
Isolation, culture and characterisation of somatic cells derived from semen and milk of endangered...
9
CSIRO PUBLISHING Reproduction, Fertility and Development, 2007, 19, 576–584 www.publish.csiro.au/journals/rfd Isolation, culture and characterisation of somatic cells derived from semen and milk of endangered sheep and eland antelope L. Nel-Themaat A,B , M. C. Gómez A,B , P. Damiani B , G. Wirtu B,C , B. L. Dresser B,D , K. R. Bondioli A,F , L. A. Lyons E , C. E. Pope B and R. A. Godke A,C A Department of Animal Sciences, Louisiana State University Agricultural Center, Baton Rouge, LA 70803, USA. B Audubon Center for Research of Endangered Species, 14001 River Road, New Orleans, LA 70131, USA. C Department of Veterinary Clinical Sciences, Louisiana State University School of Veterinary Medicine, Baton Rouge, LA 70803, USA. D Department of Biological Sciences, University of New Orleans, New Orleans, LA 70148, USA. E School ofVeterinary Medicine, University of California—Davis, Davis, CA 95616, USA. F Corresponding author. Email: [email protected] Abstract. Semen and milk are potential sources of somatic cells for genome banks. In the present study, we cultured and characterised cells from: (1) cooled sheep milk; (2) fresh, cooled and frozen–thawed semen from Gulf Coast native (GCN) sheep (Ovis aries); and (3) fresh eland ( Taurotragus oryx) semen. Cells attached to the culture surface from fresh (29%), cooled (43%) and slow-frozen (1 ◦ C/min; 14%) ram semen, whereas no attachment occurred in the fast- frozen (10 ◦ C/min) group. Proliferation occurred in fresh (50%) and cooled (100%) groups, but no cells proliferated after passage 1 (P1). Eland semen yielded cell lines (100%) that were cryopreserved at P1. In samples from GCN and cross- bred milk, cell attachment (83% and 95%, respectively) and proliferation (60% and 37%, respectively) were observed. Immunocytochemical detection of cytokeratin indicated an epithelial origin of semen-derived cells, whereas milk yielded either fibroblasts, epithelial or a mixture of cell types. Deoxyribonucleic acid microsatellite analysis using cattle-derived markers confirmed that eland cells were from the semen donor. Eland epithelial cells were transferred into eland oocytes and 12 (71%), six (35%) and two (12%) embryos cleaved and developed to morulae or blastocyst stages, respectively. In conclusion, we have developed a technique for obtaining somatic cells from semen.We have also demonstrated that semen-derived cells can serve as karyoplast donors for nuclear transfer. Additional keywords: cell culture, cryopreservation, ejaculate, epithelial, Gulf Coast native sheep, nuclear transfer. Introduction Several mammalian species are facing the threat of extinc- tion as a result of vanishing ecosystems, expansion of human populations and an increase in human-mediated interferences (Blackburn 2004). Declining population numbers of many threatened species have brought into prominence the importance of maintaining the genetic diversity of remaining small popu- lations with limited gene pools. This is extremely important in managing captive populations, in which the influx of new genetic material is generally limited (Ryder and Benirschke 1997). As a result, genome resource banking (GRB) has assumed an increas- ingly important role in the field of conservation biology. Cryop- reservation and storage of gametes (spermatozoa and oocytes) and embryos from genetically valuable animals are key compo- nents of biological resource banks (BRBs). However, gametes and embryos are finite in their use and have to be replaced constantly if used on a regular basis, which is not possible if donor animals are inaccessible or dead. Therefore, the cryop- reservation of viable somatic cell tissues is quickly assuming an increasingly important role in BRBs (Ryder 2002). Although these tissue samples are also finite, they can be propagated using specific culture conditions before being cryopreserved and stored for many years (Green 1967). The birth of the first mammal following transfer of embryos derived by somatic cell nuclear transfer (SCNT) using an adult donor cell (Wilmut et al. 1997) demonstrated that the nucleus of an adult somatic cell maintains its totipo- tency. Since then, the totipotency of somatic cells from non-domestic species has also been demonstrated by produc- ing viable cloned animals in, for example, the gaur (Lanza et al. 2000), mouflon (Loi et al. 2001), deer (http://www. cnn.com/2003/TECH/science/12/22/cloned.deer.ap/index.html/, © CSIRO 2007 10.1071/RD06153 1031-3613/07/040576
-
Upload
independent -
Category
Documents
-
view
2 -
download
0
Transcript of Isolation, culture and characterisation of somatic cells derived from semen and milk of endangered...

CSIRO PUBLISHING
Reproduction, Fertility and Development, 2007, 19, 576–584 www.publish.csiro.au/journals/rfd
Isolation, culture and characterisation of somatic cellsderived from semen and milk of endangered sheepand eland antelope
L. Nel-ThemaatA,B, M. C. GómezA,B, P. DamianiB, G. WirtuB,C, B. L. DresserB,D,K. R. BondioliA,F, L. A. LyonsE, C. E. PopeB and R. A. GodkeA,C
ADepartment of Animal Sciences, Louisiana State University Agricultural Center,Baton Rouge, LA 70803, USA.
BAudubon Center for Research of Endangered Species, 14001 River Road,New Orleans, LA 70131, USA.
CDepartment of Veterinary Clinical Sciences, Louisiana State University School of VeterinaryMedicine, Baton Rouge, LA 70803, USA.
DDepartment of Biological Sciences, University of New Orleans, New Orleans, LA 70148, USA.ESchool of Veterinary Medicine, University of California—Davis, Davis, CA 95616, USA.FCorresponding author. Email: [email protected]
Abstract. Semen and milk are potential sources of somatic cells for genome banks. In the present study, we culturedand characterised cells from: (1) cooled sheep milk; (2) fresh, cooled and frozen–thawed semen from Gulf Coast native(GCN) sheep (Ovis aries); and (3) fresh eland (Taurotragus oryx) semen. Cells attached to the culture surface fromfresh (29%), cooled (43%) and slow-frozen (1◦C/min; 14%) ram semen, whereas no attachment occurred in the fast-frozen (10◦C/min) group. Proliferation occurred in fresh (50%) and cooled (100%) groups, but no cells proliferated afterpassage 1 (P1). Eland semen yielded cell lines (100%) that were cryopreserved at P1. In samples from GCN and cross-bred milk, cell attachment (83% and 95%, respectively) and proliferation (60% and 37%, respectively) were observed.Immunocytochemical detection of cytokeratin indicated an epithelial origin of semen-derived cells, whereas milk yieldedeither fibroblasts, epithelial or a mixture of cell types. Deoxyribonucleic acid microsatellite analysis using cattle-derivedmarkers confirmed that eland cells were from the semen donor. Eland epithelial cells were transferred into eland oocytesand 12 (71%), six (35%) and two (12%) embryos cleaved and developed to morulae or blastocyst stages, respectively.In conclusion, we have developed a technique for obtaining somatic cells from semen. We have also demonstrated thatsemen-derived cells can serve as karyoplast donors for nuclear transfer.
Additional keywords: cell culture, cryopreservation, ejaculate, epithelial, Gulf Coast native sheep, nuclear transfer.
