
Ling-Ling Zheng,1 Qing Jing,2
16
© 2012 Landes Bioscience. Do not distribute. Drastic expression change of transposon-derived piRNA-like RNAs and microRNAs in early stages of chicken embryos implies a role in gastrulation Peng Shao, 1,† Jian-You Liao, 1,† Dao-Gang Guan, 1 Jian-Hua Yang, 1 Ling-Ling Zheng, 1 Qing Jing, 2 Hui Zhou 1 and Liang-Hu Qu 1, * 1 Key Laboratory of Gene Engineering of the Ministry of Education, State Key Laboratory of Biocontrol, Sun Yat-sen University; Guangzhou, China; 2 Key Laboratory of Stem Cell Biology, Institute of Health Sciences, Shanghai Institutes for Biological Sciences, Chinese Academy of Sciences; Shanghai, China † These authors contributed equally to this work. Keywords: microRNA, piRNA, regulation, gastrulation, chicken embryo Recent studies have shown that endogenous small RNAs regulate a variety of biological processes during vertebrate development; however, little is known about the role of small RNAs in regulating developmental signaling pathways during early embryogenesis. In this study, we applied Illumina sequencing to characterize an unexpected endogenous small RNA catalog and demonstrated a dramatic transition from transposon-derived piRNA-like small RNAs (pilRNAs) to microRNAs (miRNAs) in pre- and post-gastrula chicken embryos. The comprehensive expression profile of chicken miRNAs at the pre- and post-gastrula stages revealed that most known and new miRNAs were dynamically regulated during development. In addition to embryonic stem cell-related miRNAs, Gene Ontology (GO) analysis showed that miRNAs enriched in early stage chicken embryos targeted multiple signal transduction pathways associated with the reproductive process and embryogenesis, including Wnt and TGF-b, which specifies the neural fate of blastodermal cells. Intriguingly, a large cohort of pilRNAs primarily derived from the active and most abundant transposable elements (TEs) were enriched in chicken stage X blastoderms. Within stage X blastoderms, pilRNAs were specifically localized to the primordial germ cells (PGCs), indicating their post-zygotic origin. Together, these findings imply a role for small RNAs in gastrulation in early stage chicken embryos. Introduction In recent years, the important regulatory roles of small noncoding RNAs [~20–30 nucleotides (nt)] in diverse developmental pro- cesses have been uncovered in many eukaryotes. 1 Central to the ability of small noncoding RNAs to regulate these processes are the evolutionarily conserved RNAi-related pathways and three major classes of endogenous small RNAs identified in multicellular systems: microRNAs (miRNAs), Piwi-interacting small RNAs (piRNAs), and endogenous small interfering RNAs (endo-siRNAs). In animals, miRNAs and endo-siRNAs share several character- istics. Both of them are typically 21–24 nt in length, are processed by the RNase III enzyme Dicer, and function with Argonaute family proteins to trigger target RNA decay or to induce trans- lational repression. 2 Beyond these common features, miRNAs differ from endo-siRNAs in their molecular origins. miRNAs originate from short hairpin precursors and are widely expressed in various tissues and cell lines. 3 In contrast, animal endo-siRNAs are mainly derived from transposable elements (TEs), comple- mentary annealed transcripts or long hairpin RNAs (hpRNAs). 4 Furthermore, mouse endo-siRNAs are enriched in oocytes 5-7 and embryonic stem (ES) cells. 8 In contrast, piRNAs are produced through the Dicer-independent biogenesis pathway, which results in mature species that are longer (23–30 nt) than miRNAs and siRNAs. 9 Diverse piRNAs are processed from long, single- stranded precursors and classified as either repetitive or non- repetitive piRNAs based on their genomic origins. In particular, vertebrate piRNAs and their associated Piwi family members are largely restricted to the gonads and germ cells. 10 Endo-siRNAs and piRNAs are 2’O-methylated at their 3' end. 11-14 This modification is also found in plant miRNAs and siRNAs 15 but not in most animal miRNAs. Endogenous small RNAs generated through distinct biogenesis pathways play different roles during animal development. miRNAs act as developmental switches 16 and play important roles in many cellular processes, including ES cell self-renewal 17 and differentiation; 18-21 endo-siRNAs regulate the expression of target mRNAs related to microtubule dynamics in mammalian oocytes 6 and influence transposons in mouse oocytes, 6,7 Caenorhabditis elegans, 22 human cultured cells 23 and Drosophila somatic tissues 24 ; and piRNAs represent a major defense against TEs in the germ line 25 and are linked to germ cell maintenance 12 and epigenetic pathways. 26,27 Gastrulation is a key phase in the development of higher animals because fate maps of embryos are specified during *Correspondence to: Liang-Hu Qu; Email: [email protected] Submitted: 07/02/11; Revised: 10/12/11; Accepted: 10/21/11 http://dx.doi.org/10.4161/rna.9.2.18489 RESEARCH PAPER RNA Biology 9:2, 1–16; February 2012; G 2012 Landes Bioscience www.landesbioscience.com RNA Biology 1
Transcript of Ling-Ling Zheng,1 Qing Jing,2

© 2012 Landes Bioscience.
Do not distribute.
Drastic expression change of transposon-derivedpiRNA-like RNAs and microRNAs in early stages ofchicken embryos implies a role in gastrulation
Peng Shao,1,† Jian-You Liao,1,† Dao-Gang Guan,1 Jian-Hua Yang,1 Ling-Ling Zheng,1 Qing Jing,2 Hui Zhou1 and Liang-Hu Qu1,*
1Key Laboratory of Gene Engineering of the Ministry of Education, State Key Laboratory of Biocontrol, Sun Yat-sen University; Guangzhou, China; 2Key Laboratory of Stem Cell
Biology, Institute of Health Sciences, Shanghai Institutes for Biological Sciences, Chinese Academy of Sciences; Shanghai, China
†These authors contributed equally to this work.
Keywords: microRNA, piRNA, regulation, gastrulation, chicken embryo
Recent studies have shown that endogenous small RNAs regulate a variety of biological processes during vertebratedevelopment; however, little is known about the role of small RNAs in regulating developmental signaling pathwaysduring early embryogenesis. In this study, we applied Illumina sequencing to characterize an unexpected endogenoussmall RNA catalog and demonstrated a dramatic transition from transposon-derived piRNA-like small RNAs (pilRNAs) tomicroRNAs (miRNAs) in pre- and post-gastrula chicken embryos. The comprehensive expression profile of chickenmiRNAs at the pre- and post-gastrula stages revealed that most known and new miRNAs were dynamically regulatedduring development. In addition to embryonic stem cell-related miRNAs, Gene Ontology (GO) analysis showed thatmiRNAs enriched in early stage chicken embryos targeted multiple signal transduction pathways associated with thereproductive process and embryogenesis, including Wnt and TGF-b, which specifies the neural fate of blastodermal cells.Intriguingly, a large cohort of pilRNAs primarily derived from the active and most abundant transposable elements (TEs)were enriched in chicken stage X blastoderms. Within stage X blastoderms, pilRNAs were specifically localized to theprimordial germ cells (PGCs), indicating their post-zygotic origin. Together, these findings imply a role for small RNAs ingastrulation in early stage chicken embryos.
Introduction
In recent years, the important regulatory roles of small noncodingRNAs [~20–30 nucleotides (nt)] in diverse developmental pro-cesses have been uncovered in many eukaryotes.1 Central to theability of small noncoding RNAs to regulate these processes arethe evolutionarily conserved RNAi-related pathways and threemajor classes of endogenous small RNAs identified in multicellularsystems: microRNAs (miRNAs), Piwi-interacting small RNAs(piRNAs), and endogenous small interfering RNAs (endo-siRNAs).
In animals, miRNAs and endo-siRNAs share several character-istics. Both of them are typically 21–24 nt in length, are processedby the RNase III enzyme Dicer, and function with Argonautefamily proteins to trigger target RNA decay or to induce trans-lational repression.2 Beyond these common features, miRNAsdiffer from endo-siRNAs in their molecular origins. miRNAsoriginate from short hairpin precursors and are widely expressedin various tissues and cell lines.3 In contrast, animal endo-siRNAsare mainly derived from transposable elements (TEs), comple-mentary annealed transcripts or long hairpin RNAs (hpRNAs).4
Furthermore, mouse endo-siRNAs are enriched in oocytes5-7 andembryonic stem (ES) cells.8 In contrast, piRNAs are produced
through the Dicer-independent biogenesis pathway, which resultsin mature species that are longer (23–30 nt) than miRNAs andsiRNAs.9 Diverse piRNAs are processed from long, single-stranded precursors and classified as either repetitive or non-repetitive piRNAs based on their genomic origins. In particular,vertebrate piRNAs and their associated Piwi family members arelargely restricted to the gonads and germ cells.10 Endo-siRNAsand piRNAs are 2’O-methylated at their 3' end.11-14 Thismodification is also found in plant miRNAs and siRNAs15 butnot in most animal miRNAs. Endogenous small RNAs generatedthrough distinct biogenesis pathways play different roles duringanimal development. miRNAs act as developmental switches16
and play important roles in many cellular processes, including EScell self-renewal17 and differentiation;18-21 endo-siRNAs regulatethe expression of target mRNAs related to microtubule dynamicsin mammalian oocytes6 and influence transposons in mouseoocytes,6,7 Caenorhabditis elegans,22 human cultured cells23 andDrosophila somatic tissues24; and piRNAs represent a majordefense against TEs in the germ line25 and are linked to germ cellmaintenance12 and epigenetic pathways.26,27
Gastrulation is a key phase in the development of higheranimals because fate maps of embryos are specified during
*Correspondence to: Liang-Hu Qu; Email: [email protected]: 07/02/11; Revised: 10/12/11; Accepted: 10/21/11http://dx.doi.org/10.4161/rna.9.2.18489
RESEARCH PAPER
RNA Biology 9:2, 1–16; February 2012; G 2012 Landes Bioscience
www.landesbioscience.com RNA Biology 1

© 2012 Landes Bioscience.
Do not distribute.
gastrulation. The relative transparency and accessibility of thechicken (Gallus gallus) embryo make it better suited to studies ofthe process of gastrulation than other amniotes. At the egg layingstage (stage X EG&K),28 a two-layered disc somewhat like theinner cell mass (ICM) is present. The chicken stage X blastoderm(pre-gastrula stage) consists of about 40000–60000 undifferenti-ated embryonic cells that have the capacity for in vitro proliferationand differentiation into embryoid bodies.29 During gastrulation,mass migrations of these undifferentiated cells result in a massivereorganization of the embryo from a two-layered to three-layeredorganism. The chicken embryo thus provides an ideal systemfor studying both the relationship between molecular changesand maintenance of the undifferentiated stem cell phenotype ordifferentiation and germ layer specification. After approximately30 h of incubation, embryos reach the HH10 stage (post-gastrula).30 By this time, the fate maps of the chicken embryosare well determined, and the heart proper begins to develop.Attempts to understand the protein-based molecular mechanismsunderlying the control of pluripotency and differentiationcommitment in early embryonic patterning have suggested rolesfor the Wnt, bone morphogenetic protein (BMP), Nodal andfibroblast growth factor (FGF) signaling pathways.31 Althoughemerging evidence for the role of miRNAs in controlling signaltransduction pathways in animal cells32 provides some cluesregarding miRNA-mediated modulation of the developmentalprogram, detailed knowledge of which small RNAs are activatedand how these small RNAs regulate various signaling pathwaysthat contribute to early development during gastrulation remainsvery limited.
In this study, we applied Solexa/Illumina sequencing tocharacterize an unexpected small RNA population includingmiRNAs and TE-derived piRNA-like RNAs (pilRNAs) in earlychicken embryos at stage X and stage HH10 and determined theexpression of these small RNAs at diverse developmental stages.Although recent studies have identified miRNAs in adult chickensand early embryos,33-39 our work represents the systematic investi-gation of the miRNAs and TE-derived pilRNAs in pre- and post-gastrula chicken embryos. Importantly, our findings reveal anoverall transition from expression of TE-derived pilRNAs toexpression of miRNAs during early chicken development.Detailed analysis of these small RNAs provided unique insightinto the miRNA-mediated regulation of various signaling path-ways in gastrulation and the origin of TE-pilRNAs in theprimordial germ cells (PGCs) of chicken stage X blastoderms.
