
Measurement of Anaerobic Capacities in Humans: Definitions, Limitations and Unsolved Problems
16
REVIEW ARTICLE Sports Medicine 15 (5): 312-327, 1993 0112-1642/93/0005-0312/$08.00/0 © Adis International Limited. All rights reserved. SP01234 Measurement of Anaerobic Capacities in Humans Definitions, Limitations and Unsolved Problems Simon Green and Brian Dawson Department of Human Movement, University of Western Australia, Nedlands, Australia Contents 312 314 314 315 316 317 320 320 321 322 323 323 324 Summary Summary I. Anaerobic Capacity Versus Anaerobic Work Capacity 2. Oxygen Debt 2.1 Alactic Oxygen Debt 2.2 Lactic and Total Oxygen Debt 3. Peak Postexercise Blood Lactate Level 3.1 Blood Lactate: Estimating Alactic Capacity 4. Oxygen Deficit 4.1 The Variability of the Oxygen Deficit 4.2 Methodological Sources of Variance 4.3 Is the Oxygen Deficit a Valid Measure of Anaerobic Capacity? S. Conclusions 6. Future Directions Anaerobic capacity is defined as the maximal amount of adenosine triphosphate resynthesised via anaerobic metabolism (by the whole organism) during a specific mode of short-duration max- imal exercise. This review focuses on laboratory measures which attempt to quantify anaerobic capacity; it examines the evidence supporting or challenging the validity of these measures and provides research foci for future investigations. Discussion focuses on anaerobic capacity meas- ured during running and cycling, since almost all data reviewed were collected using these exercise modes. The validity of the oxygen debts (alactic and total), maximal blood lactate and oxygen deficit as measures of anaerobic capacity was examined. The total oxygen debt, now termed the excess post-exercise consumption, was used in investigations in the 1920s and 1930s to quantify an- aerobic energy production; it has since been shown to be an invalid measure of anaerobic capacity, since its magnitude is known to be influenced by factors (e.g. temperature, catecholamines, sub- strate cycling, lactate glycogenesis) other than those directly involved in anaerobic metabolism. Maximal blood lactate, a measure also used in some of those early investigations, is often used in exercise and sports physiology. Opinion on the utility of maximal blood lactate as an estimate of anaerobic (lactic) capacity is, however, divided. Despite problems interpreting the physiological meaning of maximal blood lactate levels (due primarily to acute changes in blood volume), this measure is still used in both research and athletic settings to describe anaerobic capacity. Its use is supported by (a) the high correlations observed between maximal blood lactate and short-duration exercise performance presumably dependent upon anaerobic capacity, and (b)
-
Upload
independent -
Category
Documents
-
view
1 -
download
0
Transcript of Measurement of Anaerobic Capacities in Humans: Definitions, Limitations and Unsolved Problems

REVIEW ARTICLE
Sports Medicine 15 (5): 312-327, 1993 0112-1642/93/0005-0312/$08.00/0 © Adis International Limited. All rights reserved.
SP01234
Measurement of Anaerobic Capacities in Humans Definitions, Limitations and Unsolved Problems
Simon Green and Brian Dawson Department of Human Movement, University of Western Australia, Nedlands, Australia
Contents
312 314 314 315 316 317 320 320 321 322 323 323 324
Summary
Summary I. Anaerobic Capacity Versus Anaerobic Work Capacity 2. Oxygen Debt
2.1 Alactic Oxygen Debt 2.2 Lactic and Total Oxygen Debt
3. Peak Postexercise Blood Lactate Level 3.1 Blood Lactate: Estimating Alactic Capacity
4. Oxygen Deficit 4.1 The Variability of the Oxygen Deficit 4.2 Methodological Sources of Variance 4.3 Is the Oxygen Deficit a Valid Measure of Anaerobic Capacity?
S. Conclusions 6. Future Directions
Anaerobic capacity is defined as the maximal amount of adenosine triphosphate resynthesised via anaerobic metabolism (by the whole organism) during a specific mode of short-duration maximal exercise. This review focuses on laboratory measures which attempt to quantify anaerobic capacity; it examines the evidence supporting or challenging the validity of these measures and provides research foci for future investigations. Discussion focuses on anaerobic capacity measured during running and cycling, since almost all data reviewed were collected using these exercise modes.
The validity of the oxygen debts (alactic and total), maximal blood lactate and oxygen deficit as measures of anaerobic capacity was examined. The total oxygen debt, now termed the excess post-exercise consumption, was used in investigations in the 1920s and 1930s to quantify anaerobic energy production; it has since been shown to be an invalid measure of anaerobic capacity, since its magnitude is known to be influenced by factors (e.g. temperature, catecholamines, substrate cycling, lactate glycogenesis) other than those directly involved in anaerobic metabolism.
Maximal blood lactate, a measure also used in some of those early investigations, is often used in exercise and sports physiology. Opinion on the utility of maximal blood lactate as an estimate of anaerobic (lactic) capacity is, however, divided. Despite problems interpreting the physiological meaning of maximal blood lactate levels (due primarily to acute changes in blood volume), this measure is still used in both research and athletic settings to describe anaerobic capacity. Its use is supported by (a) the high correlations observed between maximal blood lactate and short-duration exercise performance presumably dependent upon anaerobic capacity, and (b)

Anaerobic Capacities in Humans 313
the higher maximal blood lactate values observed in sprint and power athletes (who would demonstrate higher anaerobic capacities) compared with endurance athletes or untrained people. However, training-induced changes in other performance, physiological and biochemical markers of anaerobic capacity have not always been paralleled by changes in maximal blood lactate; its relatively high variability also diminishes its usefulness to athletic populations, since relatively small changes in anaerobic capacity may not be detected by a measure with such high variability. These latter findings may be partially related to the confounding influence of blood volume which often changes in response to short and long term exercise demands. Maximal blood lactate is known to be influenced by the intensity and duration of the preceding exercise bout; therefore, it is plausible that these factors may also influence the degree to which maximal blood lactate accurately reflects anaerobic capacity.
The oxygen deficit was used in several Scandinavian investigations during the early years of this century; its potential to quantify anaerobic capacity regained momentum in the early 1970s, and at present it is claimed by some to be the only measure with the potential to quantify anaerobic capacity. The validity of the oxygen deficit is supported by: (a) a quantitative similarity with anaerobic capacity determined by the change in anaerobic metabolites in muscle and blood; (b) higher values in sprint athletes than in endurance athletes and untrained people; and (c) the fact that it is increased following high-intensity training expected to improve anaerobic capacity, and changes in parallel with in vitro determinants of anaerobic capacity. Its validity, however, is based on several assumptions. One of these, that the oxygen demand at high exercise intensities can be estimated via the linear extrapolation of the submaximal V02-workload relationship, is tenuous: it may be a major source of error in determining the oxygen deficit and thus quantifying anaerobic capacity. A universal method of determining the submaximal V02-workload relationships has also not been established and warrants further investigation. Moreover, the majority of evidence which supports the validity of the oxygen deficit has been collected using untrained people. It is not known to what degree such evidence can be extrapolated to well-trained athletic populations. Therefore, future investigations should also focus on the use of the oxygen deficit in well-trained athletic populations.
In conclusion, the oxygen debts should not be used to quantify anaerobic capacities. Maximal blood lactate can only, at best, reflect anaerobic capacity; applied sport scientists should also consider the time and cost involved in measuring this variable, given that the monitoring of short and long term changes in blood volume will also be required to increase the sensitivity of maximal blood lactate to changes in anaerobic capacity. The oxygen deficit is the only measure which can potentially quantify anaerobic capacity; it has been claimed to accurately quantify anaerobic capacity. Evidence presented in this review suggests, however, that its validity, especially in athletic populations, is not clearly established and requires further examination.
A considerable research effort has focused on evaluating the power and capacity of the anaerobic metabolic systems in humans. The impetus for this research originated from studies completed early this century and was continued in the work of several European research groups between 1950 and 1970. This research effort, in part, originated from the recognition that anaerobic power and capacity are important to short-duration exercise performance characterised by the generation and/or maintenance of very high power outputs.
In contrast to anaerobic power, anaerobic capacity has proved to be a difficult metabolic con-
struct to measure. This difficulty may be partially related to the lack of a universal definition of anaerobic capacity.
Tests of an anaerobic capacity can be classified according to whether they attempt to quantify an anaerobic capacity, or provide a work estimate of an anaerobic capacity. This review focuses on the former tests with particular emphasis on both maximal blood lactate, because of its widespread but controversial use, and the oxygen deficit, because of its recent popularity within applied and theoretical sports physiology. The measures of oxygen debt are also evaluated, and the comparative util-

314
ity of all measures (i.e. oxygen deficit, oxygen debt and maximal blood lactate) are assessed.
1. Anaerobic Capacity Versus Anaerobic Work Capacity
The anaerobic resynthesis of adenosine triphosphate (ATP) occurs via the hydrolysis of high-energy phosphagens ('" P: e.g. phosphocreatine, adenine nucleotides) and the anaerobic catabolism of carbohydrate (i.e. glycogenolysis/glycolysis). The total amount of ATP replenished via ",P hydrolysis during activity has been defined as the alactic capacity (di Prampero 1981). Since lactate accounts for ",90% of all end products generated via the anaerobic catabolism of carbohydrate (Bangsbo et al. 1990), the total amount of A TP resynthesised via this pathway may be termed the lactic capacity (di Prampero 1981). The sum of these 2 capacities yields the anaerobic capacity, defined as the maximum amount of ATP resynthesised via anaerobic metabolism (by the whole organism) during a specific mode of short-duration maximal exercise.
