
Mammals Have Two Twinfilin Isoforms Whose Subcellular Localizations and Tissue Distributions Are...
10
Mammals Have Two Twinfilin Isoforms Whose Subcellular Localizations and Tissue Distributions Are Differentially Regulated* Received for publication, April 8, 2003, and in revised form, June 2, 2003 Published, JBC Papers in Press, June 14, 2003, DOI 10.1074/jbc.M303642200 Maria K. Vartiainen‡§, Elisa M. Sarkkinen‡, Tanja Matilainen¶, Marjo Salminen¶, and Pekka Lappalainen‡ From the Programs in ‡Cellular Biotechnology and ¶Developmental Biology, Institute of Biotechnology, University of Helsinki, 00014 Helsinki, Finland Twinfilin is a highly conserved actin monomer-bind- ing protein that regulates cytoskeletal dynamics in or- ganisms from yeast to mammals. In addition to the pre- viously characterized mammalian twinfilin-1, a second protein with 65% sequence identity to twinfilin-1 ex- ists in mouse and humans. However, previous studies failed to identify any actin binding activity in this pro- tein (Rohwer, A., Kittstein, W., Marks, F., and Gsch- wendt, M. (1999) Eur. J. Biochem. 263, 518 –525). Here we show that this protein, which we named twinfilin-2, is indeed an actin monomer-binding protein. Similar to twinfilin-1, mouse twinfilin-2 binds ADP-G-actin with a higher affinity (K D 0.12 M) than ATP-G-actin (K D 1.96 M) and efficiently inhibits actin filament assembly in vitro. Both mouse twinfilins inhibit the nucleotide exchange on actin monomers and directly interact with capping protein. Furthermore, the actin interactions of mouse twinfilin-1 and twinfilin-2 are inhibited by phos- phatidylinositol (4,5)-bisphosphate. Although biochemi- cally very similar, our Northern blots and in situ hybrid- izations show that these two proteins display distinct expression patterns. Twinfilin-1 is the major isoform in embryos and in most adult mouse non-muscle cell-types, whereas twinfilin-2 is the predominant isoform of adult heart and skeletal muscles. Studies with isoform-spe- cific antibodies demonstrated that although the two proteins show similar localizations in unstimulated cells, they are regulated by different mechanisms. The small GTPases Rac1 and Cdc42 induce the redistribu- tion of twinfilin-1 to membrane ruffles and cell-cell con- tacts, respectively, but do not affect the localization of twinfilin-2. Taken together, these data show that mam- mals have two twinfilin isoforms, which are differen- tially expressed and regulated through distinct cellular signaling pathways. Actin is a conserved protein that is necessary for a large number of cellular processes including cell division, movement, polarized growth, secretion, and endocytosis. In cells, actin filaments form dynamic structures that rapidly assemble and disassemble in a coordinated fashion. Actin dynamics are tightly regulated by an array of actin-binding proteins that interact with actin filaments and/or monomeric actin. The pool of unpolymerized actin in non-muscle cells can constitute up to 50% of the total cytoplasmic actin, and thus the various actin monomer-binding proteins play an important role in actin dy- namics by regulating the size, localizations, and dynamics of the cellular actin monomer pool (1). Three families of small actin monomer-binding proteins are conserved in evolution, existing in organisms as diverse as yeast and mammals. These are profilins, ADF 1 /cofilins, and twinfilins. In addition to these evolutionarily conserved pro- teins, a fourth class of small actin monomer-binding proteins, the -thymosins, exists in vertebrates. -Thymosins are small peptides that bind specifically to ATP-G-actin, prevent actin filament assembly, and thus function as actin monomer- sequestering agents in certain specialized vertebrate cell types (2). Profilins are ubiquitous actin monomer-binding proteins with a molecular weight of 12–16 kDa. Profilins have a higher affinity for ATP-actin monomers than they do for ADP-G-actin, and they also interact with phospholipids and polyproline (1). These ubiquitous proteins can affect several different aspects of actin dynamics. In the absence of free filament-barbed ends, profilins function as actin monomer-sequestering proteins, whereas in the presence of free barbed ends, profilins promote actin filament assembly (3, 4). Most profilins also enhance the nucleotide exchange on actin monomers, and at least in yeasts, Saccharomyces cerevisiae and Schizosaccharomyces pombe, this activity is essential in vivo (5, 6). ADF/cofilins are small (molecular mass 15–19 kDa) essen- tial proteins composed of a single ADF-H domain. They interact with both actin monomers and filaments and bind ADP-actin with a higher affinity than they do to ATP-actin (7, 8). ADF/ cofilins enhance actin filament dynamics by depolymerizing filaments from their pointed ends (8) and by severing actin filaments, thereby increasing the amount of filament ends (9). Furthermore, ADF/cofilins inhibit the nucleotide exchange on actin monomers (10, 11). Twinfilin was originally identified from the yeast S. cerevi- siae by its sequence homology to ADF/cofilin proteins (12, 13). Twinfilin is composed of two ADF/cofilin-like domains (ADF-H domains) that are separated by a short linker region and flanked by a short tail region. The two ADF-H domains are 20% * This study was supported by grants from the Academy of Finland, Biocentrum Helsinki, Sigrid Juse ´lius Foundation, and the European Molecular Biology Organization (EMBO) Young Investigator Program (to P. L.). The costs of publication of this article were defrayed in part by the payment of page charges. This article must therefore be hereby marked “advertisement” in accordance with 18 U.S.C. Section 1734 solely to indicate this fact. § Present address: Division of Molecular Cell Biology, School of Bio- sciences, University of Birmingham, B15–2TT, Birmingham, England. To whom correspondence should be addressed: Institute of Biotech- nology, P. O. Box 56, 00014 University of Helsinki, Finland. Tel.: 358- 9-19159499; Fax: 358-9-19159366; E-mail: pekka.lappalainen@ helsinki.fi. 1 The abbreviations used are: ADF, actin-depolymerizing factor; GST, glutathione S-transferase; CNS, central nervous system; DTT, dithio- threitol; PI(4,5)P 2, phosphatidylinositol (4,5)-bisphosphate; PNS, pe- ripheral nervous system; E, embryonic day. THE JOURNAL OF BIOLOGICAL CHEMISTRY Vol. 278, No. 36, Issue of September 5, pp. 34347–34355, 2003 © 2003 by The American Society for Biochemistry and Molecular Biology, Inc. Printed in U.S.A. This paper is available on line at http://www.jbc.org 34347 by guest on March 27, 2016 http://www.jbc.org/ Downloaded from
-
Upload
independent -
Category
Documents
-
view
1 -
download
0
Transcript of Mammals Have Two Twinfilin Isoforms Whose Subcellular Localizations and Tissue Distributions Are...

Mammals Have Two Twinfilin Isoforms Whose SubcellularLocalizations and Tissue Distributions Are Differentially Regulated*
Received for publication, April 8, 2003, and in revised form, June 2, 2003Published, JBC Papers in Press, June 14, 2003, DOI 10.1074/jbc.M303642200
Maria K. Vartiainen‡§, Elisa M. Sarkkinen‡, Tanja Matilainen¶, Marjo Salminen¶,and Pekka Lappalainen‡�
From the Programs in ‡Cellular Biotechnology and ¶Developmental Biology, Institute of Biotechnology,University of Helsinki, 00014 Helsinki, Finland
Twinfilin is a highly conserved actin monomer-bind-ing protein that regulates cytoskeletal dynamics in or-ganisms from yeast to mammals. In addition to the pre-viously characterized mammalian twinfilin-1, a secondprotein with �65% sequence identity to twinfilin-1 ex-ists in mouse and humans. However, previous studiesfailed to identify any actin binding activity in this pro-tein (Rohwer, A., Kittstein, W., Marks, F., and Gsch-wendt, M. (1999) Eur. J. Biochem. 263, 518–525). Here weshow that this protein, which we named twinfilin-2, isindeed an actin monomer-binding protein. Similar totwinfilin-1, mouse twinfilin-2 binds ADP-G-actin with ahigher affinity (KD � 0.12 �M) than ATP-G-actin (KD �1.96 �M) and efficiently inhibits actin filament assemblyin vitro. Both mouse twinfilins inhibit the nucleotideexchange on actin monomers and directly interact withcapping protein. Furthermore, the actin interactions ofmouse twinfilin-1 and twinfilin-2 are inhibited by phos-phatidylinositol (4,5)-bisphosphate. Although biochemi-cally very similar, our Northern blots and in situ hybrid-izations show that these two proteins display distinctexpression patterns. Twinfilin-1 is the major isoform inembryos and in most adult mouse non-muscle cell-types,whereas twinfilin-2 is the predominant isoform of adultheart and skeletal muscles. Studies with isoform-spe-cific antibodies demonstrated that although the twoproteins show similar localizations in unstimulatedcells, they are regulated by different mechanisms. Thesmall GTPases Rac1 and Cdc42 induce the redistribu-tion of twinfilin-1 to membrane ruffles and cell-cell con-tacts, respectively, but do not affect the localization oftwinfilin-2. Taken together, these data show that mam-mals have two twinfilin isoforms, which are differen-tially expressed and regulated through distinct cellularsignaling pathways.
