
Cuttlefish rely on both polarized light and landmarks for orientation
Upload
independentCategory
view
3download
0
1,2 3 2 1
1 C.C.MAR, Universidade do Algarve, Faculdade de Ciencias do Mar e do Ambiente, Faro, Portugal; 2 Departamento de
Biologıa Animal, Facultad de Biologıa, Universidad de La Laguna, La Laguna, Tenerife, Canary Islands, Spain; 3 Centro
Oceanografico de Canarias, Instituto Espanol de Oceanografia, Tenerife, Spain
The present work reports a characterization of mean wet
weight and moisture, the lipid class and fatty acid (FA)
composition from the total lipids (TL), of both culture and
wild eggs of the cuttlefish, Sepia officinalis, throughout the
embryonic development. Additionally, reproductive data,
such as the number of spawnings, number and mean weight
of eggs and duration of spawning period of cultured cuttle-
fish is provided. Both types of eggs were similar in mean wet
weight, moisture content, TL content and lipid composition
throughout embryonic development. Females from the cul-
tured group spawned 13 times and laid 8654 eggs in 64 days,
with a mean weight of 0.607 ± 0.179 g. A sex ratio of 1.57
(11$ for 7#) promoted an individual fecundity of 787 eggs/$
(the biggest until now on our culture facilities), which might
be related to increased bottom areas. The TL increased with
day/stage of embryonic development (P < 0.05) only in the
cultured egg group. However, no differences were found on
TL between culture and wild eggs at the same day/stage
(P > 0.05). Eggs displayed predominant levels of phospha-
tidylcholine, phosphatidylethanolamine (PE), cholesterol and
triacylglycerol at the end of embryonic development. Polar
and neutral lipids of both eggs groups remained consistently
proportional (�50% for each lipid fraction) and a significant
increase (P < 0.05) was observed in phosphatidylserine, PE
and free FA throughout the embryonic development. In
either egg type and day, 16:0, 18:0, 20:5n-3 and 22:6n-3
accounted for approximately 70 g Kg)1 of all FA and satu-
rated and n-3 totals seemed to have the same proportion in
the cuttlefish eggs. The present results suggest that lipids are
not used as energetic substrate but as structural components
in cuttlefish egg.
KEY WORDSKEY WORDS: cuttlefish, eggs, embryonic development, life cy-
cles, lipid composition, n-3 highly unsaturated fatty
acids
Received 20 September 2007, accepted 4 January 2008
Correspondence: Antnio V. Sykes, C.C.MAR, Universidade do Algarve,
Faculdade de Ciencias do Mar e do Ambiente, Campus de Gambelas, 8000-
810 Faro, Portugal. E-mail: [email protected]
Cephalopods have been defined as aquaculture candidates for
the past 20 years (Vaz-Pires et al. 2004; Sykes et al. 2006a),
and this was first acknowledged by the inclusion of a chapter
dedicated to cuttlefish in the �Bases Biologiques et ecologiques
de l�aquaculture� de Barnabe (1996). The constraints in devel-
oping cephalopod culture technology are many and have been
described by Sykes et al. (2006a) for the European cuttlefish,
Sepia officinalis. The development of culture technology for
cephalopod species faces similar problems to those found for
fish in the early years. Like in fish, one of the most important
challenges in aquaculture is larval or first stage nutrition,
whereas in most species a limited knowledge of their nutri-
tional requirements creates bottlenecks. Theoretically, wild
eggs have all the nutrients needed for both correct development
and homeostasis of the egg and larvae until their first exoge-
nous feeding. Therefore, an estimation of the use of endoge-
nous nutrients provided by the yolk reserves during embryonic
development should be used as an approach to nutritional
requirement of the embryonic and post-hatching-phase (Sar-
gent 1995; Mourente & Vazquez 1996; Rainuzzo et al. 1997;
Cejas et al. 2004). This nutrient composition and its use during
embryonic development, however, seem to be species-specific,
2009 15; 38–53. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .
doi: 10.1111/j.1365-2095.2008.00566.x
. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .
� 2008 Blackwell Publishing Ltd
No claim to original US government works
Aquaculture Nutrition

leading to differences in both quantity and quality of larvae
used during that stage (Mourente & Vazquez 1996). Energy
metabolism in fish during embryonic development is based on
protein, carbohydrates, free amino acids and lipids (Finn 1994;
Finn et al. 1995, 1996; Rønnestad et al. 1998). The choice for
the substrate varies according to species and between different
stages of embryonic development and depends on ambient/
culture conditions, physiological events and energy demands
(Rainuzzo 1993; Sargent 1995; Mourente & Vazquez 1996;
Rainuzzo et al. 1997; Almansa et al. 1999). For instance, the
lipid fraction of some fish eggs has been described as substrate
for energy metabolism and as structural components in
membrane biogenesis (Tocher & Sargent 1984; Falk-Petersen
et al. 1986; Cejas et al. 2004).
According to Boletzky (1987a), in cephalopods, �the
embryonic phase tends to be a black box, succinctly called
the egg stage�. Although there is information regarding the
distinct phases of the embryonic development in cuttlefish
(Naef 1928; Lemaire 1970), data regarding the composition,
physiology and metabolism during this phase are scarce or
inexistent. Besides, it has been assumed that cuttlefish
metabolism is mainly protein and amino acid driven (Lee
1994) and the lipid fraction represent <2% of their body
weight (Boucaud-Camou 1990); thus neglecting the study of
its lipid composition. Nevertheless, some studies have already
been conducted regarding lipid nutrition in cephalopods.
To determine the ideal lipid profile during the early life of
cephalopods, Navarro & Villanueva (2000) studied the lipid
composition of the mature ovary and late eggs and wild
hatchlings and juveniles. Additionally, the same authors also
studied the lipid composition of natural preys used in suc-
cessful cultures. From these data they stated that cephalo-
pods in their early stages of growth show a high requirement
for polyunsaturated fatty acids (PUFA), with docosahexoe-
noic acid (DHA, 22:6n-3) representing 20–30% of the total
FA of the total lipids (TL) of cuttlefish, squid and octopus
hatchlings. The importance of PUFA in larval fish nutrition
has been thoroughly researched in the last 20–30 years
(Watanabe 1993; Watanabe & Kiron 1994; Sargent et al.
1999). According to Sargent et al. (1999), teleost eggs are
generally rich in highly unsaturated FA of the n-3 series (n-3
HUFA), mainly eicosapentaenoic acid (EPA, 20:5n-3) and
DHA. Furthermore, there is strong evidence that, in fish, n-3
HUFA are crucial to female fecundity, to embryo and to
early larval development, growth and survival (Watanabe &
Kiron 1994; Sargent 1995; Silversand et al. 1996). Like in
fish, in cuttlefish n-3 HUFA are also involved in the organ-
ogenesis of structural components such as brain (Dumont
et al. 1992, 1994), and probably will have a similar degree of
importance in the eye development (Boyle et al. 2001), and
will be precursors of physiological active molecules such as
eicosanoids (Sargent 1995).
In fish, yolk reserves are obtain from vitelogenin, a phos-
pholipoprotein which are synthesized in the liver and trans-
ported to the gonads to be included in the eggs. An increase
in the liver lipid content during gonadal maturity has been
observed in fish that is related to the previous (Almansa et al.
1999). Cephalopods share some similar facts, such as, the
digestive gland (which has a similar role to the liver in fish)
usually yields a large volume of lipids during the sexual
maturity in similar way to their gonads, and these are rich in
n-3 HUFA, especially EPA and DHA (Rosa et al. 2005).
According to Blanchier & Boucaud-Camou (1984) and
Blanchier et al. (1986), there is an increase of cholesterol
(CHO) and phospholipids content of the digestive gland in
cuttlefish females at sexual maturity. However, these were
proved to be not significant. The same authors also noted a
slight increase and a marked increase in CHO and phos-
pholipids respectively, in the ovary at this time that might be
related to yolk and steroid synthesis. They suggested that the
increase in CHO must be related to membrane synthesis
during gametogenesis (Blanchier & Boucaud-Camou 1984),
while the increase in phospholipids is, according to Fujii
(1960), related to the synthesis of the vitellus. These data
suggest the importance of lipids in the egg reserves, probably
to be used along the embryonic development as energy sub-
strate and/or structural components.
In cuttlefish, the influence of temperature in the TL and LC
yolk content during the embryonic development was studied
byBouchaud&Galois (1990). The results obtained suggest the
importance of lipid content in the embryogenesis. According
to these authors, TL represents about 14% of dry weight in
eggs, despite of egg size and incubation temperature range.
However, these eggs were from the French-English Channel
population cuttlefish, displaying higher mean wet weight and
longer embryonic development times than those commonly
found in southern Portugal (for a review on this problematic,
see Perez-Losada et al. 1999, 2002, 2007; Sykes et al. 2006a).
These differences in the reproduction biology could imply
differences in the biochemical composition of the eggs. It is
therefore of interest to investigate the lipid composition of
ovulated eggs from different cuttlefish populations.Moreover,
the study of the FA composition of wild and cultured eggs is of
extreme importance as egg composition may be altered by the
breeder�s diet (Morehead et al. 2001). Therefore, a comparison
between wild and cultured cuttlefish could be useful to obtain
further insight into the nutritional requirements of embryos
and yolk sac hatchlings of the species.
. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .
� 2008 Blackwell Publishing Ltd Aquaculture Nutrition 15; 38–53
No claim to original US government works

In the present study, egg mean wet weight and moisture,
TL, lipid classes and FA of wild and cultured eggs were
examined throughout the embryonic phase until hatching.
Also, reproduction data regarding the number of spawnings,
number and mean weight of eggs and duration of spawning
period of cultured cuttlefish was studied.
The eggs used in the present study were laid in the laboratory
by both culture- and wild-captured adult individuals. Culture
spawners (n = 18; mean weight of 147.05 ± 55.09 g) were
selected from a captive F2 generation, already cultured in the
laboratory (using exclusively grass shrimp as prey), while
already reproducing wild spawners (n = 18; mean weight of
156.56 ± 51.40 g) were captured in the Ria Formosa La-
goon (south Portugal), using a small bottom trawl (6 m wide;
0.5 m high; 10-mm mesh size at the cod end) for 10 min haul
in shallow water (�4 m). Both groups were placed in white
400-L rectangular plastic tanks (with a bottom area of
1.2 · 1.0 cm2) of a culture system similar to that previously
described by Domingues et al. (2001). These tanks were
placed in a low-disturbance room of CCMAR�s Ramalhete
Field Station (Faro, south Portugal). Water flow was of
12 L h)1 for both tanks. Temperature and salinity were
measured daily for the culture-spawners tank. Oxygen was
kept at optimal values that ranged approximately 100%
saturation. Mean water temperature was 19.5 ± 1.09 �C,while mean salinity was 38.8 ± 0.42 g L)1. The conditions
of the used spawning system ensured an excellent water
quality from a physical, chemical and bacteriological point of
view. Cultured cuttlefish were fed live adult grass shrimp
(0.090 ± 0.022 g; n = 200) throughout their life cycle. Food
was captured from ponds surrounding the research facility
and supplied ad libitum.
Although the objective of the current work was to study
differences between wild and cultured eggs, only cultured
cuttlefish were conditioned before spawning. Wild specimens
were captured during the spawning season, so it was proba-
ble that the wild broodstock had started spawning before
their capture. Therefore, cultured spawners were the only
group where growth and reproduction data were acquired.
Data were collected at the beginning of the experiment, the
start of spawning and at the end of the experiment (when the
last animal died) and were used to calculate: (i) mean wet
weight (MWW); (ii) mean instantaneous growth rate (IGR)
(%body weight day)1) = (LnW2)LnW1)/t*100, where W2
and W1 are the final and initial weight respectively, Ln the
natural logarithm and t the number of days of the
time-period; (iii) total biomass (TB) =P
weight (g) of the
animals at time of death; (iv) duration of the spawning period
(DSP, days); (v) TB Increase During Spawning Period
(TBIDSP) = (TB-BBSP)/DSP, where TB is defined above,
BBSP is biomass at the beginning of the spawning period; (vi)
sex ratio = number of $/#; and (vii) individual fecundity
(F) = eggs/$. The DSP as well as the amount of times that
egg laying occurred (batches) were also recorded.