Introduction
Several mammalian species are facing the threat of extinc-tion as a result of vanishing ecosystems, expansion of humanpopulations and an increase in human-mediated interferences(Blackburn 2004). Declining population numbers of manythreatened species have brought into prominence the importanceof maintaining the genetic diversity of remaining small popu-lations with limited gene pools. This is extremely important inmanaging captive populations, in which the influx of new geneticmaterial is generally limited (Ryder and Benirschke 1997). As aresult, genome resource banking (GRB) has assumed an increas-ingly important role in the field of conservation biology. Cryop-reservation and storage of gametes (spermatozoa and oocytes)and embryos from genetically valuable animals are key compo-nents of biological resource banks (BRBs). However, gametesand embryos are finite in their use and have to be replaced
constantly if used on a regular basis, which is not possible ifdonor animals are inaccessible or dead. Therefore, the cryop-reservation of viable somatic cell tissues is quickly assuming anincreasingly important role in BRBs (Ryder 2002). Althoughthese tissue samples are also finite, they can be propagatedusing specific culture conditions before being cryopreserved andstored for many years (Green 1967).
The birth of the first mammal following transfer of embryosderived by somatic cell nuclear transfer (SCNT) using anadult donor cell (Wilmut et al. 1997) demonstrated thatthe nucleus of an adult somatic cell maintains its totipo-tency. Since then, the totipotency of somatic cells fromnon-domestic species has also been demonstrated by produc-ing viable cloned animals in, for example, the gaur (Lanzaet al. 2000), mouflon (Loi et al. 2001), deer (http://www.cnn.com/2003/TECH/science/12/22/cloned.deer.ap/index.html/,
© CSIRO 2007 10.1071/RD06153 1031-3613/07/040576

Somatic cells cultured from semen and milk Reproduction, Fertility and Development 577
accessed 17 April 2007), banteng (Janssen et al. 2004),African wildcat (Gómez et al. 2004) and water buffalo (http://news.xinhuanet.com/english/2005-03/21/content_2724026.htm,accessed 30 April 2007). These results indicated that SCNT is apotentially viable technique for the conservation of endangeredspecies.
Fibroblast cells derived mostly from skin samples are widelyused as somatic donor cells for SCNT. In addition, alternativesources of somatic cells include those derived from hair follicles(Arase et al. 1990; Kurata et al. 1994; Moll 1996), colostrum(Kishi et al. 2000) or blood (Elicha Gussin and Elias 2002).Freshly lactated milk and ejaculated semen contain large num-bers of somatic cells (Paape and Tucker 1966; Evenson andMelamed 1983). Likewise, the totipotency after SCNT of mam-mary gland epithelial cells isolated from milk colostrum hasbeen demonstrated by the birth of live cloned calves (Kishi et al.2000). Somatic cells isolated from endangered species from milkand ejaculated semen should be considered as potential donorcell sources and included in BRBs.
In the present study, attempts were made to isolate, cultureand characterise somatic cells derived from: (1) fresh, cooledand frozen–thawed semen of the Gulf Coast native (GCN) sheep(Ovis aries); (2) fresh semen of an eland antelope (Taurotragusoryx); and (3) fresh sheep milk. In addition, an effort was madeto determine the ability of eland antelope somatic cells isolatedfrom semen to dedifferentiate in eland cytoplasts after SCNT.
Materials and methodsExperimental designFor ram semen, the cryopreservation treatments were: (1) none(i.e. fresh semen); (2) cooled; (3) −1◦C/min cooling rate (slowfreezing); and (4) −10◦C/min cooling rate (fast freezing). Thesetreatments are detailed below. The variables measured were thenumber of attempts in which cells attached, proliferated, prolifer-ated until passage was performed and proliferation after passage.A total of seven replicates (collections) was performed.
For milk, two breed types (GCN and GCN × Suffolk cross-bred) were compared and the number of attempts in whichcells attached and proliferated were measured. The cell linesobtained were characterised by immunohistochemical detectionof vimentin and cytokeratin.
AnimalsThe GCN rams and ewes, as well as GCN × Suffolk cross-bred ewes, all between 2 and 5 years of age, were housedat the Louisiana State University (LSU) Agricultural CenterSheep Research Unit, whereas the eland bull was housed at theFreeport–McMoRan Audubon Species Survival Center (FMA-SSC). All sheep and eland procedures were approved by theInstitutional Animal Care and Use Committee of LSU andFMA-SSC.
ChemicalsAll chemicals, including antibodies for immunocytochemistry,were obtained from Sigma Chemical (St Louis, MO, USA),unless stated otherwise.
Semen collectionSemen was collected from two GCN rams and one 6-year-oldcommon eland antelope bull with a bodyweight of approximately530 kg. Ram semen was collected from the two 4-year-old rams(body condition scores of 3 and 4) by electroejaculation (Evansand Maxwell 1987), whereas eland semen was collected by acombination of rectal massage and electroejaculation (Wirtuet al. 2002). Each ram semen sample was divided into four treat-ments before somatic cell isolation (fresh, cooled, slow freezingand fast freezing), whereas eland somatic cells were isolatedfrom fresh semen only.
Treatment 1For fresh samples, an aliquot (0.5 mL) of fresh ram (n = 7)
or eland (n = 2) semen was diluted in the presence of seminalplasma with minimum essential medium α medium (α-MEM;Gibco BRL, Grand Island, NY, USA) containing 15% newborncalf serum (NCS; Gibco) and supplemented with 100 IU mL−1
penicillin G and 0.1 mg mL−1 streptomycin (10 µL mL−1; Cell-gro, Herndon, VA, USA). Diluted semen was incubated at 39◦Cfor no more than 5 h before somatic cell isolation, because semenwas treated according to the standard Salamon ram semen freez-ing procedure (Evans and Maxwell 1987) as if it was to be frozen.
Treatment 2After an aliquot of fresh semen sample was taken, each ejacu-
late was allowed to cool to 30◦C and then extended in Salamon’sstandard Tris-glucose extender supplemented with 15% (v/v)egg yolk and 5% (v/v) glycerol (Evans and Maxwell 1987).Additional extender was added to obtain a final concentrationof 200 × 106 spermatozoa mL−1 and a glycerol concentrationof 4–5%.Then, extended semen was slowly cooled to 4◦C for 2 h.After cooling, approximately one-third of the extended cooledsemen was used for somatic cell isolation and the remainingtwo-thirds of the sample was used for cryopreservation.
Treatment 3For slow freezing, cooled semen (one-quarter of the ejaculate)
was loaded into 1.0 mL cryovials (1 mL per cryovial) and cooledat 1◦C/min to −80◦C/min (Mr. Frosty; Nalgene, Rochester, NY,USA) before storage in liquid nitrogen (LN2).
Treatment 4For fast freezing, extended semen (one-quarter of the ejacu-
late) was loaded into 0.5 mL straws (IMV, Maple Grove, MN,USA; n = 2–5 straws, depending on ejaculate volume) and,after sealing the tip, each straw was placed into the chamberof a controlled-rate freezing unit (Model CL-863; Cryologic,Melbourne, Victoria, Australia) at 4◦C, cooled at −10◦C/min to−80◦C and plunged into LN2 for storage.
Milk collectionSheep milk samples (n = 26) were collected by hand from pureGCN (n = 5) and cross-bred (n = 10) ewes. Two collectionswere performed from each animal, but bacterial contaminationoccurred in four samples from GCN ewes and these were omit-ted from the experiment. After collection, milk was cooled to4◦C and stored for 12 h.

