
Making eggs from nectar: the role of life history and dietary carbon turnover in butterfly...
13
Making eggs from nectar: the role of life history and dietary carbon turnover in butterfly reproductive resource allocation Diane M. O’Brien, Carol L. Boggs and Marilyn L. Fogel O’Brien, D. M., Boggs, C. L. and Fogel, M. L. 2004. Making eggs from nectar: therole of life history and dietary carbon turnover in butterfly reproductive resource allocation. / Oikos 105: 279 /291. The diets of many butterflies and moths change dramatically with development: from herbivory in the larvae to nectarivory in the adults. These diets are nutritionally distinct, and thus arelikely to contribute differentially to egg manufacture. We examine the use of dietary resources in egg manufacture by four butterfly species with different patterns of oviposition and lifespan; three in the Nymphalidae (Euphydryas chalcedona, Speyeria mormonia and Heliconius charitonia ), and one in the Pieridae (Colias eurytheme ). Each species was fed two isotopically distinct adult diets based on sucrose, both of which differed from the larval hostplant in 13 C content. Egg isotopic composition was analyzed to quantify the contribution of carbon from the larval and adult diets to egg manufacture. In all four species, egg 13 C content increased to an asymptotic maximum with time, indicating that adult diet is an increasingly important source of egg carbon . The 13 C increase closely resembled that of a nectar-feeding hawkmoth, and was well-described by a model of carbon flow proposed for that species. This similarity suggests that the turnover from larval to adult dietary support of egg manufacture is conserved among nectar-feeding Lepidoptera. Species varied widely in the maximum % egg carbon that derives from the adult diet, from 44% in E. chalcedona to nearly 80% in S. mormonia . These differences were related both to the extent of oocyte provisioning prior to adult emergence, and to egg composition. A species’ lifetime use of larval vs adult resources in egg manufacture reflected both the carbon turnover of the eggs and the timing of oviposition. Thus, the extent to which dietary resources are important in egg manufacture in butterflies depends on development (egg provisioning in teneral adults), behavior (timing of oviposition) and nutritional physiology (nutrient synthesis and turnover). D. M. O’Brien and C. Boggs, Center for Conservation Biology, Dept of Biological Sciences, Stanford Univ., Stanford, CA 94305. Present address for DMO: Dept of Biological Sciences, Wellesley College, 106 Central St., Wellesley, MA 02481, USA ([email protected]). / M. Fogel, Geophysical Laboratory, Carnegie Institution of Washington, 5251 Broad Branch Rd. NW, Washington DC 20015, USA. Assumptions about how organisms use nutritional resources are fundamental to predictions about life history and evolutionary ecology (Reznick 1985, Stearns 1992, Rose and Bradley 1998). Nutrition is a key determinant of reproduction (Wheeler 1996); therefore, reproductive strategies need to be considered in light of nutrient availability (Boggs 1992, de Jong and van Noordwijk 1992). Although energy is commonly used as a currency for reproduction and survival, other nutrients, such as nitrogen (Slansky and Feeny 1977, Brodbeck et al. 1993), phosphorous (Elser et al. 1996, Markow et al. 1999), or essential carbon compounds (Douglas 1998, O’Brien et al. 2002) may pose greater constraints upon reproduction and population growth. The complexities of nutrient requirements, synthesis, storage, conversion and metabolism are poorly under- Accepted 25 September 2003 Copyright # OIKOS 2004 ISSN 0030-1299 OIKOS 105: 279 /291, 2004 OIKOS 105:2 (2004) 279
-
Upload
independent -
Category
Documents
-
view
0 -
download
0
Transcript of Making eggs from nectar: the role of life history and dietary carbon turnover in butterfly...

Making eggs from nectar: the role of life history and dietary carbon
turnover in butterfly reproductive resource allocation
Diane M. O’Brien, Carol L. Boggs and Marilyn L. Fogel
O’Brien, D. M., Boggs, C. L. and Fogel, M. L. 2004. Making eggs from nectar: the roleof life history and dietary carbon turnover in butterfly reproductive resourceallocation. �/ Oikos 105: 279�/291.
The diets of many butterflies and moths change dramatically with development: fromherbivory in the larvae to nectarivory in the adults. These diets are nutritionallydistinct, and thus are likely to contribute differentially to egg manufacture. We examinethe use of dietary resources in egg manufacture by four butterfly species with differentpatterns of oviposition and lifespan; three in the Nymphalidae (Euphydryas chalcedona,Speyeria mormonia and Heliconius charitonia ), and one in the Pieridae (Coliaseurytheme ). Each species was fed two isotopically distinct adult diets based onsucrose, both of which differed from the larval hostplant in 13C content. Egg isotopiccomposition was analyzed to quantify the contribution of carbon from the larval andadult diets to egg manufacture. In all four species, egg 13C content increased to anasymptotic maximum with time, indicating that adult diet is an increasingly importantsource of egg carbon . The 13C increase closely resembled that of a nectar-feedinghawkmoth, and was well-described by a model of carbon flow proposed for thatspecies. This similarity suggests that the turnover from larval to adult dietary supportof egg manufacture is conserved among nectar-feeding Lepidoptera. Species variedwidely in the maximum % egg carbon that derives from the adult diet, from 44% in E.chalcedona to nearly 80% in S. mormonia . These differences were related both to theextent of oocyte provisioning prior to adult emergence, and to egg composition. Aspecies’ lifetime use of larval vs adult resources in egg manufacture reflected both thecarbon turnover of the eggs and the timing of oviposition. Thus, the extent to whichdietary resources are important in egg manufacture in butterflies depends ondevelopment (egg provisioning in teneral adults), behavior (timing of oviposition)and nutritional physiology (nutrient synthesis and turnover).
D. M. O’Brien and C. Boggs, Center for Conservation Biology, Dept of BiologicalSciences, Stanford Univ., Stanford, CA 94305. Present address for DMO: Dept ofBiological Sciences, Wellesley College, 106 Central St., Wellesley, MA 02481, USA([email protected]). �/ M. Fogel, Geophysical Laboratory, Carnegie Institution ofWashington, 5251 Broad Branch Rd. NW, Washington DC 20015, USA.
Assumptions about how organisms use nutritional
resources are fundamental to predictions about life
history and evolutionary ecology (Reznick 1985, Stearns
1992, Rose and Bradley 1998). Nutrition is a key
determinant of reproduction (Wheeler 1996); therefore,
reproductive strategies need to be considered in light of
nutrient availability (Boggs 1992, de Jong and van
Noordwijk 1992). Although energy is commonly used
as a currency for reproduction and survival, other
nutrients, such as nitrogen (Slansky and Feeny 1977,
Brodbeck et al. 1993), phosphorous (Elser et al. 1996,
Markow et al. 1999), or essential carbon compounds
(Douglas 1998, O’Brien et al. 2002) may pose greater
constraints upon reproduction and population growth.
The complexities of nutrient requirements, synthesis,
storage, conversion and metabolism are poorly under-
Accepted 25 September 2003
Copyright # OIKOS 2004ISSN 0030-1299
OIKOS 105: 279�/291, 2004
OIKOS 105:2 (2004) 279

stood, especially for free-living animals with mixed or
complex diets. There is a growing appreciation for the
physiological complexity of resource allocation (Rose
and Bradley 1998, Rivero and Casas 1999, Zera and
Harshman 2001, Zhao and Zera 2002), in particular, for
hormonal controls of nutrient use (Sinervo and Svensson
1998, Zera et al. 1998). A similar focus on nutritional
physiology will further help to strengthen the connec-
tions between diet and life history.
Resource allocation in holometabolous insects is
particularly interesting because diets and energetic needs
change between life stages, causing nutrient intake and
nutrient demand to be temporally separated (Boggs
1981, 1986, Rivero et al. 2001). In these insects,
mechanisms for appropriately timed nutrient storage
and release are extremely important (Delobel et al. 1993,
Wheeler and Buck 1995, Snigirevskaya et al. 1997,
Djawdan et al. 1998, Wheeler et al. 2000, Pan and Telfer
2001). In most butterflies and many moths, diets change
dramatically from herbivory in the larvae to nectarivory
in the adults. These diets are nutritionally distinct, and
thus are likely to contribute differently to physiological
demands. Nectar is rich in sugars but relatively depau-
perate in amino acids, fatty acids, and other nutritionally
important compounds (Baker and Baker 1983). The
larval diet is the primary supply of protein, which is
stored for use during metamorphosis and oogenesis
(Wheeler et al. 2000, Pan and Telfer 2001). Butterfly
researchers have long appreciated that both larval and
adult diets are important to reproduction and are both
used in egg manufacture (Norris 1934, Dunlap-Pianka et
al. 1977, Hill 1989, Hill and Pierce 1989, Alm et al. 1990,
Boggs and Ross 1993, Boggs 1997a). Because their life
histories vary, the Lepidoptera have also provided a
model system for comparing reproductive output among
species with varying degrees and types of adult foraging
(Boggs 1986, 1990, 1992, 1997b, 2003).
