
Leaf litter fungi in a Central Amazonian forest: the influence of rainfall, soil and topography on...
12
Biodivers Conserv (2008) 17:2701–2712 DOI 10.1007/s10531-007-9247-6 1 C ORIGINAL PAPER Leaf litter fungi in a Central Amazonian forest: the inXuence of rainfall, soil and topography on the distribution of fruiting bodies Ricardo Braga-Neto · Regina Celi Costa Luizão · William Ernest Magnusson · Gabriela Zuquim · Carolina Volkmer de Castilho Received: 5 April 2007 / Accepted: 24 August 2007 / Published online: 15 September 2007 © Springer Science+Business Media B.V. 2007 Abstract Fungi are important components of tropical ecosystems, especially in the recy- cling of nutrients. However, there is little information on how fungal diversity is structured at scales suitable to plan their conservation. We tested if the distribution of fruiting bodies of litter fungi was random in the landscape (over 25 km 2 ) in a tropical evergreen forest in Central Amazonia. We used linear regressions to evaluate the inXuence of rainfall, soil characteristics and topography on morphospecies richness and composition. Fungi were collected twice in thirty 0.25 £ 250 m plots. Short-term rainfall was represented by the cumulative rainfall in the three days before each plot was surveyed. Plots were classiWed in two groups based on cumulative rainfall. Clay content in soil and rainfall inXuenced mor- phospecies richness, but responses to edaphic factors depended on rainfall. Wetter periods apparently decreased limiting moisture conditions in higher areas, allowing fungal activity and fruiting body production. Morphospecies composition was inXuenced by clay content, but inXuence on fungi was probably indirect as clay content was correlated with altitude, plant community and nitrogen availability. Our results suggest that the species of litter fungi are not randomly distributed in the landscape. Furthermore, they indicate that it is viable to conduct mesoscale evaluations of fungal diversity, if the temporal and spatial var- iation and their interaction are taken into account. Resumo (in Portuguese) Fungos são importantes componentes dos ecossistemas tropi- cais, atuando especialmente na reciclagem de nutrientes. Entretanto, existe pouca infor- mação sobre como a diversidade de fungos está estruturada em escalas adequadas para planejar sua conservação. Nós testamos se a distribuição de corpos de frutiWcação de fun- gos de liteira ocorre de forma aleatória na paisagem (em 25 km 2 ) em uma Xoresta na Amazônia Central. Utilizamos regressões lineares para avaliar a inXuência da precipitação, do solo e topograWa sobre a riqueza e composição de morfoespécies. Os fungos foram R. Braga-Neto (&) · R. C. C. Luizão · W. E. Magnusson · G. Zuquim · C. V. de Castilho Instituto Nacional de Pesquisas da Amazônia, Coordenação de Pesquisas em Ecologia, Avenida EWgênio Sales, 2239, Manaus, CP 478, CEP 69011–970, Brazil e-mail: [email protected]
Transcript of Leaf litter fungi in a Central Amazonian forest: the influence of rainfall, soil and topography on...

Biodivers Conserv (2008) 17:2701–2712 DOI 10.1007/s10531-007-9247-6
ORIGINAL PAPER
Leaf litter fungi in a Central Amazonian forest:the inXuence of rainfall, soil and topographyon the distribution of fruiting bodies
Ricardo Braga-Neto · Regina Celi Costa Luizão ·William Ernest Magnusson · Gabriela Zuquim · Carolina Volkmer de Castilho
Received: 5 April 2007 / Accepted: 24 August 2007 / Published online: 15 September 2007© Springer Science+Business Media B.V. 2007
Abstract Fungi are important components of tropical ecosystems, especially in the recy-cling of nutrients. However, there is little information on how fungal diversity is structuredat scales suitable to plan their conservation. We tested if the distribution of fruiting bodiesof litter fungi was random in the landscape (over 25 km2) in a tropical evergreen forest inCentral Amazonia. We used linear regressions to evaluate the inXuence of rainfall, soilcharacteristics and topography on morphospecies richness and composition. Fungi werecollected twice in thirty 0.25 £ 250 m plots. Short-term rainfall was represented by thecumulative rainfall in the three days before each plot was surveyed. Plots were classiWed intwo groups based on cumulative rainfall. Clay content in soil and rainfall inXuenced mor-phospecies richness, but responses to edaphic factors depended on rainfall. Wetter periodsapparently decreased limiting moisture conditions in higher areas, allowing fungal activityand fruiting body production. Morphospecies composition was inXuenced by clay content,but inXuence on fungi was probably indirect as clay content was correlated with altitude,plant community and nitrogen availability. Our results suggest that the species of litterfungi are not randomly distributed in the landscape. Furthermore, they indicate that it isviable to conduct mesoscale evaluations of fungal diversity, if the temporal and spatial var-iation and their interaction are taken into account.Resumo (in Portuguese) Fungos são importantes componentes dos ecossistemas tropi-cais, atuando especialmente na reciclagem de nutrientes. Entretanto, existe pouca infor-mação sobre como a diversidade de fungos está estruturada em escalas adequadas paraplanejar sua conservação. Nós testamos se a distribuição de corpos de frutiWcação de fun-gos de liteira ocorre de forma aleatória na paisagem (em 25 km2) em uma Xoresta naAmazônia Central. Utilizamos regressões lineares para avaliar a inXuência da precipitação,do solo e topograWa sobre a riqueza e composição de morfoespécies. Os fungos foram
R. Braga-Neto (&) · R. C. C. Luizão · W. E. Magnusson · G. Zuquim · C. V. de CastilhoInstituto Nacional de Pesquisas da Amazônia, Coordenação de Pesquisas em Ecologia, Avenida EWgênio Sales, 2239, Manaus, CP 478, CEP 69011–970, Brazile-mail: [email protected]
1 C

2702 Biodivers Conserv (2008) 17:2701–2712
amostrados em trinta parcelas de 0,25 £ 250 m em duas ocasiões. A precipitação em curtoprazo foi representada pela chuva acumulada em três dias antes da parcela ser amostrada.As parcelas foram classiWcadas em dois grupos baseados na precipitação acumulada. Oconteúdo de argila no solo e a precipitação inXuenciaram a riqueza de morfoespécies, masas respostas aos fatores edáWcos dependeram da precipitação. Períodos mais chuvososaparentemente diminuíram condições limitantes de umidade nas áreas mais elevadas,permitindo a atividade e produção de corpos de frutiWcação pelos fungos. A composição demorfoespécies foi inXuenciada pelo conteúdo de argila, mas provavelmente a inXuênciasobre os fungos foi indireta, dado que o conteúdo de argila esteve correlacionado com alti-tude, comunidade de plantas e disponibilidade de nitrogênio. Nossos resultados sugeremque as espécies de fungos de liteira não estão distribuídas aleatoriamente na paisagem.Além disso, indicam que conduzir avaliações da diversidade de fungos em mesoescala éviável, desde que a variação temporal e espacial, e sua interação, sejam consideradas.
