
McsA and B mediate the delocalization of competence proteins from the cell poles of Bacillus...
14
McsA and B mediate the delocalization of competence proteins from the cell poles of Bacillus subtilis Jeanette Hahn, Naomi Kramer, Kenneth Briley Jr and David Dubnau* Public Health Research Institute, University of Medicine and Dentistry of New Jersey, Newark, NJ, USA. Summary During the development of transformability (compe- tence), Bacillus subtilis synthesizes a set of proteins that mediate both the uptake of DNA at the cell poles and the recombination of this DNA with the resident chromosome. Most, if not all, of these Com proteins localize to the poles of the cell, where they associate with one another, and are then seen to delocalize as transformability declines. In this study, we use fluo- rescence microscopy to analyse the localization and delocalization processes. We show that localization most likely occurs by a diffusion-capture mechanism, not requiring metabolic energy, whereas delocaliza- tion is prevented in the presence of sodium azide. The kinetics of localization suggest that this process requires the synthesis of a critical protein or set of proteins, which are needed to anchor the Com protein complex to the poles. We further show that the protein kinase proteins McsA and McsB are needed for delocalization, as are ClpP and either of the AAA + (ATPases associated with a variety of cellular activi- ties) proteins ClpC or ClpE. Of these proteins, at least McsB, ClpC and ClpP localize to the cell poles of competent cells. Our evidence strongly suggests that delocalization depends on the degradation of the pos- tulated anchor protein(s) by the McsA-McsB-(ClpC or ClpE)-ClpP protease in an ATP-dependent process that involves the autophosphorylation of McsB. The extent of cell–pole association at any given time reflects the relative rates of localization and delocalization. The kinetics of this dynamic process differs for individual Com proteins, with the DNA- binding proteins SsbB and DprA exhibiting less net localization. Introduction Bacterial development is often accompanied by a pro- grammed accumulation of proteins and protein com- plexes at the poles of asymmetric cells and by the delocalization of these proteins from the poles. These movements, sometimes together with selective protein degradation, have been studied notably in the develop- mental cycle of Caulobacter crescentus and during the formation of spores (Shapiro et al., 2002), but also during the development of competence in Bacillus subtilis (Hahn et al., 2005; Kidane and Graumann, 2005; Kramer et al., 2007; Tadesse and Graumann, 2007). In B. subtilis, genetic competence is marked by the synthesis of a number of cytosolic and membrane-localized proteins that assemble into a polar-localized molecular machine dedi- cated to the internalization, processing and chromosomal integration of transforming DNA (Hahn et al., 2005; Kidane and Graumann, 2005; Kramer et al., 2007). DNA uptake during transformation consequently takes place mostly at the poles of the rod-shaped bacilli. As the cells lose the capacity to take up DNA, the competence (Com) proteins leave the poles (Hahn et al., 2005). Remarkably, at least some of the Com proteins colocalize with one another not only at the poles, but also in helical tracks that extend along the long axis of the cell. Many questions concerning the localization and delocalization of Com pro- teins remain. If the Com proteins move to the poles from their sites of synthesis, do they do so individually and then assemble to form the DNA uptake machines or do com- plexes of Com proteins move to the poles? Is pole-ward movement directed or driven by diffusion and capture? Are unknown proteins required for this localization? How is delocalization accomplished and does it require pro- tein degradation? The present study addresses these questions. We report that complexes assemble away from the poles, and that these ensembles move, probably by diffusion, to take up residence at the cell poles. In contrast to this diffusive process, delocalization seems to require metabolic energy. We show that the McsA/McsB protein kinase (Kirstein et al., 2005; 2007; 2008) is required for delocalization, and that delocalization most likely depends on the phosphorylation of McsB. We propose that an unknown anchor protein is degraded by a proteolytic complex consisting of McsA/McsB and ClpP, in complex Accepted 9 February, 2009. *For correspondence. E-mail dubnauda@ umdnj.edu; Tel. (+1) 973 854 3400; Fax (+1) 973 854 3453. Molecular Microbiology (2009) 72(1), 202–215 doi:10.1111/j.1365-2958.2009.06636.x First published online 25 February 2009 © 2009 The Authors Journal compilation © 2009 Blackwell Publishing Ltd
-
Upload
independent -
Category
Documents
-
view
1 -
download
0
Transcript of McsA and B mediate the delocalization of competence proteins from the cell poles of Bacillus...

McsA and B mediate the delocalization of competenceproteins from the cell poles of Bacillus subtilis
Jeanette Hahn, Naomi Kramer, Kenneth Briley Jrand David Dubnau*Public Health Research Institute, University of Medicineand Dentistry of New Jersey, Newark, NJ, USA.
Summary
During the development of transformability (compe-tence), Bacillus subtilis synthesizes a set of proteinsthat mediate both the uptake of DNA at the cell polesand the recombination of this DNA with the residentchromosome. Most, if not all, of these Com proteinslocalize to the poles of the cell, where they associatewith one another, and are then seen to delocalize astransformability declines. In this study, we use fluo-rescence microscopy to analyse the localization anddelocalization processes. We show that localizationmost likely occurs by a diffusion-capture mechanism,not requiring metabolic energy, whereas delocaliza-tion is prevented in the presence of sodium azide. Thekinetics of localization suggest that this processrequires the synthesis of a critical protein or set ofproteins, which are needed to anchor the Com proteincomplex to the poles. We further show that theprotein kinase proteins McsA and McsB are neededfor delocalization, as are ClpP and either of the AAA+
(ATPases associated with a variety of cellular activi-ties) proteins ClpC or ClpE. Of these proteins, at leastMcsB, ClpC and ClpP localize to the cell poles ofcompetent cells. Our evidence strongly suggests thatdelocalization depends on the degradation of the pos-tulated anchor protein(s) by the McsA-McsB-(ClpC orClpE)-ClpP protease in an ATP-dependent processthat involves the autophosphorylation of McsB. Theextent of cell–pole association at any given timereflects the relative rates of localization anddelocalization. The kinetics of this dynamic processdiffers for individual Com proteins, with the DNA-binding proteins SsbB and DprA exhibiting less netlocalization.
Introduction
Bacterial development is often accompanied by a pro-grammed accumulation of proteins and protein com-plexes at the poles of asymmetric cells and by thedelocalization of these proteins from the poles. Thesemovements, sometimes together with selective proteindegradation, have been studied notably in the develop-mental cycle of Caulobacter crescentus and during theformation of spores (Shapiro et al., 2002), but also duringthe development of competence in Bacillus subtilis (Hahnet al., 2005; Kidane and Graumann, 2005; Kramer et al.,2007; Tadesse and Graumann, 2007). In B. subtilis,genetic competence is marked by the synthesis of anumber of cytosolic and membrane-localized proteins thatassemble into a polar-localized molecular machine dedi-cated to the internalization, processing and chromosomalintegration of transforming DNA (Hahn et al., 2005;Kidane and Graumann, 2005; Kramer et al., 2007). DNAuptake during transformation consequently takes placemostly at the poles of the rod-shaped bacilli. As the cellslose the capacity to take up DNA, the competence (Com)proteins leave the poles (Hahn et al., 2005). Remarkably,at least some of the Com proteins colocalize with oneanother not only at the poles, but also in helical tracks thatextend along the long axis of the cell. Many questionsconcerning the localization and delocalization of Com pro-teins remain. If the Com proteins move to the poles fromtheir sites of synthesis, do they do so individually and thenassemble to form the DNA uptake machines or do com-plexes of Com proteins move to the poles? Is pole-wardmovement directed or driven by diffusion and capture?Are unknown proteins required for this localization? Howis delocalization accomplished and does it require pro-tein degradation? The present study addresses thesequestions. We report that complexes assemble away fromthe poles, and that these ensembles move, probably bydiffusion, to take up residence at the cell poles. In contrastto this diffusive process, delocalization seems to requiremetabolic energy. We show that the McsA/McsB proteinkinase (Kirstein et al., 2005; 2007; 2008) is required fordelocalization, and that delocalization most likely dependson the phosphorylation of McsB. We propose that anunknown anchor protein is degraded by a proteolyticcomplex consisting of McsA/McsB and ClpP, in complex
Accepted 9 February, 2009. *For correspondence. E-mail [email protected]; Tel. (+1) 973 854 3400; Fax (+1) 973 854 3453.
