
Knots in the family tree: evolutionary relationships and functions of knox homeobox genes
16
Plant Molecular Biology 42: 151–166, 2000. © 2000 Kluwer Academic Publishers. Printed in the Netherlands. 151 Knots in the family tree: evolutionary relationships and functions of knox homeobox genes Leonore Reiser 1,* , Patricia S´ anchez-Baracaldo 2 and Sarah Hake 1 1 Department of Plant and Microbial Biology, University of California, Berkeley, CA 94720, USA ( * author for correspondence; e-mail [email protected]); 2 Department of Integrative Biology and University and Jepson Herbaria, University of California, Berkeley, CA 94720, USA Key words: evolution, homeodomain protein, knox gene, meristem Abstract Knotted-like homeobox (knox) genes constitute a gene family in plants. Class I knox genes are expressed in shoot apical meristems, and (with notable exceptions) not in lateral organ primordia. Class II genes have more diverse expression patterns. Loss and gain of function mutations indicate that knox genes are important regulators of meristem function. Gene duplication has contributed to the evolution of families of homeodomain proteins in metazoans. We believe that similar mechanisms have contributed to the diversity of knox gene function in plants. Knox genes may have contributed to the evolution of compound leaves in tomato and could be involved in the evolution of morphological traits in other species. Alterations in cis-regulatory regions in some knox genes correlate with novel patterns of gene expression and distinctive morphologies. Preliminary data from the analysis of class I knox gene expression illustrates the evolution of complex patterns of knox expression is likely to have occurred through loss and gain of domains of gene expression. Introduction The primary architecture of plants derives from the shoot apical meristem, which produces leaves, inter- nodes and axillary buds. Seemingly simple differences in organ initiation from the meristem, such as leaf initiation in a spiral versus decussate phyllotaxy, can result in dramatically divergent overall morphologies. The organization and maintenance of the meristem re- mains a fundamental question in plant development. As new information emerges regarding the genetic regulation of meristem organization and function, the opportunity arises to explore the relationship between gene expression in meristems during development and in the evolution of plant form. The establishment of phylogenetic relationships provides a framework to analyze the evolution of genes and gene families and to facilitate a comparative-developmental approach. The assign- ment of phylogenetic relationships is necessary for resolving questions of homology both at the molecular and morphological level [1, 2]. Characters are con- sidered to be homologous if they are derived from a common ancestor [31]. Orthology and paralogy are distinct forms of homology. Paralogous genes arise from duplications within an organism whereas orthol- ogous genes derive at the time of divergence between taxa [21]. Assignment of orthology can be compli- cated when genes have duplicated and diverged within an organism, making their relationships to similar genes in other organisms difficult to establish [34]. To distinguish between paralogous and orthologous genes it is necessary to determine phylogenetic relationships among all members of the gene family from the or- ganisms being compared. Within the context of such a phylogeny, ancestral conditions as well as trends in de- rived characters, such as patterns of gene expression, can be inferred. Several recent phylogenetic analyses of the MADS-box family of transcription factors in plants are excellent examples of how one can look at evolution of a gene family and assess the potential for function in morphological evolution [19, 64, 82].
-
Upload
independent -
Category
Documents
-
view
0 -
download
0
Transcript of Knots in the family tree: evolutionary relationships and functions of knox homeobox genes

Plant Molecular Biology42: 151–166, 2000.© 2000Kluwer Academic Publishers. Printed in the Netherlands.
151
Knots in the family tree: evolutionary relationships and functions ofknoxhomeobox genes
Leonore Reiser1,∗, Patricia Sanchez-Baracaldo2 and Sarah Hake11Department of Plant and Microbial Biology, University of California, Berkeley, CA 94720, USA (∗author forcorrespondence; e-mail [email protected]);2Department of Integrative Biology and University andJepson Herbaria, University of California, Berkeley, CA 94720, USA
Key words:evolution, homeodomain protein,knoxgene, meristem
Abstract
Knotted-like homeobox (knox) genes constitute a gene family in plants. Class Iknoxgenes are expressed in shootapical meristems, and (with notable exceptions) not in lateral organ primordia. Class II genes have more diverseexpression patterns. Loss and gain of function mutations indicate thatknox genes are important regulators ofmeristem function. Gene duplication has contributed to the evolution of families of homeodomain proteins inmetazoans. We believe that similar mechanisms have contributed to the diversity ofknoxgene function in plants.Knox genes may have contributed to the evolution of compound leaves in tomato and could be involved in theevolution of morphological traits in other species. Alterations incis-regulatory regions in someknoxgenes correlatewith novel patterns of gene expression and distinctive morphologies. Preliminary data from the analysis of class Iknoxgene expression illustrates the evolution of complex patterns ofknoxexpression is likely to have occurredthrough loss and gain of domains of gene expression.
Introduction
The primary architecture of plants derives from theshoot apical meristem, which produces leaves, inter-nodes and axillary buds. Seemingly simple differencesin organ initiation from the meristem, such as leafinitiation in a spiral versus decussate phyllotaxy, canresult in dramatically divergent overall morphologies.The organization and maintenance of the meristem re-mains a fundamental question in plant development.As new information emerges regarding the geneticregulation of meristem organization and function, theopportunity arises to explore the relationship betweengene expression in meristems during development andin the evolution of plant form.
The establishment of phylogenetic relationshipsprovides a framework to analyze the evolutionof genes and gene families and to facilitate acomparative-developmental approach. The assign-ment of phylogenetic relationships is necessary forresolving questions of homology both at the molecular
and morphological level [1, 2]. Characters are con-sidered to be homologous if they are derived from acommon ancestor [31]. Orthology and paralogy aredistinct forms of homology. Paralogous genes arisefrom duplications within an organism whereas orthol-ogous genes derive at the time of divergence betweentaxa [21]. Assignment of orthology can be compli-cated when genes have duplicated and diverged withinan organism, making their relationships to similargenes in other organisms difficult to establish [34]. Todistinguish between paralogous and orthologous genesit is necessary to determine phylogenetic relationshipsamong all members of the gene family from the or-ganisms being compared. Within the context of such aphylogeny, ancestral conditions as well as trends in de-rived characters, such as patterns of gene expression,can be inferred. Several recent phylogenetic analysesof the MADS-box family of transcription factors inplants are excellent examples of how one can look atevolution of a gene family and assess the potential forfunction in morphological evolution [19, 64, 82].

