
Antinociceptive effect of : a Bromeliaceae from the Brazilian coastal rain forest
Upload
independentCategory
view
1download
0
ORIGINAL ARTICLE
Leaf structure of Bromelia and its significance for the evolutionof Bromelioideae (Bromeliaceae)
Raquel Fernandes Monteiro • Rafaela Campostrini Forzza •
Andre Mantovani
Received: 5 April 2010 / Accepted: 28 January 2011 / Published online: 10 March 2011
� Springer-Verlag 2011
Abstract This study investigated the leaf structure of
Bromelia and its importance for understanding the evolution
of Bromelioideae (Bromeliaceae). Because of the scarcity of
informative molecular characters in Bromeliaceae, this
study evaluates the relevance of anatomical characters for
the taxonomy and phylogeny of Bromelia and the subfamily
Bromelioideae. Anatomical studies in monocots have shown
that the combined application of anatomical and external
morphological characters from leaf structure can improve
the taxonomic delimitation of species, genera and subgen-
era, and is very informative for inferring phylogenies. The
current study analyzed the leaves of 27 species of Bromelia
and found that the most important characters for the sys-
tematics of this group are the occurrence of a water storage
hypodermis, the number of stalk cells of peltate scales, the
presence of a ribbed abaxial surface, the occurrence of pal-
isade parenchyma on the adaxial side, the shape of the cells
that surround the air lacunae, the presence of raphides and
secretory channels, and the occurrence of fibrous extensions
on the bundle sheath on minor veins. Combining our results
with those described for the family, we made a list of the
anatomical characters that can be used in phylogenetic
studies of Bromelioideae.
Keywords Foliar anatomy � Neotropics � Monocots �Systematics � Taxonomy
Anatomical characters can be useful in the phylogenetic
analysis of Bromeliaceae, especially considering that
informative molecular characters are scarce. For instance,
only 6% of the characters obtained from plastid region
sequenced markers of 48 species in 24 genera of Bromeli-
aceae were found to be informative (Schulte et al. 2009; see
also Faria 2006; Sousa et al. 2007). Anatomical features are
considered potentially useful to support taxonomic cir-
cumscriptions within the monocots as a whole (e.g., Simon
2007; Henderson and Stevenson 2007; Simpson and Burton
2006; Furness and Rudall 2000; Stevenson et al. 2000;
Tomlinson and Fisher 2000) and particularly within Bro-
meliaceae (e.g., Tomlinson 1969; Braga 1977; Sousa and
Neves 1996; Sajo et al. 1998; Aoyoma and Sajo 2003;
Arruda & Costa 2003; Scatena and Segecin 2005; Sousa
et al. 2005; Horres et al. 2007; Proenca and Sajo 2007,
2008). Within the subfamily Bromelioideae, with unclear
generic delimitation (Schulte et al. 2009, Benzing 2000),
combined anatomical and morphological characters have
already proved useful to solve taxonomic problems (e.g.,
Ramirez 1996; Brown 2000; Coffani-Nunes 2004; Sousa
2004; Sousa et al. 2007; Almeida et al. 2009).
Bromelia, a basally diverging genus of Bromelioideae
(Givinish et al. 2007; Schulte et al. 2009), includes 56
species (Luther 2008) grouped into three subgenera (Mez
1891): Distiacanthus, Karatas and Bromelia. It occurs
from central Mexico to southern Argentina, with two
centers of diversity: one in Central America extending to
the southern Andes and the other in the Brazilian Shield,
mainly in Cerrado vegetation, where the majority of its
species are found, most of them in dry or more markedly
seasonal regions (Smith and Downs 1979). Species of
Bromelia can be recognized by their leaves with curved
spines along the margins, sheaths covered with fine linear
scales, fleshy petals united into a tube by the filaments, lack
of petal appendages and flattened naked seeds (Smith and
Downs 1979). The phylogenetic study with the genus
R. F. Monteiro (&) � R. C. Forzza � A. Mantovani
Jardim Botanico do Rio de Janeiro, Rua Pacheco Leao 915,
Rio de Janeiro, RJ 22460-030, Brazil
e-mail: [email protected]
123
Plant Syst Evol (2011) 293:53–64
DOI 10.1007/s00606-011-0426-2

(Monteiro 2009) showed the occurrence of only three
synapomorphies for the genus: the fuzzy indumentum on
the abaxial surface of the leaf sheath, the oblong ovary and
the sclerenchymatous hypodermis with four layers on the
abaxial surface. Micromorphological studies with Bromelia
provide general anatomical descriptions of a few species
(Tomlinson 1969; Schmidt and Brown 2004), without
evaluating their potential for phylogenetic systematics
(Horres et al. 2007).
Despite the importance of the anatomical features in the
systematics of Bromeliaceae, there is no information
available for many of its genera. In this study we examined
the leaves of 27 species of Bromelia species with the aim of
pointing out characters that can be useful on phylogenetic
analysis of the genus and the subfamily.
Materials and methods
Twenty-seven Bromelia species were studied in an attempt
to cover all the different habitats and the distribution of the
genus (Table 1). Expanded leaves were collected in the
field and preserved in ethanol 70%. Vouchers were
deposited at the Herbarium of the Jardim Botanico do Rio
de Janeiro (RB). In addition, dry leaves from exsiccatae
were analyzed after rehydration and subsequent treatment.
Both freehand sections and resin embedding were
prepared.
Cross sections of the middle third of the leaf blade were
made freehand. One or two cross sections were replicated
for each specimen. The freehand cuts were cleared in 50%
sodium hypochlorite, stained with Safranin and Astra blue
Table 1 Species of Bromelia analyzed with their respective collection sites and voucher specimens
Taxon Provenance Voucher specimen
Subgenus Distiacanthus
B. morreniana (Regel) Mez Brazil, Amazonas G.T.Prance 16484 (INPA)
B. scarlatina (Hort. ex Herincq) E. Morren Peru, Loreto T. Plowman 7032 (F)
Subgenus Karatas
B. auriculata L.B.Sm. Brazil, Bahia B.R.Silva 1452 (JBRJ hort)
B. grandiflora Mez Brazil, Goias E.M.C.Leme 2200 (RB)
B. karatas L. Costa Rica, Puntarenas E.Bello 2903 (F)
B. lagopus Mez Brazil, Goias R.F.Monteiro 131 (RB)
B. lindevaldae Leme & E. Esteves Brazil, Goias B.R.Silva 1223 (RB)
B. macedoi L.B.Sm. Brazil, Goias R.F.Monteiro 126 (RB)
B. minima Leme & E. Esteves Brazil, Goias B.R.Silva 1253 (RB)
B. aff. tubulosa L.B.Sm. Brazil, Espırito Santo B.R.Silva 1387 (JBRJ hort), R.F.Monteiro 280 (RB)
B. tubulosa L.B.Sm. Brazil, Para M.J.Pires 1661 (INPA)
B. villosa Mez Brazil, Goias B.R.Silva 1234 (RB)
Subgenus Bromelia
B. antiacantha Bertol. Brazil, Rio de Janeiro B.R.Silva s.n. (JBRJ hort), R.F.Monteiro 263 (RB)
B. arenaria Ule Brazil, Bahia R.C.Forzza 3834 (RB)
B. balansae Mez Brazil, without locality E.M.C.Leme 2869 (RB)
B. binotii E. Morren ex Mez Brazil, Espırito Santo R.F.Monteiro 265 (RB)
B. chrysantha Jacq. Venezuela, Lara A.Fernandez 1473 (F)
B. goyazensis Mez Brazil, Goias B.R.Silva 1458 (RB)
B. hemispherica Lam. Mexico, Chiapas R.M.Laughlin 859 (F)
B. hieronymii Mez Argentina, Santiago del Estero V.Marunak 395 (RB)
B. horstii Rauh Brazil, Mato Grosso do Sul C.Fleming s.n. (RB)
B. interior L.B.Sm. Brazil, Goias R.F.Monteiro 122 (RB)
B. irwinii L.B.Sm. Brazil, Goias H.S.Irwinii 33100 (F)
B. laciniosa Mart. ex Schult. & Schult. f. Brazil, Minas Gerais B.R.Silva 1409 (JBRJ hort)
B. pinguin L. United States, Saint Croix Roadside 342 (F)
B. reversacantha Mez Brazil, Goias R.F.Monteiro 130 (RB)
B. serra Grisb. Brazil, Minas Gerais R.F.Monteiro 233 (RB)
Acronyms of herbaria are according to Thiers B (continuously updated)
54 R. F. Monteiro et al.
123
(Bukatsch 1972), and mounted under a coverslip in 50%
glycerin for light microscope viewing.
