
Looking beyond sugars: the role of amphiphilic solutes in preventing adventitious reactions in...
11
Comparative Biochemistry and Physiology Part A 131 (2002) 515–525 1095-6433/02/$ - see front matter 2002 Elsevier Science Inc. All rights reserved. PII: S1095-6433 Ž 01 . 00514-1 Looking beyond sugars: the role of amphiphilic solutes in preventing adventitious reactions in anhydrobiotes at low water contents Ann E. Oliver *, Dirk K. Hincha , John H. Crowe a, b a Section of Molecular and Cellular Biology, University of California, Davis, CA, USA a Max-Planck Institute for Molecular Plant Physiology, D-14424 Potsdam, Germany b Received 12 January 2001; received in revised form 14 June 2001; accepted 26 June 2001 Abstract Plants and animals that can survive dehydration accumulate high concentrations of disaccharides in their cells and tissues during desiccation. These sugars are necessary both for the depression of the membrane phase transition temperature of the dry lipid and for the formation of a carbohydrate glass. In the past decade, however, it has become clear that certain types of adventitious enzymatic reactions are possible at low water contents, which along with free- radical mediated damage, can cause hydrolysis of lipids and loss of membrane barrier function. Disaccharides do not necessarily prevent these types of reactions, which suggests that other compounds might also be necessary for protecting organisms from this type of degradation during anhydrobiosis. Arbutin, one possible example, accumulates in large quantities in certain resurrection plants and has been shown to inhibit phospholipase A activity at low water contents. 2 The direct effect of arbutin on membranes under stress conditions depends on the membrane lipid composition. It can serve a protective function during desiccation- or freeze y thaw-induced stress in the presence of nonbilayer-forming lipids or a disruptive function in their absence. Other possible amphiphiles, including certain naturally occurring flavonols, may serve as anti-oxidants and some might have similar lipid composition-dependent effects. Such compounds, therefore, are likely to be localized near specific membranes, where they might provide the greatest benefit at the least liability to the organism. 2002 Elsevier Science Inc. All rights reserved. Keywords: Anhydrobiosis; Membrane phase state; Disaccharides; Carbohydrate glasses; Amphiphiles; Phospholipase A ; Membrane 2 hydrolysis Abbreviations: EPC: egg phosphatidylcholine; EPE: egg phosphatidylethanolamine; FFA: free fatty acid; HES: hydroxyethyl starch; LUV: large unilamellar vesicles; MGDG: monogalactosyldiacylglycerol; T : membrane phase transition temperature; T : carbohydrate m g glass transition temperature; PLA : phospholipase A ; PLC: phospholipase C; RH: relative humidity 2 2 This paper was originally presented at a symposium on extremophiles held within the ESCPB 21st International Congress, Liege, ` Belgium, July 24–28, 2000. * Corresponding Author. Tel.: q1-530-752-1094; fax: q1-530-752-5305. E-mail address: [email protected] (A.E. Oliver).
-
Upload
independent -
Category
Documents
-
view
6 -
download
0
Transcript of Looking beyond sugars: the role of amphiphilic solutes in preventing adventitious reactions in...

Comparative Biochemistry and Physiology Part A 131(2002) 515–525
1095-6433/02/$ - see front matter� 2002 Elsevier Science Inc. All rights reserved.PII: S1095-6433Ž01.00514-1
Looking beyond sugars: the role of amphiphilic solutes inpreventing adventitious reactions in anhydrobiotes at low water
contents�
Ann E. Oliver *, Dirk K. Hincha , John H. Crowea, b a
Section of Molecular and Cellular Biology, University of California, Davis, CA, USAa
Max-Planck Institute for Molecular Plant Physiology, D-14424 Potsdam, Germanyb
Received 12 January 2001; received in revised form 14 June 2001; accepted 26 June 2001
Abstract
Plants and animals that can survive dehydration accumulate high concentrations of disaccharides in their cells andtissues during desiccation. These sugars are necessary both for the depression of the membrane phase transitiontemperature of the dry lipid and for the formation of a carbohydrate glass. In the past decade, however, it has becomeclear that certain types of adventitious enzymatic reactions are possible at low water contents, which along with free-radical mediated damage, can cause hydrolysis of lipids and loss of membrane barrier function. Disaccharides do notnecessarily prevent these types of reactions, which suggests that other compounds might also be necessary for protectingorganisms from this type of degradation during anhydrobiosis. Arbutin, one possible example, accumulates in largequantities in certain resurrection plants and has been shown to inhibit phospholipase A activity at low water contents.2
The direct effect of arbutin on membranes under stress conditions depends on the membrane lipid composition. It canserve a protective function during desiccation- or freezeythaw-induced stress in the presence of nonbilayer-forming lipidsor a disruptive function in their absence. Other possible amphiphiles, including certain naturally occurring flavonols,may serve as anti-oxidants and some might have similar lipid composition-dependent effects. Such compounds, therefore,are likely to be localized near specific membranes, where they might provide the greatest benefit at the least liability tothe organism.� 2002 Elsevier Science Inc. All rights reserved.
Keywords: Anhydrobiosis; Membrane phase state; Disaccharides; Carbohydrate glasses; Amphiphiles; Phospholipase A ; Membrane2
hydrolysis
Abbreviations: EPC: egg phosphatidylcholine; EPE: egg phosphatidylethanolamine; FFA: free fatty acid; HES: hydroxyethyl starch;LUV: large unilamellar vesicles; MGDG: monogalactosyldiacylglycerol;T : membrane phase transition temperature;T : carbohydratem g
glass transition temperature; PLA : phospholipase A ; PLC: phospholipase C; RH: relative humidity2 2� This paper was originally presented at a symposium on extremophiles held within the ESCPB 21st International Congress, Liege,`
Belgium, July 24–28, 2000.* Corresponding Author. Tel.:q1-530-752-1094; fax:q1-530-752-5305.E-mail address: [email protected](A.E. Oliver).

516 A.E. Oliver et al. / Comparative Biochemistry and Physiology Part A 131 (2002) 515–525
1. Anhydrobiosis and the importance ofdisaccharides
Several plant and animal species have evolvedthe ability to survive almost complete dehydration,often for prolonged periods. This life withoutwater, or anhydrobiosis, is dependent on a coor-dinated and controlled set of biochemical adapta-tions. Only recently, has the complexity of theoverall response to dehydration begun to be fullyappreciated(Ingram and Bartels, 1996). Severalmechanisms probably participate in protection ofliving cells from the removal of water. Theseinclude but are not limited to:(1) the downregulation of metabolism and removal of reactiveoxygen species and the free radicals they produce(Leprince et al., 1990, 2000); (2) the accumulationof certain proteins, such as the late embryogenesisabundant(LEA) (for review, see Cuming, 1999)or small heat shock proteins(sHSPs) (Almogueraand Jordano, 1992; Wehmeyer and Vierling, 2000),which appear to play some role in desiccationresistance; and(3) the maintenance of macromol-ecules in their biologically relevant structuresthrough the accumulation of disaccharides andother glass-forming compounds(Crowe et al.,1997a; Buitink et al., 1999). As an exhaustivereview of the above mechanisms is beyond thescope of this article, we will focus primarily onsmall solutes and their possible role in maintaininganhydrobiosis.