After being conditioned in the present conditions, adult-
cultured sepia took about 18 days to mature and start
spawning. Captured individuals started spawning on the first
day after being collected and placed under laboratory con-
ditions. This provided the excellent opportunity to obtain
wild eggs in the laboratory without culture interference, such
as feeding. When spawning started, eggs were collected from
both wild and culture tanks. Collection was made using a
green-plastic-rectangular-supportive net (1 cm2 holes), sus-
pended inside both wild and cultured spawners tanks. These
nets were checked daily and if eggs were present (day spawn),
they were carefully removed (as freshly laid eggs are very soft
and gelatinous) and individualized. After this, all of them
were accounted for and weighed (100%, if n < 100 or 50% if
n > 100). Also, eggs were separated based on shape and
colour. Black oval eggs were considered to be viable, while
eggs of any other shape and/or colour were considered non-
viable and discarded. Total egg numbers from both viable
and non-viable were accounted for and the percentage of
non-viable egg presence in each batch of spawned eggs
determined. The viability of both black oval eggs from both
wild and culture spawners was then investigated using 500
eggs of each group of breeders throughout the embryonic
development in the incubation tanks and at the conditions
described below.
Embryonic development of both wild and cultured eggs
(n = 1000 eggs each) was carried out in 250 L bowl-shaped
incubation tanks with clean natural seawater in a similar
seawater system described by Domingues et al. (2001) and in
the egg stage conditions described by Sykes et al. (2006b).
Water flow was of 12 L h)1. Both group of eggs were incu-
bated at a mean temperature of 21.7 ± 0.93 �C and mean
salinity of 39.2 ± 0.21 g L)1. Light intensity was 200 lux, at
the top of the water column of the incubation tanks, and
photoperiod resembled natural geographical conditions in
spring (14 hD : 10 hN).
. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .
� 2008 Blackwell Publishing Ltd Aquaculture Nutrition 15; 38–53
No claim to original US government works

While going through the embryonic development and at
every 5 days, 50 eggs from both groups were weighed and used
formoisture determination.Also, at every 10 days, samples for
biochemical analysis from both wild and cultured eggs were
taken. From these samplings, on bothwild and culture groups,
15 eggs were used to determine moisture and another 70 eggs
for later lipid determinations.
Moisture content was determined from individual egg samples
using the method of Horwitz (1980). TL was extracted with
chloroform : methanol (2 : 1 v/v) containing 0.01% of butyl-
ated hydroxytoluene (BHT) as antioxidant (Christie 1982).
The organic solventwas evaporated under a streamof nitrogen
and the lipid content determined gravimetrically. Lipid classes
were separated by one dimensional double development high-
performance thin layer chromatography (HPTLC), using
methyl acetate/isopropanol/chloroform/methanol/0.25% (w/
v) KCl (25 : 25 : 25 : 10 : 9 by vol.) as the polar solvent sys-
tem, and hexane/diethyl ether/glacial acetic acid (80 : 20 : 2 by
vol.) as the neutral solvent system. Lipid classes were quanti-
fied by charring with a copper acetate reagent followed by
calibrated scanning densitometry using a Shimadzu CS-
9001PC dual wavelength flying spot scanner (Olsen & Hen-
derson 1989). TL extracts were subjected to acid-catalysed
transmethylation for 16 h at 50 �C, using 1 mL of toluene and
2 mLof 1%sulphuric acid (v/v) inmethanol. The resultantFA
methyl esters (FAME) were purified by TLC, and visualized
under UV light with 2¢,7¢-dichlorofluorescein in 98% (v/v)
methanol, containing 0.01% BHT (Christie 1982). Prior to
transmethylation 19:0 was added to TL as internal standard.
FAME were separated and quantified by using a Shimadzu
GC-14A gas chromatograph equipped with a flame ionization
detector (250 �C) and a fused silica capillary column Supe-
lcowaxTM 10 (30 m · 0.32 mm I.D.) (Supelco, Bellefonte,
USA). Helium was used as carrier gas and samples were
applied by on-column injection at an initial temperature of
50 �C. During each analysis, the oven was programmed to rise
from 60 to 150 �C at a rate of 39 �C min)1, and then to a final
temperature of 215 �C at 2.5 �C min)1. Individual FAME
were identified by reference to authentic standards and to a
well-characterized fish oil (PUFA-3, Biosigma, Barcelona,
Spain).
Lipid samples, for each sampled day triplicate, were com-
posed of amaximum of three eggs andweight dependable. TL,
LC and FAME are expressed as lg/egg. Individual means
were calculated on a dry weight basis. BHT, potassium
chloride, potassium bicarbonate, 2¢,7¢-dichlorofluorescein
were supplied by Sigma Chemical Co. (St Louis, MO, USA).
TLC (20 · 20 cm · 0.25 mm) and HPTLC (10 · 10 cm ·0.15 mm) plates, precoated with silica gel (without fluorescent
indicator) were purchased from Macheren-Nagel (Duren,
Germany). All organic solvents for GC used were of reagent
grade and were purchased from Panreac (Barcelona, Spain).
Results are presented as means ± standard deviations (SD)
for triplicate experiments. The data were checked for normal
distribution with the one-sample Kolmogorov–Smirnoff test
(Zar 1999) as well as for homogeneity of variances with the
Levene�s test (Zar 1999) and, when necessary, arcsin trans-
formation was performed. When a normal distribution and/
or homogeneity of the variances were not achieved, non-
parametric tests were used.
A Kruskal–Wallis test (Zar 1999) was used to compare
differences in MWW between the different egg batches ob-
tained from cultured spawners, followed by a post hoc
Dunn�s test (Zar 1999) to determine which were statistically
different. A Mann–Whitney test (Zar 1999) was used to
compare differences among culture and wild eggs in terms of
MWW and moisture throughout the embryonic develop-
ment. A one-way ANOVAANOVA (Zar 1999), comparing the eggs at
different stages but within the same type of egg, was per-
formed for TL, LC and FA profiles, followed up by a pos-
teriori post hoc multi-comparison Scheffe test (Zar 1999) to
determine differences within samples. When lipid data failed
normal distribution and/or homogeneity, first arcsin trans-
formation (Fowler et al., 2002) was applied and the same
parametric test was used. After this and if data still failed to
comply to either normality and/or homogeneity, then a
Kruskal–Wallis test (Zar 1999) was used, followed up by a
posteriori post hoc multi-comparison Games–Howell test
(Zar 1999) to determine differences. A student t-test (Zar
1999) was applied to determine differences of TL, LC and
FAME between egg types at the same egg stage. If, after
arcsin transformation, data still did not comply with nor-
mality and/homogeneity, then a Mann–Whitney test (Zar
1999) was used. In all statistical tests used, P < 0.05 was
considered statistically different.
After 18 days of being set apart in the breeding tank, cul-
tured spawners MWW was 183.84 ± 72.47 g, from which #
. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .
� 2008 Blackwell Publishing Ltd Aquaculture Nutrition 15; 38–53
No claim to original US government works
displayed a MWW of 172.46 ± 45.54 g and $ of
191.08 ± 86.81 g. From these, the maximum weight re-
corded was of 325.6 g from a $, while the minimum was of
82.6 g from a #.
The cultured group of 18 cuttlefish intermittent-spawned
13 times and laid 8654 eggs in 64 days, with a mean weight
of 0.607 ± 0.179 g (Table 1). From these, 418 eggs were
considered non-viable, which corresponded to 4.83% of to-
tal egg numbers. All egg batches were found to be statisti-
cally different between them (P < 0.05). However, Dunn�s
test revealed that not every batch was different from the
other. Table 1 presents a resume of the cultured egg data
obtained, and those that were not statistically different be-
tween them (P > 0.05). We also noticed that the amount of
eggs laid over the spawning period had a normal distribu-
tion. Both wild and cultured egg batches used for the study
of weight evolution during embryogenesis displayed MWW
(g) of 0.627 ± 0.131 and 0.766 ± 0.310 respectively, at
spawning.
After the 64 DSP, culture cuttlefish MWW was
293.63 ± 131.08 g, from which # displayed a MWW of
280.69 ± 128.40 g and $ of 301.87 ± 138.29 g. From these,
the maximum and minimum weights recorded were 513.20 g
and 102.30 g, both from #s. Mean IGR during the spawning
period was 0.79% BW day)1 for # and 0.74% BW day)1 for
$. At the end of reproduction a TB of 5285.33 g was
achieved, which corresponded to a TBIDSP of 24.70% dur-
ing that time interval. A sex ratio of 1.57 (11$ for 7 #), in this
type of culture conditions, promoted an individual fecundity
of 787 eggs/$.
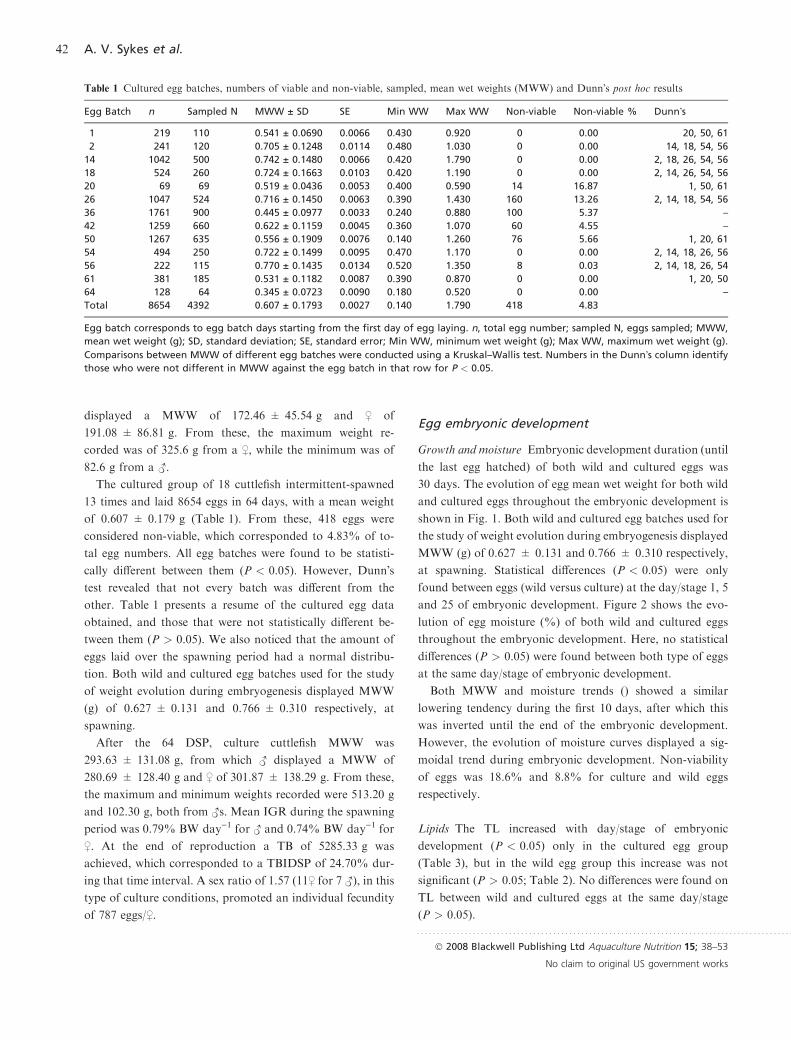
Growth and moisture Embryonic development duration (until
the last egg hatched) of both wild and cultured eggs was
30 days. The evolution of egg mean wet weight for both wild
and cultured eggs throughout the embryonic development is
shown in Fig. 1. Both wild and cultured egg batches used for
the study of weight evolution during embryogenesis displayed
MWW (g) of 0.627 ± 0.131 and 0.766 ± 0.310 respectively,
at spawning. Statistical differences (P < 0.05) were only
found between eggs (wild versus culture) at the day/stage 1, 5
and 25 of embryonic development. Figure 2 shows the evo-
lution of egg moisture (%) of both wild and cultured eggs
throughout the embryonic development. Here, no statistical
differences (P > 0.05) were found between both type of eggs
at the same day/stage of embryonic development.
Both MWW and moisture trends () showed a similar
lowering tendency during the first 10 days, after which this
was inverted until the end of the embryonic development.
However, the evolution of moisture curves displayed a sig-
moidal trend during embryonic development. Non-viability
of eggs was 18.6% and 8.8% for culture and wild eggs
respectively.
Lipids The TL increased with day/stage of embryonic
development (P < 0.05) only in the cultured egg group
(Table 3), but in the wild egg group this increase was not
significant (P > 0.05; Table 2). No differences were found on
TL between wild and cultured eggs at the same day/stage
(P > 0.05).