Results
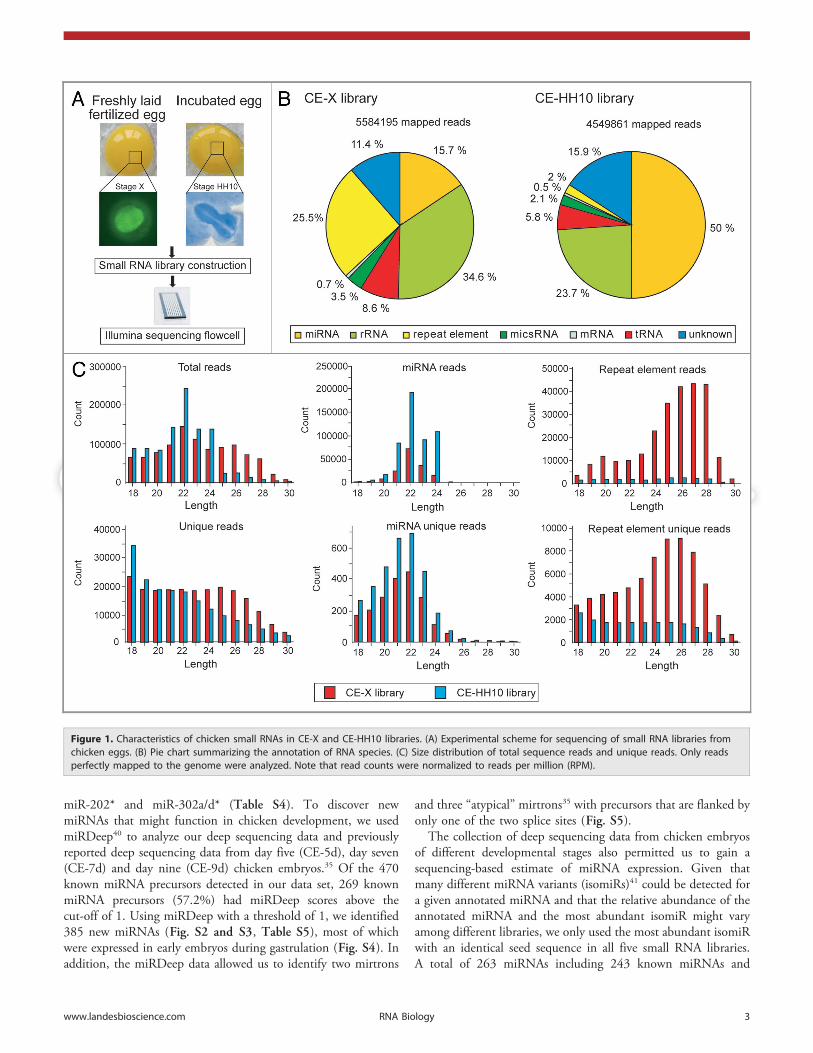
Chicken embryos express different classes of small RNAs. Toidentify small RNAs involved in the early development of chickenembryos, we constructed small RNA libraries from stage X (CE-X) and stage HH10 (CE-HH10) chicken embryos (Fig. 1A).Deep sequencing of the CE-X and CE-HH10 libraries yielded756,323 and 454,452 unique reads represented by 8,151,692and 6,545,706 total reads, respectively (Table S1). Approximately70% of the sequences obtained matched the current release of thechicken genome (galGal 3) perfectly, and these were considered
for further analysis (Fig. 1B). To investigate the diversity andabundance of the sequence reads, we analyzed the total reads andunique reads derived from the known RNA catalog and genomicrepeats. We found that the CE-X library contained significantlymore small RNAs derived from repetitive elements (~25.6%) butfewer mapped to known miRNAs (~15.6%) than the CE-HH10library. In contrast, in the CE-HH10 library, known miRNAsaccounted for ~50% of the sequence reads, whereas the smallRNA populations contained very few repetitive elements (~2.1%).These results indicate that small RNAs derived from the repetitiveelements are highly enriched in the CE-X library, whereas maturemiRNA sequences are enriched in the CE-HH10 library. Allremaining sequence reads assigned to the other annotatednoncoding RNAs (rRNAs, tRNAs, snRNAs snoRNAs and theother miscRNAs) were present at similar frequencies in bothlibraries. Although the sequence reads derived from knownmiRNAs were the most abundant component of the annotatedsmall RNA reservoir in each library, these sequence reads com-posed only a relatively small fraction (1–2%) of the total numberof unique reads. The largest fraction of the unique sequences (36–50%) mapped to un-annotated genomic regions (Table S1).
We next analyzed the size distribution of total reads andunique reads in the CE-X and CE-HH10 libraries (Fig. 1C).Analysis of the sequence reads showed that the small RNAs inthese libraries displayed unimodal length distributions. The mostabundant reads in these two libraries were ~22 nt long; this size ischaracteristic of miRNAs. When the total reads and unique readsmapped to the annotated RNA species were subtracted from theCE-X library, the 22-nt peak was replaced by a broad 25–28 ntpeak (Fig. S1); this size is characteristic of piRNAs. Consistentwith the report that vertebrate piRNAs often map to the repetitiveelements,1 this pilRNA peak was striking even among readsmapped to genomic repeats (Fig. 1C). In contrast, the pilRNApeak was not present within the CE-HH10 library even when thelength distribution of only the sequences mapped to the repeti-tive elements was analyzed, implying that the CE-HH10 smallRNA library is deficient in piRNAs. Furthermore, no significantpeaks of a size characteristic of animal siRNAs (~21 nt) werefound among repetitive elements. Therefore, our data indicatethat miRNAs and TE-derived small RNA are the two mostabundant types of small RNAs in chicken stage X blastodermalcells, while miRNAs predominate the small RNA population instage HH10 embryos.
Temporal regulation of miRNA expression in early chickenembryos. Because both of the libraries contained abundant readsof ~22-nt RNAs, we initially searched the sequences against544 annotated G. gallus miRNAs (miRBase release 16.0). Werecovered 510 known mature miRNAs (Table S2) and corrected194 known miRNA sequences and identified 143 miRNAsprocessed from the opposite arms of the known maturemiRNAs (Table S3). More than 65% (359/544) of the knownmiRNAs were expressed in pre-gastrula stage embryos. Among thetop 20 most abundant miRNAs, 12 were present in both samples(Table 1). We observed unusually high cloning recovery ratesof several miRNAs that were derived from the arm of miRNAprecursor opposite to the annotated mature miRNAs, such as
2 RNA Biology Volume 9 Issue 2
© 2012 Landes Bioscience.
Do not distribute.
miR-202* and miR-302a/d* (Table S4). To discover newmiRNAs that might function in chicken development, we usedmiRDeep40 to analyze our deep sequencing data and previouslyreported deep sequencing data from day five (CE-5d), day seven(CE-7d) and day nine (CE-9d) chicken embryos.35 Of the 470known miRNA precursors detected in our data set, 269 knownmiRNA precursors (57.2%) had miRDeep scores above thecut-off of 1. Using miRDeep with a threshold of 1, we identified385 new miRNAs (Fig. S2 and S3, Table S5), most of whichwere expressed in early embryos during gastrulation (Fig. S4). Inaddition, the miRDeep data allowed us to identify two mirtrons
and three “atypical” mirtrons35 with precursors that are flanked byonly one of the two splice sites (Fig. S5).
The collection of deep sequencing data from chicken embryosof different developmental stages also permitted us to gain asequencing-based estimate of miRNA expression. Given thatmany different miRNA variants (isomiRs)41 could be detected fora given annotated miRNA and that the relative abundance of theannotated miRNA and the most abundant isomiR might varyamong different libraries, we only used the most abundant isomiRwith an identical seed sequence in all five small RNA libraries.A total of 263 miRNAs including 243 known miRNAs and
Figure 1. Characteristics of chicken small RNAs in CE-X and CE-HH10 libraries. (A) Experimental scheme for sequencing of small RNA libraries fromchicken eggs. (B) Pie chart summarizing the annotation of RNA species. (C) Size distribution of total sequence reads and unique reads. Only readsperfectly mapped to the genome were analyzed. Note that read counts were normalized to reads per million (RPM).
www.landesbioscience.com RNA Biology 3

© 2012 Landes Bioscience.
Do not distribute.the top 20 most abundant miRNA candidates identified in thisstudy were selected for analysis (Fig. 2A, Table S6). We analyzedthe relative expression of miRNAs by normalizing each miRNAcount from each library to the total number of miRNA hairpin-matching reads for that library. This normalization techniqueprovided consistent results when used to calculate the relativeexpression of the most abundant isomiR and all isomiRs (mostabundant isomiRs plus the other isomiRs) (Spearman’s correla-tion coefficient = 0.988, Fig. S6). Furthermore, consistent witha previous report that the read count of the most abundantisomiR can be used to accurately estimate differences in miRNAexpression levels between libraries,41 the relative expression ofthe most abundant miRNA correlated well with the q-PCRmeasurements (Fig. 2E, Fig. S7, Spearman’s correlationcoefficient = 0.836).
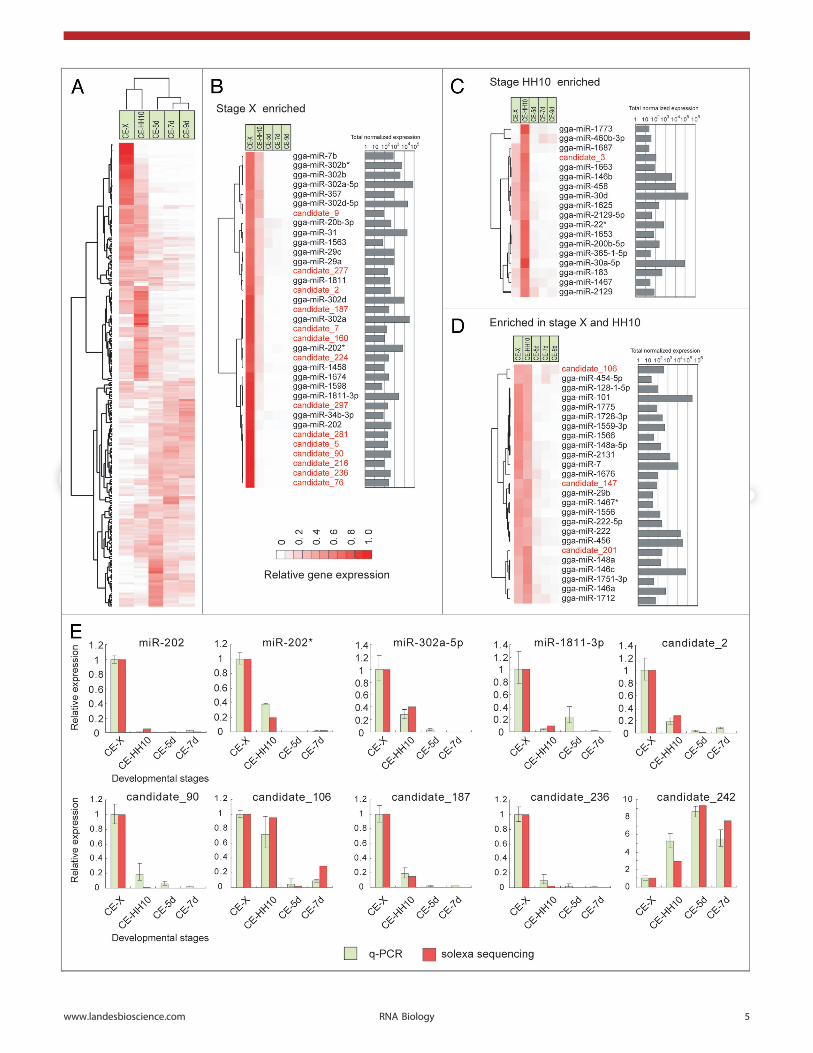
Among the 263 miRNAs selected, 137 showed significantchanges (p , 0.001) in expression between the CE-X and CE-HH10 libraries (Table S7), and only 28 miRNAs exhibited nosignificant differences in expression levels throughout the fivedevelopmental stages we examined. Nearly 40% of the 263miRNAs presented a stage-restricted expression pattern, repre-senting more than 50% of total miRNA reads across the five smallRNA libraries (Fig. 2A). Thirty-five and 18 miRNAs were expres-sed primarily (.60% of their library-normalized reads derivingfrom a single library) at stage X and stage HH10, respectively(Fig. 2B, 2C), whereas 25 miRNAs were enriched at both ofthese two stages and represented more than 75% of the totalnormalized expression levels (Fig. 2D). miR-30d and miR-101were the two most abundant miRNAs in stage X and HH10embryos, whereas some other conserved miRNAs, especially miR-302a-5p/3p and miR-302d-5p/3p, were expressed at much higherlevels at stage X but were dramatically downregulated at laterdevelopmental stages. Several members of the miR-302 familywere cloned only from undifferentiated ES cells42,43 and linked tomaintaining pluripotency and controlling the mesendodermal fatespecification.44 Although let-7 family members are present atlow levels in mammalian ES cells, several members of the let-7family exhibited a higher expression level at stage X than atstage HH10, and these miRNAs gradually accumulated asdevelopment progressed. The other three miRNAs enriched atstage X (miR-202, miR-202* and miR-1811–3p) exhibitedexpression patterns similar to that of miR-302a/d. This findingwas validated using the q-PCR assay (Fig. 2E). Although miR-202* was reported to be a germline-enriched miRNA in Xenopus,45
our data suggested that these three miRNAs were involved in earlyembryonic development. Of the top 20 most abundant newmiRNA candidates, 14 were expressed exclusively at stage X, andthe exclusive expression of six of these miRNAs at stage X wasconfirmed by q-PCR (Fig. 2E). These experiments thus demon-strated the highly dynamic expression patterns of miRNAs inchicken embryos, suggesting unique roles for these new miRNAsin blastodermal development.
