
Lacandon Maya forest management: restoration of soil fertility using native tree species
8
ecological engineering 28 ( 2 0 0 6 ) 205–212 available at www.sciencedirect.com journal homepage: www.elsevier.com/locate/ecoleng Lacandon Maya forest management: Restoration of soil fertility using native tree species Stewart A.W. Diemont a , Jay F. Martin a,∗ , Samuel I. Levy-Tacher b , Ronald B. Nigh c , Pedro Ramirez Lopez d , J. Duncan Golicher b a Ecological Engineering Group, Department of Food Agricultural and Biological Engineering, The Ohio State University, 590 Woody Hayes Dr., Columbus, OH 43210-1057, USA b Division of Conservation and Biodiversity, Department of Ecology and Terrestrial Systems, El Colegio De La Frontera Sur, San Crist´ obal de Las Casas, Chiapas, Mexico c Centro de Investigaciones y Estudios Superiores en Antropologia Social del Sureste, San Crist´ obal de Las Casas, Chiapas, M´ exico d Department of Agroecology, El Colegio De La Frontera Sur, San Crist´ obal de Las, Casas, Chiapas, Mexico article info Article history: Received 23 June 2005 Received in revised form 4 October 2005 Accepted 28 October 2005 Keywords: Rainforest restoration Indigenous knowledge Soil ecology Ochroma pyramidale Sapium lateriflorum abstract In southern Mexico, where rainforests are being degraded rapidly, the Lacandon Maya use an agroforestry system that both restores and conserves the rainforest. Their system cycles through field and fallow stages that produce food, medicines, and raw materials, and regen- erates tall secondary forest. This investigation identified plants managed by Lacandon to restore soil fertility during fallow. Through interviews, Lacandon identified 20 plants man- aged for forest restoration. Leaf litter measurements and soil samples were taken near two of these species, Ochroma pyramidale and Sapium lateriflorum. Leaf litter increased quicker beneath O. pyramidales compared to other tree species (R = 0.48, P = 0.004), and total nematode concentrations increased with distance from this tree (R = 0.71, P < 0.001). Together, these two findings indicated an inhibition of degradation that permits accelerated soil organic mat- ter accumulation. Available phosphorus (P) concentrations beneath S. lateriflorum were 16% higher than outside the canopy (P = 0.03), and increased with age of the tree, indicating P recovery from subsoil. Our research shows that the Lacandon are cognizant of the natural abilities of certain species to fulfill the restoration needs in their systems. It demonstrates that Maya agroforestry and local knowledge could contribute to efforts to conserve and restore rainforests, and reduce deforestation by accelerating fallow in tropical agriculture. © 2005 Elsevier B.V. All rights reserved. 1. Introduction Land areas of southern Mexico are being deforested and losing productivity at alarming rates. In Chiapas, Mexico, deforestation is claiming 7% of the forest each year, and erosion has moderately degraded 10% to 25%, and severely degraded 5% of the arable soil (Howard and Homer-Dixon, 1996). These problems are endemic throughout the tropics, ∗ Corresponding author. Tel.: +1 614 247 6133; fax: +1 614 292 9448. E-mail address: [email protected] (J.F. Martin). as increasing population densities stress the environment through demands on agricultural land (Lal, 1995; Alvarez and Naughton-Treves, 2003). Because these areas are experiencing high population growth and movement (Ram, 1997), these problems will be magnified in future years. Displaced and migrant populations, in particular, have had a large effect on the ecological stability of this region (Nicholson et al., 1995; Atran, 1999; Mas and Puig, 2001). Land 0925-8574/$ – see front matter © 2005 Elsevier B.V. All rights reserved. doi:10.1016/j.ecoleng.2005.10.012
Transcript of Lacandon Maya forest management: restoration of soil fertility using native tree species

Lo
SRa
Tb
Ec
d
a
A
R
R
4
A
K
R
I
S
O
S
ded1
0d
e c o l o g i c a l e n g i n e e r i n g 2 8 ( 2 0 0 6 ) 205–212
avai lab le at www.sc iencedi rec t .com
journa l homepage: www.e lsev ier .com/ locate /eco leng
acandon Maya forest management: Restorationf soil fertility using native tree species
tewart A.W. Diemonta, Jay F. Martina,∗, Samuel I. Levy-Tacherb,onald B. Nighc, Pedro Ramirez Lopezd, J. Duncan Golicherb
Ecological Engineering Group, Department of Food Agricultural and Biological Engineering,he Ohio State University, 590 Woody Hayes Dr., Columbus, OH 43210-1057, USADivision of Conservation and Biodiversity, Department of Ecology and Terrestrial Systems,l Colegio De La Frontera Sur, San Cristobal de Las Casas, Chiapas, MexicoCentro de Investigaciones y Estudios Superiores en Antropologia Social del Sureste, San Cristobal de Las Casas, Chiapas, MexicoDepartment of Agroecology, El Colegio De La Frontera Sur, San Cristobal de Las, Casas, Chiapas, Mexico
r t i c l e i n f o
rticle history:
eceived 23 June 2005
eceived in revised form
October 2005
ccepted 28 October 2005
eywords:
ainforest restoration
ndigenous knowledge
oil ecology
a b s t r a c t
In southern Mexico, where rainforests are being degraded rapidly, the Lacandon Maya use
an agroforestry system that both restores and conserves the rainforest. Their system cycles
through field and fallow stages that produce food, medicines, and raw materials, and regen-
erates tall secondary forest. This investigation identified plants managed by Lacandon to
restore soil fertility during fallow. Through interviews, Lacandon identified 20 plants man-
aged for forest restoration. Leaf litter measurements and soil samples were taken near two
of these species, Ochroma pyramidale and Sapium lateriflorum. Leaf litter increased quicker
beneath O. pyramidales compared to other tree species (R = 0.48, P = 0.004), and total nematode
concentrations increased with distance from this tree (R = 0.71, P < 0.001). Together, these two
findings indicated an inhibition of degradation that permits accelerated soil organic mat-
ter accumulation. Available phosphorus (P) concentrations beneath S. lateriflorum were 16%
chroma pyramidale
apium lateriflorum
higher than outside the canopy (P = 0.03), and increased with age of the tree, indicating P
recovery from subsoil. Our research shows that the Lacandon are cognizant of the natural
abilities of certain species to fulfill the restoration needs in their systems. It demonstrates
that Maya agroforestry and local knowledge could contribute to efforts to conserve and
restore rainforests, and reduce deforestation by accelerating fallow in tropical agriculture.