For embedding in resin, cross sections from the same
region of the leaf measuring approximately 2 cm2 were
initially fixed in glutaraldehyde, dehydrated in an increas-
ing alcohol series and included in resin hydroxyethylmet-
acrylate. Cross sections of approximately 3 lm thick were
obtained using a Spencer rotary microtome and stained
with Toluidine blue O (O’Brien and McCully 1981).
Digital photomicrographs were taken using a Coolsnap
digital camera coupled with an Olympus BX-50 optical
microscope. Cell and tissue classifications followed the
nomenclature proposed by Tomlinson (1969).
Results
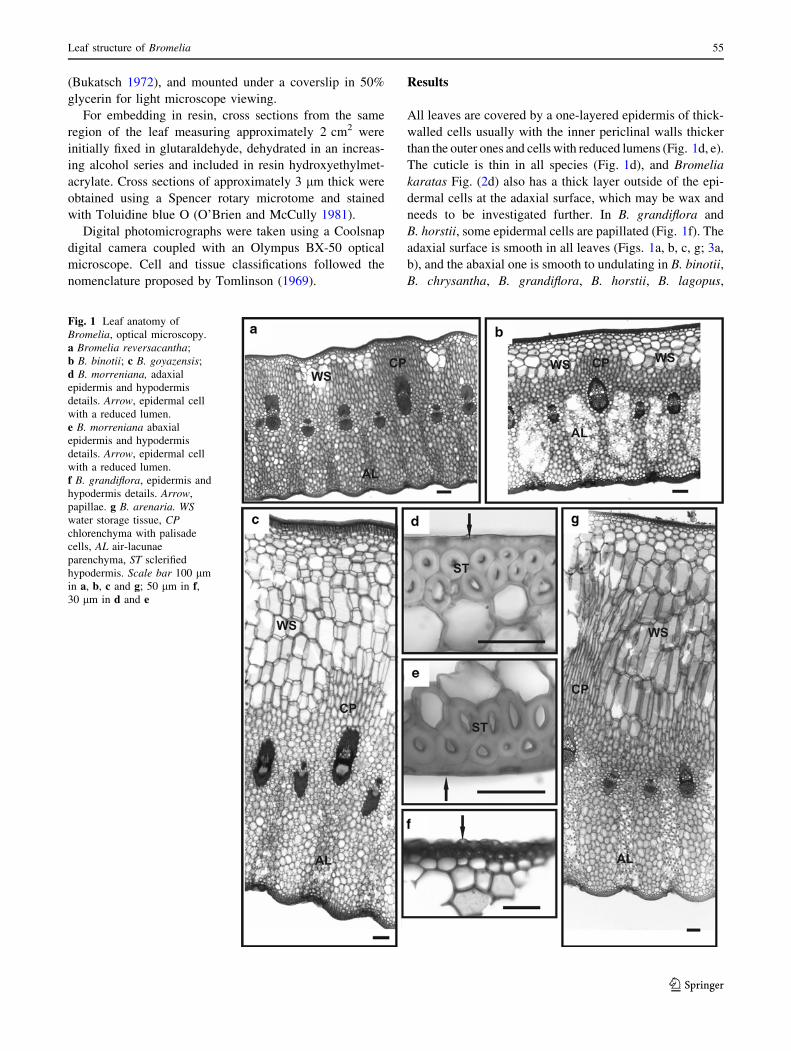
All leaves are covered by a one-layered epidermis of thick-
walled cells usually with the inner periclinal walls thicker
than the outer ones and cells with reduced lumens (Fig. 1d, e).
The cuticle is thin in all species (Fig. 1d), and Bromelia
karatas Fig. (2d) also has a thick layer outside of the epi-
dermal cells at the adaxial surface, which may be wax and
needs to be investigated further. In B. grandiflora and
B. horstii, some epidermal cells are papillated (Fig. 1f). The
adaxial surface is smooth in all leaves (Figs. 1a, b, c, g; 3a,
b), and the abaxial one is smooth to undulating in B. binotii,
B. chrysantha, B. grandiflora, B. horstii, B. lagopus,
Fig. 1 Leaf anatomy of
Bromelia, optical microscopy.
a Bromelia reversacantha;
b B. binotii; c B. goyazensis;
d B. morreniana, adaxial
epidermis and hypodermis
details. Arrow, epidermal cell
with a reduced lumen.
e B. morreniana abaxial
epidermis and hypodermis
details. Arrow, epidermal cell
with a reduced lumen.
f B. grandiflora, epidermis and
hypodermis details. Arrow,
papillae. g B. arenaria. WSwater storage tissue, CPchlorenchyma with palisade
cells, AL air-lacunae
parenchyma, ST sclerified
hypodermis. Scale bar 100 lm
in a, b, c and g; 50 lm in f,30 lm in d and e
Leaf structure of Bromelia 55
123
B. lindevaldae, B. macedoi, B. minima, B. morreniana,
B. scarlatina and B. aff. tubulosa (Figs. 1b, 3a, b), and
strongly ribbed in the remaining species (Fig. 1a, c, g).
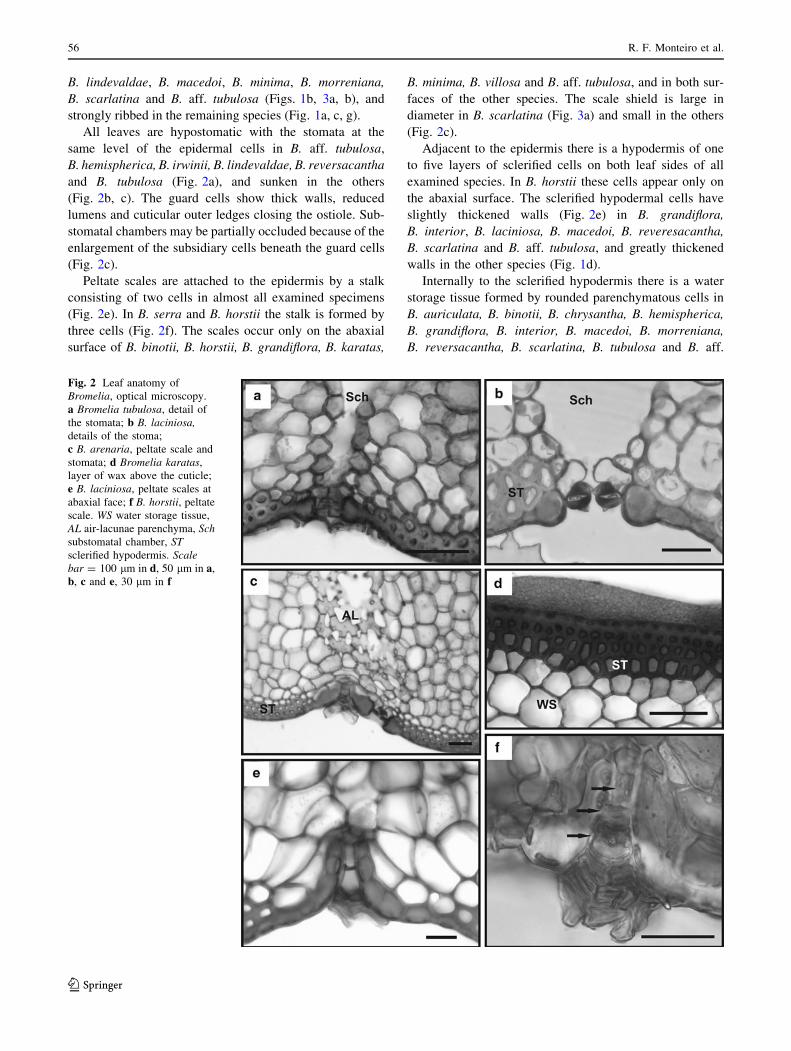
All leaves are hypostomatic with the stomata at the
same level of the epidermal cells in B. aff. tubulosa,
B. hemispherica, B. irwinii, B. lindevaldae, B. reversacantha
and B. tubulosa (Fig. 2a), and sunken in the others
(Fig. 2b, c). The guard cells show thick walls, reduced
lumens and cuticular outer ledges closing the ostiole. Sub-
stomatal chambers may be partially occluded because of the
enlargement of the subsidiary cells beneath the guard cells
(Fig. 2c).