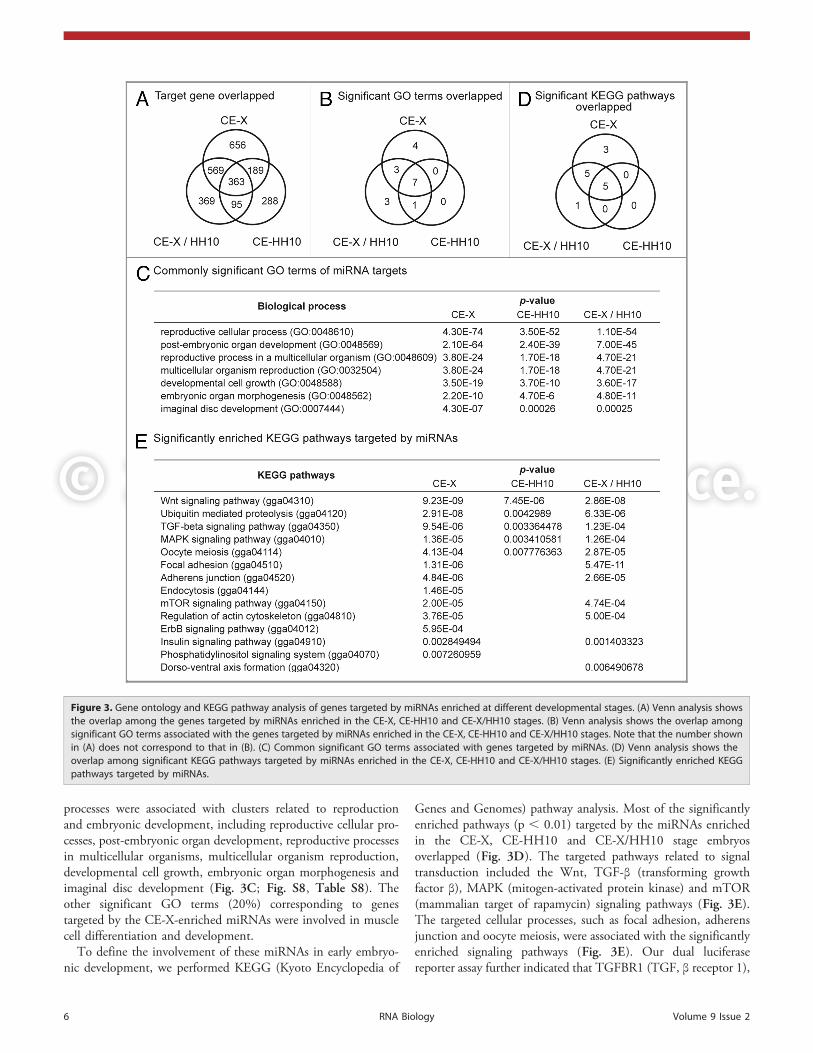
Key GO terms and signaling pathways regulated by miRNAs.To gain insight into the functional consequences of chickenmiRNAs enriched in the CE-X stage, the CE-HH10 stage orboth the CE-X and CE-HH10 stages of chicken embryos(CE-X/HH10), we used TargetScan46 to identify targets of thesemiRNAs that were conserved in vertebrates. Although thenumber of common targets predicted for miRNAs enriched inthe CE-X, CE-HH10 and CE-X/HH10 stages was relativelysmall (less than 40% of predicted targets; Fig. 3A), more than50% of the significantly overrepresented Gene Ontology (GO)47
terms (p , 0.0001) representing biological processes overlapped(Fig. 3B). Overrepresented GO terms related to biological
Table 1. The top 20 most abundant miRNAs in each sample
CE-X library CE-HH10 library
miRNA Read Percentage(%)
miRNA Read Percentage(%)
miR-101 24642.76 15.74 miR-30d 57176.69 11.42
miR-30d 9097.46 5.81 miR-101 54155.06 10.81
miR-302a-5p 7780.71 4.97 miR-30a-5p 36303.31 7.25
miR-140* 6773.22 4.33 miR-128 26904.12 5.37
miR-302a 4132.56 2.64 miR-140* 22470.80 4.49
let-7f 4060.75 2.59 miR-146c 17636.36 3.52
miR-1a 3620.04 2.31 miR-1a 14075.16 2.81
miR-146c 3352.86 2.14 miR-148a 11290.02 2.25
miR-122 2457.29 1.57 miR-302a-5p 10076.57 2.01
let-7a/j 2345.01 1.50 miR-130a 9533.26 1.90
miR-148a 2241.68 1.43 miR-103 8124.86 1.62
miR-302d-5p 2181.16 1.39 miR-130b 7094.94 1.42
miR-456 2130.48 1.36 miR-456 6105.02 1.22
miR-31 2073.53 1.32 miR-26a 5494.89 1.10
miR-130a 1645 1.05 miR-30e 5242.36 1.05
miR-128 1544.36 0.99 miR-221 4936.41 0.99
miR-103 1533.08 0.98 miR-21 3853.3 0.77
miR-21 1378.35 0.88 miR-107 3833.52 0.77
miR-302c 1312.28 0.84 miR-222 3702.31 0.74
miR-222 1216.83 0.78 miR-458 3631.54 0.73
The read number shown on the right side of each miRNA is normalized tothe total number of sequence reads that matched the chicken genomeperfectly in each library. The percentage of each miRNA among the totalknown miRNA population in the CE-X and CE-HH10 libraries is calculated.miRNAs detected in both libraries are underlined.
Figure 2 (See opposite page). Expression of chicken miRNAs. (A) The total expression profile of the chicken miRNAs (left) and the summed normalizedexpression levels (right) across all five libraries. Normalized counts for each miRNA in each library are provided (Table S6). The left panel shows theexpression patterns of chicken miRNAs enriched in stage X (B) or stage HH10 (C) embryos or embryos of both stages (D). The right panel shows thesummed normalized expression levels across all five libraries. Novel miRNAs are labeled in red. The genes and time-points were clustered with averagelinkage using the centered correlation coefficient. (E) Gene expression profiles following normalization of miRNAs compared with those determined byq-PCR assay (mean ± SD, n = 3).
4 RNA Biology Volume 9 Issue 2
© 2012 Landes Bioscience.
Do not distribute.
www.landesbioscience.com RNA Biology 5
© 2012 Landes Bioscience.
Do not distribute.
processes were associated with clusters related to reproductionand embryonic development, including reproductive cellular pro-cesses, post-embryonic organ development, reproductive processesin multicellular organisms, multicellular organism reproduction,developmental cell growth, embryonic organ morphogenesis andimaginal disc development (Fig. 3C; Fig. S8, Table S8). Theother significant GO terms (20%) corresponding to genestargeted by the CE-X-enriched miRNAs were involved in musclecell differentiation and development.
To define the involvement of these miRNAs in early embryo-nic development, we performed KEGG (Kyoto Encyclopedia of
Genes and Genomes) pathway analysis. Most of the significantlyenriched pathways (p , 0.01) targeted by the miRNAs enrichedin the CE-X, CE-HH10 and CE-X/HH10 stage embryosoverlapped (Fig. 3D). The targeted pathways related to signaltransduction included the Wnt, TGF-β (transforming growthfactor β), MAPK (mitogen-activated protein kinase) and mTOR(mammalian target of rapamycin) signaling pathways (Fig. 3E).The targeted cellular processes, such as focal adhesion, adherensjunction and oocyte meiosis, were associated with the significantlyenriched signaling pathways (Fig. 3E). Our dual luciferasereporter assay further indicated that TGFBR1 (TGF, β receptor 1),
Figure 3. Gene ontology and KEGG pathway analysis of genes targeted by miRNAs enriched at different developmental stages. (A) Venn analysis showsthe overlap among the genes targeted by miRNAs enriched in the CE-X, CE-HH10 and CE-X/HH10 stages. (B) Venn analysis shows the overlap amongsignificant GO terms associated with the genes targeted by miRNAs enriched in the CE-X, CE-HH10 and CE-X/HH10 stages. Note that the number shownin (A) does not correspond to that in (B). (C) Common significant GO terms associated with genes targeted by miRNAs. (D) Venn analysis shows theoverlap among significant KEGG pathways targeted by miRNAs enriched in the CE-X, CE-HH10 and CE-X/HH10 stages. (E) Significantly enriched KEGGpathways targeted by miRNAs.
6 RNA Biology Volume 9 Issue 2

© 2012 Landes Bioscience.
Do not distribute.
a component of TGF-β pathway, is a direct target of a newmiRNA candidate (miR-c236) (Fig. 4). Some of the signalingpathways were over-represented only among the predicted targetsof CE-X-enriched or CE-X/HH10-enriched miRNAs. Notably,the most well-represented pathway was Wnt signaling. Toinvestigate whether this most highly enriched pathway wasspecifically targeted by miRNAs enriched in the early stageembryos, we further analyzed the targeted pathways defined byKEGG analysis by randomly generating (1000 times) the samenumber of CE-X and CE-HH10 miRNA target genes asmiRNAs enriched in the CE-X and CE-HH10 libraries. Theabsence or modest enrichment of Wnt signaling inferred fromrandomly selected target genes (Fig. S9) confirmed that Wntsignaling was the most highly enriched pathway specifically andcommonly targeted by miRNAs enriched in the chickenembryos at stage X and/or stage HH10.