Table 1 Cultured egg batches, numbers of viable and non-viable, sampled, mean wet weights (MWW) and Dunn�s post hoc results
Egg Batch n Sampled N MWW ± SD SE Min WW Max WW Non-viable Non-viable % Dunn�s
1 219 110 0.541 ± 0.0690 0.0066 0.430 0.920 0 0.00 20, 50, 61
2 241 120 0.705 ± 0.1248 0.0114 0.480 1.030 0 0.00 14, 18, 54, 56
14 1042 500 0.742 ± 0.1480 0.0066 0.420 1.790 0 0.00 2, 18, 26, 54, 56
18 524 260 0.724 ± 0.1663 0.0103 0.420 1.190 0 0.00 2, 14, 26, 54, 56
20 69 69 0.519 ± 0.0436 0.0053 0.400 0.590 14 16.87 1, 50, 61
26 1047 524 0.716 ± 0.1450 0.0063 0.390 1.430 160 13.26 2, 14, 18, 54, 56
36 1761 900 0.445 ± 0.0977 0.0033 0.240 0.880 100 5.37 –
42 1259 660 0.622 ± 0.1159 0.0045 0.360 1.070 60 4.55 –
50 1267 635 0.556 ± 0.1909 0.0076 0.140 1.260 76 5.66 1, 20, 61
54 494 250 0.722 ± 0.1499 0.0095 0.470 1.170 0 0.00 2, 14, 18, 26, 56
56 222 115 0.770 ± 0.1435 0.0134 0.520 1.350 8 0.03 2, 14, 18, 26, 54
61 381 185 0.531 ± 0.1182 0.0087 0.390 0.870 0 0.00 1, 20, 50
64 128 64 0.345 ± 0.0723 0.0090 0.180 0.520 0 0.00 –
Total 8654 4392 0.607 ± 0.1793 0.0027 0.140 1.790 418 4.83
Egg batch corresponds to egg batch days starting from the first day of egg laying. n, total egg number; sampled N, eggs sampled; MWW,
mean wet weight (g); SD, standard deviation; SE, standard error; Min WW, minimum wet weight (g); Max WW, maximum wet weight (g).
Comparisons between MWW of different egg batches were conducted using a Kruskal–Wallis test. Numbers in the Dunn�s column identify
those who were not different in MWW against the egg batch in that row for P < 0.05.
. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .
� 2008 Blackwell Publishing Ltd Aquaculture Nutrition 15; 38–53
No claim to original US government works
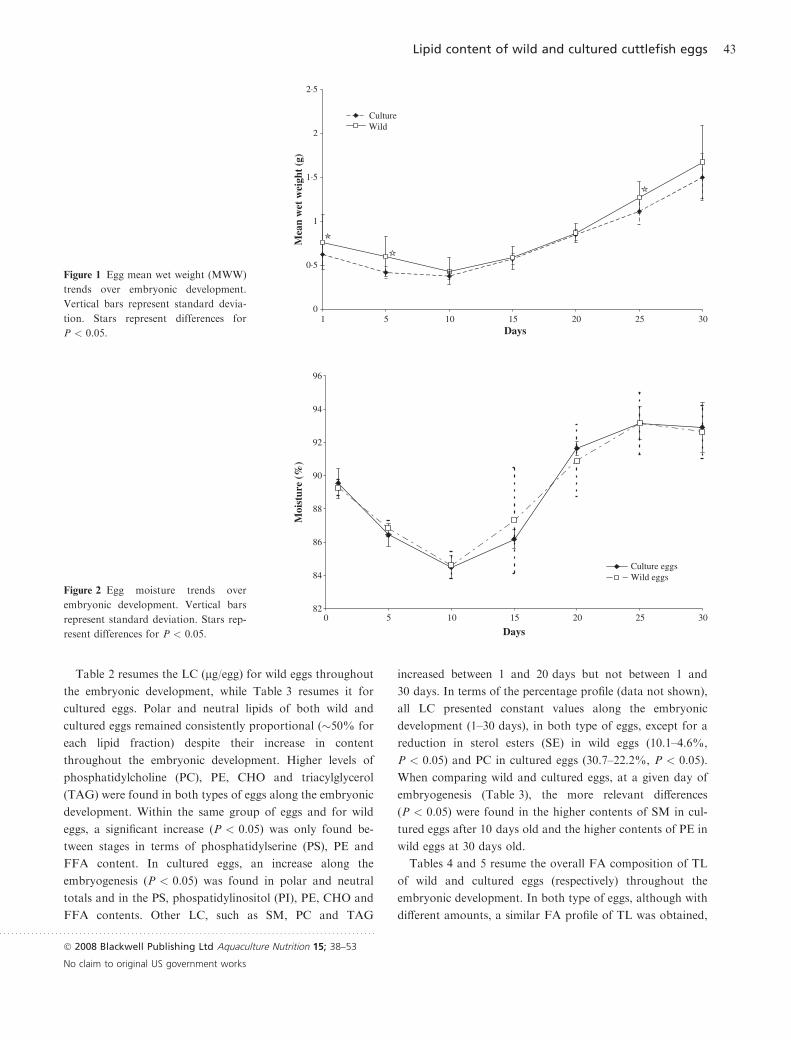
Table 2 resumes the LC (lg/egg) for wild eggs throughout
the embryonic development, while Table 3 resumes it for
cultured eggs. Polar and neutral lipids of both wild and
cultured eggs remained consistently proportional (�50% for
each lipid fraction) despite their increase in content
throughout the embryonic development. Higher levels of
phosphatidylcholine (PC), PE, CHO and triacylglycerol
(TAG) were found in both types of eggs along the embryonic
development. Within the same group of eggs and for wild
eggs, a significant increase (P < 0.05) was only found be-
tween stages in terms of phosphatidylserine (PS), PE and
FFA content. In cultured eggs, an increase along the
embryogenesis (P < 0.05) was found in polar and neutral
totals and in the PS, phospatidylinositol (PI), PE, CHO and
FFA contents. Other LC, such as SM, PC and TAG
increased between 1 and 20 days but not between 1 and
30 days. In terms of the percentage profile (data not shown),
all LC presented constant values along the embryonic
development (1–30 days), in both type of eggs, except for a
reduction in sterol esters (SE) in wild eggs (10.1–4.6%,
P < 0.05) and PC in cultured eggs (30.7–22.2%, P < 0.05).
When comparing wild and cultured eggs, at a given day of
embryogenesis (Table 3), the more relevant differences
(P < 0.05) were found in the higher contents of SM in cul-
tured eggs after 10 days old and the higher contents of PE in
wild eggs at 30 days old.
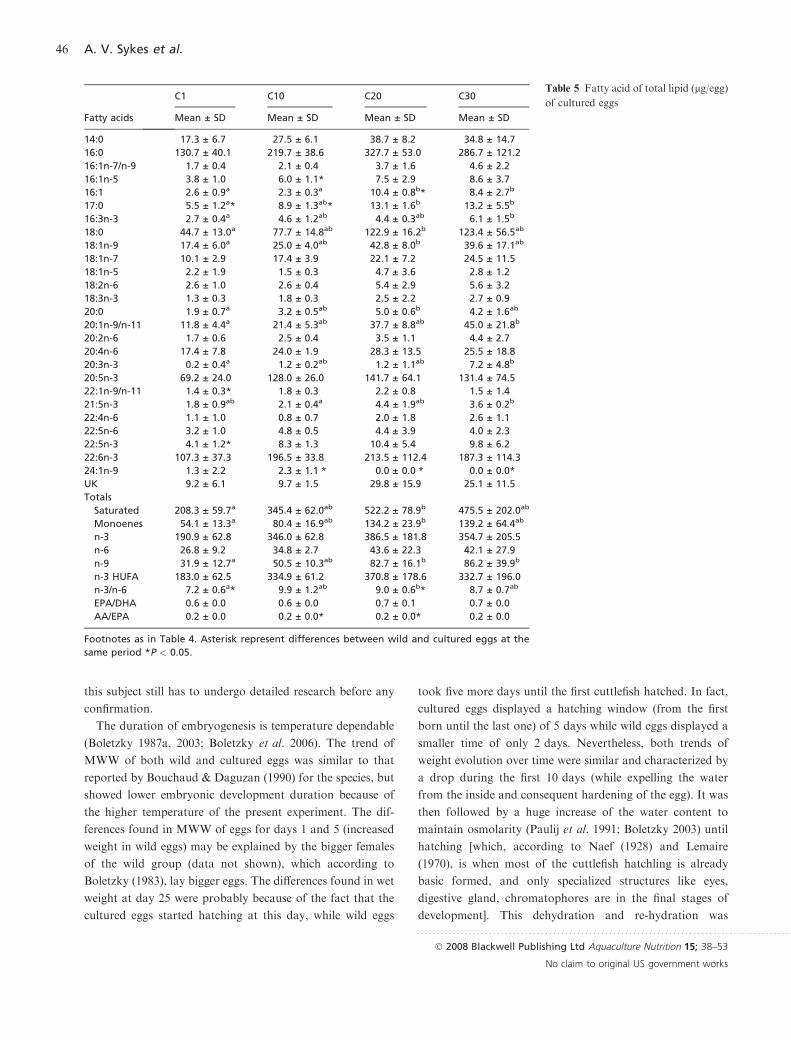
Tables 4 and 5 resume the overall FA composition of TL
of wild and cultured eggs (respectively) throughout the
embryonic development. In both type of eggs, although with
different amounts, a similar FA profile of TL was obtained,
1·5
2·5
2
1
0·5
01 5 10 15 20 25 30
Mea
n w
et w
eigh
t (g
)
CultureWild
Days
Figure 1 Egg mean wet weight (MWW)
trends over embryonic development.
Vertical bars represent standard devia-
tion. Stars represent differences for
P < 0.05.
96
94
92
90
88
86
84
8250 1510 2520 30
Days
Moi
stur
e (%
)
Culture eggsWild eggs
Figure 2 Egg moisture trends over
embryonic development. Vertical bars
represent standard deviation. Stars rep-
resent differences for P < 0.05.
. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .
� 2008 Blackwell Publishing Ltd Aquaculture Nutrition 15; 38–53
No claim to original US government works
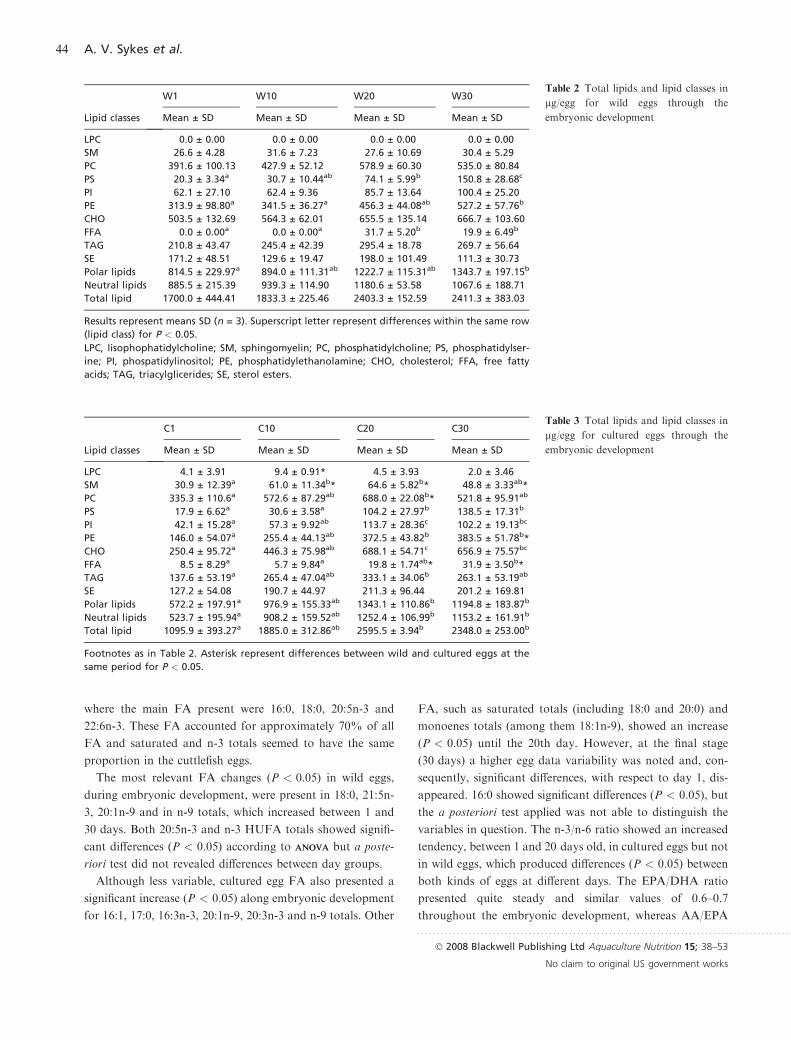
where the main FA present were 16:0, 18:0, 20:5n-3 and
22:6n-3. These FA accounted for approximately 70% of all
FA and saturated and n-3 totals seemed to have the same
proportion in the cuttlefish eggs.