Peltate scales are attached to the epidermis by a stalk
consisting of two cells in almost all examined specimens
(Fig. 2e). In B. serra and B. horstii the stalk is formed by
three cells (Fig. 2f). The scales occur only on the abaxial
surface of B. binotii, B. horstii, B. grandiflora, B. karatas,
B. minima, B. villosa and B. aff. tubulosa, and in both sur-
faces of the other species. The scale shield is large in
diameter in B. scarlatina (Fig. 3a) and small in the others
(Fig. 2c).
Adjacent to the epidermis there is a hypodermis of one
to five layers of sclerified cells on both leaf sides of all
examined species. In B. horstii these cells appear only on
the abaxial surface. The sclerified hypodermal cells have
slightly thickened walls (Fig. 2e) in B. grandiflora,
B. interior, B. laciniosa, B. macedoi, B. reveresacantha,
B. scarlatina and B. aff. tubulosa, and greatly thickened
walls in the other species (Fig. 1d).
Internally to the sclerified hypodermis there is a water
storage tissue formed by rounded parenchymatous cells in
B. auriculata, B. binotii, B. chrysantha, B. hemispherica,
B. grandiflora, B. interior, B. macedoi, B. morreniana,
B. reversacantha, B. scarlatina, B. tubulosa and B. aff.
Fig. 2 Leaf anatomy of
Bromelia, optical microscopy.
a Bromelia tubulosa, detail of
the stomata; b B. laciniosa,details of the stoma;
c B. arenaria, peltate scale and
stomata; d Bromelia karatas,
layer of wax above the cuticle;
e B. laciniosa, peltate scales at
abaxial face; f B. horstii, peltate
scale. WS water storage tissue,
AL air-lacunae parenchyma, Schsubstomatal chamber, STsclerified hypodermis. Scalebar = 100 lm in d, 50 lm in a,
b, c and e, 30 lm in f
56 R. F. Monteiro et al.
123
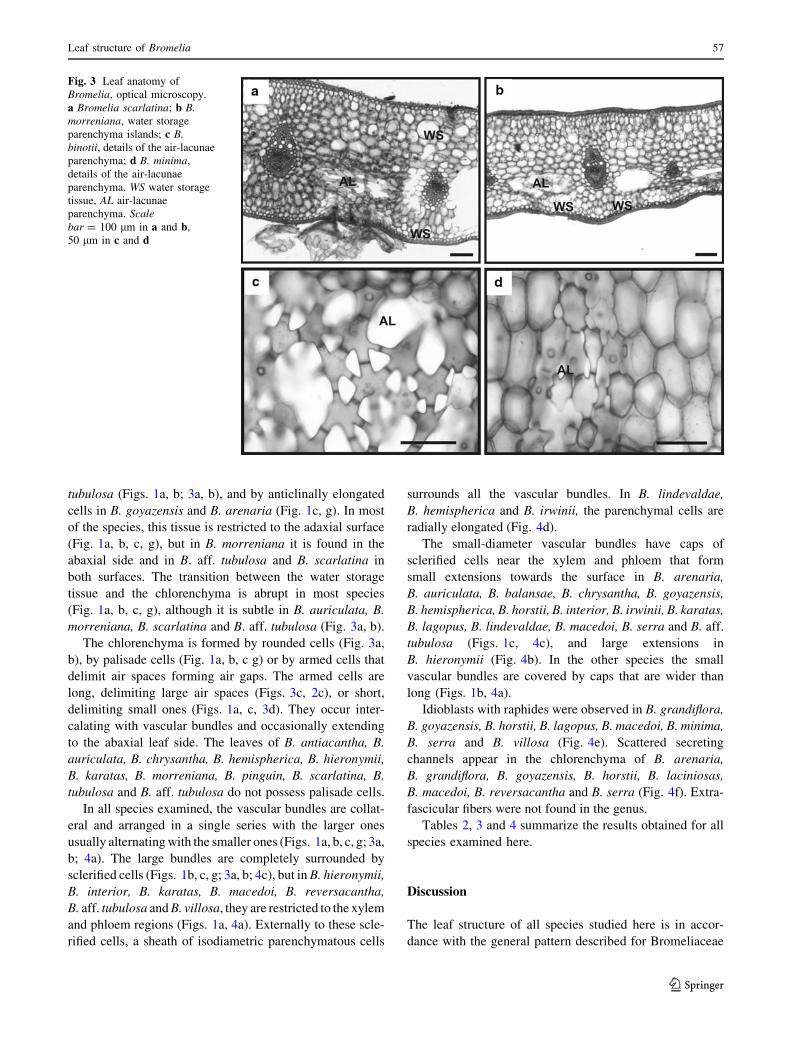
tubulosa (Figs. 1a, b; 3a, b), and by anticlinally elongated
cells in B. goyazensis and B. arenaria (Fig. 1c, g). In most
of the species, this tissue is restricted to the adaxial surface
(Fig. 1a, b, c, g), but in B. morreniana it is found in the
abaxial side and in B. aff. tubulosa and B. scarlatina in
both surfaces. The transition between the water storage
tissue and the chlorenchyma is abrupt in most species
(Fig. 1a, b, c, g), although it is subtle in B. auriculata, B.
morreniana, B. scarlatina and B. aff. tubulosa (Fig. 3a, b).
The chlorenchyma is formed by rounded cells (Fig. 3a,
b), by palisade cells (Fig. 1a, b, c g) or by armed cells that
delimit air spaces forming air gaps. The armed cells are
long, delimiting large air spaces (Figs. 3c, 2c), or short,
delimiting small ones (Figs. 1a, c, 3d). They occur inter-
calating with vascular bundles and occasionally extending
to the abaxial leaf side. The leaves of B. antiacantha, B.
auriculata, B. chrysantha, B. hemispherica, B. hieronymii,
B. karatas, B. morreniana, B. pinguin, B. scarlatina, B.
tubulosa and B. aff. tubulosa do not possess palisade cells.
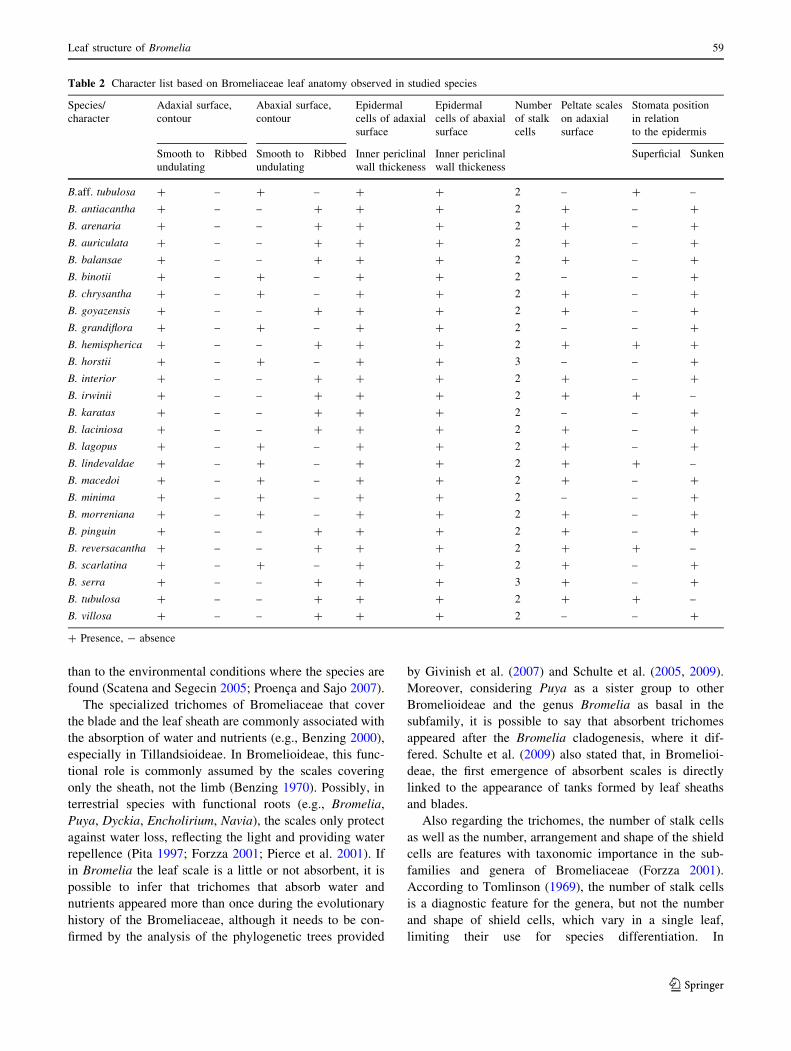
In all species examined, the vascular bundles are collat-
eral and arranged in a single series with the larger ones
usually alternating with the smaller ones (Figs. 1a, b, c, g; 3a,
b; 4a). The large bundles are completely surrounded by
sclerified cells (Figs. 1b, c, g; 3a, b; 4c), but in B. hieronymii,
B. interior, B. karatas, B. macedoi, B. reversacantha,
B. aff. tubulosa and B. villosa, they are restricted to the xylem
and phloem regions (Figs. 1a, 4a). Externally to these scle-
rified cells, a sheath of isodiametric parenchymatous cells
surrounds all the vascular bundles. In B. lindevaldae,
B. hemispherica and B. irwinii, the parenchymal cells are
radially elongated (Fig. 4d).