Molecular Microbiology (2009) 72(1), 202–215 � doi:10.1111/j.1365-2958.2009.06636.xFirst published online 25 February 2009
© 2009 The AuthorsJournal compilation © 2009 Blackwell Publishing Ltd

with either one of two AAA+ [ATPases associated withvarious cellular activities (Ogura and Wilkinson, 2001)]proteins ClpC or ClpE.
Results
Localization of ComGA
Competence genes are expressed in B. subtilis beginningat T0 (the time at which a culture departs from exponentialgrowth). In most of the experiments reported here,we have used the protein encoded by comGA, atraffic ATPase, as representative of competence (Com)proteins. ComGA is required for transformation and alsofor the assembly of the competence pseudopilus, a struc-ture formed outside the cell membrane (Chen et al.,2006). ComGA is peripherally associated with the innerface of the membrane (Chung et al., 1998) and accumu-lates at the cell poles, as the cells become maximallytransformable (Hahn et al., 2005). To follow the polarlocalization of ComGA, time-lapse experiments were con-ducted in which cells expressing ComGA–GFP undercompetence control were incubated at 37°C on anagarose pad containing conditioned competence medium(selected frames are displayed in Fig. 1 and a represen-tative time-lapse movie of additional cells may be seen inMovie S1). In this experiment, the cells were grown inbatch culture until 30 min after T0 (T0.5) and then placed onthe agarose pads. In the earliest images, the fluorescentsignals were predominantly non-polar (in about 75% ofthe competent cells) and then progressively localized tothe poles in more than 70% of these cells. Inspection ofthese and other images shows that ComGA–GFPappears first as diffuse fluorescence, or in punctate foci,
and that this fluorescence moves to the poles. In somecells, depopulation of the non-polar regions is discernible.The distribution of non-polar fluorescence is often non-uniform and in some cells is suggestive of helices (Hahnet al., 2005). Based on the examination of 18 individualcells, the average time from T0.5 to the first detection of apolar focus ranged from 0 to 20 min (6.1 � 11.5).
Interpretation of these images is complicated by severalfactors. First, continuing synthesis of ComGA–GFPoccurs during this experiment, as determined by Westernblotting on batch cultures (not shown). Second, for fluo-rescence to appear, maturation of the GFP domain musttake place following synthesis, requiring several minutes.Third, it is possible that some turnover of ComGA–GFPmay be occurring, although Western blotting after theaddition of a protein synthesis inhibitor suggests that thisprotein is stable (see below). To determine whether theprogressive accumulation of ComGA–GFP fluorescenceat the poles shown in Fig. 1 and Movie S1 representsmovement of protein, rather than its synthesis atthe poles, we used puromycin (Pm) to inhibit proteinsynthesis. Images from a typical time-lapse seriesare shown in Fig. 2 and the complete movie is presentedin Movie S2. These experiments permitted severalconclusions. First, previously synthesized ComGA–GFPcan move to the poles. We often noted non-polar fluores-cent foci that moved erratically and eventually reached apole, at which point their movement decreased. In addi-tion, the impression of movement along a helical path wasstronger in these images than in the absence of Pm,perhaps due to the absence of new ComGA–GFP syn-thesis, which would ‘fill-in’ regions vacated by moleculesthat had moved to the helix or to a pole. Finally, in thepresence of Pm, about fivefold more time was needed for
Fig. 1. Localization of ComGA–GFP. BD2899, expressing comGA–gfp, was grown in competence medium until 30 min after T0 and analiquot was placed on an agarose pad containing competence medium. Fluorescence images were collected every 30 s. Selectedfluorescence images are depicted, together with DIC images taken at the beginning and end of the experiment. The numbers in the framesrefer to minutes. The single competent cell in the field is marked with an arrow. For a similar time-lapse experiment in its entirety, seeMovie S1.
Delocalization of competence proteins 203
© 2009 The AuthorsJournal compilation © 2009 Blackwell Publishing Ltd, Molecular Microbiology, 72, 202–215

ComGA to stably occupy a pole than in its absence;32.1 � 8.7 min, based on measurements made on 12cells, compared with 6.1 � 11.5 min in the absence of Pm(P < 0.0001). The lower relative standard deviation of thismeasurement compared with that in the absence of Pm isprobably due to the greater ease of determining the end-point (localization) than when continuing synthesis ofComGA–GFP fills in non-polar areas of the cells.
We suggest that a critical anchor protein must be madefor localization to take place. When protein synthesis isinhibited early in competence development, as in theexperiment involving the addition of Pm, the concentrationof the anchor protein is low and localization is correspond-ingly slow. This low rate of localization would be expectedfor a diffusion-capture mechanism, in which the probabil-ity of capture is dependent on the concentration of polarbinding sites or of an anchor protein that forms an inter-acting surface on the Com protein complex. In rare cells,observed only in the presence of Pm, we have detectednon-polar foci that move erratically and never settle at thepole. Perhaps the concentration of the anchor protein inthese cells is insufficient to permit polar capture of thefoci. The erratic movement of the non-polar foci, as well astheir appearance and disappearance, is suggestive of adiffusion-capture mechanism. This mechanism is alsostrongly supported by the azide resistance of localization(discussed below). The slower localization observed inthe presence of Pm suggests that the probability ofcapture depends on the concentration of the hypotheticalanchor protein. The relatively high standard deviationsmeasured for the time-to-capture are also consistent witha stochastic (diffusive) process.
Delocalization is not due to Com protein degradationand requires metabolic energy, while localizationdoes not
After T2, polar Com protein foci disappear, with retentionof GFP fluorescence elsewhere in the cell, often as non-
polar foci (Hahn et al., 2005). Previous experiments didnot distinguish between the departure of protein from thepoles and the loss of polar localization due to preferentialdegradation of polar Com proteins. Although the totalfluorescent signal did not seem to decrease during delo-calization, it was possible that preferential degradation ofpolar Com protein was masked by the synthesis of newprotein. To study the delocalization of old protein, Pm andrifampicin (Rm) were added at T1, when about half of thecells already exhibited polar localization. [We have shownthat Pm acts almost immediately to inhibit protein synthe-sis (see Supporting information)]. Figure 3A shows thatpre-existing ComGA–GFP protein does delocalize. In theabsence of Pm + Rm, the per cent localization increasedslightly during the 5–10 min following the initial point at T1
and then remained essentially constant, as expected fromthe observation that net delocalization does not occur untilafter T2 (Hahn et al., 2005). During the first 2 min afterPm + Rm addition, a slight increase in polar localizationoccurred, followed by a decrease in the per cent localizedcells from 62% to 39% of the population of competence-expressing cells. The delocalized fluorescence usuallyappeared as non-polar foci and similar results wereobtained with the helicase-like competence proteinComFA–GFP in the presence of Pm + Rm (not shown). Todetermine if the delocalization of ComGA requires energy,a similar experiment was carried out with the addition ofsodium azide (40 mM) as well as Pm + Rm (Fig. 3A). Inthe presence of NaN3, not only did delocalization fail tooccur, but the per cent cells exhibiting localizationincreased. These results can be explained as follows.When no inhibitors are present, ComGA synthesis andlocalization as well as delocalization occur, and the percent cells with localized ComGA is a result of these com-peting processes. In Fig. 3A, during the 20 min followingT1, no net delocalization takes place. When Pm + Rmblocks synthesis, a slight increase in ComGA localizationat the poles takes place, reflecting movement of proteinformed prior to addition of the inhibitor. As this pool of
Fig. 2. Localization of ComGA–GFP in the presence of Pm. BD2899, expressing comGA–gfp, was grown in competence medium until30 min after T0. Pm was added to an aliquot, which was then placed on an agarose pad containing competence medium with Pm. Imageswere collected at 2 min intervals. Selected fluorescence images are shown, together with DIC images taken at the end of the experiment. Thenumbers in the frames refer to minutes. The single competent cell in the field is marked with an arrow. For a similar time-lapse experiment inits entirety, see Movie S2.