The most relevant FA changes (P < 0.05) in wild eggs,
during embryonic development, were present in 18:0, 21:5n-
3, 20:1n-9 and in n-9 totals, which increased between 1 and
30 days. Both 20:5n-3 and n-3 HUFA totals showed signifi-
cant differences (P < 0.05) according to ANOVAANOVA but a poste-
riori test did not revealed differences between day groups.
Although less variable, cultured egg FA also presented a
significant increase (P < 0.05) along embryonic development
for 16:1, 17:0, 16:3n-3, 20:1n-9, 20:3n-3 and n-9 totals. Other
FA, such as saturated totals (including 18:0 and 20:0) and
monoenes totals (among them 18:1n-9), showed an increase
(P < 0.05) until the 20th day. However, at the final stage
(30 days) a higher egg data variability was noted and, con-
sequently, significant differences, with respect to day 1, dis-
appeared. 16:0 showed significant differences (P < 0.05), but
the a posteriori test applied was not able to distinguish the
variables in question. The n-3/n-6 ratio showed an increased
tendency, between 1 and 20 days old, in cultured eggs but not
in wild eggs, which produced differences (P < 0.05) between
both kinds of eggs at different days. The EPA/DHA ratio
presented quite steady and similar values of 0.6–0.7
throughout the embryonic development, whereas AA/EPA
Table 2 Total lipids and lipid classes in
lg/egg for wild eggs through the
embryonic developmentLipid classes
W1 W10 W20 W30
Mean ± SD Mean ± SD Mean ± SD Mean ± SD
LPC 0.0 ± 0.00 0.0 ± 0.00 0.0 ± 0.00 0.0 ± 0.00
SM 26.6 ± 4.28 31.6 ± 7.23 27.6 ± 10.69 30.4 ± 5.29
PC 391.6 ± 100.13 427.9 ± 52.12 578.9 ± 60.30 535.0 ± 80.84
PS 20.3 ± 3.34a 30.7 ± 10.44ab 74.1 ± 5.99b 150.8 ± 28.68c
PI 62.1 ± 27.10 62.4 ± 9.36 85.7 ± 13.64 100.4 ± 25.20
PE 313.9 ± 98.80a 341.5 ± 36.27a 456.3 ± 44.08ab 527.2 ± 57.76b
CHO 503.5 ± 132.69 564.3 ± 62.01 655.5 ± 135.14 666.7 ± 103.60
FFA 0.0 ± 0.00a 0.0 ± 0.00a 31.7 ± 5.20b 19.9 ± 6.49b
TAG 210.8 ± 43.47 245.4 ± 42.39 295.4 ± 18.78 269.7 ± 56.64
SE 171.2 ± 48.51 129.6 ± 19.47 198.0 ± 101.49 111.3 ± 30.73
Polar lipids 814.5 ± 229.97a 894.0 ± 111.31ab 1222.7 ± 115.31ab 1343.7 ± 197.15b
Neutral lipids 885.5 ± 215.39 939.3 ± 114.90 1180.6 ± 53.58 1067.6 ± 188.71
Total lipid 1700.0 ± 444.41 1833.3 ± 225.46 2403.3 ± 152.59 2411.3 ± 383.03
Results represent means SD (n = 3). Superscript letter represent differences within the same row
(lipid class) for P < 0.05.
LPC, lisophophatidylcholine; SM, sphingomyelin; PC, phosphatidylcholine; PS, phosphatidylser-
ine; PI, phospatidylinositol; PE, phosphatidylethanolamine; CHO, cholesterol; FFA, free fatty
acids; TAG, triacylglicerides; SE, sterol esters.
Table 3 Total lipids and lipid classes in
lg/egg for cultured eggs through the
embryonic developmentLipid classes
C1 C10 C20 C30
Mean ± SD Mean ± SD Mean ± SD Mean ± SD
LPC 4.1 ± 3.91 9.4 ± 0.91* 4.5 ± 3.93 2.0 ± 3.46
SM 30.9 ± 12.39a 61.0 ± 11.34b* 64.6 ± 5.82b* 48.8 ± 3.33ab*
PC 335.3 ± 110.6a 572.6 ± 87.29ab 688.0 ± 22.08b* 521.8 ± 95.91ab
PS 17.9 ± 6.62a 30.6 ± 3.58a 104.2 ± 27.97b 138.5 ± 17.31b
PI 42.1 ± 15.28a 57.3 ± 9.92ab 113.7 ± 28.36c 102.2 ± 19.13bc
PE 146.0 ± 54.07a 255.4 ± 44.13ab 372.5 ± 43.82b 383.5 ± 51.78b*
CHO 250.4 ± 95.72a 446.3 ± 75.98ab 688.1 ± 54.71c 656.9 ± 75.57bc
FFA 8.5 ± 8.29a 5.7 ± 9.84a 19.8 ± 1.74ab* 31.9 ± 3.50b*
TAG 137.6 ± 53.19a 265.4 ± 47.04ab 333.1 ± 34.06b 263.1 ± 53.19ab
SE 127.2 ± 54.08 190.7 ± 44.97 211.3 ± 96.44 201.2 ± 169.81
Polar lipids 572.2 ± 197.91a 976.9 ± 155.33ab 1343.1 ± 110.86b 1194.8 ± 183.87b
Neutral lipids 523.7 ± 195.94a 908.2 ± 159.52ab 1252.4 ± 106.99b 1153.2 ± 161.91b
Total lipid 1095.9 ± 393.27a 1885.0 ± 312.86ab 2595.5 ± 3.94b 2348.0 ± 253.00b
Footnotes as in Table 2. Asterisk represent differences between wild and cultured eggs at the
same period for P < 0.05.
. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .
� 2008 Blackwell Publishing Ltd Aquaculture Nutrition 15; 38–53
No claim to original US government works
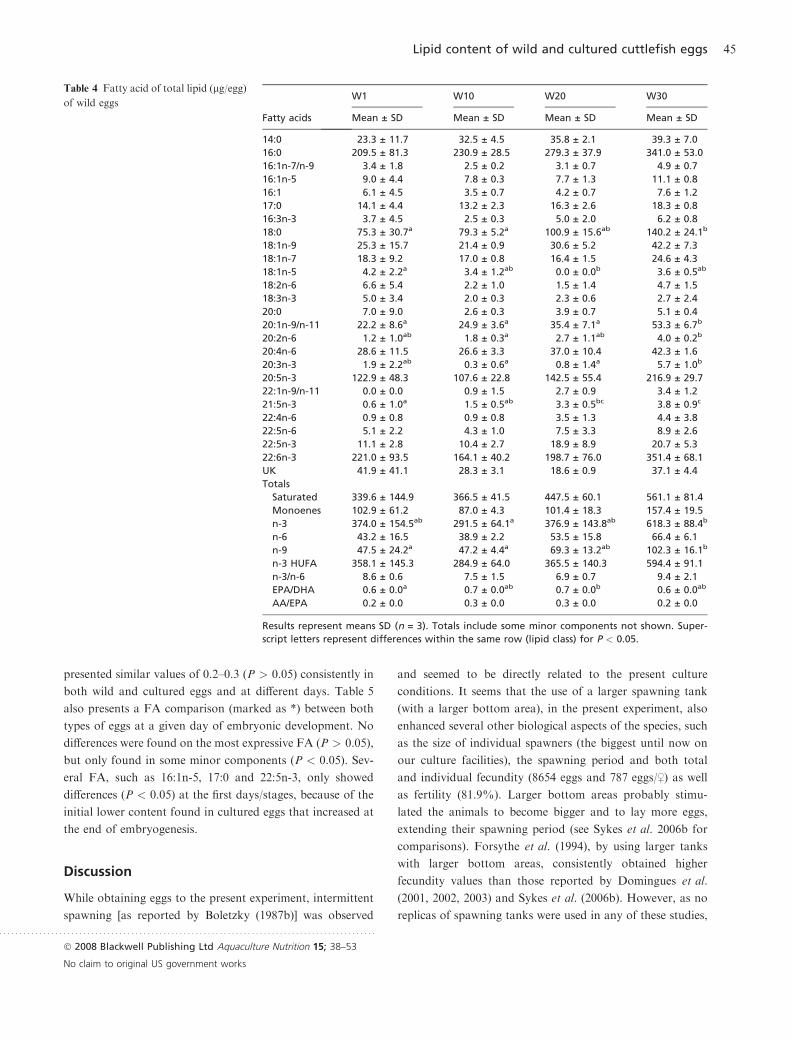
presented similar values of 0.2–0.3 (P > 0.05) consistently in
both wild and cultured eggs and at different days. Table 5
also presents a FA comparison (marked as *) between both
types of eggs at a given day of embryonic development. No
differences were found on the most expressive FA (P > 0.05),
but only found in some minor components (P < 0.05). Sev-
eral FA, such as 16:1n-5, 17:0 and 22:5n-3, only showed
differences (P < 0.05) at the first days/stages, because of the
initial lower content found in cultured eggs that increased at
the end of embryogenesis.
While obtaining eggs to the present experiment, intermittent
spawning [as reported by Boletzky (1987b)] was observed
and seemed to be directly related to the present culture
conditions. It seems that the use of a larger spawning tank
(with a larger bottom area), in the present experiment, also
enhanced several other biological aspects of the species, such
as the size of individual spawners (the biggest until now on
our culture facilities), the spawning period and both total
and individual fecundity (8654 eggs and 787 eggs/$) as well
as fertility (81.9%). Larger bottom areas probably stimu-
lated the animals to become bigger and to lay more eggs,
extending their spawning period (see Sykes et al. 2006b for
comparisons). Forsythe et al. (1994), by using larger tanks
with larger bottom areas, consistently obtained higher
fecundity values than those reported by Domingues et al.