As mentioned, one important adaptation is theaccumulation of disaccharides in the cells andtissues of the organism during drying. In general,anhydrobiotic animals, such as tardigrades, nema-todes and yeast cells accumulate trehalose(Westhand Ramlov, 1991; Womersly, 1990; Crowe et al.,1984, 1997a), whereas the dehydration resistanttissues of higher plants,(i.e. seeds and pollengrains) accumulate sucrose(Hoekstra et al., 1989;Koster and Leopold, 1988; Hoekstra, 1986). Thereare exceptions, however, as some resurrectionplants, such asMyrothamnus flabellifolia accu-mulate a combination of sucrose and trehalose(Bianchi et al., 1993; Drennan et al., 1993).
According to one model, the disaccharides fulfilltwo important roles in protecting the membranesof the dry organism: reduction of the drymembrane phase transition temperature(T ) andm
formation of a carbohydrate glass with a highmelting temperature. Both of these are necessary,
but neither is sufficient by itself for full protectionof membranes during drying and rehydration.
When unprotected membranes are dehydrated,the water molecules that help to maintain spacingbetween the phospholipid headgroups are removed,allowing a closer approach of the lipid moleculesand an increase in the membraneT by as muchm
as 708C. This elevation of the dry membraneTm
is especially important for membranes that have asub-ambientT under hydrated conditions, as ism
the case for most biological membranes. In thesecircumstances, theT shifts during dehydration,m
which causes a phase change from liquid crystal-line to gel phase lipid. Subsequently, as water isadded back during rehydration,T shifts back tom
the hydrated value and the membrane undergoesanother phase transition, resulting in transient leak-age of soluble cell contents through the membrane(Leslie et al., 1994; Crowe et al., 1997b).
In contrast, when membranes are dried in thepresence of trehalose or sucrose, the water replace-ment model suggests that the sugar moleculesprevent the close approach of lipids through aninteraction with the phospholipid headgroups,thereby preventing the elevation ofT duringm
drying (Crowe and Crowe, 1988b; Leslie et al.,1995). As water is added back to such a system,no phase transition occurs and the membraneretains its barrier function(Harrigan et al., 1990;Hoekstra et al., 1992; Crowe et al., 1994). Fouriertransform infrared spectroscopy(FTIR) and nucle-ar magnetic resonance spectroscopy(NMR) databoth provide evidence in support of this directinteraction, most likely by hydrogen bonding,between the sugars and the phospholipid head-groups(Tsvetkova et al., 1998).
The second important protective function pro-vided by disaccharides, as well as other carbohy-drates and proteins, is vitrification at low watercontents. The carbohydrate glass is a solid-like,amorphous material that prevents many diffusionlimited deterioration processes, such as membranefusion. Vitrification is a common feature of car-bohydrates, but the temperature at which the glassdevitrifies (T ) depends on many factors. In gen-g
eral,T increases with increasing molecular weightg
and decreasing water content(Slade and Levine,1991). The presence of the carbohydrate glass isnecessary during storage in the dry state, becausestorage above theT of a system,(i.e. devitrifica-g
tion) can lead to fusion, membrane leakage(Sunet al., 1996) and loss of cell viability(Sun and

517A.E. Oliver et al. / Comparative Biochemistry and Physiology Part A 131 (2002) 515–525
Leopold, 1994, 1997; Buitink et al., 1998). Inter-estingly, recent evidence indicates that the LEAproteins, which accumulate in some plant tissuesduring drying, might also participate in glass for-mation and improve glass stability(Wolkers et al.,1999, 2001).
There has been some discussion in the literature(Koster et al., 1994, 2000; Crowe et al., 1994,1996, 1997b) as to whether carbohydrate vitrifi-cation is not only necessary but sufficient forpreservation of dry membranes. This suggestion isbased on the model of Wolfe and Bryant, anelegant analysis of the physical forces involved indehydrating membranes, especially in multilamel-lar vesicles (MLVs) (Bryant and Wolfe, 1992;Wolfe and Bryant, 1999). This theory explains theeffect of sugars on the phase behavior of mem-branes during dehydration, using non-specificosmotic and volumetric effects(Koster et al.,2000). This theory can explain most aspects of thedehydrating system and it is likely that bothmodels can contribute to our overall understandingof membranes under conditions of water deficit.There remains one piece of experimental data,however, that we find difficult to explain entirelyby non-specific effects. If vitrification were suffi-cient for full protection, the carbohydrate polymersHES and dextran should be excellent protectiveexcipients due to their highT values. The poly-g
mers, however, do not depress the dry membraneT and do not protect vesicles from leaking theirm
contents due to drying and rehydration(Koster etal., 2000; Crowe et al., 1994, 1997b). The expla-nation forwarded for this lack of protection is thatthe polymers are likely to be excluded from inter-lamellar spaces due to their size(Wolfe andBryant, 1999; Koster et al., 2000). It is difficultto see how this could explain their failure toprotect unilamellar vesicles(ULVs), however,especially in light of the finding that the polymersdo prevent fusion in the ULV samples(Crowe etal., 1996, 1997b). In order to prevent fusion, thepolymers must be able to prevent close approachof the ULV membranes,(i.e. separate one vesiclefrom another), and thus their sequestration into abulk phase and their exclusion from the spacesvery near to the membranes are difficult to rec-oncile with the experimental data.
In our opinion, therefore, the failure of mostcarbohydrate polymers to prevent membrane leak-age during drying and rehydration most likelyresults from the inability of the polymers to interact
directly with the membrane headgroups(Crowe etal., 1994, 1997b; Tsvetkova, et al., 1998) and drivedown the dry membraneT . Conversely, glucose,m
which can interact directly with the lipid head-groups, but has a lowT value and thus devitrifiesg
at relatively low temperatures, cannot preventfusion caused during dehydration(Crowe et al.,1997b). But while neither glucose nor HES alonecan protect vesicles during drying, a 1:1 combi-nation of glucose and HES is able to both decreasethe dry membraneT and form a stable glass, thusm
inhibiting fusion and leakage(Crowe et al.,1997b). Both the direct interaction and the glassystate appear to be necessary, therefore, for fullprotection of membranes during drying and rehy-dration (Crowe et al., 1994, 1997b; Tsvetkova etal., 1998). This finding, then, accounts for theevolutionary success of disaccharides in anhydro-biotes. Disaccharides are able to fulfill both impor-tant functions: (1) interacting with the lipidheadgroups to depress the dry membraneT ; andm
(2) formation of a stable glass with a relativelyhigh glass transition temperature.
Interestingly, some plant polysaccharides, name-ly fructans (polyfructoses), are able to interactwith lipid headgroups in a similar manner todisaccharides. In the dry state, fructans can depressT and stabilize dry bilayers(Hincha et al., 2000).m
Fructans are also accumulated in many plant spe-cies under stress conditions and may play animportant role in plant desiccation tolerance(Pol-lock and Cairns, 1991; Pilon-Smits et al., 1995;Vijn and Smeekens, 1999).
2. The limitations of protection by sugars
Although the disaccharides are of great impor-tance in the protection of anhydrobiotic tissues andorganisms(for some interesting exceptions, seeHoekstra et al., 1992), there appears to be moreto the story. While the sugars can enable survivalof the initial drying and rehydration events, theyare not sufficient for survival during prolongedperiods in the dry state. Seed storage provides aninstructive example. Even with their full comple-ment of carbohydrates and even under optimalconditions of low temperature and moisture, seedviability decreases during storage following a neg-ative sigmoidal curve(Roberts, 1979; Priestly,1986).
