
Local administration of calcitriol positively influences bone remodeling and maturation during...
46
Local administration of calcitriol positively influences bone remodeling and maturation during restoration of mandibular bone defects in rats Hongrui Liu, Jian Cui, Wei Feng, Shengyu Lv, Juan Du, Jing Sun, Xi- uchun Han, Zhenming Wang, Xiong Lu, Yimin, Kimimitsu Oda, Norio Amizuka, Minqi Li PII: S0928-4931(14)00862-5 DOI: doi: 10.1016/j.msec.2014.12.064 Reference: MSC 5149 To appear in: Materials Science & Engineering C Received date: 10 July 2014 Revised date: 26 November 2014 Accepted date: 17 December 2014 Please cite this article as: Hongrui Liu, Jian Cui, Wei Feng, Shengyu Lv, Juan Du, Jing Sun, Xiuchun Han, Zhenming Wang, Xiong Lu, Yimin, Kimimitsu Oda, Norio Amizuka, Minqi Li, Local administration of calcitriol positively influences bone remodeling and maturation during restoration of mandibular bone defects in rats, Materials Science & Engineering C (2014), doi: 10.1016/j.msec.2014.12.064 This is a PDF file of an unedited manuscript that has been accepted for publication. As a service to our customers we are providing this early version of the manuscript. The manuscript will undergo copyediting, typesetting, and review of the resulting proof before it is published in its final form. Please note that during the production process errors may be discovered which could affect the content, and all legal disclaimers that apply to the journal pertain.
Transcript of Local administration of calcitriol positively influences bone remodeling and maturation during...

�������� ����� ��
Local administration of calcitriol positively influences bone remodeling andmaturation during restoration of mandibular bone defects in rats
Hongrui Liu, Jian Cui, Wei Feng, Shengyu Lv, Juan Du, Jing Sun, Xi-uchun Han, Zhenming Wang, Xiong Lu, Yimin, Kimimitsu Oda, NorioAmizuka, Minqi Li
PII: S0928-4931(14)00862-5DOI: doi: 10.1016/j.msec.2014.12.064Reference: MSC 5149
To appear in: Materials Science & Engineering C
Received date: 10 July 2014Revised date: 26 November 2014Accepted date: 17 December 2014
Please cite this article as: Hongrui Liu, Jian Cui, Wei Feng, Shengyu Lv, Juan Du, JingSun, Xiuchun Han, Zhenming Wang, Xiong Lu, Yimin, Kimimitsu Oda, Norio Amizuka,Minqi Li, Local administration of calcitriol positively influences bone remodeling andmaturation during restoration of mandibular bone defects in rats, Materials Science &Engineering C (2014), doi: 10.1016/j.msec.2014.12.064
This is a PDF file of an unedited manuscript that has been accepted for publication.As a service to our customers we are providing this early version of the manuscript.The manuscript will undergo copyediting, typesetting, and review of the resulting proofbefore it is published in its final form. Please note that during the production processerrors may be discovered which could affect the content, and all legal disclaimers thatapply to the journal pertain.

ACC
EPTE
D M
ANU
SCR
IPT
ACCEPTED MANUSCRIPT
1
Local administration of calcitriol positively influences bone remodeling and
maturation during restoration of mandibular bone defects in rats
Hongrui Liu§1
, Jian Cui§1
, Wei Feng1, Shengyu Lv
1, Juan Du
1, Jing Sun
1, Xiuchun
Han1, Zhenming Wang
2, Xiong Lu
2, Yimin
3, Kimimitsu Oda
4, Norio Amizuka
5, Minqi
Li1
1Department of Bone Metabolism, School of Stomatology Shandong University,
Shandong Provincial Key Laboratory of Oral Biomedicine, Jinan, China 2Key Lab of
Advanced Technologies of Materials, Ministry of Education, School of Materials
Science and Engineering, Southwest Jiaotong University, Chengdu, Sichuan, China 3
Department of Advanced Medicine, Graduate School of Medicine, Hokkaido
University, Sapporo, Japan 4Division of Biochemistry, Niigata University Graduate
School of Medical and Dental Sciences, Niigata, Japan and 5Department of
Developmental Biology of Hard Tissue, Graduate School of Dental Medicine,
Hokkaido University, Sapporo, Japan
Address for correspondence:
Minqi Li, DDS, PhD
Department of Bone Metabolism
School of Stomatology Shandong University
Shandong Provincial Key Laboratory of Oral Biomedicine

ACC
EPTE
D M
ANU
SCR
IPT
ACCEPTED MANUSCRIPT
2
Wenhua West Road 44-1, Jinan 250012, China
Tel & Fax: +86-531-88382095
E-mail: [email protected]
§ We regard these authors equally contributed to this article.
Abstract
The aim of this study was to investigate the influence of calcitriol on osteoinduction
following local administration into mandibular bone defects. Calcitriol-loaded
absorbable collagen membrane scaffolds were prepared using the polydopamine
coating method and characterized by scanning electron microscopy. Composite
scaffolds were implanted into rat mandibular bone defects in the following groups: no
graft material (control), bare collagen membrane (CM group), collagen membrane
bearing polydopamine coating (DOP/CM group), and collagen membrane bearing
polydopamine coating absorbed with calcitriol (CAL/DOP/CM group). At 1, 2, 4 and
8 weeks post-surgery, the osteogenic potential of calcitriol was examined by
histological and immunohistochemical methods. Following in vivo implantation,
calcitriol-loaded composite scaffolds underwent rapid degradation with pronounced
replacement by new bone and induced reunion of the bone marrow cavity. Calcitriol
showed strong potential in inhibiting osteoclastogenesis and promotion of osteogenic
differentiation at weeks 1, 2. Furthermore, statistical analysis revealed that the newly
formed bone volume in the CAL/DOP/CM group was significantly higher than other
groups at weeks 1, 2. At weeks 4, 8, the CAL/DOP/CM group showed more
mineralized bone and uniform collagen structure. These data suggest that local

ACC
EPTE
D M
ANU
SCR
IPT
ACCEPTED MANUSCRIPT
3
administration of calcitriol is promising in promoting osteogenesis and mineralization
for restoration of mandibular bone defects.
Keywords: calcitriol, osteoclast, osteoblast, mineralization, mandibular bone defect
Running title: Calcitriol promotes repair of mandibular bone defects
Introduction
Achieving satisfactory bone regeneration in vivo remains an important goal in
orthopedic and dental clinical applications. Growth-factor-based tissue engineering
technologies have been widely studied, with much of this research focused on bone
morphogenetic proteins (BMPs), which have favorable osteogenic potential (Han et al.
2014; Muzio et al. 2014; Sigurdsson et al. 2001; Tatakis et al. 2002; Wang et al. 2014).
Despite that, their use is going to be limited due to several drawbacks including their
rapid degradation, high costs, safety and efficacy concerns, osteolysis, ectopic bone
formation, and soft tissue swelling (Arrabal et al. 2013; Ehnert et al. 2012; Oryan et al.
2014). Recently, it was reported that some drugs and bioactive substances which have
stable and srong activity, low costs and high biological safety could also be used to
regulate tissue growth (Jang et al. 2003; Kim et al. 2006). Calcitriol, also called
1,25-dihydroxyvitamin D3 (1, 25(OH)2D3), is the active form of vitamin D3 and is

ACC
EPTE
D M
ANU
SCR
IPT
ACCEPTED MANUSCRIPT
4
involved in various physiological processes, including calcium homeostasis, bone
metabolism and immune response (Inada et al. 2008). It functions by binding to a
single vitamin D receptor (VDR), which is a member of the nuclear hormone receptor
superfamily (Feldman et al. 2005). Vitamin D-deficiency causes impairment of bone
mineralization, resulting in rickets in infants and osteomalacia in adults (Suda et al.
2003). Moreover, mice lacking VDR exhibit impaired bone formation (Yoshizawa et
al. 1997). Conversely, administration of vitamin D to ovariectomized or rachitic
animals can relieve impaired bone mineralization (Shiraishi et al. 2002). In addition,
clinical studies show that vitamin D treatment can reduce bone fracture risk in
age-related bone loss, postmenopausal osteoporosis and drug-induced osteoporosis
(Avenell et al. 2005; de Nijs et al. 2004; Nawata et al. 2005). Besides the overall
positive effects on bone, calcitriol has also been shown to regulate transcription of the
collagen gene in osteoblasts (Harrison et al. 1989) and is a potent transcriptional
activator of genes encoding alkaline phosphatase (ALP), osteocalcin (OCN) and
osteopontin (OPN) (Noda et al. 1990). Moreover, osteoblasts have shown increased
activity after 1, 25(OH)2D3 treatment in vitro (Bosetti et al. 2007).
Based on these studies, we aimed to investigate local administration of calcitriol
for the restoration of mandibular bone defects in rats. However, the lack of a suitable
local delivery system has previously impeded in vivo application of calcitriol. An
appropriate scaffold with good biocompatibility and biodegradability is therefore
required. More importantly, an optimal amount of calcitriol to generate osteoinduction
needs to be released into the target site over a prolonged period of time. Absorbable