Actin is a conserved protein that is necessary for a largenumber of cellular processes including cell division, movement,polarized growth, secretion, and endocytosis. In cells, actin
filaments form dynamic structures that rapidly assemble anddisassemble in a coordinated fashion. Actin dynamics aretightly regulated by an array of actin-binding proteins thatinteract with actin filaments and/or monomeric actin. The poolof unpolymerized actin in non-muscle cells can constitute up to50% of the total cytoplasmic actin, and thus the various actinmonomer-binding proteins play an important role in actin dy-namics by regulating the size, localizations, and dynamics ofthe cellular actin monomer pool (1).
Three families of small actin monomer-binding proteins areconserved in evolution, existing in organisms as diverse asyeast and mammals. These are profilins, ADF1/cofilins, andtwinfilins. In addition to these evolutionarily conserved pro-teins, a fourth class of small actin monomer-binding proteins,the �-thymosins, exists in vertebrates. �-Thymosins are smallpeptides that bind specifically to ATP-G-actin, prevent actinfilament assembly, and thus function as actin monomer-sequestering agents in certain specialized vertebrate celltypes (2).
Profilins are ubiquitous actin monomer-binding proteinswith a molecular weight of 12–16 kDa. Profilins have a higheraffinity for ATP-actin monomers than they do for ADP-G-actin,and they also interact with phospholipids and polyproline (1).These ubiquitous proteins can affect several different aspects ofactin dynamics. In the absence of free filament-barbed ends,profilins function as actin monomer-sequestering proteins,whereas in the presence of free barbed ends, profilins promoteactin filament assembly (3, 4). Most profilins also enhance thenucleotide exchange on actin monomers, and at least in yeasts,Saccharomyces cerevisiae and Schizosaccharomyces pombe,this activity is essential in vivo (5, 6).
ADF/cofilins are small (molecular mass � 15–19 kDa) essen-tial proteins composed of a single ADF-H domain. They interactwith both actin monomers and filaments and bind ADP-actinwith a higher affinity than they do to ATP-actin (7, 8). ADF/cofilins enhance actin filament dynamics by depolymerizingfilaments from their pointed ends (8) and by severing actinfilaments, thereby increasing the amount of filament ends (9).Furthermore, ADF/cofilins inhibit the nucleotide exchange onactin monomers (10, 11).
Twinfilin was originally identified from the yeast S. cerevi-siae by its sequence homology to ADF/cofilin proteins (12, 13).Twinfilin is composed of two ADF/cofilin-like domains (ADF-Hdomains) that are separated by a short linker region andflanked by a short tail region. The two ADF-H domains are 20%
* This study was supported by grants from the Academy of Finland,Biocentrum Helsinki, Sigrid Juselius Foundation, and the EuropeanMolecular Biology Organization (EMBO) Young Investigator Program(to P. L.). The costs of publication of this article were defrayed in part bythe payment of page charges. This article must therefore be herebymarked “advertisement” in accordance with 18 U.S.C. Section 1734solely to indicate this fact.
§ Present address: Division of Molecular Cell Biology, School of Bio-sciences, University of Birmingham, B15–2TT, Birmingham, England.
� To whom correspondence should be addressed: Institute of Biotech-nology, P. O. Box 56, 00014 University of Helsinki, Finland. Tel.: 358-9-19159499; Fax: 358-9-19159366; E-mail: [email protected].
1 The abbreviations used are: ADF, actin-depolymerizing factor; GST,glutathione S-transferase; CNS, central nervous system; DTT, dithio-threitol; PI(4,5)P2, phosphatidylinositol (4,5)-bisphosphate; PNS, pe-ripheral nervous system; E, embryonic day.
THE JOURNAL OF BIOLOGICAL CHEMISTRY Vol. 278, No. 36, Issue of September 5, pp. 34347–34355, 2003© 2003 by The American Society for Biochemistry and Molecular Biology, Inc. Printed in U.S.A.
This paper is available on line at http://www.jbc.org 34347
by guest on March 27, 2016
http://ww
w.jbc.org/
Dow
nloaded from

homologous to ADF/cofilin and to each other (12). Althoughtwinfilin shares similar building blocks to ADF/cofilins, thesetwo proteins have profound differences in their biochemicalproperties. Whereas ADF/cofilins bind both actin monomersand filaments, twinfilins interact only with monomeric actinand do not promote filament depolymerization (13–15). Similarto ADF/cofilins, twinfilins bind ADP-G-actin with a higheraffinity than they do to ATP-G-actin and have slower koff ratesfrom actin monomers and prevent filament assembly muchmore efficiently than ADF/cofilins (16, 17). Twinfilins also in-teract with heterodimeric capping proteins, and their physio-logical function may be to localize actin monomers to the cel-lular sites of rapid actin dynamics (16, 18).
In multicellular organisms, there are typically several iso-forms of both ADF/cofilin and profilin. The three mammalianADF/cofilin isoforms (cofilin-1, cofilin-2, and ADF) are ex-pressed in distinct tissues, and their affinities for actin mono-mers and their effects on actin filament depolymerization andnucleation differ. This finding suggests that the three mamma-lian ADF/cofilin isoforms have evolved to fulfill the specificrequirements for actin dynamics in differentiated cell types(19, 20). Also the three profilin isoforms in mammals (profilin I,profilin IIa, and profilin IIb) show cell type-specific expressionpatterns (21, 22) and display differences in their affinities foractin and their other ligands: polyproline and phospholipids(21–24). To examine whether there are also several twinfilinisoforms in mammals, we carried out searches on mouse andhuman sequence databases. These searches revealed onemouse and one human protein with �65% sequence identitiesto previously characterized mouse and human twinfilin-1.Here, we show that the mouse protein, which we named twin-filin-2, is an actin monomer-binding protein with similar bio-chemical activities to other known twinfilins. Our analysis alsorevealed that the two mouse twinfilin isoforms have distinctexpression patterns. Twinfilin-1 is expressed with variable lev-els in most cell types of the developing and adult mouse,whereas elevated levels of twinfilin-2 expression was detectedin maturing skeletal and heart muscles. Furthermore, ourstudies demonstrated that the subcellular localizations of thetwo mouse twinfilin isoforms are regulated through differentsignaling pathways.
EXPERIMENTAL PROCEDURES
Plasmid Construction—The DNA fragment corresponding to mousetwinfilin-2 cDNA was amplified by PCR from an embryonic days(E)7–20 mouse embryo cDNA library. The oligonucleotides introducedNcoI and HindIII sites at the 5� and 3� ends of the PCR product,respectively. The PCR fragment was digested with NcoI and HindIIIand ligated into the pGAT2 vector (25) to create plasmid pPL68. Theplasmid for expressing mouse twinfilin-1 as a glutathione S-transferase(GST) fusion protein (pPL78) has been described previously (14). Forexpression of His-tagged versions of twinfilin-1 and twinfilin-2 in Esch-erichia coli, the plasmids pPL78 and pPL68 were digested with NcoIand HindIII and the cDNA fragments were cloned into the pHAT2vector (25) to create plasmids pPL144 and pPL182. For in situ hybrid-izations, the full-length mouse twinfilin-1 and twinfilin-2 cDNAs weresubcloned into pBSIIKS vector to yield plasmids pPL184 and pPL185.The vectors for expressing mouse �1�2 and chicken �1�1-capping pro-teins in E. coli were kindly provided by Drs. John Cooper (WashingtonUniversity, St. Louis, MO) and Takashi Obinata (Chiba University,Chiba, Japan), and have been described previously (16, 26). The plas-mids for expressing Rho family GTPases in mammalian cells have beendescribed previously (14).