The relative contributions ofthe alactic and lactic capacities to anaerobic capacity generated during exhaustive exercise over several minutes approximate 20 and 80%, respectively (Bangsbo et al. 1990; Saltin 1987). The relative importance of these metabolic capacities to exercise performance will differ according to the intensity and duration of the performance: the alactic capacity would be important to activities characterised by the maintenance of maximum power output over "'5 to 15 sec, whereas the lactic capacity and the mechanisms that determine it become more important as the duration of the maximal activity is increased (up to several minutes). Anaerobic capacity per se would only be important in those events during which it is nearly completely maximised (for example, 800m run, 1000m cycle pursuit, sustained sprint finishes over slightly longer distances). For athletes competing in such events, it is important that the anaerobic capacities are measured accurately to correctly assess the athlete's functional state and to subsequently recommend the appropriate training.
Mechanical work output which is primarily de-
Sports Medicine 15 (5) 1993
pendent on the anaerobic capacity may be termed anaerobic work capacity [a term originally proposed by Maison & Broeker (1941 )]. Anaerobic work capacity can be defined as the total amount of work performed during an exhaustive work bout which is of a sufficient duration to maximise the anaerobic ATP yield, given that this A TP yield exceeds that from oxidative metabolism.
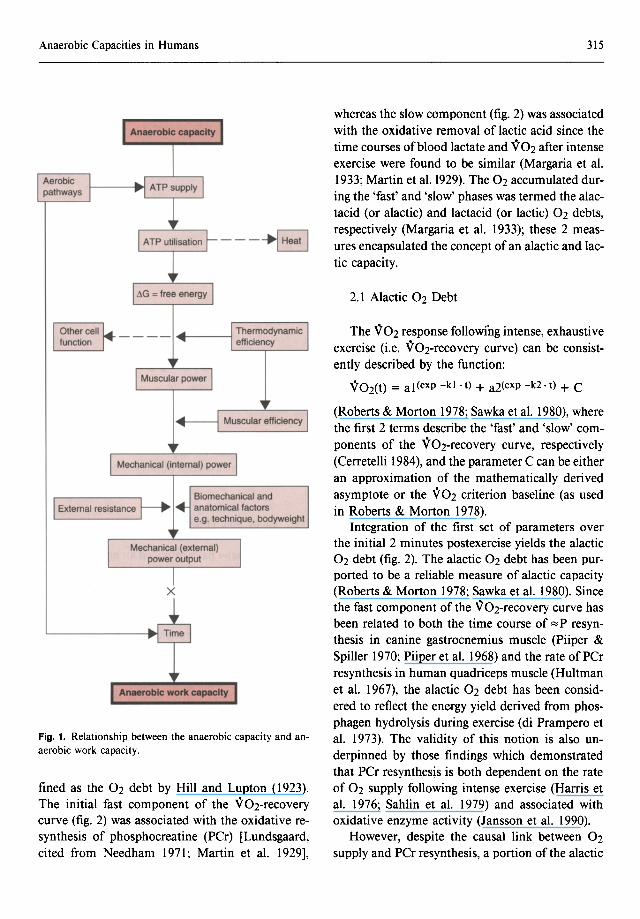
The physical basis underlying the conceptual distinction between the anaerobic capacity and anaerobic work capacity is represented schematically in figure I, and provides the basis for the classification of tests of anaerobic capacity. That is, tests attempt to provide either a quantitative estimate of an anaerobic capacity, or yield a work estimate of an anaerobic capacity, i.e. measure an anaerobic work capacity. Both anaerobic and aerobic metabolism supply the chemical input (i.e. free energy liberated via the hydrolysis of A TP required for the cross-bridge cycling, muscle tension development and ultimately muscle power). For any given chemical input, muscle power will depend on muscle efficiency [a term defined by Woledge (1989) as 'the ratio of work done during one muscle contraction to the total free energy dissipation during contraction, relaxation and recovery of the muscles to their original state']. The subsequent translation of muscle power to mechanical, or external, power output is determined by factors which influence biomechanical efficiency during the movement. The anaerobic work capacity is the product of the time over which this mechanical power output is maintained. For a given anaerobic capacity, the relative contribution of aerobic metabolism to A TP supply will also influence the anaerobic work capacity by affecting the time over which a given power output is sustained; alternatively, if the duration (time) of exercise is held constant, a greater aerobic contribution to A TP supply will increase the anaerobic work capacity. Thus, estimates of anaerobic work capacity are dependent on several factors and can only, at best, reflect anaerobic capacity.
2. Oxygen Debt
The total amount of oxygen consumed during recovery (i.e. area between a resting V02 baseline and the V02-recovery curve) was originally de-
Anaerobic Capacities in Humans
---~
x
Fig. 1. Relationship between the anaerobic capacity and anaerobic work capacity.
fined as the 02 debt by Hill and Lupton (1923). The initial fast component of the V02-recovery curve (fig. 2) was associated with the oxidative resynthesis of phosphocreatine (PCr) [Lundsgaard, cited from Needham 1971; Martin et al. 1929],
315
whereas the slow component (fig. 2) was associated with the oxidative removal of lactic acid since the time courses of blood lactate and V02 after intense exercise were found to be similar (Margaria et al. 1933; Martin et al. 1929). The 02 accumulated during the 'fast' and 'slow' phases was termed the alactacid (or alactic) and lactacid (or lactic) 02 debts, respectively (Margaria et al. 1933); these 2 measures encapsulated the concept of an alactic and lactic capacity.
2.1 Alactic 02 Debt
The V02 response following intense, exhaustive exercise (i.e. V02-recovery curve) can be consistently described by the function:
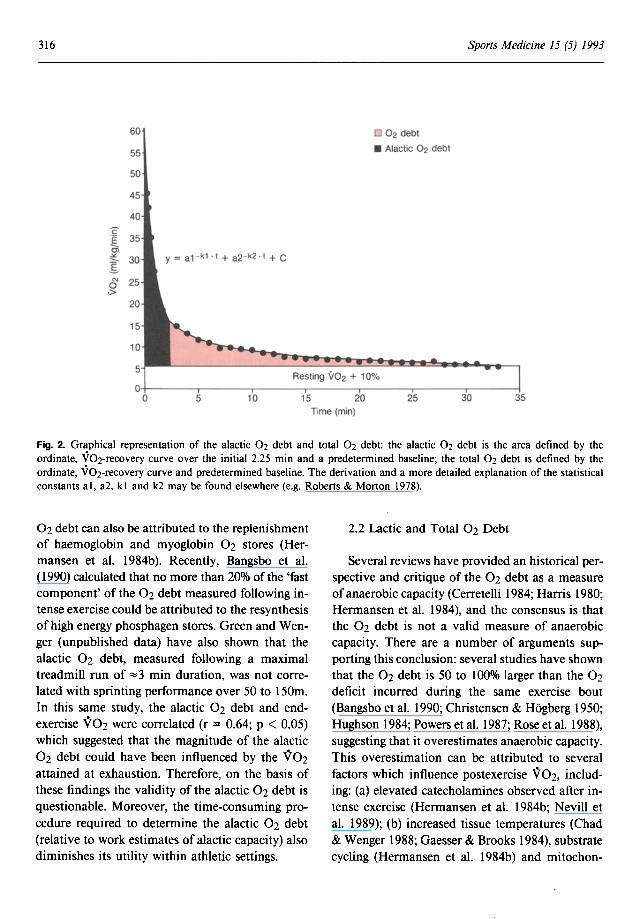
V02(t) = al (exp -kl . t) + a2(exp -k2· t) + C
(Roberts & Morton 1978; Sawka et al. 1980), where the first 2 terms describe the 'fast' and 'slow' components of the V02-recovery curve, respectively (Cerretelli 1984), and the parameter C can be either an approximation of the mathematically derived asymptote or the V02 criterion baseline (as used in Roberts & Morton 1978).
Integration of the first set of parameters over the initial 2 minutes postexercise yields the alactic 02 debt (fig. 2). The alactic 02 debt has been purported to be a reliable measure of alactic capacity (Roberts & Morton 1978; Sawka et al. 1980). Since the fast component of the V02-recovery curve has been related to both the time course of ~ P resynthesis in canine gastrocnemius muscle (Piiper & Spiller 1970; Piiper et al. 1968) and the rate of PCr resynthesis in human quadriceps muscle (Hultman et al. 1967), the alactic 02 debt has been considered to reflect the energy yield derived from phosphagen hydrolysis during exercise (di Prampero et al. 1973). The validity of this notion is also underpinned by those findings which demonstrated that PCr resynthesis is both dependent on the rate of 02 supply following intense exercise (Harris et al. 1976; Sahlin et al. 1979) and associated with oxidative enzyme activity (Jansson et al. 1990).
However, despite the causal link between 02 supply and PCr resynthesis, a portion of the alactic
316
c :[ Ol ~
I N o .>
Sports Medicine 15 (5) 1993
60 0 02 debt
• Alactic 02 debt
y = a1 - k1 . t + a2- k2 . t + C
Time (min)
Fig. 2. Graphical representation of the alactic 02 debt and total 02 debt: the alactic 02 debt is the area defined by the ordinate, V02-recovery curve over the initial 2.25 min and a predetermined baseline; the total 02 debt is defined by the ordinate, V02-recovery curve and predetermined baseline. The derivation and a more detailed explanation of the statistical constants ai, a2, kl and k2 may be found elsewhere (e.g. Roberts & Morton 1978).
