
A Novel Isoform of Acetylcholinesterase Exacerbates Photoreceptors Death after Photic Stress
Upload
ua-birminghamCategory
view
2download
0
Loss of the Major Isoform of Phosphoglucomutase Results in AlteredCalcium Homeostasis in Saccharomyces cerevisiae*
(Received for publication, September 22, 1999, and in revised form, November 2, 1999)
Lianwu Fu‡, Attila Miseta‡§, Dacia Hunton¶, Richard B. Marchase¶, and David M. Bedwell‡i
From the Departments of ‡Microbiology and ¶Cell Biology, University of Alabama at Birmingham,Birmingham, Alabama 35294
Phosphoglucomutase (PGM) is a key enzyme in glu-cose metabolism, where it catalyzes the interconversionof glucose 1-phosphate (Glc-1-P) and glucose 6-phos-phate (Glc-6-P). In this study, we make the novel obser-vation that PGM is also involved in the regulation ofcellular Ca21 homeostasis in Saccharomyces cerevisiae.When a strain lacking the major isoform of PGM(pgm2D) was grown on media containing galactose assole carbon source, its rate of Ca21 uptake was 5-foldhigher than an isogenic wild-type strain. This increasedrate of Ca21 uptake resulted in a 9-fold increase in thesteady-state total cellular Ca21 level. The fraction ofcellular Ca21 located in the exchangeable pool in thepgm2D strain was found to be as large as the exchange-able fraction observed in wild-type cells, suggesting thatthe depletion of Golgi Ca21 stores is not responsible forthe increased rate of Ca21 uptake. We also found thatgrowth of the pgm2D strain on galactose media is inhib-ited by 10 mM cyclosporin A, suggesting that activation ofthe calmodulin/calcineurin signaling pathway is re-quired to activate the Ca21 transporters that sequesterthe increased cytosolic Ca21 load caused by this highrate of Ca21 uptake. We propose that these Ca21-relatedalterations are attributable to a reduced metabolic fluxbetween Glc-1-P and Glc-6-P due to a limitation of PGMenzymatic activity in the pgm2D strain. Consistent withthis hypothesis, we found that this “metabolic bottle-neck” resulted in an 8-fold increase in the Glc-1-P levelcompared with the wild-type strain, while the Glc-6-Pand ATP levels were normal. These results suggest thatGlc-1-P (or a related metabolite) may participate in thecontrol of Ca21 uptake from the environment.
Saccharomyces cerevisiae shares many features of Ca21 ho-meostasis and signaling pathways with mammals (1). Likeother eukaryotes, yeast maintain a low resting free Ca21 con-centration in their cytosol in the range of 50–200 nM (2–5).Considering that the total cellular Ca21 content is in the mil-limolar range, more than 99% of the total cell-associated Ca21
is either bound to proteins within the cytosol or compartmen-talized within intracellular organelles. This remarkably tightcontrol of free cytosolic Ca21 is a universal phenomenon among
living organisms that apparently relates to the low solubility ofCa21 complexes of many phosphorylated metabolites (6). Thistight control of free cytosolic Ca21 is also an essential feature ofCa21 signaling, where Ca21-sensing proteins like calmodulinutilize a transient increase in the steady-state cytosolic Ca21
concentration to activate various signal transduction pathways(7).
The great majority of cellular Ca21 is stored within intracel-lular organelles in S. cerevisiae (8). The vacuole is believed tocontain the largest (;90%) reservoir of Ca21, where it isthought to reside primarily in a complex with polyphosphate (9,10). In contrast, the endoplasmic reticulum (ER)1 and the Golgiapparatus hold Ca21 in a more readily mobilized form, which,together with cytosolic Ca21, is often referred to as the ex-changeable Ca21 pool (11). Cytosolic Ca21 can be transportedinto the Golgi apparatus by the Ca21-ATPase Pmr1p (12–15)and possibly by another putative Ca21 and Mn21 transporter,Ccc1p (16–18). Consistent with its capacity to store Ca21, arecent study demonstrated that the Golgi apparatus can play asignificant role in maintaining cellular Ca21 homeostasis un-der conditions where vacuolar Ca21 storage is compromised (4).Cytosolic Ca21 can also be sequestered into the vacuolethrough the action of the Ca21-ATPase Pmc1p and the vacuolarH1/Ca21 exchanger Vcx1p/Hum1p (17, 19, 20). Of these twotransporters, Vcx1p has been shown to play a predominant rolein pumping cytosolic Ca21 into the vacuole and restoring basalcytosolic Ca21 conditions following an increase in cytosolicCa21 (5, 17, 20). The expression and function of each of theseCa21 transporters is regulated by calcineurin, a highly con-served protein phosphatase that is activated by Ca21/calmod-ulin (4, 17, 20). While mitochondria have been shown to play animportant role in Ca21 signaling in mammalian cells, there iscurrently little or no evidence that mitochondria play any rolein Ca21 sequestration in S. cerevisiae (21, 22).
A number of stimuli can trigger a transient increase in thecytosolic Ca21 concentration in yeast. These stimuli includevarious nutrients, mating pheromones, or a hypotonic shock (3,23). While glucose is frequently considered to be primarily acarbon and energy source in yeast, it can also activate severalsignaling pathways in yeast, including the RAS-cAMP path-way, phosphatidylinositol turnover, Ca21 influx and efflux, andthe glucose repression/de-repression pathway (24). It is unclearwhether these distinct pathways are regulated by glucose, aglucose derivative, a glycosylated protein, or by some othermechanism. Eilam and co-workers (25, 26) have shown thatglucose-stimulated Ca21 uptake is not controlled by changes in
* This work was supported by Grant JDF 99502 from the JuvenileDiabetes Foundation International. The costs of publication of thisarticle were defrayed in part by the payment of page charges. Thisarticle must therefore be hereby marked “advertisement” in accordancewith 18 U.S.C. Section 1734 solely to indicate this fact.
§ Present address: Dept. of Clinical Chemistry, University MedicalSchool, 7624 Pecs, Hungary.
i To whom correspondence should be addressed: Dept. of Microbiol-ogy, Bevill Biomedical Research Bldg., Rm. 432, 845 19th St. S., Uni-versity of Alabama at Birmingham, Birmingham, AL 35294-2170. Tel.:205-934-6593; Fax: 205-975-5482; E-mail: [email protected].
1 The abbreviations used are: ER , endoplasmic reticulum; WT, wild-type; Glc-1-P, glucose 1-phosphate; Glc-6-P, glucose 6-phosphate; CsA,cyclosporin A; MES, 2-(N-morpholino)ethane sulfonic acid; Bis-Tris,bis(2-hydroxyethyl)iminotris(hydroxymethyl)methane; CpY, carboxy-peptidase Y; PGM, phosphoglucomutase; CIF, calcium influx factor;SM, synthetic medium.
THE JOURNAL OF BIOLOGICAL CHEMISTRY Vol. 275, No. 8, Issue of February 25, pp. 5431–5440, 2000© 2000 by The American Society for Biochemistry and Molecular Biology, Inc. Printed in U.S.A.
This paper is available on line at http://www.jbc.org 5431

either the ATP level or by a cAMP-dependent pathway. Inter-estingly, Glc-6-P has been shown to stimulate the ATP-depend-ent transport of Ca21 into mammalian microsomes (27, 28),indicating that glucose metabolites may directly influenceCa21 transport mechanisms in higher eukaryotes.
The biochemical pathways required to utilize glucose as acarbon and energy source are highly conserved from bacteria tohumans. A key enzyme involved in this process is phosphoglu-comutase (PGM; EC 2.7.5.1), which catalyzes the interconver-sion of Glc-1-P and Glc-6-P. PGM acts at a key metabolictrafficking point that controls the synthesis and degradation ofcarbohydrates. The direction of metabolic flow through PGMdepends on the available carbon source. Both the degradationof glycogen and the metabolism of galactose as carbon sourcerequire PGM activity to convert Glc-1-P to Glc-6-P. The Glc-6-Pproduced can then be utilized in glycolysis and the pentose-phosphate shunt. Conversely, PGM mediates the conversion ofGlc-6-P to Glc-1-P for sugar nucleotide (and ultimately, cellwall and glycogen) synthesis in cells grown on glucose or non-fermentable carbon sources. S. cerevisiae contains genes encod-ing two isoforms of PGM. Expression of the PGM2 gene, encod-ing the major isoform of PGM, is greatly increased in galactose-containing media. In contrast, the PGM1 gene expresses theminor isoform of PGM in a relatively low, constitutive manner(29, 30). Yeast strains that lack both isoforms of PGM areunable to grow when galactose is the sole carbon and energysource (31). However, strains lacking both isoforms of PGM cansurvive when grown on glucose, apparently because the con-version of Glc-6-P to Glc-1-P can be supported to a small extentby related enzymes such as phosphomannomutase andN-acetylglucosamine-1-phosphate mutase (31, 32).
In the current study, we found that a mutant strain lackingthe major isoform of PGM (pgm2D) has a much higher rate ofCa21 uptake than an isogenic wild-type strain when grown ongalactose media. This increased rate of Ca21 uptake results ina 9-fold increase in total cellular Ca21, but does not reduce thefraction of Ca21 located in the exchangeable pool. Consistentwith this observation, we could not detect any defect in ER andGolgi functions that have previously been shown to result froman inability to sequester Ca21 into these compartments in apmr1D strain (12, 13, 33). Interestingly, this Ca21 hyper-accu-mulation phenotype is observed only when the pgm2D strain isgrown in media containing galactose as the sole carbon source,
and correlates with the accumulation of Glc-1-P under thesegrowth conditions. These results suggest that Glc-1-P (or aderivative) may regulate Ca21 uptake from the environment.