Proteins—The expression and purification of mouse twinfilin-1 andtwinfilin-2 as GST fusion proteins from the plasmids pPL68 and pPL78were carried out as described previously (14). For expressing His-tagged mouse twinfilins, 6000 ml of E. coli BL21(DE3) cells transformedwith plasmid pPL144 (twinfilin-1) or pPL182 (twinfilin-2) were grownin Luria broth medium to an optical density of 0.6 at 600 nm andexpression was induced with 0.2 mM isopropyl-�-D-thiogalactopyrano-side. Cells were harvested 3 h after induction, washed once with 20 mM
Tris (pH 7.5), resuspended to 40 ml of 20 mM imidazole, 50 mM NaCl, 10mM Tris (pH 7.5), lysed by sonication, and centrifuged for 30 min at30,000 � g. The supernatant was loaded onto a nickel-nitrilotriaceticacid-agarose column (Qiagen) and eluted with a linear 20–250 mM
imidazole gradient. The peak fractions containing the His-tagged twin-filins were concentrated to 1.5 ml in Centricon 10-kDa cutoff devicesand loaded onto a Superdex-75 HiLoad gel filtration column (Amer-sham Biosciences), which had been equilibrated with 10 mM Tris (pH7.5), 50 mM NaCl. The peak fractions containing twinfilins were pooled,concentrated in Centricon 10-kDa cutoff devices to a final concentrationof 100–300 �M, frozen in liquid N2, and stored at �70 °C. Chicken �1�1
and mouse �1�2-capping proteins were produced in E. coli BL21(DE3)cells transformed with pET expression plasmids containing the cDNAsfor � and � subunits of capping protein. Cells were grown in 6000 ml ofLuria broth medium to an optical density of 0.6 at 600 nm, and expres-sion was induced with 0.2 mM isopropyl-�-D-thiogalactopyranoside.Cells were harvested by 3 h after induction, washed once with 20 mM
Tris (pH 7.5), resuspended in 20 mM Tris (pH 7.5), and lysed by soni-cation. The cell lysate was centrifuged in a Beckmann SS-34 rotor at10,000 rpm for 30 min at 4 °C, and the supernatant was further cen-trifuged in a Beckman Ti45 rotor at 30,000 rpm for 30 min at 4 °C. Thesupernatant from the second centrifugation was applied onto a Q-Sepharose Fast Flow anion exchange column (Amersham Biosciences)equilibrated with 20 mM Tris (pH 7.5), and the proteins were elutedwith a 0–1 M NaCl gradient. The fractions containing the cappingprotein were pooled, concentrated to a volume of �2 ml, diluted 1:20 in20 mM KH2PO4 (pH 7.0), and applied onto a hydroxyapatite column(Bio-Rad) that had been equilibrated in 20 mM KH2PO4 (pH 7.0). Pro-teins were eluted with a linear 20–250 mM KH2PO4 gradient, and thefractions containing capping protein were pooled, concentrated to 1.5ml, and applied onto a Superdex-75 column equilibrated in 10 mM Tris(pH 7.5), 50 mM NaCl. The peak fractions containing the cappingprotein were concentrated to 100–300 �M and stored at �70 °C. Rabbitmuscle actin was prepared from acetone powder as described previously(27), and the recombinant mouse cofilin-1 was purified as described byVartiainen et al. (19). The human platelet actin and pyrene-labeledactin were from the Cytoskeleton Inc.
Actin Monomer-binding Assays—The change in the fluorescence of7-chloro-4-nitrobenz-2-oxa-1,3-diazole (NBD)-labeled G-actin was usedto monitor the binding of twinfilins to actin monomers. Actin waslabeled by NBD-Cl as described previously (28, 29). ADP-actin wasprepared by incubating NBD-actin with hexokinase-agarose beads(Sigma) and glucose for 2 h at �4 °C (30). The final concentration ofactin in these assays was 0.2 �M, and the twinfilin concentration wasvaried from 0.05 to 14 �M. Experiments were carried out at roomtemperature in F-buffer (2 mM Tris-HCl (pH 8.0), 0.1 mM CaCl2, 0.1 mM
DTT, 0.2 mM ADP or ATP, 1 mg/ml bovine serum albumin, 2 mM MgCl2,0.1 M KCl). The data were analyzed as described previously (17). Therate of nucleotide exchange on actin monomers in the absence andpresence of twinfilin was measured as described previously (10).
Actin Assembly Assays—Kinetics of actin filament assembly wasmonitored by pyrene fluorescence with excitation at 365 nm and emis-sion at 407 nm using a BioLogic MOS-250 fluorescence spectrophotom-eter. 54 �l of 3.9 �M actin (1:6 pyrene actin:human platelet actin) inG-buffer (5 mM Tris (pH 7.5), 0.2 mM ATP, 0.2 mM DTT, 0.2 mM CaCl2)was mixed with 7 �l of G-buffer or 15/30 �M twinfilin-1 or twinfilin-2 inthe presence or absence of PI(4,5)P2 (Matreya Inc). Polymerization wasinduced by the addition of 7 �l of 10� initiation mixture (1 M KCl, 20mM MgCl2, 5 mM ATP).
Native Gel Electrophoresis—The interaction of mouse twinfilins withPI(4,5)P2 was examined by native gel electrophoresis as described foryeast twinfilin (16). Final concentrations of twinfilin in these assayswere 4 �M, and the PI(4,5)P2 concentrations were 0, 20, or 30 �M.Twinfilin-capping protein interactions were studied on 6% native poly-acrylamide gels. Purified mouse twinfilins and chicken �1�1- or mouse�1�2-capping protein (either alone or in a mixture with each other) wasdiluted to desired concentrations in 10 mM Tris (pH 7.5), 50 mM NaCl,0.5 mM DTT and incubated for 60 min at room temperature. 15-�laliquots of the protein samples were then mixed with 5 �l of loadingbuffer (125 mM Tris (pH 8.8), 250 mM NaCl, 2.5 mM DTT, 50% glycerol)and loaded onto a 6% polyacrylamide gel. The gel was run for 150 minat 100 V by using buffer (25 mM Tris, 194 mM glycine (pH 8.5), 0.5 mM
DTT), and the proteins were detected either by Coomassie Blue stainingor by Western blotting with anti-twinfilin antibodies.
Northern Blotting—Twinfilin-1 and twinfilin-2 cDNA probes wereprepared as described previously (14). Both twinfilin probes were hy-bridized to commercial mouse embryo and adult mouse multiple tissueNorthern blots (Clontech) according to manufacturer’s instructions.
Mammals Have Two Twinfilin Isoforms34348
by guest on March 27, 2016
http://ww
w.jbc.org/
Dow
nloaded from

The Northern blot filters were then exposed on a PhosphorImagerscreen for 120 min (with the exception of 600 min in the case of embryoblot for twinfilin-2).
RNA in Situ Hybridization—Radioactive in situ hybridization assayswere performed on sections from paraffin-embedded mouse embryos oradult tissues as described previously (31). The 35S-labeled antisenseand sense RNA probes for twinfilin-1 and twinfilin-2 were transcribedfrom plasmids pPL184 and pPL185, respectively.
Antibodies—Generation of rabbit anti-mouse twinfilin-1 antibodyhas been described previously (14). However, because our studiesshowed that the antibody also cross-reacted with mouse twinfilin-2, thisantibody was further purified by removing the twinfilin-2-recognizingfraction by a twinfilin-2 affinity column. Chicken anti-mouse twinfilin-2IgY was generated in Agrisera Inc (Vannas, Sweden) by immunizingchicken with recombinant His-tagged twinfilin-2. For affinity purifica-tion, recombinant twinfilin-2 was immobilized to CNBr-activatedSepharose 4B beads (Amersham Biosciences) and the chicken antibodywas purified from 2 ml of serum according to manufacturer’s instruc-tions. Because this antibody also cross-reacted with twinfilin-1, thefraction recognizing twinfilin-1 was removed by a twinfilin-1 affinitycolumn.
Cell Culture, Immunofluorescence, and Western Blotting—NIH 3T3and Neuro2A cells were maintained in Dulbecco’s modified Eagle’smedium supplemented with 10% fetal bovine serum (Invitrogen), 2 mM
L-glutamine, 100 units/ml penicillin, and 100 �g/ml streptomycin. Forimmunofluorescence studies, Neuro2A cells were grown on glass cover-slips coated with poly-L-ornithine (1 mg/ml). For studies with dominantactive RhoA, Rac1, and Cdc42, NIH 3T3 cells were transfected with 1.5�g of plasmids using the Roche Applied Science FuGENE 6 transfectionsystem according to manufacturer’s instructions. Immunofluorescencemicroscopy was performed as described previously (14). Anti-twinfilin-1antibody was used at a 1:500 dilution and anti-twinfilin-2 at a 1:300dilution. Rhodamine-phalloidin (Molecular Probes) was used at a 1:300dilution. For Western blotting, cell lysates were prepared as describedpreviously (14) and the total protein concentrations were measuredusing Bradford reagent (Sigma) according to manufacturer’s instruc-tions. Western blotting was performed according to instructions fromAgrisera Inc. Anti-twinfilin-1 antibody was used at a 1:1000 dilution,anti-twinfilin-2 was used at 1:500, and anti-cofilin-1 was used at1:1000. The horseradish peroxidase-conjugated anti-chicken and anti-rabbit secondary antibodies (Jackson Inc.) were used at a 1:15,000dilution.
