant a l’agressivitat de l’infiltrat enanimals molt joves, efecte atribuït a la capacitat
proinflamatòria de la citocina. Sorprenentment, elmodel suposadament resistent va manifestar unainsulitis severa, també en animals no diabètics, i de lamateixa composició (figura 5) que el model transgènicNOD. En animals diabètics, com és lògic, es vadetectar un augment de la destrucció insular i unadisminució —gairebé desaparició— del nombre decèl·lules beta.
Figura 4. Immunofluorescència doble en pàncrees d’un animal
RIP-IFN-beta no diabètic. En vermell, tinció per insulina; en verd,
marcador específic. a) pa leucocitari, b) macròfags, c) dendrítiques,
d) cèl·lules B, e) cèl.lules T, f) MHC classe I, i g) MHC classe II.
Figura 5. Immunofluorescència en pàncrees d’un animal NOR RIP-
IFN-beta no diabètic. En vermell, insulina, i en verd, marcador
específic. a) pa leucocitari, b) macròfags, c) cèl·lules T CD4+, d)
cèl·lules T CD8+, e) cèl·lules B, f) MHC classe I, i g) IFN-beta humà.
Detecció d’IFN-beta humà en sèrum, pàncrees itimus. Efecte en la tolerància centralEl producte del transgèn (citocina) es va detectar(mitjançant ELISA) en el sèrum dels animals transgè-nics però no en els controls. La concentració d’interfe-ró beta humà en sèrum va ser de 48 UI/ml ± 9(mitjana± SD) i lògicament indetectable en els controlsno transgènics.
Els experiments per quantificar els nivells de molècu-les autoantigèniques es van dur a terme mitjançant latècnica RT-PCR a temps real, una de les méssensibles del moment. Com era d’esperar, es vadetectar RNA corresponent a preproinsulina I (la formamenys expressada i representada dels dos gensd’insulina que tenen els ratolins) únicament en pàncre-
RECERCA BIOMÈDICA | 217

es i no en timus ni en fetge. El nivell d’expressió no vadiferir significativament entre animals transgènics icontrols, tot confirmant que el transgèn no afectava lasíntesi d’insulina (taula 3). La preproinsulina II (genmés expressat i representat) es va detectar enpàncrees, timus i fetge sense diferències entretransgènics i controls. RNA corresponent a IFN-betahumà es va detectar i quantificar en pàncrees i timusdels individus transgènics positius. L’expressió debeta-2 microglobulina en pàncrees va ser significativa-ment més alta en animals transgènics que en controls(p<0,05), probablement com a efecte de l’interferó.Per contra, l’expressió de beta-2 microglobulina altimus va ser la mateixa en individus transgènics queen controls.
En resum, es conclou que l’expressió d’IFN beta entimus no altera l’expressió d’MHC de classe I ni altresmolècules autoantigèniques, la qual cosa fa pensarque la tolerància tímica (central) no està afectada. Pelque fa als illots, les dades de l’expressió d’insulinarefermen les dades metabòliques i demostren que lasíntesi i secreció d’insulina no està afectada peltransgèn en si. Aquestes dades també van a favor delcaràcter autoimunne de la malaltia.
Per tot això, podem dir que els illots de Langerhansdels ratolins transgènics tenen un perfil d’expressiónormal d’insulina I, insulina II i glucagó, mentre que lad’MHC de classe I està augmentada. Contràriament,el microambient tímic sembla no estar afectat perl’expressió del transgèn.
Taula 2. Quantificació d’expressió de gens en timus, pàncrees i
fetge en transgènics RIP-IFN-beta i controls. Els valors correspo-
nen al quocient entre el nombre de còpies del producte respecte
de ß-actina. *p<0,05
Subpoblacions limfocitàries
Les dades de citometria de flux ens indiquen que nohi ha diferències significatives en les subpoblacionsdels animals transgènics respecte dels controls pel
que fa als percentatges de cèl·lules B i T (CD4 i CD8).
Transferència d’autoimmunitat per part d’esplenò-cits i limfòcits infiltrants de RIP-huIFNbeta NOD aScid-NODMitjançant la transferència limfocitària de la malaltias’ha demostrat el seu caràcter autoimmune. Es vanutilitzar com a animals donants individus diabèticsRIP-IFN-beta NOD, just al moment de l’inici clínic dela malaltia i que no haguessin estat tractats ambinsulina exògena. Com a receptors es van utilitzarratolins NOD Scid (immunodeficients que no tenencèl·lules T ni B). Un cop processada la melsa iseparats els esplenòcits, se’n va avaluar la viabilitat ies van transferir entre 10-20x106 esplenòcits frescosper receptor. Posteriorment, els animals es van traslla-dar a una sala de quarantena, on es va fer unseguiment cada dos dies (glicèmia i glucosúria) perdeterminar l’evolució de la possible diabetis (figura 6).En cas de diagnosticar-se la malaltia, els animals esvan sacrificar i es van extreure els seus òrgans per aestudis d’immunohistoquímica.
Figura 5. Esquema de la transferència de diabetis autoimmune en
el ratolí transgènic RIP-IFN-beta.
S’ha pogut determinar la transferència de la insulitis ide la diabetis en el 100% dels animals receptors.L’anàlisi histològica dels òrgans indica que la majoriadels illots dels animals diabètics es troben infiltrats(grau d’infiltració severa) i com que el receptor Scid noté cèl.lules B ni T, la procedència de l’infiltrat és deldonant.
Aquest fet demostra la capacitat de les cèl·lules delsistema immune d’aquests animals per transferir lamalaltia en receptors que no expressen IFN-beta enels seus illots, és a dir, que no tinguin afectat l’òrgandiana. Podem afirmar, doncs, que es tracta d’unamalaltia autoimmunitària causada per la presènciad’un un interferó de tipus 1 en la cèl·lula beta.
Conclusions1. Els ratolins trangènics RIP-IFN-beta (CD-1)desenvolupen diabetes a partir de les 8 setmanesd’edat.
2. La glicèmia, insulinèmia, contingut d’insulinapancreàtica i metabolisme de la glucosa (test detolerància) dels animals transgènics, és normalrespecte dels controls en tots els backgroundsgenètics.
FUNDACIÓ LA MARATÓ DE TV3

3. Els illots dels animals transgènics tenen unadimensió superior als controls a causa d’un nombrede cèl·lules més gran. No hi ha diferencies en la taxade replicació, grau d’apoptosi ni mida de les cèl·lulesbeta en els dos grups.
4. Els ratolins transgènics RIP-IFN-beta (CD-1) sanspresenten una insulitis lleu amb predomini de la periin-sulitis. L’infiltrat està format principalment per cèl·lulesT (CD4 i CD8 en proporció 3:1), cèl·lules B imacròfags. Les cèl·lules dendrítiques només s’obser-ven en els illots no infiltrats (prèviament a l’infiltrat). Lamajoria dels illots dels animals transgènics hiperex-pressen MHC de classe I però no de classe II.Aquestes característiques recorden els illots humansal principi de la malaltia.
5. Existeixen almenys 11 microsatèl·lits que enspermeten diferenciar la colònia CD-1 de les soquesNOD i NOR i aquests són de gran utilitat en el procésde canvi de background genètic.
6. L’expressió del transgèn en background NOD iNOR ha resultat en un important augment en laincidència de la malaltia, acceleració del moment depresentació i un atac més agressiu contra els illotsd’aquests animals. La composició cel·lular de l’infiltratés qualitativament similar al model original peròquantitativament més gran.
7. La citocina antiviral IFN-beta té un paper determi-nant en el desenvolupament de la diabetis en el modeltransgènic en els tres backgrounds genètics en estudi.
8. Els limfòcits d’animals diabètics poden transferir lamalaltia a animals sans no transgènics amb igualbackground genètic, fenomen que confirma elcaràcter autoimmunitari de la diabetis.
3. Rellevància i possibles implicacions clíniquesdels resultats finals obtingutsEls treballs d’aquest projecte han contribuït a laposada en marxa del nou estabulari lliure de patògens(SPF, Specific Pathogen Free), com també a laintroducció de les colònies de ratolins Non ObeseDiabetic (NOD) i Non Obese Resistant (NOR) ambmanteniment i cria, per primera vegada a Catalunya.Aquests models són els més utilitzats, en l’àmbitinternacional, per a l’estudi de la diabetis tipus 1,arran de la seva semblança amb el model humà i lamanca d’accessibilitat al teixit diana (illots) enpacients. Al nostre estabulari s’ha establert aquestacolònia (manteniment i cria) des de l’any 2000, comtambé els models transgènics generats i detallats enaquesta memòria. Aquest fet és rellevant perquè obreles portes a altres possibles projectes de recerca queutilitzin aquests models i a futures col·laboracions ambaltres grups.