This loss of viability is correlated with anaccumulation of free fatty acids(FFAs) in the cell
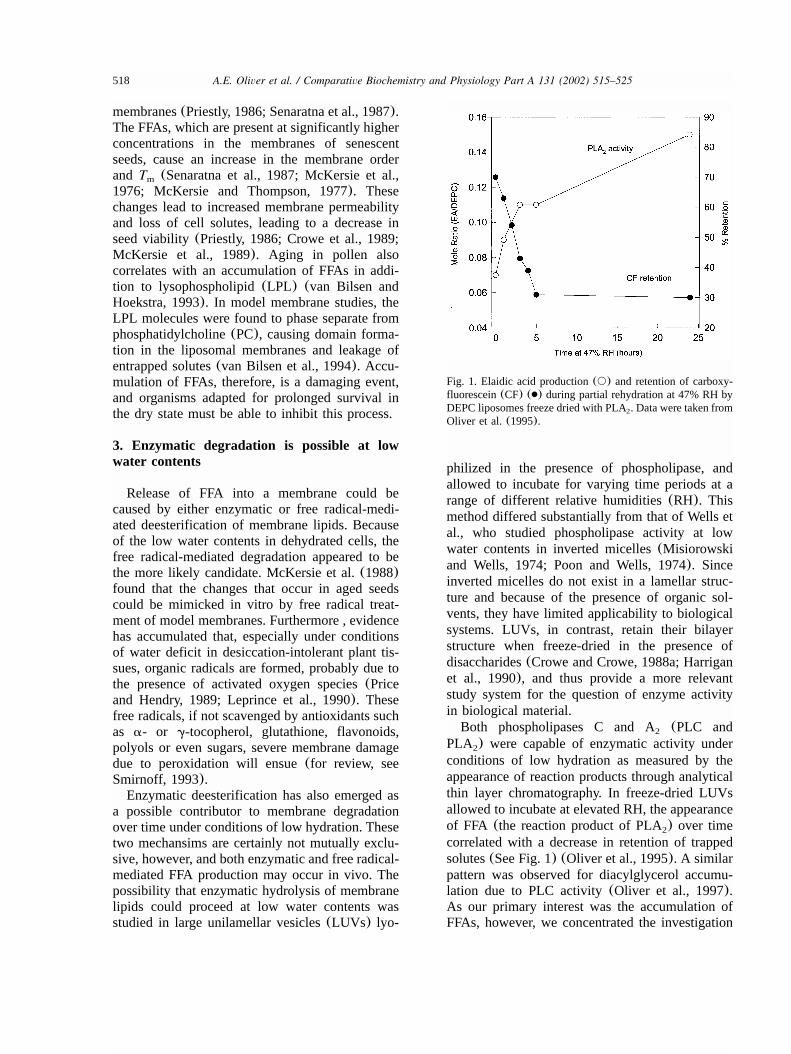
518 A.E. Oliver et al. / Comparative Biochemistry and Physiology Part A 131 (2002) 515–525
Fig. 1. Elaidic acid production(s) and retention of carboxy-fluorescein(CF) (●) during partial rehydration at 47% RH byDEPC liposomes freeze dried with PLA . Data were taken from2
Oliver et al.(1995).
membranes(Priestly, 1986; Senaratna et al., 1987).The FFAs, which are present at significantly higherconcentrations in the membranes of senescentseeds, cause an increase in the membrane orderand T (Senaratna et al., 1987; McKersie et al.,m
1976; McKersie and Thompson, 1977). Thesechanges lead to increased membrane permeabilityand loss of cell solutes, leading to a decrease inseed viability(Priestly, 1986; Crowe et al., 1989;McKersie et al., 1989). Aging in pollen alsocorrelates with an accumulation of FFAs in addi-tion to lysophospholipid(LPL) (van Bilsen andHoekstra, 1993). In model membrane studies, theLPL molecules were found to phase separate fromphosphatidylcholine(PC), causing domain forma-tion in the liposomal membranes and leakage ofentrapped solutes(van Bilsen et al., 1994). Accu-mulation of FFAs, therefore, is a damaging event,and organisms adapted for prolonged survival inthe dry state must be able to inhibit this process.
3. Enzymatic degradation is possible at lowwater contents
Release of FFA into a membrane could becaused by either enzymatic or free radical-medi-ated deesterification of membrane lipids. Becauseof the low water contents in dehydrated cells, thefree radical-mediated degradation appeared to bethe more likely candidate. McKersie et al.(1988)found that the changes that occur in aged seedscould be mimicked in vitro by free radical treat-ment of model membranes. Furthermore , evidencehas accumulated that, especially under conditionsof water deficit in desiccation-intolerant plant tis-sues, organic radicals are formed, probably due tothe presence of activated oxygen species(Priceand Hendry, 1989; Leprince et al., 1990). Thesefree radicals, if not scavenged by antioxidants suchas a- or g-tocopherol, glutathione, flavonoids,polyols or even sugars, severe membrane damagedue to peroxidation will ensue(for review, seeSmirnoff, 1993).
Enzymatic deesterification has also emerged asa possible contributor to membrane degradationover time under conditions of low hydration. Thesetwo mechansims are certainly not mutually exclu-sive, however, and both enzymatic and free radical-mediated FFA production may occur in vivo. Thepossibility that enzymatic hydrolysis of membranelipids could proceed at low water contents wasstudied in large unilamellar vesicles(LUVs) lyo-
philized in the presence of phospholipase, andallowed to incubate for varying time periods at arange of different relative humidities(RH). Thismethod differed substantially from that of Wells etal., who studied phospholipase activity at lowwater contents in inverted micelles(Misiorowskiand Wells, 1974; Poon and Wells, 1974). Sinceinverted micelles do not exist in a lamellar struc-ture and because of the presence of organic sol-vents, they have limited applicability to biologicalsystems. LUVs, in contrast, retain their bilayerstructure when freeze-dried in the presence ofdisaccharides(Crowe and Crowe, 1988a; Harriganet al., 1990), and thus provide a more relevantstudy system for the question of enzyme activityin biological material.
Both phospholipases C and A(PLC and2
PLA ) were capable of enzymatic activity under2
conditions of low hydration as measured by theappearance of reaction products through analyticalthin layer chromatography. In freeze-dried LUVsallowed to incubate at elevated RH, the appearanceof FFA (the reaction product of PLA) over time2
correlated with a decrease in retention of trappedsolutes(See Fig. 1) (Oliver et al., 1995). A similarpattern was observed for diacylglycerol accumu-lation due to PLC activity(Oliver et al., 1997).As our primary interest was the accumulation ofFFAs, however, we concentrated the investigation

519A.E. Oliver et al. / Comparative Biochemistry and Physiology Part A 131 (2002) 515–525
Fig. 2. Line drawings of the chemical structures for arbutin(A) and rutin(B).
on characterizing PLA activity in terms of water2
requirements and effects of substrate properties.Quantitation of the water contents in equilibrat-
ed samples indicated that PLA activity in freeze-2
dried dielaidoylphosphatidylcholine (DEPC)LUVs proceeded when water was present at aslittle as two moleculesymolecule lipid (Oliver etal., 1997). In addition, retention of trapped solutesdecreased over time when DEPC LUVs lyophili-zed with PLA were incubated at a relative humid-2
ity of only 20% (Oliver et al., 1997). As ambientrelative humidities in nature are quite likely toexceed 20%, this finding indicates that FFA accu-mulation in seeds or other desiccation-toleranttissues, along with its concomitant effect onmembrane leakage, could conceivably proceed viaan enzymatic mechanism.