ACC
EPTE
D M
ANU
SCR
IPT
ACCEPTED MANUSCRIPT
5
collagen membrane, a well-documented barrier membrane in guided bone
regeneration (GBR) (Chiapasco and Zaniboni 2009; Hämmerle et al. 2002), has been
used as a growth-factor-combined scaffold. Type-I collagen, as the main component
of collagen membrane, is a natural and ubiquitous protein. Its inherent properties
include cell attachment and bioactivity, natural biodegradability, amenability to
chemical modifications, mechanical cross-linking and self-assembly into a fibrillar gel
under physiological conditions, making collagen advantageous for tissue engineering
and regenerative medicine applications (Drzewiecki et al. 2014). In addition, it also
showed significant strength and rigidity which can be attributed to its three
polypeptide chains structure (Drzewiecki et al. 2014). Numerous animal studies have
shown that collagen membrane is crucially important in retaining BMP levels and
ensuring maintenance of space for new bone formation (Khan et al. 2012). Moreover,
it is reported that loading of collagen membrane with rhBMP-2 shows favorable bone
regenerative ability (Jung et al. 2003). In view of these results, we selected collagen
membrane as the scaffold for loading calcitriol. However, to achieve enhanced
immobilization of calcitriol, this carrier material requires additional processing.
Various techniques are available for immobilizing biomolecules onto scaffolds,
including gamma ray irradiation and plasma treatment (Cottam et al. 2009; Nagai et al.
2006; Vallières et al. 2007; Xia et al. 2010). However, these methods have limitations
in terms of penetration depth and possible cleavage of polymeric chains (Nyström et
al. 2010). Recently, a versatile surface modification method, by simply dip-coating
with dopamine solution, was reported, and can be applied to almost all solid materials
from metals to synthetic polymers (Lee et al. 2007). Dopamine can undergo oxidative
polymerization and form a stable layer adherent to the surface of biomaterials under
basic conditions. Previous studies have applied the polydopamine-coated method to

ACC
EPTE
D M
ANU
SCR
IPT
ACCEPTED MANUSCRIPT
6
immobilizing animated methoxy-polyethylene glycol, trypsin, bovine serum albumin
(BSA), and antibodies (Lynge et al. 2011). In addition, it is reported that growth
factors, such as vascular endothelial growth factor and BMP-2, immobilized onto
titanium surfaces following polydopamine coating, promote proliferation of
endothelial cells and differentiation of rat mesenchymal stem cells, respectively
(Lynge et al. 2011). Most importantly, polydopamine has shown excellent
biocompatibility and low cytotoxicity, making it a favorable platform (Cui et al. 2012;
Yang et al. 2012).
In this study, we loaded calcitriol onto collagen membranes using the
polydopamine-coating method and evaluated the osteogenic capability of calcitriol by
applying the composite scaffold into mandibular bone defects in rats.
Materials and Methods
Immobilization of calcitriol on collagen membrane and SEM observation
Dopamine solution (2mg/ml) was prepared by dissolving 3,4-dihydroxyphenylamine
(Sigma-Aldrich, St. Louis, MO, USA) in Tris-HCl buffer (10 mM, pH 8.5), in
accordance with a previous study (Yang et al. 2012). Calcitriol (Selleck Chemicals,
Houston, TX, USA) was then dissolved in a small amount of ethanol and diluted with
dopamine solution to a final concentration of 5×10-4
g/ml referring to published
studies (Bosetti et al. 2014; Yoon et al. 2007). Collagen membranes purchased by
ZhengHai Biotechnology Co., Ltd. (Yantai, Shandong, China) were cut at 4×3×1 mm
and incubated with 100 μl of calcitriol solution at 4°C overnight. Tris-HCl buffer and

ACC
EPTE
D M
ANU
SCR
IPT
ACCEPTED MANUSCRIPT
7
dopamine solution were used as controls. All steps were performed in the dark.
Scanning electron microscopy (SEM; S-2250N, Hitachi, Tokyo, Japan) was used
to observe the surface morphology of the original collagen membrane,
polydopamine-coated collagen membrane and collagen membrane bearing
polydopamine coating absorbed with calcitriol. For this purpose, samples were
mounted on aluminum stubs and sputter-coated with gold particles to a thickness of
10-15 nm.
Animal mandibular bone defect model and tissue preparation
All animal experiments were conducted in accordance with the Guidelines for
Animal Experimentation of the School and Hospital of Stomatology, Shandong
University. Ninety-six Wistar rats (200-250 g, male) were anesthetized with 10%
chloral hydrate (0.4 ml/100 g body weight). After an extra-oral incision parallel to the
inferior border of the mandible, subcutaneous tissues and masseter muscle were
dissected. Bilateral mandible defects were then made around the first molar with a
fissure bur at low speed with copious saline irrigation. The mandibular wound
window was approximately 5×4×1 mm, with the anterior margin 1 mm distal to the
front of the mandible, and the coronal margin was approximately 1 mm apical of the
crest of the alveolar bone (Fig. 1D). Collagen membranes immersed in different
solutions were then implanted into the defects. Soft tissues were then sutured, and
antibiotics were administered during the first 3 postoperative days.
Four groups were established to treat bone defects (n=6 animals per group per
time point, Table 1): (i) no graft material (control group); (ii) bare collagen membrane
(CM group); (iii) collagen membrane bearing polydopamine coating (DOP/CM

ACC
EPTE
D M
ANU
SCR
IPT
ACCEPTED MANUSCRIPT
8
group); (iv) collagen membrane bearing polydopamine coating absorbed with
calcitriol (CAL/DOP/CM group).
At 1, 2, 4 and 8 weeks post-surgery, animals were anesthetized and fixed with a
transcardiac perfusion of 4% paraformaldehyde in 0.1 M phosphate-buffered saline
(PBS; pH 7.4). Following fixation, mandibles were then demineralized in 10%
EDTA-2Na solution for 4 weeks at 4°C. After dehydration through a graded
ascending ethanol series, the tissues were embedded in paraffin. Serial horizontal
5-µm-thick sections were cut in the apical-to-coronal direction of the tooth.
Histological examination and image analysis
Hematoxylin and eosin staining and Masson’s trichrome stainings were
performed to identify the osteogenic status of the different groups. Stained sections
were observed and digital images were taken with a light microscope (Olympus
BX-53, Tokyo, Japan). With the aid of Image-Pro Plus 6.2 software (Media
Cybernetics, Silver Spring, MD), newly formed bone volume, expressed as a
percentage (area of newly formed bone/area of original wound (defect)*100%), was
measured at an original magnification of ×40. Specifically, bone defect area was
defined from histologically visible cut edge of cortical bone and newly formed bone
was identified by their woven structure.
Three tissue sections were selected from each of three levels throughout the
whole defect (the apical, middle and coronal levels), and three non-overlapping
images of each section were analyzed to obtain the mean value. Ultimately, the newly
formed bone volume of each group was expressed as the average level of the parallel
samples.