Els resultats obtinguts en aquest estudi contribueixena completar el coneixement del paper de l’IFN-beta enla immunologia de la diabetis. Les nostres dadesrefermen la hipòtesi vírica com a mecanisme desenca-denant de la malaltia i demostren que un factor compot ser una citocina antiviral pot trencar la tolerànciaperifèrica i causar autoimmunitat. D’altra banda, es
demostra que el factor genètic de predisposició oprotecció a la malaltia és important, però que per sisol no determina la presentació de l’autoimmunitatcontra la cèl·lula beta. Les dades podrien aplicar-se ala immunoteràpia específica basada en l’administracióde bloquejants de citocines (anticossos monoclonals,receptors solubles...) en el model animal i, a la llarga,en humans (immunoteràpia en una fase concreta de laprediabetis), tot i que caldrien estudis complementaris.
4. PublicacionsRevisions
Alba A, Verdaguer J, Vives-Pi M. Diabetes tipo 1: autoinmuni-
dad frente a la célula beta. Endocrinologia (en premsa, 2004)
Articles originalsA. Alba, J. Carrillo, MC Puertas, R. Planas, X. Pastor, R.
Ampudia, F. Bosch, R. Pujol-Borrell, J. Verdaguer, M. Vives-Pi.
Acceleration of type 1 diabetes in NOD and NOR transgenic mice
expressing IFN-beta in islet beta cells. (enviat per ser publicat).
A.Alba, J. Carrillo, MC. Puertas, R. Planas, X. Pastor, R.
Ampudia, C. Mora, R. Pujol-Borrell, J. Verdaguer, M. Vives-Pi.
PCR-based microsatellite analysis and genetic monitoring for
differentiation of mice strains Non Obese Diabetic and Non Obese
Resistant after backcrosses with transgenic mice. (en preparació).
RECERCA BIOMÈDICA | 219

FUNDACIÓ LA MARATÓ DE TV3
MEMÒRIA FINAL
1. Resum del projecte originalLa diabetis mellitus tipus 2 és una malaltia crònica queafecta aproximadament 100 milions de persones.L’existència d’una alta incidència de diabetis tipus 2en certes poblacions i en familiars directes depacients diabètics de tipus 2, com també l’altaconcordança en bessons, són grans evidències quefan pensar que hi ha factors genètics que confereixensusceptibilitat a aquesta malaltia. Una característicacentral de la diabetis mellitus tipus 2 és la resistènciaa la insulina detectada en múscul esquelètic, la qualrepresenta un marcador primerenc i heretat del fenotipdiabètic de tipus 2. Al nostre laboratori hem identificatrecentment dos gens que poden tenir un paperimportant en les alteracions associades a la diabetismellitus: 3.1 i 44.2. Ambdós gens mostren unaexpressió abundant en teixit muscular, i aquestapateix una reducció significativa en el múscul esquelè-tic de pacients amb diabetis mellitus tipus 2. Calassenyalar que el gen 44.2, el cDNA del qual encarano coneixem, no ha estat descrit prèviament;igualment, es desconeix la funcionalitat biològica delgen 3.1. A partir d’aquests resultats, el nostre projecte
pretén determinar la implicació d’aquests gens en lafisiopatologia de la diabetis mellitus tipus 2, avaluar-neel paper funcional, i també continuar la identificaciód’altres gens associats a la resistència a la insulina enel múscul esquelètic de pacients diabètics de tipus 2.Aquest objectiu es pretén desenvolupar en les fasessegüents:
a) Anàlisi funcional del gen 3.1 en múscul i determina-ció de l’impacte de la repressió d’aquest gen en ladiabetis mellitus tipus 2.
b) Identificació del cDNA complet de 44.2, determina-ció del seu paper en la diabetis mellitus tipus 2 ianàlisi funcional.
c) Clonatge i identificació de gens expressats demanera diferencial en el múscul de pacients diabèticsde tipus 2 respecte de no diabètics.
En resum, el nostre projecte pretén comprendre labase molecular de la fisiopatologia de la diabetismellitus, identificar els gens implicats en la resistènciaa la insulina, i també descobrir noves dianes farmaco-lògiques que permetin el desenvolupament de
Títol del projecte
identificació i anàlisi funcional de gens implicats en la fisiopatologia de la diabetis mellitus tipus 2
Investigador principalAntonio Zorzano Olarte
InstitucióFacultat de Biologia. UB
131.415 €

teràpies futures que es puguin emprar per al tracta-ment i/o la prevenció de la diabetis mellitus tipus 2.
Els objectius específics d’aquest projecte són elssegüents:1. Anàlisi funcional del gen 3.1 en múscul i determi-nació de l’impacte de la seva repressió en la diabetismellitus tipus 2En aquest punt es pretén determinar el paper biològicdel gen 3.1 en condicions normals, i també l’impactede reprimir-lo en l’activitat del múscul. Per aconseguir-ho, s’obtindrà: a) informació sobre la localitzaciótissular del seu producte gènic, per a la qual cosas’obtindran anticossos contra la proteïna 3.1, i esdeterminarà l’expressió muscular en pacients obesos ien subjectes amb resistència a la insulina; b) s’analit-zaran els presumibles canvis metabòlics obtinguts encèl·lules musculars L6E9 —que en condicionsnormals no expressen 3.1— després de la sobreex-pressió d’aquest gen, c) s’obtindrà la seqüència de laregió 5’ no codificant propera a l’inici de transcripciódel gen 3.1 en ratolí (el cDNA de ratolí es troba en lesbases de dades), i d) es produiran ratolins KO per algen 3.1 i s’avaluarà l’impacte sobre la sensibilitat a lainsulina.
2. Identificació del cDNA complet del gen 44.2,repercussió en la diabetis mellitus tipus 2 i anàlisi dela funció gènicaEn aquest apartat es duran a terme estudis semblantsals de la secció anterior amb l’única diferència queprimerament s’ha de fer el clonatge i la seqüenciaciódel cDNA complet del gen 44.2, atès que actualmentnomés hem identificat al voltant d’1 kb de l’extrem 3’.Els estudis previstos seran els següents: a) clonatge iseqüenciació del cDNA complet del gen humà de44.2, b) seqüenciació del gen murí de 44.2 i anàlisi dela regió cromosòmica propera a l’origen de transcrip-ció, c) localització tissular del producte gènic de 44.2 iexpressió en pacients obesos i en subjectes ambresistència a la insulina, d) s’analitzaran els presumi-bles canvis metabòlics obtinguts en cèl·lulesmusculars L6E9 —que expressen 44.2 endogen—després de la sobreexpressió d’aquest gen o d’ungen antisentit. Com que en aquest punt encaramanca molta informació bàsica, no és previsible quedurant el temps del projecte puguem generar ratolinsKO per al gen 44.2; amb tot, aquest és un objectiuprioritari.
3. Clonatge i identificació de gens expressats demanera diferencial en el múscul de pacientsdiabètics de tipus 2 respecte dels no diabèticsPer fer-ho emprarem preparacions d’RNA provinentsde múscul esquelètic de rates controls, obesesZucker (fa/fa) i obeses diabètiques (rates ZDF/Drt-fa).És altament esperable que l’ús de tècniques dedifferential display permeti la detecció de més gensamb expressió alterada al múscul durant la diabetis.Els gens identificats d’aquesta manera passaran a unsegon nivell d’estudi, en què s’analitzarà si la diabetishumana (tipus 2) afecta la seva expressió muscular.Els gens amb expressió diferencial en múscul depacients diabètics tipus 2 seran identificats per seqüen-ciació parcial i se n’obtindran els clons complets en elcas de ser identificats gens desconeguts.
2. ResultatsEls resultats més rellevants obtinguts durant aquestprojecte són els que s’esmenten tot seguit. Cal dirque el gen que anomenem inicialment 3.1, actualmentes coneix com mitofusina-2 o amb l’abreviatura Mfn2,i així ens hi referirem en aquest resum. Igualment, elgen 44.2, actualment l’anomenem DOR i ho faremtambé d’aquesta manera en el resum. Com aconseqüència de la nostra activitat de recerca, hempublicat 11 articles científics.