578 Reproduction, Fertility and Development L. Nel-Themaat et al.
Somatic cell isolation, culture and cryopreservationCryopreserved semen straws and cryovials were thawed in awater bath at 37◦C for 1 or 3 min, respectively, and trans-ferred to 15-mL centrifuge tubes immediately before processing.Cooled semen and milk samples were processed within 12 hof collection. Fresh (0.5 mL), cooled (one-third of extendedejaculate) and frozen–thawed semen (two to five cryovials orstraws) and cooled milk (15–50 mL) were washed in 10 mLCa2+- and Mg2+-free Dulbecco’s phosphate-buffered saline(DPBS; Gibco) and centrifuged (500g) for 10 min. Pelletedsamples of each semen treatment were resuspended in 2 mLα-MEM supplemented with 15% NCS, 1500 IU mL−1 peni-cillin, 5 mg mL−1 streptomycin, 250 µg mL−1 gentamicin and500 IU mL−1 polymixin B, placed in a 60-mm tissue culturedish (Falcon 1007; Becton Dickinson, Franklin Lakes, NJ, USA),covered with a layer of Type 1 collagen (from calf skin) and cul-tured at 38.5◦C in 5% CO2 in air. Pelleted milk was resuspendedin 2 mL Dulbecco’s modified Eagle’s medium (DMEM; Gibco)supplemented with 15% fetal bovine serum (FBS), 100 IU mL−1
penicillin and 0.1 mg mL−1 streptomycin, placed in 60-mmtissue culture dishes and cultured at 38.5◦C in 10% CO2 in air.
After 24 h incubation, dishes from each treatment were rinsedonce and culture medium was replaced every 3–4 days. Disheswere searched periodically for the presence of attached cellsusing an inverted microscope (100×). When cell colonies cov-ered approximately 0.5 cm2 of the dish surface (proliferated),they were disaggregated. Somatic cells obtained from semensamples were disaggregated by 15 min of incubation in PBS at38.5◦C, followed by 20 min incubation in 200 IU mL−1 colla-genase (because preliminary studies showed that epithelial cellsmay be sensitive to trypsin). Somatic cells obtained from milkwere dissociated by incubation in 0.25% trypsin until all the cellswere dissociated from the surface of the culture dish.
Dissociated cells were centrifuged (500g) for 10 min, resus-pended in α-MEM (semen somatic cells) or DMEM (milksomatic cells) and cultured in 35-mm collagen-coated dishesor 75-cm2 tissue culture flasks for sperm or milk-derived cells,respectively (passaged). When cells from each sample reachedconfluence, cells were passaged once or twice before beingresuspended in tissue culture medium (α-MEM or DMEM) with15% NCS and 10% (v/v) dimethyl sulfoxide (DMSO) and cooledat 1◦C/min to −80◦C (Mr. Frosty) before storage in LN2.
Cell characterisation and microsatellite analysisCell characterisation was performed on cells that were obtainedduring preliminary studies from fresh and cooled GCN ramsemen and from fresh eland semen during the current exper-iment. A fraction of cells from each established cell line wascultured on a microscope coverslip.Then, immunohistochemicaldetection of cell types was performed by fluorescent labellingof cytokeratin and vimentin to identify epithelial and fibroblastcells, respectively (Katska et al. 2002). Briefly, cells were cul-tured (as described above) on sterilised microscope coverslipsor Biocoat collagen-coated culture slides (Becton Dickson) andfixed in 3.7% paraformaldehyde, permeabilised for 10 min in1% Triton X-100, diluted in PBS (PBS-T), rinsed twice in 0.1%PBS–Tween 20 (PBS-T20) and stabilised with 0.1 m glycine
solution. Primary monoclonal mouse antibodies were: (1) 1 : 100anti-β-tubulin (positive control); (2) 1 : 100 anti-vimentin (forlabelling fibroblast cells); and (3) 1 : 200 anti-pan cytokeratin(for labelling epithelial cells).
Incubation of somatic cells with the primary antibodies wasperformed on a shaking platform overnight at room temperature(RT). After incubation, somatic cells were rinsed three times for10 min per rinse in PBS-T20.
A negative control was also included by labelling only with asecondary antibody that was added and incubated overnight ona shaking platform at RT. The β-tubulin- and vimentin-labelledslides were incubated in anti-mouse IgG-tetramethylrhodamineisothiocyanate (TRITC) conjugate, whereas the negative con-trol and cytokeratin-labelled slides were incubated in anti-mouseIgG-fluorescein isothiocyanate conjugate, both at 1 : 64 dilu-tions in PBS-T20. The next day, slides were rinsed three timesfor 10 min per rinse in PBS-T20 before mounting in glycerolcontaining 0.5 µg mL−1 Hoechst 33342. Under epifluorescencemicroscopy, epithelial cells labelled with cytokeratin fluorescedgreen, whereas fibroblast cells labelled for vimentin fluorescedred.
To determine the genetic status of the established cell lines,DNA was extracted from a blood sample and epithelial cellsfrom the semen sample of the eland sperm donor using theDNAeasy Kit (Qiagen, Valencia, CA, USA). Deoxyribonu-cleic acid was also isolated from a blood sample of a second,unrelated eland. Fourteen microsatellite markers (BM1818,BM1824, BM2113, BRR, CYP21, ETH10, ETH225, INRA023,RM006, RM067, SPS115, TGLA122, TGLA126, TGLA227)that were derived from cattle were tested for efficacy in theeland (www.isag.org.uk/ISAG/all/02_PVpanels_LPCGH.doc;Creighton et al. 1992; Kossarek et al. 1993a, 1993b; Bishop et al.1994). These markers were labelled fluorescently for analysiswithABI instrumentation (Applied Biosystems, Foster City, CA,USA). Then, approximately 10 ng genomic eland DNA (2 µLextracted DNA) was used per 10 µL polymerase chain reaction(PCR) reaction. The PCR parameters included an initial 3 mindenaturation at 94◦C followed by 35 cycles of 1 min denaturationat 94◦C, annealing for 1 min at 58◦C and a 72◦C extension for1 min, followed by a final extension at 72◦C for 10 min. Afterthe initial amplification, the products were denatured at 95◦Cfor 3 min; the PCR products were separated on an ABI Prism377 DNA Analyzer after the addition of 2 µL formamide load-ing dye with GeneScan 350 ROX (GeneScan, Foster City, CA,USA) size standard to approximately 0.1 µL of the PCR product.The allele sizes and genotypes were determined using STRandsoftware (Toonen and Hughes 2001).
Oocyte maturation, nuclear transfer and embryo cultureTo obtain eland oocytes, two females were treated withaltrenogest orally (0.044 mg/kg Regumate; Intervet, Millsboro,DE, USA) for 7 days and, on the last day of altrenogesttreatment, prostaglandin F2α (25 mg, i.m.; Lutalyse; Pharma-cia and Upjohn, Kalamazoo, MI, USA) was administered.Ovarian follicular stimulation was induced using porcinefollicle-stimulating hormone (FSH; equivalent to 400 mg NIH;Folltropin; Bionche Animal Health Canada, Belleville, Ontario,Canada) dissolved in 30% polyvinyl pyrrolidone.