MATERIALS AND METHODS
Strains Used—Yeast strains used in this study are listed in Table I.Sc252 and Sc451 were kindly provided by J. Hopper (30). W303–1C andYRP032 were gifts from H. K. Rudolph (33). Yeast strains YDB0171,YDB0200, YDB0322, and YDB0201 were generated using a one-stepgene replacement method (34). Briefly, the PGM2 gene in YDB0171,YDB0322, and YDB0201 was disrupted by the insertion of the LEU2gene. A 1.5-kilobase pair HpaI/HindIII DNA fragment containing mostof the structural gene of PGM2 in pDB0197 was removed by the inser-tion of a BamHI/HindIII fragment containing the LEU2 gene frompJJ282 (35). The PGM1 genes in YDB0200 and YDB0201 were dis-rupted by the insertion of the URA3 gene. A 2-kilobase pair fragment ofthe PGM1 gene was generated by polymerase chain reaction usingwild-type genomic DNA as template. The forward primer used wasDB225 (59-CAA GAC TCG AGA AGG GCG CAT CAC) and DB226(59-GAT CTT GAA TTC CTG TAC GGC TCT GC). These primers con-tain XhoI and EcoRI restriction endonuclease sites, respectively (un-derlined). The polymerase chain reaction product was digested withEcoRI and XhoI and cloned into a pBluescript KS II (1) plasmid. The730-base pair HindIII segment of the PGM1 gene was replaced by theHindIII fragment containing URA3 gene from pJJ244 (35). The EcoRI/XhoI fragment containing the disrupted pgmD1::URA3 was then usedto transform yeast. In all cases, the correct gene replacements wereconfirmed by Southern blot analysis (data not shown). YDB0310 andYDB0312 were generated by backcrossing the Sc451 strain with YR122(36) by standard methods (37).
Culture Medium—Bacteria strains were grown on standard media(38). Media for yeast growth were prepared as described (37). YP mediaand synthetic media (SM) was supplemented with 2% glucose (YPD orSMD) or 2% galactose (YPGal or SMGal) All media containing Ca21 orEGTA were routinely buffered with 40 mM MES-Tris, pH 5.5 or 6.5 asspecified. In all experiments, cultures were grown for a minimum of 5–6generations to an A600 of #1.0.
Measurement of Total Cellular Ca21 Levels, Cytosolic Free Ca21
Concentration, the Rate of Ca21 Uptake, and the Efflux of Ca21—Totalcellular Ca21 levels were measured as described earlier (4). Briefly,50–100 OD600 units of yeast cells grown in YPD or YPGal were har-vested and washed. The cell pellets were dried in a Savant SpeedVacsystem and resuspended in 1 M HCl. The total calcium levels were thenmeasured by using a flame photometry method and expressed in molesper kilogram of dry mass.
The cytosolic free Ca21 was measured by using an aequorin-basedmethod as described (4, 23). Briefly, the cells of the wild-type andpgm2D strains containing the apoaequorin-expressing plasmid weregrown in SMGal media supplemented with the required amino acids.
TABLE IYeast strains used in this study
Strains Relevantgenotype Complete genotype Reference
Sc252 (SJ21R) Wild-type MATa leu2–3 112, ura3–52, ade1 MEL (30)Sc451 (21R-5UD) pgm2D MATa leu2–3 112, ura3–52, ade1, MEL
pgm2D<URA3(30)
YDB0171 pgm2D MATa leu2–3 112, ura3–52, ade1, MELpgm2D<LEU2
This study
YDB0200 pgm1D MATa leu2–3 112, ura3–52, ade1, MELpgm1D<URA3
This study
YDB0201 pgm1D/pgm2D MATa leu2–3 112, ura3–52, ade1, MELpgm2D<LEU2, pgm1D<URA3
This study
YDB0312 pmr1D MATa leu2–3 112, ura3–52, ade2–101, his3-D200, pmr1-D1<LEU2
This study
YDB0310 pgm2D/pmr1D MATa leu2–3 112, ura3–52, ade2–101, his3-D200, pgm2D<URA3, pmr1-D1<LEU2
This study
W303–1C prc1–1 MATa ade2–1oc, ura3–1, his3–1115, leu2–3112, trp1–1, can1–100, prc1–1
(33)
YRP023 pmr1D/prc1–1 MATa ade2–1oc, ura3–1, his3–1115, leu2–3112, trp1–1, can1–100, prc1–1,pmr1D<HIS3
(33)
YDB0322 pgm2D/prc1–1 MATa ade2–1oc, ura3–1, his3–1115, leu2–3112, trp1–1, can1–100, prc1–1,pgm2D<LEU2
This study
Altered Calcium Homeostasis in a Phosphoglucomutase Mutant5432

The cells were then harvested and loaded with coelenterazine. Forglucose- or galactose-stimulated Ca21 uptake, the cells were loadedwith coelenterazine, then were incubated without a carbon source for2 h in SM medium supplemented with 2 mM EGTA. To confirm that theglucose- or galactose-stimulated increase in cytosolic Ca21 was depend-ent upon extracellular Ca21, cells were incubated in the presence orabsence of 5 mM CaCl2 for 10 min prior to the addition of glucose orgalactose. As previously reported (3), the increase in cytosolic Ca21 wasstrictly dependent upon extracellular Ca21. In these experiments, ae-quorin luminescence was measured using a Berthold Lumat 9050 lu-minometer. The data were recorded and transferred to a personalcomputer for analysis. The cytosolic free Ca21 was calculated using astandard curve (4).
The rate of Ca21 uptake was measured as described before (14). Thecells were harvested from an exponential phase culture, washed threetimes with distilled water, and resuspended in a buffer containing 40mM MES-Tris, pH 5.5, and 20 mM D-glucose. The Ca21 uptake experi-ment was then initiated by the addition of [45Ca21] to a concentration of1 mCi/ml. At the times indicated, aliquots were filtered through a0.45-mm Gelman filter pre-washed with buffer containing 20 mM MgCl2and 0.2 mM LaCl3. The cells were then rapidly washed three times withthe same washing solution, and cell-associated [45Ca21] was determinedby scintillation counting. For the measurement of [45Ca21] uptake inglucose-starved cells, strains were grown in SMGal supplemented withrequired amino acids and then starved for carbon source for 2 h. Thecells were then harvested, washed three times with water, and resus-pended in 40 mM MES-Tris buffer, pH 5.5. Ca21 uptake was initiated bythe addition of 1% glucose and 1 mCi of [45Ca21]. At the indicated times,cells were filtered and washed, and the cell-associated radioactivity wasdetermined by scintillation counting. Nonspecific binding of [45Ca21] tothe cells at the zero time point was subtracted.
The Ca21 exchange experiments were done as described previously(4, 19). Wild-type and pgm2D strains were grown in YPGal for 5–6generations in the presence of 10 mCi/ml [45Ca21] before harvest atlogarithmic phase. The cells were harvested by centrifugation at roomtemperature, washed with YP medium, and then resuspended in YPGalplus 20 mM CaCl2. After incubating at 30 °C for the time indicated,aliquots of cells were harvested by rapid filtration, washed, and pro-cessed for scintillation counting as described above.
Phosphoglucomutase Assay—To assay for PGM activity, frozen cellpellets were resuspended to 50 OD600 units/ml in assay buffer (50 mM
Bis-Tris, pH 6.0, 150 mM NaCl, 1 mM phenylmethylsulfonyl fluoride).Cells were disrupted by mechanical agitation with glass beads in amini-bead beater (Biospec Products). Assays were carried out on totalcellular homogenates at 22 °C using a coupled enzyme assay as de-scribed (29). Specific activity is expressed as micromoles of productformed/min/mg of protein. Protein concentrations were determined bythe method of Bradford (39), with bovine serum albumin used to gen-erate the standard curve.
Measure of the Metabolite Levels—Extracts were made using amethod published previously (40). An aliquot of exponentially growingyeast culture was filtered under vacuum through a 0.45-mm Gelmanfilter. The wet weight of cells was rapidly determined, and the cellswere then frozen in liquid nitrogen. 3 ml of 3 M HClO4 was added to thefrozen cells, and they were then ground in the presence of glass beadsin a mortar cooled with liquid nitrogen. The extract was centrifuged at4 °C for 10 min at 10,000 3 g, and the supernatant was neutralized bythe addition of 5 M KClO4. Glc-6-P levels in the extract were measuredusing an assay coupled with Glc-6-P dehydrogenase (41). Glc-1-P levelswere measured similarly, with the addition of PGM. For the calculationof concentrations of each metabolite, it was assumed that 1.67 g of wetyeast contain 1 ml of cell volume (40).
The levels of ATP, ADP, and AMP were measured by high perform-ance liquid chromatography analysis. Briefly, yeast extracts were re-solved using a Keystone Partisil-10 SAX column using a linear salt andpH gradient from 5 to 750 mM (NH4)H2PO4 and from pH 2.8 to 3.7,respectively, at a flow rate of 2 ml/min over 40 min. External standardsof known concentration were used to identify and quantitate specificpeaks that corresponded to each metabolite. The energy charge of eachstrain was calculated as the sum of the ATP concentration plus one-halfthe ADP concentration divided by the sum of the concentrations of ATP,ADP, and AMP.