204 J. Hahn, N. Kramer, K. Briley Jr and D. Dubnau �
© 2009 The AuthorsJournal compilation © 2009 Blackwell Publishing Ltd, Molecular Microbiology, 72, 202–215

unlocalized protein is exhausted, a net decrease in theper cent cells with polar foci occurs due to delocalization,unobscured by further synthesis. When NaN3 is present,delocalization is blocked and all of the ComGA can becaptured at the poles, resulting in an increase in the percent of localized cells. Western blotting revealed thatduring the course of incubation with Pm, no detectabledegradation of ComGA–GFP took place (Fig. 3B). Weconclude from these experiments that delocalization of‘old’ ComGA occurs without degradation and that delocal-ization requires metabolic energy. This conclusion issubject to the caveat that azide is an inhibitor of SecA(Oliver et al., 1990). However, the energy requirement fordelocalization is strongly supported by the involvement ofATP requiring proteins, described below. It is interestingthat in the presence of NaN3 the per cent of localized cellsactually increased, consistent with the idea that localiza-
tion does not require metabolic energy, proceedinginstead by diffusion and capture.
McsA and B are required for delocalization
We noticed that a clpC knockout strain expressingComGA–GFP failed to exhibit delocalization. Because theclpC deletion in this strain also affected the immediateupstream open reading frame (Pan et al., 2001), encodingthe protein kinase McsB (Fig. 4A), we tested a non-polarmcsB mutant for delocalization and observed a strikingphenotype (Fig. 4B). First, at T2 localization was morecomplete than in the wild-type strain, with nearly all thedetectable fluorescence confined to one or two foci ineach cell, almost invariably at a cell pole. In contrast, asnoted previously, wild-type cells exhibiting localizationalso possessed visible fluorescence located away fromthe poles, either distributed diffusely or localized to one ormore non-polar foci. Second, in the mcsB mutant,ComGA–GFP failed to delocalize after T2, as seen in theT6 images in Fig. 4B, and in fact localization appeared toincrease. In four experiments, in each of which at least100 competent cells were counted, the wild-type andmcsB strains exhibited an average of 78 � 18% and98 � 1.4% of competent cells with clear polar localizationat T2. At T5, the per cent of localized cells had decreasedto an average of 29 � 8.7% in the wild-type strain andwas 96 � 2.3% in the mcsB background. CtsR is arepressor, acting on the promoter upstream from theoperon containing mcsB (Fig. 4A) and in its absence wemight expect that all four genes in the operon, includingmcsB, would be overexpressed. In accordance with this,we noted that in a ctsR knockout, localization of ComGA–GFP is decreased; few polar foci are observed and thefluorescence is distributed throughout the cell, althoughoften with a punctate appearance (Fig. 4B). This is notsurprising, because an increase in the cellular concentra-tion of McsB due to the absence of the repressor mightreasonably be expected to confer the reverse of the mcsBknockout phenotype, whatever the mechanism involved.Figure 3A includes a control experiment with the mcsBstrain BD3594 (filled symbols) identical to that performedwith the wild-type strain (open symbols). As expected, nodelocalization was detected with this strain during the20 min following T1, showing that the delocalizationobserved with the wild-type strain in the presence of Pmwas not an artefact caused by the inhibitor or a processunrelated to the role of McsB.
The mcsB phenotype is competence-specific
Several constructs tested the specificity of the mcsBeffect on Com protein delocalization. When GFP is
0 10 20
100
50
Time (min)
Loca
lized
cel
ls (%
)
A
B
0 10 20 30 (min)
Fig. 3. Kinetics of ComGA–GFP localization and delocalization.A. BD2899, expressing comGA–gfp, grown in competence medium,was taken at T1 and incubated further in the presence of Pm + Rm(open circles), Pm + Rm + NaN3 (open squares) and with noinhibitor added (open triangles). The same experiment wasperformed with the isogenic mcsB strain BD3594 and the resultsare displayed with the equivalent closed symbols. The per cent ofcompetence-expressing cells showing polar localization offluorescence was determined by counting at least 100competence-expressing cells per sample.B. Western blot of samples taken at the indicated times followingaddition of Pm + Rm, using anti-GFP antiserum to visualize nativeComGA.
Delocalization of competence proteins 205
© 2009 The AuthorsJournal compilation © 2009 Blackwell Publishing Ltd, Molecular Microbiology, 72, 202–215
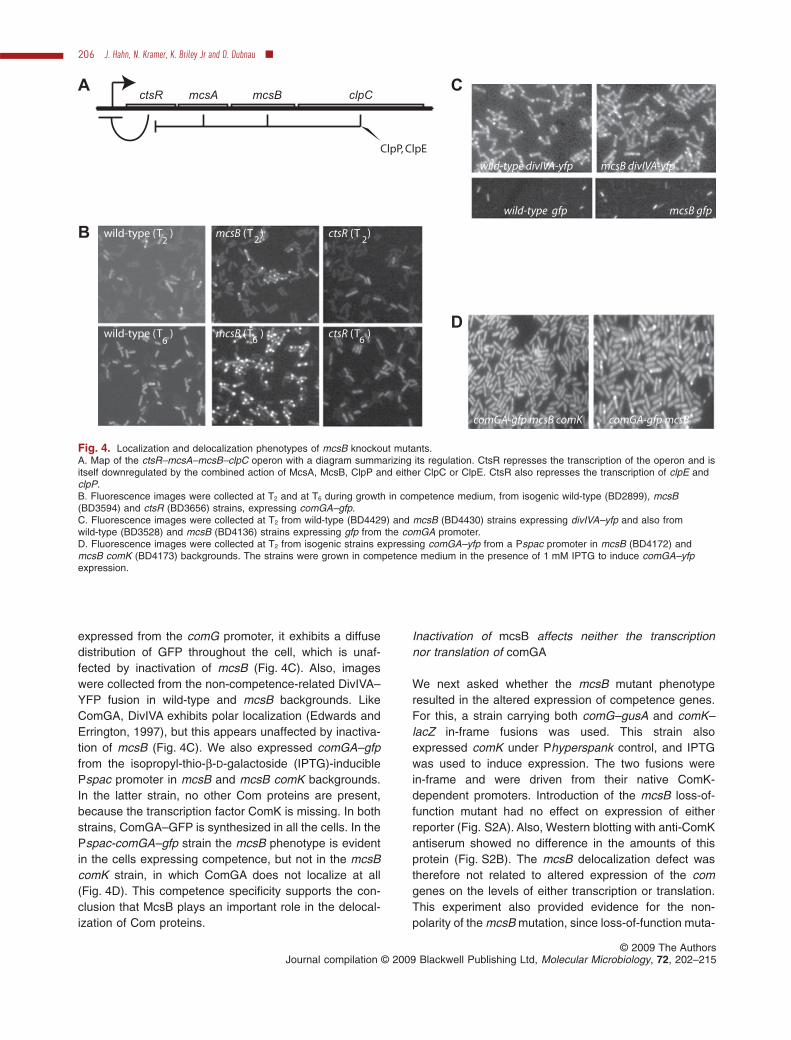
expressed from the comG promoter, it exhibits a diffusedistribution of GFP throughout the cell, which is unaf-fected by inactivation of mcsB (Fig. 4C). Also, imageswere collected from the non-competence-related DivIVA–YFP fusion in wild-type and mcsB backgrounds. LikeComGA, DivIVA exhibits polar localization (Edwards andErrington, 1997), but this appears unaffected by inactiva-tion of mcsB (Fig. 4C). We also expressed comGA–gfpfrom the isopropyl-thio-b-D-galactoside (IPTG)-induciblePspac promoter in mcsB and mcsB comK backgrounds.In the latter strain, no other Com proteins are present,because the transcription factor ComK is missing. In bothstrains, ComGA–GFP is synthesized in all the cells. In thePspac-comGA–gfp strain the mcsB phenotype is evidentin the cells expressing competence, but not in the mcsBcomK strain, in which ComGA does not localize at all(Fig. 4D). This competence specificity supports the con-clusion that McsB plays an important role in the delocal-ization of Com proteins.