Somatic cells cultured from semen and milk Reproduction, Fertility and Development 579
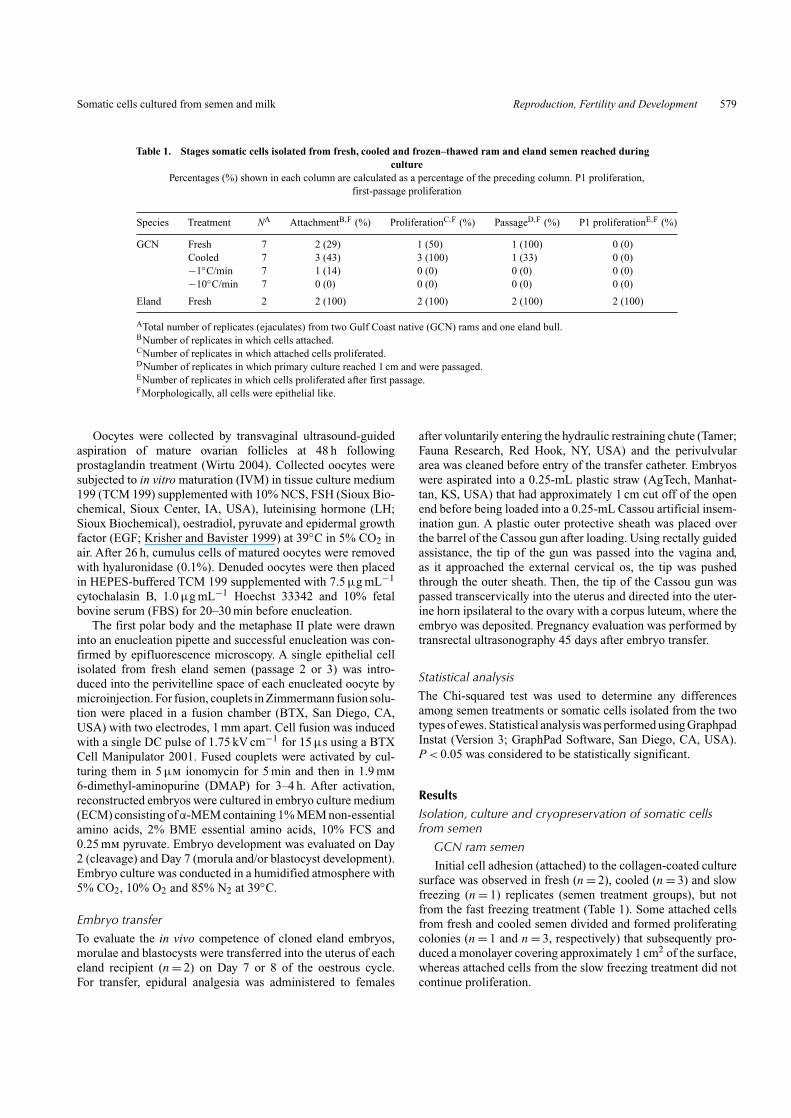
Table 1. Stages somatic cells isolated from fresh, cooled and frozen–thawed ram and eland semen reached duringculture
Percentages (%) shown in each column are calculated as a percentage of the preceding column. P1 proliferation,first-passage proliferation
Species Treatment NA AttachmentB,F (%) ProliferationC,F (%) PassageD,F (%) P1 proliferationE,F (%)
GCN Fresh 7 2 (29) 1 (50) 1 (100) 0 (0)Cooled 7 3 (43) 3 (100) 1 (33) 0 (0)−1◦C/min 7 1 (14) 0 (0) 0 (0) 0 (0)−10◦C/min 7 0 (0) 0 (0) 0 (0) 0 (0)
Eland Fresh 2 2 (100) 2 (100) 2 (100) 2 (100)
ATotal number of replicates (ejaculates) from two Gulf Coast native (GCN) rams and one eland bull.BNumber of replicates in which cells attached.CNumber of replicates in which attached cells proliferated.DNumber of replicates in which primary culture reached 1 cm and were passaged.ENumber of replicates in which cells proliferated after first passage.FMorphologically, all cells were epithelial like.
Oocytes were collected by transvaginal ultrasound-guidedaspiration of mature ovarian follicles at 48 h followingprostaglandin treatment (Wirtu 2004). Collected oocytes weresubjected to in vitro maturation (IVM) in tissue culture medium199 (TCM 199) supplemented with 10% NCS, FSH (Sioux Bio-chemical, Sioux Center, IA, USA), luteinising hormone (LH;Sioux Biochemical), oestradiol, pyruvate and epidermal growthfactor (EGF; Krisher and Bavister 1999) at 39◦C in 5% CO2 inair. After 26 h, cumulus cells of matured oocytes were removedwith hyaluronidase (0.1%). Denuded oocytes were then placedin HEPES-buffered TCM 199 supplemented with 7.5 µg mL−1
cytochalasin B, 1.0 µg mL−1 Hoechst 33342 and 10% fetalbovine serum (FBS) for 20–30 min before enucleation.
The first polar body and the metaphase II plate were drawninto an enucleation pipette and successful enucleation was con-firmed by epifluorescence microscopy. A single epithelial cellisolated from fresh eland semen (passage 2 or 3) was intro-duced into the perivitelline space of each enucleated oocyte bymicroinjection. For fusion, couplets in Zimmermann fusion solu-tion were placed in a fusion chamber (BTX, San Diego, CA,USA) with two electrodes, 1 mm apart. Cell fusion was inducedwith a single DC pulse of 1.75 kV cm−1 for 15 µs using a BTXCell Manipulator 2001. Fused couplets were activated by cul-turing them in 5 µm ionomycin for 5 min and then in 1.9 mm6-dimethyl-aminopurine (DMAP) for 3–4 h. After activation,reconstructed embryos were cultured in embryo culture medium(ECM) consisting of α-MEM containing 1% MEM non-essentialamino acids, 2% BME essential amino acids, 10% FCS and0.25 mm pyruvate. Embryo development was evaluated on Day2 (cleavage) and Day 7 (morula and/or blastocyst development).Embryo culture was conducted in a humidified atmosphere with5% CO2, 10% O2 and 85% N2 at 39◦C.
Embryo transferTo evaluate the in vivo competence of cloned eland embryos,morulae and blastocysts were transferred into the uterus of eacheland recipient (n = 2) on Day 7 or 8 of the oestrous cycle.For transfer, epidural analgesia was administered to females
after voluntarily entering the hydraulic restraining chute (Tamer;Fauna Research, Red Hook, NY, USA) and the perivulvulararea was cleaned before entry of the transfer catheter. Embryoswere aspirated into a 0.25-mL plastic straw (AgTech, Manhat-tan, KS, USA) that had approximately 1 cm cut off of the openend before being loaded into a 0.25-mL Cassou artificial insem-ination gun. A plastic outer protective sheath was placed overthe barrel of the Cassou gun after loading. Using rectally guidedassistance, the tip of the gun was passed into the vagina and,as it approached the external cervical os, the tip was pushedthrough the outer sheath. Then, the tip of the Cassou gun waspassed transcervically into the uterus and directed into the uter-ine horn ipsilateral to the ovary with a corpus luteum, where theembryo was deposited. Pregnancy evaluation was performed bytransrectal ultrasonography 45 days after embryo transfer.
Statistical analysisThe Chi-squared test was used to determine any differencesamong semen treatments or somatic cells isolated from the twotypes of ewes. Statistical analysis was performed using GraphpadInstat (Version 3; GraphPad Software, San Diego, CA, USA).P < 0.05 was considered to be statistically significant.