Miscellaneous—Protein concentrations were determined with aHewlett-Packard 8452A diode array spectrophotometer using calcu-
lated extinction coefficients at A280 for mouse twinfilin-1 (� � 36,400M�1 cm�1), mouse twinfilin-2 (� � 31,300 M�1 cm�1), mouse cofilin-1 (� �13,490 M�1 cm�1), and chicken �1�1-capping protein (� � 78,270 M�1
cm�1) and at A290 for actin (� � 26,600 M�1 cm�1).
RESULTS
Twinfilin is an actin monomer-binding protein that regulatescytoskeletal dynamics in yeast (13), fruitfly (15), and mousecells (14). In our data base searches, we identified a new twin-filin-related protein in human and mouse and named it twin-filin-2. The human and mouse twinfilin-2 are 97% identical atthe amino acid level and show �65% sequence identity to thepreviously characterized human and mouse twinfilin-1 (14).The mammalian twinfilin-2 proteins appear to have similardomain structures to other known twinfilins, and the residuesthat were shown to be important for actin monomer binding inyeast twinfilin (16) and mouse twinfilin-1 (32) are well con-served in twinfilin-2 (Fig. 1). However, the human homologueof twinfilin-2 (A6-related protein) had been previously identi-fied as a binding partner of the catalytic domain of proteinkinase C�, and these earlier studies failed to detect any actin-related activities in this protein. These data suggested thatdespite the strong sequence homology to other twinfilins, itwould not be an actin-binding protein (33).
Mouse Twinfilin-2 Is an Actin Monomer-binding Protein—Toexamine the biochemical activities of mouse twinfilin-2, weexpressed it as a GST fusion protein in E. coli cells. GST wassubsequently removed by thrombin digestion, and the recom-binant protein was further purified by gel-filtration chromatog-raphy. The purified recombinant mouse twinfilin-2 was fullysoluble and monomeric according to its elution position from agel-filtration column.
We recently showed that the fluorescence of muscle NBD-G-actin is modulated upon binding to mouse twinfilin-1, therebyproviding a means to determine the affinity of twinfilin-1/actinmonomer complex (17). Therefore, we examined whethermouse twinfilin-2 would also affect the fluorescence of NBD-G-actin. The assays were carried out under physiological ionic
FIG. 1. Multiple sequence align-ment of mouse (mm) and human (hs)twinfilins. Residues that have beenshown to be important for actin monomerbinding in yeast twinfilin (16) or mousetwinfilin-1 (32) are indicated above thesequences with asterisks. The accessionnumbers for the sequences are humantwinfilin-1 (Protein Information Resourceaccession number A55922); mouse twin-filin-1 (GenBankTM accession numberAY267188); human twinfilin-2 (Gen-BankTM accession number Y17168); andmouse twinfilin-2 (GenBankTM accessionnumber AY267189).
Mammals Have Two Twinfilin Isoforms 34349
by guest on March 27, 2016
http://ww
w.jbc.org/
Dow
nloaded from

concentrations and by using NBD-labeled platelet actin. Thebinding of twinfilin-2 resulted in a 50% enhancement in thefluorescence of NBD-G-actin, indicating that twinfilin-2 alsointeracts with actin monomers (Fig. 2). The KD values of twin-filin-2 for ADP-G-actin and ATP-G-actin were 0.12 and 1.96�M, respectively. A similar assay with twinfilin-1 provided KD
values of 0.09 �M (ADP-G-actin) and 1.77 �M (ATP-G-actin).Therefore, both mouse twinfilins bind ADP-G-actin with � 20-fold higher affinity than the ATP-G-actin, and the affinities ofboth proteins for actin monomers are very similar to eachother. Although both twinfilins appear to bind platelet ADP-G-actin with similar affinities (KD � 0.09–0.12 �M) than pre-viously reported for muscle ADP-G-actin/twinfilin-1 complex(KD � 0.05 �M) (17), the affinities of these proteins for plateletATP-G-actin (KD � 1.77–1.96 �M) (Fig. 2B) are significantlylower than the affinity of twinfilin-1 for muscle ATP-G-actin(KD � 0.47 �M) (17).
We also used an actin co-sedimentation assay to examine thepossible interactions of mouse twinfilins with actin filaments.Neither twinfilin-1 nor twinfilin-2 co-sedimented with actinfilaments, providing further support for the previous conclu-sions that twinfilins do not interact with F-actin (13, 15). How-ever, both twinfilin-1 and twinfilin-2 increased the amount ofmonomeric actin in F-actin co-sedimentation assays, indicatingthat they inhibit actin filament assembly in vitro (data notshown).
Previous studies on yeast twinfilin (13) showed that thisprotein decreases the rate of nucleotide exchange of G-actin.
The ability of mouse twinfilin-1 and twinfilin-2 to inhibit theexchange of �-ATP to ATP on actin monomers was measured byusing 1 �M G-actin and 0–3 �M twinfilins. The results showedthat both twinfilin-1 and twinfilin-2 efficiently inhibit the spon-taneous nucleotide exchange on G-actin in a concentration-de-pendent manner (Fig. 3).
Interactions of Mouse Twinfilin Isoforms with PI(4,5)P2 andCapping Protein—Phosphoinositides, especially PI(4,5)P2,modulate the activities of many actin-binding proteins such asADF/cofilin (34) and profilin (35). Previous studies showed thatyeast twinfilin also interacts with phosphoinositides and thatPI(4,5)P2 inhibits its actin monomer-sequestering activity invitro (16). To examine whether mouse twinfilins also interactwith PI(4,5)P2, we carried out a native gel electrophoresisassay (Fig. 4A). In this assay, 4 �M mouse twinfilin-1 or twin-filin-2 were loaded onto a native gel either alone or in a mixturewith 20 or 30 �M PI(4,5)P2 and run toward the anode by theirown charges. As shown in Fig. 4A, purified twinfilins migrateas a single sharp band, but when mixed with PI(4,5)P2 beforeloading on a gel, twinfilins migrate as a smear. This shows thatboth mouse twinfilins directly interact with PI(4,5)P2.
To examine the effects of PI(4,5)P2 on the actin bindingactivity of mouse twinfilins, we carried out a pyrene-actinassembly assay (Fig. 4B). Both mouse twinfilin-1 and twin-filin-2 inhibit the nucleation/polymerization of pyrene-actin ina concentration-dependent manner. However, when twinfilinswere preincubated with PI(4,5)P2 before the assay, they weresignificantly less effective in inhibiting the pyrene-actin assem-bly. 3 �M twinfilin-1 and twinfilin-2 with 30 �M PI(4,5)P2 in-hibit actin assembly as efficiently as 1.5 �M twinfilin withoutPI(4,5)P2, suggesting that 30 �M PI(4,5)P2 down-regulates theactivity of twinfilin by �50% (Fig. 4B). It is important to notethat higher concentrations of PI(4,5)P2 decreased twinfilin ac-tivity even further, but they also affected the pyrene-actinsignal (data not shown).
In addition to actin and phospholipids, yeast twinfilin bindscapping protein (Cap1/2p). This interaction appears to be es-sential for the correct subcellular localization of twinfilin inyeast cells (16). Furthermore, mouse twinfilin-1 was shown toalso interact with �1�2-capping protein in vitro (16). To exam-ine whether mouse twinfilin-2 also interacts with capping pro-tein and whether the two mouse twinfilins would have anyspecificity toward certain capping protein isoforms, we carriedout a native gel electrophoresis assay with purified twinfilinsand chicken �1�1 and mouse �1�2-capping proteins. �1�1 is the
FIG. 2. Binding of mouse twinfilins (Twf) to actin monomers.The increase in fluorescence of 0.2 �M NBD-labeled MgADP-G-actin (A)or MgATP-actin (B) was measured at different concentrations of twin-filin-1 or twinfilin-2 under physiological ionic conditions. Symbols aredata, and lines are fitted binding curves for a complex with 1:1 stoichi-ometry. Each data point on the graph is a mean value of three inde-pendent experiments. Dissociation constants (KD) derived from thebinding curves are indicated in the figure.
FIG. 3. Mouse twinfilins (Twf) inhibit the nucleotide exchangeon actin monomers. The concentrations of twinfilin-1 (triangles) andtwinfilin-2 (squares) are shown on the x axis, and the rates of nucleotideexchange are indicated on the y axis.