We use natural variation in 13C to identify egg carbon
deriving from the larval and adult diets in butterflies
with nectivorous adults, but differing lifespans and age-
specific fecundities. 13C content differs between plants
with C3 and C4 photosynthesis, but also can vary among
plants sharing a photosynthetic mode (O’Leary 1981,
1988). Thus, it can be a powerful tracer of carbon flow
from multiple plant sources. Natural abundances of light
stable isotopes are a well-established tracer of nutrient
flow, within both organisms (Metges et al. 1990, Hobson
et al. 1997, Schmidt et al. 1999) and ecosystems
(Rounick and Winterbourn 1986, Peterson and Fry
1987, Ostrom et al. 1997, Wolf and Martınez del Rio
2000). In a previous study, O’Brien and colleagues (2000)
used carbon isotopes to trace the use of larval and adult
diets in reproduction by a hawkmoth. Females were fed
one of two sucrose-based adult diets made from beet
(C3) or cane (C4) sugar, both of which differed from the
natural larval hostplant in 13C content. This approach
enabled them to trace carbon use in eggs with high
accuracy, and they proposed a descriptive carbon turn-
over model to describe how egg nutrients were incorpo-
rated into eggs (O’Brien et al. 2000).
In this study, we apply a similar approach to
investigate the dietary sources of reproductive nutrients
in four nectar-feeding butterfly species, three from the
family Nymphalidae and one from the family Pieridae.
The species selected exhibit a range of variation in
lifespan and age-specific fecundity, and differ in degree
of shared phylogenetic history. We investigate whether
the turnover pattern modeled in hawkmoth eggs is
exhibited in other nectar-feeding Lepidoptera. If so, we
evaluate whether the extent of adult diet use or the
rapidity of larval to adult diet turnover vary according to
differences in life history. We predict that the reliance on
adult dietary carbon in egg manufacture will be higher in
those species whose reproductive effort takes place later
in the adult lifespan.
This study uses sucrose solution as an experimental
proxy for the adult nectar diet, as it elicts a strong
feeding response from butterflies (Romeis and Wackers
2000) and is easy to acquire in different isotopic forms.
This proxy is a nutritional oversimplication, especially as
some butterflies have been shown to detect and slightly
prefer nectars containing amino acids (Alm et al. 1990,
Rusterholz and Erhardt 1998, Mevi-Schutz and Erhardt
2002). We do not suggest that other dilute components
of nectar could not play some nutritional role in
oviposition. This study might slightly underestimate
the importance of the adult diet in species that capitalize
on nectar amino acids. However, as the vast majority of
carbon provided by the adult diet is in the form of sugar
(Baker and Baker 1983), sugar is a reasonable proxy
given our carbon isotope mass-balance approach.
Methods
Study organismsEuphydryas chalcedona
Euphydryas chalcedona (Nymphalidae: Nymphalinae;
Melitaeini) is a univoltine butterfly distributed through-
out western North America. Individuals were field
collected as 5th and 6th instar larvae from Scrophularia
californicus and Mimulus aurantiacus at the Silver Creek
Hills Butterfly Reserve near San Jose, California, USA.
Larvae were placed in mesh bags on potted S. californi-
cus for the remainder of their larval development, and
bags were moved as needed to ensure a constant supply
of fresh leaves. Pupae were stored at 208C until adults
emerged. Average female forewing length was 29.89/0.4
mm. Females mated either the day of emergence or the
day following. Once mated, females were placed in wire
mesh cages (20�/20�/30 cm) and provided fresh S.
californicus leaves for oviposition. Females oviposited
280 OIKOS 105:2 (2004)

daily or every other day in clusters of typically 100�/200
eggs (all other species laid eggs singly). Females were
handfed twice daily. Of the 10 females to successfully
mate and start the experiment, 2 were removed because
they either waited unusually long to oviposit, or didn’t
eat consistently. The remaining 8 butterflies were used in
both isotope analyses and feeding and fecundity mea-
surements.
Speyeria mormonia
Speyeria mormonia (Nymphalidae: Heliconiinae: Heli-
coniini: Argynniti) is a temperate zone univoltine
butterfly that occurs in montane and subalpine habitat
in western North America. Adult females were captured
during August 1999 near Gothic, Gunnison County,
Colorado, USA, and oviposited in captivity. Speyeria
mormonia overwinters as unfed first instar larvae;
therefore, their hatchlings were placed in humid vials
and stored at 48C. The following February (2000), larvae
were warmed to room temperature and placed in mesh
bags tied over leaves of potted violets (Viola soraria ).
Bags were moved as needed to provide larvae with a
constant supply of fresh leaves. Pupae were stored at
208C until adults emerged. Average female forewing
length was 26.99/0.6 mm (nearly identical to that of
field-caught females, 26.69/1.1 mm, Boggs 1987). Newly
emerged females were hand-mated and placed in cylind-
rical cages made from lantern globes and lined with wax
paper (15 cm diameter�/20 cm height). Females were
provided fresh violet leaves for oviposition, and were fed
twice daily. Of 15 healthy experimental females, two were
removed mid-way through because excessive wing da-
mage made them difficult to hold for feeding. Thus, N�/
13 females for fecundity and lifespan estimates, 5 of
which were hand-fed quantitatively.
Heliconius charitonia
H. charitonia (Nymphalidae: Heliconiinae: Heliconini:
Heliconiiti) is a sub-tropical butterfly, which cannot
diapause. Adults feed on both nectar and pollen. A free-
flying greenhouse colony of H. charitonia is maintained
at Stanford University, founded from individuals col-
lected in Costa Rica and Yucatan, Mexico. Adults in the
colony oviposit on potted Passiflora caerula and feed
from potted Lantana camara and Psiguria umbrosa .
Honey water feeders supplement these natural sources of
nectar and pollen. Experimental females were collected
as pupae from the colony and mated upon emergence.
Average female forewing length was 38.69/0.6 mm.
H. charitonia requires more space than the other
butterflies to fly and oviposit normally; therefore, mated
females were placed individually into 1.8�/1.0�/1.0
meter cages and each was given a potted Passiflora
caerula plant for oviposition. Females were provided
experimental sugar solutions ad lib in feeders colored
to resemble Psiguria umbrosa flowers, and sugar solu-
tions were changed daily. Although H. charitonia
normally supplement their diet with pollen, females in
this experiment were not provided with pollen. Eggs
were collected daily. Nine females initiated oviposition
and were used in isotope measurements; of those, two
died prematurely (N�/7 for lifespan and fecundity
estimation).
Colias eurytheme
Colias eurytheme (Pieridae) is a temperate zone butterfly,
which does not diapause. Colias eurytheme eggs were
provided by Ward Watt from his colony at Stanford
University, which derived from females collected near
Tracy, California, USA. Larvae were reared on hydro-
ponically grown Vicia villosa (Watt, unpubl.), and pupae
were housed at 208C until emergence. Newly emerged
adult females were hand-mated the same day. Average
female forewing length was 28.79/0.05 mm. Mated
females were housed in 20�/20�/30 cm mesh cages
and provided potted vetch plants for oviposition.
Females were hand-fed twice daily. Of 18 females that
started the experiment, 4 laid fewer than 500 eggs, lived
fewer than 10 days, or skipped multiple days of
oviposition in the first week. These butterflies were
considered non-representative and were removed. Of 14
healthy ovipositing females used for isotope analyses,
two were prematurely removed due to injury or escape
and were not included in our estimates of lifespan and
fecundity (N�/12).
Experimental methods
Experiments were carried out between February and
May, 2000 (except where noted), at the Herrin Green-
houses at Stanford University. Greenhouses were main-
tained on a 16:8 L:D photoperiod, with daytime and
nighttime temperatures of 278C and 15.58C, respectively.
All butterflies were fed 30% sucrose solution (wt/wt) as
adults. Approximately half of the individuals received
beet sugar solution (C3) and half received cane sugar
solution (C4). A single batch was mixed for each sugar
solution, and stored as frozen aliquots until fed to the
butterflies. A subset of the experimental females were fed
quantitatively from 25 or 50 ml Hamilton graduated
syringes, as described elsewhere (Boggs and Ross 1993),
to evaluate their intake quantitatively under the experi-
mental conditions. Eggs were collected daily into labeled
0.5 ml Eppendorf tubes, which were placed open into a
drying oven at 508C. After 3 days, tubes were removed
from the oven, sealed, and stored at room temperature
until isotope analysis.
OIKOS 105:2 (2004) 281

d13C analysis
Isotope ratios are given a d13C�/(Rsample/Rstandard
�/1)�/1000, in which R�/13C/12C and the standard is
PDB limestone. Dried egg samples weighing 0.30 to 0.35
mg were analyzed for d13C using a CHNS analyzer (CE
Instruments, model NC2500) interfaced to a Finnigan
Delta Plus XL isotope ratio mass spectrometer via the
Finnigan Conflo II interface. The CHNS analyzer also
measured %C and %N for each sample. Acetanilide
standards (0.2�/0.25 mg.) were run with each batch of
samples (�/1 standard per 5 samples), and were used to
correct for consistent variation in d13C among batches
run at different times. The standard deviation of
acetanilide d13C within a batch was typically B/0.15�.