02 debt can also be attributed to the replenishment of haemoglobin and myoglobin 02 stores (Hermansen et al. I 984b). Recently, Bangsbo et al. (1990) calculated that no more than 20% of the 'fast component' ofthe 02 debt measured following intense exercise could be attributed to the resynthesis of high energy phosphagen stores. Green and Wenger (unpublished data) have also shown that the alactic 02 debt, measured following a maximal treadmill run of "'3 min duration, was not correlated with sprinting performance over 50 to 150m. In this same study, the alactic 02 debt and endexercise V02 were correlated (r = 0.64; p < 0.05) which suggested that the magnitude of the alactic 02 debt could have been influenced by the V02 attained at exhaustion. Therefore, on the basis of these findings the validity of the alactic 02 debt is questionable. Moreover, the time-consuming procedure required to determine the alactic 02 debt (relative to work estimates of alactic capacity) also diminishes its utility within athletic settings.
2.2 Lactic and Total 02 Debt
Several reviews have provided an historical perspective and critique of the 02 debt as a measure of anaerobic capacity (Cerretelli 1984; Harris 1980; Hermansen et al. 1984), and the consensus is that the 02 debt is not a valid measure of anaerobic capacity. There are a number of arguments supporting this conclusion: several studies have shown that the 02 debt is 50 to 100% larger than the 02 deficit incurred during the same exercise bout (Bangsbo et al. 1990; Christensen & Hogberg 1950; Hughson 1984; Powers et al. 1987; Rose et al. 1988), suggesting that it overestimates anaerobic capacity. This overestimation can be attributed to several factors which influence postexercise V02, including: (a) elevated catecholamines observed after intense exercise (Hermansen et al. 1984b; Nevill et al. 1989); (b) increased tissue temperatures (Chad & Wenger 1988; Gaesser & Brooks 1984), substrate cycling (Hermansen et al. 1984b) and mitochon-

Anaerobic Capacities in Humans
drial respiration (Gaesser & Brooks 1984) after exercise; (c) the type of substrate used (Chad & Wenger 1988); and (d) the resynthesis of glycogen from lactate (La') following intense exercise (Astrand et al. 1986; Hermansen & Vaage 1977; Nordheim & V 011estad 1990) which occurs at an energetic cost twice that released during La' formation (Martin 1987). Hagberg et al. (1990), using MCArdle's disease patients, demonstrated a complete dissociation between La' production during exercise and the slow 'lactacid' component of the V02-recovery curve to provide conclusive evidence of the spurious use of the lactic 02 debt to describe the lactic capacity. When the abovementioned problems inherent in the alactic 02 debt are also considered, it can be concluded that the total 02 debt does not quantify anaerobic capacity.
3. Peak Postexercise Blood Lactate Level
Margaria, di Prampero and colleagues (e.g. Margaria et al. 1963, 1964, 1971) had consistently used blood La' (Lab) values to estimate the energy liberated via La' production. Di Prampero (1981) had argued that the net accumulation of La' within the body during an exercise bout is quantitatively related to the level of La' within the blood several minutes after exercise, if assumptions are made relating to the diffusion space for La' and the distribution kinetics of La' throughout this space. When these assumptions are met, the peak postexercise blood La' (Labmax) is considered to provide an accurate and reliable quantitative measure of lactic capacity, as long as the recovery mode and site of blood sampling are standardised (di Prampero 1981).
The effect of the site of blood sampling on Labmax has been verified by other investigators (Foxdal et al. 1990; Graham 1984). Lactate values determined from femoral blood samples can be twice as high as those values determined from cephalic or antecubital venous samples (Graham 1984). Finger capillary Lab has also been reported to be "'8% higher than the Lab determined from forearm venous samples (Foxdal et al. 1990) but
317
similar to values determined from arterialised earlobe samples (Telford, unpublished observations).
It has also been established that La' removal from the circulation is increased during active, as opposed to passive, recovery (Belcastro & Bonen 1975; Hermansen & Stensvold 1972; McGrail et al. 1978). Maximal Lab is also influenced by preexercise glycogen levels (Hepburn & Maughan 1982; Jacobs 1987; Klausen et al. 1975) and acid-base balance changes induced by prior exercise (Kindermann & Kullmer 1985; Klausen et al. 1972). Therefore, the site used for blood sampling must be consistent between testing sessions, and blood sampling must be made during a passive recovery. Before testing sessions, efforts should be made to ensure that the subjects have fully recovered from previous training sessions (48 to 72 hours) and that this recovery period is standardised for all testing periods. Tests which will induce prolonged disturbances in body acid-base status should not be implemented before lactic capacity tests, or if multiple tests must be administered during one testing session, the sequence of tests must be standardised.
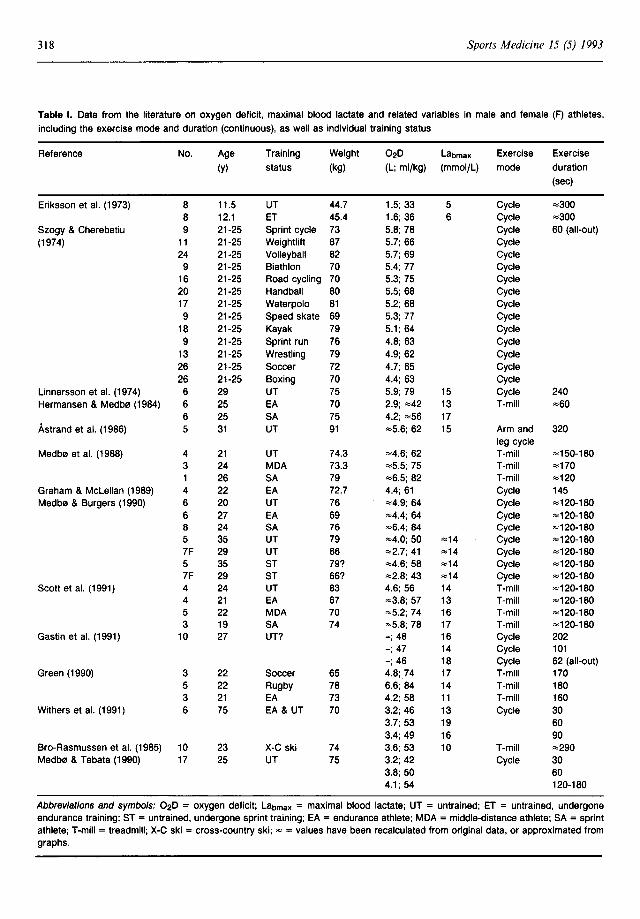
Maximal Lab has been used to provide both a qualitative (Lacour et al. 1990; Mero 1988; Ohkuwa et al. 1984; Paterson et al. 1986) and quantitative (Camus & Thys 1991; di Prampero 1981) measure of anaerobic capacity in athletic and nonathletic populations. Significant relationships between Labmax and performances dependent on anaerobic capacity have been demonstrated: Labmax was significantly correlated with running performances over 400m (r = -0.76 to -0.89) [Berg & Keul 1985; Fujitsuka et al. 1982; Lacour et al. 1990; Ohkuwa et al. 1984] and 800m (r = -0.71) [Lacour et al. 1990], treadmill running over 30 to 60 sec (r = 0.67 to 0.86) [Cheetham & Williams 1987; Cheetham et al. 1986; Fujitsuka et al. 1982] and total work output on the Wingate test (r = 0.55) [Tamayo et al. 1984]. Higher Labmax values have been reported for sprint and power athletes than endurance athletes or untrained individuals (Komi et al. 1977) [table I], and Labmax has also been significantly correlated (r = 0.24), albeit weakly, with the proportion of type II fibres in skeletal muscle of athletes (Komi et al. 1977). Maximal Lab values as
318 Sports Medicine 15 (5) 1993
Table I. Data from the literature on oxygen deficit, maximal blood lactate and related variables in male and female (F) athletes,
including the exercise mode and duration (continuous), as well as individual training status
Reference No. Age Training Weight 02D Labmax Exercise Exercise
(y) status (kg) (L; ml/kg) (mmol/L) mode duration
(sec)
Eriksson et al. (1973) 8 11.5 UT 44.7 1.5; 33 5 Cycle ",,300
8 12.1 ET 45.4 1.6; 36 6 Cycle ",,300
Szogy & Cherebetiu 9 21-25 Sprint cycle 73 5.8; 78 Cycle 60 (all-out)
(1974) 11 21-25 Weightlift 87 5.7; 66 Cycle 24 21-25 Volleyball 82 5.7; 69 Cycle
9 21-25 Biathlon 70 5.4; 77 Cycle 16 21-25 Road cycling 70 5.3; 75 Cycle 20 21-25 Handball 80 5.5; 68 Cycle 17 21-25 Waterpolo 81 5.2; 68 Cycle 9 21-25 Speed skate 69 5.3; 77 Cycle
18 21-25 Kayak 79 5.1; 64 Cycle 9 21-25 Sprint run 76 4.8; 63 Cycle
13 21-25 Wrestling 79 4.9; 62 Cycle 26 21-25 Soccer 72 4.7; 65 Cycle 26 21-25 Boxing 70 4.4; 63 Cycle
Linnarsson et al. (1974) 6 29 UT 75 5.9; 79 15 Cycle 240 Hermansen & Medb0 (1984) 6 25 EA 70 2.9; ",,42 13 T-mill ",,60
6 25 SA 75 4.2; ""56 17 Astrand et al. (1986) 5 31 UT 91 ""5.6; 62 15 Arm and 320
leg cycle Medb0 et al. (1988) 4 21 UT 74.3 ""4.6; 62 T-mill ""150-180
3 24 MDA 73.3 ""5.5; 75 T-mill ",,170
26 SA 79 ""6.5; 82 T-mill ""120 Graham & McLellan (1989) 4 22 EA 72.7 4.4; 61 Cycle 145 Medb0 & Burgers (1990) 6 20 UT 76 ""4.9; 64 Cycle ""120-180
6 27 EA 69 ""4.4; 64 Cycle ""120-180 8 24 SA 76 ""6.4; 84 Cycle ""120-180 5 35 UT 79 ""4.0; 50 ""14 Cycle ""120-180 7F 29 UT 66 ""2.7; 41 ""14 Cycle ""120-180 5 35 ST 79? ""4.6; 58 ""14 Cycle ""120-180 7F 29 ST 66? ""2.8; 43 ""14 Cycle ""120-180
Scott et al. (1991) 4 24 UT 83 4.6; 56 14 T-mill ""120-180 4 21 EA 67 ""3.8; 57 13 T-mill ""120-180 5 22 MDA 70 ""5.2; 74 16 T-mill ""120-180 3 19 SA 74 ""5.8; 78 17 T-mill ""120-180
Gastin et al. (1991) 10 27 UT? -; 48 16 Cycle 202 -; 47 14 Cycle 101 -; 46 18 Cycle 62 (all-out)
Green (1990) 3 22 Soccer 65 4.8; 74 17 T-mill 170 5 22 Rugby 78 6.6; 84 14 T-mill 180 3 21 EA 73 4.2; 58 11 T-mill 160
Withers et al. (1991) 6 75 EA & UT 70 3.2; 46 13 Cycle 30 3.7; 53 19 60 3.4; 49 16 90
Bro-Rasmussen et al. (1985) 10 23 X-C ski 74 3.6; 53 10 T-mill ",,290 Medb0 & Tabata (1990) 17 25 UT 75 3.2; 42 Cycle 30
3.8; 50 60 4.1; 54 120-180
Abbreviations and symbols: 02D = oxygen deficit; Labmax = maximal blood lactate; UT = untrained; ET = untrained, undergone endurance training; ST = untrained, undergone sprint training; EA = endurance athlete; MDA = middle-distance athlete; SA = sprint athlete; T-mill = treadmill; X-C ski = cross-country ski; "" = values have been recalculated from original data, or approximated from graphs.