Inactivation of mcsB affects neither the transcriptionnor translation of comGA
We next asked whether the mcsB mutant phenotyperesulted in the altered expression of competence genes.For this, a strain carrying both comG–gusA and comK–lacZ in-frame fusions was used. This strain alsoexpressed comK under Phyperspank control, and IPTGwas used to induce expression. The two fusions werein-frame and were driven from their native ComK-dependent promoters. Introduction of the mcsB loss-of-function mutant had no effect on expression of eitherreporter (Fig. S2A). Also, Western blotting with anti-ComKantiserum showed no difference in the amounts of thisprotein (Fig. S2B). The mcsB delocalization defect wastherefore not related to altered expression of the comgenes on the levels of either transcription or translation.This experiment also provided evidence for the non-polarity of the mcsB mutation, since loss-of-function muta-
ctsRA
clpCmcsBmcsA
ClpP, ClpE
B wild-type (T )2
wild-type (T )6
mcsB (T )2
mcsB (T )6
ctsR (T )2
ctsR (T )6
wild-type divIVA-yfp mcsB divIVA-yfp
C
D
comGA-gfp mcsBcomGA-gfp mcsB comK
wild-type gfp mcsB gfp
Fig. 4. Localization and delocalization phenotypes of mcsB knockout mutants.A. Map of the ctsR–mcsA–mcsB–clpC operon with a diagram summarizing its regulation. CtsR represses the transcription of the operon and isitself downregulated by the combined action of McsA, McsB, ClpP and either ClpC or ClpE. CtsR also represses the transcription of clpE andclpP.B. Fluorescence images were collected at T2 and at T6 during growth in competence medium, from isogenic wild-type (BD2899), mcsB(BD3594) and ctsR (BD3656) strains, expressing comGA–gfp.C. Fluorescence images were collected at T2 from wild-type (BD4429) and mcsB (BD4430) strains expressing divIVA–yfp and also fromwild-type (BD3528) and mcsB (BD4136) strains expressing gfp from the comGA promoter.D. Fluorescence images were collected at T2 from isogenic strains expressing comGA–yfp from a Pspac promoter in mcsB (BD4172) andmcsB comK (BD4173) backgrounds. The strains were grown in competence medium in the presence of 1 mM IPTG to induce comGA–yfpexpression.
206 J. Hahn, N. Kramer, K. Briley Jr and D. Dubnau �
© 2009 The AuthorsJournal compilation © 2009 Blackwell Publishing Ltd, Molecular Microbiology, 72, 202–215

tions in clpC, the gene immediately downstream of mcsB,would be expected to exhibit a dramatic increase in comgene expression (Dubnau and Roggiani, 1990). However,Western blotting with anti-ComGA antiserum did show anincreased concentration of this protein, presumably due toits greater stability in the mcsB background (Fig. S2C).The same has been observed for ComFA by Westernblotting, and we will return to this point below, inDiscussion.
Inactivation of mcsB affects the delocalization ofadditional Com proteins
To test the generality of the mcsB delocalization pheno-type, we studied several additional Com protein fusions toYFP (Fig. 5). The mcsB delocalization defect was appar-ent for ComFA, YjbF, DprA and SsbB (Fig. 5). The patternobserved for DprA in the mcsB + background was differentfrom that of the other Com proteins, because at T2, local-ization was observed in fewer of the competent cells thanwas the case for YjbF, ComFA, SsbB and ComGA.However, the effect of the mcsB mutation on the localiza-tion of YFP–DprA at T2 was quite pronounced and thisdramatic difference persisted until T5. Although SsbB wasmore localized when mcsB was inactivated, it was not asdramatically affected as the other Com proteins and theapparently enhanced stability noted by Western blottingfor ComGA and ComFA in the mcsB mutant was notobserved for SsbB (Fig. S2D). ComGA, YjbF and DprA failto localize when expressed in non-competent cells.Because SsbB can associate with the poles indepen-dently of other Com proteins (Kramer et al., 2007), wesuspected that it might be needed for them to localize.However, an ssbB knockout strain appeared unaffected inthe localization of ComGA–GFP (not shown).
The failure-to-delocalize phenotype of mcsA and mcsBmutants is not mediated by an effect on the repressorprotein CtsR
mcsB and its immediate upstream neighbour mcsA(Fig. 4A) have been shown by the Turgay lab to encodeproteins that bind to ClpC, targeting CtsR for degradationunder heat-shock conditions (Kirstein et al., 2005; 2007).McsB is an auto-kinase and its kinase activity is stimu-lated by the presence of McsA. The complex of McsA andB phosphorylates CtsR, reversing its binding to DNA, andthese proteins also act as adapter proteins, targeting thedegradation of CtsR by complexes of ClpP with ClpC. Ithas been reported that not only ClpC, but also the relatedAAA+ protein ClpE are required for the full in vivo degra-dation of CtsR during heat shock (Miethke et al., 2006),suggesting that ClpE, like ClpC, may associate with McsAand B. In fact, it has been demonstrated in vitro that McsA
and McsB can target CtsR for degradation by ClpE+ClpP(J. Kirstein and K. Turgay, pers. comm.). CtsR is the re-pressor of the ctsR–mcsA–mcsB–clpC operon (Fig. 4A),as well as of clpP and clpE (Krüger and Hecker, 1998;Derré et al., 1999; 2000). As noted above, we observedthat in a ctsR mutant strain, the localization of ComGA–GFP is decreased (Fig. 4B). This observation raises thepossibility that McsA and B are over-produced in theabsence of CtsR, leading to decreased localization.However, it is also possible that CtsR itself regulates polarlocalization, while inactivation of mcsB acts indirectlythrough its effect on CtsR. By this logic, the absence ofMcsA or B would result in excess CtsR, which would thenmediate increased localization, either directly, or by regu-lating some other gene. To exclude a role for CtsR accu-mulation in the failure-to-delocalize phenotype, weconstructed a strain in which mcsB and ctsR were bothinactivated. If the mcsB phenotype resulted from excessCtsR, the double mutant would reverse the failure-to-delocalize phenotype. Clearly, the opposite is true; inac-tivation of ctsR failed to reverse the delocalizationphenotype of the mcsB mutant (Fig. 6) and we concludethat the delocalization phenotype is not mediated by aneffect on CtsR.