1) Anàlisi funcional del gen Mfn2 en múscul ideterminació de l’impacte de la seva repressió enel metabolisme mitocondrialA) Resum global de la seccióEn molts tipus cel·lulars i especialment en el múscul,les mitocòndries formen filaments allargats o un reticleramificat. En aquest estudi hem identificat que laproteïna Mfn2 (mitofusina-2), una proteïna demembrana mitocondrial, participa en la fusió mitocon-drial en cèl·lues de mamífer, s’indueix durant lamiogènesi i contribueix al manteniment i l’operació dela xarxa mitocondrial. La repressió d’Mfn2 va provocarla fragmentació morfològica i funcional de la xarxamitocondrial en unitats més petites i independents.D’una manera concomitant, la repressió d’Mfn2 vaprovocar la reducció de l’oxidació de glucosa, delpotencial de membrana mitocondrial, de la respiraciócel·lular i del leak de protons en la mitocòndria. Tambéhem demostrat que aquest mecanisme dependentd’Mfn2 de control del metabolisme mitocondrial s’alte-ra en obesitat per disminució en l’expressió d’Mfn2.En resum, les nostres dades indiquen que l’expressiód’Mfn2 és crucial en el metabolisme mitocondrial permitjà del manteniment de l’arquitectura de la xarxamitocondrial, i que la reduïda expressió d’Mfn2 pot serque expliqui algunes de les alteracions metabòliquesassociades a l’obesitat.
B) ResultatsMfn2 s’expressa en abundància en múscul i s’indueixen la miogènesi
En un cribratge dissenyat amb l’objectiu d’identificargens expressats de manera diferencial en ratesobeses Zucker per mitjà de la tecnologia de sostrac-ció de cDNA PCR-select, vam aïllar un clon de 321parells de bases (que vam anomenar clon C31), elqual va hibridar amb una espècie d’ARNm de 4.7-kbmRNA en múscul esquelètic. La seqüència de cDNAdel clon C31 va ser idèntica a la seqüència de rataU41803 del GenBankTM entre els nucleòtids 2098 i2419, i 79% idèntica a la seqüència de cDNA humàD86987 sotmesa per Nagase et al. al GenBankTM iidentificada recentment com mitofusina (Mfn2).L’alineament múltiple d’Mfn2 humana amb altresproductes gènics va mostrar un 95% d’identitat ambla proteïna Mfn2 de rata (U41308), un 94% d’identitatamb Mfn2 de ratolí (número d’accés GenBankTMAY028170), i un 47% d’identitat amb Mfn2 deDrosophila melanogaster (número d’accésGenBankTM AF355475). Es va trobar menor identitatamb els productes gènics de fzo de D. melanogaster(31% d’identitat) i de Saccharomyces cerevisiae (8%d’identitat). La distribució tissular de l’ARNm d’Mfn2humà es va examinar per anàlisi de Northern blot en
RECERCA BIOMÈDICA | 221

assajos d’alta severitat. Es va observar una bandad’ARNm de 4.7 kb que va hibridar amb el clon de 321parells de bases. Els transcrits van ser més abundantsen múscul esquelètic i cor, i en menor abundància esvan detectar en cervell, ronyó i fetge (figura 1A). Unperfil semblant de distribució es va detectar en teixitsde rata (resultats no mostrats).
Sobre la base de l’abundància relativa de l’ARNmd’Mfn2 en múscul esquelètic, vam examinar si la sevaexpressió es regulava durant la miogènesi. Elsmioblastos L6E9 gairebé no expressen Mfn2, i es vadetectar una marcada inducció en els nivells d’ARNmd’Mfn2 durant la diferenciació a miotubs (figura 1B).
Es va detectar una banda de 110-kDa en extractes decor, múscul esquelètic, teixit adipós marró i ronyó enrates amb un anticòs policlonal contra Mfn2; aquestabanda es va rentar específicament amb un excés depèptid immunogènic. La Mfn2 endògena estàaugmentada en fraccions mitocondrials de cor, teixitadipós marró i ronyó, fet que concorda amb lasobreexpressió d’Mfn2 en mitocòndria. En aquestescondicions, podem dir que la miogènesi causa lainducció d’Mfn2 en cèl·lules L6E9.
La inducció d’Mfn2 és paral·lela al desenvolupamentde la xarxa mitocondrial i a la seva funció com unsistema connectat de manera intralumenal
La inducció de l’expressió del gen d’Mfn2 en miotubsL6E9 és paral·lela al desenvolupument d’una extensaxarxa mitocondrial. L’evidència del funcionament de laxarxa mitocondrial com un sistema connectat intralu-menalment en cèl·lules musculars es va obtenir permicroirradiació amb làser. En cèl·lules mioblàstiques,una petita part del filament mitocondrial és desenergit-zat quan un dels clústers compostos per mitocòndriesés danyat per làser. Com a conseqüència de lapèrdua del potencial de membrana mitocondrial, lamitocòndria no es pot tenyir amb JC-1. Com acontrast, la il·luminació d’un únic filament mitocondrialen miotubs L6E9 causa una pèrdua generalitzada delpotencial de membrana mitocondrial per mitjà de laxarxa mitocondrial, tot afectant àrees situades llunydel punt d’irradiació. Aquestes dades indiquen l’exis-tència d’una extensa connectivitat intralumenal queforma la xarxa mitocondrial en miotubs.
Mfn2 controla la generació i el funcionament de laxarxa mitocondrialPer determinar el rol d’Mfn2 en la xarxa mitocondrial,vam fer estudis en què l’expressió d’Mfn2 erasobreexpressada o reprimida. La sobreexpressiód’Mfn2 després d’una transfecció transitòria vadestruir la xarxa en mioblastos L6E9 i les mitocòndriesvan formar clústers al voltant del nucli. Les mitocòn-dries en fibroblastos 10T1/2 també solen estarorganitzades en filaments, però rarament es veuencom orgànuls aïllats. La sobreexpressió d’Mfn2després d’una transfecció transitòria també causa laformació de clústers al voltant del nucli. La microscò-pia electrònica de transferència va revelar que aquestaformació de clústers no és conseqüència de lageneració de mitocòndries gegants, sinó que corres-pon a agregats mitocondrials amb una morfologia
ultraestructural normal. Tant la Mfn2 endògena com lasobreexpressada va ser localitzada en la mitocòndriaper mitjà d’assajos bioquímics i immunofluorescència.Aquestes dades indiquen que la Mfn2 exigeix unesproteïnes addicionals per completar el procés de fusiómitocondrial complet. Per determinar l’impacte de larepressió de l’expressió d’Mfn2, vam inhibir l’expressiód’Mfn2 en cèl·lules musculars. Per fer-ho, vamgenerar un vector adenoviral que codifica per a unaseqüència antisentit de ratolí per a Mfn2 (AS), i es vaemprar per injectar miotubs L6E9. La infeccióadenoviral amb AS va reduir els nivells d’Mfn2 enextractes mitocondrials (un 50% aproximadament) iabsència d’alteracions en l’abundància de porina.Aquest efecte és específic perquè la infecció amb unadenovirus no rellevant, que codifica per a b -galacto-sidasa, no té efecte en l’expressió d’Mfn2. Tant elsmiotubs controls com els que estaven infectats ambun adenovirus irrellevant presentaven una xarxamitocondrial amb filaments llargs que van longitudinal-ment per tota la fibra muscular. En contrast, lescèl·lules transfectades amb l’adenovirus AS mostravendiscontinuïtats en la xarxa mitocondrial. La quantifica-ció ens va indicar que el 82% dels filaments mitocon-drials eren més llargs de 5 m m en les cèl·lulescontrols, mentre que només el 22% dels filamentsmitocondrials totals mostraven aquesta longitud en lescèl·lules AS. La il·luminació de les mitocòndries en elsmiotubs AS està associada a la pèrdua del potencialde membrana mitocondrial que afecta una extensiómolt reduïda de la xarxa mitocondrial comparada ambles cèl·lules controls, tot donant més força a la hipòte-si d’una fragmentació de la xarxa mitocondrial enclústers funcionalment independents.