1. Introduction
Land areas of southern Mexico are being deforested andlosing productivity at alarming rates. In Chiapas, Mexico,
eforestation is claiming 7% of the forest each year, androsion has moderately degraded 10% to 25%, and severelyegraded 5% of the arable soil (Howard and Homer-Dixon,996). These problems are endemic throughout the tropics,
∗ Corresponding author. Tel.: +1 614 247 6133; fax: +1 614 292 9448.E-mail address: [email protected] (J.F. Martin).
925-8574/$ – see front matter © 2005 Elsevier B.V. All rights reserved.oi:10.1016/j.ecoleng.2005.10.012
© 2005 Elsevier B.V. All rights reserved.
as increasing population densities stress the environmentthrough demands on agricultural land (Lal, 1995; Alvarez andNaughton-Treves, 2003). Because these areas are experiencinghigh population growth and movement (Ram, 1997), these
problems will be magnified in future years.Displaced and migrant populations, in particular, havehad a large effect on the ecological stability of this region(Nicholson et al., 1995; Atran, 1999; Mas and Puig, 2001). Land

r i n g
206 e c o l o g i c a l e n g i n e emanagement of these groups tends to take the form of cattlepasture (Mas and Puig, 2001; Durand and Lazos, 2004) or short-term milpa (O’Brien, 1998). Uncontrolled grazing leaves theland compacted and incapable of production, even for graz-ing, after a very short time (Garciaoliva et al., 1994; Durand andLazos, 2004). Following crop and cattle production, these areasdo not return to a mature, enriched forest, but to degradedgrass and brush vegetation. These lands have very little pro-ductive use and relatively low biodiversity (Miller, 1999). Landsdevastated by inappropriate use intensify the demand fornew lands, leading to further deforestation and social conflict(Nicholson et al., 1995; Howard and Homer-Dixon, 1996).
Ecosystem management and restoration in this area ofthe world is a complex social, economic, and ecological prob-lem (Nicholson et al., 1995). Tools that offer monetary incen-tives or sustenance may be important to any long-term solu-tion to rainforest loss and restoration (Nicholson et al., 1995;Foroughbakhch et al., 2001; Li, 2004). Numerous researchersstate the importance of recording indigenous knowledge tobetter understand sustainable land management (Fox et al.,2000; Long and Zhou, 2001; Hardwick et al., 2004). Becauseindigenous swidden agroforestry systems can be produc-tive (Long and Nair, 1999) while maintaining their ecologicalintegrity (Wang and Young, 2003), swidden practices couldcontribute to better land management in the tropics (De Clerckand Negreros-Castillo, 2000; Fox et al., 2000). The LacandonMaya, an indigenous group that has met subsistence needswhile maintaining both secondary and primary forests forcenturies in southern Mexico, combine forest restoration anddomestic production (Nations and Nigh, 1980; Levy, 2000;Diemont and Martin, 2005).
The Lacandon land management system cycles throughthree field stages starting with the milpa, progressing to theacahual (low secondary forest), and then to tall secondary for-est, before returning to the milpa (Nations and Nigh, 1980;McGee, 2002). Neighboring primary forest is conserved tomaintain a biodiverse seedbank (Quintana-Ascencio et al.,1996). Ecological succession drives the conversion betweenfield stages (Levy and Aguirre Rivera, in press). From the view-point of ecological succession, the milpa represents early suc-cessional grasses, the acahual represents the shrub or earlywoody stage, and the forest is the climax stage. The milpa, orearly successional stage, is a polyculture field that includes20–30 cultivated species. The acahual and forest stages arealso productive, offering over 50 plant species used by theLacandon (Nations and Nigh, 1980). By selecting for certainspecies and managing the natural succession of the acahualand forest stages, the Lacandon are able to restore soil fertilityand regenerate secondary forest following the milpa stage inless than 20 years (Diemont and Martin, 2005).
Lacandon Maya agroforestry is ecological engineering asdescribed by Odum et al. (1963), where the technology avail-able from natural systems is dominant, and human engi-neering is supplementary rather than primary. The Lacandonrely on the regenerative capacity of nature. They seed certainplants during the fallow and eliminate others, but in general
allow the system to develop without intervention, permittingforcing functions such as sun, wind, and rain to drive the sys-tem (Diemont et al., 2006). Furthermore, the Lacandon systemis as Mitsch and Jorgensen (1989) have described ecological2 8 ( 2 0 0 6 ) 205–212
engineering—designed for the benefit of both humans andthe environment. At all stages of successional development,the Lacandon recover harvestable foods, medicines, and rawmaterials (Nations and Nigh, 1980). This production does notcome at the cost of ecosystem health. The system largely self-designs and develops in biodiversity and complexity (Nationsand Nigh, 1980; Levy, 2000); numerous animals are drawn tothe richness of this ecosystem (Nations and Nigh, 1980).