ResultsIsolation, culture and cryopreservation of somatic cellsfrom semen
GCN ram semenInitial cell adhesion (attached) to the collagen-coated culture
surface was observed in fresh (n = 2), cooled (n = 3) and slowfreezing (n = 1) replicates (semen treatment groups), but notfrom the fast freezing treatment (Table 1). Some attached cellsfrom fresh and cooled semen divided and formed proliferatingcolonies (n = 1 and n = 3, respectively) that subsequently pro-duced a monolayer covering approximately 1 cm2 of the surface,whereas attached cells from the slow freezing treatment did notcontinue proliferation.
580 Reproduction, Fertility and Development L. Nel-Themaat et al.
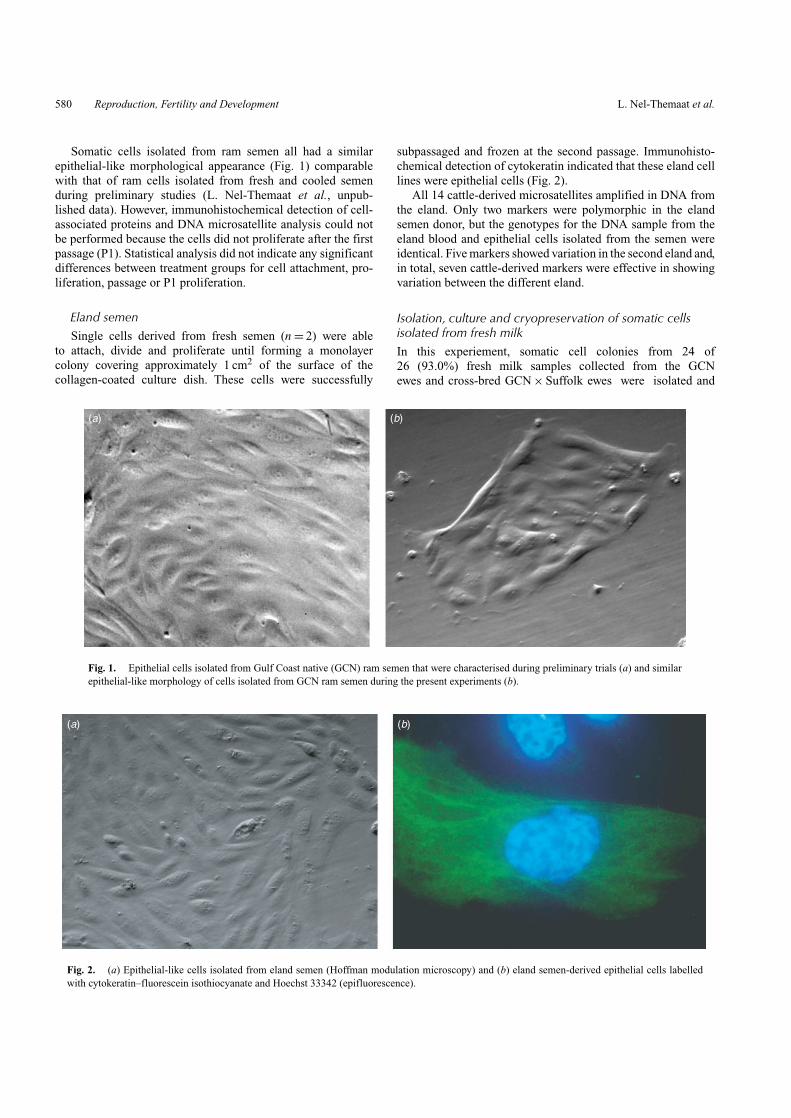
Somatic cells isolated from ram semen all had a similarepithelial-like morphological appearance (Fig. 1) comparablewith that of ram cells isolated from fresh and cooled semenduring preliminary studies (L. Nel-Themaat et al., unpub-lished data). However, immunohistochemical detection of cell-associated proteins and DNA microsatellite analysis could notbe performed because the cells did not proliferate after the firstpassage (P1). Statistical analysis did not indicate any significantdifferences between treatment groups for cell attachment, pro-liferation, passage or P1 proliferation.
Eland semenSingle cells derived from fresh semen (n = 2) were able
to attach, divide and proliferate until forming a monolayercolony covering approximately 1 cm2 of the surface of thecollagen-coated culture dish. These cells were successfully
(a) (b)
Fig. 1. Epithelial cells isolated from Gulf Coast native (GCN) ram semen that were characterised during preliminary trials (a) and similarepithelial-like morphology of cells isolated from GCN ram semen during the present experiments (b).
(a) (b)
Fig. 2. (a) Epithelial-like cells isolated from eland semen (Hoffman modulation microscopy) and (b) eland semen-derived epithelial cells labelledwith cytokeratin–fluorescein isothiocyanate and Hoechst 33342 (epifluorescence).
subpassaged and frozen at the second passage. Immunohisto-chemical detection of cytokeratin indicated that these eland celllines were epithelial cells (Fig. 2).
All 14 cattle-derived microsatellites amplified in DNA fromthe eland. Only two markers were polymorphic in the elandsemen donor, but the genotypes for the DNA sample from theeland blood and epithelial cells isolated from the semen wereidentical. Five markers showed variation in the second eland and,in total, seven cattle-derived markers were effective in showingvariation between the different eland.
Isolation, culture and cryopreservation of somatic cellsisolated from fresh milkIn this experiement, somatic cell colonies from 24 of26 (93.0%) fresh milk samples collected from the GCNewes and cross-bred GCN × Suffolk ewes were isolated and
Somatic cells cultured from semen and milk Reproduction, Fertility and Development 581
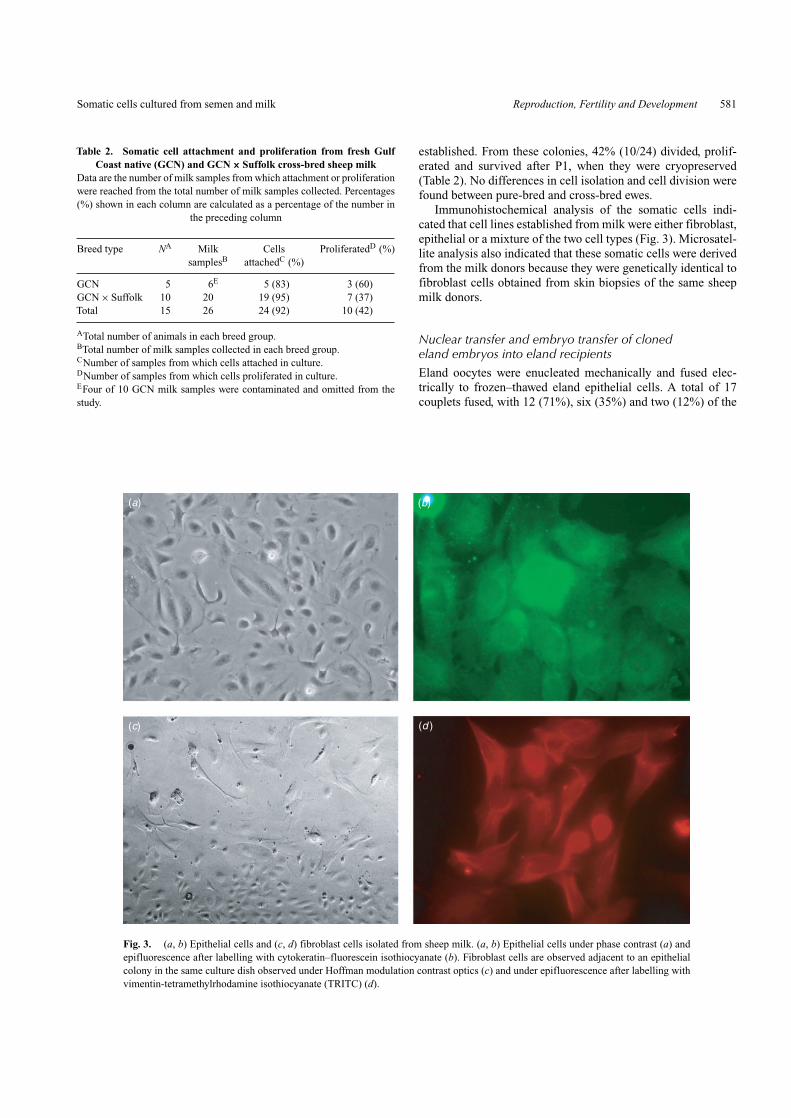
Table 2. Somatic cell attachment and proliferation from fresh GulfCoast native (GCN) and GCN × Suffolk cross-bred sheep milk
Data are the number of milk samples from which attachment or proliferationwere reached from the total number of milk samples collected. Percentages(%) shown in each column are calculated as a percentage of the number in
the preceding column
Breed type NA Milk Cells ProliferatedD (%)samplesB attachedC (%)
GCN 5 6E 5 (83) 3 (60)GCN × Suffolk 10 20 19 (95) 7 (37)Total 15 26 24 (92) 10 (42)
ATotal number of animals in each breed group.BTotal number of milk samples collected in each breed group.CNumber of samples from which cells attached in culture.DNumber of samples from which cells proliferated in culture.EFour of 10 GCN milk samples were contaminated and omitted from thestudy.