In the experiments described above, trehalosewas present in the freeze-dried samples at a con-centration of 2 gyg lipid. The sugar was necessaryto inhibit fusion between the liposomes, althoughit was not necessary to protect the protein. PLA2
is an extremely stable enzyme with seven disulfidelinkages preserving its native structure(Dijkstra etal., 1978) and freeze-drying it without sugar doesnot inactivate it(Oliver et al., 1995). For theseexperiments, however, the sugar was necessary topreserve the unilamellar nature of the vesicles,because sequestration of lipid in the internal lamel-lae due to the formation of multilamellar vesicles,caused a dramatic decrease in FFA production(Oliver et al., 1995). This finding raises twoimportant points. Firstly, similar to PLA activity2
in fully hydrated systems(Dennis, 1973; op denKamp, et al., 1974; Verger and de Haas, 1976;Wilschut et al., 1978; Kensil and Dennis, 1985),the physical state of the substrate(including struc-ture, charge, and headgroup spacing) is importantin determining PLA activity in mostly dehydrated2
systems as well(Oliver et al., 1995). This indicatesthat PLA activity in hydrated and mostly dehy-2
drated systems share many common features andunderscores the possibility that enzymatic hydrol-ysis of membrane lipids could contribute to theaccumulation of FFAs in senescent seeds andpollen grains. Secondly, the presence of disaccha-rides, although necessary for protection ofmembrane structure, does not inhibit the enzymeactivity. Thus, it is likely that other compoundsmight be necessary to fulfill that role in vivo forthe long-term protection of anhydrobiotic tissuesand organisms in the dry state.
4. Arbutin inhibits PLA activity at low water2
contents
The glycosylated hydroquinone, arbutin(4-hydroxyphenyl-b-D-glucopyranoside), might fulfillsuch a role. Arbutin, which consists of a phenolmolecule with a glucose moiety in the para posi-tion (See Fig. 2a), is found in extremely highconcentrations in certain resurrection plants ableto survive almost complete dehydration for extend-ed periods of time. For example, inM. flabellifolia,which can survive in the dry state for years, arbutinrepresents up to 25% of the dry weight of theleaves(Suau et al., 1991; Bianchi et al., 1993).The physiological role and sub-cellular location ofarbutin are not known, but it is thought to benecessary for survival in the dry state. Based onthe structural similarity between arbutin and thePLA inhibitor p-bromophenacyl bromide, we2
investigated the possibility that arbutin might becapable of PLA inhibition.2
Indeed, arbutin specifically inhibited PLA2
under conditions of limiting hydration, while ithad a negligible effect on the activities of phos-pholipases B or C(Oliver et al., 1996). WhenLUVs were lyophilized in the presence of PLA2
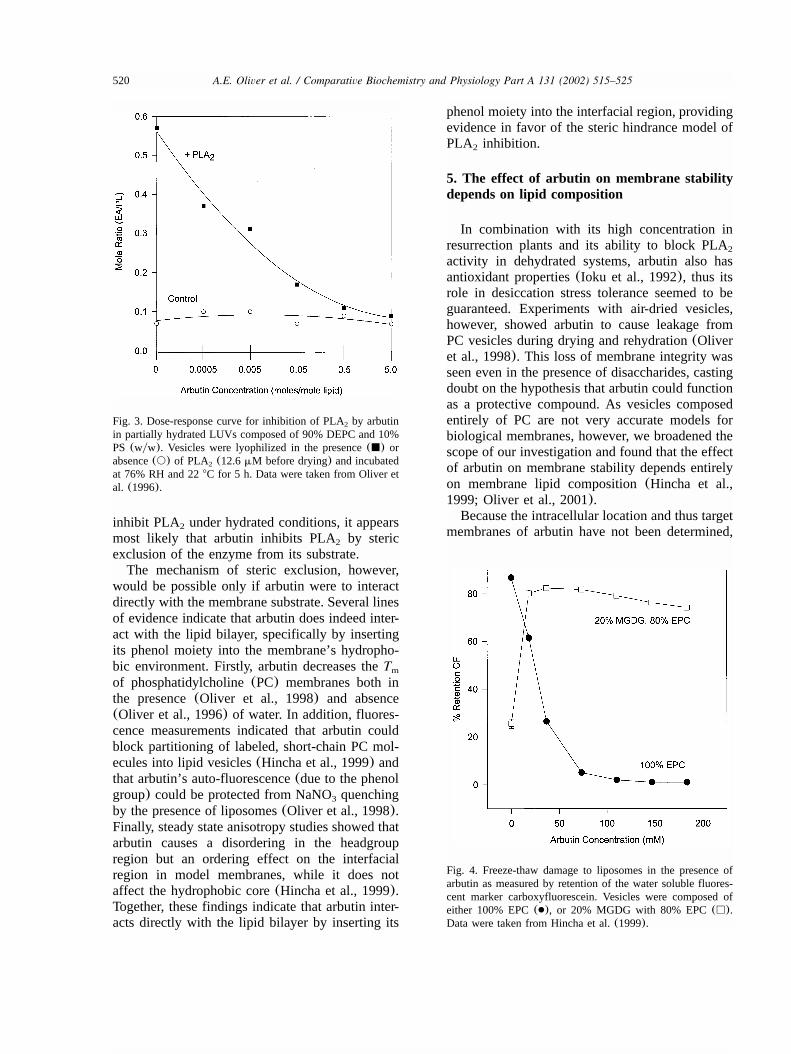
and varying amounts of arbutin, then allowed topartially rehydrate at 76% RH for 5 h, full enzymeinhibition was seen at an arbutin to lipid ratio of0.5 molymol (See Fig. 3) (Oliver et al., 1996).As no direct interaction between PLA and arbutin2
could be detected and because arbutin did not
520 A.E. Oliver et al. / Comparative Biochemistry and Physiology Part A 131 (2002) 515–525
Fig. 3. Dose-response curve for inhibition of PLA by arbutin2
in partially hydrated LUVs composed of 90% DEPC and 10%PS (wyw). Vesicles were lyophilized in the presence(j) orabsence(s) of PLA (12.6mM before drying) and incubated2
at 76% RH and 228C for 5 h. Data were taken from Oliver etal. (1996).
Fig. 4. Freeze-thaw damage to liposomes in the presence ofarbutin as measured by retention of the water soluble fluores-cent marker carboxyfluorescein. Vesicles were composed ofeither 100% EPC(●), or 20% MGDG with 80% EPC(h).Data were taken from Hincha et al.(1999).
inhibit PLA under hydrated conditions, it appears2
most likely that arbutin inhibits PLA by steric2
exclusion of the enzyme from its substrate.The mechanism of steric exclusion, however,
would be possible only if arbutin were to interactdirectly with the membrane substrate. Several linesof evidence indicate that arbutin does indeed inter-act with the lipid bilayer, specifically by insertingits phenol moiety into the membrane’s hydropho-bic environment. Firstly, arbutin decreases theTm
of phosphatidylcholine(PC) membranes both inthe presence(Oliver et al., 1998) and absence(Oliver et al., 1996) of water. In addition, fluores-cence measurements indicated that arbutin couldblock partitioning of labeled, short-chain PC mol-ecules into lipid vesicles(Hincha et al., 1999) andthat arbutin’s auto-fluorescence(due to the phenolgroup) could be protected from NaNO quenching3
by the presence of liposomes(Oliver et al., 1998).Finally, steady state anisotropy studies showed thatarbutin causes a disordering in the headgroupregion but an ordering effect on the interfacialregion in model membranes, while it does notaffect the hydrophobic core(Hincha et al., 1999).Together, these findings indicate that arbutin inter-acts directly with the lipid bilayer by inserting its
phenol moiety into the interfacial region, providingevidence in favor of the steric hindrance model ofPLA inhibition.2
5. The effect of arbutin on membrane stabilitydepends on lipid composition
In combination with its high concentration inresurrection plants and its ability to block PLA2
activity in dehydrated systems, arbutin also hasantioxidant properties(Ioku et al., 1992), thus itsrole in desiccation stress tolerance seemed to beguaranteed. Experiments with air-dried vesicles,however, showed arbutin to cause leakage fromPC vesicles during drying and rehydration(Oliveret al., 1998). This loss of membrane integrity wasseen even in the presence of disaccharides, castingdoubt on the hypothesis that arbutin could functionas a protective compound. As vesicles composedentirely of PC are not very accurate models forbiological membranes, however, we broadened thescope of our investigation and found that the effectof arbutin on membrane stability depends entirelyon membrane lipid composition(Hincha et al.,1999; Oliver et al., 2001).