Previous studies have identified plants that the Lacandonmay be using to restore soil fertility (Levy, 2004; Levy andGolicher, 2004). The presence of Ochroma pyramidale Urban,has been related to greater leaf-litter (Levy, 2004; Levy andGolicher, 2004) and soil organic matter accumulation (Levy andGolicher, 2004), indicating that the Lacandon manage their fal-low to accelerate regeneration of soil fertility. An evaluation ofsoil chemistry, soil nematode populations, and plant commu-nity in all field stages of the Lacandon system revealed sevenplants that correlated positively with improved soil conditions(SAW Diemont, JF Martin, and MF Quigley, unpublished data inreview). Hampea stipitata S. Watson, found in the acahual, corre-lated with elevated soil organic matter concentrations. Sapiumlateriflorum, found in secondary and primary forest, correlatedwith elevated ammonium and nitrate concentrations. Cecropiaobtusifolia Bertol., discovered in milpa, acahual and secondaryforest, and Piper auritum H.B.K., in acahual and secondary for-est, correlated with higher bacterivorous nematode concen-trations. These results encouraged additional research intothe plants the Lacandon are using during the fallow to restoresecondary forest. The objectives of this study were to: (1)through interviews, determine if the Lacandon have selectedfor certain species to accelerate the regeneration of soil fertil-ity; (2) better identify the contribution of selected species (i.e.O. pyramidale) to soil fertility; (3) identify the mechanism bywhich selected species regenerate soil fertility; and (4) evalu-ate the potential for these methods to be used to restore andconserve tropical rainforests.
2. Materials and methods
Interviews and soil sampling were conducted in LacanjaChansayab, Chiapas, Mexico. Lacanja has a population ofapproximately 400 and is one of three major communitiesof the Lacandon Maya. Lacanja is located in Chiapas, thesouthern-most Mexican state, at 16◦45′ 30′ ′N and 91◦08′ 30′ ′Wand at an elevation of 400 m. The predominate soil type is Luvi-sol (INEGI, 1982), with a clayey texture and neutral pH. Thesurrounding ecosystem is tall moist forest, and annual rain-fall is 2500 cm (Guillen-Trujillo, 1998).
2.1. Interviews
Interviews were conducted with five Lacandon farmers todetermine what plants they regarded as important for fertilityregeneration during the fallow stages of the system. Farmerswere asked first to identify what plants were good for fertil-
ity regeneration in the fallow. They were asked if soil thatresulted in the area where the plant was located would begood, whether the plant leaf litter produced good compost,whether the milpa stage that may follow in the location would
e c o l o g i c a l e n g i n e e r i n g
Table 1 – Mean (standard deviation) of the age (years)and diameter at breast height (DBH) (cm) of the Sapiumlateriflorum and Ochroma pyramidale trees sampled
Tree n Age (year) DBH (cm)
Sapium lateriflorumAcahual 3 3.0(1.0) 26.7(16.3)Secondary Forest 5 16.4(6.2) 78.0 (46.8)Primary forest 1 40 142Total 9 14.6(12.4) 68.0 (50.5)Lattice 1 25 145
Ochroma pyramidaleMilpa 2 1 18.0(1.4)Acahual 3 3.7 (0.6) 24.0 (6.1)
parownft
2
BsSwsamf
ibTtu
wpttatAsTesabVicF
Secondary forest 4 11.5(2.5) 62.8 (7.1)Total 9 6.6 (5.5) 39.9 (22.5)Lattice 1 8 30
roduce healthy cultivated species (i.e. maize). Farmers werelso asked if the plants had alternative uses aside from fertilityegeneration (i.e. construction materials). Plants identified byne farmer were subsequently asked of the following farmersho where interviewed. Species that received more than oneegative response to any of these questions from subsequent
armers were discarded from the list of fertility enhancingrees.
.2. Field sampling
ased on interviews and literature review, two trees were cho-en for soil sampling and analysis: Ochroma pyramidale andapium lateriflorum. Soil sampling under Sapium lateriflorumas conducted in November 2004 during the wet season. Soil
ampling under Ochroma pyramidale was conducted in Febru-ry 2005 during the intermediate wet-dry season. Trees inilpa, acahual, secondary forest, and primary forest were used
or analysis.Three small, medium, and large trees of each species were
dentified for sampling. Approximate age and diameter atreast height (DBH) of each tree were determined (Table 1).ree age was determined by a Lacandon farmer familiar withree size relative to age and the specific history of the individ-al trees sampled.
One transect was laid out through each tree, and samplingas conducted at seven locations along each transect. Sam-ling locations along transects were evenly spaced so thathe sampling location second to the end at each side of eachransect was at the canopy edge. The end sampling locationslong each transect were outside the tree canopy. The cen-er point of each transect was located next to the tree trunk.
lattice was laid out beneath the canopy of one tree withixteen sampling locations at the intersection of lattice lines.he midpoint of lattice edge lines intersected with the canopydge. Four additional points within the lattice were randomlyelected. Non-sampled trees with a DBH greater than 15 cmnd with a canopy over a sampling location were identifiedy Lacandon name and cross-referenced for Latin binomials.
oucher specimens for all plants not previously collected anddentified in Nations and Nigh (1980) or Levy et al. (2002) wereollected and deposited in the herbarium at El Colegio de larontera Sur, San Cristobal de Las Casas, Mexico.
2 8 ( 2 0 0 6 ) 205–212 207
Within a 1 m2 circular sampling area at each samplinglocation, leaf litter depth was measured at three points andaveraged. Soil moisture and pH were evaluated using a Kelwaysoil acidity and moisture meter. The detrital layer was thenremoved, and eight replicate 2.5 cm diameter cores weretaken from 0–20 cm soil depth and pooled. Each soil samplewas analyzed for soil organic matter (SOM) (Walkley andBlack, 1934), total nitrogen (N) (semi-microkjeldhal), availablephosphorus (P) (Olsen et al., 1954), and texture (Bouyoucos,1951). Nematodes were extracted from 20 g of the pooled soilfrom each sampling location for analysis. Soil samples fornematode extraction were kept cool and extracted over 48 husing the Baermann wet funnel technique (McSorley andWelter, 1991). Nematodes were heat fixed. Extract was storedin 2% formaldehyde, and all nematodes in the bottom 10 mLof extract were identified to trophic level at 90× magnification(Parmelee and Alston, 1986; Edwards, 1991; Dominguez etal., 2003; Arancon et al., 2003). Nematodes were identified asplant parasite, fungivore, bacterivore, and omnivore-predatortrophic groups according to Parmelee and Alston (1986).Omnivore/predator was reported as omnivore, as in Aranconet al. (2003).