152
Table 1. Some representative class I and class IIknoxgenes from monocots and dicots. Partially sequenced genes suchas ESTs were not included. a, domiant; b, recessive; NR, not reported.
Gene Plant Expression Mutant phenotype(s) References
Class I-Monocotskn1 maize Apical Da: knots on leaves, [27, 39, 44, 78]
meristems blade-sheath boundary
displacement
Rb: reduced tassel
branches, fewer spikelets,
arrested shoot development.
rs1 maize Leaf D: displaced blade-sheath [8, 72]
base/internode boundary, sheath
Roots mesophyll overgrowth
knox8 maize Apical NR [39]
meristems
knox4 maize Leaf D (Gn1): displaced [23, 24]
base/internode blade-sheath boundary,
Roots sheath mesophyll
overgrowth
knox3 maize Leaf NR [39]
base/internode
knox5 maize NRa D (Lg4)? blade into [25, 44]
sheath transformation
knox11 maize NR D (Lg4)? blade into [25, 44]
sheath transformation
lg3 maize NR D (Lg3): blade into [25, 44]
sheath transformation
knox 10 maize NR NR [44]
OSH1 rice Apical meristems NR [55]
Oskn2 rice Apical NR [63]
meristems
Oskn3 rice Leaf NR [63]
base/internode
OSH15 rice leaf dwarf plants with [67]
base/internode shortened internodes,
cell identity defects
HvKnox3 barley Apical D (Hooded): ectopic inflorescences [60]
meristems form on awns
Class I-DicotsSTM Arabidopsis Apical R: arrested shoot [6, 20, 49, 50]
meristems, development, abnormal
ovary wall floral organ number
ATK1/KNAT2 Arabidopsis Apical NR [17, 47]
meristems
KNAT1 Arabidopsis Hypocotyl, NR [16, 48]
between organ
primordia
SBHI soybean Embryos, NR [51]
meristems
153
Table 1.Continued.
Gene Plant Expression Mutant phenotype(s) References
NTH15 tobacco apical NR [81]
meristems,
anthers
Knap1 apple stems NR [88]
Knap2 apple stems NR [88]
Tkn1 tomato meristems, NR [32]
leaves
Tkn2/Let6 tomato meristems, D: supercompound [15, 41, 62]
leaves, ovules leaves, ectopic shoots,
reduced apical
dominance, fasciated
inflorescence
AmSTM Antirrhinum apical NR [87]
meristem
Class I-GymnospermsHBK1 Picea albies apical NR [79]
meristem
Class II-Monocotsknox1 maize roots NR [43]
knox2 maize shoots, leaves, NR [43]
roots, embryos
knox6 maize shoots, leaves, NR [43]
roots, embryos
knox7 maize NR NR [43]
Class II-Dicotsknat3 Arabidopsis flowers NR [73, 74]
knat4 Arabidopsis leaves, roots, NR [74]
stems, etc.
knat5 Arabidopsis highest in roots NR [74]
Let12 tomato meristems, NR [41]
leaves, floral
organs, ovules
BNHD1 Brassica NR NR [11]
Knap3 apple NR NR [88]
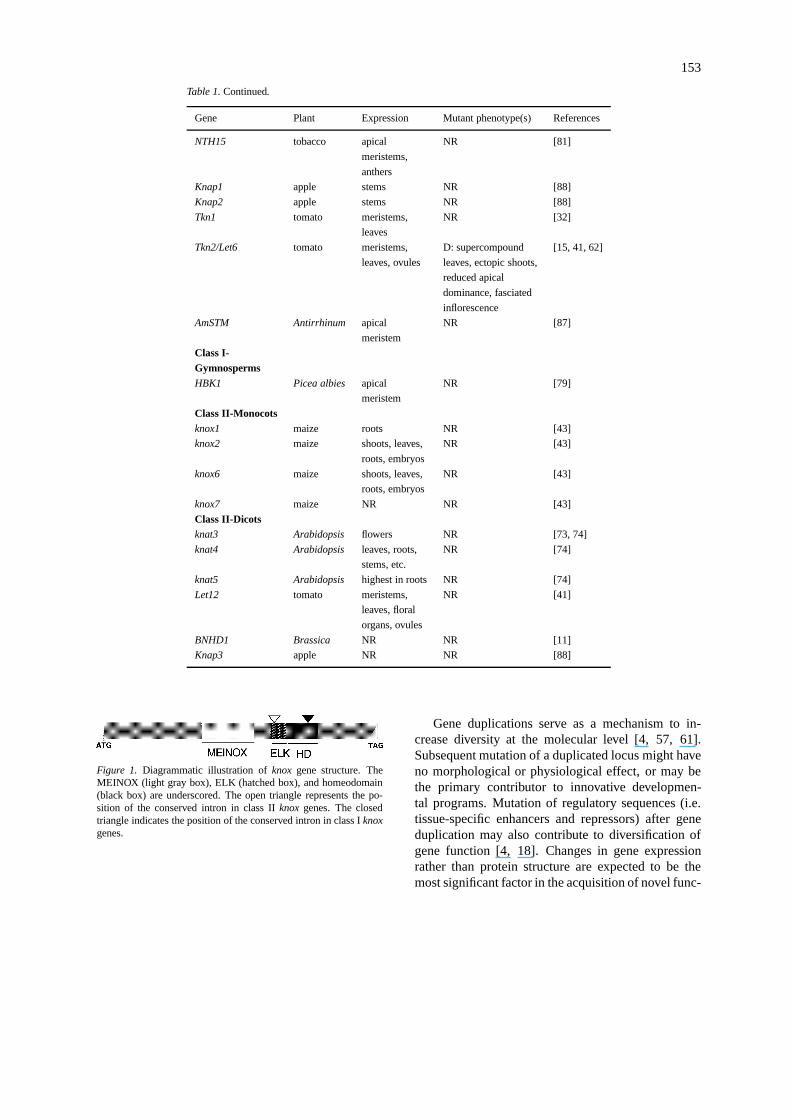
Figure 1. Diagrammatic illustration ofknox gene structure. TheMEINOX (light gray box), ELK (hatched box), and homeodomain(black box) are underscored. The open triangle represents the po-sition of the conserved intron in class IIknox genes. The closedtriangle indicates the position of the conserved intron in class Iknoxgenes.
Gene duplications serve as a mechanism to in-crease diversity at the molecular level [4, 57, 61].Subsequent mutation of a duplicated locus might haveno morphological or physiological effect, or may bethe primary contributor to innovative developmen-tal programs. Mutation of regulatory sequences (i.e.tissue-specific enhancers and repressors) after geneduplication may also contribute to diversification ofgene function [4, 18]. Changes in gene expressionrather than protein structure are expected to be themost significant factor in the acquisition of novel func-

154
tions [57]. Because coding regions may be broughtinto a new regulatory context via acquisition/deletionof regulatory elements or as a consequence of muta-tions in regulatory sequences, genes can be expressedat a new time or in a new position. In the case ofa transcription factor, expression in novel tissues cangenerate new morphologies, either through inductionof a developmental program, or through interactionswith new target genes. In addition to changes in thepattern of gene expression, mutations within proteincoding regions can result in a novel function.
The contribution of homeodomain transcriptionfactors to the evolution of form has been well docu-mented in animals [36, 37, 42, 65]. The homeoboxencodes a 61 amino acid motif (the homeodomain)that acts as a sequence-specific DNA-binding domain.Homeobox genes are found in plants, fungi and ani-mals and they are required for cell fate determinationas well as pattern definition and boundary specifica-tion [45, 56]. Homeodomain proteins also containmotifs that mediate interactions with other proteinsand these interactions are known to affect their activi-ties as transcriptional activators or repressors [52, 53].Duplication and diversification of an ancient home-obox gene cluster is associated with alterations of thebody plan of metazoans [5, 38]. Possibly, the modularnature of homeodomain proteins makes them partic-ularly amenable to rearrangements, with selection foradaptive characteristics leading to functional diversity[4].
Knotted-like homeobox (knox) genes belong to alarger superfamily, the Three Amino acid Loop Exten-sion (TALE) family [13]. In addition to the conservedELK and homeodomains, these proteins have a con-served motif termed the MEINOX domain that mayfunction in protein-protein interactions (Figure 1) [9,13, 14]. The predicted amphipathic helical region ofthe ELK domain, first described for the KNOX familyof proteins, could also act as a protein-protein inter-action domain [85].TALE genes are found in plants,animals, and fungi, indicating a common ancestralorigin [9, 13].
In plants,knoxgenes fall into two classes (Table 1).Each class is distinguished by similarity of residueswithin the homeodomain and in the positioning ofintrons (Figure 1) [43]. Phylogenetic analysis of theknoxfamily in plants places class I and class II genesinto different clades (Figure 2) [9, 10]. Since bothclades contain representatives from monocots and di-cots, a common ancestor of each class must have been
present before monocot and dicot lineages diverged[10].
The phylogeny presented in Figure 2 was gen-erated using nucleotide sequences from the home-odomain only. Other phylogenies ofknoxgenes havebeen generated using different regions of sequencefrom an overlapping data set ofknoxgenes [10]. Manyof the basic features of these phylogenies are the same,yet there are differences in the level of resolution ofeach tree and in their overall topologies. Basic pat-terns of gene expression ofknoxgenes fall into groupswithin the phylogenetic division that separates theminto two classes. Members of the class I group areexpressed in meristems and excluded from developingorgan primordia (with one exception discussed be-low). In contrast to class I genes, class II genes areexpressed in various locations throughout the plant,including lateral organs (Table 1). Loss-of-functionmutations implicate class Iknox genes in determi-nation of cell fate and patterning in the meristem.Gain-of-function mutations illustrate their capabilityto alter plant morphology profoundly. In this reviewwe evaluate theknoxgene family from both a devel-opmental and evolutionary perspective. We describecharacters that define subgroups within the family ofknoxgenes and suggest that duplication and diversi-fication has generated gene families whose membersshare related but unique functions. We also considerhow changes inknoxgene expression are manifestedin various species.
Most of the identified class Iknox genes areexpressed throughout the meristem but not inlateral organs
Meristem-specific gene expression is characteristic ofall class I knox genes. Within the meristem, theirpatterns are distinct as illustrated schematically in Fig-ure 3. Expression of some class I genes, includingkn1 from maize, is detected throughout the meristemand excluded from lateral organs (Figure 3A).kn1 isexpressed in vegetative, inflorescence and floral meris-tems [39].kn1 mRNA accumulation is first detectedin the developing maize embryo at about the time ofcoleoptile initiation [78].kn1 gene expression is notdetected in determinate lateral organ primordia.kn1is not expressed in the scutellum which is consid-ered the grass cotyledon, nor in the coleoptile whichis considered by some to be the ensheathing base ofthe scutellum [89]. Althoughkn1 is continuously ex-