Analysis of Invertase Glycosylation and CpY* Degradation—Invert-ase glycosylation was determined from cells labeled with [35S] methio-nine/cysteine (Expre35S35S; NEN Life Science Products). Immunopre-cipitation was carried out using an invertase antibody (a gift from ScottEmr) as described previously (42). To amplify the invertase signal,strains containing pRB58 (43), a high copy invertase-expressing plas-
mid, were grown in SMGal medium supplemented with the requiredamino acids. 10 A600 units of cells were used for the initial immunopre-cipitation, followed by a second immunoprecipitation using the sameantibody. CpY* degradation was assayed by a pulse-chase experimentas described previously (33), except the cells were grown in SMGalmedium.
RESULTS
Growth Phenotype of a pgm2D Yeast Strain—PGM is a keyenzyme in glucose and galactose metabolism. In S. cerevisiaethere are two isoforms of PGM that are subject to distinctpatterns of regulation (29, 30). To better understand the phys-iological roles of PGM, we constructed strains containing singlepgm1D or pgm2D knockouts, as well as a strain containingknockouts of both genes (pgm1D/pgm2D). Initially, we com-pared the growth of these strains on YPD and YPGal media.Consistent with the results reported by Boles et al. (31), thepgm1D, pgm2D, and pgm1D/pgm2D strains could all grow aswell as the WT strain in YPD medium (data not shown). OnYPGal medium, the pgm1D strain grew as well as the WTstrain, while the growth of the pgm2D mutant was somewhatreduced. In contrast, the pgm1D/pgm2D strain could not growon YPGal media, confirming that PGM activity is essential forgrowth when galactose is the sole carbon source (31).
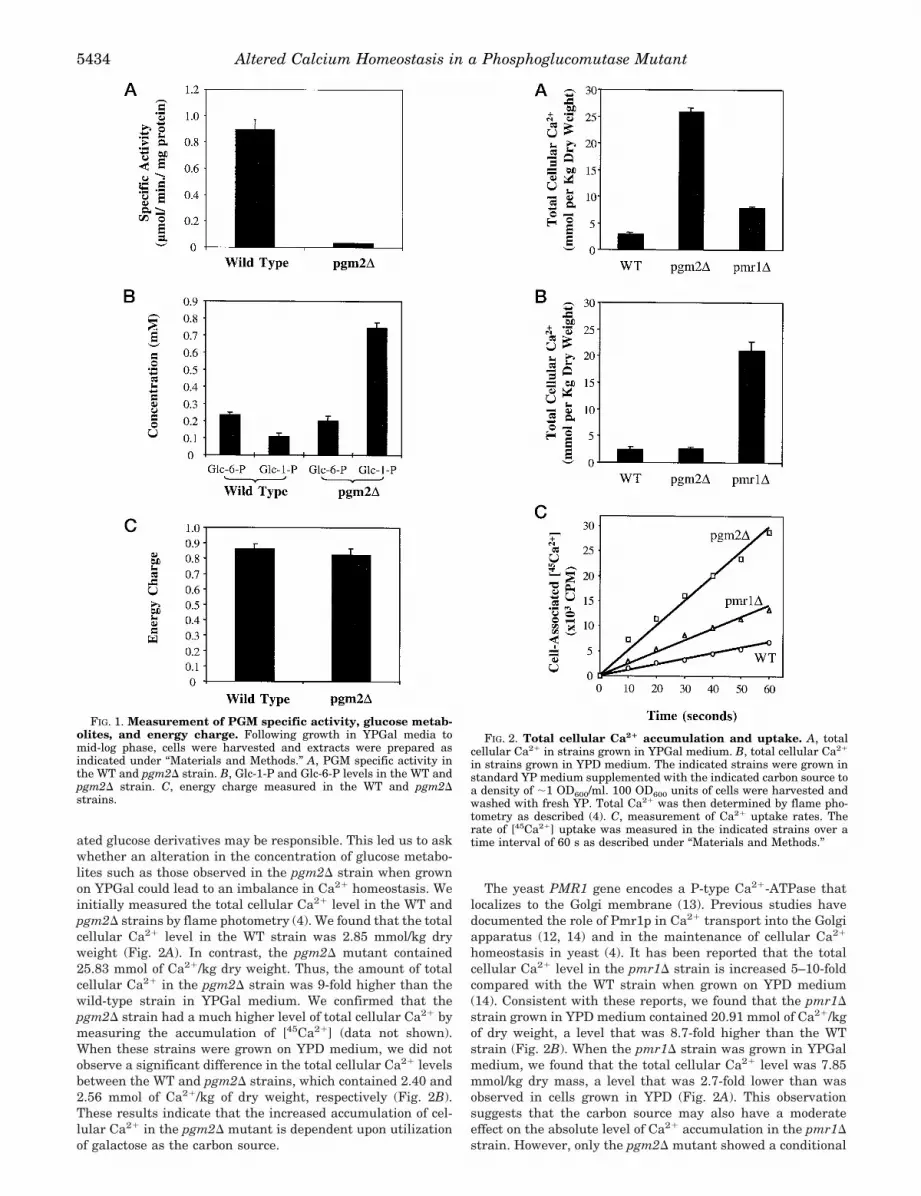
The pgm2D Strain Accumulates Glc-1-P When Grown onGalactose Media—PGM catalyzes the interconversion of Glc-6-P and Glc-1-P. To determine how the pgm1D and pgm2Dmutations affect the levels of sugar metabolites and energymetabolism in cells grown on galactose media, we measuredthe specific activity of PGM and the concentrations of Glc-1-Pand Glc-6-P in strains containing deletions of each PGM gene.As shown in Fig. 1A, inactivation of the PGM2 gene greatlyreduced the enzymatic activity of PGM in galactose-growncells. The specific activity of PGM in the WT strain was about0.9 mmol/min/mg of protein, while the pgm2D mutant wasabout 0.03 mmol/min/mg of protein. Thus, the pgm2D mutanthas 30-fold less PGM activity than the WT strain.
We next measured the levels of Glc-6-P and Glc-1-P in thesestrains. We found that the steady-state levels of Glc-6-P andGlc-1-P in the WT strain were 0.23 and 0.11 mM, respectively,when the cells were grown in galactose-containing media.While the pgm2D strain had a level of Glc-6-P that was notsignificantly different from the WT strain (0.20 mM), the Glc-1-P level in this strain was increased to 0.75 mM (Fig. 1B).Hence, the intracellular Glc-1-P concentration was 7-foldhigher in the pgm2D mutant than in the WT strain. Theseresults suggest that the limitation of PGM enzymatic activitycaused by the pgm2D mutation results in a metabolic bottle-neck that restricts the conversion of Glc-1-P to Glc-6-P. Likethe Glc-6-P level, the cellular energy charge in these strainswas also found to be similar (Fig. 1C), indicating that thepgm2D mutation did not cause a reduction in the ATP levelwhen galactose was utilized as the carbon source. Since thepgm2D strain exhibits a moderately slower growth rate thanthe WT strain, these results suggest that cellular growth maybe limited by the ability to generate a normal level of Glc-6-P(and ultimately, ATP). In contrast, no significant differences inthe levels of Glc-1-P, Glc-6-P, or the energy charge were meas-ured in these strains when they were grown in YPD media(data not shown).