2.3. Analysis
Statistical analysis was conducted using Systat 10.2 computersoftware. Sampling points from beneath and outside the treecanopy were compared using independent sample Student’st-tests. Relationships between factors were determined usingPearson linear regression. Sampling locations under canopytrees of the same species as the tree under consideration wereremoved. Outliers (|Studentized residuals| > 2.75) and pointswith large leverage (leverage > 0.3) were removed. Where tran-sect and lattice data were pooled for regressions, the ratioof dependent and independent factors of lattice and transectpoints were compared using independent sample Student’s t-tests to determine that differences were not present that mayskew results. Where a difference was present, results werereported for transect points only.
3. Results
3.1. Interviews
Interviews with Lacandon farmers yielded 20 plants thatwere considered to be important for fertility regeneration(Table 2). Of these plants, 19 are useful to the Lacandon insome way apart from soil restoration. Uses included food,construction materials, furniture, arts and crafts, medicines,canoes, and food for wild birds. Due to uniformly strong inter-view responses and past research (Levy, 2004; and Levy andGolicher, 2004), Ochroma pyramidale and Sapium lateriflorumwere chosen for more in-depth assessment. O. pyramidale isa cultivated species that is planted in the milpa, and acahual indense patches. S. lateriflorum is not planted by the Lacandonbecause seeds are readily disbursed by birds, and an abundant
amount of these plants can therefore be found in the sec-ondary forest. Although other species are considered weedyand removed by the Lacandon, S. lateriflorum is never removedfrom an acahual or secondary forest.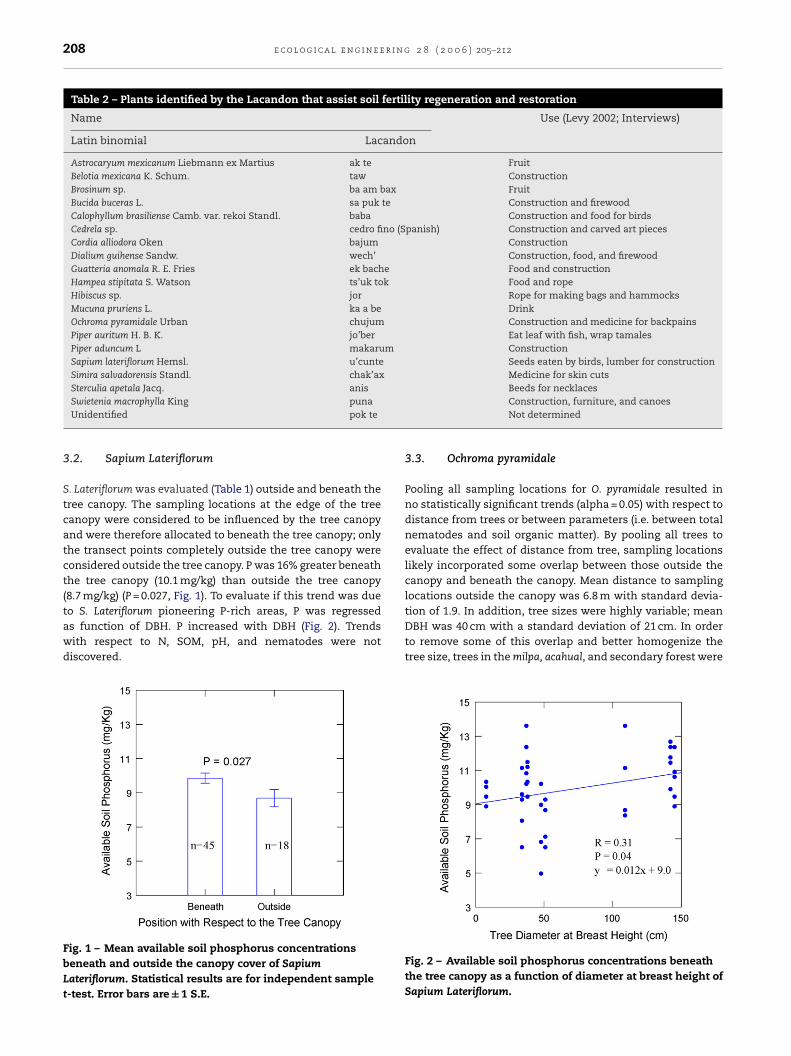
208 e c o l o g i c a l e n g i n e e r i n g 2 8 ( 2 0 0 6 ) 205–212
Table 2 – Plants identified by the Lacandon that assist soil fertility regeneration and restoration
Name Use (Levy 2002; Interviews)
Latin binomial Lacandon
Astrocaryum mexicanum Liebmann ex Martius ak te FruitBelotia mexicana K. Schum. taw ConstructionBrosinum sp. ba am bax FruitBucida buceras L. sa puk te Construction and firewoodCalophyllum brasiliense Camb. var. rekoi Standl. baba Construction and food for birdsCedrela sp. cedro fino (Spanish) Construction and carved art piecesCordia alliodora Oken bajum ConstructionDialium guihense Sandw. wech’ Construction, food, and firewoodGuatteria anomala R. E. Fries ek bache Food and constructionHampea stipitata S. Watson ts’uk tok Food and ropeHibiscus sp. jor Rope for making bags and hammocksMucuna pruriens L. ka a be DrinkOchroma pyramidale Urban chujum Construction and medicine for backpainsPiper auritum H. B. K. jo’ber Eat leaf with fish, wrap tamalesPiper aduncum L makarum ConstructionSapium lateriflorum Hemsl. u’cunte Seeds eaten by birds, lumber for constructionSimira salvadorensis Standl. chak’ax Medicine for skin cutsSterculia apetala Jacq. anis Beeds for necklaces
Swietenia macrophylla King punaUnidentified pok te3.2. Sapium Lateriflorum
S. Lateriflorum was evaluated (Table 1) outside and beneath thetree canopy. The sampling locations at the edge of the treecanopy were considered to be influenced by the tree canopyand were therefore allocated to beneath the tree canopy; onlythe transect points completely outside the tree canopy wereconsidered outside the tree canopy. P was 16% greater beneaththe tree canopy (10.1 mg/kg) than outside the tree canopy(8.7 mg/kg) (P = 0.027, Fig. 1). To evaluate if this trend was due
to S. Lateriflorum pioneering P-rich areas, P was regressedas function of DBH. P increased with DBH (Fig. 2). Trendswith respect to N, SOM, pH, and nematodes were notdiscovered.Fig. 1 – Mean available soil phosphorus concentrationsbeneath and outside the canopy cover of SapiumLateriflorum. Statistical results are for independent samplet-test. Error bars are ± 1 S.E.