Because the intracellular location and thus targetmembranes of arbutin have not been determined,

521A.E. Oliver et al. / Comparative Biochemistry and Physiology Part A 131 (2002) 515–525
Fig. 5. Diagrammatic sketch of the ‘shape concept’ of lipid morphology(Tilcock, 1986; Cullis et al., 1986). PC membranes(A) tendto remain in the bilayer phase, because the molecules are roughly cylindrical in shape, favoring a planar orientation. Lipids are drawninside boxes to emphasize their cylindrical nature. In the presence of arbutin(B), PC membranes may be destabilized. Because arbutininserts between the headgroups with its phenol moiety, it is likely to disrupt the ideal packing of the membrane lipids. In membranescomposed of PC and a nonbilayer-forming lipid, such as MGDG or PE(C), the conical shape of the nonbilayer-forming lipid causesan inverted curvature stress on the bilayer. If this stress is strong enough, the membrane will transform to Hex phase. The effect ofII
arbutin on a membrane containing nonbilayer-forming lipids(D) is stabilization of the bilayer phase. As arbutin inserts between theheadgroups, it may be able to equalize the relative volumes of the headgroup and fatty acyl tail regions of the conical lipids, relievingthe inverted curvature stress on the membrane.
isolated chloroplast thylakoid membranes werechosen as a well-characterized possibility to studythe effects of arbutin on membrane stability. Theeffect of arbutin on isolated spinach chloroplastthylakoids and on vesicles containing the thylakoidlipid monogalactosyldiacylglycerol(MGDG) wasmonitored after a freeze–thaw cycle. Althoughfreezing and drying are not identical stress vectors(Crowe et al., 1990), there is an extensive bodyof evidence which indicates that dehydration stresscaused by sequestration of free water as ice playsan important role in freezing-induced injury(Webband Steponkus, 1993; Bronshteyn and Steponkus,1993). Therefore, freezing can provide importantinformation relevant to the effect of arbutin onmembrane stability under conditions of waterdeficit.
Notably, arbutin protected isolated thylakoidmembranes from freeze–thaw damage as measuredby Western blot analysis of release of the internalsoluble protein plastocyanin(Hincha et al., 1999).Furthermore, in vesicles composed entirely of eggPC, arbutin caused freeze-induced damage in a
dose-dependent manner, with only 20 mM arbutincausing complete leakage of entrapped carboxyflu-orescein(CF). In contrast, when 20% MGDG(wyw) was included in the vesicle membranes, arbu-tin instead protected the vesicles in a dose-depend-ent fashion. Only 5 mM arbutin was required toincrease CF retention from 25 to 80%(See Fig.4) (Hincha et al., 1999). Thus, the effect of arbutincould be changed from cryotoxic to cryoprotectivedepending on lipid composition.
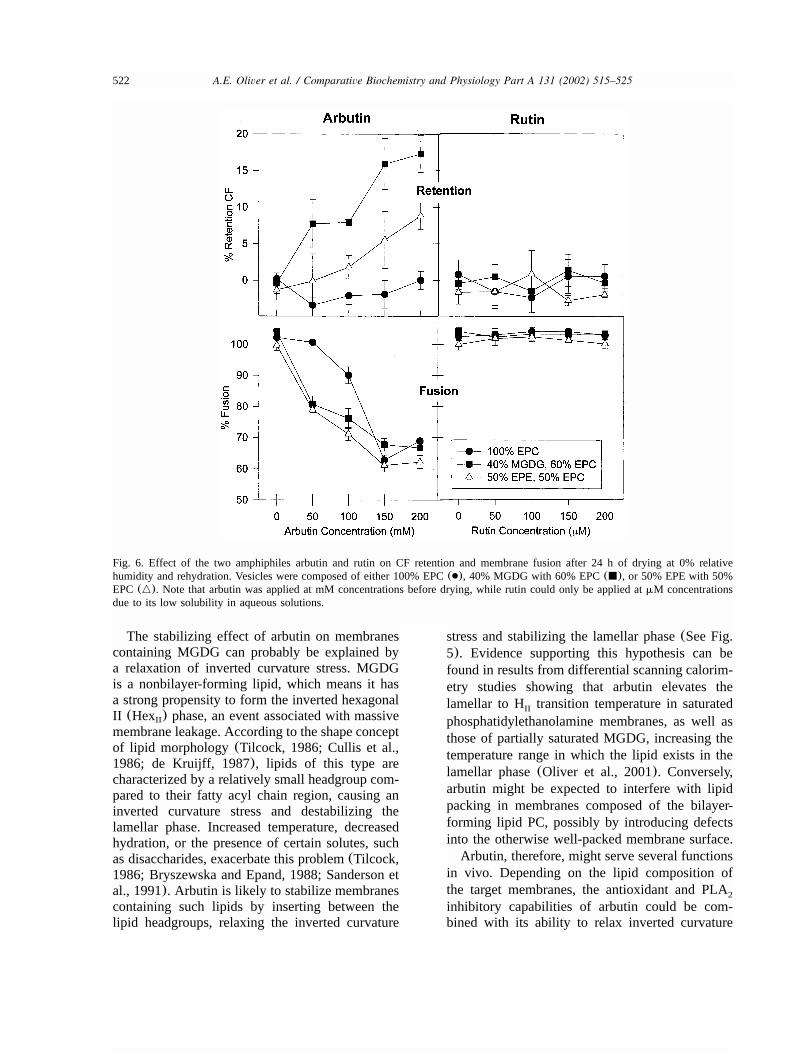
Recent results have indicated that the lipid-dependent effects of arbutin can be seen inresponse to desiccation stress as well. In air-driedLUVs, arbutin destabilizes EPC vesicles, but sta-bilizes vesicles containing EPC with 40% MGDG(wyw). In fact, in the vesicles containing EPCand MGDG, the most protective excipient wasfound to be the combination of sucrose, arbutin,and trehalose in a 3:2:1 mass ratio, the samecompliment of sugars found in driedM. flabelli-folia leaves(Oliver et al., 2001; Bianchi et al.,1993).
522 A.E. Oliver et al. / Comparative Biochemistry and Physiology Part A 131 (2002) 515–525
Fig. 6. Effect of the two amphiphiles arbutin and rutin on CF retention and membrane fusion after 24 h of drying at 0% relativehumidity and rehydration. Vesicles were composed of either 100% EPC(●), 40% MGDG with 60% EPC(j), or 50% EPE with 50%EPC(n). Note that arbutin was applied at mM concentrations before drying, while rutin could only be applied atmM concentrationsdue to its low solubility in aqueous solutions.