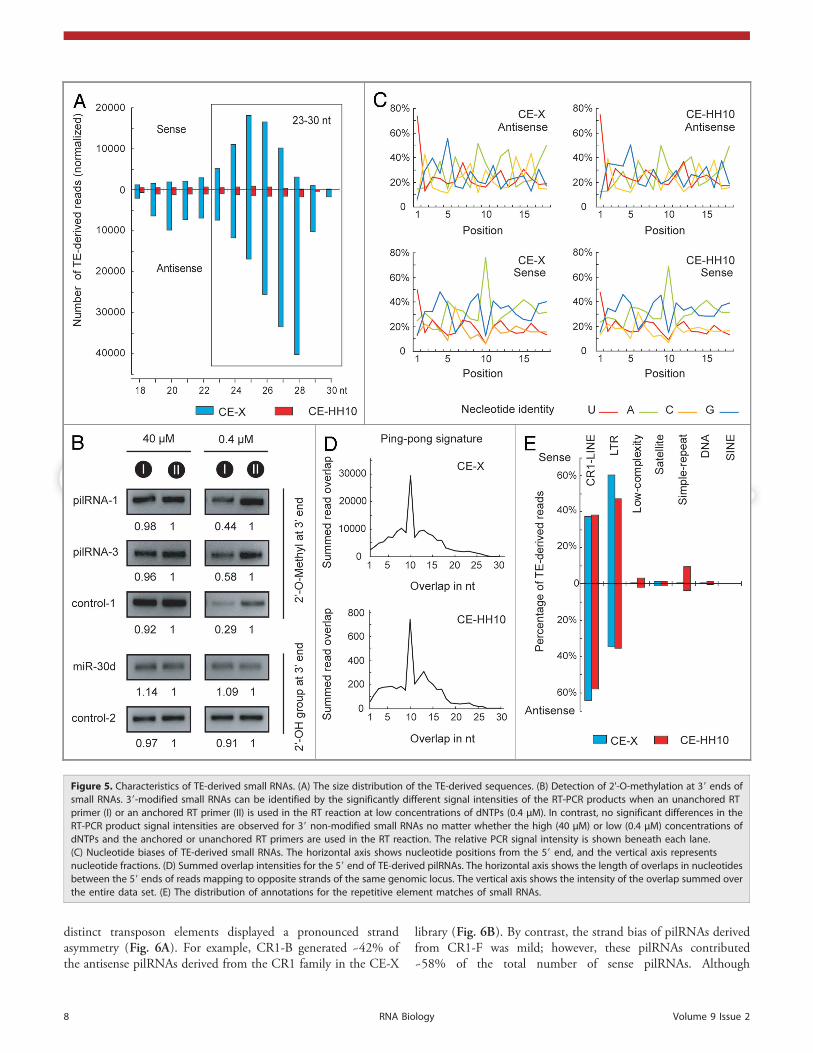
TE-derived pilRNAs in chicken embryos. Repetitive genomicelements, and particularly TEs, are the major source of piRNAsin vertebrate12,48-50 and fruit fly germ lines.51,52 The size distri-bution of TE-derived reads in our libraries showed that 23–30-ntRNAs were prominent (~80% of the TE-derived reads) in theCE-X library but underrepresented in the CE-HH10 library(Fig. 5A). Approximately 70% of the TE-derived small RNAswere antisense to TEs, and their size distribution peaked at 27–28 nt; however, the size distribution peak shifted to 25–26 ntwhen the sense-TE-derived reads were analyzed. This size distri-bution was consistent with the lengths of piRNAs observed inother animals. The predominant class of TE-derived RNAsranging from 23–30 nt in size was thus referred to as pilRNAs.
Our 2’-O-methylation assay detected modified 3' ends in thechicken pilRNAs, while within the same samples, the miRNAtermini were unmodified (Fig. 5B, Fig. S10).
The pilRNA populations were quite diverse because 25% ofthe RNAs were cloned only once and most of the pilRNAs(67%) matched multiple genomic loci. The top 20 TE-derivedpilRNAs (Table S9) accounted for approximately 20% of thetotal number of pilRNAs. Our RT-qPCR assay confirmed theirprominent expression in stage X chicken embryos (Fig. S11). Ofthe TE-derived pilRNAs, the antisense-oriented species presenteda marked uracil bias (~74%) at their 5' end, whereas the sense-oriented species showed a strong tendency (~70%) for adenosineat position 10 (Fig. 5C). A strong signal corresponding to a 10-ntoverlap of the 5' ends of the sense and antisense pilRNAs wasdetected in the CE-X library (Fig. 5D). In contrast, the 10-ntoverlapping signal was greatly reduced in the CE-HH10 sample.This unique observation of complementary pilRNA pairs isreminiscent of a piRNA amplification loop mechanism proposedby Brennecke and colleagues,52 suggesting that the ping-pongmodel of piRNA biogenesis is also conserved in chickens.
The TE-derived pilRNAs in our libraries preferentiallyoriginated from long-terminal repeats (LTR) and the CR1 familyof non-LTR retrotransposons (Fig. 5E). Upon further analysis, itbecame obvious that chicken pilRNAs displayed bias towardparticular families within transposon classes (Table S10). Forinstance, the CR1-B and CR1-F subfamilies produced, respect-ively, 28.5% and 28.29% of the pilRNAs derived from the CR1family. In contrast, the other subfamilies of CR1 generated fewerreads. As observed in zebrafish, chicken pilRNAs derived from
Figure 4. miR-c236 directly targets the chicken TGFBR1 39-UTR. (A) Quantitative analyses of the miR-c236 expression in the CE-X and CE-HH10 by theRT-PCR assay, respectively. The size of RT-PCR product is in line with our expectation. U6 snRNA was used as a loading control. (B) Schema of miR-c236binding site in the TGFBR1 39-UTR sequence of Gallus gallus (Gga, NM_204246), Homo sapiens (Hsa, NM_004612) and Mus musculus (Mmu, NM_009370).The mutated sequences in the TGFBR1 39-UTR fragment is underlined. The putative target site are shown in bold. (C) Target validation using a reporterassay. HEK293T cells were cotransfected with TGFBR1 39-UTR-wt with either miR-c236-mimic or miR-NC-mimic (left), and TGFBR1 39-UTR-mut with eithermiR-c236-mimic or miR-NC-mimic (right). Data are means of at least three independent experiments done in duplicate; error bars indicated the SD**p , 0.0001, compared with miR-NC-mimic transfected cells. NC, negative control.
www.landesbioscience.com RNA Biology 7
© 2012 Landes Bioscience.
Do not distribute.
distinct transposon elements displayed a pronounced strandasymmetry (Fig. 6A). For example, CR1-B generated ~42% ofthe antisense pilRNAs derived from the CR1 family in the CE-X
library (Fig. 6B). By contrast, the strand bias of pilRNAs derivedfrom CR1-F was mild; however, these pilRNAs contributed~58% of the total number of sense pilRNAs. Although
Figure 5. Characteristics of TE-derived small RNAs. (A) The size distribution of the TE-derived sequences. (B) Detection of 2'-O-methylation at 39 ends ofsmall RNAs. 39-modified small RNAs can be identified by the significantly different signal intensities of the RT-PCR products when an unanchored RTprimer (I) or an anchored RT primer (II) is used in the RT reaction at low concentrations of dNTPs (0.4 μM). In contrast, no significant differences in theRT-PCR product signal intensities are observed for 39 non-modified small RNAs no matter whether the high (40 μM) or low (0.4 μM) concentrations ofdNTPs and the anchored or unanchored RT primers are used in the RT reaction. The relative PCR signal intensity is shown beneath each lane.(C) Nucleotide biases of TE-derived small RNAs. The horizontal axis shows nucleotide positions from the 59 end, and the vertical axis representsnucleotide fractions. (D) Summed overlap intensities for the 59 end of TE-derived pilRNAs. The horizontal axis shows the length of overlaps in nucleotidesbetween the 59 ends of reads mapping to opposite strands of the same genomic locus. The vertical axis shows the intensity of the overlap summed overthe entire data set. (E) The distribution of annotations for the repetitive element matches of small RNAs.
8 RNA Biology Volume 9 Issue 2

© 2012 Landes Bioscience.
Do not distribute.
TE-derived pilRNAs mainly originated from the antisense strand,some TEs preferentially generated sense pilRNAs. For example,the ERVK family generated over eight times more sense pilRNAsthan antisense pilRNAs. In particular, the RSV-LTR-int elementexclusively produced sense pilRNAs. As observed in Drosophilaand mammals and in contrast to findings in Xenopus,45,50 thechicken DNA transposons generated only a minor fraction ofthe TE-derived small RNAs (~0.2%). In addition, the chickenpilRNAs were nearly absent from the short interspersed nucleo-tide elements (SINEs).
pilRNA expression in PGCs of chicken stage X blastoderms.piRNAs are usually identified in germ cells or gonads. Becausethe blastoderm is mainly composed of undifferentiated ES cellsand a small number of primordial germ cells (PGCs), which areparticularly dense in the central zone of the area pellucida in stageX embryos,53 the observation of abundant pilRNAs in chickenblastoderms drove us to examine their expression pattern in moredetail. We first performed whole-mount in situ hybridization(WISH) using locked nucleic acid (LNA) oligonucleotides asprobes to detect the most abundant pilRNA species (pilRNA-1)
Figure 6. Profound strand bias of pilRNAs. (A) pilRNAs mapped to diverse transposable elements. The upper panel shows a heat map indicating thestrand bias of TE-derived pilRNAs with respect to canonical transposon sequences. Transposons are grouped into long-terminal repeat (LTR) elementsand long interspersed nuclear elements (LINE). Blue intensities denote the degree of the strand bias. The lower panel shows the number of normalizedreads (reads per million, RPM) mapping to individual transposon elements indicated as in the upper panel. (B) Distribution of pilRNAs on the LINEretrotransposon CR1-F and CR1-B consensus sequences is shown for the indicated small RNA libraries. The number of pilRNAs was normalized to RPM.
www.landesbioscience.com RNA Biology 9

© 2012 Landes Bioscience.
Do not distribute.in our libraries. These experiments demonstrated that thepilRNAs were localized to the central zone of the area pellucida(Fig. 7A). Next, we probed samples for the two most specificmarkers for PGCs, Vasa and Dazl54 to investigate the distributionof PGCs in the chicken blastoderm (Fig. 7B, 7C). The locali-zation of both of these mRNA transcripts largely mirrored that ofthe pilRNAs; however, consistent with previous findings, thechicken ES cell-specific marker cENS-1 was expressed throughoutthe area pellucida (Fig. 7D).55 The similar localization of pilRNAsand PGC-specific markers indicated that the pilRNAs detected inthe chicken blastoderm were mainly produced in the PGCs.