Construction, furniture, and canoesNot determined
3.3. Ochroma pyramidale
Pooling all sampling locations for O. pyramidale resulted inno statistically significant trends (alpha = 0.05) with respect todistance from trees or between parameters (i.e. between totalnematodes and soil organic matter). By pooling all trees toevaluate the effect of distance from tree, sampling locationslikely incorporated some overlap between those outside thecanopy and beneath the canopy. Mean distance to samplinglocations outside the canopy was 6.8 m with standard devia-
tion of 1.9. In addition, tree sizes were highly variable; meanDBH was 40 cm with a standard deviation of 21 cm. In orderto remove some of this overlap and better homogenize thetree size, trees in the milpa, acahual, and secondary forest wereFig. 2 – Available soil phosphorus concentrations beneaththe tree canopy as a function of diameter at breast height ofSapium Lateriflorum.

e c o l o g i c a l e n g i n e e r i n g 2 8 ( 2 0 0 6 ) 205–212 209
Fig. 3 – (a–f) Leaf litter (a), soil organic matter (transect only) (b), total nitrogen (c), total nematodes (d), bacterivore nematodes( m Oc
afiMt
etllm
e), and omnivore nematodes (f) as a function of distance fro
nalyzed separately (Table 1). For example, in the secondaryorest, the distance to the sampling point outside the canopyncreased to 7.8, while the standard deviation decreased to 1.0.
ean DBH increased to 63, and standard deviation decreasedo 6.1 (Table 1).
Considering only trees in the secondary forest revealed sev-ral trends (Fig. 3). To compensate for reduced n in examining
rees in the secondary forest, transect and lattice samplingocations were pooled (see 2.3 Analysis). Log-transformed leafitter depth decreased as a function of distance from O. Pyra-idale (Fig. 3a). Mean leaf litter at the tree was over 6 cm and
hroma pyramidale trees in the secondary forest.
decreased to less than 3 cm outside the tree canopy. SOM (tran-sect only) (Fig. 3b), total N (Fig. 3c), and P (R = 0.047, P = 0.78)did not exhibit any significant trends. Nematodes displayedtrends opposite to that of leaf litter depth. Total nematodes(Fig. 3d) increased from 8 per 20 g soil at the trunk to over60 at a distance of 14 m. Bacterivorous nematodes (Fig. 3e)increased by an order of magnitude from 4 per 20 g soil at the
trunk to nearly 40 outside the tree canopy. Omnivorous (Fig. 3f)nematodes increased from 5 per 20 g soil at the trunk to nearly15 outside the canopy. Trees in the milpa and acahual did notexhibit any trends with respect to soil nematodes or chemistry.
r i n g
210 e c o l o g i c a l e n g i n e e4. Discussion
The numerous plants identified by five Lacandon farmers asbeing important for soil fertility (Table 2) and the results of thisstudy lend support to the hypothesis by Levy (2004) and Levyand Golicher (2004) that the Lacandon are actively enhanc-ing the soil of their fallow stages through plant management.Whereas other trees are removed during milpa and fallowstages (Nations and Nigh, 1980; Diemont and Martin, 2005),the trees identified by the Lacandon are either cultivated (e.g.Ochroma pyramidale) or are left in the fallow following aeo-lian or animal seed disbursal (e.g. Sapium lateriflorum and Piperspp.). That most of these plants are also useful (Table 2) sug-gests that production does not need to come at the cost offertility regeneration, or vice versa (Nations and Nigh, 1980;Diemont and Martin, 2005). Lacandon farmers are able torestore the rainforest during the fallow, while at the same timeproducing numerous useful crops (Nations and Nigh, 1980).Levy (2002) recorded over 400 plants that are useful to theLacandon that occur in the fallow and primary forest. Theproductivity of secondary vegetation allows the Lacandon tomaintain their lands in fallow for longer periods of over twentyyears (Diemont and Martin, 2005) and conserve primary for-est for medicines, raw materials, food and ecosystem services(Nations and Nigh, 1980; Levy, 2002). In comparison, the fallowduration has been reduced to less than 10 years in many swid-den systems in the Yucatan, Mexico (Weisbach et al., 2002).Furthermore, Lacandon plant management to enhance soilfertility may shorten the time needed to restore mature for-est to sites previously used for agriculture (Levy and Golicher,2004). Reducing the time needed to restore soil fertility andenhancing the productivity of fallows are both important waysfor protecting primary forest. Determining management tech-niques that allow simultaneous restoration of soil fertilityand productivity, as does the Lacandon system, could reducedeforestation throughout the world.
Traditional Lacandon farmers do not use fertilizer (Nationsand Nigh, 1980; Levy and Aguirre, 1999; Diemont and Martin,2005). In a geographic area where phosphorous is typicallyunavailable, P could become a limiting factor in agriculturalproduction (Bautista-Cruz and del Castillo, 2005). Whereas Ncan be introduced from the atmosphere by N fixing legumi-nous herbs and trees (Badejo, 1998; Okogun et al., 2000), Pcan only enter from runoff and plant extraction from the soilor sub-soil. Consequently, the elevated levels of P beneathS. lateriflorum (Table 2) and the increasing P concentrationswith DBH (Fig. 2) support the conclusion that this plant, whichis managed by the Lacandon, serves as a “phosphorus pump”(Perezllorens et al., 1993; Badejo, 1998). Vejre and Hoppe (1998)describe such a pumping effect of K, P, Ca, and Mg, whereminerals in the lower soil profiles were brought to surface bydeeper tree roots in a forest in Denmark. Similarly, Christantyet al. (1997) found that the nutrient pumping provided by Phyl-lostachys sp. in an Indonesian agroforestry system was vital tomaintaining fertility of the surface soils.