(a) (b)
(c) (d )
Fig. 3. (a, b) Epithelial cells and (c, d) fibroblast cells isolated from sheep milk. (a, b) Epithelial cells under phase contrast (a) andepifluorescence after labelling with cytokeratin–fluorescein isothiocyanate (b). Fibroblast cells are observed adjacent to an epithelialcolony in the same culture dish observed under Hoffman modulation contrast optics (c) and under epifluorescence after labelling withvimentin-tetramethylrhodamine isothiocyanate (TRITC) (d).
established. From these colonies, 42% (10/24) divided, prolif-erated and survived after P1, when they were cryopreserved(Table 2). No differences in cell isolation and cell division werefound between pure-bred and cross-bred ewes.
Immunohistochemical analysis of the somatic cells indi-cated that cell lines established from milk were either fibroblast,epithelial or a mixture of the two cell types (Fig. 3). Microsatel-lite analysis also indicated that these somatic cells were derivedfrom the milk donors because they were genetically identical tofibroblast cells obtained from skin biopsies of the same sheepmilk donors.
Nuclear transfer and embryo transfer of clonedeland embryos into eland recipientsEland oocytes were enucleated mechanically and fused elec-trically to frozen–thawed eland epithelial cells. A total of 17couplets fused, with 12 (71%), six (35%) and two (12%) of the

582 Reproduction, Fertility and Development L. Nel-Themaat et al.
Table 3. Embryo development following nuclear transfer of eland epithelial-like cells isolatedfrom semen into enucleated common eland oocytes
Percentages (%) are calculated as a percentage of the number of couplets produced
Eland Fused Cleaved (%) Morulae (%) Blastocysts (%) Embryosrecipient ID couplets (n) transferred (%)
123 10 6 (60) 3 (30) 1 (10) 4 (40)140 7 6 (86) 3 (43) 1 (14) 4 (57)Total 17 12 (71) 6 (35) 2 (12) 8 (47)
embryos cleaving and developing to the morula or blastocyststages, respectively (Table 3).
In total, three morulae (Day 7) and one blastocyst (Day 7)-stage cloned eland embryos were transferred into each of twoeland recipients. No pregnancies were detected at transrectalultrasonography on Day 45 after embryo transfer.
Discussion
In the present study, we successfully isolated somatic cells frommilk and ejaculated semen. To our knowledge, this is the firstreport of somatic cell isolation from semen for in vitro cultureand nuclear transfer.Although we showed that cell isolation fromsemen is possible, cell propagation rates were low. Cells isolatedfrom fresh, cooled and slow-frozen ram semen samples attachedto the surface of the tissue culture dish, whereas cells isolatedfrom fast-frozen semen did not attach. These results are similarto those observed in preliminary studies (L. Nel-Themaat et al.,unpublished data), in which cells were isolated successfully fromfresh and cooled samples, but not from conventionally cryopre-served semen. Therefore, it is possible that the cryopreservationmethod that was used for the ram semen may be detrimental tosomatic cells.
Although DMSO is the most commonly used cryoprotectantfor freezing cultured cells, glycerol is an acceptable cryoprotec-tant for these cells (Luyet and Gonzales 1952; Mazur et al. 1972;Ashwood-Smith and Lough 1975).The most likely reason for ourfailure to isolate live epithelial cells from fast-frozen semen isthe differences in the cooling and warming rates normally usedfor freezing semen and somatic cells. Cooling rates for somaticcells are in the range of 1–5◦C/min and are followed by rapidthawing rates of 150–200◦C/min (Ashwood-Smith 1980). Thefast-frozen semen treatment used a much faster freezing rate(10◦C/min), which is likely not optimal for somatic cell survival.
Furthermore, cryopreservation may sensitise cells to otherstressors, such as dilution of the cryopreservation medium andcentrifugation (Lovelock 1953; Farrant and Morris 1973), oreven to unfavourable culture conditions that cells would nor-mally withstand. The procedure used during the present experi-ment included dilution and three washing steps to ensure that thecryoprotectant was removed and to minimise contamination. It islikely that some epithelial cells survived the freeze–thaw cycle,but were damaged during post-thaw processing. Likewise, a per-centage of ram cells possibly survived both cryopreservationand post-thaw processing but, due to suboptimum culture con-ditions, were unable to attach to the substrate. Therefore, further
studies should be focused on optimising the culture conditionsspecifically for semen-derived epithelial cells.
In preliminary experiments, somatic cells isolated from elandand sable antelope semen were identified as epithelial cells. Theexact origin of the somatic cells is not known, but is possiblya mixture of tubule epithelial cells that were sloughed off fromthe accessory glands, seminiferous tubes and the vas deferens.In the present study, the morphology of ram cells was similarin appearance to that of the previously identified ram, eland andsable antelope epithelial cells (Figs 1, 2).
A multitude of factors affects the growth of epithelial cellsin vitro. Epithelial cells require more complex culture systemsthan fibroblast cells, such as substrate modification, additionalmedium supplementation and different dissociation techniques(Freshney and Freshney 2002). In our experiments, collagen-coated dishes were used to support growth in vitro. Althoughattachment and some proliferation were observed, suboptimalculture conditions may have contributed to the low cell prolifer-ation. Another aspect that may be affecting the initial attachmentis the presence of motile spermatozoa. Sperm motility was main-tained for over 24 h after the initiation of culture and this mayhave physically prevented cells from attaching to the culturesurface. Therefore, further experiments should be conducted toimprove the culture system and enhance separation of somaticcells from spermatozoa before culture. This may enhance cellattachment to the substrate and support the proliferation ofepithelial cells.