The small-diameter vascular bundles have caps of
sclerified cells near the xylem and phloem that form
small extensions towards the surface in B. arenaria,
B. auriculata, B. balansae, B. chrysantha, B. goyazensis,
B. hemispherica, B. horstii, B. interior, B. irwinii, B. karatas,
B. lagopus, B. lindevaldae, B. macedoi, B. serra and B. aff.
tubulosa (Figs. 1c, 4c), and large extensions in
B. hieronymii (Fig. 4b). In the other species the small
vascular bundles are covered by caps that are wider than
long (Figs. 1b, 4a).
Idioblasts with raphides were observed in B. grandiflora,
B. goyazensis, B. horstii, B. lagopus, B. macedoi, B. minima,
B. serra and B. villosa (Fig. 4e). Scattered secreting
channels appear in the chlorenchyma of B. arenaria,
B. grandiflora, B. goyazensis, B. horstii, B. laciniosas,
B. macedoi, B. reversacantha and B. serra (Fig. 4f). Extra-
fascicular fibers were not found in the genus.
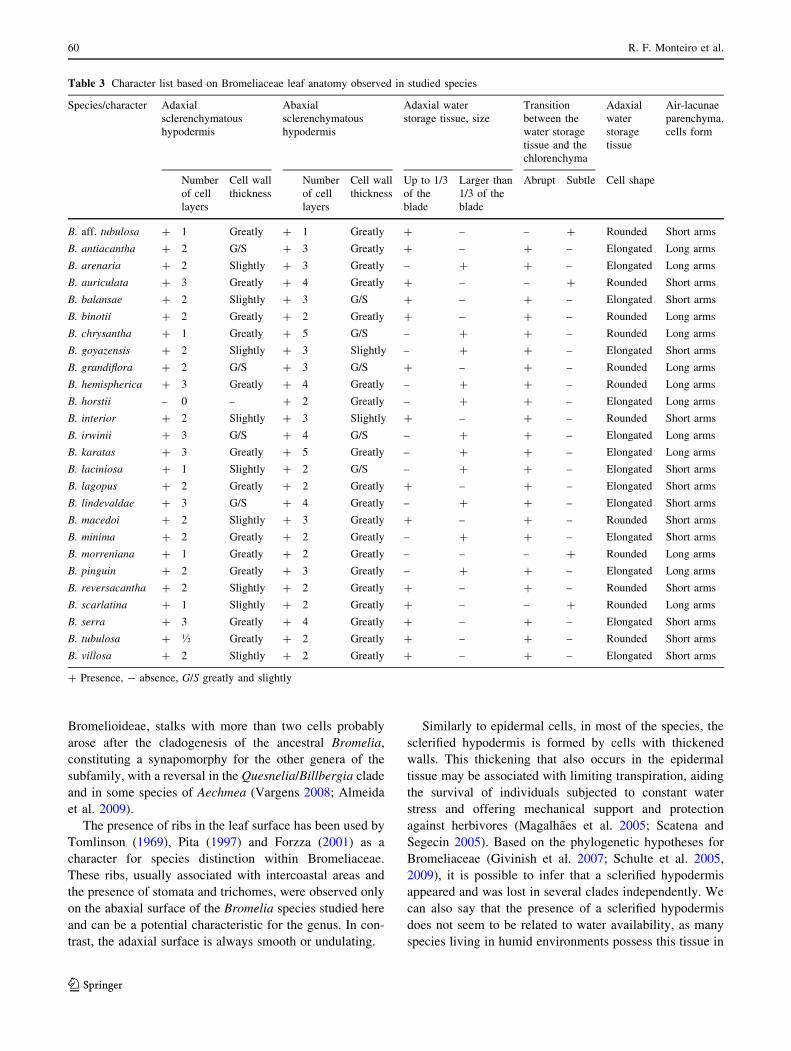
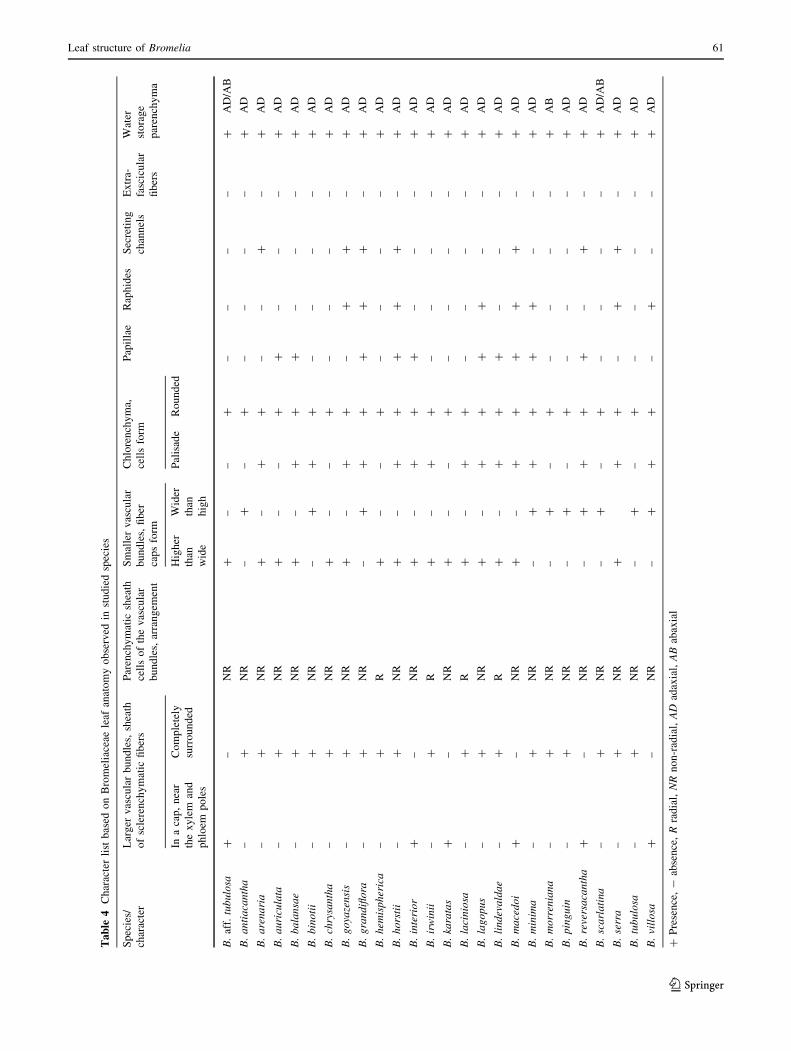
Tables 2, 3 and 4 summarize the results obtained for all
species examined here.