Anaerobic Capacities in Humans
high as 32 mmoljL reported for elite oarsmen (McNaughton & Cedaro 1991) are consistent with the idea that the lactic and anaerobic capacity of these athletes would be among the highest of all athletes. Moreover, Labmax has been shown to improve as a result of supra-V02max training regimens (Cunningham & Faulkner 1969; Houston & Thomson 1977; Jacobs et al. 1987; Ready et al. 1981; Sharp et al. 1986; Sjodin et al. 1976) and seasonal competition (Green & Houston 1975) expected to improve lactic capacity. Maximal Lab has also been shown to be reliable (r = 0.87) when measured ~7 min after a short-duration, exhaustive cycle test (Crielaard et al. 1986).
Despite these findings, however, the validity of Labmax is controversial and the accuracy with which it reflects anaerobic capacity in high performance athletes has been challenged (Saltin 1987, 1990; Schnabel et al. 1979). Saltin (1987) has questioned the validity of the assumptions (see di Prampero 1981) underlying the use of Labmax to quantify lactic capacity: it was suggested that since a large portion of the La' produced has been metabolised after exercise before an equilibrium in La' distribution between. muscle and blood has been achieved, Labmax cannot provide an accurate quantitative estimate of lactic capacity. Rieu et al. (1988) also demonstrated that the original assumption made by Margaria et al. (1963) relating to the free and rapid diffusion of lactate throughout the water compartments of the body was not valid for intermittent exercise; Rieu et al. (1988) showed that the Lab values observed throughout intermittent exercise could only be approximated by calculation if the time for La' to reach equilibrium throughout the body compartments was assumed to be much longer than that originally proposed by Margaria et al. (1963). These investigators suggested that the 'caloric equivalent' for Lab of 3ml 02/mmol La' calculated by Margaria et al. (1963) must be interpreted with caution.
The kinetics of Lab, and consequently the magnitude of Labmax following submaximal exercise, are dependent on the intensity and duration of the exercise (Freund et al. 1986, 1989). Although a similar dependence of Labmax on the intensity and
319
duration of the exercise at sUpra-V02max intensities has not been demonstrated directly, there is evidence to suggest that Lab kinetics may differ between maximal exercise of varying intensity and duration (Rieu et al. 1988). Gastin et al. (1991) recently demonstrated that Labmax values following a 1 min all out cycle test were higher than the corresponding values following maximal constant-load cycle bouts of ~ 1.5 to 3.5 min duration, although the 02 deficits incurred during all tests were similar. Koziris and Montgomery (1991) also showed that Labmax was significantly greater after an allout 90 sec test compared with a maximal constantload test of the same duration, despite the fact that mean power outputs were similar between these tests. These findings suggest that all out tests may yield higher Labmax values than those observed after constant-load tests, and that these differences may be related to the exercise intensity and/or duration.
As for all blood-borne metabolites, differences in blood volume will also influence the Lab levels at any given rate of intracellular accumulation of La' and La' efflux. This effect is especially important when interpreting Labmax values before and after training considering the lability of plasma volume to short and long term exercise (Green et al. 1987; Harrison 1986), detraining or tapering (Coyle et al. 1986; Sheply et al. 1990) and heat acclimatisation (Harrison 1986).
Bro-Rasmussen et al. (1985) demonstrated that training at altitude (2800m) increased the muscle buffer values, 02 deficit and treadmill running performance in well trained crosscountry skiers, although the observed 8% reduction in Labmax failed to parallel these changes indicative of an increased anaerobic capacity. Other studies have demonstrated that high intensity training fails to alter Labmax despite improvements in high intensity performance and/or other metabolic determinants of anaerobic capacity (Cheetham & Williams 1987; Medb0 & Burgers 1990; Nevill et al. 1989; Roberts et al. 1982). Labmax was not correlated with mean power output on the Wingate test (Goslin & Graham 1985; Taunton et al. 1981) and short-distance running performance in well trained athletic popu-

320
lations (Green & Wenger, unpublished observations; Ohkuwa et al. 1984).
These findings may be partially attributed to the high variability of Labmax: coefficients of variation of7.2% (Fujitsuka et al. 1982), 16.5% (Margaria et al. 1963) and 21 % (Graham & Andrew 1973) have been reported for Labmax. The higher coefficients of variation were reported for Labmax measured after exhaustive exercise oflonger duration, whereas the lower value reported by Fujitsuka et al. (1982) was determined following a I min exhaustive run. This lower value is similar to those variabilities reported for V02max (Katch et al. 1982; Kuipers et al. 1985) and anaerobic work capacity measures (Coggan & Costill 1984). It would be of interest to determine the variation in Labmax elicited following exercise of a duration required to maximise anaerobic capacity. All these findings, however, demonstrate the potential problems inherent in using Labmax to accurately estimate lactic capacity on repeated occasions.
3.1 Blood Lactate: Estimating Alactic Capacity
Schnabel and Kindermann (1983) proposed a method of measuring alactic capacity using the change in Lab response to a nonexhaustive, supraV02max treadmill run of 40 sec duration. The assumptions underlying this method were that (a) the majority of the A TP used during this work bout was resynthesised via anaerobic processes, and (b) the magnitude of "'P hydrolysis was inversely and directly proportional to the anaerobic catabolism of carbohydrate - that is, a smaller change in Lab (ilLab) was indicative of a higher alactic capacity. In general, sprint trained athletes (400 to 800m) revealed smaller ilLab values than marathon runners or controls, suggesting that sprinters had higher alactic capacities than marathoners. However, there were no differences in ilLab values between athletes who competed over distances of 400 to 5OO0m, when higher alactic capacities were expected in those athletes who compete over the shorter distances. The validity of this method to estimate alactic capacity is also undermined by the probable influence of the exercise-V02 kinetics on the Lab
Sports Medicine 15 (5) 1993
response, the energetic contribution of the body 02 stores, as well as those abovementioned problems associated with measuring Labmax.
4. Oxygen Deficit
Although the concept of the oxygen deficit (020) was introduced early this century by Krogh and Lindhard (1919/20), it was not until the 1970s that the potential of the 020 to quantify the anaerobic A TP yield during maximal exercise was revealed (e.g. Karlsson & Saltin 1971). In the latter half of the 1980s, several investigations supported the utility of the 020 as a quantitative measure of the anaerobic ATP yield (e.g. Medb0 & Tabata 1989; Medb0 et al. 1988). In a recent review, Saltin (1990) claimed that the 020 is the only method with the potential to quantify anaerobic capacity.