Turnover model
We parameterize the pattern of dietary carbon allocation
with the model of O’Brien et al. (O’Brien et al. 2000).
Briefly, the model assumes two classes of egg nutrients:
one that shifts from larval to adult sources as the
butterfly feeds, and one that retains an unchanging
larval isotopic signature. We label these the ‘‘mixing’ and
‘non-mixing’’ carbon sources, respectively. The propor-
tion of egg carbon in these two nutrient classes is given
by pmax, which represents the maximum contribution of
adult dietary carbon to eggs:
egg d13C�(d13Cmixing)(pmax)�(d13Cnon-mixing)(1�pmax)
(1)
Because d13Cnon-mixing will be constant, whereas d13
Cmixing will change over time, the isotopic composition
of the two nutrient pools is modeled as:
d13Cnon-mixing�d13Clarval C (2)
d13Cmixing�d13Clarval C�((d13Cdiet�fa)�d13Clarval C)
� ( 1�e�rt) (3)
Thus, egg
d13C�Cd13Clarval C�((d13Cdiet�fa)�d13Clarval C)
� ( 1�e�rt)) (pmax) �Cd13Clarval C)(1�pmax) (4)
where
pmax�/the maximum % of egg carbon to derive from the
adult diet
r�/the fractional turnover rate of carbon from larval to
adult sources
fa�/the isotopic fractionation associated with incorpor-
ating adult dietary carbon into eggs
d13Clarval C�/the isotopic composition of egg carbon
deriving from the larval diet.
d13Cdiet�/the isotopic composition of the adult diet
(known)
t�/days since adult emergence (known)
By applying this model to our data, we can estimate
the values of the parameters in bold. In particular, we
are interested in pmax, which describes the maximum
contribution of adult dietary carbon to eggs, and r,
which indicates the rate at which butterflies transition
from larval to adult sources of egg carbon.
Calculating % contribution of the adult diet
The magnitude of the difference in egg d13C between C3
vs C4 �/ fed females on a given day reflects how much of
the carbon in those eggs comes from the adult diet. We
term the proportional contribution of the adult diet to
eggs ‘‘p’’, and quantify it as follows (O’Brien et al. 2002):
p�(d13CC4 egg�d13CC3 egg)=(d13CC4 diet�d13CC3 diet)
(5)
We use the lines of best fit through the data from C3
and C4 fed females to calculate an average daily ‘‘p’’ for
each species.
Amino acid composition
Egg amino acid compositions were analyzed by post-
column ninhydrin-based amino acid analyses at the
Molecular Structure Facility at the University of Cali-
fornia, Davis (Smith 1997). This technique provides data
on 16 of the 20 amino acids found in proteins: cysteine
and tryptophan are partially or completely destroyed by
the analysis, and glutamine and asparagine become
indistinguishable from glutamic acid and aspartic acid,
respectively, after acid hydrolysis (Smith 1997).
Oocyte provisioning at adult emergence
Asking whether adult-derived nutrients are used in egg
manufacture implies that at least some oocytes are not
mature at the time of adult emergence. In order to
evaluate the extent of oocyte provisioning during
metamorphosis, we dissected the ovaries of newly
emerged females and calculated the volume of the
most mature and least mature oocytes assuming a
cylindrical egg. We used the same method to measure
the volume of newly laid eggs for all species, except
Euphydyras chalcedona , which has chorionated eggs in
the ovaries at emergence, which therefore cannot change
in volume. Oocyte volume as a % of the volume of newly
laid eggs was then calculated for both the most and least
mature eggs in the ovaries. Because females emerge with
a full complement of oocytes, these measurements
represent the first and last eggs potentially laid, and
hence the endpoints of eggs used in our isotopic analysis.
Note that Heliconius charitonia exhibits oogenesis as an
adult only when fed pollen (Dunlap-Pianka et al. 1977);
282 OIKOS 105:2 (2004)

hence its ovarian dynamics here resemble the other
species in that new oocytes are not produced.
Statistical analyses
All statistical analyses were performed in JMP version
5.0 (SAS Institute, Inc.) Means are given9/SD unless
otherwise noted. Species differences in lifespan, fecund-
ity, egg weight, and egg C/N were evaluated with
Analysis of Variance (ANOVA). Variation among spe-
cies in meal size over time was tested with Analysis of
Covariance (ANCOVA). Within each species, changes in
egg mass and C/N over time were evaluated using
ANCOVA to control for individual differences. With
each application of ANOVA or ANCOVA, means were
compared using Tukey’s HSD contrasts and residuals
were tested for normality using the Shapiro-Wilks test.
Non-linear fitting was iteratively performed using least
squares minimization until estimates converged. Briefly,
non-linear fitting applies a model with unknown para-
meters to a set of known data and modifies the
parameters from pre-set values to achieve the best fit
(via least-squares minimization). We tested whether
nutrient allocation differed between species pairs by
evaluating whether the data were best described by a
single carbon model or separate models, as proposed by
Ratkowsky and outlined in Motulsky and Ransas
(Motulsky and Ransnas 1987). In these comparisons,
we used p�/0.008 (Bonferroni correction) as the sig-
nificance threshold to account for multiple tests.
Results
Lifespan and fecundity
Average adult lifespan in this study ranged from 15 days
for Euphydryas chalcedona to 41 days for Heliconius
charitonia , and average lifetime fecundity ranged from
138 eggs in H. charitonia to 1065 eggs in E. chalcedona
(Fig. 1, Table 1, species were different at PB/0.0001).
Age-specific fecundity (mean9/SE) is presented for each
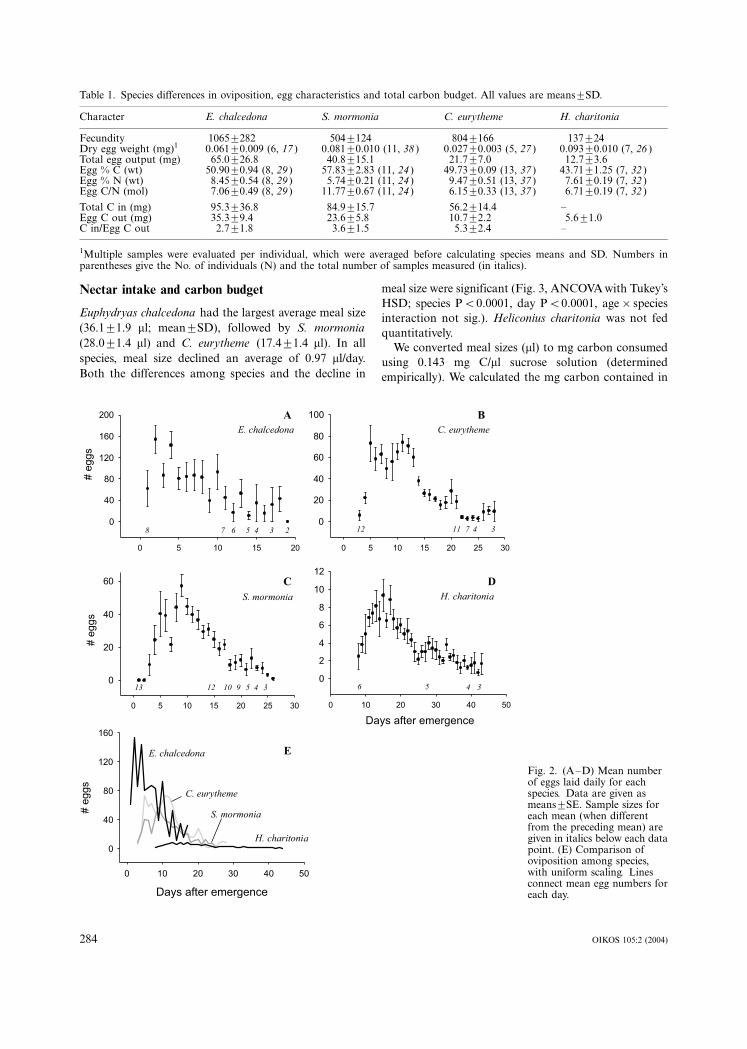
species in Fig. 2. E. chalcedona began to deposit eggs in
large clusters immediately after mating, typically laying
�/500 eggs within the first 5 days of adult life. In
contrast, H. charitonia began to oviposit more than a
week after emergence and mating, laid fewer eggs daily,
and lived for weeks longer than the other three butterfly
species. H. charitonia is a species that normally supple-
ments its nectar diet with pollen, (Dunlap-Pianka et al.
1977, Boggs 1979), but still lays relatively few eggs per
day (�/10�/15, Dunlap-Pianka et al. 1977, Boggs 1997a).