Discussion
Dramatic transition between miRNAs and pilRNAs duringgastrulation. The expression of small RNAs, and especiallymiRNAs, has been extensively investigated at different develop-mental stages in animals; however, little is known about theirexpression patterns and functions in gastrulation. Here, we reporta comprehensive miRNA expression profile in chicken embryosat a variety of developmental stages and demonstrate a com-pletely opposite expression pattern of miRNAs and pilRNAs atpre- and post-gastrula stages. miRNAs were dramatically upregu-lated during early chicken development from stage X to stageHH10, suggesting that these miRNAs are involved in deve-lopmental switches. In sharp contrast to miRNAs, pilRNAswere highly abundant in the small RNA population of chickenstage X blastoderm cells and significantly diminished at the
post-gastrula stage. A similar change in the expression of smallRNA classes was also found during mouse pre-implantationdevelopment.56 Compared with the small RNA population in themouse blastocyst, TE-derived pilRNAs are more abundant inchicken blastoderms, and the TE-derived endo-siRNA compon-ent is underrepresented (~0.02% of the total small RNApopulation). These findings reflect the different requirementsfor siRNAs and piRNAs/pilRNAs in pre-gastrula embryos of thesetwo species.
miRNAs contribute to the establishment and maintenanceof ES cell identity.57 Our data revealed that the complexity andexpression level of miRNAs rapidly increases during the trans-ition from stage X blastoderms to stage HH10 embryos, whereasthe ES cell-specific miRNAs, the miR-302 cluster, and many newmiRNAs identified in this study are markedly downregulatedfollowing gastrulation. Several ES cell-specific miRNA clusters,including miR-302 and the mouse miR-290 family (humanmiR-371–373), have been characterized in mouse and humanES cells.42,43 These miRNAs share a similar seed sequence andlikely play a role in ES cell biology, such as promoting ES cellproliferation and cell cycle G-S1 transition.58 Given that mousemiR-290–296 (human miR-371–373) are absent in the chickenand other non-mammalian vertebrates, the miR-302 family isproposed to be the key regulator of tetrapod ES cells. In thisstudy, the miR-302 family was highly enriched (~16%) in thesmall RNA population of chicken blastodermal cells but down-regulated in post-gastrula embryos, suggesting that these EScell-enriched miRNAs play important roles in maintaining“stemness.” Interestingly, the comparison of the top 40miRNAs in the chicken blastoderm and the mouse blastocystrevealed that only a few of these miRNAs overlap (data notshown), indicating that different sets of miRNAs are requiredin chicken and mouse pre-gastrula embryos. In contrast, chickenstage X blastoderms shared a common set of miRNA specieswith human ES cells,41 such as the miR-302 family, miR-30dand let-7a, suggesting that chicken and human ES cells mightshare a conserved pathway through which miRNAs mediatethe maintenance of a pluripotent state. It should be noted thatmiR-101 was the most abundant miRNA cloned from stage Xblastoderms and was downregulated during the other develop-mental stages, whereas this miRNA is repressed in mammalianES cells and blastocysts.56 Although the function of miR-101 inES cell regulation is unclear, the recent report that miR-101 hasprofound effects on cancer cells and ES cells by regulating theepigenetic factor EZH259 might shed light on its function inearly development because EZH2 is required for gastrulation andES cell derivation.60
In addition to the ES cell-related miRNAs, miRNAs enrichedin the early chicken embryos (before stage HH10) were linkedto early embryogenesis, including reproductive processes andembryonic organogenesis (Fig. 3; Table S8). These miRNAscommonly targeted various signaling pathways that are impor-tant for gastrulation. Notably, the Wnt signaling pathway wasthe most significantly over-represented KEGG pathway that wasnegatively regulated by miRNAs at the pre- and post-gastrulastages. Inhibition of Wnt signaling by miRNAs at different
Figure 7. Expression patterns of pilRNAs (A), Cvh (B), Dazl (C) and cENS-1(D) in chicken stage X blastoderms as revealed by ISH of whole-mountembryos. The localization of pilRNA-1 and PGC-specific markers wasindicated with the white dashed circle. Representative data are shownfrom three to four independent experiments. Bars, 1 mm.
10 RNA Biology Volume 9 Issue 2

© 2012 Landes Bioscience.
Do not distribute.
embryonic stages plays distinct roles during embryogenesis.Blocking Wnt signaling initiates neural differentiation in stageX medial epiblast cells,61 and continued inhibition of Wntsignaling induces normal heart formation.62 Our study indicatedthat miRNAs are involved in the inhibition of Wnt signalingin early chicken embryos and promote the neural fate of thestage X blastodermal cells. The TGF-β signaling pathway, whichmust be downregulated for neural induction,63 was also targetedby these miRNAs (Fig. 4). Consistent with the developmentalrequirement for normal neural induction, the FGF signalingpathway, which is crucial for the suppression of TGF-β signalingand for acquisition of neural fate,63 was not over-represented inthe KEGG pathways targeted by the miRNAs. Notably, severalmesoderm differentiation-associated miRNAs, such as the miR-30family and miR-1, were highly expressed in chicken blastodermalcells. miR-30 family miRNAs have been shown to be involved inthe epithelial-to-mesenchymal transition,64 and the enhancedexpression of miR-30d can promote cancer cell migration andinvasion.65 The upregulation of miR-30 family members in CE-HH10 embryos and downregulation in late stage embryosindicates that miR-30 has an essential function in gastrulation,during which the morphology of the embryos is dramaticallyrestructured by cell migration. miR-1 is a muscle-specific miRNAthat promotes mesoderm differentiation in mouse ES cells.66 Itshigh expression in chicken blastodermal cells may be essential forthe development of the heart, which is the first organ to formand function in vertebrate embryos. Our data suggest that thecells in the growing blastoderms remain undifferentiated butthat the developmental fate of many of these cells is alreadyspecified, if not completely determined.
A subset of piRNAs was proposed to be involved in trans-lational regulation, mRNA stability and epigenetic control, andtransposon silencing during early embryo development. Althoughthe activity of TE elements in chicken embryos is not currentlywell understood, the dynamic expression patterns of TE-derivedsmall RNAs may reflect the activity state of TEs and the necessityfor regulation of the expression of these “selfish genetic elements”at early development stages. Our study suggests that transposonactivity is under tight control of the small RNA-based defensesystem not only in the germ cells but also in the undifferentiatedcells of pre-gastrula embryos.
pilRNAs detected in PGCs of chicken stage X blastoderms.Endogenous small RNAs including piRNAs play important rolesin early embryogenesis. Although Drosophila and vertebratepiRNAs are generally detected in the germ cells or gonads, thenucleotide characteristics and length distribution of the TE-derived small RNA sequences indicated abundant pilRNAs inthe chicken stage X blastodermal extracts. Considering thatundifferentiated stem cells share some characteristics with thegerm line stem cells from which piRNAs are generally derived andthat a recent study has shown that PGCs appear sporadicallyamong poly-ingressing cells in the stage X blastoderm,67 wefurther applied WISH to detect the expression of pilRNAs inthe blastodermal cells. The similar localization of pilRNAsand PGC-specific markers revealed by the WISH experimentindicated that chicken pilRNAs of the stage X blastoderm were
derived from the PGCs; however, we could not rule out thepossibility that chicken pluripotent ES cells might also producepilRNAs. In fact, maternally derived Piwi proteins have beendetected not only in PGCs and germ cells but also in earlyembryos,12 and piRNAs were identified in mouse early embryosduring pre-implantation development,56 human ES cells,41
Planarian neoblasts68 and even the central nervous system.69
Because piRNAs are maternally inherited,70 the frequency ofpiRNAs in the small RNA population progressively decreasesover the course of development.56 In our study, highly abundantpilRNAs were predominantly expressed in the chicken PGCs.Considering that the chicken PGCs are derived from the chickenstage X epiblast, the pilRNAs detected in PGCs might bepost-zygotically produced rather than maternally deposited.Extensive characterization of the piRNA status of the PGCs andthe other pluripotent undifferentiated cells might improve theunderstanding of the biological function of these small RNAsduring early development.
TE-derived piRNAs expressed in germ cells preferentiallytarget certain transposons, such as LTR and LINE elements, bymechanisms whose details remain elusive. Our study revealedthat the amount of small RNAs derived from transposableelements seems to be associated with the abundance and activityof transposons in the chicken genome. Over 98% of the smallRNAs were derived from the CR1 LINE and LTR families, whichare the two most abundant elements in the chicken genome.Notably, small RNAs that originated from CR1-B and CR1-F,the most recently active CR1 elements in the chicken genome,71
were more abundant than those derived from the other CR1elements in the chicken embryos. Similar findings have beenreported for the abundant small RNAs derived from GGERVLsubfamilies, the endogenous retrovirus-like elements that maystill be active in the genome.71 Consistent with the absence ofSINEs in the chicken genome, the proportion of pilRNAstargeting SINE was extremely low. By contrast, SINEs are themost numerous repetitive genomic sequences and are still activein the mouse genome, and the fraction of SINE-derived smallRNAs has been shown to increase during mouse fetal deve-lopment.26 Our results imply that the TE-derived pilRNAspreferentially control the most abundant and active transposableelements in the genome.
Differences in piRNA biogenesis between mammals andnon-mammalian vertebrates. piRNAs are found in germ cells andgonads of diverse animals, including mammals, frogs, zebrafishand fruit flies. In this study, abundant TE-derived pilRNAswere unexpectedly cloned from chicken blastodermal cells. Thisfinding allowed us to compare the differences in sequence char-acteristics and biogenesis among vertebrate piRNAs. Despite thedifferences in sequence content, piRNAs/pilRNAs of chickensand other vertebrates (mice, frogs, and zebrafish) share manytypical features, including length distribution, nucleotide bias,genomic derivation and 3' modification. Based on several lines ofevidence, two mechanisms for piRNA biogenesis have beenproposed: primary piRNA processing in the ovarian somaticcell (OSC) line,72,73 and the ping-pong amplification cycle ingerm cells, with sense and antisense piRNAs with overlapping
www.landesbioscience.com RNA Biology 11

© 2012 Landes Bioscience.
Do not distribute.
5' ends separated by precisely 10 nt.52,74 The ping-pongamplification cycle has been detected in fetal mice, zebrafish,Xenopus,12,26,45,49,50 indicating a conserved mechanism of piRNAbiogenesis in vertebrates. Although the strong signature of ping-pong amplification found in the chicken further supports thismodel, appreciable differences between the mammalian andnon-mammalian vertebrate systems can be detected when thecharacteristics of primary (1U but no 10A) and secondary (10Abut no 1U) piRNAs are analyzed (Fig. 8). In mammals, the sense-oriented TE-piRNAs are the primary species, whereas antisense-oriented species are secondary; however, in chickens, theantisense-oriented TE-pilRNAs belong to the primary species,and the sense-oriented pilRNAs are secondary. We further got thesimilar results in the other non-mammalian vertebrates, includingthe frog and zebrafish (data not shown). The mechanism drivingthe differential processing of primary and secondary piRNAsbetween mammals and non-mammalian vertebrates may berelated to differences in the content of TEs in the genome, asmammalian genomes contain significantly high TE content, butnon-mammalian vertebrate genomes contain far fewer mobileelements. As revealed by the size distribution of pilRNAs inFigure 5A, the distinct sizes of the sense and antisense readsindicate that different proteins are involved in chicken pilRNAbiogenesis; however, only one PIWI protein (Piwi) gene has beendetected in the current chicken genome so far. The exactmechanism by which the ping-pong amplification cycle occurs inthe chicken remains unknown. Notably, Piwi-associated piRNAbiogenesis can also occur in a ping-pong-independent pathway(primary processing) in soma. Nearly half of the chicken pilRNAsidentified in our study showed few ping-pong signatures. Whetherthe primary processing pathway exists in undifferentiated cellssuch as PGCs remains an open question, and further studies arenecessary to completely delineate the biogenesis of piRNAs.
Methods
Library construction and sequencing. Fertilized chicken eggswere obtained from Guangdong Academy of AgriculturalSciences, China. Chicken embryos were collected at develop-mental stage X and stage HH10.30 After removal of the amnion,embryos were rinsed in DEPC-treated 0.75% NaCl solution andimmediately processed for RNA isolation using TRIzol reagent(Invitrogen) according to the manufacturer’s instructions. TheRNA concentration and purity were determined photometricallyby measuring absorbance at 260 nm and A260/A280 ratio usinga NanoDrop ND-1000 spectrophotometer (Nanodrop Techno-logies). Small RNAs ranging from 18 to 30 nt were gel purifiedand ligated to 5' and 3' adaptor oligonucleotides. Ligationproducts were gel purified, reverse transcribed, and amplifiedusing corresponding primers. Sequences of the adapters andprimers were as published (Illumina). Samples were sequencedusing an Illumina 1G Genome Analyzer by Beijing GenomicsInstitute (BGI) in Shenzhen, China.