Phosphorylation of McsB is needed for delocalizationof ComGA
The phosphatase YwlE has been shown to mediate thedephosphorylation of McsB-PO4 as well as other proteins(Mijakovic et al., 2003; Kirstein et al., 2005). If phospho-rylation were involved in the McsB-dependent delocaliza-tion of Com proteins, we would expect that an increasedcellular concentration of YwlE would mimic the mcsBphenotype by decreasing the level of McsB~PO4. To testthis, we expressed ywlE from the IPTG-inducible Pspankpromoter. In the presence of inducer, a clear increase inthe localization of ComGA–GFP was observed, similar tothe effect of mcsB inactivation (Fig. 7A). A ywlE loss-of-function mutant exhibited no obvious alteration in thepattern of ComGA–GFP localization (not shown), sug-gesting that under these conditions the cells do not limitthe level of McsB-PO4 or that some other phosphataseaccomplishes this task. A further prediction of the hypoth-esis that phosphorylation of McsB is involved in delocal-ization is that inactivation of mcsA would present a similarphenotype, because the protein encoded by this gene isneeded to activate the autophosphorylation of McsB(Kirstein et al., 2005). This prediction was confirmedusing a non-polar mcsA knockout, and the delocalizationphenotypes of mcsA and mcsB knockouts are in factindistinguishable (Fig. 7B). These experiments stronglysuggest that phosphorylation of McsB is needed fordelocalization.
Delocalization of competence proteins 207
© 2009 The AuthorsJournal compilation © 2009 Blackwell Publishing Ltd, Molecular Microbiology, 72, 202–215

ClpC, ClpE and ClpP contribute to delocalization
These results, together with the above-mentioned role ofMcsA and B as adaptor proteins (Kirstein et al., 2007),
suggest that they may facilitate Com protein delocaliza-tion by degrading one or more unknown proteins that areneeded to anchor the Com proteins to the cell poles,consistent with the working model described above
Fig. 5. Effect of mcsB knockout on localization of ComFA, SsbB, DprA and YjbF. Fluorescence images were collected at T2 and T5 fromrespective isogenic wild-type and mcsB strains expressing ssbB–yfp (BD2988 and BD4121), comFA–yfp (BD3799 and BD4119), yfp–yjbF(BD4183 and BD4200) and yfp–dprA (BD4184 and BD4197). The yfp–yjbF and yfp–dprA fusions were expressed from Pspank promoters andthe strains were grown in competence medium with 1 mM IPTG.
208 J. Hahn, N. Kramer, K. Briley Jr and D. Dubnau �
© 2009 The AuthorsJournal compilation © 2009 Blackwell Publishing Ltd, Molecular Microbiology, 72, 202–215

derived from the time-lapse experiments conducted in thepresence and absence of Pm. If anchor protein degrada-tion were the explanation for the mcs phenotype, wewould expect loss-of-function mutants of clpP, clpC andclpE to exhibit delocalization defects similar to those ofthe mcsA and mcsB mutants. Although the individual clpEand clpC mutants had minor or no effects, a clpC clpEdouble mutant had a strong phenotype, consistent with
the likely abilities of McsA and B to act with either of theseAAA+ proteins (Fig. 8). To test the effect of clpP inactiva-tion, we used an spx clpP double mutant, since clpP isneeded for the full expression of competence while inac-tivation of spx bypasses this effect (Nakano et al., 2001;2002). Knockout of spx alone did not exhibit a delocaliza-tion defect (not shown), while the clpP spx strain exhibiteda clear defect in delocalization (Fig. 8). As also shown inFig. 8, inactivation of ctsR did not bypass the phenotypeof a double clpC clpE mutant, just as it failed to reversethat of the mcsB mutant. These data strongly support thehypothesis that degradation of one or more anchor pro-teins is needed for delocalization and is also consistentwith the energy requirement documented above, becauseAAA+ proteins require ATP hydrolysis to function andbecause McsB kinase activity is ATP-dependent.
McsB, ClpC and ClpP localize to the poles ofcompetent cells
If these proteins act to degrade a presumptive anchor,they presumably do so at the cell poles. We would there-fore expect them to exhibit polar localization and to colo-calize with Com proteins. To investigate this, fusions ofYFP to the C-termini of ClpC, McsB and ClpP, expressedfrom their native promoters, were used in a strain that alsoexpressed comGA–CFP to identify the competence-expressing cells (Fig. 9). In such cells, the ClpC, ClpP andMcsB fusions were localized at the poles and colocalizedwith ComGA–CFP. Although the McsB–YFP fusion doesnot interfere with heat-shock induction (Kirstein et al.,2008), it is not fully functional in the present assay, sincein its presence, the ComGA–CFP fusion exhibited a mcsBphenotype. This is noticeable in Fig. 9A where localizationis seen to persist at T5. Polar localization of McsB–YFPalmost never occurred in the non-competent cells, whichexhibited faint diffuse fluorescence, consistent with pub-lished results (Kirstein et al., 2008). Western blotting dem-onstrated that the amount of McsB protein is constantthroughout growth in competence medium (not shown).We do not know whether the apparent increase in McsB–YFP fluorescence in the competent cells is due to theconcentration of this protein at the poles, or to an increasein its synthesis or stability in the competent subfraction.Regardless of the mechanism, it appears that McsB istargeted to the poles in response to competence, possiblydue to its affinity for one or more Com proteins. ClpC–YFPsometimes localized to the poles of non-competent cells,as previously reported (Kain et al., 2008; Kirstein et al.,2008; Simmons et al., 2008), but was almost always local-ized in competent cells where it colocalized with ComGA(Fig. 9B). In other experiments, we have determined thatClpC and McsB accumulate at the cell poles indepen-dently; ClpC–YFP is at the poles of a mcsB knockout
Fig. 6. Inactivation of ctsR does not reverse the mcsB phenotype.Fluorescence images were collected at T5 from isogenic strainsexpressing comGA–gfp in wild-type (BD2899), mcsB (BD3594) andmcsB ctsR (BD4541) backgrounds.
Delocalization of competence proteins 209
© 2009 The AuthorsJournal compilation © 2009 Blackwell Publishing Ltd, Molecular Microbiology, 72, 202–215

Fig. 7. Phenotypes of ywlE-overexpressingand mcsA knockout mutants.A. Fluorescence images are shown taken atT5, from a strain expressing comGA–yfp andcarrying Pspank–ywlE (BD4770). The nativeywlE locus was intact in this strain. Imagesare shown from this culture grown with andwithout 1 mM IPTG to induce expression ofywlE.B. Fluorescence images are shown, taken atT5, from isogenic strains expressingcomGA–gfp in wild-type (BD2899), mcsB(BD3594) and mcsA (BD3655) backgrounds.
wild-type clpC clpE
clpC clpE clpP spx clpC clpE ctsR
Fig. 8. Phenotypes of ComGA–GFP localization in clp mutants. Fluorescence images are shown, taken at T5, of isogenic strains expressingcomGA–gfp in wild-type (BD2899 or BD3142), clpC (BD3579), clpE (BD4252), clpC clpE (BD4261), clpP spx (BD4530) and clpC clpE ctsR(BD4262) backgrounds. Note that the wild-type comparison with BD4530 is BD3142, expressing comGA–cfp. In many experiments we haveseen no difference in the patterns of ComGA–GFP or ComGA–CFP localization.