ACC
EPTE
D M
ANU
SCR
IPT
ACCEPTED MANUSCRIPT
9
Immunochemical examination for ALP, Runx2, Type I collagen and OPN
Serial 5-µm-thick paraffin sections were used for immunolocalization of ALP,
Runx2, Type I collagen and OPN. Briefly, dewaxed paraffin sections were pretreated
with 0.3% hydrogen peroxidase for 30 min to inhibit endogenous peroxidase activity
prior to treatment with 1% BSA in PBS (1% BSA-PBS) for 20 min. Sections were
then incubated for 2 h at room temperature with: 1) rabbit antiserum against rat
tissue-nonspecific ALP, generated by Oda et al. (1999) at a dilution of 1:100; 2)
mouse anti-Runx2 antibody (MBL, Nagoya, Japan) at a dilution of 1:50; 3) rabbit
anti-collagen I antibody (Abcam, Cambridge, MA, USA) at a dilution of 1:200; or 4)
rabbit anti-OPN antibody (Abcam, Cambridge, MA, USA) at a dilution of 1:50. After
a PBS rinse, sections were incubated with horseradish peroxidase-conjugated
secondary antibodies (DaKo, Glostrup, Denmark) at a dilution of 1:100 for 1 h at
room temperature. The immunoreaction was visualized with diaminobenzidine
(Sigma-Aldrich, St. Louis, MO, USA). Double-staining of ALP and tartrate-resistant
acid phosphatase (TRAP) was performed as previously reported (Li et al. 2013).
Staining was assessed by light microscopy after faintly counterstaining with methyl
green. Primary antibodies were replaced with PBS for negative controls.
Stained sections were imaged with an Olympus BX53 microscope. Specifically,
three tissue sections were selected from each of the three levels (middle, coronal and
apical). Osteoclast numbers were counted, and the mean optical density of ALP and
Runx2 were measured in three randomly selected non-overlapping microscopic fields
from each section by Image-Pro Plus 6.2 software. In brief, regions of interest (ROI)
were manually selected in a color cube based manner.

ACC
EPTE
D M
ANU
SCR
IPT
ACCEPTED MANUSCRIPT
10
Statistical analysis
All values are presented as means ± standard deviation (SD). Statistical analysis
was performed using SPSS software, with differences between groups being assessed
by analysis of variance and considered statistically significant when P<0.05.
Results
Characterization of the calcitriol loaded collagen membrane scaffolds
Using SEM, the surface of the original collagen membrane was observed to be
highly porous with good pore interconnectivity (Fig. 1A). For the
polydopamine-coated collagen membrane, pore number was significantly reduced,
owing to coverage of the polydopamine layer over the surface pores (Fig. 1B). SEM
micrographs of the CAL/DOP/CM group revealed a certain amount of calcitriol, in
spherical shape, distributed on the surface of the collagen membrane (Fig. 1C).
Histological and statistical analysis for bone regeneration
At 1 week post-surgery, new bone nodules emerged. Although fibrous tissue
predominantly occupied the bone defect in the control group, a few areas of new bone
were found scattered above the soft tissue (Fig. 2A). In the CM group, remnants of the
collagen membrane were arranged dispersedly, with some new bone nodules detected
at the bottom of the cavity (Fig. 2B). The DOP/CM group showed a more intact and
regularly assembled collagen membrane, as well as small bone nodules (Fig. 2C). The
CAL/DOP/CM group presented the least amount of collagen membrane residue and
the largest amount of new bone nodules. Furthermore, some regions of the collagen
membrane had been partially replaced by new bone (Fig. 2D).

ACC
EPTE
D M
ANU
SCR
IPT
ACCEPTED MANUSCRIPT
11
At 2 weeks post-surgery, a meshwork of thin bone with large bone marrow
cavities could be identified. An abundance of fibrous tissue remained in the control
group (Fig. 2E). However, the collagen membrane had been mostly degraded in the
CM group and obvious new bone was detected (Fig. 2F). The DOP/CM group was
characterized by considerable collagen residues, with new bone adjacent to the defect
edge (Fig. 2G). The CAL/DOP/CM group showed the most abundant amount of new
bone, occupying the defect up to two-thirds the height of the previous alveolar ridge
(Fig. 2H).
Masson’s trichrome staining was further performed to distinguish between
connective tissue, mineralized and non-mineralized bone. Generally, primary bone
mainly made up of collagen type I is stained into blue while highly mineralized bone
is stained into red. At week 1, control group showed blue-stained fibrous tissue and a
few new bone nodules (red) (Fig. 3A). In the CM and DOP/CM group, the new bone
was mostly blue stained while CAL/DOP/CM group emerged a certain amount of
mineralized bone (red) (Fig. 3B-D). At week 2, primary bone nodules (blue)
distributed dispersedly among fibrous tissue in control group (Fig. 3E). In other three
groups, primary bone (blue) and highly mineralized bone (red) both were observed
(Fig. 3F-H). They staggered mutually forming in an ordered structure especially in
CAL/DOP/CM group (Fig. 3H).
At 4 weeks post-surgery, the new bone featured thicker trabeculae. The control
group showed fibrous healing characteristics, namely fibrous tissue invasion into the
defect with new bone failing to attach to the pre-existing bone (Fig. 4A). In the other
three groups, new bone almost covered the entire bone defect area (Fig. 4B–D). As
shown in Fig. 4, the CM and DOP/CM groups contained more woven bone (Fig.4B
and C) while the CAL/DOP/CM group showed highly mineralized bone matrix (Fig.

ACC
EPTE
D M
ANU
SCR
IPT
ACCEPTED MANUSCRIPT
12
4D).
At 8 weeks post-surgery, new bone was highly mineralized and featured compact
bone. Fibrous tissue was still visible in the control group (Fig.4E). The bone marrow
cavities in the CM and DOP/CM groups were much larger than those in the
CAL/DOP/CM group, which were closer to the cortical bone (Fig. 4F–H).
Masson’s trichrome staining showed consistent results with HE staining at week
4. In control group, abundant fibrous tissue (blue) occupied in the defect area (Fig.
5A). CM group and DOP/CM group showed defects containing more primary bone
(blue) and less mineralized bone (red) (Fig. 5B,C). Conversely, there was mass of
mineralized bone (red) with tiny primary bone (blue) in CAL/DOP/CM group (Fig.
5D). At week 8, control group still showed blue stained fibrous tissue (Fig. 5E).
CAL/DOP/CM group has more mineralized bone (red) and less primary bone (blue)
compared with CM group and DOP/CM group (Fig. 5F-H).
As shown in the schema graph (Fig. 6A), the control group displayed fibrous
healing characteristics, the DOP/CM group was characterized by delayed degradation
of the collagen membrane, while the CAL/DOP/CM group showed accelerated
collagen membrane degradation and bone regeneration. Statistical analysis revealed
that at week 1, newly formed bone volume increased successively in the order of the
control, DOP/CM, CM and CAL/DOP/CM groups. At week 2, newly formed bone
volume in the CAL/DOP/CM group was highest, followed by the CM group. Control
and DOP/CM groups were the lowest with no significant difference between each
other. At weeks 4 and 8, the control group exhibited the least amount of bone, with no
significant difference in bone volume among the other three groups (Fig. 6B).
Immunohistochemical examination and statistical analysis

ACC
EPTE
D M
ANU
SCR
IPT
ACCEPTED MANUSCRIPT
13
At 1 week post-surgery, the control, CM and DOP/CM groups all showed large
populations of TRAP positive osteoclasts adjacent to the bottom of the defects.
However, osteoclasts were barely detected in the CAL/DOP/CM group (Fig. 7A–D).
In contrast, abundant ALP positive osteoblasts were found in the same area in the
CAL/DOP/CM group, with fewer numbers seen in other three groups (Fig. 7A–D).
Consistently, there were significantly more Runx2-positive osteoblasts in the
CAL/DOP/CM group compared with the other three groups (Fig. 7E–H).
Not surprisingly, at 2 weeks post-surgery, the number of osteoclasts decreased in
all groups. However, there remained a faint inhibitory effect when calcitriol was used
(Fig. 7I–L). As expected, Runx2 was highly expressed only in the CAL/DOP/CM
group (Fig. 7M–P) while ALP immunoreactivity was faintly detected in all groups
(Fig. 7I–L).
Statistic analysis further revealed that ALP and Runx2 immunoreactivity both
increased dramatically after calcitriol administration at week 1 (Fig. 8A and B). At
week 2, a significant increase in Runx2 was observed in the CAL/DOP/CM group
with no significant difference in ALP activity among groups (Fig. 8A and B). In
addition, there was reduction in osteoclastogenesis, which could be attributed to
calcitriol administration both at weeks 1 and 2 (Fig. 8C).
At week 4, the control and CAL/DOP/CM groups showed uniform collagen
structure (Fig. 9A and D). Uneven immunostaining was seen in the CM and DOP/CM
groups (Fig. 9B and C). Cement lines showed obvious OPN immunoreactivity in the
CM and DOP/CM groups (Fig. 9F and G). However, in the control and
CAL/DOP/CM groups, OPN was only faintly expressed (Fig. 9E and H). At week 8,
similar results were observed. Newly formed bone was more highly matured in the
control and CAL/DOP/CM groups compared with the other two groups (Fig. 9I–P).