Bioinformatic analysis of sequencing data. To simplify thesequencing data, all identical sequence reads in each small RNAlibrary were grouped and converted into sequence tags, with thenumber of reads for each sequence and tag reflecting relativeabundance and diversity, respectively. Only reads perfectlymatching the UCSC galGal3 chicken genome assembly75 wereused for further analysis. Sequences were annotated based on theiroverlap with publicly available genome annotations downloadedfrom NCBI (www.ncbi.nih.gov/Genbank/index.html, rRNA),76
Ensembl (www.ensembl.org/index.html, miscRNAs),77 snoRNA-LBMEdb (wwwsnorna.biotoul.fr/index.php, snoRNA),78 UCSC(http://genome.ucsc.edu, miscRNAs and transposons/repeats),75
Repbase (www.girinst.org/repbase/index.html, transposons/repeats),79 miRBase (microrna.sanger.ac.uk/sequences/index.shtml,
Figure 8. Comparison of the schematic ping-pong pair in mammals and non-mammalian vertebrates. The 10-nt overlap between the 59 ends of senseand antisense piRNAs are generated by the ping-pong mechanism. In mammals, the sense-oriented piRNAs are primary species that have a bias foruridine at position 1 and do not have nucleotide bias at position 10, and antisense-oriented piRNAs are secondary species that have a bias for adenine atposition 10 and do not have a bias at position 1. In the non-mammalian vertebrates, however, the primary and secondary piRNAs/pilRNAs have oppositenucleotide biases. N, any nucleotide base.
12 RNA Biology Volume 9 Issue 2

© 2012 Landes Bioscience.
Do not distribute.
release 16, miRNA).80 The annotation order was miRNA,mitochondrial RNA, rRNA, snoRNA, scaRNA, tRNA, snRNA,RNaseP, SRP RNA, Xist RNA, 7SK RNA, tel RNA, H19RNA, Y RNA, RNaseMRP, mRNA. The read count of eachunique sequence was normalized to reads per million (RPM)based on the total read count of the mapped data.
miRNA gene prediction. New miRNAs were predicted usingthe recently described algorithm miRDeep.40 Prediction wasperformed according to the miRDeep user manual. The data setused to predict miRNA comprised pooled reads from the smallRNA libraries of the stage X, HH10, and day five, seven andnine embryos.35 Only new miRNA candidates with total non-normalized read counts greater than 10 in at least one library orwith at least one read in at least two libraries were furtheranalyzed. We discarded the following miRNA candidates: (1) themature forms of candidates that were present at more than fivegenomic loci or that overlapped with known RNAs, (2) themature forms of candidates with lengths shorter than 19 nt orlonger than 25 nt, and (3) candidates with GC contents higherthan 85%. RNA secondary structures were predicted usingRNAfold, Vienna RNA Package.81
miRNA expression analysis. Deep sequencing data from thisstudy (stage X and HH10 embryos) and previous studies (five,seven, and nine day old chicken embryos)35 were used for expres-sion analysis. miRNA expression patterns were analyzed follow-ing a modified version of the protocol described by Ruby et al.82
For each library, the total number of miRNA hairpin-matchingreads was calculated as a normalization factor. The copy numberof each miRNA within each library was normalized to the totalnumber of miRNA hairpin-matching reads for that library.Relative expression analysis was used to determine the expressionbiases of individual miRNAs at different developmental stages.The normalized count of a particular miRNA in a particularlibrary was divided by the sum of normalized counts for thatmiRNA across all libraries. As a result, the value of the expressionprofile for an miRNA in a particular library ranged from 0 to 1.The application Cluster was used for hierarchical clusteringof miRNAs using average linkage correlation. Total expressionanalysis was performed to evaluate and compare the expressionlevels of individual miRNAs. The normalized counts of eachmiRNA across all libraries were summed and multiplied by 106.The resulting value corresponded to the number of reads for agiven miRNA per million reads matching miRNA hairpins.
Real-Time PCR and RT-PCR. Expression levels of knownand candidate mature miRNAs and pilRNAs were quantifiedby SYBR Green-based real-time RT-PCR using a SYBRPrimeScriptTM RT-PCR kit (TaKaRa). Small RNA-specificprimers were designed as described previously.83 Reverse trans-cription (RT) reactions were performed using 400 ng of totalRNA. Real-time PCR reactions were performed as follows: onecycle of 95°C for 30 sec followed by 40 cycles of 95°C for 5 secand 60°C for 20 sec. The expression levels of small RNAs rela-tive to U6 snRNA were determined using the 2 -ΔΔCт method.84
Error bars represent standard deviations. Comparative real-timePCR was performed in triplicate and included no-templatecontrols.
Analysis of miRNA targets, gene ontology (GO) terms andKEGG pathways. The conserved targets of known and newmiRNAs enriched in stage X and/or HH10 embryos werepredicted by TargetScan (www.targetscan.org/, Release 5.1).46
Only miRNAs enriched in CE-X, CE-HH10 or both stages (CE-X/HH10) were included in the target analysis. The ENSEMBLID for each target gene was obtained from the Ensembl databaseusing the BioMart data mining tool (www.ensembl.org/index.html). The significantly enriched biological process-related GO(Gene Ontology)47 terms in the lists of miRNA target geneswere identified using agriGO (bioinfo.cau.edu.cn/agriGO/index.php).85 Because our data set was moderate in size, a hypergeo-metric distribution analysis was applied, and the significancelevel was set at 0.05. The miRNA targets were also subjected toKEGG pathway annotation using the DAVID gene annotationtool (david.abcc.ncifcrf.gov/).86
Dual Luciferase reporter assay. The dual-luciferase reporterswere derived from the psiCHECK-2 vector (Promega). The3' untranslated region (UTR) fragments of chicken TGFBR1containing a wild-type (wt) miR-c236 binding site (TGFBR1–3'UTR-wt) or a mutated site complementary to the seed regionof miR-c236 (TGFBR1–3'UTR-mut) were synthesized andcloned into the 3' UTR of the Renilla luciferase (hRluc) gene.HEK 293T cells were plated in 48-well plates (4 × 104 in eachwell) and then cotransfected with 100 ng of reporter plasmid(psiCHECK-3'UTR) and 20 nM of the synthetic miR-c236mimic or negative control (GenePharma), using Lipofectamine2000 (Invitrogen) according to the manufacturer's protocol.Luciferase activities were measured 30 h post-transfection usingthe Dual-Luciferase® Reporter Assay System (Promega) onGloMaxTM 96 Microplate Luminometer (Promega). The Renillaluciferase activity was normalized to the Firefly luciferaseexpression (Rluc/Fluc) for each sample. Data shown are theMean ± SD from at least three independent experiments donein duplicate.
Detection of 2'-O-methyl modification at the 3' ends of smallRNAs. 2’-O-methylated nucleotides were detected using primerextension by reverse transcriptase at low dNTP concentrations asdescribed previously87 with some modification (Dong and Qu,manuscript in preparation). Briefly, the small RNA was ligatedto a 3' RNA oligonucleotide adaptor using T4 RNA ligase(TaKaRa). The ligation product was reverse-transcribed (RT)into cDNA with or without anchored RT primers that weredesigned to anchor the modified nucleotide at low (0.4 μM) orhigh (40 μM) dNTP concentrations. The cDNA was amplifiedby PCR with specific primers under the same reaction condi-tions. Equivalent loading of PCR products were separated on2.5% agarose gels and detected by GelRed dye (Biotium) stainingwith visualization by UV trans-illumination. PCR signal inten-sities were analyzed using Bio-Rad Quantity One software.
ISH of whole-mount chicken embryos. Digoxigenin end-labeled (5') LNA-modified DNA oligonucleotide probes com-plementary to the pilRNAs were purchased from Exiqon.Digoxigenin-labeled ddUTP was added to the 3' end of theLNAs using a DIG Oligonucleotide 3'-End Labeling Kit, 2ndGeneration (Roche). RT-PCR products of the chicken vasa
www.landesbioscience.com RNA Biology 13

© 2012 Landes Bioscience.
Do not distribute.
homolog (Cvh) transcript (400 bp), deleted in azoospermia-like(Dazl) transcript (355 bp), and ENS-1 transcript (451 bp) wereobtained using specific primers. To detect mRNA, antisense andsense cRNA probes were synthesized using a PCR-amplifiedtemplate containing only the partial gene sequence and RNApolymerase promoter sequence. Digoxigenin-labeled antisenseand sense cRNA probes were prepared with T7 RNA polymeraseby using a DIG RNA Labeling Mix (Roche) according to themanufacturer’s instructions.
ISH of whole-mount chicken embryos for small RNAs wasperformed as previously described.88 In brief, chicken embryoswere fixed in 4% paraformaldehyde in PBS overnight at 4°C.The samples were washed with PBS containing 1% Tween 20(PBT). After the dehydration and rehydration steps, the preparedembryos were transferred to a prehybridization solution (50%formamide, 5 × SSC, 2% blocking powder, 0.1% Tween-20,0.1% CHAPS, 50 μg/ml yeast RNA, 5 mM EDTA, 50 μg/mlheparin) at 62°C. The LNA probe (2.5 pmol) was added to 1 mlfresh prehybridization buffer, and hybridization was performedat 62°C for 48 h. Embryos were washed 3 × 20 min with a highsalt (2 × SSC, 0.1% CHAPS) wash and 3 × 20 min with 0.2 ×SSC, 0.1% CHAPS. Embryos were then rinsed twice in KTBT(50 mM Tris, pH 7.5, 150 mM NaCl, 10 mM KCl, 1% Tween-20). Embryos were pretreated with 20% sheep serum in KTBTat 4°C for 3 h and incubated with anti-DIG antibody (1:5000)overnight at 4°C. Embryos were then washed with KTBT at 4°Covernight and transferred into fresh NTMT (100 mM NaCl,100 mM Tris pH 9.5, 50 mM MgCl2, 0.1% Tween-20). Colorreactions (NBT/BCIP in NTMT) were performed for 1–6 hat room temperature until a signal or the background became
visible. Embryos were photographed using a Zeiss SteREO(Lumar V12) stereomicroscope. For the most part, ISH formRNAs was conducted using the protocol described for detect-ing pilRNAs, except that Tween-20 was replaced in all solutionswith the same concentration of Triton X-100. In addition,30 ng of antisense DIG-labeled cRNA probes were added to thehybridization solution.
Oligonucleotides. Oligonucleotides used in this study are listedin Table S11.
Accession numbers. The short read sequence data from thisstudy have been submitted to NCBI Gene Expression Omnibus(GEO) (www.ncbi.nlm.nih.gov/geo/) under accession numberGSE28668.
Disclosure of Potential Conflicts of Interest
No potential conflicts of interest were disclosed.
Acknowledgments
We would like to thank Wenqing Li, Hui Xu, Yin Zhang for theirtechnical assistance. This study was supported by the NationalBasic Research Program of China (No. 2011CB811300 to L.H.Q.), the National Natural Science Foundation of China (No.31000571 to P.S. and No. 30830066 to L.H.Q.), the ChinaPostdoctoral Science Foundation (No. 20100480816 to J.Y.L.)and the Guangdong Provincial Natural Science Foundation ofChina (No. 10451027501005659 to P.S.).