The stabilizing effect of arbutin on membranescontaining MGDG can probably be explained bya relaxation of inverted curvature stress. MGDGis a nonbilayer-forming lipid, which means it hasa strong propensity to form the inverted hexagonalII (Hex ) phase, an event associated with massiveII
membrane leakage. According to the shape conceptof lipid morphology(Tilcock, 1986; Cullis et al.,1986; de Kruijff, 1987), lipids of this type arecharacterized by a relatively small headgroup com-pared to their fatty acyl chain region, causing aninverted curvature stress and destabilizing thelamellar phase. Increased temperature, decreasedhydration, or the presence of certain solutes, suchas disaccharides, exacerbate this problem(Tilcock,1986; Bryszewska and Epand, 1988; Sanderson etal., 1991). Arbutin is likely to stabilize membranescontaining such lipids by inserting between thelipid headgroups, relaxing the inverted curvature
stress and stabilizing the lamellar phase(See Fig.5). Evidence supporting this hypothesis can befound in results from differential scanning calorim-etry studies showing that arbutin elevates thelamellar to H transition temperature in saturatedII
phosphatidylethanolamine membranes, as well asthose of partially saturated MGDG, increasing thetemperature range in which the lipid exists in thelamellar phase(Oliver et al., 2001). Conversely,arbutin might be expected to interfere with lipidpacking in membranes composed of the bilayer-forming lipid PC, possibly by introducing defectsinto the otherwise well-packed membrane surface.
Arbutin, therefore, might serve several functionsin vivo. Depending on the lipid composition ofthe target membranes, the antioxidant and PLA2
inhibitory capabilities of arbutin could be com-bined with its ability to relax inverted curvature

523A.E. Oliver et al. / Comparative Biochemistry and Physiology Part A 131 (2002) 515–525
stress and stabilize the lamellar phase of sensitivemembranes.
6. Possible effects of other amphiphiles onmembrane stability
Arbutin, of course, is not the only amphiphilicsubstance present in living cells. An interestingfeature of all such compounds is that they aremainly present in solution in fully hydrated cellsbecause of their partial hydrophilicity. The distri-bution of any given solute between membranesand the aqueous phase will be determined by thestructure and hydrophobicity of its components. Inaddition, it has recently been shown that thehydration state of the cells will also determine thepartitioning of the solutes between the aqueouscompartments and the membranes(Golovina etal., 1998). In the dry state, amphiphiles are expect-ed to be almost exclusively partitioned into thecellular membranes. This partitioning may fluidizebilayers in the dry state, thus decreasingT of them
dry membranes(Golovina et al., 1998; Hoekstraet al., 1997). A decrease inT seems to be am
general effect of many phenolic compounds, suchas salicylic acid or even phenol itself, even in theabsence of drying(van Dael and Ceuterickx, 1984;Casal et al., 1987). This decrease inT is expectedm
to reduce phase-transition leakage during rehydra-tion. Nevertheless, increased leakage has beenobserved in model systems(Golovina et al., 1998).
Quite obviously, not all amphiphiles are able tostabilize membranes under stress conditions. Thisis illustrated in Fig. 6 by comparing the effects ofarbutin and rutin on leakage and fusion duringdrying of liposomes. Rutin is a glycosylated fla-vonoid (with quercetin as the aglycon) (See Fig.2b), which has only a comparatively low solubilityin water. While arbutin was able to stabilizeliposomes containing nonbilayer lipids, asdescribed in detail above, rutin had no beneficialeffects for the membranes under these conditions.
Nevertheless, substances like rutin may havebeneficial effects during long-term desiccation, asthey have been shown to be powerful anti-oxidants(Terao et al., 1994; Ratty et al., 1988). In addition,their effects on membrane stability may also beinfluenced by the presence of other amphiphilesand by sugars. From our current state of knowl-edge, we would hypothesize that anhydrobioteshave evolved to contain a spectrum of differentprotective solutes that may be active against dif-
ferent stresses associated with dehydration. Carefulanalysis is necessary to disentangle the effects ofdifferent solutes and their possible interactions,which allow organisms to survive long-termanhydrobiosis.
References
Almoguera, C., Jordano, J., 1992. Developmental and environ-mental concurrent expression of sunflower dry-seed-storedlow-molecular-weight heat-shock protein and Lea mRNAs.Plant Mol. Biol. 19, 781–792.
Bianchi, G., Gamba, A., Limiroli, R., Pozzi, N., Elster, R.,Salamini, F., Bartels, D., 1993. The unusual sugar compo-sition in leaves of the resurrection plantMyrothamnusflabellifolia. Physiol. Plant. 87, 223–226.
Bronshteyn, V.K., Steponkus, P.L., 1993. Calorimetric studiesof freeze-induced dehydration of phospholipid vesicles.Biophys. J. 65, 1853–1865.
Bryant, G., Wolfe, J., 1992. Interfacial forces in cryobiologyand anhydrobiology. Cryobiol.-Lett. 13, 23–36.
Bryszewska, M., Epand, R.M., 1988. Effects of sugar alcoholsand disaccharides in inducing the hexagonal phase andaltering membrane properties: implications for diabetes mel-litus. Biochim. Biophys. Acta 943, 485–492.
Buitink, J., Claessens, M.M.A.E., Hemminga, M.A., Hoekstra,F.A., 1998. Influence of water content and temperature onmolecular mobility and intracellular glasses in seeds andpollen. Plant Physiol. 118, 531–541.
Buitink, J., Hemminga, M.A., Hoekstra, F.A., 1999. Character-ization of molecular mobility in seed tissues: an electronparamagnetic resonance spin probe study. Biophys. J. 76,3315–3322.
Casal, H.L., Martin, A., Mantsch, H.H., 1987. Infrared spec-troscopic characterization of the interaction of lipid bilayerswith phenol, salicylic acid ando-acetylsalicylic acid. Chem.Phys. Lipids 43, 47–53.
Crowe, J.H., Carpenter, J.F., Crowe, L.M., Anchordoguy, T.J.,1990. Are freezing and dehydration similar stress vectors?A comparison of modes of interaction of stabilizing soluteswith biomolecules. Cryobiology 27, 219–231.
Crowe, J.H., Crowe, L.M., 1988a. Factors affecting the stabil-ity of dry liposomes. Biochim. Biophys. Acta 939, 327–334.
Crowe, L.M., Crowe, J.H., 1988b. Trehalose and dry dipalmi-toylphosphatidylcholine revisited. Biochim. Biophys. Acta946, 193–201.
Crowe, J.H., Crowe, L.M., Carpenter, J.F., Petrelski, S., Hoek-stra, F.A., de Araujo, P., Panek, A.D., 1997a. Anhydrobiosis:cellular adaptation to extreme dehydration. In: Dantzler,W.H. (Ed.), Handbook of Physiology, Vol. II. OxfordUniversity Press, Oxford, pp. 1445–1477.
Crowe, J.H., Crowe, L.M., Chapman, D., 1984. Preservationof membranes in anhydrobiotic organisms: the role oftrehalose. Science 223, 701–703.
Crowe, J.H., Hoekstra, F.A., Nguyen, K.N., Crowe, L.M.,1996. Is vitrification involved in depression of the phasetransition temperature in dry phospholipids? Biochim. Bio-phys. Acta 1280, 187–196.
Crowe, J.H., Leslie, S.B., Crowe, L.M., 1994. Is vitrificationsufficient to preserve liposomes during freeze-drying? Cry-obiology 31, 355–366.