The results for Ochroma pyramidale indicate that nematodepopulations are inhibited by O. pyramidale leaf litter (Fig. 3).Leaf litter concentration decreased with distance from thetree, whereas nematodes were found to increase (Fig. 3).
2 8 ( 2 0 0 6 ) 205–212
Nematodes would generally decrease in number with dis-tance from the tree along with leaf litter depth (Fig. 3a) if aninhibitory effect were not present. Ferris and Matute (2003)and Matlack (2001) found that the abundance of bacterivoreswere elevated with the addition of organic matter. Bjornlundand Christensen (2005) found that leaf litter addition resultedin bacterivorous nematode spikes followed by fungivorousnematode dominance as the leaf litter was broken down. Inother successional swidden systems, similar to the Lacandonsystem, fallow length, and thus greater leaf litter accumula-tion, appeared to increase the abundance of nematodes (Pateet al., 2000; Villenave et al., 2001).
The correlation between nematodes, litter, and distancefrom trees was not noted in younger trees (e.g. trees less thanfour years old), but only in trees found in the secondary forest;this result indicates that the inhibition may be a cumulativeeffect. Chemical analysis of O. pyramidale showed an abun-dance of soluble saccharides compared to starch (Marencoet al., 2001). O’Dowd (1979) found that O. pyramidale pro-duces nectar on the leaves of young trees. The abundanceof foliar nectar or other secondary compounds depositedover the course of early succession may inhibit nematodepopulations beneath the canopy. In addition, Molofsky andAugspurger (1992) found that establishment of other speciesof tree seedlings underneath O. pyramidale was limited duringseed germination, not seed deposition, which may indicate apossible allelopathic effect.
Leaf litter and thus organic matter beneath O. pyramidaleare abundant (Levy, 2004; Levy and Golicher, 2004), an unusualsituation in tropical rainforests where conditions favor rapidbacterial and fungal decomposition (Attignon et al., 2004).Dense planting of O. pyramidale may allow Lacandon farmers toaccumulate and store leaf litter and organic matter for futureuse. O. pyramidale is an early succession species, which wouldpermit the next successional stage to have an abundance ofsoil organic matter available at the outset. That no trend wasnoted for soil organic matter as a function of distance fromthe tree despite the abundance of leaf litter closer to the treefurther supports this idea. Levy and Golicher (2004) discoveredhigher soil organic matter near O. pyramidale trees comparedto where O. pyramidale was not present, but this difference waspatchy and inconsistent between plots.
Other species represented on the list of fertility-enhancingtrees (Table 2) may contribute to the accumulation of leaf litterduring the fallow. Piper auritum contains Safrol in its leaves,which has been shown to have high cytotoxic effects, andPiperacae in general are notoriously allelopathic (Anaya-Lang,1979, Gupta et al., 1996, Xuan et al., 2004). The Lacandon favorO. pyramidale in their fallows for fertility enhancement, butfarmers stated in the interviews that these trees will only growin a quarter of their fallow lands. Therefore, a mosaic of othertrees that have similar functions may be important for man-aging restoration.
5. Conclusions
Lacandon Maya indigenous agroforesty uses numerous treesfor soil restoration during fallow. These trees also producefood, medicines, and raw materials. Results indicate that one

i n g
ormasefaf
A
WKPOba
r
A
A
A
A
A
B
B
B
B
C
D
D
D
e c o l o g i c a l e n g i n e e r
f these trees, Sapium laterijlorum, acts as a phosphorus pump,estoring this nutrient to the soil during fallow. Ochroma pyra-idale appears to be slowing organic matter degradation and
llowing storage of organic matter in leaf litter. Future researchhould verify these results with experimental testing andxamine the use of other trees identified by the Lacandonor soil fertility regeneration. Lacandon land managementppears to offer new tools for rainforest restoration and soilertility enhancement.
cknowledgements
e wish to thank Manuel Castellanos, Kin Bor, Adolfo Chan’in, Poncho Kin, Jorge Paneagua, Vicente Paneagua, Enriqueaneagua, and Kin Paneagua. Financial support from thehio Agricultural Research and Development Center, the Ful-right Program, and NSF Grant OISE-0431230 is gratefullycknowledged.
e f e r e n c e s
lvarez, N.L., Naughton-Treves, L., 2003. Linking nationalagrarian policy to deforestation in the Peruvian Amazon: acase study of Tambopata, 1986–1997. Ambio 32 (4), 269–274.
naya-Lang, A.L., 1979. Consideraciones sobre el potencialaleleopatico de la vegetacion secundaria. In: Gomez-Pompa,A., Vazquez-Yanes, C., Silvia del Amo, R., Butanda, C. (Eds.),Regeneration de Selvas. Mexico CECSA, Mexico City, Mexico,pp. 428–445.
rancon, N., Galvis, P., Edwards, C., Yardim, E., 2003. The trophicdiversity of nematode communities in soils treated withvermicompost. Pedobiologia 47, 736–740.
tran, S., 1999. Managing the Maya commons: the value of localknowledge. In: Nzarea, V.D. (Ed.), Ethnoecology: SituatedKnowledge/located Lives. University of Arizona Press, Tucson,pp. 190–214.
ttignon, S.E., Weibel, D., Lachat, T., Sinsin, B., Nagel, P., Peveling,R., 2004. Leaf litter breakdown in natural and plantationforests of the Lama forest reserve in Benin. Appl. Soil Ecol. 27(2), 109–124.
adejo, M.A., 1998. Agroecological restoration of savannaecosystems. Ecol. Eng. 10 (2), 209–219.