(2001, 2002, 2003) and Sykes et al. (2006b). However, as no
replicas of spawning tanks were used in any of these studies,
Table 4 Fatty acid of total lipid (lg/egg)of wild eggs
Fatty acids
W1 W10 W20 W30
Mean ± SD Mean ± SD Mean ± SD Mean ± SD
14:0 23.3 ± 11.7 32.5 ± 4.5 35.8 ± 2.1 39.3 ± 7.0
16:0 209.5 ± 81.3 230.9 ± 28.5 279.3 ± 37.9 341.0 ± 53.0
16:1n-7/n-9 3.4 ± 1.8 2.5 ± 0.2 3.1 ± 0.7 4.9 ± 0.7
16:1n-5 9.0 ± 4.4 7.8 ± 0.3 7.7 ± 1.3 11.1 ± 0.8
16:1 6.1 ± 4.5 3.5 ± 0.7 4.2 ± 0.7 7.6 ± 1.2
17:0 14.1 ± 4.4 13.2 ± 2.3 16.3 ± 2.6 18.3 ± 0.8
16:3n-3 3.7 ± 4.5 2.5 ± 0.3 5.0 ± 2.0 6.2 ± 0.8
18:0 75.3 ± 30.7a 79.3 ± 5.2a 100.9 ± 15.6ab 140.2 ± 24.1b
18:1n-9 25.3 ± 15.7 21.4 ± 0.9 30.6 ± 5.2 42.2 ± 7.3
18:1n-7 18.3 ± 9.2 17.0 ± 0.8 16.4 ± 1.5 24.6 ± 4.3
18:1n-5 4.2 ± 2.2a 3.4 ± 1.2ab 0.0 ± 0.0b 3.6 ± 0.5ab
18:2n-6 6.6 ± 5.4 2.2 ± 1.0 1.5 ± 1.4 4.7 ± 1.5
18:3n-3 5.0 ± 3.4 2.0 ± 0.3 2.3 ± 0.6 2.7 ± 2.4
20:0 7.0 ± 9.0 2.6 ± 0.3 3.9 ± 0.7 5.1 ± 0.4
20:1n-9/n-11 22.2 ± 8.6a 24.9 ± 3.6a 35.4 ± 7.1a 53.3 ± 6.7b
20:2n-6 1.2 ± 1.0ab 1.8 ± 0.3a 2.7 ± 1.1ab 4.0 ± 0.2b
20:4n-6 28.6 ± 11.5 26.6 ± 3.3 37.0 ± 10.4 42.3 ± 1.6
20:3n-3 1.9 ± 2.2ab 0.3 ± 0.6a 0.8 ± 1.4a 5.7 ± 1.0b
20:5n-3 122.9 ± 48.3 107.6 ± 22.8 142.5 ± 55.4 216.9 ± 29.7
22:1n-9/n-11 0.0 ± 0.0 0.9 ± 1.5 2.7 ± 0.9 3.4 ± 1.2
21:5n-3 0.6 ± 1.0a 1.5 ± 0.5ab 3.3 ± 0.5bc 3.8 ± 0.9c
22:4n-6 0.9 ± 0.8 0.9 ± 0.8 3.5 ± 1.3 4.4 ± 3.8
22:5n-6 5.1 ± 2.2 4.3 ± 1.0 7.5 ± 3.3 8.9 ± 2.6
22:5n-3 11.1 ± 2.8 10.4 ± 2.7 18.9 ± 8.9 20.7 ± 5.3
22:6n-3 221.0 ± 93.5 164.1 ± 40.2 198.7 ± 76.0 351.4 ± 68.1
UK 41.9 ± 41.1 28.3 ± 3.1 18.6 ± 0.9 37.1 ± 4.4
Totals
Saturated 339.6 ± 144.9 366.5 ± 41.5 447.5 ± 60.1 561.1 ± 81.4
Monoenes 102.9 ± 61.2 87.0 ± 4.3 101.4 ± 18.3 157.4 ± 19.5
n-3 374.0 ± 154.5ab 291.5 ± 64.1a 376.9 ± 143.8ab 618.3 ± 88.4b
n-6 43.2 ± 16.5 38.9 ± 2.2 53.5 ± 15.8 66.4 ± 6.1
n-9 47.5 ± 24.2a 47.2 ± 4.4a 69.3 ± 13.2ab 102.3 ± 16.1b
n-3 HUFA 358.1 ± 145.3 284.9 ± 64.0 365.5 ± 140.3 594.4 ± 91.1
n-3/n-6 8.6 ± 0.6 7.5 ± 1.5 6.9 ± 0.7 9.4 ± 2.1
EPA/DHA 0.6 ± 0.0a 0.7 ± 0.0ab 0.7 ± 0.0b 0.6 ± 0.0ab
AA/EPA 0.2 ± 0.0 0.3 ± 0.0 0.3 ± 0.0 0.2 ± 0.0
Results represent means SD (n = 3). Totals include some minor components not shown. Super-
script letters represent differences within the same row (lipid class) for P < 0.05.
. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .
� 2008 Blackwell Publishing Ltd Aquaculture Nutrition 15; 38–53
No claim to original US government works
this subject still has to undergo detailed research before any
confirmation.
The duration of embryogenesis is temperature dependable
(Boletzky 1987a, 2003; Boletzky et al. 2006). The trend of
MWW of both wild and cultured eggs was similar to that
reported by Bouchaud & Daguzan (1990) for the species, but
showed lower embryonic development duration because of
the higher temperature of the present experiment. The dif-
ferences found in MWW of eggs for days 1 and 5 (increased
weight in wild eggs) may be explained by the bigger females
of the wild group (data not shown), which according to
Boletzky (1983), lay bigger eggs. The differences found in wet
weight at day 25 were probably because of the fact that the
cultured eggs started hatching at this day, while wild eggs
took five more days until the first cuttlefish hatched. In fact,
cultured eggs displayed a hatching window (from the first
born until the last one) of 5 days while wild eggs displayed a
smaller time of only 2 days. Nevertheless, both trends of
weight evolution over time were similar and characterized by
a drop during the first 10 days (while expelling the water
from the inside and consequent hardening of the egg). It was
then followed by a huge increase of the water content to
maintain osmolarity (Paulij et al. 1991; Boletzky 2003) until
hatching [which, according to Naef (1928) and Lemaire
(1970), is when most of the cuttlefish hatchling is already
basic formed, and only specialized structures like eyes,
digestive gland, chromatophores are in the final stages of
development]. This dehydration and re-hydration was
Table 5 Fatty acid of total lipid (lg/egg)of cultured eggs
Fatty acids
C1 C10 C20 C30
Mean ± SD Mean ± SD Mean ± SD Mean ± SD
14:0 17.3 ± 6.7 27.5 ± 6.1 38.7 ± 8.2 34.8 ± 14.7
16:0 130.7 ± 40.1 219.7 ± 38.6 327.7 ± 53.0 286.7 ± 121.2
16:1n-7/n-9 1.7 ± 0.4 2.1 ± 0.4 3.7 ± 1.6 4.6 ± 2.2
16:1n-5 3.8 ± 1.0 6.0 ± 1.1* 7.5 ± 2.9 8.6 ± 3.7
16:1 2.6 ± 0.9a 2.3 ± 0.3a 10.4 ± 0.8b* 8.4 ± 2.7b
17:0 5.5 ± 1.2a* 8.9 ± 1.3ab* 13.1 ± 1.6b 13.2 ± 5.5b
16:3n-3 2.7 ± 0.4a 4.6 ± 1.2ab 4.4 ± 0.3ab 6.1 ± 1.5b
18:0 44.7 ± 13.0a 77.7 ± 14.8ab 122.9 ± 16.2b 123.4 ± 56.5ab
18:1n-9 17.4 ± 6.0a 25.0 ± 4.0ab 42.8 ± 8.0b 39.6 ± 17.1ab
18:1n-7 10.1 ± 2.9 17.4 ± 3.9 22.1 ± 7.2 24.5 ± 11.5
18:1n-5 2.2 ± 1.9 1.5 ± 0.3 4.7 ± 3.6 2.8 ± 1.2
18:2n-6 2.6 ± 1.0 2.6 ± 0.4 5.4 ± 2.9 5.6 ± 3.2
18:3n-3 1.3 ± 0.3 1.8 ± 0.3 2.5 ± 2.2 2.7 ± 0.9
20:0 1.9 ± 0.7a 3.2 ± 0.5ab 5.0 ± 0.6b 4.2 ± 1.6ab
20:1n-9/n-11 11.8 ± 4.4a 21.4 ± 5.3ab 37.7 ± 8.8ab 45.0 ± 21.8b
20:2n-6 1.7 ± 0.6 2.5 ± 0.4 3.5 ± 1.1 4.4 ± 2.7
20:4n-6 17.4 ± 7.8 24.0 ± 1.9 28.3 ± 13.5 25.5 ± 18.8
20:3n-3 0.2 ± 0.4a 1.2 ± 0.2ab 1.2 ± 1.1ab 7.2 ± 4.8b
20:5n-3 69.2 ± 24.0 128.0 ± 26.0 141.7 ± 64.1 131.4 ± 74.5
22:1n-9/n-11 1.4 ± 0.3* 1.8 ± 0.3 2.2 ± 0.8 1.5 ± 1.4
21:5n-3 1.8 ± 0.9ab 2.1 ± 0.4a 4.4 ± 1.9ab 3.6 ± 0.2b
22:4n-6 1.1 ± 1.0 0.8 ± 0.7 2.0 ± 1.8 2.6 ± 1.1
22:5n-6 3.2 ± 1.0 4.8 ± 0.5 4.4 ± 3.9 4.0 ± 2.3
22:5n-3 4.1 ± 1.2* 8.3 ± 1.3 10.4 ± 5.4 9.8 ± 6.2
22:6n-3 107.3 ± 37.3 196.5 ± 33.8 213.5 ± 112.4 187.3 ± 114.3
24:1n-9 1.3 ± 2.2 2.3 ± 1.1 * 0.0 ± 0.0 * 0.0 ± 0.0*
UK 9.2 ± 6.1 9.7 ± 1.5 29.8 ± 15.9 25.1 ± 11.5
Totals
Saturated 208.3 ± 59.7a 345.4 ± 62.0ab 522.2 ± 78.9b 475.5 ± 202.0ab
Monoenes 54.1 ± 13.3a 80.4 ± 16.9ab 134.2 ± 23.9b 139.2 ± 64.4ab
n-3 190.9 ± 62.8 346.0 ± 62.8 386.5 ± 181.8 354.7 ± 205.5
n-6 26.8 ± 9.2 34.8 ± 2.7 43.6 ± 22.3 42.1 ± 27.9
n-9 31.9 ± 12.7a 50.5 ± 10.3ab 82.7 ± 16.1b 86.2 ± 39.9b
n-3 HUFA 183.0 ± 62.5 334.9 ± 61.2 370.8 ± 178.6 332.7 ± 196.0
n-3/n-6 7.2 ± 0.6a* 9.9 ± 1.2ab 9.0 ± 0.6b* 8.7 ± 0.7ab
EPA/DHA 0.6 ± 0.0 0.6 ± 0.0 0.7 ± 0.1 0.7 ± 0.0
AA/EPA 0.2 ± 0.0 0.2 ± 0.0* 0.2 ± 0.0* 0.2 ± 0.0
Footnotes as in Table 4. Asterisk represent differences between wild and cultured eggs at the
same period *P < 0.05.
. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .
� 2008 Blackwell Publishing Ltd Aquaculture Nutrition 15; 38–53
No claim to original US government works

previously described by Bouchaud & Daguzan (1990), with
similar values of moisture content between 85% and 95%
(Fig. 2) but it is not temperature dependable. In fact, the
trend of the moisture curves obtained for both wild and
cultured eggs, in the present experiment, was similar to those
obtained by these authors at similar embryonic development
temperatures. While undergoing its development, the ceph-
alopod embryos use yolk as food, to obtain energy and
structural components. Its amount and quality is of para-
mount importance to achieve a successful embryonic and
post-embryonic development. The sepia yolk has been
described as consisting of soft globules, surrounded by a
membrane and composed by a water soluble glyco-lipo-
protein (Ito et al. 1962; Blanchier 1981). According to Ito &
Fujii (1962), the glyco-lipo-protein has 20% lipid (with 65%
phospholipid and minor or none CHO contents) and a high
amount of carbohydrate (12.6%). In cuttlefish, yolk is
available in the outer and inner yolk sacs. According to
Boletzky (1989, 2003), the outer yolk sac is an �organ� which
plays several functions such as respiratory and circulatory
(until the proper formation of both gills and hearts) and also
acts as a stirrer, that allow the continuous movement of the
perivitelline fluid around the embryo. The epithelium of the
inner yolk sac is functional from early stages, while the outer
yolk sac epithelium becomes progressively active with the
completion of the outer (ectodermic) cover (Boletzky 1987a).
Newly born hatchlings generally have yolk left in the inner
yolk sac, the amount of which varies greatly (Boucher-Ro-
doni et al. 1987), and its quantity may influence the matu-
ration of central nervous system (CNS; Dickel et al. 1997).
During embryonic development and until hatching, the yolk
platelets are �digested� and assimilated by the growing
embryo through its blood sinuses, which exist in the external
yolk sac (Boletzky 1975).
In the present study, after deposition by females, both wild
and cultured egg batches used for the study of weight evo-
lution during embryogenesis displayed MWW(g) of
0.626 ± 0.131 and 0.766 ± 0.310 respectively (Fig. 1); while
Bouchaud & Galois (1990) reported a MWW(g) of
1.31 ± 0.12. Several factors may be the cause for such a
difference in egg weight and we will address this issue by
discussing the influence of these factors in egg weight,
duration of embryonic development and lipid content.
It is known that bigger females lay bigger eggs (Boletzky
1983) and that English Channel cuttlefish females, such as
those used in the Bouchaud & Galois (1990) study, are bigger
in nature because of the increased life cycle [data from
Richard (1971) compared with that of Sykes et al. (2006b)].