Keywords Beta diversity · Biodiversity inventories · Community ecology · Environmental gradients · Fruiting body production · Litter decomposition ·Mesoscale · Nutrient cycling · Patterns of distribution · Tropical fungi
Abbreviations INPA (Instituto Nacional de the Portuguese acronym for National Institute of
Pesquisas da Amazônia) Amazonian ResearchRFAD (Reserva Florestal the Portuguese acronym for Adolpho Ducke
Adolpho Ducke) Forest ReserveRAPELD Term coined by Magnusson et al. (2005) applied to
their sampling methodology, useful both forrapid access to biodiversity and long-termresearch
PELD (Pesquisas Ecológicas de the Portuguese acronym for Long Term EcologicalLonga Duração) Research Program
PPBio (Programa de Pesquisas the Portuguese acronym for Biodiversity Researchem Biodiversidade) Program
COL1 First collection occasion for all plots, from 29 Juneto 18 September 2005
COL2 Second collection occasion for all plots, from 20September to 22 January 2006
PCoA Principal Coordinates AnalysisMR Morphospecies richness, given by the total number
of morphospecies collectedMC Morphospecies composition, based on the incidence
of morphospecies in each plotCPCRH (Coordenação de Pesquisas the Portuguese acronym for Climate and Water
em Clima e Recursos Hídricos) Resources Research Coordination
Introduction
Fungi are important components of tropical systems (Hawksworth and Colwell 1992), andare responsible for most nutrient and carbon cycling (Swift 1982). Saprotrophic litter fungicontribute substantially to these processes (Hedger 1985) because Wne litter production
1 C

Biodivers Conserv (2008) 17:2701–2712 2703
represents the bulk of the input of biomass to the decomposition system in tropical forests(Luizão 1989). Resource distribution in time and space strongly inXuence the activity of lit-ter fungi, as diVerent species have diVerent degrees of resource selectivity, either taxon-selectivity or component-selectivity (Hedger 1985; Rayner et al. 1985). The resource repre-sents both the substratum and the source of organic nutrients for these fungi, and its dura-bility aVects directly mycelial longevity (Rayner et al. 1985). The fast rate ofdecomposition of Wne litter on the ground in tropical forests (Swift et al. 1979; Hedger1985) necessitates rapid colonization by fungi for capture of resources and subsequently,rapid exit during the colonization of newer resources.
Topography and soil properties inXuence the spatial distribution of plant communities inCentral Amazonia (Costa et al. 2005; Kinupp and Magnusson 2005). However, there is lit-tle information on how those aVect the distribution of fungi in the landscape. Most speciesof fungi in the Agaricales are believed to have restricted spatial distributions (Lodge et al.1995), but owing to the small scale of sampling and the lack of standardized methodsamong studies, there is still little understanding of how the diversity is spatially organizedat diVerent scales. Lodge et al. (1995) concluded that the diversity of saprobic fungi ishigher in areas with low latitudes, medium to low altitudes, moderate to high rainfall, highhabitat diversity, and to a lesser extent in areas with higher host diversity and resourceabundance. These characteristics are widely present in Amazonia, leading to an expectationof high fungal diversity in evergreen tropical forests, independent of the site. However, thescale of analysis of Lodge et al. (1995) was several times greater than scales where conser-vation decisions usually take place, and it is diYcult to interpolate their conclusions todetermine whether the diversity of fungi is non-randomly distributed in the landscape atsmaller scales. Also, although mean annual rainfall may be high in Amazonia, lack of rain-fall may limit fungus fruiting body production in dryer years, which may occur due to thewarming of PaciWc or Atlantic Ocean superWcial waters (Fearnside 2006).
Although the occurrence of fruiting bodies does not necessarily reXect the activity anddistribution of mycelia (Rayner et al. 1985), the observation and collection of fruiting bod-ies have been used to determine macrofungal species occurrence in space and time (Vogtet al. 1992; Lodge and Cantrell 1995; Lodge et al. 2004; O’Dell et al. 2004). However,temporal variation in the detectability of litter fungi sporocarps confuses the perception ofspatial variation in the occurrence of species. One of the impediments to landscape studiesof fungi is the diYculty of identifying species in the Weld, even when fruiting bodies arepresent. Microscopic examination is necessary to separate many species, especially those inTricholomataceae (Singer 1986), and speciWc determination of large collections by special-ists may take years or decades. These problems are exacerbated in tropical areas, wheremany species may be undescribed. However, given the importance of fungi to ecosystemprocesses, these problems should not be used to exclude fungi from biodiversity surveys.