155
Figure 2. Phylogenetic analysis ofknoxgenes. Nucleotide sequence of the homeobox was used to align sequences for analysis which were thenanalyzed using PAUP∗ 4.0 [80]. The tree represents a strict consensus of the three most parsimonious trees. We did a heuristic search. Bootstrapanalysis was performed with 100 replicates. Branches without values assigned had less than 50% bootstrap support. Accession numbers/sourcesfor the sequences are as follows:Ceratopteris(Hasebe and Banks, personal communication),KNAT1(Arabidopsis thaliana, U14174),kn1(Zeamays, X61308),Oshkn2(Oryza sativa, AF050180),HvKnox3(Hordeum vulgare, AF022390),rs1 (Zea mays, L44133),OSH1(Oryza sativa,D16507),SBH1(Glycine max, 13663),Tkn2/Let6(Lycopersicon esculentum, AF000141),STM(A. thaliana, U32344),KNAT2(A. thaliana,U14175),POTH1(Solanum tuberosum, U65648),Let12(L. esculentum, AF000142),NTH23(Nicotiana tabacum, AB004797),Knap3(Malusdomestica, Z71980),KNAT3 (A. thaliana, X92392),KNAT4 (A. thaliana, X92393),KNAT5 (A. thaliana, X92394),Mrg1a (Mus musculus,U68383),CEH-25 (Caenorhabditis elegans, AJ000533),X-Meis1-3(Xenopus laevis, U68388).knox3, knox8, knox4, lg3, knox11, knox5,knox1, knox2, knox6, knox7 Zea mays: sequences were obtained within our lab (Kerstetter, Vollbrecht, Lowe and Hake, unpublished results).
pressed in the shoot apical meristems it is negativelyregulated in some cells within the meristem where thenext leaf primordium is initiated. This region corre-sponds to the plastochron 0 (P0) leaf which has yet toundergo the anticlinal divisions that form the moundcharacteristic of a P1 leaf primordium [46].
All of our kn1 loss-of-function alleles display asimilar phenotype [44]. In the tassel, which bears themale florets, fewer branches and spikelets are formed,resulting in a sparse appearance. Within the femalereproductive structure, the ear, loss-of-function plantsmake fewer branches and florets. Loss ofkn1functionalso causes extra silks and abnormalities during ovuledevelopment. Other less penetrant phenotypes are as-sociated with recessive mutations ofkn1, such as theintitation of multiple leaves from the same node. Thisphenotype has been interpreted as a failure to repressdifferentiation of cells within the meristem [44]. Amore dramatic phenotype is detected in some inbredlines where ca. 50% of the seedlings fail to progress
Figure 3. Schematic representation of the three major types of geneexpression seen forknoxgenes. The meristem outlined has a disti-chous phyllotaxy. A.kn1-like expression throughout the meristemand excluded from P0 and the L1 layer. B.rs1-like gene expressionin the meristem is restricted to positions between the lateral organs.C. knoxgene pattern showing expression throughout the meristemincluding the L1 and P0 is similar toknoxexpression in tomato.
beyond the coleoptile stage (Vollbrechtet al., manu-script in preparation). In the permissive inbred lines,germinated kernels elaborate a coleoptile and ceasefurther development (Figure 4, compare panels A and

156
B). Instead of a meristematic dome, these arrestedshoots have a flattened area that occupies the spacewhere the shoot apical meristem should be (Vollbrechtet al., manuscript in preparation). Taken together,the expression pattern ofkn1and the loss-of-functionphenotypes indicate a function forkn1 in meristemmaintenance.
Why do presumably null alleles that lack a func-tional kn1 gene show such a variable, background-dependent phenotype? One possible explanation isthat loss ofkn1 function in maize is compensated forby the activity of a duplicate locus whose activity isvariable in different inbred lines. Maize is thought tobe an ancient allotetraploid and a large percentage ofthe genome is duplicated [28, 33]. Although no du-plicate factor forkn1 has been identified, the regionof chromosome 1L wherekn1 resides is duplicatedon chromosome 5S [33]. An alternative explanation isthat the redundant function is supplied by one or morerelated class Iknoxgenes. It is equally probable thatthe lack of an absolute requirement forkn1function insome backgrounds is due to the activity of a gene orgenes that are unrelated tokn1.
TheSHOOTMERISTEMLESS(STM) gene ofAra-bidopsisshares many similarities withkn1. Like kn1,STM is expressed specifically in meristems and isdown-regulated in the P0 [50].STMmRNA first ac-cumulates in a single cell of the globular embryo andexpression gradually expands to fill the space betweenthe cotyledon primordia [49].STMexpression remainsin the meristem throughout vegetative and floral de-velopment, and later includes the ovary wall [50].HomozygousSTM mutant seedlings lack a visibleshoot apical meristem and have cotyledons with fusedpetioles [6, 20]. One interpretation of theSTMphe-notype suggests that in the absence ofSTM function,a meristem is never formed [6, 49]. Another possibil-ity is that STMprevents precocious determination ofthe meristem after it is initiated. Meristems terminateprematurely in partial loss of function or weak alle-les ofSTM[20]. These weak alleles are also defectivein flower development, producing fused and extra flo-ral organs. These data also suggest a role forSTM inmeristem maintenance.
In manyArabidopsisecotypes theSTMphenotypecan also be suppressed (K. Barton, personal com-munication) implying that a background-dependentredundant function can compensate for loss ofSTMfunction. Are similar genes acting to modify the lossof knoxgene function in meristems in both maize andArabidopsisor are the mechanisms different between
these two species? Establishing the molecular identityof theseknoxgene modifiers will answer this question.
Expression patterns of otherknoxgenes in maizeandArabidopsisare similar to that ofkn1 andSTM.knox8 expression in the maize apex overlaps com-pletely with that ofkn1 [39]. KNAT2 (also known asAKT1) mRNA is detected in the vegetative and flo-ral meristems ofArabidopsis[17, 48]. Based uponanalysis of promoter::β-glucoronidase (GUS) fusions,expression ofKNAT2/AKT1 in the vegetative meris-tem is restricted to the L3 layer [47]. Therefore,KNAT2/AKT1is expressed in a subset of cells that alsoexpressSTM. These genes may share similar functionswith kn1 and STM and thus may be candidates forredundant factors.
Based upon similarity of protein sequence andgene expression patterns it has been suggested thatkn1 andSTMare orthologues. Phylogenetic analysisshows thatSTM is not theArabidopsisorthologue ofkn1because it resolves into a separate clade (Figure 2)[10]. So far orthologues ofSTMhave been only foundin dicots while orthologues ofkn1are found in mono-cots. Incomplete representation of gene families couldalso lead to uncertainty in establishing orthology. Pos-sibly not all of theknoxgenes have been identified inArabidopsisand maize.
The orthology of STM to the dicot genes,Tkn2/Let6 (tomato) andSBH1(soybean) is well sup-ported by phylogenetic analysis (Figure 2) [10]. Adifferent, more limited analysis also placesNTH15,a knox gene from tobacco, in the same group withTkn2/Let6, STMandSBH1[81]. STM, Tkn2/Let6andNTH15 are all expressed in a similar pattern in themeristem with the exception that the tomato gene isalso expressed in leaves (see below).
Orthologues ofkn1 have been identified in othermonocots. mRNA accumulation ofOSH1(rice) andHvKnox3 (barley) is restricted to the meristem andexcluded from organ primordia [51, 55, 60]. No loss-of-function mutations have been described for thesegenes. Phylogenetic analysis supports the hypothesisthatOSH1andHvKnox3are orthologues ofkn1(Fig-ure 2; [10]). Some grasses share extensive regions ofsimilarity at the molecular level within their individualgenomes and between genomes [58]. This synteny ofgrass genomes provides additional evidence to supportthe orthology ofkn1, HvKnox3andOSH1[10].