The pgm2D Strain Contains an Elevated Level of Total Cel-lular Ca21—Previous studies have shown that glucose cantransiently stimulate Ca21 uptake in glucose-deprived yeastcells (3, 26). In addition, glucose can induce phosphatidylinosi-tol turnover and activation of the plasma membrane H1-ATPase in glucose-deprived cells (24). While the mechanism bywhich glucose activates these (and other) signaling pathways isnot well understood, it is thought that glucose or phosphoryl-
Altered Calcium Homeostasis in a Phosphoglucomutase Mutant 5433
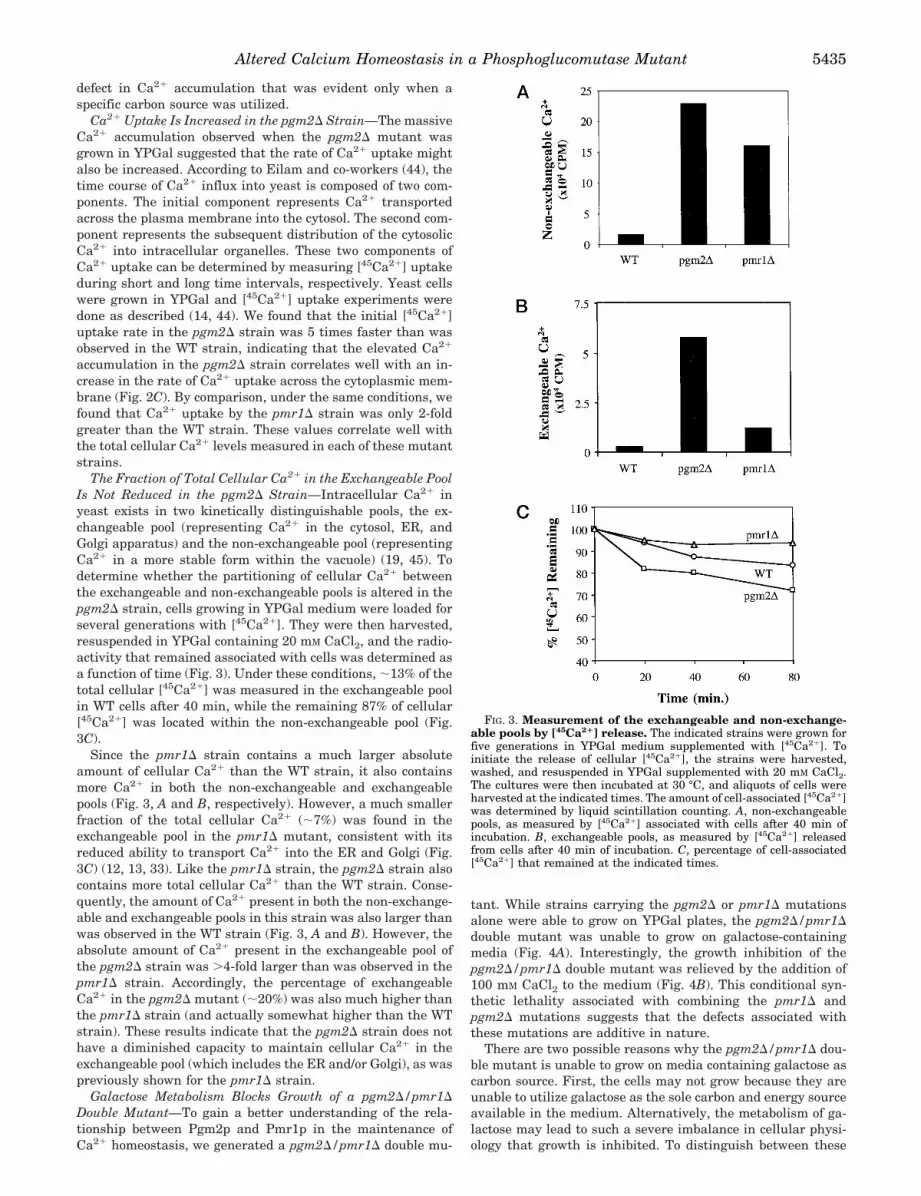
ated glucose derivatives may be responsible. This led us to askwhether an alteration in the concentration of glucose metabo-lites such as those observed in the pgm2D strain when grownon YPGal could lead to an imbalance in Ca21 homeostasis. Weinitially measured the total cellular Ca21 level in the WT andpgm2D strains by flame photometry (4). We found that the totalcellular Ca21 level in the WT strain was 2.85 mmol/kg dryweight (Fig. 2A). In contrast, the pgm2D mutant contained25.83 mmol of Ca21/kg dry weight. Thus, the amount of totalcellular Ca21 in the pgm2D strain was 9-fold higher than thewild-type strain in YPGal medium. We confirmed that thepgm2D strain had a much higher level of total cellular Ca21 bymeasuring the accumulation of [45Ca21] (data not shown).When these strains were grown on YPD medium, we did notobserve a significant difference in the total cellular Ca21 levelsbetween the WT and pgm2D strains, which contained 2.40 and2.56 mmol of Ca21/kg of dry weight, respectively (Fig. 2B).These results indicate that the increased accumulation of cel-lular Ca21 in the pgm2D mutant is dependent upon utilizationof galactose as the carbon source.
The yeast PMR1 gene encodes a P-type Ca21-ATPase thatlocalizes to the Golgi membrane (13). Previous studies havedocumented the role of Pmr1p in Ca21 transport into the Golgiapparatus (12, 14) and in the maintenance of cellular Ca21
homeostasis in yeast (4). It has been reported that the totalcellular Ca21 level in the pmr1D strain is increased 5–10-foldcompared with the WT strain when grown on YPD medium(14). Consistent with these reports, we found that the pmr1Dstrain grown in YPD medium contained 20.91 mmol of Ca21/kgof dry weight, a level that was 8.7-fold higher than the WTstrain (Fig. 2B). When the pmr1D strain was grown in YPGalmedium, we found that the total cellular Ca21 level was 7.85mmol/kg dry mass, a level that was 2.7-fold lower than wasobserved in cells grown in YPD (Fig. 2A). This observationsuggests that the carbon source may also have a moderateeffect on the absolute level of Ca21 accumulation in the pmr1Dstrain. However, only the pgm2D mutant showed a conditional
FIG. 1. Measurement of PGM specific activity, glucose metab-olites, and energy charge. Following growth in YPGal media tomid-log phase, cells were harvested and extracts were prepared asindicated under “Materials and Methods.” A, PGM specific activity inthe WT and pgm2D strain. B, Glc-1-P and Glc-6-P levels in the WT andpgm2D strain. C, energy charge measured in the WT and pgm2Dstrains.
FIG. 2. Total cellular Ca21 accumulation and uptake. A, totalcellular Ca21 in strains grown in YPGal medium. B, total cellular Ca21
in strains grown in YPD medium. The indicated strains were grown instandard YP medium supplemented with the indicated carbon source toa density of ;1 OD600/ml. 100 OD600 units of cells were harvested andwashed with fresh YP. Total Ca21 was then determined by flame pho-tometry as described (4). C, measurement of Ca21 uptake rates. Therate of [45Ca21] uptake was measured in the indicated strains over atime interval of 60 s as described under “Materials and Methods.”
Altered Calcium Homeostasis in a Phosphoglucomutase Mutant5434
defect in Ca21 accumulation that was evident only when aspecific carbon source was utilized.
Ca21 Uptake Is Increased in the pgm2D Strain—The massiveCa21 accumulation observed when the pgm2D mutant wasgrown in YPGal suggested that the rate of Ca21 uptake mightalso be increased. According to Eilam and co-workers (44), thetime course of Ca21 influx into yeast is composed of two com-ponents. The initial component represents Ca21 transportedacross the plasma membrane into the cytosol. The second com-ponent represents the subsequent distribution of the cytosolicCa21 into intracellular organelles. These two components ofCa21 uptake can be determined by measuring [45Ca21] uptakeduring short and long time intervals, respectively. Yeast cellswere grown in YPGal and [45Ca21] uptake experiments weredone as described (14, 44). We found that the initial [45Ca21]uptake rate in the pgm2D strain was 5 times faster than wasobserved in the WT strain, indicating that the elevated Ca21
accumulation in the pgm2D strain correlates well with an in-crease in the rate of Ca21 uptake across the cytoplasmic mem-brane (Fig. 2C). By comparison, under the same conditions, wefound that Ca21 uptake by the pmr1D strain was only 2-foldgreater than the WT strain. These values correlate well withthe total cellular Ca21 levels measured in each of these mutantstrains.
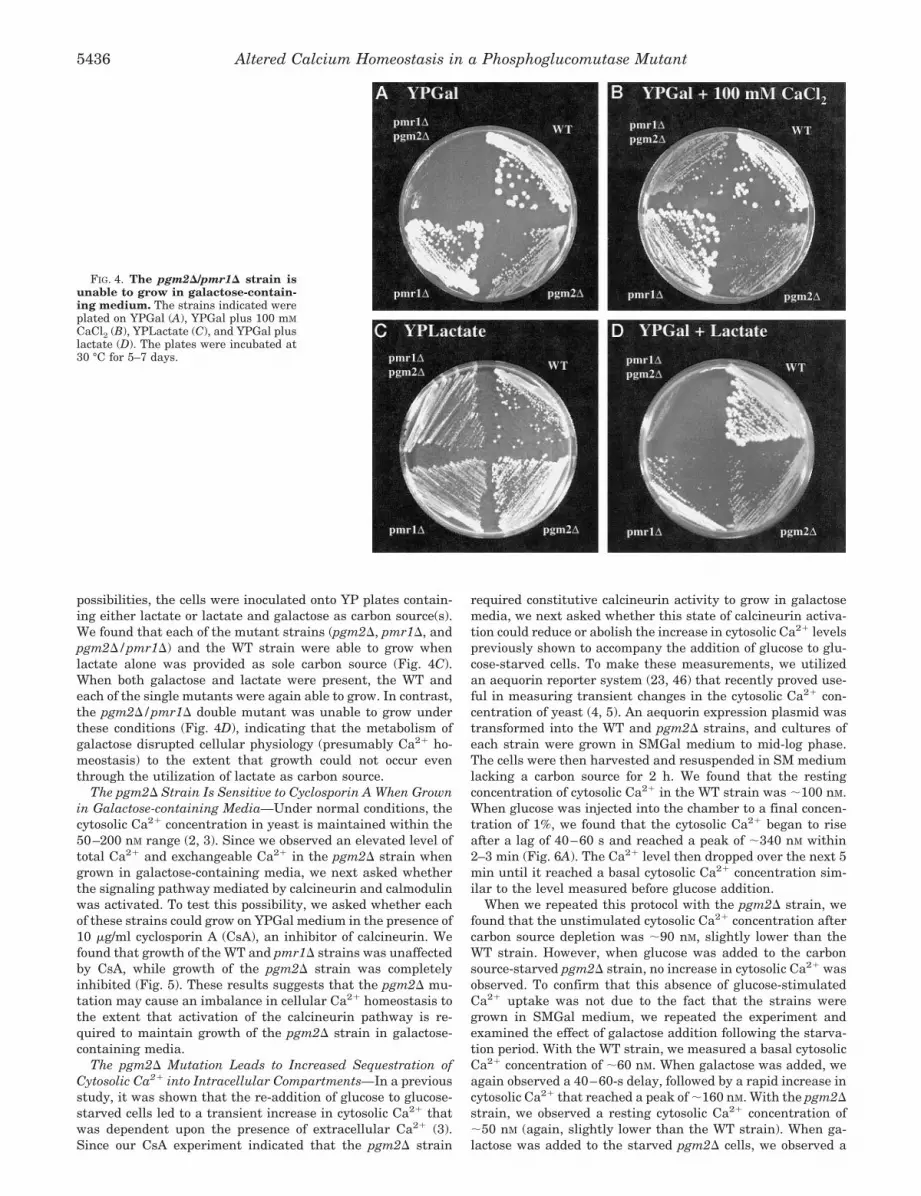
The Fraction of Total Cellular Ca21 in the Exchangeable PoolIs Not Reduced in the pgm2D Strain—Intracellular Ca21 inyeast exists in two kinetically distinguishable pools, the ex-changeable pool (representing Ca21 in the cytosol, ER, andGolgi apparatus) and the non-exchangeable pool (representingCa21 in a more stable form within the vacuole) (19, 45). Todetermine whether the partitioning of cellular Ca21 betweenthe exchangeable and non-exchangeable pools is altered in thepgm2D strain, cells growing in YPGal medium were loaded forseveral generations with [45Ca21]. They were then harvested,resuspended in YPGal containing 20 mM CaCl2, and the radio-activity that remained associated with cells was determined asa function of time (Fig. 3). Under these conditions, ;13% of thetotal cellular [45Ca21] was measured in the exchangeable poolin WT cells after 40 min, while the remaining 87% of cellular[45Ca21] was located within the non-exchangeable pool (Fig.3C).