210 J. Hahn, N. Kramer, K. Briley Jr and D. Dubnau �
© 2009 The AuthorsJournal compilation © 2009 Blackwell Publishing Ltd, Molecular Microbiology, 72, 202–215
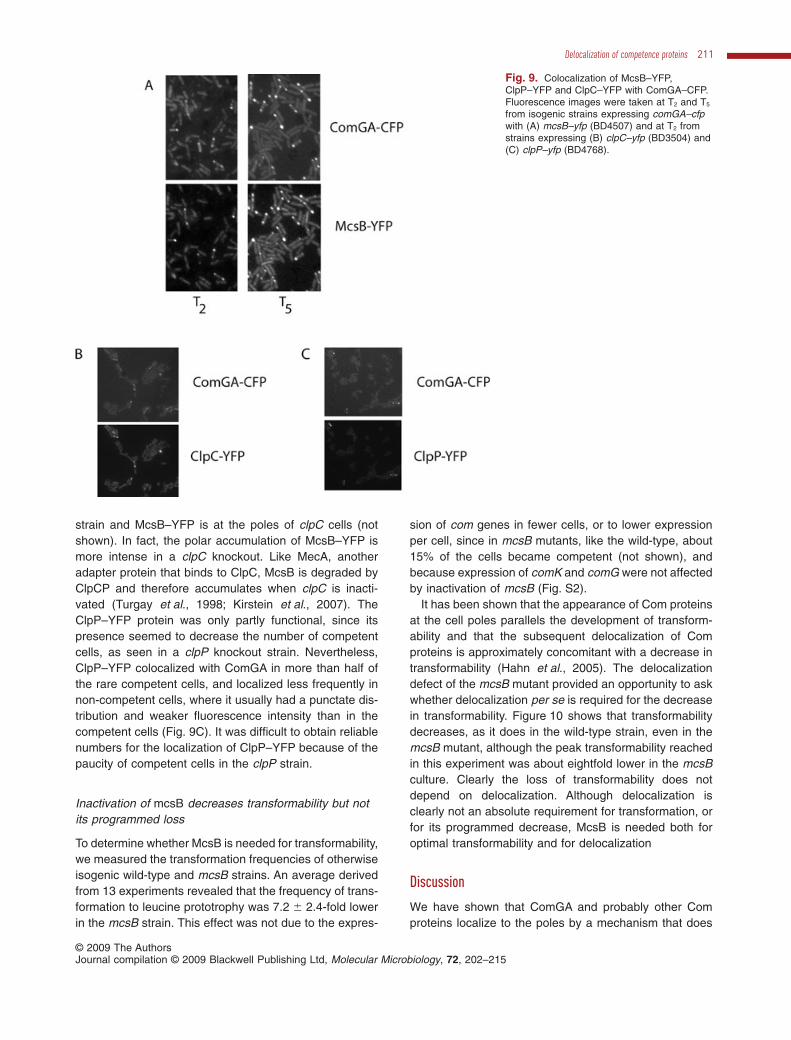
strain and McsB–YFP is at the poles of clpC cells (notshown). In fact, the polar accumulation of McsB–YFP ismore intense in a clpC knockout. Like MecA, anotheradapter protein that binds to ClpC, McsB is degraded byClpCP and therefore accumulates when clpC is inacti-vated (Turgay et al., 1998; Kirstein et al., 2007). TheClpP–YFP protein was only partly functional, since itspresence seemed to decrease the number of competentcells, as seen in a clpP knockout strain. Nevertheless,ClpP–YFP colocalized with ComGA in more than half ofthe rare competent cells, and localized less frequently innon-competent cells, where it usually had a punctate dis-tribution and weaker fluorescence intensity than in thecompetent cells (Fig. 9C). It was difficult to obtain reliablenumbers for the localization of ClpP–YFP because of thepaucity of competent cells in the clpP strain.
Inactivation of mcsB decreases transformability but notits programmed loss
To determine whether McsB is needed for transformability,we measured the transformation frequencies of otherwiseisogenic wild-type and mcsB strains. An average derivedfrom 13 experiments revealed that the frequency of trans-formation to leucine prototrophy was 7.2 � 2.4-fold lowerin the mcsB strain. This effect was not due to the expres-
sion of com genes in fewer cells, or to lower expressionper cell, since in mcsB mutants, like the wild-type, about15% of the cells became competent (not shown), andbecause expression of comK and comG were not affectedby inactivation of mcsB (Fig. S2).
It has been shown that the appearance of Com proteinsat the cell poles parallels the development of transform-ability and that the subsequent delocalization of Comproteins is approximately concomitant with a decrease intransformability (Hahn et al., 2005). The delocalizationdefect of the mcsB mutant provided an opportunity to askwhether delocalization per se is required for the decreasein transformability. Figure 10 shows that transformabilitydecreases, as it does in the wild-type strain, even in themcsB mutant, although the peak transformability reachedin this experiment was about eightfold lower in the mcsBculture. Clearly the loss of transformability does notdepend on delocalization. Although delocalization isclearly not an absolute requirement for transformation, orfor its programmed decrease, McsB is needed both foroptimal transformability and for delocalization
Discussion
We have shown that ComGA and probably other Comproteins localize to the poles by a mechanism that does
Fig. 9. Colocalization of McsB–YFP,ClpP–YFP and ClpC–YFP with ComGA–CFP.Fluorescence images were taken at T2 and T5
from isogenic strains expressing comGA–cfpwith (A) mcsB–yfp (BD4507) and at T2 fromstrains expressing (B) clpC–yfp (BD3504) and(C) clpP–yfp (BD4768).
Delocalization of competence proteins 211
© 2009 The AuthorsJournal compilation © 2009 Blackwell Publishing Ltd, Molecular Microbiology, 72, 202–215

not depend on metabolic energy and apparently pro-ceeds by diffusion and capture. Because Com proteinscolocalize even when non-polar (Hahn et al., 2005), andbecause the present experiments show that they moveto the poles from non-polar locations, we conclude thatat least some assembly of the Com machinery occursaway from the poles and that these complexes diffuseuntil they are captured at the poles. In some cells wehave seen large foci form, presumably due to the coa-lescence of individual assemblies. These foci sometimesmove to the poles and sometimes seem to disappear,perhaps due to disassembly. Although binding anduptake of transforming DNA occur preferentially at thepoles, we have detected less frequent transformationevents along the lateral surfaces of the rod-shaped cells(Hahn et al., 2005), consistent with non-polar assemblyof the Com machinery. It is intriguing that the non-polarfluorescence of the Com fusion proteins is helically dis-tributed and we are tempted to suggest that the pole-ward movement occurs along these helical paths.Indeed some of our movies can be interpreted in thisway (see Movie S2). However, it is also possible that themovement is not processive and that the helical tracksrepresent temporary binding sites. The Com proteinsmay attach and detach from these sites continually untilthey finally settle at the poles. Only high-speed videomicroscopy with sufficient spatial and temporal resolu-tion can address this question.
It appears that localization is a dynamic process, inthe sense that both localization and delocalization occurduring competence development, with McsA, McsB,ClpP and ClpC or ClpE required for delocalization. Thus,in the mcsB knockout strain, in which delocalization isblocked, localization is more complete and seems to
occur faster than in the wild-type strain. Also, in thepresence of NaN3, which also blocks delocalization,more net localization takes place than in its absence(Fig. 3). The extent of localization is thus the result ofcompeting processes and it is notable that following T2,the net localization declines. We do not know what regu-lates this equilibrium and what signals are involved.Perhaps the amounts of McsB and McsA or their phos-phorylation states are regulated temporally. Another tem-porally regulated process is responsible for the decreasein transformability that occurs after T2, because thisdecrease occurs independently of McsB (Fig. 10).Although the regulation of the proteins involved in delo-calization is complex (Fig. 4A), the repressor of all of thegenes involved (CtsR) is not directly required for thedelocalization process (Figs 6 and 8), a ctsR loss-of-function mutant exhibits reduced polar localization(Fig. 4B). This can easily be explained by over-production of mcsA, mcsB, clpP, clpC and clpE, consis-tent with the epistatic effects of each of these knockoutson the ctsR mutation.