Mfn2 controla el metabolisme mitocondrialTot seguit vam observar si la reducció de l’expressiód’Mfn2 causava alteracions en el metabolismemitocondrial. La repressió de l’expressió d’Mfn2mediada per l’expressió antisentit d’Mfn2 en miotubsL6E9 va reduir l’oxidació de glucosa un 30% (figura2A). Aquest efecte era específic i la infecció amb unvector adenoviral que codifica per a la ß-galactosidasano va alterar l’oxidació de glucosa (figura 2A). Enaquestes condicions, la repressió d’Mfn2 va reduir elpotencial de membrana mitocondrial (figura 2B).També hem transfectat d’una manera estable cèl·lules10T1/2 amb la seqüència antisentit d’Mfn2 de ratolí.Els nivells d’mRNA i de proteïnes d’Mfn2 eren mésbaixos en els clons antisentit que en les cèl·lules notransfectades o en les cèl·lules transfectades controls(els nivells d’Mfn2 varien en un rang del 32% al 67%dels valors controls en els clons AS). La repressiód’Mfn2 en cèl·lules 10T1/2 també redueix l’oxidacióde glucosa a un 30%. A més, les cèl·lules AS presen-ten una reducció d’un 30% en el consum d’oxigen.En aquestes condicions, el consum acoblat d’oxigenes mostra inalterat, mentre que la respiració lligada alproton leak mitocondrial es mostra significativamentreduïda en cèl·lules AS. Aquestes dades indiquen quela reducció de l’expressió d’Mfn2 causa alteracions enel metabolisme mitocondrial caracteritzat per unareducció del consum d’oxigen cel·lular i una depressióde l’oxidació de la glucosa.
L’expressió d’Mfn2 està reprimida en obesitat
FUNDACIÓ LA MARATÓ DE TV3

El múscul esquelètic en l’obesitat humana estàcaracteritzat per alteracions metabòliques, incloent-hiresistència a la insulina, acumulació de triglicèridsintracel·lulars, processos d’oxidació i termogènesiinduïda per glucosa defectuosa. En alguns modelsd’obesitat en animals, com les rates obeses Zucker oels ratolins ob/ob, el seu múscul esquelètic presentaun perfil metabòlic caracteritzat per una reducció enl’entrada i l’oxidació de la glucosa, alteració delrepartiment dels àcids grassos que s’incorporen alstriglicèrids o oxidats, resistència a la insulina i reducciódel consum d’oxigen. De fet, els mecanismes pelsquals l’obesitat mena a una oxidació deficient delssubstrats musculars es manté sense explicació i sónconcomitants amb activitats mitocondrials normalsd’enzims crucials, com són la piruvat deshidrogenasa,citrat sintasa, succinat deshidrogenasa o ?-hidroxiace-til-CoA deshidrogenasa.
Basant-nos en el rol de la Mfn2 en el metabolismemitocondrial, en vam determinar l’expressió enobesitat. L’expressió de l’mRNA d’Mfn2 en ratesobeses Zucker és un 34% més baixa que en lesprimes. A més, vam estudiar l’expressió del gend’Mfn2 en múscul esquelètic d’humans obesos. Elssubjectes humans eren normoglicèmics i hiperinsulinè-mics i mostraven una disminució del 50% en lacaptació de la glucosa estimulada per insulina(4.9+0.8 i 10.1+1.1mg/kg/min per a subjectes obesosi prims, respectivament) (27). Es va purificar RNA totalde músculs de vastus lateralis, i s’analitzà per mitjàd’una PCR transcriptasa-competitiva-quantitativa permesurar els nivells d’mRNA d’Mfn2. L’expressió era un36% més baixa en els subjectes obesos. Hi ha unarelació temporal negativa lineal entre els nivellsd’mRNA d’Mfn2 i l’índex de massa corporal quantotes les dades estan barrejades.
Després vam examinar si la xarxa mitocondrial esmodifica en el múscul esquelètic en obesitat. Ambaquest objectiu es van obtenir músculs de ratesobeses Zucker i es van processar per microscòpiaelectrònica, i el grau de xarxa mitocondrial es vadonar com la ràtio entre el volum mitocondrial/unitatde superfície mitocondrial. No es van trobar diferèn-cies en el volum total mitocondrial entre rates controlsi obeses, tot indicant que la massa total mitocondrialestà inalterada. Com a contrast, la densitat de lesmitocòndries era més alta en el grup obès (8.2 + 0.5 i11.2 + 0.6 orgànuls/10 µm2 en control i obesos,respectivament, p<0.001). Això suggereix un midamitocondrial més petita i una xarxa mitocondrialfragmentada en obesitat en múscul esquelètic.Seguint aquest punt de vista, la superfície totalmitocondrial era un 42% més alta en el grupd’obesos, i per això el volum mitocondrial per unitatd’àrea era més baix (25%) en obesitat.
2. Identificació del gen DOR, i implicacions en lasíndrome metabòlicaA) Resum global de la seccióLa resistència a la insulina i una acumulació excessivade greix precedeixen el desenvolupament de lasíndrome metabòlica i de diabetis tipus 2. La predis-posició genètica a desenvolupar resistència a la insuli-na encara està en gran part indefinida, però les
observacions de la disminució de la sensibilitat a lainsulina entre parents de subjectes amb diabetis detipus 2 suggereix una associació genètica entre lasensibilitat a la insulina en teixits diana i l’acció de lainsulina. Hem identificat un nou gen, anomenat DOR,que està reprimit en múscul esquelètic en obesitat i endiabetis tipus 2. DOR és un coactivador nuclear queactiva l’activitat transcripcional del receptor del’hormona tiroïdal, receptors de glucocorticoides iPPARg de manera dependent de lligand. Una variantcomuna en el promotor de DOR (-23G/C, rs 2378256)té influència sobre la transcripció de DOR in vitro.Variacions en aquest SNP estan associades, ensubjectes humans, a una reducció en l’expressió deDOR en múscul esquelètic, i mesures de tot el cos desensibilitat a insulina i adipositat. Els nostres resultatsindiquen que DOR és un nou coactivador dereceptors nuclears que tenen un rol important en elmanteniment de la senyalització de la insulina.
B) ResultatsPer identificar els gens responsables de les alteracionsen múscul esquelètic en diabetis associada a obesitati resistència a la insulina, vam fer un cribratge de gensexpressats diferencialment en rates diabètiquesZucker (ZDF) i rates primes no diabètiques per PCR-select cDNA substraction (Clontech PCR-Select). Esva purificar RNA Poly(A)+ del múscul gastrocnemius.Després d’obtenir una llibreria de cDNA "substracted",vam aïllar diversos clons emprant un cribratge diferen-cial per PCR-selection. Un d’aquests clons conteniauna inserció de 260 parells de bases (bp) i vapermetre la detecció d’un mRNA de 4.5 kb (anomenatDOR; Diabetes-and Obesity-Regulated gene) endiversos teixits. Emprant aquest cDNA de 260 bp, esva fer un cribratge en una llibreria de cDNA de corhumà (Stratagene, Palo Alto) i es van aïllar 5 clons, undels quals contenia el cDNA sencer del gen humà deDOR. Els cDNA murí i de rata de DOR es van amplifi-car per seqüenciar-los.