autista-Cruz, A., del Castillo, R.F., 2005. Soil changes duringsecondary succession in a tropical montane cloud forest area.Soil Sci. Soc. Am. J. 69 (3), 906–914.
jornlund, L., Christensen, S., 2005. How does litter quality andsite heterogeneity interact on decomposer food webs of asemi-natural forest? Soil Biol. Biochem. 37 (2), 203–213.
ouyoucos, G.J., 1951. A recalibration of the hydrometer methodfor making mechanical analysis of soil. Agron. J. 43, 434–438.
hristanty, L., Kimmins, J.P., Mailly, D., 1997. ‘Without bamboo,the land dies’: A conceptual model of the biogeochemical roleof bamboo in an Indonesian agroforestry system. Forest Ecol.Manage. 91 (1), 83–91.
e Clerck, F.A.J., Negreros-Castillo, P., 2000. Plant species oftraditional Mayan homegardens of Mexico as analogs formultistrata agroforests. Agroforestry Syst. 48, 303–317.
iemont, S.A.W., Martin, J.F., 2005. Management impacts on the
trophic diversity of nematode communities in an indigenousagroforestry system of Chiapas, Mexico. Pedobiologia 49,325–334.iemont, S.A.W., Martin, J.F., Levy-Tacher, S.I., 2006. Emergyevaluation of Lacandon Maya indigenous swidden
2 8 ( 2 0 0 6 ) 205–212 211
agroforestry in Chiapas, Mexico. Agroforestry Syst. 66,23–42.
Dominguez, J., Parmelee, R.W., Edwards, C.A., 2003. Interactionsbetween Eisenia andrei (Oligochaeta) and nematodepopulations during vermicomposting. Pedobiologia 47, 53–60.
Durand, L., Lazos, E., 2004. Colonization and tropicaldeforestation in the Sierra Santa Marta, southern Mexico.Environ. Conserv. 31 (1), 11–21.
Edwards, C.A., 1991. Methods for assessing populations ofsoil-inhabiting invertebrates. In: Crossely, D.A., Coleman, D.C.,Hendrix, P.F., Cheng, W., Edwards, C.A. (Eds.), ModernTechniques on Soil Ecology. Elsevier, New York,pp. 145–176.
Ferris, H., Matute, M.M., 2003. Structural and functionalsuccession in the nematode fauna of a soil food web. Appl.Soil Ecol. 23, 93–110.
Foroughbakhch, F., Hauad, L.A., Cespedes, A.E., Ponce, E.E.,Gonzalez, N., 2001. Evaluation of 15 indigenous andintroduced species for reforestation and agroforestry innortheastern Mexico. Agroforestry Syst. 51 (3), 213–221.
Fox, J., Truong, D.M., Rambo, A.T., Tuyen, N.P., Cuc, L.T., Leisz, S.,2000. Shifting cultivation: A new old paradigm for managingtropical forests. Bioscience 50, 521–528.
Garciaoliva, F., Casar, I., Morales, P., Maass, J.M., 1994.Forest-to-pasture conversion influences on soilorganic-carbon dynamics in a tropical deciduous forest.Oecologia 99 (3–4), 392–396.
Guillen-Trujillo, H.A., 1998. Sustainability of Ecotourism andTraditional Agricultural Practices in Chiapas, Mexico. Ph.D.dissertation, Environmental Engineering Sciences, Universityof Florida, Gainesville, 246 pp.
Gupta, M.P., Monge, A., Karikas, G.A., deCerain, A.L., Solis, P.N.,deLeon, E., Trujillo, M., Suarez, O., Wilson, F., Montenegro, G.,Noriega, Y., Santana, A.I., Correa, M., Sanchez, C., 1996.Screening of Panamanian medicinal plants for brine shrimptoxicity, crown gall tumor inhibition, cytotoxicity and DNAintercalation. Int. J. Pharmacogn. 34 (1), 19–27.
Hardwick, K., Healey, J.R., Elliott, S., Blakesley, D., 2004. Researchneeds for restoring seasonal tropical forests in Thailand:accelerated natural regeneration. New Forests 27, 285–302.
Howard, P., Homer-Dixon, T., 1996. Project on Environment,Population and Security. In: Environmental Scarcity andViolent Conflict: The Case of Chiapas, Mexico. AmericanAssociation for the Advancement of Science and theUniversity of Toronto, Washington, DC.
INEGI (Instituto National de Estadistica Geografica e Informatica),1982. Carta Edafologica, Las Margaritas, E15-12. EstadosUnidos Mexicanos.
Lal, R., 1995. Erosion-crop productivity relationships for soils ofAfrica. Soil Sci. Soc. Am. J. 59 (3), 661–667.
Levy T., S.I., Aguirre R., R.J., 1999. Conceptuacion etnobotanica(experiencia de un estudio en la Lacandona). Revista deGeografia Agricola, 29, 83–114.
Levy T., S.I., 2000. Sucesion causada por roza-tumba-quema en lasselvas de Lacanha, Chiapas. Doctoral Dissertation, Institucionde Ensefianza e Investigation en Ciencias Agricolas, Institutode Recursos Naturales. Montecillo, Texcoco, Mexico.
Levy T., S.I, Aguirre R, J.R., Martınez R., M.M. Duran F., Y.A., 2002.Caracterizacion del uso tradicional de la flora espontanea enla comunidad Lacandona de Lacanha, Chiapas, Mexico,Interciencia, 27, 512–520.
Levy T., S.I. 2004. El papel de Ochroma pyramidale Cav en laagricultura tradicional Lacandona. Report to Secretaria del
Medio Ambiente y Recursos Naturales (Semarnat) of Mexico,San Cristobal de las Casas, Mexico.Levy T., S.I., Golicher, J.D., 2004. How predictive is traditionalecological knowledge? The case of the Lacandon Maya fallowenrichment system. Interciencia, 29, 496–503.

r i n g
212 e c o l o g i c a l e n g i n e eLevy T., S.I., Aguirre R., J.R., in press. Successional pathwaysderived from different vegetation use patterns by LacandonMaya. J. Sustainable Agric.