Since the pmr1D strain contains a much larger absoluteamount of cellular Ca21 than the WT strain, it also containsmore Ca21 in both the non-exchangeable and exchangeablepools (Fig. 3, A and B, respectively). However, a much smallerfraction of the total cellular Ca21 (;7%) was found in theexchangeable pool in the pmr1D mutant, consistent with itsreduced ability to transport Ca21 into the ER and Golgi (Fig.3C) (12, 13, 33). Like the pmr1D strain, the pgm2D strain alsocontains more total cellular Ca21 than the WT strain. Conse-quently, the amount of Ca21 present in both the non-exchange-able and exchangeable pools in this strain was also larger thanwas observed in the WT strain (Fig. 3, A and B). However, theabsolute amount of Ca21 present in the exchangeable pool ofthe pgm2D strain was .4-fold larger than was observed in thepmr1D strain. Accordingly, the percentage of exchangeableCa21 in the pgm2D mutant (;20%) was also much higher thanthe pmr1D strain (and actually somewhat higher than the WTstrain). These results indicate that the pgm2D strain does nothave a diminished capacity to maintain cellular Ca21 in theexchangeable pool (which includes the ER and/or Golgi), as waspreviously shown for the pmr1D strain.
Galactose Metabolism Blocks Growth of a pgm2D/pmr1DDouble Mutant—To gain a better understanding of the rela-tionship between Pgm2p and Pmr1p in the maintenance ofCa21 homeostasis, we generated a pgm2D/pmr1D double mu-
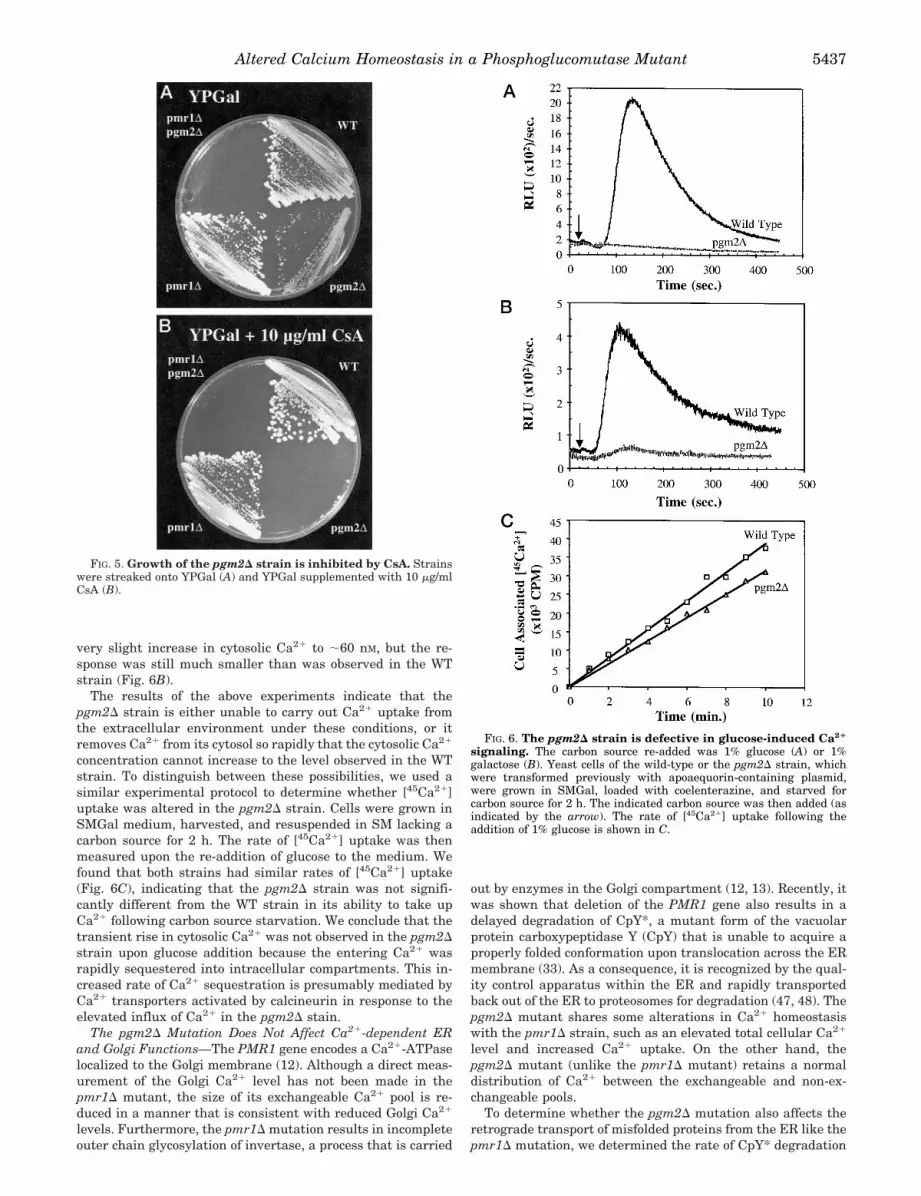
tant. While strains carrying the pgm2D or pmr1D mutationsalone were able to grow on YPGal plates, the pgm2D/pmr1Ddouble mutant was unable to grow on galactose-containingmedia (Fig. 4A). Interestingly, the growth inhibition of thepgm2D/pmr1D double mutant was relieved by the addition of100 mM CaCl2 to the medium (Fig. 4B). This conditional syn-thetic lethality associated with combining the pmr1D andpgm2D mutations suggests that the defects associated withthese mutations are additive in nature.
There are two possible reasons why the pgm2D/pmr1D dou-ble mutant is unable to grow on media containing galactose ascarbon source. First, the cells may not grow because they areunable to utilize galactose as the sole carbon and energy sourceavailable in the medium. Alternatively, the metabolism of ga-lactose may lead to such a severe imbalance in cellular physi-ology that growth is inhibited. To distinguish between these
FIG. 3. Measurement of the exchangeable and non-exchange-able pools by [45Ca21] release. The indicated strains were grown forfive generations in YPGal medium supplemented with [45Ca21]. Toinitiate the release of cellular [45Ca21], the strains were harvested,washed, and resuspended in YPGal supplemented with 20 mM CaCl2.The cultures were then incubated at 30 °C, and aliquots of cells wereharvested at the indicated times. The amount of cell-associated [45Ca21]was determined by liquid scintillation counting. A, non-exchangeablepools, as measured by [45Ca21] associated with cells after 40 min ofincubation. B, exchangeable pools, as measured by [45Ca21] releasedfrom cells after 40 min of incubation. C, percentage of cell-associated[45Ca21] that remained at the indicated times.
Altered Calcium Homeostasis in a Phosphoglucomutase Mutant 5435
possibilities, the cells were inoculated onto YP plates contain-ing either lactate or lactate and galactose as carbon source(s).We found that each of the mutant strains (pgm2D, pmr1D, andpgm2D/pmr1D) and the WT strain were able to grow whenlactate alone was provided as sole carbon source (Fig. 4C).When both galactose and lactate were present, the WT andeach of the single mutants were again able to grow. In contrast,the pgm2D/pmr1D double mutant was unable to grow underthese conditions (Fig. 4D), indicating that the metabolism ofgalactose disrupted cellular physiology (presumably Ca21 ho-meostasis) to the extent that growth could not occur eventhrough the utilization of lactate as carbon source.
