
Towards an Ecological Understanding of Dinoflagellate Cyst Functions
Upload
independentCategory
view
0download
0
Review of Palaeobotany and Palynology 158 (2009) 52–71
Contents lists available at ScienceDirect
Review of Palaeobotany and Palynology
j ourna l homepage: www.e lsev ie r.com/ locate / revpa lbo
Last Glacial to Holocene hydrology of the Marmara Sea inferred from a dinoflagellatecyst record
Laurent Londeix a,⁎, Yannick Herreyre a, Jean-Louis Turon a, William Fletcher a,b
a Université Bordeaux 1, UMR 5805 CNRS ‘EPOC’, avenue des Facultés, F-33405 Talence Cedex, Franceb Institut für Geowissenschaften, Goethe Universität, Frankfurt am Main, Germany
⁎ Corresponding author. Fax: +33 556 84 08 48.E-mail address: [email protected] (L. Lo
0034-6667/$ – see front matter © 2009 Elsevier B.V. Adoi:10.1016/j.revpalbo.2009.07.004
a b s t r a c t
a r t i c l e i n f oArticle history:Received 22 January 2009Received in revised form 17 July 2009Accepted 20 July 2009Available online 25 July 2009
Keywords:organic-walled dinoflagellate cystslate QuaternarypalaeohydrologyMarmara SeaBlack Sea connectionFennoscandian Ice Sheet deglaciationM1 sapropelHeinrich Event 1.
Thehistoryof the connectionbetween theMediterranean andBlack seas since theLastGlacialMaximum(LGM) isdocumented through the study of an organic-walled dinoflagellate cysts (dinocysts) record at high temporalresolution from theMarmara Sea (MD01-2430 core). LateGlacial dinocyst assemblages indicate that theMarmaraSea was a mildly brackish (caspibrackish) lake with mean annual sea-surface salinities (SSS) around 7–14 psuwhich probably experienced low seasonal contrasts in SSS; winter and summer sea-surface temperature (SST)were probably around 3–5 and 10–13 °C, respectively. This climatic and hydrological context remained effectivelyunchanged up to 20.5 cal. ka BP. Prior to the end of the LGM, a clearwarming preceding a strongmeltwater influxhas been observed for the first time in the Marmara Sea as early as ~20.5 cal. ka BP. The meltwater episodepresents a two-step history: 1) aweak phase (~20.0 to 18.1 cal. ka BP)which corresponds to a possiblemelting ofhinterland permafrost; and 2) a strong phase with important meltwater inflow originating from melting of theFennoscandian Ice Sheet and arriving via the Black Sea. Occuring from ~18.1 to 15.9 cal. ka BP, the second phaseimplies that Black Sea sea-level temporarily exceeded the Bosphorus sill at that time (ca Heinrich Event 1). Theintroduction of meltwater into the Marmara Sea caused very high seasonal contrasts in Marmara Sea surface-water salinity and temperature. During the Bølling/Allerød, the Marmara Sea was again a caspibrackish lakeexperiencing relatively cold conditions (with lowseasonal contrast inSST) and enhancedmoisture as recordedonthe continent. Inflowing Mediterranean waters filled the Marmara Sea bottom-waters ca 12.2 cal. ka BP(approximatively the beginning of the Younger Dryas), then reached the sea-surface at ~11.7 cal. ka BP. YoungerDryas SSTs were colder than during the Bølling/Allerød, with a likely enhancement of seasonal SST contrasts. Atthe beginningof theHolocene, SSTsmay have been slightly cooler than today,withminimumSSS ranging from13to 17.5 psu (maximum 27 psu) and likely with high seasonality in both SST and SSS (possibly up to 19 °C and17 psu). During the deposition of the Holocene sapropel M1, the Marmara Sea experienced enhanced primaryproductivity and dysoxic/anoxic conditions in the benthic zone. Dinocyst assemblages from theM1 time intervaldo not provide evidence of an enhanced outflow from the Black Sea. Following the connection of theMediterranean to the Black Sea (~9 cal. ka BP), several distinct climatic and hydrological phases occurred,including a short cooling (~7.5 to 5.8 cal. ka BP; possibly linked to a strengthening of the Siberian High [Kotthoff,U., Pross, J., Müller, U.C., Peyron, O., Schmiedl, G., Schulz, H., Bordon, A., 2008. Climate dynamics in the borderlandsof the Aegean Sea during formation of sapropel S1 deduced from a marine pollen record. Quaternary ScienceReviews, 27: 832–845.]), a thermal optimum (~5.8 to 4.7 cal. ka BP), and the installation of the modernhydrological system (~4.7 cal. ka BP). The timing of this sequence appears close to that recorded in the Black Sea,supporting the idea that the two basins would have operated in tandem.
© 2009 Elsevier B.V. All rights reserved.
1. Introduction
Sea-level rise during the last deglaciation led to a connectionbetween the Mediterranean and Black seas via the Marmara Sea (e.g.Nevesskii, 1967; Ross et al., 1970; Serebryanny, 1982; Pirazzoli, 1996;Ryan et al., 1997; Ryan and Pitman, 1999). The question of the precise
ndeix).
ll rights reserved.
nature and timing of this connection remains even today matter ofdebate, and has given rise to several competing hypotheses, including,for example, the ‘Flood Hypothesis’ (Ryan and Pitman, 1999) and the‘Outflow Hypothesis’ (Aksu et al., 1999).
According to the ‘Flood Hypothesis’, Mediterranean waters floodedcatastrophically into the depressed Black Sea basin when a presumedsediment dam in the Bosphorus channel was breached. Ryan et al. (1997)and Ryan and Pitman, (1999) have proposed a connection at ca. 7.5 ka ago(later corrected to 9.4 cal. ka BP; Ryan et al., 2003), and have claimed thatthis eventprovides thehistorical basis for thebiblical storyofNoah's Flood.

Fig.1. (A)Map of Europe showing present-daymajor rivers of the drainage basin of theMarmara, Black and Caspian seas, in relationwith reconstructed LGM ice coverage [after Ehlersand Gibbard (2004) and Svendssen et al. (2004)]. (B) Locationmap of theMarmara Sea and the southwestern Black Sea, showing the Bosphorus and Dardanelles straits with rivers ofthe present-day drainage area; position of the MD01-2430 and of the cores used for the age model are indicated by circles.
53L. Londeix et al. / Review of Palaeobotany and Palynology 158 (2009) 52–71
According to the ‘Outflow Hypothesis’, the Black Sea reached a −40 mbedrock sill depth in the Strait of Bosphorus first, initiating a downslopecascade into the risingMarmara Sea from10 to 9 14C ka1 [~11.2 to 10.0 cal.ka BP] and leading to the construction of an outflow delta at the southernexit of the strait (Hiscott et al., 2002; Aksu et al., 2002b). This latterhypothesis does not involve a catastrophic flood (Hiscott et al., 2007). Inorder to test these hypotheses, numerous studies have aimed toreconstruct the late Quaternary hydrological evolution of the MarmaraandBlack seas (seeMajoret al., 2006;Bahret al., 2006;Yanko-Hombachetal., 2007; Bahr et al., 2008 for reviews). Nevertheless, illuminating thishistory depends critically on knowledge on the evolution of the depths ofthe sills surrounding the Marmara Sea (Dardanelles and Bosphorus)during the last several millennia (e.g. Yaltirak et al., 2000; Yaltirak andAlpar, 2002; Koral, 2007).
Subsequent to the opening of the Mediterranean–Black Seaconnection, sapropel-like sediments were deposited in the MarmaraSea (Stanley and Blanpied,1980; Ünlüata and Özsoy,1986; Evans et al.,1989; Aksu et al., 1995; Lane-Serff et al., 1997; Çağatay et al., 1999,2000; Aksu et al., 2002b; Sperling et al., 2003). Among severalinterpretations (see Section 7.5) the associated temporary reductionin bottom-water ventilation has been considered to have resultedfrom runoff from the melting of the Northern European ice sheets
1 Conventional radiocarbon ages BP with no reservoir correction.
released 9.5–7 14C ka BP [~10.6 to 7.6 cal. ka BP] through the Black Sea(e.g. Ryan, 1972; Lane-Serff et al., 1997). These complex aspects ofpostglacial history make the Marmara Sea a remarkable area to studyboth the timing and nature of Fennoscandian Ice Sheet (FIS) decay andthe climatic evolution of the easternmost Mediterranean Sea.
The “Marmara Sea Gateway” (including the narrow straits of theDardanelles and Bosphorus, and the deep intervening Marmara Sea)provides a set of natural flow valves and sediment traps that inprinciple should contain a record of the re-connection of theMediterranean and Black seas (Aksu et al., 2002c). In tracing theevolution of surface-water conditions using quantitative analysis ofdinoflagellate cyst assemblages from a Marmara Sea core, the presentwork has the following two main objectives: to address the ‘flood‘versus ‘outflow’ hypotheses concerning the re-connection of the Blackand Mediterranean seas, and to decipher the timing and sequence ofevents involved in the demise of the FIS during the last deglaciationand the associated impact(s) on the hydrography of Black andMarmara Sea system.
2. Environmental setting
The Marmara Sea is an embanked semi-closed marine intraconti-nental basin of up to 1280 m water depth (Ünlüata et al., 1990),located in the eastern part of the Mediterranean region (Fig. 1A). Thebasin is situated in an area of strong seismic instability associated withstrain on the North Anatolian Fault (e.g. Gutzwiller, 1923; Le Pichon

54 L. Londeix et al. / Review of Palaeobotany and Palynology 158 (2009) 52–71
et al., 2001). The Marmara Sea is a transitional zone, connected todayto the Black Sea by the Istanbul Strait (Bosphorus) and to the AegeanSea by the Çanakkale Strait (Dardanelles) (Fig. 1B). These two straitshave sill depths of approximately 35–45 m and 55–75 m, respectively.The topographic restriction resulting from the two sills leads atpresent to the development of a steady two-layer flow system into theSea of Marmara. The cooler (5–15 °C), lower salinity (17–20 psu) BlackSea water forms a westward-flowing surface layer, 20–25 m thick(Ünlüata and Özsoy, 1986). Warmer (15–20 °C), saline (38–39 psu)Mediterranean water enters the Strait of Dardanelles beneath the lowsalinity layer in the northeastern Aegean Sea and flows northeast(Özsoy et al., 1995; Polat and Tuğrul, 1996) to occupy the entireMarmara basin below a pronounced halocline (Alavi, 1988; Taneret al., 2001). This stratification of the upper part of the water bodyhampers efficient circulation and, by preventing ventilation of thedeeper water column, promotes organic-matter preservation and thedevelopment of benthic communities adapted to suboxic to dysoxicconditions (0.1–1.3 mg/l O2) in the deep basins (Ünlüata and Özsoy,1986; Aksu et al., 2002c). Significant fluviatile input into the Sea ofMarmara comes only from the south via the three rivers Biga, Gönenand Kocasu, which are chiefly responsible for nutrient and allochto-nous organic-matter supply. As the hydrological contribution of theserivers is weak (~6 km3/year) compared with the ~300 km3/year BlackSea outflow (Özsoy et al., 1995), the estuarine circulation is dominatedby the latter. Mean sea-surface temperatures are around 5 °C and 26 °Cin winter and summer, respectively.
3. Previous studies on eastern Mediterranean dinocysts
Organic-walled dinoflagellate cysts (dinocysts) from the eastern-most Mediterranean region were previously studied by Wall et al.(1973), who noted the strong endemism of the cyst flora andproduced a working taxonomic framework for the region. Thisframework has been expanded by further studies in the Marmara,Black and Caspian seas by Eaton (1996), Mudie et al. (2001), Rochon etal. (2002) and Marret et al. (2004). The evolution of the dinocystassemblages from the ‘Paratethyan’ late Quaternary (i.e. recentMarmara, Black, Caspian and Aral seas) has been documented byWall and Dale (1974), Mudie et al. (2001, 2002a, 2004), Kouli et al.(2001), Popescu (2003), Atanassova (2005) and Marret et al. (inpress), but few studies deal with their modern repartition in surfacesediments (Mudie et al., 2002a; Marret et al., 2004). While severalstudies concern the ecology of the Quaternary ‘Paratethyan’ endemictaxa (Wall and Dale, 1974; Kouli et al., 2001; Mudie et al., 2001, 2002a;Rochon et al., 2002; Mudie et al., 2004; Marret et al., 2004) very fewhave yet attempted to correlate such assemblages of physico-chemicalconditions in surface-water masses in a quantitative manner (Mudieet al., 2001). Moreover, a high resolution palaeoclimatic sequencecovering the last 22 ka has not been yet studied on the basis ofdinocyst assemblages.
4. Materials and methods
4.1. Material
This study is based on IMAGES core MD01-2430 obtained duringthe NO Marion-Dufresne (Institut Polaire Français) cruise MD 123,Geosciences, Leg Marmacore (2001). This Calypso piston core wasretrieved in the western Marmara Sea at 40° 47.81′ N, 27° 43.51′ E at awater depth of 580 m (Fig. 1), and measures 28.9 m long.
The sequence studied here concerns only the uppermost 7.5mof thecore. Lithologically, this section consists of relatively homogeneous, greyto greenish calcareous clayey silt, with a dark-coloured layer depositedbetween220and360 cm. This section also contains a greyish-brownashlayer at 691–698 cm,which is considered furtherwith respect to the agecontrol of the core in Section 6 ‘Age determination’.
4.2. Organic-walled dinoflagellate cysts
Fifty-five samples were taken for palynological analysis from coreMD01-2430; with an average sampling interval of 10 cm, representinga temporal resolution of 180 to 370 years. Samples were preparedusing a standard method (e.g. de Vernal et al., 1998; Head et al., 2005)that excludes oxidizing reagents in order to prevent destruction of themore acid-sensitive organic-walled cysts (such as peridinioid cysts;e.g. Turon, 1984; Schrank, 1988; Marret, 1993). Prior to acidtreatments, Lycopodium tablets were added to permit estimation ofpalynomorph concentrations (Stockmarr,1971). Calculation of relativeabundances (percentages) is based on the total sum of dinocysts (250cysts, where possible).
Dinoflagellate cyst nomenclature follows Fensome and Williams(2004), and followsWall et al. (1973), Eaton (1996),Mudie et al. (2001),Rochon et al. (2002) and Marret et al. (2004) for the ‘Paratethyan’ taxa.Most of these endemic taxa present a wide intraspecific variability, andsome particular taxonomic attributions are discussed in Appendix A.Other aquatic palynomorphs were also counted, such as freshwateralgae (Pediastrum, Concentricystes, Cymatiosphaera,…), acritarch-typespecimens, tintinnids and benthic foraminiferal linings.
To characterize the dinocyst assemblages, select indices have beencalculated. Diversity is expressed as the H(s) index: −∑ [(n/N)×Ln(n/N)], where n is the number of specimens recorded for each taxon,and N the total dinocyst count for each sample (Shannon andWeaver,1949). Dominance corresponds to (n′+n″)/N, where n′ is the numberof specimens of the most abundant species, n″ the number ofspecimens of the second most abundant species, and N the totaldinocyst count for each sample (e.g. Goodman, 1979).
To reconstruct qualitative SST changes at the core site, ‘thermophilictaxa’ have been grouped (Fig. 3). These taxa are: Tuberculodiniumvancampoae, Tectatodinium pellitum, Spiniferites mirabilis, Spiniferitesdelicatus, Lingulodinium machaerophorum, Polysphaeridium zoharyi,Operculodinium israelianum, Stelladinium stellatum (e.g. Wall et al., 1977;de Vernal et al., 1992; Marret and Zonneveld, 2003; LL bibliographicdatabase).
5. Results
All the samples processed contain an abundant and often diversedinocystflora. At least 40dinoflagellate cyst specieswere recorded,withcyst concentrations ranging from 123 to 66,617 cysts per cm3 of drysediment (Fig. 3). Relative frequencies of these taxa showa successionofseveral distinct associations,which are indicative of variations inpalaeo-climatic conditions and surface-water characteristics. On the whole,assemblages are generally dominated by Spiniferites, Operculodiniumand Lingulodinium genera, and six major intervals can be identified inthe dinocyst results (Fig. 2). The concentrations of some non-pollenaquatic palynomorphs encountered (e.g. Pediastrum, Cymatiosphaeraalgae) are also shown in Fig. 3.
5.1. Dinocyst interval A (samples from 710 to 630 cm)
This interval is characterized by the dominance of the endemicspecies Spiniferites cruciformis (around 80%; Fig. 2). The other significantspecies in this interval are the ‘Paratethyan’ taxa Caspidinium rugosum,Invertocysta? sp. A, Invertocysta? sp. B and the cosmopolitan speciesOperculodinium centrocarpum and Spiniferites belerius (e.g. Turon, 1984;Rochon et al., 1999; Marret and Zonneveld, 2003). Reworked palyno-morph abundances (predominantly bisaccate pollen of indistinct age)appear relatively high, although thismust be confirmed in the context ofdetailed pollen analysis of the core. Individual cysts of S. cruciformisdisplay a verywide range ofmorphotypes, that we regrouped accordingto both the length of the processes (forms 1 to 5 of Mudie et al., 2001)and the shape of the central body (from ovoidal to cruciform). No clear