Supplemental Materials
Supplemental materials can be found at:www.landesbioscience.com/journals/rnabiology/article/18489
References1. Ghildiyal M, Zamore PD. Small silencing RNAs: an
expanding universe. Nat Rev Genet 2009; 10:94-108;PMID:19148191; http://dx.doi.org/10.1038/nrg2504
2. Carthew RW, Sontheimer EJ. Origins and Mechan-isms of miRNAs and siRNAs. Cell 2009; 136:642-55; PMID:19239886; http://dx.doi.org/10.1016/j.cell.2009.01.035
3. Bartel DP. MicroRNAs: genomics, biogenesis, mechan-ism, and function. Cell 2004; 116:281-97; PMID:14744438; http://dx.doi.org/10.1016/S0092-8674(04)00045-5
4. Okamura K, Lai EC. Endogenous small interferingRNAs in animals. Nat Rev Mol Cell Biol 2008; 9:673-8;PMID:18719707; http://dx.doi.org/10.1038/nrm2479
5. Watanabe T, Takeda A, Tsukiyama T, Mise K, OkunoT, Sasaki H, et al. Identification and characterization oftwo novel classes of small RNAs in the mouse germline:retrotransposon-derived siRNAs in oocytes and germ-line small RNAs in testes. Genes Dev 2006; 20:1732-43; PMID:16766679; http://dx.doi.org/10.1101/gad.1425706
6. Tam OH, Aravin AA, Stein P, Girard A, MurchisonEP, Cheloufi S, et al. Pseudogene-derived small inter-fering RNAs regulate gene expression in mouse oocytes.Nature 2008; 453:534-8; PMID:18404147; http://dx.doi.org/10.1038/nature06904
7. Watanabe T, Totoki Y, Toyoda A, Kaneda M,Kuramochi-Miyagawa S, Obata Y, et al. EndogenoussiRNAs from naturally formed dsRNAs regulatetranscripts in mouse oocytes. Nature 2008; 453:539-43; PMID:18404146; http://dx.doi.org/10.1038/nature06908
8. Babiarz JE, Ruby JG, Wang Y, Bartel DP, Blelloch R.Mouse ES cells express endogenous shRNAs, siRNAs,and other Microprocessor-independent, Dicer-dependentsmall RNAs. Genes Dev 2008; 22:2773-85; PMID:18923076; http://dx.doi.org/10.1101/gad.1705308
9. Kim VN, Han J, Siomi MC. Biogenesis of small RNAsin animals. Nat Rev Mol Cell Biol 2009; 10:126-39;PMID:19165215; http://dx.doi.org/10.1038/nrm2632
10. Thomson T, Lin H. The biogenesis and function ofPIWI proteins and piRNAs: progress and prospect.Annu Rev Cell Dev Biol 2009; 25:355-76; PMID:19575643; http://dx.doi.org/10.1146/annurev.cellbio.24.110707.175327
11. Horwich MD, Li C, Matranga C, Vagin V, Farley G,Wang P, et al. The Drosophila RNA methyltransferase,DmHen1, modifies germline piRNAs and single-stranded siRNAs in RISC. Curr Biol 2007; 17:1265-72; PMID:17604629; http://dx.doi.org/10.1016/j.cub.2007.06.030
12. Houwing S, Kamminga LM, Berezikov E, CronemboldD, Girard A, van den Elst H, et al. A role for Piwi andpiRNAs in germ cell maintenance and transposonsilencing in Zebrafish. Cell 2007; 129:69-82; PMID:17418787; http://dx.doi.org/10.1016/j.cell.2007.03.026
13. Kirino Y, Mourelatos Z. Mouse Piwi-interacting RNAsare 2'-O-methylated at their 3' termini. Nat Struct MolBiol 2007; 14:347-8; PMID:17384647; http://dx.doi.org/10.1038/nsmb1218
14. Ohara T, Sakaguchi Y, Suzuki T, Ueda H, Miyauchi K,Suzuki T. The 3' termini of mouse Piwi-interactingRNAs are 2'-O-methylated. Nat Struct Mol Biol 2007;14:349-50; PMID:17384646; http://dx.doi.org/10.1038/nsmb1220
15. Yang Z, Ebright YW, Yu B, Chen X. HEN1 recognizes21-24 nt small RNA duplexes and deposits a methylgroup onto the 2' OH of the 3' terminal nucleotide.Nucleic Acids Res 2006; 34:667-75; PMID:16449203;http://dx.doi.org/10.1093/nar/gkj474
16. Stefani G, Slack FJ. Small non-coding RNAs in animaldevelopment. Nat Rev Mol Cell Biol 2008; 9:219-30;PMID:18270516; http://dx.doi.org/10.1038/nrm2347
17. Wang Y, Medvid R, Melton C, Jaenisch R, Blelloch R.DGCR8 is essential for microRNA biogenesis andsilencing of embryonic stem cell self-renewal. NatGenet 2007; 39:380-5; PMID:17259983; http://dx.doi.org/10.1038/ng1969
18. Kanellopoulou C, Muljo SA, Kung AL, Ganesan S,Drapkin R, Jenuwein T, et al. Dicer-deficient mouseembryonic stem cells are defective in differentiationand centromeric silencing. Genes Dev 2005; 19:489-501; PMID:15713842; http://dx.doi.org/10.1101/gad.1248505
19. Murchison EP, Partridge JF, Tam OH, Cheloufi S,Hannon GJ. Characterization of Dicer-deficient murineembryonic stem cells. Proc Natl Acad Sci USA 2005;102:12135-40; PMID:16099834; http://dx.doi.org/10.1073/pnas.0505479102
20. Tay Y, Zhang J, Thomson AM, Lim B, Rigoutsos I.MicroRNAs to Nanog, Oct4 and Sox2 coding regionsmodulate embryonic stem cell differentiation. Nature2008; 455:1124-8; PMID:18806776; http://dx.doi.org/10.1038/nature07299
14 RNA Biology Volume 9 Issue 2

© 2012 Landes Bioscience.
Do not distribute.
21. Xu N, Papagiannakopoulos T, Pan G, Thomson JA,Kosik KS. MicroRNA-145 regulates OCT4, SOX2,and KLF4 and represses pluripotency in human em-bryonic stem cells. Cell 2009; 137:647-58; PMID:19409607; http://dx.doi.org/10.1016/j.cell.2009.02.038
22. Sijen T, Plasterk RH. Transposon silencing in theCaenorhabditis elegans germ line by natural RNAi.Nature 2003; 426:310-4; PMID:14628056; http://dx.doi.org/10.1038/nature02107
23. Yang N, Kazazian HH, Jr. L1 retrotransposition issuppressed by endogenously encoded small interferingRNAs in human cultured cells. Nat Struct Mol Biol2006; 13:763-71; PMID:16936727; http://dx.doi.org/10.1038/nsmb1141
24. Chung WJ, Okamura K, Martin R, Lai EC. Endo-genous RNA interference provides a somatic defenseagainst Drosophila transposons. Curr Biol 2008;18:795-802; PMID:18501606; http://dx.doi.org/10.1016/j.cub.2008.05.006
25. Aravin AA, Hannon GJ, Brennecke J. The Piwi-piRNApathway provides an adaptive defense in the transposonarms race. Science 2007; 318:761-4; PMID:17975059;http://dx.doi.org/10.1126/science.1146484
26. Aravin AA, Sachidanandam R, Bourc'his D, Schaefer C,Pezic D, Toth KF, et al. A piRNA pathway primed byindividual transposons is linked to de novo DNAmethylation in mice. Mol Cell 2008; 31:785-99;PMID:18922463; http://dx.doi.org/10.1016/j.molcel.2008.09.003
27. Kuramochi-Miyagawa S, Watanabe T, Gotoh K,Totoki Y, Toyoda A, Ikawa M, et al. DNA methylationof retrotransposon genes is regulated by Piwi familymembers MILI and MIWI2 in murine fetal testes.Genes Dev 2008; 22:908-17; PMID:18381894; http://dx.doi.org/10.1101/gad.1640708
28. Eyal-Giladi H, Kochav S. From cleavage to primitivestreak formation: a complementary normal table and anew look at the first stages of the development of thechick. I. General morphology. Dev Biol 1976; 49:321-37; PMID:944662; http://dx.doi.org/10.1016/0012-1606(76)90178-0
29. Pain B, Clark ME, Shen M, Nakazawa H, Sakurai M,Samarut J, et al. Long-term in vitro culture andcharacterisation of avian embryonic stem cells withmultiple morphogenetic potentialities. Development1996; 122:2339-48; PMID:8756279
30. Hamburger V, Hamilton HL. A series of normal stagesin the development of the chick embryo. 1951. DevDyn 1992; 195:231-72; PMID:1304821; http://dx.doi.org/10.1002/aja.1001950404
31. Heisenberg CP, Solnica-Krezel L. Back and forthbetween cell fate specification and movement duringvertebrate gastrulation. Curr Opin Genet Dev 2008;18:311-6; PMID:18721878; http://dx.doi.org/10.1016/j.gde.2008.07.011
32. Inui M, Martello G, Piccolo S. MicroRNA control ofsignal transduction. Nat Rev Mol Cell Biol 2010;11:252-63; PMID:20216554; http://dx.doi.org/10.1038/nrm2868
33. Xu H, Wang X, Du Z, Li N. Identification ofmicroRNAs from different tissues of chicken embryoand adult chicken. FEBS Lett 2006; 580:3610-6;PMID:16750530; http://dx.doi.org/10.1016/j.febslet.2006.05.044
34. Burnside J, Ouyang M, Anderson A, Bernberg E, Lu C,Meyers BC, et al. Deep sequencing of chicken micro-RNAs. BMCGenomics 2008; 9:185; PMID:18430245;http://dx.doi.org/10.1186/1471-2164-9-185
35. Glazov EA, Cottee PA, Barris WC, Moore RJ,Dalrymple BP, Tizard ML. A microRNA catalog ofthe developing chicken embryo identified by a deepsequencing approach. Genome Res 2008; 18:957-64;PMID:18469162; http://dx.doi.org/10.1101/gr.074740.107
36. Shao P, Zhou H, Xiao ZD, He JH, Huang MB, ChenYQ, et al. Identification of novel chicken microRNAsand analysis of their genomic organization. Gene 2008;418:34-40; PMID:18511220; http://dx.doi.org/10.1016/j.gene.2008.04.004
37. Bannister SC, Tizard ML, Doran TJ, Sinclair AH,Smith CA. Sexually dimorphic microRNA expressionduring chicken embryonic gonadal development. BiolReprod 2009; 81:165-76; PMID:19357368; http://dx.doi.org/10.1095/biolreprod.108.074005
38. Hicks JA, Tembhurne PA, Liu HC. Identification ofmicroRNA in the developing chick immune organs.Immunogenetics 2009; 61:231-40; PMID:19198828;http://dx.doi.org/10.1007/s00251-009-0355-1
39. Rathjen T, Pais H, Sweetman D, Moulton V,Munsterberg A, Dalmay T. High throughput sequen-cing of microRNAs in chicken somites. FEBS Lett2009; 583:1422-6; PMID:19328789; http://dx.doi.org/10.1016/j.febslet.2009.03.048
40. Friedländer MR, Chen W, Adamidi C, Maaskola J,Einspanier R, Knespel S, et al. Discovering microRNAsfrom deep sequencing data using miRDeep. Nat Bio-technol 2008; 26:407-15; PMID:18392026; http://dx.doi.org/10.1038/nbt1394
41. Morin RD, O'Connor MD, GriffithM, Kuchenbauer F,Delaney A, Prabhu AL, et al. Application of massivelyparallel sequencing to microRNA profiling and dis-covery in human embryonic stem cells. GenomeRes 2008; 18:610-21; PMID:18285502; http://dx.doi.org/10.1101/gr.7179508
42. Houbaviy HB, Murray MF, Sharp PA. Embryonicstem cell-specific MicroRNAs. Dev Cell 2003; 5:351-8;PMID:12919684; http://dx.doi.org/10.1016/S1534-5807(03)00227-2
43. Suh MR, Lee Y, Kim JY, Kim SK, Moon SH, Lee JY,et al. Human embryonic stem cells express a uniqueset of microRNAs. Dev Biol 2004; 270:488-98; PMID:15183728; http://dx.doi.org/10.1016/j.ydbio.2004.02.019
44. Rosa A, Spagnoli FM, Brivanlou AH. The miR-430/427/302 family controls mesendodermal fate specifica-tion via species-specific target selection. Dev Cell 2009;16:517-27; PMID:19386261; http://dx.doi.org/10.1016/j.devcel.2009.02.007