El gen DOR humà codifica una proteïna de 220residus amb un pes molecular predit de 24 kDa i ambun pI 6.6. L’única proteïna homòloga eraTEAP/SIP/p53DINP1, que s’ha descrit que regulal’apoptosi dependent de p53. La identitat total entreles proteïnes de DOR humà i TEAP/SIP/p53DINP1 ésdel 36%. Els polipèptids de DOR humà, rata i ratolíestan ben conservats; l’humà i el de ratolí mostrenuna identitat del 84%, entre l’humà i el de rata n’hi haun 83% i entre el de rata i el de ratolí hi ha un 85%d’identitat. DOR conté una càrrega positiva molt fortaen la seva regió C-terminal, que se suposa que formauna estructura en hèlice alfa. DOR expressat enbacteris es troba en extractes solubles, però no enfraccions enriquides en membranes. La proteïna deDOR en humans, rata i ratolí conté un motiu putatiuLXXLL o caixa NR, la qual media interaccions entrereceptors i coactivadors. La proteïna de DOR humanamostra cinc llocs putatius de fosforilació per a lacaseïnaquinasa en la regió N-terminal de la molècula ien l’extrem C-terminal mostra uns llocs de fosforilacióper a la proteïna C quinasa. DOR també té unsdominis rics en leucines al mig de la proteïna quepodrien participar en la interacció entre proteïnes.La distribución del mRNA de DOR fue analizada
RECERCA BIOMÈDICA | 223

mediante Northern blot de alta astringencia en lasmuestras de RNA obtenidas de tejidos humanos y derata. Los mRNA de 4.5 kb hibridaban con el clon de260 pb. En humanos, los tránscritos eran predomi-nantes en músculo esquelético, corazón y cerebro;también se detectó una baja expresión en tejidoadiposo y otros tejidos. Estos datos son consistentescon los datos obtenidos en rata, en la que se observauna expresión máxima de mRNA de DOR en músculoesquelético y corazón así como también unaexpresión sustancial en tejido adiposo blanco, riñón,cerebro y hígado. La expresión de DOR, determinadapor Northern blot, en músculo esquelético de ratasdiabéticas ZDF se redujo en un 77%, corroborando elensayo de hibridación por sustracción. Este efecto esespecífico y la expresión de otros tránscritos (b -actina) resultó inalterada. Basándonos en estasobservaciones, también analizamos la expresión deDOR en músculo esquelético de sujetos no diabéticosy de diabéticos de tipo 2. En una serie experimental,15 controles (índice de masa corporal=23+1 kg/m2),12 sujetos obesos (índice de masa corporal=34+1kg/m2) y 15 diabéticos de tipo 2 (índice de masacorporal=31+1 kg/m2), se seleccionaron sujetos deedades similares. Los sujetos obesos y los diabéticosde tipo 2 eran hiperinsulinémicos y mostraban unabaja absorción de glucosa estimulada por insulina(4.9+0.5, 3.6+0.5 y 10.1+1.1 mg.kg-1.min-1 paraobesos y diabéticos de tipo 2, respectivamente). Sepurificó RNA total de músculos vastus lateralis desujetos y se realizó un análisis mediante PCR-real-timepara medir los niveles de mRNA de DOR. Laexpresión era un 70% más baja en los sujetos obesosy en los diabéticos comparados con los sujetoscontrol. En otra serie experimental se seleccionaron 9sujetos control (índice de masa corporal= 26+1kg/m2) y 8 diabéticos de tipo 2 no obesos (índice demasa corporal= 28+1 kg/m2) también eran sujetos deedades parecidas. Los diabéticos de tipo 2 mostraronuna reducción de un 31% en la absorción de glucosaestimulada por insulina. La expresión de DOR de losmúsculos de vastus lateralis era un 25% más baja enpacientes diabéticos de tipo 2. Como en las ratasdiabéticas ZDF, así la reducción de la expresión deDOR en músculo es una característica de los estadosde resistencia a la insulina en humanos
La distribució de l’mRNA de DOR va ser analitzadaper mitjà de Northern blot d’alta astringència en lesmostres d’RNA obtingudes de teixits humans i derata. Els mRNA de 4.5 kb hibridaven amb el clon de260 pb. En humans, els transcrits eren predominantsen múscul esquelètic, cor i cervell; també es detectàuna expressió baixa en teixit adipós i en altres teixits.Aquestes dades són consistents amb les dadesobtingudes en rata, en què s’observa una expressiómàxima d’mRNA de DOR en múscul esquelètic i cor, itambé una expressió substancial en teixit adipósblanc, ronyó, cervell i fetge. L’expressió de DOR,determinada per Northern blot, en múscul esquelèticde rates diabètiques ZDF es va reduir un 77%, totconfirmant l’assaig d’hibridació per sostracció. Aquestefecte és específic i l’expressió d’altres transcrits (b -actina) va resultar inalterada. Basant-nos en aquestesobservacions, també vam analitzar l’expressió deDOR en múscul esquelètic de subjectes no diabètics i
de diabètics de tipus 2. En una sèrie experimental, 15controls (índex de massa corporal 23+1 kg/m2), 12subjectes obesos (índex de massa corporal =34+1kg/m2) i 15 diabètics de tipus 2 (índexs de massacorporal =31+1 kg/m2), es van seleccionar subjectesd’edats semblants. Els subjectes obesos i elsdiabètics de tipus 2 eren hiperinsulinèmics i mostravenuna baixa absorció de glucosa estimulada per insulina(4.9+0.5, 3.6+0.5 i 10.1+1.1 mg.kg-1.min-1 per aobesos i diabètics de tipus 2, respectivament). Es vapurificar RNA total de músculs vastus lateralis desubjectes i es va fer una anàlisi per mitjà de PCR-real-time per mesurar els nivells d’mRNA de DOR.L’expressió era un 70% més baixa en els subjectesobesos i en els diabètics comparats amb els subjec-tes controls. En una altra sèrie experimental es vanseleccionar 9 subjectes controls (índex de massacorporal= 26+1 kg/m2) i 8 diabètics de tipus 2 noobesos (índex de massa corporal = 28+1 kg/m2)també eren subjectes d’edats semblants. Els diabèticsde tipus 2 van mostrar una reducció d’un 31% enl’absorció de glucosa estimulada per insulina.L’expressió de DOR dels músculs de vastus lateralisera un 25% més baixa en pacients diabètics de tipus2. Com en les rates diabètiques ZDF, la reducció del’expressió de DOR en múscul és una característicadels estats de resistència a la insulina en humans.
Hi ha dues línies d’evidència que suggereixen queDOR participa en la regulació transcripcional: laprimera és un lloc putatiu LXXLL que podria mediar eninteraccions entre el receptor nuclear i coactivadors, ien la segona, DOR mostra homologia amb la proteïnanuclear TEAP/SIP/p53DINP1, el qual modula la funcióde p53. Per avaluar si DOR regula l’activitat delreceptor d’hormona nuclear, es van transfectar transi-tòriament cèl·lules HeLa amb receptor de l’hormonatiroïdal TRa1. Aquest receptor s’uneix a un fragmentde DNA anomenat enhancer de GLUT4. La cotrans-fecció en cèl·lules HeLa de l’hormona tiroïdal Tralfa1,DOR i l’enhancer de GLUT4 específic de músculfusionat amb el gen reporter CAT, va provocar queTRalfa1 transactivés l’enhancer de GLUT4 6 vegades,mentre que DOR en solitari només té un efecte petit(figura 3A). La cotransfecció de DOR i TRa1 remarcal’activitat transcripcional de l’enhancer de GLUT4muscular d’una manera dosidependent (figura 3B).Aquest efecte és específic de l’expressió de DORperquè la transfecció amb un vector buit o amb unplasmidi que codifica per a una proteïna irrellevant nocausa cap efecte. L’efecte de DOR és inhibit per lamutació de la caixa LXXLL i depèn de la presència dellligand T3 (figura 3A). DOR també activa de maneradosidependent la transactivació del receptor deglucocorticoïdes en presència de lligand (dexametaso-na) i és inhibit per la mutació en la caixa LXXLL(figures 3C i D). A més, DOR també augmenta latransactivació de PPARgd’una manera dosidependenti dependent de lligand (figures 3E i F).
Tot seguit vam passar a determinar si DOR estàlocalitzat al nucli cel·lular. Per fer-ho, vam generar unaproteïna de fusió de DOR amb una proteïna verdafluorescent (GFP) i l’expressió es va limitar al nucli. Encanvi, la proteïna nadiua GFP es va trobar tant alcitosol com al nucli. Això es va confirmar per mitjà de
FUNDACIÓ LA MARATÓ DE TV3

l’anàlisi de Western blot d’extractes rics en nuclis. Perdeterminar si DOR i el receptor d’hormona nuclearinteraccionen físicament, es van produir proteïnesquimèriques de fusió TRa1-GST, GR-GST i DOR-His.TRa1-GST però no GST s’uneix a DOR i la interacciófísica en assajos pull-down és independent de lapresència de T3 en el medi. D’una manera semblant,GR-GST s’uneix a DOR en assajos pull-downindependent de dexometasona.
Vam concloure que DOR està localitzat al nucli i queté l’habilitat d’unir receptors hormonals tiroïdals ireceptors de glucocorticoides i després activar l’activi-tat transcripcional dels receptors d’hormona tiroïdal,PPARg i receptors de glucocorticoides, la qual cosasuggereix una funció com a coactivador nuclear. Ésprobable que la funció com a coactivador dependentde lligand tingui com a causa el fet que la unió deDOR al receptor d’hormona nuclear és regulada encondicions in vivo per mecanismes absents enassajos pull-down. A més, DOR estimula la transacti-vació de l’enhancer específic de múscul GLUT4 permitjà de receptors d’hormona tiroïdal, la qual cosasuggereix un rol de sensibilització a la insulina.