Mammals Have Two Twinfilin Isoforms34350
by guest on March 27, 2016
http://ww
w.jbc.org/
Dow
nloaded from
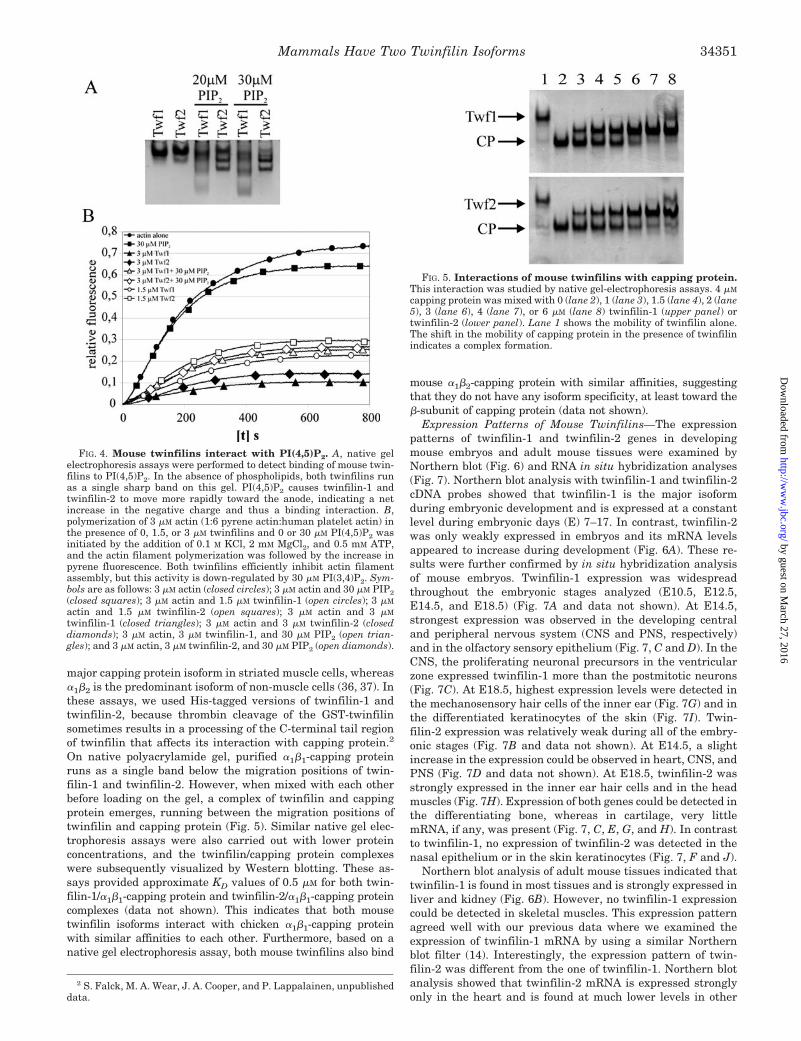
major capping protein isoform in striated muscle cells, whereas�1�2 is the predominant isoform of non-muscle cells (36, 37). Inthese assays, we used His-tagged versions of twinfilin-1 andtwinfilin-2, because thrombin cleavage of the GST-twinfilinsometimes results in a processing of the C-terminal tail regionof twinfilin that affects its interaction with capping protein.2
On native polyacrylamide gel, purified �1�1-capping proteinruns as a single band below the migration positions of twin-filin-1 and twinfilin-2. However, when mixed with each otherbefore loading on the gel, a complex of twinfilin and cappingprotein emerges, running between the migration positions oftwinfilin and capping protein (Fig. 5). Similar native gel elec-trophoresis assays were also carried out with lower proteinconcentrations, and the twinfilin/capping protein complexeswere subsequently visualized by Western blotting. These as-says provided approximate KD values of 0.5 �M for both twin-filin-1/�1�1-capping protein and twinfilin-2/�1�1-capping proteincomplexes (data not shown). This indicates that both mousetwinfilin isoforms interact with chicken �1�1-capping proteinwith similar affinities to each other. Furthermore, based on anative gel electrophoresis assay, both mouse twinfilins also bind
mouse �1�2-capping protein with similar affinities, suggestingthat they do not have any isoform specificity, at least toward the�-subunit of capping protein (data not shown).
Expression Patterns of Mouse Twinfilins—The expressionpatterns of twinfilin-1 and twinfilin-2 genes in developingmouse embryos and adult mouse tissues were examined byNorthern blot (Fig. 6) and RNA in situ hybridization analyses(Fig. 7). Northern blot analysis with twinfilin-1 and twinfilin-2cDNA probes showed that twinfilin-1 is the major isoformduring embryonic development and is expressed at a constantlevel during embryonic days (E) 7–17. In contrast, twinfilin-2was only weakly expressed in embryos and its mRNA levelsappeared to increase during development (Fig. 6A). These re-sults were further confirmed by in situ hybridization analysisof mouse embryos. Twinfilin-1 expression was widespreadthroughout the embryonic stages analyzed (E10.5, E12.5,E14.5, and E18.5) (Fig. 7A and data not shown). At E14.5,strongest expression was observed in the developing centraland peripheral nervous system (CNS and PNS, respectively)and in the olfactory sensory epithelium (Fig. 7, C and D). In theCNS, the proliferating neuronal precursors in the ventricularzone expressed twinfilin-1 more than the postmitotic neurons(Fig. 7C). At E18.5, highest expression levels were detected inthe mechanosensory hair cells of the inner ear (Fig. 7G) and inthe differentiated keratinocytes of the skin (Fig. 7I). Twin-filin-2 expression was relatively weak during all of the embry-onic stages (Fig. 7B and data not shown). At E14.5, a slightincrease in the expression could be observed in heart, CNS, andPNS (Fig. 7D and data not shown). At E18.5, twinfilin-2 wasstrongly expressed in the inner ear hair cells and in the headmuscles (Fig. 7H). Expression of both genes could be detected inthe differentiating bone, whereas in cartilage, very littlemRNA, if any, was present (Fig. 7, C, E, G, and H). In contrastto twinfilin-1, no expression of twinfilin-2 was detected in thenasal epithelium or in the skin keratinocytes (Fig. 7, F and J).
Northern blot analysis of adult mouse tissues indicated thattwinfilin-1 is found in most tissues and is strongly expressed inliver and kidney (Fig. 6B). However, no twinfilin-1 expressioncould be detected in skeletal muscles. This expression patternagreed well with our previous data where we examined theexpression of twinfilin-1 mRNA by using a similar Northernblot filter (14). Interestingly, the expression pattern of twin-filin-2 was different from the one of twinfilin-1. Northern blotanalysis showed that twinfilin-2 mRNA is expressed stronglyonly in the heart and is found at much lower levels in other
2 S. Falck, M. A. Wear, J. A. Cooper, and P. Lappalainen, unpublisheddata.
FIG. 4. Mouse twinfilins interact with PI(4,5)P2. A, native gelelectrophoresis assays were performed to detect binding of mouse twin-filins to PI(4,5)P2. In the absence of phospholipids, both twinfilins runas a single sharp band on this gel. PI(4,5)P2 causes twinfilin-1 andtwinfilin-2 to move more rapidly toward the anode, indicating a netincrease in the negative charge and thus a binding interaction. B,polymerization of 3 �M actin (1:6 pyrene actin:human platelet actin) inthe presence of 0, 1.5, or 3 �M twinfilins and 0 or 30 �M PI(4,5)P2 wasinitiated by the addition of 0.1 M KCl, 2 mM MgCl2, and 0.5 mM ATP,and the actin filament polymerization was followed by the increase inpyrene fluorescence. Both twinfilins efficiently inhibit actin filamentassembly, but this activity is down-regulated by 30 �M PI(3,4)P2. Sym-bols are as follows: 3 �M actin (closed circles); 3 �M actin and 30 �M PIP2(closed squares); 3 �M actin and 1.5 �M twinfilin-1 (open circles); 3 �M
actin and 1.5 �M twinfilin-2 (open squares); 3 �M actin and 3 �M
twinfilin-1 (closed triangles); 3 �M actin and 3 �M twinfilin-2 (closeddiamonds); 3 �M actin, 3 �M twinfilin-1, and 30 �M PIP2 (open trian-gles); and 3 �M actin, 3 �M twinfilin-2, and 30 �M PIP2 (open diamonds).
FIG. 5. Interactions of mouse twinfilins with capping protein.This interaction was studied by native gel-electrophoresis assays. 4 �M
capping protein was mixed with 0 (lane 2), 1 (lane 3), 1.5 (lane 4), 2 (lane5), 3 (lane 6), 4 (lane 7), or 6 �M (lane 8) twinfilin-1 (upper panel) ortwinfilin-2 (lower panel). Lane 1 shows the mobility of twinfilin alone.The shift in the mobility of capping protein in the presence of twinfilinindicates a complex formation.