157
Figure 4. kn1loss-of-function mutant and normal sibling. A. Normal seedling 2 weeks after germination. These plants had made an average of3 visible leaves. B.kn1-E1seedling in W23 inbred background 2 weeks after germination. Only the coleoptile (co) emerged from the kernel.No other apical organs were produced.
Other Class I knox genes are expressed in specificdomains within the meristem
mRNA of a second group of class Iknoxgenes tendsto be excluded from the central region of the meristemand instead accumulates in the periphery, between lat-eral organs (Figure 3B).rough sheath1(rs1), knox4andknox3are all expressed in meristem cells whichappear to correspond to the internode in maize [39,72]. Later expression in the inflorescence and floralmeristems resembles that ofkn1. No recessive loss-of-function mutant phenotype has been described foreitherrs1, knox4or knox3. knox4andrs1 map to du-plicated regions of the maize genome and are likely tobe duplicate factors [28, 33, 58]. If the duplication wasrecent (i.e. at the time of the duplication of the maizegenome), thenknox4andrs1 may not have had timeto diverge significantly and they may share redundantfunctions. In that case, a mutant phenotype may onlybe apparent in thers1/knox4double-mutant combina-tion. Clonal analysis of bothrs1 andknox4(Gnarley)dominant gain-of-functionmutations indicates thatrs1acts non-autonomouslywhereas part ofknox4action is
partially cell-autonomous which may indicate that atleast some functional divergence has occurred [8, 23].
Rice is a true diploid and therefore should haveonly single copies of eachknoxgene. At least eightknox genes have been isolated from rice cDNA li-braries [66].Oskn3andOSH15, which were clonedindependently [63, 68], are identical to each other innucleotide sequence and very similar tors1 [63]. Inyoung embryos,Oskn3/OSH15is expressed through-out the meristem. Later in development, expres-sion is restricted to the boundary between the lat-eral organs similar to the expression ofrs1/knox4 inmaize. Four independent loss-of-function mutationsof OSH15were isolated which are defective in inter-node elongation [67]. Within the internodes of mutantplants, epidermal and hypodermal cellular morphol-ogy is defective; the cells are abnormally shaped andsome cell types are absent. These data suggest thatOSH15has a role in specifying cell fate in the riceinternode.OSH15maps to a region of the genomethat is syntenous with the maize locations ofrs1 andknox4 [3]. Oskn3/OSH15is likely an orthologue ofknox4/rs1. If so, it should fall into theknox4/rs1

158
clade in a phylogentic analysis. Within the contextof our own phylogeny this relationship remains un-resolved, possibly due to limitations of informationwhen only the homeodomain is used [10]. Other phy-logenies clearly show thatrs1/knox4are in the sameclade, however these analyses did not includeOSH15[10]. If gene function was conserved between maizeand rice, then thers1/knox4loss-of-function double-mutant-phenotype might resemble that of null mutantsin OSH15.
In Arabidopsis, KNAT1 mRNA accumulates invegetative meristems at the base of the point of leafinsertion and in the cortex of the shoot apex [48]. Thispattern of expression is similar to that ofrs1/knox4andknox3in maize.KNAT1RNA is not found in theincipient lateral organ primordia, nor the inflorescenceor floral meristem [48]. Based on sequence similar-ity and expression patternsKNAT1 is most similar toknox4andrs1. However, phylogenetic analysis placesKNAT1in a separate clade suggesting thatKNAT1rep-resents an independent lineage (Figure 2) [10]. If thisis true, then perhapsKNAT1andrs1/knox4are exam-ples of convergence in expression pattern. At this time,we can only say that the pattern of gene expressionin the vegetative apex is similar, not thatrs1/KNAT1function in the same way. Because maize andAra-bidopsismeristems are different,KNAT1 expressionmay mark a different domain than that defined byrs1/knox4expression. Our data suggest thatKNAT1and theCeratopteris knoxgene are orthologues. In thiscase, the divergence of class I and class II genes iseven more ancient than we had previously supposed.Another phylogenetic analysis that does not includethe fern sequences indicatesTkn1to be an orthologueof KNAT1[10].
Class I knox genes in tomato are expressed inmeristems and leaves
In contrast to other class Iknox genes, genes frommembers of the tomato family are expressed in in-cipient leaf primordia as well as in meristems (seeFigure 3C). Tkn1 and Tkn2/Let6 mRNAs are notdown-regulated in the P0 and continue to be expressedin immature leaves [15]. The orthology ofTkn2/Let6andSTMis well supported whileTkn1orthology withKNAT1has been indicated but is less well supported[10]. Tkn2/Let6andTkn1, however, have a pattern ofgene expression unlike eitherSTMor KNAT1.
Of all theknoxgene family members, the ones intomato are most unique in their patterns of gene ex-pression. What are the consequences of this pattern ofknoxgene expression in tomatoes? The class I genesTkn1andTkn2/Let6are not down-regulated in the leafprimordia. Interestingly, the related geneNTH15fromtobacco is negatively regulated in the P0 [81]. Thissuggests that the difference in expression pattern oc-curred after the divergence of tobacco from the lineleading to tomato. It has been suggested thatknoxgeneexpression in leaves may have contributed to the evo-lution of compound leaves in tomato [40, 75]. Thispossibility can be tested by superimposing leaf shapeand the patterns of gene expression on a phylogenyof Tkn1/Tkn2orthologues in selected members of theSolanaceae that have different leaf morphologies.
How has the expression ofknox genes evolved?Derived and ancestral character states can be recon-structed when these characters are superimposed uponthe phylogeny [12]. Using the related TALE home-odomain proteinsBELL1 (BEL1) andARABIDOPSISTHALIANA HOMEOBOX 1(ATH1) as an outgroup,root-specific expression was mapped onto aknoxhomeodomain phylogeny [10]. These results indicatethat root-specific expression was lost twice inknoxgene evolution; once in the lineage leading to class Iknoxgenes and once inATH1. The analysis also showsa gain of root specific expression in the lineage leadingto knox4/rs1.
We have mapped meristem expression patternsonto a knox gene tree usingkn1/STM-like orKNAT1/rs1-like patterns of gene expression as char-acter states (Figure 5). The tree represents a portionof a larger phylogenetic analysis that included all se-quencedknoxgenes fromArabidopsisand maize usingCEH-25, Xmeis1–3andMrg1a genes as the outgroup(Sánchez-Baracaldo, unpublished results). As shownin Figure 5,KNAT1, 2 andSTMare paralogous geneswithin Arabidopsisas arekn1, rs1/knox4, knox3andknox8in maize. That is, each group arose from dupli-cations that occurred after speciation. Consequently,each group is orthologous to the other. Based upon ourphylogeny, several models could explain the evolutionof gene expression patterns in maize andArabidopsis.As stated above, we ascribe the similarity of expres-sion pattern betweenKNAT1andknox4/rs1 to parallelevolution. Within the maize group, reconstruction ofcharacter states forknox3, knox4/rs1 and knox8 isequivocal. Theknox3, knox4/rs1 pattern could havearisen early and have been lost in the lineage leading

159
Figure 5. Character state analysis of meristem-specific patterns of expression. Only class I representatives from maize andArabidopsisareshown. This figure represents a branch of a larger tree consisting of only maize andArabidopsisgenes. The tree was generated from nucleotidesequence of the homeodomain using PAUP∗ 4.0 [80], with the same outgroups as in Figure 1. This was the single and most parsimonioustree. Character analysis was performed using MacClade (Version 3) and character states are unordered. We reconstructed the node states usinginformation from extant genes as there is no comparable information available for closely related genes. The most related genes are fromanimals (see Bharathanet al. [10]), and other plant homeobox genes are more distantly related. We did not feel confident that including theexpression patterns of these genes in an outgroup analysis was appropriate.
to knox8. It is equally possible thatknox3, knox4/rs1patterns of expression arose independently.
Neither of the analyses described above is proofof the ancestral state forknox gene expression pat-terns. What they do provide are hypotheses that canbe tested as new sequence and expression pattern dataemerge. One hypothesis based upon our characterreconstruction is thatkn1-type meristem-specific ex-pression represents the ancestral state. We need to fillin the gaps for expression pattern and sequence in themodel systemsArabidopsisand maize as well as moredistantly related taxa.
Recent work has begun to address the gaps in ourunderstanding ofknoxgenes in non-flowering plants.
A knoxgene from spruce (HBK1) was recently clonedand is similar to class Iknoxgenes suggesting that thedivergence of class I and class II genes occurred beforethe split between gymnosperms and angiosperms [79].In situ localization ofHBK1 mRNA in the meristemshowed expression in the internal cell layers and not inthe needles. Threeknoxgenes that are similar tokn1have been cloned from the fernCeratopteris(Hasebeand Banks, personal communication). Based on RNAgel blots, two of theseknoxgenes are highly expressedin the shoot apical region of the sporophyte and mod-erately expressed in immature leaves (fiddleheads). Atruncated form of oneCeratopteris knoxgene is alsoexpressed in the gametophyte (Hasebe and Banks, per-