ACC
EPTE
D M
ANU
SCR
IPT
ACCEPTED MANUSCRIPT
14
Discussion
In this study, we investigated the potential role of calcitriol in bone tissue
engineering using a rat mandibular bone defect model. At the early stage of bone
defect repair (weeks 1 and 2), calcitriol exhibited positive effects on bone remodeling
by reducing osteoclastogenesis and promoting osteoblast differentiation, leading to
accelerated bone regeneration. During the subsequent bone maturation phase (weeks 4
and 8), calcitriol regulated collagen maturation and promoted bone mineralization
(Fig. 10).
Tratrate-resistant acid phosphatase (TRAP) , as a glycosylated metalloenzyme, is
highly expressed in osteoclasts and widely used as a specific marker of osteoclasts in
bone (Cole and Walters. 1987; Minkin. 1982). With respect to osteoclastogenesis, our
results that calcitriol inhibit TRAP-positive osteoclasts are consistent with some
previous studies, in which calcitriol inhibited osteoclast formation. Specifically,
calcitriol was reported to suppress the expression of c-fos and nuclear factor of
activated T cells c1 (NFATc1), both of which are key regulators of osteoclast
formation (Sakai et al. 2009; Takasu et al. 2006). However, other studies have shown
that calcitriol strongly induces both receptor activator of NF-B ligand (RANKL) and
macrophage colony-stimulating factor (M-CSF) in vitro and in vivo and induces bone
resorption (Sato et al. 2007; Suda et al. 1999). More interestingly, in
thyroparathyroidectomized rats infused with parathyroid hormone (PTH),
systemically administered calcitriol inhibited PTH-induced bone resorption at a
physiological dose and stimulated bone resorption at a toxic dose (Ueno et al. 2003).

ACC
EPTE
D M
ANU
SCR
IPT
ACCEPTED MANUSCRIPT
15
Taken together, calcitriol may function by both stimulating and inhibiting bone
resorption depending on the dose and methods of administration. Furthermore,
calcitriol has opposing effects in osteoblast differentiation, which may be due to
differences in animal species and reagents (Hicok et al. 1998; Li et al. 2008; Lohmann
et al. 2000). For example, Li et al. evaluated the in vitro effects of 1,25(OH)2D3 on
osteoblasts from three different species and found that in rat cells, the active vitamin
D increased cell proliferation, inhibited cell apoptosis and increased alkaline
phosphatase (ALP) activity. In mouse cells, however, it initiated cell apoptosis and
inhibited ALP activity. In human cells, although cell proliferation and apoptosis was
while ALP activity was enhanced (Li et al. 2008). In the current study, calcitriol
enhanced both ALP- and Runx2-immunoreactivity at week 1 and promoted Runx2
expression at week 2. These findings further confirm that ALP and Runx2 exhibit
differential expression during osteoblast differentiation (Reichert et al. 2013).
Enhanced bone regeneration observed at weeks 1 and 2 may be attributed to
inhibited bone resorption and induced bone formation. The detailed mechanisms of
calcitriol in bone formation is still the ongoing controversy. Phex, a marker of the
mature osteoblasts, and Runx2, a key regulator of bone formation in vivo, were
regulated by calcitriol (Drissi et al. 2002; Ecarot et al. 1999). A recent study showed
that calcitriol binding to VDR regulates BMP2 gene expression by binding to the
BMP2 promoter region and inducing DNA methylation and histone modification (Fu
et al. 2013). Another noteworthy problem is that during normal bone remodeling,
bone deposition usually depends on the accurate coordination between osteoclasts and

ACC
EPTE
D M
ANU
SCR
IPT
ACCEPTED MANUSCRIPT
16
osteoblasts (Wright et al. 2005). The current study identified a discrepancy between
inhibition of osteoclastogenesis and promotion of osteogenic differentiation. This
phenomenon can perhaps be partly interpreted by our previous findings (Saito et al.
2013), in which eldecalcitol, a second-generation vitamin D analog, induced focal
bone formation in absence of osteoclasts, referred to as ‘bone mini modeling’.
Another possible explanation for this discrepancy is that osteoclastogenesis occurred
at earlier stages which escaped our detection. The specific mechanism for this remains
to be elucidated.
A previous study demonstrated that calcitriol directly regulates collagen
cross-linking in an osteoblast-like cell line (MC3T3-E1), likely by up-regulating gene
expression of specific LH and LOX enzymes (Nagaoka et al. 2008). This partially
explains our findings, that calcitriol induced collagen maturation and promoted bone
mineralization at weeks 4 and 8. The mechanism of calcitriol in promoting bone
mineralization is complex. In addition to its systemic effects on mineral ion
homeostasis, calcitriol has been proposed to have a direct effect on mineralization
through the VDR on skeletal cells (Chen et al. 2013). Furthermore, calcitriol can
stimulate mineralization of human osteoblast cultures (Miyahara et al. 2002; van Driel
et al. 2006) and modulate expression of various mineralization-related genes, such as
OCN (BGLAP) and OPN (SPP1) (Barthel et al. 2007; Shen and Christakos 2005).
However, high-dose 1,25(OH)2D3 is reported to inhibit osteoblast mineralization in
vitro (Yamaguchi and Weitzmann 2012).
The physiological effect of calcitriol is closely related to its dosage. We chose

ACC
EPTE
D M
ANU
SCR
IPT
ACCEPTED MANUSCRIPT
17
the dose at 5×10-4
g/ml referring to published studies which decare using calcitriol at
10-4
g/ml level with scaffolds showed good potential to implement osteoinduction and
osteogenesis in vivo (Bosetti et al. 2014; Yoon et al. 2007). In our present study,
calcitriol showed more significant effect on osteogenesis at week-1 and week-2 than
week-4 and week-8 owing to dose attenution. Therefore, it needs more study to find
more appropriate dose of calcitriol.
In the current study, another interesting phenomenon was that polydopamine
delayed the degradation of collagen membrane. As described in a previously
published paper, polydopamine coating is stable and not easily degraded. It acts as an
impermeable cap, preventing the release of Ca2+
and PO43-
from BCP scaffolds and
slowing their degradation (Jia et al. 2013). It is therefore not surprising that
polydopamine formed a stable layer adherent on the surface of the collagen membrane,
protecting it from degradation. However, the exact mechanism by which calcitriol
accelerates degradation of the collagen membrane requires further study.
Calcitriol is liposoluble and it could be very hard to immoblize it on scaffolds
without modification method. That is the reason why we use poly-dopamine coating
instead of using calcitriol only. In our present study, CAL/DOP/CM group exhibited
positive effects on bone remodeling and mineralization while DOP/CM group showed
no silimar action. It gives a strong evidence that calcitriol only can promote
osteogenesis. From another aspect, polydopamine delayed the degradation of collagen
membrane while calcitriol accelerated it completely offseting the effect of
polydopamine. Therefore, we could have observed promoted osteogenesis and more
obviously accelerated degradation of collagen membrane if there was CAL only group.

ACC
EPTE
D M
ANU
SCR
IPT
ACCEPTED MANUSCRIPT
18
The follow-up study to confirm this phenomenon depends on some new technologies.
For GBR, a barrier film is often placed covering the bone defect to create a
relatively closed environment and prevent fibrous tissue invading into the bone defect.
Based on this theory, it is not difficult to understand that the control group showed
fibrous healing characteristics in our study. In addition, the collagen carrier for
calcitriol was mostly resorbed by 2 weeks, but further studies are necessary to identify
materials that can provide longer-term scaffolding, to support continuing bone
formation in a mechanically challenging environment (Khan et al. 2012).
Conclusions
Our results suggest that local administration of calcitriol accelerates bone
formation and promotes bone maturation, thus providing important experimental data
for the favorable use of calcitriol for bone regeneration. However, considering the
bidirectional actions of calcitriol, more effort needs to be made towards finding the
best characteristics of calcitriol for bone tissue engineering purposes.
Acknowledgements
This study was partially supported by the National Nature Science Foundation of
China (grant No. 81271965; 81470719; 81311140173) and Specialized Research
Fund for the Doctoral Program of Higher Education (grant No. 20120131110073) to
Li M.