Our preliminary surveys showed that about 70 % of species in a tropical forest in Cen-tral Amazonia can be sorted to morphospecies based only on macroscopic characteristics,and these species represent the majority (about 90%) of sporocarps encountered. Currently,these materials are identiWed only by herbarium accession numbers, but a photographicguide to morphospecies (Braga-Neto 2007) permits comparison with other sites. We inves-tigated whether temporal variation in rainfall and spatial variation associated with soil andtopography were correlated with the distribution of fruiting bodies of common species oflitter decomposer fungi over an area of 25 km2 in evergreen tropical forest in CentralAmazonia during a dry year.
1 C

2704 Biodivers Conserv (2008) 17:2701–2712
Study site
The study was conducted at Reserva Florestal Adolpho Ducke (RFAD), located on the out-skirts of Manaus, Brazil (02°55� S 59°59� W; Fig. 1). The reserve protects about 10,000 ha(10 £ 10 km) of terra Wrme forests, but is in contact with urban areas on the southern andwestern borders. The highly leached RFAD soils belong to the Alter do Chão geologicalformation (Chauvel et al. 1987; Sombroek 2000). The topography is variable, intermin-gling ridge tops, slopes and lowlands, which may be Xooded partially after intense rainfall,especially during the wet season. The altitude varies from 40 to 110 m above sea level. InRFAD, variations of soil texture are strongly associated with topography (Chauvel et al.1987; Mertens 2004). Rainfall is concentrated between November and June, with highestprecipitation in March and April. There are about three months with less than 100 mm ofrainfall, between July and September (Marques Filho et al. 1981). The annual rainfall is2436 § 332 mm (mean § standard deviation), with amplitude of 1300–2900 mm (from1975 to 2004). Mean temperature is about 26°C, and varies little through the year. The dryseason in 2005 had little rainfall due to the warming of Atlantic Ocean superWcial waters(Fearnside 2006; Trenberth and Shea 2006).
Methods
Sampling design
Following the RAPELD methodology (Magnusson et al. 2005), we used thirty0.25 £ 250 m plots, maintained by the Brazilian Long Term Ecological Research Program(PELD) and the Program for Biodiversity Research (PPBio) at RFAD. Plots are orientatedalong the topographic contours, which reduces the variation in factors correlated withaltitude, such as soils (Chauvel et al. 1987; Mertens 2004). The 30 plots are distributed on agrid system of 5 £ 5 km, and plots were at least one kilometer apart. The utilization of longthin plots increases the number of species registered in comparison with square or circularplots of the same area (Krebs 1998; Cantrell 2004).
Fig. 1 Location of Reserva Florestal Adolpho Ducke (RFAD, 02°55� S 59°59� W) and the grid system insidethe reserve. Dots on the trail system indicate the location of permanent plots. Lines represent topographic con-tours, ranging from 40 to 110 m above sea level; darker lines indicate higher areas
1 C

Biodivers Conserv (2008) 17:2701–2712 2705
Data collection
Fungi
The fungi were collected between June 2005 and January 2006. Each of the 30 plots wasvisited twice. The Wrst collection (COL1) occurred from 29 June to 18 September 2005,and the second (COL2), from 20 September to 22 January 2006. The number of daysbetween sampling occasions for each plot was 109 § 17.3 (mean § standard deviation).Marasmioid, crepidotoid and collybioid fungi connected to Wne litter components weresampled. Only fruiting bodies visible at the surface were collected; the litter was not turnedover. The specimens were photographed in situ whenever feasible. Fruiting bodies weremeasured and described macroscopically, and subsequently dried. For most specimens,photographs and macroscopic data were available for each sample. Specimens were classi-Wed into morphospecies. The descriptions were used to construct a preliminary guide tomorphospecies of litter fungi at RFAD (Braga-Neto 2007). Some species of fungi cannotbe distinguished macroscopically, and only data for morphospecies that could be consis-tently grouped on macroscopic characters (about 70%) were used in the analyses presentedhere. All fruiting bodies collected were deposited in the INPA Herbarium.
Environmental variables
Rainfall data were collected at the RFAD Meteorological Station, which is located outsideof the grid system, between June 2005 and January 2006. Altitude and soil variables datawere obtained from the PPBio database. Variations in soil texture are strongly associatedwith topography in the study region (Chauvel et al. 1987). Within the RFAD, as elsewherein Central Amazonia (Luizão et al. 2004), clay content was strongly correlated with altitude(r = 0.977) and sand content (r = ¡0.999), such that clay predominated in higher areas andsand in the lowlands. Moreover, clay content predicted much of the spatial variation in thecommunities of herbs (Costa et al. 2005) and shrubs (Kinupp and Magnusson 2005). Thus,clay content was chosen to represent the correlated edaphic variables, but the inXuence onfungi may be indirect.
Data analysis
We tested possible correlation in fungus composition between the two sampling periods andspatial auto-correlation of morphospecies richness and composition (Legendre and Legen-dre 1998). Mantel tests were used to evaluate spatial auto-correlation, using the Euclideandistance for richness and spatial distance and the Sørensen index for community composi-tion (incidence data). Additionally, we tested the spatial auto-correlation of clay content insoil using the Euclidean distances of standardized clay data against the spatial distance.