160
sonal communication). We have used an antibody toKN1 to immunolocalize KNOX proteins in represen-tatives of non-flowering plant groups. Our preliminaryresults show that KNOX proteins are found throughoutthe apical regions of bothGnetumand Lycopodiumand do not appear to be restricted to specific celltypes (Sánchez-Baracaldo, unpublished results). Weare using PCR to cloneknoxgenes fromGnetumandLycopodiumin order to determine their phylogeneticrelationships and patterns of gene expression withgene-specific probes.
Class II knox genes in maize andArabidopsishavea more general pattern of expression
The pattern of mRNA accumulation for a number ofclass IIknoxgenes has been determined. In general,class II genes have a more diverse pattern of geneexpression than class I genes (see Table 1). Geneexpression ofknox1, -6 and -7 was assayed withnorthern blots [43].knox6 and -7 are expressed inleaves, stems, inflorescences, meristems and rootswhileknox1is expressed only in roots. InArabidopsis,three class II genes,KNAT3, 4 and5, have been iden-tified to date, and are expressed in a wide variety oftissues [74]. mRNA accumulation was determined bynorthern analysis.KNAT3 andKNAT4 are expressedin most tissues whileKNAT5 is primarily found inroots. Analysis of aKNAT3promoter-GUSfusion in-dicates thatKNAT3 is expressed in lateral organs, butnot in meristems [73]. The sequence of tomatoLet12ismost similar toKNAT3andKNAT4[41]. Like KNAT4,Let12mRNA accumulation appears to be ubiquitous atthe organ level.In situ localization of mRNA in floralorgans indicatesLet12 is expressed in all four floralwhorls and the ovule [41]. No mutant phenotypes havebeen associated with loss of function of any class IIknoxgenes.
The variation in gene expression patterns for classII genes suggests diverse roles for members of thisfamily. Perhaps our inability to identify a mutationin any of these genes is due to redundant functionsshared by members of this class that mask a loss offunction phenotype. Alternatively, loss of function ofsome class II genes may have either a subtle effector such a strong effect on development that they arelethal.
Effects of gain of function ofknox genes on plantmorphology
Dominant mutations ofknox genes in maize leavesare characterized by ectopic expression of the mu-tant gene, resulting in perturbation of the blade-sheathboundary. Maize leaves are divided into the distalblade and proximal sheath region. The ligule (an epi-dermal outgrowth) and auricle are positioned at theboundary between the blade and sheath.Kn1, Rs1,Liguleless3(Lg3) andGnarley1(Gn1) mutations sharea characteristic displacement of sheath and ligule [8,23]. Other features distinguish each class of muta-tion. Kn1mutations are characterized by vein clearingand knot formation over lateral veins which is inter-preted as another shift of sheath character onto theblade (see Figure 6A) [27]. All dominantKn1 mu-tations are associated with ectopic expression ofkn1mRNA and protein along the vasculature in youngleaves [30, 77]. In addition to alterations of the blade-sheath boundary,Rs1 mutants have reduced statureand excessive proliferation of auricle mesophyll cells[8]. In contrast to their normal siblings,rs1 mRNA isdetectable in leaf primordia as early as P2 (the sec-ond visible leaf primordium) inRs1-Omutant plants[72]. Transposon-induced revertants ofRs1-Owereobtained and mapped, but the genetic nature of thedominant phenotype is not known.Gn1mutants, likeRs1, display shortening of the internodes, prolifera-tion of sheath mesophyll cells as well as displacementof the blade-sheath boundary (Figure 6B).Gn1mapsto the same position asknox4, the putative duplicatefactor for rs1 [22]. Gn1 ectopically expressesknox4mRNA and protein in P2–P8/9 leaves suggesting thata mutation inknox4 is responsible for the dominantGn1phenotype [24]. A polymorphism 5′ to theknox4gene cannot be genetically uncoupled from theGn1mutant phenotype [24]. Another dominant leaf muta-tion, Lg3-O, is a class I knox gene [43].Lg3-O alsotransforms blade, auricle and ligule tissue into sheath-like tissue [25, 26, 59].Lg3 is most closely related tothe duplicated lociknox5andknox11(Figure 1).
Expression of proximal characters in the distal leafblade can be interpreted in a variety of ways [30,59]. A consistent theme is the failure to acquire aspecific developmental fate. That is, ectopic expres-sion ofknoxgenes interferes with normal processes ofdifferentiation.

161
Figure 6. Phenotypes of plants misexpressingknoxgenes. A. Leaf blade ofKn1plant showing knots forming on the lamina. B.Gn1mutant leaf.Excessive cellular proliferation of the sheath mesophyll causes buckling and contortions of the sheath(s). The blade (b) is relatively unaffected.C. Flower from aHoodedbarley plant showing ectopic floral meristems (arrowheads) forming on the awn. D. Leaf lobing and ectopic meristems(arrowheads) on leaves ofArabidopsisplants expressing 35S::KNAT1. E. Main axis of a tobacco plant expressing a 35S::KNAT2 transgene.Leaves are very reduced and malformed. F. Tobacco leaf from a transgenic plant overexpressingKNAT1with ectopic meristems. G. A singleleaf from a 35S::kn1 tomato plant showing many additional orders of compounding.
Ectopic expression of theHvknox3gene in barleyalters floral development
Hooded, a dominant mutation of barley, is causedby the duplication of intron sequences in the class Iknox geneHvknox3 [60]. Ectopic Hvknox3expres-sion in the lemma, which is the first lateral organ ofthe floret, causes formation of florets on the awn (anoutgrowth of the lemma; see Figure 6C). AHoodedphenocopy is induced whenkn1 expression in barleyis introduced under the control of a ubiquitin promoter
[91]. Williams-Carrier suggested that, in response toectopicknoxgene expression, the upper leaf zone ofthe lemma forms an ectopic inflorescence from whichthe flowers are produced [91]. In this regard overex-pression ofkn1 in barley resembles that of otherknoxgene overexpressors that make ectopic meristems onleaves (see below). Why barley forms ectopic meris-tems in the presence ofHvKnox3is unknown but couldbe dependent upon interactions with genes expressedin the awn.