An unidentified protein limits the rate of capture and isprogressively synthesized or activated as the cellsdevelop competence. Delocalization requires the action ofMcsA, McsB, ClpP and either ClpC or ClpE, presumablyto degrade a protein or proteins required to secure theCom proteins at the cell poles. It is tempting to regard therate-limiting capture protein and the McsB target as oneand the same. However, binding of the Com proteins tothe pole presumably requires at least two interacting sur-faces, one at the pole and another on the Com complex,and either of these surfaces may be formed by the rate-limiting protein or the proteolysis target. However, one lineof reasoning suggests that the McsB proteolysis targetmay be part of the Com protein complex itself, rather thana protein forming a binding site at the poles. Althoughinactivation of mcsB does not affect the transcription ortranslation of comK or comG, the amounts of ComGA andComFA are increased (Fig. S2), suggesting that theseproteins are stabilized in the mutant. Similarly, these pro-teins are stabilized when expressed in conjunction withother Com proteins. Our observations therefore suggestthat McsB targets a protein that facilitates the assembly ofthe Com machinery and is required to maintain its integ-rity, perhaps acting as a scaffold. We also suspect thatthis rate-limiting scaffold protein is under ComK control,because its amount appears to increase during the devel-opment of competence. It is worth pointing out that about100 genes are upregulated in the presence of ComK(Berka et al., 2002; Hamoen et al., 2002; Ogura et al.,2002), and that the roles of many of their encoded pro-teins remain undefined. These are all candidates for ascaffold protein. Clearly a fuller understanding of theseprocesses will require the experimental identification of
Fig. 10. Transformability of isogenic wild-type (BD2899, squares)and mcsB (BD3594, circles) strains during growth in competencemedium. Note the 10-fold difference in the scales on the ordinates.
212 J. Hahn, N. Kramer, K. Briley Jr and D. Dubnau �
© 2009 The AuthorsJournal compilation © 2009 Blackwell Publishing Ltd, Molecular Microbiology, 72, 202–215

proteins needed for the polar localization of the Comproteins.
The requirement for McsA, McsB, ClpP and eitherClpC or ClpE for delocalization implies strongly that thetarget protein is degraded using a mechanism similar tothat used to degrade CtsR, the only documented sub-strate for the McsAB adaptor proteins. Both ClpC andClpE bind to McsB (Kirstein et al., 2005; 2007) and ClpCand ClpE both are required for the regulation of CtsR(Miethke et al., 2006). These results are consistent withthe data in this report and suggest strongly that McsBacts as an adaptor protein for both ATPases. Thus wepropose that McsB, activated by its partner McsA, trans-fers one or more phosphoryl groups to the rate-limitingprotein, targeting it for degradation by the ClpC-ClpP orClpE-ClpP proteases. Over-production of the YwlE phos-phatase which is known to act on McsB~P (Kirsteinet al., 2005) mimics the phenotype conferred by inacti-vation of mcsA or mcsB, lending strong support to thismodel. Also highly suggestive are the localizations of thedegradation proteins. At least McsB, ClpP and ClpC arepresent near the poles of competent cells and all threeseem to colocalize with ComGA. Particularly intriguing isthe observation that McsB is only targeted to the polesin competent cells. The localization we have observedfor ClpC is consistent with three previous studies (Kainet al., 2008; Kirstein et al., 2008; Simmons et al., 2008),showing ClpC at or near the poles in some, but not allcells. One of these studies examined cells in a mediumthat would permit the development of competence(Simmons et al., 2008). In this medium, ClpC andClpE were associated with the nucleoids in mostcells, but were at the poles in 11–17% of the cells. Thepresent study would suggest that these cells might becompetent.
Clearly McsB is not required for transformation,although in its absence the yield of recoverable transfor-mants is depressed about sevenfold. The lack of an abso-lute requirement for McsB argues that the competencemachinery can assemble and function correctly in itsabsence. What might be the reason for the decrease intransformability? Although the Com proteins establishmultiple contacts with one another in competent cells,the competence machinery is most likely not a staticassembly. For instance, DprA is not as completely asso-ciated with the poles as are other Com proteins in mcsB +
cells (Fig. 5), although it does establish contacts withother Com proteins (Kramer et al., 2007). DprA is a DNA-binding protein and clearly interacts with the incomingsingle-stranded DNA following uptake. Indeed it has beenshown to protect this DNA from degradation (Berge et al.,2003; Mortier Barriere et al., 2007). It is likely that DprAassociates with the uptake apparatus during transport ofDNA across the membrane, so as to be positioned to
contact the incoming strands, but then departs to accom-pany these DNA strands to the chromosome where itfacilitates recombination, possibly by helping the DNAinteract with RecA (Mortier Barriere et al., 2007). It makessense therefore, that the association of DprA with theuptake apparatus is relatively loose, as observed. In themcsB knockout, DprA associates dramatically with theother Com proteins at the cell poles (Fig. 5), presumablybecause accumulation of the postulated scaffold proteinfavours this association. If as we suggest, loose associa-tion of DprA with the uptake proteins were important forrecombination, the lowered rate of transformation of themcsB mutant would be easily explained. More generally,we suggest that the complex must be dynamic to functionoptimally.
From the current state of knowledge, a picture emergesof complex regulated movement of proteins to particularcellular locations, targeted and regulated proteolysis anddynamic assembly disassembly processes that lead tothe proper development and loss of transformability.
Experimental procedures
Strains and microbiological methods
All the B. subtilis strains used in this study were derived fromthe 168 derivative BD630 (his leu met) (Table S1). Strainswere constructed by combining mutant constructs usingtransformation by standard methods, always with selectionfor the antibiotic resistance markers listed in Table S1. Theindividual mutant constructs were made as described in thereferences given in the table, except for the Phyperspank–ywlE, clpC–yfp and comGA–yfp constructs, which aredescribed in Supporting information. Strains were grown tothe indicated growth stages in competence medium (Albanoet al., 1987). When needed, Pm and Rm were added at200 mg ml-1.
Microscopy
For microscopy, cells taken at the indicated times weredeposited either on polylysine-coated slides or on 1.2%agarose pads, containing competence medium and inhibitorsas indicated. Images were collected in a Nikon 90i micro-scope using a TIRF Plan Neo-Fluor 100¥ oil immersion objec-tive (NA 1.45). Filter sets appropriate for each fluorophorewere from Semrock Optical. For the time-lapse experiments,a home-built temperature-controlled chamber that enclosesthe entire microscope was used (constructed by K.B.), andmaintained at 37°C. Chambers for time-lapse were con-structed from Chambered Cover glasses (Laboratory-Tek) asdescribed in Supporting information.
Western blotting
Western blots were carried out as described previously(Kramer et al., 2007), using polyclonal antisera raised in
Delocalization of competence proteins 213
© 2009 The AuthorsJournal compilation © 2009 Blackwell Publishing Ltd, Molecular Microbiology, 72, 202–215

rabbits. Rabbit anti-SsbB and anti-McsB antisera were kindgifts from Patrice Pollard and Kursad Turgay respectively.
Acknowledgements
This work was supported by NIH Grant GM057720. We thankU. Gerth, Qi Pan, R. Losick, L. Hamoen, P. Polard, M. Dias,D. Rudner, M. Nakano and P. Zuber for donating strains andantisera and N. Mirouze for the Pm luciferase experiment. Weare also grateful to all the members of the Dubnau andNeidtich labs for frequent and valuable discussions. We par-ticularly thank K. Turgay and J. Kirstein for donating strainsand antisera prior to publication and for sharing ideas andinformation.
References
Albano, M., Hahn, J., and Dubnau, D. (1987) Expression ofcompetence genes in Bacillus subtilis. J Bacteriol 169:3110–3117.
Berge, M., Mortier-Barriere, I., Martin, B., and Claverys, J.P.(2003) Transformation of Streptococcus pneumoniae relieson DprA- and RecA-dependent protection of incoming DNAsingle strands. Mol Microbiol 50: 527–536.