Rainfall was represented by the cumulative rainfall in the three days before each plotwas surveyed, a period assumed to allow fruit body production in response to rainfall. Plotswere grouped in two rainfall categories. The plots with more than 10 mm of three-daycumulative rainfall were considered as high-rainfall collection plots (N = 15) and the onessampled after a three-day period in which cumulative rainfall was less than 10 mm wereconsidered as low-rainfall collection plots (N = 45). No plot classiWed in the high-rainfallgroup was repeated. However, in the low-rainfall group 15 plots were sampled twice, andone of the sampling occasions was randomly excluded for each of these plots to avoid tem-poral pseudoreplication. Hence, the same plot was not counted twice in either treatment.
1 C
2706 Biodivers Conserv (2008) 17:2701–2712
Richness (number of morphospecies) and morphospecies composition were used indiVerent analyses to investigate the distribution of the litter fungus community. The multi-dimensionality of the community composition was reduced by principal coordinates analy-sis (PCoA) based on incidence data (presence or absence of morphospecies in each plot),using the Sørensen index as a community composition distance measure. Gotelli and Elli-son (2004) recommend the use of PCoA when the objective is to preserve the original mul-tivariate distances between observations in the reduced space, especially in analyses ofpresence-absence data. The cumulative variance explained by the Wrst three axes wasabout 51% for plots surveyed in low-rainfall periods, and about 57% for plots surveyed inhigh-rainfall periods. We excluded three plots with zero morphospecies in the compositionanalysis.
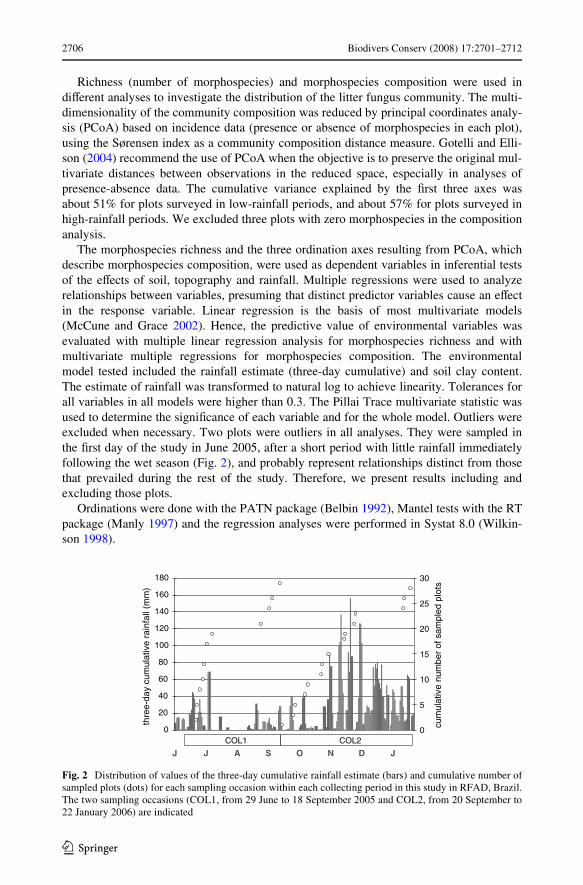
The morphospecies richness and the three ordination axes resulting from PCoA, whichdescribe morphospecies composition, were used as dependent variables in inferential testsof the eVects of soil, topography and rainfall. Multiple regressions were used to analyzerelationships between variables, presuming that distinct predictor variables cause an eVectin the response variable. Linear regression is the basis of most multivariate models(McCune and Grace 2002). Hence, the predictive value of environmental variables wasevaluated with multiple linear regression analysis for morphospecies richness and withmultivariate multiple regressions for morphospecies composition. The environmentalmodel tested included the rainfall estimate (three-day cumulative) and soil clay content.The estimate of rainfall was transformed to natural log to achieve linearity. Tolerances forall variables in all models were higher than 0.3. The Pillai Trace multivariate statistic wasused to determine the signiWcance of each variable and for the whole model. Outliers wereexcluded when necessary. Two plots were outliers in all analyses. They were sampled inthe Wrst day of the study in June 2005, after a short period with little rainfall immediatelyfollowing the wet season (Fig. 2), and probably represent relationships distinct from thosethat prevailed during the rest of the study. Therefore, we present results including andexcluding those plots.
Ordinations were done with the PATN package (Belbin 1992), Mantel tests with the RTpackage (Manly 1997) and the regression analyses were performed in Systat 8.0 (Wilkin-son 1998).
Fig. 2 Distribution of values of the three-day cumulative rainfall estimate (bars) and cumulative number ofsampled plots (dots) for each sampling occasion within each collecting period in this study in RFAD, Brazil.The two sampling occasions (COL1, from 29 June to 18 September 2005 and COL2, from 20 September to22 January 2006) are indicated
0
20
40
60
80
100
120
140
160
180
J J A S O N D J
thre
e-da
y cu
mul
ativ
e ra
infa
ll (m
m)
0
5
10
15
20
25
30
cum
ulat
ive
num
ber
of s
ampl
ed p
lots
COL1 COL2
1 C

Biodivers Conserv (2008) 17:2701–2712 2707
Results
Sampling eVort and rainfall pattern
Most plots were sampled in the dry and early wet seasons of 2005 (Fig. 2). A total of 1,193fruit bodies were recorded and classiWed into 87 morphospecies. During the Wrst samplingperiod (COL1), 635 fruit bodies were assigned to 61 morphospecies. In the second period(COL2), 558 fruit bodies were classiWed into 58 morphospecies. There was no diVerenceamong sampling periods for the observed number of morphospecies (t = 1.290, P = 0.203)or the number of fruiting bodies (t = 0.363, P = 0.718). Only 32 morphospecies were foundin both sampling periods.