Fig. 2. Summary diagram showing provisional stratigraphic framework, dinocyst zonation and percentage diagram of major dinocyst taxa versus depth in MD01-2430. Arrows on the right-hand side show levels with direct or indirect datings(uncorrected 14C ages BP).
55L.Londeix
etal./
Reviewof
Palaeobotanyand
Palynology158
(2009)52
–71

Fig. 3. Distribution of dinoflagellate cysts and other palynomorphs in MD01-2430.
56 L. Londeix et al. / Review of Palaeobotany and Palynology 158 (2009) 52–71
relationship between this complex classification and surface-watercharacteristics, however, has been detected.
5.2. Dinocyst interval B (samples from 620 to 450 cm)
Besides S. cruciformis, the dominant taxa in this interval are Lin-gulodinium machaerophorum, Operculodinium centrocarpum and Spi-niferites ramosus, i.e. three eurybiotic species (e.g. Wall et al., 1977;Edwards and Andrle, 1992; Marret and Zonneveld, 2003).
Two phases can be distinguished in this interval: •1) In the firstphase, B. tepikiense, L. machaerophorum, T. vancampoae are moreabundant than in the second. They are found in association withthermophilic species such as Tectatodinium pellitum, Spiniferitesdelicatus or Spiniferites mirabilis (Marret and Zonneveld, 2003) andalso with the cool temperate to subpolar species Pentapharsodiniumdalei (Matthiessen, 1995; Rochon et al., 1999; Marret et al., 2004; deVernal et al., 2005); •2) In the second phase, peak concentrations of C.rugosum are recorded.
One of the more striking features of dinocyst interval ‘B’ is themodest representation (up to 6–7%) of the subtropical/tropical coastalspecies Tuberculodinium vancampoae (e.g. Edwards and Andrle, 1992;Wall et al., 1977; Marret and Zonneveld, 2003). This indication ofwarming is supported by the coeval relative abundance of otherthermophilic taxa (Fig. 3). Nevertheless, these taxa record a slightdecrease during the ‘B2’ phase.
With the exception of S. cruciformis, we note that the dominanttaxa in this interval tolerate high seasonal contrasts in both SSS andSST.
5.3. Dinocyst interval C (samples from 440 to 360 cm)
This interval is characterized by a second local maximum in S.cruciformis frequencies, accompanied by the maximum developmentof Pyxidinopsis psilata, another endemic taxon (up to 7%; Fig. 2) and amarked increase in dinocyst concentrations. C. rugosum is present atthe base and at the top of the interval, albeit at low frequencies. We

57L. Londeix et al. / Review of Palaeobotany and Palynology 158 (2009) 52–71
also note the very low relative abundance of O. centrocarpum (Fig. 2)and the virtual absence of thermophylic taxa (Fig. 3). A remarkablefeature in the middle of interval C is the sharp peak in theindeterminate Peridinioid cyst A (Fig. 2, Plate I.15). In contrast todecreasing concentrations of reworked palynomorphs (reaching verylow values in the second part of the interval), the abundance of thefreshwater algae Pediastrum increases over the interval (Fig. 3).
5.4. Dinocyst interval D (samples from 350 to 333 cm)
In this interval, Pyxidinopsis sp. A and Spiniferites belerius attainmax-imum abundances. Furthermore, cool species such as Bitectatodiniumtepikiense, Spiniferites elongatus, and Nematosphaeropsis labyrinthus areabundant in addition to the ubiquitous and eurybiotic Spiniferitesramosus. It is noteworthy that N. labyrinthus was not present in thesequence prior to that interval; it is considered a typical oceanic species(Matthiessen, 1995; Rochon et al., 1999; Marret et al., 2004; de Vernalet al., 2005) which may be associated with nutrient-rich, cool waters(Turon and Londeix, 1988; Devillers and de Vernal, 2000). Severalauthors have noticed the transitional role of N. labyrinthus duringthe course of severe hydrological changes, particularly during thetransitions between glacial and interglacial conditions (Turon, 1984;Eynaud et al., 2004; Penaud et al., 2008). Even if poorly represented,heterotrophic taxa (and particularly Brigantedinium spp.) show here aclear increase in relative abundance (Fig. 2), suggesting increasedprimary production (e.g. Lewis et al., 1990; Radi and de Vernal,2004) and/or oxygen depletion of the bottom-waters (Zonneveldet al., 2007).
5.5. Dinocyst interval E (samples from 330 to 250 cm)
Interval ‘E’ corresponds approximately to the sapropel layer M1.Dinoflagellate cyst concentrations are here the highest of thesequence, and show two distinct peaks. Operculodinium centrocarpum(notably morphotypes with short processes) displays peak frequen-cies and dominates the dinocyst assemblages. O. centrocarpum isaccompanied by the eurybiotic taxa Spiniferites ramosus, Lingulodi-nium machaerophorum and Polysphaeridium zoharyi.
5.6. Dinocyst interval F (samples from 240 cm to top)
On the whole, O. centrocarpum, S. ramosus and L. machaerophorumare the dominant species of this interval. Three sub-intervals can bedistinguished:
1) The first, characterized by high percentages of P. dalei and B.tepikiense and, to a lesser extent, S. elongatus and Ataxiodiniumchoane, concomitant with a slight decrease in the thermophilictaxa (Fig. 3). Dinocyst concentrations are low, when freshwateralgae and reworked palynomorphs are almost absent (Fig. 3).
2) The second, showing a slight increase in dinocyst concentrations,as well as enhanced frequencies of L. machaerophorum, O.centrocarpum and Tectatodinium pellitum, and reduced frequenciesof P. dalei, B. tepikiense, S. elongatus and heterotrophic species(peridinioids only). The dinocyst assemblages are similar to thoseof interval ‘E’, but are suggestive of lower primary productionwithin a warmer context. These changes occurred in conjunctionwith a slight reduction in specific diversity (Fig. 3).
3) The third, with the highest percentages of the whole sequence forS. ramosus, as well as for minor taxa such as Spiniferites mirabilis,Nematosphaeropsis labyrinthus and Islandinium? cf. minutum. Thecool species P. dalei is relatively abundant among the cyst flora,apparently concomitant with the relatively high abundance ofthermophilic taxa. Benthic foraminiferal lining concentrationsbecome significant (Fig. 3). The assemblages of this sub-intervalappear almost identical to those of the present-day Marmara Sea.
6. Age determination
Based on direct and indirect datings, a provisional age model isproposed for coreMD01-2430 (Fig. 5). The dark layer at 220–360 cm isassigned to the M1 sapropel (e.g. Aksu et al., 2002b; Hiscott et al.,2002) that was dated between 10,500 and 6400 14C conventionalyears BP (Çağatay et al., 2000; Aksu et al., 2002a; Kaminski et al.,2002). Because of its stratigraphical position, the 691–698 cm ashlayer (‘afl-1’ of Beck et al., 2007) is considered to represent the Y-2tephra from the Cape Riva eruption of the Santorini volcano. The latterwas dated between 18,050 and 18,880 14C conventional years BP(Pichler & Friedrich, 1976; Eriksen et al., 1990; Wulf et al., 2002); amean value of 18,465±415 14C years BP is employed here.
The precise correlation of dinoflagellate cyst assemblages (Fig. 4)allows chronological data available from neighbouring cores MAR98-12 and MAR97-11 (Mudie et al., 2002a) to be incorporated in thepresent study. Intercore correlation and depth-to-time transformationwere achieved through linear interpolation procedures betweenconsecutive markers: Table 1 presents AMS 14C ages (not correctedof reservoir-effect) available from cores MAR98-12 and MAR 97-11(Aksu et al., 2002b; Mudie et al., 2007). Because of the uncertaintiesassociatedwith the age determinationmethod employed in this study,and because an accurate direct radiocarbon based age model isforthcoming (Vidal et al., in press), our data will be presented versusdepth, nevertheless some ages will be used to aid the comparison ofour results with other findings from the same time interval.
The basal age of the studied interval of core MD01-2430 isapproximately 19 14C ka BP. Sedimentation rates are indicated on Fig. 5.
Unless otherwise indicated, all dates presented here are conventional14C ages (i.e. uncalibrated andnot reservoir-effect corrected) because thereservoir age is not actually known for the Marmara Sea (Major et al.,2002; Bahr et al., 2005). However, to facilitate stratigraphic comparisons,provisionally calibrated calendar ages are provided (Table 1; betweensquarebrackets in the text), reservoir-effect corrected, using theprogramCalib 5.1b (Stuiver and Reimer, 1993; Stuiver et al., 2005) which use astandard marine reservoir of around 400 years.
7. Discussion — evolution of the environmental context
Variations indinocyst assemblages fromMD01-2430 are very similarto those previously studied from theMarmara Seawithin the same timeinterval (Mudie et al., 2001, 2002a, 2004), allowing confident correla-tions and the drawing-up of a (provisional) chronostratigraphicalframework (cf. above). The present sequence provides the longest,continuous dinocyst record for the late Quaternary of the Marmara Sea.Moreover, the high resolution sampling should allow us to evaluate ingreater detail the transitions between the various events previouslyobserved in other studies.
7.1. Interval ‘A’=lower Last Glacial Maximum (~19 to 17.2 14C ka [~22.4to 20.5 cal. ka BP]): a caspibrackish lake
In modern surface sediments, S. cruciformis has been recorded onlyin the Black Sea (Mudie et al., 2001) and the Caspian Sea (Marret et al.,2004). This species corresponds to present-day environments withcontinuous low sea-surface salinities (SSS=12.5–19 psu) throughoutthe year. Sea-surface temperatures (SST) range from5 to 10.5 °C duringwinter to 21–27 °C during summer. Using SSS estimates applied toS. cruciformis relative abundances from a late Quaternary sequence ofthe Black Sea, Mudie et al. (2001) established that the mean annualsalinity tolerance of this species lies in the range 15–22 psu. When S.cruciformis percentages are higher than ~70%, these authors note thatmean annual SSS lies between 5–7 and 14–16 psu with winter andsummer SST around 3–5 and 10–13 °C, respectively (op. cit.). Thatcorroborates the occurrence of not fully fresh water (Aksu et al.,2002b), even if the Kouli et al. (2001) record of S. cruciformis in late