162
Phenotypes due toknox gene mutations andoverexpression in dicots are different frommonocots
The effects ofknoxgene overexpression in dicots aredistinct from those in monocots and may be attribut-able to differences in competence of tissues to respondto, or modulate the function of, KNOX proteins [90].TransgenicArabidopsisplants that constitutively ex-pressknox genes have highly lobed leaves and canhave ectopically placed meristems in the sinuses ofthese lobes (Figure 6D) [16, 48]. Ectopic expres-sion in tobacco of severalknox genes driven by theconstitutive cauliflower mosaic virus (CaMV) 35Spromoter result in leaf phenotypes such as rumpling,reduced lamina, and formation of ectopic shoots onthe leaves (Figure 6E, F) [55, 63, 69, 76, 81]. Adramatic response to ectopicknoxgene expression isseen in tomato. Transgenic plants overexpressing bothendogenous and heterologous gene constructs makesupercompound leaves (Figure 6G). Other phenotypesin tomato include reduced apical dominance, reducedlaminae, and ectopic shoot development [32, 40].
The way in which genes are misexpressed mayaffect the phenotype. The tomato dominant mutationCurl (Cu) is characterized by the formation of super-compound leaves and epiphyllic shoots among otherphenotypes [62]. TheMouse ears(Me) mutation in-duces changes in leaf and overall architecture that aresimilar to but distinct fromCu [62]. Cu andMe areboth genetically linked toTkn2. Tkn2mRNA showsa five-fold increase in expression inCu mutant plantscompared to normal sibs suggesting that levels ratherthan the pattern ofTkn2 gene expression are altered[62]. Two labs have shown a correlation betweenthe Me phenotype and altered spatial regulation ofTkn2/Let6 [15, 62]. In Me, the Tkn2 coding regionis fused to a constitutively expressed gene. The re-sult is high-level expression of a fusion mRNA thatis ectopically localized [15, 62]. It was suggested thatectopic expression versus overexpression contributesto the differences in phenotype betweenMe and Cu[62].
Within each species there are often a range of phe-notypes in the transgenic plants expressing differentknox transgenes. This could be due to variation intransgene expression or differences in genetic back-grounds of the transformants. Differences might alsoreflect unique specificities of individualknox genes[40, 90]. Each KNOX protein may interact with dif-
ferent protein partners and thus target distinct DNAsequences.
What makes certain tissues competent to respondto knoxgenes? Why do ectopic meristems form alongthe midvein of the lamina in tobacco, in the sinusesof lobed leaves inArabidopsisand on the awn of bar-ley flowers? Why doeskn1 make knots in maize butnot barley leaves? Other genes must influence the ef-fect of ectopicknox gene expression. For example,CaMV 35S::Tkn1does not correct the phenotypes ofLanceolate, solanifolia and other tomato leaf shapemutants indicating that overexpression ofknoxgenesalone is not sufficient to induce the formation ofcompound leaves in tomato [32].
Gain and loss of function ofknoxgenes can affectoverall plant stature, leaf shape, and floral develop-ment in a variety of species. From these data, we caninfer potential sites where changes inknoxgene ex-pression may have influenced the evolution of plantdevelopmental processes. Dramatic effects ofknoxexpression on leaf morphology led to a hypothesisthat knoxgenes may be involved in the evolution ofcompound leaves in tomato [32, 75]. How general isthe effect ofknox expression in developing leaves?Compound leaves arose multiple times via differentmechanisms. Therefore, not all cases of compoundleaves would necessarily be attributable to changes inknox gene expression. For example, in pea, the de-gree of leaf compounding is controlled byunifoliata,a leafyhomologue [35].
The dominant mutations ofknox genes in maizeand tomato are good examples of how mutations canalter expression domains and consequently affect mor-phogenesis. For example, theKn1-Oallele is a tandemduplication ofkn1 in which a second coding region isassociated with a novel 5′-upstream region [54, 84].Many of the otherKn1 alleles characterized to dateare associated with insertions of transposable elementsinto the large third intron [29]. These transposon inser-tions point to at least two potential regulatory domainswithin the kn1 genomic region that determinekn1expression in leaves. Based upon the phenotype of cer-tain dominant and revertantKn1alleles we suspect thepresence of leaf silencing elements at the 5′ end of thegene and in the large third intron [29, 54]. Mutationof either of these regulatory domains or of second-siteloci that interact with these domains could alter thespatial or temporal pattern ofkn1gene expression.
But what does the analysis ofknoxgene expres-sion evolution tell us about morphological evolution inplants? While thecis-regulatory changes such as those

163
described forknoxgenes in maize and tomato clearlyalter plant morphology, there is no firm evidenceimplicating knox genes in morphological evolution.Changes in the regulation ofknoxgene expression mayor may not be due to specific changes at theknoxlocuswhich alter its regulation. Baum made the very impor-tant point that ectopic expression of a specific genedoes not by itself define the innovative force in theevolution of form [7]. It is first necessary to have phy-logenetic evidence that observed differences in geneexpression are meaningful in terms of morphologicalevolution. Then it is appropriate to identifycisor transeffects on gene expression to provide mechanistic ex-planations for a change that defines the proximatecause of novelty in gene expression.
Mechanisms to repressknox gene expression maybe similar in monocots and dicots
An emerging theme in the expression of class Iknoxgenes is their negative regulation in the meristem priorto organ initiation. To date, with the exception oftomato, all class Iknoxgenes are excluded from lateralorgan primordia. A significant feature of all the ectopicexpression studies is that despite the fact that the trans-genes are presumably under the control of constitutivepromoters, the transgene is not detected uniformlythroughout the plants. While it is possible that thesedifferences are a consequence of transgene silencingin specific domains, the most probable explanation ispost-transcriptional regulation ofknox accumulation[91].
The recessiverough sheath2mutation in maize hasa phenotype similar to that of dominantknox genemutations [71]. Ectopic expression ofrs1, knotted1,and liguleless3can be detected as early as P1 inrs2mutant leaf primoridia. Therefore,rs2 acts to nega-tively regulateknox genes in immature leaves [71].However, ectopicknoxgene expression is not seen inP0 leaves and is not uniform throughout the leaves.The ability to negatively regulateknoxgenes in the P0and later stages of development inrs2 mutants maybe due to the presence and function of a duplicatefactor forrs2. Other loci may be required, such asnar-rowsheath(ns) and leafbladeless(lbl), both of whichare thought to participate in founder cell recruitment[70, 83]. Double mutants betweenns and rs2 wereadditive, indicating that these genes act in separatepathways to restrictknoxgene expression in the leafprimordia [71].rs2 was cloned and the predicted pro-
tein encodes a myb-like transcription factor similar tothephantastica(phan) gene fromAntirrhinum[71, 83,87]. Lateral organs inphan mutants are abaxializedand have radial symmetry [86]. Thephanexpressiondomain complements that of anAntirrhinum kn1-likegene, consistent with a role in delimiting the pa-rameters ofknox gene expression [87]. These datasuggest that a common mechanism to regulateknoxgene expression exists inAntirrhinumand maize.
Concluding remarks
Knox genes are known to play important roles inmeristems and are likely to function in other tissues.In situ localization shows differences in patterns ofgene expression that likely reflect novel functions forduplicated genes. Class II genes have more diversepatterns of expression suggesting that members of thisgroup have functions outside of the meristem.Cis- andtrans-regulatory changes inknoxgene expression arecorrelated with alterations in plant morphology. Dupli-cations and diversification ofknoxgenes have likelycontributed to changes in organization of meristemsand features of lateral organs.
Acknowledgements
We thank Neelima Sinha for providing the image ofthe 35S::kn1tomato plant leaf in Figure 6G and GeetaBharathan for allowing us to preview unpublished re-sults. We are indebted to Toby Kellogg and the anony-mous reviewers whose comments greatly enhancedthis review. Our thanks to Brent Mishler for manydiscussions and invaluable assistance with the phylo-genetic analysis and to D.L. Swofford for the beta testversion of PAUP∗4.0. We are grateful to Paul Bethke,George Chuck, Naomi Ori, Mark Running, and Kris-ten Shepard for comments on this manuscript. Workin S.H.’s lab onknoxgenes is funded by USDA grant9701255. L.R. is supported by NIH post-doctoral fel-lowship 18330-03. P.S-B. is funded by NSF GRANTDEB-9801245 and a Rimo Bacigalupi Fellowship.
References
1. Abouheif E: Developmental genetics and homology: a heirar-chical approach. Trends Evol Ecol 12: 405–408 (1997).
2. Abouheif E, Akam M, Dickinson W, Holland PWH, MeyerA, Patel N, Raff R, Roth VL, Wray GA: Homology anddevelopmental genes. Trends Genet 13: 432–433 (1997).