The pgm2D Strain Is Sensitive to Cyclosporin A When Grownin Galactose-containing Media—Under normal conditions, thecytosolic Ca21 concentration in yeast is maintained within the50–200 nM range (2, 3). Since we observed an elevated level oftotal Ca21 and exchangeable Ca21 in the pgm2D strain whengrown in galactose-containing media, we next asked whetherthe signaling pathway mediated by calcineurin and calmodulinwas activated. To test this possibility, we asked whether eachof these strains could grow on YPGal medium in the presence of10 mg/ml cyclosporin A (CsA), an inhibitor of calcineurin. Wefound that growth of the WT and pmr1D strains was unaffectedby CsA, while growth of the pgm2D strain was completelyinhibited (Fig. 5). These results suggests that the pgm2D mu-tation may cause an imbalance in cellular Ca21 homeostasis tothe extent that activation of the calcineurin pathway is re-quired to maintain growth of the pgm2D strain in galactose-containing media.
The pgm2D Mutation Leads to Increased Sequestration ofCytosolic Ca21 into Intracellular Compartments—In a previousstudy, it was shown that the re-addition of glucose to glucose-starved cells led to a transient increase in cytosolic Ca21 thatwas dependent upon the presence of extracellular Ca21 (3).Since our CsA experiment indicated that the pgm2D strain
required constitutive calcineurin activity to grow in galactosemedia, we next asked whether this state of calcineurin activa-tion could reduce or abolish the increase in cytosolic Ca21 levelspreviously shown to accompany the addition of glucose to glu-cose-starved cells. To make these measurements, we utilizedan aequorin reporter system (23, 46) that recently proved use-ful in measuring transient changes in the cytosolic Ca21 con-centration of yeast (4, 5). An aequorin expression plasmid wastransformed into the WT and pgm2D strains, and cultures ofeach strain were grown in SMGal medium to mid-log phase.The cells were then harvested and resuspended in SM mediumlacking a carbon source for 2 h. We found that the restingconcentration of cytosolic Ca21 in the WT strain was ;100 nM.When glucose was injected into the chamber to a final concen-tration of 1%, we found that the cytosolic Ca21 began to riseafter a lag of 40–60 s and reached a peak of ;340 nM within2–3 min (Fig. 6A). The Ca21 level then dropped over the next 5min until it reached a basal cytosolic Ca21 concentration sim-ilar to the level measured before glucose addition.
When we repeated this protocol with the pgm2D strain, wefound that the unstimulated cytosolic Ca21 concentration aftercarbon source depletion was ;90 nM, slightly lower than theWT strain. However, when glucose was added to the carbonsource-starved pgm2D strain, no increase in cytosolic Ca21 wasobserved. To confirm that this absence of glucose-stimulatedCa21 uptake was not due to the fact that the strains weregrown in SMGal medium, we repeated the experiment andexamined the effect of galactose addition following the starva-tion period. With the WT strain, we measured a basal cytosolicCa21 concentration of ;60 nM. When galactose was added, weagain observed a 40–60-s delay, followed by a rapid increase incytosolic Ca21 that reached a peak of ;160 nM. With the pgm2Dstrain, we observed a resting cytosolic Ca21 concentration of;50 nM (again, slightly lower than the WT strain). When ga-lactose was added to the starved pgm2D cells, we observed a
FIG. 4. The pgm2D/pmr1D strain isunable to grow in galactose-contain-ing medium. The strains indicated wereplated on YPGal (A), YPGal plus 100 mM
CaCl2 (B), YPLactate (C), and YPGal pluslactate (D). The plates were incubated at30 °C for 5–7 days.
Altered Calcium Homeostasis in a Phosphoglucomutase Mutant5436
very slight increase in cytosolic Ca21 to ;60 nM, but the re-sponse was still much smaller than was observed in the WTstrain (Fig. 6B).
The results of the above experiments indicate that thepgm2D strain is either unable to carry out Ca21 uptake fromthe extracellular environment under these conditions, or itremoves Ca21 from its cytosol so rapidly that the cytosolic Ca21
concentration cannot increase to the level observed in the WTstrain. To distinguish between these possibilities, we used asimilar experimental protocol to determine whether [45Ca21]uptake was altered in the pgm2D strain. Cells were grown inSMGal medium, harvested, and resuspended in SM lacking acarbon source for 2 h. The rate of [45Ca21] uptake was thenmeasured upon the re-addition of glucose to the medium. Wefound that both strains had similar rates of [45Ca21] uptake(Fig. 6C), indicating that the pgm2D strain was not signifi-cantly different from the WT strain in its ability to take upCa21 following carbon source starvation. We conclude that thetransient rise in cytosolic Ca21 was not observed in the pgm2Dstrain upon glucose addition because the entering Ca21 wasrapidly sequestered into intracellular compartments. This in-creased rate of Ca21 sequestration is presumably mediated byCa21 transporters activated by calcineurin in response to theelevated influx of Ca21 in the pgm2D stain.
The pgm2D Mutation Does Not Affect Ca21-dependent ERand Golgi Functions—The PMR1 gene encodes a Ca21-ATPaselocalized to the Golgi membrane (12). Although a direct meas-urement of the Golgi Ca21 level has not been made in thepmr1D mutant, the size of its exchangeable Ca21 pool is re-duced in a manner that is consistent with reduced Golgi Ca21
levels. Furthermore, the pmr1D mutation results in incompleteouter chain glycosylation of invertase, a process that is carried
out by enzymes in the Golgi compartment (12, 13). Recently, itwas shown that deletion of the PMR1 gene also results in adelayed degradation of CpY*, a mutant form of the vacuolarprotein carboxypeptidase Y (CpY) that is unable to acquire aproperly folded conformation upon translocation across the ERmembrane (33). As a consequence, it is recognized by the qual-ity control apparatus within the ER and rapidly transportedback out of the ER to proteosomes for degradation (47, 48). Thepgm2D mutant shares some alterations in Ca21 homeostasiswith the pmr1D strain, such as an elevated total cellular Ca21
level and increased Ca21 uptake. On the other hand, thepgm2D mutant (unlike the pmr1D mutant) retains a normaldistribution of Ca21 between the exchangeable and non-ex-changeable pools.
To determine whether the pgm2D mutation also affects theretrograde transport of misfolded proteins from the ER like thepmr1D mutation, we determined the rate of CpY* degradation
FIG. 5. Growth of the pgm2D strain is inhibited by CsA. Strainswere streaked onto YPGal (A) and YPGal supplemented with 10 mg/mlCsA (B).
FIG. 6. The pgm2D strain is defective in glucose-induced Ca21
signaling. The carbon source re-added was 1% glucose (A) or 1%galactose (B). Yeast cells of the wild-type or the pgm2D strain, whichwere transformed previously with apoaequorin-containing plasmid,were grown in SMGal, loaded with coelenterazine, and starved forcarbon source for 2 h. The indicated carbon source was then added (asindicated by the arrow). The rate of [45Ca21] uptake following theaddition of 1% glucose is shown in C.
Altered Calcium Homeostasis in a Phosphoglucomutase Mutant 5437
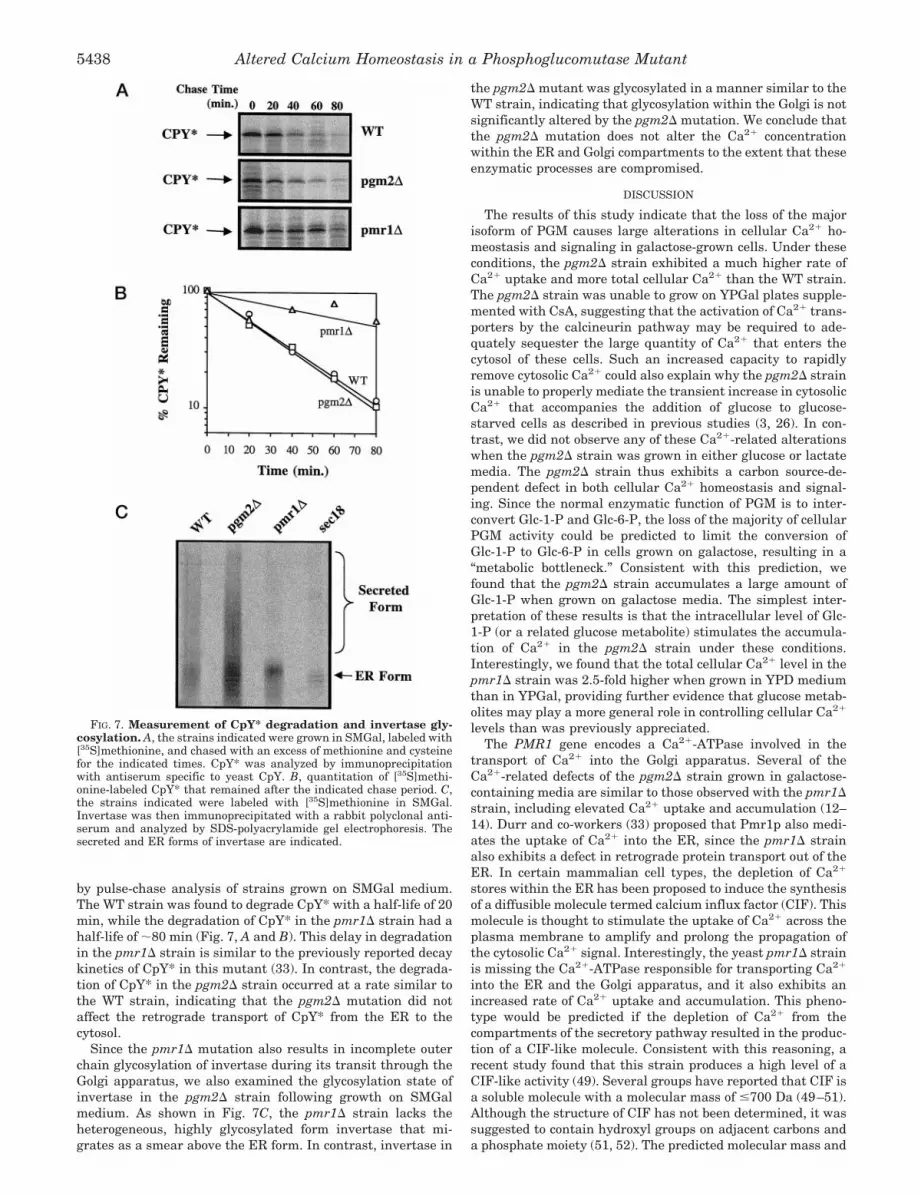
by pulse-chase analysis of strains grown on SMGal medium.The WT strain was found to degrade CpY* with a half-life of 20min, while the degradation of CpY* in the pmr1D strain had ahalf-life of ;80 min (Fig. 7, A and B). This delay in degradationin the pmr1D strain is similar to the previously reported decaykinetics of CpY* in this mutant (33). In contrast, the degrada-tion of CpY* in the pgm2D strain occurred at a rate similar tothe WT strain, indicating that the pgm2D mutation did notaffect the retrograde transport of CpY* from the ER to thecytosol.