Morphospecies richness was not correlated among sampling occasions (r = 0.061,P = 0.747), nor was it spatially auto-correlated. Morphospecies composition was not spa-tially auto-correlated, but it was weakly correlated between sampling occasions (r = 0.176,P = 0.001), implying non-independence in the detection of fruiting bodies of most morpho-species between sampling occasions. Mantel tests revealed spatial auto-correlation for claycontent (r = 0.115, P = 0.017) when all 30 plots are considered in the low-rainfall group,but the relationship is so weak that it does not inXuence the interpretation of results.
Responses of litter fungi community to rainfall and clay content
For collections made in the low-rainfall periods (three-day cumulative rainfall <10 mm),morphospecies richness (MR) was related to the predictor variables (MR = constant +rainfall + clay, F2, 24 = 8.250, P = 0.002, R2 = 0.358, N = 27); both rainfall (P = 0.002) andclay content (P = 0.040) contributed signiWcantly to the model. Fewer morphospecies werefound in dry periods with less rainfall (Fig. 3a) and in collections on clay soils (Fig. 3b).For collections made in the high-rainfall periods (three-day cumulative rainfall > 10 mm),MR was also signiWcantly inXuenced by the model (MR = constant + rainfall + clay,F2, 11 = 4.456, P = 0.038, R2 = 0.347, N = 14). Rainfall (P = 0.046) contributed signiW-cantly to the model (Fig. 3c), but clay content did not (P = 0.198, Fig. 3d).
Clay content did not contribute signiWcantly to the model (P = 0.076) for low-rainfallgroup when all the plots, which were outliers were included (MR = constant +rainfall + clay, F2, 27 = 3.129, P = 0.060, R2 = 0.128, N = 30). The outlying plots all hadmuch higher morphospecies richness than expected for plots collected in low-rainfallperiods. Two were sampled immediately following the rainy season, which generally endsin May–June in the region, so soil and leaf-litter water potentials were still high, despite thelack of rainfall in the previous three days. Excluding these plots, the relationship betweenclay content and morphospecies richness was signiWcant for plots collected in the low-rain-fall periods.
For collections made in low-rainfall periods, morphospecies composition (MC) wasinXuenced by the model (MC = constant + rainfall + clay, F6, 44 = 3.699, P = 0.005,R2 = 0.267, N = 26), but only clay content (P < 0.001) contributed to the model; there wasno evidence for an eVect of rainfall (P = 0.678). There was also some evidence for an eVectof the variables in collections in high-rainfall periods (MC = constant + rainfall + clay, F6,
22 = 2.474, P = 0.056, R2 = 0.301, N = 15) and the pattern was similar to that in the dryperiods, with low probability for the null hypothesis associated with clay (P = 0.051) andno evidence for an eVect of rainfall (P = 0.223).
1 C

2708 Biodivers Conserv (2008) 17:2701–2712
Discussion
The community of litter fungi seemed to be spatially structured over 25 km2 along rainfalland edaphic gradients. Our results indicated that litter fungal fruiting body production in atropical forest varied predictably in space and time, suggesting that litter fungi species arenot randomly distributed at mesoscales. Rainfall and clay content in soil predict morpho-species richness and composition, but responses of the number of morphospecies toedaphic factors depended on rainfall. Relationships between morphospecies richness andsoil texture were not constant along the rainfall gradient. Clay content, which was corre-lated with topographic position, inXuenced morphospecies richness only in low-rainfallperiods, while in the high-rainfall periods richness was not related to soil characteristics.
Rainfall is a primary determinant of the availability of water in forest ecosystems, butthe gradient of moisture in the RFAD landscape seemed to depend on drainage patterns,which is mainly determined by topography. Low clay content is associated with lowlandsand proximity to watercourses in the study site (Chauvel et al. 1987). These areas are rela-tively wetter, especially in the driest months of the year. This trend may favor mycelialactivity, fruiting body production, and the chance of observation of species. The absorptivenutrition of fungi implies dependence on substrate moisture conditions. Daily and seasonalvariances are determinants of litter fungi mycelial activity and fruiting body production intropical forests (Hedger 1985). Though optimal moisture conditions vary among species,litter fungal fruiting body production is expected to be lower in periods with low humidity(Singer and Araujo 1979).
The absence of a signiWcant correlation between the number of morphospecies and claycontent for the high-rainfall periods may be related to a decrease in topographic inXuence
Fig. 3 Partial plots of the eVects of rainfall and clay content on morphospecies richness for low-rainfall(three-day cumulative rainfall <10 mm) periods (a, b), and for high-rainfall (three-day cumulative rainfall >10 mm) periods (c, d)
-50 -40 -30 -20 -10 0 10 20 30 40 50
-50 -40 -30 -20 -10 0 10 20 30 40 50
-4-3-2-101234
-1.0 -0.5 0.0 0.5 1.0 1.5-3
-2
-1
0
1
2
3
4
-1.0 -0.5 0.0 0.5 1.0-5-4-3-2-10123
-3
-2
-1
0
1
2
3
Low-rainfallPeriod
(< 10 mm)
High-rainfallPeriod
(> 10 mm) Mor
psoh
cepie
sci
Renhss
trap(i
)la
Log(Rainfall)(partial)
Clay Content(partial)
a b
dc
1 C

Biodivers Conserv (2008) 17:2701–2712 2709
over the moisture gradient, presumably caused by less limiting moisture conditions drivenby higher rainfall. On the other hand, rainfall inXuences on morphospecies richness wereconsistent among rainfall groups. We found an expected positive relation between morpho-species richness and three-day cumulative rainfall for both rainfall groups, since litter fun-gal fruit body production depends on short-term rainfall (Singer and Araujo 1979).However, the magnitude of the eVect is likely to be related to the fact that we sampled dur-ing a relative dry year, and probably the eVects are weaker in wetter years.