524 A.E. Oliver et al. / Comparative Biochemistry and Physiology Part A 131 (2002) 515–525
Crowe, J.H., McKersie, B.D., Crowe, L.M., 1989. Effects offree fatty acids and transition temperature on the stabilityof dry liposomes. Biochim. Biophys. Acta 979, 7–10.
Crowe, J.H., Oliver, A.E., Hoekstra, F.A., Crowe, L.M., 1997b.Stabilization of dry membranes by mixtures of hydroxyethylstarch and glucose: the role of vitrification. Cryobiology 35,20–30.
Cullis, P.R., Hope, M.J., Tilcock, C.P.S., 1986. Lipid polymor-phism and the roles of lipids in membranes. Chem. Phys.Lipids 40, 127–144.
Cuming, A.C., 1999. LEA Proteins. In: Shewry, P.R., Casey,R. (Eds.), Seed Proteins, Kluwer Academic Publishers,Dordrecht, Boston, pp. 753–780.
de Kruijff, B., 1987. Polymorphic regulation of membranelipid composition. Nature 329, 587–588.
Dennis, E.A., 1973. Phospholipase A activity towards phos-2
phatidylcholine in mixed micelles: surface dilution kineticsand the effect of thermotropic phase transitions. Arch.Biochem. Biophys. 158, 485–493.
Dijkstra, B.W., Drenth, J., Kalk, K.H., Vandermaelen, P.J.,1978. Three-dimensional structure and disulfide bond con-nections in bovine pancreatic phospholipase A . J. Mol.2
Biol. 124, 53–60.Drennan, P.M., Smith, M.T., Goldsworthy, D., van Staden, J.,
1993. The occurrence of trehalose in the leaves of thedesiccation-tolerant angiospermMyrothamnus flabellifoliusWelw. J. Plant Physiol. 142, 493–496.
Golovina, E.A., Hoekstra, F.A., Hemminga, M.A., 1998. Dry-ing increases intracellular partitioning of amphiphilic sub-stances into the lipid phase. Plant Physiol. 118, 975–986.
Harrigan, P.R., Madden, T.D., Cullis, P.R., 1990. Protection ofliposomes during dehydration or freezing. Chem. Phys.Lipids 52, 139–149.
Hincha, D.K., Hellwege, E.M., Heyer, A.G., Crowe, J.H.,2000. Plant fructans stabilize phosphatidylcholine liposomesduring freeze-drying. Eur. J. Biochem. 267, 535–540.
Hincha, D.K., Oliver, A.E., Crowe, J.H., 1999. Lipid compo-sition determines the effects of arbutin on the stability ofmembranes. Biophys. J. 77, 2024–2034.
Hoekstra, F.A., Crowe, J.H., Crowe, L.M., 1989. Membranebehavior in drought and its physiological significance. In:Taylorson, R.B.(Ed.), Recent Advances in the Developmentand Germination of Seeds, Plenum Press, New York, pp.71–88.
Hoekstra, F.A., 1986. Water content in relation to stress inpollen. In: Leopold, A.C.(Ed.), Membranes, Metabolism,and Dry Organisms, Comstock Pub. Assoc, Ithaca, NY, pp.102–122.
Hoekstra, F.A., Crowe, J.H., Crowe, L.M., van Roekel, T.,Vermeer, E., 1992. Do phospholipids and sucrose determinemembrane phase transitions in dehydrating pollen species?Plant, Cell Environ. 15, 601–606.
Hoekstra, F.A., Wolkers, W.F., Buitink, J., Golovina, E.A.,Crowe, J.H., Crowe, L.M., 1997. Membrane stabilization inthe dry state. Comp. Biochem. Physiol. 117A, 335–341.
Ingram, J., Bartels, D., 1996. The molecular basis of dehydra-tion tolerance in plants. Annu. Rev. Plant Physiol. PlantMol. Biol. 47, 377–403.
Ioku, K., Terao, J., Nakatani, N., 1992. Antioxidative activityof arbutin in a solution and liposomal suspension. Biosci.Biotechnol. Biochem. 56, 1658–1659.
Kensil, C.R., Dennis, E.A., 1985. Action of cobra venomphospholipase A on large unilamellar vesicles: comparison2
with small unilamellar vesicles and multilayers. Lipids 20,80–83.
Koster, K.L., Lei, Y.P., Anderson, M., Martin, S., Bryant, G.,2000. Effects of vitrified and nonvitrified sugars on phos-phatidylcholine fluid-to-gel phase transitions. Biophys. J.78, 1932–1946.
Koster, K.L., Leopold, A.C., 1988. Sugars and desiccationtolerance in seeds. Plant Physiol. 88, 829–832.
Koster, K.L., Webb, M.S., Bryant, G., Lynch, D.V., 1994.Interactions between soluble sugars and POPC during dehy-dration: vitrification of sugars alters the phase behavior ofthe phospholipid. Biochim. Biophys. Acta 1193, 143–150.
Leprince, O., Deltour, R., Thorpe, P.C., Atherton, N.M., Hen-dry, G.A.F., 1990. The role of free radicals and radicalprocessing systems in loss of desiccation tolerance in ger-minating maize (Zea mays L.). New Phytologist 116,573–580.
Leprince, O., Harren, F.J.M., Buitink, J., Alberda, M., Hoek-stra, F.A., 2000. Metabolic dysfunction and unabated respi-ration precede the loss of membrane integrity duringdehydration of germinating radicles. Plant Physiol. 122,597–608.
Leslie, S.B., Teter, S.A., Crowe, L.M., Crowe, J.H., 1994.Trehalose lowers membrane phase transitions in dry yeastcells. Biochim. Biophys. Acta 1192, 7–13.
Leslie, S.B., Israeli, E., Lighthart, B., Crowe, J.H., Crowe,L.M., 1995. Trehalose and sucrose protect both membranesand proteins in intact bacteria during drying. Appl. Environ.Microbiol. 61, 3592–3597.
McKersie, B.D., Crowe, J.H., Crowe, L.M., 1989. Free fattyacid effects on leakage, phase properties and fusion of fullyhydrated model membranes. Biochim. Biophys. Acta 982,156–160.
McKersie, B.D., Senaratna, T., Walker, M.A., Kendall, E.J.,Hetherington, P.R., 1988. Deterioration of membranes duringaging in plants: evidence for free radical mediation. In:Nooden, L.D., Leopold, A.C.(Eds.), Senescence and Agingin Plants, Academic Press, Inc, San Diego, pp. 441–464.
McKersie, B.D., Thompson, J.E., 1977. Lipid crystallizationin senescent membranes from cotyledons. Plant Physiol. 59,803–807.
McKersie, B.D., Thompson, J.E., Brandon, J.K., 1976. X-raydiffraction evidence for decreased lipid fluidity in senescentmembranes from cotyledons. Can. J. Bot. 54, 1074–1078.
Misiorowski, R.L., Wells, M.A., 1974. Phospholipase A in2
reversed micelles of phosphatidylcholine in diethylether:effect of water and cations. Biochemistry 13, 4921–4927.
Oliver, A.E., Crowe, L.M., de Araujo, P.S., Fisk, E., Crowe,J.H., 1996. Arbutin inhibits PLA in partially hydrated2
model systems. Biochim. Biophys. Acta 1302, 69–78.Oliver, A.E., Fisk, E., Crowe, L.M., de Araujo, P.S., Crowe,
J.H., 1995. Phospholipase A activity in dehydrated systems:2
effect of the physical state of the substrate. Biochim.Biophys. Acta 1267, 92–100.