Berka, R.M., Hahn, J., Albano, M., Draskovic, I., Persuh, M.,Cui, X., et al. (2002) Microarray analysis of the Bacillussubtilis K-state: genome-wide expression changes depen-dent on ComK. Mol Microbiol 43: 1331–1345.
Chen, I., Provvedi, R., and Dubnau, D. (2006) A macromo-lecular complex formed by a pilin-like protein in competentBacillus subtilis. J Biol Chem 281: 21720–21727.
Chung, Y.S., Breidt, F., and Dubnau, D. (1998) Cell surfacelocalization and processing of the ComG proteins, requiredfor DNA binding during transformation of Bacillus subtilis.Mol Microbiol 29: 905–913.
Derré, I., Rapoport, G., and Msadek, T. (1999) CtsR, a novelregulator of stress and heat shock response, controls clpand molecular chaperone gene expression in gram-positive bacteria. Mol Microbiol 31: 117–131.
Derré, I., Rapoport, G., and Msadek, T. (2000) The CtsRregulator of stress response is active as a dimer and spe-cifically degraded in vivo at 37 degrees C. Mol Microbiol38: 335–347.
Dubnau, D., and Roggiani, M. (1990) Growth medium-independent genetic competence mutants of Bacillussubtilis. J Bacteriol 172: 4048–4055.
Edwards, D.H., and Errington, J. (1997) The Bacillus subtilisDivIVA protein targets to the division septum and controlsthe site specificity of cell division. Mol Microbiol 24: 905–915.
Hahn, J., Maier, B., Haijema, B.J., Sheetz, M., and Dubnau,D. (2005) Transformation proteins and DNA uptake localizeto the cell poles in Bacillus subtilis. Cell 122: 59–71.
Hamoen, L.W., Smits, W.K., de Jong, A., Holsappel, S., andKuipers, O.P. (2002) Improving the predictive value of thecompetence transcription factor (ComK) binding site inBacillus subtilis using a genomic approach. Nucleic AcidsRes 30: 5517–5528.
Kain, J., He, G.G., and Losick, R. (2008) Polar localizationand compartmentalization of ClpP proteases during growth
and sporulation in Bacillus subtilis. J Bacteriol 190: 6749–6757.
Kidane, D., and Graumann, P.L. (2005) Intracellular proteinand DNA dynamics in competent Bacillus subtilis cells. Cell122: 73–84.
Kirstein, J., Zuhlke, D., Gerth, U., Turgay, K., and Hecker, M.(2005) A tyrosine kinase and its activator control the activityof the CtsR heat shock repressor in B. subtilis. EMBO J 24:3435–3445.
Kirstein, J., Dougan, D.A., Gerth, U., Hecker, M., and Turgay,K. (2007) The tyrosine kinase McsB is a regulated adaptorprotein for ClpCP. EMBO J 26: 2061–2070.
Kirstein, J., Strahl, H., Moliere, N., Hamoen, L.W., andTurgay, K. (2008) Localization of general and regulatoryproteolysis in Bacillus subtilis cells. Mol Microbiol 70: 682–694.
Kramer, N., Hahn, J., and Dubnau, D. (2007) Multiple inter-actions among the competence proteins of Bacillus subtilis.Mol Microbiol 65: 454–464.
Krüger, E., and Hecker, M. (1998) The first gene of theBacillus subtilis clpC operon, ctsR, encodes a negativeregulator of its own operon and other class III heat shockgenes. J Bacteriol 180: 6681–6688.
Miethke, M., Hecker, M., and Gerth, U. (2006) Involvement ofBacillus subtilis ClpE in CtsR degradation and proteinquality control. J Bacteriol 188: 4610–4619.
Mijakovic, I., Poncet, S., Boel, G., Maze, A., Gillet, S., Jamet,E., et al. (2003) Transmembrane modulator-dependentbacterial tyrosine kinase activates UDP-glucosedehydrogenases. EMBO J 22: 4709–4718.
Mortier Barriere, I., Velten, M., Dupaigne, P., Mirouze, N.,Pietrement, O., McGovern, S., et al. (2007) A key presyn-aptic role in transformation for a widespread bacterialprotein: DprA conveys incoming ssDNA to RecA. Cell 130:824–836.
Nakano, M.M., Hajarizadeh, F., Zhu, Y., and Zuber, P. (2001)Loss-of-function mutations in yjbD result in ClpX- andClpP-independent competence development of Bacillussubtilis. Mol Microbiol 42: 383–394.
Nakano, M.M., Nakano, S., and Zuber, P. (2002) Spx (YjbD),a negative effector of competence in Bacillus subtilis,enhances ClpC–MecA–ComK interaction. Mol Microbiol44: 1341–1349.
Ogura, M., Yamaguchi, H., Kobayashi, K., Ogasawara, N.,Fujita, Y., and Tanaka, T. (2002) Whole-genome analysisof genes regulated by the Bacillus subtilis competencetranscription factor ComK. J Bacteriol 184: 2344–2351.
Ogura, T., and Wilkinson, A.J. (2001) AAA+ superfamilyATPases: common structure – diverse function. GenesCells 6: 575–597.
Oliver, D.B., Cabelli, R.J., Dolan, K.M., and Jarosik, G.P.(1990) Azide-resistant mutants of Escherichia coli alter theSecA protein, an azide-sensitive component of the proteinexport machinery. Proc Natl Acad Sci USA 87: 8227–8231.
Pan, Q., Garsin, D.A., and Losick, R. (2001) Self-reinforcingactivation of a cell-specific transcription factor by proteoly-sis of an anti-sigma factor in B. subtilis. Mol Cell 8: 873–883.
Shapiro, L., McAdams, H.H., and Losick, R. (2002) Generat-ing and exploiting polarity in bacteria. Science 298: 1942–1946.
214 J. Hahn, N. Kramer, K. Briley Jr and D. Dubnau �
© 2009 The AuthorsJournal compilation © 2009 Blackwell Publishing Ltd, Molecular Microbiology, 72, 202–215

Simmons, L.A., Grossman, A.D., and Walker, G.C. (2008)Clp and Lon proteases occupy distinct subcellular positionsin Bacillus subtilis. J Bacteriol 190: 6758–6768.
Tadesse, S., and Graumann, P.L. (2007) DprA/Smf proteinlocalizes at the DNA uptake machinery in competent Bacil-lus subtilis cells. BMC Microbiol 7: 105.
Turgay, K., Hahn, J., Burghoorn, J., and Dubnau, D. (1998)Competence in Bacillus subtilis is controlled by regulatedproteolysis of a transcription factor. EMBO J 17: 6730–6738.
Supporting information
Additional supporting information may be found in the onlineversion of this article.
Please note: Wiley-Blackwell are not responsible for thecontent or functionality of any supporting materials suppliedby the authors. Any queries (other than missing material)should be directed to the corresponding author for thearticle.
Delocalization of competence proteins 215
© 2009 The AuthorsJournal compilation © 2009 Blackwell Publishing Ltd, Molecular Microbiology, 72, 202–215









![Ligand Kedge X-ray Absorption Spectroscopy and DFT Calculations on [Fe 3 S 4 ] 0,+ Clusters: Delocalization, Redox, and Effect of the Protein Environment](https://static.fdokumen.com/doc/165x107/6314290dfc260b71020f6e78/ligand-kedge-x-ray-absorption-spectroscopy-and-dft-calculations-on-fe-3-s-4-0.jpg)



![Control Systems I - Lecture 6: Poles and Zeros [10pt] Readings:](https://static.fdokumen.com/doc/165x107/63346a354e43a4bcd80d3699/control-systems-i-lecture-6-poles-and-zeros-10pt-readings.jpg)