ACC
EPTE
D M
ANU
SCR
IPT
ACCEPTED MANUSCRIPT
19
References
Arrabal PM, Visser R, Santos-Ruiz L, Becerra J, Cifuentes M (2013) Osteogenic
molecules for clinical applications: improving the BMP-collagen system. Biol Res 46:
421-429
Avenell A, Gillespie WJ, Gillespie LD, O'Connell DL (2005) Vitamin D and vitamin
D analogues for preventing fractures associated with involutional and
post-menopausal osteoporosis. Cochrane Database Syst Rev 20: CD000227
Barthel TK, Mathern DR, Whitfield GK, Haussler CA, Hopper HA 4th, Hsieh JC,
Slater SA, Hsieh G, Kaczmarska M, Jurutka PW, Kolek OI, Ghishan FK, Haussler
MR (2007) 1,25-Dihydroxyvitamin D3/VDR-mediated induction of FGF23 as well as
transcriptional control of other bone anabolic and catabolic genes that orchestrate the
regulation of phosphate and calcium mineral metabolism. J Steroid Biochem Mol Biol
103: 381-388
Bosetti M, Boccafoschi F, Leigheb M, Cannas MF (2007) Effect of different growth
factors on human osteoblasts activities: a possible application in bone regeneration
fortissue engineering. Biomol Eng 24: 613-618
Bosetti M, Fusaro L, Nicolì E, Borrone A, Aprile S, Cannas M (2013) Poly-L-lactide
acid-modified scaffolds for osteoinduction and osteoconduction. J Biomed Mater Res
A. doi: 10.1002/jbm.a.35016
Chen J, Dosier CR, Park JH, De S, Guldberg RE, Boyan BD, Schwartz Z (2013)
Mineralization of three-dimensional osteoblast cultures is enhanced by the interaction
of 1α,25-dihydroxyvitamin D3 and BMP2 via two specific vitamin D receptors. J

ACC
EPTE
D M
ANU
SCR
IPT
ACCEPTED MANUSCRIPT
20
Tissue Eng Regen Med. doi: 10.1002/term.1770
Chiapasco M, Zaniboni M (2009) Clinical outcomes of GBR procedures to correct
peri-implant dehiscences and fenestrations: a systematic review. Clin Oral Implants
Res 20. Suppl 4:113-123
Cole AA, Walters LM (1987)
Tartrate-resistant acid phosphatase in bone and cartilage following decalcification and
cold-embedding in plastic. J Histochem Cytochem 35: 203-206.
Cottam E, Hukins DW, Lee K, Hewitt C, Jenkins MJ (2009) Effect of sterilisation by
gamma irradiation on the ability of polycaprolactone (PCL) to act as a scaffold
material. Med Eng Phys 31: 221-226
Cui J, Yan Y, Such GK, Liang K, Ochs CJ, Postma A, Caruso F (2012)
Immobilization and intracellular delivery of an anticancer drug using mussel-inspired
polydopamine capsules. Biomacromolecules 13: 2225-2228
de Nijs RN, Jacobs JW, Algra A, Lems WF, Bijlsma JW (2004) Prevention and
treatment of glucocorticoid-induced osteoporosis with active vitamin D3 analogues: a
review withmeta-analysis of randomized controlled trials including organ
transplantation studies. Osteoporos Int 15: 589-602
Drissi H, Pouliot A, Koolloos C, Stein JL, Lian JB, Stein GS, van Wijnen AJ (2002)
1,25-(OH)2-vitamin D3 suppresses the bone-related Runx2/Cbfa1 gene promoter. Exp
Cell Res 274: 323–333
Drzewiecki KE, Parmar AS, Gaudet ID, Branch JR, Pike DH, Nanda V, Shreiber DI
(2014) Methacrylation Induces Rapid, Temperature-Dependent, Reversible

ACC
EPTE
D M
ANU
SCR
IPT
ACCEPTED MANUSCRIPT
21
Self-Assembly of Type-I Collagen. Langmuir. [Epub ahead of print]
Ecarot B, Desbarats M (1999) 1,25-(OH)2D3 down-regulates expression of Phex,
a marker of the mature osteoblast. Endocrinology 140: 1192–1199
Ehnert S, Zhao J, Pscherer S, Freude T, Dooley S, Kolk A, Stöckle U, Nussler
AK, Hube R (2012) Transforming growth factor β1 inhibits bone morphogenic
protein (BMP)-2 and BMP-7 signaling via upregulation of Ski-related novel protein N
(SnoN): possible mechanism for the failure of BMP therapy? BMC Med 10: 101
Feldman D, Pike JW, Glorieux F (2005) Vitamin D. Elsevier Academic Press,
Burlington
Fu B, Wang H, Wang J, Barouhas I, Liu W, Shuboy A, Bushinsky DA, Zhou D, Favus
MJ (2013) Epigenetic regulation of BMP2 by 1,25-dihydroxyvitamin D3 through
DNA methylation and histone modification. PLoS One 8: e61423
Hämmerle CH, Jung RE, Feloutzis A (2002) A systematic review of the survival of
implants in bone sites augmented with barrier membranes (guided bone regeneration)
in partially edentulous patients. J Clin Periodontol 29 Suppl 3: 226-231
Han F, Zhou F, Yang X, Zhao J, Zhao Y, Yuan X (2014) A pilot study of conically
graded chitosan-gelatin hydrogel/PLGA scaffold with dual-delivery of TGF-β1
and BMP-2 for regeneration of cartilage-bone interface. J Biomed Mater Res B Appl
Biomater. doi: 10.1002/jbm.b.33314
Harrison JR, Petersen DN, Lichtler AC, Mador AT, Rowe DW, Kream BE (1989)
1,25-Dihydroxyvitamin D3 inhibits transcription of type I collagen genes in the rat
osteosarcoma cell line ROS17/2.8. Endocrinology 125: 327-333

ACC
EPTE
D M
ANU
SCR
IPT
ACCEPTED MANUSCRIPT
22
Hicok KC, Thomas T, Gori F, Rickard DJ, Spelsberg TC, Riggs BL (1998)
Development and characterization of conditionally immortalized osteoblast precursor
cell lines from human bone marrow stroma. J Bone Miner Res 13: 205-217
Inada M, Tsukamoto K, Hirata M, Takita M, Nagasawa K, Miyaura C (2008) Novel
vitamin D3 analogs, 1alpha, 25(OH)2D(3)-26, 23-lactam (DLAMs), antagonize bone
resorption viasuppressing RANKL expression in osteoblasts. Biochem Biophys Res
Commun 372: 434-439
Jang JW, Lee B, Han CW, Lee IW, Lee HB, Khang G (2003) Preparation and
characterization of ipriflavone loaded PLGA scaffolds for tissue engineered bone.
Polymer 27: 226
Jia X, Ma ZY, Zhang GX, Hu JM, Liu ZY, Wang HY, Zhou F (2013) Polydopamine
film coated controlled-release multielement compound fertilizer based on
mussel-inspired chemistry. J Agric Food Chem 61: 2919-2924
Jung RE, Glauser R, Schärer P, Hämmerle CH, Sailer HF, Weber FE (2003) Effect of
rhBMP-2 on guided bone regeneration in humans. Clin Oral Implants Res 14:
556-568
Khan SN, Toth JM, Gupta K, Glassman SD, Gupta MC (2012) Early and Mid Term
Histological Events During Single Level Posterolateral Intertransverse Process Fusion
With Rhbmp-2/Collagen Carrier and a Ceramic Bulking Agent In a Non-Human
Primate Model: Implications for BoneGraft Preparation. J Spinal Disord Tech. [Epub
ahead of print]
Kim SH, Park KS, Kim SH, Jang JW, Han CH, Kim MS (2006) Effects of SIS/PLGA

ACC
EPTE
D M
ANU
SCR
IPT
ACCEPTED MANUSCRIPT
23
porous scaffolds and muscle-derived stem cell on the formation of tissue engineered
bone. Polymer 30: 14
Lee H, Dellatore SM, Miller WM, Messersmith PB (2007) Mussel-inspired surface
chemistry for multifunctional coatings. Science 318: 426-430
Li M, Hasegawa T, Hogo H, Tatsumi S, Liu Z, Guo Y, Sasaki M, Tabata C, Yamamoto
T, Ikeda K, Amizuka N (2013) Histological examination on osteoblastic activities in
the alveolar bone of transgenic mice with induced ablation of osteocytes. Histol
Histopathol 28: 327-335
Li Y, Bäckesjö CM, Haldosén LA, Lindgren U (2008) Species difference exists in the
effects of 1alpha,25(OH)(2)D(3) and its analogue
2-methylene-19-nor-(20S)-1,25-dihydroxyvitamin D(3) (2MD) on osteoblastic cells. J
Steroid Biochem Mol Biol 112: 110-116
Lohmann CH, Bonewald LF, Sisk MA, Sylvia VL, Cochran DL, Dean DD, Boyan BD,
Schwartz Z (2000) Maturation state determines the response of osteogenic cells to
surface roughness and 1,25-dihydroxyvitamin D3. J Bone Miner Res 15: 1169-1180
Lynge ME, van der Westen R, Postma A, Städler B (2011) Polydopamine--a
nature-inspired polymer coating for biomedical science. Nanoscale 3: 4916-4928
Minkin C (1982) Bone acid phosphatase: tartrate-resistant acid phosphatase as a
marker of osteoclast function. Calcif. Tissue Int 34: 285–290.
Miyahara T, Simoura T, Osahune N, Uchida Y, Sakuma T, Nemoto N, Kozakai A,
Takamura T, Yamazaki R, Higuchi S, Chiba H, Iba K, Sawada N (2002) A highly
potent 26,27-Hexafluoro-1a,25-dihydroxyvitamin D3 on calcification in