Results from the microsatellite analysis on eland epithelialcells correspond to those from preliminary experiments withram semen, where semen-derived ram epithelial cells were geno-typically identical to fibroblast cells from the semen donor.It is important to confirm the species of origin, because eggyolk could potentially be a source of chicken somatic cellcontamination in the semen extender.
We found that success rates in isolating and propagatingsomatic cells from milk far exceeded results obtained withsemen. One possible explanation is that the volume of milk usedwas typically between 30 and 40 mL, whereas the average vol-ume of the ram ejaculate was between 1 and 2 mL. Therefore,we expected that the starting cell numbers harvested from milkshould be larger than that of cells from semen. Furthermore,epithelial cells, fibroblast cells and a mixture of cell types wereisolated from milk. The presence of fibroblast cells in severalof the isolated cultures and the uniform fibroblast cell culturesthat were occasionally obtained should increase the likelihoodof obtaining somatic cell attachment and proliferation from milk

Somatic cells cultured from semen and milk Reproduction, Fertility and Development 583
because the culture system is already optimised for this cell type.In mixed cell cultures, the fibroblast cells may actually serve asfeeder cells for the epithelial cells.
One successful method that is widely used for enhancingepithelial culture conditions is coculture with inactivated fibrob-last cells (Macpherson and Bryden 1971). In addition to servingas an attachment substrate and a source of continuously secretedgrowth factor, feeder cells may inactivate the inhibitory action ofserum-derived transforming growth factor (TGF)-β on epithe-lial cells (Tucker et al. 1984; Moses et al. 1985; Masui et al.1986). Therefore, this effect could be partially responsible forthe higher success rates in milk cell proliferation. In caseswhere only fibroblast cells attached, low initial cell numbersmay be detrimental to proliferation. In contrast, epithelial cellsappear tolerant to low initial numbers and proliferate muchfurther than fibroblasts with similarly low starting cell num-bers. Highly specific culture procedures have been describedfor human mammary epithelial cells (Stampfer et al. 2002) andincorporating these techniques to culture cells isolated from milkshould improve proliferation rates. Although our system was notoptimised for this specific cell type, we demonstrated that milkfrom non-dairy animals could be a source of somatic cells forin vitro culture. Ideally, the cell type of each primary culture thatis obtained from milk or semen should be identified and the cul-ture system modified accordingly based on existing knowledgeof fibroblast and epithelial cell culture.
The DNA microsatellite analysis confirmed that cells fromsemen and milk were from the respective donors, which impliesthat cells could be used for innumerable genetic studies and beincorporated in GRBs. In addition, bovine microsatellite mark-ers proved to be effective in the eland. Ultimately, isolated cellscould serve as karyoplast donors during nuclear transfer proce-dures. Our nuclear transfer results indicated that eland epithelialcells isolated from semen could direct early embryonic develop-ment after transfer into eland cytoplasts. Although two embryosdeveloped to blastocysts in vitro, no pregnancies were estab-lished after the transfer of cloned eland blastocysts into elandrecipients.
Culture methods have not been optimised for in vitro-produced eland embryos (Wirtu 2004). To our knowledge, elandpregnancies have not been established following in vitro embryoproduction. Therefore, the inability to establish pregnanciescould be attributed to suboptimal conditions that these embryoswere exposed to before embryo transfer. Even in domestic cat-tle, where extensive research has been ongoing for years andin vitro culture to the blastocyst stage has been relatively suc-cessful (approximately 40%), success rates of producing liveoffspring following nuclear transfer in many species (i.e. cowsand sheep) remain less than 4% (Wilmut et al. 2002). There-fore, it is not surprising that pregnancies were not establishedfrom the eland, of which still so little is known. Future researchshould focus on optimising in vitro protocols specifically foreland embryo production and culture.
In conclusion, we have described a novel technique forobtaining somatic cells from semen and their subsequent usefor in vitro cell culture. The isolation of somatic cells fromcryopreserved semen could have valuable implications for criti-cally endangered species. Potentially, a clone could be produced
from a deceased but genetically valuable animal using cellsobtained from frozen semen and, subsequently, additional off-spring produced by naturally breeding the resulting clonedoffspring.
AcknowledgementsThe authors extend their gratitude to Tray Harding and the LSU AgriculturalCenter Sheep Research Station team for providing the sheep and helping withsample collection. The authors also acknowledge Dr Alex Cole, ContessaKnight and the animal keepers from the Audubon Research Center SpeciesSurvival Center for helping with the eland semen collections.
ReferencesArase, S., Sadamoto, Y., Kato, S., Fujie, K., Nakanishi, H., and Takeda, K.
(1990). Three-dimensional culture of plucked human hair follicles insidethe collagen gel matrix. Nippon Hifuka Gakkai Zasshi 100, 879–882.
Ashwood-Smith, M. J. (1980). Low temperature preservation of cells, tissuesand organs. In ‘LowTemperature Preservation in Medicine and Biology’.(Eds M. J. Ashwood-Smith and J. Farrant.) pp. 19–44. (Pitman Medical:Kent.)
Ashwood-Smith, M. J., and Lough, P. (1975). Cryoprotection of mam-malian cells in tissue culture with methanol. Cryobiology 12, 517–518.doi:10.1016/0011-2240(75)90034-6
Bishop, M. D., Kappes, S. M., Keele, J. W., Stone, R. T., Sunden, S. L.,Hawkins, G. A., Toldo, S. S., Fries, R., Grosz, M. D., and Yoo, J. (1994).A genetic linkage map for cattle. Genetics 136, 619–639.
Blackburn, H. D. (2004). Development of national animal genetic resourceprograms. Reprod. Fertil. Dev. 16, 27–32. doi:10.1071/RD03075
Creighton, P., Eggen, A., Fries, R., Jordan, S. A., Hetzel, J.,Cunningham, E. P., and Humphries, P. (1992). Mapping of bovinemarkers CYP21, PRL, and BOLA DRBP1 by genetic linkage analysisin reference pedigrees. Genomics 14, 526–528. doi:10.1016/S0888-7543(05)80260-5
Elicha Gussin, H. A., and Elias, S. (2002). Culture of fetal cells from mater-nal blood for prenatal diagnosis. Hum. Reprod. Update 8, 523–527.doi:10.1093/HUMUPD/8.6.523
Evans, G., and Maxwell, W. M. C. (1987). ‘Artificial Insemination of Sheepand Goats.’ (Butterworths: Sydney.)
Evenson, D. P., and Melamed, M. R. (1983). Rapid analysis of normaland abnormal cell types in human semen and testis biopsies by flowcytometry. J. Histochem. Cytochem. 31, 248–253.
Farrant, J., and Morris, G. J. (1973). Thermal shock and dilution shock as thecauses of freezing injury. Cryobiology 10, 134–140. doi:10.1016/0011-2240(73)90019-9
Freshney, R. I., and Freshney, M. G. (Eds) (2002). ‘Culture of EpithelialCells.’ (Wiley-Liss: New York.)
Gómez, M. C., Pope, C. E., Giraldo,A., Lyons, L.A., Harris, R. F., King,A. L.,Cole,A., Godke, R.A., and Dresser, B. L. (2004). Birth ofAfrican wildcatcloned kittens born from domestic cats. Cloning Stem Cells 6, 247–258.