45. Armisen J, Gilchrist MJ, Wilczynska A, Standart N,Miska EA. Abundant and dynamically expressedmiRNAs, piRNAs, and other small RNAs in thevertebrate Xenopus tropicalis. Genome Res 2009;19:1766-75; PMID:19628731; http://dx.doi.org/10.1101/gr.093054.109
46. Lewis BP, Burge CB, Bartel DP. Conserved seedpairing, often flanked by adenosines, indicates thatthousands of human genes are microRNA targets. Cell2005; 120:15-20; PMID:15652477; http://dx.doi.org/10.1016/j.cell.2004.12.035
47. Ashburner M, Ball CA, Blake JA, Botstein D, Butler H,Cherry JM, et al. Gene ontology: tool for the unifica-tion of biology. The Gene Ontology Consortium. NatGenet 2000; 25:25-9; PMID:10802651; http://dx.doi.org/10.1038/75556
48. Aravin AA, Sachidanandam R, Girard A, Fejes-Toth K,Hannon GJ. Developmentally regulated piRNA clus-ters implicate MILI in transposon control. Science2007; 316:744-7; PMID:17446352; http://dx.doi.org/10.1126/science.1142612
49. Houwing S, Berezikov E, Ketting RF. Zili is requiredfor germ cell differentiation and meiosis in zebrafish.EMBO J 2008; 27:2702-11; PMID:18833190; http://dx.doi.org/10.1038/emboj.2008.204
50. Lau NC, Ohsumi T, Borowsky M, Kingston RE,Blower MD. Systematic and single cell analysis ofXenopus Piwi-interacting RNAs and Xiwi. EMBO J2009; 28:2945-58; PMID:19713941; http://dx.doi.org/10.1038/emboj.2009.237
51. Vagin VV, Sigova A, Li C, Seitz H, Gvozdev V,Zamore PD. A distinct small RNA pathway silencesselfish genetic elements in the germline. Science2006; 313:320-4; PMID:16809489; http://dx.doi.org/10.1126/science.1129333
52. Brennecke J, Aravin AA, Stark A, Dus M, Kellis M,Sachidanandam R, et al. Discrete small RNA-generatingloci as master regulators of transposon activity in Droso-phila. Cell 2007; 128:1089-103; PMID:17346786;http://dx.doi.org/10.1016/j.cell.2007.01.043
53. Ginsburg M, Eyal-Giladi H. Primordial germ cells ofthe young chick blastoderm originate from the centralzone of the area pellucida irrespective of the embryo-forming process. Development 1987; 101:209-19;PMID:3446474
54. Lavial F, Acloque H, Bachelard E, Nieto MA, SamarutJ, Pain B. Ectopic expression of Cvh (Chicken Vasahomologue) mediates the reprogramming of chickenembryonic stem cells to a germ cell fate. Dev Biol 2009;330:73-82; PMID:19324033; http://dx.doi.org/10.1016/j.ydbio.2009.03.012
55. Acloque H, Risson V, Birot AM, Kunita R, Pain B,Samarut J. Identification of a new gene family speci-fically expressed in chicken embryonic stem cells andearly embryo. Mech Dev 2001; 103:79-91; PMID:11335114; http://dx.doi.org/10.1016/S0925-4773(01)00336-7
56. Ohnishi Y, Totoki Y, Toyoda A, Watanabe T,Yamamoto Y, Tokunaga K, et al. Small RNA classtransition from siRNA/piRNA to miRNA during pre-implantation mouse development. Nucleic Acids Res2010; 38:5141-51; PMID:20385573; http://dx.doi.org/10.1093/nar/gkq229
57. Martinez NJ, Gregory RI. MicroRNA gene regulatorypathways in the establishment and maintenance ofESC identity. Cell Stem Cell 2010; 7:31-5; PMID:20621047; http://dx.doi.org/10.1016/j.stem.2010.06.011
58. Wang Y, Baskerville S, Shenoy A, Babiarz JE, BaehnerL, Blelloch R. Embryonic stem cell-specific microRNAsregulate the G1-S transition and promote rapid pro-liferation. Nat Genet 2008; 40:1478-83; PMID:18978791; http://dx.doi.org/10.1038/ng.250
59. Varambally S, Cao Q, Mani RS, Shankar S, Wang X,Ateeq B, et al. Genomic loss of microRNA-101 leads tooverexpression of histone methyltransferase EZH2 incancer. Science 2008; 322:1695-9; PMID:19008416;http://dx.doi.org/10.1126/science.1165395
60. O'Carroll D, Erhardt S, Pagani M, Barton SC, SuraniMA, Jenuwein T. The polycomb-group gene Ezh2 isrequired for early mouse development. Mol Cell Biol2001; 21:4330-6; PMID:11390661; http://dx.doi.org/10.1128/MCB.21.13.4330-4336.2001
61. Wilson SI, Rydstrom A, Trimborn T, Willert K, NusseR, Jessell TM, et al. The status of Wnt signallingregulates neural and epidermal fates in the chickembryo. Nature 2001; 411:325-30; PMID:11357137;http://dx.doi.org/10.1038/35077115
62. Marvin MJ, Di Rocco G, Gardiner A, Bush SM, LassarAB. Inhibition of Wnt activity induces heart formationfrom posterior mesoderm. Genes Dev 2001; 15:316-27; PMID:11159912; http://dx.doi.org/10.1101/gad.855501
63. Wilson SI, Graziano E, Harland R, Jessell TM, EdlundT. An early requirement for FGF signalling in theacquisition of neural cell fate in the chick embryo. CurrBiol 2000; 10:421-9; PMID:10801412; http://dx.doi.org/10.1016/S0960-9822(00)00431-0
64. Joglekar MV, Patil D, Joglekar VM, Rao GV, ReddyDN, Mitnala S, et al. The miR-30 family microRNAsconfer epithelial phenotype to human pancreatic cells.Islets 2009; 1:137-47; PMID:21099261; http://dx.doi.org/10.4161/isl.1.2.9578
www.landesbioscience.com RNA Biology 15

© 2012 Landes Bioscience.
Do not distribute.
65. Yao J, Liang L, Huang S, Ding J, Tan N, Zhao Y, et al.MicroRNA-30d promotes tumor invasion and meta-stasis by targeting Galphai2 in hepatocellular carcin-oma. Hepatology 2010; 51:846-56; PMID:20054866
66. Ivey KN, Muth A, Arnold J, King FW, Yeh RF, FishJE, et al. MicroRNA regulation of cell lineages inmouse and human embryonic stem cells. Cell StemCell 2008; 2:219-29; PMID:18371447; http://dx.doi.org/10.1016/j.stem.2008.01.016
67. Aramaki S, Sato F, Kato T, Soh T, Kato Y, HattoriMA. Molecular cloning and expression of dead endhomologue in chicken primordial germ cells. CellTissue Res 2007; 330:45-52; PMID:17610087; http://dx.doi.org/10.1007/s00441-007-0435-1
68. Friedländer MR, Adamidi C, Han T, Lebedeva S,Isenbarger TA, Hirst M, et al. High-resolution profilingand discovery of planarian small RNAs. Proc Natl AcadSci USA 2009; 106:11546-51; PMID:19564616;http://dx.doi.org/10.1073/pnas.0905222106
69. Lee EJ, Banerjee S, Zhou H, Jammalamadaka A, ArcilaM, Manjunath BS, et al. Identification of piRNAs inthe central nervous system. Rna 2011. In Press. PMID:21515829; http://dx.doi.org/10.1261/rna.2565011
70. Brennecke J, Malone CD, Aravin AA, SachidanandamR, Stark A, Hannon GJ. An epigenetic role for mater-nally inherited piRNAs in transposon silencing. Science2008; 322:1387-92; PMID:19039138; http://dx.doi.org/10.1126/science.1165171
71. Consortium ICGS. Sequence and comparative ana-lysis of the chicken genome provide unique perspec-tives on vertebrate evolution. Nature 2004; 432:695-716; PMID:15592404; http://dx.doi.org/10.1038/nature03154
72. Li C, Vagin VV, Lee S, Xu J, Ma S, Xi H, et al.Collapse of germline piRNAs in the absence ofArgonaute3 reveals somatic piRNAs in flies. Cell2009; 137:509-21; PMID:19395009; http://dx.doi.org/10.1016/j.cell.2009.04.027
73. Saito K, Inagaki S, Mituyama T, Kawamura Y, Ono Y,Sakota E, et al. A regulatory circuit for piwi by the largeMaf gene traffic jam in Drosophila. Nature 2009;461:1296-9; PMID:19812547; http://dx.doi.org/10.1038/nature08501
74. Gunawardane LS, Saito K, Nishida KM, Miyoshi K,Kawamura Y, Nagami T, et al. A slicer-mediatedmechanism for repeat-associated siRNA 5' end forma-tion in Drosophila. Science 2007; 315:1587-90;PMID:17322028; http://dx.doi.org/10.1126/science.1140494
75. Kuhn RM, Karolchik D, Zweig AS, Wang T, SmithKE, Rosenbloom KR, et al. The UCSC GenomeBrowser Database: update 2009. Nucleic Acids Res2009; 37:D755-61; PMID:18996895; http://dx.doi.org/10.1093/nar/gkn875
76. Maglott D, Ostell J, Pruitt KD, Tatusova T. EntrezGene: gene-centered information at NCBI. NucleicAcids Res 2011; 39:D52-7; PMID:21115458; http://dx.doi.org/10.1093/nar/gkq1237
77. Hubbard TJ, Aken BL, Ayling S, Ballester B, Beal K,Bragin E, et al. Ensembl 2009. Nucleic Acids Res 2009;37:D690-7; PMID:19033362; http://dx.doi.org/10.1093/nar/gkn828
78. Lestrade L, Weber MJ. snoRNA-LBME-db, a compre-hensive database of human H/ACA and C/D boxsnoRNAs. Nucleic Acids Res 2006; 34:D158-62;PMID:16381836; http://dx.doi.org/10.1093/nar/gkj002
79. Jurka J, Kapitonov VV, Pavlicek A, Klonowski P,Kohany O, Walichiewicz J. Repbase Update, a databaseof eukaryotic repetitive elements. Cytogenet GenomeRes 2005; 110:462-7; PMID:16093699; http://dx.doi.org/10.1159/000084979
80. Griffiths-Jones S, Saini HK, van Dongen S, Enright AJ.miRBase: tools for microRNA genomics. Nucleic AcidsRes 2008; 36:D154-8; PMID:17991681; http://dx.doi.org/10.1093/nar/gkm952
81. Hofacker IL. RNA secondary structure analysis usingthe Vienna RNA package. Curr Protoc Bioinformatics2004; Chapter 12:Unit 12 2; PMID: 18428716.
82. Ruby JG, Stark A, Johnston WK, Kellis M, Bartel DP,Lai EC. Evolution, biogenesis, expression, and targetpredictions of a substantially expanded set of Droso-phila microRNAs. Genome Res 2007; 17:1850-64;PMID:17989254; http://dx.doi.org/10.1101/gr.6597907
83. Chen C, Ridzon DA, Broomer AJ, Zhou Z, Lee DH,Nguyen JT, et al. Real-time quantification of micro-RNAs by stem-loop RT-PCR. Nucleic Acids Res 2005;33:e179; PMID:16314309; http://dx.doi.org/10.1093/nar/gni178
84. Livak KJ, Schmittgen TD. Analysis of relative geneexpression data using real-time quantitative PCR andthe 2(-Delta Delta C(T)). Methods 2001; 25:402-8; PMID:11846609; http://dx.doi.org/10.1006/meth.2001.1262
85. Du Z, Zhou X, Ling Y, Zhang Z, Su Z. agriGO: a GOanalysis toolkit for the agricultural community. NucleicAcids Res 2010; 38(Suppl):W64-70; PMID:20435677;http://dx.doi.org/10.1093/nar/gkq310
86. Huang W, Sherman BT, Lempicki RA. Systematicand integrative analysis of large gene lists usingDAVID bioinformatics resources. Nat Protoc 2009;4:44-57; PMID:19131956; http://dx.doi.org/10.1038/nprot.2008.211
87. Maden BE, Corbett ME, Heeney PA, Pugh K, AjuhPM. Classical and novel approaches to the detectionand localization of the numerous modified nucleotidesin eukaryotic ribosomal RNA. Biochimie 1995; 77:22-9; PMID:7599273; http://dx.doi.org/10.1016/0300-9084(96)88100-4
88. Darnell DK, Kaur S, Stanislaw S, Konieczka JH,Yatskievych TA, Antin PB. MicroRNA expressionduring chick embryo development. Dev Dyn 2006;235:3156-65; PMID:17013880; http://dx.doi.org/10.1002/dvdy.20956
16 RNA Biology Volume 9 Issue 2




