Mammals Have Two Twinfilin Isoforms 34351
by guest on March 27, 2016
http://ww
w.jbc.org/
Dow
nloaded from
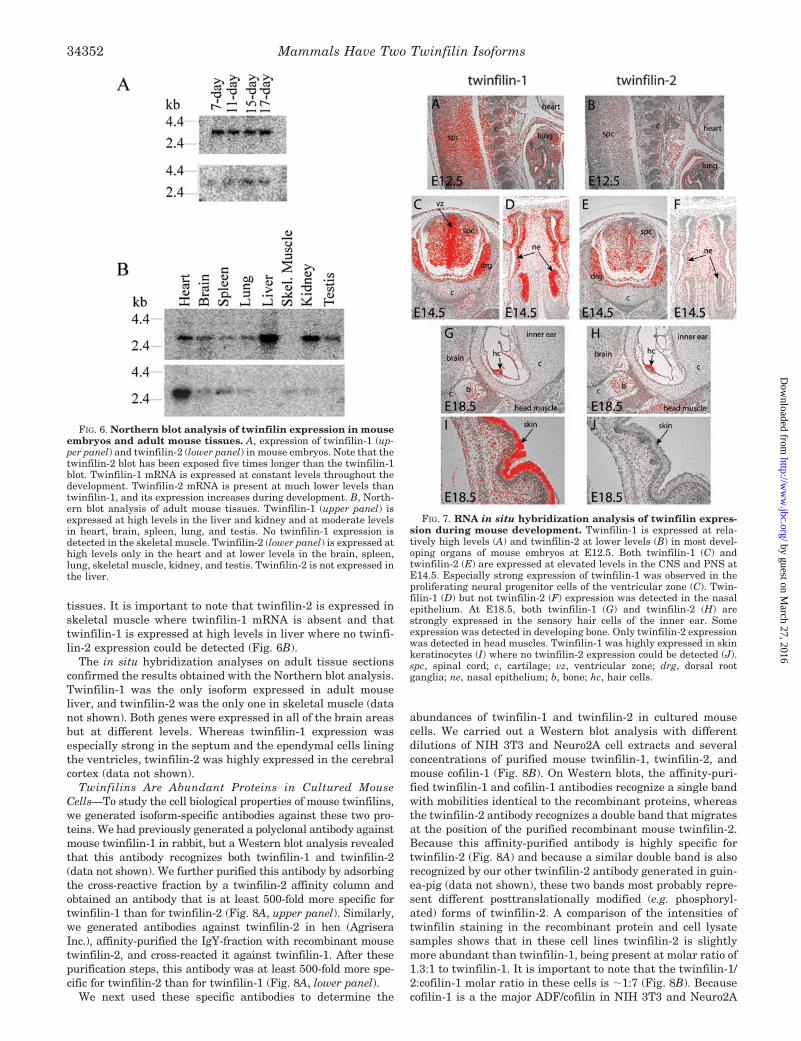
tissues. It is important to note that twinfilin-2 is expressed inskeletal muscle where twinfilin-1 mRNA is absent and thattwinfilin-1 is expressed at high levels in liver where no twinfi-lin-2 expression could be detected (Fig. 6B).
The in situ hybridization analyses on adult tissue sectionsconfirmed the results obtained with the Northern blot analysis.Twinfilin-1 was the only isoform expressed in adult mouseliver, and twinfilin-2 was the only one in skeletal muscle (datanot shown). Both genes were expressed in all of the brain areasbut at different levels. Whereas twinfilin-1 expression wasespecially strong in the septum and the ependymal cells liningthe ventricles, twinfilin-2 was highly expressed in the cerebralcortex (data not shown).
Twinfilins Are Abundant Proteins in Cultured MouseCells—To study the cell biological properties of mouse twinfilins,we generated isoform-specific antibodies against these two pro-teins. We had previously generated a polyclonal antibody againstmouse twinfilin-1 in rabbit, but a Western blot analysis revealedthat this antibody recognizes both twinfilin-1 and twinfilin-2(data not shown). We further purified this antibody by adsorbingthe cross-reactive fraction by a twinfilin-2 affinity column andobtained an antibody that is at least 500-fold more specific fortwinfilin-1 than for twinfilin-2 (Fig. 8A, upper panel). Similarly,we generated antibodies against twinfilin-2 in hen (AgriseraInc.), affinity-purified the IgY-fraction with recombinant mousetwinfilin-2, and cross-reacted it against twinfilin-1. After thesepurification steps, this antibody was at least 500-fold more spe-cific for twinfilin-2 than for twinfilin-1 (Fig. 8A, lower panel).
We next used these specific antibodies to determine the
abundances of twinfilin-1 and twinfilin-2 in cultured mousecells. We carried out a Western blot analysis with differentdilutions of NIH 3T3 and Neuro2A cell extracts and severalconcentrations of purified mouse twinfilin-1, twinfilin-2, andmouse cofilin-1 (Fig. 8B). On Western blots, the affinity-puri-fied twinfilin-1 and cofilin-1 antibodies recognize a single bandwith mobilities identical to the recombinant proteins, whereasthe twinfilin-2 antibody recognizes a double band that migratesat the position of the purified recombinant mouse twinfilin-2.Because this affinity-purified antibody is highly specific fortwinfilin-2 (Fig. 8A) and because a similar double band is alsorecognized by our other twinfilin-2 antibody generated in guin-ea-pig (data not shown), these two bands most probably repre-sent different posttranslationally modified (e.g. phosphoryl-ated) forms of twinfilin-2. A comparison of the intensities oftwinfilin staining in the recombinant protein and cell lysatesamples shows that in these cell lines twinfilin-2 is slightlymore abundant than twinfilin-1, being present at molar ratio of1.3:1 to twinfilin-1. It is important to note that the twinfilin-1/2:cofilin-1 molar ratio in these cells is �1:7 (Fig. 8B). Becausecofilin-1 is a the major ADF/cofilin in NIH 3T3 and Neuro2A
FIG. 6. Northern blot analysis of twinfilin expression in mouseembryos and adult mouse tissues. A, expression of twinfilin-1 (up-per panel) and twinfilin-2 (lower panel) in mouse embryos. Note that thetwinfilin-2 blot has been exposed five times longer than the twinfilin-1blot. Twinfilin-1 mRNA is expressed at constant levels throughout thedevelopment. Twinfilin-2 mRNA is present at much lower levels thantwinfilin-1, and its expression increases during development. B, North-ern blot analysis of adult mouse tissues. Twinfilin-1 (upper panel) isexpressed at high levels in the liver and kidney and at moderate levelsin heart, brain, spleen, lung, and testis. No twinfilin-1 expression isdetected in the skeletal muscle. Twinfilin-2 (lower panel) is expressed athigh levels only in the heart and at lower levels in the brain, spleen,lung, skeletal muscle, kidney, and testis. Twinfilin-2 is not expressed inthe liver.
FIG. 7. RNA in situ hybridization analysis of twinfilin expres-sion during mouse development. Twinfilin-1 is expressed at rela-tively high levels (A) and twinfilin-2 at lower levels (B) in most devel-oping organs of mouse embryos at E12.5. Both twinfilin-1 (C) andtwinfilin-2 (E) are expressed at elevated levels in the CNS and PNS atE14.5. Especially strong expression of twinfilin-1 was observed in theproliferating neural progenitor cells of the ventricular zone (C). Twin-filin-1 (D) but not twinfilin-2 (F) expression was detected in the nasalepithelium. At E18.5, both twinfilin-1 (G) and twinfilin-2 (H) arestrongly expressed in the sensory hair cells of the inner ear. Someexpression was detected in developing bone. Only twinfilin-2 expressionwas detected in head muscles. Twinfilin-1 was highly expressed in skinkeratinocytes (I) where no twinfilin-2 expression could be detected (J).spc, spinal cord; c, cartilage; vz, ventricular zone; drg, dorsal rootganglia; ne, nasal epithelium; b, bone; hc, hair cells.
Mammals Have Two Twinfilin Isoforms34352
by guest on March 27, 2016
http://ww
w.jbc.org/
Dow
nloaded from

cells3 and ADF/cofilins are highly abundant proteins present ina 1:2–1:10 ratio to actin in eukaryote cells (1), these data showthat twinfilins are also abundant proteins in these mammaliancells.