It is also known that temperature plays a major role in
cuttlefish growth and life cycle (Domingues et al. 2001, 2002;
Sykes et al. 2006b) and that both geographical locations
(south Portugal and English Channel) have different tem-
peratures all year round. Higher temperature could explain
the lower egg weights that have been consecutively found in
the Faro populations [data from the current study and Sykes
et al. (2006b)]. For instance, eggs obtained by Sykes et al.
(2006b) during consecutive generations displayed bigger eggs
from females cultured at lower temperatures than those from
females cultured at higher temperatures, and in here the
premise �bigger females lay bigger eggs� is still confirmed.
However, the existence of contradictory data between both
English Channel and south Portugal populations also may be
as a result of the different genetic substructure found (Wol-
fram et al. 2006), which would imply different physiological
adaptations and eggs nutritionally different in content and
development. Sykes et al. (2006a) discussed, in its recent re-
view on the potential of cuttlefish culture, that there might
exist sub-speciation in the species, based on several reported
data on biological parameters such as egg and hatchling
weights, duration of the life cycle, etc.
Similarly, embryonic development is known to be tem-
perature dependent (Boletzky 1983; Bouchaud & Daguzan
1990). However, the geographical variations were not taken
into account until now and, although the period of embry-
onic development increases with lower temperatures, the
values diverge between Faro and Caen (see Bouchaud &
Galois 1990; Sykes et al. 2006b). A longer incubation period
demands for higher and/or different yolk content. In fact,
Bouchaud & Galois (1990) described higher yolk-TL con-
sumption at 12 �C (82% of initial yolk TL) than at 24 �C(only 26%). Thus, the embryo will have different metabolic
needs and rates at lower temperatures that also promote
higher available dissolved oxygen. However, at higher culture
temperatures there is an increased need for oxygen uptake by
the embryo (Wolf et al. 1985). Therefore, different physiol-
ogies may be applied in accordance. This is even more evi-
dent when, according to Bouchaud & Galois (1990),
cuttlefish embryos consume only 26% of their egg-yolk
during the embryonic development at 24 �C, while at 15 �Cegg-yolk consumption is close to 78%. Also, Bouchaud &
Daguzan (1990) reported data regarding the conversion of
the yolk in hatchling dry weight, where eggs cultured at 12 �Cwould convert as much as 91%, while those cultured at 21 �Cwould only convert 49%. So, at 15 �C, the embryo needs to
cope with a longer embryonic development, yolk must pro-
vide enough food (for growth and energetic demands) and
oxygen demands are more easily met due the high solubility,
thus not requiring so much outer yolk sac area/volume
. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .
� 2008 Blackwell Publishing Ltd Aquaculture Nutrition 15; 38–53
No claim to original US government works

available. According to the energetic budgets of Bouchaud
(1991), at 15 �C, 41% of egg-yolk are used for growth rate
and only 10% for catabolic processes (respiration and
excretion), while at 24 �C growth rates are smaller (15%) and
there is an increased catabolism (52%). In practice, English
Channel eggs cultured at 15 �C use about 7.8% of their yolk
as energy, while those cultured at 24 �C use as much as 13%.
All these data would be related with the smaller hatchlings
obtained at higher temperatures. It seems that lower tem-
peratures (12–15 �C), which promote extended incubation
because of lower growth rates but also higher use of yolk, will
also imply the requirement of the egg for higher and/or dif-
ferent nutritional content. Depending on the temperature,
life cycle and embryogenesis duration in cuttlefish are
adapted [lower temperatures promote extended periods; see
Sykes et al. (2006b) for data], and therefore geographical
locations probably stimulate physiological differences that
are reflected in the egg content and metabolism. Therefore,
studies that will use both Faro and Caen eggs cultured
through their embryogenesis at similar temperatures and
covering a wide range of temperatures needed to determine if
embryonic development follows the same time periods and if
hatchlings share similar weight at hatching. Also, the quan-
tity, quality and the use of the yolk should be investigated.
When we analysed the lipid content and composition in
both egg groups (from wild and cultured spawners),
throughout embryonic development, they showed similar li-
pid and moisture content. It was noteworthy an increase in
TL, and certain LC and FA, but different live feed (cultured
spawners were only fed live grass shrimp while wild spawners
usually feed on a wide variety of prey) did not promoted a
significantly different lipid profiles between eggs.
Sargent (1995) and Rainuzzo et al. (1997) stated that, in
fish, lipids are major sources of metabolic energy throughout
the embryonic development. However the TL amounts and
the LC used varies with the fish species. Normally, there is a
PC and TAG catabolism and a PS and PE synthesis. In the
present experiment, no lipid consumption, either in percent-
age or in absolute value (lg/egg), during embryonic devel-
opment was observed. The maintenance of lipid contents of
this study is in agreement with Bouchaud & Galois (1990),
who observed maintenance of TL between newly-laid eggs
and hatchlings. However, percentage values from the present
study (2–3%DW of TL, data not shown) were clearly lower
than that of the previous authors (14% DW). Also, our egg
absolute TL values were lower at spawning (�1.5 mg/egg;
mean from values of Tables 2 and 3) than those of the Bou-
chaud & Galois (1990) study (�8.5 mg/egg). Therefore the
differences found cannot be explained only by the elimination
of the outer gummy layers from the eggs, made by the latter
authors and not performed in the present study. From the
present data, we understand that the lipid value cannot be
independent from egg size and constant geographically, as
stated earlier by Bouchaud & Galois (1990) and, therefore,
this value is different between different populations of the
same species. The reason for the different amount of lipids in
cuttlefish eggs from Faro and Caen could be related with
differences in metabolism, temperature influence or genetic
sub-speciation, as mentioned before in this discussion.
According to Richard (1971), the egg size greatly influences
the amount of viteline reserves. Viteline assimilation provides
energy for breathing and, through catabolic processes, is used
in the formation of new tissues. Therefore, bigger eggs will
probably mean different content (in %) either in protein, lipid
and carbohydrate, and/or in their individual content [e.g. it
may refer to an increased lipid content and different LC of
eggs from the English Channel, that are used during
embryogenesis at similar temperatures (laboratory induced)
obtained naturally in the present study, which are not normal
in that geographical area], as seen when comparing data from
the latter authors and that of present study.
In the present study, SD obtained for both TL and LC
were quite high and seemed to reflect not only different
amounts of lipid content per egg (promoted by either dif-
ferent females and/or the individual content of each egg), but
also the fact that each female may lay eggs for several hours,
thus leading to different embryonic development times (to a
maximum of 24 h). The increase or maintenance of the egg�s
lipid fraction, in its total weight, points out to a possible
alternative use of the protein and/or carbohydrate fraction as
energy during this phase, which would be in accordance with
the findings of Hochachka (1994). This author stated that
cephalopod muscle mitochondria work under oxygen limi-
tations, because of the lack of several mechanism to upre-
gulate the capacities for oxygen flux from blood to
mitochondria, such as lack of intracellular myoglobin-ana-
logue and low-intracellular lipid concentrations (presented
that high-lipid concentrations supplied a higher oxygen sol-
ubility and diffusion). This situation would favour an oxy-
gen-efficient metabolic organization which maximizes the
ATP yield per mole of oxygen. In this sense, the oxidation of
carbohydrates and amino acids are more oxygen efficient
than lipids. This is also in accordance with O�Dor et al.
(1984), that states that cephalopods use protein as building
blocks for growth and carbohydrate as energy fuel, the later
being stored in small amounts as muscle glycogen. Similar
results were obtained by Castro et al. (1992), for starved
juvenile-adult cuttlefish. In the present study, the verified
. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .
� 2008 Blackwell Publishing Ltd Aquaculture Nutrition 15; 38–53
No claim to original US government works

lipid increase (although not statistical significant in wild eggs)
may perhaps be explained by de novo synthesis of some of its
components during embryogenesis, although further studies
are necessary to confirm this.
Both polar and neutral lipid fractions seemed to be equal
in the egg content (�50% each). A similar 1 : 1 ratio of PL
and NL was found by Silversand et al. (1996) in turbot eggs.
The absolute value (lg/egg) of several lipid classes such as
PS, PI, PE and CHO increased along the embryonic devel-
opment. Similar results were reported by Vazquez et al.
(1994), in Solea senegalensis eggs, for the PS and PI; this
could be explained by de novo synthesis of these LC, given
their importance as membrane constituents. Navarro & Vi-
llanueva (2000) reported similar findings for hatchlings and
Almansa et al. (2006) for juvenile and maturing cuttlefish.
Phospholipids and CHO are known to have a structural role
as components of cell membranes (Sargent et al. 1995a).
Phosphoglycerides (especially PC) are used not only for cell
division and organogenesis but also used as fuel by some fish
species (Sargent 1995), while neutral lipids (especially TAG
and SE) are generally considered the most important energy
reserves in marine fish eggs and larvae (Almansa et al. 1999).
Triacylglycerol content described in the present study
corresponded to >10% of TL. That is in agreement with
Almansa et al. (2006), who found low levels of TAG in the
mantle of maturing cuttlefish, despite the high level of this
LC content in fish used as diet. Bouchaud & Galois (1990)
reported a small decrease in TAG content between newly-laid
eggs and hatchlings of cuttlefish, which was not observed in
the current study in either group. It is known that TAG may
have a dual application: a metabolic, where it can be used as
a source for ATP production through oxidation, and/or
structural, where it can be a source of FA for polar lipid (PL)
biosynthesis (Sargent et al. 1995a). However, the statistical
differences found in TAG (only for cultured eggs) did not
allow us to determine a trend. Nonetheless, it was notewor-
thy the lower SD found in both groups at day 20. Finally, the
higher contents of PE present in wild eggs respect to cultured
eggs at 30 days old suggest a possible deficiency of this LC,
which has a relevant role in the nervous system and eye
formation.
The high levels of CHO obtained for both type of eggs in
the present study was similar to those found by Almansa
et al. (2006) and Domingues et al. (2004) in cuttlefish mantle.
CHO is an important LC as a component of cell membranes
with relevant properties (Crockett & Hazel 1997) and, in
cuttlefish, CHO (either endo or exogenous) is a precursor for
steroid biosynthesis (Blanchier 1981). In addition, it was re-
ported earlier by Goad (1981) that cuttlefish gonads possess
all seven enzymes required to produce the steroid interme-
diates that allow the biosynthesis of testosterone. Besides the
earlier Zandee�s (1967) findings regarding an absence of
endogenous synthesis of CHO for the species, we believe that
CHO should be included in the elaboration of a future diet
and must be present in the egg yolk for a correct develop-
ment. In fact, grass shrimp, which was used as optimal prey
during the first 30 DAH and the rest of the life cycle (Sykes
et al. 2006b), presents high levels of CHO. It is relevant that
Ito & Fujii (1962) reported the non-existence of CHO in
cuttlefish high-density lipoproteins (HDL) isolated from the
egg yolk. However, our results and previous data by Bou-
chaud & Galois (1990), found an 8–9% of CHO (%TL) in
cuttlefish eggs. Evidence for de novo sterol biosynthesis in the
species had been acknowledged by Kanazawa (2001). These
differences may perhaps be explained by the inclusion of
CHO in other egg-yolk components different from the HDL
studied by the later authors. Also, the presence of CHO, in
an esterified form, as SE (CHO esterified with a FA), would
explain the difficult of detection of CHO content in the HDL
by the experimental method used by Ito & Fujii (1962), al-
though these hypotheses should be confirmed with further
studies.
Lipid content of the eggs is supplied by the female, ob-
tained from diet and/or its lipid storage. Lee (1994) suggested
that if the lipid fraction of the mantle is so low, then the lipid
storage is made at the digestive gland. Blanchier & Boucaud-
Camou (1982) studied the lipid composition of the digestive
gland in wild-mature cuttlefish and found differences of TL
between males and females (9%WW and 13%WW respec-
tively). This is in accordance with the findings of Boucaud-
Camou (1971), who described the lipid fraction of the
digestive gland to be rich in TAG, sterols, sterol and wax
esters, plus phospholipids (PE and PC). Blanchier & Bou-
caud-Camou (1982) also found a similar LC content, for
both sexes, to be characterized by phospholipids, sterols,
FFA, TAG, wax and SE. Nevertheless, the authors stated
that the available lipid (in the digestive gland) does not
change in terms of percentage, but do not discard its use in
lipid and lipoprotein synthesis in the gonads. It must be
considered that cuttlefish presents an intermittent spawning,
which would allow the females to replace the lipids used
and taken from the digestive gland through food ingested
during the spawning period. As information regarding oocyte
lipid incorporation is inexistent, studies regarding the lipid
pathways involved in digestive gland storage and release
into gonad development and oocytes in females at matur-
ing and spawning age need to be conducted to clarify this
issue.
. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .
� 2008 Blackwell Publishing Ltd Aquaculture Nutrition 15; 38–53
No claim to original US government works

In terms of FA, no major differences were found between
wild and cultured eggs throughout the embryonic develop-
ment. Although 16:0, EPA and DHA are the most commonly
found FA in cephalopod flesh, whole body, digestive gland,
CNS and photoreceptors, cuttlefish eggs also displayed high
levels of 18:0 and intermediate levels of both 18:1n-9 and
20:1n-9. Dumont et al. (1992) studied the FA profiles in the
CNS of cuttlefish and found a similar predominance of
DHA, EPA, 16:0 and 18:0 in its CNS. The present results
revealed high levels of the n-3 HUFA (especially EPA and
DHA) in cuttlefish eggs, thus confirming the results already
obtained by Navarro & Villanueva (2000) that the dietary
requirements for n-3 HUFA are critical in early cephalopod
developmental stages because of the high demand for mem-
brane synthesis (Henderson & Sargent 1985; Sargent
et al.2002). DHA plays an important role in the synthesis,
maintenance and functional integrity of the structure of cell
membranes in fish, especially in brain and retina, where it can
account for up to 40% of the total FA (Sargent 1995). The
importance of DHA in general cephalopods and in S. offi-
cinalis specifically may be classified as extreme if one takes
into account the exclusivity of both their brain and retinas.
EPA is a precursor of active eicosanoids such as prosta-
glandins. Nonetheless, despite the growing importance of
EPA and DHA in cuttlefish nutrition, one should not fall in
the same mistake of not considering AA as an essential FA.
The importance of AA as the major eicosanoid precursor for
normal growth and development has been demonstrated in
fish (Bell et al. 1995, 2003; Sargent et al. 1995a,b; Bell &
Sargent 2003) and Almansa et al. (2006) suggested an n-6
HUFA metabolism in juvenile and maturing juvenile cuttle-
fish, where AA could be involved in a possible biosynthesis
pathway of 22:4n-6 and 22:5n-6.
When we analysed the variation of FA along the
embryonic development, it was relevant the increased of
several FA such as n-9 totals and particularly 20:1n-9,
found in both wild and cultured eggs. This tendency could
be as a result of de novo synthesis of these FA, which is a
usual pathway in marine species (Sargent 1995). In wild
eggs, despite being statistically non-significant, an increase
in 20:5n-3 was also noted. The synthesis of this FA is not as
usual as the one observed for monoenes and especially in a
carnivorous species as cuttlefish, which must obtain 20:5n-3
from the diet (Sargent 1995; Sargent et al. 2002). However,
no data regarding both elongation and desaturation (met-
abolic pathways) of FA are currently available for cepha-
lopods, and therefore additional studies are necessary to
provide further insight and confirmation of the current
data.
In conclusion, the present authors suggest that the dis-
crepancy in results obtained here (in terms of egg lipid con-
tent and possible lipid metabolism) might be related to the
existence of sub-species or at least two different populations
(see Sykes et al. 2006a). If this is proven to be true, then the
optimal embryonic development temperature of 15–18 �Csuggested by Bouchaud (1991) for the English Channel might
not be the best for the south Portugal population. One pos-
sible way to determine nutritional and physiological differ-
ences in eggs between populations, based on the above
assumptions, would be to determine both total amount of
energy per individual (J or cal/ind.) of south Portugal (as it
will indicate the energy available to a developing embryo),
and the caloric value (J or cal mg)1) of those eggs. Surely the
energy available in bigger eggs will be substantially higher
than in smaller eggs, as previously observed by Bouchaud
(1991), as the embryos will have higher energetic demands as
demonstrated above. Another way would be the study of
nutrient contents and metabolism in geographically different
eggs (embryo plus yolk) and hatchlings. This would allow a
better judgment of what really happens in terms of different
egg weights and if the egg energetic reserves are or not
completely exhausted before hatching.
The present results suggest that lipids are not used as ener-
getic substrate in cuttlefish eggs of south Portugal. The FA
extracted from eggs of wild cuttlefish were highly unsaturated,
thus indicating that embryos and hatchlings have nutritional
requirements for PUFA, especially EPA and DHA. An
increasing trendwas found during embryonic development for
lipid and FA content which was not statistically significant in
many components because of the high SDof lipids within eggs.
The present data suggest a possible anabolism of several LC
such as PS, PI, PE and CHO, which could be explained given
their importance for the cell membrane formation. Regarding
that, further studies, which should involve separation of cho-
rion, embryo and yolk, need to be conducted to provide ex-
tended knowledge of lipid metabolism at this stage and prove
the above. We believe, however, that lipid importance at
the reproduction, egg and hatchling stages must be under-
lined because of their structural function. The present
results provide new insight in what should be the lipid
requirements for a prepared diet for the hatchling stage, which
should have a balanced PL and NL fractions and have similar
FA profiles as the ones presented here.
The authors would like to thank the Fundacao para a Ci-
encia e a Tecnologia (FCT) from the Portuguese government,
. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .
� 2008 Blackwell Publishing Ltd Aquaculture Nutrition 15; 38–53
No claim to original US government works

and the Agencia de Inovacao (project AQUASEPIA) for
the financial support for this research. This work was also
funded by a FCT PhD grant (SFRH/BD/12409/2003) to
Antonio Sykes.
Almansa, E., Perez, M.J., Cejas, J.R., Badıa, P., Villamandos, J.E. &
Lorenzo, A. (1999) Influence of broodstock seabream (Sparus
aurata L.) dietary fatty acids on egg quality and egg fatty acid
composition throughtout the spawning season. Aquaculture, 170,
323–336.
Almansa, E., Domingues, P.M., Sykes, A.V., Tejera, N., Lorenzo, A.
& Andrade, J.P. (2006) The effects of feeding with shrimp or fish
fry on growth and mantle lipid composition of juvenile and adult
cuttlefish (Sepia officinalis). Aquaculture, 256, 403–413.
Barnabe, G. (1996) Bases biologiques et ecologiques de l�aquaculture,p. 519. Editorial Acribia, S.A. Zaragoza, Spain.
Bell, J.G. & Sargent, J.R. (2003) Arachidonic acid in aquaculture
feeds: current status and future opportunities. Aquaculture, 218,
491–499.
Bell, J.G., Castell, J.D., Tocher, D.R., MacDonald, F.M. & Sargent,
J.R. (1995) Effects of different arachidonic acid: docosahexaenoic
acid ratios on phospholipid fatty acid compositions and prosta-
glandin production in juvenile turbot (Scophthalmus maximus).
Fish Physiol. Biochem., 14, 139–151.
Bell, J.G., McEvoy, L.A., Estevez, A., Shields, R.J. & Sargent, J.R.
(2003) Optimizing lipid nutrition in first-feeding larvae. Aquacul-
ture, 227, 211–220.
Blanchier, B. (1981) Etude des lipids totaux et des steroids dans la
glande digestive et la gonade chez la Seiche Sepia officinalis L. These
Docteur de Specialite (3e Cycle), p. 172. Universite de Caen, Caen,
France.
Blanchier, B. & Boucaud-Camou, E. (1982) Contenu lipidique de la
glande digestive de Sepia officinalis L. a maturite sexuelle. Mal-
acologia, 22, 691–696.
Blanchier, B. & Boucaud-Camou, E. (1984) Lipids in the digestive
gland and the gonad of immature and mature Sepia officinalis
(Mollusca: Cephalopoda). Mar. Biol., 80, 39–43.
Blanchier, B., Boucaud-Camou, E. & Silberzahn, P. (1986) Com-
parative study of sterol composition of the digestive gland and the
gonad of Sepia officinalis L. (Mollusca: Cephalopoda). Comp.
Biochem. Physiol., 83, 599–602.
Boletzky, S.v. (1975) A contribution to the study of yolk absorption
in the cephalopoda. Z. Morph. Tiere, 80, 229–246.
Boletzky, S.v. (1983) Sepia officinalis. In: Cephalopod Life Cycles
(Boyle, P.R. ed.), Vol. I. – Species Accounts, pp. 31–52. Academic
Press, London.
Boletzky, S.v. (1987a) Embryonic phase. In: Cephalopod Life
Cycles (Boyle, P.R. ed.), Vol. II., pp. 5–31. Academic Press,
London.
Boletzky, S.v. (1987b) Fecundity variation in relation to intermittent
or chronic spawning in the cuttlefish, Sepia officinalis (Mollusca,
Cephalopoda). Bull. Mar. Sci., 40, 382–387.
Boletzky, S.v. (1989) Recent studies on spawning, embryonic devel-
opment, and hatching in the cephalopoda. Adv. Mar. Biol., 25, 85–
115.
Boletzky, S.v. (2003) Biology of early life stages in cephalopod
molluscs. Adv. Mar. Biol., 44, 143–203.
Boletzky, S.v., Erlwein, B. & Hofmann, D.K. (2006) The Sepia egg: a
showcase of cephalopod embryology. Vie Milieu, 56, 191–201.
Boucaud-Camou, E. (1971) Counstituants lipidiques du foie de Sepia
officinalis. Mar. Biol., 8, 66–69.
Boucaud-Camou, E. (1990) La Seiche, un animal d�avenir, La Peche
Maritime. La Peche maritime, 69, 321–329.
Bouchaud, O. (1991) Energy consumption of the cuttlefish
Sepia officinalis L. (Mollusca: Cephalopoda) during embryonic
development, preliminary results. Bull. Mar. Sci., 49, 333–340.
Bouchaud, O. & Daguzan, J. (1990) Etude experimentale de l�influ-ence de la temperature sur le deroulement du developpement em-
bryonnaire de la Seiche Sepia officinalis L. (Cephalopode,
Sepioidae). Cah. Biol. Mar., 31, 131–145.
Bouchaud,O.&Galois, R. (1990)Utilization of egg-yolk lipids during
the embryonic development of Sepia officinalis L. in relation to
temperature of the water. Comp. Biochem. Physio., 97B, 611–615.
Boucher-Rodoni, R., Boucaud-Camou, E. & Mangold, K. (1987)
Feeding and digestion. In: Cephalopod Life Cycles (Boyle, P.R.
ed.), Vol. 2, pp. 85–108. Academic Press, London.
Boyle, P.R., Noble, L., Emery, A.M., Craig, S., Black, K.D. &
Overnell, J. (2001) Development and hatching in cephalopod eggs:
a model system for partitioning environmental and genetic effects
on development. In: Animal Development Biology – Environment
and Animal Development: Genes, Life History and Plasticity
(Atkinson, D. & Thorndyke, M. eds.). pp. 251–267. Oxford BIOS
Scientific, Oxford.
Castro, B.G., Garrido, J.L. & Sotelo, C.G. (1992) Changes in
composition of digestive gland and mantle muscle of the cuttlefish
Sepia officinalis during starvation. Mar. Biol., 114, 11–20.
Cejas, J.R., Almansa, E., Jerez, S., Bolanos, A., Felipe, B. & Lore-
nzo, A. (2004) Changes in lipid class and fatty acid composition
during development in white seabream (Diplodus sargus) eggs and
larvae. Comp. Biochem. Physiol., 139, 209–216.
Christie, W.W. (1982) Lipid Analysis, 2nd edn. Pergamon Press,
Oxford.
Crockett, E.L. & Hazel, J.R. (1997) Cholesterol affects physical
properties and (Na+,K+)-ATPase in basolateral membranes of
renal and intestinal epithelia from thermally acclimated rainbow
trout. J. Comp. Physiol., 167, 344–351.
Dickel, L., Chichery, M.P. & Chichery, R. (1997) Postembryonic
maturation of the vertical lobe complex and early development of
predatory behavior in the cuttlefish (Sepia officinalis). Neurobio.
Learn. Mem., 67, 150–160.
Domingues, P.M., Sykes, A. & Andrade, J.P. (2001) The use of
artemia or mysids as food for hatchlings of the cuttlefish Sepia
officinalis Linnaeus, 1758; effects on growth and survival
throughout the life cycle. Aquacult. Int., 9, 319–331.
Domingues, P.M., Sykes, A. & Andrade, J.P. (2002) The effects of
temperature in the life cycle of two consecutive generations of the
cuttlefish Sepia officinalis (Linnaeus, 1758), cultured in the Algarve
(south Portugal). Aquacult. Int., 10, 207–220.