At submaximal exercise intensities, the steadystate V02 is assumed to reflect the total rate of energy expenditure during exercise. On the assumption that the energy expenditure increases linearly with increasing exercise intensity, the 02 demand (i.e. V02) at supra-V02max power outputs can be estimated by linear extrapolation of the V02 power output relationship determined at submaximal power outputs (Hermansen & Medb0 1984). During supra-V02max exercise, the 020 is calculated as the area between the estimated 02 demand and the V02 asymptote over the exercise duration; it is often expressed in 02 equivalents (i.e. ml/kg or L) and is considered to reflect that energy liberated via the hydrolysis of ",P, the anaerobic catabolism of carbohydrate, as well as the use of body oxygen stores present in the lungs, body fluid, haemoglobin and myoglobin (Astrand et al. 1986). Approximately 400 to 600mi of endogenous 02 is used during short duration, maximal exercise, and contributes to the 020 to a minor, but not negligible, extent (Astrand et al. 1986).
Oxygen deficits reported in the literature are displayed in table I. Mean 020 values reported for exercising humans vary from 33 ml/kg in prepubescent males (Eriksson et al. 1973) to 80-85 ml/ kg in sprint athletes (Medb0 & Burgers 1990; Medb0 et al. 1988; Scott et al. 1991). Larger 020

Anaerobic Capacities in Humans
values (i.e. "=' 130 ml/kg) have been reported in trained thoroughbred horses with individual values as high as 168 ml/kg (Rose et al. 1988), values which are much greater than the maximal 020 predicted for humans (i.e. "=' 100 ml/kg: di Prampero 1981; Saltin 1987).
The 020 of endurance athletes (i.e. 50 to 65 ml/ kg: Graham & McLellan 1989; Hermansen & Medb0 1984; Medb0 & Burgers 1990; Scott et al. 1991; Withers et al. 1991) are of a similar magnitude to those reported for untrained subjects (Medb0 & Burgers 1990; Medb0 et al. 1988; Scott et al. 1991; Withers et al. 1991). Athletes with well developed anaerobic and aerobic capabilities (for example, middle-distance runners) incur 02Ds which lie within the range of values for endurance and sprint athletes (i.e. "='75 ml/kg) [Medb0 et al. 1988; Scott et al. 1991].
Comparing data in table I reveals that a considerable interstudy variation in 020 exists when individuals of a similar training status are examined. This variation could be attributed to several factors: the exercise mode and exercising muscle mass; the exercise duration; technological and biological variability; and methodological differences between studies.
Anaerobic capacity is dependent on the amount of muscle mass recruited during exercise, which in turn is a function of the mode of exercise. That 020 values incurred during treadmill running or arm-leg ergometry are consistently higher than those measured during cycling may be explained by the greater musculature recruited during running and arm-leg exercise as opposed to cycling (A strand et al. 1986; Medb0 & Tabata 1990).
The duration of exercise may also partially account for the disparity in 020 values reported between studies. Scandinavian investigators (Hermansen & Medb0 1984; Medb0 et al. 1988) originally demonstrated that increasing the exercise duration (within a period approximating 2 to 3 minutes) increased the 020, with further increases in duration failing to further increase it. Oxygen deficits incurred during maximal constantload exercise lasting less than 2 minutes (for example, Hermansen & Medb0 1984) may not be
321
maximal and would, therefore, be lower than those incurred during longer exercise (e.g. Medb0 et al. 1988; Scott et al. 1991). However, recent evidence demonstrated that the 020 was maximised within 1 min during all-out, as opposed to constant-load, exercise (Gastin et al. 1991; Withers et al. 1991). Gastin et al. (1991) also showed that the 020 incurred during 60 sec of all out cycling or 2 to 3min of maximal constant-load cycling was similar; the range of 020 values cited in this study was, however, lower than those reported elsewhere when similar subjects and ergometers were used (Graham & Mclellan 1989; Medb0 & Burgers 1990; Medb0 & Tabata 1989; Szogy & Cherebetiu 1974). It remains to be verified whether this difference is because of the exercise protocol and/or type of person used; it is also plausible that the degree to which the 02 demand, and thus the 020, in those studies examining all-out (and thus higher intensity) exercise was underestimated to a greater extent (see section 4.2).
4.1 The Variability of the Oxygen Deficit
The coefficient of variation for the 020 measured in 4 endurance cyclists on 4 separate occasions ranged from 7.9 to 12.5% (Graham & McLellan 1989), although no information was provided concerning the sources of variability (i.e. biological or technological). These values are higher than the corresponding statistics reported for performance on short term, maximal cycling tests of 30 to 100 sec duration (Coggan & Costill 1984) and V02max (Katch et al. 1992; Kuipers et al. 1985). In addition, the test-retest coefficient for the 020 measured during cycling at 120% V02max (i.e. r = 0.73: Pate et al. 1983) is lower than that observed for the 020 incurred during 30 sec (r = 0.96), 60 sec (r = 0.95) and 90 sec all out cycling (r = 0.84: Withers et al. 1991), maximal work outputs during all out cycling over 10 to 90 sec (r ~ 0.89: Bar-Or 1987; McKenna et al. 1987; Simoneau et al. 1983) and anaerobic mechanical power tests (r ~ 0.97: Beckenholdt & Mayhew 1983). The reasons for the greater variance associated with the 020 are not well defined, although they may be related to both

322
the longer exercise duration and associated motivational problems experienced by subjects - note the decline in correlations with increasing exercise duration in Withers et al. (1991) - and/or methodological sources of variance.
4.2 Methodological Sources of Variance
The measurement of the 020 at supra-V02max power outputs is based on 2 primary assumptions: (a) the 02 demand of supra-V02max exercise can be extrapolated from submaximal V02 measurements; and (b) the 02 demand is constant during this type of exercise. Violation of the first assumption is likely to occur: muscular efficiency at submaximal intensities is likely to be higher than those at supra-V02max power outputs (Hansen et al. 1988; Luhtanen et al. 1989, 1990), which would result in an underestimate of the 02 demand and 020. It is not clear whether violations of the second assumption would occur and, if so, whether the changes in the 02 demand during exercise would be of sufficient magnitude to undermine the validity of 020.
Assumption (a) reflects the requirement to estimate the 02 demand from measures of submaximal work economy. In several of the earlier studies which measured the 020 (Karlsson & Saltin 1970, 1971; Pate et al. 1983; Szogy & Cheribetiu 1974) the exercise 02 demand was determined assuming a common value of work economy ranging from 19.5 to 22.5%. It has since been demonstrated that individual work economies (i.e. the regression of V02 on power output) at submaximal workloads may vary by 16% (Medb0 et al. 1988), suggesting that a high degree of error may be incurred when 02 deficits are determined using a common value for work economy. Medb0 et al. (1988) suggested that the 02 demand should be estimated according to the individual V02-power output relationship. These investigators measured 'steadystate' V02 values during 40 separate submaximal tests to determine the V02-power output relationship. Since this method is very time-consuming, an alternative method using a common y-intercept (5 ml/kg/min) and a single V02 measured at a speed
Sports Medicine 15 (5) 1993
equal to 90% V02max was recommended (Medb0 et al. 1988); Scott et al. (1991) recently applied a similar method to determine 020 in track athletes.
However, at high power outputs (i.e. 90%
V02max) substantial Lab accumulation would occur and probably diminish the accuracy with which V02 reflects the 02 demand (Vandewalle et al. 1989). Moreover, since V02 does not achieve a steady-state at power outputs greater than a 'critical' submaximal intensity (Poole et al. 1990; Whipp & Wasserman 1986), it is unlikely that steady-state conditions, and thus an accurate estimate of the energy cost of the exercise, can be realised at such high intensities.
In an attempt to minimise the anaerobic A TP contribution to submaximal work and develop a more practical method of measuring the 020, Green & Wenger (unpublished observations) determined a V02-speed regression line based on 5 V02 values obtained at 5 treadmill speeds below the ventilatory threshold, on 2 occasions. These investigators found that the 02 demands determined from the 2 V02-speed regression lines were not correlated (r = 0.20; p > 0.05), and that a 10%
difference in 020 values was observed when either regression line was used to estimate the 02 demand. The variation in the slope and y-intercept of the V02-speed relation was much higher than the corresponding variations reported by Medb0 et al. (1988); these larger values could be partially explained by the smaller range of speeds across which V02 was measured; they may also relate to alterations in physiological factors which determine V02 at any given workload, such as muscle glycogen levels and the relative mix of carbohydrate and fat used to sustain submaximal work, as well as anxiety and/or fatigue levels.
Recent data have demonstrated that running economy (V02 at 3.33 m/sec) can be estimated with minimal variability (CV = 1.3%) in trained athletes when the testing conditions (i.e. time of day, footwear) are strictly controlled and subjects are tested in a 'non-fatigued state' (Morgan et al. 1991). Similar rigour must therefore be applied when using submaximal V02 measures to estimate the 02 demand reliably, since the estimation of the 02 de-

Anaerobic Capacities in Humans
mand is a likely source of error when measuring the 02D.
The period over which the V02 response to submaximal workloads should be averaged and used to measure the exercise energy cost has also not been established. For example, Medb0 and colleagues (Medb0 & Burgers 1990; Medb0 & Tabata 1989; Medb0 et al. 1988) averaged V02 measured over 8 to 10 min of each submaximal workload, whereas Astrand et al. (l986) selected a 6 min period to determine V02-workload values across a similar range of intensities. Since V02 drift occurs at high submaximal workloads (Poole et al. 1990; Whipp & Wasserman 1986), the slope of the V02-workload regression, for a given individual, would be expected to be higher when derived from V02 values averaged across 8 to 10 min as opposed to 6 min. Such methodological differences in determining the oxygen demand of supra-V02max exercise could result in substantially different 02D values.