Discussion
The leaf structure of all species studied here is in accor-
dance with the general pattern described for Bromeliaceae
Fig. 3 Leaf anatomy of
Bromelia, optical microscopy.
a Bromelia scarlatina; b B.morreniana, water storage
parenchyma islands; c B.binotii, details of the air-lacunae
parenchyma; d B. minima,
details of the air-lacunae
parenchyma. WS water storage
tissue, AL air-lacunae
parenchyma. Scalebar = 100 lm in a and b,
50 lm in c and d
Leaf structure of Bromelia 57
123

by Tomlinson (1969) and corresponds to type IV of the
classification for the subfamily Bromelioideae suggested
by Horres et al. (2007), which recognizes four groups of
species based on leaf anatomy. In this type of leaves, the
vascular bundles are completely immersed in the chloren-
chyma, the extra-vascular fibers are absent, and both the
adaxial water storage tissue and air gaps in the air lacunae
are developed. Only B. aff. tubulosa, B. scarlatina and B.
morreniana do not completely agree with this type as their
water storage tissue is formed by small groups of paren-
chymatous cells adjacent to the abaxial surface.
The epidermal cells are U-shaped with the inner peri-
clinal wall thickened in all Bromelia studied here, cor-
roborating the description of Tomlinson (1969) for other
representatives of the family. Epidermal cells with uniform
thickness along the entire wall and a large lumen seem to
be a rare character within the Bromeliaceae as it does not
appear in any representatives studied by Aoyoma and Sajo
(2003), Scatena and Segecin (2005), Proenca and Sajo
(2007), Almeida (2006) and Vargens (2008).
The sunken stomata often hidden by the scales, as
observed here for the Bromelia species, is probably asso-
ciated with a reduction in transpiration rates, as suggested
by Tomlinson (1969). A sunken stomata was observed for
other Bromeliaceae from xeric environments (Pita 1997;
Forzza 2001, Arruda and Costa 2003; Proenca and Sajo
2007), although it can also appear in some epiphytic
Aechmea, Billbergia, Hohenbergia, Neoregelia, Quesnelia
and Wittrockia of humid habitats (Sajo et al. 1998; Sousa
et al. 2005; Almeida 2006; Faria 2006; Vargens 2008). In
Bromeliaceae, the stomata are rarely superficial, although
this was observed in Vriesea species of xeric environments
by Arruda and Costa (2003). Thus, the stomata position
appears to be more related to the phylogeny of the group
Fig. 4 Leaf anatomy of
Bromelia, optical microscopy.
a Bromelia villosa, details of the
vascular bundles;
b B. hieronymii, details of the
vascular bundles; c B. serra,
details of the vascular bundles;
d B. irwinii, details of the
vascular bundles; e B. balansae,
raphides; f B. laciniosa,secreting channel. R raphides,
SC secreting channel. Scalebar = 100 lm in a, b, c and d;
50 lm in f; 30 lm in e
58 R. F. Monteiro et al.
123
than to the environmental conditions where the species are
found (Scatena and Segecin 2005; Proenca and Sajo 2007).
The specialized trichomes of Bromeliaceae that cover
the blade and the leaf sheath are commonly associated with
the absorption of water and nutrients (e.g., Benzing 2000),
especially in Tillandsioideae. In Bromelioideae, this func-
tional role is commonly assumed by the scales covering
only the sheath, not the limb (Benzing 1970). Possibly, in
terrestrial species with functional roots (e.g., Bromelia,
Puya, Dyckia, Encholirium, Navia), the scales only protect
against water loss, reflecting the light and providing water
repellence (Pita 1997; Forzza 2001; Pierce et al. 2001). If
in Bromelia the leaf scale is a little or not absorbent, it is
possible to infer that trichomes that absorb water and
nutrients appeared more than once during the evolutionary
history of the Bromeliaceae, although it needs to be con-
firmed by the analysis of the phylogenetic trees provided
by Givinish et al. (2007) and Schulte et al. (2005, 2009).
Moreover, considering Puya as a sister group to other
Bromelioideae and the genus Bromelia as basal in the
subfamily, it is possible to say that absorbent trichomes
appeared after the Bromelia cladogenesis, where it dif-
fered. Schulte et al. (2009) also stated that, in Bromelioi-
deae, the first emergence of absorbent scales is directly
linked to the appearance of tanks formed by leaf sheaths
and blades.
Also regarding the trichomes, the number of stalk cells
as well as the number, arrangement and shape of the shield
cells are features with taxonomic importance in the sub-
families and genera of Bromeliaceae (Forzza 2001).
According to Tomlinson (1969), the number of stalk cells
is a diagnostic feature for the genera, but not the number
and shape of shield cells, which vary in a single leaf,
limiting their use for species differentiation. In
Table 2 Character list based on Bromeliaceae leaf anatomy observed in studied species
Species/
character
Adaxial surface,
contour
Abaxial surface,
contour
Epidermal
cells of adaxial
surface
Epidermal
cells of abaxial
surface
Number
of stalk
cells
Peltate scales
on adaxial
surface
Stomata position
in relation
to the epidermis
Smooth to
undulating
Ribbed Smooth to
undulating
Ribbed Inner periclinal
wall thickeness
Inner periclinal
wall thickeness
Superficial Sunken
B.aff. tubulosa ? – ? – ? ? 2 – ? –
B. antiacantha ? – – ? ? ? 2 ? – ?
B. arenaria ? – – ? ? ? 2 ? – ?
B. auriculata ? – – ? ? ? 2 ? – ?
B. balansae ? – – ? ? ? 2 ? – ?
B. binotii ? – ? – ? ? 2 – – ?
B. chrysantha ? – ? – ? ? 2 ? – ?
B. goyazensis ? – – ? ? ? 2 ? – ?
B. grandiflora ? – ? – ? ? 2 – – ?
B. hemispherica ? – – ? ? ? 2 ? ? ?
B. horstii ? – ? – ? ? 3 – – ?
B. interior ? – – ? ? ? 2 ? – ?
B. irwinii ? – – ? ? ? 2 ? ? –
B. karatas ? – – ? ? ? 2 – – ?
B. laciniosa ? – – ? ? ? 2 ? – ?
B. lagopus ? – ? – ? ? 2 ? – ?
B. lindevaldae ? – ? – ? ? 2 ? ? –
B. macedoi ? – ? – ? ? 2 ? – ?
B. minima ? – ? – ? ? 2 – – ?
B. morreniana ? – ? – ? ? 2 ? – ?
B. pinguin ? – – ? ? ? 2 ? – ?
B. reversacantha ? – – ? ? ? 2 ? ? –
B. scarlatina ? – ? – ? ? 2 ? – ?
B. serra ? – – ? ? ? 3 ? – ?
B. tubulosa ? – – ? ? ? 2 ? ? –
B. villosa ? – – ? ? ? 2 – – ?
? Presence, - absence
Leaf structure of Bromelia 59
123
Bromelioideae, stalks with more than two cells probably
arose after the cladogenesis of the ancestral Bromelia,
constituting a synapomorphy for the other genera of the
subfamily, with a reversal in the Quesnelia/Billbergia clade
and in some species of Aechmea (Vargens 2008; Almeida
et al. 2009).
The presence of ribs in the leaf surface has been used by
Tomlinson (1969), Pita (1997) and Forzza (2001) as a
character for species distinction within Bromeliaceae.
These ribs, usually associated with intercoastal areas and
the presence of stomata and trichomes, were observed only
on the abaxial surface of the Bromelia species studied here
and can be a potential characteristic for the genus. In con-
trast, the adaxial surface is always smooth or undulating.