Subcellular Localizations of Twinfilin-1 and Twinfilin-2—Immunofluorescence microscopy studies with the isoform-spe-cific antibodies showed that twinfilin-1 and twinfilin-2 havesimilar subcellular localizations to each other in unstimulatedNeuro 2A and NIH 3T3 cells. In Neuro 2A cells (a mouseneuroblastoma cell line), both twinfilin-1 and twinfilin-2 showstrong perinuclear staining but also localize to the F-actin richfilopodia (Fig. 9, A–C). Also in NIH 3T3 fibroblast cells, bothproteins show punctate cytoplasmic localization that is espe-cially prominent around the nucleus but are also enriched atactin-rich cell processes (Fig. 9, D–I). However, in NIH 3T3cells, twinfilin-1 appears to accumulate more strongly to thesecell processes than twinfilin-2, suggesting that in this cell type,twinfilin isoforms have slight differences in their subcellularlocalizations.
Rho GTPases are members of the Ras superfamily of mono-meric GTP-binding proteins whose main function is to linkmembrane receptors to the assembly and organization of theactin cytoskeleton. The most well characterized members of
this family are RhoA, Cdc42, and Rac1 (38). Previous studieswith an antibody that recognized both twinfilin isoforms sug-gested that the localization of twinfilins in NIH 3T3 cells maybe regulated by the small GTPases Rac1 and Cdc42 (14). There-fore, we examined the effects of these Rho GTPases on thelocalization of twinfilin-1 and twinfilin-2 by using the isoform-specific antibodies. Surprisingly, we observed that Rac1 andCdc42 have distinct effects on these two proteins. Twinfilin-1co-localizes very strongly with the activated Cdc42(V12) at thecell-cell contacts, whereas twinfilin-2 did not concentrate atthese sites in NIH 3T3 cells (Fig. 9, D–E). Furthermore, in cellsexpressing the activated form of Rac1(V12), twinfilin-1 concen-trated strongly to the Rac1-induced membrane ruffles, whereastwinfilin-2 was localized equally throughout the lamellipodium(Fig. 9, G–I). We further confirmed that the twinfilin-1-richsites induced by Cdc42(V12) are cell-cell contacts and not di-viding cells by �-catenin staining (data not shown). In conclu-sion, these results suggest that the subcellular localizations ofthe two mammalian twinfilin isoforms are regulated by differ-ent signaling pathways.
DISCUSSION
Twinfilin is a ubiquitous regulator of actin dynamics found inorganisms from yeast to mammals (18). Here, we identified andcharacterized a second mouse twinfilin isoform, which shows65% sequence identity to previously characterized mouse twin-filin-1. The protein, which we named twinfilin-2, is a mouseorthologue of human “A6-related protein” that was recentlyidentified by Rohwer et al. (33). Interestingly, these authorsreported that the recombinant GST fusion of human A6-relatedprotein did not have any actin binding activity and therefore3 M. K. Vartiainen, P. Hilpela, and P. Lappalainen, unpublished data.
FIG. 8. Both twinfilins are abundant proteins in NIH 3T3 andNeuro2A cells. A, three different concentrations of recombinant mousetwinfilin-1 and twinfilin-2 proteins were run on polyacrylamide gels,and the proteins were visualized by Western blotting using anti-twin-filin-1 and anti-twinfilin-2 antibodies. These antibodies are isoform-specific, because each of them recognizes only one mouse twinfilinisoform. B, mouse cell extracts and known concentrations of recombi-nant mouse twinfilin-1, twinfilin-2, and cofilin-1 were run on polyacryl-amide gels, and the proteins were visualized by Western blotting usingisoform-specific twinfilin-1/2 and cofilin-1 antibodies. The twinfilin-1:twinfilin-2:cofilin-1 molar ratio in these cells is �1:1.3:15. It is impor-tant to note that twinfilin-2 from NIH 3T3 and Neuro 2A cells runs asa double band on this gel, suggesting that a posttranslationally modi-fied form of this protein exists in these cells (see “Results” for furtherdetails).
FIG. 9. Regulation of twinfilin-1 and twinfilin-2 localization incultured mammalian cells. A–C, localization of twinfilin isoforms inNeuro2A cells. Twinfilin-1 (B) and twinfilin-2 (C) visualized with anti-twinfilin-1 and anti-twinfilin-2 antibodies show punctate cytoplasmicstaining but are also localized to the actin filament-rich filopodia visu-alized by rhodamine-phalloidin (A). D–I, effects of Rho family GTPaseson mouse twinfilin localization in NIH 3T3 cells. Cells were transfectedwith the active Cdc42(V12) (D–F) or Rac1(V12) (G–I). Localization ofthe Cdc42(V12) and Rac1(V12) green fluorescent fusion proteins areshown in panels D and G, respectively. Cells were stained with anti-twinfilin-1 antibody (E and H) and anti-twinfilin-2 antibodies (F and I).Twinfilin-1 co-localizes with the active Cdc42 at the cell-cell contacts(arrows in panel E), whereas twinfilin-2 (panel F) is not found at thesesites. The activated form of Rac1 induces the localization of twinfilin-1to the membrane ruffles (arrows in panel H), but it does not affect thelocalization of twinfilin-2 (panel I).
Mammals Have Two Twinfilin Isoforms 34353
by guest on March 27, 2016
http://ww
w.jbc.org/
Dow
nloaded from

suggested that despite high sequence homology to twinfilins,this protein would exhibit a different function. Because ourdirect actin monomer binding, filament sedimentation, actinassembly, and G-actin nucleotide exchange assays clearly dem-onstrate that mouse twinfilin-2 binds actin monomers with ahigh affinity that is similar to those of mouse twinfilin-1 andother known twinfilins, we conclude that twinfilin-2 (A6-re-lated protein) is indeed a functional member of the twinfilinprotein family. We speculate that the lack of actin bindingactivity in the study by Rohwer et al. (33) may have resultedeither from the lack of proper actin monomer-binding assays orfrom the inactivity of their recombinant GST fusion protein. Itis possible that the GST at the N terminus of the recombinantprotein in the study by Rohwer et al. (33) inhibits the actinbinding activity of twinfilin-2 (in our studies the GST wasremoved from the recombinant proteins before biochemicalassays).
Our Northern blot and in situ hybridization analyses sug-gested that either twinfilin-1 or twinfilin-2 is expressed in mostcell types of mouse embryos and adult mice. Therefore, twin-filins appear to be involved in some central cellular processesthat are common to all of the mammalian cell types. Similar tothe mammalian ADF/cofilin and profilin isoforms, the twotwinfilin isoforms show distinct expression patterns. Twin-filin-1 is the major isoform during development and in adultnon-muscle cells, whereas twinfilin-2 is the predominant iso-form of heart and skeletal muscles. Similarly, one of the mam-malian ADF/cofilin isoforms, cofilin-2, is predominant in mus-cle, whereas the two other isoforms, cofilin-1 and ADF, areexpressed strongly in non-muscle cells (19, 39). It is importantto note that both mouse twinfilins are especially abundant inthe mechanosensory hair cells of the inner ear. These cells haveactin-based apical projections called stereocilia that displaystructural and functional similarity to Drosophila mechanosen-sory bristles. In a Drosophila strain that carries a mutation inthe twinfilin gene, the actin bundles forming the bristles areseverely misorganized (15). Therefore, high levels of twinfilinsin mammalian inner ear hair cells may be necessary to main-tain proper stereocilia structure and function.
Although the two mammalian twinfilin isoforms have dis-tinct expression patterns, we could not detect any significantdifferences in their affinities for actin monomers or cappingprotein. Furthermore, our studies did not reveal any specificitytoward certain actin or capping protein isoforms for either ofthe mouse twinfilins (data not shown). Our previous studies(13) demonstrated that yeast twinfilin inhibits the nucleotideexchange on actin monomers and that the activity of yeasttwinfilin can be down-regulated by PI(4,5)P2 in vitro (16). Here,we show that both mouse twinfilins inhibit the nucleotide ex-change on actin monomers and bind PI(4,5)P2 similarly toyeast twinfilin, suggesting that these functions are universalfor all of the members of the twinfilin family. Although wecould not detect any significant biochemical differences be-tween mouse twinfilin-1 and twinfilin-2, it is possible that thetwo mouse proteins have differences in some other aspects ofactin dynamics that can not be measured by currently availablemethods (e.g. the kinetics of actin monomer dissociation fromtwinfilin after interactions with capping protein). Further-more, at least one of these proteins may have other ligandsthan actin, capping protein, and PIP2. For example, it is pos-sible that twinfilin-2 may interact with certain sarcomere pro-teins to promote its specific function in the muscle cells.
Our quantitative Western blotting analysis showed thattwinfilins are abundant proteins, at least in cultured mouseNeuro 2A and NIH 3T3 cells. In these cells, twinfilins are foundin �1:7 ratio to the major ADF/cofilin isoform, cofilin-1. In
yeast cells, the twinfilin:cofilin ratio was reported to be 1:2.5(16). Therefore, the high abundance of twinfilins in these twomammalian cell lines, together with their high affinities forADP-G-actin, suggest that twinfilins may interact with a rela-tively large proportion of actin monomers in cells and thus havea profound effect on the localization and dynamics of the cel-lular actin monomer pool.