Speyeria mormonia fell between H. charitonia and E.
chalcedona in the timing of its reproductive effort, its
lifespan, and its fecundity. Colias eurytheme, from the
family Pieridae, was also intermediate; it achieved
greater fecundity than S. mormonia, but did so with
smaller eggs (see below). The data are consistent with
previously published life history studies on the three of
these species for which records are available (Stern and
Smith 1960, Dunlap-Pianka et al. 1977, Boggs 1986,
1990).
Egg weight and composition
Egg weight and % carbon and nitrogen are presented for
each species in Table 1. Egg weight declined slightly
(between 1�/5 mg dry weight/day) but significantly with
age in C. eurytheme, E. chalcedona and S. mormonia
(ANCOVA, Table 2), consistently with many other
butterfly species (Boggs 1986, Karlsson 1987, Wickman
and Karlsson 1987). Speyeria mormonia eggs had a
markedly lower protein content compared to the other
species, as indicated by their high egg C/N (Table 1). Egg
C/N increased slightly over time in H. charitonia
(ANCOVA, Day effect P�/0.0152, slope�/0.014/day),
but did so in no other species.
Oocyte provisioning at adult emergence
Euphydryas chalcedona , with an average of 489/6 (n�/3
newly emerged females) oocytes mature and chorionated
at emergence, had its most mature egg equal to 100% of
the volume of a laid egg. The other species showed lower
percentages: 7% (n�/2 newly emerged females) for S.
mormonia and 4% for both C. eurytheme (n�/2) and H.
charitonia (n�/1). The volume of least mature oocytes
was invariant within species and similar among species,
at 0.0008 mm3 for S. mormonia and C. eurytheme and
0.0005 mm3 for E. chalcedona and H. charitonia . These
volumes are 0.1% of that of a laid egg for E. chalcedona
and H. charitonia , 0.2% for S. mormonia and 0.4% for
C. eurytheme.
Fig. 1. Species differences in lifespan and fecundity for thisexperiment. Shown are means9/SD, sample sizes are given inparentheses next to the species names.
OIKOS 105:2 (2004) 283
Nectar intake and carbon budget
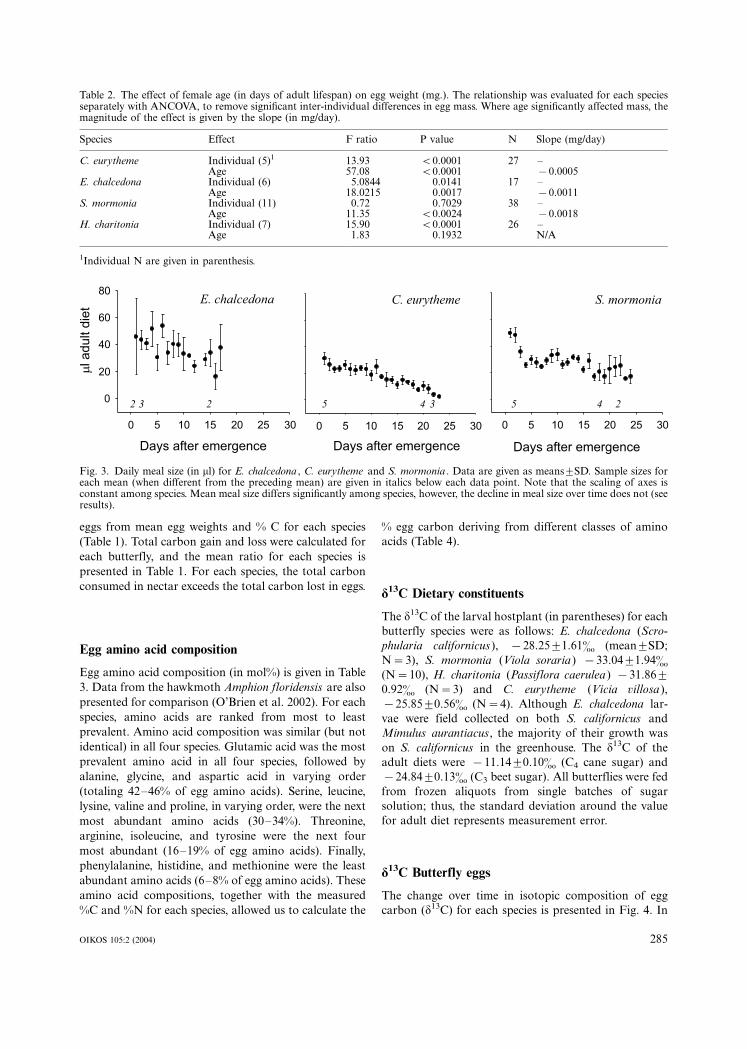
Euphydryas chalcedona had the largest average meal size
(36.19/1.9 ml; mean9/SD), followed by S. mormonia
(28.09/1.4 ml) and C. eurytheme (17.49/1.4 ml). In all
species, meal size declined an average of 0.97 ml/day.
Both the differences among species and the decline in
meal size were significant (Fig. 3, ANCOVA with Tukey’s
HSD; species PB/0.0001, day PB/0.0001, age�/species
interaction not sig.). Heliconius charitonia was not fed
quantitatively.
We converted meal sizes (ml) to mg carbon consumed
using 0.143 mg C/ml sucrose solution (determined
empirically). We calculated the mg carbon contained in
Table 1. Species differences in oviposition, egg characteristics and total carbon budget. All values are means9/SD.
Character E. chalcedona S. mormonia C. eurytheme H. charitonia
Fecundity 10659/282 5049/124 8049/166 1379/24Dry egg weight (mg)1 0.0619/0.009 (6, 17 ) 0.0819/0.010 (11, 38 ) 0.0279/0.003 (5, 27 ) 0.0939/0.010 (7, 26 )Total egg output (mg) 65.09/26.8 40.89/15.1 21.79/7.0 12.79/3.6Egg % C (wt) 50.909/0.94 (8, 29 ) 57.839/2.83 (11, 24 ) 49.739/0.09 (13, 37 ) 43.719/1.25 (7, 32 )Egg % N (wt) 8.459/0.54 (8, 29 ) 5.749/0.21 (11, 24 ) 9.479/0.51 (13, 37 ) 7.619/0.19 (7, 32 )Egg C/N (mol) 7.069/0.49 (8, 29 ) 11.779/0.67 (11, 24 ) 6.159/0.33 (13, 37 ) 6.719/0.19 (7, 32 )
Total C in (mg) 95.39/36.8 84.99/15.7 56.29/14.4 �/
Egg C out (mg) 35.39/9.4 23.69/5.8 10.79/2.2 5.69/1.0C in/Egg C out 2.79/1.8 3.69/1.5 5.39/2.4 �/
1Multiple samples were evaluated per individual, which were averaged before calculating species means and SD. Numbers inparentheses give the No. of individuals (N) and the total number of samples measured (in italics).
Fig. 2. (A�/D) Mean numberof eggs laid daily for eachspecies. Data are given asmeans9/SE. Sample sizes foreach mean (when differentfrom the preceding mean) aregiven in italics below each datapoint. (E) Comparison ofoviposition among species,with uniform scaling. Linesconnect mean egg numbers foreach day.
284 OIKOS 105:2 (2004)
eggs from mean egg weights and % C for each species
(Table 1). Total carbon gain and loss were calculated for
each butterfly, and the mean ratio for each species is
presented in Table 1. For each species, the total carbon
consumed in nectar exceeds the total carbon lost in eggs.
Egg amino acid composition
Egg amino acid composition (in mol%) is given in Table
3. Data from the hawkmoth Amphion floridensis are also
presented for comparison (O’Brien et al. 2002). For each
species, amino acids are ranked from most to least
prevalent. Amino acid composition was similar (but not
identical) in all four species. Glutamic acid was the most
prevalent amino acid in all four species, followed by
alanine, glycine, and aspartic acid in varying order
(totaling 42�/46% of egg amino acids). Serine, leucine,
lysine, valine and proline, in varying order, were the next
most abundant amino acids (30�/34%). Threonine,
arginine, isoleucine, and tyrosine were the next four
most abundant (16�/19% of egg amino acids). Finally,
phenylalanine, histidine, and methionine were the least
abundant amino acids (6�/8% of egg amino acids). These
amino acid compositions, together with the measured
%C and %N for each species, allowed us to calculate the
% egg carbon deriving from different classes of amino
acids (Table 4).
d13C Dietary constituents
The d13C of the larval hostplant (in parentheses) for each
butterfly species were as follows: E. chalcedona (Scro-
phularia californicus ), �/28.259/1.61� (mean9/SD;
N�/3), S. mormonia (Viola soraria ) �/33.049/1.94�(N�/10), H. charitonia (Passiflora caerulea ) �/31.869/
0.92� (N�/3) and C. eurytheme (Vicia villosa ),
�/25.859/0.56� (N�/4). Although E. chalcedona lar-
vae were field collected on both S. californicus and
Mimulus aurantiacus, the majority of their growth was
on S. californicus in the greenhouse. The d13C of the
adult diets were �/11.149/0.10� (C4 cane sugar) and
�/24.849/0.13� (C3 beet sugar). All butterflies were fed
from frozen aliquots from single batches of sugar
solution; thus, the standard deviation around the value
for adult diet represents measurement error.
d13C Butterfly eggs
The change over time in isotopic composition of egg
carbon (d13C) for each species is presented in Fig. 4. In
Table 2. The effect of female age (in days of adult lifespan) on egg weight (mg.). The relationship was evaluated for each speciesseparately with ANCOVA, to remove significant inter-individual differences in egg mass. Where age significantly affected mass, themagnitude of the effect is given by the slope (in mg/day).