Since the pmr1D mutation also results in incomplete outerchain glycosylation of invertase during its transit through theGolgi apparatus, we also examined the glycosylation state ofinvertase in the pgm2D strain following growth on SMGalmedium. As shown in Fig. 7C, the pmr1D strain lacks theheterogeneous, highly glycosylated form invertase that mi-grates as a smear above the ER form. In contrast, invertase in
the pgm2D mutant was glycosylated in a manner similar to theWT strain, indicating that glycosylation within the Golgi is notsignificantly altered by the pgm2D mutation. We conclude thatthe pgm2D mutation does not alter the Ca21 concentrationwithin the ER and Golgi compartments to the extent that theseenzymatic processes are compromised.
DISCUSSION
The results of this study indicate that the loss of the majorisoform of PGM causes large alterations in cellular Ca21 ho-meostasis and signaling in galactose-grown cells. Under theseconditions, the pgm2D strain exhibited a much higher rate ofCa21 uptake and more total cellular Ca21 than the WT strain.The pgm2D strain was unable to grow on YPGal plates supple-mented with CsA, suggesting that the activation of Ca21 trans-porters by the calcineurin pathway may be required to ade-quately sequester the large quantity of Ca21 that enters thecytosol of these cells. Such an increased capacity to rapidlyremove cytosolic Ca21 could also explain why the pgm2D strainis unable to properly mediate the transient increase in cytosolicCa21 that accompanies the addition of glucose to glucose-starved cells as described in previous studies (3, 26). In con-trast, we did not observe any of these Ca21-related alterationswhen the pgm2D strain was grown in either glucose or lactatemedia. The pgm2D strain thus exhibits a carbon source-de-pendent defect in both cellular Ca21 homeostasis and signal-ing. Since the normal enzymatic function of PGM is to inter-convert Glc-1-P and Glc-6-P, the loss of the majority of cellularPGM activity could be predicted to limit the conversion ofGlc-1-P to Glc-6-P in cells grown on galactose, resulting in a“metabolic bottleneck.” Consistent with this prediction, wefound that the pgm2D strain accumulates a large amount ofGlc-1-P when grown on galactose media. The simplest inter-pretation of these results is that the intracellular level of Glc-1-P (or a related glucose metabolite) stimulates the accumula-tion of Ca21 in the pgm2D strain under these conditions.Interestingly, we found that the total cellular Ca21 level in thepmr1D strain was 2.5-fold higher when grown in YPD mediumthan in YPGal, providing further evidence that glucose metab-olites may play a more general role in controlling cellular Ca21
levels than was previously appreciated.The PMR1 gene encodes a Ca21-ATPase involved in the
transport of Ca21 into the Golgi apparatus. Several of theCa21-related defects of the pgm2D strain grown in galactose-containing media are similar to those observed with the pmr1Dstrain, including elevated Ca21 uptake and accumulation (12–14). Durr and co-workers (33) proposed that Pmr1p also medi-ates the uptake of Ca21 into the ER, since the pmr1D strainalso exhibits a defect in retrograde protein transport out of theER. In certain mammalian cell types, the depletion of Ca21
stores within the ER has been proposed to induce the synthesisof a diffusible molecule termed calcium influx factor (CIF). Thismolecule is thought to stimulate the uptake of Ca21 across theplasma membrane to amplify and prolong the propagation ofthe cytosolic Ca21 signal. Interestingly, the yeast pmr1D strainis missing the Ca21-ATPase responsible for transporting Ca21
into the ER and the Golgi apparatus, and it also exhibits anincreased rate of Ca21 uptake and accumulation. This pheno-type would be predicted if the depletion of Ca21 from thecompartments of the secretory pathway resulted in the produc-tion of a CIF-like molecule. Consistent with this reasoning, arecent study found that this strain produces a high level of aCIF-like activity (49). Several groups have reported that CIF isa soluble molecule with a molecular mass of #700 Da (49–51).Although the structure of CIF has not been determined, it wassuggested to contain hydroxyl groups on adjacent carbons anda phosphate moiety (51, 52). The predicted molecular mass and
FIG. 7. Measurement of CpY* degradation and invertase gly-cosylation. A, the strains indicated were grown in SMGal, labeled with[35S]methionine, and chased with an excess of methionine and cysteinefor the indicated times. CpY* was analyzed by immunoprecipitationwith antiserum specific to yeast CpY. B, quantitation of [35S]methi-onine-labeled CpY* that remained after the indicated chase period. C,the strains indicated were labeled with [35S]methionine in SMGal.Invertase was then immunoprecipitated with a rabbit polyclonal anti-serum and analyzed by SDS-polyacrylamide gel electrophoresis. Thesecreted and ER forms of invertase are indicated.
Altered Calcium Homeostasis in a Phosphoglucomutase Mutant5438

structural characteristics are consistent with the features ex-pected if CIF were related to (or derived from) a sugar phos-phate or sugar nucleotide (53). This raises the possibility thatGlc-1-P may be a metabolic precursor in the CIF biosyntheticpathway.
While the increased rate of Ca21 uptake and total cellularCa21 levels in the pgm2D strain could be attributable to a storedepletion mechanism that involves a CIF-like signaling mole-cule, other aspects of the phenotype of the pgm2D strain areinconsistent with such a model. First, our data indicate thatthe pgm2D strain, unlike the pmr1D strain, contains a fractionof total cellular Ca21 in the exchangeable pool that is at least aslarge (if not larger) than the exchangeable pool in the WTstrain. This suggests that the level of Ca21 contained in the ERand the Golgi apparatus is not significantly reduced. Consist-ent with this, we did not observe any evidence of reduced Ca21
levels within the ER or Golgi apparatus of the pgm2D strain infunctional assays that measured retrograde transport of CpY*from the ER, or invertase glycosylation in the Golgi apparatus.These results indicate that the elevated rate of Ca21 uptake inthe pgm2D strain is not coupled to a defect in filling the Ca21
stores within the ER and/or the Golgi apparatus. Instead, itappears that the pmr1D and pgm2D mutations induce theuptake of Ca21 in mechanistically distinct ways. Since thepgm2D strain accumulates a high level of Glc-1-P, we hypoth-esize that Glc-1-P accumulation may be coupled to elevatedCa21 uptake by some mechanism. It is possible that the accu-mulation of Glc-1-P may bypass the need to deplete ER or GolgiCa21 stores prior to increased Ca21 uptake. This could occur ifan increased concentration of Glc-1-P stimulates the synthesisof CIF in the absence of store depletion, or if Glc-1-P directlystimulates the rate of Ca21 accumulation into one or moreintracellular compartments (such as the ER, Golgi apparatus,or vacuole). This latter possibility would not be unprecedented,since it has been shown that Glc-6-P can stimulate ATP-de-pendent Ca21 uptake into isolated mammalian microsomes(27, 28). The existence of a Glc-6-P transporter in mammalianER membranes is well established. While Glc-6-P transporteractivity in the ER is coupled to Glc-6-P phosphatase activity inthe ER lumen of gluconeogenic tissues such as liver, the muchbroader tissue distribution of the Glc-6-P transporter suggeststhat it may also participate in another general cellular processsuch as Ca21 homeostasis (54). Consistent with this, it hasbeen shown that neutrophils and monocytes from patients withGlc-6-P transporter deficiency are unable to efficiently seques-ter Ca21 (55). As would be predicted from our model thatGlc-1-P can also function in this way, it was recently found thatGlc-1-P can also stimulate the ATP-dependent uptake of Ca21
into isolated cardiac microsomes.2
To better understand how Pgm2p and Pmr1p each influencecellular Ca21 homeostasis, we constructed a pgm2D/pmr1Dstrain. To our surprise, the pgm2D/pmr1D double mutant wasunable to grow on YPGal medium, even when an alternatecarbon source (lactate) was present. This suggests that themetabolism of galactose in the pgm2D/pmr1D double mutantmay cause such a severe imbalance in ion homeostasis that cellgrowth is inhibited. This finding again suggests that the pmr1Dand pgm2D mutations may increase Ca21 uptake in distinctways, since the phenotypic consequences of combining thesetwo mutations were more severe than the phenotype of strainscarrying either mutation alone. Although our results indicatethat the free Ca21 level within the ER and the Golgi apparatusis normal in the pgm2D strain, we cannot exclude the possibil-ity that the elevated Ca21 uptake associated with the pgm2D
mutation is caused by the inability to fill another intracellularcompartment (or subcompartment) with Ca21. If this were thecase, the combination of the pgm2D and pmr1D mutationscould further compromise the ability to adequately fill theeffected compartments when grown in standard YPGal me-dium (which contains only 0.3 mM Ca21). Our finding that theaddition of 100 mM CaCl2 can restore the growth of the pgm2D/pmr1D strain on this growth medium is consistent with such amodel. Clearly, further studies are required to determine themechanism that couples glucose metabolism to Ca21 homeo-stasis. The elucidation of this process may provide importantnew insights into the control of Ca21 homeostasis in yeast.Ultimately, it may also lead to a better understanding of hu-man diseases such as diabetes that are also caused by defectsin glucose metabolism.