Oliver, A.E., Fisk, E., Crowe, L.M., de Araujo, P.S., Crowe,J.H., 1997. Evidence of phospholipase activity in phospho-lipid bilayers under conditions of low hydration. J. PlantPhysiol. 150, 661–667.
Oliver, A.E., Hincha, D.K., Crowe, J.H., 2001. The effect ofarbutin on membrane integrity during drying is mediated by

525A.E. Oliver et al. / Comparative Biochemistry and Physiology Part A 131 (2002) 515–525
stabilization of the lamellar phase in the presence of non-bilayer-forming lipids. Chem. Phys. Lipids 111, 37–57.
Oliver, A.E., Hincha, D.K., Crowe, L.M., Crowe, J.H., 1998.Interactions of arbutin with dry and hydrated bilayers.Biochim. Biophys. Acta 1370, 87–97.
op den Kamp, J.A.F., de Gier, J., van Deenen, L.L.M., 1974.Hydrolysis of phosphatidylcholine liposomes by pancreaticphospholipase A at the transition temperature. Biochim.2
Biophys. Acta 345, 253–256.Pilon-Smits, E.A.H., Ebskanmp, M.J.M., Paul, M.J., Jeuken,
M.J.W., Weisbeek, P.J., Smeekens, S.C.M., 1995. Improvedperformance of transgenic fructan-accumulating tobaccounder drought stress. Plant Physiol. 107, 125–130.
Pollock, C.J., Cairns, A.J., 1991. Fructan metabolism in grassesand cereals. Ann. Rev. Plant Physiol. Plant Mol. Biol. 42,77–101.
Poon, P.H., Wells, M.A., 1974. Physical studies of egg phos-phatidylcholine in diethyl ether-water solution. Biochemistry13, 4928–4936.
Price, A.H., Hendry, G.A.F., 1989. Stress and the role ofactivated oxygen scavengers and protective enzymes inplants subjected to drought. Biochem. Soc. Trans. 17,493–494.
Priestly, D.A., 1986. Seed Aging Implications for Seed Storageand Persistence in Soil. Cornell Univ. Press, Ithaca, NY.
Ratty, A.K., Sunamoto, J., Das, N.P., 1988. Interaction offlavonoids with 1,1-diphenyl-2-picrylhydrazyl free radical,liposomal membranes and soybean lipoxygenase-1. Bioch-em. Pharmacol. 37, 989–995.
Roberts, E.H., 1979. Seed deterioration and loss of viability.Adv. Res. Technol. Seeds 4, 25–42.
Sanderson, P.W., Lis, L.J., Quinn, P.J., Williams, W.P., 1991.The Hofmeister effect in relation to membrane lipid phasestability. Biochim. Biophys. Acta 1067, 43–50.
Senaratna, T., McKersie, B.D., Borochov, A., 1987. Desicca-tion and free radical mediated changes in plant membranes.J. Exp. Bot. 38, 2005–2014.
Slade, L., Levine, H., 1991. Beyond water activity: recentadvances based on an alternative approach to the assessmentof food quality and safety. Crit. Rev. Food Sci. Nutr. 30,115–360.
Smirnoff, N., 1993. The role of active oxygen in the responseof plants to water deficit and desiccation. New Phytol. 125,27–58.
Suau, R., Cuevas, A., Valpuesta, V., Reid, M.S., 1991. Arbutinand sucrose in the leaves of the resurrection plantMyro-thamnus flabellifolia. Phytochemistry 30, 2555–2556.
Sun, W.Q., Leopold, A.C., 1994. Glassy state and seed storagestability: a viability equation analysis. Ann. Bot. 74,601–604.
Sun, W.Q., Leopold, A.C., 1997. Cytoplasmic vitrification andsurvival of anhydrobiotic organisms. Comp. Biochem. Phy-siol. 117A, 327–333.
Sun, W.Q., Leopold, A.C., Crowe, L.M., Crowe, J.H., 1996.Stability of dry liposomes in sugar glasses. Biophys. J. 70,1769–1776.
Terao, J., Piskula, M., Yao, Q., 1994. Protective effect ofepicatechin, epicatechin gallate and quercetin on lipid per-oxidation in phospholipid bilayers. Arch. Biochem. Biophys.308, 278–284.
Tilcock, C.P.S., 1986. Lipid polymorphism. Chem. Phys. Lip-ids 40, 109–125.
Tsvetkova, N.M., Phillips, B.L., Crowe, L.M., Crowe, J.H.,Risbud, S.H., 1998. Effect of sugars on headgroup mobilityin freeze-dried dipalmitoylphosphatidylcholine bilayers: sol-id-state P NMR and FTIR studies. Biophys. J. 75,31
2947–2955.van Bilsen, D.G.J.L., Hoekstra, F.A., 1993. Decreased
membrane integrity in agingTypha latifolia L. pollen.Accumulation of lysolipids and free fatty acids. Plant Phy-siol. 101, 675–682.
van Bilsen, D.G.J.L., Hoekstra, F.A., Crowe, L.M., Crowe,J.H., 1994. Altered phase behavior in membranes of agingdry pollen may cause imbibitional leakage. Plant Physiol.104, 1193–1199.
van Dael, H., Ceuterickx, P., 1984. The interaction of phenolwith lipid bilayers. Chem. Phys. Lipids 35, 171–181.
Verger, R., de Haas, G.H., 1976. Interfacial enzyme kineticsof lipolysis. Ann. Rev. Biophys. Bioeng. 5, 77–117.
Vijn, I., Smeekens, S., 1999. Fructan: more than a reservecarbohydrate? Plant Physiol. 120, 351–359.
Webb, M.S., Steponkus, P.L., 1993. Freeze-induced membraneultrastructural alterations in Rye(Secale cereale) leaves.Plant Physiol. 101, 955–963.
Wehmeyer, N., Vierling, E., 2000. The expression of smallheat shock proteins in seeds responds to discrete develop-mental signals and suggests a general protective role indesiccation tolerance. Plant Physiol. 122, 1099–1108.
Westh, P., Ramlov, H., 1991. Trehalose accumulation in thetardigradeAdorybiotus coronifer during anhydrobiosis. J.Exp. Zool. 258, 303–311.
Wilschut, J.C., Regts, J., Westenberg, H., Scherphof, G., 1978.Action of phopholipase A on phosphatidylcholine bilayers.2
Effects of the phase transition, bilayer curvature and struc-tural defects. Biochim. Biophys. Acta 508, 185–196.
Wolfe, J., Bryant, G., 1999. Freezing, drying andyor vitrifi-cation of membrane-solute-water systems. Cryobiology 39,103–129.
Wolkers, W.F., McCready, S., Brandt, W.F., Lindsey, G.G.,Hoekstra, F.A., 2001. Isolation and characterization of a D-7 LEA protein from pollen that stabilizes glasses in vitro.Biochim. Biophys. Acta 1544, 196–206.
Wolkers, W.F., Tetteroo, F., Alberda, M., Hoekstra, F.A., 1999.Changed properties of the cytoplasmic matrix associatedwith desiccation tolerance of dried carrot somatic embryos.An in situ Fourier transform infrared spectroscopic study.Plant Physiol. 120, 153–163.
Womersly, C., 1990. A reconsideration of diversity of adapta-tion in nematode anhydrobiotes in relation to their environ-ments. In: Veeck, J.A., Dickson, D.W.(Eds.), Vistas onNematology, Academic Press, New York, pp. 165–173.




