Species Effect F ratio P value N Slope (mg/day)
C. eurytheme Individual (5)1 13.93 B/0.0001 27 �/
Age 57.08 B/0.0001 �/0.0005E. chalcedona Individual (6) 5.0844 0.0141 17 �/
Age 18.0215 0.0017 �/0.0011S. mormonia Individual (11) 0.72 0.7029 38 �/
Age 11.35 B/0.0024 �/0.0018H. charitonia Individual (7) 15.90 B/0.0001 26 �/
Age 1.83 0.1932 N/A
1Individual N are given in parenthesis.
Fig. 3. Daily meal size (in ml) for E. chalcedona , C. eurytheme and S. mormonia . Data are given as means9/SD. Sample sizes foreach mean (when different from the preceding mean) are given in italics below each data point. Note that the scaling of axes isconstant among species. Mean meal size differs significantly among species, however, the decline in meal size over time does not (seeresults).
OIKOS 105:2 (2004) 285

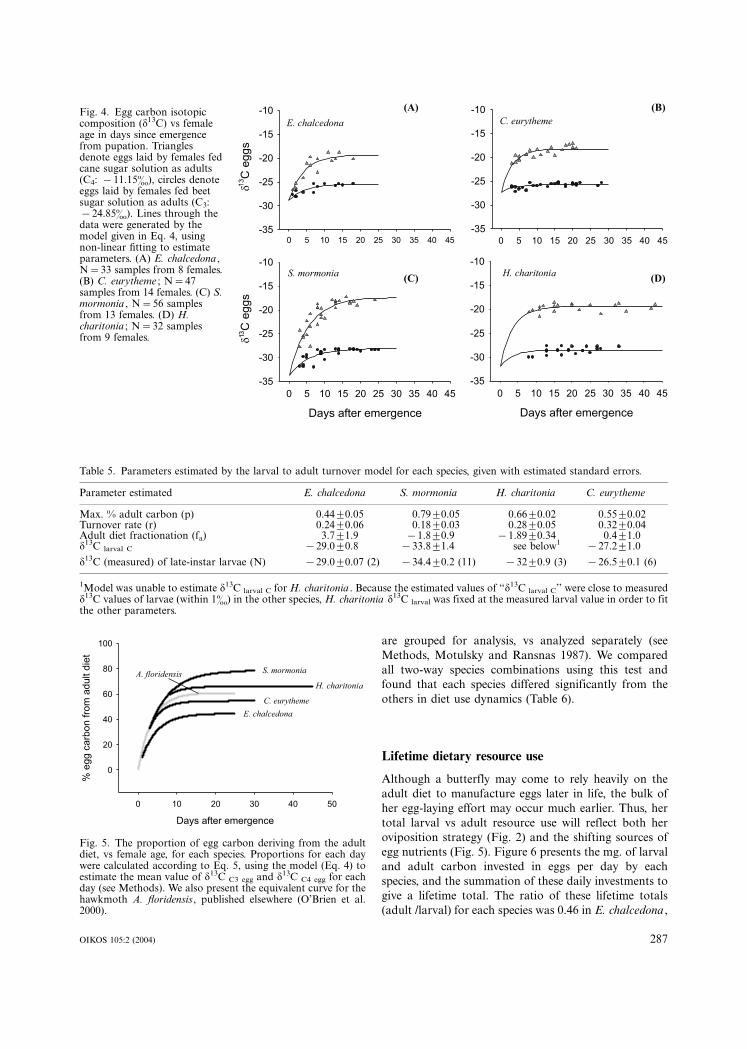
all four species, egg d13C reflected the d13C of the C3 or
C4 adult diet (triangles vs circles, Fig. 4). Egg d13C
increased asymptotically over time (Fig. 4), as found
elsewhere (O’Brien et al. 2000). The extent of the
increase depended on the species, and on the lag between
emergence and oviposition. For example, larval to adult
carbon turnover in H. charitonia stabilized prior to the
onset of oviposition on day 8, and thus egg d13C changed
little if at all (Fig. 4). In E. chalcedona, S. mormonia and
C. eurytheme, eggs laid within several days of emergence
were predominantly larval in carbon composition, and
rapidly shifted to a more adult-derived isotopic signal.
The data were well described by the turnover model
outlined in the Methods (lines of best fit in Fig. 4,
parameters given in Table 5).
Use of adult diet in egg provisioning
The daily proportion of egg carbon deriving from the
adult diet is summarized in Fig. 5 for each species.
Species varied widely in the maximum percentage of egg
carbon that derives from the adult diet (from 44% in E.
chalcedona to nearly 80% in S. mormonia , Fig. 5, Table
5). In contrast, species were all relatively similar in their
turnover rate (r), the inverse of the time required to
reach half turnover (Fig. 5). The turnover rate indicates
how rapidly the shift from larval to adult sources occurs.
These varied from 0.18 day�1 in S. mormonia (most
rapid turnover) to 0.32 day�1 in C. eurytheme (least
rapid, Table 5).
Comparing allocation patterns among species
In order to evaluate whether species differed significantly
in their patterns of adult and larval resource use, we
tested whether the parameters fit to each species’ data by
the turnover model could also adequately describe that
of the other species. In order to do this we used a test
that compares whether the goodness of fit of a non-
linear model significantly worsens when two sets of data
Table 3. Amino acid composition of butterfly eggs. Each column gives averaged data from 2 individuals, with inter-individualdifferences less than 0.1% on average. Data are ranked from highest to lowest prevalence for each species. Glx�/glutamic acid�/
glutamine, Asx�/aspartic acid�/asparagine. Horizontal lines separate groups of amino acids that share similar prevalence (but invarying order).
E. chalcedona S. mormonia H. charitonia C. eurytheme A. floridensis1
aa mol% aa mol% aa mol% aa mol% aa mol%
glx 13.0 glx 12.5 glx 13.0 glx 12.5 gly 15.6asx 10.5 gly 11.4 gly 11.2 asx 10.1 ala 11.6ala 9.7 asx 9.4 asx 11.0 gly 9.8 glx 11.1gly 8.7 ala 8.7 ala 10.3 ala 9.6 asx 8.2
leu 7.2 pro 7.6 ser 7.8 ser 7.6 leu 7.0ser 7.1 val 7.2 leu 6.6 leu 7.1 val 6.0val 6.1 leu 6.9 lys 5.7 lys 6.1 pro 5.9lys 6.1 ser 6.3 pro 5.5 val 5.8 ser 5.7pro 5.7 lys 5.4 val 5.0 pro 5.8 tyr 5.0
thr 4.7 ileu 4.9 tyr 4.7 thr 4.7 lys 4.4arg 4.7 tyr 4.5 thr 4.3 arg 4.6 thr 4.3ileu 4.6 thr 4.3 arg 4.1 ileu 4.4 arg 3.8tyr 4.3 arg 3.7 ileu 3.5 tyr 4.3 ileu 3.8
phe 3.3 phe 3.1 phe 2.9 phe 3.3 phe 2.8his 2.3 his 2.0 his 2.0 his 2.3 his 2.4met 1.9 met 1.6 met 2.0 met 1.8 met 1.7
1Data originally published in (O’Brien et al. 2002).
Table 4. Calculations of egg protein and EAA content, based on egg amino acid composition, %C and %N.
E. chalcedona S. mormonia H. charitonia C. eurytheme
g N g�1 egg1 0.085 0.057 0.076 0.095g protein g�1 N2 5.713 5.817 5.716 5.697g protein g�1 egg3 0.486 0.331 0.434 0.541
g C g�1 egg1 0.51 0.58 0.44 0.497g EAA C g�1 protein2 0.292 0.301 0.283 0.302g EAA C g�1 C4 0.278 0.172 0.279 0.329
1Data are from the Elemental Analyzer (Table 1).2Calculated from the amino acid composition data (Table 3).3Calculated from the above two lines.4Calculated from the above three lines.
286 OIKOS 105:2 (2004)
are grouped for analysis, vs analyzed separately (see
Methods, Motulsky and Ransnas 1987). We compared
all two-way species combinations using this test and
found that each species differed significantly from the
others in diet use dynamics (Table 6).