Although the two twinfilin isoforms show relatively similarlocalizations in unstimulated cells, only the subcellular local-ization of twinfilin-1 appears to be regulated by Rac1 andCdc42. These two small GTPases are central regulators ofpolarized growth and motility in non-muscle cells (38), suggest-ing that twinfilin-1 may be involved in these cellular functions.The role of twinfilin-1 in polarized growth is further supportedby its strong expression in developing neurons, skin, and olfac-tory sensory epithelium. In contrast, the predominant isoformin muscle cells, twinfilin-2, is not regulated by the smallGTPases Cdc42 and Rac1. Twinfilin-2 may therefore be in-volved in the maintenance of sarcomere structure and perhapsdoes not promote any Rac1 and Cdc42-induced morphologicaland motile processes in muscle cells.
In conclusion, our data shows that mammals have two iso-forms of an actin monomer-binding protein, twinfilin. Twin-filin-1 is the major isoform in non-muscle cells, whereas twin-filin-2 is strongly expressed in heart and skeletal muscle cells.In addition, high levels of both proteins seem to be required inthe inner ear sensory hair cells. Although biochemically simi-lar, the subcellular localizations of twinfilin-1 and twinfilin-2appear to be regulated through different intracellular signalingpathways. In the future, it will be important to elucidate the invivo roles of these two mammalian twinfilin isoforms by geneticmethods and to reveal the molecular mechanisms by which theactivities and localizations of these two abundant actin-bindingprotein isoforms are regulated in mammalian cells.
REFERENCES
1. Pollard, T. D., Blanchoin, L., and Mullins, R. D. (2000) Annu. Rev. Biophys.Biomol. Struct. 29, 545–576
2. Huff, T., Muller, C. S., Otto, A. M., Netzker, R., and Hannappel, E. (2001) Int.J. Biochem. Cell Biol. 33, 205–220
3. Pantaloni, D., and Carlier, M. F. (1993) Cell 75, 1007–10144. Vinson, V. K., De La Cruz, E. M., Higgs, H. N., and Pollard, T. D. (1998)
Biochemistry 37, 10871–108805. Lu, J., and Pollard, T. D. (2001) Mol. Biol. Cell 12, 1161–11756. Wolven, A. K., Belmont, L. D., Mahoney, N. M., Almo, S. C., and Drubin, D. G.
(2000) J. Cell Biol. 150, 895–9037. Maciver, S. K., and Weeds, A. G. (1994) FEBS Lett. 347, 251–2568. Carlier, M.-F., Laurent, V., Santolini, J., Melki, R., Didry, R., Xia, G. X., Hong,
Y., Chua, N. H., and Pantaloni, D. (1997) J. Cell Biol. 136, 1307–13229. Blanchoin, L., and Pollard, T. D. (1999) J. Biol. Chem. 274, 15538–15546
10. Hawkins, M., Pope, B., Maciver, S. K., and Weeds, A. G. (1993) Biochemistry32, 9985–9993
11. Hayden, S. M., Miller, P. S., Brauweiler, A., and Bamburg, J. R. (1993)Biochemistry 32, 9994–10004
12. Lappalainen, P., Kessels, M. M., Cope, M. J. T. V., and Drubin, D. G. (1998)Mol. Biol. Cell 9, 1951–1959
13. Goode, B. L., Drubin, D. G., and Lappalainen, P. (1998) J. Cell Biol. 142,723–733
14. Vartiainen, M., Ojala, P. J., Auvinen, P., Peranen, J., and Lappalainen, P.(2000) Mol. Cell Biol. 20, 1772–1783
15. Wahlstrom, G., Vartiainen, M., Yamamoto, L., Mattila, P. K., Lappalainen, P.,and Heino, T. I. (2001) J. Cell Biol. 155, 787–795
16. Palmgren, S., Ojala, P. J., Wear, M. A., Cooper, J. A., and Lappalainen, P.(2001) J. Cell Biol. 155, 251–260
17. Ojala, P. J., Paavilainen, V. O., Vartiainen, M. K., Tuma, R., Weeds, A. G., andLappalainen, P. (2002) Mol. Biol. Cell 13, 3811–3821
18. Palmgren, S., Vartiainen, M., and Lappalainen, P. (2002) J. Cell Sci. 115,881–886
19. Vartiainen, M. K., Mustonen, T., Mattila, P. K., Ojala, P. J., Thesleff, I.,Partanen, J., and Lappalainen, P. (2002) Mol. Biol. Cell 13, 183–194
20. Yeoh, S., Pope, B., Mannherz, H. G., and Weeds, A. (2002) J. Mol. Biol. 315,911–925
21. Lambrechts, A., Braun, A., Joncheere, V., Aszodi, A., Lanier, L. M., Robbens,J., Van Colen, I., Vandekerckhove, J., Fassler, R., and Ampe, C. (2000) Mol.Cell Biol. 20, 8209–8219
22. Di Nardo, A., Gareus, R., Kwiatkowski, D., and Witke, W. (2000) J. Cell Sci.113, 3795–3803
Mammals Have Two Twinfilin Isoforms34354
by guest on March 27, 2016
http://ww
w.jbc.org/
Dow
nloaded from

23. Gieselmann, R., Kwiatkowski, D. J., Janmey, P. A., and Witke, W. (1995) Eur.J. Biochem. 229, 621–628
24. Lambrechts, A., van Damme, J., Goethals, M., Vandekerckhove, J., and Ampe,C. (1995) Eur. J. Biochem. 230, 281–286
25. Peranen, J., Rikkonen, M., Hyvonen, M., and Kaariainen, L. (1996) Anal.Biochem. 236, 371–373
26. Soeno, Y., Abe, H., Kimura, S., Maruyama, K., and Obinata, T. (1998) J. Mus-cle Res. Cell Motil. 19, 639–646
27. Pardee, J. D., and Spudich, J. A. (1982) Methods Enzymol. 85, 164–18128. Detmers, P., Weber, A., Elzinga, M., and Stephens, R. E. (1981) J. Biol. Chem.
256, 99–10529. Weeds, A. G., Harris, H., Gratzer, W., and Gooch, J. (1986) Eur. J. Biochem.
161, 77–8430. Pollard, T. D. (1986) J. Cell Biol. 103, 2747–275431. Rice, D. B., Aberg, T., Chan, Y., Tang, Z., Kettunen, P. J., Pakarinen, L.,
Maxon, R. E., and Thesleff, I. (2000) Development 127, 1845–185532. Paavilainen, V. O., Merckel, M. C., Falck, S., Ojala, P. J., Pohl, E., Wilmanns,
M., and Lappalainen, P. (2002) J. Biol. Chem. 277, 43089–4309533. Rohwer, A., Kittstein, W., Marks, F., and Gschwendt, M. (1999) Eur. J. Bio-
chem. 263, 518–52534. Yonezawa, N., Nishida, E., Iida, K., Yahara, I., and Sakai, H. (1990) J. Biol.
Chem. 265, 8382–838635. Lassing, I., and Lindberg, U. (1985) Nature 314, 472–47436. Hart, M. C., Korshunova, Y. O., and Cooper, J. A. (1997) Cell Motil. Cytoskel-
eton 38, 120–13237. Schafer, D. A., Korshunova, Y. O., Schroer, T. A., and Cooper J. A. (1994)
J. Cell Biol. 127, 453–46538. Etienne-Manneville, S., and Hall, A. (2002) Nature 420, 629–63539. Ono, S., Minami, N., Abe, H., and Obinata, T. (1994) J. Biol. Chem. 269,
15280–15286
Mammals Have Two Twinfilin Isoforms 34355
by guest on March 27, 2016
http://ww
w.jbc.org/
Dow
nloaded from

LappalainenMaria K. Vartiainen, Elisa M. Sarkkinen, Tanja Matilainen, Marjo Salminen and Pekka
Distributions Are Differentially RegulatedMammals Have Two Twinfilin Isoforms Whose Subcellular Localizations and Tissue
doi: 10.1074/jbc.M303642200 originally published online June 14, 20032003, 278:34347-34355.J. Biol. Chem.
10.1074/jbc.M303642200Access the most updated version of this article at doi:
Alerts:
When a correction for this article is posted•
When this article is cited•
to choose from all of JBC's e-mail alertsClick here
http://www.jbc.org/content/278/36/34347.full.html#ref-list-1
This article cites 38 references, 21 of which can be accessed free at
by guest on March 27, 2016
http://ww
w.jbc.org/
Dow
nloaded from




