Similarly to epidermal cells, in most of the species, the
sclerified hypodermis is formed by cells with thickened
walls. This thickening that also occurs in the epidermal
tissue may be associated with limiting transpiration, aiding
the survival of individuals subjected to constant water
stress and offering mechanical support and protection
against herbivores (Magalhaes et al. 2005; Scatena and
Segecin 2005). Based on the phylogenetic hypotheses for
Bromeliaceae (Givinish et al. 2007; Schulte et al. 2005,
2009), it is possible to infer that a sclerified hypodermis
appeared and was lost in several clades independently. We
can also say that the presence of a sclerified hypodermis
does not seem to be related to water availability, as many
species living in humid environments possess this tissue in
Table 3 Character list based on Bromeliaceae leaf anatomy observed in studied species
Species/character Adaxial
sclerenchymatous
hypodermis
Abaxial
sclerenchymatous
hypodermis
Adaxial water
storage tissue, size
Transition
between the
water storage
tissue and the
chlorenchyma
Adaxial
water
storage
tissue
Air-lacunae
parenchyma,
cells form
Number
of cell
layers
Cell wall
thickness
Number
of cell
layers
Cell wall
thickness
Up to 1/3
of the
blade
Larger than
1/3 of the
blade
Abrupt Subtle Cell shape
B. aff. tubulosa ? 1 Greatly ? 1 Greatly ? – – ? Rounded Short arms
B. antiacantha ? 2 G/S ? 3 Greatly ? – ? – Elongated Long arms
B. arenaria ? 2 Slightly ? 3 Greatly – ? ? – Elongated Long arms
B. auriculata ? 3 Greatly ? 4 Greatly ? – – ? Rounded Short arms
B. balansae ? 2 Slightly ? 3 G/S ? – ? – Elongated Short arms
B. binotii ? 2 Greatly ? 2 Greatly ? – ? – Rounded Long arms
B. chrysantha ? 1 Greatly ? 5 G/S – ? ? – Rounded Long arms
B. goyazensis ? 2 Slightly ? 3 Slightly – ? ? – Elongated Short arms
B. grandiflora ? 2 G/S ? 3 G/S ? – ? – Rounded Long arms
B. hemispherica ? 3 Greatly ? 4 Greatly – ? ? – Rounded Long arms
B. horstii – 0 – ? 2 Greatly – ? ? – Elongated Long arms
B. interior ? 2 Slightly ? 3 Slightly ? – ? – Rounded Short arms
B. irwinii ? 3 G/S ? 4 G/S – ? ? – Elongated Long arms
B. karatas ? 3 Greatly ? 5 Greatly – ? ? – Elongated Long arms
B. laciniosa ? 1 Slightly ? 2 G/S – ? ? – Elongated Short arms
B. lagopus ? 2 Greatly ? 2 Greatly ? – ? – Elongated Short arms
B. lindevaldae ? 3 G/S ? 4 Greatly – ? ? – Elongated Short arms
B. macedoi ? 2 Slightly ? 3 Greatly ? – ? – Rounded Short arms
B. minima ? 2 Greatly ? 2 Greatly – ? ? – Elongated Short arms
B. morreniana ? 1 Greatly ? 2 Greatly – – – ? Rounded Long arms
B. pinguin ? 2 Greatly ? 3 Greatly – ? ? – Elongated Long arms
B. reversacantha ? 2 Slightly ? 2 Greatly ? – ? – Rounded Short arms
B. scarlatina ? 1 Slightly ? 2 Greatly ? – – ? Rounded Long arms
B. serra ? 3 Greatly ? 4 Greatly ? – ? – Elongated Short arms
B. tubulosa ? � Greatly ? 2 Greatly ? – ? – Rounded Short arms
B. villosa ? 2 Slightly ? 2 Greatly ? – ? – Elongated Short arms
? Presence, - absence, G/S greatly and slightly
60 R. F. Monteiro et al.
123
Ta
ble
4C
har
acte
rli
stb
ased
on
Bro
mel
iace
aele
afan
ato
my
ob
serv
edin
stu
die
dsp
ecie
s
Sp
ecie
s/
char
acte
r
Lar
ger
vas
cula
rb
un
dle
s,sh
eath
of
scle
ren
chy
mat
icfi
ber
s
Par
ench
ym
atic
shea
th
cell
so
fth
ev
ascu
lar
bu
nd
les,
arra
ng
emen
t
Sm
alle
rv
ascu
lar
bu
nd
les,
fib
er
cap
sfo
rm
Ch
lore
nch
ym
a,
cell
sfo
rm
Pap
illa
eR
aph
ides
Sec
reti
ng
chan
nel
s
Ex
tra-
fasc
icu
lar
fib
ers
Wat
er
sto
rag
e
par
ench
ym
a
Ina
cap
,n
ear
the
xy
lem
and
ph
loem
po
les
Co
mp
lete
ly
surr
ou
nd
ed
Hig
her
than
wid
e
Wid
er
than
hig
h
Pal
isad
eR
ou
nd
ed
B.
aff.
tub
ulo
sa?
–N
R?
––
?–
––
–?
AD
/AB
B.
an
tia
can
tha
–?
NR
–?
–?
––
––
?A
D
B.
are
na
ria
–?
NR
?–
??
––
?–
?A
D
B.
au
ricu
lata
–?
NR
?–
–?
?–
––
?A
D
B.
ba
lan
sae
–?
NR
?–
??
?–
––
?A
D
B.
bin
oti
i–
?N
R–
??
?–
––
–?
AD
B.
chry
san
tha
–?
NR
?–
–?
––
––
?A
D
B.
go
yaze
nsi
s–
?N
R?
–?
?–
??
–?
AD
B.
gra
nd
iflo
ra–
?N
R–
??
??
??
–?
AD
B.
hem
isp
her
ica
–?
R?
––
?–
––
–?
AD
B.
ho
rsti
i–
?N
R?
–?
??
??
–?
AD
B.
inte
rio
r?
–N
R?
–?
??
––
–?
AD
B.
irw
inii
–?
R?
–?
?–
––
–?
AD
B.
kara
tas
?–
NR
?–
–?
––
––
?A
D
B.
laci
nio
sa–
?R
?–
??
––
––
?A
D
B.
lag
op
us
–?
NR
?–
??
??
––
?A
D
B.
lin
dev
ald
ae
–?
R?
–?
??
––
–?
AD
B.
ma
ced
oi
?–
NR
?–
??
??
?–
?A
D
B.
min
ima
–?
NR
–?
??
??
––
?A
D
B.
mo
rren
ian
a–
?N
R–
?–
?–
––
–?
AB
B.
pin
gu
in–
?N
R–
?–
?–
––
–?
AD
B.
reve
rsa
can
tha
?–
NR
–?
??
?–
?–
?A
D
B.
sca
rla
tin
a–
?N
R–
?–
?–
––
–?
AD
/AB
B.
serr
a–
?N
R?
??
–?
?–
?A
D
B.
tub
ulo
sa–
?N
R–
?–
?–
––
–?
AD
B.
vill
osa
?–
NR
–?
??
–?
––
?A
D
?P
rese
nce
,-
abse
nce
,R
rad
ial,
NR
no
n-r
adia
l,A
Dad
axia
l,A
Bab
axia
l
Leaf structure of Bromelia 61
123

their leaves (e.g., Sousa et al. 2005, Almeida 2006; Faria
2006), and others in non-xeric settings also do (e.g., Forzza
2001). The number of layers of hypodermis cells is variable
inside the genera in Bromeliaceae (Tomlinson 1969; Sousa
et al. 2005; Almeida 2006; Faria 2006; Vargens 2008).
Near the sclerified hypodermis a multilayered paren-
chymatous tissue provides water storage, making the leaves
more or less succulent (Mantovani 1999; Proenca and Sajo
2004; Scatena and Segecin 2005). The cells of this tissue
can be isodiametric or vary from isodiametric to anticlinal
elongated in the inner layers, as observed in B. antiacan-
tha. Occasionally, these cells show shrinkage, possibly due
to water loss (Krauss 1949), and, because they are located
just above the photosynthetic layers, Brighigna et al.
(1984) argued that this region protects the chlorenchyma
from high light levels in order to optimize the photosyn-
thesis and reduce the risk of photoinhibition.
An abaxial water storage tissue was only observed in B.
aff. tubulosa, B. morreniana and B. scarlatina, and seems
to be unusual in other basal genera of Bromelioideae (Terry
et al. 1997; Sajo et al. 1998; Schulte et al. 2005; Givinish
et al. 2004; 2007). Apparently, this is a derived character
that appeared several times in different genera of the
subfamily.
Chlorenchyma endowed with palisade cells, as observed
here for the Bromelia, is an uncommon feature in the
family, and up to the present has been found only in Puya,
Brocchinia, Pitcairnia, Encholirum and Acanthostachys
(Tomlinson 1969; Downs 1974; Forzza 2001). Considering
the phylogenetic hypotheses for the family (Givinish et al.
2007; Schulte et al. 2005; 2009), we can infer that palisade
tissue had appeared several times independently. However,
the presence of this feature in both Puya and Bromelia
suggests that it can occur in other Bromelioideae and had
appeared again in Acanthostachys.
The aeration channels delimited by armed parenchy-
matous cells, as observed here for Bromelia, are common
in Bromeliaceae (Aoyoma and Sajo 2003; Scatena and
Segecin 2005; Proenca and Sajo 2007). According to
Tomlinson (1969) and Gilmartin et al. (1989), these
channels facilitate leaf ventilation. In these channels, the
cell shape is constant for the same leaf region in the same
species and can be useful to distinguish the Bromelia
(Braga 1977), because the arm size of the cells varies from
species to species.