Soil texture inXuences on litter fungus community composition
Morphospecies composition was inXuenced by clay content in soil within the low-rainfallgroup, and approached signiWcance for the high-rainfall plots. These results suggest thatfruit body production by the litter fungus community is spatially structured along edaphicgradients over 25 km2, although there is no evidence of these eVects on the mycelial distri-bution. Given the strong correlation with altitude (Chauvel et al. 1987), clay content seemsto reXect the landscape moisture gradient, hence inXuencing the distribution of litter fungusmorphospecies, as species are supposed to diVer in their abilities to deal with moisture(Hedger 1985). Moreover, clay content inXuences plant community spatial distribution(Costa et al. 2005; Kinupp and Magnusson 2005), is positively correlated with leaf litter Nconcentrations (Luizão et al. 2004) and is supposed to be negatively associated with ecto-mycorrhizal fungi (Singer 1978; Singer and Araujo 1979) in Central Amazonia.
The inXuence of plant community composition on the spatial distribution of litter fungiis often considered to be high, due to taxon selectivity (Singer 1976; Cornejo et al. 1994;Lodge et al. 1995; Polishook et al. 1996), although quantitative data on this issue for basid-iomycete fungi are scarce. Santana et al. (2005) found that matching of decomposer fungito the leaf substratum by the host plant family or chemical/physical traits of the sourceplant in which they were dominants increased rates of decomposition. At large scales, spa-tial distribution patterns of Marasmius spp. and Marasmiellus spp. fungi do coincide withpatterns of phanerogam plants (Redhead 1989). In RFAD, plants are not randomly distrib-uted in space, but spatially structured by topographic and edaphic factors (Costa et al.2005; Kinupp and Magnusson 2005). As litter fungal species possess some degree of taxonselectivity (Singer 1976), one expects that they are inXuenced by the distribution of theirhosts. Additionally, litter fungi species are expected to have diVerent demands for resourcenutrient composition (Hedger 1985). The litter layer N concentration may be lower in thevalley plots, where soils have low clay content, as elsewhere in Central Amazonia (Luizãoet al. 2004). However, leaf litter nutrient concentrations may be related to soil nutrient sta-tus rather than topography (Wood et al. 2006).
In a nearby site, Singer and Araujo (1979) observed more species of litter fungi on claysoils, attributing the pattern to negative inXuence of competitive ectomycorrhizal fungi insandy soils (Singer 1978). However, we found the opposite pattern. The number of mor-phospecies of litter fungi detected was higher in sandy sites. Besides the diYculties in thecomparison of results due to diVerent sampling designs, drier environmental conditionsmay have limited the detectability of litter fungi species in sites with higher clay contentduring our study.
The correlation of morphospecies composition between sampling occasions may reXectseasonality in fruiting body production, and implies non-independence in the detection offruiting bodies of most morphospecies between sampling occasions. Non-independence isa problem with repeated measures analysis, because time series are likely to be correlated,such that current values are a function of values observed in the past (Gotelli and Ellison
1 C

2710 Biodivers Conserv (2008) 17:2701–2712
2004). However, temporal correlation between samples does not mean one cannot incorpo-rate time series data into conventional statistical analyses. Temporal replicate sampling isvery appropriate to study how ecological systems change trough time, and many conserva-tion models designed to predict future conditions are derived more reliably from replicateddata in time (Ferraz et al. 2007). Some alternatives to repeated measures analysis may beespecially useful to avoid pseudoreplication problems (Gotelli and Ellison 2004). Ourapproach to avoid problems related to lack of independence of the data and to maximizedata use was to classify all plots collected in two groups (based on 10 mm of three-daycumulative rainfall), not to count the plot twice in either group, and to analyze the dataseparately.
The further apart in time the samples are separated from one another the more likelythey function as independent replicates (Gotelli and Ellison 2004). In our study, the numberof days between the sampling occasions for each plot was similar, although it was not suY-cient to guarantee independence. Future sampling designs on leaf litter fungus shouldconsider the interval between samples of this study (about four months), aiming to avoidtemporal correlation among samples. This information may be extremely useful for biodi-versity inventory planning, as it may be possible to take into account diVerent speciesdetectabilities while reducing sampling costs in time and Wnancial resources.
Caveats and implications for fungal conservation
Although no diVerences in abundance or morphospecies richness between the Wrst and sec-ond sampling periods were found, the low proportion of shared morphospecies suggeststhat an increase in rainfall in the early wet season of 2005 triggered fruiting body produc-tion of diVerent species, an indication of diVerential responses to seasonal/moistureregimes. However, most morphospecies were infrequent or rare, and the detectability ofmost litter fungal species seems to be low. Repeated surveys of the same plots will beneeded to detect most species.
This study has shown that rainfall, probably through its eVect on litter water potential,can strongly aVect apparent habitat speciWcity in fungi. Although we showed only temporaleVects associated with an uncommonly dry year (Fearnside 2006), spatial eVects in areaswith lower rainfall, typical of much of the Amazon basin, are likely. Therefore, it is impor-tant that studies of spatial variation in fruiting body production take into account the tempo-ral variation caused by weather. However, fungi are likely to be responding to waterpotential in the litter rather than rainfall per se. The distribution of plots in mesoscale(25 km2) incorporates more environmental heterogeneity than sampling at smaller scalesand it is important to improve the quality of moisture estimates in space, either by directlymeasuring moisture in leaf litter or by including more rainfall collecting stations within thegrid system.