Acknowledgments—We thank Dr. James Hopper, Dr. Hans K. Ru-dolph, and Dr. Todd R. Graham for providing strains and plasmids; Dr.Scott Emr for antiserum; and Dr. Richard Kellermayer for helpfuldiscussions and for critically reading the manuscript.
REFERENCES
1. Cunningham, K. W., and Fink, G. R. (1994) J. Exp. Biol. 196, 157–1662. Iida, H., Yagawa, Y., and Anraku, Y. (1990) J. Biol. Chem. 265, 13391–133993. Nakajima-Shimada, J., Iida, H., Tsuji, F. I., and Anraku, Y. (1991) Proc. Natl.
Acad. Sci. U. S. A. 88, 6878–68824. Miseta, A., Fu, L., Kellermayer, R., Buckley, J., and Bedwell, D. M. (1999)
J. Biol. Chem. 274, 5939–59475. Miseta, A., Kellermayer, R., Aiello, D. P., Fu, L., and Bedwell, D. M. (1999)
FEBS Lett. 451, 132–1366. Williams, R. J. (1990) Adv. Exp. Med. Biol. 269, 7–167. Putney, J. W., Jr., and Bird, G. S. J. (1993) Cell 75, 199–2018. Halachmi, D., and Eilam, Y. (1989) FEBS Lett. 256, 55–619. Eilam, Y., Lavi, H., and Grossowicz, N. (1985) J. Gen. Microbiol. 131, 623–629
10. Ohsumi, Y., Kitamoto, K., and Anraku, Y. (1988) J. Bacteriol. 170, 2676–268211. Cunningham, K. W., and Fink, G. R. (1994) J. Cell Biol. 124, 351–36312. Antebi, A., and Fink, G. R. (1992) Mol. Biol. Cell 3, 633–65413. Rudolph, H. K., Antebi, A., Fink, G. R., Buckley, C. M., Dorman, T. E., LeVitre,
J., Davidow, L. S., Mao, J. I., and Moir, D. T. (1989) Cell 58, 133–14514. Halachmi, D., and Eilam, Y. (1996) FEBS Lett. 392, 194–20015. Sorin, A., Rosas, G., and Rao, R. (1997) J. Biol. Chem. 272, 9895–990116. Fu, D., Beeler, T., and Dunn, T. (1994) Yeast 10, 515–52117. Pozos, T. C., Sekler, I., and Cyert, M. S. (1996) Mol. Cell. Biol. 16, 3730–374118. Lapinskas, P. J., Lin, S. J., and Culotta, V. C. (1996) Mol. Microbiol. 21,
519–52819. Cunningham, K. W., and Fink, G. R. (1994) J. Cell Biol. 124, 351–36320. Cunningham, K. W., and Fink, G. R. (1996) Mol. Cell. Biol. 16, 2226–223721. Pinton, P., Brini, M., Bastianutto, C., Tuft, R. A., Pozzan, T., and Rizzuto, R.
(1998) Biofactors 8, 243–25322. Uribe, S., Rangel, P., and Pardo, J. P. (1992) Cell Calcium 13, 211–21723. Batiza, A. F., Schulz, T., and Masson, P. H. (1996) J. Biol. Chem. 271,
23357–2336224. Thevelein, J. M., and Hohmann, S. (1995) Trends Biochem. Sci. 20, 3–1025. Eilam, Y., and Othman, M. (1990) J. Gen. Microbiol. 136, 861–86626. Eilam, Y., Othman, M., and Halachmi, D. (1990) J. Gen. Microbiol. 136,
2537–254327. Benedetti, A., Fulceri, R., Romani, A., and Comporti, M. (1988) J. Biol. Chem.
263, 3466–347328. Chen, P. Y., Csutora, P., Veyna-Burke, N. A., and Marchase, R. B. (1998)
Diabetes 47, 874–88129. Fu, L., Bounelis, P., Dey, N., Browne, B. L., Marchase, R. B., and Bedwell,
D. M. (1995) J. Bacteriol. 177, 3087–309430. Oh, D., and Hopper, J. E. (1990) Mol. Cell. Biol. 10, 3373–337731. Boles, E., Liebetrau, W., Hofmann, M., and Zimmermann, F. K. (1994) Eur.
J. Biochem. 220, 83–9632. Hofmann, M., Boles, E., and Zimmermann, F. K. (1994) Eur. J. Biochem. 221,
741–74733. Durr, G., Strayle, J., Plemper, R., Elbs, S., Klee, S. K., Catty, P., Wolf, D. H.,
and Rudolph, H. K. (1998) Mol. Biol. Cell 9, 1149–116234. Rothstein, R. J. (1983) Methods Enzymol. 101, 202–20935. Jones, J. S., and Prakash, L. (1990) Yeast 6, 363–36636. Harmsen, M. M., Langedijk, A. C., van Tuinen, E., Geerse, R. H., Raue, H. A.,
and Maat, J. (1993) Gene (Amst) 125, 115–12337. Rose, M. D., Winston, F., and Hieter, P. (1990) Methods in Yeast Genetics: A
Laboratory Course Manual, Cold Spring Harbor Laboratory Press, ColdSpring Harbor, NY
38. Miller, J. H. (1992) A Short Course in Bacterial Genetics., Cold Spring HarborLaboratory Press, Cold Spring Harbor, NY
39. Bradford, M. M. (1976) Anal. Biochem. 72, 248–25440. Saez, M. J., and Lagunas, R. (1976) Mol. Cell. Biochem. 13, 73–7841. Bergmeyer, H. U., Bergmeyer, J., and Grassl, M. (1984) Methods of Enzymatic
analysis, 3rd Ed., Vol. 6, pp. 163–199, Verlag Chemie, Weinheim, Germany42. Bedwell, D. M., Klionsky, D. J., and Emr, S. D. (1987) Mol. Cell. Biol. 7,
4038–404743. Carlson, M., and Botstein, D. (1982) Cell 28, 145–15444. Eilam, Y., and Chernichovsky, D. (1987) J. Gen. Microbiol. 133, 1641–16492 R. Marchase, unpublished results.
Altered Calcium Homeostasis in a Phosphoglucomutase Mutant 5439

45. Dunn, T., Gable, K., and Beeler, T. (1994) J. Biol. Chem. 269, 7273–727846. Allen, D. G., Blinks, J. R., and Prendergast, F. G. (1977) Science 195, 996–99847. Finger, A., Knop, M., and Wolf, D. H. (1993) Eur. J. Biochem. 218, 565–57448. Hiller, M. M., Finger, A., Schweiger, M., and Wolf, D. H. (1996) Science 273,
1725–172849. Csutora, P., Su, Z., Kim, H. Y., Bugrim, A., Cunningham, K. W., Nuccitelli, R.,
Keizer, J. E., Hanley, M. R., Blalock, J. E., and Marchase, R. B. (1999) Proc.Natl. Acad. Sci. U. S. A. 96, 121–126
50. Thomas, D., and Hanley, M. R. (1995) J. Biol. Chem. 270, 6429–6432
51. Randriamampita, C., and Tsien, R. Y. (1993) Nature 364, 809–81452. Randriamampita, C., and Tsien, R. Y. (1995) J. Biol. Chem. 270, 29–3253. Kim, H. Y., Thomas, D., and Hanley, M. R. (1995) J. Biol. Chem. 270,
9706–970854. Hiraiwa, H., Pan, C. J., Lin, B., Moses, S. W., and Chou, J. Y. (1999) J. Biol.
Chem. 274, 5532–553655. Kilpatrick, L., Garty, B. Z., Lundquist, K. F., Hunter, K., Stanley, C. A.,
Baker, L., Douglas, S. D., and Korchak, H. M. (1990) J. Clin. Invest. 86,196–202
Altered Calcium Homeostasis in a Phosphoglucomutase Mutant5440
Copyright © 2022 FDOKUMEN




