Domingues, P.M., Poirier, R., Dickel, L., Almansa, E., Sykes, A. &
Andrade, J.P. (2003) Effects of culture density and live prey on
growth and survival of juvenile cuttlefish, Sepia officinalis. Aqua-
cult. Int., 11, 225–242.
Domingues, P.M., Sykes, A., Sommerfield, A., Almansa, E., Lore-
nzo, A. & Andrade, J.P. (2004) Growth and survival of cuttlefish
(Sepia officinalis) of different ages fed crustaceans and fish. Effects
of frozen and live prey. Aquaculture, 290, 239–254.
Dumont, E., Chichery, M.P., Chichery, R. & Nouvelot, A. (1992)
Analysis of the fatty acids in the central nervous system of Sepia
officinalis. Comp. Biochem. Physiol., 101B, 341–344.
Dumont, E., Chichery, M.P., Nouvelot, A. & Chichery, R. (1994)
Variations of the lipid constituents in the central nervous system of
. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .
� 2008 Blackwell Publishing Ltd Aquaculture Nutrition 15; 38–53
No claim to original US government works

the cuttlefish (Sepia officinalis) during aging. Comp. Biochem.
Physiol., 108A, 315–323.
Falk-Petersen, S., Falk-Petersen, I.-B., Sargent, J.R. & Haug, T.
(1986) Lipid class and fatty acid composition of eggs from the
Atlantic Halibut (Hippoglossus hippoglossus). Aquaculture, 52,
207–211.
Finn, R.N. (1994) Physiological Energetics of Developing Marine Fish
Embryos and Larvae. Dr. Scient. Thesis, University of Bergen,
Bergen, Norway, 206 pp.
Finn, R.N., Rønnestad, I. & Fyhn, H.J. (1995) Respiration, nitrogen
and energy metabolism of developing yolk-sac larvae of Atlantic
halibut (Hippoglossus hipoglossus L.). Comp. Biochem. Physiol.,
IIIA, 647–671.
Finn, R.N., Fyhn, H.J., Henderson, R.J. & Evjen, M.S. (1996) The
sequence of catabolic substrate oxidation and enthalpy balance of
developing embryos and yolk-sac larvae of Turbot (Scophthalmus
maximus L.). Comp. Biochem. Physiol. Vol., 115A, 133–151.
Forsythe, J.W., DeRusha, R.H. & Hanlon, R.T. (1994) Growth,
reproduction and life span of Sepia officinalis (Cephalopoda:
Mollusca) cultured through seven consecutive generations. J. Zool.
Lond., 233, 175–192.
Fowler, J., Cohen, L. & Jarvis, P. (2002) Practical statistics for field
biology, 2nd ed. Wiley and Sons, West Sussex, England, 259 p.
Fujii, T. (1960) Comparative biochemical studies on the egg-yolk
proteins of various animal species. Acta Embryol. Morph. Exp., 3,
260–285.
Goad, L.J. (1981) Sterol biosynthesis and metabolism in marine
invertebrates. Pure Appl. Chem., 51, 837–852.
Henderson, R.J. & Sargent, J.R. (1985) Fatty acid metabolism in
fish. In: Nutrition and Feeding in Fish (Cowey, C.B., Mackie, A.M.
& Bell, J.G. eds), pp. 349–364. Academic Press, London.
Hochachka, P.W. (1994) Oxygen efficient design of cephalopod
muscle metabolism. Mar. Fresh. Behav. Physiol., 25, 61–67.
Horwitz, W. (1980) Methods of Analysis, 13th edn. Association of
Official Analytical Chemists, Washington DC.
Ito, Y. & Fujii, T. (1962) Chemical composition of the egg-yolk
lipoproteins. J. Biochem., 52, 221–222.
Ito, Y., Fujii, T. & Otake, M. (1962) On a peptide constituent of the
egg-yolk lipo-proteins. J. Biochem., 52, 223–225.
Kanazawa, A. (2001) Sterols in marine invertebrates. Fish. Sci., 67,
997–1007.
Lee, P.G. (1994) Nutrition of cephalopods: fuelling the system. Mar.
Freshw. Behav. Physiol., 25, 35–51.
Lemaire, J. (1970) Table de development embryonnaire de Sepia
officinalis L. (Mollusque Cephalopode). Bull. Soc. Zool. France,
95, 773–782.
Morehead, D.T., Hart, P.R., Dunstan, G.A., Brown, N. & Pank-
hurst, N.W. (2001) Differences in egg quality of wild stripped
trumpeter (Latris lineata) and captive striped trumpeter that were
fed different diets. Aquaculture, 192, 39–53.
Mourente, G. & Vazquez, R. (1996) Changes in the content of total
lipid, lipid classes and their fatty acids of developing eggs and
unfed larvae of the Senegal sole, Solea senegalensis Kaup. Fish
Physiol. Biochem., 15, 221–235.
Naef, A. (1928) Die cephalopoden (Embryologie). In: Fauna e Flora
del Golfo di Napoli (Bardi, G. ed.), pp. 1–357. Friedlander & John
(Berlin). Stazione Zoologica di Napoli, Italia.
Navarro, J.C. & Villanueva, R. (2000) Lipid and fatty acid compo-
sition of early stages of cephalopods: an approach to their lipid
requirements. Aquaculture, 183, 161–177.
O�Dor, R.K., Mangold, K., Boucher-Rodoni, R., Wells, M.J. &
Wells, J. (1984) Nutrient absortion, storage and remobilizaion in
Octopus vulgaris. Mar. Behav. Physiol., 11, 239–258.
Olsen, R.E. & Henderson, R.J. (1989) The rapid analysis of
neutral and polar marine lipids using double-development HPTLC
and scanning densitometry. J. Exp. Mar. Biol. Ecol., 129,
189–197.
Paulij, W.P., Herman, P.M.J., Roozen, M.E.F. & Denuce, J.M.
(1991) The influence of photoperiodicity on hatching of Sepia of-
ficinalis. J. Mar. Biol. Ass. U.K., 71, 665–678.
Perez-Losada, M., Guerra, A. & Sanjuan, A. (1999) Allozyme dif-
ferentiation in the cuttlefish Sepia officinalis (Mollusca: Cephalo-
poda) from the NE Atlantic and Mediterranean. Heredity, 83,
280–289.
Perez-Losada, M., Guerra, A., Carvalho, G.R., Sanjuan, A. & Shaw,
P.W. (2002) Extensive population subdivision of the cuttlefish
Sepia officinalis (Mollusca: Cephalopoda) around the Iberian
Peninsula indicated by microsatellite DNA variation. Heredity, 89,
417–424.
Perez-Losada, M., Nolte, M.J., Crandal, K.A. & Shaw, P.W. (2007)
Testing hypothesis of population structuring in the Northeast
Atlantic Ocean and Mediterranean Sea using the common cuttle-
fish Sepia officinalis. Mol. Ecol., 16, 2667–2679.
Rainuzzo, J.R. (1993) Lipids in Early Stages of Marine Fish.
Dr. Scient. Thesis, University of Trondheim, Trondheim,
Norway.
Rainuzzo, J.R., Reitan, K.I. & Olsen, Y. (1997) The significance of
lipids at the early stages of marine fish: a review. Aquaculture, 155,
103–115.
Richard, A. (1971) Contribuition a l�etude experimentale de la crois-
sance et de la maturation sexuelle de Sepia officinalis L. (Mollusque,
Cephalopode), p. 264. These no. 248, University of Lille.
Rønnestad, I., Koven, W., Tandler, A., Harel, M. & Fyhn, H.J.
(1998) Utilization of yolk fuels in developing eggs and larvae of
European sea bass (Dicentrarchus labrax). Aquaculture, 162, 157–
170.
Rosa, R., Pereira, J. & Nunes, M.L. (2005) Biochemical composition
of cephalopods with different life strategies, with special reference
to giant squid, Architeuthis sp. Mar. Biol., 146, 739–751.
Sargent, J.R. (1995) Origins and functions of egg lipids: nutritional
implications. In: Broodstock Management and Egg and Larval
Quality (Bromage, N.R. & Roberts, J.R. eds), pp. 353–372. Oxford
University Press, Cambridge.
Sargent, J.R., Bell, M.V., Bell, J.G., Henderson, R.J. & Tocher, D.R.
(1995a) Origins and functions of n-3 polyunsaturated fatty acids in
marine organisms. In: Phospholipids: Characterization, Metabolism
and Novel Biochemical Applications (Ceve, G. & Paltauf, F. eds),
pp. 248–259. American Oil Chemical Society Press, Champaign,
IL.
Sargent, J.R., Bell, J.G., Bell, M.V., Henderson, R.J. & Tocher, D.R.
(1995b) Requirement criteria for essential fatty acids. J. Appl.
Ichthyol., 11, 183–198.
Sargent, J., McEvoy, L., Estevez, A., Bell, G., Bell, M., Henderson,
J. & Tocher, D. (1999) Lipid nutrition of marine fish during early
development: current status and future directions. Aquaculture,
179, 217–229.
Sargent, J.R., Tocher, D.R. & Bell, J.G. (2002) The lipids. In: Fish
Nutrition, 3rd edn (Halver, J.E. & Hardy, R.W. eds), pp. 181–257.
Academic Press, New York.
Silversand, C., Norberg, B. & Haux, C. (1996) Fatty-acid composi-
tion of ovulated eggs from wild and cultured turbot Scophthalmus
maximus in relation to yolk and oil globule lipids. Mar. Biol., 125,
269–278.
Sykes, A.V., Domingues, P.M., Correia, M. & Andrade, J.P. (2006a)
Cuttlefish culture – state of the art and future trends. Vie Milieu,
56, 129–137.
. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .
� 2008 Blackwell Publishing Ltd Aquaculture Nutrition 15; 38–53
No claim to original US government works

Sykes, A.V., Domingues, P.M. & Andrade, J.P. (2006b) Effects of
using live grass shrimp (Palaemonetes varians) as the only source of
food for the culture of cuttlefish, Sepia officinalis (Linnaeus, 1758).
Aquacult. Int., 14, 551–568.
Tocher, D.R. & Sargent, J.R. (1984) Analyses of lipids and fatty
acids in ripe roes of some northwest marine fish. Lipids, 19, 492–
499.
Vaz-Pires, P., Seixas, P. & Barbosa, A. (2004) Aquaculture potential
of the common octopus (Octopus vulgaris Cuvier, 1797): a review.
Aquaculture, 238, 221–238.
Vazquez, R., Gonzalez, S., Rodrıguez, A. & Mourente, G. (1994)
Biochemical composition and fatty acid content of fertilizad
eggs, yolk sac stage larvae and first-feeding larvae of the
Senegal sole (Solea senegalensis Kaup). Aquaculture, 119, 273–
286.
Watanabe, T. (1993) Importance of docosahexanoic acid in marine
larval fish. Aquaculture, 176, 181–188.
Watanabe, T. & Kiron, V. (1994) Prospects in larval fish dietetics.
Aquaculture, 124, 223–251.
Wells (1958) Factors affecting reaction to mysis by newly-hatched
Sepia. Behaviour, 13, 96–111.
Wolf, G., Verheyn, E., Vlaeminck, A., Lemaire, J. & Decleir, W.
(1985) Respiration of Sepia officinalis during embryonic and early
juvenile life. Mar. Biol., 90, 35–39.
Wolfram, K., Mark, F.M., John, U., Lucassen, M. & Portner, H.O.
(2006) Microsatellite DNA variation indicates low levels of genetic
differentiation among cuttlefish (Sepia officinalis L.) populations in
the English Channel and the Bay of Biscay. Comp. Biochem.
Physiol., Part D, 1, 375–383.
Zandee, D.J. (1967) Absence of cholesterol synthesis in Sepia offici-
nalis L. Arch. Internat. de Physiol. Bioch., 75, 487.
Zar, J.H. (1999) Biostatistical analysis. In: Biostatistical Analysis, 4th
edn (Ryu, T. ed.), p. 663. Prentice-Hall Inc., Upper Saddle River,
NJ.
. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .
� 2008 Blackwell Publishing Ltd Aquaculture Nutrition 15; 38–53
No claim to original US government works
Copyright © 2022 FDOKUMEN




