58 L. Londeix et al. / Review of Palaeobotany and Palynology 158 (2009) 52–71
Quaternary lacustrine sediments from North Greece, leads to considera wider possible range of salinity for this species. Indeed, given thatregional sea-level was around ~100m lower thanpresent (Smith et al.,1995; Aksu et al., 1999) and that the Dardanelles and Bosphorus sillswere situated at ca −78 m and −40, respectively, at this time (Aksuet al., 2002b)], theMarmara Seawas an isolated lacustrine system (e.g.Stanley and Blanpied, 1980; Çağatay et al., 1999; Aksu et al., 2002b;Reichel and Halbach, 2007).
Independently of SSS, the abundance of S. cruciformis undoubtedlyindicates a narrow seasonal contrast in SSS, but a large probableseasonal contrast in SST, potentially of up to 17 °C. The very lowdinocyst concentrations denote a weak primary production, a findingthat is in agreement with the characteristics of an isolated system (e.g.Çağatay et al., 2000; Caner and Algan, 2002; Yaltirak and Alpar, 2002).Although the erosion of the emerged margins (Aksu et al., 1999;Çağatay et al., 2000; Meriç and Algan, 2007) may account for the highamount of remorked palynomorphs, the absence of fresh water algaeindicates very weak river inputs (unless their scarcity reflects climaticconditions unfavorable to their development). This climatic andhydrological context remained constant from at least the base of thesequence (~19 14C ka BP) to ca 17.2 14C ka BP (between the samples650 and 600 cm)[~22.4 to 20.5 cal. ka BP].
7.2. Interval ‘B’=upper Last Glacial Maximum and HE1 (~17.2 to 13.9/13.8 14C ka BP [~20.5 to 15.9 cal. ka BP]): warming, with subsequentmeltwater outflow from the Black Sea
Increased relative abundance of thermophilic species in thisinterval indicate a significant increase in sea-surface temperature,particularly during the interval ‘B1’ (620 to 520 cm; ~17.2 to ~15.3 14Cka BP [~20.5 to 18.1 cal. ka BP]). As T. vancampoae has not been foundin modern surface sediments (with two exceptions) in areas with SSTbelow 12.7 °C in winter and 14.5 °C in summer (maximumtemperature range: 7.1 and 29.5 °C winter–summer SST; Marret andZonneveld, 2003; Zonneveld and Susek, 2007), this species isregarded as being thermophilic. It has been found in brackish aswell as in fully marine environments with SSS ranging between 0 and38 psu (Bradford and Wall, 1984; McMinn, 1991). In addition to itswide SSS tolerance, T. vancampoae withstands a seasonal SSS contrastof up to 27–39 psu (McMinn, 1991). Although it is always found closeto the coast, T. vancampoae is not restricted to shallow sites (Martínez-Hernández and Hernández-Campos, 1991; Marret, 1994; Marret andde Vernal, 1997; Marret and Zonneveld, 2003). Its highest relativeabundances are found in oligotrophic environments (Marret andZonneveld, 2003). The modest presence of T. vancampoae during theupper Last Glacial Maximum (LGM) indicates that winter and summerSSTs were above 12.7° and 14.5 °C, respectively, consistent withgeochemical changes recorded in the Black Sea (Major et al., 2006)and European geomorphological systems and warmer climate
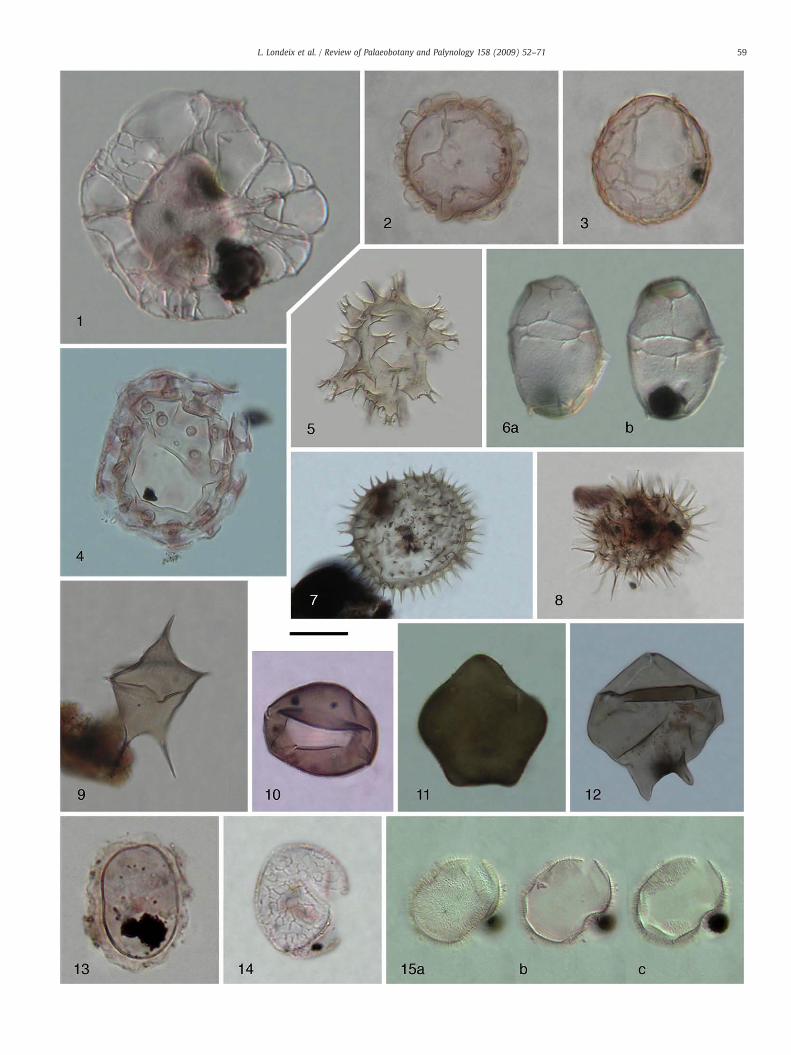
Plate I. Dinoflagellate cysts from the late Quaternary of Marmara Sea. Same magnification,
1. Seriliodinium explicatum, 450 cm.2. Ataxiodinium cf. choane, 280 cm.3. Ataxiodinium sp. A, 280 cm.4. Tuberculodinium vancampoae, 560 cm.5. Xandarodinium xanthum, 240 cm.6. Impagidinium sp. A; a — upper focus lateral view, b — low focus lateral view,7. Selenopemphix quanta, 280 cm.8. Selenopemphix? cf. quanta, 230 cm.9. Stelladinium stellatum, 230 cm.10. Brigantedinium cariacoense, 50 cm.11. Quinquecuspis concreta, 280 cm.12. Lejeunecysta sabrina, 280 cm.13. Cyst of Scrippsiella trifida, 230 cm.14. Pyxidinopsis reticulata, 230 cm.15. Peridinioid cyst A; a — antapical view, b — equatorial cross section, c — apica
occurring ~3 ka prior to the Bølling warming (Kalicki and Sanko,1998; Huhmann et al., 2004; C. Schluchter cited by Major et al., 2006).
Modern C. rugosum is only known from Caspian Sea (Marret et al.,2004) from environments with very high seasonal contrast in SST, lowseasonal contrast in SSS and influence of fresh meltwaters providedduring spring from the decay of northern seasonal sea ice cover.Moreover, we can note that C. rugosum has a morphology extremelyclose to that of Gonyaulax apiculata (sensu Evitt et al., 1985), a modernspecies recorded in some freshwater alpine lakes supplied by meltedice (e.g. Pénard, 1891; Bourrelly and Couté, 1980; Evitt et al., 1985;Hickel and Pollingher, 1986; Jacquet et al., 2007). Maximumfrequencies of C. rugosum begin at or after ~15.3 14C ka BP [~18.1 cal.ka BP](510 cm), consistent with meltwater inputs via the Black Sea(e.g. Major et al., 2002, 2006; Knies et al., 2007) which are themselvesa consequence of the synchronous disintegration of the Eurasian icesheets (e.g. Grosswald, 1980, 1998; Denton et al., 1999; Grosswald andHughes, 2002). Maximum frequencies of C. rugosum correspond to asharp rise in Black Sea sea-level (e.g. Serebryanny, 1982; Pirazzoli,1996; Konikov et al., 2007; Shmuratko, 2007) and to the overflow ofwater into theMarmara Sea (e.g. Hiscott et al., 2007; Knies et al., 2007;Reichel and Halbach, 2007). Nevertheless, if the abundance of C.rugosum is indicative of meltwater input, the relative abundance of C.rugosum on and after ~16.8 14C ka BP [~20.0 cal. ka BP](600 cm) raisesa problem asmeltwater could not come from the Black Sea as sea-levelis considered below the Bosphorus sill at this time (e.g. Konikov et al.,2007; Shmuratko 2007). As no Late Glacial glaciers are known in thedrainage basin of the Marmara Sea, a meltwater influx may be linkedto an increase in melting of hinterland permafrost (Dobiński, 2005).
The introduction of meltwater into theMarmara Sea led to a strongseasonal contrasts in SSS and SST expressed by the decline of S.cruciformis, which was replaced by species able to accept both suchseasonal contrasts (e.g. O. centrocarpum, S. ramosus, L. machaero-phorum, T. vancampoae); seasonal contrast in SSS was neverthelessprobably lower than 17 psu (the tolerance threshold of L. machaer-ophorum). Such strong seasonal contrasts corroborate the finding ofstrong gradients leading to stratification, stagnation and deoxygena-tion (Reichel and Halbach, 2007).
Overall, the MD01-2430 sequence records a two-step meltingepisode, with:
1) Aweak phase, from~16.8 to ~15.3 14C ka BP [~20.0 to 18.1 cal. ka BP],corresponding to the highest SST of interval ‘B’ and associated withpossible melting of hinterland permafrost. Within the uncertaintiesof the provisional age model, these episodes appear synchronouswith the early warming observed at the end of the LGM, from 21 cal.ka BP (ca 18.1 14C ka BP), in the Greenland air–temperature record(Stuiver andGrootes, 2000; Johnsen et al., 2001) and in severalNorthAtlantic temperature records (e.g., Jones and Keigwin, 1988; Bardet al., 2000; Alley et al., 2002; Pailler and Bard, 2002) as well as withconsiderable continental freshwater fluxes routed through the
scale bar=20 µm.
500 cm.
l view, 390 cm.
59L. Londeix et al. / Review of Palaeobotany and Palynology 158 (2009) 52–71

Fig. 4. Correlation of dinoflagellate cyst records from cores MAR98-12, MAR97-11 and MD01-2430. This correlation enables the application of chronological data available from coresMAR98-12 andMAR97-11 (Mudie et al., 2002a) to theMD01-2430 core. Additional dates are deduced from stratigraphical interpretation of distinctive layers in coreMD01-2430. The14C ages are not corrected for the reservoir age, which is not known.
60 L. Londeix et al. / Review of Palaeobotany and Palynology 158 (2009) 52–71
English Channel between ~20 and 17 cal. ka BP (ca 17 and 14.5 14C kaBP; Ménot et al., 2006; Eynaud et al., 2007; Toucanne et al., 2008).
2) A higher-magnitude second phase with important meltwateroutflow from the Black Sea, occurring from ~15.3 to ~13.9/13.814C ka BP [~18.1 to 15.9 cal. ka BP]. This phase corresponds todinocyst interval ‘B2’ and an increased erosion rate, and appearscontemporaneouswith the disintegration of large icemasses in theEurasian continental interior (e.g. Grosswald, 1998; Denton et al.,1999; Grosswald and Hughes, 2002), as well as with the beginningof HE1 in the open Atlantic (e.g. Elliot et al., 1998; Bard et al., 2000).
Table 1Data used for age determination for the investigated core MD01-2430.
Core Depth (cm) Data Dated material Uncorrradioc
MD01-2430 0 Sediment surface 0230 M1-interruption 6,400691–698 Y-2 tephra Terrestrial charcoal 18,465
MAR98-12 50 14C AMS dating Bivalve fragment 4,200±130 14C AMS dating Nuculacea spp. 10,660
MAR97-11 79 14C AMS dating Turritella spp. 10,79092 14C AMS dating White mussels 12,970174 14C AMS dating Small oyster 14,940204 14C AMS dating White mussels 15,590
References: (a) Çağatay et al. (2000), Aksu et al. (2002a), Kaminski et al. (2002); (b) Pichler &et al. (2007). Dates from carbonates were calibrated with Marine04 from Calib 5.1b; dates f
7.3. Interval ‘C’=ca Bølling/Allerød (~13.9/13.8 to 10.9/10.7 14C ka BP[~15.9 to 12.2 cal. ka BP]): the last Marmara lake
The decrease of C. rugosum at about 13.9 14C ka BP [~15.9 cal. ka BP](450–440 cm) indicates a decrease in meltwater supply and whichparallels with the weakening of the streamflow of the Seim river (aDnepr emissary) at 14 14C ka BP (Borisova et al., 2006). The subsequentslight rise of C. rugosum from ~11.9 to ~10.9 14C ka BP [~14.0 to 12.4 cal.ka BP](380–360 cm) is further consistent with a retreat of theFennoscandian Ice Sheet during the Allerød (Sejrup et al., 2000;
ected conventionalarbon dates (yr BP)
Reference MD01-2430depth (cm)
Deduced dates (cal. yr BP)calibrated from Calib 5.1b
0 0(a) 230 6,880
±415 (b) 695 21,877100 (c) 171 4,279
±130 (c) 351 12,003±70 (c) 357 12,219±80 (c) 402 14,519±90 (c) 496 17,543±90 (c) 530 18,620
Friedrich (1976), Eriksen et al. (1990),Wulf et al. (2002); (c) Aksu et al. (2002b), Mudierom terrestrial material (Y-2 layer only) were calibrated with IntCal04 from Calib 5.1b.