ACC
EPTE
D M
ANU
SCR
IPT
ACCEPTED MANUSCRIPT
24
SV40-transformed human fetal osteoblastic cells. Calcif Tissue Int 70: 488-495
Muzio G, Martinasso G, Baino F, Frairia R, Vitale-Brovarone C, Canuto RA (2014)
Key role of the expression of bone morphogenetic proteins in increasing the
osteogenic activity of osteoblast-like cells exposed to shock waves and seeded on
bioactive glass-ceramic scaffolds for bone tissue engineering. J Biomater Appl. doi:
10.1177/0885328214541974
Nagai M, Hayakawa T, Makimura M, Yoshinari M (2006) Fibronectin immobilization
using water-soluble carbodiimide on poly-L-lactic acid for enhancing initial fibroblast
attachment. J Biomater Appl 21: 33-47
Nagaoka H, Mochida Y, Atsawasuwan P, Kaku M, Kondoh T, Yamauchi M (2008)
1,25(OH)2D3 regulates collagen quality in an osteoblastic cell culture system.
Biochem Biophys Res Commun 377: 674-678
Nawata H, Soen S, Takayanagi R, Tanaka I, Takaoka K, Fukunaga M, Matsumoto T,
Suzuki Y, Tanaka H, Fujiwara S, Miki T, Sagawa A, Nishizawa Y, Seino Y (2005)
Guidelines on the management and treatment of glucocorticoid-induced osteoporosis
of the Japanese Society for Bone and Mineral Research (2004). J Bone Miner Metab
23: 105-109
Noda M, Vogel RL, Craig AM, Prahl J, DeLuca HF, Denhardt DT (1990)
Identification of a DNA sequence responsible for binding of the
1,25-dihydroxyvitamin D3 receptor and 1,25-dihydroxyvitamin D3 enhancement of
mouse secreted phosphoprotein 1 (SPP-1 or osteopontin) gene expression. Proc Natl
Acad Sci USA 87: 9995-9999

ACC
EPTE
D M
ANU
SCR
IPT
ACCEPTED MANUSCRIPT
25
Nyström D, Malmström E, Hult A, Blakey I, Boyer C, Davis TP, Whittaker MR (2010)
Biomimetic surface modification of honeycomb films via a "grafting from" approach.
Langmuir 26: 12748-12754
Oryan A, Alidadi S, Moshiri A, Bigham-Sadegh A (2014) Bone morphogenetic
proteins: A powerful osteoinductive compound with non-negligible side effects and
limitations. Biofactors 40: 459-481
Reichert JC, Schmalzl J, Prager P, Gilbert F, Quent VM, Steinert AF, Rudert M, Nöth
U (2013) Synergistic effect of Indian hedgehog and bone morphogenetic protein-2
gene transfer to increase the osteogenicpotential of human mesenchymal stem cells.
Stem Cell Res Ther 4: 105
Saito H, Takeda S, Amizuka N (2013) Eldecalcitol and calcitriol stimulates 'bone
minimodeling,' focal bone formation without prior bone resorption, inrat trabecular
bone. J Steroid Biochem Mol Biol 136: 178-182.
Saito K, Tanaka A, Taniguchi T, Suda T, Miyamoto T, Toyama Y (2009) 1-Alpha,
25-dihydroxy vitamin D3 inhibits osteoclastogenesis through IFN-beta-dependent
NFATc1 suppression. J Bone Miner Metab 27: 643-652
Sakai S, Takaishi H, Matsuzaki K, Kaneko H, Furukawa M, Miyauchi Y, Shiraishi A,
Sato M, Nakamichi Y, Nakamura M, Sato N, Ninomiya T, Muto A, Nakamura H,
Ozawa H, Iwasaki Y, Kobayashi E, Shimizu M, DeLuca HF, Takahashi N, Udagawa
N (2007) New 19-nor-(20S)-1alpha,25-dihydroxyvitamin D3 analogs strongly
stimulate osteoclast formation both in vivo and in vitro. Bone 40: 293-304
Shen Q, Christakos S (2005) The vitamin D receptor, Runx2, and the Notch signaling

ACC
EPTE
D M
ANU
SCR
IPT
ACCEPTED MANUSCRIPT
26
pathway cooperate in the transcriptional regulation of osteopontin. J Biol Chem 280:
40589-40598
Shiraishi A, Higashi S, Masaki T, Saito M, Ito M, Ikeda S, Nakamura T (2002) A
comparison of alfacalcidol and menatetrenone for the treatment of bone loss in an
ovariectomized rat model of osteoporosis. Calcif Tissue Int 71: 69-79
Sigurdsson TJ, Nguyen S, Wikesjo UM (2001) Alveolar ridge augmentation with
rhBMP-2 and bone-to-implant contact in induced bone. Int J Periodontics Restorative
Dent 21: 461-473
Suda T, Takahashi N, Udagawa N, Jimi E, Gillespie MT, Martin TJ (1999)
Modulation of osteoclast differentiation and function by the new members of the
tumor necrosis factor receptor and ligand families. Endocr Rev 20: 345-357
Suda T, Ueno Y, Fujii K, Shinki T (2003) Vitamin D and bone. J Cell Biochem 88:
259-266
Takasu H, Sugita A, Uchiyama Y, Katagiri N, Okazaki M, Ogata E, Ikeda K (2006)
c-Fos protein as a target of anti-osteoclastogenic action of vitamin D, and synthesis of
new analogs. J Clin Invest 116: 528-535
Tatakis DN, Koh A, Jin L, Wozney JM, Rohrer MD, Wikesjö UM (2002) Peri-implant
bone regeneration using recombinant human bone morphogenetic protein-2 in a
canine model: a dose-response study. J Periodontal Res 37:93-100
Ueno Y, Shinki T, Nagai Y, Murayama H, Fujii K, Suda T (2003) In vivo
administration of 1,25-dihydroxyvitamin D3 suppresses the expression of RANKL
mRNA in bone of thyroparathyroidectomized rats constantly infused with PTH. J Cell

ACC
EPTE
D M
ANU
SCR
IPT
ACCEPTED MANUSCRIPT
27
Biochem 90: 267-277
Vallières K, Petitclerc E, Laroche G (2007) Covalent grafting of fibronectin onto
plasma-treated PTFE: influence of the conjugation strategy on fibronectin biological
activity. Macromol Biosci 7: 738-745
van Driel M, Koedam M, Buurman CJ, Roelse M, Weyts F, Chiba H, Uitterlinden AG,
Pols HA, van Leeuwen JP (2006) Evidence that both 1alpha,25-dihydroxyvitamin D3
and 24-hydroxylated D3 enhance human osteoblast differentiation and mineralization.
J Cell Biochem 99: 922-935
Wang Z, Wang K, Lu X, Li M, Liu H, Xie C, Meng F, Jiang O, Li C, Zhi W (2014)
BMP-2 encapsulated polysaccharide nanoparticle modified biphasic calcium
phosphate scaffolds for bone tissueregeneration. J Biomed Mater Res A. doi:
10.1002/jbm.a.35282
Wright LM, Maloney W, Yu X, Kindle L, Collin-Osdoby P, Osdoby P (2005) Stromal
cell-derived factor-1 binding to its chemokine receptor CXCR4 on precursor cells
promotes the chemotactic recruitment, development and survival of human osteoclasts.
Bone 36: 840-853
Xia Y, Boey F, Venkatraman SS (2010) Surface modification of poly (L-lactic acid)
with biomolecules to promote endothelialization. Biointerphases 5: FA32-40
Yamaguchi M, Weitzmann MN (2012) High dose 1,25(OH)2D3 inhibits osteoblast
mineralization in vitro. Int J Mol Med 29: 934-938
Yang K, Lee JS, Kim J, Lee YB, Shin H, Um SH, Kim JB, Park KI, Lee H, Cho SW
(2012) Polydopamine-mediated surface modification of scaffold materials for human