Regarding the secreting channels of Bromeliaceae,
Tomlinson (1969) describes them as formed by cells with a
mucilaginous content in Aechmea and Billbergia. How-
ever, for Bromelia species they are reported here for the
first time. As recent studies do not corroborate the presence
of such channels in Aechmea and Billbergia (Aoyoma and
Sajo 2003; Sousa et al. 2005; Vargens 2008), it is possible
to say that they are rare in the family and are an important
taxonomic characteristic for differentiating Bromelia from
the other Bromelioideae. Although raphides are common
within the family, it is possible to separate species of
Aechmea (Aoyoma and Sajo 2003; Sousa et al. 2005) from
those of Bromelia by the presence or lack of them.
The vascular bundles are arranged in a single series, as
in other species of the family (Tomlinson 1969; Downs
1974; Pita 1997; Forzza 2001; Arruda and Costa 2003;
Aoyoma and Sajo 2003, Scatena and Segecin 2005 ; Sousa
et al. 2005; Proenca and Sajo 2007), and can be completely
surrounded by sclerified cells or present fiber caps adjacent
to xylem and phloem. This variation was also found in
several other groups of the family (e.g., Tomlinson 1969;
Aoyoma and Sajo 2003) and seems to have arisen inde-
pendently several times during the evolutionary history of
the group. However, this character seems important for
supporting small clades within Bromelia (Monteiro 2009)
and may also be useful in the taxonomy of other genera.
The small vascular bundles often show small fiber
extensions towards the abaxial surface in Bromelia species.
Large vascular fiber extensions as well as their complete
absence are rare in the genus. This feature may have been
lost several times independently during the evolution of the
family because these fibers do not appear in many groups
of different sides of the phylogenetic tree, like all studied
Tillandsioideae (Tomlinson 1969; Arruda and Costa 2003;
Segecin and Scatena 2005; Proenca and Sajo 2007;
Givinish et al. 2007). Each vascular bundle is also sur-
rounded by an outer parenchymatous sheath whose cells,
with slightly thickened walls and a wide lumen, are in a
slightly radiating arrangement in three Bromelia. The
radiating arrangement of the parenchymatous sheath was
also reported for other genera, such as Aechmea, Brew-
caria, Brocchinia, Billbergia, Encholirium, Fernesea,
Hohenbergia, Pitcairnia, Portea, Puya and Quesnelia
(Sajo et al. 1998; Forzza 2001; Aoyoma and Sajo 2003;
Sousa et al. 2005; Almeida 2006). Considering the known
phylogenetic relationships among these genera (Terry et al.
1997; Givinish et al. 2004, 2007; Barfuss et al. 2005,
Schulte et al. 2005), we can assume that this is a highly
homoplastic feature.
Fibers not associated with vascular bundles are unusual
in Bromeliaceae, occurring only in derived Bromelioideae
genera, like Aechmea, Canistrum and Quesnelia, and being
rare in Tillandsioideae (Tomlinson 1969; Proenca and Sajo
2007). Taking into account the basal position of Bromelia
(Givinish et al. 2004, 2007; Schulte et al. 2005, 2009), the
absence of extra-fascicular fibers in these genera and in
Puya seems to be a plesiomorphic state within the
subfamily.
The four layers of abaxial sclerenchymatous hypodermis
are the only anatomic synapomorphy for Bromelia
(Monteiro 2009). The results presented here show that,
62 R. F. Monteiro et al.
123

although we can recognize groups of species of Bromelia
using leaf anatomy, no characteristic is unique to any of the
three subgenera proposed for Bromelia by Mez (1891).
Based on the present results as well as on those previ-
ously described, the leaf anatomy seems to provide features
useful in the phylogeny and the delimitation of the Bro-
meliaceae taxa. Therefore, micromorphology must be
emphasized alongside the macromorphology in the sys-
tematics of Bromeliaceae and of monocots over molecular
characters to produce better results for phylogenies as a
rewarding approach, as recently shown by Horres et al.
(2007) and Faria (2006).
Acknowledgments The authors would like to thank Anna Karla
Venda for assistance with the cuttings, Renato de Mello-Silva, Andrea
Ferreira da Costa, Cassia Monica Sakuragui, Daniela Zappi and Graca
Sajo for the criticisms and suggestions, and Conselho Nacional de
Desenvolvimento Cientıfico e Tecnologico (CNPq) and Coordenad-
oria de Aperfeicoamento do Pessoal de Nıvel Superior (CAPES) for
fellowships.
References
Almeida VR (2006) Filogenia e circunscricao de Quesnelia Gaudch.
(Bromeliaceae–Bromelioideae). Master dissertation, Universid-
ade Federal do Rio de Janeiro, Rio de Janeiro, Rio de Janeiro,
Brasil
Almeida VR, Costa AF, Mantovani A, Goncalves-Esteves V, Arruda
RCO, Forzza RC (2009) Morphological phylogenetics of
Quesnelia (Bromeliaceae, Bromelioideae). Syst Bot 34(4):660–
672
Aoyoma EM, Sajo MG (2003) Estrutura foliar de Aechmea Ruiz &
Pav. subgenero Lamproccocus (Beer) Baker e especies relacio-
nadas. Revista Brasileira de Botanica 26:461–473
Arruda RCO, Costa AF (2003) Foliar anatomy of five Vriesea sect.
Xyphion (Bromeliaceae) species. Selbyana 24(2):180–189
Barfuss MHJ, Samuel R, Till W, Stuessy TF (2005) Phylogenetic
relationships in subfamily Tillandsioideae (Bromeliaceae) based
on DNA sequence data from seven plastid regions. Am J Botany
92(2):337–351
Benzing DH (1970) Foliar permeability and the absorbtion of
minerals and organic nitrogen by certain tank bromeliads. Bot
Gaz 131:23–31
Benzing DH (2000) Bromeliaceae: profile of an adaptive radiation.
Cambridge University Press, Cambridge
Braga MMN (1977) Anatomia foliar de Bromeliaceae da Campina.
Acta Amazonica 7:1–74
Brighigna L, Fiordi AC, Palandri MR (1984) Structural characteristics
of mesophyll in some Tillandsia species. Phytomorphology
34:191–200
Brown GK (2000) Dados Moleculares em Bromeliaceae. In: Leme
EMC (ed) Nidularium: Bromelias da Mata Atlantica. Ed.
Salamandra, Rio de Janeiro, pp 198–201
Bukatsch F (1972) Bermerkungen zur Doppelfarbung Astrablau-
Safranin. Mikrokosmos 61:255
Coffani-Nunes JV (2004) Revisao Taxonomica e Filogenia de PorteaBrong. ex K. Koch (Bromelioideae-Bromeliaceae). Ph.D. dis-
sertation, Universidade de Sao Paulo
Downs RJ (1974) Anatomy and physiology. In: Smith LB, Downs RJ
(eds) Pitcairnioideae (Bromeliaceae). Flora Neotropica, mono-
graph14. Hafner, New York, pp 2–28
Faria APG (2006) Revisao taxonomica e filogenia de Aechmea Ruiz
& Pav. subg. Macrochordion (De Vriese) Baker, Bromelioi-
deae—Bromeliaceae. Ph.D. dissertation, Universidade Federal
do Rio de Janeiro
Forzza RC (2001) Filogenia da tribo Puyeae Wittm. e revisao
taxonomica do genero Encholirium Mart. ex Schult. & Schult. f.
(Pitcairnioideae—Bromeliaceae). Ph.D. dissertation, Universi-
dade de Sao PauloFurness CA, Rudall PJ (2000) The systematic significance of
simultaneous cytokinesis during microsporogenesis in monocot-
yledons. In: Wilson KJ, Morrison DA (eds) Monocots: system-
atics and evolution. CSTRO, Australia, pp 189–193
Gilmartin AJ, Brown GK, Varadarajan GS, Neighbours M (1989)
Status of Glomeropitcairnia within evolutionary history of
Bromeliaceae. Syst Bot 14:339–348
Givinish TJ, Millam KC, Evans TM, Hall JC, Pires JC, Barry PE,
Sytsam KJ (2004) Ancient vicariance or recent long-distance
dispersal? Inferences about phylogeny and South American-
African disjunctions in Rapateaceae and Bromeliaceae based on
ndhF sequence data. Intern J Plant Sci 165:35–54
Givinish TJ, Millam KC, Barry PE, Sytsam KJ (2007) Phylogeny,
adaptive radiation and historical biogeography of Bromeliaceae
inferred from ndhF sequence data. In: Colombus JT, Friar EA,
Porter JM, Prince LM, Simpson MG (eds) Monocots: compar-
ative biology and evolution—Poales. Rancho Santa Ana Botanic
Garden, California, pp 3–26
Henderson FM, Stevenson DW (2007) A phylogenetic study of
Arecaceae based on seedling morphological and anatomical data.