4.3 Is the Oxygen Deficit a Valid Measure of Anaerobic Capacity?
On the basis of the relatively large coefficients of variation reported (Graham & McLellan 1989) and potential difficulties in estimating the exercise 02 demand accurately, the accuracy with which the 02D measures anaerobic capacity could be questioned. The lack of correlation between 02D and running performance over 400 to 1500m (r = 0.05 to 0.21) in a heterogeneous group of athletes demonstrated that better performances on tasks partially determined by anaerobic capacity (or underlying mechanisms) are not always associated with a larger 02D (Green & Wenger, unpublished observations). However, in this same study the investigators demonstrated that a skill-related factor was probably a major determinant of performance. When a factor related to running technique has been kept relatively constant by using only track athletes (200 to 10 OOOm), a significant correlation between 02D and 300m sprinting performance has been observed (r = 0.76) [Scott et al. 1991].
There are many findings which indirectly sup-
323
port the validity of the 02D. Total work output during exhaustive exercise over 60 sec of all-out cycling (Szogy & Cherebetiu 1974) or 2 to 3 min of treadmill running (Scott et al. 1991) has been significantly correlated with 02D (r = 0.97 and r = 0.66) incurred during the same work bout. Sprint-trained athletes have consistently displayed greater 02D values than endurance-trained athletes during exhaustive treadmill tests (Hermansen & Medb0 1984; Medb0 & Sejersted 1985; Scott et al. 1991), a finding consistent with the greater anaerobic potential (i.e. buffer values, glycolytic enzyme activities) observed in sprint athletes (e.g. Komi et al. 1977; Parkhouse et al. 1985). The 02D can be improved by high-intensity training (Medb0 & Burgers 1990) and training at altitude (Bro-Rasmussen et al. 1985; Mizuno et al. 1990) in untrained and trained subjects, respectively. Bro-Rasmussen et al. (1985) also demonstrated that the magnitude of increase in 02D (12%) following a brief sojourn at altitude (2800m) was paralleled by a similar increase in muscle buffer values (6 to 8%) and treadmill running performance (11%) despite no change in V02max. These findings may also be partially explained, however, by changes in factors related to the assumptions on which 020 is based.
Recent studies have provided direct evidence supporting the validity of the 02D: Bangsbo et al. (1990) demonstrated that the 02D (mmol A TP) incurred during maximal one-legged exercise was almost identical to the A TP yield calculated from metabolite changes and the estimated clearance of La' from skeletal muscle during the exercise; similar findings have been reported for all-out cycling exercise (Withers et al. 1991). This latter finding was, however, based on data obtained from 2 untrained and 4 club-level endurance cyclists: 02D was only shown to be quantitatively similar to a crude estimate of anaerobic capacity, and not related to anaerobic capacity.
5. Conclusions
There is sufficient evidence to demonstrate that 02 debts do not accurately quantify anaerobic capacities, whereas the validity of Labmax as an es-

324
timate of lactic capacity is questionable. Researchers (e.g. di Prampero 1981; Saltin 1990) have expressed opposing views regarding the validity of Labmax, and its use by applied sports scientists is not universal. The degree to which Labmax accurately reflects lactic capacity may depend on the duration of the maximal exercise, and thus the question still remains open as to whether or not Labmax can, under certain exercise conditions, reflect lactic capacity with the accuracy required in elite athletic populations.
Recent research has supported the potential of the 02D to accurately quantify anaerobic capacity. If the assumption is made that, at present, the 02D is the most accurate measure of anaerobic capacity, an insight into the validity of Labmax can be established from those studies which have reported both 02D and Labmax data (table I). These studies reveal that the relative magnitudes of the 02D and Labmax values across a range of athletic and nonathletic people are similar: higher 02Ds are associated with higher Labmax values, and vice versa. However, several studies have reported a failure of Labmax to parallel the training-induced increases in 02D, buffering capacity and exercise performance (Medb0 & Burgers 1990; Mizuno et al. 1990). Short and long term changes in blood volume will always confound interpretation of Labmax data. Moreover, Labmax determined after a 2.5- to 3-minute maximal treadmill run was not correlated with the 02D (r = 0.09; p > 0.05), although Labmax determined after an all out 400m sprint was correlated with the 02D (r = 0.70; p < 0.05: Green, unpublished observations). This latter finding suggests that Labmax may be a more accurate indicator of lactic capacity when it is determined after maximal exercise of brief duration (Le. "" I min).
6. Future Directions
The use of Labmax with high performance athletes requires closer scrutiny, since it is an invasive and expensive procedure; the effects of exercise intensity and duration on Labmax and associated variabilities need to be addressed. Further research is also required to establish the utility of 02D within
Sports Medicine 15 (5) 1993
homogeneous, high performance athletic populations, with particular emphasis on reducing its variability to approximate values of normal biological variation. This would also be important to investigators using 02D to examine the metabolic and physiological bases of anaerobic capacity. Efforts to minimise variability must, however, recognise that the variance in 02D is alwalys likely to be higher than the other physiologicaljperformance measures (e.g. V02, work output) since it will be derived from multiple measures with their corresponding errors. Recent evidence indicates that approximately 2 min of maximal exercise is required to maximise the 02D during constant-load tests, but further research may reveal that the individual variation in the time required to achieve this maximal value may be large and specific to various athletic populations. If the duration required to maximise the 02D is similar to the duration of the competitive event, then there is a requirement to measure the 02D under exercise conditions (i.e. intensity and duration) specific to the athlete's chosen event. This principle of specificity may also apply to the 'pacing strategy' employed by the athlete, i.e. sustained, all-out efforts are rare in athletics and therefore, measuring the 020 under such conditions may yield values different from those incurred during the competitive event.
The method used to predict the 02 demand of intense exercise also needs closer scrutiny. Despite the support for the validity of the 02D as a measure of anaerobic capacity, evidence suggests that its validity is not established and that, at present, its use in quantifying anaerobic capacity should be conducted with caution.
References
Astrand po, Hultman E, Juhlin-Dannfeldt A, Reynolds G. Disposal of lactate during and after strenuous exercise in humans. Journal of Applied Physiology 61: 33-343, 1986
Bangsbo J, Gollnick PD, Graham TE, Juel C, Kiens B, et al. Anaerobic energy production and 02 deficit-debt relationship during exhaustive e"ercise in humans. Journal of Physiology 422: 539-559, 1990
Bar-Or O. Wingate test: update on reliability and validity. Sports Medicine 4: 381-394, 1987
Beckenholdt SE, Mayhew JL. Specificity among anaerobic power tests in male athletes. Journal of Sports Medicine and Physical Fitness 23: 326-332, 1983

Anaerobic Capacities in Humans
Belcastro AN, Bonen A. Lactic acid removal rates during controlled and uncontrolled recovery exercise. Journal of Applied Physiology 39: 932-936, 1975
Berg A, Keul J. Comparative performance diagnostics of anaerobic exertion in laboratory and field exercise of decathletes. International Journal of Sports Medicine 6: 244-253, 1985
Bro-Rasmussen T, Mizuno M, Mygind E, Juel C, Lortie G. Increased short term performance and buffer capacity in well trained cross-country skiers after training at altitude. Clinical Physiology 5: 139, 1985
Camus G, Thys H. An evaluation of the maximal anaerobic capacity in man. International Journal of Sports Medicine 12: 349-355, 1991
Cerretelli P. Oxygen debt: definition, role and significance. In Marconnet et al. (Eds) Medicine and sport sciences: physiological chemistry of training and detraining, Vol. 17, pp. 68-80, Karger, Basel, 1984
Chad K E, Wenger HA. The effect of exercise duration on the exercise and post-exercise oxygen consumption. Canadian Journal of Sports Sciences 13: 204-207, 1988
Cheetham ME, Boobis H, Brooks S, Williams C. Human muscle metabolism during sprinting. Journal of Applied Physiology 61: 54-60, 1986
Cheetham ME, Williams C. High intensity training and treadmill sprint performance. British Journal of Sports Medicine 21: 14-17, 1987