ACC
EPTE
D M
ANU
SCR
IPT
ACCEPTED MANUSCRIPT
28
neural stem cell engineering. Biomaterials 33: 6952-6964
Yoon SJ, Park KS, Kim MS, Rhee JM, Khang G, Lee HB (2007)
Repair of diaphyseal bone defects with calcitriol-loaded PLGA scaffolds and marrow
stromal cells. Tissue Eng 13: 1125-1133.
Yoshizawa T, Handa Y, Uematsu Y, Takeda S, Sekine K, Yoshihara Y, Kawakami T,
Arioka K, Sato H, Uchiyama Y, Masushige S, Fukamizu A, Matsumoto T, Kato S
(1997) Mice lacking the vitamin D receptor exhibit impaired bone formation, uterine
hypoplasia and growth retardation after weaning. Nat Genet 16: 391-396
Figure Captions
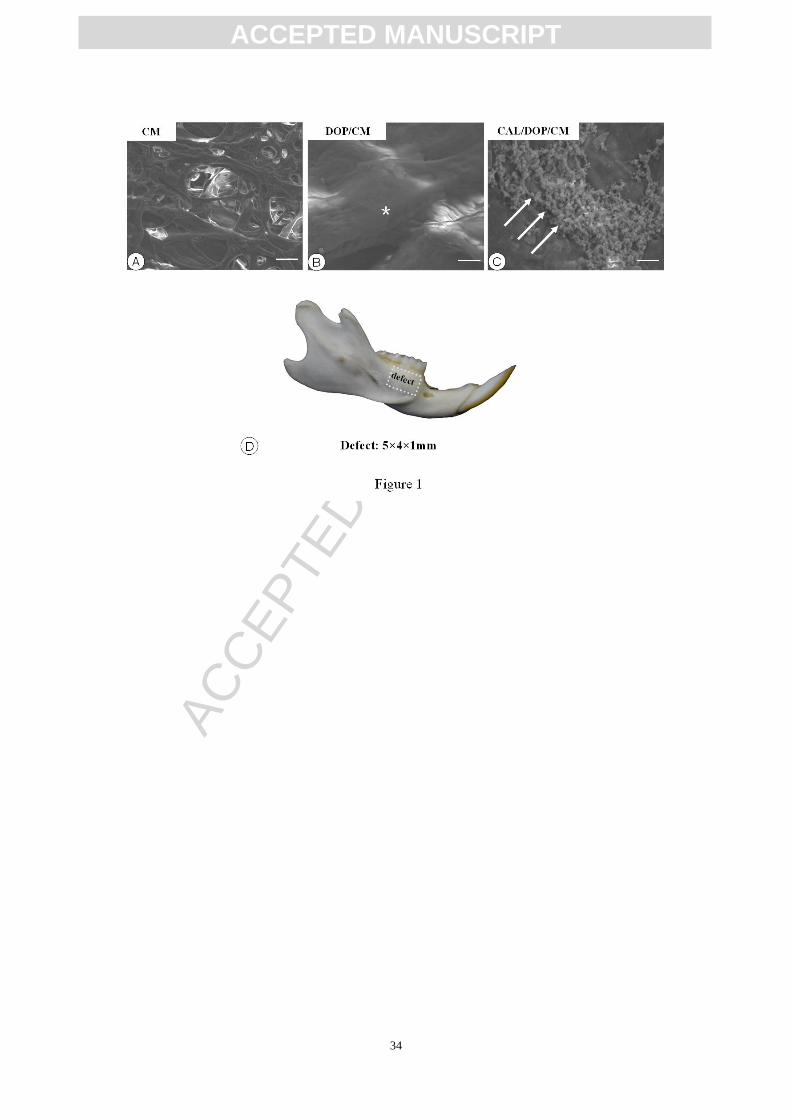
Fig. 1 SEM micrographs and diagram representing mandibular bone defect
SEM micrographs of the original collagen membrane (A), polydopamine-modified
membrane (B) and calcitriol-polydopamine composite membrane (C). The diagram
represents the surgical window in the buccal surface of the first molar (D). The
surface of the original collagen membrane was highly porous with good pore
interconnectivity (A). The polydopamine-coated collagen membrane displayed
coverage of the polydopamine layer over the membrane surface (asterisk, B).
Calcitriol granules were observed distributed on the surface of the scaffolds (white
arrows, C). Bar, A–C: 1μm
Fig. 2 Hematoxylin and eosin staining of bone defects at weeks 1 and 2
Low-magnification images of bone defects at weeks 1 (A-D) and 2 (E-H). In the
control group, fibrous tissue occupied the bone defect with a small number of new

ACC
EPTE
D M
ANU
SCR
IPT
ACCEPTED MANUSCRIPT
29
bone nodules (A). In the CM group, remnants of the collagen membrane were
arranged dispersedly and some new bone nodules were detected at the bottom of the
cavity (B). More intact and regularly assembled collagen membrane, as well as small
bone nodules, were seen in the DOP/CM group (C). The CAL/DOP/CM group
showed the least collagen membrane residue and largest amount of new bone (D). At
week 2, there was still much fibrous tissue in the control group (E). The collagen
membrane was degraded completely in the CM group and obvious new bone was
detected (F). The DOP/CM group was characterized by considerable collagen residue
while new bone was limited adjacent to the defect edge (G). The CAL/DOP/CM
group showed the most amount of new bone, almost filling the entire bone defect (H).
CB: cortical bone; NB: newly formed bone; M: collagen membrane; F: fibrous tissue.
Bar, A–H: 250 μm.
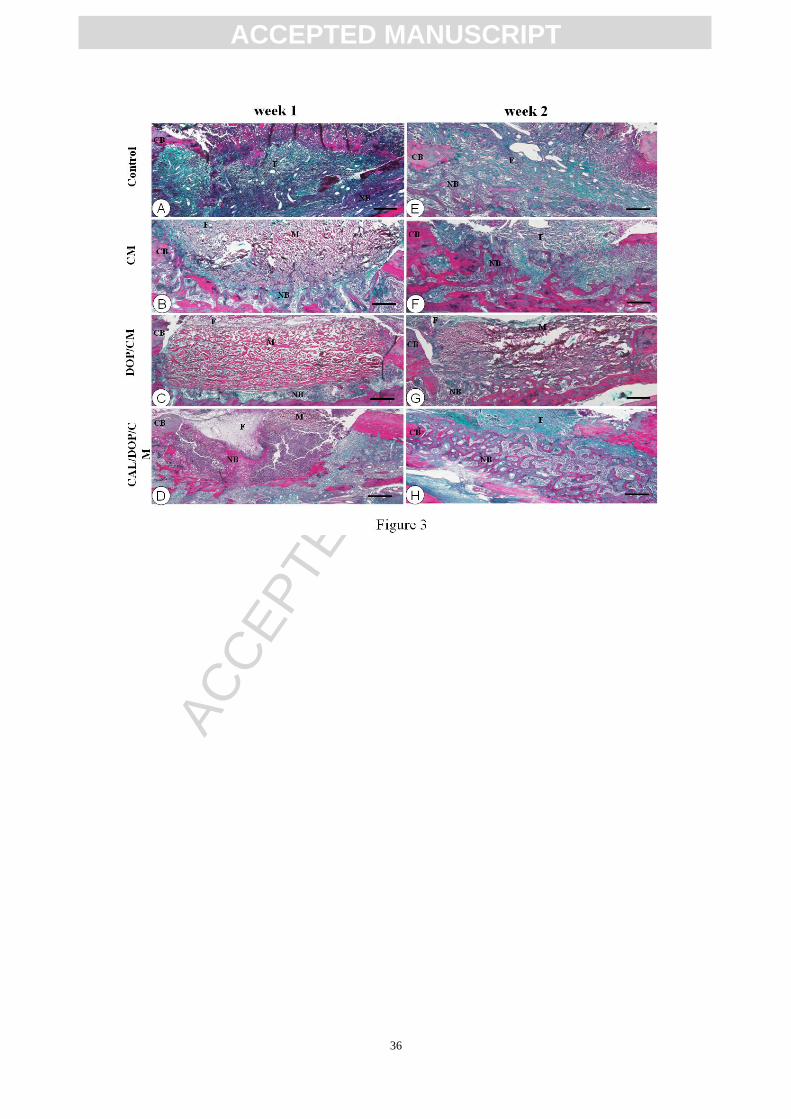
Fig. 3 Masson’s trichrome staining of bone defects at weeks 1 and 2
Overview images of bone defects at weeks 1 (A-D) and 2 (E-H) showed by Masson’s
trichrome staining. At week 1, control group showed blue-stained fibrous tissue and a
few new bone nodules (red) (A). In the CM and DOP/CM group, the new bone was
mostly blue stained while CAL/DOP/CM group emerged a certain amount of
mineralized bone (red) (B-D). At week 2, primary bone nodules (blue) distributed
dispersedly among fibrous tissue in control group (E). In other three groups, primary
bone (blue) and highly mineralized bone (red) both were observed (F-H). They
staggered mutually forming in an ordered structure especially in CAL/DOP/CM group
(H). CB: cortical bone; NB: newly formed bone; M: collagen membrane; F: fibrous
tissue. Bar, A–H: 250 μm.