Fig. 5. Age–depth curve for core MD01-2430 as constructed using the correlation ofdinocyst records from cores MAR98-12, MAR97-11 and MD01-2430. The 14C ages arenot corrected for the reservoir age, which is not actually known. Select averagesedimentation rates are provided (italics).
61L. Londeix et al. / Review of Palaeobotany and Palynology 158 (2009) 52–71
Knies et al., 2007) and with the freshwater deposit ‘C2’ from the BlackSea (Major et al., 2002; Ryan et al., 2003). These findings suggest thatthe ~2‰ drop in ∂18O recorded by Reichel and Halbach (2007) mayhave resulted from a decrease in salinity rather than an increase in thetemperature of marine surface water entering the sea of Marmarabetween ~13 and 11.5 cal. ka BP (~11.4 to 10.2 14C ka BP; op. cit.).
The second peak in S. cruciformis reflects a return to low seasonalcontrasts in SSS with mean values probably close to that occurring atthe base of the sequence (between 5 and 16 psu). The high amount ofreworked palynomorphs suggests that the erosion rate remainedsignificant but nevertheless decreased rapidly at ~13 14C ka BP[~14.6 cal. ka BP]. At this time, the abundance of the freshwater algaePediastrum increased sharply, indicating increased river inputs. Thesechanges may reflect a rise in base level (leading to reduced erosion)and/or a wetter climate in the hinterland (higher freshwater input).
The very low proportion of thermophilic dinocyst species agreeswith cold conditions at this time (Filipova-Marinova et al., 2004;Mudie et al., 2007), perhaps in association with stable SSTsthroughout the year (absence of O. centrocarpum). Primary productionappears strongly enhanced (distinct increase in dinocyst concentra-tions), probably due to improved hydrological circulation. Thisincrease in primary productivity is nearly synchronous with theincreased bottom-water oxygenation recorded between ~13 and 11.514C ka BP by Reichel and Halbach (2007).
The significance of the sharp peak of unknown peridinioids cysts(centered 390 cm) is presently unclear but is associated with amarked decrease in the freshwater algae Pediastrum. According to itsstratigraphical position (~12.4 14C ka BP [~13.9 cal. ka BP]) near the
Older Dryas, it may reflect a drier (with reduced river input) and/orcooler climate at that time.
For the time interval ~14.7–11.314C kaBPMarmara Sea times,Hiscottet al. (2002, 2008) surmise an evaporative basin with evaporationexceeding precipitation and river runoff, leading to a lowstand sea-level.However, the overwhelming dominance of the stenohaline taxa S.cruciformis, decrease in reworked cysts, increase in freshwater algae, andimproved primary production do not agree with this assertion.
7.4. Interval ‘D’=Younger Dryas (~10.9/10.7 to 10.0/9.9 14C ka [~12.2 to11.2 cal. ka BP]): entrance of Mediterranean water into the Marmara Sea
Modern marine conditions with the influence of Mediterraneanwater on bottom-waters of the Marmara basin are detected ca 10.8 14Cka BP [~12.2 cal. ka BP] in the record of benthic foraminiferal linings. Asurface water influence is detected slightly later when abundances ofthe dinocyst N. labyrinthus (a fairly marine species) increased, ca 10.414C ka BP [~11.7 cal. ka BP] (340/350 cm). These dates for the entranceof Mediterranean water into the Marmara Sea are slightly youngerthat those provided by Aksu et al. (2002b) or Kaminski et al. (2002),perhaps due to uncertainty in the age model used, and imply atransition from ‘caspibrackish’ to marine environmental conditionssomewhat faster than the ~1.5 ka estimated by Reichel & Halbach(2007).
Sea-surface salinity is likely to have been relatively stable (B.tepikiense does not tolerate seasonal contrast in SSS higher than 3 psu)and probably no lower than 27 psu (cf. Section 5.5). While theabundance of the caspibrackish species S. cruciformis declinedprogressively in the Marmara Sea (Mudie et al., 2002a; this work), itremained fairly abundant in the Black Sea at that time (Mudie et al.,2002a; Atanassova, 2005). This contrast may indicate a weakerinfluence of the Black Sea outflow, or even a possible deconnection ofthe Marmara Sea from the Black Sea (e.g. Ryan et al., 1997; Sperling etal., 2003). Despite the apparently reduced influence of freshwaterinput (sharp decrease in freshwater algae amount, disappearance ofreworked palynomorphs) in the Marmara Sea which is linked to adecrease in precipitation (e.g. Bottema, 1995; Bottema et al., 1995;Aksu et al., 2002c; Mudie et al., 2002b; Filipova-Marinova et al., 2004;Mudie et al., 2007; Eriş et al., 2007), the dinocyst assemblages do notprovide evidence for high sea-surface salinities.
The maximum occurrence of the cool temperate to subpolar taxa B.tepikiense and S. elongatus indicate YoungerDryas SSTcolder thanduringthe Bølling/Allerød in agreement with pollen records from thesurrounding region (e.g. Bottema, 1995; Bottema et al., 1995; Mudieet al., 2002b; Filipova-Marinova et al., 2004; Mudie et al., 2007).However, the relative abundance of thermophilic species increasedslightly from the Allerød to Younger Dryas; an intriguing findingpreviously also detected by Sperling et al. (2003) in alkenone SSTestimates. This contrasting behaviour probably reflects strong seasonalcontrasts in SST and also suggests that (perhaps due to the inflow ofMediterraneanwaters)Marmara SSTs actually increased fromAllerød toYounger Dryas while atmospheric conditions became colder, implyingan opposition between hydrological and atmospherical phenomena atthis time. This opposition is not observed in the Black Sea (Bahr et al.,2008), reinforcing the assumption of a decoupling of the Black andMarmara sea basins (op. cit.).
Increased abundances of both N. labyrinthus and heterotrophictaxa, together with a rise in benthic foraminiferal linings, testify toinvigoration of circulation and enhanced bottom-water oxygenation.
7.5. Interval ‘E’=lower Holocene (~10.0/9.9 to 7.1/6.8 14C ka BP [~11.2 to7.5 cal. ka BP]): M1 sapropel deposition
In modern assemblages, O. centrocarpum is generally abundantwhen seasonality in SST is important (up to 19 °C), and the speciestolerates all SSSs and seasonal salinity contrasts (e.g. Wall et al., 1977;

62 L. Londeix et al. / Review of Palaeobotany and Palynology 158 (2009) 52–71
Matsuoka, 1987; McMinn, 1991; Pospelova et al., 2005). The associa-tion of cool temperate to subpolar taxa (Pentapharsodinium dalei andBitectatodinium tepikiense) and thermophilic species (for which thecombined sum is higher than in the preceding interval) also suggests astrong seasonal contrast in SST. In contrast to the Aegean Sea, no clearevidence of strong SST increase (i.e. 10 °C, Hiscott et al., 2002) hasbeen recorded here, and as inferred by Aksu et al. (2002b), surfacewaters may have been slightly cooler than today.
O. centrocarpum with short or aberrant processes is considered tobemore abundant in subnormal to hypohaline SSS (e.g. de Vernal et al.,1989; Nehring, 1994; Dale, 1996; Matthiessen and Brenner, 1996;Nehring, 1997; Ellegaard, 2000; Brenner, 2001a,b, 2005). P. dalei, P.psilata, B. tepikiense or S. belerius have never been recorded in modernenvironments with SSS lower than 13 to 15 psu (cf. de Vernal et al.,2001 [n=677 data base]; Leroy, 2001;Marret et al., 2004). Conversely,N. labyrinthus is absent when SSS are lower than 17.5 to 27 psu (Wallet al., 1977; de Vernal et al., 2001). Seeing that N. labyrinthus is absentin interval ‘E’ (and not in surrounding intervals), minimum SSS for thatinterval is therefore estimated to fall in the range from 13 to 15 to17.5 psu (maximum 27 psu). The decrease in species richness (Fig. 3)also suggests decreased SSS relative to the preceding interval (Dale,1996; Head et al., 2005).
Themajor species present in this interval tolerate seasonal contrastin SSS of up to 17 psu (L. machaerophorum) or 39 psu (O. centrocarpum,S. ramosus). Although less abundant, species such as B. tepikiense or S.belerius, however, appear not to tolerate seasonal contrast in SSShigher than 3 psu (Grøsfjeld and Harland, 2001). Unless the tolerancerange of these latter species is wider than that recorded in modernsurface sediments, the association of species in this interval withstrongly contrasting tolerances of SSS seasonality may possibly beexplained by strong inter-annual variability.
The speciesO. centrocarpum is quite resistant to benthic degradation(e.g. Turon,1984; Zonneveld et al., 2001, 2007); thus, its abundance hereis considered to reflect a real increase in the production of cysts ratherthan increased preservation in the benthic zone. This point isemphasised by the fact that the concentration of oxidation-sensitivecysts (e.g. heterotrophic dinocysts; Zonneveld et al., 2007) remainsrelatively low. The prasinophyte algae Cymatiosphaera (Boalche andParke,1971; Tappan,1980) is commonlyassociatedwithneritic andevenestuarine environments (e.g. Takahashi, 1971; Tappan, 1980; Wall andDale, 1974; Mudie, 1992; Solomon et al., 2000), and is often associatedwith high primary productivity (de Vernal and Mudie, 1989). In thiscontext, the abundance of bothCymatiosphaera and dinocysts in interval‘E’may reflect enhancedprimary productivity due to the influxof coastalwaters.
A sharp decrease in dinocyst concentrations and O. centrocarpumpercentages is recorded around 7.8 14C ka BP [~8.5 cal. ka BP](270 cm),the timing of which is reminiscent of the 8.2 cal. ka BP (~7.7 14C ka BP)cold event widely known from the Northern Hemisphere (see Alleyand Àgústsdóttir, 2005; Rohling and Pälike, 2005 for reviews). Whileany decrease in thermophylic species is observed (Fig. 3) a slightincrease in the cool temperate species S. elongatus is centered on thattime (Fig. 2), as in the Aegean sea dinocyst assemblages at 8.3 cal. ka
Plate II. Dinoflagellate cysts from the late Quaternary of Marmara Sea. Same magnification
1. Spiniferites cruciformis forma 1, 560 cm.2. Spiniferites cruciformis forma 2, 590 cm.3. Spiniferites cruciformis forma 3, 590 cm.4. Spiniferites bentorii, 50 cm.5. Spiniferites mirabilis, 230 cm.6. Spiniferites delicatus, 230 cm.7. Spiniferites hyperacanthus, 360 cm.8. Spiniferites ramosus ramosus, 230 cm.9. Spiniferites septentrionalis, 170 cm.10. Cyst of Pentapharsodinium dalei, 290 cm.11. Spiniferites belerius, 280 cm.12. Spiniferites elongatus, 560 cm.
BP (Marino, 2008). In the Marmara Sea, the ‘8.2 event’ should havebeen an hydrological perturbation (particularly in productivity) ratherthan a fair cooling event.
The near disappearance of benthic foraminiferal linings during thedeposition of sapropel M1 suggests dysoxic/anoxic benthic conditions,in agreement with the observations of Kaminski et al. (2002) andTolun et al. (2002). Several debated hypotheses exist regarding theorigin of sapropel M1, with different studies suggesting that thesapropel resulted from either: a) freshwater outflow from the BlackSea (e.g. Stanley & Blanpied, 1980; Aksu et al., 1999, 2002b; Hiscottet al., 2002; Hiscott and Aksu, 2002); b) increases in local rainfall andrunoff, contributing to the creation of a freshwater lid and anoxia(Roberts et al., 2008); or, c) global transgression, via the Mediterra-nean Sea, leading to enhanced productivity (Sperling et al., 2003).
If the Black Sea outflowwere strengthened to the point of initiatingthe M1 deposition (~10.5 14C ka BP; Hiscott et al., 2002; Aksu et al.,2002b), it would be rather surprising not to observe closer similaritiesbetween the cyst floras of the Black Sea (Mudie et al., 2001, 2002a;Marret et al., in press) and the Marmara Sea (Mudie et al., 2002a; thiswork), particularly with respect to the endemic taxa S. cruciformis andP. Psilata. These taxawere very abundant in the Black Sea until ~7.2 14Cka BP (Marret et al., in press) but appear to have been virtually absentfrom the Marmara Sea by ~9.6 14C ka BP (Mudie et al., 2002a; thiswork). In addition, the scarcity of freshwater algae and reworkedpalynomorphs amounts in the M1 sapropelic layer suggests a weakinput from surrounding rivers, as formerly noticed by Aksu et al.(2002c), and contradicts the hypothesis of enhanced runoff (Robertset al., 2008); nevertheless, SSS appears to have been relatively low.
The conditions characterizing the M1 deposition did not actuallyinitiate at the beginning of the dinocyst interval ‘E’ (~9.9 14C ka BP[~11.1 cal. ka BP], 330 cm), but instead slightly afterwards. Interval ‘E’began with a moderate primary productivity (moderate dinocystconcentrations), moderate SST seasonal contrasts and, probably,effective bottom-water oxygenation (as suggested by a peak in benthicforaminiferal linings). The onset of increased productivity started ca9.4 14C ka BP [~10.5 cal. ka BP] (310/320 cm). While other hypothesescannot be fully ruled out, the dinocyst evidence suggests thatenhanced productivity was a key factor in the generation of sapropelM1.
7.6. Interval ‘F’=upper Holocene (~7.1/6.8 14C ka BP [~7.5 cal. ka BP] toPresent): initiation of the modern circulation
After the end of M1 sapropel deposition, the Marmara Seaexperienced periods of unstable hydrological conditions. From ~6.8to 5.6 14C ka BP [~7.5 to 5.8 cal. ka BP] (240 to 210 cm; sub-interval‘F1’) dinocyst concentrations (i.e. primary productivity) decreased tovalues close those of the present-day, while maximum in B. tepikiense,P. dalei, S. elongatus and A. choane point to a substantial cooling withregard to theM1 deposition period. Indeed, the relative abundances ofthese latter taxa indicate winter SST below 12 °C but summer SSTpotentially up to 25 °C (cf. P. dalei; e.g. Rochon et al., 1999; Marret andZonneveld, 2003). In addition, the abrupt reduction in river inputs (as
, scale bar=20 µm.