164
3. Ahn S, Tanksley SD: Comparative linkage maps of the riceand maize genomes. Proc Natl Acad Sci USA 90: 7980–7984(1993).
4. Averof M, Akam M: Hox genes and the diversification of in-sect and crustacean body plans. Nature 376: 420–423 (1995).
5. Averof M, Dawes R, Ferrier D: Diversification of arthropodHOX genes as a paradigm for the evolution of gene functions.Cell Dev Biol 7: 539–551 (1996).
6. Barton MK, Poethig RS: Formation of the shoot apical meris-tem inArabidopsis thaliana: an analysis of development in thewild type and in the shoot meristemless mutant. Development119: 823–831 (1993).
7. Baum DA: The evolution of development. Curr Opin PlantBiol 1: 79–86 (1998).
8. Becraft PW, Freeling M: Genetic analysis ofRough sheath1developmental mutants of maize. Genetics 136: 295–311(1994).
9. Bharathan G, Janssen B-J, Kellogg EA, Sinha N: Did home-odomain proteins duplicate before the origin of angiosperms,fungi and metazoa? Proc Natl Acad Sci USA 94: 13749–13753 (1997).
10. Bharathan G, Janssen B-J, Kellogg EA, Sinha N: Phylogeneticrelationships and evolution of the KNOTTED class of planthomeodomain proteins. Mol Biol Evol, in press (1999).
11. Boivin R, Hamel F, Beauseigle D, Bellemare G: Stage-specifictranscription of the homeobox geneBnhd1 in young tissuesand flowers ofBrassica napus. Biochim Biophys Acta 1219:201–204 (1994).
12. Brooks D, McLennan: Phylogeny, Ecology and Behavior.University of Chicago Press, Chicago (1991).
13. Bürglin TR: Analysis of TALE superclass homeobox genes(MEIS, PBC, Iriquois, TGIF) reveals a novel domain con-served between plants and animals. Nucl Acids Res 25:4173–4180 (1997).
14. Burglin TR: The PBC domain contains a MEINOX domain:coevolution of Hox and TALE homeobox genes. Dev GenesEvol 208: 113–116 (1998).
15. Chen J-J, Janssen B-J, Williams A, Sinha N: A gene fusion ata homeobox locus: alterations in leaf shape and implicationsfor morphological evolution. Plant Cell 9: 1289–1304 (1997).
16. Chuck G, Lincoln C, Hake S:KNAT1 induces lobed leaveswith ectopic meristems when overexpressed inArabidopsis.Plant Cell 8: 1277–1289 (1996).
17. Dockx J, Quaedvlieg N, Keultjes G, Kock P, Weisbeek P,Smeekens S: The homeobox geneATK1 of Arabidopsisthaliana is expressed in the shoot apex of the seedling andin flowers and inflorescence stems of mature plants. Plant MolBiol 28: 723–737 (1995).
18. Doebley J, Lukens L: Transcriptional regulators and the evo-lution of plants. Plant Cell 10: 1075–1082 (1998).
19. Doyle J: Evolution of a plant homeotic multigene family:toward connecting molecular systematics and molecular de-velopmental genetics. Syst Biol 43: 307–328 (1994).
20. Endrizzi K, Moussian B, Haecker A, Levin JZ, Laux T: TheSHOOT MERISTEMLESSgene is required for maintenance ofundifferentiated cells inArabidopsisshoot and floral meris-tems and acts at a different regulatory level than the meristemgenesWUSCHELandZWILLE. Plant J 10: 101–113 (1996).
21. Fitch WM: Distinguishing homologous from analogous pro-teins. Syst Zool 19: 99–113 (1970).
22. Foster T, Hake S: It’s a Gnarley one! (Gn1). Maize Genet CoopNewsl 68: 2 (1994).
23. Foster T, Veit B, Hake S: Mosaic analysis of the dominantmutantGnarley1-Rreveals distinct lateral and transverse sig-
naling pathways during maize leaf development. Development126: 305–313 (1999).
24. Foster T, Yamaguchi J, Wong B, Veit B, Hake S:Gnarley1isa dominant mutation in theknox4homeobox gene that affectscell identity and cell shape. Plant Cell, in press (1999).
25. Fowler JE, Freeling M: Genetic analysis of mutations that altercell fates in maize leaves: dominantLigulelessmutations. DevGenet 18: 198–222 (1996).
26. Fowler JE, Meuhlbauer GJ, Freeling M: Mosaic analysis oftheLiguleless3mutant phenotype in maize by coordinate sup-pression of mutator insertion alleles. Genetics 143: 489–503(1996).
27. Freeling M, Hake S: Developmental genetics of mutants thatspecify Knotted leaves in maize. Genetics 111: 617–634(1985).
28. Gaut BS, F. DJ: DNA sequence evidence for the segmentalallotetraploid origin of maize. Proc Natl Acad Sci USA 94:6809–6814 (1997).
29. Greene B, Walko R, Hake S:Mutator insertions in an intronof the maizeknotted1gene result in dominant suppressiblemutations. Genetics 138: 1275–1285 (1994).
30. Hake S, Char BR, Chuck G, Foster T, Long J, Jackson D:Homeobox genes in the functioning of plant meristems. PhilTrans R Soc Lond 350: 45–51 (1995).
31. Hall BK: Homology: The Hierarchical Basis of ComparativeMorphology. Academic Press, San Diego (1994).
32. Hareven D, Gutfinger T, Parnis A, Eshed Y, Lifschitz E: Themaking of a compound leaf: genetic manipulation of leafarchitecture in tomato. Cell 84: 735–744 (1996).
33. Helentjaris T, Weber D, Wright S: Identification of the ge-nomic locations of duplicate nucleotide sequences in maizeby analysis of restriction fragment length polymorphisms.Genetics 118: 353–363 (1988).
34. Hillis DM: Homology in molecular biology. In: Hall BK (ed),Homology, the Hierarchical Basis of Comparative Morphol-ogy, pp. 339–368. Academic Press, San Diego (1994).
35. Hofer J, Turner L, Hellens R, Ambrose M, Matthews P,Michael A, Ellis N: UNIFOLIATA regulates leaf and flowermorphogenesis in pea. Curr Biol 7: 581–587 (1997).
36. Holland P: Homeobox genes and vertebrate evolution. BioEs-says 14: 267–273 (1992).
37. Holland PW, Garcia-Fernandez J:Hox genes and chordateevolution. Dev Biol 173: 382–395 (1996).
38. Holland PW, Garcia-Fernandez J, Williams NA, Sidow A:Gene duplications and the origins of vertebrate development.Development (Suppl): 125–133 (1994).
39. Jackson D, Veit B, Hake S: Expression of maizeKNOT-TED1 related homeobox genes in the shoot apical meristempredicts patterns of morphogenesis in the vegetative shoot.Development 120: 405–413 (1994).
40. Janssen B-J, Lund L, Sinha N: Overexpression of a home-obox gene,Let6, reveals indeterminate features of the tomatocompound leaf. Plant Physiol 117: 771–786 (1998).
41. Janssen B-J, Williams A, Chen J-J, Mathern J, Hake S,Sinha N: Isolation and characterization of two knotted-likehomeobox genes from tomato. Plant Mol Biol 36: 417–425(1998).
42. Kenyon C: If birds can fly, why can’t we? Homeotic genes andevolution. Cell 78: 175–180 (1994).
43. Kerstetter R, Vollbrecht E, Lowe B, Veit B, Yamaguchi J, HakeS: Sequence analysis and expression patterns divide the maizeknotted1-like homeobox genes into two classes. Plant Cell 6:1877–1887 (1994).