ACC
EPTE
D M
ANU
SCR
IPT
ACCEPTED MANUSCRIPT
30
Fig. 4 Hematoxylin and eosin staining of bone defects at weeks 4 and 8
Low-magnification images of bone defects at weeks 4 (A-D) and 8 (E-H). At week 4,
the control group showed fibrous healing characteristics, with new bone failing to
attach to the pre-existing bone because of invading fibrous tissue (A). In the other
three groups, new bone almost covered the entire bone defect area: the CM and
DOP/CM groups presented woven bone, while the CAL/DOP/CM group showed
highly mineralized bone (B-D). At week 8, new bone was highly mineralized and
featured compact bone. Fibrous tissue was still visible in the bone defect of the
control group (E). The bone marrow cavities were much larger in the CM and
DOP/CM groups than those in CAL/DOP/CM group, which was closer to the cortical
bone (F-H). CB: cortical bone; NB: newly formed bone; F: fibrous tissue. Bar, A–H:
250 μm.
Fig. 5 Masson’s trichrome staining of bone defects at weeks 4 and 8
Overview images of bone defects at weeks 4 (A-D) and 8 (E-H). In control group,
abundant fibrous tissue (blue) occupied in the defect area (A). CM group and
DOP/CM group showed defects containing more primary bone (blue) and less
mineralized bone (red) (B,C). Conversely, there was mass of mineralized bone (red)
with tiny primary bone (blue) in CAL/DOP/CM group (D). At week 8, control group
still showed blue stained fibrous tissue (E). CAL/DOP/CM group has more
mineralized bone (red) and less primary bone (blue) compared with CM group and
DOP/CM group (F-H).
Fig. 6 Schema graph of different groups and statistical analysis
The schema graph illustrates the results of the histological analysis (A). Statistical

ACC
EPTE
D M
ANU
SCR
IPT
ACCEPTED MANUSCRIPT
31
analysis for newly formed bone volume (B). The control group showed fibrous
healing characteristics, the DOP/CM group was filled by collagen membrane, and the
CAL/DOP/CM group featured less collagen membrane and more new bone (A). At
week 1, following the sequence of less to more bone volume, the order was control,
DOP/CM, CM and CAL/DOP/CM groups. At week 2, the newly formed bone volume
in the CAL/DOP/CM group was the highest, followed by the CM group. Control and
DOP/CM groups were the lowest, with no significant differences between each other.
At week 4 and 8, the control group exhibited the least amount of bone while there was
no significant difference among the other three groups (B). (n=6; * P< 0.05; ** P<
0.001), Bars indicate ±SD.
Fig. 7 Effect of calcitriol on osteoblasts and osteoclasts at the early stage of bone
repair
Double staining for ALP (brown) and TRAP (red) in bone defect at weeks 1 (A-D)
and 2 (I-L). Immunohistochemistry for Runx2 (brown) in the same visual field at
weeks 1 (E-H) and 2 (M-P). At week 1, the control, CM and DOP/CM groups all
showed large populations of TRAP positive osteoclasts adjacent to the bottom of the
defects, which were barely detected in the CAL/DOP/CM group (A-D). In contrast,
abundant ALP positive osteoblasts (asterisk) were found in the same area in the
CAL/DOP/CM group, with few observed in other three groups (A-D). Consistently,
there were significantly more Runx2 positive cells (asterisk) in the CAL/DOP/CM
group (H). At week 2, the number of osteoclasts decreased in all groups, but a visible
inhibitory effect remained when calcitriol was used (L). Runx2 was highly expressed
(asterisk) only in the CAL/DOP/CM group (P) while ALP immunoreactivity was
faintly detected in all groups (I-L). oc: osteoclast. Bars, A–H: 25 μm.

ACC
EPTE
D M
ANU
SCR
IPT
ACCEPTED MANUSCRIPT
32
Fig. 8 Statistical analysis for ALP and RUNX2 activity and TRAP positive
osteoclast number
Statistical analysis for ALP activity (A), Runx2 activity (B) and TRAP positive cell
number (C). ALP and Runx2 immunoreactivity both increased dramatically after
calcitriol administration at week 1 (A, B). At week 2, Runx2 still showed a significant
increase in CAL/DOP/CM group while there was no significant difference in ALP
activity (A, B). In addition, osteoclast number was significantly reduced in the group
of calcitriol administration both at weeks 1 and 2 (C). (n=6; ** P< 0.001). Bars
indicate ± SD.
Fig. 9 Effect of calcitriol on bone maturation and mineralization
The immunohistochemistry for Type I collagen (brown) at weeks 4 (A–D) and 8 (I–L).
Immunohistochemistry for OPN (brown) in the same visual field at weeks 4 (E–H)
and 8 (M–P). At week 4, the control and CAL/DOP/CM groups showed uniform
collagen structure (asterisk) (A, D). Uneven immunostaining was seen in the CM and
DOP/CM groups (B, C). Cement lines showed obvious OPN immunoreactivity in the
CM and DOP/CM groups (black arrows, F, G). In the control and CAL/DOP/CM
groups, OPN was only faintly expressed (E, H). At week 8, similar results were
observed, newly formed bone in the control and CAL/DOP/CM groups was more
highly matured than in the other two groups (I–P). Bar, A–H: 25 μm.
Fig. 10 Hypothetical scheme illustrating the influence of calcitriol on restoration
of mandibular bone defects
During initial bone remodeling, calcitriol inhibits osteoclastogenesis and promotes

ACC
EPTE
D M
ANU
SCR
IPT
ACCEPTED MANUSCRIPT
33
osteoblast differentiation. During subsequent bone maturation, calcitriol regulates
collagen maturation and promotes bone mineralization.
ACC
EPTE
D M
ANU
SCR
IPT
ACCEPTED MANUSCRIPT
34

ACC
EPTE
D M
ANU
SCR
IPT
ACCEPTED MANUSCRIPT
35
ACC
EPTE
D M
ANU
SCR
IPT
ACCEPTED MANUSCRIPT
36

ACC
EPTE
D M
ANU
SCR
IPT
ACCEPTED MANUSCRIPT
37

ACC
EPTE
D M
ANU
SCR
IPT
ACCEPTED MANUSCRIPT
38

ACC
EPTE
D M
ANU
SCR
IPT
ACCEPTED MANUSCRIPT
39

ACC
EPTE
D M
ANU
SCR
IPT
ACCEPTED MANUSCRIPT
40

ACC
EPTE
D M
ANU
SCR
IPT
ACCEPTED MANUSCRIPT
41

ACC
EPTE
D M
ANU
SCR
IPT
ACCEPTED MANUSCRIPT
42

ACC
EPTE
D M
ANU
SCR
IPT
ACCEPTED MANUSCRIPT
43

ACC
EPTE
D M
ANU
SCR
IPT
ACCEPTED MANUSCRIPT
44
Table 1
Experimental groups

ACC
EPTE
D M
ANU
SCR
IPT
ACCEPTED MANUSCRIPT
45
Highlights (for review)
More information on collagen material was added in the revised manuscript.
Masson-Goldner trichrome stain was performed for histomorphometry.
More specific information on calcitriol was supplemented in Discussion section.
The MOD of ALP and Runx2 was explained in more detail.
The inhibition of osteoclastogenesis was described more accurately in the second
paragraph of the discussion.




