63L. Londeix et al. / Review of Palaeobotany and Palynology 158 (2009) 52–71

64 L. Londeix et al. / Review of Palaeobotany and Palynology 158 (2009) 52–71
shown by the near absence of freshwater algae and reworkedpalynomorphs) agrees with a reduced seasonal contrast in SSS asrevealed by B. tepikiense and A. choane tolerances (cf. de Vernal et al.,2001 [n=677 data base]; LL bibliographic database) on the order of3 psu or less. Nevertheless, taking into account the dominant speciesin this interval, SSS during the early part of interval ‘F’must have beenclose to that of the underlying interval. Thus, the decreased primaryproductivity recorded during the beginning of interval ‘F’ may belinked to reduced river input, and/or weakened hydrological circula-tion. The sub-interval ‘F1’ parallels the cooling observed from 7.1 to4.4 cal. ka BP in the northern borderlands of the Aegean Sea (Kotthoffet al., 2008) which has been interpreted as a strengthening of theSiberian High (op. cit.).
A modest increase in primary productivity occurred centered on ca5.1 14C ka BP [~7.5 cal. ka BP](200/190 cm; interval ‘F2’). Enhancedproductivity is accompanied by a clear signal of warming that may beconsidered a type of climatic optimum (withmaximum frequencies ofthermophillic species). This warm optimum parallels the climaticamelioration observed between 5.4 and 4.5 14C ka BP in pollen recordsfrom SWCrimea (Cordova and Lehman, 2005) and in the SWBlack Sea(Marret et al., in press). While the interval ‘F2’ corresponds to anepisode of enhanced primary productivity, it appears too old to beconnected to the Marmara Sea “upper sapropel” (4.75–3.2 14C yr BP;Çağatay et al., 2000; Tolun et al., 2002).
By 4.5 14C ka BP [~4.7 cal. ka BP] (170 cm, base of interval ‘F3’) thenear disappearance of the stenohaline tomildly brackish taxon P. psilata(Wall et al., 1973; Mudie et al., 2001; Leroy et al., 2007) associated withan increase in benthic foraminiferal linings indicates a more effectivebottom-water oxygenation. This, in turn, suggests a reduction in BlackSea outflow and a weakening of the halocline respectively (Kaminskiet al., 2002). From this point onwards, dinoflagellate cyst assemblagesare similar up to the topof the sequence, suggesting theestablishmentoffairly stable climatic and hydrological conditions comparable to thepresent-day situation, i.e. characterised by the outflow of brackishsurfacewater from the Black Sea, and the inflow of saline bottom-waterfrom the Mediterranean Sea. The combined presence of thermophilicand cool temperate to subpolar species suggests strong seasonality inSSTs.
7.7. A remark on similarities to Baltic Sea dinocyst assemblages
We note significant similarities between some Marmara Seadinocyst assemblages and certain Holocene or Eemian assemblagesfrom the Baltic Sea (Brenner, 2005; Head et al., 2005; Yu and Berglund,2007). Perhaps most striking is the presence of the thermophilic taxaTuberculodinium vancampoae during the Eemian deglaciation from theBaltic Sea (Head et al., 2005) as detected during the late LGM from theMarmara Sea.
We also note that the Baltic postglacial transition records a longer icefree period during summer and autumn marked by the abundance ofGonyaulax apiculata, with the subsequent dominance of O. centrocar-pum+L.machaerophorum (Brenner, 2005), interpreted as an admixtureof saline bottom-waters at the sea surface. That transition is similar totheMarmara lateGlacial toHolocene transitionwhere assemblageswith
Plate III. Dinoflagellate cysts from the late Quaternary of Marmara Sea. Same magnification
1. Pyxidinopsis psilata, 560 cm.2. Pyxidinopsis psilata, slightly cruciform shape, 390 cm.3. Pyxidinopsis? sp. A, 350 cm.4. Pyxidinopsis? sp. A, 390 cm.5. Caspidinium rugosum, 450 cm.6. Caspidinium rugosum, 500 cm.7. Invertocysta? sp. A; a — ventral view, b — dorsal view, 500 cm.8. Pterocysta cruciformis, 450 cm.9. Achomosphaera cf. andalousiense, 230 cm.10. Invertocysta? sp. B, 470 cm.11. Dubridinium caperatum, 280 cm.
Caspidinium rugosum are followed by assemblageswith dominance ofO.centrocarpum+L. machaerophorum.
Furthermore, the coeval dominance of O. centrocarpum+ L.machaerophorum+Spiniferites spp. recorded in the late Holocene ofthe Marmara Sea has also been recorded during the Holocene‘Littorina Sea’ stage from the Baltic Sea and interpreted by Yu andBerglund (2007) as the entrance of relatively warm and nutrient-richmarine water in a weak brackish-water context.
These similarities in dinocyst assemblages are likely to reflectcommon features of the hydrological evolution of two semi-enclosedseas under strong meltwater influence during the last (and penulti-mate) deglaciation, and merit further investigation in the future.
8. Conclusions
The dinocyst record presented here has a centennial-scale resolu-tion and provides the longest sequence currently available from theMarmara Sea. This continuous, high resolution record permits theevaluation of previous observations regarding the evolution of watermasses related to the connection between the Black and Mediterra-nean seas. The goodmatch between dinocyst records fromMD01-2430and two neighbouring cores has enabled the construction of aprovisional agemodel. Dinocyst assemblages include several abundant‘Paratethyan’ endemic taxa for which ecological interpretation ishampered by the paucity of recent records and modern analogues.Nevertheless, this study contributes to the advancement of ourecological knowledge of these taxa and allows for improved environ-mental interpretations (i.e. for C. rugosum).
During the LGM, the Marmara Sea was a ‘caspibrackish’ lake withmean annual SSS around 7–14 psu and very low seasonal contrasts inSSS; winter and summer SST were probably around 3–5 and 10–13 °C,respectively. This climatic and hydrological context remained effectivelyunchanged up to 20.5 cal. ka BP. A clear warming preceding a strongmeltwater influxhas beenobserved for thefirst time in theMarmara Seaas soon as ~20.5 cal. ka and around 18.1 cal. ka BP, respectively. Themeltwater episode presents a two-step history: 1) a weak phase (~20.0to 18.1 cal. ka BP) which corresponds to a possible melting of hinterlandpermafrost; 2) a strong phase with important meltwater outflow fromBlack Sea, occurring from ~18.1 to 15.9 cal. ka BP, which implies thatBlack Sea sea-level temporarily exceeded the Bosphorus sill at that time(ca Heinrich Event 1). The introduction of meltwater into the MarmaraSea led to strong SSS and SST seasonal contrasts.
During the Bølling/Allerød, the Marmara Sea was again acaspibrackish lake experiencing relatively cold conditions (with lowseasonal contrast in SST). Increased river inputs contradict the viewthat theMarmara Seamay have been an evaporative basin at that time(Hiscott et al., 2002, 2008).
Inflowing Mediterranean waters filled the Marmara Sea bottom-waters ca 12.2 cal. ka BP (approximatively the beginning of theYounger Dryas), then reached the sea-surface at ~11.7 cal. ka BP,implying a transition from ‘caspibrakish’ to marine environmentalconditions faster than the ~1.5 ka estimated by Reichel and Halbach(2007). Younger Dryas SSTs were colder than during the Bølling/Allerød, with a likely enhancement of seasonal SST contrasts.
, scale bar=20 µm.

65L. Londeix et al. / Review of Palaeobotany and Palynology 158 (2009) 52–71

66 L. Londeix et al. / Review of Palaeobotany and Palynology 158 (2009) 52–71
At the beginning of the Holocene, SSTs may have been slightlycooler than today, with minimum SSS ranging from 13 to 17.5 psu(maximum 27 psu) and likely with high seasonality in both SST andSSS (possibly up to 19 °C and 17 psu). The Marmara Sea experiencedenhanced primary productivity (perhaps due to an influx of coastalwaters) and dysoxic/anoxic conditions in the benthic zone. Compar-ison of Marmara and Black Sea dinocyst assemblages from the M1time interval do not provide evidence of an enhanced outflow fromthe Black Sea. The sequence of distinct dinocyst associations suggeststhat following the connection of the Mediterranean to the Black Sea(~8.4 14C ka BP; Ryan et al.,1997, 2003;Major et al., 2006; Hiscott et al.,2007), several distinct climatic and hydrological phases occurred,including a short cooling [~7.5 to 5.8 cal. ka BP; possibly linked to astrengthening of the Siberian High (Kotthoff et al., 2008)], a thermaloptimum (~5.8 to 4.7 cal. ka BP), and the installation of the modernhydrological system (~4.7 cal. ka BP). The timing of this sequenceappears close to that recorded in the Black Sea by Major et al. (2002)and Ryan et al. (2003), supporting the idea that the two basins wouldhave operated in tandem (Major et al., 2002).
On the whole, SST seasonality was strong throughout the wholesequence, except during the Bølling-Allerød. In contrast, SSS season-ality was weak, except during the end of the LGM, the HE1 timeinterval (particularly from ~18.1 to 15.9 cal. ka BP) and during thedeposition of the M1 sapropel. These periods of high seasonal contrastin SSS were probably linked to seasonal freshwater inputs, whichappear particularly strengthened by meltwater supply during the HE1time interval.
Quantitative estimates of past sea-surface environmental para-meters were not possible using transfer functions based on the best-analogue methods (e.g. Hutson, 1980; Guiot, 1990; de Vernal et al.,2001, 2005) because of the prevalence of non-analogue situationsregarding the composition of dinocyst assemblages. Nevertheless,transfer functions based on the mutual climatic range principle (e.g.Iversen, 1944; Fauquette et al., 1998; Pross et al., 2000; Londeix et al.,2007) might be a way to be followed with dinocysts assemblages.
Finally, it may prove rewarding to investigate further thesimilarities in dinocyst assemblages from the Marmara Sea and theHolocene or Eemian of the Baltic Sea (Brenner, 2005; Head et al.,2005; Yu and Berglund, 2007) in order to understand better thenature and timing of past deglaciations in regions strongly influencedby the decay of the Fennoscandian ice sheet.
Acknowledgements
The Marmacore Cruise was supported by the French INSU-CNRS,and the French Ministry of Foreign Affairs (MAE). Post-cruiselaboratory investigations were funded by the involved Laboratoriesassociated with CNRS Marmacore. Scientific Team is very grateful toIPEV and to Yvon Balut for the successful management of the giantpiston corer. We acknowledge the captain of the NO Marion-Dufresneand the whole crew. We are grateful to Marie-Hélène Castéra for thepreparation of palynological slides in the EPOC laboratory. The criticaland constructive comments of two anonymous reviewers greatlyimproved the manuscript; they are gratefully acknowledged.
Appendix A. Taxonomy
List of the dinoflagellate cyst taxa encountered in core MD01-2430,with some taxonomic comments:
Achomosphaera cf. andalousiense Jan du Chêne, 1977 emend Jan duChêne and Londeix, 1988; tips of the processes conformwith the typematerial, but base of most of them larger and sometime fenestrate.
Ataxiodinium choane Reid, 1974Ataxiodinium cf. choane Reid, 1974; low pericoel.Ataxiodinium sp. A; pericoel faintly developed.Bitectatodinium tepikiense Wilson, 1973
Brigantedinium cariacoense (Wall, 1967) Reid, 1977Brigantedinium simplex (Wall, 1967) Reid, 1977Brigantedinium spp.; include all round brown specimens not iden-
tified at the specific level due to poor orientation.Caspidinium rugosum Marret in Marret et al., 2004Dubridinium caperatum Reid, 1977Invertocysta? sp. A; circumcavate cyst with low parasutural ridge
and consistent openings in the outer layer.Invertocysta? sp. B; circumcavate cyst with low parasutural ridge
and a densely fenestrated outer layer.Impagidinium aculeatum (Wall, 1967) Lentin and Williams, 1981Impagidinium sp. A; elongate body with very low septa.Islandinium cf. minutum (Harland and Reid in Harland et al., 1980)
Head et al., 2001Lejeunecysta sabrina (Reid, 1977) Bujak, 1984Lingulodinium machaerophorum (Deflandre and Cookson, 1955)
Wall, 1967; numerous specimens with atypic process morphologies(with processes low, stout, bulbous tip, etc.) have also been recorded.
Nematosphaeropsis labyrinthus (Ostenfeld, 1903) Reid, 1974Operculodinium centrocarpum sensu Wall Dale, 1966; numerous
atypic forms (with processes low to very low, truncated, stout, etc.)have also been recorded.
Operculodinium israelianum (Rossignol, 1962) Wall, 1967cyst of Pentapharsodinium dalei Indelicato and Loeblich, 1986Peridinioid cyst A; proximate cyst densely covered by delicate
short processes.Polysphaeridium zoharyi (Rossignol, 1962) Bujak et al., 1980Pterocysta cruciformis Rochon et al., 2003Pyxidinopsis psilata (Wall and Dale in Wall et al., 1973) Head,
1994a; name here given only to the specimens close to the holotype.Pyxidinopsis reticulata McMinn and Sun Xuekun, 1994Pyxidinopsis? sp. A; corrugate periphragm, cyst sometime with a
cruciform shape. This allocation is used here for specimens close to‘Tectatodinium psilatum’ sensu Wall et al., 1973 (Plate II, fig. 5) thatwe considered as a separate species from Pyxidinopsis psilata sensustricto.
Quinquecuspis concreta (Reid, 1977) Harland, 1977bScrippsiella trifida Lewis, 1991Selenopemphix quanta (Bradford, 1975) Matsuoka, 1985aSelenopemphix? cf. quanta; processes slender and longer than for
Selenopemphix quanta sensu stricto.Seriliodinium explicatum Eaton, 1996Spiniferites belerius Reid, 1974Spiniferites bulloides (Deflandre & Cookson, 1955) Sarjeant, 1970Spiniferites bentorii (Rossignol, 1964) Wall & Dale, 1970Spiniferites cruciformisWall and Dale inWall et al., 1973; specimens
of S. cruciformis show very variable development of the periphragmand processes (Wall et al., 1973; Mudie et al., 2001, 2004), from theholotype morphology to specimens with detached endocyst and/orextremely reduced processes (cf. Mudie et al., 2001, 2004). Thetentatives to link these morphotypes to SSS and/or SST trends lead toany significant result (Mudie et al., 2004; this work).
Spiniferites delicatus Reid, 1974Spiniferites elongatus Reid, 1974Spiniferites ‘granular’; because the original descriptions of granu-
lar Spiniferites taxa are generally inaccurate, the very few speci-mens encountered in MD01-2430 core are grouped as Spiniferites‘granular’.
Spiniferites hyperacanthus (Deflandre & Cookson) Reid, 1974Spiniferites membranaceus (Rossignol, 1964) Sarjeant, 1970Spiniferites mirabilis (Rossignol, 1964) Sarjeant, 1970Spiniferites ramosus multiplicatus (Rossignol, 1964) Lentin &
Williams, 1973; in S. ramosus multiplicatus intergonal processes arenot systematically present on each sutural ridge, since they are inS. hyperacanthus. That is why the synonymisation of these two taxaproposed by Matsuoka (1985) is not followed here.

67L. Londeix et al. / Review of Palaeobotany and Palynology 158 (2009) 52–71
Spiniferites ramosus ramosus (Ehrenberg, 1838) Mantell, 1854Spiniferites septentrionalis Harland, 1977b; Achomosphaera anda-
lousiense sensu stricto has a spherical, psilate central body without
Plate IV. Dinoflagellate cysts from the late Quaternary of Marmara Sea. Same magnification
1. Operculodinium centrocarpum centrocarpum sensu Wall and Dale (1966); a —
2. Operculodinium centrocarpum forma 1, with aberrant processes; a — ventral v3. Operculodinium centrocarpum, very short processes, 290 cm.4. Operculodinium centrocarpum, short processes, 290 cm.5. Tectatodinium pellitum, 280 cm.6. Bitectatodinium tepikiense, 230 cm.7. Operculodinium centrocarpum forma 2, with aberrant processes; a — dorsal vie8. Nematosphaeropsis labyrinthus, 230 cm.9. Lingulodinium machaerophorum, 560 cm.
septa, when Spiniferites septentrionalis has a scabrate ovoid centralbody with faint ridge or septa. In addition, processes tips aretrabeculate in A. andalousiense and somewhat fenestrate in S.
, scale bar=20 µm (except for 1b, 2b, 7b; scale bar=10 µm).
dorsal view, b — magnification of processes, 230 cm.iew, b — magnification of stout processes, 230 cm.
w, b — magnification of stout and sometime bifid processes, 290 cm.