Lifetime dietary resource use
Although a butterfly may come to rely heavily on the
adult diet to manufacture eggs later in life, the bulk of
her egg-laying effort may occur much earlier. Thus, her
total larval vs adult resource use will reflect both her
oviposition strategy (Fig. 2) and the shifting sources of
egg nutrients (Fig. 5). Figure 6 presents the mg. of larval
and adult carbon invested in eggs per day by each
species, and the summation of these daily investments to
give a lifetime total. The ratio of these lifetime totals
(adult /larval) for each species was 0.46 in E. chalcedona ,
Fig. 4. Egg carbon isotopiccomposition (d13C) vs femaleage in days since emergencefrom pupation. Trianglesdenote eggs laid by females fedcane sugar solution as adults(C4: �/11.15�), circles denoteeggs laid by females fed beetsugar solution as adults (C3:�/24.85�). Lines through thedata were generated by themodel given in Eq. 4, usingnon-linear fitting to estimateparameters. (A) E. chalcedona ,N�/33 samples from 8 females.(B) C. eurytheme ; N�/47samples from 14 females. (C) S.mormonia , N�/56 samplesfrom 13 females. (D) H.charitonia ; N�/32 samplesfrom 9 females.
Table 5. Parameters estimated by the larval to adult turnover model for each species, given with estimated standard errors.
Parameter estimated E. chalcedona S. mormonia H. charitonia C. eurytheme
Max. % adult carbon (p) 0.449/0.05 0.799/0.05 0.669/0.02 0.559/0.02Turnover rate (r) 0.249/0.06 0.189/0.03 0.289/0.05 0.329/0.04Adult diet fractionation (fa) 3.79/1.9 �/1.89/0.9 �/1.899/0.34 0.49/1.0d13C larval C �/29.09/0.8 �/33.89/1.4 see below1 �/27.29/1.0
d13C (measured) of late-instar larvae (N) �/29.09/0.07 (2) �/34.49/0.2 (11) �/329/0.9 (3) �/26.59/0.1 (6)
1Model was unable to estimate d13C larval C for H. charitonia . Because the estimated values of ‘‘d13C larval C’’ were close to measuredd13C values of larvae (within 1�) in the other species, H. charitonia d13C larval was fixed at the measured larval value in order to fitthe other parameters.
Fig. 5. The proportion of egg carbon deriving from the adultdiet, vs female age, for each species. Proportions for each daywere calculated according to Eq. 5, using the model (Eq. 4) toestimate the mean value of d13C C3 egg and d13C C4 egg for eachday (see Methods). We also present the equivalent curve for thehawkmoth A. floridensis, published elsewhere (O’Brien et al.2000).
OIKOS 105:2 (2004) 287
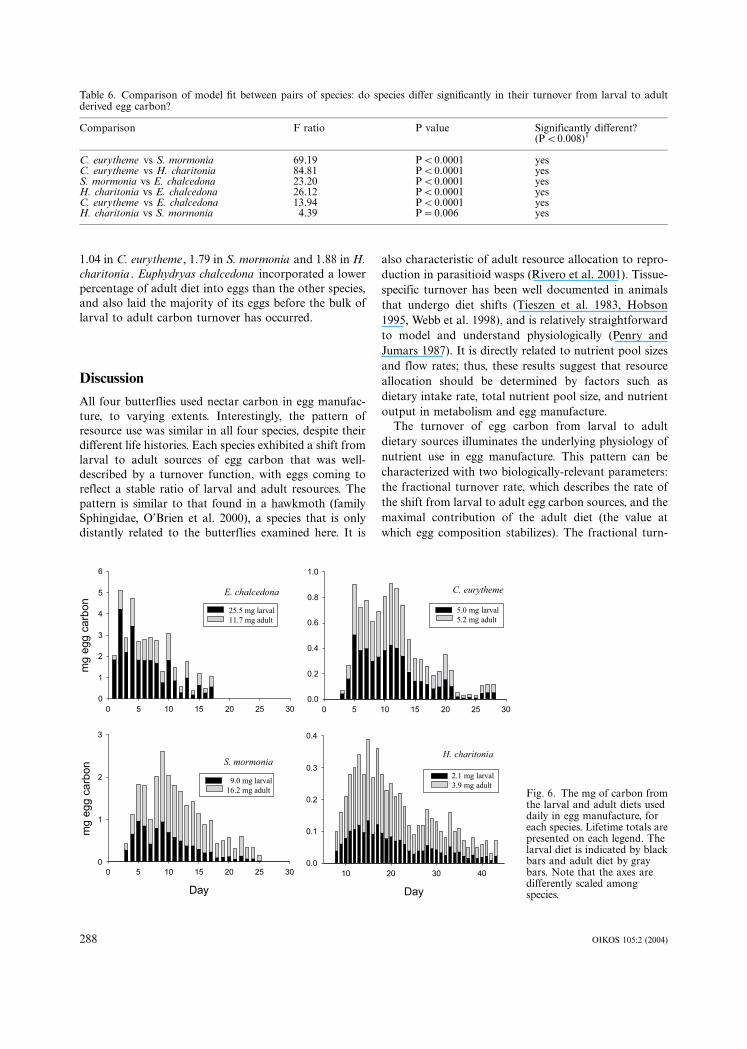
1.04 in C. eurytheme, 1.79 in S. mormonia and 1.88 in H.
charitonia . Euphydryas chalcedona incorporated a lower
percentage of adult diet into eggs than the other species,
and also laid the majority of its eggs before the bulk of
larval to adult carbon turnover has occurred.
Discussion
All four butterflies used nectar carbon in egg manufac-
ture, to varying extents. Interestingly, the pattern of
resource use was similar in all four species, despite their
different life histories. Each species exhibited a shift from
larval to adult sources of egg carbon that was well-
described by a turnover function, with eggs coming to
reflect a stable ratio of larval and adult resources. The
pattern is similar to that found in a hawkmoth (family
Sphingidae, O’Brien et al. 2000), a species that is only
distantly related to the butterflies examined here. It is
also characteristic of adult resource allocation to repro-
duction in parasitioid wasps (Rivero et al. 2001). Tissue-
specific turnover has been well documented in animals
that undergo diet shifts (Tieszen et al. 1983, Hobson
1995, Webb et al. 1998), and is relatively straightforward
to model and understand physiologically (Penry and
Jumars 1987). It is directly related to nutrient pool sizes
and flow rates; thus, these results suggest that resource
allocation should be determined by factors such as
dietary intake rate, total nutrient pool size, and nutrient
output in metabolism and egg manufacture.
The turnover of egg carbon from larval to adult
dietary sources illuminates the underlying physiology of
nutrient use in egg manufacture. This pattern can be
characterized with two biologically-relevant parameters:
the fractional turnover rate, which describes the rate of
the shift from larval to adult egg carbon sources, and the
maximal contribution of the adult diet (the value at
which egg composition stabilizes). The fractional turn-
Table 6. Comparison of model fit between pairs of species: do species differ significantly in their turnover from larval to adultderived egg carbon?
Comparison F ratio P value Significantly different?(PB/0.008)1
C. eurytheme vs S. mormonia 69.19 PB/0.0001 yesC. eurytheme vs H. charitonia 84.81 PB/0.0001 yesS. mormonia vs E. chalcedona 23.20 PB/0.0001 yesH. charitonia vs E. chalcedona 26.12 PB/0.0001 yesC. eurytheme vs E. chalcedona 13.94 PB/0.0001 yesH. charitonia vs S. mormonia 4.39 P�/0.006 yes
Fig. 6. The mg of carbon fromthe larval and adult diets useddaily in egg manufacture, foreach species. Lifetime totals arepresented on each legend. Thelarval diet is indicated by blackbars and adult diet by graybars. Note that the axes aredifferently scaled amongspecies.
288 OIKOS 105:2 (2004)

over rate was quite consistent across all species studied
to date; from 3�/5 days to achieve 50% turnover.
Another factor contributing to dietary resource use is
how much sugar-derived dietary carbon a species can or
will use in their eggs. This parameter, termed ‘‘pmax’’,
varied widely among the different species in this study.
At one extreme, E. chalcedona eggs never contained
more than 44% adult carbon, whereas S. mormonia eggs
came to comprise nearly 80% adult carbon. All of the
butterflies appeared to consume sufficient carbon to
make up for losses to eggs, although a caveat is that
carbon lost in respiration and excretion was not
quantified in this study. Egg composition is likely to
play a role in determining the extent to which adult
dietary carbon is used. Eggs are biochemically complex,
containing primarily protein and lipid, but also glycogen
granules, hydrocarbons, and nucleic acids (Nation 2002).
Approximately half of the carbon in egg protein derives
from essential amino acids (Table 4). These amino acids
have been shown to derive exclusively from the larval
diet in Amphion floridensis (O’Brien et al. 2002).
Essential fatty acids (and the fatty acids that derive
from them) also occur in lepidopteran eggs, estimated
for one species as 33% of all egg fatty acids (Forte et al.
2002). Thus, requirements for essential nutrients not
abundant in nectar diets limit the extent to which a
species can use adult dietary nutrients in egg manufac-
ture. For example, pollen-feeding H. charitonia have
access to essential amino acids (O’Brien et al. 2003), and
would be expected to rely more heavily on the adult diet
than the exclusively sucrose-fed H. charitonia in this
study.