SigniWcant eVorts have been made to improve global fungal diversity estimates (Hawks-worth 1991, 2001), incorporating available data on biogeographic distributions, levels ofendemism and host speciWcity (Schmit et al. 2005; Mueller et al. 2006; Schmit and Mueller2006). However, scarcity of available information in tropical regions, especially in Amazo-nia, will preclude the test of these estimates and fungal conservation planning as long aslarge-scale inventories are not accomplished. Most of the unknown fungal diversity isexpected to occur in the tropics (Lodge et al. 1995; Hawksworth and Rossman 1997;Hawksworth 2001), and most species distribution ranges are believed to be small (Lodgeet al. 1995; Mueller et al. 2006). Lack of methodological standardization between studiesrestricts the understanding of fungal diversity spatial structure at scales important for
1 C

Biodivers Conserv (2008) 17:2701–2712 2711
biological conservation. Both large and small scale datasets are needed to assess fungalspecies spatial distributions and levels of endemism (Mueller et al. 2006). Our results showthat it is viable to conduct mesoscale evaluations of fungal diversity, as long as the tempo-ral and spatial variation and their interaction are taken into account.
Acknowledgements Ricardo Braga-Neto acknowledges the support of a Brazilian National ResearchCouncil (CNPq) graduate scholarship. Brazilian Long Term Ecological Research (PELD) guaranteed Weldsupport and equipment acquisition. The European Network for Research in Global Change (ENRICH) pro-vided support during the preparation of this manuscript. Topographic and edaphic data were obtained fromPELD and PPBio (Programa de Pesquisas em Biodiversidade). Eleusa Barros, Tânia Pimentel and Jane Mer-tens contributed to soil data acquisition. Rainfall data were obtained from CPCRH/INPA (Coordenação dePesquisas em Clima e Recursos Hídricos). We thank Dennis Desjardin, Carla Puccinelli and Débora Druckerfor their contributions; and D. Jean Lodge, Roberto Garibay Orijel and two anonymous reviewers for revisingearlier versions of the manuscript.
References
Belbin L (1992) PATN: Pattern Analysis Package. CSIRO Division of Wildlife and Ecology, CanberraBraga-Neto R (2007) Guia de morfoespécies de fungos de liteira da Reserva Ducke. Programa de Pesquisas
em Biodiversidade (PPBio/INPA). Available from: http://ppbio.inpa.gov.br/Port/inventarios/guias/Guia_fungos_RFAD.pdf
Cantrell SA (2004) A comparison of two sampling strategies to assess discomycete diversity in wet tropicalforests. Caribb J Sci 40:8–16
Chauvel A, Lucas Y, Boulet R (1987) On the genesis of the soil mantle of the region of Manaus, CentralAmazonia, Brazil. Experientia 43:234–241
Cornejo FH, Varela A, Wright SJ (1994) Tropical forest litter decomposition under seasonal drought: nutrientrelease, fungi and bacteria. Oikos 70:183–190
Costa FRC, Magnusson WE, Luizão RCC (2005) Mesoscale distribution patterns of Amazonian understoreyherbs in relation to topography, soil and watersheds. J Ecol 93:863–878
Fearnside PM (2006) A vazante na Amazônia e o aquecimento global. Revista Ciência Hoje 39:76–78Ferraz G, Nichols JD, Hines JE, StouVer PC, Bierregaard Jr. RO, Lovejoy TE (2007) A large-scale defores-
tation experiment: eVects of patch area and isolation on Amazon birds. Science 315:238–241Gotelli NJ, Ellison AM (2004) A primer of ecological Statistics. Sinauer Associates, Inc., MassachusettsHawksworth DL (1991) The fungal dimension of biodiversity: magnitude, signiWcance, and conservation.