Plate V. Algae from the late Quaternary of Marmara Sea. Same magnification, scale bar=20 µm.
1. Multiplicasphaeridium-type, two arms, 560 cm.1. Multiplicasphaeridium-type, three arms, 560 cm.1. Multiplicasphaeridium-type, four arms, 560 cm.1. Pediastrum indet., 350 cm.
68 L. Londeix et al. / Review of Palaeobotany and Palynology 158 (2009) 52–71
sptentrionalis. For these reasons, the synonymisation of these two taxaproposed by Jean du Chêne and Londeix (1988) is not followed here.
Some Spiniferites specimens with very atypic morphologies (failedprocesses, presence of some faint trabeculae, development of a velum,etc.) have been encountered; it seemed vain to determinate them atthe species level.
Stelladinium stellatum (Wall and Dale, 1968) Reid, 1977Tectatodinium pellitum Wall, 1967Tuberculodinium vancampoae (Rossignol, 1962) Wall, 1967Xandarodinium xanthum Reid, 1977In addition, the following new combination is proposed:Impagidinium inaequalis (Wall and Dale in Wall et al., 1973) comb.
nov.=Spiniferites inaequalisWall and Dale inWall et al., 1973 (Plate III,fig. 7–8; Plates IV and V).
References
Aksu, A.E., Yaşar, D., Mudie, P.J., Gillespie, H., 1995. Late glacial–Holocene paleoclimaticand paleoceanographic evolution of the Aegean Sea: micropaleontological andstable isotopic evidence. Marine Micropaleontology 25, 1–28.
Aksu, A.E., Hiscott, R.N., Yasar, D., 1999. Oscillating Quaternary water levels of theMarmara Sea and vigorous outflow into the Aegean Sea from the Marmara Sea–Black Sea drainage corridor. Marine Geology 153 (1–4), 275–302.
Aksu, A.E., Hiscott, R.N., Yasar, D., Işler, F.I., Marsh, S., 2002a. Seismic stratigraphy ofLate Quaternary deposits from the southwestern Black Sea shelf: evidence fornon-catastrophic variations in sea-level during the last ~10000 yr. Marine Geology190 (1–2), 61–94.
Aksu,A.E., Hiscott, R.N., Kaminski,M.A.,Mudie, P.J., Gillespie, H., Abrajano, T., Yasar, D., 2002b.Last glacial–Holocene paleoceanography of the Black Sea and Marmara Sea: stableisotopic, foraminiferal and coccolith evidence. Marine Geology 190 (1–2), 119–149.
Aksu, A.E., Hiscott, R.N., Mudie, P.J., Rochon, A., Kaminski, M.A., Abrajano, T., Yaşar, D.,2002c. Persistent Holocene outflow from the Black sea to the Eastern Mediterra-nean contradicts Noah's flood hypothesis. GSA Today 4–10.
Alavi, S.N., 1988. Late Holocene deep-sea benthic foraminifera from the Sea of Marmara.Marine Micropaleontology 13 (3), 213–237.
Alley, R.B., Àgústsdóttir, A.M., 2005. The 8 k event: cause and consequences of amajor Holocene abrupt climate change. Quaternary Science Reviews 24 (10–11),1123–1149.
Alley, R.B., Brook, E.J., Anandakrishnan, S., 2002. A northern lead in the orbital band: north–south phasing of Ice-Age events. Quaternary Science Reviews 21 (1–3), 431–441.
Atanassova, J., 2005. Palaeoecological setting of the western Black Sea area during thelast 15000 years. The Holocene 15 (4), 576–584.
Bahr, A., Lamy, F., Arz, H., Kuhlmann, H., Wefer, G., 2005. Late glacial to Holoceneclimate and sedimentation history in the NW Black Sea. Marine Geology 214 (4),309–322.
Bahr, A., Arz, H.W., Lamy, F., Wefer, G., 2006. Late glacial to Holocene paleo-environmental evolution of the Black Sea, reconstructed with stable oxygenisotope records obtained on ostracod shells. Earth and Planetary Science Letters241 (3–4), 863–875.
Bahr, A., Lamy, F., Arz, H.W., Major, C., Kwiecien, O., Wefer, G., 2008. Abrupt changes oftemperature and water chemistry in the late Pleistocene and early Holocene BlackSea. Geochemistry, Geophysics, Geosystems 9 (1), 1–16.

69L. Londeix et al. / Review of Palaeobotany and Palynology 158 (2009) 52–71
Bard, E., Rosteck, F., Turon, J.-L., Gendreau, S., 2000. Hydrological impact of Heinrichevents in the subtropical Northeast Atlantic. Science 289, 1321–1324.
Beck, C., Mercier de Lépinay, B., Schneider, J.-L., Cremer, M., Çağatay, N., Wendenbaum,E., Boutareaud, S., Ménot-Combes, G., Schmidt, S., Weber, O., Eris, K., Armijo, R.,Meyer, B., Pondard, N., Marmacore Cruise Party, Turon J.-L., Labeyrie L., Cortijo E.,Gallet Y., Bouquerel H., Gorur N., Gervais A., Castera M.-H., Londeix L., de RességuierA. & Jaouen A., 2007. Late quaternary co-seismic sedimentation in the Sea ofMarmara's deep basins. Sedimentary Geology 199 (1–2), 65–89.
Boalche, G.T., Parke, M., 1971. The prasinophycean genera (Chorophyta) possibly relatedto fossil genera, in particular the genus Tasmanides. Proceeding of the 2nd PlanktonConference, vol. 1. Roma, pp. 99–105. 1970.
Borisova, O., Sidorchuk, A., Panin, A., 2006. Palaeohydrology of the Seim River basin,Mid-Russian Upland, based on palaeochannel morphology and palynological data.Catena 66 (1–2), 53–73.
Bottema, S., 1995. The Younger Dryas in the eastern Mediterranean. Quaternary ScienceReviews 14, 883–891.
Bottema, S., Woldring, H., Aytugfl, B., 1995. Late Quaternary vegetation history ofNorthern Turkey. Paleohistoria 17, 53–143.
Bourrelly, P., Couté, A., 1980. Trois dinophycées d'eau douce observées en microscopie àbalayage. Cryptogramie. Algologie 1, 161–169.
Bradford, M.R., Wall, D.A., 1984. The distribution of Recent organic-walled dinoflagellatecysts in thePersianGulf, Gulf ofOman,andnorthwesternArabian Sea. PaleontographicaAbt. B, vol. 192, pp. 16–84.
Brenner, W.W., 2001a. Distribution of organic walled microfossils within single laminaefrom Gotland Basin and their environmental evidence. Baltica 14, 34–39.
Brenner, W.W., 2001b. Organic walled microfossils from the central Baltic Sea,indicators of environmental change and base for ecostratigraphic correlation.Baltica 14, 40–51.
Brenner,W.W., 2005. Holocene environmental historyof theGotlandBasin (Baltic Sea)— amicropalaeontological model. Palaeogeography, Palaeoclimatology, Palaeoecology220, 227–241.
Çağatay, M.N., Algan, O., Sakinç, M., Eastoe, C.J., Egesel, L., Balkis, N., Ongan, D., Caner, H.,1999. Amid-late Holocene sapropelic sediment unit from the southernMarmara seashelf and its palaeoceanographic significance. Quaternary Science Reviews 18 (4–5),531–540.
Çağatay, M.N., Görür, N., Algan, O., Eastoe, C., Tchapalyga, A., Ongan, D., Kuhn, T., Kuşcu,I., 2000. Late Glacial–Holocene palaeoceanography of the Sea of Marmara: timing ofconnections with the Mediterranean and the Black Seas. Marine Geology 167 (3–4),191–206.
Caner, H., Algan, O., 2002. Palynology of sapropelic layers from theMarmara Sea. MarineGeology 190 (1–2), 35–46.
Cordova, C.E., Lehman, P.H., 2005. Holocene environmental change in southwesternCrimea (Ukraine) in pollen and soil records. The Holocene 15, 263–277.
Dale, B., 1996. Dinoflagellate cyst ecology: modeling and geological applications. In:Jansonius, J., McGregor, D.C. (Eds.), Palynology: Principles and Applications, vol. 3.AASP Foundation, Salt Lake City, UT, pp. 1249–1275.
de Vernal, A., Mudie, P.J., 1989. Late Pliocene to Holocene palynostratigraphy at ODP645, Baffin Bay. In: Srivastava, S.P., Arthur, M., Clement, B., et al. (Eds.), Proceedingsof the Ocean Drilling Program: Scientific Results, vol. 105, pp. 387–399.
de Vernal, A., Goyette, C., Rodriques, C.G., 1989. Contribution palynostratigraphique(dinokystes, pollen et spores) à la mer de Champlain: coupe de Saint-CésaireQuébec. Canadian Journal of Earth Sciences 26, 2450–2464.
de Vernal, A., Londeix, L., Mudie, P., Harland, R., Morzadec-Kerfourn, M.-T., Turon, J.-L.,Wrenn, J.H., 1992. Quaternary organic-walled dinoflagellate cysts of the North AtlanticOceanandadjacent seas: ecostratigraphyandbiostratigraphy. In:Head,M.J.,Wrenn, J.H.(Eds.), Neogene and Quaternary Dinoflagellate Cysts and Acritarchs. AmericanAssociation of Stratigraphic Palynologists Foundation, Dallas, pp. 289–328.
de Vernal, A., Rochon, A., Turon, J.-L., Matthiessen, J., 1998. Organic-walled dino-flagellate cysts: palynological tracers of sea-surface conditions in middle to highlatitude marine environments. Geobios 30, 905–920.
de Vernal, A., Henry, M., Matthiessen, J., Mudie, P.J., Rochon, A., Boessenkool, K.P.,Eynaud, F., GrQsfjeld, K., Guiot, J., Hamel, D., Harland, R., Head, M.J., Kunz-Pirrung,M., Levac, E., Loucheur, V., Peyron, O., Pospelova, V., Radi, T., Turon, J.-L., Voronina, E.,2001. Dinoflagellate cyst assemblages as tracers of sea-surface conditions in thenorthern North Atlantic, Arctic and sub-Arctic seas: the new n=677 data base andits application for quantitative paleoceanographic reconstruction. Journal ofQuaternary Science 16, 681–698.
de Vernal, A., Eynaud, F., Henry, M., Hillaire-Marcel, C., Londeix, L., Mangin, S.,Matthiessen, J., Marret, F., Radi, T., Rochon, A., Solignac, S., Turon, J.L., 2005.Reconstruction of sea-surface conditions at middle to high latitudes of the NorthernHemisphere during the last glacial maximum (LGM) based on dinoflagellate cystassemblages. Quaternary Science Reviews 24, 897–924.
Denton, G.H., Heusser, C.J., Lowell, T.V., Moreno, P.I., Anderson, B.G., Heusser, L.E.,Schlüchter, C., Marchant, D.R., 1999. Interhemispheric linkage of paleoclimateduring the last glaciation. Geografiska Annaler 81, 107–153.
Devillers, R., de Vernal, A., 2000. Distribution of dinoflagellate cysts in surfacesediments of the North Atlantic in relation to nutrient content and productivityin surface waters. Marine Geology 166, 103–124.
Dobiński, W., 2005. Permafrost of the Carpathian and Balkan Mountains, eastern andsoutheastern Europe. Permafrost and Periglacial Processes 16 (4), 395–398.
Eaton, G.L., 1996. Seriliodinium, a new Late Cenozoic dinoflagellate from the Black Sea.Review of Palaeobotany and Palynology 91, 151–169.
Edwards, L.E., Andrle, A.S., 1992. Distribution of selected dinoflagellate cysts in modernmarine sediments. In: Head, M.J., Wrenn, L.H. (Eds.), Neogene and QuaternaryDinoflagellate Cysts and Acritarchs. American Association of Stratigraphic Palynol-ogists Foundation, Dallas, TX, pp. 259–288.
Ehlers, J., Gibbard, P.L. (Eds.), 2004. Quaternary Glaciations: Extent and Chronology. In:Europe. Developments in Quaternary Science, vol. 1. Elsevier, Oxford.
Ellegaard, M., 2000. Variations in dinoflagellate cyst morphology under conditions ofchanging salinity during the last 2000 years in the Limfjord, Denmark. Review ofPalaeobotany and Palynology 109, 65–81.
Elliot, M., Labeyrie, L., Bond, G., Cortijo, E., Turon, J.-L., Tisnerat, N., Duplessy, J.-C., 1998.Millenial-scale iceberg discharges in the Irminger Basin during the last glacial period:relationship with the Heinrich events and environmental settings. Paleoceanography13 (5), 433–446.
Eriksen, U., Friedrich, W.L., Buchardt, B., Tauber, H., Thomson, M.S., 1990. The StronghyleCaldera: geological, palaeontological and stable isotope evidence from radiocarbondated stromatolites from Santorini. In: Hardy, D.A., et al. (Ed.), Thera and theAegean World III. In: Earth Sciences, vol. 2. Thera Foundation, London, pp. 139–150.
Eriş, K.K., Ryan, W.B.F., Çağatay, M.N., Sancar, U., Lericolais, G., Ménot, G., Bard, E., 2007.The timing and evolution of the post-glacial transgression across the Sea ofMarmara shelf south of Istanbul. Marine Geology 243, 57–76.
Evans, G., Erten, H., Alavi, S.N., Von Gunten, H.R., Ergin, M., 1989. Superficial deep-watersediments of the eastern Marmara basin. Geo-Marine Letters 9, 27–36.
Evitt, W.R., Gocht, H., Netzel, H., 1985. Gonyaulax cysts from lake Zürich sediments.Review of Palaeobotany and Palynology 45, 35–46.
Eynaud, F., Turon, J.L., Duprat, J., 2004. Comparison of the Holocene and Eemianpalaeoenvironments in the South-Icelandic basin: dinoflagellate cysts as proxies forthe North Atlantic surface circulation. Review of Palaeobotany and Palynology 128,55–79.
Eynaud, F., Zaragosi, S., Scourse, J.D., Mojtahid, M., Bourillet, J.F., Hall, I.R., Penaud, A.,Locascio, M., Reijonen, A., 2007. Deglacial laminated facies on the NW Europeancontinental margin: the hydrographic significance of British–Irish Ice Sheetdeglaciation and Fleuve Manche paleoriver discharges. Geochemistry GeophysicsGeosystems 8 (6), 1–19.
Fauquette, S., Guiot, J., Suc, J.-P., 1998. A method for climatic reconstruction of theMediterranean Pliocene using pollen data. Palaeogeography, Palaeoclimatology,Palaeoecology 144, 183–201.
Fensome, R.A., Williams, G.L., 2004. The Lentin andWilliams Index of fossil dinoflagellates2004 edition. AASP Contributions Series, vol. 42. 909 pp.
Filipova-Marinova, M., Christova, R., Bozilova, E., 2004. Palaeoeological conditions in theBulgarian Black Sea area during the Quaternary. Journal of EnvironmentalMicropaleontology, Microbiology and Meiobenthology 1, 135–154.
Goodman, D.K., 1979. Dinoflagellate “communities” from the Lower Eocene NanjemoyFormation of Maryland, U.S.A. Palynology 3, 169–190.
Grøsfjeld, K., Harland, R., 2001. Distribution of modern dinoflagellate cysts from inshoreareas along the coast of southern Norway. Journal of Quaternary Science 16, 651–659.
Grosswald,M.G.,1980. LateWeichselian ice sheet of Northern Eurasia. QuaternaryResearch13 (1), 1–32.
Grosswald, M.G., 1998. Late-Weichselian ice sheets in Arctic and Pacific Siberia.Quaternary International 45–46, 3–18.
Grosswald, M.G., Hughes, T.J., 2002. The Russian component of an Arctic ice sheetduring the Last Glacial Maximum. Quaternary Science Reviews 21, 121–146.
Guiot, J., 1990. Methodology of paleoclimatic reconstruction from pollen in France.Palaeogeography, Palaeoclimatology, Palaeoecology 80, 49–69.
Gutzwiller, O., 1923. Beitrage zur Geologie der Umgebung von Merfete (MuÅs refte) amMarmara Meer. Basel University, Basel. 25 pp.
Head, M.J., Seidenkrantz, M.-S., Janczyk-Kopikowa, Z., Marks, L., Gibbard a, P.L., 2005.Last Interglacial (Eemian) hydrographic conditions in the southeastern Baltic Sea,NE Europe, based on dinoflagellate cysts. Quaternary International 130, 3–30.
Hickel, B., Pollingher, U., 1986. On the morphology and ecology of Gonyaulax apiculata(Penard) Entz from the Selenter See (West Germany). Archiv fuer HydrobiologieSupplement 73 (2), 227–232 (Algological Studies 43).
Hiscott, R.N., Aksu, A.E., 2002. Late Quaternary history of theMarmara Sea and Black Seafrom high-resolution seismic and gravity-core studies. Marine Geology 190 (1–2),261–282.
Hiscott, R.N., Aksu, A.E., Yaşar, D., Kaminski,M.A.,Mudie, P.J., Kostylev, V.E.,MacDonald, J.C.,Işler, F.I., Lord, A.R., 2002. Deltas south of the Bosphorus Strait record persistent BlackSea outflow to the Marmara Sea since ~10 ka. Marine Geology 190 (1–2), 95–118.
Hiscott, R.N., Aksu, A.E., Mudie, P.J., Kaminski, M.A., Abrajano, T., Yaşar, D., Rochon, A.,2007. The Marmara Sea Gateway since ~16 ky BP: non-catastrophic causesof paleoceanographic events in the Black Sea at 8.4 and 7.15 ky BP. In: Yanko-Hombach, V., Gilbert, A.S., Panin, N., Dolukhanov, P.M. (Eds.), The Black SeaFlood Question: Changes in Coastline, Climate, and Human Settlement. Springer,pp. 89–117.
Hiscott, R.N., Aksu, A.E., Mudie, P.J., 2008. Comment on “The timing and evolution of thepost-glacial transgression across the Sea of Marmara shelf south of İstanbul” by Erişet al., Marine Geology 243, 57–76. Marine Geology 248, 228–236.
Huhmann, M., Kremenetski, K.V., Hiller, A., Bruckner, H., 2004. Late Quaternarylandscape evolution of the upper Dnister valley, western Ukraine. Palaeogeography,Palaeoclimatalogy, Palaeoecology 209, 51–71.
Hutson,W.H., 1980. The Agulhas current during the late Pleistocene: analysis of modernfaunal analogs. Science 207, 64–66.
Iversen, J., 1944. Viscum, Hedera and Ilex as climate indicators. A contribution to thestudy of the Postglacial temperature climate. Geologiska Föreningens i StockholmFörhandlingar 66, 463–483.
Jacquet, S., Paolini, G., Lazzarotto, J., Perga, M., Domaizon, I., Druart, J.-C., Humbert, J.-F.,Perney, P., Girel, C., Laine, L., 2007. Suivi de la qualité des eaux du lac du Bourgetpour l'année 2006. Rapport 2007. 110 pp.
Jan du Chêne, R., Londeix, L., 1988. Données nouvelles sur Achomosphaera andalousienseJan du Chêne 1977, kyste de dinoflagellé fossile. Bulletin des Centres de RecherchesExploration-Production Elf-Aquitaine 12 (1), 237–250.