Christensen EH, Hogberg P. Steady-state, 02-deficit and 02- debt at severe work. Arbeitsphysiologie 14: S251-S254, 1950
Coggan AR, Costill DL. Biological and technological variability of three anaerobic ergometer tests. International Journal of Sports Medicine 5: 142-145, 1984
Coyle EF, Hemmert MK, Coggan AR. Effects of detraining on cardiovascular responses to exercise: role of blood volume. Journal of Applied Physiology 60: 95-99, 1986
Crielaard JM, Merken P, Franchimont P, Pirnay F, Petit J-M. Evaluation de la capacite anaerobique lactique en athIetisme. Medecine du Sport 60: 239-244, 1986
Cunningham DA, Faulkner JA. The effect of training on aerobic and anaerobic metabolism. Medicine and Science in Sports and Exercise I: 65-69, 1969
di Prampero PE. Energetics of muscular exercise. Reviews of Physiology Biochemistry and Pharmacology 89: 144-222, 1981
di Pram pero PE, Peeters L, Margaria R. Alactic 02 debt and lactic acid production after exhausting exercise in man. Journal of Applied Physiology 34: 628-632, 1973
Eriksson BO, Gollnick PO, Saltin B. Muscle metabolism and enzyme activities after training in boys 11-13 years old. Acta Physiologica Scandinavica 87: 485-497, 1973
Foxdal P, Sjodin B, Rudstam H, Ostman C. Ostman B, et al. Lactate concentration differences in plasma, whole blood, capillary finger blood and erythrocytes during submaximal graded exercise in humans. European Journal of Applied Physiology 61: 218-222, 1990
Freund H, Oyono-Enguelle S, Heitz A, Marbach J, Ott C, et al. Work rate-<lependent lactate kinetics after exercise in humans. Journal of Applied Physiology 61: 932-939, 1986
Freund H, Oyono-Enguelle S, Heitz A, Marbach J, Ott C, et al. Effect of exercise duration on lactate kinetics after short muscular exercise. European Journal of Applied Physiology 58: 534-542, 1989
Fujitsuka NT, Yamamoto T, Ohkuwa T, Saito M, Miyamura M. Peak blood lactate after short periods of treadmill running. European Journal of Applied Physiology 48: 289-296, 1982
Gaesser GA, Brooks GA. Metabolic bases of excess post-exercise oxygen consumption: a review. Medicine and Science in Sports Exercise 16: 29-43, 1984
Gastin P, Krzeminski K, Costill DL, MCConnell G. Accumulated oxygen deficit during isokinetic all-out and constant load supramaximal exercise. 8th Biennial Conference on Cardiovas-
325
cular and Respiratory Responses to Exercise in Health and Disease, University of Sydney, 1991
Goslin BR, Graham TE. A comparison of 'anaerobic' components of 02 debt and the Wingate Test. Canadian Journal of Applied Sports Sciences 10: 134-140, 1985
Graham KS, Mclellan TM. Variability of time to exhaustion and oxygen deficit in supramaximal exercise. Australian Journal of Science and Medicine in Sport 21: 11-14, 1989
Graham TE. Measurement and interpretation of lactate. In Lollgen & Mellerowicz (Eds) Progress in ergometry: quality control and test criteria, pp. 51-66, Springer Verlag, Berlin, 1984
Graham TE, Andrew GM. The variability of repeated measurements of oxygen debt in man following a maximal treadmill exercise. Medicine and Science in Sports and Exercise 5: 73-78, 1973
Green HJ, Houston ME. Effect of a season of ice hockey on energy capacities and associated function. Medicine and Science in Sports and Exercise 7: 299-303, 1975
Green HJ, Hughson RL, Thomson lA, Sharratt MT. Supramaximal exercise after training induced hypervolemia I. Gas exchange and acid-base balance. Journal of Applied Physiology 62: 1944-1961, 1987
Green S. The relationships between blood-borne and gas-exchange descriptors of anaerobic capacity and short-distance running performances, M.Sc. Thesis, University of Victoria, Canada, 1990
Hagberg JM, King DS, Rogers MA, MO!ltain SJ, Jilka SM, et al. Exercise and recovery ventilatory and V02 responses of patients with MCArdle's disease. Journal of Applied Physiology 68: 1393-1398, 1990
Hansen JE, Casaburi R, Cooper DM, Wasserman K. Oxygen uptake as related to work rate increment during cycle ergometer exercise. European Journal of Applied Physiology 57: 140-145, 1988
Harris P. Oxygen debt does not exist. In Moret et al. (Eds) lactate, physiologic, methodologic and pathologic approach, pp. 67-72, Springer Verlag, Berlin, 1980
Harris RC, Edwards RHT, Hultman E, Nordesjo LO, Nylind B, et al. The time course of phosphoryl creatine resynthesis during recovery of the quadriceps muscle in man. Ptlugers Archiv -European Journal of Physiology 367: 137-142, 1976
Harrison MH. Heat and exercise: effects on blood volume. Sports Medicine 3: 214-223, 1986
Hepburn D, Maughan RJ. Glycogen availability as a limiting factor in the performance of isometric exercise. Journal of Physiology 325: 52P-53P, 1982
Hermansen L, Grandmontagne M, Maehlum S, Ingnes I. Postexercise elevation of resting oxygen uptake - possible mechanisms and physiological significance. In Marconnet et al. (Eds) Medicine and sports sciences: physiological chemistry of training and detraining, Vol. 17, pp. 119-129, Karger, Basel, 1984b
Hermansen L, Medbe JI. The relative significance of aerobic and anaerobic processes during maximal exercise of short duration. In Marconnet et al. (Eds) Medicine and sport sciences: physiological chemistry of training and detraining, Vol. 17, pp. 56-67, Karger, Basel, 1984
Hermansen L, Orheim A, Sejersted OM. Metabolic acidosis and changes in water and electrolyte balance in relation to fatigue during maximal exercise of short duration. International Journal of Sports Medicine 5: 110-115, 1984a
Hermansen L, Stensvold I. Production and removal of lactate during exercise in man. Acta Physiologica Scandinavica 36: 191-200, 1972
Hermansen L, Vaage O. Lactate disappearance and glycogen synthesis in human muscle after maximal exercise. American Journal of Physiology 233: 422-429, 1977
Hill A V, Lupton H. Muscular exercise, lactic acid, and the supply and utilization of oxygen. Quarterly Journal of Medicine 16: 135-171, 1923

326
Houston ME, Thomson JA. The response of endurance-adapted adults to intense anaerobic training. European Journal of Applied Physiology 36: 207-213, 1977
Hughson RL. Alterations in the oxygen deficit-oxygen debt relationship with beta-adrenergic receptor blockade in man. Journal of Physiology 349: 375-387, 1984
Hultman E, Bergstrom J, McLennon-Anderson N. Breakdown and resynthesis of phosphory1creatine and adenosine-triphosphate in connection with muscular work in man. Scandinavian Journal of Clinical and Laboratory Investigation 19: 56-66, 1967
Jacobs I. Influence of carbohydrate stores on maximal human power output. In Macleod et al. (Eds) Exercise: benefits, limits and adaptation, pp. 104-115, E. and F.N. Spon, London, 1987
Jacobs I. Esbjornsson M, Sylven C, Holm I, Jansson E. Sprint training effects on muscle myoglobin, enzymes, fiber type, and blood lactate. Medicine and Science in Sports and Exercise 19: 368-374, 1987
Jansson E, Dudley GA, Norman B, Tesch PA. Relationship of recovery from intense exercise to the oxidative potential of skeletal muscle. Acta Physiologica Scandinavica 139: 147-152, 1990
Karlsson J, Saltin B. Lactate, A TP and CP in working muscles. Journal of Applied Physiology 29: 598-602, 1970
Karlsson J, Saltin B. Oxygen deficit and muscle metabolites in intermittent exercise. Acta Physiologica Scandinavica 82: 115-122, 1971
Katch VL, Sady SS, Freedson P. Biological variability of maximal aerobic power. Medicine and Science in Sports and Exercise 14: 21-25, 1982
Kindermann W, Kullmer T. Effect of previous exercise on the blood lactate in anaerobic exercise. International Journal of Sports Medicine 6: 241-253, 1985
Klausen K, Knuttgen HG, Forster HV. Effect of pre-existing high blood lactate concentration on maximal exercise performance. Scandinavian Journal of Clinical and Laboratory Investigation 30: 415-419,1972
Klausen K, Piehl K, Saltin B. Muscle glycogen stores and capacity for anaerobic work. In Howald & Poortmans (Eds) Metabolic adaptations to prolonged physical exercise, pp. 127-139, Birkhauser, Basel, 1975
Komi PV, Rusko H, Vos V, Vihko V. Anaerobic performance capacity in athletes. Acta Physiologica Scandinavica 100: 107-114, 1977
Koziris LP, Montgomery DL. Blood lactate concentration following intermittent and continuous cycling tests of anaerobic capacity. European Journal of Applied Physiology 63: 273-277. 1991
Krogh A, Lindhard J. The changes in respiration at the transition from work to rest. Journal of Physiology 53: 431-437, 1919/ 20
Kuipers H, Verstappen F, Keizer H, Geurten P. Van Kranenburg G. Variability of aerobic performance in the laboratory and its physiological correlates. International Journal of Sports Medicine 6: 197-210, 1985
Lacour JR, Bouvat E, Barthelemy Jc. Post-competition blood lactate concentrations as indicators of anaerobic energy expenditure during 400m and 800m races. European Journal of Applied Physiology 61: 172-176, 1990
Linnarsson D, Karlsson J, Fagraeus L, Saltin B. Muscle metabolites and oxygen deficit with exercise in hypoxia and hyperoxia. Journal of Applied Physiology 36: 399-402, 1974