Mycol Res 95:641–655Hawksworth DL (2001) The magnitude of fungal diversity: the 1.5 million species estimate revisited. Mycol
Res 105:1422–1432Hawksworth DL, Colwell RR (1992) Microbial Diversity: biodiversity amongst micro-organisms and its rel-
evance. Biodivers Conserv 1:221–226Hawksworth DL, Rossman AY (1997) Where are all the undescribed fungi? Phytopathology 87:888–891Hedger J (1985) Tropical agarics: resource relations and fruiting periodicity. In: Moore D, Casselton LA,
Wood DA, Frankland JC (eds) Developmental biology of higher fungi. Cambridge University Press,Cambridge
Kinupp VF, Magnusson WE (2005) Spatial patterns in the understorey shrub genus Psychotria in CentralAmazonia: eVects of distance and topography. J Trop Ecol 21:363–374
Krebs CJ (1998) Ecological methodology. Benjamin Cummings, New YorkLegendre P, Legendre L (1998) Numerical ecology. Elsevier Science BV, AmsterdamLodge DJ, Cantrell S (1995) Fungal communities in wet tropical forests: variation in time and space. Can J
Bot 73: S1391–S1398Lodge DJ, Chapela I, Samules G et al. (1995) A survey of patterns of diversity in non-lichenized fungi. Mitt-
eilungen der Eidgenössischen Forschungsanstalt für Wald, Schnee und Landschaft 70:157–173Lodge DJ, Ammirati JF, O’Dell TE, Mueller GM (2004) Collecting and describing macrofungi. In: Mueller
GM, Bills GF, Foster MS (eds) Biodiversity of fungi: Inventory and monitoring methods. Elsevier Aca-demic Press, San Diego
Luizão FJ (1989) Litter production and mineral element input to the forest Xoor in a Central AmazonianForest. GeoJournal 19:407–417
1 C

2712 Biodivers Conserv (2008) 17:2701–2712
Luizão RCC, Luizão FJ, Paiva RQ, Monteiro TF, Sousa L, Kruijt B (2004) Variation of carbon and nitrogencycling processes along a topographic gradient in a Central Amazonian forest. Global Change Biol10:592–600
Magnusson WE, Lima AP, Luizão RCC et al. (2005) RAPELD: uma modiWcação do método de Gentrypara inventários de biodiversidade em sítios para pesquisa ecológica de longa duração. BiotaNeotropica 5:19–24. Available from: http://www.scielo.br/scielo.php?script=sci_arttext&pid=S1676–06032005000300002&lng=en&nrm=iso
Manly BFJ (1997) Randomization, bootstrap and Monte-Carlo methods in biology. Chapman and Hall,London
Marques-Filho AO, Ribeiro MNG, Santos JM (1981) Estudos climatológicos da Reserva Florestal Ducke,Manaus, AM. IV - Precipitação. Acta Amazonica 4:759–768
McCune B, Grace JB (2002) Analysis of Ecological Communities. MjM Software Design, OregonMertens J (2004) The characterization of selected physical and chemical soil properties of the surface soil lay-
er in the “Reserva Ducke”, Manaus, Brazil, with emphasis on their spatial distribution. Dissertation,Humboldt-Universität Zu Berlin
Mueller GM, Schmit JP, Leacock PR, Buyck B, Cifuentes J, Desjardin DE, Halling RE, Hjorstam K, Itur-riaga T, Larsson K-H, Lodge DJ, May TW, Minter D, Rajchenberg M, Redhead SA, Ryvarden L,Trappe JM, Watling R, Wu Q-X (2006) Global diversity and distribution of macrofungi. BiodiversConserv 16:37–48
O’Dell TE, Lodge DJ, Mueller GM (2004) Approaches to sampling macrofungi. In: Mueller GM, Bills GF,Foster MS (eds) Biodiversity of fungi: Inventory and monitoring methods. Elsevier Academic Press,San Diego
Polishook JD, Bills GF, Lodge DJ (1996) Microfungi from decaying leaves of two rain forest trees in PuertoRico. J Indust Microbiol 17:284–294
Rayner ADM, Watling R, Frankland JC (1985) Resource relations – an overview. In: Moore D, CasseltonLA, Wood DA, Frankland JC (eds). Developmental biology of higher fungi. Cambridge UniversityPress, Cambridge
Redhead SA (1989) A biogeographical overview of the Canadian mushroom Xora. Can J Bot 67:3003–3062Santana M, Lodge DJ, Lebow P (2005) Relationships of host recurrence in fungi to rates of tropical leaf
decomposition. Pedobiologia 49:549–564Schmit JP, Mueller GM, Leacock PR, Mata JL, Wu Q-X, Huang Y-Q (2005) Assessment of tree species rich-
ness as a surrogate for macrofungal species richness. Biol Conserv 121:99–110Schmit JP, Mueller GM (2006) An estimate of the lower limit of global fungal diversity. Biodivers Conserv
16:99–111Singer R (1976) A Monograph of the Neotropical species of the Marasmieae (excepting the Oudemansielli-
nae), Basidiomycetes–Tricholomataceae. Flora Neotropica 17:1–347Singer R (1978) Origins of the deWciency of Amazonian soils––a new approach. Acta Amazonica 8:315–316Singer R (1986) The Agaricales in modern taxonomy, 4th edn. Koeltz ScientiWc Books, KoenigsteinSinger R, Araujo IJS (1979). A comparison of litter decomposing and ectomycorrhizal Basidiomycetes in
latosol-terra-Wrme rain forest and white podzol campinarana. Acta Amazonica 9:25–41Sombroek WG (2000) Amazon land forms and soils in relation to biological diversity. Acta Amazonica
30:81–100Swift MJ, Heal OW, Anderson JM (1979) Decomposition in terrestrial ecosystems. Oxford Blackwell Publi-
cations, OxfordSwift MJ (1982) Basidiomycetes as components of forest ecosystems. In: Frankland JC, Hedger J, Swift MJ
(eds). Decomposer basidiomycetes: their biology and ecology. Cambridge University Press, CambridgeTrenberth KE, Shea DJ (2006) Atlantic hurricanes and natural variability in 2005. Geophysical Research
Letters 33:L120704Vogt KA, BloomWeld J, Ammirati JF, Ammirati SR (1992) Sporocarp production by Basidiomycetes, with
emphasis on forest ecosystems. In: Carroll GC, Wicklow DT (eds) The fungal community, 2nd edn.Marcel Dekker Inc, New York
Wilkinson L (1998). Systat, Version 8.0. SPSS Inc., ChicagoWood TA, Lawrence D, Clark DA (2006) Determinants of leaf litter nutrient cycling in a tropical rain forest:
soil fertility versus topography. Ecosystems 9:1–11
1 C









![Flowering and Fruiting Characteristics of Eight Pawpaw [Asimina triloba (L.) Dunal] Selections in Kentucky](https://static.fdokumen.com/doc/165x107/6338663121271f736b017418/flowering-and-fruiting-characteristics-of-eight-pawpaw-asimina-triloba-l-dunal.jpg)