70 L. Londeix et al. / Review of Palaeobotany and Palynology 158 (2009) 52–71
Johnsen, S.J., Dahl-Jensen, D., Gundestrup, N., Steffensen, J.P., Clausen, H.B., Miller, H.,Masson-Delmotte, V., Sveinbjörnsdottir, A.E., White, J., 2001. Oxygen isotope andpalaeotemperature records from six Greenland ice-core stations: Camp Century,Dye-3, GRIP, GISP2, Renland and NorthGRIP. Journal of Quaternary Science 16 (4),299–307.
Jones, G.A., Keigwin, L.D., 1988. Evidence from Fram Strait (78° N) for early deglaciation.Nature 336, 56–59.
Kalicki, T., Sanko, A.F., 1998. Palaeohydrological changes in the Upper Dneper Valley,Belarus, during the last 20,000 years. In: Benito, G., et al. (Ed.), Palaeohydrology andEnvironmental Change. Wiley, Chichester, England, pp. 125–135.
Kaminski, M.A., Aksu, A., Box, M., Hiscott, R.N., Filipescu, S., Al-Salameen, M., 2002. LateGlacial to Holocene benthic foraminifera in the Marmara Sea: implications for BlackSea–Mediterranean Sea connections following the last deglaciation.Marine Geology190 (1–2), 165–202.
Knies, J., Vogt, C., Matthiessen, J., Nam, S.-I., Ottesen, D., Rise, L., Bargel, T., Eilertsen, R.S.,2007. Re-advance of the Fennoscandian Ice Sheet during Heinrich Event 1. MarineGeology 240 (1–4), 1–18.
Konikov, E., Likhodedova, O., Pedan, G., 2007. Paleogeographic reconstructions of sea-level change and coastline migration on the northwestern Black Sea shelf over thepast 18 kyr. Quaternary International 167–168, 49–60.
Koral, H., 2007. Modes, rates and geomorphological consequences of active tectonics inthe Marmara Region, NW Turkey—a critical overview based on seismotectonic fieldobservations. Quaternary International 167–168, 149–161.
Kotthoff, U., Pross, J., Müller, U.C., Peyron, O., Schmiedl, G., Schulz, H., Bordon, A., 2008.Climate dynamics in the borderlands of the Aegean Sea during formation ofsapropel S1 deduced from a marine pollen record. Quaternary Science Reviews 27,832–845.
Kouli, K., Brinkhuis, H., Dale, B., 2001. Spiniferites cruciformis: a fresh water dinoflagellatecyst? Review of Palaeobotany and Palynology 113 (4), 273–286.
Lane-Serff, G., Rohling, E., Bryden, H., Charnock, H., 1997. Postglacial connection of theBlack Sea to the Mediterranean and its relation to the timing of sapropel formation.Paleoceanography 12 (2), 169–174.
Le Pichon, X., Sengör, A.M.C., Demirbag, E., Rangin, C., Imren, C., Armijo, R., Görür, N.,Çağatay, N., Mercier de Lepinay, B., Meyer, B., Saatcilar, R., Tok, B., 2001. The activeMain Marmara Fault. Earth and Planetary Science Letters 192, 595–616.
Leroy, V., 2001. Traceurs palynologiques des flux biogéniques et des conditionshydrographiques en milieu marin côtier: exemple de l'étang de Berre. DEAdissertation, Géosciences de l'Environnement, Université Aix-Marseille. 31 pp.
Leroy, S.A.G., Marret, F., Gibert, E., Chalié, F., Reyss, J.-L., Arpe, K., 2007. River inflow andsalinity changes in the Caspian Sea during the last 5500 years. Quaternary ScienceReviews 26 (25–28), 3359–3383.
Lewis, J., Dodge, J.D., Powell, A.J., 1990. Quaternary dinoflagellate cysts from theupwelling system offshore Peru, Hole 686B, ODP Leg 112. Proceeding of the ODPScientific Results 112, 323–327.
Londeix, L., Benzakour, M., Suc, J.-P., Turon, J.-L., 2007. Messinian palaeoenvironmentsand hydrology in Sicily (Italy): the dinoflagellate cyst record. Geobios 40, 233–250.
Major, C., Ryan, W., Lericolais, G., Hajdas, I., 2002. Constraints on Black Sea outflow tothe Sea of Marmara during the last glacial–interglacial transition. Marine Geology190 (1–2), 19–34.
Major, C.O., Goldstein, S.L., Ryan,W.B.F., Lericolais, G., Piotrowski, A.M., Hajdas, I., 2006. Theco-evolution of Black Sea level and composition through the last deglaciation and itspaleoclimatic significance. Quaternary Science Reviews 25 (17–18), 2031–2047.
Marino, G., 2008. Palaeoceanography of the interglacial eastern Mediterranean Sea. Ph.D. Utrecht University, LPP Contributions Series, 24: 145 p.
Marret, F., 1993. Les effets de l'acétolyse sur les assemblages des kystes de dinoflagellés.Palynosciences 2, 267–272.
Marret, F., 1994. Distribution of dinoflagellate cysts in recentmarine sediments from theeast Equatorial Atlantic (Gulf of Guinea). Review of Palaeobotany and Palynology84, 1–22.
Marret, F., de Vernal, A.,1997. Dinoflagellate cyst distribution in surface sediments of thesouthern Indian Ocean. Marine Micropaleontology 29, 367–392.
Marret, F., Zonneveld, K.A.F., 2003. Atlas of modern organic-walled dinoflagellate cystdistribution. Review of Palaeobotany and Palynology 125, 1–200.
Marret, F., Leroy, S., Chalie, F., Gasse, F., 2004. New organic-walled dinoflagellate cystsfrom recent sediments of Central Asian seas. Reviewof Palaeobotany and Palynology129 (1–2), 1–20.
Marret, F.,Mudie, P., Aksu, A., Richard,N., Hiscott, R.N., 2009. AHolocene dinocyst record ofa two-step transformation of the Neoeuxinian brackish water lake into the Black Sea.Quaternary International 197 (1-2), 72–86.
Martínez-Hernández, E., Hernández-Campos, H.E., 1991. Distribución de quistes dedinoflagelados y acritarcas en sedimentos Holocénicos del Golfo de California.Paleontologia Mexicana 57, 1–133.
Matsuoka, K., 1985. Organic-walled dinoflagellate cysts from surface sediments ofNagasaki Bay and Senzaki Bay, west Japan. Faculty of Liberal Arts, NagasakiUniversity, Natural Science, Bulletin 25 (2), 21–115.
Matsuoka, K., 1987. Organic-walled dinoflagellate cysts from surface sediments ofAkkeshi Bay and Lake Saroma, North Japan. Faculty of Liberal Arts, NagasakiUniversity, Natural Science, Bulletin 28, 35–123.
Matthiessen, J., 1995. Distribution patterns of dinoflagellate cysts and other organic-walledmicrofossils in recent Norwegian–Greenland Sea sediments. Marine Micropaleontol-ogy 24, 307–334.
Matthiessen, J., Brenner, W., 1996. Chlorococcalalgen und Dinoflagellaten–Zysten inrezenten Sedimenten des Greifswalder Boddens (sqdliche Ostsee). SenckenbergianaMaritima 27, 33–48.
McMinn, A.,1991. Recent dinoÅíagellate cysts from estuaries on the central coast of NewSouth Wales, Australia. Micropaleontology 37, 269–287.
Ménot, G., Bard, E., Rostek, F.,Weijers, J.W.H., E.C., H., Schouten, S., SinningheDamsté, J.S.,2006. Early reactivation of European Rivers during the last deglaciation. Science 313,1623–1625.
Meriç, E., Algan, O., 2007. Paleoenvironments of the Marmara Sea (Turkey) Coasts frompaleontological and sedimentological data. Quaternary International 167–168,128–148.
Mudie, P.J., 1992. Circum-arctic Quaternary and Neogene marine palynofloras:paleoecology and statistical analysis. In: Head, M.J., Wrenn, L.H. (Eds.), Neogeneand Quaternary Dinoflagellate Cysts and Acritarchs, vol. X. American Association ofStratigraphic Palynologists Foundation, Dallas, pp. 347–390.
Mudie, P.J., Aksu, A.E., Yas-ar, D., 2001. Late Quaternary dinoflagellate cysts from theBlack, Marmara and Aegean seas: variations in assemblages, morphology andpaleosalinity. Marine Micropalaeontology 43, 155–178.
Mudie, P.J., Rochon, A., Aksu, A.E., Gillespie, H., 2002a. Dinoflagellate cysts andfreshwater algae and fungal spores as salinity indicators in Late Quaternary coresfrom Marmara and Black Seas. Marine Geology 190, 203–231.
Mudie, P.J., Rochon, A., Aksu, A.E., 2002b. Pollen stratigraphy of Late Quaternary coresfromMarmara Sea: land–sea correlation and paleoclimatic history. Marine Geology190, 233–260.
Mudie, P.J., Rochon, A., Aksu, A.E., Gillespie, H., 2004. Late glacial, Holocene and moderndinoflagellate cyst assemblages in the Aegean–Marmara–Black Sea corridor:statistical analysis and re-interpretation of the early Holocene Noah's Floodhypothesis. Review of Palaeobotany and Palynology 128, 143–167.
Mudie, P.J., Marret, F., Aksu, A.E., Hiscott, R.N., Gillespie, H., 2007. Palynological evidencefor climatic change, anthropogenic activity and outflow of Black Sea water duringthe late Pleistocene and Holocene: centennial- to decadal-scale records from theBlack and Marmara Seas. Quaternary International 167–168, 73–90.
Nehring, S., 1994. Spatial distribution of dinoflagellate resting cysts in Recent sedimentsof Kiel Bight Germany (Baltic Sea). Ophelia 39, 137–158.
Nehring, S., 1997. Dinoflagellate resting cysts from Recent German coastal sediments.Botanica Marina 40, 307–324.
Nevesskii, Y.N., 1967. Sedimentation Processes in the Nearshore Zone. Nauka, Moscow.255 pp., in Russian.
Özsoy, E., Latif, M.A., Tuğrul, S., Ünlüsata, Ü., 1995. Ex-changes with the MediterraneanÅíuxes, and boundary mixing processes in the Black Sea. Bulletin de l'InstitutOcéanographique de Monaco, Spec., vol. 15, pp. 1–26.
Pailler, D., Bard, E., 2002. High frequency palaeoceanographic changes during the past140 000 yr recorded by the organic matter in sediments of the Iberian Margin.Palaeogeography, Palaeoclimatology, Palaeoecology 181 (4), 431–452.
Pénard, E., 1891. Les Péridiniacées du Leman. Bulletin de la Société Botanique de Genève6, 1–72.
Penaud, A., Eynaud, F., Turon, J.-L., Zaragosi, S., Marret, F., Bourillet, J.-F., 2008.Interglacial variability (MIS 5 and MIS 7) and dinoflagellate cyst assemblages in theBay of Biscay (North Atlantic). Marine Micropaleontology 68 (1–2), 136–155.
Pichler, H., Friedrich, W., 1976. Radiocarbon dates of Santorini volcanics. Nature 262,373–374.
Pirazzoli, P.A., 1996. Chapter Four: Deglacial sea-level changes, Sea-level changes: thelast 20 000 years. John Wiley, Chichester, pp. 81–99.
Polat, C.M., Tuğrul, S., 1996. Chemical exchange between the Mediterranean and BlackSea via the Turkish Straits. In: Briand, F. (Ed.), Dynamics of Mediterranean Straitsand Channels: Bulletin de l'Institut Océanographique de Monaco, Spec. No. 17,CIESME Sci. Ser., vol. 2, pp. 167–186.
Popescu, S.-M., 2003. Holocene Black Sea Environments according to Palynology, SeattleAnnual Meeting. Geological Society of America, Seattle, p. 462.
Pospelova, V., Chmura, G.L., Boothman, W.S., Latimer, J.S., 2005. Spatial distribution ofmodern dinoflagellate cysts in polluted estuarine sediments from Buzzards Bay(Massachusetts, USA) embayments. Marine Ecology. Progress Series 292, 23–40.
Pross, J., Klotz, S., Mosbrugger, V., 2000. Reconstructing palaeotemperatures for theEarly and Middle Pleistocene using the mutual climatic range method based onplant fossils. Quaternary Science Reviews 19, 1785–1799.
Radi, T., de Vernal, A., 2004. Dinocyst distribution in surface sediments from thenortheastern Pacific margin (40–60°N) in relation to hydrographic conditions,productivity and upwelling. Review of Palaeobotany Palynology 128, 169–193.
Reichel, T., Halbach, P., 2007. An authigenic calcite layer in the sediments of the Sea ofMarmara—a geochemical marker horizon with paleoceanographic significance.Deep Sea Research. Part 2. Topical Studies in Oceanography 54, 1201–1215.
Roberts, N., Jones, M.D., Benkaddour, A., Eastwood,W.J., Filippi, M.L., Frogley, M.R., Lamb,H.F., Leng,M.J., Reed, J.M., Stein,M., Stevens, L., Valero-Garcés, B., Zanchetta, G., 2008.Stable isotope records of Late Quaternary climate and hydrology from Mediterra-nean lakes: the ISOMED synthesis. Quaternary Science Reviews 27, 2426–2441.
Rochon, A., de Vernal, A., Turon, J.L., Matthiessen, J., Head, M.J., 1999. Distribution ofrecent dinoflagellate cysts in surface sediments from the North Atlantic Ocean andadjacent seas in relation to sea-surface parameters. American Assocociation ofStratigraphic Palynologist. AASP Control Series 35, 1–152.
Rochon, A., Mudie, P.J., Aksu, A.E., Gillespie, H., 2002. Pterocysta gen. nov.: a newdinoflagellate cyst from Pleistocene glacialstage sediments of the Black andMarmara seas. Palynology 26, 95–105.
Rohling, E.J., Pälike, H., 2005. Centennial-scale climate cooling with a sudden cold eventaround 8,200 years ago. Nature 434, 975–979.
Ross, D.A., Degens, E.T., MacIlvaine, J., 1970. Black Sea: recent sedimentary history.Science 170, 163–165.
Ryan, W.B.F., 1972. The stratigraphy of late Quaternary sediments in the easternMediterranean. In: Stanley, D.J. (Ed.), The Mediterranean Sea: A Natural Laboratory.Dowden, Hutchinson, Ross, Stroudsbourg, PA, pp. 149–169.
Ryan, W.B.F., Pitman III, W.C., 1999. Noah's Flood: the New Scientific Discoveries aboutEvents that Changed History. Simon and Schuster, New York. 319 pp.