Li, W.H., 2004. Degradation and restoration of forest ecosystemsin China Forest. Forest Ecol. Manage. 201 (1), 33–41.
Long, A.J., Nair, P.K.R., 1999. Trees outside forests: agro-,community, and urban forestry. New Forests 17, 145–174.
Long, C.L., Zhou, Y.L., 2001. Indigenous community forestmanagement of Jinuo people’s swidden agroecosystems insouthwest China. Biodiversity Conserv. 10, 753–767.
Marenco, R.A., Goncalves, J.D., Vieira, G., 2001. Photosynthesisand leaf nutrient contents in Ochroma pyramidale(Bombacaceae). Photosynthetica 39 (4), 539–543.
Mas, J., Puig, H., 2001. Deforestation methods in southwesternCampeche (Mexico). Can. J. Forest Res. 31 (7), 1280–1288.
Matlack, G.R., 2001. Factors determining the distribution of soilnematodes in a commercial forest landscape. Forest Ecol.Manage. 146, 129–143.
McGee, R.J., 2002. Watching Lacandon Maya, Allyn and Bacon,Boston, p. 194.
McSorley, R., Welter, E.E., 1991. Comparisons of soil extractionmethods for nematodes and micro-arthropods. In: Crossely,D.A., Coleman, D.C., Hendrix, P.F., Cheng, W., Edwards, C.A.(Eds.), Modern Techniques on Soil Ecology. Elsevier, NewYork,pp. 201–208.
Miller, P.M., 1999. Effects of deforestation on seed banks in atropical deciduous forest of western Mexico. J. Trop. Ecol. 15,179–188.
Mitsch, W.J., Jorgensen, S.E., 1989. Ecological Engineering: AnIntroduction to Ecotechnology. Wiley, New York.
Molofsky, J., Augspurger, C.K., 1992. The effect of leaf litter onearly seedling establishment in a tropical forest. Ecology 73(1), 68–77.
Nations, J.D., Nigh, R., 1980. Evolutionary potential of LacandonMaya sustained-yield tropical forest agriculture. J. Anthropol.Res. 36, 1–30.
Nicholson, C.F., Blake, R.W., Lee, D.R., 1995. Livestock,deforestation, and policy-making - intensification of cattleproduction systems in Central-America revisited. J. Dairy Sci.78 (3), 719–734.
O’Brien, K., 1998. Sacrificing the Forest: Environmental and SocialStruggles in Chiapas. Westview Press, Boulder.
O’Dowd, D.J., 1979. Foliar nectar production and ant activity on aneotropical tree, Ochroma pyramidale. Oecologia 43, 233–248.
Odum, H.T., Siler, W.L., Beyers, R.J., Armstrong, N., 1963.Experiments with engineering of marine ecosystems,Publications of the Institute of Marine Science, 9. TheUniversity of Texas Marine Science Institute, Port Aransas,Texas, pp. 373–403.
2 8 ( 2 0 0 6 ) 205–212
Okogun, J.A., Sanginga, N., Mulongoy, K., 2000. Nitrogencontribution of five leguminous trees and shrubs to alleycropped maize in Ibadan, Nigeria. Agroforestry Syst. 50 (2),123–136.
Olsen, S.R., Cole, C.V., Watanabe, F.S., Dean, L.A., 1954. Estimationof available phosphorus in soils by extraction with sodiumbicarbonate. U.S. Dep. Agric. Circ., 939.
Parmelee, R.W., Alston, D.G., 1986. Nematode trophic structure inconventional and no-tillage agroecosystems. J. Nematol. 18,403–407.
Pate, E., Ndiaye-Faye, N., Thioulouse, J., Villenave, C., Bongers, T.,Cadet, P., Debouzie, D., 2000. Successional trends in thecharacteristics of soil nematode communities in cropped andfallow lands in Senegal (Sonkorong). Appl. Soil Ecol. 14 (1),5–15.
Perezllorens, J.L., Devisscher, P., Nienhuis, P.H., Niell, F.X., 1993.Light-dependent uptake, translocation and foliar release ofphosphorus by the intertidal seagrass Zostera noltii Hornem.J. Exp. Marine Biol. Ecol. 166 (2), 165–174.
Quintana-Ascencio, P.F., Gonzalez-Espinosa, M., Ramirez-Marcial,N., Dominguez-Vazquez, G., Martinez-Ico, M., 1996. Soil seedbank and regeneration of tropical rain forest from milpa fieldsat the Selva Lacandona, Chiapas, Mexico. Biotropica 28 (2),192–209.
Ram, R., 1997. Tropics and economic development: an empiricalinvestigation. World Develop. 25 (9), 1443–1452.
Vejre, H., Hoppe, C., 1998. Distribution of Ca, K, Mg, and P in acidforest soils in plantations of Picea abies—evidence of thebase-pump effect. Scandinavian J. Forest Res. 13 (3),265–273.
Villenave, C., Bongers, T., Ekschmitt, K., Djigal, D., Chotte, J.L.,2001. Changes in nematode communities followingcultivation of soils after fallow periods of different length.Appl. Soil Ecol. 17, 43–52.
Walkley, A., Black, L.A., 1934. An examination of the Degtjareffmethod for determining soil organic matter, and a proposedmodification of the chromic acid titration method. Soil Sci. 37,29–38.
Wang, Z.J., Young, S.S., 2003. Differences in bird diversity betweentwo swidden agricultural sites in mountainous terrain,Xishuangbanna, Yunnan, China. Biol. Conserv. 110,231–243.
Weisbach, C., Tiessen, H., Jimenez-Osornio, J.J., 2002. Soil fertility
during shifting cultivation in the tropical Karst soils ofYucatan. Agronomie 22 (3), 253–263.Xuan, T.D., Eiji, T., Shinkichi, T., Khanh, T.D., 2004. Methods todetermine allelopathic potential of crop plants for weedcontrol. Allelopathy J. 13 (2), 149–164.




