El gen humà de DOR conté 5 exons, malgrat que unsplicing alternatiu du a 5 variants de transcripció. En eltranscrit més llarg, la regió codificadora de la proteïnacomença a l’exó 3. La reseqüenciació de la seqüènciaexònica en 20 individus espanyols no diabètics,juntament amb la inspecció de CELERA i les bases dedades públiques no va permetre identificar cap variantcomuna dintre de la seqüència codificant de DOR,però va revelar una variant comuna (DOR1,rs2378256) localitzada 23pb a 5’ de l’inici detranscripció (predit i confirmat per 5’RACE) dintre de laseqüència fortament conservada entre humans iratolins (87% d’identitat). Per examinar l’efecte de lavariació de DOR1 en l’activitat del promotor es vanclonar fragments de diverses longituds del promotorde DOR amb un al·lel G o bé C en DOR1 upstreamdel gen reporter de la luciferasa. Es van provar 4construccions, en total: una contenia el gen reporterperò cap element promotor, una altra contenia elpromotor d’SV40 per controlar el gen reporterd’expressió, una altra contenia un fragment de 2.3 kbamb l’al·lel C de DOR1 i una darrera contenia l’al·lel Gde DOR1 per conduir l’expressió del gen reporter.L’activitat luciferasa estava clarament reduïda en l’al·lelG respecte del C, tant en fibroblastos 10T1/2 com encèl·lules musculars L6E9 en els constructes de 2.3 i2kb. La reducció de l’activitat de la luciferasa en l’al·lelG varia des d’un 42% en cèl·lules 10T1/2 a un 88%en cèl·lules musculars L6E9. Aquests efectes es vanobservar amb quantitats diferents de cDNA deconstructes del reporter, amb un rang de 0.3 a 1.3µg. Per eliminar la possibilitat que aquests efectesfossin causats per diferències no detectades prèvia-ment en la seqüència, vam intercanviar el fragment derestricció de 220pb (posició -77/+146) que contenia elpolimorfisme de DOR1 entre els constructes de l’al·lelG i l’al·lel C. El clon que anteriorment contenia un al·lelC, ara conté un al·lel G (Cà G) i viceversa (Gà C). Vamtrobar que l’activitat transcripcional de l’al·lel C eraidèntica al grup Gà C i que l’activitat en l’al·lel Gtambé era semblant a la del grup Cà G. Per determi-
nar més clarament el paper de DOR1 en l’expressiógènica de DOR, vam obtenir biòpsies de múscul iDNA de subjectes homes sans (BMI= 24+0.5, edat=48+2). Els subjectes CC de DOR1 mostraven nivellsalts d’mRNA de DOR, els quals eren significativamentmés grans que els nivells detectats en subjectes GG.En tots els casos, aquestes dades ens indiquen queel polimorfisme de DOR1 determina el grau d’expres-sió de DOR.
Sobre la base d’aquestes dades funcionals i perestablir si la reducció de l’expressió en múscul deDOR vista en obesos i/o diabètics podia contribuir a laresistència a la insulina, vam buscar si hi havia algunacorrelació entre la variació de DOR1 i els fenotipshumans en subjectes europeus. Per fer aquestsestudis, ens vam concentrar en la sensibilitat a lainsulina com el fenotip intermedi més rellevant i enmesuraments de l’adipositat en diabetis tipus 2 comun possible fenotip distal.
En tres poblacions (espanyola, anglesa i polonesa),vam trobar evidències que els individus homozigots,per a l’al·lel G tenien una sensibilitat reduïda a la insuli-na. En 163 homes espanyols de mitjana edat, els quetenien el genotip GG (n=54) eren més resistents a lainsulina (P=0.016) basat en valors de Si (índexs desensibilitat a la insulina) derivats d’un test de tolerànciaa la glucosa intravenosa.
En sintonia amb la resistència a la insulina, aquestsindividus GG també tenien uns nivells significativamentmés alts de triglicèrids (P=0.04), i també de nivellsd’àcid úric (P=0.04). En la mostra de subjectes sansanglesos de l’Oxford Biobank (419 dones, 354 homes),el genotip GG es va associar amb la resistència a lainsulina (mesuraments mitjançant el model homeostàticde valoració [HOMA]) en dones (P=0.01) però no enhomes (P=0.63). Per acabar, en una mostra de 96subjectes polonesos no diabètics (55 dones, 41homes), els nivells d’insulina post-OGTT eren significati-vament superiors (P=0.016, en total, P=0.018 enhomes, P=0.27 en dones) en individus GG.
Hi havia certes proves per a una associació entre elgenotip del DOR1 i mesuraments d’adipositat. Peraixò, en la població espanyola els subjectes GGmostraven un increment més gran en el BMI durantun període de 7 anys (P=0.019, taula 1). En donesangleses, el genotip GG s’associa amb un plecsuprailíac més gran (P=0.009), subescapular (P=0.02)i de bíceps (P=0.04), amb un comportament similar enhomes però no significatiu (taula 1). En els polonesos,les dones GG tenien el percentatge més alt de greixcorporal (P=0.08, dades no mostrades). Quan elsmesuraments del BMI van ser emprats per classificartots els subjectes com a prims (<25kgm-2) o ambsobrepès (<25kgm-2), el genotip GG estava significati-vament associat a un excés d’adipositat (Cochran-Mantel-Haenszel test, p=0.008).
En cap dels estudis hi havia cap relació entre el genotipde DOR1 i les concentracions de glucosa en dejuni.Les freqüències del genotip de DOR1 eren similars en582 subjectes diabètics de tipus 2 i en 556 controls.L’associació entre el genotip GG de DOR1 i la
RECERCA BIOMÈDICA | 225

reducció de la sensibilitat a la insulina en tres estudisseparats és consistent amb les dades funcionals queindiquen que l’al·lel G redueix la transcripció de DOR iimplica una expressió reduïda de DOR en el desenvo-lupament de la resistència a la insulina. Les associa-cions amb increment de l’adipositat exigeixen unaconfirmació en cohorts més grans, però són consis-tents amb una contribució de la variació de DOR a unincrement de l’adipositat. Un error per detectar unaassociació significativa entre mesuraments de glicèmiai la diabetis tipus 2 podria reflectir un "poder" estadís-tic inadequat i la complexitat del fenotip.
Respecte del paper específic de DOR, hem demostratque és un coactivador nuclear i que afavoreix l’activitattranscripcional del receptor de l’hormona tiroïdal, elreceptor de glucocorticoides i PPARg de modelliganddependent i caixa NR-dependent. In vitro, DORs’uneix al receptor d’hormona tiroïdal o al receptor deglucocorticoides de manera lliganddependent, totsuggerint que tant la interacció funcional exigeix unióde lligand o que en condicions in vivo, DOR s’uneix areceptors nuclears hormonals, amb una unió prèvia delligand. Basant-nos en l’activitat de DOR en estudis detransfecció transitòria, vam postular que DORconfereix especificitat als efectes del receptord’hormona tiroïdal, PPARg i receptors de glucocorti-coides en múscul esquelètic, teixit adipós, cor icervell. Hem demostrat més específicament que DORestimula un enhancer de GLUT4 específic de músculsconduït per receptors d’hormones tiroïdals. En conclu-sió, la repressió de DOR també podria regular lasensibilitat a la insulina mitjançant la modulació dePPARg o l’activitat del receptor de glucocorticoides encèl·lules adiposes.
Diverses observacions suggereixen que la variació deDOR contribueix a la variació de la sensibilitat a lainsulina i l’adipositat en homes: a) l’expressió de DORes redueix en múscul esquelètic de pacients obesos idiabètics de tipus 2; b) la variant del promotor deDOR1 té influència sobre la transcripció in vitro i estàassociada a l’expressió de DOR en múscul in vivo; i c)la mateixa variant està associada a la sensibilitat a lainsulina i l’adipositat. L’associació de les dadessuggereixen que els canvis en l’expressió de DOR nosón només una conseqüència de la resistència a lainsulina, sinó que també la reducció de l’expressió deDOR està associada a la variació de DOR1, quecontribueix al desenvolupament de la resistència a lainsulina i, potencialment, a l’adipositat. Per descomp-tat, aquestes troballes en DOR1 no exclouen lapossibilitat que d’altres en DOR o en gens veïnstambé contribueixin a la variació fenotípica humana.Genotipacions addicionals de variants dins o al voltantde DOR indiquen que és una regió de diversitathaplotípica limitada i amb un fort desequilibri delligament. Les disminucions en l’expressió de DOR enrates diabètiques ZDF suggereixen que l’expressió deDOR està regulada addicionalment per la resistència ala insulina o, més probablement, que la variació enl’expressió de DOR és una conseqüència de lasenyalització deficient de leptina, la qual contribueix ala resistència a la insulina i a la diabetis en aquestmodel.