71L. Londeix et al. / Review of Palaeobotany and Palynology 158 (2009) 52–71
Ryan, W.B.F., Pitman, W.C., Major, C.O., Shimkus, K., Moskalenko, V., Jones, G.A.,Dimitrov, P., Gorur, N., Sakinc, M., Yuce, H., 1997. An abrupt drowning of the BlackSea shelf. Marine Geology 138 (1–2), 119–126.
Ryan, W.B.F., Major, C.O., Lericolais, G., Goldstein, S.L., 2003. Catastrophic flooding of theBlack Sea. Annual Review of Earth and Planetary Sciences 31, 525–554.
Schrank, E., 1988. Effects of chemical processing on the preservation of peridinoiddinoflagellates: a case from the Late Cretaceous of NE Africa. Review ofPalaeobotany and Palynology 56, 123–140.
Sejrup, H.P., Larsen, E., Landvik, J., King, E.L., Haflidason, H., Nesje, A., 2000. Quaternaryglaciations in southern Fennoscandia: evidence from southwestern Norway and thenorthern North Sea region. Quaternary Science Reviews 19, 667–685.
Serebryanny, L.R., 1982. Postglacial Black Sea coast fluctuations and their comparisonwith the glacial history of the Caucasian highmountain region. In: Kaplin, P.A., Klige,R.K., Chepalyga, A.L. (Eds.), Sea and Oceanic Level Fluctuations for 15000 years (inRussian). Nauka, Moscow, p. 161-167.
Shannon, C.F., Weaver,W.,1949. Themathematical theory of communication. Universityof Illinois Press, Urbana. 322 pp.
Shmuratko, V.I., 2007. The post-glacial transgression of the Black Sea. In: Yanko-Hombach,V., Gilbert, A.S., Panin, N., Dolukhanov, P.M. (Eds.), The Black Sea Flood Question:Changes in Coastline, Climate, and Human Settlement. Springer, pp. 221–250.
Smith, A.D., Taymaz, T., Oktay, F.Y., Yüce, H., Alpar, B., Başaran, H., Jackson, J.A., Kara, S.,Şimşek, M., 1995. High-resolution seismic profiling in the Sea of Marmara(northwest Turkey): late Quaternary sedimentation and sea-level changes.Geological Society of America Bulletin 107 (8), 923–936.
Solomon, S., Mudie, P.J., Cranston, R., Hamilton, T., Thibaudeau, S.A., Collins, E.S., 2000.Characterisation of marine and lacustrine sediments in a drowned thermokarstembayment, Richards Island, Beaufort Sea, Canada. International Journal of EarthSciences 89, 503–521.
Sperling, M., Schmiedl, G., Hemleben, C., Emeis, K.C., Erlenkeuser, H., Grootes, P.M.,2003. Black Sea impact on the formation of eastern Mediterranean sapropel S1?Evidence from the Marmara Sea. Palaeogeography, Palaeoclimatology, Palaeoecol-ogy 190, 9–21.
Stanley, D.J., Blanpied, C., 1980. Late Quaternary water exchange between the easternMediterranean and the Black Sea. Nature 285, 537–541.
Stockmarr, J., 1971. Tablets with spores used in absolute pollen analysis. Pollen et Spores13, 615–621.
Stuiver, M., Grootes, P.M., 2000. GISP2 oxygen isotope ratios. Quaternary Research 53 (3),277–284.
Stuiver, M., Reimer, P.J., 1993. Extended 14C data base and revised CALIB 3.0 14C agecalibration program. In: M. Stuiver, A. Long and R.S. Kra, Editors, Calibration 1993.Radiocarbon, 35 (1): 215–230.
Stuiver, M., Reimer, P.J., Reimer, R., 2005. Calib radiocarbon calibration, execute version5.0.2 html Available from: http://calib.qub.ac.uk/calib.
Svendssen, J.I., Alexanderson, H., Astakhov, I., Demidov, I., Dowdeswell, J.A., Funder, S.,Gataullin, V., Henriksen, M., Hjort, C., Houmark-Nielsen, M., 2004. Late Quaternaryice sheet history of northern Eurasia. Quaternary Science Reviews 23, 1229–1271.
Takahashi, K., 1971. Microfossils from the Pleistocene sediments of the Ariake Sea area,west Kyushu. Pal. Soc. Japan Trans. Proc, vol. 81, pp. 11–26.
Taner, F., Eryilmaz, M., Yucesoy Eryilmaz, F., Erkurt, E.A., Turkiye, Mersin, 2001. Biomasspotential in the Black Sea, Aegean Sea, as Biogas source. World Energy Council.
Tappan, H., 1980. The paleobiology of plant protests. In: Freeman, W.H., Co (Eds.). SanFrancisco.
Tolun, L., Çağatay, M.N., Carrigan, W.J., 2002. Organic geochemistry and origin of LateGlacial–Holocene sapropelic layers and associated sediments in Marmara Sea.Marine Geology 190 (1–2), 47–60.
Toucanne, S., Zaragosi, S., Bourillet, J.F., Naughton, F., Cremer, M., Eynaud, F., Dennielou,B., 2008. Activity of the turbidite levees of the Celtic–Armorican margin (Bay ofBiscay) over the last 30,000 years: imprints of the last European deglaciation andHeinrich events. Marine Geology 247, 84–103.
Turon, J.-L., 1984. Le palynoplancton dans l'environnement actuel de l'Atlantique Nord-Oriental. Évolution climatique et hydrologique depuis le dernier maximumglaciaire. Mémoires de l'Institut de Géologie du Bassin d'Aquitaine 17, 1–313.
Turon, J.L., Londeix, L., 1988. Les assemblages de kystes de dinoflagellés enméditerranéeoccidentale (mer d'Alboran) mise en évidence de l'évolution des paléoenvironne-ments depuis le dernier maximum glaciaire. Bulletin des Centres de RecherchesExploration–Production Elf-Aquitaine 12 (1), 313–344.
Ünlüata, Ü., Özsoy, E., 1986. Oceanography of the Turkish Straits: Health of the TurkishStraits: I. Oxygen deficiency of the Sea of Marmara. Report of the Institute of MarineSciences, METU, p. 88. Erdemli, Içel.
Ünlüata, Ü., Oguz, T., Latif, M.A., Özsoy, E., 1990. On the physical oceanography of theTurkish Straits. In: Pratt, L.J. (Ed.), The Physical Oceanography of Sea Straits, NATO/ASI Series. InKluwer, Dordrecht, pp. 25–60.
Vidal, L., Ménot-Combes, G., Joly, C., Bruneton, H., Rostek, F., Çağatay, M.N., Major, C.,Bard, E., in press. Hydrology of the Sea of Marmara over the last 23 ka: implicationsfor timing of the Black Sea connections and sapropel deposition. Paleoceanography.
Wall, D., Dale, B., 1966. “Living fossils” in western Atlantic plankton. Nature 211 (5053),1025–1026.
Wall, D., Dale, B., 1974. Dinoflagellates in Late Quaternary deep-water sediments ofBlack Sea. In: The Black Sea Geology, chemistry, and biology. Memoir of theAmerican Association of Petroleum Geologist 20, 364–380.
Wall, D., Dale, B., Harada, K., 1973. Description of new fossil dinoflagellates from the lateQuaternary of the Black Sea. Micropaleontology 19 (1), 18–31.
Wall, D., Dale, B., Lohmann, G.P., Smith, W.K., 1977. The environmental and climaticdistributionofdinoflagellate cysts inmodernmarine sediments fromregions in thenorthand south Atlantic oceans and adjacent seas. Marine Micropaleontology 2, 121–200.
Wulf, S., Kraml, M., Kuhn, T., Schwarz, M., Inthorn, M., Keller, J., Kuscu, I., Halbach, P.,2002. Marine tephra from the Cape Riva eruption (22 ka) of Santorini in the Sea ofMarmara. Marine Geology 183 (1–4), 131–141.
Yaltirak, C., Alpar, B., 2002. Evolution of the middle strand of North Anatolian Fault andshallow seismic investigation of the southeastern Marmara Sea (Gemlik Bay). MarineGeology 190 (1–2), 307–327.
Yaltirak, C., Alpar, B., Sakinç, M., Yüce, H., 2000. Origin of the Strait of Canakkale(Dardanelles): regional tectonics and the Mediterranean–Marmara incursion.Marine Geology 164 (3–4), 139–156.
Yanko-Hombach, V., Gilbert, A.S., Dolukhanov, P., 2007. Controversy over the great floodhypotheses in the Black Sea in light of geological, paleontological, and archaeologicalevidence. Quaternary International 167–168, 91–113.
Yu, S.-Y., Berglund, B.E., 2007. A dinoflagellate cyst record of Holocene climate andhydrological changes along the southeastern Swedish Baltic coast. QuaternaryResearch 67, 215–224.
Zonneveld, K.A.F., Susek, E., 2007. Effects of temperature, light and salinity on cystproduction and morphology of Tuberculodinium vancampoae (the resting cyst ofPyrophacus steinii). Review of Palaeobotany and Palynology 145, 77–88.
Zonneveld, K.A.F., Hoek, R.P., Brinkhuis, H.,Willems, H., 2001. Geographical distributions oforganic-walled dinoflagellate cysts in surficial sediments of the Benguela upwellingregion and their relationship to upper ocean conditions. Progress in Oceanography 48(1), 25–72.
Zonneveld, K.A.F., Bockelmann, F., Ulrike Holzwarth, U., 2007. Selective preservation oforganic-walled dinoflagellate cysts as a tool to quantify past net primary productionand bottomwater oxygen concentrations. Marine Geology 237, 109–126.
Copyright © 2022 FDOKUMEN




