For the species studied here, the percentage of egg
carbon deriving from essential amino acids is presented
in Table 4. This percentage approximates the minimum
possible carbon contribution from the larval diet (if
essential amino acids were the only carbon compound
not synthesized from sugar). We plotted these values for
each species against the % egg carbon that is exclusively
larval in origin (1�/pmax) (Fig. 7), including data
previously published from the hawkmoth A. floridensis
(O’Brien et al. 2000). This relationship provides some
insight into the differences among species in carbon
utilization. S. mormonia, H. charitonia, A. floridensis
and (to a lesser degree) C. eurytheme all fall fairly close
to the line y�/x, indicating that they are incorporating
nearly the maximum amount of adult dietary carbon
possible, given their individual egg composition. E.
chalcedona is notably different: it seems to incorporate
much less adult dietary carbon than it theoretically
‘‘could’’, based on egg composition.
The difference in E. chalcedona may lie in the extent of
egg provisioning that takes place prior to adult emer-
gence. Based on ovarian dissections, E. chalcedona
provisions its eggs at a much higher level prior to adult
emergence than do the other three species. These data
suggest two diet use ‘‘strategies’’ here: one in which a
butterfly makes as much of its eggs from adult diet as is
biochemically possible (S. mormonia, H. charitonia,C.
eurytheme and A. floridensis ), and the other in which the
butterfly foregoes the potential carbon influx from the
adult diet in favor of egg provisioning during pupal
development, and more rapid reproduction (E. chalce-
dona ). The smallest oocytes (corresponding to the last
eggs to be laid) were similarly sized among all four
species at emergence, suggesting at least the possibility
that E. chalcedona could use more adult resources to
provision the eggs laid latest in life. However, E.
chalcedona concentrates oviposition in the first week of
adult life; therefore, it is likely that even the least mature
oocytes are provisioned in early adulthood, before much
turnover to adult nutrients has occurred.
S. mormonia has a higher proportion of non-protein
egg nutrient than the other butterflies (high C/N), and
exhibits the highest % adult dietary carbon in its eggs. A
possible explanation for the high egg C/N is glycogen.
Glycogen stores are converted into the colligative anti-
freezes glycerol and sorbitol in freeze-avoiding Lepidop-
tera (Chino 1957, Storey 1990, Pullin et al. 1991).
Because S. mormonia overwinters as unfed, 1st instar
hatchlings at high altitude in the Rocky Mountains,
antifreeze production is likely to be a factor in over-
winter survivorship. Thus, we predict that S. mormonia
eggs are especially glycogen rich, causing nectar feeding
to be especially important to egg provisioning in this
species.
By combining the data on egg carbon turnover with
age-specific fecundity, we can calculate the total con-
tribution of the larval and adult diets to egg manufac-
ture, summed across a female’s lifetime (Fig. 6). The
larval diet supplied the majority of carbon for egg
Fig. 7. The proportion of egg carbon for each species that isexclusively larval in origin, plotted against the proportion of eggcarbon that derives from essential amino acids. The lineindicates y�/x. Species that fall close to y�/x are deriving amaximum of egg raw materials from the adult diet; those thatfall far away rely to a greater degree on larval resources thanbiochemical composition requires.
OIKOS 105:2 (2004) 289

manufacture in E. chalcedona (69%), the butterfly that
oviposits immediately after mating. In C. eurytheme,
input from the larval and the adult diet is fairly even (49
vs 51%), whereas in both S. mormonia and H. charitonia
the adult diet supplies the majority of egg carbon (65%).
One might expect S. mormonia to resemble C. eurytheme
in carbon allocation because their timing of oviposition
is similar; however, S. mormonia uses more adult dietary
carbon on a per egg basis. This difference has a
significant impact on lifetime resource use. Thus, the
extent to which dietary resources are important in egg
manufacture in butterflies depends on development (egg
provisioning in teneral adults), behavior (timing of
oviposition) and nutritional physiology (nutrient synth-
esis and turnover). Further understanding of this inter-
play is critical if we are to interpret variation in nutrient
use in light of reproductive strategies.
Acknowledgements �/ We would like to thank Jesse Boyett, ZoeChafe, Becky Chaplin, Liz Forwand, Sharon Komarow, JanetLarsen, Margie Mayfield, Sarah Oehm, Joanne White andGwen Yoshimura for their help in butterfly rearing and care,and Ward Watt for providing Colias eurytheme. We also thankKlaus Fischer for his helpful comments on this manuscript.This research was supported by NSF Grant IBN 98�/83044.
References
Alm, J., Ohnmeiss, T. E., Lanza, J. et al. 1990. Preference ofcabbage white butterflies and honey bees for nectar thatcontains amino acids. �/ Oecologia 84: 53�/57.
Baker, H. G and Baker, I. 1983. Floral nectar sugar constituentsin relation to pollinator type. �/ In: Jones, C. E. and Little,R. J. (eds), A handbook of experimental pollination biology.Van Nostrand-Reinhold, New York.
Boggs, C. L. 1979. Resource allocation and reproductivestrategies in several heliconiine butterflies. �/ Univ. of Texas,Austin.
Boggs, C. L. 1981. Nutritional and life-history determinants ofresource allocation in holometabolous insects. �/ Am. Nat.117: 692�/709.
Boggs, C. L. 1986. Reproductive strategies of female butterflies:variation in and constraints on fecundity. �/ Ecol. Entomol.11: 7�/15.
Boggs, C. L. 1987. Within-population variation in the demo-graphy of Speyeria mormonia (Lepidoptera; Nymphalidae).�/ Holarct. Ecol. 10: 175�/184.
Boggs, C. L. 1990. A general model of the role of male-donatednutrients in female insects’ reproduction. �/ Am. Nat. 136:598�/617.
Boggs, C. L. 1992. Resource allocation: exploring connectionsbetween foraging and life history. �/ Funct. Ecol. 6: 508�/
518.Boggs, C. L. 1997a. Dynamics of reproductive allocation
fromjuvenile and adult feeding: radiotracer studies.�/ Ecology 78: 192�/202.
Boggs, C. L. 1997b. Reproductive allocation from reserves andincome in butterfly species with differing adult diets.�/ Ecology 78: 181�/191.
Boggs, C. L. 2003. Environmental variation, life histories, andallocation. �/ In: Boggs, C. L., Watt, W. B. and Ehrlich, P. R.(eds), Butterflies: ecology and evolution taking flight.Chicago Univ. Press, pp. 185�/206.
Boggs, C. L. and Ross, C. L. 1993. The effect of adult foodlimitation of life history traits in Speyeria mormonia(Lepidoptera: Nymphalidae). �/ Ecology 74: 433�/441.
Brodbeck, B. V., Mizell III, R. F. and Andersen, P. C. 1993.Physiological and behavioral adaptations of three species ofleafhoppers in response to the dilute nutrient content ofxylem fluid. �/ J. Insect Physiol. 39: 73�/81.
Chino, H. 1957. Conversion of glycogen to sorbitol and glycerolin the diapause egg of the Bombyx silkworm. �/ Nature 180:606�/607.
de Jong, G. and van Noordwijk, A. J. 1992. Aquisition andallocation of resources: genetic (co)variances, selection, andlife histories. �/ Am. Nat. 139: 749�/770.
Delobel, B., Rahbe, Y., Nardon, C. et al. 1993. Biochemical andcytological survey of tyrosine storage proteins in Coleop-tera: diversity of strategies. �/ Insect Biochem. Mol. Biol. 23:355�/365.
Djawdan, M., Chippindale, A. K., Rose, M. R. et al. 1998.Metabolic reserves and evolved stress resistance in Droso-phila melanogaster. �/ Physiol. Zool. 71: 584�/594.
Douglas, A. E. 1998. Nutritional interactions in insect�/micro-bial symbioses: aphids and their symbiotic bacteria Buch-nera. �/ Annu. Rev. Entomol. 43: 17�/37.
Dunlap-Pianka, H., Boggs, C. L. and Gilbert, L. E. 1977.Ovarian dynamics in heliconiine butterflies: programmedsenescence versus eternal youth. �/ Science 197: 487�/490.
Elser, J. J., Dobberfuhl, D. R., MacKay, N. A. et al. 1996.Organism size, life history, and N:P stoichiometry.�/ BioScience 46: 674�/684.
Forte, S. N., Ferrero, A. A. and Alonso, T. S. 2002. Content andcomposition of phosphoglycerols and neutral lipids atdifferent developmental stages of the eggs of the codlingmoth, Cydia pomonella (Lepidoptera: Tortricidae). �/ Arch.Insect Biochem. Physiol. 50: 121�/130.
Hill, C. J. 1989. The effect of adult diet on the biology ofbutterflies 2. The common crow butterfly, Euploea corecorinna . �/ Oecologia 81: 258�/266.