In: Colombus JT, Friar EA, Porter JM, Prince LM, Simpson MG
(eds) Monocots: comparative biology and evolution—Poales.
Rancho Santa Ana Botanic Garden, California, pp 251–264
Horres R, Schulte K, Weising K, Zizka G (2007) Systematics of
Bromelioideae (Bromeliaceae)—evidence from molecular and
anatomical studies. In: Colombus JT, Friar EA, Porter JM, Prince
LM, Simpson MG (eds) Monocots: comparative biology and
evolution—Poales. Rancho Santa Ana Botanic Garden, Califor-
nia, pp 27–43
Krauss BH (1949) Anatomy of vegetative organs of the pineapple
Ananas comosus (L.) Merr. Bot Gaz 10:330–404
Luther HE (2008) An alphabetical list of bromeliad binomies. The
Bromeliad Society International, 11th edn. The Marie Selby
Botanical Gardens, Sarasota
Magalhaes N, Mantovani A, Resende B, Leitao G, Teixeira ML
(2005) First report on host plants and feeding habits of the leaf
beetle Acentroptera pulchella Guerin-Meneville (Chrysomeli-
dae, Hispinae). In: Konstantinov A, Penev L, Tischechki A (eds)
Contributions to systematics and biology of insects: papers
celebrating the 80th birthday of I.K. Lopatin. Pensoft Publishers,
Moscow, pp 153–157
Mantovani A (1999) Leaf morphophysiology and distribution of
epiphytic aroids along a vertical gradient in a Brazilian rain
forest. Selbyana 20(2):241–249
Mez C (1891) Bromeliaceae. In: Martius CFP von, Eichler AW,
Urban I (eds), Flora brasiliensis, vol. 3, part. 3, Munchen, Wien,
Leipzig, pp 173–634
Monteiro RF (2009) Estudos anatomicos e filogeneticos em BromeliaL. (Bromeliaceae, Bromeliodeae). Master dissertation, Jardim
Botanico do Rio de Janeiro, Rio de Janeiro
O’Brien TP, McCully ME (1981) The study of plant structure:
principles and selected methods. Thermarcarphi, Melbourne
Pierce S, Maxwell K, Griffiths H, Winter K (2001) Hydrophobic
trichome layers and epicuticular wax powders in Bromeliaceae.
Am J Bot 88:1371–1389
Pita PB (1997) Estudos anatomicos de orgaos vegetativos de Dyckia e
Encholirium (Bromeliaceae) da Serra do Cipo (Minas Gerais,
Brasil). Master dissertation, Universidade de Sao Paulo
Leaf structure of Bromelia 63
123

Proenca SL, Sajo MG (2004) Estrutura foliar de especies de AechmeaRuiz & Pav. (Bromeliaceae) do estado de Sao Paulo, Brasil. Acta
Botanica Brasilica 18(2):319–331
Proenca SL, Sajo MG (2007) Anatomia foliar de bromelias em areas
de cerrado do estado de Sao Paulo, Brasil. Acta Botanica
Brasilica 21(3):657–673
Proenca SL, Sajo MG (2008) Anatomy of the floral scape of
Bromeliaceae. Revista Brasileira de Botanica 31:399–408
Ramirez IM (1996) Systematics, phylogeny and chromosome number
evolution in Cryptanthus (Bromeliaceae). Ph.D. dissertation,
University of Missouri
Sajo MG, Machado SR, Camello-Guerreiro SM (1998) Aspectos
estruturais de folha de bromelia e suas implicacoes no agrupa-
mento de especies. In: Pereira MV (ed) Bromelias da Mata
Atlantica: Canistropsis. Salamandra, Rio de Janeiro, pp 102–111
Scatena VL, Segecin E (2005) Anatomia foliar de Tillandsia L.
(Bromeliaceae) dos Campos Gerais, Parana, Brasil. Revista
Brasileira Botanica 28(3):635–649
Schmidt RE, Brown GK (2004) Leaf anatomy in Bromeliaceae
subfamily Bromelioideae-I historical perspectives. Vidalia
2(2):11–22
Schulte K, Horres R, Zizka G (2005) Molecular phylogeny of
Bromelioideae and its implications on biogeography and the
evolution of CAM in the family (Poales, Bromeliaceae).
Senckengergiana biologica 85:113–125
Schulte K, Barfuss MHJ, Zizka G (2009) Phylogeny of Bromelioideae
(Bromeliaceae) inferred from nuclear and plastid DNA loci
reveals the evolution of the tank habit within the subfamily. Mol
Phylogenet Evol 51:327–339
Simon BK (2007) Grass phylogeny and classification: conflict of
morphology and molecules. In: Colombus JT, Friar EA, Porter
JM, Prince LM, Simpson MG (eds) Monocots: comparative
biology and evolution—Poales. Rancho Santa Ana Botanic
Garden, Claremont, pp 259–266
Simpson MG, Burton DH (2006) Systematic floral anatomy of
Pontederiaceae In: Colombus JT, Friar EA, Porter JM, Prince
LM, Simpson MG (eds) Monocots: comparative biology and
evolution—excluding Poales. Rancho Santa Ana Botanic Gar-
den, Claremont, pp 499–519
Smith LB, Downs JR (1979) Bromelioideae (Bromeliaceae). Flora
Neotropica Monograph 14. Hafner Press, New York
Sousa GM (2004) Revisao taxonomica de Aechmea Ruiz & Pavon
subg. Chevaliera (Gaudich. ex Beer) Baker (Bromelioideae—
Bromeliaceae). Ph.D. dissertation, Universidade de Sao Paulo
Sousa RCOS, Neves LJ (1996) Leaf anatomy of four Tillandsiaspecies. Bromelia 3:28–39
Sousa GM, Estelita MEM, Wanderley MGL (2005) Anatomia foliar
de especies brasileiras de Aechmea subg. Chevaliera (Gaudich.
ex Beer) Baker, Bromelioideae-Bromeliaceae. Revista Brasileira
Botanica 28(3):603–613
Sousa LOF, Wendt T, Brown GK, Tuthill GE, Evans TM (2007)
Monophyly and phylogenetic relationships in Lymania (Brome-
lioideae-Bromeliaceae) based on morphology and chloroplast
DNA sequences. Syst Bot 32(2):264–270
Stevenson DW, Davis JI, Freudenstein JV, Hardy CR, Simmons MP,
Specht CD (2000) A phylogenetic analysis of monocotyledons
based on morphological and molecular characters sets, with
comments on the placement of Acorus and Hydatellaceae. In:
Wilson KL, Morriso DA (eds) Monocots: systematics and
evolution. CSTRO, Austria, pp 17–24
Terry RG, Brown GK, Olmstead RG (1997) Examination of
subfamilial phylogeny in Bromeliaceae using comparative
sequencing of the plastid locus ndhF. Am J Botany 84(5):
664–670
Thiers B (continuously updated) Index herbariorum. A global
directory of public herbaria associated staff. New York Botanical
Garden’s Virtual Herbarium. http://sweetgum.nybg.org/ih/.
Accessed 17 March 2010
Tomlinson PB (1969) Commelinales-Zingiberales. In: Metcalf CR
(ed) Anatomy of the monocotyledons. Claredon Press, Oxford,
pp 193–294
Tomlinson PB, Fisher JB (2000) Stem vascular in climbing mono-
cotyledons: a comparative approach. In: Wilson KL, Morriso DA
(eds) Monocots: systematics and evolution. CSTRO, Austria,
pp 89–100
Vargens FAC (2008) Anatomia Foliar de Billbergia Thunb.
(Bromelioideae-Bromeliaceae). Master dissertation, Universid-
ade Federal do Rio de Janeiro
64 R. F. Monteiro et al.
123
Copyright © 2022 FDOKUMEN




