Luhtanen P, Rahkila P, Rusko H, Viitasalo JT. Mechanical work and efficiency in ergometer cycling at aerobic and anaerobic thresholds. Acta Physiologica Scandinavica 131: 331-337, 1989
Luhtanen P, Rahkila P, Rusko H, Viitasalo JT. Mechanical work and efficiency in treadmill running at aerobic and anaerobic thresholds. Acta Physiologica Scandinavica 139: 153-159, 1990
Maison GL, Broeker AC. Training in human muscle working with
Sports Medicine 15 (5) 1993
and without blood supply. American Journal of Physiology 132: 391-404, 1941
Margaria R, Aghemo P, Sassi G. Lactic acid production in supramaximal exercise. Pflugers Archiv 326: 152-161, 1971
Margaria R, Cerretelli P, di Prampero PE, Massari C, Torelli G. Kinetics and mechanism of oxygen debt contraction in man. Journal of Applied Physiology 18: 371-377, 1963
Margaria R, Cerretelli P, Mangili F. Balance and kinetics of anaerobic energy release during strenuous exercise in man. Journal of Applied Physiology 19: 623-628, 1964
Margaria R, Edwards HT, Dill DB. The possible mechanism of contracting and paying the oxygen debt and the role of lactic acid in muscular contraction. American Journal of Physiology 106: 689-714, 1933
Martin BR. Metabolic regulation: a molecular approach. Blackwell Scientific Publications, 1987
Martin EG, Field J, Hall VE. Metabolism following anoxemia: I. Oxygen consumption and blood lactates after experimentally induced exercise. American Journal of Physiology 88: 407, 1929
McGrail JC, Bonen A, Belcastro AN. Dependence of lactate removal on muscle metabolism in man. European Journal of Applied Physiology 39: 91-97, 1978
McKenna MJ, Green RA, Shaw PF, Meyer AD. Tests of anaerobic power and capacity. Australian Journal of Science and Medicine in Sport 19: 13-17,1987
McNaughton LR, Cedaro R. The effect of sodium bicarbonate on rowing ergometer performance in elite rowers. Australian Journal Science Medicine and Sport 23: 669, 1991
Medbe JI, Burgers S. Effects of training on the anaerobic capacity. Medicine and Science in Sports and Exercise 22: 501-507, 1990
Medbe JI, Mohn AC, Tabata J, Bahr R, Vaage 0, et al. Anaerobic capacity determined by maximal accumulated 02 deficit. Journal of Applied Physiology 64: 50-60, 1988
Medbe JI, Tabata I. Relative importance of aerobic and anaerobic energy release during short-lasting exhausting bicycle exercise. Journal of Applied Physiology 67: 1881-1886, 1989
Mero A. Blood lactate production and recovery from anaerobic exercise in trained and untrained boys. European Journal of Applied Physiology 57: 660-666, 1988
Mizuno M, Juel C, Bro-Rasmussen T, Mygind E, Schibye B, et al. Limb skeletal muscle adaptation in athletes after training at altitude. Journal of Applied Physiology 68: 496-502, 1990
Morgan DW, Martin PE, Krahenbuhl GS, Baldini FD. Variability in running economy and mechanics among trained male runners. Medicine and Science in Sports and Exercise 23: 378-383, 1991
Needham DM. The biochemistry of muscular contraction in its historical development, Cambridge University Press, London, 1971
Nevill ME, Boobis LH, Brooks S, Williams C. Effect of training on muscle metabolism during treadmill sprinting. Journal of Applied Physiology 67: 2376-2382, 1989
Nordheim K, Vollestad MK. Glycogen and lactate metabolism during low-intensity exercise in man. Acta Physiologica Scandinavica 139: 475-484, 1990
Ohkuwa T, Kats Y, Katsumata K, Nakao T, Miyamura M. Blood lactate and glycerol after 400m and 3000m runs in sprint and long distance runners. European Journal of Applied Physiology 53: 213-218, 1984
Pate RR, Goodyear V, Dover J, Dorociak J, McDaniel J. Maximal 02 deficit: a test of anaerobic capacity. Medicine and Science in Sports and Exercise 15: 121-122, 1983
Paterson DH, Cunningham DA, Bumstead LA. Recovery 02 and blood lactic acid: longitudinal analysis in boys aged II to 15 years. European Journal of Applied Physiology 55: 93-99, 1980
Piiper J, di Prampero PE, Cerretelli P. Oxygen debt and high energy phosphates in gastrocnemius muscle of the dog. American Journal of Physiology 215: 523-531, 1968
Piiper J, Spiller P. Repayment of 02 debt and resynthesis of high

Anaerobic Capacities in Humans
energy phosphates in gastrocnemius muscle of the dog. Journal of Applied Physiology 28: 657-662, 1970
Poole DC, Ward SA, Whipp BJ. The effects of training on the metabolic and respiratory profile of high-intensity cycle ergometer exercise. European Journal of Applied Physiology 59: 421-429, 1990 . . . .
Powers SK, Beadle RE, Lawler J, Thompson D. Oxyg~~ deflci'toxygen debt relationships in ponies during submaximal treadmill exercise. Respiration Physiology 57: 251-263, 1987
Ready AE, Eynon RB, Cunningham DA. Effect of interval training and detraining on anaerobic fitness in women. Canadian Journal of Applied Sports Science 6: 114-118, 1991
Rieu M, Duvallet A, Scharapan L, Thieulart L, Ferry A. Blood lactate accumulation in intermittent supramaximal exercise. European Journal of Applied Physiology 57: 235-245, 1988
Roberts AD, Billeter R, Howald H. Anaerobic muscle enzyme changes after interval training. International Journal of Sports Medicine 3: 18-21, 1982
Roberts AD, Morton AR. Total and alactic oxygen debts after supramaximal work. European Journal of Applied Physiology 38: 281-289, 1978
Rose RJ, Hodgson DR, Kelso TB, McCutcheon U, Reid TA, et al. Maximum 02 uptake, 02 debt and deficit, and muscle metabolites in Thoroughbred horses. Journal of Applied Physiology 64: 781-788, 1988
Sahlin K, Harris RC, Hultman E. Resynthesis of creatine phosphate in human muscle after exercise in relation to intramuscular pH and availability of oxygen. Scandinavian Journal of Clinical and Laboratory Investigation 39: 551-558, 1979
Saltin B. The physiological and biochemical basis of aerobic and anaerobic capacities in man: effect of training and range of adaptation. [n Maehlum et al. (Eds) An update on sports medicine: proceedings from the Second Scandinavian Conference on Sports Medicine, pp. 16-59, Oslo, 1987
Saltin B. Anaerobic capacity: past, present, and prospective. In Taylor et al. (Eds) Biochemistry of exercise VII, International Series on Sport Sciences. Vol. 21, pp. 387-412, Human Kinetics Publishers, Champaign, 1990
Sawka MN. Tahamont MV, Fitzgerald PI, Miles DS, Knowlton RG. Alactic capacity and power: reliability and interpretation. European Journal of Applied Physiology 45: 109-116, 1980
Schnabel A. Kindermann W, Keul J, Schmitt WM. Bearteilung der anaeroben Ausdauer ('Stehvermogen') im Labor. Leistungssport 9: 503-507, 1979
Schnabel A, Kindermann W. Assessment of anaerobic capacity in runners. European Journal of Applied Physiology 52: 42-46, 1983
Scott CB, Roby FB, Lohman TG, Bunt Jc. The maximally ac-
327
cumulated oxygen deficit as an indicator of anaerobic capacity. Medicine and Science in Sports and Exercise 23: 618-624, 1991
Sharp RL, Costill DL, Fink WJ, King DS. Effects of eight weeks of bicycle ergometer sprint training on human muscle buffer capacity. [nternational Journal of Sports Medicine 7: l3-17, 1986
Sheply B, MacDougall D, Cipriano N, Sutton [, Coates G. Physiological effects of tapering in highly trained athletes. Medicine and Science in Sports and Exercise 22: S134, 1990
Simoneau JA, Lortie G, Boulay MR, Bouchard C. Tests of anaerobic alactacid and lactacid capacities: description and reliability. Canadian Journal of Applied Sports and Sciences 8: 266-270, 1983
Sjodin B, Thorstensson A, Frith K, Karlsson l. Effect of physical training on LDH activity and LDH isozyme patterns in human skeletal muscle. Acta Physiologica Scandina vica 97: 150-l57, 1976
Szogy A, Cherebetiu G. Minutentest auf dem Fahrradergometer wr Bestimmung der anaeroben Kapazitat. European Journal of Applied Physiology 33: 171-176, 1974
Tamayo M, Sucec A, Phillips W, Bicono M. Laubach L, et al. The Wingate anaerobic power test, peak blood lactate and maximal oxygen debt in volleyball players. Abstract. Medicine and Science in Sports and Exercise 16: 126, 1984
Taunton JE, Maron H, Wilkinson JG. Anaerobic performance in middle and long-distance runners. Canadian Journal of Applied Sports Science 6: 109-113, 1981
Vandewalle H, Kapitaniak B, Grun S, Raveneau S, Monod H. Comparison between a 30s all-out test and a time-work test on a cycle ergometer. European Journal of Applied Physiology 58: 375-381, 1989
Whipp BJ, Wasserman K. Effect of anaerobiosis on the kinetics of 02 uptake during exercise. Federation Proceedings 45: 2942-2947, 1986
Withers RT, Sherman WM, Clark DU, Esselbach PC, Nolan SR, et al. Muscle metabolism during 30, 60 and 90s of maximal cycling on an air-braked ergometer. European Journal of Applied Physiology 63: 354-362, 1991
Woledge RC. Energy transformation in living muscle. In Wieser & Grainger (Eds) Energy transformations in cells and organisms. Proceedings of the 10th Conference of the European Society for Comparative Physiology and Biochemistry, pp. 36-45, Georg Thieme Verlag, Stuttgart, 1989
Correspondence and reprints: Simon Green. Department of Human Movement. University of Western Australia, Nedlands, WA 6009, Australia.




