Els nostres resultats suggereixen que DOR és unaproteïna que participa en la fisiopatologia de la síndro-me metabòlica i a més referma la idea que els coacti-vadors nuclears tenen un rol crucial en la diabetishumana o en les alteracions del balanç energètic.Malgrat que només s’ha parat una atenció limitada alrol dels coreguladors com a causa de desenvolupa-ment de diabetis, els seus nivells podrien explicar elsdiversos fenotips observats en poblacions humanes.Els coactivadors actuen en el pas d’amplificació del’expressió gènica i, en conseqüència, les petitesdiferències derivades dels nivells de coactivador són labase de variacions més grans en respostes biològi-ques com les referents a l’acció de la insulina o laregulació metabòlica.
3. Identificació de gens expressats de maneradiferencial en pacients diabètics de tipus 2 respectede no diabèticsA) ResumLa proteïna d’adhesió vascular-1 (VAP-1), que ésidèntica a l’amino oxidasa sensible a la semicarbazida(SSAO), és una proteïna de membrana amb duesfuncions: una amb propietats d’adhesió i una altraamb activitat amino oxidasa. Una forma soluble deVAP-1 es troba en sèrum, on les concentracionsestan augmentades en diabetis i obesitat. In vitro,VAP-1 soluble facilita la capacitat d’adhesió delimfòcits que està implicada en complicacions cardio-vasculars associades a diabetis i obesitat. Enambdues, el teixit d’origen de VAP-1/SSAO soluble ésdesconegut. Nosaltres hem examinat si el teixitadipós, en el qual hi ha una expressió abundant deVAP-1/SSAO, és una font de VAP-1 soluble.
Tant els animals diabètics com els obesos tenen unincrement de l’activitat de SSAO en plasma associadaa la proteïna VAP-1. També trobem que els adipòcits3T3-L1 i els explants de teixit adipós humà alliberenuna forma soluble de VAP-1/SSAO, que deriva de lamembrana. L’alliberament de la forma soluble de VAP-1 es veu facilitada per l’exposició d’adipòcits murins ihumans a TNFa, però resulta bloquejat per batimastat,un inhibidor de metal·loproteases. L’ablació parcial delteixit adipós redueix l’activitat de SSAO en sèrum derates normals i diabètiques. Les cèl·lules adiposes sónuna font de VAP-1/SSAO soluble alliberat per mitjà deproteòlisi de la forma de membrana. L’alliberament deSSAO està regulat per TNFa i insulina. Per l’allibera-ment de VAP-1/SSAO, les cèl·lules adiposes podriencontribuir a l’aterogènesi i a la disfunció vascularassociada amb la diabetis i l’obesitat.
B) Resultats
La diabetis i obesitat incrementen la quantitat de VAP-1/SSAO en plasma
Rates diabètiques induïdes per estreptozotocina, unmodel de diabetis tipus 1, tenen incrementada l’activi-tat de SSAO en plasma. També vam trobar que elsratolins diabètics de tipus 1 tenien un nivell d’activitatde SSAO plasmàtica més alt (figura 4b). L’ús de l’anti-còs TK7-88 contra un ratolí de VAP-1 que és capaçd’immunoprecipitar la proteïna VAP-1 d’adipòcit deratolí (figura 4c) mostrava una activitat SSAO més alta
FUNDACIÓ LA MARATÓ DE TV3

en els immunoprecipitats de plasma de ratolinsdiabètics respecte dels controls (figura 4b). Ambaquestes condicions, l’anticòs TK7-88 va immunopre-cipitar el 79±7% de l’activitat total de SSAO enplasma. També vam trobar un augment de l’activitatde SSAO en plasma en rates diabètiques Goto-Kakizaki (figura 4a).
Per determinar si l’obesitat/resistència a la insulinaaltera l’activitat de SSAO en absència de diabetis,vam comparar l’activitat de SSAO en rates de 3mesos Zucker genèticament obeses (fa/fa) amb lesrates controls primes (Fa/?). Les rates obeses Zuckerpresentaven resistència a la insulina però no diabetis, iles concentracions de glucosa en plasma en els dosgrups no eren diferents (8.5±0.5 i 7.8±0.4 mmol/l perals animals obesos i prims, respectivament). Enaquestes condicions, l’activitat SSAO en plasmaaugmentà en el grup obès d’una manera significativa.
Adipòcits 3T3-L1 i explants humans de teixit adipósalliberen activitat SSAO en el medi
L’activitat SSAO va ser detectada en el medi, totaugmentant de manera tempodependent (figura 5a).Després d’1 h de contacte amb adipòcits diferenciats,el medi mostrava una activitat SSAO no significativa.Després de 24 h, es va detectar activitat SSAO(141±19 pmol·h—1·plate—1), que es va doblar endos dies (278±13 pmol·h—1·plate—1). Els valors dela Km de l’oxidació de la bencilamina catalitzada pelmedi i les preparacions de membranes (43±8 µmol/l i25±5 µmol/l, respectivament) (figura 5b) indiquen queles dues activitats estan íntimament relacionades.Respecte dels adipòcits humans estudiats, vam trobarque després de 48 h, el medi dels explants humansmés rellevants mostraven també una activitat SSAOincrementada (figura 5c).
Els adipòcits 3T3-L1 produeixen una forma soluble deVAP-1/SSAO diferent de la forma integral de membrana
Paral·lelament a l’increment dependent de temps del’activitat de la SSAO soluble, la quantitat de VAP-1/SSAO present en el medi també s’incrementava.SSAO va ser detectat en poc menys d’1 h. Desprésd’una incubació de 48 h, la quantitat de proteïnas’incrementa un 220% (n=4) respecte del valor a les24 h. Com la forma de membrana, la proteïna solubleés dimèrica en condicions no reductores, peròmonomèrica després del tractament amb ditiotreitol.Els adipòcits indiferenciats, que no expressen SSAOde superfície, no alliberen cap activitat enzimàtica deSSAO detectable o proteïna SSAO.
Una possible contaminació del medi recollit i concen-trat de membranes d’adipòcits que contenen SSAOens va fer pensar d’eliminar aquest medi, perquè: I) noes detectava en aquest medi ni GLUT4 ni caveolina-1,mentre que les dues proteïnes es detectaven enextractes de 3T3-L1 (figura 3a), i II) com s’observa encondicions reductores, SSAO en el medi era aproxi-madament 2 M més petita que en els adipòcits.
També vam determinar si l’activitat de SSAO i laproteïna de VAP-1 estaven associades al medi de
cultiu de les cèl·lules 3T3-L1. La immunoprecipitacióde VAP-1 provinent del medi de cultiu de les cèl·lules3T3-L1 mostrava que, en aquestes condicions, mésd’un 70% de l’activitat de SSAO també era present enels immunoprecipitats. A més, vam trobar una fortacorrelació (r=0.7, p<0.001) entre la quantitat de VAP-1i l’activitat de SSAO detectada en 19 medis de cultiude 3T3-L1 obtinguts d’experiments separats.
La SSAO soluble i la forma integral de membranapresenten nivells similars d’N-glicosilació i sialització
La SSAO secretada pels adipòcits 3T3-L1 també estàN-glicosilada i sialitzada. Els tractaments amb endogli-cosidasa F o amb sialidasa provoquen un canviidèntic en la motilitat tant en la forma de membrana isoluble de SSAO (corresponent a canvis de 3 i 15 Mrdesprés del tractament amb endoglicosidasa F i lasialidasa, respectivament).