Green, A. E. (1967). Viability of cell cultures following extended preserva-tion in liquid nitrogen. Proc. Soc. Exp. Biol. Med. 124, 1302–1307.
Janssen, D. L., Edwards, M. L., Koster, J. A., Lanza, R. P., andRyder, O. A. (2004). Postnatal management of chryptordich bantengcalves cloned by nuclear transfer utilizing frozen fibroblast culturesand enucleated cow ova. Reprod. Fertil. Dev. 16, 224. [Abstract]doi:10.1071/RDV16N1AB206
Katska, L., Bochenek, M., Kania, G., Rynska, B., and Smorag, Z. (2002).Flow cytometric cell cycle analysis of somatic cells primary cul-tures established for bovine cloning. Theriogenology 58, 1733–1744.doi:10.1016/S0093-691X(02)01043-9
Kishi, M., Itagaki, Y., Takakura, R., Imamura, M., Sudo, T., Yoshinari, M.,Tanimoto, M., Yasue, H., and Kashima, N. (2000). Nuclear

584 Reproduction, Fertility and Development L. Nel-Themaat et al.
transfer in cattle using colostrum-derived mammary gland epithelialcells and ear-derived fibroblast cells. Theriogenology 54, 675–684.doi:10.1016/S0093-691X(00)00382-4
Kossarek, L. M., Grosse, W. M., Finco, T. S., and McGraw, R. A.(1993a). Rapid communication: bovine dinucleotide repeat polymor-phism RM002. J. Anim. Sci. 71, 3173.
Kossarek, L. M., Su, X., Grosse, W. M., Finlay, O., Barendse, W., Hetzel, D. J.,Dietz, A. B., Womack, J. E., and McGraw, R. A. (1993b). Rapid com-munication: bovine dinucleotide repeat polymorphism RM067. J. Anim.Sci. 71, 3178.
Krisher, R. L., and Bavister, B. D. (1999). Enhanced glycolysis after matura-tion of bovine oocytes in vitro is associated with increased developmentalcompetence. Mol. Reprod. Dev. 53, 19–26. doi:10.1002/(SICI)1098-2795(199905)53:1<19::AID-MRD3>3.0.CO;2-U
Kurata, S., Itami, S., Terashi, H., and Takayasu, S. (1994). Successful trans-plantation of cultured human outer root sheath cells as epithelium. Ann.Plast. Surg. 33, 290–294. doi:10.1097/00000637-199409000-00009
Lanza, R. P., Cibelli, J. B., Diaz, F., Moraes, C. T., Farin, P. W., Hammer, C. J.,West, M. D., and Damiani, P. (2000). Cloning of an endangered species(Bos gaurus) using interspecies nuclear transfer. Cloning 2, 79–90.doi:10.1089/152045500436104
Loi, P., Ptak, G., Barboni, B., Fulka, J. Jr, Cappai, P., and Clinton, M. (2001).Genetic rescue of an endangered mammal by cross-species nuclear trans-fer using post-mortem somatic cells. Nat. Biotechnol. 19, 962–964.doi:10.1038/NBT1001-962
Lovelock, J. E. (1953). The haemolysis of human red blood cells by freezingand thawing. Biochim. Biophys. Acta 10, 414–426. doi:10.1016/0006-3002(53)90273-X
Luyet, B. J., and Gonzales, F. (1952). Protective action of glycerol againstfreezing injury in embryonic tissues of chick. Biodynamica 7, 101–106.
Macpherson, I., and Bryden,A. (1971). Mitomycin C treated cells as feeders.Exp. Cell Res. 69, 240–241. doi:10.1016/0014-4827(71)90335-1
Masui, T., Wakefiels, J. F., Lechner, M. A., LaVech, M. B., Sporn, M. B.,and Harris, C. C. (1986). Type beta transforming growth factor isthe primary differentiation-inducing serum factor for normal humanbronchial epithelial cells. Proc. Natl Acad. Sci. USA 83, 2438–2442.doi:10.1073/PNAS.83.8.2438
Mazur, P., Leibo, S. P., and Chu, E. H. Y. (1972). A two-factor hypothesisof freezing injury. Exp. Cell Res. 71, 345–355. doi:10.1016/0014-4827(72)90303-5
http://www.publish.csiro.au/journals/rfd
Moll, I. (1996). Differential epithelial outgrowth of plucked and microdis-sected human hair follicles in explant culture. Arch. Dermatol. Res. 288,604–610.
Moses, H. L., Tucker, E. B., Leof, R. J., Coffey, R. J., Halper, J., andShipley, G. D. (1985). Type beta transforming growth factor is a growthstimulator and growth inhibitor. Cancer Cells 3, 67–71.
Paape, M. J., and Tucker, H. A. (1966). Somatic cell content variation infraction-collected milk. J. Dairy Sci. 49, 265–267.
Ryder, O. A. (2002). Cloning advances and challenges for conservation.Trends Biotechnol. 20, 231–232. doi:10.1016/S0167-7799(02)01954-6
Ryder, O. A., and Benirschke, K. (1997). The potential use of ‘cloning’ inthe conservation effort. Zoo Biol. 16, 295–300. doi:10.1002/(SICI)1098-2361(1997)16:4<295::AID-ZOO1>3.0.CO;2-5
Stampfer, M. R., Yaswen, P., and Taylor-Papadimitriou, J. (2002). Cultureof human epithelial cells. In ‘Culture of Epithelial Cells’. (Eds R. I.Freshney and M. G. Freshney.) pp. 95–135. (Wiley-Liss: New York.)
Toonen, R. J., and Hughes, S. (2001). Increased throughput for fragmentanalysis on an ABI PRISM 377 automated sequencer using a membranecomb and STRand software. Biotechniques 31, 1320–1324.
Tucker, R. F., Shipley, G. D., Moses, H. L., and Holley, R. W. (1984).Growth inhibitor from BSC-1 cells closely related to platelet typebeta transforming growth facor. Science 226, 705–707. doi:10.1126/SCIENCE.6093254
Wilmut, I., Schnieke, A. E., McWhir, J., Kind, A. J., and Campbell, K. H. S.(1997). Viable offspring derived from fetal and adult mammalian cells.Nature 385, 810–813. doi:10.1038/385810A0
Wilmut, I., Beaujean, N., de Sousa, P. A., Dinnyes, A., King, T. J.,Paterson, L. A., Wells, D. N., and Young, L. E. (2002). Somatic cellnuclear transfer. Nature 419, 583–586. doi:10.1038/NATURE01079
Wirtu, G. (2004). Developing embryo technologies for the eland ante-lope (Taurotragus oryx). Ph.D. Thesis, Louisiana State University andAgricultural and Mechanical College, USA.
Wirtu, G., Pope, C. E., Lee, B. C., Cole, A., Vaccaro, J., Sarrat, E. L., Bavis-ter, B. D., Godke, R. A., and Dresser, B. L. (2002). Collection of semenby rectal massage from an eland (Taurotragu oryx) bull. In ‘Proceedingsof the 2nd International Symposium on Assisted Reproductive Technol-ogy (ART) for the Conservation and Genetic Management of Wildlife’.pp. 318–320. (Omaha Zoo: Nebraska.)
Manuscript received 8 November 2006, accepted 19 March 2007




