Hill, C. J. and Pierce, N. E. 1989. The effect of adult die on thebiology of butterflies 1. The common imperial blue,Jalmenus evagoras. �/ Oecologia 81: 249�/257.
Hobson, K. A. 1995. Reconstructing avian diets using stable-carbon and nitrogen isotope analysis of egg components:patterns of isotopic fractionation and turnover. �/ Condor97: 752�/762.
Hobson, K. A., Hughes, K. D. and Ewins, P. J. 1997. Usingstable-isotope analysis to identify endogenous and exogen-ous sources of nutrients in eggs of migratory birds:applications to Great Lakes contaminants research. �/ Auk114: 467�/478.
Karlsson, B. 1987. Variation in egg weight, oviposition rate andreproductive reserves with female age in a natural popula-tion of the speckled wood butterfly, Pararge aegeria . �/ Ecol.Entomol. 12: 473�/476.
Markow, T. A., Raphael, B., Dobberfuhl, D. et al. 1999.Elemental stoichiometry of Drosophila and their hosts.�/ Funct. Ecol. 13: 78�/84.
Metges, C., Kempe, K. and Schmidt, H. L. 1990. Dependence ofthe carbon-isotope contents of breath carbon dioxide, milk,serum, and rumen fermentation products on the d13C valueof food in dairy cows. �/ Brit. J. Nutr. 63: 187�/196.
Motulsky, H. J. and Ransnas, L. A. 1987. Fitting curves to datausing nonlinear regression: a practical and nonmathematicalreview. �/ FASEB J. 1: 365�/374.
Mevi-Schutz, J. and Erhardt, A. 2002. Can Inachis io detectnectar amino acids at low concentrations? �/ Physiol.Entomol. 27: 256�/260.
Nation, J. L. 2002. Insect physiology and biochemistry. �/ CRCPress.
Norris, M. J. 1934. Contributions towards the study of insectfertility. III. Adult nutrition, fecundity, and longevity in thegenus Ephesia (Lepidoptera, Phycitidae). �/ Proc. Zool. Soc.Lond. 1934: 333�/360.
O’Brien, D. M., Boggs, C. L. and Fogel, M. L. 2003. Pollenfeeding in Heliconius charitonia : isotopic evidence foressential amino acid transfer from pollen to eggs. �/ Proc.R. Soc. Lond.: B 270: 2631�/2636.
290 OIKOS 105:2 (2004)

O’Brien, D. M., Fogel, M. L. and Boggs, C. L. 2002. Renewableand nonrenewable resources: amino acid turnover andallocation to reproduction in Lepidoptera. �/ Proc. NatlAcad. Sci. USA 99: 4413�/4418.
O’Brien, D. M., Schrag, D. P. and Martınez del Rio, C. 2000.Allocation to reproduction in a hawkmoth: a quantitativeanalysis using stable carbon isotopes. �/ Ecology 81: 2822�/
2831.O’Leary, M. H. 1981. Carbon isotope fractionation in plants.
�/ Phytochemistry 20: 553�/567.O’Leary, M. H. 1988. Carbon isotopes in photosynthesis.
�/ BioScience 38: 328�/336.Ostrom, P. H., Colunga Garcia, M. and Gage, S. H. 1997.
Establishing pathways of energy flow for insect predatorsusing stable isotope ratios: field and laboratory evidence.�/ Oecologia (Berlin) 109: 108�/113.
Pan, M. L. and Telfer, W. H. 2001. Storage hexamer utilizationin two lepidopterans: difference correlated with the timing ofegg formation. �/ J. Insect Sci. 1.2: 9.
Penry, D. L. and Jumars, P. A. 1987. Modeling animal guts aschemical reactors. �/ Am. Nat. 129: 69�/96.
Peterson, B. J. and Fry, B. 1987. Stable isotopes in ecosystemstudies. �/ Annu. Rev. Ecol. Syst. 18: 293�/320.
Pullin, A. S., Bale, J. S. and Fontaine, X. L. R. 1991.Physiological aspects of diapause and cold tolerance duringoverwintering in Pieris brassicae. �/ Physiol. Entomol. 16:447�/456.
Reznick, D. 1985. Costs of reproduction: an evaluation of theempirical evidence. �/ Oikos 44: 247�/267.
Rivero, A. and Casas, J. 1999. Incorporating physiology intoparasitoid behavioral ecology: the allocation of nutritionalresources. �/ Res. Popul. Ecol. 41: 39�/45.
Rivero, A., Giron, D. and Casas, J. 2001. Lifetime allocation ofjuvenile and adult nutritional resources to egg production ina holometabolous insect. �/ Proc. R. Soc. Lond. B 268:1231�/1237.
Romeis, J. and Wackers, F. L. 2000. Feeding responses by femalePieris brassicae butterflies to carbohydrates and aminoacids. �/ Physiol. Entomol. 25: 247�/253.
Rose, M. R. and Bradley, T. J. 1998. Evolutionary physiology ofthe cost of reproduction. �/ Oikos 83: 443�/451.
Rounick, J. S. and Winterbourn, M. J. 1986. Stable carbonisotopes and carbon flow in ecosystems. �/ Bioscience 36:171�/177.
Rusterholz, H. P. and Erhardt, A. 1998. Do peacock butterflies(Inachis io L. ) detect and prefer nectar amino acids andother nitrogenous compounds? �/ Oecologia 117: 536�/542.
Schmidt, O., Scrimgeour, C. M. and Curry, J. P. 1999. Carbonand nitrogen stable isotope ratios in body tissue and mucusof feeding and fasting earthworms (Lumbricus festivus ).�/ Oecologia 118: 9�/15.
Sinervo, B. and Svensson, E. 1998. Mechanistic and selectivecauses of life history trade-offs and plasticity. �/ Oikos 83:432�/442.
Slansky, F. J. and Feeny, P. 1977. Stabilization of the rate ofnitrogen accumulation by larvae of the cabbage butterfly onwild and cultivated food plants. �/ Ecol. Monogr. 47: 209�/
228.Smith, A. J. 1997. Postcolumn amino acid analysis. �/ Methods
Mol. Biol.: protein sequencing protocols 64: 139�/146.Snigirevskaya, E. S., Hays, A. R. and Raikhel, A. S. 1997.
Secretory and internalization pathways of mosquito yolkprotein precursors. �/ Cell Tissue Res. 290: 129�/142.
Stearns, S. 1992. The evolution of life histories. �/ Oxford Univ.Press.
Stern, V. M. and Smith, R. F. 1960. Factors affecting eggproduction and oviposition in populations of Colias philo-dice eurytheme Boisduval (Lepidoptera: Pieridae). �/ Hil-gardia 29: 411�/454.
Storey, K. B. 1990. Biochemical adaptation for cold hardiness ininsects. �/ Philos. Trans. R. Soc. Lond. Ser. B-Biol. Sci. 326:635�/654.
Tieszen, L. L., Boutton, T. W., Tesdahl, K. G. et al. 1983.Fractionation and turnover of stable isotopes in animaltissues: implications for d13C analysis of diet. �/ Oecologia57: 32�/37.
Webb, S. C., Hedges, R. E. M. and Simpson, S. J. 1998. Dietquality influences the delta C-13 and delta N-15 of locustsand their biochemical components. �/ J. Exp. Biol. 201:2903�/2911.
Wheeler, D. 1996. The role of nourishment in oogenesis.�/ Annu. Rev. Entomol. 41: 407�/431.
Wheeler, D. E. and Buck, N. A. 1995. Storage proteins in antsduring development and colony founding. �/ J. InsectPhysiol. 41: 885�/894.
Wheeler, D. E., Tuchinskaya, I., Buck, N. A. et al. 2000.Hexameric storage proteins during metamorphosis and eggproduction in the diamondback moth; Plutella xylostella(Lepidoptera). �/ J. Insect Physiol. 46: 951�/958.
Wickman, P. O. and Karlsson, B. 1987. Changes in egg colour,egg weight and oviposition rate with the number of eggs laidby wild females of the small heath butterfly, Coenonymphapamphilus. �/ Ecol. Entomol. 12: 109�/114.
Wolf, B. O. and Martınez del Rio, C. 2000. Use of saguaro fruitby white-winged doves: isotopic evidence of a tight ecolo-gical association. �/ Oecologia 124: 536�/543.
Zera, A. J. and Harshman, L. G. 2001. The physiology of lifehistory trade-offs in animals. �/ Annu. Rev. Ecol. Syst. 32:95�/126.
Zera, A. J., Potts, J. and Kobus., K. 1998. The physiology of life-history trade-offs: experimental analysis of a hormonallyinduced life-history trade-off in Gryllus assimilis. �/ Am.Nat. 152: 7�/23.
Zhao, Z. and Zera, A. J. 2002. Differential lipid biosynthesisunderlies a trade-off between reproduction and flight-cap-ability in a wing-polymorphic cricket. �/ Proc. Natl Acad.Sci. USA 99: 16829�/16834.
OIKOS 105:2 (2004) 291




