165
44. Kerstetter RA, Laudencia-Chingcuanco D, Smith LG, HakeS: Loss of function mutations in the maize homeobox gene,knotted1, are defective in shoot meristem maintenance. Devel-opment 124: 3045–3054 (1997).
45. Krumlauf R:Hox genes in vertebrate development. Cell 78:191–201 (1994).
46. Lane B, Freeling M: The maize leaf. In: Freeling M, WalbotV (eds), The Maize Handbook, Springer-Verlag, New York(1994).
47. Laufs P, Dockx J, Kronenberger J, Traas J: MGOUN1 andMGOUN2: two genes required for primordium initiation atthe shoot apical and floral meristems inArabidopsis thaliana.Development 125: 1253–1260 (1998).
48. Lincoln C, Long J, Yamaguchi J, Serikawa K, Hake S: Aknotted1-like homeobox gene inArabidopsisis expressed inthe vegetative meristem and dramatically alters leaf morphol-ogy when overexpressed in transgenic plants. Plant Cell 6:1859–1876 (1994).
49. Long J, Barton MK: The development of apical embryonicpattern inArabidopsis. Development 125: 3027–3035 (1998).
50. Long JA, Moan EI, Medford JI, Barton MK: A member of theKNOTTED class of homeodomain proteins encoded by theSHOOTMERISTEMLESS gene ofArabidopsis. Nature 379:66–69 (1996).
51. Ma H, McMullen MD, Finer JJ: Identification of a homeobox-containing gene with enhanced expression during soybean(Glycine maxL.) somatic embryo development. Plant MolBiol 24: 465–473 (1994).
52. Mann R, Chan S-K: Extra specificty fromextradenticle: thepartnership between HOX and PBX/EXD homeodomain pro-teins. Trends Genet 12: 258–262 (1996).
53. Mann RS, Affolter M: Hox proteins meet more partners. CurrOpin Genet Dev 8: 423–429 (1998).
54. Mathern J, Hake S:Mu element-generated gene conversions inmaize attenuate the dominantKnotted1phenotype. Genetics147: 305–314 (1997).
55. Matsuoka M, Ichikawa H, Saito A, Tamda Y, Fujimura T,Kano-Murakami Y: Expression of a rice homeobox genecauses altered morphology of transgenic plants. Plant Cell 5:1039–1048 (1993).
56. McGinnis W, Krumlauf R: Homeobox genes and axial pattern-ing. Cell 68: 283–302 (1992).
57. Meyer A: The evolution of body plans: HOM/hoxcluster evo-lution, model systems, and the importance of phylogeny. In:Harvey PH (ed), New Uses for New Phylogenies, pp. 99–113.Oxford University Press, London (1996).
58. Moore GK, Devos KM, Z W, Gale MD: Grasses, line up andform a circle. Curr Biol 5: 737–739 (1995).
59. Muehlbaur GJ, Fowler JE, Freeling M: Sectors expressing thehomeobox geneliguless3implicate a time-dependent mecha-nism for cell fate acquisition along the proximal-distal axis ofthe maize leaf. Development 124: 5097–5106 (1997).
60. Müller K, Romano N, Gerstner O, Garcia-Maroto F, Pozzi C,Salamini F, Rohde W: The barleyHoodedmutation caused bya duplication in a homeobox gene intron. Nature 374: 727–730(1995).
61. Ohno S: Evolution by Gene Duplication. Springer-Verlag,Heidelberg (1970).
62. Parnis A, Cohen O, Gutfinger T, Hareven D, Zamir D, Lif-schitz E: The dominant developmental mutants of tomato,Mouse-Earand Curl, are associated with distinct modes ofabnormal transcriptional regulation of aKnottedgene. PlantCell 9: 2143–2158 (1997).
63. Postma-Haarsma AD, Verwoert IIGS, Stronk OP, Koster J,Lamer G, EM, Hoge HC, Meijer AH: Characterization of theKNOX class homeobox genesOskn2andOskn3identified ina collection of cDNA libraries covering the early stages of riceembryogenesis. Plant Mol Biol 39: 257–271 (1999).
64. Purugganan M, Rounsley S, Schmidt R, Yanofsky M: Molec-ular evolution of flower development: diversification of a plantMADS-box regulatory gene family. Genetics 140: 345–356(1995).
65. Ruddle FH, Bartels JL, Bentley KL, Kappen C, Murtha MT,Pendelton JW: Evolution ofHox genes. Annu Rev Genet 28:423–442 (1994).
66. Sato Y, Sentoku N, Matsuoka M: Analysis of the functionsof rice homeobox genes. In: Shimamoto K (ed), MolecularBiology of Rice, pp. 119–135. Springer-Verlag, Tokyo (1999).
67. Sato Y, Sentoku N, Miura Y, Hirochicka H, Kitano H, Mat-suoka M: Loss-of-function mutations in the rice homeoboxgene,OSH15, cause defects in the development of hypodermalschlerenchyma in internodes resulting in dwarf plants. EMBOJ 18: 992–1002 (1999).
68. Sato Y, Sentoku N, Nagato Y, Matsuoka M: Isolation and char-acterization of a rice homeobox geneOSH15. Plant Mol Biol38: 983–998 (1998).
69. Sato Y, Tamaoki M, Murakami T, Yamamoto N, Kano-Murakami Y, Matsuoka M: Abnormal cell divisions in leafprimordia caused by the expression of the rice homeoboxgeneOSH1lead to altered morphology of leaves in transgenictobacco. Mol Gen Genet 251: 13–22 (1996).
70. Scanlon MJ, Schneeberger RG, Freeling M: The maize mu-tant narrow sheath fails to establish leaf margin identity in ameristematic domain. Development 122: 1683–1691 (1996).
71. Schneeberger R, Tsantis M, Freeling M, Langdale JA: Therough sheath2gene negatively regulates homeobox gene ex-pression during maize leaf development. Development 125:2857–2865 (1998).
72. Schneeberger RG, Becraft PW, Hake S, Freeling M: Ectopicexpression of theknox homeobox generough sheath1alterscell fate in the maize leaf. Genes Dev 9: 2292–2304 (1995).
73. Serikawa KA, Martinez-Laborda A, Kim H-S, Zambryski P:Localization of expression ofKNAT3, a class 2knotted1-likegene. Plant J 11: 853–861 (1997).
74. Serikawa KA, Martinez-Laborda A, Zambryski P: Threeknotted1-like homeobox genes inArabidopsis. Plant Mol Biol32: 673–683 (1996).
75. Sinha N: Simple or compound leaves: reduction or multiplica-tion? Trends Plant Sci 2: 396–402 (1997).
76. Sinha NR, Williams RE, Hake S: Overexpression of the maizehomeo box gene,KNOTTED-1, causes a switch from deter-minate to indeterminate cell fates. Genes Dev 7: 787–795(1993).
77. Smith LG, Greene B, Veit B, Hake S: A dominant mutationin the maize homeobox gene,Knotted-1, causes its ectopicexpression in leaf cells with altered fates. Development 116:21–30 (1992).
78. Smith LG, Jackson D, Hake S: The expression ofKnotted1marks shoot meristem formation during maize embryogenesis.Dev Genet 16: 344–348 (1995).
79. Såndas-Larsson A, Svenson M, Liao H, Engstrom P: A home-obox gene with potential developmental control function in themeristem of the coniferPicea abies. Proc Natl Acad Sci USA95: 15118–15122 (1998).
80. Swofford DL: Phylogenetic Analysis Using Parsimony. Illi-nois Natural History Survey, Champaign, IL (1991).

166
81. Tamaoki M, Kusuba S, Kano-Murakami Y, Matsuoka M:Ectopic expression of a tobacco homeobox gene,NTH15,dramatically alters leaf morphology and hormone levels intransgenic tobacco. Plant Cell Physiol 38: 917–927 (1997).
82. Theiben G, Kim J, Saedler H: Classification and phylogenyof the MADs box multigene family suggest defined roles ofMADS box gene subfamilies in the morphological evolutionof eukaryotes. J Mol Evol 43: 484–516 (1996).
83. Timmermans MCP, Schutes NP, Jankovsky JP, Nelson T:Leaf-bladeless1is required for dorsiventrality of lateral organs inmaize. Development 125: 2813–2823 (1998).
84. Veit B, Vollbrecht E, Mathern J, Hake S: A tandem duplicationcauses theKn1-Oallele ofKnotted, a dominant morphologicalmutant of maize. Genetics 125: 623–631 (1990).
85. Vollbrecht E, Veit B, Sinha N, Hake S: The developmentalgeneKnotted-1is a member of a maize homeobox gene family.Nature 350: 241–243 (1991).
86. Waites R, Hudson A:phantastica, a gene required for dor-siventrality in leaves ofAntirrhinum majus. Development 121:2143–2154 (1995).
87. Waites R, Selvadurai HRN, Oliver IR, Hudson A: ThePHAN-TASTICAgene encodes a MYB transcription factor involvedin growth and dorsiventrality of lateral organs inAntirrhinum.Cell 93: 779–789 (1998).
88. Watillon B, Kettmann R, Boxus P, Burney A:Knotted1-likehomeobox genes are expressed during apple tree (Malus do-mesticaL. Borkh) growth and development. Plant Mol Biol33: 757–763 (1997).
89. Weatherwax P: Position of the scutellum and homology of thecoleoptile in maize. Bot Gaz 69: 179–182 (1920).
90. Williams RW: Plant homeobox genes: many functions stemfrom a common motif. BioEssays 20: 280–282 (1998).
91. Williams-Carrier RE, Lie YS, Hake S, Lemaux PG: Ectopicexpression of the maizekn1 gene phenocopies theHoodedmutant of barley. Development 124: 3737–3745 (1997).




