L’alliberament de SSAO pels adipòcits és dependentde l’activitat metal·loproteasa
Tot seguit vam estudiar si l’alliberament de SSAO pelsadipòcits resultava d’un procés proteolític. També vamestudiar l’efecte de la matriu de l’inhibidor demetal·loproteases en els nivells de SSAO secretada.Mentre que en les concentracions de SSAO demembranes d’adipòcits, no patien cap efecte amb eltractament, la presència de batismat reduïa l’allibera-ment de SSAO a 20±1% del control. L’activitat en elmedi també es va reduir un 20% respecte del control.L’alliberament de l’activitat de SSAO per explants deteixit adipós humà va ser completament inhibida perbatmastat.
L’alliberament de SSAO pels adipòcits està facilitadaper TNFa
Per a un estudi més a fons de la possible regulació del’alliberament de SSAO soluble per factors implicatsen la fisiopatologia del teixit adipós i diabetis, es vancultivar adipòcits 3T3-L1 en presència o absència dediverses hormones o metabòlits durant 48 h.Normalment, els adipòcits 3T3-L1 es cultiven en unmedi ric en glucosa (25 mmol/l glucosa). Per al nostrepropòsit, els adipòcits es van incubar durant 4 dies enun medi que només tenia 5 mmol/l de glucosa.L’alliberament de SSAO no va quedar afectat. Comque vam veure un augment de SSAO en ratesobeses, vam cultivar 3T3-L1 durant 48 h en presènciade 5 i 50 ng/ml de leptina. Aquesta citoquina no vamodificar l’alliberament de VAP-1/SSAO. La citoquinaTNFa va fer minvar el contingut de SSAO en extractesd’adipòcits, però es va trobar un augment de laproteïna VAP-1/SSAO i també de la seva activitatenzimàtica en el medi d’incubació. Com en lescondicions control, l’alliberament de VAP-1/SSAOinduïda per TNFa era dependent de l’activitat de l’acti-vitat de la matriu de metal·loproteases: un 63% del’alliberament de SSAO es va inhibir per batismatat.Igual que en les cèl·lules 3T3-L1, TNFa causa l’esti-mulació de l’alliberament de SSAO per explantshumans. També vam determinar si la producció deperòxid d’hidrogen per SSAO podria tenir algunainfluència en l’alliberament de VAP-1/SSAO. Les
RECERCA BIOMÈDICA | 227

cèl·lules tractades amb bencilamina (50 µmol/l), soleso en combinació amb TNFa, no van mostrar capefecte de la bencilamina per si sola, malgrat queinhibeix parcialment l’alliberament de VAP-1/SSAOinduïda per TNFa.
L’ablació parcial del teixit adipós intern de rata redueixla quantitat de SSAO en el plasma
El nostre examen de l’impacte en plasma de VAP-1/SSAO del teixit adipós en rates vives Wistar vamostrar que en el grup de rates a què s’havia practi-cat l’ablació, l’activitat de SSAO va minvar un 85% iera, significativament, respecte del grup no operat(p=0.037). Les diferències entre els animals a quès’havia fet l’ablació va esdevenir més gran 3 diesdesprés de la injecció d’estreptozotocina.
3. Rellevància i possibles implicacions clíniquesdels resultats finals obtingutsEls gens mitofusina-2 i DOR semblen molt rellevantsdes del punt de vista de la fisiopatologia de la diabetisi/o de l’obesitat.
Així, la mitofusina-2 es reprimeix en múscul esquelèticde pacients amb diabetis tipus 2, com també ensubjectes obesos. A més, la repressió d’Mfn2condueix a una disminució en la capacitat de lacèl·lula per oxidar glucosa i per mantenir potencial demembrana mitocondrial normal. Actualment estemanalitzant la possibilitat que la mitofusina-2 sigui ungen de susceptibilitat a la diabetis tipus 2.
DOR és un gen que codifica per a un coactivadornuclear i coactiva receptors nuclears d’hormones,com PPARg, el receptor d’hormones tiroïdals i elreceptor de glucocorticoides. Hem descrit l’existènciade variants funcionals del gen de DOR en la poblacióeuropea i que un genotip determinat sembla queexerceix susceptibilitat al desenvolupament deresistència a la insulina i a l’acumulació de greix.Per últim, el gen SSAO/VAP-1 codifica per a una
FUNDACIÓ LA MARATÓ DE TV3

proteïna bifuncional amb activitat amino oxidasa i ambpropietats de proteïna d’adhesió cel·lular. Aquestaproteïna s’expressa molt abundantment en l’adipòcit ies localitza en la membrana plasmàtica. Determinatsestímuls, com TNFa, provoquen el trencamentd’aquesta proteïna i l’alliberament subsegüent del’anomenada forma soluble de SSAO/VAP-1. Ladiabetis i l’obesitat menen a un increment en laconcentració circulant de proteïna soluble SSAO/VAP-1 com a conseqüència d’un increment en el seualliberament en teixit adipós.
Les implicacions clíniques són tan notables queaquests resultats han dut a la sol·licitud de trespatents diferents, la qual cosa ha estat gestionat perl’Oficina de Patents de la UB.
4. PublicacionsN. Morin, J.M. Lizcano, E. Fontana, L. Martí, F. Smih, P. Rouet,
D. Prevot, A. Zorzano, M. Unzeta, C. Carpéné. Semicarbazide-
sensitive amine oxidase substrates stimulate glucose transport
and inhibit lipólisis in human adipocytes. Journal of Pharmacology
and Experimental Therapeutics 297, 563-572, 2001.
L. Martí, A. Abella, C. Carpéné, M. Palacín, X. Testar, A.
Zorzano. Combined treatment with benzylamine and low doses of
vanadate enhances glucose tolerance and reduces hyperglycemia
in streptozotocin-induced diabetic rats. Diabetes 50, 2061-2068,
2001.
Consenso del grupo de trabajo "Resistencia a la Insulina" de la
Sociedad Española de Diabetes. La resistencia a la insulina y su
implicación en múltiples factores de riesgo asociados a diabetes
tipo 2. Medicina Clínica 119, 458-463, 2002.
A. Zorzano, A. Abella, L. Martí, C. Carpéné, M. Palacín, X.
Testar. Semicarbazide-sensitive amine oxidase activity exerts insulin-
like effects on glucose metabolism and insulin-signaling pathways in
adipose cells. Biochim. Biophys. Acta 1647, 3-9, 2003.
A. Abella, L. Martí, M. Camps, M. Claret, J. Fernández-Alvarez,
R. Gomis, A. Gumà, N. Viguerie, C. Carpéné, M. Palacín, X.
Testar, A. Zorzano. SSAO/VAP-1 activity exerts an anti-diabetic
action in Goto-Kakizaki rats. Diabetes 52, 1004-1013, 2003.
D. Bach, S. Pich, F.X. Soriano, N. Vega, B. Baumgartner, J.
Oriola, J.R. Daugaard, J. Lloberas, M. Camps, J.R. Zierath, R.
Rabasa-Lhoret, H. Wallberg-Henriksson, M. Laville, M. Palacín, H.
Vidal, F. Rivera, M. Brand, A. Zorzano. Mitofusin-2 determines
mitochondrial architecture and mitochondrial metabolism: a novel
regulatory mechanism altered in obesity. J. Biol. Chem. 278,
17190-17197, 2003.
Zorzano A. Terapia génica en endocrinología: una aproxima-
ción realista. Endocrinología 50, 244 — 249, 2003.
A. Abella, L. Martí, C. Carpéné, M. Palacín, X. Testar, Zorzano A.
Stimulation of glucose transport by semicarbazide-sensitive amine
oxidase activity in adipocytes from diabetic rats. J. Physiol.
Biochem. 59, 225-232, 2003.
Stolen CM, Madanat R, Marti L, Kari S, Yegutkin GG, Sariola
H, Zorzano A, Jalkanen S. Semicarbazide-sensitive amine oxidase
overexpression has dual consequences: insulin mimicry and
diabetes-like complications. FASEB J. 18, 702-704, 2004.
Abella A, García-Vicente S, Viguerie N, Ros-Baro A, Camps M,
Palacin M, Zorzano A, Martí L. Adipocytes release a soluble form
of VAP-1/SSAO by a metalloprotease-dependent process and in
a regulated manner. Diabetologia 47, 429-38, 2004.
Visentin V, Prevot D, De Saint Front VD, Morin-Cussac N,
Thalamas C, Galitzky J, Valet P, Zorzano A, Carpene C. Alteration
of Amine Oxidase Activity in the Adipose Tissue of Obese
Subjects. Obes. Res. 12, 547-555, 2004.
RECERCA BIOMÈDICA | 229




















